
Submitted:
12 May 2023
Posted:
22 May 2023
You are already at the latest version
Abstract
Starch content in wheat is an important characteristic for various purposes. Starch is composed of two components known as amylose and amylopectin, and the proportion between these two components plays a crucial role in determining the properties and structure of starch. One of the key genes involved in starch biosynthesis in wheat is granule bound starch synthase I (GBSSI), also known as the waxy protein, which synthesizes amylose in the endosperm. A mutant cultivar, named Gunji-3, was created by crossing the waxy type wheat (Shinmichal 1) with the wild type bread wheat (Keumkang). When comparing the Wx-B1 allele of the waxy protein gene in Gunji-3 with that of the waxy type wheat, there were 12 SNPs and one deletion difference. Additionally, when comparing amino acid sequences with Wx-B1 alleles, differences at two positions are found, indicating that the mutant carries new Wx-B1 allele which is named as Wx-B1o. In terms of the physicochemical properties of Gunji-3 starch, it had a lower amylose content than the parental wheat varieties and showed higher protein content and greater swelling power. These newly introduced waxy type wheat could provide important basis not only for understanding the starch characteristics of wheat but also for various breeding programs.
Keywords:
Starch
; Amylose
; Amylopectin
; GBSSI
; mutation
1. Introduction
Despite the importance of seed storage proteins in wheat for food quality, wheat carbohydrate is still the key nutritional factor and feeding a plethora of population in the world with a total of 215.9 million hectares harvested [1]. However, its complex chromosomal and genetic architecture mainly due to polyploidy hampers wheat researchers from investigating the molecular basis of nutrient biosynthesis. Modern agricultural environment, two main wheat species are cultivated. One is durum or pasta wheat (Triticum turgidum ssp. durum; 2n=4x=28, AABB) and common or bread wheat (T. aestivum ssp. aestivum; 2n=6x=42, AABBDD).
Wheat starch is mainly composed of amylopectin and amylose. Amylopectin is a major component of starch granule, which has α(1-4)-linked glucose linear chains and α(1-6)-linked branch points. The clustered branches of amylopectin chains tend to form packed structures, resulting in the crystalline domains of the starch granules. On the other hand, amylose is composed of α(1-4)-linked glucose linear chains but free amylose is often complexed with lipids, acting as resistant starch (RS) which has a number of dietary advantages.
Four main genes are involved in the biosynthesis of starch in wheat. The biosynthesis of amylopectin involves three enzymes, namely starch synthases (SSs), starch branching enzymes (SBEs), and debranching enzymes (DBEs). In contrast, the production of amylose in the endosperm is solely attributed to the granule bound starch synthase I (GBSSI) enzyme [2]. GBSSI is also recognized as the waxy (Wx) protein and possesses the genes Wx-A1, Wx-B1, and Wx-D1. These three genes are located on wheat chromosomes 7AS, 4AL (translocated from the original 7BS), and 7DS, respectively. Three copies of GBSSI in hexaploid wheats cause partial waxy or complete waxy phenotypes. Seib [3] reported that the possible combinations of null alleles in the subgenomes of common wheat (Wx-A1, Wx-B1, and Wx-D1) can produce eighty types of isogenic lines differing in starch properties. Single, double, or triple mutations at the Wx loci of common wheat can result in distinct properties due to small variations in molecular weight (Wx-A1: 62.8 kDa, Wx-B1: 56.7 kDa, and Wx-D1: 58.7 kDa), as determined by protein separation using sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) [4]. The GBSSI gene sequence in wheat was first reported by Clark and Robertson [5] who isolated a waxy cDNA sequence of 2,186 bp, with an open reading frame (ORF) which is 1,845 bp in length without its subgenomic information. Later Briney and Wilson [6] identified that the cloned sequence was Wx-A1 using PCR markers in Australian wheat. In the late 20th century, three subgenomic copies of waxy genes in Chinese Spring (a reference cultivar) were characterized [7], revealing their slight variations in length at genomic DNA level. In their report, it was stated that the sizes of the open reading frames (ORFs) for Wx-A1, Wx-B1, and Wx-D1 were 2,781, 2,794, and 2,862 bp, respectively. Additionally, the sequences showed a similarity of over 95% in the multiple alignment. All three subgenomic copies have similar genic structures with 12 exons and 11 introns, also showing high similarity to the Wx genic structure in barley which are included in the same subfamily, Pooideae [8].
The detailed information toward the gene structures and sequences of waxy genes accelerated the characterization of a variety of mutant alleles at a molecular level. The first null alleles were reported from two waxy common wheat cultivars [9]. One cultivar (cv. Kanto 107) was a double mutant which does not have Wx-A1 and Wx-B1 whereas the other (cv. Bai Huo) was a single mutant (Wx-D1 null). Since then a number of mutant alleles were identified and characterized in common wheat and tetraploid wheat species. According to a search of published literature, the Wx-A1 gene appears to have the highest degree of variation when compared to the other Wx genes. A total of 18, 14, and six mutant alleles have been discovered for Wx-A1, Wx-B1, and Wx-D1, respectively. These mutant alleles exhibit a variety of changes in their nucleotide sequences, which can affect the activity of the corresponding enzyme. The allelic variations are previously reviewed by Guzman and Alvarez [10]. Here, we will focus on the variations in Wx-B1 gene which highly affect the content of amylose in endosperm [11,12]. A total of six allelic variations have been reported for the Wx-B1 gene in hexaploid wheat to date, including the wild type. Those five mutants show clear difference in nucleotide sequences compared to the wild type (Wx-B1a). Vrinten and Nakamura [9] identified the first mutant allele in hexaploid wheat, of which the entire coding sequences are deleted (Wx-B1b). Over 10 years later, Yamamori and Guzmán [13] found the second mutant which has at least four single nucleotide polymorphisms (SNPs) in three exons (Wx-B1c). Two additional mutations were reported in indian dwarf wheat (Wx-B1k) and club wheat (T. compactum, Wx-B1m) that have insertion and deletion in their coding sequences, respectively [14]. Wx-B1l is the latest mutant allele identified in bread wheat, characterized by a single nucleotide deletion in the second exon, resulting in an alteration of the open reading frame (ORF) [15].
In the current study, we report a novel mutant allele for Wx-B1 gene (putatively named as Wx-B1o) in bread wheat. The mutant cultivar ‘Gunji-3’ was selected in an F8 population generated by crossing Korean bread wheat ‘Keumkang and Korean waxy wheat ‘Shinmichal 1’. The DNA banding pattern of ‘Gunji-3’ was overlapped with that of ‘Shinmichal 1’; however, the mutant cultivar shows much lower amylose content than ‘Shinmichal 1’ and the physicochemical properties of wheat flour was a bit different. We cloned and sequenced three waxy genes in ‘Gunji-3’ and characterized mutant alleles at the nucleotide level.
2. Materials and Methods
2.1. Materials
In 2010, a cross was made between Keumkang, a popular Korean wheat cultivar with normal amylose content, and Shinmichal 1, a waxy wheat cultivar created from Alchan//Kanto107/BaiHuo, resulting in a total of 127 F1 plants. These F1 plants were then bred using the single seed descent method, and the resulting generations were rapidly advanced from F2 to F6 within three years. In the F5 generation, 12 waxy wheat lines were chosen by screening allelic variations at the Wx loci. Amylose content was measured in the F11 generation, leading to the identification of Gunji-3, which had lower amylose content compared to Shinmichal 1. In 2016, a replicated experiment was conducted at the experimental farm of Chonbuk National University (Jeonju, Korea) using Shinmichal 1, Keumkang, and Gunji-3, each planted in triplicate. The experimental plots were arranged in three rows, each 4 m long and spaced 25 cm apart. Prior to sowing, fertilizer was applied in the ratio of 5:7:5 kg/10a for nitrogen, phosphorus, and potassium, respectively, and the plot was harvested in 2017. Additionally, rigorous measures were implemented to control weeds, insects, and diseases throughout the experiment. The evaluation of flour and starch characteristics was conducted by obtaining a sufficient quantity of grain from three plots and drying the harvested grain under conditions of 22°C temperature and 14% humidity before the evaluation.
2.2. Identification of a novel Wx-B1 allele
To identify allelic variations in the Wx-1 loci, one plant per line/cultivar grown in a temperature-controlled greenhouse was sampled. After two weeks of growth post-germination, leaves were collected and immediately stored in liquid nitrogen until use, then stored at -80°C. Genomic DNA (gDNA) was extracted from 100mg of young leaf tissue using a genomic DNA prep kit (Solgent Co., Korea) according to the manual, and the DNA was quantified using a Biodrop (Biodrop Ltd, UK). Primers for the Wx-1 gene loci were designed based on the mutant or non-mutant waxy allele described by Nakamura et al [16]. In addition, for the confirmation of Wx-1 isoforms in the samples, the isolation and purification of starch granules from the starch granule, as well as the preparation of waxy proteins, were carried out based on the methods described by Seo et al [17]. Electrophoresis of agarose gels for starch was performed using the method described by Jegasothy et al [18].
2.3. Cloning of Wx-B1 and genome walking
Amplification of the Wx-B1 coding region was performed on previously extracted gDNA using the method of Guzman and Alvarez [19]. For the design of primers, the International Wheat Genome Sequencing Consortium Chinese Spring (IWGSC CS) RefSeq v2.1 and Triticum aestivum gene for starch synthase (GBSSI), complete cds (AB019623.1) sequences registered at NCBI were used, and cloning were performed in six parts (Table 1). A 20 μl PCR reaction mixture was prepared containing 4 μl of Taq 5X Master Mix (New England Biolabs) which included 1.5 mM MgCl2, 0.2 mM dNTP each, and 25 units/ml Taq polymerase. Also 50 ng of gDNA and 100 pmol each of the forward and reverse primers were used, and the remaining volume was nuclease free water. PCR cycling parameters and primer sequences are shown in Table 1. Aliquots of the PCR products were separated and visualized by electrophoresis on 1% agarose gels.
Compared to DNA 100 bp plus ladder (SmartGene, Korea) by electrophoresis, PCR products with significant length were purified and sequenced by Macrogen Sequencing service (Macrogen, Korea). After that, overlapping sequences were removed and aligned with Wx-B1a by Geneious software [20]. Furthermore, the sequence was analyzed by BLASTN [21] on the IWGSC CS RefSeq v2.1 assembly.
