
Submitted:
17 May 2023
Posted:
18 May 2023
You are already at the latest version
Abstract
Reproductive isolating barriers are a crucial element in the speciation process. There are various types of barriers, and within those that act early in the speciation process, the behavioral one can play an important role in isolating populations or closely related species. It has been shown that the high diversity of some taxa correlates with the variation of sexual signals, which facilitate isolation. Here it is explored whether chemical signals may contribute to the high diversity of Liolaemus lizards, which has more than 290 species. Population chemical discrimination was investigated in two co-distributed species, L. tenuis and L. lemniscatus. The studied populations were closely located and likely belonged to the same genetic lineages. Lizards of both species discriminated between scents of their own and different populations, and in most cases, scents from their population were preferred. Results suggest a fast evolution of scents, and their recognition, and there for the ethological barriers associated with the chemical modality may evolve fast. However, because the studied species differed in the strength of the exhibited population chemical discrimination, ethological barriers evolve at different rates across species. It can be concluded that ethological barriers involving the chemical modality may disrupt species' cohesion, contributing to Liolaemus diversification, in synergy or not with geographical barriers.
Keywords:
geographic barrier
; homotypic
; heterotypic
; Liolaemus lemniscatus
; Liolaemus tenuis
; olfaction
; scents
; speciation
; tongue flick
1. Introduction
A critical step in the speciation process is the occurrence of reproductive barriers that limit gene flow between populations or incipient species e.g., [1,2,3]. Such barriers are classified based on when they restrict gene exchange, with premating barriers precluding copulation [2], which tend to evolve early in the process of reproductive isolation [4]. Among different premating barriers, behavioral ones, also called ethological or sexual isolation barriers [2], have been proposed to play a pivotal role in isolating populations or closely related species [5]. Thus, in the context of sexual selection, signals involved in mate attraction and male-male competition can play a significant role in speciation [6,7,8], as supported by many studies involving different taxa and sensory modalities e.g., [9,10,11,12]. Remarkably, in some taxa, high speciation rates may result from high divergence in sexual signals e.g., [13]. This includes the Laupala crickets, the genus with the highest speciation rate among arthropods, with a high divergence in song characteristics across taxa [14]. Similarly, the increased diversification of African cichlid fishes has been linked to variation in male colorations involved in mate selection [15,16].
Liolaemus, a genus with more than 290 lizard species [17,18], is one of the most diverse tetrapod genera in the world, distributed in the southern part of South America [18,19]. Its high diversity has been related to the Andean uplift that fragmented species distribution promoting speciation [19,20], and different genetic analyses support the role of geographic barriers, i.e., rivers and mountains, in this diversification [21,22,23]. In fact, the diversity of different South American taxa has also been associated with changes in landscape, climate, and drainage from the Andean formation e.g., [24,25,26]. However, no other genus in the Andean mountains has a similar diversity to Liolaemus, even the genera belonging to the Liolaemidae family [18]. Therefore, geographic isolation per se does not explain the Liolaemus diversity, and other factors, such as reproductive barriers involving organismal traits, i.e., sexual signals, may play a prominent role in Liolaemus speciation [27]. In this context, the working hypothesis of this study is that populations of Liolaemus species may show differences in sexual signals allowing population discrimination, which may break species cohesion [28]. If so, ethological isolation may contribute to Liolaemus diversity.
Chemical signals, or scents, are relevant in the speciation process of many different taxa [29]. In Liolaemus, further than allowing individual recognition [30,31,32], scents may facilitate isolation among sympatric congeneric species [30,33,34]. The composition of precloacal secretions, a pheromone source in Liolaemus lizards [35,36,37], differ between sexes [38], among males of a population, between populations, and among congeneric species [39,40], suggesting that scents may evolve fast [33,34]. Thus, considering that scents of nearby populations can differ [40], possibly populations may discriminate between homo- and heterotypic conspecific scents, i.e., from the same and different populations, respectively, and most likely, homotypic scents would be selected e.g., [41,42], disrupting the species cohesion in the absence of geographic barriers.
This study tested the working hypothesis by determining population scent recognition of closely located populations of two Liolaemus species, L. tenuis and L. lemniscatus. These species are co-distributed in most of their distributional ranges, which may have shared vicariant events evolving along similar demographic trajectories [43]. Thus, by studying these species together, the effects of factors like environmental conditions that modulate scent composition e.g., [44,45,46], are controlled.
2. Materials and Methods
Lizards from L. tenuis and L. lemniscatus were collected from the south bank of the Maipo River in central Chile (Figure 1), a well-recognized geographic barrier for many lizard species [22,47,48,49]. The aim was to study populations that might experience gene flow. The studied populations have no genetic analyses, although data suggest they are likely from a single genetic lineage [22,43].
2.1. Experiment I. Recognition between homo- vs. heterotypic competitors
Animals and their maintenance. In January 2001, males of both species were collected from different populations (Figure 1); 19 L. tenuis [nine from Codegua (34.03335ºS; 70.56668ºW; Datum WGS84, 765 m), three from Las Vizcachas (33.61560°S; 70.51593ºW; 890m), and seven from Pirque (33.72283°S; 70.49030°W; 850m)] and 16 L. lemniscatus [eight from Las Vizcachas, and eight from Codegua]. In the laboratory, lizards were placed in an indoor vivarium with a glass roof, exposing them to the sun and natural photoperiod (13:11 L:D). Additionally, the vivarium was equipped with halogen lamps to maintain similar temperatures to those recorded on a typical summer day (between 12 ºC to 36 ºC). Lizards were housed individually in plastic enclosures (37 × 30 × 15 cm) with a 3 cm sand-layer substrate. Enclosures had a lid of plastic mesh and were equipped with a bowl to keep water permanently and a rock to provide shelter and a basking place. Food (mealworms) was supplied three times a week, dusted with vitamins.
Experimental design. Animals remained for a week in their enclosures allowing habituation to experimental conditions and scent releasing, as enclosures were used as a source of substrate-borne scents [33]. For the experiments, a focal lizard was taken from its enclosure and kept in a cloth bag for 10 min to reduce handling associated stress [33]; after that, the bag was placed in the experimental enclosure, allowing the lizard to move freely. The experimental enclosure corresponded to one used by homo- or heterotypic males. Each lizard was tested in a counterbalanced design with scents from different populations. The enclosure resident was removed with the rock and the bowl for water just before the trial. The focal population of L. tenuis was Codegua, which was tested with homo- and heterotypic scents from Pirque and Las Vizcachas. In the case of L. lemniscatus, both populations, Codegua and Las Vizcachas, were tested with scents from different populations.
Once the focal lizard was in the experimental enclosure, the latency to the first tongue flick was recorded as the time elapsed since the lizard was placed in the enclosure until it made the first tongue flick [50]. After this display, the lizard's behavior was filmed for 10 min with an 8-mm digital video camera placed 50 cm above the enclosure. At the end of each trial, the lizard's cloacal temperature was recorded to ensure that individuals were at the species selected body temperatures [~35- 36°C; 51] during the experiment. Finally, focal and scent-donor lizards were placed back in their enclosures. Lizards were subjected to one trial per day and had an inter-trial of at least two days.
From the videos, the following variables were recorded [32,50]: 1- Number of tongue flicks. The number of times the lizard protruded and retracted the tongue, which touched the air or a surface. Tongue flicks allow lizards to explore the chemical environment (vomerolfaction) by collecting scents (volatile and non-volatile compounds) and delivering them to the vomeronasal organ [52]. 2- Motion time. Total time that the lizard moved, which included adjustments of body posture, head movements (scanning), displacements of the body's center of gravity, and displays that occurred at too low frequency to be analyzed independently (marking behaviors, tail waving, slow motion, defecation, and digging). 3- Head displays. Time exhibiting up and down head movements combined or not with forelimb flexions.
2.2. Experiment II. Recognition between homo- vs. heterotypic potential sexual partners
In December 2003, individuals of both sexes from two populations of each species were collected; L. tenuis [22 from Codegua (11♂, 11♀) and 19 from Las Vizcachas (10♂, 9♀)] and L. lemniscatus [35 from Codegua (21♂, 14♀) and 18 from Las Vizcachas (11♂, 7♀)]. In the laboratory, lizards were maintained as before, except that the rocks of the maintenance enclosures were replaced by inverted clay pots, which were later used as scent sources.
