
Submitted:
16 May 2023
Posted:
18 May 2023
You are already at the latest version
Abstract
The article presents data on phylogeny, genome size, and ploidy of species of the genus Kalidium Moq. in the flora of Kazakhstan. Genus Kalidium belongs to the tribe Salicornieae of the subfamily Salicornioideae of the family Chenopodiaceae and unites 8 species, the main range of which covers the Iranian-Turanian and Central Asian deserts. There are 4 species in the flora of Kazakhstan: K. foliatum, K. caspicum, K. schrenkianum and the recently described K. juniperinum. Populations of species of the genus Kalidium in the saline deserts of Kazakhstan occupy large areas, often forming monodominant communities. Sometimes there is a joint growth of 2 and very rarely 3 species of the genus. During the period of fieldwork (2021-2022), populations were identified in which these species grew together with a predominance, in most cases, of K. caspicum. Samples of representatives from 15 populations were collected for research. Selected plant samples were studied by flow cytometry to determine plant ploidy. Sequencing of nrITS and two chloroplast fragments were used to build a phylogenetic tree, including sequences from the NCBI database., A phylogenetic tree of species of the genus Kalidium was compiled which considers previously published data. In the valley of the middle reaches of the Syrdarya River tetraploid populations of K. caspicum were found. A hybrid between K. foliatum and K. caspicum was found in the Ili River valley (Almaty region, Uigur district). To identify phylogenetic processes at the intraspecific level, the SCoT (Start codon targeted) fingerprinting method was used.
Keywords:
Chenopodiaceae
; Salicornioideae
; Salicornieae
; Kazakhstan
; hybrid
; tetraploid
1. Introduction
The genus Kalidium Moq. belongs to the tribe Salicornieae, subfamily Salicornioideae [1,2] of the largest family, Chenopodiaceae (Amaranthaceae s. lato), in the Kazakhstan part of the North Turan deserts [3]. Species of the genus Kalidium are euhalophytes [4] or halo-succulents, which are halophytes with relatively fleshy, succulent stems or leaves [5,6]. Until recently, the genus was divided into six species: Kalidium foliatum (Pall.) Moq., K. caspicum (L.) Ung.-Sternb., K. cuspidatum (Ung.-Sternb.) Grubov, K. gracile Fenzl, K. schrenkianum Bunge and K. wagenitzii (Aellen) Freitag & G. Kadereit. In 2020, the subspecies K. cuspidatum var. sinicum A.J. Li [7] was reclassified to species K. sinicum (A. J. Li) by H.C. Fu and Z.Y. Chu [8], and at the end of 2022, an eighth species of this genus, K. juniperinum Sukhor. & Lomon [9], was added. The main range of the genus covers the Irano–Turanian and Central Asian deserts [1,10].
Members of the genus Kalidium are small or dwarf glabrous shrubs, mostly with reduced, semi-amplex leaf laminae. The species K. foliatum, K. wagenitzii, and K. juniperinum are found with roll-shaped (terete) and succulent leaf laminae up to 1.2 cm long [9]. The peduncles consist of three submerged flowers with one or two stamens and united segments with four or five teeth. The fruit has a parenchymatous pericarp and a thin, yellow or brownish seed coat with a fine papillary surface [9].
Species of the genus Kalidium play an important role in maintaining the balance of grassland ecosystems and preventing soil erosion [11]. Comparative studies have shown that, as the dominant species in desert areas, species of the genus are highly tolerant to saline and alkaline soils as well as to drought [11,12]. As succulents, they are mainly used as winter fodder for camels, horses, and sheep [13]. In addition, Wang and Jia [14] showed that ethanol crude extracts of aerial parts of K. foliatum have high antibacterial activity.
A review of the literature has shown that most of the scientific work relating to the study of Kalidium species is aimed at investigating the topical phenomenon of salt tolerance. Among the species particularly well adapted to saltwater habitats are K. foliatum and K. caspicum [14,15,16,17,18,19,20,21].
A few works are related to studies of the genus’s systematics. Some papers contain information on the scope of the tribe and subfamily, in which two tribes are combined into one: Salicornieae [1,2]. A recently published paper by Chinese scientists provides information on the DNA barcoding of species of the genus Kalidium and substantiates the independence of the species K. sinicum [8]. This paper presents the results of a study of species of the genus Kalidium from Kazakhstan, also at the molecular genetic level, but from other regions of the world, allowing for comparative analysis.
For one species, K. foliatum, a full-genome analysis of chloroplast DNA has already been carried out, revealing that it is phylogenetically related to two species of Salicornia, S. bigelovii Torr. and S. brachiata Roxb. [22].
In most cases, species of the genus Kalidium are used as an outgroup for various research works on other genera of the tribe Salicornieae [23,24,25]. There are also papers on the morphology of pollen from the Salicornieae tribe, including classification [26].
According to data in the literature, the genus Kalidium is generally represented by three species in Kazakhstan: K. caspicum, K. foliatum, and K. schrenkianum [27,28,29,30,31,32]. However, in 2022, A.P. Sukhorukov and M.N. Lomonosova described a new species: K. juniperinum Sukhor. & Lomon., occurring mostly in the central and northern parts of Kazakhstan [9]. Unfortunately, this work was published after we had conducted our molecular genetic analysis, so we did not analyze K. juniperinum material.
The relevance of this study lies in the fact that populations of Kalidium species in the saline deserts of Kazakhstan occupy large areas, while very few other plants can grow under such conditions. It should be noted that species of the genus form monodominant communities in most cases, with only occasional cooccurrences of two and very rarely three species. Two species cannot usually dominate together, except under special circumstances where there is exposure to external, usually anthropogenic, influences. Additionally, according to our observations, two species growing together will hybridize. However, given the specific morphological structure of the Kalidium species, it is virtually impossible to visually identify hybrid plants in the field.
In the desert part of the Syrdarya valley, the dominant species of the genus Kalidium that form large communities include K. caspicum and K. foliatum. During the fieldwork period (2021–2022), populations were identified in which both species occurred, with K. caspicum predominating in most cases. Monopopulations dominated only by K. caspicum and covering a large area were also observed. In these populations, individuals of K. caspicum were well developed and reached a larger size than in the other populations but did not differ morphologically.
The original plant samples were studied by flow cytometry. One of the most common uses of this method is to study hybridogenic processes manifested as polyploidy and aneuploidy [33]. The discovery of such hybrid and polyploid specimens in our samples necessitated additional analyses. It was decided to use molecular genetics techniques, analyzing internal transcribed spacer (ITS) and chloroplast fragments to more accurately identify hybrid and polyploid samples.
One of the most important molecular methods in the study of relatedness in supraspecific systematics is the comparison and analysis of aligned DNA sequences of individual genome fragments and plant plastomes. Analysis of ITS ribosomal DNA (ITS nrDNA) is the most popular for genome fragment studies (nuclear DNA), while the plastome uses a wide range of genes and introns [34].
2. Results
As a result of expeditionary trips, samples of Kalidium representatives from 15 populations were collected for the research. This article, however, presents the results of flow cytometry studies of the samples examined in only 11 populations. Unfortunately, the pattern obtained from the analysis of samples from four populations (including a sample of K. schrenkianum species from the Almaty region) did not allow it to be interpreted accordingly. We believe this is due to the accumulation of significant amounts of metabolites in the cells of these plant samples because of the late period (September) of their collection.
Based on a summary and critical analysis of the data we obtained during our studies, including sequences from the NCBI database using QGIS 2.14 software (https://qgis.org), a point distribution map of the samples studied was produced (Figure 1).