2.4. Characterization of a novel waxy mutant
The wheat grains were processed using an experimental mill from Bühler, in accordance with the AACC International Approved Method 26-31.01 [22]. Flour yield was determined by calculating the ratio of break and reduction flours to the total weight of the grains fed into the mill. To analyze the particle size distribution of the flour, the LS13320 multi-wavelength laser particle size analyzer (Beckman Coulter, Inc., Brea, CA, USA) was used, following the guidelines of Approved Method 55-40.01. Flour color was measured using an 11-mm measurement aperture colorimeter (CM-2002, Minolta Camera, Osaka, Japan), and the whiteness index was calculated in accordance with the method described by Nguimbou et al (2013) [23]. Moisture, ash, and protein content of the samples were determined in accordance with the protocols described in AACC International Approved Methods 44-15.02, 08-01.01, and 46-30.01, respectively [22]. The amylose content, damaged starch content, and total starch content were measured using enzymatic assay kits (MegaZyme Pty., Ltd., NSW, Australia), following the methods outlined by Gibson et al. [24,25] and McCleary et al. [26], respectively. The content of arabinoxylan was determined following the method described by Douglas [27]. The SDS-sedimentation test was conducted in accordance with the guidelines of the approved method 56-60.01 [22]. The mixing time, mixing tolerance, and optimum water absorption of wheat flour were determined using a 10 g mixograph (National Mfg. Co., Lincoln, NE, USA) following the approved method 54-40.02 [22].
To fractionate starch, 100 g (db) of flour was processed using the method outlined by Czuchajowska and Pomeranz [28]. The particle size of starch was measured using a laser diffraction particle size analyzer (Model LS13320, Beckman Coulter, Inc., Brea, CA, USA) with starch dispersed in 99.5% 2-propanol, following the manufacturer's instructions. The particle size distribution was determined by measuring the cumulative volume of starch granules in 0.1 µm intervals ranging from 0 to 40 µm. The microstructure of the starch was observed using a scanning electron microscope (SEM, SN-3000, Hitachi, Tokyo, Japan) following the procedure described by Kasemsuwan et al [29]. The cold water retention capacity of starch was determined using the method described by Sollars [30]. The swelling volume and power were measured using the methods of Yamamori et al [31] and Sasaki and Matsuki [32], respectively.
The pasting properties of starch were analyzed using a Micro Visco-Amylo-Graph (manufactured by Brabender OHG, Straben, Germany) following standard procedures. For this analysis, 10.0g (db) of starch was suspended in a 0.1% AgNO3 solution (100ml). The suspension was then heated from 30 to 95°C at a rate of 7.5°C/min, followed by an additional five minutes of heating at 95°C. Subsequently, it was cooled at a rate of 5.0°C/min to 50°C. The sample was maintained at 50°C while being stirred at 110 rpm for two minutes. The viscosity of the starch sample was measured in Brabender units, and the values for holding strength, peak viscosity, and final viscosity were determined. The breakdown value was calculated by subtracting the holding strength from the widely used peak viscosity and final viscosity values. After determining the pasting properties of the starch using the Micro Visco-Amylo-Graph, the starch gel was prepared and stored in a container with a diameter of 60mm and a height of 20 mm at 4°C for 24 hours. The gel consistency of the starch gel was measured using a TA-XT2 Texture Analyzer (manufactured by Stable Micro Systems, Cambridge, UK) through compression testing. This involved placing the compressed starch gel on a flat metal plate and measuring it with a flat plastic plunger with a diameter of 20mm while applying a 25% compression at a speed of 1.0mm/s.
500 mg (db) of starch was equilibrated in a 90% humidity chamber for 16 hours. Diffractograms were obtained by scanning from 5° 2θ to 50° 2θ at a scanning speed of 8°/min and a scanning step of 0.03°. X-ray diffractometry (XRD) was utilized to determine the relative degree of crystallinity of the starch. The analysis was conducted using X’pert powder (PANalytical Inc., Westborough, MA, USA). The degree of crystallinity (%) of the starch was quantitatively estimated from the crystalline and amorphous regions on the diffractograms, following the method described by Hayakawa et al [33].
The thermal properties of the starch samples were analyzed using a differential scanning calorimeter (DSC) from Pyris1 Perkin-Elmer Co., located in Waltham, MA, USA. Temperature and enthalpy calibration were performed using an indium standard. To prepare the samples, accurately weighed 10 mg (db) of starch was mixed with 20 µl of distilled water in a stainless-steel capsule. The capsule was sealed and allowed to equilibrate at 24°C for 24 hours before conducting the DSC analysis. The sample temperature was gradually increased from 20 to 180°C at a rate of 10°C per minute, with an inert material (aluminum oxide) and water (in a 1:2 ratio) used as a reference in a separate capsule. The onset temperature (To) and peak temperature (Tp) of each endotherm were determined using Pyris Manager data processing software. The transition enthalpy (ΔH) was calculated by determining the peak area and expressed in Joules per gram (J/g) of dry matter.
For the preparation of white salted noodles, we utilized the optimal water absorption for noodle dough as established in a previous study [34]. A commercially available wheat flour suitable for noodle production, which required 34% water absorption to achieve consistent, non-sticky, and smooth dough, was used as a reference to compare with other types of flour in determining the optimal water absorption for noodle production. A pin mixer (National Mfg. Co., USA) was used to mix 100 g of flour with a specific amount of sodium chloride solution for 4 minutes at a head speed of 86 rpm. The flour had a moisture content of 14% (wet basis). To prepare noodles with varying levels of water absorption, we adjusted the concentration of the sodium chloride solution to achieve a consistent 2.0% sodium chloride content in the noodle dough. Crumbly dough was processed using the rollers of a noodle machine (Ohtake Noodle Machine Mfg. Co., Japan) set at a 3-mm gap and operating at 65 rpm to create a dough sheet. The dough sheet was folded and passed through the sheeting rollers multiple times. Specifically, the folding and sheeting process was repeated twice, followed by a 1-hour resting period for the dough sheet. Subsequently, the dough sheet was passed through the sheeting rollers 3 times, with the gap progressively reduced to 2.40 mm, 1.85 mm, and 1.30 mm, respectively. Following the final sheeting step, the thickness of the dough sheet was promptly measured using a Peacock Dial Thickness Gauge G (Ozaki Mfg. Co., Japan), a micrometer-based device that enables accurate thickness measurements. To evaluate the color of the dough sheet, it was cut into portions of 5 × 10 cm. Color measurements were performed using a colorimeter (CM-2002, Minolta Camera, Osaka, Japan) equipped with an 11-mm measurement aperture. The whiteness index of noodle dough sheet was calculated according to Nguimbou et al [23]. The dough sheet was fed through cutting rolls with a number 12 blade, resulting in the production of noodle strands with a cross-sectional dimension of 3 × 2 mm and a length of approximately 30 cm.
Twenty grams of fresh noodles were cooked in 500 mL of boiling distilled water for 18 minutes and then rinsed with cold water. Two replicates of the cooked noodles were analyzed using Texture Profile Analysis (TPA) within 5 minutes of cooking. TPA was performed using a TA-XT2 Texture Analyzer (Stable Micro Systems, UK). Five strands of cooked noodles were arranged in parallel on a flat metal plate and subjected to two crosswise compressions, each to 70% of their original height. A 3.175-mm metal blade was used for the compression test, which was conducted at a crosshead speed of 1.0 mm/sec. Springiness, hardness, and cohesiveness were determined from the force-time curves obtained through TPA, following the method described by Park et al [35].
2.5. Statistical analysis
Statistical analyses including Fisher's least significant difference test (LSD), analysis of variance (ANOVA), and pairwise t-tests were conducted using SAS software (SAS Institute, Cary, NC, USA). Flour properties were evaluated through three replications, while starch characteristics were assessed at least 10 times to ensure statistical validity.
3. Results
3.1. Analysis of the waxy protein polymorphism and PCR
Figure 1 shows the PCR amplification of the waxy allele and the SDS-PAGE results of the waxy protein for Gunji-3 and its parental varieties. As mentioned above, the PCR amplification results showed that the mutant cultivar (Gunji-3) exhibited the same DNA band pattern as Shinmichal 1 (Shinmichal 1; Wx-A1b, Wx-B1b, Wx-D1b) (Figure 1a). Keumkang is a wild-type bread wheat that expresses all three types of Wx protein, whereas Shinmichal 1, a waxy type, was null for all three types of Wx protein. Additionally, the mutant cultivar Gunji-3 showed the same results as Shinmichal 1 (Figure 1b). However, Gunji-3 showed a much lower amylose content compared to Shinmichal 1 (Table 2). Thus, to analyze the Wx-B1 gene which is the gene most related to the amylose content in the endosperm [11,12], cloning and sequencing of the waxy genes were performed.
3.2. Sequence analysis of the waxy genes
To understand the low amylose content of the mutant cultivar (Gunji-3; DH185), we tried to confirm the sequence of the waxy genes. Based on the sequence of the waxy gene registered in the NCBI, the sequence of the waxy gene of DH185 was characterized. As a result, the sequences of Wx-A1 and Wx-D1 in DH185 matched Shinmichal 1 (Wx-A1b, Wx-D1b respectively) but not Wx-B1 (Figure 2). Therefore, using Wx-B1a and IWGSC RefSeq v2.1 as a reference, the sequence of Wx-B1 of DH185 was confirmed through cloning and sequencing.
To identify the sequence of the Wx-B1 gene in the mutant cultivar DH185, the Wx-B1a allele sequence of Chinese Spring was used as a reference for comparison. The cloning was conducted in six parts, and the primers and sizes for each part are listed in Table 1. As a result, a 2,805 bp genetic sequence for the Wx-B1 gene was identified in the mutant cultivar. Alignment of this sequence with the Wx-B1a allele revealed 12 SNPs and one deletion (Figure 3). Furthermore, a comparison of the amino acid sequences between Wx-B1a and the mutant cultivar showed a distinct difference from the 740th amino acid (Figure 4B). Additional comparison with other Wx-B1 alleles using BLASTN showed significant homology with Triticum aestivum (Table 3).