Experiments were performed in a round enclosure of 50 cm in diameter with a layer of 2 cm clean sand at the bottom. The arena was divided virtually into two halves, each containing a clay pot, one from a homo- and the other from a heterotypic potential sexual partner to the focal lizard. The lizard was filmed for 20 min after the first tongue flick. From the videos, five variables were quantified: time spent at each side of the experimental enclosure, time spent on the two clay pots, the number of tongue flicks performed on each side of the enclosure, number of tongue flicks directed to each clay pot, and finally, time moving on each side of the experimental enclosure. After each trial, enclosures were washed with soap, and the sand was discarded and replaced.
2.4. Experiment III. Scent preference for homo- vs. heterotypic potential sexual partners
In February 2004, new individuals of both sexes from two populations of each species were collected; L. tenuis [11 from Codegua (5♂, 6♀) and 18 from Las Vizcachas (8♂, 10♀)] and L. lemniscatus [16 from Codegua (8 ♂, 8♀) and 13 from Las Vizcachas (8♂, 5♀)]. In the laboratory, lizards were maintained as in Experiment II. To assess scent preferences, a focal lizard remained for 4.5 hrs in the round enclosure described above. Every 30 min, an observer behind a blind determined at which side of the enclosure the focal lizard was located. Scent preference was determined by calculating the percentage of time that lizards were at each side of the enclosure.
Individuals were kept in good conditions in the laboratory, and at the end of each of the three experiments, they were released at their capture sites.
2.5. Statistics
Data for each experiment by species were analyzed using Generalized Linear Models for repeated measures (scent origin; homo- vs. heterotypic), followed by Fisher LSD tests. For Experiment I, analyses assessed the effect of scent origin on four responses. However, the head display was not analyzed for L. tenuis since it was displayed in low frequency but was included in motion time. The normality of the head display in L. lemniscatus was improved by log-transforming the data. The analyses of L. lemniscatus also included the effect of the population (Codegua vs. Las Viscachas) and its interaction with scent origin.
For Experiments II and III, tests assessed the effects of scent origin, the sex and population of the focal lizard, and their interactions on the recorded variables. The percentages of time associated with homo- and heterotypic scents (Experiment III) were transformed using the square root of the arcsine.
Statistical analyses were performed with StatSoft, Inc (2001)
3. Results
3.1. Experiment I. Recognition between homo- vs. heterotypic competitors
Liolaemus tenuis. Scent origin did not modulate the latency to the first tongue flick nor motion time (Table 1A; Figure 2A and 2C, respectively), but it did affect the number of tongue flicks; lizards directed more tongue flicks toward hetero- than homotypic scents, and the scents of both heterotypic populations were similarly examined (Figure 2B).
Liolaemus lemniscatus. Scent origin was the only factor with an effect (Table 1B). First, males showed shorter latency to the first tongue-flick to hetero- than to homotypic male scents (Figure 3A). Second, males exhibited head displays for an extended period with homotypic scents (Figure 3D). None of the other variables, i.e., number of tongue flicks and motion time, were affected by scent origin (Figure 3B and 3C, respectively).
3.2. Experiment II Recognition between homo- vs. heterotypic potential sexual partners
Liolaemus tenuis. Scent origin affected all the recorded variables (Table 2); lizards, independently of their sex or population, spent more time, tongue flicked more, and moved more in the area with heterospecific scents (Figure 4A,B). In addition, sex and population modulated the time spent moving (Table 2); males moved more than females (2.09 ± 0.17 vs. 1.54 ± 0.12 min, respectively), and lizards from Codegua moved more than those from Las Viscachas (2.01 ± 0.17 vs. 1.58 ± 0.12 min, respectively).
Liolaemus lemniscatus. Scent origin modulated the time spent on the pot and the number of tongue flicks directed to the pot but only in interaction with the sex of the focal lizards (Table 3); females used less and directed fewer tongue flicks to pots with heterotypic scents than males (Figure 4C). However, the a posteriori tests could not exclude that both sexes behave similarly to pots with different scents (Figure 4C).
3.3. Experiment III. Scent preference for homo- vs. heterotypic potential sexual partners
Individuals of L. tenuis, independent of their sex and population, spent more time with homotypic scents (Table 3; Figure 5), while scent preferences of L. lemniscatus were influenced by the origin of the focal lizard; individuals from Las Viscachas preferred homotypic scents and individuals from Codegua preferred heterotypic scents (Table 3; Figure 5).
4. Discussion
The studied Liolaemus species showed discrimination between homo- and heterotypic scents, preferring homotypic scents in most cases. This suggests scent differences across populations, probably mirrored by differences in chemoreception. Because the studied populations were closely located and most likely from a unique genetic lineage [22,43], these findings indicate that behavioral barriers involving chemical modality may evolve relatively fast and may facilitate reproductive isolation even without geographic barriers. The two species, however, differed in the strength of their responses toward different populations and, therefore, in the degree of reproductive isolation conveyed by the chemical modality.
1- Recognition between homo- vs. heterotypic competitors (Experiment I)
Signals involved in competitor deterrence, as well as those implicated in mate attraction, can play a role in ethological isolation [6,8,11,53]. Thus, reproductive isolation has been studied by assessing the responses triggered by competitors from different origins e.g., [12,54]. In this study, males of both species discriminated between potential competitors from different populations, suggesting that scents may contribute to population isolation e.g., [42].
The higher rate of tongue flick made by L. tenuis toward non-homotypic scents suggests that these scents were perceived as different from those of the own population. These scents, however, were not as different as those from congeneric males, which elicited less exploration than homotypic scents [33,55]. Potentially, populations share a “species signature” [33], allowing recognition of non-homotypic males as conspecifics. On the other hand, since heterotypic populations were treated similarly, scents may also provide a "population signature” e.g., [56], inducing a dichotomous response, where any heterotypic c scent is treated as different, independently of how much they differ from the “population signature”.
The scents from different populations triggered similar tongue-flick rates in L. lemniscatus. This may be because volatile scent compounds were assessed and discriminated by olfaction rather than vomerolfaction. In fact, scent evaluation by olfaction has been proposed for different Liolaemus species [32,35], including for L. lemniscatus [50]. The olfactory assessment triggered a faster exploration of heterotypic scents with the tongue and a longer exhibition of head displays toward homotypic male scents, indicating that these individuals were perceived as more threatening competitors [32,33].
2- Recognition and preference for homo- vs. heterotypic potential sexual partners (Experiments II and III)
In L. tenuis, the simultaneous exposition to scents from potential mates of different populations triggered a higher exploration (Experiment II) and a preference for homotypic partners (Experiment III); conspecifics from the own population were more attractive. These results suggest that these populations experience ethological isolation barriers mediated by the chemical modality and that gene flow may be restricted e.g., [42]. Consistently, genetic analyses of L. tenuis along its geographic distribution have revealed that the species might constitute a diverse cryptic-species complex [22,47,57]. Therefore, genetic studies should integrate scent analyses and discrimination e.g., [58].
In L. lemniscatus, the simultaneous exposure to scents from potential partners of the two populations did not determine clear-cut results; weak or nonexistent population recognition was observed during the short scent exposure (20 min, Experiment II) while the prolonged exposure (4.5 hrs, Experiment III) revealed population preferences. During the brief exposition, the sexes explored the heterotypic scents differently, with females spending less time investigating than males. However, because the sexes behaved similarly with homo- and heterotypic scents, data suggest a lack of population discrimination. Remarkably, the prolonged scent exposure revealed population discrimination, although preferences were asymmetric; both populations were more attracted to Las Viscachas scents, suggesting that populations may have only a weak reproductive barrier. Altogether, competitor (male) scents allowed population discrimination (Experiment I) in L. lemniscatus, but not scents from potential sexual partners, at least after a short exposure. Possibly, lizards may be more tolerant to deviation from the "population signature" when these are from potential sexual partners (Experiment II), requiring a prolonged scent exposure to exhibit preferences (Experiment III). Therefore, L. lemniscatus may have relatively strong species cohesion, and gene flow across populations may only be slightly restricted.