2.1. Flow cytometry
2.2. Molecular phylogeny
Internal transcribed spacer (nrITS) and chloroplast (trnQ-rpS16 and trnL(UAG)-rpL32) fragments were sequenced for 15 populations of three species: K. caspicum, K. foliatum, and K. schrenkianum, as shown in Figure 3 and Figure 4. The ITS fragments for populations B10, B11, B12, and B13 were each made of two samples, and for B14 of three samples.
According to the ITS phylogenetic tree (Figure 2), the species of the genus Kalidium are divided into two large groups. The first group includes K. foliatum, K. gracile, K. wagenitzii, K. sinicum (K. cuspidatum var. sinicum), and K. cuspidatum var. cuspidatum. The second group comprises the species K. caspicum and K. schrenkianum (Figure 2). Similarly, in the first group, the species K. sinicum clearly (100%) diverges from the other species, K. foliatum, K. gracile, and K. cuspidatum. Furthermore, among the three species mentioned above, bootstrap support was 83%, and between K. cuspidatum and K. gracile 73%.
Most of the K. foliatum specimens we studied (B02, B05, B06, and B13.2) were arranged quite predictably, although specimen B13 was positioned among the specimens of K. caspicum, relatively distant from K. foliatum in the ITS phylogenetic tree. Most taxa form clear monophyletic clades in the ITS tree, but some sequences from GenBank have a questionable position. That concern accessions KU975203 K. schrenkianum; HM131637 – K. cuspidatum; DQ340146 – Kalidiopsis wagenitzii.
Combined plastid tree (trnQ-rps16 + rpl32-trnL) only with own sequences and with Halochnemum strobilaceus and Halostachys belangeriana as outgroup show only two well-supported clades (Figure 3): K. caspicum including K. schrenkianum and K. foliatum. Caspicum clade is divided into three subclades with good support, more or less by geographic origin. It is interesting that the plastid sequences of K. schrenkianum accession B15 are similar with sequences of three accessions of K. caspicum (B04, B14 and B12).
Figure 2.