3.3. Analysis of the flour characteristic and starch structure of the waxy-type mutant (Gunji-3) and parental wheat
3.3.1. Flour characteristics
The flour characteristics of Gunji-3, Shinmichal 1, and Keumkang are summarized in Table 4. The flour yield value for Gunji-3 was between Shinmichal 1 and Keumkang (71.23 ± 0.74%, 63.60 ± 0.08%, and 73.43 ± 0.22%, respectively), while Gunji-3 showed a significantly similar whiteness index (89.86 ± 0.06%) to Shinmichal 1 and Keumkang (90.02 ± 0.07% and 88.44 ± 0.06%, respectively). However, the ash amount of the three wheat cultivars was not significantly different (0.43 ± 0.01, 0.44 ± 0.01, and 0.45 ± 0.01, respectively). The average particle size of Gunji-3 was not significantly different from those of Shinmichal 1 and Keumkang. The total starch of Gunji-3 was lower compared to the other wheat cultivars which showed a similar total starch (70.82 ± 0.24% versus 72.13 ± 0.19% and 72.16 ± 0.16%), whereas all the wheat cultivars showed no significant differences in the damaged starch content.
The SDS-sedimentation volume and protein content of Gunji-3 were higher than those of Shinmichal 1 and Keumkang. Gunji-3 showed an approximately 2% higher protein content than that of Keumkang as a non-waxy wheat (15.27 ± 0.15% and 13.27 ± 0.15%, respectively). Additionally, for the SDS-sedimentation volume, Gunji-3 showed a higher volume than that of Shinmichal 1 as a waxy-wheat (53.67 ± 0.58 mL and 39.50 ± 0.50 mL, respectively) and had a similar value compared to Keumkang as a non-waxy wheat (53.67 ± 0.58 mL and 59.50 ± 0.50 mL, respectively). In the mixograph, Gunji-3 showed a similar mixing time and tolerance to that of Shinmichal 1 and higher values compared to Keumkang; however, there was no difference in the water absorption.
3.3.2. Noodle dough sheet characteristics and texture of the cooked noodles.
The noodle dough sheet characteristics and texture of the cooked noodle of Gunji-3, Shinmichal 1, and Keumkang are summarized in Table 5. Gunji-3 showed a higher water absorption and lower thickness and whiteness index compared to the other wheat cultivars. Furthermore, in the comparison of the texture of cooked noodles between Gunji-3 and the two parental varieties, Gunji-3 exhibited the lowest values for hardness and cohesiveness. The springiness of Gunji-3 was lower than that of Keumkang and similar to that of Shinmichal 1.
3.3.3. Starch characteristics
The size distribution of starch granules and SEM images are presented in Table 2 and Figure 5a, respectively. The A-type of the starch granule of Gunji-3 was smaller than those of Shinmichal 1 and Keumkang (19.43 ± 0.39 μm, 22.17 ± 0.74 μm, and 20.33 ± 0.88 μm, respectively), while the B-type of the starch granule of all the wheat cultivars was not significantly different. For the amylose content, Shinmichal 1, the waxy wheat, showed a lower amylose level than that of Keumkang, a non-waxy wheat (7.45 ± 0.05% and 27.67 ± 0.26%, respectively). Gunji-3 showed the lowest amylose content at approximately a quarter of the Shinmichal1 amylose content, the waxy wheat (2.30 ± 0.15% and 7.45 ± 0.05%, respectively). As a result of the amylose content, Gunji-3 had the highest percentage for the water retention capacity.
The swelling properties of Gunji-3 showed the same characteristic of waxy wheat based on Shinmichal 1, i.e., the swelling volume and swelling power of Gunji-3 were higher than those of Keumkang (swelling volume: 8.30 ± 0.11 ml and 4.23 ± 0.06 ml & swelling power: 19.43 ± 0.06 g and 9.82 ± 0.07 g, respectively). For the pasting properties, Gunji-3 showed a similar value for the gelatinization Tm to that of Shinmichal 1; however, the value for the peak viscosity (295.00 ± 1.00 BU) was between Shinmichal 1 (403.33 ± 1.53 BU) and Keumkang (208.33 ± 1.00 BU) (Figure 5b). Additionally, Gunji-3 showed totally different characteristics for the holding strength and final viscosity compared to Shinmichal 1 and Keumkang. In the X-ray diffraction and gelatinization characteristics, Gunji-3 showed the same characteristics as the waxy wheat Shinmichal 1.
4. Discussion
Due to the current reduction in crop yields caused by various biotic and abiotic stresses and the increase in the global population, there is a growing demand for sustainable agriculture and nutritionally superior crop production. This has led to an increased desire for new genetic variations and the discovery of many genetic sources through them, among which wheat is currently being developed through various breeding programs with many varieties [36].
One of the most important things for such wheat is the starch content. Starch acts as a storage polysaccharide in many cereals and is one of the major constituents present in the wheat endosperm [37]. Wheat starch has performance-determining properties in many food applications and is closely related to the wheat shelf life and nutrients [10]. The quality of starch is closely related to the ratio between the amylose and the amylopectin forming the starch, in which a high amylose wheat starch makes it have unique functional properties [38,39].
The synthesis of GBSS I (Granule-Bound Starch Synthase I) controls the amylose content, which is determined by the Wx-A1, Wx-B1, and Wx-D1 genes [3]. The wild type Keumkang wheat used in this study has Wx-A1a, Wx-B1a and Wx-D1a in the Wx loci, and the waxy type Shinmichal 1 has Wx-A1b, Wx-B1b and Wx-D1b. (Figure 1a). This is consistent with previous findings of differences in the wheat Wx allele between wild-type and waxy-type wheat [40]. These wild-type to waxy-type mutations prevent Wx protein production through allelic variations of the Wx loci, and waxy wheat is null in all three Wx proteins [41]. In this context, Shimichal 1 had Wx-A1b, Wx-B1b, and Wx-D1b in the Wx sequence, and Gunji-3 had the same. In addition, the waxy protein confirmed by electrophoresis was not detected in both varieties (Figure 1b). However, Gunji-3 showed a much lower amylose content than ‘Shinmichal 1’ (Table 2). In addition, a previous study has shown that the starch of waxy wheat and non-waxy wheat has a similar amylopectin structure, and there is no significant difference in the expression of the starch granule proteins involved in amylopectin synthesis [37]. Based on these results, it can be implied that Gunji-3 is a mutant cultivar with a modified Wx protein.
To determine whether the low amylose content observed in the mutant cultivar Gunji-3 (DH185) compared to its parent cultivars was due to a mutation in the Wx protein, the sequences of the Wx genes were analyzed. It was found that the sequences of Wx-A1 and Wx-D1 in DH185 matched those of Wx-A1b and Wx-D1b, respectively, but the sequence of Wx-B1 in DH185 was different from that of Wx-B1b (Figure 2). To identify the precise sequence of Wx-B1 in DH185, the gene was cloned and sequenced in six parts using Wx-B1a as a reference (n=3). Through this process, the Wx-B1 gene in DH185 corresponding to 2,805 bp was identified. Comparison of the amino acid sequences of the newly identified Wx-B1 allele with those of previously known Wx-B1 alleles revealed differences at positions 39th (proline to alanine) and 45th (threonine to valine) (Figure 6), indicating the presence of a new allele, which was designated as Wx-B1o. As a result, it has been confirmed that the new mutant cultivar (DH185) is a new type of waxy wheat that is different from the previous ones.
The physicochemical properties of starch are important factors that significantly affect the noodle dough quality and cooked noodle texture [42]. Studies on starch properties are continuously performed to improve the quality of cooked noodles [32,42,43,44]. Springiness and cohesiveness of cooked noodles are affected by the ratio of the amylose and amylopectin in the starch and by different types of starch from various crops [45,46].
Based on the amylose content, wheat is generally classified into non-waxy (about 25-28%) and waxy types (about 0-3%). Moreover, there is partial waxy wheat in which the amylose content is from 16-22% [47]. Gunji-3 with low amylose is a waxy wheat with a lower amylose content than that of Shinmichal 1 which is produced as a partial waxy wheat [48]. The amylose content is negatively correlated with the swelling power [49]. In this study, we identified the same result, i.e., Gunji-3 showed a lower amylose content compared to Shinmichal 1 and Keumkang and a higher swelling power than those wheat varieties. On the other hand, the amylose content positively correlates with the hardness of the noodles [49,50]. Our result shows that the higher amylose content is, the higher hardness of cooked noodles is, i.e., Gunji-3 with the lowest amylose content among the three wheat varieties shows a lower hardness in the cooked noodles.
Flour possessing high swelling power, peak viscosity, and breakdown rate, and low gelatinization temperature is preferred for the production of premium white-salted noodles [51,52,53,54]. The noodle hardness has a negative correlation with the swelling power, peak viscosity, and final viscosity [49]. The noodle hardness, swelling power and final viscosity were similar to previous results, but the peak viscosity was different. The peak viscosity for Gunji-3 was between Shinmichal 1 and Keumkang. Gunji-3 had a much higher protein content than that of the parental wheat, while the SDS-sedimentation volume of Gunji-3 was similar to that of Keumkang and higher than that of Shinmichal 1. Other studies showed a higher protein content in wheat positively correlates with the SDS-sedimentation volume [55]. There is a negative relationship between the protein content and the optimum water absorption of noodle dough, but in this study, each wheat showed a significantly different protein content while the water absorption of the noodle dough was similar for the wheat varieties [56].
5. Conclusions
A novel waxy allele was identified in an organism resulting from a cross between wild-type and waxy-type wheat cultivars. Furthermore, the mutant cultivar exhibited a lower amylose content compared to the parental cultivar, while the seed protein content was higher, indicating a contrasting pattern. This finding, in combination with the new allele and diverse genetic backgrounds, presents a potential application in a wide range of breeding programs for related traits. Furthermore, it is expected that the ratio between amylose and amylopectin can be modulated to modify and improve the characteristics of the starch, depending on the intended usage of the wheat flour [15].
Acknowledgments
This research was financially supported by the “Cooperative Research Program for Agriculture Science and Technology Development (Project No. PJ01514904)” Rural Development Administration, South Korea.
References
- FAOSTAT, F., Forestry Database. 2019.
- Jeon, J.S., et al., Starch biosynthesis in cereal endosperm. Plant Physiol Biochem, 2010. 48(6): p. 383-92.
- Seib, P.A., Reduced-amylose wheats and Asian noodles. Cereal Foods World, 2000. 45: p. 504-512.
- Yamamori, M., T. Nakamura, and A. Kuroda, Variations in the content of starch-granule bound protein among several Japanese cultivars of common wheat (Triticum aestivum L.). Euphytica, 1992. 64(3): p. 215-219.