3- Liolaemus diversity and ethological isolation associated with the chemical sensory modality
Scents may evolve fast in lizards e.g., [44], showing differences across populations e.g., [40,59,60,61]. However, these differences do not necessarily determine population recognition e.g., [41,62] and may play a minor role in the evolution of reproductive isolation [63]. There are, however, also evidence for lizards from different genera showing population scent variations mirrored by population discrimination, driving the proposition that such variation may promote reproductive isolation, i.e., Podarcis [41,62,64], Psammodromus [65] and Aspidoscelis [42]. Of these genera, Aspidoscelis has the highest diversity, with 44 recognized species [17] and some with many subspecies [66]. This diversity possibly is determined by hybridization and introgression across taxa [66]. Therefore, ethological isolation may not contribute to the diversification of this genus. So then, although population scent recognition may allow reproductive isolation in these genera, this does not determine a high diversification of these taxa.
In Liolaemus, considering that the studied populations by species were closely located, without geographic barriers that prevented gene flow, and most likely belonging to a unique genetic lineage [22,43], the exhibited population discrimination suggest that scents, and their recognition, may evolve fast e.g., [33]. In this context, ethological barriers associated with the chemical modality may evolve fast, although at different rates across species, contributing to Liolaemus diversification, even without geographic barriers. This may explain the restricted geographic distribution of various Liolaemus species [18].
5. Conclusions
Different taxa show significant variation in speciation rates [2,27], and unraveling what causes such variation is one fundamental question in speciation research [67] but difficult to tackle, as speciation is a multi-cause process [2]. The present study provides a starting point to investigate how ethological barriers involving the chemical modality may disrupt species' cohesion, and facilitate speciation in Liolaemus, in synergy or not with geographical barriers e.g., [19]. However, new studies including species from different clades are needed to test the proposed role of the ethological “chemical” barriers in Liolaemus diversity. Furthermore, clades may differ in the relevance of chemical barriers, considering that some studies in Liolaemus show hybridizations and introgressions e.g., [68,69,70], implying that chemorecognition may not be relevant for all species.
Funding
Financial support was provided by IFS (International Foundation of Science, Sweden) 2933-1/2.
Acknowledgments
This study followed the current Chilean laws governing animal capture and maintenance imposed by Servicio Agrícola y Ganadero (Permits N° 2320; 3327). I thank Marco Mangiacotti for the invitation to participate in the special issue "Ecology and Evolution of Chemical Communication in Lizards”, Daniel Benitez and Julio Labra for their help during fieldwork, Eduardo Aguilera for his invaluable assistance with filming, animal care, and fieldwork, Hermann M. Niemeyer and Mario Penna for allow using their facilities to develop this study, Juan Contardo for kindly make the map of the population localities., and Thomas F. Hansen for very insightful comments on an early version of the manuscript,.
References
- Matute, D.R.; Cooper, B.S. Comparative studies on speciation: 30 years since Coyne and Orr. Evolution, 2021, 75, 764–778. [Google Scholar] [CrossRef] [PubMed]
- Coyne, J.A.; Orr, H.A. Speciation. Sinauer Associates: Sunderland, 2004.
- Mayr, E. Systematics and the Origin of Species. Columbia University Press New York, 1942.
- Gavrilets, S.; Vose, A. Dynamic patterns of adaptive radiation: Evolution of mating preferences. In: Butlin, R.K., Bridle, J.R., Schluter, D., eds. Speciation and Patterns of Diversity. Cambridge, United Kingdom: Cambridge University Press, 2009; pp. 102-126.
- Butlin, R.K.; Ritchie, M.G. Behaviour and speciation. In: Slater, P.J.B., Halliday, T.R., Barrett, P., eds. Behaviour and Evolution New York: Cambridge University Press, 1994; pp. 43-79.
- Panhuis, T.M.; Butlin, R.K.; Zuk, M.; Tregenza, T. Sexual selection and speciation. Trends Ecol. Evol., 2001, 16, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Mendelson, T.C.; Safran, R.J. Speciation by sexual selection: 20 years of progress. Trends Ecol. Evol., 2021, 36, 1153–1163. [Google Scholar] [CrossRef] [PubMed]
- West-Eberhard, M.J. Sexual selection, social competition, and speciation. Q. Rev. Biol., 1983, 58, 155–183. [Google Scholar] [CrossRef]
- Roriz, A.K.P.; Japyassú, H.F.; Cáceres, C.; Vera, M.T.; Joachim-Bravo, I.S. Pheromone emission patterns and courtship sequences across distinct populations within Anastrepha fraterculus (Diptera-Tephritidae) cryptic species complex. Bull. Entomol. Res., 2019, 109, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Roberts, N.S.; Mendelson, T.C. Identifying female phenotypes that promote behavioral isolation in a sexually dimorphic species of fish Etheostoma zonale. Curr. Zool., 2021, 67, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Uy, J.A.C.; Irwin, D.E.; Webster, M.S. Behavioral isolation and incipient speciation in birds. Annual Rev. Ecol. Evol. System., 2018, 49, 1–24. [Google Scholar] [CrossRef]
- Lipshutz, S.E.; Overcast, I.A.; Hickerson, M.J.; Brumfield, R.T.; Derryberry, E.P. Behavioural response to song and genetic divergence in two subspecies of white-crowned sparrows (Zonotrichia leucophrys). Mol. Ecol., 2017, 26, 3011–3027. [Google Scholar] [CrossRef]
- Mason, N.A.; Burns, K.J.; Tobias, J.A.; Claramunt, S.; Seddon, N.; Derryberry, E.P. Song evolution, speciation, and vocal learning in passerine birds. Evolution, 2017, 71, 786–796. [Google Scholar] [CrossRef]
- Mendelson, T.C.; Shaw, K.L. Rapid speciation in an arthropod. Rapid speciation in an arthropod, 2005, 433, 375–376. [Google Scholar] [CrossRef]
- Knight, M.E.; Turner, G.F. Laboratory mating trials indicate incipient speciation by sexual selection among populations of the cichlid fish Pseudotropheus zebra from Lake Malawi. P. Roy. Soc. Lond. B Bio., 2004, 271, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Maan, M.E.; Seehausen, O.; Söderberg, L.; Johnson, L.; Ripmeester, E.A.P.; Mrosso, H.D.J.; Taylor, M.I.; van Dooren, T.J.M.; van Alphen, J.J.M. Intraspecific sexual selection on a speciation trait, male coloration, in the Lake Victoria cichlid Pundamilia nyererei. P Roy. Soc. Lond. B Bio., 2004, 271, 2445–2452. [Google Scholar] [CrossRef] [PubMed]
- Uetz, P.; Freed, P.; Aguilar, R.; Hošek, J. (2023). The Reptile Database. Access 15-06-2023.
- Abdala, C.; Laspiur, A.; Scrocchi, G.; Semhan, R.; Lobo, F.; Valladares, P. Las Lagartijas de la Familia Liolaemidae. Sistemática, Distribución e Historia Natural de una de las Familias de Vertebrados más Diversas del Cono Sur de Sudamérica. V1, V2. RIL Editores, Universidad de Tarapacá: Santiago, 2021.