ITS tree of the genus Kalidium. The joint presence of Bayesian with a probability greater than 0.98 and bootstrap support greater than 95% is indicated by a black dot. The putative Cuspidatum group is marked in blue and the Caspicum group in red. For visual convenience, the species boundaries are highlighted in grey. The specimens we studied are shown in bold. Hybrid B13 Kalidium foliatum is marked in yellow. Samples with unclear locations in the tree are highlighted in green. When running the data through the JModeltest program, the following data is obtained: GTR+G, -lnL 1871.40999, AIC 4232.819980.
Figure 2.
ITS tree of the genus Kalidium. The joint presence of Bayesian with a probability greater than 0.98 and bootstrap support greater than 95% is indicated by a black dot. The putative Cuspidatum group is marked in blue and the Caspicum group in red. For visual convenience, the species boundaries are highlighted in grey. The specimens we studied are shown in bold. Hybrid B13 Kalidium foliatum is marked in yellow. Samples with unclear locations in the tree are highlighted in green. When running the data through the JModeltest program, the following data is obtained: GTR+G, -lnL 1871.40999, AIC 4232.819980.
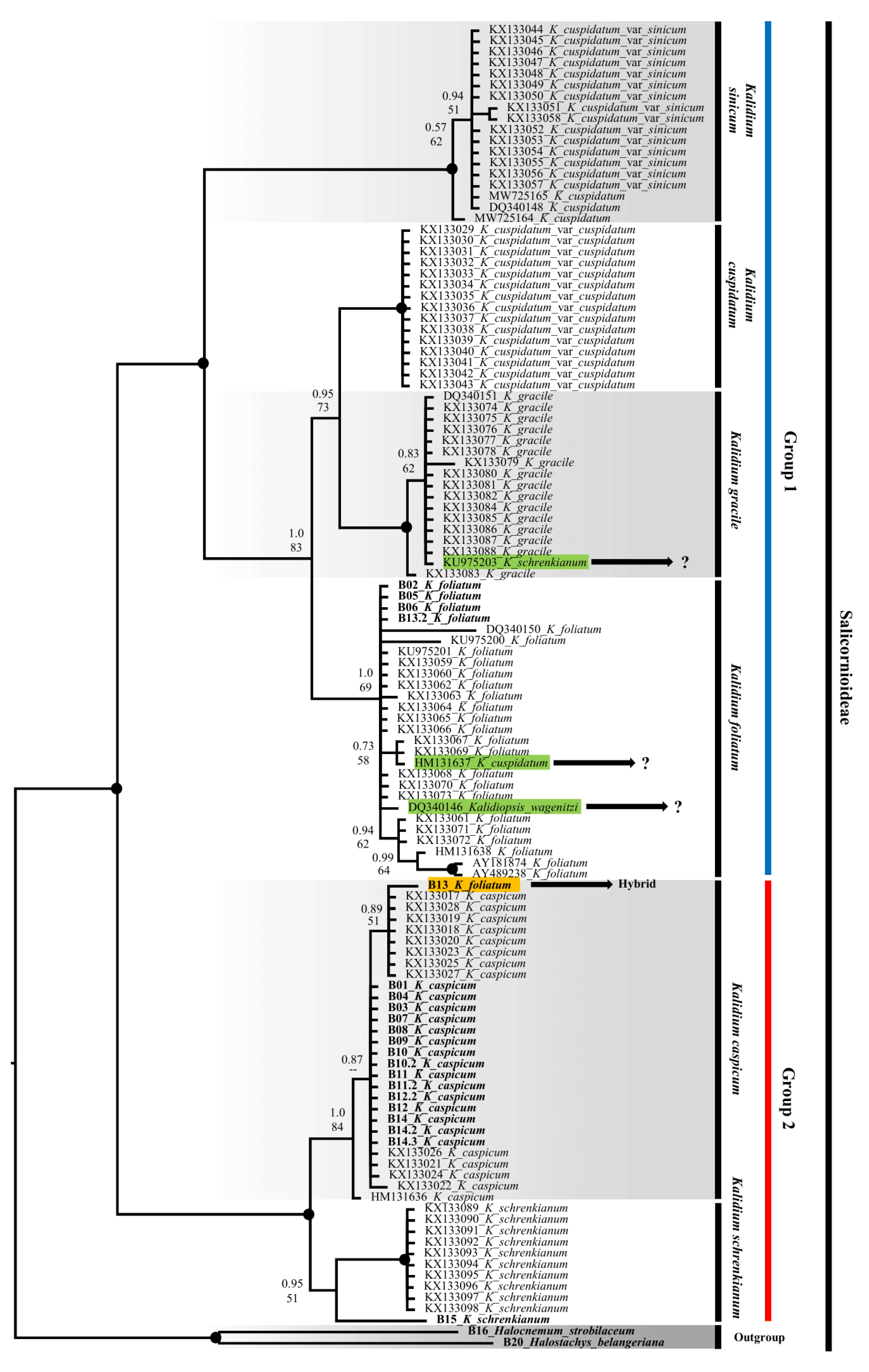
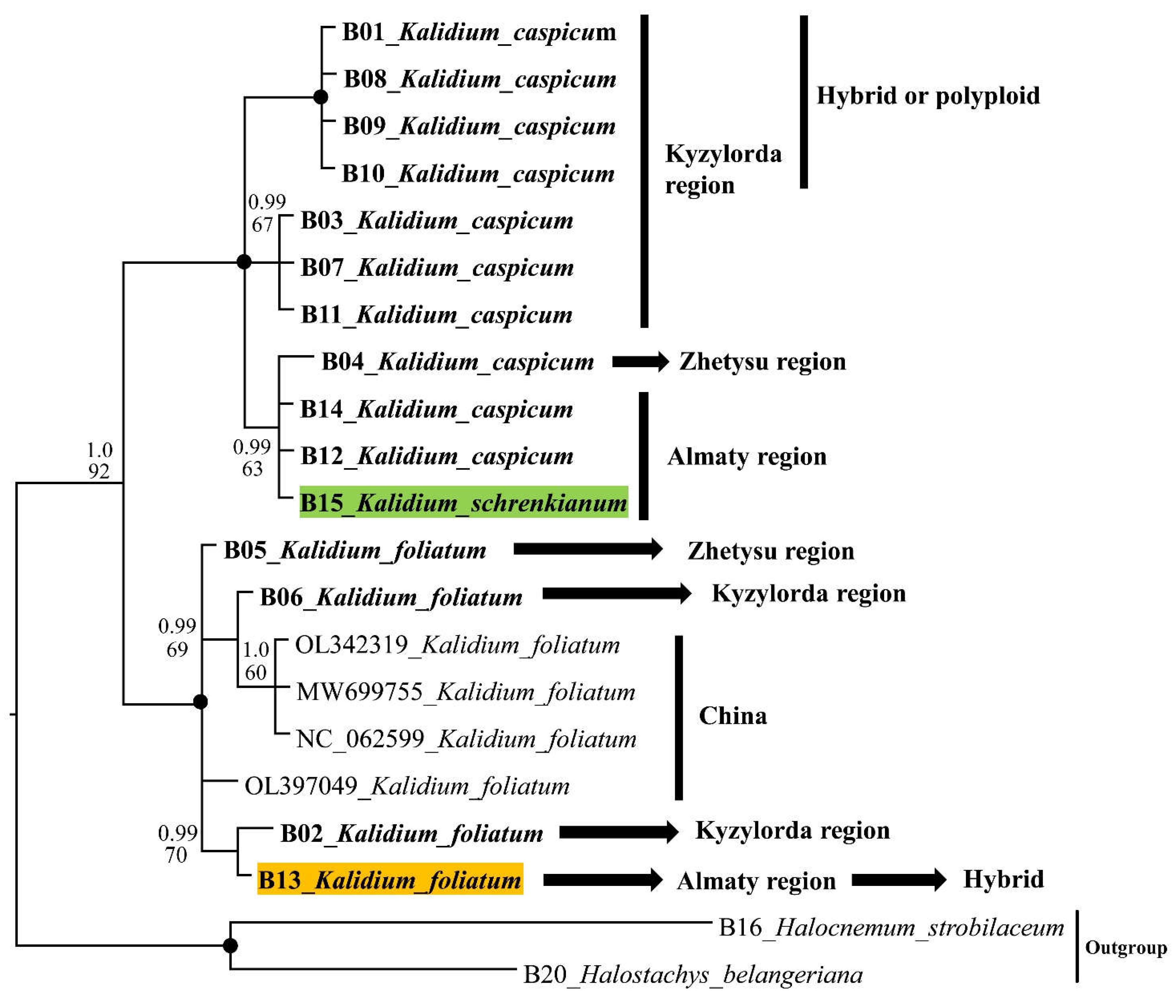
Figure 3.
Plastiden Tree (trnQ-rps16 and rpl32-trnL) of the genus Kalidium. The joint presence of Bayesian with probability greater than 0.98 and bootstrap support greater than 95% is indicated by a black dot. Bold type indicates the specimens that we have isolated. Hybrid B13 Kalidium foliatum is marked in yellow. Specimen B15 of Kalidium schrenkianum, whose position in the tree is not clear, is highlighted in green. The arrows and lines show the areas from where these samples were taken.
Figure 3.
Plastiden Tree (trnQ-rps16 and rpl32-trnL) of the genus Kalidium. The joint presence of Bayesian with probability greater than 0.98 and bootstrap support greater than 95% is indicated by a black dot. Bold type indicates the specimens that we have isolated. Hybrid B13 Kalidium foliatum is marked in yellow. Specimen B15 of Kalidium schrenkianum, whose position in the tree is not clear, is highlighted in green. The arrows and lines show the areas from where these samples were taken.
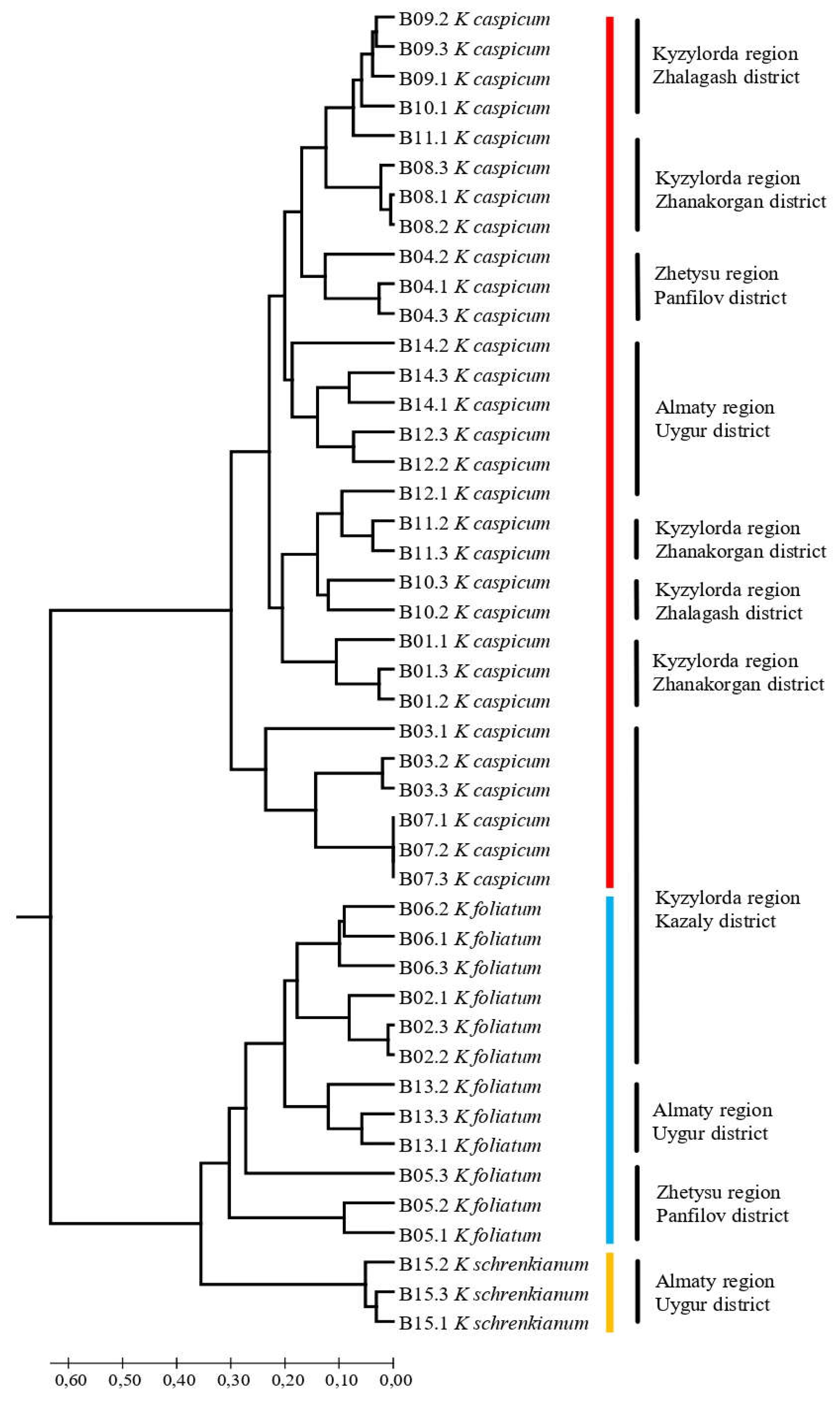
The following data were obtained by running the data through the JModeltest software: TVM+G, -lnL 2301.19646, AIC 4698.392920.
