
Submitted:
19 May 2023
Posted:
22 May 2023
You are already at the latest version
Abstract
Inflammation is the response of the innate immune system to any type of injury. Although acute inflammation is critical for survival, dysregulation of innate immune response leads to chronic in-flammation. Many synthetic anti-inflammatory drugs have side effects, and thus, natural an-ti-inflammatory compounds are still needed. Cannabis sativa L. may provide a good source of in-flammation-reducing molecules. Here, we tested the anti-inflammatory properties of cannabis ex-tracts and pure cannabinoids in lipopolysaccharide (LPS)-induced inflammation in human THP-1 macrophages. We found that pre-treatment with cannabidiol (CBD), delta-9-tetrahydrocannabinol (THC), or extracts containing high levels of CBD or THC reduce the level of induction of various cytokines. The CBD was more efficient than THC, and the extracts were more efficient than pure cannabinoids. Finally, IL-6, IL-10 and monocyte chemoattractant protein-1 (MCP-1) were most sensitive to pre-treatments with CBD and THC, while IL-1β, IL-8 and TNF-α were less responsive. Thus, our work demonstrates the potential of use of cannabinoids or/and cannabis extracts for the reduction of inflammation and establishes IL-6 and MCP-1 as the sensitive markers for the analysis of the effect of cannabinoids on inflammation in macrophages.
Keywords:
cannabidiol
; delta-9-hydrocannabinol
; CBD
; THC
; THP-1 macrophages
; Cannabis sativa
; inflammation
; lipopolysaccharide
1. Introduction
The positive impact of cannabis on human health has been known for many years [1]. C. sativa plant contains many compounds, but the most well-known are cannabinoids, terpenes, and flavonoids. Cannabinoids act via the endocannabinoid system (ECS), which consists of receptors like cannabinoid 1 (CB1), cannabinoid 2 (CB2), transient receptor potential channels of the vanilloid subtype 1 and 2 (TRPV1, TRPV2), G protein-coupled receptors 18, 55, 119 (GPR18, GPR55, GPR119), endocannabinoids like 2-arachidonoylglycerol and anandamide (2-AG, AEA), and their biosynthetic and degradation enzymes. Homeostasis maintenance is the main function of ECS [2]. The most studied phytocannabinoids produced by C. sativa plant are cannabidiol (CBD) and Δ9-tetrahydrocannabinol (THC). The effect of cannabis extracts is frequently different from single cannabinoids, with extracts often being more efficient – this is known as an entourage effect. This phenomenon is analogous to the orchestra when the complex interaction between minor cannabinoids, flavonoids, and terpenes are more potent in comparison to each individual compound [3]. The main cannabinoid receptors, CB1 and CB2, are highly expressed on immune cells, such as macrophages, T lymphocytes, and B cells, as well as in lungs and the bronchial tissue [4,5]. Therefore, cannabis extracts, as well as their active ingredients, can modulate the function of immune system and inflammation in general.
Inflammation is a body’s vital defensive mechanism. Epithelial cells and macrophages are often the first line of defence against bacteria, viruses, and toxins [6]. Bacteria express pathogen-associated molecular patterns (PAMPs) that are recognized by pathogen recognition receptors (PRRs) present on immune and non-immune cells. Lipopolysaccharide (LPS), being a component of gram-negative bacterial cell wall, is an example of PAMPs [7]. Chronic inflammation plays an important role in the pathogenesis of many chronic conditions, like inflammatory bowel diseases, obesity, diabetes mellitus, atherosclerosis, asthma, Alzheimer’s disease, and many others [8]. The uncontrolled and continuous release of pro-inflammatory cytokines is associated with organ damage and chronic disease progression [9]. The most frequently induced cytokines are TNF-α, IL-1β, IL-1, IL-6, IL-8, IL-10, monocyte chemoattractant protein-1 (MCP-1), and IL-8. The pro-inflammatory cytokines (for instance, MCP-1 and IL-8) induce chemotaxis of neutrophils, monocytes, T cells, and dendritic cells to the site of infection and stimulate phagocytosis [10,11]. TNF-α and IL-1β are key mediators of the inflammatory response [12,13]. IL-6 stimulates inflammation and induces synthesis of acute phase proteins [14]. The anti-inflammatory cytokines, like IL-10, function to limit a host immune response to pathogens, tissue homeostasis maintenance, the prevention of autoimmune conditions development [15]. The balance between pro-inflammatory and anti-inflammatory cytokines is crucial for proper tissue homeostasis.
The anti-inflammatory effects of cannabis extracts, as well as its active ingredients, are well documented [16]. For instance, cannabis users suffering from multiple sclerosis had lower levels of pro-inflammatory cytokines (TNF-α, IL-1, IL-6, IFN-γ), while higher levels of anti-inflammatory cytokine (IL-10) as compared to non-cannabis users [17]. Cannabis extracts high in CBD and THC suppressed the expression of IL-6, IL-1β, MCP-1, and TNF-α genes induced by TNF-α/IFN-γ in human 3D EpiDermFT tissue [18]. Cannabis extract high in CBD was demonstrated to be more potent than pure CBD in suppressing inflammation by preventing TNF-α production, reducing pain and paw swelling in the zymosan-induced inflammation mice model [19]. Another study demonstrated that high CBD cannabis extract was superior to CBD in reducing IL-8 and IL-6 levels in vitro on the A549 alveolar epithelial cell line [20]. In addition, cannabis extracts high in CBD and THC were shown to be more effective than pure CBD and THC in graft versus host disease in allogeneic bone marrow transplantation model by reducing inflammation and improving survival rate [21]. Our previous work also demonstrated that cannabis extracts were efficient in reducing inflammation in skin fibroblasts, lung fibroblasts and human 3D tissues [18,22,23].
In this work, we attempted to study the effects of cannabis extracts on the lipopolysaccharides (LPS)-induced cytokine release in human THP-1 leukemia monocytes. We aimed to establish the model of induction of inflammation and hypothesized that cannabinoids will prevent strong induction of inflammation. We further hypothesized that extracts differ from pure cannabinoids in their anti-inflammatory properties. Here, we found that inflammation was induced efficiently at 3 h after application of LPS and was partially prevented by exposure to CBD and various cannabis extracts. We also noted that IL-6, IL-10 and monocyte chemoattractant protein-1 (MCP-1) were more sensitive to pre-treatments with cannabinoids, as compared to IL-1β, IL-6 and TNF-α.
2. Results
2.1. Analysis of cannabinoids content using HPLC
We first tested the CBD and THC concentrations of three extracts we planned to use in the study. We found that extracts #14 and #178 were high in THC, while #131 was a high CBD extract (Table 1). We also calculated the amount of CBD and THC (in µM) in the extracts at the concentration of 7 µg/mL. This concentration was found to be not toxic to normal skin and lung fibroblasts [23].