- Esquerré, D.; Brennan, I.G.; Catullo, R.A.; Torres-Pérez, F.; Keogh, J.S. How mountains shape biodiversity: The role of the Andes in biogeography, diversification, and reproductive biology in South America’s most species-rich lizard radiation (Squamata: Liolaemidae). Evolution, 2019, 73, 214–230. [Google Scholar] [CrossRef] [PubMed]
- Esquerré, D.; Ramírez-Álvarez, D.; Pavón-Vázquez, C.J.; Troncoso-Palacios, J.; Garín, C.F.; Keogh, J.S.; Leaché, A.D. Speciation across mountains: Phylogenomics, species delimitation and taxonomy of the Liolaemus leopardinus clade (Squamata, Liolaemidae). Mol. Phylogenet. Evol., 2019, 139 106524. [CrossRef]
- Torres-Pérez, F.; Mendez, M.A.; Benavides, E.; Moreno, R.A.; Lamborot, M.; Palma, R.E.; Ortiz, J.C. Systematics and evolutionary relationships of the mountain lizard Liolaemus monticola (Liolaemini): How morphological and molecular evidence contributes to reveal hidden species diversity. Biol. J. Linnean Soc., 2009, 96, 635–650. [Google Scholar] [CrossRef]
- Muñoz-Mendoza, C.; D'Elía, G.; Panzera, A.; Méndez T, M.A.; Villalobos-Leiva, A.; Sites, J.W.; Victoriano, P.F. Geography and past climate changes have shaped the evolution of a widespread lizard from the Chilean hotspot. Mol. Phylogenet. Evol., 2017, 116, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Cianferoni, F.; Yáñez, R.P.; Eduardo, R.E.; Garin, C.F.; Torres-Pérez, F. Deep divergences within Liolaemus nigroviridis (Squamata, Liolaemidae) lineages associated with sky islands in central Chile. Zootaxa, 2013, 3619. [CrossRef]
- Boschman, L.M.; Condamine, F.L. Mountain radiations are not only rapid and recent: Ancient diversification of South American frog and lizard families related to Paleogene Andean orogeny and Cenozoic climate variations. Global Planet. Change, 2022, 208, 103704. [Google Scholar] [CrossRef]
- Vallejos Garrido, P.; Pino, K.; Espinoza-Aravena, N.; Pari, A.; Inostroza-Michael, O.; Toledo-Muñoz, M.; Castillo-Ravanal, B.; Romero-Alarcón, V.; Hernández, C.; Palma, R.; Rodríguez-Serrano, E. The importance of the Andes in the evolutionary radiation of Sigmodontinae (Rodentia, Cricetidae), the most diverse group of mammals in the Neotropics. Sci. Rep., 2023, 13. [CrossRef]
- Chaves, J.A.; Weir, J.T.; Smith, T.B. Diversification in Adelomyia hummingbirds follows Andean uplift. Mol. Ecol., 2011, 20, 4564–4576. [Google Scholar] [CrossRef]
- Singhal, S.; Colli, G.R.; Grundler, M.R.; Costa, G.C.; Prates, I.; Rabosky, D.L. No link between population isolation and speciation rate in squamate reptiles. P. Natl. Acad. Sci. -Biol., 2022, 119, e2113388119. [Google Scholar] [CrossRef]
- Barker, M.J.; Wilson, R.A. Cohesion, gene flow, and the nature of species. J. Philos., 2010, 107, 61–79 http://www.jstor.org/stable/25700483. [Google Scholar] [CrossRef]
- Smadja, C.; Butlin, R.K. On the scent of speciation: The chemosensory system and its role in premating isolation. Heredity, 2009, 102, 77–97. [Google Scholar] [CrossRef]
- Labra, A. Multi-contextual use of chemosignals by Liolaemus lizards. In: Hurst, J.L., Beynon, R.J., Roberts, S.C., Wyatt, T.D., eds. Chemical Signals in Vertebrates XI. New York, USA: SpringerLink, 2008; pp. 357-365.
- Labra, A. Comunicación, el mediador de las interacciones sociales en animales. In: Ebensperger, L.A., Labra, A., eds. Comportamiento Social de la Fauna Nativa de Chile. Santiago, Chile: Ediciones UC, 2020; pp. 263-273.
- Labra, A. Chemoreception and the assessment of fighting abilities in the lizard Liolaemus monticola. Ethology, 2006, 112, 993–999. [Google Scholar] [CrossRef]
- Labra, A. Chemical stimuli and species recognition in Liolaemus lizards. J. Zool., 2011, 285, 215–221. [Google Scholar] [CrossRef]
- Labra, A. The chemical-speciation hypothesis in Liolaemus: A response to Pincheira-Donoso. J. Zool., 2012, 288, 234–236. [Google Scholar] [CrossRef]
- Valdecantos, S.; Labra, A. Testing the functionality of precloacal secretions from both sexes in the South American lizard, Liolaemus chiliensis. Amphibia-Reptilia, 2017, 38, 209–216. [Google Scholar] [CrossRef]
- Labra, A. Sistemas de comunicación en reptiles. In: Vidal, M.A., Labra, A., eds. Herpetología de Chile. Santiago, Chile: Science Verlag, 2008; pp. 547-577.
- Labra, A.; Brann, J.H.; Fadool, D.A. Heterogeneity of voltage-and chemosignal-activated response profiles in vomeronasal sensory neurons. J. Neurophysiol., 2005, 94, 2535–2548. [Google Scholar] [CrossRef] [PubMed]
- García-Roa, R.; Carreira, S.; López, P.; Martín, J. Genders matters: Sexual differences in chemical signals of Liolaemus wiegmannii lizards (Iguania, Liolaemidae). Biochem. Syst. Ecol., 2016, 69, 108–114. [Google Scholar] [CrossRef]
- Escobar, C.A.; Labra, A.; Niemeyer, H.M. Chemical composition of precloacal secretions of Liolaemus lizards. J. Chem. Ecol., 2001, 27, 1677–1690. [Google Scholar] [CrossRef]
- Escobar, C.; Escobar, C.A.; Labra, A.; Niemeyer, H.M. Chemical composition of precloacal secretions of two Liolaemus fabiani populations: Are they different? J. Chem. Ecol., 2003, 29, 629–638. [Google Scholar] [CrossRef]
- Runemark, A.; Gabirot, M.; Svensson, E. Population divergence in chemical signals and the potential for premating isolation between islet-and mainland populations of the Skyros wall lizard (Podarcis gaigeae). J. Evol. Biol., 2011, 24, 795–809. [Google Scholar] [CrossRef]
- Raya-García, E.; Suazo-Ortuño, I.; Campos-García, J.; Martín, J.; Alvarado-Díaz, J.; Mendoza-Ramírez, E. Chemical signal divergence among populations influences behavioral discrimination in the whiptail lizard Aspidoscelis lineattissimus (Squamata: Teiidae). Behav. Ecol. Sociobiol., 2020, 74, 144. 10.1007/s00265–020. [Google Scholar] [CrossRef]
- Victoriano, P.F.; Ortiz, J.C.; Benavides, E.; Adams, B.J.; Sites, J.W. Comparative phylogeography of codistributed species of Chilean Liolaemus (Squamata: Tropiduridae) from the central-southern Andean range. Mol. Ecol., 2008, 17, 2397–2416. [Google Scholar] [CrossRef] [PubMed]
- Baeckens, S.; Martín, J.; García-Roa, R.; Pafilis, P.; Huyghe, K.; Van Damme, R. Environmental conditions shape the chemical signal design of lizards. Funct. Ecol., 2018, 32, 566–580. [Google Scholar] [CrossRef]
- Zozaya, S.M.; Teasdale, L.C.; Moritz, C.; Higgie, M.; Hoskin, C.J. Composition of a chemical signalling trait varies with phylogeny and precipitation across an Australian lizard radiation. J. Evol. Biol., 2022, 35, 919–933. [Google Scholar] [CrossRef] [PubMed]
- Campos, S.M.; Pruett, J.A.; Soini, H.A.; Zúñiga-Vega, J.J.; Goldberg, J.K.; Vital-García, C.; Hews, D.K.; Novotny, M.V.; Martins, E.P. Volatile fatty acid and aldehyde abundances evolve with behavior and habitat temperature in Sceloporus lizards. Behav. Ecol., 2020, 31, 978–991. [Google Scholar] [CrossRef] [PubMed]
- Victoriano, P.F. Phylogeography of Chilean lizards: Histories of genetic diversification on the western slope of southern Andes. In: Morando, M., Avila, L.J., eds. Lizards of Patagonia. Diversity, Systematics, Biogeography and Biology of the Reptiles at the End of the World. Switzerland: Springer, 2020; pp. 255-291.