Figure 4.
UPGMA tree based on SCoT data of the three species of the genus Kalidium with area and region of selected samples. Origin of the accession see in supplement A.
Figure 4.
UPGMA tree based on SCoT data of the three species of the genus Kalidium with area and region of selected samples. Origin of the accession see in supplement A.
2.3. SCoT results
For SCoT analysis, 3 samples were collected from each population. Thus, with a total of 15 populations studied, a total of 45 samples were analyzed. Initially, 10 SCoT primers were used: SCoT2, SCoT4, SCoT11, SCoT12, SCoT13, SCoT14, SCoT16, SCoT17, SCoT21, SCoT23. However, only 6 primers gave good results: SCoT11, SCoT12, SCoT13, SCoT14, SCoT21, SCoT23 (Appendix С). A UPGMA tree of the three species of the genus Kalidium was constructed from the resulting matrix using the MEGA 7.0 software (Figure 4). In the UPGMA tree with SCoT matrix data, the accessions are clearly classified by origin. In addition, SCoT analysis of the UPGMA tree showed some variation in the proximity data for samples B10.1, B11.1, and B12.1. Thus, it appears that samples B10.1 and B11.1 are close to the B09 population and B12.1 to the B11 population.
The three accessions of K. schrenkianum are classified as a sister group to K. foliated. Also, in the PCA analysis based on SCoT data (Supplements E; F; G) species are clearly separated.
3. Discussion
According to the phylogenetic tree (ITS), the species of the genus Kalidium are divided into two large groups. The first group includes K. foliatum, K. gracile, K. wagenitzii, K. sinicum (K. cuspidatum var. sinicum), and K. cuspidatum var. cuspidatum. The second group comprises the species K. caspicum and K. schrenkianum (Figure 3). These figures are in good agreement with those cited by Liang and Wu [13]. Similarly, in the first group, the species K. sinicum clearly (100%) diverges from the other species, K. foliatum, K. gracile, and K. cuspidatum. Furthermore, among the three species mentioned above, bootstrap support was 83%, and between K. cuspidatum and K. gracile 73%. These values are close to those obtained by Liang and Wu [13].
Recall that two variations, K. cuspidatum var. sinicum and K. cuspidatum var. cuspidatum, were previously identified within K. cuspidatum [43]. The first of these is now regarded as an independent species of K. sinicum by Liang and Wu [13]. Because the program allocated the specimens named K. cuspidatum (MW725164, MW725165, DQ340148) in the group to K. sinicum, they most likely belong to K. cuspidatum var. sinicum, while the specimens of K. cuspidatum var. cuspidatum represent K. cuspidatum proper.
According to the ITS tree, K. gracile is closely related to K. cuspidatum. Unfortunately, we did not see the herbarium specimen K. schrenkianum (KU975203) from the Kyzylorda deserts, which was found by H. Freitag and S. Rilke (26500) in the vicinity of Novokazalinsk (now Aitekebi) in the Kazaly district of Kazakhstan [1,2]. Therefore, we cannot explain why this specimen fell into the K. gracile species group.
We now turn to the results obtained for K. foliatum, which has the most extensive range of all species in the genus, and accordingly has several questions related to its phylogeny. Most of the K. foliatum specimens we studied (B02, B05, B06, and B13.2) were arranged quite predictably, although specimen B13 was positioned among the specimens of K. caspicum, relatively distant from K. foliatum in the phylogenetic tree (ITS). However, it should be considered that our K. foliatum specimens were growing on the edge of a large population of K. caspicum, near a road slope where there was a change in microrelief.
To clarify the results obtained, we sequenced another sample of K. foliatum (13.2) from this population, resulting in its placement in its own sample group. When the results of ITS and the chloroplast tree were compared (Figure 4), B13 appeared to be in the K. foliatum group, revealing the hybrid nature of this sample.
Without further research, it is not possible to explain why the only K. wagenitzii, DQ340146 Kalidiopsis wagenitz, H. Freitag 28801 [1,2], became included in the K. foliatum sample group in the resulting phylogenetic tree. A detailed study of this issue is highly relevant, as K. wagenitzii is considered endemic to the territory of Turkey.
Another specimen, K. cuspidatum (HM131638), once used as an outgroup [24], fell into the K. foliatum group.
In the second group of species, K. caspicum and K. schrenkianum, analysis of specimens of K. schrenkianum showed a standard result, with little intraspecific variation (ITS), most likely related to the geographical location of their habitat (Liang and Wu 2017) [13]. When the chloroplast DNA sequences were analyzed, the K. schrenkianum sample (B15) was found to be a member of the K. caspicum group (Figure 4). It should be noted, however, that the population of K. schrenkianum (which consisted of no more than 20 specimens) grew within a large population of K. caspicum. This circumstance suggests a hybrid origin of the specimen K. schrencianum (B15). Unfortunately, we did not have enough material in our own collections and resources in the NCBI database to confirm or disprove this version.
Analysis of K. caspicum by ITS fragments showed a standard arrangement in the phylogenetic tree (Figure 2), but analysis by chloroplast fragments showed that the species was distributed according to collection sites (Figure 3). Given that the specimens studied were collected from different areas located at different hypsometric heights, they differed quite well in the phylogenetic tree. Thus, samples of K. caspicum (B01, B03, B07, B08, B09, B10, and B11) collected in the Kyzylorda region grew at absolute altitudes between 60 m and 160 m above sea level. Samples B12 and B14, from the Almaty region, grew at altitudes between 530 m and 620 m, while B04, from the Zhetysu region, grew at an altitude of 1010 m.
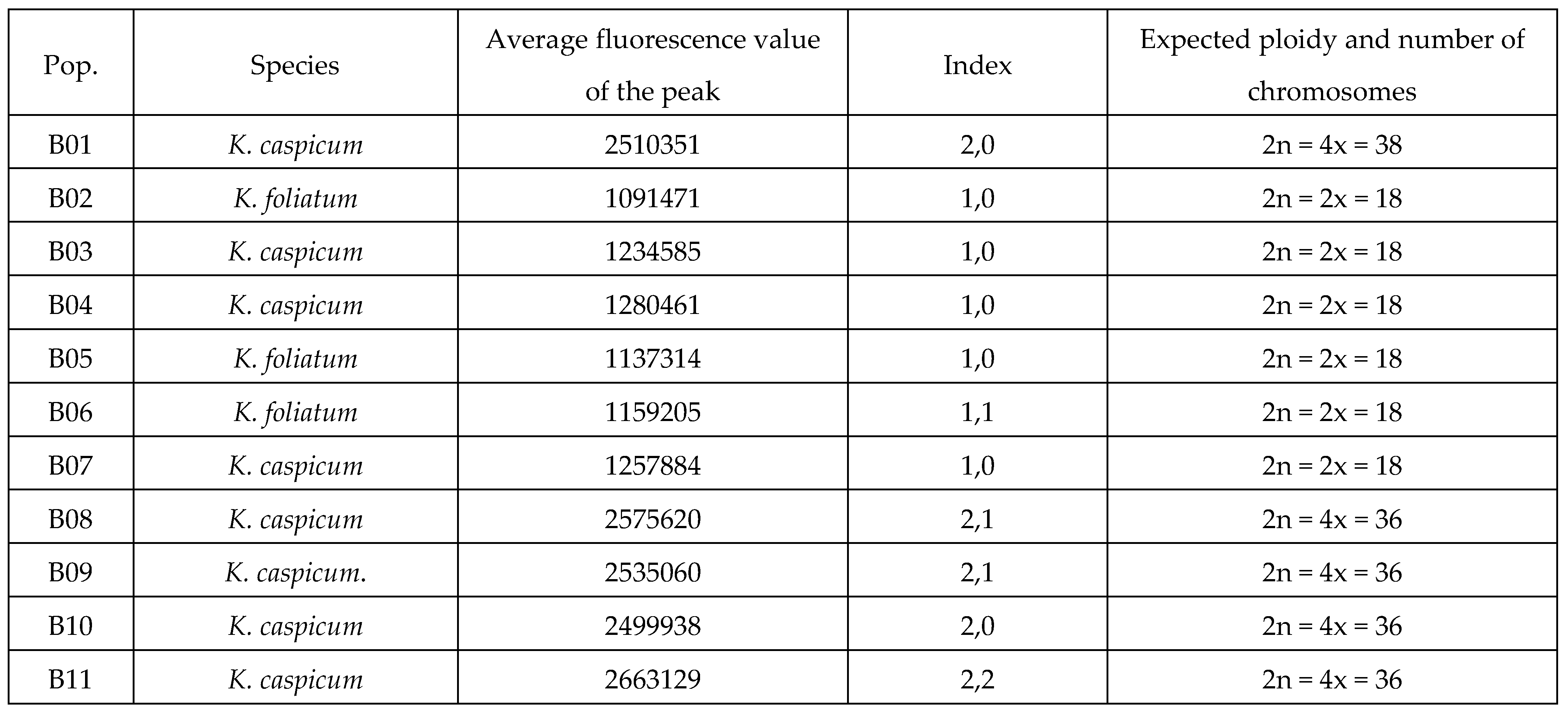
The tetraploid specimens B01, B08, B09, and B10 were the most interesting. They were combined into one group (Table 2), proving the presence of polyploidization within K. caspicum. Only sample B11 did not show the expected result (not being in the tetraploid group), which we attribute to a possible technical error in selecting a sample for analysis from this population.