We hypothesized that terpene composition and concentration may have an effect on anti-inflammatory properties and thus tested the terpenes profile (Table 2). Cultivar #14 was dominant in trans-caryophyllene and limonene, cultivar #131 was dominant in -3-carene, α-pinene, β-pinene and limonene, while cultivar #178 was high in α-pinene, β-pinene, β-myrcene and α-bisabolol.
2.2. Analysis of time of induction of inflammation by LPS
To establish the inflammation model, we tested the time of the induction of pro-inflammatory cytokines. We exposed human THP-1 macrophages to LPS for 3 and 6 h; we used 0.5 μg/mL of LPS as this concentration was shown to be effective in induction of inflammation in cell models [24]. We then analyzed the expression of NFκB and p-NFκB using Western blotting. Our aim was to observe upregulation in p-NFκB expression and the corresponding downregulation of NFκB expression to assume that our in vitro inflammation model is working. The quantification of p-NFκB/NFκB ratio showed that the desired inflammatory response was attained at 3 h post-LPS stimulation (Figure 1).
After determining that 3 h is the best time for the induction of inflammation by the LPS, we attempted to find out which sample is better for the analysis, the cell culture supernatant (media) or the cell lysate. We exposed cells to LPS for 3 h, harvested media and cell lysates separately and performed a multiplex ELISA experiment. We found that LPS media samples (3 h of treatment) had much higher levels of IL-1β, TNF-α and IL-10 cytokines in comparison to LPS cell lysate samples (Figure S1). It was concluded that media samples are better for the cytokines analysis, that is why we used cell media for Multiplex ELISA in the analysis of the effect of cannabinoids on inflammation.
2.3. Treatment with selected concentration of LPS, THC, CBD and extracts do not reduce the viability of macrophages
To test the effect of cannabis extracts and pure cannabinoids, CBD and THC, on the reduction of inflammation triggered by LPS, we first tested the viability THP-1 macrophages in response to LPS (0.5 µg/mL), CBD (5 µM), THC (5 µM) or of aforementioned extracts (7 μg/mL). We found that the viability of THP-1 macrophages was not changed in response to any treatment (Figure S2). The cell morphology also did not change (data not shown). This is important, because the level of inflammation can change due to the impact of active ingredients on cell viability.
2.4. Cannabis sativa extracts attenuate TNF-α, IL-1β, IL-8, MCP-1, IL-10, and IL-6 production in LPS-stimulated THP-1 macrophages
As explained in the methods, after differentiation, macrophages were pre-treated with cannabinoids or cannabis extracts, and LPS was added 30 min later. Cell supernatants were evaluated for the levels of cytokines 3 h after LPS exposure.
LPS treatment drastically increased the level of all cytokines. CBD pre-treatment significantly decreased the levels of IL-1β, while high CBD extract #131 did not (Figure 2A). Pre-treatment with THC decreased IL-1β but the change was not significant, while high THC-containing extracts #14 and #178 decreased it significantly (Figure 2B). The overall increase in IL-6 was lower compared to other cytokines in all tested groups. Although both pure cannabinoids and cannabis extracts significantly reduced IL-6 levels, the cannabis extracts were more efficient (Figure 2C, D). Pure THC did not affect the levels IL-8 post-LPS; however, pure CBD, high CBD and high THC extracts significantly reduced IL-8 levels. (Figure 2E, F). Curiously, pure CBD or THC did not prevent the increase in MCP-1, while extracts have done this very effectively (Figure 2G, H). The increase in TNF-α was significantly decreased by CBD, but not by THC, while extracts again were more efficient (Figure 2I, J). CBD prevented the increase in IL-10, while THC did not, and again, the extracts have done this efficiently.
We summarized our findings in the Table 3 below. It appears that CBD is more efficient than THC in the prevention of the increase in cytokine release. Moreover, it is apparent that the extracts are more efficient than CBD or THC, and this was especially apparent in THC group comparison.
We found that high THC extracts (#14 and #178) were more efficient than high CBD extract #131 in reducing the levels of IL-1β, IL-6 and TNF-α after LPS stimulation (Figure 2). In contrast, high CBD extract was better than high THC extracts in preventing the increase in IL-10 levels (Figure 2). The responses of IL-8 and MCP-1 were comparable in all extracts.
3. Materials and Methods
3.1. Main Reagents
The Δ9-THC (Cat#T4764) and CBD (Cat#C-045) were purchased from Sigma-Aldrich (Oakville, Canada). The 1 mg/mL stock solutions of cannabinoids were made by dissolving in methanol and then were kept at −20 °C.
Lipopolysaccharide (Cat#L4391) was purchased from Sigma. The sterile PBS was used for dissolving LPS to make 1 mg/mL stock solution. Phorbol-12-myristate-13-acetate (PMA) was purchased from Sigma-Aldrich (Cat#524400, Oakville, Canada). The DMSO (Dimethyl sulfoxide anhydrous, Life Technologies) was used to make 1 mg/mL stock solution. Then, the sterile PBS was used to make 5 μg/mL stock solution. Trypan Blue Solution (Cat#15250061), 0.4%, was purchased from ThermoFisher Scientific (Cat#15250061).
3.2. Plant Growth and Extract Preparation
All cannabis plants were grown at the University of Lethbridge in the licensed facility. Cannabis sativa cultivars #14 and #131 were used for the experiments. Per each cultivar, four plants were grown at 22 °C, 18 h light 6 h dark for the period of 4 weeks. For the flowering promotion, plants were grown in the chambers with 12 h light/12 h dark regime. Then, the flowers were harvested and dried. First, flowers from four plants were combined and after used for the extraction. Three grams of powdered plant tissue was used for the extraction per each studied cultivar. The prepared material was placed into a 250 mL Erlenmeyer flask, then 100 mL of ethyl acetate was added into each flask. Next, those flasks covered in tin foil were incubated overnight at 21 °C on a shaker at 120 rpm. The extracts were filtrated with subsequent concentration with the help of a rotary vacuum evaporator and transferred to a tared 3-dram vial. The solvent was eliminated by evaporating to dryness in an oven overnight at 50 °C. The DMSO was used for the dissolving of crude extracts to make 60 mg/mL concentration, and extracts were kept at −20 °C. The complete culture media was used to make the desired concentration of extracts (7 μg/mL). Next, the extracts were filtered with 0.22 μm filter [18].
3.3. High Performance Liquid Chromatography (HPLC) Analysis of Cannabinoids
Agilent Technologies 1200 Series HPLC system was used for analyzing the levels of THC and CBD in each extract. Software ChemStation LC 3D Rev B.04.02 (Agilent Technologies) was used for the data acquisition, control of the instrument, and integration. 9:1 methanol/chloroform (% v/v) was used as a mobile phase of the HPLC. Calibration of standards and sample analysis was done by using 2 μL of the injection volume. The detection of compound peaks was performed for 230 nm and 280 nm. Per each cannabis cultivar, two samples with two technical replicates per each treatment were analyzed [18]. LoQ (wt%): THC – 0.073, THCA – 0.06, CBD – 0.065, CBDA – 0.1.