- Lamborot, M.; Alvarez-Sarret, E. Karyotypic variation within and between populations of Liolaemus monticola (Tropiduridae) separated by the Maipo River in the Coastal Range of Central Chile. Herpetologica, 1993, 49, 435–449 http://wwwjstororg/stable/3892638. [Google Scholar]
- Lamborot, M.; Eaton, L. The Maipo River as a biogeographical barrier to Liolaemus monticola (Tropiduridae) in the mountain ranges of central Chile. J. Zool. Syst. Evol. Res., 1997, 35, 105–111. [Google Scholar] [CrossRef]
- Labra, A.; Niemeyer, H.M. Variability in the assessment of snake predation risk by Liolaemus lizards. Ethology, 2004, 110, 649–662. [Google Scholar] [CrossRef]
- Labra, A.; Pienaar, J.; Hansen, T.F. Evolution of thermal physiology in Liolaemus lizards: Adaptation, phylogenetic inertia, and niche tracking. Am. Nat., 2009, 174, 204–220. [Google Scholar] [CrossRef]
- Halpern, M.; Martínez-Marcos, A. Structure and function of the vomeronasal system: an update. Prog. Neurobiol., 2003, 70, 245–318. [Google Scholar] [CrossRef]
- Price, T. Sexual selection and natural selection in bird speciation. Philos. T. Roy. Soc. B, 1998, 353, 251–260. [Google Scholar] [CrossRef]
- Weir, J.T.; Price, T.D. Song playbacks demonstrate slower evolution of song discrimination in birds from Amazonia than from temperate North America. PLoS Biol., 2019, 17, e3000478. [Google Scholar] [CrossRef] [PubMed]
- Labra, A.; Escobar, C.A.; Niemeyer, H.M. Chemical discrimination in Liolaemus lizards: Comparison of behavioral and chemical data. In: Marchelewska-Koj, A., Lepri, J.J., Müller-Schwarze, D., eds. Chemical Signals in Vertebrates IX. New York, USA: Kluwer Academic/Plenum Publishers, 2001; pp. 439-444.
- Wyatt, T. Pheromones and signature mixtures: Defining species-wide signals and variable cues for identity in both invertebrates and vertebrates. J. Comp. Physiol. A, 2010, 196, 685–700. [Google Scholar] [CrossRef] [PubMed]
- Panzera, A.; Leaché, A.D.; D'Elía, G.; Victoriano, P.F. Phylogenomic analysis of the Chilean clade of Liolaemus lizards (Squamata: Liolaemidae) based on sequence capture data. PeerJ 2017, 5, e3941. [Google Scholar] [CrossRef] [PubMed]
- Zozaya, S.M.; Higgie, M.; Moritz, C.; Hoskin, C.J. Are pheromones key to unlocking cryptic lizard diversity? Am. Nat., 2019, 194, 168–182. [Google Scholar] [CrossRef] [PubMed]
- Mangiacotti, M.; Fumagalli, M.; Scali, S.; Zuffi, M.A.; Cagnone, M.; Salvini, R.; Sacchi, R. Inter-and intra-population variability of the protein content of femoral gland secretions from a lacertid lizard. Curr. Zool., 2016, 63, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Ortega, J.; Martín, J.; Crochet, P.A.; López, P.; Clobert, J. Seasonal and interpopulational phenotypic variation in morphology and sexual signals of Podarcis liolepis lizards. PLoS ONE, 2019, 14, e0211686. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez, A.; Menke, M.; Quezada, G. Gustavo Jiménez-Uzcátegui; Schulz, S.; Steinfartz, S. Diversity of compounds in femoral secretions of Galápagos iguanas (genera: Amblyrhynchus and Conolophus), and their potential role in sexual communication in lek-mating marine iguanas (Amblyrhynchus cristatus). PeerJ, 2017, 5:e3689. [Google Scholar] [CrossRef]
- Gabirot, M.; López, P.; Martín, J. Differences in chemical sexual signals may promote reproductive isolation and cryptic speciation between Iberian wall lizard populations. Intern. J. Evol. Biol., 2012, 2012, ID 698520. [Google Scholar] [CrossRef]
- MacGregor, H.E.A.; Lewandowsky, R.A.M.; d'Ettorre, P.; Leroy, C.; Davies, N.W.; While, G.M.; Uller, T. Chemical communication, sexual selection, and introgression in wall lizards. Evolution, 2017, 71, 2327–2343. [Google Scholar] [CrossRef]
- Martín, J.; López, P. Interpopulational differences in chemical composition and chemosensory recognition of femoral gland secretions of male lizards Podarcis hispanica: Implications for sexual isolation in a species complex. Chemoecology, 2006, 16, 31–38. [Google Scholar] [CrossRef]
- Martín, J.; López, P.; Iraeta, P.; Díaz, J.A.; Salvador, A. Differences in males’ chemical signals between genetic lineages of the lizard Psammodromus algirus promote male intrasexual recognition and aggression but not female mate preferences. Behav. Ecol. Sociobiol., 2016, 70, 1657–1668. [Google Scholar] [CrossRef]
- Barley, A.J.; Nieto-Montes de Oca, A.; Reeder, T.W.; Manríquez-Morán, N.L.; Monroy, J.C.; Hernández-Gallegos, O.; Thomson, R.C. Complex patterns of hybridization and introgression across evolutionary timescales in Mexican whiptail lizards (Aspidoscelis). Mol. Phyl. Evol., 2019, 132, 284–295. [Google Scholar] [CrossRef] [PubMed]
- Butlin, R.; Debelle, A.; Kerth, C.; Snook, R.R.; Beukeboom, L.W.; Cajas, R.C.; Diao, W.; Maan, M.E.; Paolucci, S.; Weissing, F.J. What do we need to know about speciation? Trends Ecol. Evol., 2012, 27, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Esquerré, D.; Keogh, J.S.; Demangel, D.; Morando, M.; Avila, L.J.; Sites Jr, J.W.; Ferri-Yáñez, F.; Leaché, A.D. Rapid radiation and rampant reticulation: Phylogenomics of South American Liolaemus lizards. Syst. Biol., 2022, 71, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Morando, M.; Avila, L.J.; Baker, J.; Sites, J.W. Phylogeny and phylogeography of the Liolaemus darwinii complex (Squamata : Liolaemidae): Evidence for introgression and incomplete lineage sorting. Evolution, 2004, 58, 842–861. [Google Scholar] [CrossRef]
- Morando, M.; C.D., M.; Minoli, I.; Pérez, C.H.F.; Sites, J.W.J.; Avila, L.J. Diversification and evolutionary histories of Patagonian steppe lizards. In: Morando, M., Avila, L.J., eds. Lizards of Patagonia. Diversity, Systematics, Biogeography and Biology of the Reptiles at the End of the World. Cham, Switzerland: SpringerNature Switzerland, 2020; pp. 217–254.
Figure 1.
Geographic localization of the populations of Liolaemus tenuis and L. lemniscatus in central Chile.
Figure 1.
Geographic localization of the populations of Liolaemus tenuis and L. lemniscatus in central Chile.
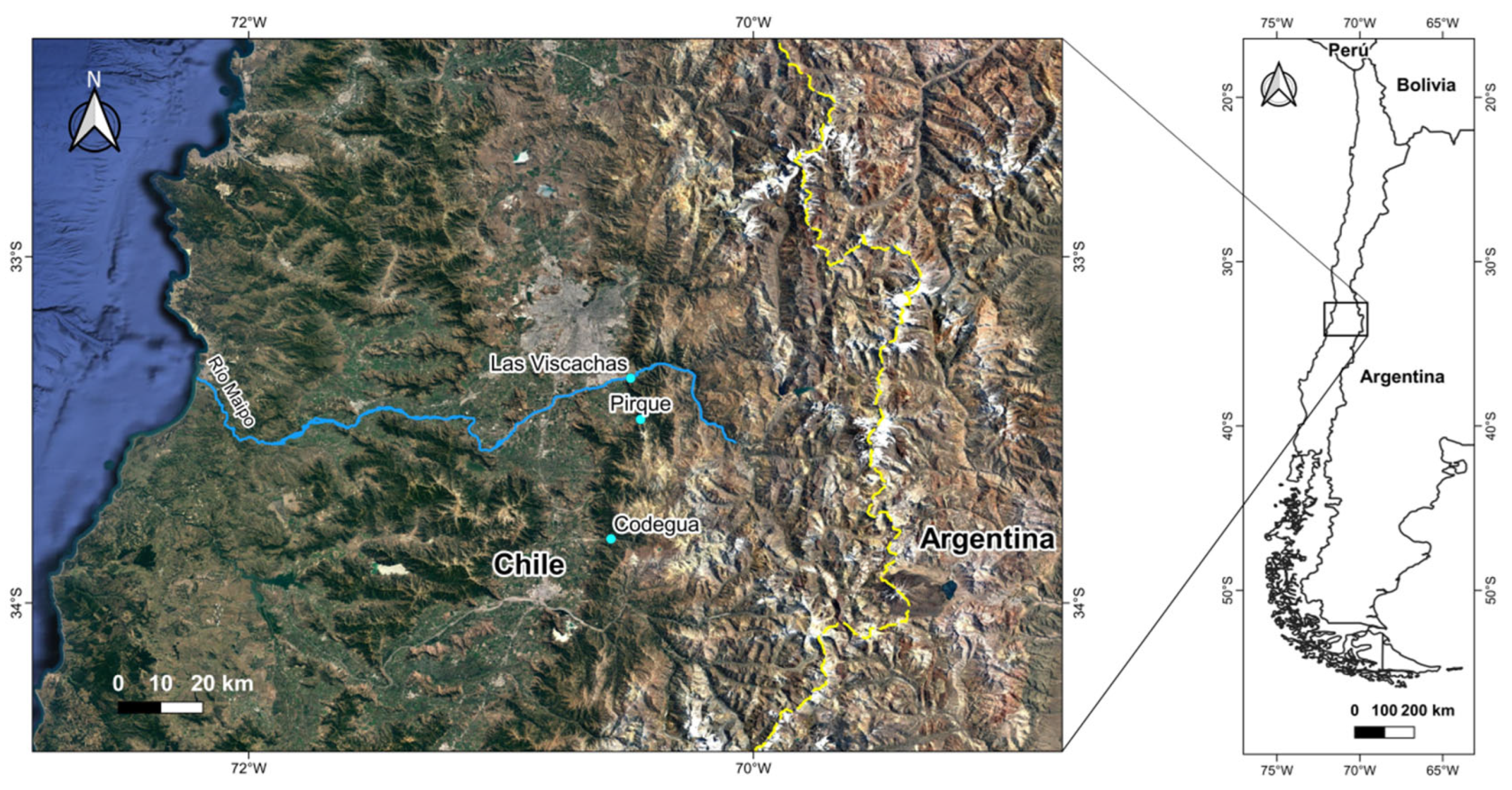
Figure 2.