When discussing the results of the SCoT analysis, in which K. schrenkianum was seen to stay close to K. foliatum, attention must be drawn to the fact that the phylogenetic tree data (ITS and chloroplast DNA) show this species to be close to K. caspicum (Figure 4).
Otherwise, all three species studied differed well from each other. Of the submitted specimens in population B07, all appeared to be identical, while in the other populations, small differences between the studied specimens were recorded.
The PCA show the interspecific arrangement of the species studied. They show, in particular, a clear distinction between the three species, proximity between K. schrenkianum and K. foliatum, and proximity between K. foliatum and K. caspicum (Supplement E).
A cross-population analysis of K. caspicum showed that populations B03, B07, and B08 were distinct (Supplement F). Samples of these populations were selected for research in the Kyzylorda region. The B03 and B07 populations were located much farther away from the other populations, namely in the Aral and Kazaly districts (Supplement F).
The geographical location of populations B03, B07, and B08 corresponds, more or less, to PCA 9A and 9B of the point map in Supplement F. Although Supplement F shows population B08 as distant from populations B03 and B07, the SCoT analysis histogram (Supplement F) shows that it is closer to these populations than B09 and B10, on the other side of the Syrdarya.
Comparison of the map of population locations in general with data on the confinement of specifically tetraploid species indicates their concentration in the middle reaches of the Syrdarya River (within the Zhalagash and Zhanakorgan districts), where solonchak deserts are most widely represented. This may also be due to an increase in air temperature in the western direction.
Unfortunately, due to the small number of K. foliatum populations collected in nature and analyzed, we cannot yet reliably explain the results shown in the histograms and map (Supplement G).
4. Materials and Methods
The objects of the study are species of the genus Kalidium growing in the territory of Kazakhstan: K. caspicum, K. foliatum, and K. schrenkianum (not considering the recently described species, K. juniperinum Sukhor. & Lomon.), as shown in Figure 5.
Classical botanical methods (route reconnaissance, ecological-systematic, and ecological-geographical) were used in the research process. The herbarium collection was carried out according to the method of Skvortsov [44], and fundamental summaries were used to identify the material collected: Flora of Kazakhstan [28], Illustrated Plant Identifier of Kazakhstan [29], and Central Asian Plant Identifier [30]. The names of plant species were taken from the International Plant Names Index (IPNI) database.
The material was collected within the Zhetysu, Almaty, and Kyzylorda regions of Kazakhstan. The largest amount of analyzed material was collected in Kyzylorda oblast, where the largest populations of the study objects occurred. All points of our collections, as well as those cited by other researchers in the National Center for Biotechnology Information (NCBI) database, are presented in Supplements A and B.
4.1. Flow cytometry
The relative DNA content was determined by flow cytometry techniques using propidium iodide by internal standardization. The samples, leaves dried with silica gel, were ground using a blade in a 1 ml Tris-MgVl2 buffer solution with the composition 0.2 M tris base, 4 mM MgCl2, 0.5% Triton X-100, 50 µg/ml RNase, 50µg/ml propidium iodide, pH 7.5 [45]. Fluorescence data of isolated nuclei were detected using a Cytoflex flow cytometer (Beckman Coulter, USA) with a 488 nm laser source. Histograms were visualized and processed using CytExpert software (Beckman Coulter, USA). The standards Glycine max (L.) Merr. ‘Polanka’ 2C=2.5 pg, Petroselinum crispum (Mill.) Fuss ‘Champion moss’ 2C=4.46 pg, and Pisum sativum L. ‘Ctirad’ 2C=9.09 pg were used as internal standards for DNA content determination. DNA content (2C, pg) was calculated according to the following formula: 2C, pg (Sample Peak Average / Standard Peak Average) * 2C Standard
The ploidy of the test samples was determined by external standardization without changing the cytometer settings, using the index of the difference between the mean peak values of the test species with known or suspected ploidy:
Index = Sample Peak Average / Standard Peak Average
4.2. Molecular genetics methods
Extraction of DNA from leaves dried with silica gel was carried out using the NucleoSpin Plant II Mini kit (MACHEREY-NAGEL GmbH & Co. KG). The ITS fragment was amplified using ITS-A [46] and ITS-4 [47] primers. The chloroplast fragments trnQ-rpS16 and trnL(UAG)-rpL32 were amplified using the primers described by Shaw et al. [48]. The Polymerase Chain Reaction (PCR) mix consisted of 1 µl DNA, 1 µl primer, 10 µl Red HS Taq 2x Mix, and 8 µl distilled water.
4.3.. Amplification and sequencing
To perform the PCR, the amplification protocol for Red Mix was used, involving a 20 µl reaction mixture with 2×HS Taq Mix Red (Biozym Scientific GmbH, Germany), where the mix was 1 µl each of direct primer and reverse primer, 10 µl Red Mix, and 8 µl . The amplified products were tested by electrophoresis on a 1.5% agarose gel stained with ethidium bromide. The DNA fragments were visualized under UV light on a Gel I X20 Lmager (INTAS Science Imaging Instruments GmbH, Germany) and documented using a Mitsubishi P93D printer (Mitsubishi Elec. Corp., Japan). The PCR products were sent to Microsynth SeqLab (Göttingen, Germany; www. microsynth.seqlab.de) for sequencing. The sequences from all individuals were manually edited in Chromas Lite 2.1 (Technelysium Pty Ltd., South Brisbane, QLD, Australia) and aligned with ClustalX [49]; the alignment was manually corrected using MEGA7 [50].
4.4. Phylogenetic analyses
Both datasets (nrITS and the cpDNA markers) were analyzed separately through Fitch parsimony with the heuristic search option in PAUP version 4.0 b10 [51] with MULTREES, TBR branch swapping, and 100 replicates of random addition sequence. Gaps were treated as missing data. The consistency index (CI) was calculated to estimate the amount of homoplasy in the character set [52]. The most parsimonious trees returned by the analysis were summarized in one consensus tree using the strict consensus method. Bootstrap support (BS)—were performed using 1,000 pseudoreplicates to assess the support of the clades [53]. Bayesian phylogenetic analyses were also performed using MrBayes 3.1.23 [54]. The sequence evolution model was chosen by following the Akaike information criterion (AIC) obtained from jModelTest2 [55]. Two independent analyses with four Markov chains were run for 10 million generations, sampling trees every 100 generations. The first 25% of the trees were discarded as burn-in. The remaining 150,000 trees were combined into a single dataset, and a majority-rule consensus tree was obtained, along with posterior probabilities (PP).