3.4. Terpene Analysis
We used 2 g of flower samples. Terpene analysis was performed by Health Canada certified provider, Canvas Lab (Vancouver, BC, Canada) using 8610C GC coupled with a flame ionization detector (GC-FID, SRI Instruments). After calibration, the set of terpenes was used as standards (Restek Cat.# 34095). LoQs an LoDs were set at less than 0.01%. Diagrams with retention times are available upon request. Two samples per each cannabis cultivar were analyzed.
3.5. Cell Cultures and Treatments
The THP-1 cells were purchased from American Type Culture Collection (ATCC, Rockville, MD, USA) [25]. This suspension cell line was grown in 100 mm petri dish in the Roswell Park Memorial Institute Medium (RPMI-1640, Cat# 350-000-CL, WISENT INC., Quebec, Canada). The complete growth media was made by adding the 10% heat-inactivated Premium Grade Fetal Bovine Serum (Cat# 97068-085, VWR International LLC, Radnor, USA), according to the ATCC recommendation. The incubation environment for the cells was a humidified atmosphere of 5% CO2 at 37 °C. PMA (50 ng/mL, 48 h) was used for the terminal differentiation of monocytes into macrophages. Next, cells were given fresh media for one day. The verification of successful transformation was done by evaluating cell adhesion and spreading under the light microscope [26]. 0.5 μg/mL of LPS for 3 h was used for the inflammation induction [27]. Cannabis extracts (7 μg/mL) or single cannabinoids (5 μM) were added 20-30 min prior to LPS.
3.6. Cell viability assay using trypan blue
Trypan blue assay was used for the determination of THP-1 macrophages’ viability and morphology. First, cells were washed with sterile PBS, then trypsinized. The fresh media was added to THP-1 trypsinized macrophages. After that, the cells were centrifuged at 1,500 rpm for 5 min at 20 °C. The supernatant was discarded, and the pellet was resuspended by fresh media. The ratio of aliquot cells to trypan blue solution was 1:1. The automated cell counter, LUNA I (Logos Biosystems), was used for counting. The number of viable cells was determined by trypan blue exclusion. The results were shown as a percent of viability.
3.7. Multiplex ELISA
Multiplex ELISA was performed for cytokine assessment. Cell culture supernatants were centrifuged at 3,000 × g at 4 °C for 3 min prior to aliquoting to remove debris. The samples were stored at −70 °C. All prepared samples were shipped to Eve Technologies (Calgary, Alberta, Canada) on dry ice. Human Cytokine Array Proinflammatory Focused 13-plex (HDF13) discovery assay was used for measuring the levels of cytokines upon the first thaw. The results were analyzed using BioPlex 200 [28].
3.8. Statistical analysis
One way analysis of variance (ANOVA), followed by Tukey post-hoc multiple comparison test, was performed in GraphPad Prism 6.0 software (La Jolla, CA, USA) [29]. A p-value ˂ 0.05 was considered statistically significant.
4. Discussion
In this work, we have established an LPS-induced model of induction of inflammation in THP-1 macrophages. We found that pro-inflammatory cytokines are induced the most at 3 h post exposure to LPS. We then demonstrated that pre-treatment with CBD, THC or extracts high in one of these two cannabinoids substantially decrease the induction of cytokines post-LPS. CBD was more efficient than THC, and extracts were more efficient than single cannabinoids. IL-6, IL-8 and MCP-1 were most sensitive to pre-treatments with cannabinoids, while IL-1β, IL-10 and TNF-α were much less responsive.
Several other models of inflammation were used in the past, including U937 [30], and Mono Mac 6 [31] cell lines. U937 cells are monocytes derived from tissue origin of human histiocytic lymphoma. These cells can also be transformed into macrophage-like phenotype and be stimulated with LPS for inflammation induction [32,33]. The response of U937 cells to LPs was not as efficient as in our work; only 7 out of 34 inflammation-related genes were induced [34]. Stimulation of Mono Mac 6 cell line, a human monocytic line, with different triggers only upregulated TNF-α, IL-1α/β, and IL-6 [35]. Another type of cells, that can be used in inflammation is human peripheral blood mononuclear cell (PBMC) derived macrophages. These primary cells are considered superior to cell lines, as they more resemble in vivo settings. The main disadvantages of primary cells are their short lifespan and heterogenic response among donors which arises during cell differentiation from progenitors [36]. A comparison between the response of THP-1 cells and PBMC derived macrophages to LPS revealed a close correlation in inflammatory gene expression, making a THP-1 cell line a good model for studying LPS-induced changes [34].
Many studies have demonstrated the effects of single cannabinoids, such as THC and CBD, on inflammation [37]. Other components of the plant (such as minor cannabinoids, terpenes, terpenoids, flavonoids, and others) may act synergistically with cannabinoids and can be useful from a therapeutic point of view [19]. The modulating effect of these compounds is known as "an entourage effect"; such modulation is typically positive which means that the medicinal effect of the whole plant extract is more significant than the effect of isolated compounds [3,38].
IL-1β is the most prominent pro-inflammatory cytokine which plays a crucial role in inflammasome signaling [39]. IL-6, a pro-inflammatory cytokine, also plays an important role in the inflammatory response of many chronic inflammatory conditions [40]. TNF-α cytokine stimulates inflammation and is one of the most studied and most important pro-inflammatory cytokines [41]. IL-8 and MCP-1 induce chemotaxis of granulocytes to the site of infection and stimulate phagocytosis [42,43]. Our cannabis extracts significantly downregulated the levels of several of these cytokines, with IL-6, IL-8 and MCP-1 being affected the most. The results of this study were similar to other studies, where it was shown that cannabinoids and cannabis extracts efficiently inhibit inflammation by suppressing the levels of pro-inflammatory cytokines. For example, cannabis users that suffered from multiple sclerosis had significantly lower levels of many pro-inflammatory cytokines, like TNF-α, IL-1, IL-6, IL-12, IFN-γ, while higher levels of the anti-inflammatory cytokine, IL-10, in comparison to non-cannabis users [44]. Another study reported that cannabis extract high in CBD was superior to pure CBD in reducing the IL-6 and IL-8 levels effectively in an alveolar epithelial cell line, A549, while cannabis extract high in THC on the same cell line showed only minor anti-inflammatory activity and was more cytotoxic. This study was using comparable concentration of extracts - 5 µg/mL, while in our study, we used 7 µg/mL [20]. In a 3D EpiDermFT tissue inflammation model, it was shown that the extracts high in CBD and THC were effective in reducing the expression of IL-6, IL-1β, MCP-1, and TNF-α [18]. Another study demonstrated that cannabis extract high in CBD was more effective than pure CBD in zymosan-induced inflammation mice model. The extract effectively reduced pain and paw swelling, prevented TNF-α production, and overcame the bell-shaped dose response of CBD [19]. High CBD and high THC extracts were shown to be superior to the pure CBD and THC in reducing inflammation in graft versus host disease in allogeneic bone marrow transplantation model [21].
