
Submitted:
15 May 2023
Posted:
18 May 2023
You are already at the latest version
Abstract
The objective of this study was to evaluate the effect of the addition of atomized black Maca in the cryopreservation medium of epididymal spermatozoa of alpaca species. Fifteen pares of testes and epididymis from alpacas of reproductive age were collected from a local (Huancavelica) slaughterhouse and transported in NaCl 0,9% at 10°C to the laboratory. Each sample of epididymal sperm was divided into six groups: 1) Fresh, 2) Yolk medium (YM), 3) YM+10mg/ml Maca, 4) YM+20mg/ml Maca, 5) YM+30mg/ml Maca, and 6) YM+ Resveratrol (antioxidant positive control). Sperm cryopreservation was performed through the slow-freezing method. Markers associated with functionality such as motility, vitality and plasma membrane integrity. Cellular markers associated with oxidative damage, such as DNA fragmentation, total ROS production and lipid peroxidation, were analyzed before and after freezing. The results showed that the supplementation with black Maca at 20mg/ml improved the sperm motility, viability and integrity of the membrane.
Similarly, total ROS production decreased with Maca at 10 mg/ml (25 ± 2.23%) and 20mg/ml (20 ± 2.52%) (P<0.05). However, DNA fragmentation and lipid peroxidation were similar in all groups. These results suggest that Maca at 20mg/ml has a protective effect during the freezing/thawing process of epididymal sperm of alpaca species.
Keywords:
Freezing
; black Maca
; sperm parameters
; alpaca
; sperm
1. Introduction
Peru is the world's leading producer of alpacas (Vicugna pacos) and the second of llamas (Lama glama). [1]. Due to the economic importance that these species represent several groups have focused the on the conservation and freezing of semen of South American camelids in Peru. Specifically, the alpaca species has been used as a model for cryopreservation of semen, in vitro fertilization (IVF), and artificial insemination s; however, the expected outcomes have not been reached such as in other domestic species [2]
Gamete freezing, specifically cryopreservation of mammalian spermatozoa, is an important tool in assisted reproduction, safeguarding the species' genetic pedigree, it allows long-term preservation of genetic resources, and/or preservation of endangered species [3]. Nevertheless, cryopreservation of alpaca spermatozoa has not been used extensively because of its limiting step of semen collection and poor freezability [4]. associated with specific semen characteristics, such as low sperm count, a high percentage of abnormalities, and the viscosity of seminal plasma [5].
During cryopreservation, the sperm are exposed to different stressors, such as the formation of ice crystals, toxicity of the cryoprotectants and changes of temperature during chilling, freezing, and rewarming [6]. These changes lead to osmotic and thermal shock and oxidative stress, which are among the main biophysical factors affecting sperm viability [7]. Oxidative stress refers to the increase of intracellular concentrations of reactive oxygen species (ROS) [8]. Spermatozoa are susceptible to oxidative stress through all semen processing, which is why it is considered one of the main factors associated with fertilization failures [9].
Although a controlled ROS production is essential for activation of sperm motility, capacitation, and fertilization [10]. During a pathophysiological imbalance, however, ROS accumulate and react with biomolecules (i.e., membrane phospholipids, enzymes, chromatin), impairing structure and cellular functions [8,11]. ROS can induce lipid peroxidation and DNA fragmentation, affect sperm motility and their ability to fertilize [10]. Lipid peroxidation associated to OS decrease sperm motility and viability, acrosome integrity, and mitochondrial function [12].
However, it has been shown that detrimental effects of ROS imbalance can be partially prevented by adding antioxidants during in vitro manipulation of spermatozoa [13]. Therefore, antioxidant therapy by the addition of antioxidants to semen extenders has been a strategy to improve the quality of frozen-thawed sperm [14]. For instance, methionine, vitamin C (ascorbic acid), and vitamin E (α-tocopherol) have been used in semen extenders to protect the cells against cryoinjury [15]. However, dilution or cooling of these antioxidants during the cryopreservation process could reduce their benefits related to antioxidant defense.
In this scenario, a natural source of additives to preserve and enhance sperm function during semen storage is plant extracts [16]. Plants produce a wide range of chemical compounds with bioactive properties, including alkaloids, phenolic compounds, and terpenoids. For instance, resveratrol, a natural polyphenolic compound, has been extensively used as a suitable antioxidant supplement for semen extenders in human and domestic species [17]. Furthermore, these natural metabolites can have antimicrobial activities and act as ROS scavengers. Therefore, plant extracts represent a good alternative to conventional antioxidants used for cryogenic storage of semen.
Maca (Lepidium meyenii Walpers or Lepidium peruvianum Chacón), from the Brassicaceae family, it is a native Peruvian plant that grows in the Central Andes between 4,000 to 4,500 meters above sea level. The root of this plant has been used as folk medicine to increase fertility due to its high sucrose content [18] and phytochemicals [19,20,21] Several health benefits have been reported to its bioactive compounds, such as improving sexual performances in animals [22] and in humans [23], spermatogenesis [21,24], energizing anti-stress and antioxidant effects [25,26,27], natural cytoprotection [28,29] and cytoprotection against oxidative stress conditions [25,30].
In addition to the high concentration of antioxidants, L. meyenii contains several secondary metabolites that help keep ROS levels low, which is why the cytoprotective effects of Maca are associated with its antioxidant activity by increasing the enzymatic activity of superoxide dismutase. and through the direct elimination of free radicals [31]. Studies have shown that Maca secondary metabolites participate in the hydroxyl radical scavenging and free radical scavenging capacity of 2,2-diphenyl-1-picrylhydrazyl, thus increasing superoxide dismutase and glutathione activity [32].
Thus, regarding the valuable cytoprotective and antioxidant capacities of Maca, the purpose of this study was to evaluate the effects of atomized extract of Maca during cryopreservation of epidydimal alpaca spermatozoa.
2. Materials and Methods
2.1. Sample Collection
Fifteen pairs of alpaca testicles and epididymal of alpacas from between 4 to 6 years old were obtained from the Municipal slaughterhouse, Huancavelica (3000-3700 m.s.n.m) Peru. The samples were stored in 0,9% NaCl at 10°C and transported for 22 hours to the laboratory. The sample collections were carried out once every two weeks for four months.
2.2. Obtaining the Epididymal Sperm
At the laboratory, we proceeded to isolate from the epididymal tail region, which tail regions were used, which were removed and cut into small pieces in 1ml of 0,9% NaCl 9% at 37°C. Then the suspension containing the spermatozoa was transferred to 1.5 ml plastic tubes, and they were placed in an incubator at 37°C for fifteen minutes to allow the spermatozoa to recover. After this time, the sample will be separated into two parts. The first was to determine the conventional parameters such as motility, viability and integrity of the plasma membrane; then, the sample was washed with PBS using centrifugation to remove tissue remains, leaving only the sperm. To carry out the cryopreservation of the spermatozoa, the exclusion criterion was that the motility value was more significant than or equal to 70%.