Two species from the tribe Salicornieae were selected as an outgroup: Halocnemum strobilaceum (Pall.) M. Bieb. and Halostachys belangeriana (Moq.) Botsch.
4.5. The start codon targeted (SCoT) method
To assess the genetic polymorphism, population samples of the genus Kalidium were tested with 10 SCoT primers, resulting in the selection of six primers showing polymorphism for further analysis.
The PCR was performed in a Professional Thermocycler (Biometra, Germany), with the following program: pre-precipitation - 01:30 min at 94 °C, then 36°ycles (00:45 min - +94 °C, 00:45 min - +50 °C, 1:30 min - +72 °C), final step - 6:00 min. - +72 ºC and 90:00 min at 12 ºC. The DNA was separated in an electrophoresis chamber using agarose gels with an agarose concentration of 1.5% in a TVE buffer using ethidium bromide. The duration of electrophoresis was 3.5–4 h at an electric field voltage of 85 V. The DNA was visualized using INTAS Science Imaging with Intas GDS software [56].
A 100 bp-DNA EXTENDED ladder was used as a DNA standard. The electrophoresis results were analyzed by the presence (1) or absence (0) of bands in the gel, followed by matrix generation; IBM SPSS Statistics was used to carry out PCA analysis of the data. A dendrogram showing the degree of similarity between the populations studied and the genetic distance was constructed using MEGA7.0 software [50]. To do this, the numeric values 1 and 0 in the matrix were replaced by alphabetic values (1 to A and 0 to G), and the sample names were formatted in Fasta format. Matrix of SCoT data see in Supplement C.
The matrix of SCoT data was also analyzed in the SPSS program (Version 28 https://www.ibm.com/products/spss-statistics accessed on 20.12.2022). A PCA analysis was executed with this program. The PCA is based on a correlation matrix of characters (Supplement C) using the Pearson correlation coefficient.
5. Conclusions
As a result of field studies of the phytocenotic role of Kalidium species in the composition of vegetation of Kyzylorda region it was found that among the halophytic plant communities confined to the Syrdarya river valley, phytocenoses with the participation of Kalidium species are widely represented. At the same time, it is K. caspicum that acts as a dominant. The analysis of samples of K. caspicum taken in the middle course of Syrdarya river (Zhanakorgan and Shili districts) by flow cytometry revealed tetraploid populations, occupying considerable space and differing by the larger size of plants.
At the same time, K. foliatum, both in the Syrdarya river valley and in the Ili river valley, are found much less frequently, and K. schrenkianum in the Syrdarya river valley does not occur at all, occupying more eastern territories.
According to the phylogenetic tree of species of the genus Kalidium compiled (taking into account the NCBI database):
- The species of the genus Kalidium are divided into 2 large groups. The first group: K. foliatum, K. gracile, K. wagenitzii, K. sinicum and K. cuspidatum. The second group: K. caspicum and K. schrenkianum. A promising direction for further study of the genus is additional research on the isolation of superspecific categories.
- specimens previously attributed to Kalidium cuspidatum variations (K. cuspidatum var. sinicum and K. cuspidatum var. cuspidatum) represent two independent taxa: K. cuspidatum var. sinicum is understood as K. sinicum, and K. cuspidatum var. cuspidatum, as K. cuspidatum.
A specimen of hybrid (between K. foliatum and K. caspicum) origin was found in the Ili River valley, according to molecular genetic studies.
Data Availability Statement
All the data are presented in figures, tables, and Supporting Information.
Acknowledgments
This research has been funded by the Science Committee of the Ministry of Education and Science of the Republic of Kazakhstan (Grant No. AR09258929 - Prospects for using the correlation between the composition of the anthropophilic element of the flora of the desert part of the Syrdarya River valley and the type of land disturbance for forecast purposes” (2021-2023) (leader Vesselova P.V.).
Conflicts of Interest
The authors have no conflict of interest in any manner.
References
- Kadereit, G.; Mucina, L.; Freitag, H. Phylogeny of Salicornioideae (Chenopodiaceae): diversification, biogeography, and evolutionary trends in leaf and flower morphology. Taxon 2006, 55, 6. [Google Scholar] [CrossRef]
- Piirainen, M.; Liebisch, O.; Kadereit, G. Phylogeny, biogeography, systematics and taxonomy of Salicornioideae (Amaranthaceae/Chenopodiaceae) – A cosmopolitan, highly specialized hygrohalophyte lineage dating back to the Oligocene. Taxon 2017, 66, 109–132. [Google Scholar] [CrossRef]
- Vesselova, P.V.; Kudabayeva, G.M. Comparative analysis of floristic lists of the desert part of the Syr Darya valley. Ukrainian Journal of Ecology 2018, 8, 427–432. [Google Scholar]
- Chapman, V.J. The New Perspective in the Halophytes. Q. Rev. Biol. 1942, 17, 291–311. [Google Scholar] [CrossRef]
- Shreve, F. Vegetation of the Sonoran Desert. Carnegie Institution of Washington Publication 1951, 591, 1–192. [Google Scholar]
- Wilder, B.T.; Felger, R.S.; Morales, H.R. Succulent Plant Diversity Of the Sonoran Islands, Gulf Of California, Mexico. Haseltonia 2008, 14, 127–160. [Google Scholar] [CrossRef]
- Kung, H.-W.; Chu, G.-L.; Tsien, C.-P.; Li, A.-J.; Ma, C.-G. The Chenopodiaceae in China. Acta Phytotaxonomica Sinica. (In Chinese). 1978, 16, 99–123. [Google Scholar]
- Zhao, Y.-Z.; Zhao, L.-Q.; Cao, R. Flora Intramongolica. 3rd ed. Vol. 1. Huhhot: Typis Intramongolicae Popularis. 2020, 779 pp.