- Clark, J.R., M. Robertson, and C.C. Ainsworth, Nucleotide sequence of a wheat (Triticum aestivum L.) cDNA clone encoding the waxy protein. Plant Mol Biol, 1991. 16(6): p. 1099-101.
- Briney, A., et al., A PCR-based marker for selection of starch and potential noodle quality in wheat. Molecular Breeding, 1998. 4(5): p. 427-433.
- Murai, J., T. Taira, and D. Ohta, Isolation and characterization of the three Waxy genes encoding the granule-bound starch synthase in hexaploid wheat. Gene, 1999. 234(1): p. 71-9.
- Yan, L., et al., The genes encoding granule-bound starch synthases at the waxy loci of the A, B, and D progenitors of common wheat. Genome, 2000. 43(2): p. 264-72.
- Vrinten, P., T. Nakamura, and M. Yamamori, Molecular characterization of waxy mutations in wheat. Mol Gen Genet, 1999. 261(3): p. 463-71.
- 1Guzman, C. and J.B. Alvarez, Wheat waxy proteins: polymorphism, molecular characterization and effects on starch properties. Theor Appl Genet, 2016. 129(1): p. 1-16.
- 1Araki, E., H. Miura, and S. Sawada, Differential effects of the null alleles at the three Wx loci on the starch-pasting properties of wheat. Theoretical and Applied Genetics, 2000. 100(7): p. 1113-1120.
- Yamamori, M., et al., Genetic elimination of a starch granule protein, SGP-1, of wheat generates an altered starch with apparent high amylose. Theoretical and Applied Genetics, 2000. 101(1): p. 21-29.
- Yamamori, M. and C. Guzmán, SNPs and an insertion sequence in five Wx-A1 alleles as factors for variant Wx-A1 protein in wheat. Euphytica, 2013. 192(3): p. 325-338.
- Ayala, M., et al., Molecular characterization of waxy alleles in three subspecies of hexaploid wheat and identification of two novel Wx-B1 alleles. Theor Appl Genet, 2015. 128(12): p. 2427-35.
- Guzmán, C., et al., Molecular characterization of two novel null waxy alleles in Mexican bread wheat landraces. Journal of Cereal Science, 2015. 62: p. 8-14.
- Nakamura, T., et al., Rapid classification of partial waxy wheats using PCR-based markers. Genome, 2002. 45(6): p. 1150-6.
- Seo, Y.-W., B.-H. Hong, and Y.-W. Ha, Identification of granule bound starch synthase (GBSS) isoforms in wheat. KOREAN JOURNAL OF CROP SCIENCE, 1998. 43(2): p. 89-94.
- Jegasothy, H., M. Wootton, and R.J. Fairclough, Agarose gel electrophoresis of wheat starch. Journal of Cereal Science, 2000. 31(1): p. 75-78.
- Guzmán, C. and J.B. Alvarez, Molecular characterization of a novel waxy allele (Wx-A u 1a) from Triticum urartu Thum. ex Gandil. Genetic resources and crop evolution, 2012. 59: p. 971-979.
- Kearse, M., et al., Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics, 2012. 28(12): p. 1647-1649.
- Camacho, C., et al., BLAST plus : architecture and applications. Bmc Bioinformatics, 2009. 10.
- Dubat, A., A new AACC International approved method to measure rheological properties of a dough sample. Cereal Foods World (CFW), 2010. 55(3): p. 150.
- Nguimbou, R.M., et al., Effect of Cross-Section Differences and Drying Temperature on the Physicochemical, Functional and Antioxidant Properties of Giant Taro Flour. Food and Bioprocess Technology, 2013. 6(7): p. 1809-1819.
- Gibson, T.S., V.A. Solah, and B.V. McCleary, A procedure to measure amylose in cereal starches and flours with concanavalin A. Journal of Cereal Science, 1997. 25(2): p. 111-119.
- Gibson, T.S., H. Alqalla, and B.V. Mccleary, An Improved Enzymatic Method for the Measurement of Starch Damage in Wheat-Flour. Journal of Cereal Science, 1992. 15(1): p. 15-27.
- McCleary, B.V., T.S. Gibson, and D.C. Mugford, Measurement of total starch in cereal products by amyloglucosidase-alpha-amylase method: Collaborative study. Journal of Aoac International, 1997. 80(3): p. 571-579.
- Douglas, S.G., A rapid method for the determination of pentosans in wheat flour. Food Chemistry, 1981. 7(2): p. 139-145.
- Czuchajowska, Z. and Y. Pomeranz, Protein-Concentrates and Prime Starch from Wheat Flours. Cereal Chemistry, 1993. 70(6): p. 701-706.
- Kasemsuwan, T., et al., Characterization of the Dominant Mutant Amylose-Extender (Ael-5180) Maize Starch. Cereal Chemistry, 1995. 72(5): p. 457-464.
- Sollars, W.F., Fractionation and reconstitution techniques for studying water-retention properties of wheat flours. 1973.
- Yamamori, M., et al., Resistant starch and starch pasting properties of a starch synthase IIa-deficient wheat with apparent high amylose. Australian Journal of Agricultural Research, 2006. 57(5): p. 531-535.
- Sasaki, T. and J. Matsuki, Effect of wheat starch structure on swelling power. Cereal Chemistry, 1998. 75(4): p. 525-529.
- Hayakawa, K., et al., Quality characteristics of waxy hexaploid wheat (Triticum aestivum L.): Properties of starch gelatinization and retrogradation. Cereal Chemistry, 1997. 74(5): p. 576-580.
- Park, C.S. and B.K. Baik, Flour characteristics related to optimum water absorption of noodle dough for making white salted noodles. Cereal Chemistry, 2002. 79(6): p. 867-873.
- Park, C.S., B.H. Hong, and B.K. Baik, Protein quality of wheat desirable for making fresh white salted noodles and its influences on processing and texture of noodles. Cereal Chemistry, 2003. 80(3): p. 297-303.
- Trethowan, R.M. and A. Mujeeb-Kazi, Novel germplasm resources for improving environmental stress tolerance of hexaploid wheat. Crop science, 2008. 48(4): p. 1255-1265.
- Van Hung, P., et al., Waxy and high-amylose wheat starches and flours—characteristics, functionality and application. 2006. 17(8): p. 448-456.
- Zeng, M., et al., Sources of variation for starch gelatinization, pasting, and gelation properties in wheat. Cereal Chemistry, 1997. 74(1): p. 63-71.
- Fredriksson, H., et al., The influence of amylose and amylopectin characteristics on gelatinization and retrogradation properties of different starches. Carbohydrate polymers, 1998. 35(3-4): p. 119-134.
- Nakamura, T., et al., Identification of three Wx proteins in wheat (Triticum aestivum L.). Biochemical genetics, 1993. 31: p. 75-86.
- Graybosch, R.A., Waxy wheats: Origin, properties, and prospects. Trends in Food Science & Technology, 1998. 9(4): p. 135-142.
- Zhang, K., et al., Effect of different starch acetates on the quality characteristics of frozen cooked noodles. 2022. 10(3): p. 678-688.
- Ragaee, S. and E.-S.M.J.F.c. Abdel-Aal, Pasting properties of starch and protein in selected cereals and quality of their food products. 2006. 95(1): p. 9-18.
- Singh, S., et al., Relationship of granule size distribution and amylopectin structure with pasting, thermal, and retrogradation properties in wheat starch. 2010. 58(2): p. 1180-1188.
- Heo, H., et al., Influence of amylose content on cooking time and textural properties of white salted noodles. 2012. 21: p. 345-353.
- Huang, Y.-C. and H.-M.J.J.o.F.E. Lai, Noodle quality affected by different cereal starches. 2010. 97(2): p. 135-143.
- Van Hung, P., T. Maeda, and N. Morita, Waxy and high-amylose wheat starches and flours—characteristics, functionality and application. Trends in Food Science & Technology, 2006. 17(8): p. 448-456.
- Heo, H., et al., A new waxy wheat cultivar" Shinmichal1" with stress tolerance. 2007. 39(3): p. 385-386.
- Li, Q., et al., A molecular explanation of wheat starch physicochemical properties related to noodle eating quality. 2020. 108: p. 106035.
- Abdel-Aal, E.S., et al., Physicochemical and structural characteristics of flours and starches from waxy and nonwaxy wheats. 2002. 79(3): p. 458-464.
- Crosbie, G.J.J.o.c.s., The relationship between starch swelling properties, paste viscosity and boiled noodle quality in wheat flours. 1991. 13(2): p. 145-150.
- Konik, C.M., et al., Contribution of starch and non-starch parameters to the eating quality of Japanese white salted noodles. 1992. 58(3): p. 403-406.
- McCormick, K., J. Panozzo, and S.J.A.J.o.A.R. Hong, A swelling power test for selecting potential noodle quality wheats. 1991. 42(3): p. 317-323.
- Oh, N., et al., Starch damage on the quality characteristics of dry noodles'. 1985. 62(6): p. 441-446.
- De Villiers, O., E.J.S.A.J.o.P. Laubscher, and Soil, Use of the SDSS test to predict the protein content and bread volume of wheat cultivars. 1995. 12(4): p. 140-142.
- Oh, N., et al., Noodles. V. Determination of optimum water absorption of flour to prepare oriental noodles. 1986. 63(2): p. 93-96.
Figure 1.
The expression patterns of the waxy genes and proteins in three wheat cultivars: (a) Gel electrophoresis of the polymerase chain reaction amplification of the waxy gene; (b) One-dimensional SDS-PAGE patterns of the waxy protein. M, molecular size marker; K, Keumkang (wild type); S, Shinmichal 1 (waxy type); G, Gunji-3 (waxy mutant).
Figure 1.
The expression patterns of the waxy genes and proteins in three wheat cultivars: (a) Gel electrophoresis of the polymerase chain reaction amplification of the waxy gene; (b) One-dimensional SDS-PAGE patterns of the waxy protein. M, molecular size marker; K, Keumkang (wild type); S, Shinmichal 1 (waxy type); G, Gunji-3 (waxy mutant).