2.3. Sperm Cryopreservation and Maca Supplementation
The medium used for cryopreservation was Tris Yolk Medium (YM) with 10% fetal bovine serum (FBS) containing 0.25M Fructose and 10% Dimethyl sulfoxide additionally it was supplemented with the atomized hydroalcoholic (methanolic) extract of commercial black Maca powder (JUVENS® Cayenatur, Lima, Peru) as described by the Research Circle of Plants with Effect on Health (Grant no. 010-2014-FONDECYT). This medium was made into four treatments where black Maca was added at three concentrations of 10mg/ml, 20mg/ml and 30mg/ml, and 5mg/ml Resveratrol (Sigma, St. Louis, MO, USA) as antioxidant control.
2.4. Evaluation of Sperm Parameters
2.4.1. Sperm Motility
Sperm motility was evaluated according to the criteria of the World Health Organization [33]. 10µl of the sperm sample was placed on a slide previously tempered at 37ºC and observed under a field microscope. Rapidly plus slowly progressive motility with forward movement was considered Progressive motility. The values were expressed in percentages based on 100 spermatozoa from the observed field.
2.4.2. Sperm Viability
On a slide previously tempered at 37ºC, 5µl of the sperm sample was analyzed and then 5µl of eosin Y (tempered at 37ºC) was added, both drops were mixed, and the viability was evaluated after 2 minutes. It was placed on a coverslip and observed under a light microscope at 400X magnification. It was carried out using the vital dye Eosin Y at 0.5% to evaluate the viability of the spermatozoa and find the ratio of live and dead (orange head) spermatozoa.
2.4.3. Plasma Membrane Integrity
The evaluation was through the hypoosmotic test, also known as the HOST test, a hypoosmotic solution was prepared with a final osmolarity of 100mOsm. 30μl of the sperm preparation was incubated with 270μl of the hypoosmotic solution for 30 minutes at 37°C. The proportion of spermatozoa showing the typical curling of the tail (caused by swelling) concerning the total number of spermatozoa evaluated was evaluated. The result was expressed as a percentage of spermatozoa with an intact membrane, that is, the percentage of spermatozoa that responded to the hypoosmotic test.
2.5.1. Sperm Mitochondrial Activity
It was analyzed using Nitro Blue Tetrazolium (NBT) to measure mitochondrial activity. A 10μL drop of spermatozoa homogenate was mixed with 10μL of 1X NBT on a slide pre-heated to 37°C on a heated stage. The oxidation-reduction system was placed in an anaerobic chamber for 45 minutes at 37°C. Spermatozoas were evaluated under a microscope with the immersion objective 100X; the mitochondrial activity was determined according to the classification in percentages proposed by Hrudka [34].
2.5.2. DNA Fragmentation Analysis
The evaluation was carried out using the commercial terminal deoxynucleotidyl transferase-mediated dUTP nick-end labelling (TUNEL) kit (In situ cell death detection Kit, Fluorescein Roche® Mannheim, BW, Germany). The sample's treatment was described by Santiani [35] with some modifications. A sperm suspension of 2x106 spermatozoa /ml was used to be then fixed in 4% formaldehyde for 15 minutes at 4°C. Subsequently, two washes were carried out with phosphate saline solution (PBS). Next, the samples were permeabilizated, resuspending in 100μl with 0.5% Triton X-100 diluted in 0.1% Sodium Citrate for 30 minutes at room temperature. The permeabilization solution was removed by washing by centrifugation in PBS.
The samples under study were resuspended in 50μl of TUNEL working solution and incubated for 60 minutes in a humid and dark chamber at room temperature. After this time, they were resuspended in 500μl of PBS and washed at 500 g for 6 minutes. The pellets were resuspended in 400 µl of PBS and 2μl of propidium iodide (2.4mM stock solution). The samples were conserved in a dark chamber at 4°C before being evaluated by fluorescence microscopy. The fragmentation index was defined as live fluorescent spermatozoa were counted for TUNEL over spermatozoa without propidium iodide fluorescence (Figure 1).
2.5.3. Evaluation of ROS Production
The evaluation of the intracellular ROS using the reactive oxygen species assay kit (Beyotime Institute of Biotechnology) was applied to measure the level of intracellular ROS following the manufacturer's instructions. 2',7'-Dichlorofluorescein diacetate (H₂DCFDA) is a cell-permeable, non-fluorescent reagent deacetylated by cellular esterases to a non-fluorescent 2',7'-dichlorofluorescein (H₂DCF) after diffusion into the cell. ROS then oxidizes the H₂DCF to a highly fluorescent 2',7'-dichlorofluorescein (DCF). The samples were conserved in a dark chamber and evaluated by fluorescence microscopy [36].
2.6. Cryopreservation Method
The straws were sealed and placed in the Thermo-controlled freezing system using the CRYOBATH system directed by the Cryogenesis Version 4.0 (Cryologics) software, the freezing protocol being as follows: The initial temperature was 4°C, the samples were kept at that temperature for five minutes. The second period was the drop in temperature from 4°C to 2°C at a rate of 3°C/min, maintaining 2°C for one minute. The third period was from 2°C to -30°C with a freezing rate of 5°C/min and finally from -30°C to -80°C with a cool rate of 8°C/min. After this point, the straws were placed in liquid nitrogen until thawed [37].
2.7. Statistical Analysis
Data were analyzed using the Shapiro-Wilk test and ANOVA for distribution. Data (mean ± SD) were compared using multiple comparisons t-test (non-parametric test). The Wilcoxon Test was used to analyze the NBT data that did not present a normal distribution. Statistical analysis and graphical representation were performed using R version R-studio 1.4.17173
Results
3.1. Effect of Maca on Sperm Parameters
Sperm motility, membrane function, viability, acrosome integrity as well as DNA integrityy, lipid peroxidation and total levels of ROS are showed in Table 1.
3.2. Effect of Maca on Sperm Motility
First, we evaluated how sperm motility was affected by the cryopreservation process and Maca supplementation. The addition of 10 mg/ml and 20 mg/ml of Maca concentrations improved sperm motility compared to the control group (YM). Sperm motility at 20mg/ml of Maca was greater (40 ± 2.45%) than Resveratrol (27 ± 1.20%) (P<0.05).