- Sukhorukov, P.; Kushunina, M.A.; Lomonosova, M.N. A new Kalidium species (Amaranthaceae s. l.) from northern Central Asia. Turczaninowia 2022, 25, 24–33. [Google Scholar] [CrossRef]
- Sukhorukov, A.P.; Kushunina, M.A. Taxonomic revision of Chenopodiaceae in Nepal. Phytotaxa 2014, 191, 10–44. [Google Scholar] [CrossRef]
- Zhao, K.F.; Fan, H.; Jiang, X.Y.; et al. Improvement and utiliza- tion of saline soil by planting halophytes. Chinese Journal of Applied & Environmental Biology 2002, 8, 31–35. (in Chinese). [Google Scholar]
- Zhou, Z.Y.; Yan, S.Y.; Qin, Y.; et al. The character of shrub di- versity in arid desert regions in Alashan. Journal of Arid Land Resources and Environment 2009, 23, 146–150. [Google Scholar] [CrossRef]
- Liang, X.H.; Wu, Y.X. Identification of Kalidium species (Chenopodiaceae) by DNA barcoding. Sciences in Cold and Arid Regions 2017, 9, 0089–0096. [Google Scholar] [CrossRef]
- Wang, G.; Jia, J. Effects of NaCl on physiology and leaf ultrastructure in the halophyte Kalidium foliatum. Nordic Journal of Botany 2015, 33, 232–238. [Google Scholar] [CrossRef]
- Tobe, K. Seed Germination and Radicle Growth of a Halophyte, Kalidium capsicum (Chenopodiaceae). Annals of Botany 2000, 85, 391–396. [Google Scholar] [CrossRef]
- Tobe, K.; Li, X.; Omasa, K. Effects of sodium, magnesium and calcium salts on seed germination and radicle survival of a halophyte, Kalidium caspicum (Chenopodiaceae). Aust. J. Bot. 2002, 50, 163–169. [Google Scholar] [CrossRef]
- Song, J.; Feng, G.; Zhang, F. Salinity and Temperature Effects on Germination for Three Salt-resistant Euhalophytes, Halostachys caspica, Kalidium foliatum and Halocnemum strobilaceum. Plant and Soil 2006, 279, 201–207. [Google Scholar] [CrossRef]
- Qu, X.; Baskin, J.M.; Wang, L.; Huang, Z. Effects of cold stratification, temperature, light and salinity on seed germination and radicle growth of the desert halophyte shrub, Kalidium caspicum (Chenopodiaceae). Plant Growth Regulation 2007, 54, 241–248. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, D.Y.; Huang, Z.Y.; Tian, C.Y. Factors influencing seed germination of Kalidium caspicum (Chenopodiaceae), a halophytic desert shrub of Xinjiang, China. Seed Sci. and Technol. 2009, 37, 281–290. [Google Scholar] [CrossRef]
- Gong, D.H.; Wang, G.Z.; Si, W.T.; Zhou, Y.; Liu, Z.; Jia, J. Effects of Salt Stress on Photosynthetic Pigments and Activity of Ribulose-1,5-bisphosphate Carboxylase/Oxygenase in Kalidium foliatum. Russian Journal of Plant Physiology 2018, 65, 98–103. [Google Scholar] [CrossRef]
- Wang, Z.G.; Zhang, P.X.; Shao, Y.T.; Xu, T.T.; Jia, X.Y.; Zhang, X.Q.; Jia, J. Molecular Cloning and the Expression Pattern of a Phospholipid Hydroperoxide Glutathione Peroxidase in Kalidium foliatum under NaCl Treatment. Russian Journal of Plant Physiology 2020, 67, 750–757. [Google Scholar] [CrossRef]
- Wang, J.; Wu, S.; Wang, M. The complete chloroplast genome of halophyte Kalidiumfoliatum (Pall. ) Moq., a dominant species of desert grassland, Mitochondrial DNA Part B. 2022, 7, 544–545. [Google Scholar] [CrossRef] [PubMed]
- Schutze, P.; Freitag, H.; Weising, K. An integrated molecular and morphological study of the subfamily Suaedoideae Ulbr. (Chenopodiaceae). Plant Syst. Evol. 2003, 239, 257–286. [Google Scholar] [CrossRef]
- Wen, Z.-B.; Zhang, M.-L.; Zhu, G.-L.; Sanderson, S.C. Phylogeny of Salsoleae s. l. (Chenopodiaceae) based on DNA sequence data from ITS, psbB–psbH, and rbcL, with emphasis on taxa of northwestern China. Plant Syst. Evol. 2010, 288, 25–42. [Google Scholar] [CrossRef]
- Xue, J.J.; Zhang, M.L. Monophyly and infrageneric variation of Corispermum L. (Chenopodiaceae), evidence from sequence data psbB-psbH, rbcL and ITS. Journal of Arid Land 2011, 3, 240–253. [Google Scholar] [CrossRef]
- Lu, K.-Q.; Li, M.; Wang, G.-H.; Xu, L.-S.; Ferguson, D.K.; Trivedi, A.; Wang, Y.-F. A new pollen classification of Chenopodiaceae for exploring and tracing desert vegetation evolution in the eastern central Asia. Journal of Systematics and Evolution 2018. [CrossRef]
- Ilyin, M.M. The genus Kalidium Moq. Flora of the USSR. - М., 1936, VI, p. 166-168. [in Russian].
- Goloskokov, V.P. and Polyakov, P.P. The genus Kalidium Moq. Alma-Ata. 1960, III, p. 243-246. [in Russian].
- Goloskokov, V.P. and Polyakov, P.P. The genus Kalidium Moq. Illustrated Plant Identifier of Kazakhstan. Alma-Ata. ed. of the Kazakh SSR Academy of Sciences 1969, Т 1, p. 300-301. [in Russian].
- Pratov, U. The genus Kalidium Moq. Plant Identifier of Plants of Central Asia. Tashkent. FAN Publishing House. 1972, III, p. 70-72. [in Russian].
- Abdulina, S.A. Рoд Kalidium Moq. Checklist of vascular plants of Kazakhstan. Almaty. 1998, p. 77. (in Russian).
- Baitenov, M.S. Genus Kalidium Moq. Flora of Kazakhstan. Genus complex flora. Almaty. 2001, 2, p. 72. (in Russian).
- Skaptsov, M.V. and Kutsev, M.G. Possibilities of flow cytometry in modern plant science. "Problems of botany of South Siberia and Mongolia" - XIII International Scientific-Practical Conference. Barnaul. 2014, p. 204-207. (in Russian)
- Friesen, N. Molecular methods used in plant systematics. Barnaul. AzBuka, 64 p. (in Russian). 2007. [Google Scholar]
- Lomonosova, M.N. and Krasnikov, A.A. Chromosome numbers in some members of the Chenopodiaceae. Bot. Zhurn. 1993, 78, 158–159. [Google Scholar]
- Lomonosova, M.N. and Krasnikov, A.A. Chromosome numbers of some Chenopodiaceae representatives of the flora of Russia. Bot. Zhurn. 2006, 91, 1757–1759. [Google Scholar]
- Krasnikov, A. and Schaulo, D. Karyological study of the Tuva Republic flora: a summary. Turczaninowia 2004, 7, 82–95. [Google Scholar]
- Marhold, K. IAPT/IOPB chromosome data 13. Taxon 2012, 61, 892–893. [Google Scholar] [CrossRef]
- Marhold, K. IAPT/IOPB chromosome data 18. Taxon 2014, 63, 1387–1393. [Google Scholar] [CrossRef]
- Marhold, K.; Kucera, J.; Alexeeva, T.V.; Andriyanova, E.A.; Ankova, T.V.; Astashenkov, A.Y.; … Zavgorodnyaya, O.Y. IAPT chromosome data 32. Taxon 2020. [CrossRef]
- Lomonosova, M.N.; Krasnikov, A.A.; Krasnikova, S.A. Chromosome numbers of Chenopodiaceae family members of the Kazakhstan flora. Bot. Zhurn. 2003, 88, 134–135. [Google Scholar]
- Ghaffari, S.M.; Saydrasi, L.; Ebrahimzadeh, H.; Akhani, H. Chromosome numbers and karyotype analyses of species of subfamily Salicornioideae (Chenopodiaceae) from Iran. Iran. J. Bot. 2006, 12, 128–135. [Google Scholar]
- Zhu, G.; Sergei, L.M.; Stevan, E. Chenopodiaceae: Flora of China. 2003, 5, 351–414.
- Skvortsov, A.K. Herbarium: A Manual on Methods and Techniques. Prof. L.I. Prilipko M. Nauka, 1977. 199 p. (in Russian) [Google Scholar]
- Pfosser, М.; Amon, A.; Lelley, Т.; Heberle-Bors, E. Evaluation of sensitivity of flow cytometry in detecting aneuploidy in wheat using disomic and ditelosomic wheat-rye addition lines. Cytometry 1995, 21, 387–393. [Google Scholar] [CrossRef]
- Blattner, F.R. Direct amplification of the entire ITS region from poorly preserved plant material using recombinant PCR. In. Biotechniques 1999, 27, 1180–1186. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Shinsky JJ, White TJ, editors. PCR Protocols: A Guide to Methods and Applications. 1990, 315–322.