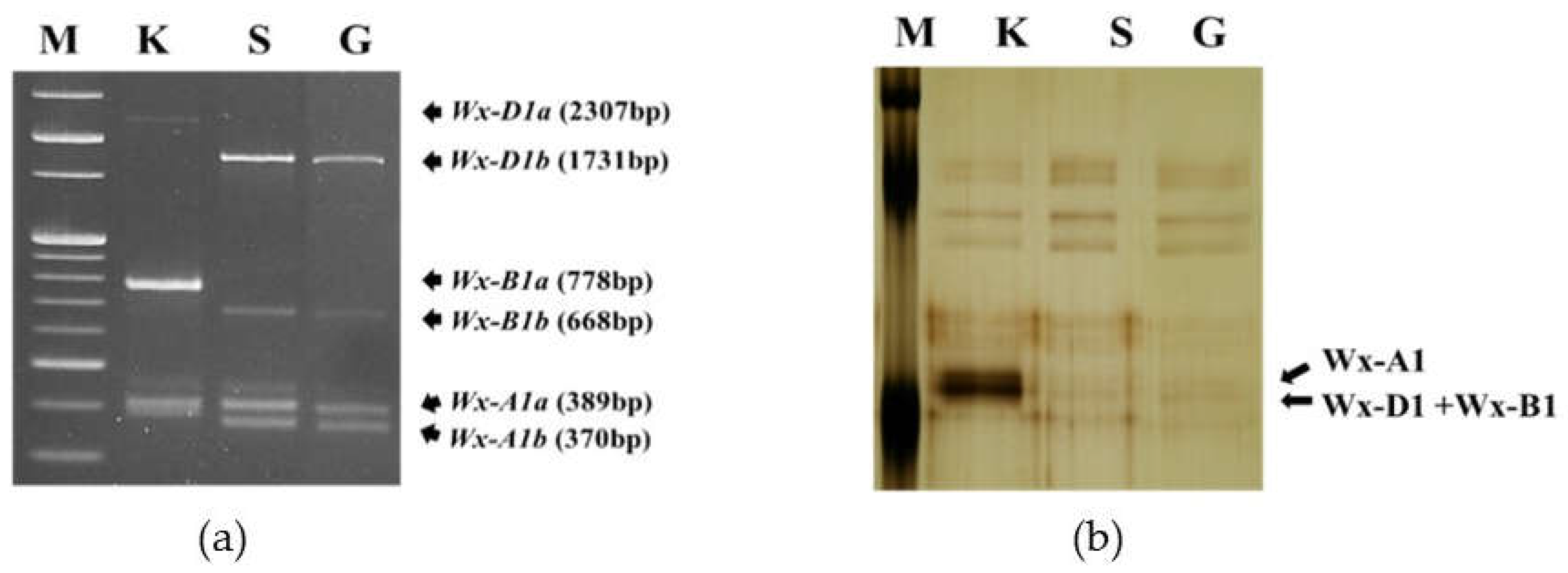
Figure 2.
Alignment of the waxy gene sequence of DH185.
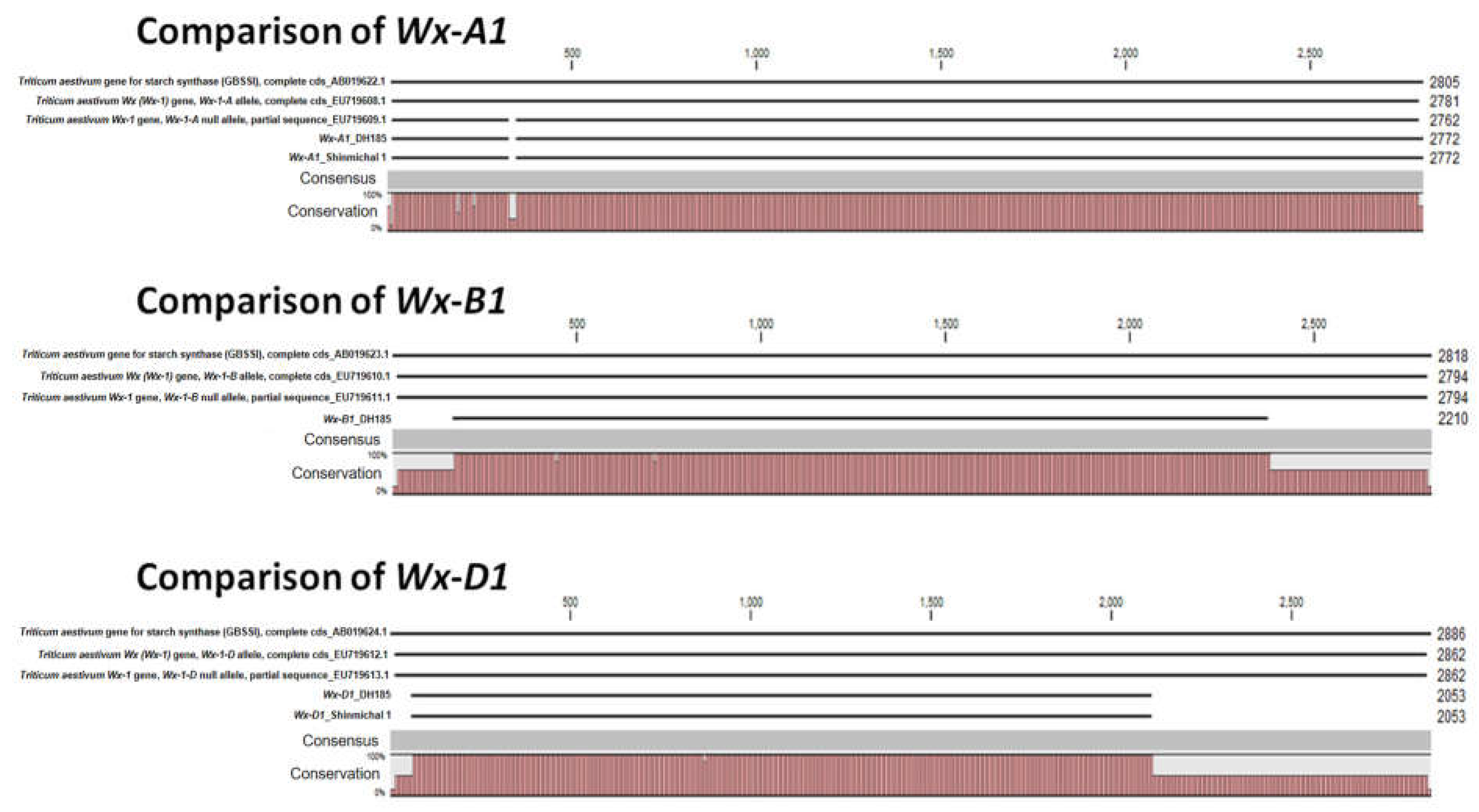
Figure 3.
Alignment of the genomic DNA sequences between DH185 (Gunji-3) and Wx-B1a.
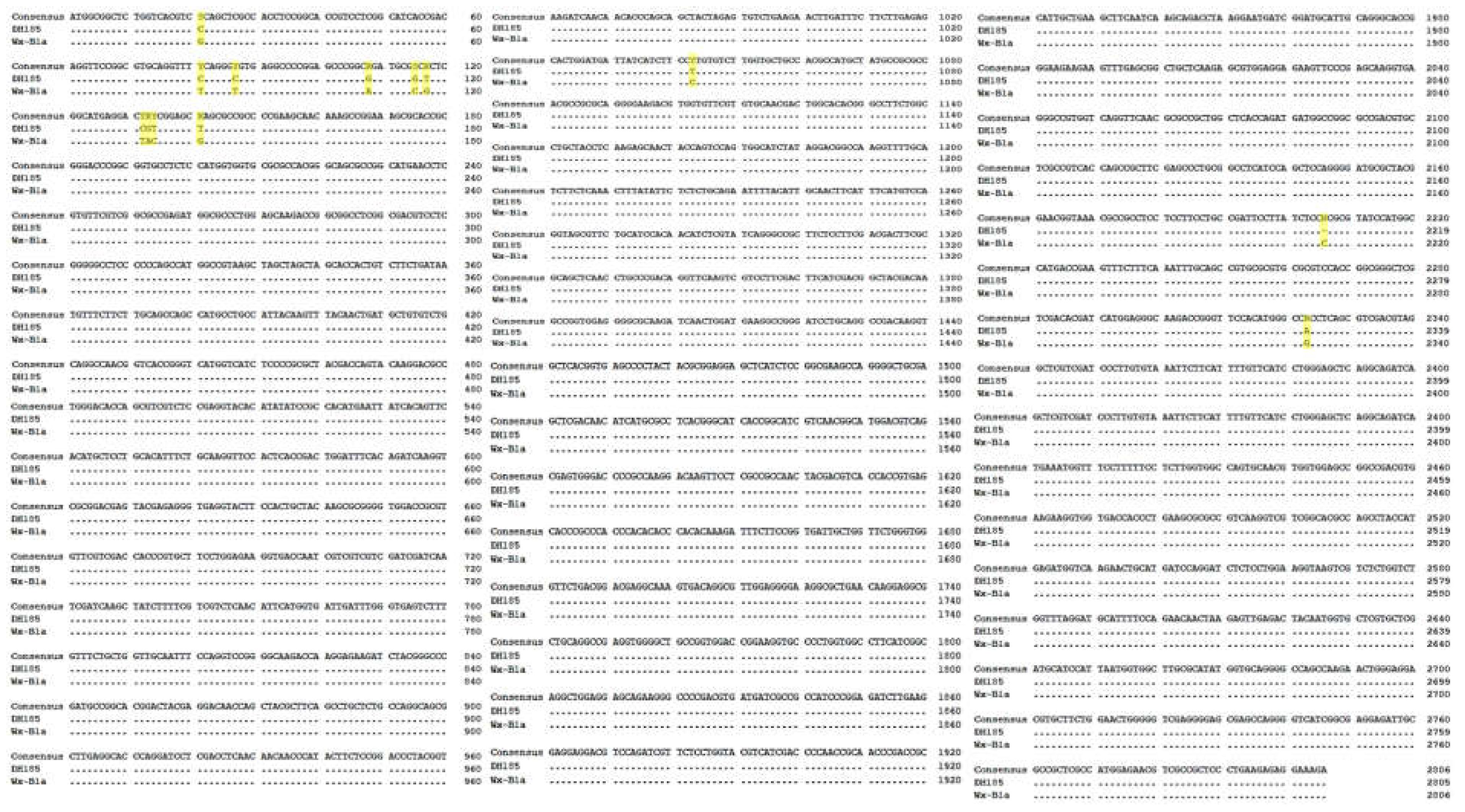
Figure 4.
