
Submitted:
22 May 2023
Posted:
23 May 2023
You are already at the latest version
Abstract
Acinetobacter baumannii (AB) is a bacterium that causes infections, particularly in immunocompromised patients. Treatment is challenging due to biofilm formation by AB strains, which hinders antibiotic effec-tiveness and promotes drug resistance. The aim of our study was to analyze the biofilm-producing capacity of AB isolates from various forms of infections in relation to biofilm-related genes and their drug resistance. We tested one hundred isolates for biofilm formation using the crystal violet microplate method. Drug re-sistance analyses were performed based on EUCAST and CLSI guidelines, and biofilm genes were detected using PCR. All tested strains were found to form biofilms, with 50% being ICU strains and 72% classified as strong biofilm-producers. Among these, 87% were extensively drug-resistant (XDR) and 2% were extra extensively drug-resistant (E-XDR). The most common gene set was bap, bfmS, csuE, and ompA, found in 57% of all isolates. Our research has shown that, regardless of the form of infection, biofilm-forming strains can be expected among AB isolates. The emergence of E-XDR and XDR strains among non-ICU infections highlights the necessity for the rational use of antibiotics to stop or limit the further acquisition of drug re-sistance by A. baumannii.
Keywords:
Acinetobacter baumannii
; biofilm
; extensively drug resistance
; biofilm associated genes
1. Introduction
Acinetobacter baumannii (AB) is an opportunistic pathogen dominant in Central and Southern Europe in healthcare associated infections (HAIs). It is considered by the WHO as a critical-priority pathogen for which there is an urgent need to search for new therapeutic solutions, primarily due to acquired resistance mechanisms [1,2]. It causes a variety of infections as pneumonia in ventilated patients, bacteraemia or urinary tract infections [3,4].
According to European Antimicrobial Resistance Surveillance Network (EARS Net), the prevalence of Acinetobacter isolates resistant to at least one of the antimicrobial groups varied greatly according to country, ranging from 0% to 98.2%. In 2020, 3 countries (Ireland, the Netherlands and Norway) reported a prevalence of less than 1%, while in 21 European countries it was above 50% - the highest level of resistant isolates is recorded in Eastern and Southern Europe, including Poland [5].
In polish hospitals, especially in ICUs, the dominance of extensively drug-resistant (XDR) AB, defined as those strains that were susceptible to no more than two antimicrobial classes has been noted for many years and it is one of the major therapeutic problems associated to Gram-negative bacilli. The prevalence of AB isolates resistant to carbapenems reaches 80%, while 60% are resistant to carbapenems, fluoroquinolones and aminoglycosides together [5,6,7,8].
Multidrug resistance in Acinetobacter baumannii is associated with numerous mechanisms - enzymatic degradation, modification of antibiotics, reduction of membrane permeability and increased efflux. Carbapenem resistance of AB is conferred by carbapenem-hydrolyzing class D oxacillinases (OXA): OXA-23-like, OXA-24/40-like, OXA-58-like and intrinsic OXA-51-like. Permeability to beta-lactams, efflux pumps, and ISAba1 elements located upstream of the blaOXA-51-like gene also contributes to carbapenem resistance. Often the production of carbapenemases coexists with overexpression of efflux pumps [9].
The connection between biofilm and antibiotic resistance is of a considerable interest to biomedical researchers. The ability to form a biofilm, which is possessed by a large percentage of Acinetobacter baumannii strains (significantly higher than in the case of other Acinetobacter species [10]) - is considered to be one of the main factors of virulence, and also directly contributes to the antibiotic resistance of bacteria increasing tolerance to drugs and acting as a barrier against the penetration of antimicrobial agents or altering their metabolism and action [11].A. baumannii outside HAIs can also cause community-acquired infections, but still little is known about the main natural reservoirs of this pathogen [12]. Extra-hospital reservoirs of AB such as natural habitat, animals, food, high-touch surface in cities and the routes of transmission of this pathogen within community and between community and hospital environment are being investigated. The presence of AB has been demonstrated both in the urban environment on frequently touched surfaces as well as soil, water, plants or food of animal origin [13,14]. Among the strains isolated from food, biofilm forming strains and multidrug-resistant strains were also found, which may be a potential reservoir of new genes of carbapenemases carried on plasmids [13,15,16].