The data we obtained on IL-1β levels was quite interesting. This was the only cytokine that decreased in response to CBD more efficiently than in response to the high CBD extract or THC/high THC extracts. It remains to be shown whether this is a specific response to our extracts or the nature of the response of this cytokine to cannabinoids.
It was curious also to note that CBD and THC had no effect on MCP-1 levels while our extracts significantly downregulated high MCP-1 levels post-LPS. Our data and reports by others demonstrated that MCP-1 can indeed be downregulated by cannabis extracts [45]. MCP-1 was also downregulated in human Epithelial cells (BEAS-2B and NHBE), macrophages (U937), and lung fibroblast cells (HFL-1) exposed to LPS in response to CBD [46]. In contrast, exposure of THP-1 to LPs with subsequent treatment with increasing concentrations of CBD did not reduce the MCP-1 levels [47]. Therefore, it appears that decrease in MCP-1 expression in response to pure cannabinoids may be cell-specific.
Another interesting result was the effect of cannabinoids on IL-10 levels. IL-10 is an anti-inflammatory cytokine, which protects cells against profound inflammation. We observed that LPS induced this cytokine, and pure CBD as well as all extracts reduced it; in contrast, THC slightly increased it. Some studies demonstrated that cannabinoids and cannabis extracts enhance the levels of anti-inflammatory cytokines. For example, in the murine model of bone marrow transplantation, it was demonstrated that pure THC and CBD, as well as cannabis extracts high in CBD and THC, reduced inflammation, by reducing IL-17 secretion and enhancing IL-10 secretion [21]. Another study performed on mouse primary bone marrow-derived macrophages stimulated by LPS showed that flavonoids, luteolin and quercetin, increased the levels of IL-10 secretion [48]. It was also shown that THC reduced inflammation on endotoxemic mice model by significantly upregulating the plasma level of anti-inflammatory cytokine, IL-10, while suppressing the pro-inflammatory cytokine, MCP-1 [49]. Along this line, it was reported that CBD and THC suppressed the secretion of IL-17 but elevated the secretion of IL-10 by mouse-derived encephalitogenic T cell line [50]. CBD was reported to significantly reduce the plasma levels of pro-inflammatory cytokines (IFN-γ, TNF-α), along with the increase in the levels of anti-inflammatory cytokines (IL-10, IL-4) on diabetic mice model [51].
Since LPS was used to induce the inflammatory response, it was expected that it would also induce anti-inflammatory cytokines to counteract high levels of pro-inflammatory cytokines, which is required for cell survival. For effective clearance of pathogens, the pro-inflammatory response is crucial, while excessive inflammatory response causes tissue damage. Hence, human body always maintains equilibrium by activating counteractive pathways to bring down pro-inflammatory response. Since levels of pro-inflammatory cytokines were elevated by LPS in our study, we observed the corresponding increase in the IL-10 levels in LPS group to counteract the increase in pro-inflammatory cytokines.
We hypothesize that since our extracts and CBD were able to significantly downregulate the levels of several pro-inflammatory cytokines, they reduced the levels of IL-10 possibly to maintain the equilibrium. On the other hand, THC was not able to reduce the levels of pro-inflammatory cytokines as effectively as CBD and hence, did not change the high levels of IL-10 post-LPS. It is also possible that cannabis extracts inhibit all cytokines, regardless of their nature.
5. Conclusions and Limitations of the Study
We established the THP-1 macrophage model for the analysis of anti-inflammatory properties of cannabinoids and cannabis extracts. We found that studied cannabis extracts significantly prevented the increase in the levels of pro-inflammatory cytokines induced by LPS. It would be important in the future to treat cells with extracts after the induction of inflammation, since, in general, there is a need to treat inflammation rather than to prevent it. We may also want to compare the effect of extracts to well-known anti-inflammatory agents such as dexamethasone. For better understanding of our results, it would be very beneficial to look at key transcription factors and target genes by performing RNA sequencing, transcriptome profiling, followed by the analyses of relevant protein expression. The most important consideration in the future is to perform this study in vivo and to confirm the immunomodulatory activity of the extracts.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1. Analysis of inflammation in cell lysate and cell media using multiplex ELISA. Pilot experiment (n = 1) showing IL-1β, TNF-α and IL-10 cytokines levels by multiplex ELISA in cell media and cell lysate of LPS-treated (3 h) and control samples. Figure S2. Analysis of cell viability using trypan blue assay in THP-1 macrophages in response to CBD (A) and THC (B). Results are expressed as means ± SD of three independent experiments. Figure S3. Comparison of cytokine induction between different extracts. A – IL-1β; B – IL-6; C – TNF-α; D – IL-10; E – MCP-1; F – IL-8. Results are presented as mean of calculated concentration [pg/mL] ± SD. Statistical analysis was performed by ANOVA followed by Tukey post-hoc multiple comparison test. Significant differences between groups are marked with: *p < 0.05, **p < 0.01, ***p < 0.001, **** p < 0.0001.
Author Contributions
S.S., I.K. and O.K. developed the idea and study concept. M.Z. and S.S. designed the experiments. M.Z., N.P. and S.S. conducted the experiments. M.Z., S.S., I.K. and O.K. analyzed and interpreted data. M.Z. prepared tables and prepared a first draft of the manuscript. I.K., S.S. and O.K. edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by MITACS, grants IT11447, IT15089.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are available in the main text or the Supplementary Materials.
Acknowledgments
We thank Olena Shymanovska for the technical assistance.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Sample Availability
Not available.