Mean (+SE) values of three behaviors recorded in males of Liolaemus tenuis from Codegua when exposed to male conspecific scents from the same population (homotypic), and two heterotypic populations, Pirque and Las Viscachas. A- Latency to the first tongue flick, B- Number of tongue flicks, and C- Motion time (s). * = p < 0.05.
Figure 2.
Mean (+SE) values of three behaviors recorded in males of Liolaemus tenuis from Codegua when exposed to male conspecific scents from the same population (homotypic), and two heterotypic populations, Pirque and Las Viscachas. A- Latency to the first tongue flick, B- Number of tongue flicks, and C- Motion time (s). * = p < 0.05.
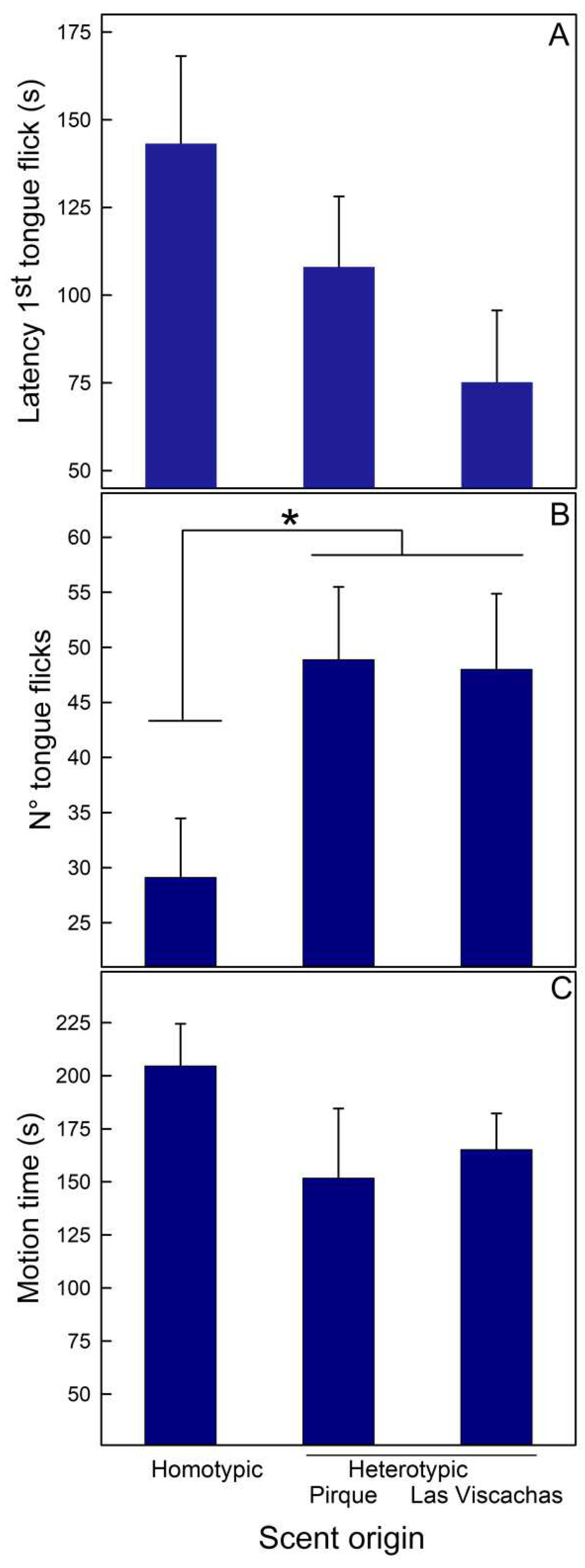
Figure 3.
Mean (+SE) values of four behaviors recorded in males of Liolaemus lemniscatus from two populations (Codegua and Las Viscachas) when they were exposed to scents of homotypic or heterotypic conspecific males. Data from both populations were pooled. A- Latency to the first tongue flick, B- Number of tongue flicks, C- Motion time (s), and D- Head display (s). * = p < 0.05.
Figure 3.
Mean (+SE) values of four behaviors recorded in males of Liolaemus lemniscatus from two populations (Codegua and Las Viscachas) when they were exposed to scents of homotypic or heterotypic conspecific males. Data from both populations were pooled. A- Latency to the first tongue flick, B- Number of tongue flicks, C- Motion time (s), and D- Head display (s). * = p < 0.05.
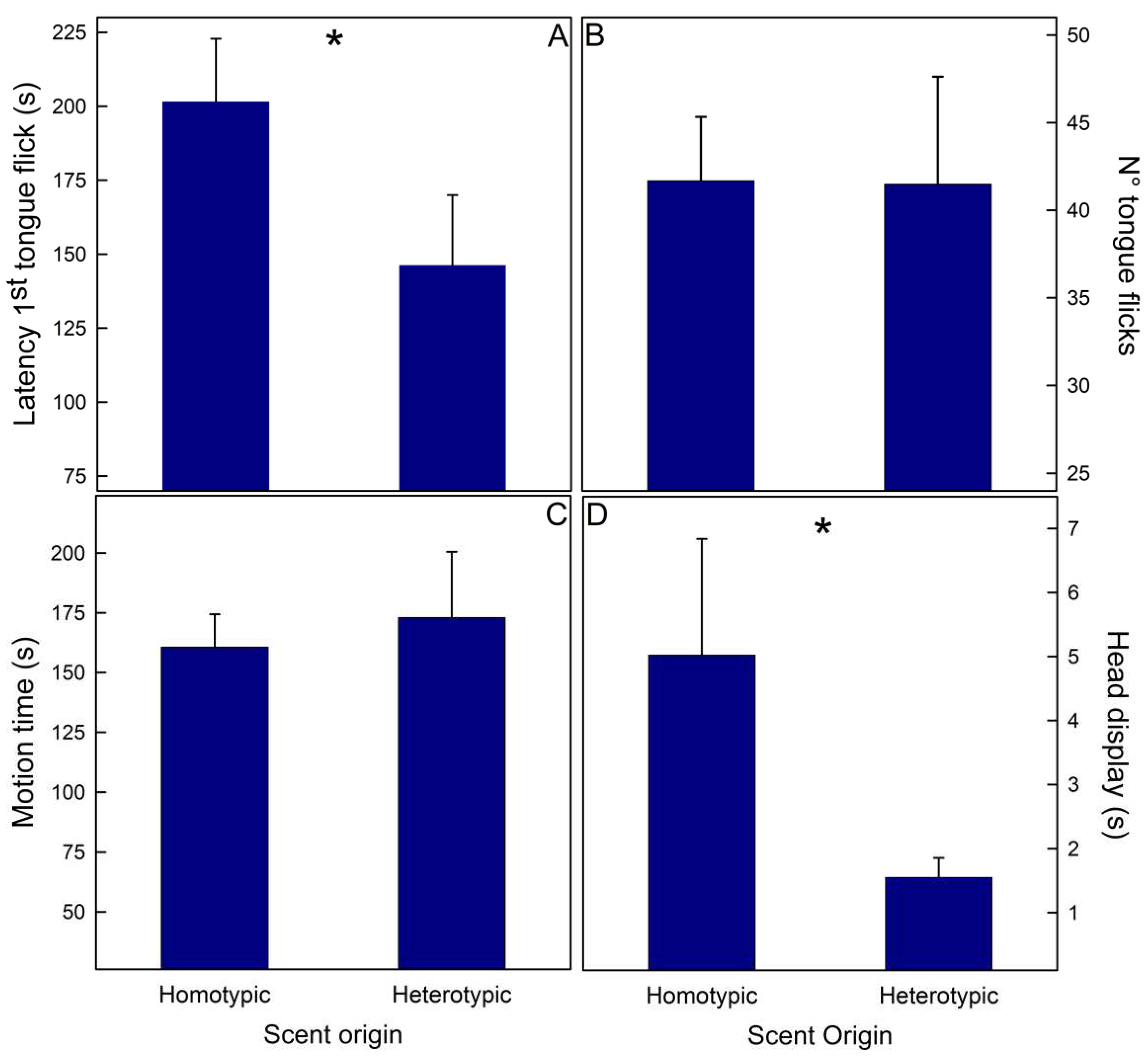
Figure 4.