Molecular characterization of the novel Wx-B1o allele: (a) Diagrammatic representation of the Wx-B1a and Wx-B1o. Green regions encode the waxy protein. The mint-colored portion was not analyzed; (b) Alignment of deduced amino-acid sequences to compare modified region between DH185 (Gunji-3) and Wx-B1a. The red arrow indicates the 740th amino acid position. The green-highlighted part is the non-matching part.
Figure 4.
Molecular characterization of the novel Wx-B1o allele: (a) Diagrammatic representation of the Wx-B1a and Wx-B1o. Green regions encode the waxy protein. The mint-colored portion was not analyzed; (b) Alignment of deduced amino-acid sequences to compare modified region between DH185 (Gunji-3) and Wx-B1a. The red arrow indicates the 740th amino acid position. The green-highlighted part is the non-matching part.
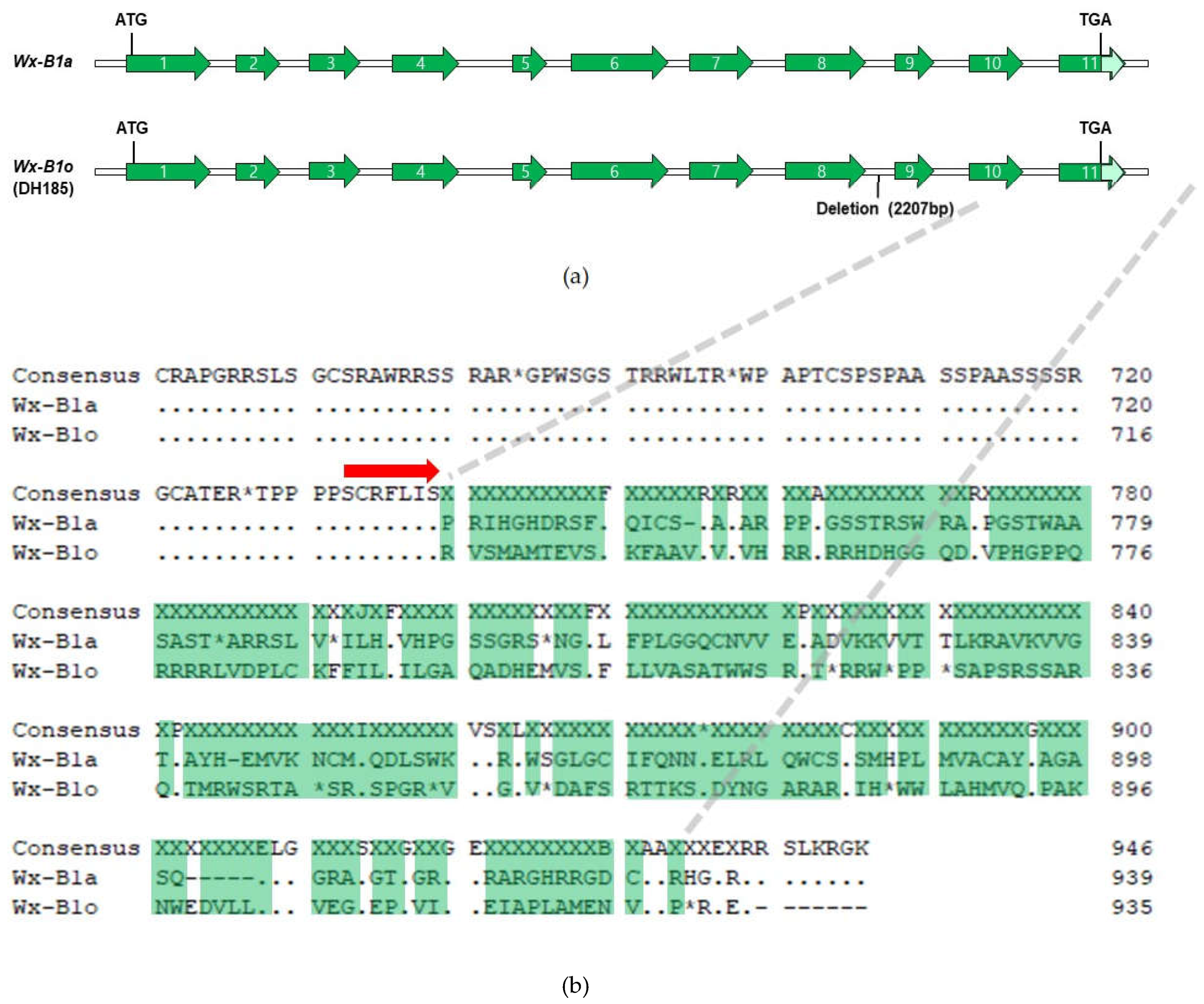
Figure 5.
Comparison of starch structure and viscosity among the three wheat varieties: (a) Scanning electron micrographs of starch isolated from a waxy bread wheat mutant (Gunji-3), waxy wheat (cv. Shinmichal 1) and wild type wheat genotypes (cv. Keumkang); (b) Micro Visco-Amylo-Graph pasting curves of starch isolated from a waxy bread wheat mutant (Gunji-3), waxy wheat (cv. Shinmichal 1) and wild type wheat genotypes (cv. Keumkang). Scale bar = 20 µm.
Figure 5.
Comparison of starch structure and viscosity among the three wheat varieties: (a) Scanning electron micrographs of starch isolated from a waxy bread wheat mutant (Gunji-3), waxy wheat (cv. Shinmichal 1) and wild type wheat genotypes (cv. Keumkang); (b) Micro Visco-Amylo-Graph pasting curves of starch isolated from a waxy bread wheat mutant (Gunji-3), waxy wheat (cv. Shinmichal 1) and wild type wheat genotypes (cv. Keumkang). Scale bar = 20 µm.
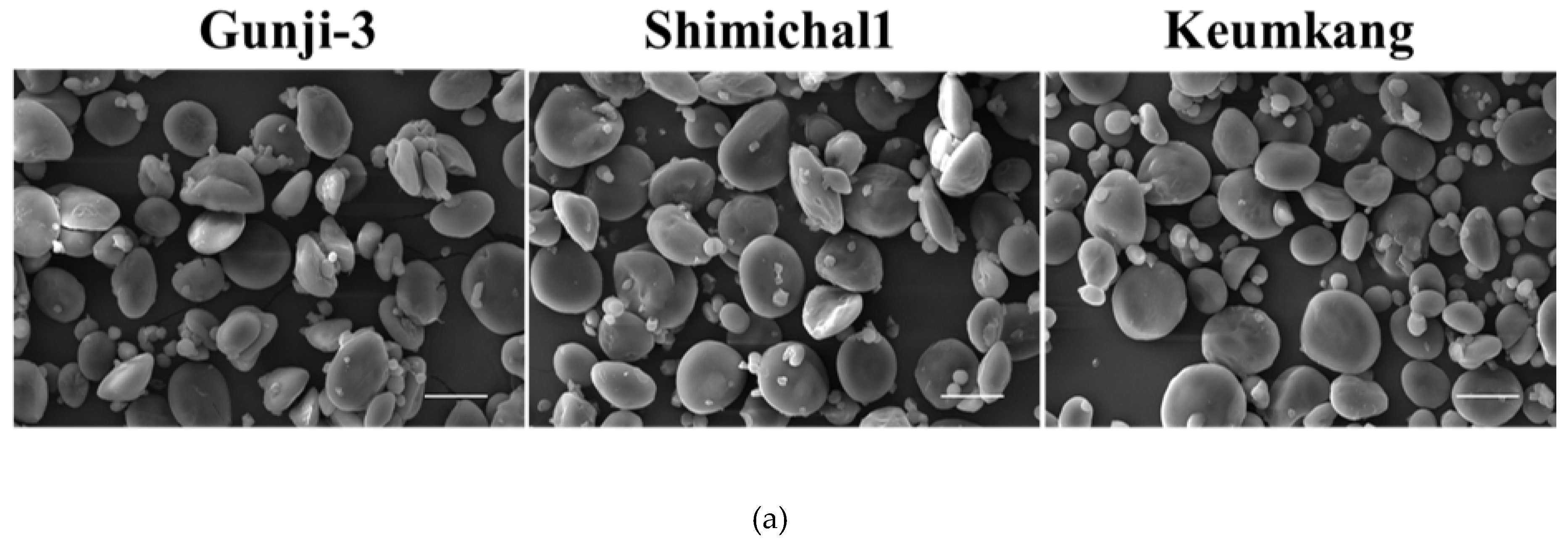
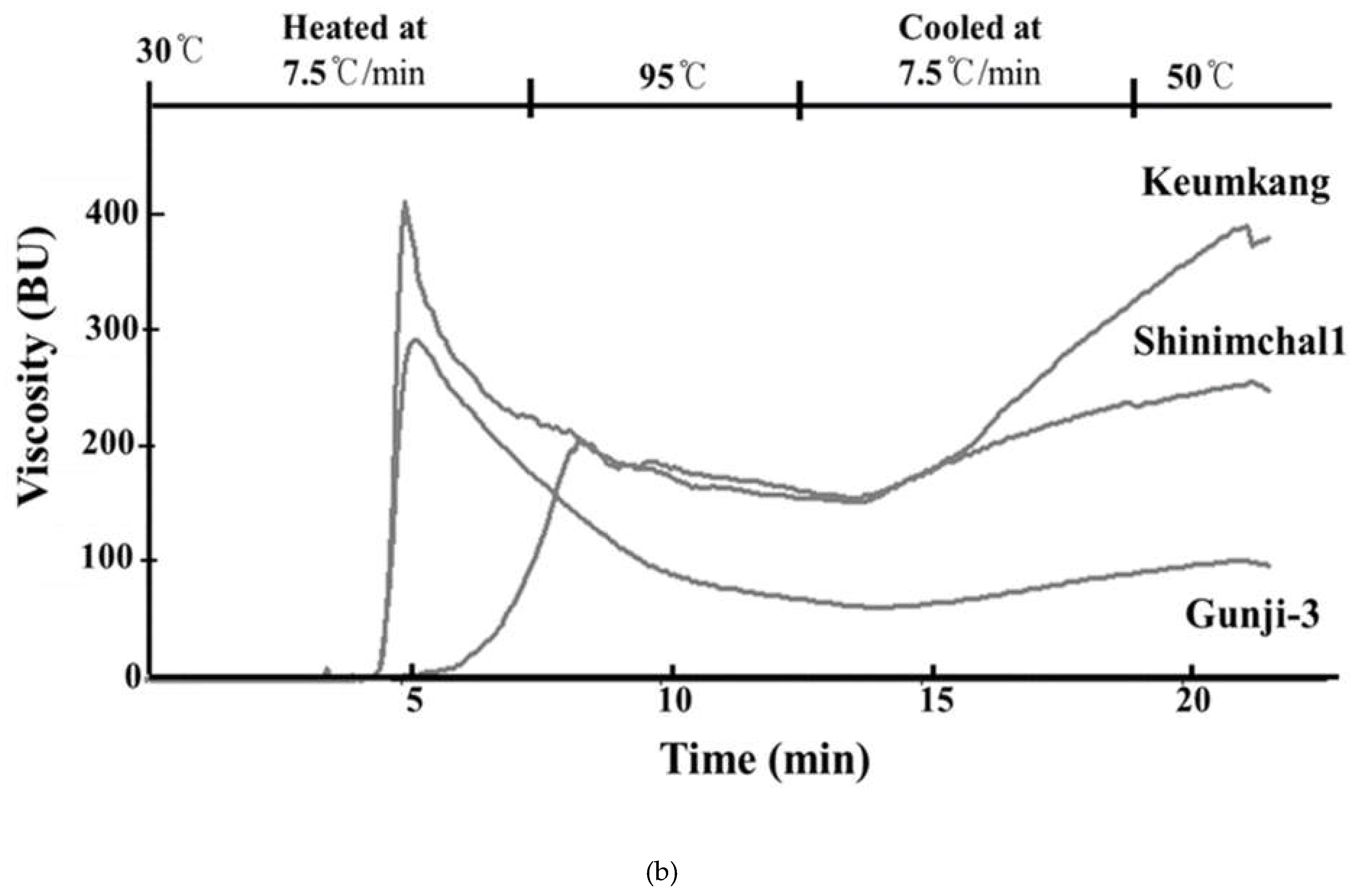
Figure 6.