References
- Cohen, K.; Weizman, A. and Weinstein, A. Positive and Negative Effects of Cannabis and Cannabinoids on Health. Clin Pharmacol Therapeutics 2019, 105(5), 1139–1147. [CrossRef]
- Cristino, L.; Bisogno, T. and Di Marzo, V. Cannabinoids and the expanded endocannabinoid system in neurological disorders. Nature Rev Neurology 2020, 16(1), 9–29. [CrossRef]
- Russo, E. B. The case for the entourage effect and conventional breeding of clinical cannabis: No ‘Strain,’ no gain”. Front. Plant Sci. 2019, 9, 1–8. [CrossRef]
- Chiurchiù, V. Endocannabinoids and Immunity. Cannabis Cannabinoid Res. 2016, 1(1), 59–66. [CrossRef]
- Galiègue, S.; Mary, S.; Marchand, J.; Dussossoy, D.; Carrière, D.; Carayon, P.; Bouaboula, M.; Shire, D.; Le Fur, G.; Casellas, P. Expression of central and peripheral cannabinoid receptors in human immune tissues and leukocyte subpopulations. Eur J Biochem. 1995, 232(1), 54-61. [CrossRef]
- Iwasaki, A.; Foxman, E. F.; Molony, R. D. Early local immune defences in the respiratory tract. Nature Rev. Immunology 2017, 17(1), 7–20. [CrossRef]
- Amarante-Mendes, G. P.; Adjemian, S.; Branco, L. M.; Zanetti, L. C.; Weinlich, R. and Bortoluci, K. R. Pattern recognition receptors and the host cell death molecular machinery. Front. Immunol. 2018, 9, 2379. [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9(6), 7204–7218. [CrossRef]
- Tisoncik, J. R.; Korth, M. J.; Simmons, C. P.; Farrar, J.; Martin, T. R.; Katze, M. G. Into the eye of the cytokine storm. Microbiol. Mol. Biol. Rev. 2012, 76(1), 16-32. [CrossRef]
- Deshmane, S. L.; Kremlev, S.; Amini, S. and Sawaya, B. E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interferon Cytokine Research 2009, 29(6), 313–325. [CrossRef]
- Benakanakere, M. R.; Finoti, L. S.; Tanaka, U.; Grant, G. R.; Scarel-Caminaga, R. M. and Kinane, D. F. Investigation of the functional role of human Interleukin-8 gene haplotypes by CRISPR/Cas9 mediated genome editing. Sci. Rep. 2016, 6(1), 1–11. [CrossRef]
- Zelová, H. and Hošek, J. TNF-α signalling and inflammation: Interactions between old acquaintances. Inflammation Research 2013, 62(7), 641–651. [CrossRef]
- Martin-Sanchez, F.; Diamond, C.; Zeitler, M.; Gomez, A.I.; Baroja-Mazo, A.; Bagnall, J.; Spiller, D.; White, M.; Daniels, M.J.D.; Mortellaro, A. et al. Inflammasome-dependent IL-1β release depends upon membrane permeabilisation. Cell Death Differ. 2016, 23(7), 1219–1231. [CrossRef]
- Tanaka, T.; Narazaki, M. and Kishimoto, T. Il-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6(10), 16295–16296. [CrossRef]
- Iyer, S. S. and Cheng, G. Role of interleukin 10 transcriptional regulation in inflammation and autoimmune disease. Crit. Rev. Immunol. 2012, 32(1), 23–63. [CrossRef]
- Nagarkatti, P.; Pandey, R.; Rieder, S. A.; Hegde, V. L. and Nagarkatti, M. Cannabinoids as novel anti-inflammatory drugs. Future Medicinal Chemistry 2009, 1(7), 1333–1349. [CrossRef]
- Mustafa, W.; Elgendy, N.; Salama, S.; Jawad, M. and Eltoukhy, K. The Effect of Cannabis on the Clinical and Cytokine Profiles in Patients with Multiple Sclerosis. Mult. Scler. Int. 2021, 2021, 1–10. [CrossRef]
- Kovalchuk, A.; Wang, B.; Li, D.; Rodriguez-Juarez, R.; Ilnytskyy, S.; Kovalchuk, I. and Kovalchuk, O. Fighting the Storm: Could Novel Anti-Tnfá And Anti-Il-6 C. Sativa Cultivars Tame Cytokine Storm in COVID-19? Aging (Albany. NY). 2021, 13(2), 1571–1590. [CrossRef]
- Gallily, R.; Yekhtin, Z. and L. Ondřej Hanuš, L. Overcoming the Bell-Shaped Dose-Response of Cannabidiol by Using Cannabis Extract Enriched in Cannabidiol. Pharmacol. Pharm. 2015, 6, 75–85. [CrossRef]
- Anil, S.M.; Shalev, N.; Vinayaka, A.C.; Nadarajan, S.; Namdar, D.; Belausov, E.; Shoval, I.; Mani, K.A.; Mechrez, G. and Koltai, H. Cannabis compounds exhibit anti-inflammatory activity in vitro in COVID-19-related inflammation in lung epithelial cells and pro-inflammatory activity in macrophages. Sci. Rep. 2021, 11(1), 1462. [CrossRef]
- Khuja, I.; Yekhtin, Z.; Or, R. and Almogi-Hazan, O. Cannabinoids Reduce Inflammation but Inhibit Lymphocyte Recovery in Murine Models of Bone Marrow Transplantation. Int. J. Mol. Sci. Artic. 2019, 20(3), 668. [CrossRef]
- Wang, B.; Li, D.; Fiselier, A.; Kovalchuk, I. and Kovalchuk, O. New AKT-dependent mechanisms of anti-COVID-19 action of high-CBD Cannabis sativa extracts. Cell Death Discov. 2022, 8, 110 (2022). [CrossRef]
- Wang, B.; Kovalchuk, A.; Li, D.; Rodriguez-Juarez, R.; Ilnytskyy, Y.; Kovalchuk, I. and Kovalchuk O. In search of preventive strategies: novel high-CBD Cannabis sativa extracts modulate ACE2 expression in COVID-19 gateway tissues. Aging (Albany NY). 2020, 12(22), 22425-22444. [CrossRef]
- Suryavanshi, S.V.; Zaiachuk, M.; Pryimak, N.; Kovalchuk, I.; Kovalchuk, O. Cannabinoids Alleviate the LPS-Induced Cytokine Storm via Attenuating NLRP3 Inflammasome Signaling and TYK2-Mediated STAT3 Signaling Pathways In Vitro. Cells 2022, 11, 1391. [CrossRef]
- Chanput, W.; Mes, J. J. and Wichers, H. J. THP-1 cell line: An in vitro cell model for immune modulation approach. International Immunopharmacology 2014, 23(1), 37–45. [CrossRef]
- Gatto, F.; Cagliani, R.; Catelani, T.; Guarnieri, D.; Moglianetti, M.; Pompa, P.P. and Bardi, G. PMA-induced THP-1 macrophage differentiation is not impaired by citrate-coated platinum nanoparticles. Nanomaterials, 2017, 7(10), 155. [CrossRef]
- Liu, X.; Yin, S.; Chen, Y.; Wu, Y.; Zheng, W.; Dong, H.; Bai, Y.; Qin, Y.; Li, J.; Feng, S. et al. LPS-induced proinflammatory cytokine expression in human airway epithelial cells and macrophages via NF-κB, STAT3 or AP-1 activation. Mol. Med. Rep. 2018, 17(4), 5484–5491. [CrossRef]