Mean (+SE) values of behaviors exhibited by lizards in enclosures with scents from homo- and heterotypic potential sexual partners. Liolaemus tenuis. A- Time (min) spent on each side of the enclosure, on the pot with scents, and moving in each side of the enclosure. B- Number of tongue flicks displayed at each side of the enclosure and directed to the pot. Data from both populations and sexes were pooled. C- Liolaemus lemniscatus. Effect of the interaction between Scent origin and Sex upon the Time (min) spent in the pot with scents from different origins (left) and the number of tongue flicks directed to those pots (right). Data from both populations were pooled. * = p < 0.05.
Figure 4.
Mean (+SE) values of behaviors exhibited by lizards in enclosures with scents from homo- and heterotypic potential sexual partners. Liolaemus tenuis. A- Time (min) spent on each side of the enclosure, on the pot with scents, and moving in each side of the enclosure. B- Number of tongue flicks displayed at each side of the enclosure and directed to the pot. Data from both populations and sexes were pooled. C- Liolaemus lemniscatus. Effect of the interaction between Scent origin and Sex upon the Time (min) spent in the pot with scents from different origins (left) and the number of tongue flicks directed to those pots (right). Data from both populations were pooled. * = p < 0.05.
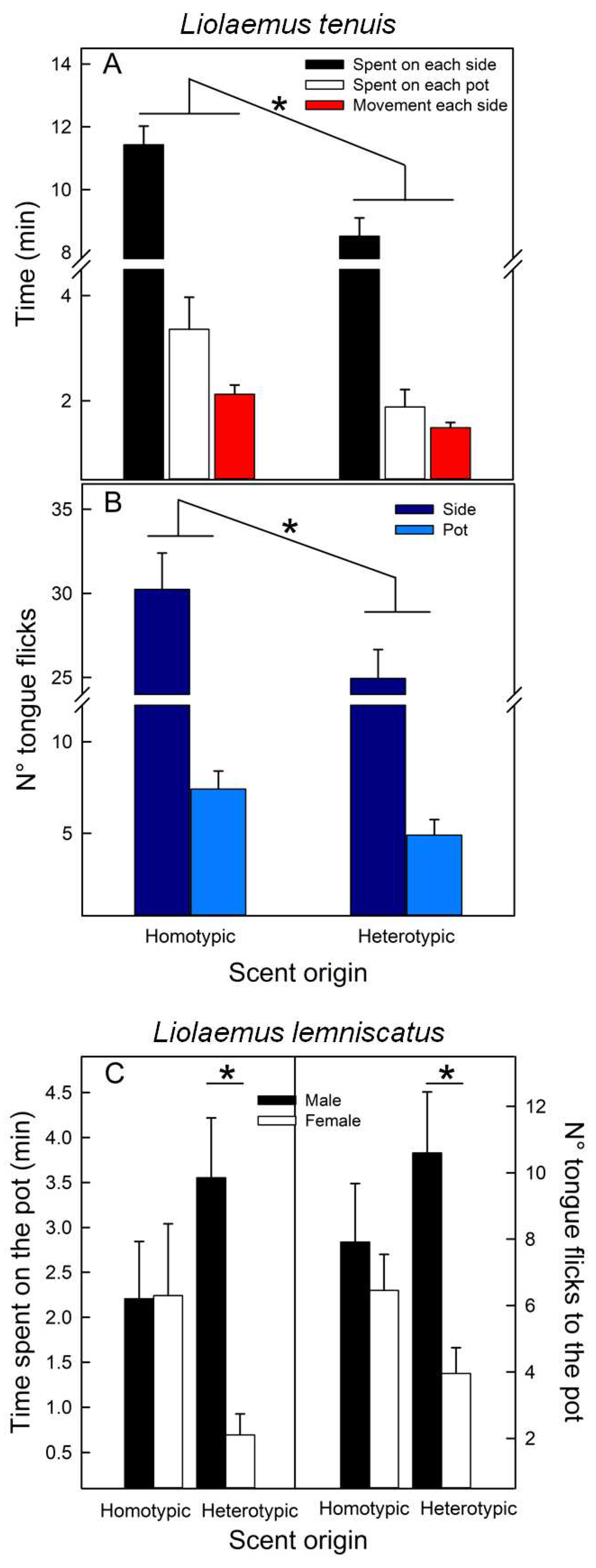
Figure 5.
Mean untransformed (+SE) values of the Permanence (%) at each side of the enclosure contained clay pots with scents from homotypic and heterotypic conspecifics, of Liolaemus tenuis and L. lemniscatus, from two populations, Codegua and Las Viscachas. * = p < 0.05.
Figure 5.
Mean untransformed (+SE) values of the Permanence (%) at each side of the enclosure contained clay pots with scents from homotypic and heterotypic conspecifics, of Liolaemus tenuis and L. lemniscatus, from two populations, Codegua and Las Viscachas. * = p < 0.05.