- Shaw, J.; Lickey, E.B.; Schilling, E.E.; Small, R.L. Comparison of whole chloroplast genome sequences to choose noncoding regions for phylogenetic studies in angiosperms: the tortoise and the hare III. American Journal of Botany 2007, 94, 275–288. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The clustal X window interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Swofford, D. –L. PAUP*: Phylogenetic analysis using parsimony (* and other methods). version 4 (Sunderland, Massachusetts: Sinauer Associates). 2002.
- Kluge, A.G.; Farris, J.S. Quantitative phyletics and the evolution of anurans. Syst. Zool. 1969, 18, l–32. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, R. and Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 77. [Google Scholar] [CrossRef]
- Bertrand, C.; Collard, Y.; Mackill, D.J. Start Codon Targeted (SCoT) Polymorphism: A Simple, Novel DNA Marker Technique for Generating Gene-Targeted Markers in Plants. Plant Mol. Biol. Rep. 2009, 27, 86–93. [Google Scholar] [CrossRef]
Figure 1.
Distribution map of the samples examined.
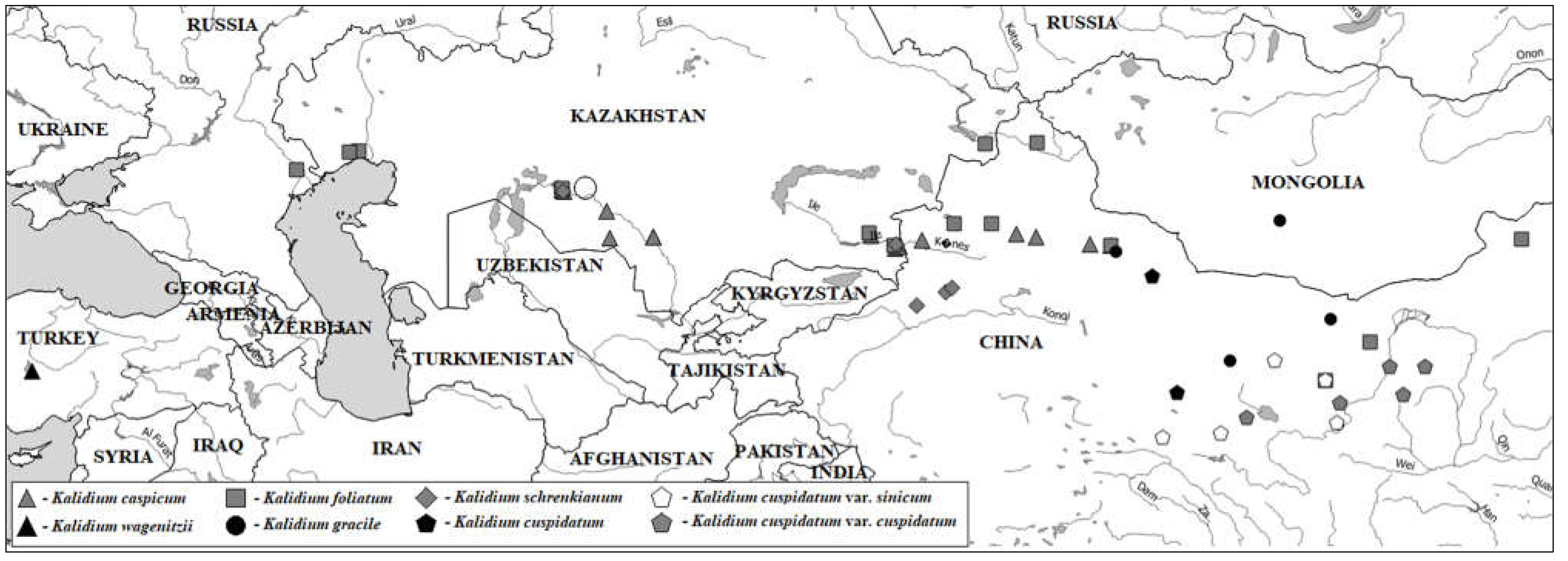
Figure 5.
Objects of the study: A-C – Kalidium caspicum Ung.-Sternb.; D-F – K. foliatum Moq.; G-I – K. schrenkianum Bunge ex Ung.-Sternb. (all data on the collection points is provided in Supplement A).
Figure 5.
Objects of the study: A-C – Kalidium caspicum Ung.-Sternb.; D-F – K. foliatum Moq.; G-I – K. schrenkianum Bunge ex Ung.-Sternb. (all data on the collection points is provided in Supplement A).


Table 1.
DNA content in the nuclei of Kalidium species and expected ploidy based on cytometry data.
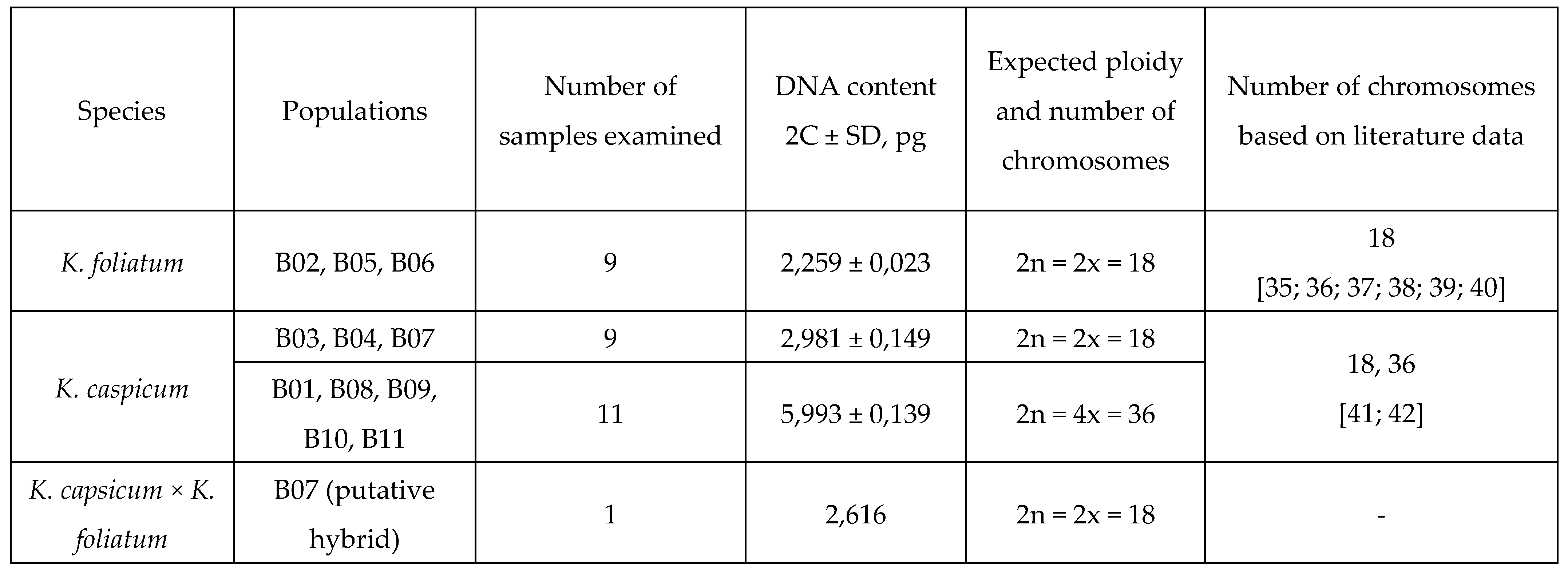 |
Table 2.
Ploidy data of Kallidium species by external standardization without changing the cytometer settings.
Table 2.
Ploidy data of Kallidium species by external standardization without changing the cytometer settings.
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