In the hospital environment biofilm formation promoting long-term persistence of AB on abiotic surfaces [17]. Under unfavorable environmental conditions, AB cells in the biofilm can become dormant and metabolically inactive, allowing it to survive environmental stress [18]. The ability to form a biofilm is facilitating colonization of patients and, consequently, infection. Infections connected with biofilm-forming AB strains associated with medical devices, primarily in central venous catheter-related bloodstream infections (CVC-BSI) have been confirmed [19]. Very often infections with biofilm-forming strains of AB are manifested by ventilator-associated (VAP). [20]. The ability to increase environment contamination, combined with the multidrug resistance of this microorganism, may lead not only to the development of infection, its severity, but also to clonal spread and result in outbreaks in hospital wards [21,22].
Biofilms as organized multicellular communities of bacteria are surrounded by self-produced exopolysaccharide matrices. Both the ability to form biofilms and genes involved in this process has been studied extensively in recent years. The formation and development of the biofilm involve many virulence factors such as the outer membrane protein A (OmpA), biofilm associated protein (Bap), chaperon-usher pilus (Csu), extracellular exopolysaccharide (EPS), and two-component regulatory system (BfmS/BfmR) [10,17,23]. Csu pili are adhesive organelles and are required to induce in the initial adhering of biofilm, promoting the maturation of biofilm and maintaining the structure of mature biofilm. BfmR/BfmS system coordinate the genes expression from Csu cluster [20]. The OmpA protein and the extracellular exopolysaccharide also act as adhesins. The Bap protein (Biofilm-Associated Protein Bap), in turn, is a key component of the mature biofilm and is involved in their various stages of formation.
The aim of our research was to investigate the biofilm-forming ability of Acinetobacter baumannii strains recovered from bloodstream infections, pneumonia, skin and soft tissue infections and colonizing patients in relation to presence of biofilm-related genes and antimicrobial resistance.
2. Materials and Methods
2.1. Bacterial isolates
The studied AB strains were isolated from clinical materials collected from patients hospitalized in 2019-2021 in hospitals of the Silesian Province in southern Poland (Katowice and Sosnowiec) and from materials collected as part of screening tests at the University Hospital in Krakow. The collection was stored in the laboratory of the Department of Microbiology at -70oC.
One hundred AB isolates were randomly selected applying the principle of one isolate from one patient, taking into account three clinical forms of infection such as bloodstream infection (BSI), pneumonia (PNEU), skin and soft tissue infections (SSTI) and colonization. Patients were hospitalized in ICU and non-ICU units (general surgery, orthopedics, neurology, internal medicine, palliative medicine). In this way, 25 isolates from BSI, 25 from PNEU, 25 from SSTI and 25 from patients without symptoms of infection (colonization) were collected.
The strains were collected in accordance with the consent of the bioethics committee of the Jagiellonian University KBET 1072.6120.274.2021 and KBET 1072.6120.2.2021.
Isolates were identified using automated systems (MALDI-TOFF identification; Maldi Biotyper, Bruker or MALDI TOF MS Vitek MS Home bioMérieux, depending on the hospital laboratory. In addition, the reference strain: Acinetobacter baumannii ATCC® 19606™, intensively producing biofilm, was used in the study as a positive control in the experiment on the production of biofilm by bacterial isolates [24].