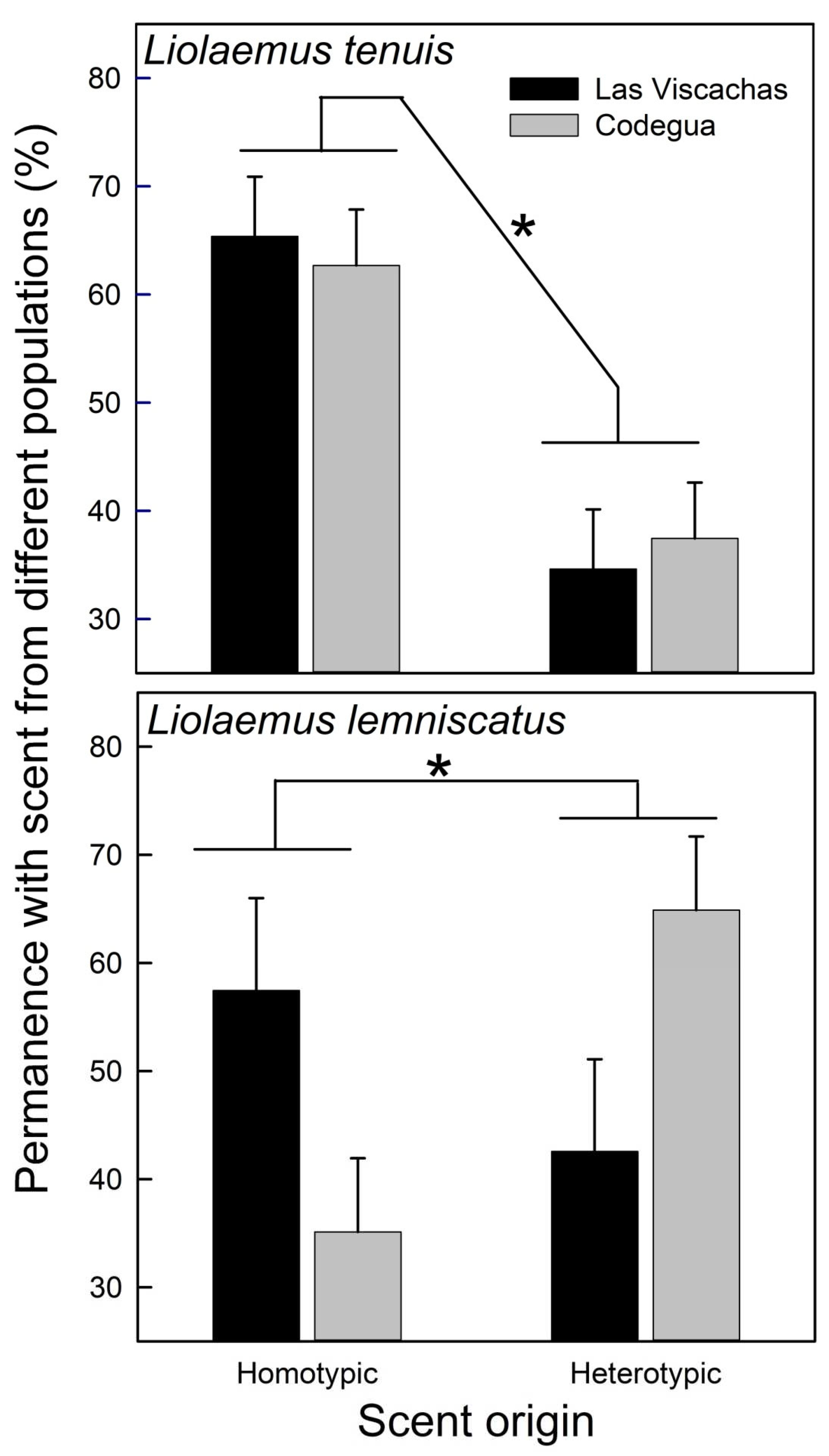

Table 1.
Results of the General Linear Models for repeated measures to test male scent discrimination of homo- and heterotypic competitors (Scent origin) in two species. A. Liolaemus tenuis. Three responses of the focal population (Codegua) were exhibited with scents from the own population and two others. Sample size: nine. B. Liolaemus lemniscatus. The analyses included the effect of Population (Codegua and Las Viscachas, the two focal populations) and its interaction with Scent origin upon four responses. Total sample size: 16, eight from each population. Shown values are the F-statistics [degree of freedom] (p-value); statistically significant tests (p < 0.05) are shown in bold.
Table 1.
Results of the General Linear Models for repeated measures to test male scent discrimination of homo- and heterotypic competitors (Scent origin) in two species. A. Liolaemus tenuis. Three responses of the focal population (Codegua) were exhibited with scents from the own population and two others. Sample size: nine. B. Liolaemus lemniscatus. The analyses included the effect of Population (Codegua and Las Viscachas, the two focal populations) and its interaction with Scent origin upon four responses. Total sample size: 16, eight from each population. Shown values are the F-statistics [degree of freedom] (p-value); statistically significant tests (p < 0.05) are shown in bold.
| A- Liolaemus tenuis F[2,16] (p) | ||||||
| Variable | Scent origin | |||||
| Latency 1st tongue flick (s) | 1.909 (0.181) | |||||
| N° tongue flicks | 4.757 (0.024) | |||||
| Motion time (s) | 1.048 (0.374) | |||||
| B- Liolaemus lemniscatus F[1,14](p) | ||||||
| Variable | Scent origin | Population | Scent origin*Population | |||
| Latency 1st tongue flick (s) | 5.147 (0.039) | 0.098 (0.756) | 0.476 (0.502) | |||
| N° tongue flicks | 0.001 (0.978) | 0.321 (0.580) | 1.778 (0.204) | |||
| Motion time (s) | 0.311 (0.586) | 0.186 (0.673) | 1.872 (0.193) | |||
| Head displays (s) | 6.494 (0.023) | 1.284 (0.276) | 1.984 (0.181) | |||
| A- Liolaemus tenuis F[1,38](p) | ||||||
| Scent origin | Population | Sex | Scent origin*Pop. | Scent origin*Sex | ||
| Time on each side (min) | 5.968 (0.019) | 0.432 (0.515) | 3.304 (0.077) | 0.001 (0.982) | 0.386 (0.538) | |
| Time on each pot (min) | 4.261 (0.046) | 0.501(0.483) | 3.578(0.066) | 0.029 (0866) | 0.371 (0.546) | |
| Tongue flicks each side | 4.110 (0.050) | 0.522(0.474) | 0.247(0.622) | 0.456 (0.504) | 0.754 (0.391) | |
| Tongue flicks each pot | 4.902 (0.033) | 0.306 (0.583) | 0.000 (0.980) | 0.523 (0.474) | 0.247 (0.622) | |
| Motion each side (min) | 12.854 (<0.001) | 4.472 (0.041) | 5.485 (0.025) | 0.649 (0.425) | 0.242 (0.626) | |
Table 2.
Results of the General Linear Models for repeated measures to test responses to homo- and heterotypic potential sexual partner (Scent origin). Analyses included the Sex (male vs. female) and Population (Codegua vs. Las Viscachas) of the focal populations and their interactions modulating five responses. A. Liolaemus tenuis. B. Liolaemus lemniscatus. Shown values are the F-statistics [degree of freedom] (p-value); statistically significant tests (p < 0.05) are shown in bold.
Table 2.
Results of the General Linear Models for repeated measures to test responses to homo- and heterotypic potential sexual partner (Scent origin). Analyses included the Sex (male vs. female) and Population (Codegua vs. Las Viscachas) of the focal populations and their interactions modulating five responses. A. Liolaemus tenuis. B. Liolaemus lemniscatus. Shown values are the F-statistics [degree of freedom] (p-value); statistically significant tests (p < 0.05) are shown in bold.
| B- Liolaemus lemniscatus F[1,54](p) | |||||
| Scent origin | Population | Sex | Scent origin*Pop. | Scent origin*Sex | |
| Time on each side (min) | 0.373 (0.544) | 1.049 (0.310) | 0.119 (0.731) | 0.086 (0.771) | 1.583 (0.214) |
| Time on each pot (min) | 0.103 (0.750) | 0.570(0.453) | 4.454 (0.039) | 0.428 (0.516) | 4.566 (0.037) |
| Tongue flicks each side | 0.738 (0.394) | 1.223 (0.274) | 0.601 (0.442) | 0.186 (0.668) | 1.127 (0.293) |
| Tongue flicks each pot | 0.005 (0.945) | 0.633 (0.430) | 3.871 (0.054) | 0.001 (0.971) | 5.233 (0.026) |
| Motion each side (min) | 0.130 (0.719) | 0.785 (0.380) | 1.535 (0.221) | 0.485 (0.489) | 0.551 (0.461) |
Table 3.
Results of the General Linear Models for repeated measures to test preference for homo- and heterotypic potential sexual partner (Scent origin). Analyses included the Sex (male vs. female) and Population (Codegua vs. Las Viscachas) of the focal individuals of the two populations and the factor interactions upon scent preference. Shown values are the F-statistics [degree of freedom] (p-value); statistically significant tests (p < 0.05) are shown in bold.
Table 3.
Results of the General Linear Models for repeated measures to test preference for homo- and heterotypic potential sexual partner (Scent origin). Analyses included the Sex (male vs. female) and Population (Codegua vs. Las Viscachas) of the focal individuals of the two populations and the factor interactions upon scent preference. Shown values are the F-statistics [degree of freedom] (p-value); statistically significant tests (p < 0.05) are shown in bold.
| Variables | Liolaemus tenuis F[1,26](p) |
Liolaemus lemniscatus F[1,26](p) |
|---|---|---|
| Scent origin | 11.953 (0.002) | 0.262(0.613) |
| Population | 1.934 (0.176) | 0.000 (1.000) |
| Sex | 1.934 (0.176) | 0.000 (1.000) |
| Scent origin * Population | 0.048 (0.827) | 4.837(0.037) |
| Scent origin * Sex | 1.867 (0.184) | 1.814 (0.190) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