3.3. Effect of Maca on the plasma membrane integrity
The membrane integrity was significantly affected by the cryopreservation process (Fig 1); the addition of 10 mg/ml, 20 mg/ml, and 30 mg/ml of Maca improved the plasma membrane integrity compared to fresh samples. Moreover, 20mg/ml of Maca increased the viability after thawing (58.33±7.78%) compared to the control Tris-Yolk medium (P<0.05), but at levels similar to the group supplemented with Resveratrol (57.08 ± 2.57%) (Table 1).
3.4. Effect of Maca on Sperm Viability
Sperm viability was also affected by the cryopreservation process Table 1. Spermatozoa cryopreserved in YM supplemented with 20 mg/ml showed similar viability after thawing (54.58 ± 5.41%) to the group supplemented with Resveratrol (55.83 ±8.75%). In addition, 20 mg/ml of Maca and Resveratrol increased viability when compared with the control Tris-Yolk Medium (TYM) (p<0.05). Finally, fresh semen had the highest proportion of viable sperm (Table 1).
3.5. Effect of Maca on Sperm Oxidative Stress Activity
Total reactive oxygen species (ROS) emission from alpaca spermatozoa (Figure 1), increased after cryostorage in YM (40±12.6%) with respect to the initial pre-freezing (fresh) samples (29.2±10.7 %) (P<0.05). The group treated with Maca at 20mg/ml showed a significant decrease in total sperm ROS emission of (19.2 ± 7.33%) with values similar to resveratrol used as positive control (22.5±7.23%).
3.6. Effect of Maca on Sperm DNA Fragmentation
DNA fragmentation (Figure 3) was increased after freezing/thawing when compared to the Yolk medium supplemented with Maca or with the Resveratrol, which shows damage at the DNA level as a result of cryopreservation. After freezing, no significant differences (P >0.05) were observed among groups (Table 1 and Figure 2)
3.7. Effect of Maca on Lipid Peroxidation
Mitochondrial activity index and function for each sample were assesed based on the classification of formazan deposits (Figure 4), obtained from the number of cells classified in each category (Compact: number of cells classified in each category (Compact: “C”, Subcompact: “SC”, Focal: “F”, Residual: “R”). No significant differences (P>0.05) were found among all groups.
3.8. Effect of Maca on Cytochemical Activity of Oxidoreductases
Cytochemical activity rates were identified according to the amount of formazan deposited in the mitochondrial sheath (Fig. 4), where the Compact (strong and uniform staining) and Subcompact (medium to strong deposit) patterns were associated with the standard and substandard rates. Respectively, both were classified as Intensive Reactions. On the other hand, Focal (few localized deposits) and Diffuse (weak and/or absent reaction) patterns were related to Low and Residual rates as reduced reactions. The results of the comparison of sperm cytochemical activity percentages are summarized in Table 2. Among the treatments with Maca, an intensive reaction was found at 10mg/ml (61.6%), close to the values of the fresh group (70.2%) and Maca 30mg/ml (51.3%). Maca at 20mg/ml presented (38.2%), similar to the positive antioxidant control (35%), this being the lowest value (P<0.05).
On the other hand, the post-freezing groups, Yolk medium, Maca at 10mg/ml and 30mg/ml, showed a significant increase in the average Reduced Reaction (0.65%), (0.55%) and (0.608%), a higher value was found with the positive antioxidant control (2.217%) respectively about the other groups, Maca at 10mg/ml and 20mg/ml presented values of (0.55% and 0.408%) respectively, evidencing poor mitochondrial function. (P<0.05).
4. Discussion
The present study showed the effects of the supplementation of cryopreservation media with Maca (L. meyenii) on the functionality parameters of frozen epididymal alpaca sperm. Several studies have shown that Maca improved seminal quality in humans and stallions related to its high antioxidant power [38].
In spermatozoa, the plasma membrane, the mitochondrial membrane and the acrosomal membrane contain polyunsaturated fatty acids and are, therefore, very susceptible to stressors, especially during freezing procedures. Cryopreservation is one of the factors that increase cell damage by ROS and have negative effects on sperm parameters [39]. Also, during cryopreservation and thawing, and even during storage in liquid nitrogen, there is damage to the sperm due to the biophysical changes that occur in this process [40]
The components of black Maca have been extensively studied in recent years due to their antioxidant capacity. The presence of glucosinolates and macamides allows Maca to protect cells from oxidative stress damage [41]. In addition, there are various factors, such as the Maca hypocotyls colour or the use of different cryopreservation methods, processing and types of extraction (methanol, chloroform, DMSO and water) of Maca that can modify the concentrations of bioactive metabolites and influence its antioxidant capacity [42]
In this study, sperm motility was significantly affected by the cryopreservation process. However, adding Maca to the media at concentrations of 10 mg/ml, 20 mg/ml, and 30 mg/ml improved sperm motility. In addition, the alpaca spermatozoa recovered from the epididymis and post-freezing have low motility in relation to ejaculated semen. Various works on epididymal sperm cryopreservation have shown variable values of sperm viability from 21 to 32% post-thawing [4,37,44]. It has been observed that alpaca spermatozoa, whose initial motility was 46%, decreased to 24% after the freezing process, with motility being the most affected parameter in this process [37,44].
However, some researchers have chosen to study the phosphorylation pattern of spermatozoa, and it is speculated that immediately after thawing, cryopreserved semen would show a phosphorylation pattern similar to that observed in fresh semen subjected to capacitation, so it could also be an additional factor for increased sperm motility [45].
The in vitro protective effect of Lepidium meyenii on frozen-thawed bovine spermatozoa was studied, and it was found in terms of the morphology and motility test that these improved at concentrations of 1 and 10 µg/mL, respectively, but an adverse effect was also observed in morphology at the highest concentration of 100 µg/ml. However, the best activity of the Maca extract was observed on sperm DNA integrity, which was effective at the three concentrations evaluated after 24 h of incubation [46].
According to the results of the DNA fragmentation of the alpaca spermatozoa, there were no differences between the media with post-freezing black Maca. In addition, a study was observed in male rats, with damage induced in spermatogenesis by acetate, and these were treated with Maca and showed an improvement in spermatogenesis, as well as an increase in the motile sperm count and a sperm count with normal morphology. On the contrary, they found an effect in the concentration of 100 µg/mL in TUNEL, which would show a hormetic effect of the extract on these parameters [29].
Hormesis is defined as an adaptive response to stressors that results in a biphasic dose-response relationship, such that low-dose stimulation results in a beneficial adaptation while high-dose stimulation produces a toxic effect. [47]. Furthermore, this deleterious effect could be followed by an excess concentration of antioxidants that reduce too many oxidants that, in low concentration, are necessary for sperm functions [48].