Comparison of amino acid sequences between Wx-B1 (Wx-B1o) of DH185 and Wx-B1 alleles. The red box indicates the amino acids that differ only in Wx-B1o compared to other Wx-B1 alleles.
Figure 6.
Comparison of amino acid sequences between Wx-B1 (Wx-B1o) of DH185 and Wx-B1 alleles. The red box indicates the amino acids that differ only in Wx-B1o compared to other Wx-B1 alleles.
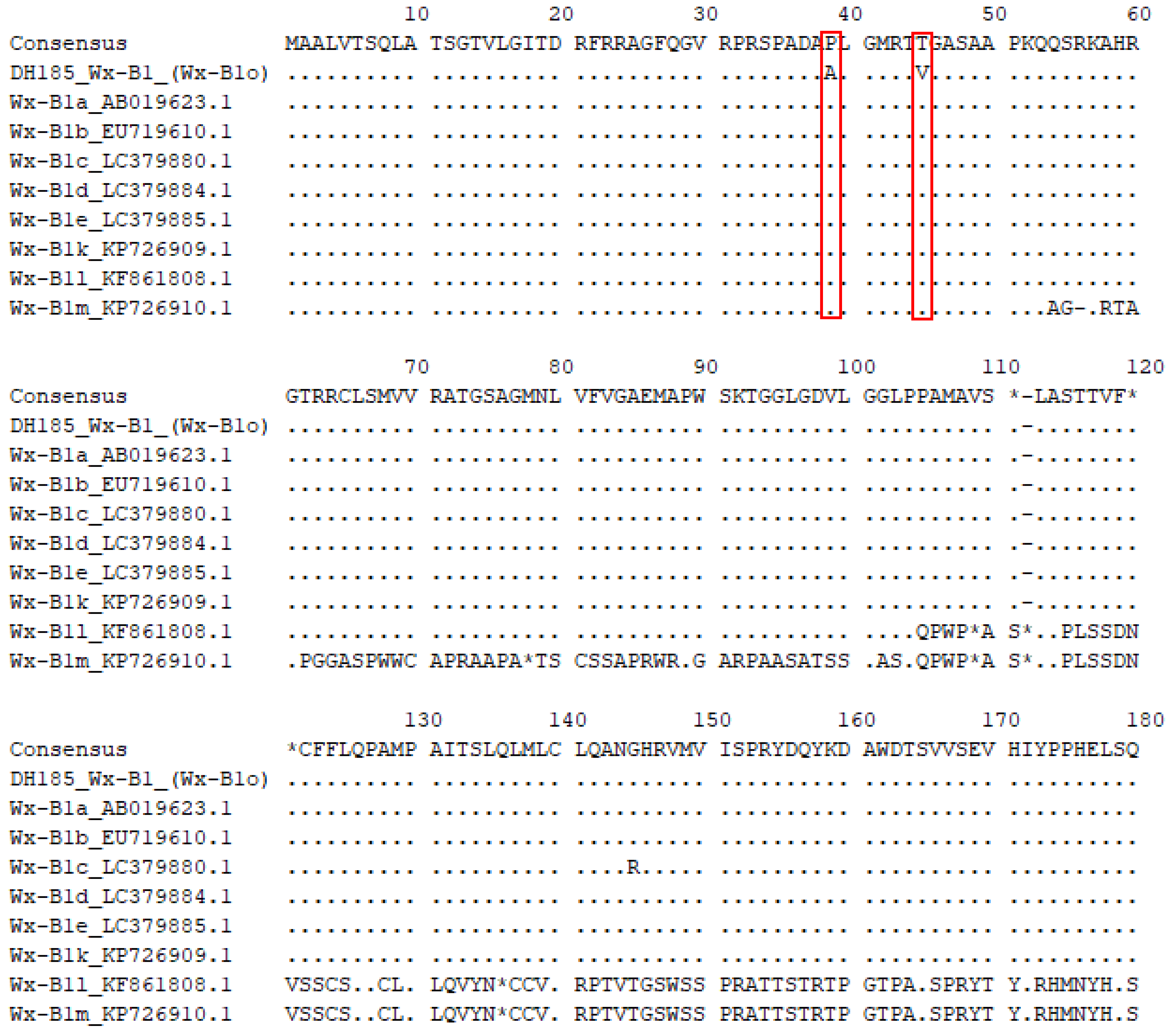

Table 1.
Description of PCR primer pairs for the amplification.
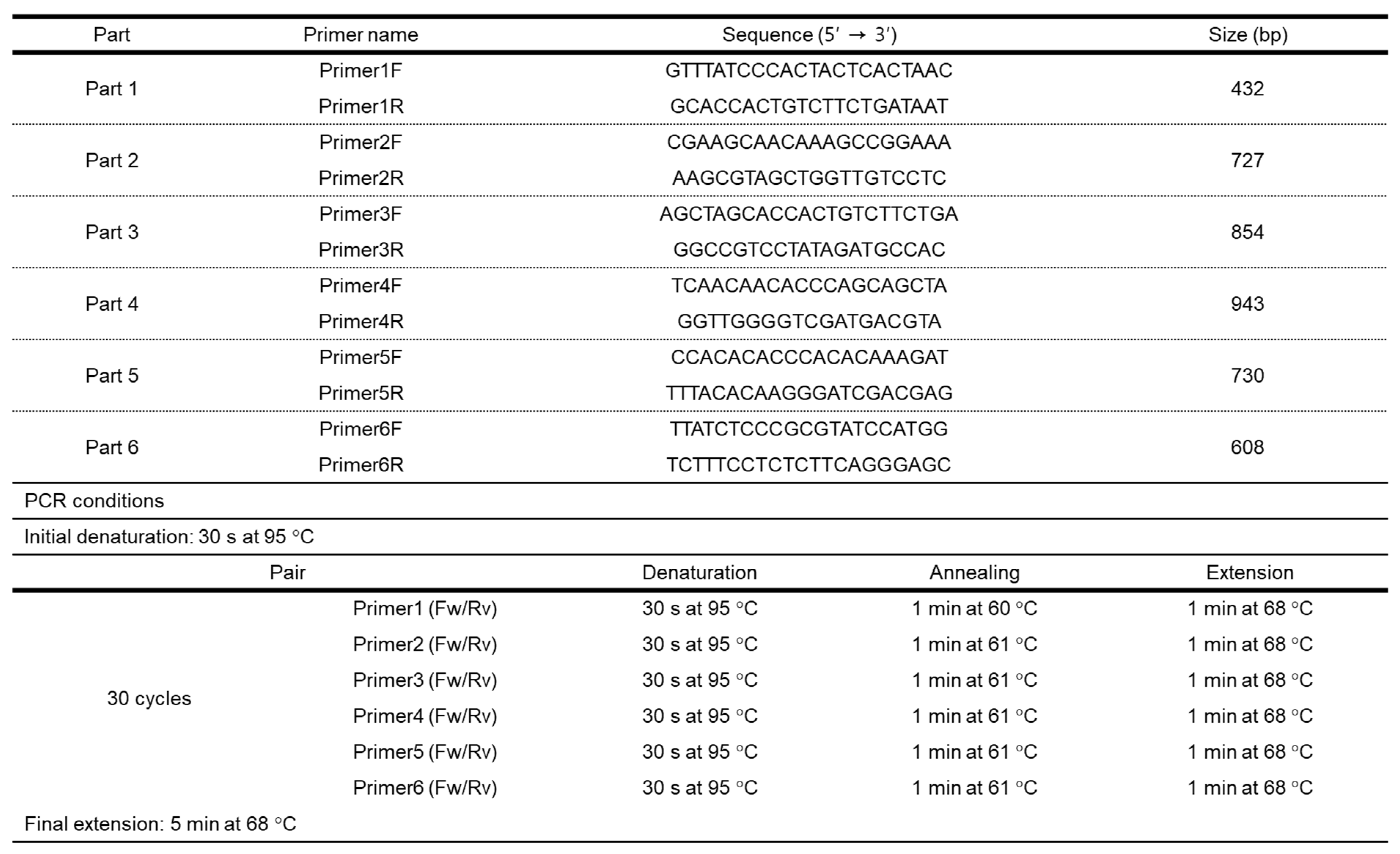
Table 2.
Comparison of starch characteristics between the waxy-type mutant (Gunji-3) and parental wheat varieties revealed the following results a.
Table 2.