- Juknat, A.; Gao, F.; Coppola, G.; Vogel, Z. and Kozela, E. miRNA expression profiles and molecular networks in resting and LPS-activated BV-2 microglia—Effect of cannabinoids. PLoS One 2019, 14(2), p. e0212039. [CrossRef]
- Andrade, C. The P value and statistical significance: Misunderstandings, explanations, challenges, and alternatives. Indian J. Psycholog. Med. 2019, 41(3), 210–215. [CrossRef]
- Harikrishnan, H.; Jantan, I.; Haque, M. A. and E. Kumolosasi. Anti-inflammatory effects of Phyllanthus amarus Schum. & Thonn. through inhibition of NF-κB, MAPK, and PI3K-Akt signaling pathways in LPS-induced human macrophages. BMC Complement. Altern. Med. 2018, 18(1), 224. [CrossRef]
- Kong, Q.; Six, D.A.; Liu, Q.; Gu, L.; Wang, S.; Alamuri, P.; Raetz, C.R.; Curtiss, R. 3rd. Phosphate groups of lipid a are essential for Salmonella enterica serovar typhimurium virulence and affect innate and adaptive immunity. Infect. Immun. 2012, 80(9), 3215–3224, 2012. [CrossRef]
- Yao, W.; Yang, L.; Dai, F.; Tang, L. and Le, Y. Macrophage differentiation induced by PMA is mediated by activation of RhoA/ROCK signaling. J. Toxicol. Sci. 2017, 42(6), 763-771. [CrossRef]
- Vogel, C.F.; Garcia, J.; Wu, D.; Mitchell, D.C.; Zhang, Y.; Kado, N.Y.; Wong, P.; Trujillo, D.A.; Lollies, A.; Bennet, D. et al. Activation of inflammatory responses in human U937 macrophages by particulate matter collected from dairy farms: an in vitro expression analysis of pro-inflammatory markers. Environ Health. 2012, 11, 17. [CrossRef]
- Sharif, O.; Bolshakov, V.N.; Raines, S.; Newham, P. and Perkins, N.D. Transcriptional profiling of the LPS induced NF-κB response in macrophages. BMC Immunol. 2007, 8, 1. [CrossRef]
- Neustock, P.; Brand, J.M.; Kruse, A. and Kirchner, H. Cytokine production of the human monocytic cell line Mono Mac 6 in comparison to mature monocytes in peripheral blood mononuclear cells. Immunobiology, 1993, 188(3), 293–302. [CrossRef]
- Hume Underhill, D.A.; Wainwright, B.J.; Aderem, A.; Timothy Ravasi, S.; Wells, C. and Forest, A. Probability of Individual Inducible Genes from Gene-Autonomous Transcriptional System: Macrophage Heterogeneity Arises Generation of Diversity in the Innate Immune. J. Immunol. Ref. 2021, 168, 44–50. [CrossRef]
- Kovalchuk, O. and Kovalchuk, I. Cannabinoids as anticancer therapeutic agents. Cell Cycle, 2020, 19(9), 961–989. [CrossRef]
- Ferber, S.G.; Namdar, D.; Hen-Shoval, D.; Eger, G.; Koltai, H.; Shoval, G.; Shbiro, L.; Weller, A. The ‘Entourage Effect’: Terpenes Coupled with Cannabinoids for the Treatment of Mood Disorders and Anxiety Disorders. Curr. Neuropharmacol. 2019, 18(2), 87–96. [CrossRef]
- Martín-Sánchez, F.; Diamond, C.; Zeitler, M.; Gomez, A.I.; Baroja-Mazo, A.; Bagnall, J.; Spiller, D.; White, M.; Daniels, M.J.; Mortellaro, A. et al. Inflammasome-dependent IL-1β release depends upon membrane permeabilisation. Cell Death Differ. 2016, 23(7), 1219–1231. [CrossRef]
- Gabay, C. Interleukin-6 and chronic inflammation. Arthritis Res. Ther. 2006, 8(2), S3. [CrossRef]
- Zelová, H. and Hošek, J. TNF-α signalling and inflammation: Interactions between old acquaintances. Inflammation Research 2013, 62(7), 641–651. [CrossRef]
- Lopez-Castejon, G. and Brough, D. Understanding the mechanism of IL-1β secretion. Cytokine Growth Factor Reviews 2011, 22(4), 189–195. [CrossRef]
- Viedt, C.; Dechend, R.; Fei, J.; Hänsch, G.M.; Kreuzer, J.; Orth, S.R. MCP-1 Induces Inflammatory Activation of Human Tubular Epithelial Cells: Involvement of the Transcription Factors, Nuclear Factor-B and Activating Protein-1. J. Am. Soc. Nephrol. 2002, 13(6), 1534-47. [CrossRef]
- Mustafa, W.; Elgendy, N.; Salama, S.; Jawad, M. and Eltoukhy, K. The Effect of Cannabis on the Clinical and Cytokine Profiles in Patients with Multiple Sclerosis. Mult. Scler. Int., 2021, 2021, 1–10. [CrossRef]
- Aswad, M.; Hamza, H.; Pechkovsky, A.; Zikrach, A.; Popov, T.; Zohar, Y.; Shahar, E.; Louria-Hayon, I. High-CBD Extract (CBD-X) Downregulates Cytokine Storm Systemically and Locally in Inflamed Lungs. Front Immunol., 2022, 13, 875546. [CrossRef]
- Muthumalage, T. and Rahman, I. Cannabidiol differentially regulates basal and LPS-induced inflammatory responses in macrophages, lung epithelial cells, and fibroblasts. Toxicol. Appl. Pharmacol., 2019, 382, 114713. [CrossRef]
- Yeisley, D.J.; Arabiyat, A.S.; Hahn, M.S. Cannabidiol-Driven Alterations to Inflammatory Protein Landscape of Lipopolysaccharide-Activated Macrophages In Vitro May Be Mediated by Autophagy and Oxidative Stress. Cannabis Cannabinoid Res. 2021, 6(3), 253-263. [CrossRef]
- Comalada, M.; Ballester, I.; Bailón, E.; Sierra, S.; Xaus, J.; Gálvez, J.; de Medina, F.S.; Zarzuelo, A. Inhibition of pro-inflammatory markers in primary bone marrow-derived mouse macrophages by naturally occurring flavonoids: Analysis of the structure-activity relationship. Biochem Pharmacol. 2006, 72(8), 1010-21. [CrossRef]
- Joffre, J.; Yeh, C.C.; Wong, E.; Thete, M.; Xu, F.; Zlatanova, I.; Lloyd, E.; Kobzik, L.; Legrand, M.; Hellman, J. Activation of CB 1 R Promotes Lipopolysaccharide-Induced IL-10 Secretion by Monocytic Myeloid-Derived Suppressive Cells and Reduces Acute Inflammation and Organ Injury. J. Immunol. 2020, 204(12), 3339–3350. [CrossRef]
- Kozela, E.; Juknat, A.; Health, M.; Kaushansky, N. and Rimmerman, N. Cannabinoids Decrease the Th17 Inflammatory Autoimmune Phenotype Mechanisms of Anti-depressive Treatments View project Cell death induced by delta-aminolevulinic acid in astrocytes View project. Artic. J. Neuroimmune Pharmacol., 2013, 8(5), 1265-76. [CrossRef]
- Weiss, L.; Zeira, M.; Reich, S.; Har-Noy, M.; Mechoulam, R.; Slavin, S.; Gallily, R. Cannabidiol lowers incidence of diabetes in non-obese diabetic mice. Autoimmunity, 2006, 39(2), 143–151. [CrossRef]
Figure 1.