Regarding lipid peroxidation, no significant differences were found between the post-freezing treatments. Similar results were observed in horses fed with Maca in a period between 30 and 120 days; the spermatozoa were stored at 5C and later evaluated at 24, 48, and 72 hours. Even when other sperm parameters did decrease, no difference was detected. significant increase in lipid peroxidation [49].
Accordingly, the loss of motility in equine spermatozoa after freezing was observed, but without significant differences in lipid peroxidation. This is because damage to the biological membranes of spermatozoa can cause fusion and loss of saturated and unsaturated fatty acids, which could explain the lower presence of peroxidized lipids [50]. Regarding lipid peroxidation and fragmentation, its positive relationship with sperm motility and ROS emission is still not clear. However, for the success of in vitro and in vivo fertilization, good sperm motility is crucial for reproductive success.
5. Conclusion
In conclusion, the supplementation with 20mg/ml of black Maca in the media improves sperm parameters such as sperm motility, sperm viability and membrane integrity, it also decreases the emission of total sperm ROS after freezing showing the natural antioxidant property of Black Maca. Thus, Maca rises as a valid alternative as a cytoprotective supplement for the cryopreservation medium of epidydimal sperm of alpaca species. Further studies will be focused on the use of Maca-supplemented cryostored sperm on fertility capacities using either, in vitro (IVF) or in vivo (AI) models.
Author Contributions
Conceptualization, G.L. and M.V.; methodology, G.L, M.V. and M.G.; formal analysis, G.L; data curation, M.V.; writing—original draft preparation, D.V., J.Q. and LA; investigation, G.L. and M.V.; writing—review and editing, G.L, LA, and MV.; funding acquisition, M.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by PROCIENCIA - CONCYTEC-Basic Research Project, 2019-01-Agreement No. 428- 2019-FONDECYT
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article or Supplementary Material.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Quina, E. Inseminación artificial de alpacas en un contexto de crianza campesina. Descosur 2017.
- Landeo L, Zuñiga M, Gastelu T, Artica M, Ruiz J, Silva M, Ratto MH. Oocyte Quality, In Vitro Fertilization and Embryo Development of Alpaca Oocytes Collected by Ultrasound-Guided Follicular Aspiration or from Slaughterhouse Ovaries. Animals 2022, 12, 1102. [Google Scholar] [CrossRef] [PubMed]
- O'Flaherty, C. , Beorlegui, N., & Beconi, M. T. Participation of superoxide anion in the capacitation of cryopreserved bovine sperm. International journal of andrology 2003, 26, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Santiani A, Huanca W, Sapana R, Huanca T, Sepúlveda N, Sánchez R. Effects on the quality of frozen-thawed alpaca (Lama pacos) semen using two different cryoprotectants and extenders. Asian J. Androl. 2005, 7, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Casaretto, C. , Martínez Sarrasague, M., Giuliano, S., Rubin de Celis, E., Gambarotta, M., Carretero, I., & Miragaya, M. Evaluation of Lama glama semen viscosity with a cone-plate rotational viscometer. Andrologia, 2012, 44, 335–341. [Google Scholar] [CrossRef]
- O’Neill, H. C. , Nikoloska, M., Ho, H., Doshi, A., & Maalouf, W.Improved cryopreservation of spermatozoa using vitrification: comparison of cryoprotectants and a novel device for long-term storage. Journal of assisted reproduction and genetics, 2019, 36, 1713–1720. [Google Scholar] [CrossRef]
- Rosato, M. & Laffaldano, N.. Cryopreservation of rabbit semen: comparing the effects of different cryoprotectants, cryoprotectant-free vitrification, and the use of albumin plus osmoprotectants on sperm survival and fertility after standard vapor freezing and vitrification. Theriogenology, 2013, 79, 508–516. [Google Scholar] [CrossRef]
- Agarwal, A. , Makker, K., Sharma, R. Clinical relevance of oxidative stress in male factor infertility: an update. Am J Reprod Immunol 2008, 59, 2–11. [Google Scholar] [CrossRef]
- Du Plessis, S. , Makker K., Desai N.& Agarwal A. The impact of oxidative stress on in vitro fertilization. Expert Review Of Obstetrics & Gynecology. 2008, 3, 539–554. [Google Scholar] [CrossRef]
- Aitken, R. J. , Gibb, Z., Baker, M. A., Drevet, J., & Gharagozloo, P. Causes and consequences of oxidative stress in spermatozoa. Reproduction, Fertility and Development 2016, 28, 1–10. [Google Scholar] [CrossRef]
- Aitken, R.J. , 2017. Reactive oxygen species as mediators of sperm capacitation and pathological damage. Mol Reprod Dev 2017, 84, 1039–1052. [Google Scholar] [CrossRef] [PubMed]
- Treulen F, Arias ME, Aguila L, Uribe P, Felmer R. Cryopreservation induces mitochondrial permeability transition in a bovine sperm model. Cryobiology. 2018, 83, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Treulen F, Aguila L, Arias ME, Jofré I, Felmer R. Impact of post-thaw supplementation of semen extender with antioxidants on the quality and function variables of stallion spermatozoa. Anim Reprod Sci. 2019, 201, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Shojaeian, K. , Nouri, H., Kohram, H. Does MnTBAP ameliorate DNA fragmentation and in vivo fertility of frozen-thawed Arabian stallion sperm? Theriogenology 2018, 108, 16–21. [Google Scholar] [CrossRef]