Comparison of starch characteristics between the waxy-type mutant (Gunji-3) and parental wheat varieties revealed the following results a.
| Characteristics | Gunji-3 | Shinmichal 1 | Keumkang | |
|---|---|---|---|---|
| Starch Granule Size (µm) | ||||
| A-type | 19.43 ± 0.39b | 22.17 ± 0.74a | 20.33 ± 0.88b | |
| B-type | 4.17 ± 0.06a | 4.09 ± 0.12a | 4.22 ± 0.22a | |
| Amylose (%) | 2.30 ± 0.15c | 7.45 ± 0.05b | 27.67 ± 0.26a | |
| Cold water retention capacity (%) | 84.66 ± 1.05a | 75.91 ± 0.69b | 66.07 ± 1.05c | |
| Swelling properties | ||||
| Swelling volume (ml) | 8.30 ± 0.11a | 8.40 ± 0.10a | 4.23 ± 0.06b | |
| Swelling power (g) | 19.43 ± 0.06a | 18.35 ± 0.04b | 9.82 ± 0.07c | |
| Pasting properties | ||||
| Gelatinization Tm (℃) | 64.97 ± 0.12b | 64.13 ± 0.71b | 75.10 ± 0.17a | |
| Peak viscosity (BU) | 295.00 ± 1.00b | 403.33 ± 1.53a | 208.33 ± 1.00c | |
| Tm at Peak viscosity (℃) | 69.87 ± 0.21b | 67.27 ± 0.50c | 92.77 ± 0.38a | |
| Holding strength (BU) | 63.33 ± 2.08c | 159.00 ± 1.00a | 145.33 ± 2.52b | |
| Final viscosity (BU) | 94.33 ± 1.53c | 243.67 ± 2.08b | 336.67 ± 2.52a | |
| Breakdown (BU) | 231.67 ± 2.89b | 244.33 ± 2.31a | 63.00 ± 2.00c | |
| Setback (BU) | 31.00 ± 1.00c | 84.67 ± 1.53b | 191.33 ± 4.62a | |
| Gel hardness (N) | 0.38 ± 0.01b | 0.49 ± 0.03b | 17.56 ± 0.23a | |
| X-ray diffraction | ||||
| Patterns | A | A | A | |
| Degree of crystallinity (%) | 18.68 ± 0.16a | 17.40 ± 0.24a | 11.92 ± 0.01b | |
| Gelatinization characteristicsb | ||||
| Onset temperature (°C, To) | 57.91 ± 1.20a | 56.95 ± 0.47ab | 54.80 ± 1.36b | |
| Peak temperature (°C, Tp) | 64.08 ± 0.11a | 64.04 ± 0.10a | 59.71 ± 0.21b | |
| Transition enthalpy (J/g, ΔH) | 146.19 ± 0.32a | 143.60 ± 5.74a | 124.68 ± 0.32b |
aStatistical analysis was performed to determine significant differences between mean values in the same row, denoted by different letters, using the 5% level of significance (P < 0.05). bGelatiniaztion characteristics were measured by differential scanning calorimetry.
Table 3.
The BLASTN results for the comparison of DH185 Wx-B1 with other Wx-B1 alleles.
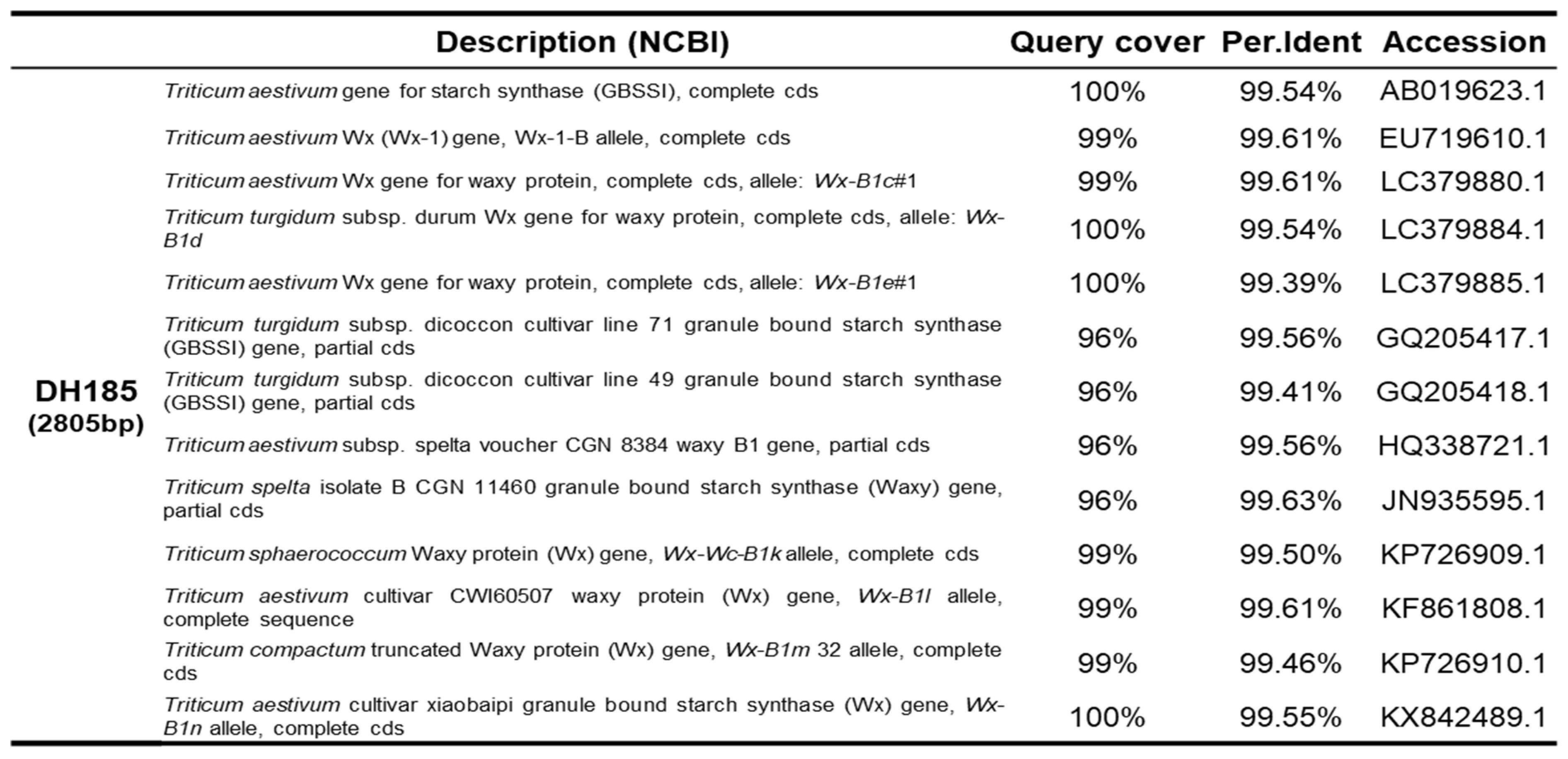
Table 4.
Comparison of flour characteristics between the waxy-type mutant (Gunji-3) and parental wheat varieties revealed the following results a.
Table 4.
Comparison of flour characteristics between the waxy-type mutant (Gunji-3) and parental wheat varieties revealed the following results a.
| Characteristics | Gunji-3 | Shinmichal 1 | Keumkang | |
|---|---|---|---|---|
| Flour yield (%) | 71.23 ± 0.74b | 63.60 ± 0.08c | 73.43 ± 0.02a | |
| Ash (%) | 0.43 ± 0.01a | 0.44 ± 0.01a | 0.45 ± 0.01a | |
| Whiteness index | 89.86 ± 0.06ab | 90.02 ± 0.07a | 88.44 ± 0.06b | |
| Average of particle size (μm) | 79.77 ± 0.46a | 81.73 ± 1.67a | 77.61 ± 0.66b | |
| Total starch (%) | 70.82 ± 0.24b | 72.13 ± 0.19a | 72.16 ± 0.16a | |
| Damaged starch (%) | 4.01 ± 0.01a | 4.12 ± 0.06a | 3.96 ± 0.02a | |
| Arabinoxylan (%) | 1.74 ± 0.01b | 1.87 ± 0.01a | 1.55 ± 0.04c | |
| Protein (%) | 15.27 ± 0.15a | 11.19 ± 0.14c | 13.27 ± 0.15b | |
| SDS-sedimentation volume (mL) | 53.67 ± 0.58a | 39.50 ± 0.50b | 59.50 ± 0.50a | |
| Mixograph absorption (%) | 64.33 ± 0.58a | 65.00 ± 1.00a | 65.17 ± 0.29a | |
| Mixing time (min) | 2.40 ± 0.10b | 2.60 ± 0.01b | 4.03 ± 0.06a | |
| Mixing tolerance (mm) | 19.67 ± 0.58b | 21.67 ± 0.58b | 24.67 ± 1.53a |
aStatistical analysis was performed to determine significant differences between mean values in the same row, denoted by different letters, using the 5% level of significance (P < 0.05).
Table 5.
The comparative analysis of the noodle dough sheet characteristics and texture properties of cooked noodles using the waxy-type mutant (Gunji-3) and parental wheat varieties yielded the following results a.
Table 5.
The comparative analysis of the noodle dough sheet characteristics and texture properties of cooked noodles using the waxy-type mutant (Gunji-3) and parental wheat varieties yielded the following results a.
| Characteristics | Gunji-3 | Shinmichal 1 | Keumkang | |
|---|---|---|---|---|
| Noodle dough sheet | ||||
| Water absorption (%) | 41.00a | 37.00b | 34.00c | |
| Thickness (mm) | 1.80 ± 0.03b | 1.88 ± 0.01a | 1.89 ± 0.02a | |
| Whiteness index | 76.01 ± 0.01b | 77.76 ± 0.01a | 76.01 ± 0.01b | |
| Texture of cooked noodles | ||||
| Hardness (N) | 1.07 ± 0.01c | 1.55 ± 0.11b | 3.35 ± 0.11b | |
| Springiness (Ratio) | 0.83 ± 0.01b | 0.85 ± 0.02b | 0.88 ± 0.02a | |
| Cohesiveness (Ratio) | 0.63 ± 0.01c | 0.70 ± 0.01a | 0.66 ± 0.01b |
aStatistical analysis was performed to determine significant differences between mean values in the same row, denoted by different letters, using the 5% level of significance (P < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