Changes in p-NFκB/NFκB ratio in response to LPS. A. Representative Western blots of p-NFκB and NFκB with GAPDH bands in response to LPS (at 3 h and 6 h) for four independent samples per each treatment are shown. B. Average ratio between p-NFκB and NFκB for control (vehicle), 3 h and 6 h post-treatment with LPS. Data are expressed as mean (n=4) ± SD. Significance of the differences (***, p < 0.001) was evaluated by one-way ANOVA followed by Tukey post-hoc multiple comparison test.
Figure 1.
Changes in p-NFκB/NFκB ratio in response to LPS. A. Representative Western blots of p-NFκB and NFκB with GAPDH bands in response to LPS (at 3 h and 6 h) for four independent samples per each treatment are shown. B. Average ratio between p-NFκB and NFκB for control (vehicle), 3 h and 6 h post-treatment with LPS. Data are expressed as mean (n=4) ± SD. Significance of the differences (***, p < 0.001) was evaluated by one-way ANOVA followed by Tukey post-hoc multiple comparison test.
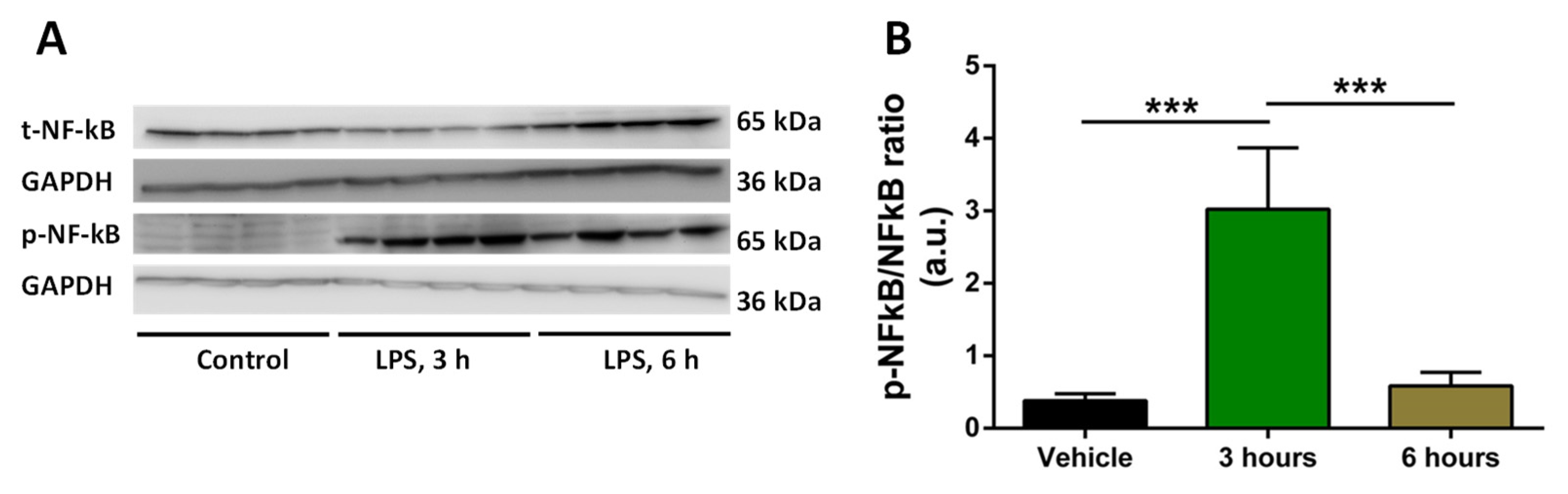
Figure 2.
Analysis of cytokine levels using ELISA in response to CBD, THC or extracts. A – IL-1β in CBD sample; B – IL-1β in THC sample; C – IL-6 in CBD sample; D – IL-6 in THC sample; E – IL-8 in CBD sample; F – IL-8 in THC sample; G – MCP-1 in CBD sample; H – MCP-1 in THC sample; I – TNF-α in CBD sample; J – TNF-α in THC sample; K – IL-10 in CBD sample; L – IL-10 in THC sample. Results are presented as mean of calculated concentration (pg/mL) ± SD of four independent experiments performed in duplicates and quantified by one-way ANOVA followed by Tukey post-hoc multiple comparison test. Significant differences between groups are marked with: *p < 0.05, **p < 0.01, ***p < 0.001, **** p < 0.0001.
Figure 2.
Analysis of cytokine levels using ELISA in response to CBD, THC or extracts. A – IL-1β in CBD sample; B – IL-1β in THC sample; C – IL-6 in CBD sample; D – IL-6 in THC sample; E – IL-8 in CBD sample; F – IL-8 in THC sample; G – MCP-1 in CBD sample; H – MCP-1 in THC sample; I – TNF-α in CBD sample; J – TNF-α in THC sample; K – IL-10 in CBD sample; L – IL-10 in THC sample. Results are presented as mean of calculated concentration (pg/mL) ± SD of four independent experiments performed in duplicates and quantified by one-way ANOVA followed by Tukey post-hoc multiple comparison test. Significant differences between groups are marked with: *p < 0.05, **p < 0.01, ***p < 0.001, **** p < 0.0001.