- Amidi, F. , Pazhohan, A., Shabani Nashtaei, M., Khodarahmian, M., & Nekoonam, S. The role of antioxidants in sperm freezing: a review. Cell and tissue banking 2016, 17, 745–756. [Google Scholar] [CrossRef]
- Ros-Santaella JL, Pintus E. Plant Extracts as Alternative Additives for Sperm Preservation. Antioxidants (Basel). 2021, 10, 772. [Google Scholar] [CrossRef]
- Pasquariello R, Verdile N, Brevini TAL, Gandolfi F, Boiti C, Zerani M, Maranesi M. The Role of Resveratrol in Mammalian Reproduction. Molecules. 2020, 25, 4554. [Google Scholar] [CrossRef]
- Wang S, Zhu F. Chemical composition and health effects of maca (Lepidium meyenii). Food Chem. 2019, 288, 422–443. [Google Scholar] [CrossRef]
- León, J. (1964). The" Maca"(Lepidium meyenii), a little known food Plant of Peru. Economic botany 1964, 18, 122–127. [Google Scholar] [CrossRef]
- Piacente, S. , Carbone, V., Plaza, A., Zampelli, A., & Pizza, C. Investigation of the tuber constituents of maca (Lepidium meyenii Walp. ). Journal of agricultural and food chemistry 2022, 50, 5621–5625. [Google Scholar] [CrossRef]
- Gonzales GF, Cordova A, Vega K, Chung A, Villena A, Góñez C. Effect of Lepidium meyenii (Maca), a root with aphrodisiac and fertility-enhancing properties, on serum reproductive hormone levels in adult healthy men. J Endocrinol. 2003, 176, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Zheng BL, He K, Kim CH, Rogers L, Shao Y, Huang ZY, et al. . Effect of a lipidic extract from Lepidium meyenii on sexual behavior in mice and rats. Urology. 2000, 55, 598–602. [Google Scholar] [CrossRef] [PubMed]
- Zenico T, Cicero AF, Valmorri L, Mercuriali M, Bercovich E. Subjective effects of Lepidium meyenii (Maca) extract on well-being and sexual performances in patients with mild erectile dysfunction: a randomised, double-blind clinical trial. Andrologia., 2009, 41, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Gonzales C, Rubio J, Gasco M, Nieto J, Yucra S, Gonzales GF. Effect of short-term and long-term treatments with three ecotypes of Lepidium meyenii (MACA) on spermatogenesis in rats. J Ethnopharm. 2006, 103, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Sandoval M, Okuhama NN, Angeles FM, Melchor VV, Condezo LA, Lao J, et al. . Antioxidant activity of the cruciferous vegetable Maca (Lepidium meyenii). Food Chem. 2002, 79, 207–213. [Google Scholar] [CrossRef]
- Lee K-J, Dabrowski K, Sandoval M, Miller MJ. Activity-guided fractionation of phytochemicals of maca meal, their antioxidant activities and effects on growth, feed utilization, and survival in rainbow trout (Oncorhynchus mykiss) juveniles. Aquaculture. 2005, 244, 293–301. [Google Scholar] [CrossRef]
- Zha S, Zhao Q, Chen J, Wang L, Zhang G, Zhang H, et al. . Extraction, purification and antioxidant activities of the polysaccharides from maca (Lepidium meyenii). Carbohydrate polymers. 2014, 111, 584–587. [Google Scholar] [CrossRef] [PubMed]
- Valdivia M, Yarasca K, Lévano G, Vásquez J, Temoche H, Torres L, et al. Effect of Lepidium meyenii (maca) on testicular function of mice with chemically and physically induced subfertility. Andrologia. 2016, 48, 927–934. [Google Scholar] [CrossRef]
- Rubio, J. , Riqueros, M.I., Gasco, M., Yucra, S., Miranda, S. and Gonzales, G.F. Lepidium meyenii (Maca) reversed the lead acetate induced-damage on reproductive function in male rats. Food Chem. Toxicol. 2006, 44, 1114–1122. [Google Scholar] [CrossRef]
- Gonzales-Castañeda C, Rivera V, Chirinos AL, Evelson P, Gonzales GF. Photoprotection against the UVB-induced oxidative stress and epidermal damage in mice using leaves of three different varieties of Lepidium meyenii (maca). Int J Dermatol. 2011, 50, 928–938. [Google Scholar] [CrossRef]
- Dzięcioł, M. , Wróblewska, A., & Janda-Milczarek, K. Comparative Studies of DPPH Radical Scavenging Activity and Content of Bioactive Compounds in Maca (Lepidium meyenii) Root Extracts Obtained by Various Techniques. Applied Sciences. 2023, 13, 4827. [Google Scholar] [CrossRef]
- N. Inoue, C. Farfan and G. F. Gonzales, Effect of butanolic fraction of yellow and black maca (Lepidium meyenii) on the sperm count of adult mice, Andrology, 2016, 48, 915–921. [CrossRef] [PubMed]
- World Health Organization "WHO Laboratory Manual for the examination and processing of human semen”, 2010 ,5ta edición.
- Hrudka, F. Cytochemistry of oxidoreductases in spermatozoa: The technique revisited. Andrologia, 1979, 11, 337–353. [Google Scholar] [CrossRef] [PubMed]
- Santiani, A. , Evangelista, S., Cheuquemán, C., von Baer, A., Risopatrón, J., & Sánchez, R.. Evaluación de la integridad de ADN mediante citometría de flujo en espermatozoides de alpaca criopreservados con análogos de superóxido dismutasa. Revista de Investigaciones Veterinarias del Perú, 2012, 23, 182–191. [Google Scholar]
- Nova, Z. , Skovierova, H., Strnadel, J., Halasova, E., & Calkovska, A. Short-term versus long-term culture of A549 cells for evaluating the effects of lipopolysaccharide on oxidative stress, surfactant proteins and cathelicidin LL-37. International Journal of Molecular Sciences, 2020, 21, 1148. [Google Scholar] [CrossRef]
- Canorio Pariona, N. , Paredes Arnedo, F., & Valdivia Cuya, M. Agentes crioprotectores alternativos para el congelamiento lento de espermatozoides epididimarios de alpaca (Vicugna pacos). Revista de Investigaciones Veterinarias del Perú 2015, 26, 434–443. [Google Scholar] [CrossRef]
- Buyanbadrakh, E. , Hong, H.S., Lee, K.W., Huang, W.Y. and Oh, J.H. Anti-oxidant activity, macamide B content and muscle cell protection of Maca (Lepidium meyenii) extracted using ultrasonication-assisted extraction. Microbiol. Biotechnol. Lett. 2020, 48, 129–13. [Google Scholar] [CrossRef]
- Ezzati, M. , Shanehbandi, D., Hamdi, K., Rahbar, S., & Pashaiasl, M. Influence of cryopreservation on structure and function of mammalian spermatozoa: An overview. Cell and Tissue Banking. 2020, 21, 1–15. [Google Scholar] [CrossRef]
- Grötter, L. G. , Cattaneo, L., Marini, P. E., Kjelland, M. E., & Ferré, L. B.. Recent advances in bovine sperm cryopreservation techniques with a focus on sperm post-thaw quality optimization. Reproduction in Domestic Animals. 2019, 54, 655–665. [Google Scholar] [CrossRef]
- Ciani F, Cocchia N, Del Prete C, Palumbo V, Carotenuto D, Pasolini MP, Tafuri S. Sperm cromatin integrity in stallions with Lepidium meyenii (Maca) dietary supplementation. Reprod Domest Anim. 2017, 52, 7. [Google Scholar]
- Tafuri, S.; Cocchia, N.; Carotenuto, D.; Vassetti, A.; Staropoli, A.; Mastellone, V.; Peretti, V.; Ciotola, F.; Albarella, S.; Del Prete, C.; et al. Chemical analysis of Lepidium meyenii (Maca) and its effects on redox status and on reproductive biology in stallions. Molecules 2019, 24, 1981. [Google Scholar] [CrossRef] [PubMed]
- Mamani-Mango, G. , Moina Gonzales, M., Ramos Hidalgo, M., Mendoza Mallma, J., Ruiz Bejar, J., Rivas Palma, V., & Mellisho Salas, E. Effect of extender and freezing rate on quality parameters and in vitro fertilization capacity of alpaca spermatozoa recovered from cauda epididymis. Biopreservation and biobanking, 2019, 17, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Juárez, J. , & Santiani, A. Determinación del porcentaje de viabilidad espermática mediante citometría de flujo durante el proceso de criopreservación en espermatozoides obtenidos de epidídimo de alpaca. Revista de Investigaciones Veterinarias del Perú, 2019, 30, 1175–1183. [Google Scholar] [CrossRef]
- Naresh, S. , & Atreja, S. K.. The protein tyrosine phosphorylation during in vitro capacitation and cryopreservation of mammalian spermatozoa. Cryobiology, 2015, 70, 211–216. [Google Scholar] [CrossRef]
- Leiva-Revilla, J. , Rolón, M., Siyadatpanah, A., de Lourdes Pereira, M., & Nissapatorn, V. First study of in vitro protective effect of Lepidium meyenii (Maca) on frozen–thawed bovine spermatozoa. Veterinary World 2022, 15, 1481. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J. , Mattson, M.P. and Calabrese, V. Resveratrol commonly displays hormesis: Occurrence and biomedical significance. Hum. Exp. Toxicol., 29(12): 980–1015 fragmentation. Journal of Assisted Reproduction and Genetics. 2010, 24, 561–569. [Google Scholar] [CrossRef]
- Rashki Ghaleno, L. , Alizadeh, A., Drevet, J. R., Shahverdi, A., & Valojerdi, M. R. Oxidation of sperm DNA and male infertility. Antioxidants, 2021, 10, 97. [Google Scholar] [CrossRef]
- Del Prete, C. , Tafuri, S., Ciani, F., Pasolini, M. P., Ciotola, F., Albarella, S.,... & Cocchia, N. Influences of dietary supplementation with Lepidium meyenii (Maca) on stallion sperm production and on preservation of sperm quality during storage at 5 C. Andrology. 2018, 6, 351–361. [Google Scholar] [CrossRef]
- Restrepo, G. , Varela, E., Duque, J. E., Gómez, J. E., & Rojas, M. Freezing, vitrification, and freeze-drying of equine spermatozoa: Impact on mitochondrial membrane potential, lipid peroxidation, and DNA integrity. Journal of equine veterinary science, 2019, 72, 8–15. [Google Scholar] [CrossRef]
Figure 1.
Effect of Lepidium meyenii on sperm oxidative stress (ROS) activity. Percentage of sperm with a high amount of ROS. Fresh, Yolk medium (YM), YM supplemented with Maca 10mg/ml, YM supplemented with Maca 20mg/ml, YM supplemented with Maca 30mg/ml and YM supplemented with resveratrol . Significant difference; *: p <= 0.05, **: p <= 0.01, ***: p <= 0.00 between Control and each treatment group.
Figure 1.
Effect of Lepidium meyenii on sperm oxidative stress (ROS) activity. Percentage of sperm with a high amount of ROS. Fresh, Yolk medium (YM), YM supplemented with Maca 10mg/ml, YM supplemented with Maca 20mg/ml, YM supplemented with Maca 30mg/ml and YM supplemented with resveratrol . Significant difference; *: p <= 0.05, **: p <= 0.01, ***: p <= 0.00 between Control and each treatment group.
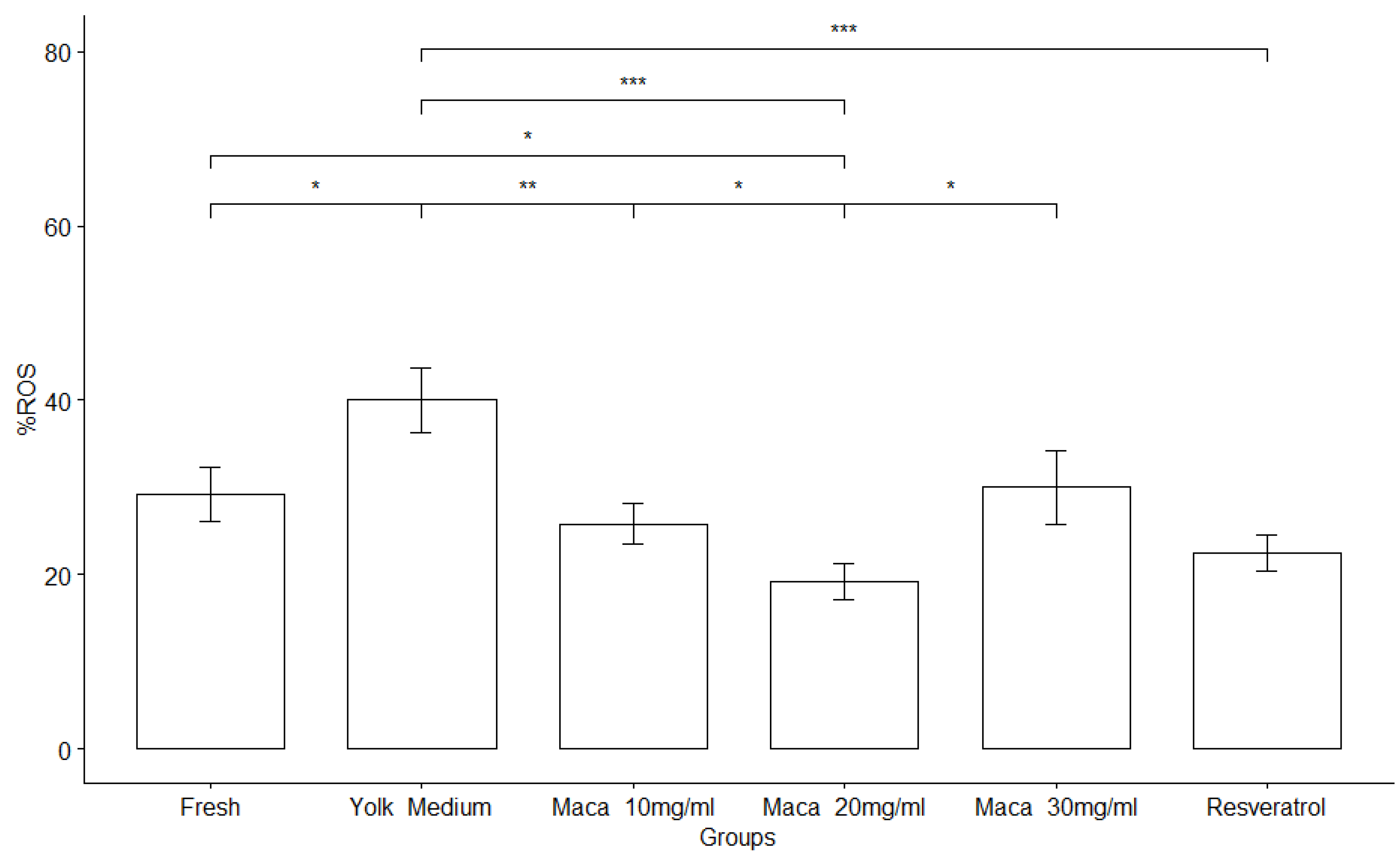
Figure 2.