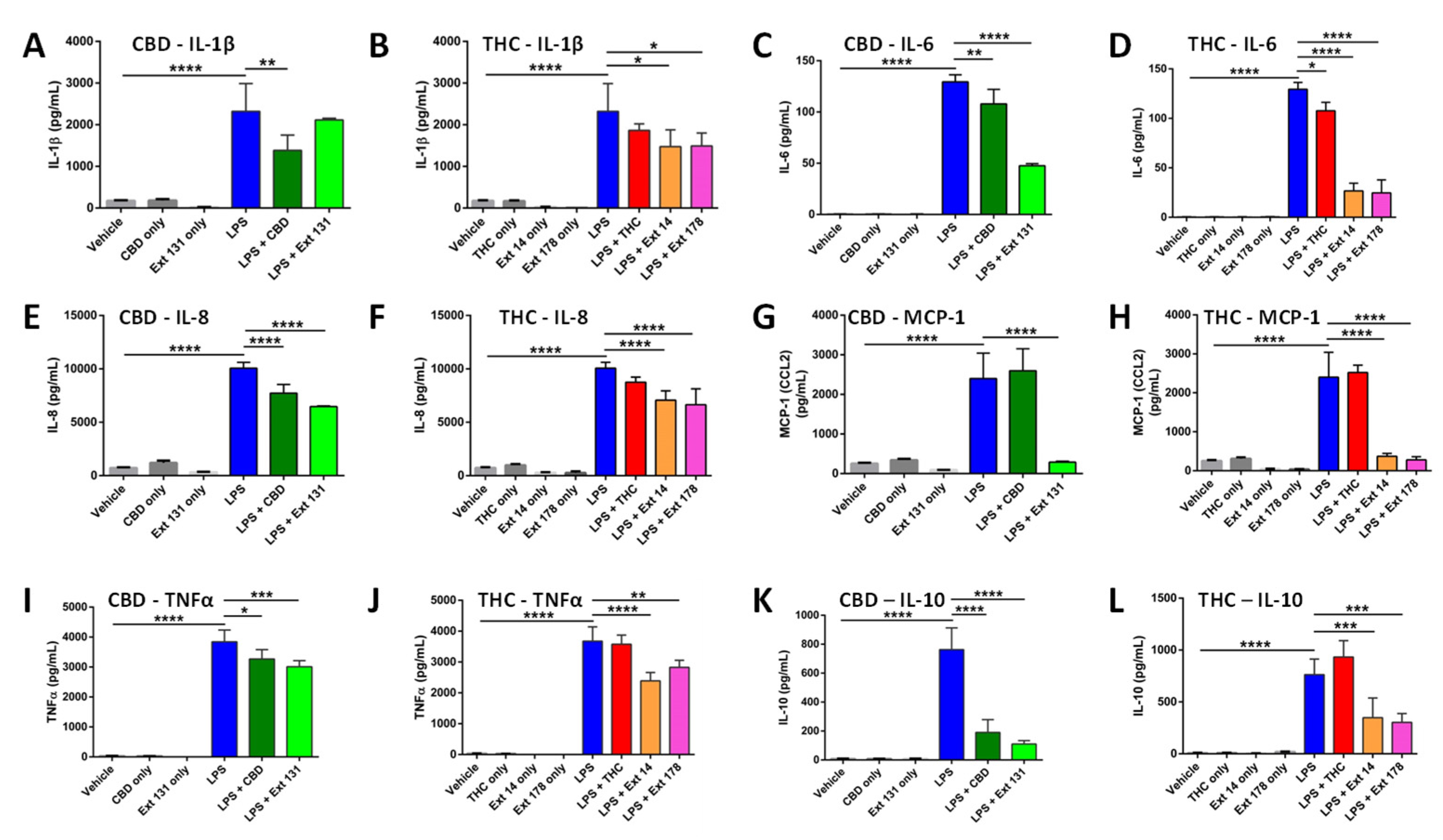

Table 1.
Concentrations of THC and CBD in extracts of selected cannabis cultivars.
| Total THC, % | Total CBD, % | THC (μM) | CBD (μM) | |
|---|---|---|---|---|
| #14 | 33.35±2.75 | 2.81±0.23 | 7.42±0.56 | 0.63±0.04 |
| #131 | 2.11±0.19 | 19.65±2.05 | 0.47±0.03 | 4.37±0.33 |
| #178 | 33.98±3.12 | 1.01±0.09 | 7.56±0.64 | 0.22±0.02 |
Concentrations of THC and CBD in the extracts #14, #131 and #178, in %, and concentration of THC and CBD in 7 µg/mL of the extract, in μM.
Table 2.
Composition of terpenes and their concentrations in each extract as analyzed by GC-FID.
| Terps in mg/g | #131 | #178 | #14 |
|---|---|---|---|
| α-Pinene | 0.295±0.055 | 0.649±0.12 | 0.048±0.008 |
| β-Pinene | 0.212±0.046 | 0.245±0.042 | 0.068±0.012 |
| β-Myrcene | ND | 0.361±0.064 | 0.124±0.042 |
| Limonene | 0.262±0.052 | 0.003±0.001 | 0.263±0.062 |
| Terpinolene | 0.025±0.005 | 0.008±0.002 | 0.004±0.001 |
| Linalool | 0.058±0.009 | 0.029±0.005 | 0.193±0.062 |
| α-Bisabolol | 0.003±0.001 | 0.244±0.06 | 0.061±0.02 |
| trans-Caryophyllene | 0.04±0.008 | 0.076±0.014 | 0.545±0.08 |
| α-Humulene | ND | 0.04±0.009 | 0.136±0.03 |
| trans-Nerolidol | ND | 0.008±0.002 | 0.187±0.04 |
| cis-Nerolidol | 0.001±0.001 | 0.003±0.001 | ND |
| Camphene | 0.022±0.006 | 0.025±0.001 | 0.015±0.005 |
| β-Ocimene | ND | 0.089±0.016 | ND |
| Fenchone isomers | ND | 0.003±0.001 | 0.004±0.001 |
| δ-3-Carene | 0.506±0.12 | 0.001±0.001 | ND |
| α-Terpinene | 0.006±0.002 | 0.573±0.11 | ND |
| Eucalyptol | 0.002±0.001 | ND | ND |
| γ-Terpinene | ND | 0.001±0.001 | ND |
| β-Cymene | ND | 0.048±0.009 | ND |
| Camphor isomers | ND | 0.118±0.04 | ND |
| isopulegol | 0.01±0.004 | 0.016±0.004 | ND |
| Caryophyllene oxide | 0.017±0.005 | 0.098±0.003 | ND |
| Valencene | ND | 0.01±0.003 | ND |
| Geraniol | 0.005±0.002 | 0.004±0.002 | ND |
| Guaiol | 0.029±0.005 | 0.116±0.04 | ND |
| trans-P-Ocimene | 0.016±0.004 | ND | ND |
| α-Humulene | 0.025±0.006 | ND | 0.136±0.034 |
| Fenchyl Alcohol | ND | ND | 0.036±0.007 |
| Borneol isomers | ND | ND | 0.012±0.003 |
| α -Terpineol | ND | ND | 0.052±0.009 |
| Total Terpene | 1.534± 0.32 | 2.768± 0.46 | 1.75± 0.25 |
ND – not determined.
Table 3.
Summary of the effects of extracts, CBD and THC on studied cytokines.
| Analyzed cytokines | CBD | THC | #131 | #14 | #178 |
|---|---|---|---|---|---|
| IL-1β | ↓ | = | = | ↓ | ↓ |
| IL-6 | ↓ | ↓ | ↓ | ↓ | ↓ |
| IL-8 | ↓ | = | ↓ | ↓ | ↓ |
| IL-10 | ↓ | = | ↓ | ↓ | ↓ |
| MCP-1 | = | = | ↓ | ↓ | ↓ |
| TNF-α | ↓ | = | ↓ | ↓ | ↓ |
↓ shows the decrease, while = shows no significant change.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