Relative DNA fragmentation by treatment Percentage of index DNA fragmentation per treatment. Fresh, Yolk medium, Maca 10mg/ml, Maca 20mg/ml, Maca 30mg/ml and resveratrol. Significant difference; *: p <= 0.05, **: p <= 0.01, ***: p <= 0.00 between Control and each treatment group.
Figure 2.
Relative DNA fragmentation by treatment Percentage of index DNA fragmentation per treatment. Fresh, Yolk medium, Maca 10mg/ml, Maca 20mg/ml, Maca 30mg/ml and resveratrol. Significant difference; *: p <= 0.05, **: p <= 0.01, ***: p <= 0.00 between Control and each treatment group.
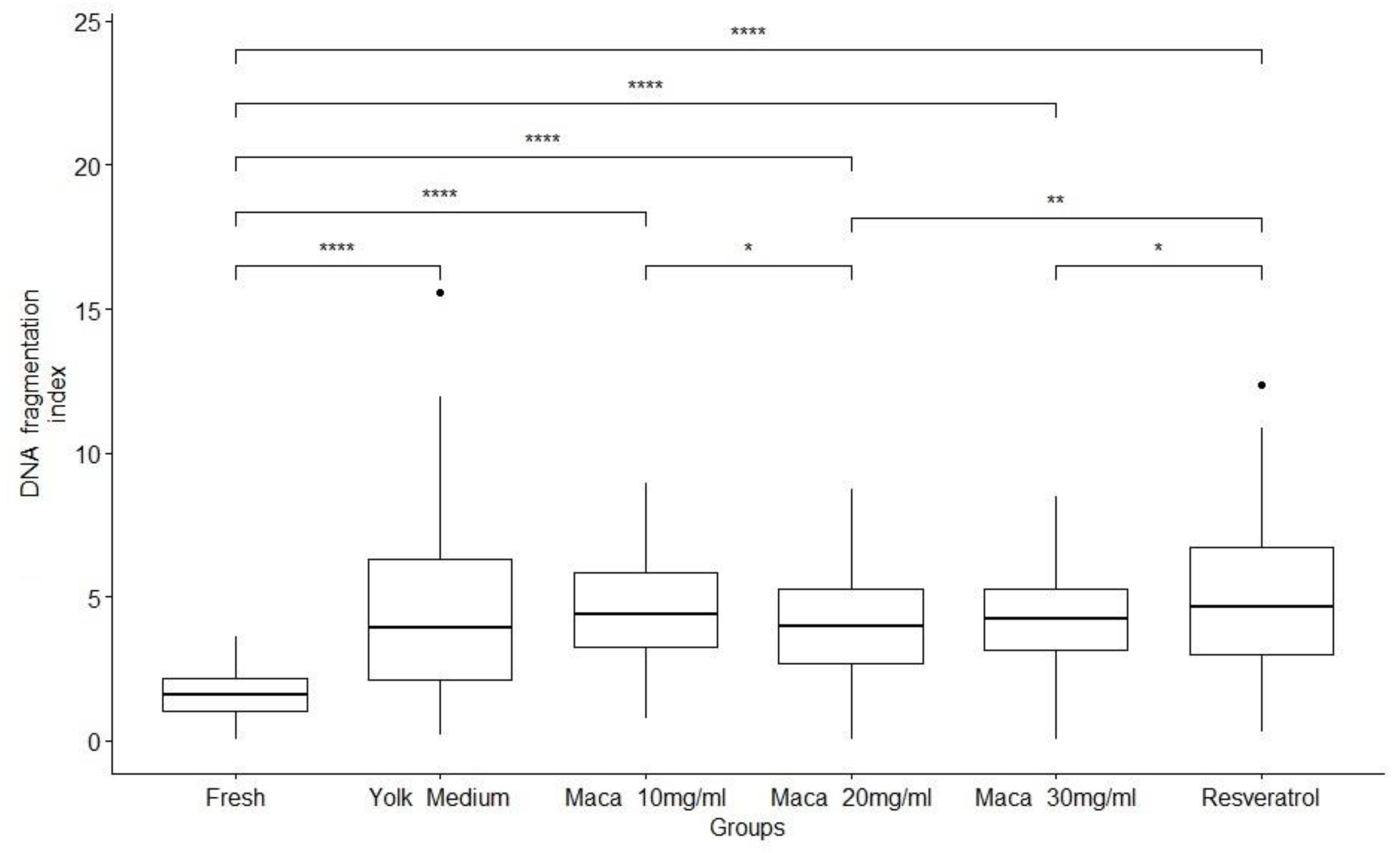
Figure 3.
DNA fragmentation of frozen/thawed epididymal alpaca sperm. (a) Bright field. (b) Fragmented TUNEL-positive spermatozoa (intense green), (c) Sperm positive for propidium iodide (red). Total magnification: 400x.
Figure 3.
DNA fragmentation of frozen/thawed epididymal alpaca sperm. (a) Bright field. (b) Fragmented TUNEL-positive spermatozoa (intense green), (c) Sperm positive for propidium iodide (red). Total magnification: 400x.
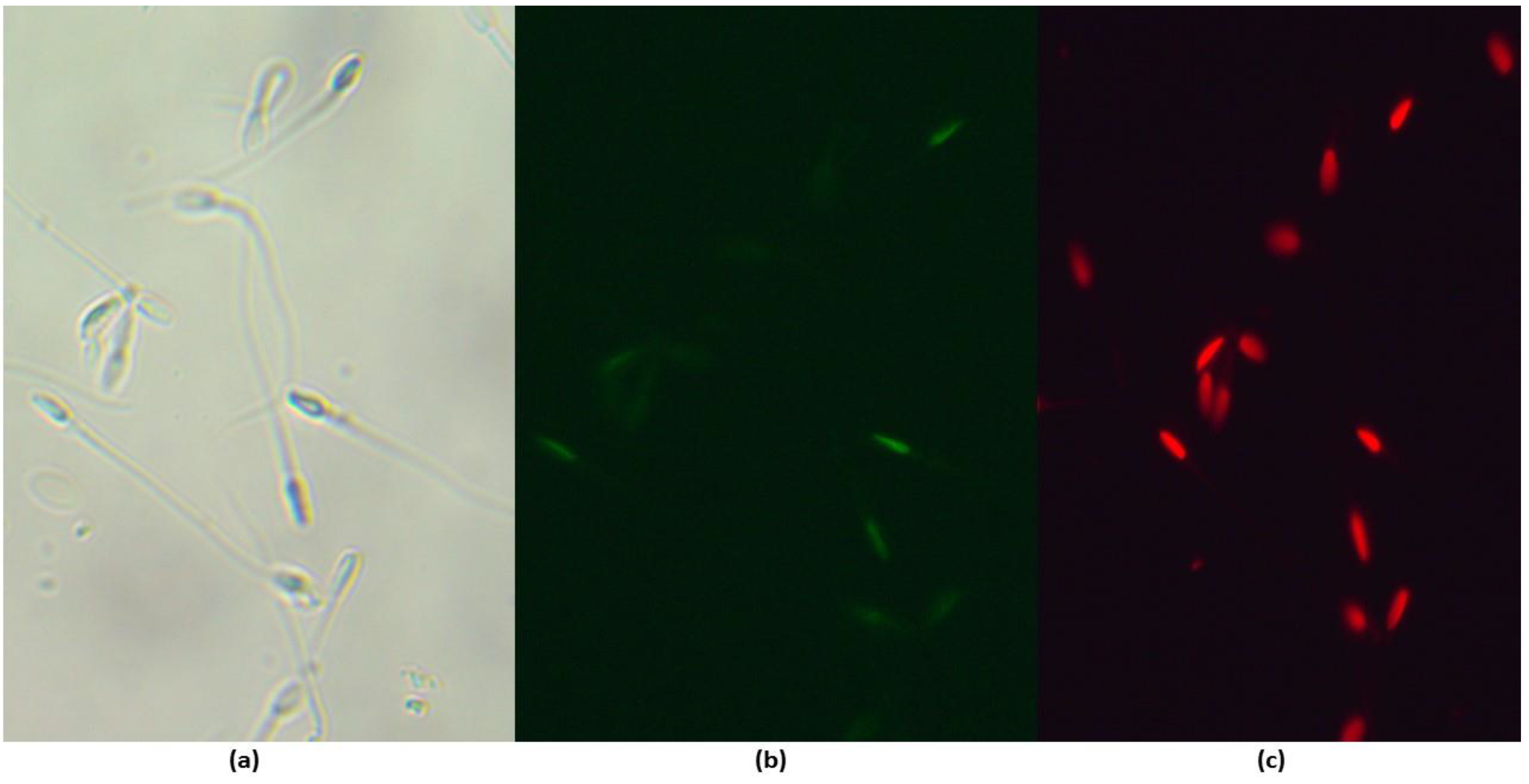
Figure 4.
Cytochemical reaction of endogenous reductases in the middle sperm piece. Four formazan deposits were identified. A) C (Compact), B) Sc (Sub-compact), C) F (Focal), and D) D (Diffuse) in alpaca sperm mitochondria visualized with NBT technique. Photographs taken by 800X optical microscopy.
Figure 4.
Cytochemical reaction of endogenous reductases in the middle sperm piece. Four formazan deposits were identified. A) C (Compact), B) Sc (Sub-compact), C) F (Focal), and D) D (Diffuse) in alpaca sperm mitochondria visualized with NBT technique. Photographs taken by 800X optical microscopy.


Table 1.
Sperm parameters before (Fresh) and after freezing/thawing (*).
| Groups | Motility % | Viability % | HOST% | DNA fragmentation index |
ROS % | Mitochondrial activity index |
|---|---|---|---|---|---|---|
| Fresh | 75 ± 2.67a | 61.25 ± 8.29a | 69.16 ± 8.21a | 1.64 ± 0.83a | 29.2 ± 10.7a |
0.892±0.255a |
| *YM | 25 ± 1.49b | 46.25 ± 5.69b | 51.25 ± 7.72b | 4.47 ± 3.10a | 40 ± 12.6b | 0,910 ± 0,277a |
| *Maca 10mg/ml | 32.5±1.71c | 47.08 ± 6.89b | 50.08 ± 6.20b | 4.55 ± 1.91a | 25.8 ±7.93c | 0,896 ± 0,255a |
| *Maca 20mg/ml | 40 ± 2.45d | 54.58 ± 5.41c | 58.33 ± 7.78c | 3.95 ± 1.88a | 19.2 ± 7.33c | 0,882 ± 0,171ª |
| *Maca 30mg/ml | 22.5 ±1.83b | 48.33 ±6.85b | 52.91 ± 3.34b | 4.16 ± 1.77a | 30 ± 14.8c | 0,765± 0,188a |
| *Resveratrol | 27 ± 1.20b | 55.83 ±8.75c | 57.08 ± 2.57c | 4.90 ± 2.65a | 22.5 ± 7.23c | 0.872 ± 0.098a |
Significant difference P<0.05; between the control and each treatment group. YM: Tris-Yolk medium. Trist-Yolk medium without and supplement with different concentration of Maca (*). a,b,c,d Different letters between rows indicate significant between groups.
Table 2.
Rates of Cytochemical Activity of Oxidoreductases on the fresh and freezing/thawing (*).
| Rate of Activity (%) | ||||||
|---|---|---|---|---|---|---|
| Standar | Sub-standar | Low | Residual | Intensive Reaction (St + Sub-st) |
Reduced Reaction (Low + Res) |
|
| Fresh | 38 ± 19.4 | 13.5± 9.42 | 0.900 ±0.65 | 0 | 51.5a | 0.9a |
| *TYM | 44.2± 32.8 | 26.0 ± 19.2 | 0.500 ±0.79 | 0.150± 0.28 | 70.2b | 0.65b |
| *Maca 10mg/ml | 47.9 ± 32.1 | 13.7 ± 9.50 | 0.550 ±1.50 | 0.008 ±0.03 | 61.6b | 0.55c |
| *Maca 20mg/ml | 21.3 ± 5.61 | 16.9 ± 22.4 | 0.375 ±0.45 | 0.033± 0.12 | 38.2c | 0.408c |
| *Maca 30mg/ml | 40.3 ± 21.2 | 11.0 ± 8.44 | 0.525 ±0.73 | 0.083±0.15 | 51.3a | 0.608d |
| *Resveratrol | 23.5 ± 12.4 | 11.5 ± 9.66 | 2.22 ±0.46 | 0.017±0.06 | 35c | 2.217e |
a,b,c,,d,e Different letters between rows indicate significant between groups.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