2.2. Antimicrobial Susceptibility Testing
Antibiotic susceptibility was determined based on the results of an automated system MIDITECH Analyser v.12 for ampicillin/sulbactam; piperacilin/tazobactam; cefoperazone/sulbactam; imipenem; meropenem; ciprofloxacin; amikacin; gentamicin; tobramycin and trimethoprim/sulfamethoxazole. The results were interpreted using the clinical breakpoints defined in the latest EUCAST guidelines – v. 13.0 [25] For cefoperazone/sulbactam the interpretation was made based on the manufacturer's instructions for the ATCC reference strain and ampicilin/sulbactam and piperacilin/tazobactam based on CLSI (Clinical and Laboratory Standards Institute) guidelines [26]. For the levofloxacin, the Kirby-Bauer disc diffusion method on Mueller-Hinton agar plates was used and the results were interpreted according to EUCAST v. 13.0; . In addition, antibiotic resistance to tigecycline was determined for all strains using the MIC Test Strip (TGC 0.016-256 mg/L; Liofilchem Diagnostic; Italy), results were interpreted in accordance with EUCAST recommendations using MIC for not-species related breakpoints. The MIC for colistin was confirmed by the microdilution method (MIC STRIPPED PLATES COL; Diagnostics, Slovakia), the results were interpreted in line with the manufacturer's instructions according to EUCAST v. 13.0;. Based on the obtained results, the isolates were classified in terms of multidrug resistance as non-multidrug-resistant (nMDR), multidrug-resistant (MDR), extensively drug-resistant (XDR) [27] and E-XDR extra extensively drug-resistant. MDR strains were defined as those strains that were nonsusceptible to one antimicrobial in at least three different antimicrobial classes. XDR strains were defined as those strains that were susceptible to no more than two antimicrobial classes [23]. E-XDR strains was defined as strains resistant to all antibiotics tested in this work. Strains showing intermediate susceptibility to any of the antibiotics were interpreted as non-susceptible and counted in the group of resistant strains.
The pattern of antimicrobial resistance has been defined as the set of antibiotics to which at least two strains are resistant. For all strains The Multi-Antibiotic Resistant (MAR) index was calculated. MAR Index is the number of antibiotics that an isolate is resistant to divided by the total number of antibiotics utilized in the study [28].
2.3. Detection of carbapenemase genes
The Genomic Mini AX Bacteria Kit (A&A Biotechnology, Gdansk, Poland), was used to extract genomic DNA from AB isolates following the manufacturer's protocol. The concentration and purity of the isolated DNA was assessed using a NanoDrop Lite spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). DNA extracted from pure cultures was stored at -20°C for a further study.
The most common carbapenemase genes in AB in Poland were detected: blaOXA-23, blaOXA-40, blaOXA-58, blaNDM [7].Detection of carbapenemase genes was carried out according out following the protocol described by Cerezales et al. [29]. In the multiplex polymerase chain reaction (PCR) blaOXA-23 (718 bp), blaOXA-40 (413 bp), blaNDM (517 bp) , and blaOXA-58 (303 bp) genes were identified [Table 1]. PCR amplification was performed using the Color OptiTaq PCR Master Mix (EURx Ltd., Poland) in a final volume of 25 μl with a final primer concentration of 0.1 μM for each primer. Bacterial DNA functioned as the template. PCR was conducted with an initial denaturation step of 3 minutes at 94°C, followed by 30 cycles of 30 seconds at 94°C, 15 seconds at 58°C, and 1 minute at 72°C for amplification and a a final extension step of 5 minutes at 72°C. PCR products were analyzed by gel electrophoresis.
2.4. Assessment of biofilm formation
2.4.1. Quantitative of biofilm formation assessment
Quantification of biofilm formation was performed as previously described by Stepanovic [30] with some modifications. Contrary to the original work, plates with a smaller number of wells (24 wells) were used to increase the area of biofilm formation, and absorbance was measured by dry staining and fixation of the biofilm formed and measuring its thickness at 225 points of each well using a TECAN Infinite® 200 plate reader PRO.
Briefly, a bacterial suspension of approx. 0.5 MacFarland (1.5X108 CFU/ml) in saline was prepared from colonies on Tryptic Soy Agar (TSA) plates (TSA, Becton Dickinson, Franklin Lakes, NJ, USA). Twenty microliters of the prepared bacterial suspension were mixed with 1980 μL of Tryptic Soy Broth (TSB) (TSB, Becton Dickinson, Franklin Lakes, NJ, USA) and applied to 24-well flat-bottomed plates (Costar® Corning HTL SA, Warsaw, Poland) yielding a titre of approximately 1.5X106 CFU/ml.
The plate was incubated at 37°C without shaking for 20 hours. Liquid medium alone (TSB) was used as a negative control. ATCC 19606 strain was used as a positive control. After incubation, the medium with unbound cells was gently removed, then carefully rinsed three times with PBS and fixed with methanol for 30s. The plate was dried overnight at 37°C upside down, and then the biofilm was stained with crystal violet (1000 µl) for ~15 min at room temperature. After this time, the dye was poured off and rinsed with distilled water until the water in the wells was colorless. Again, it was dried overnight at 37°C upside down. After the plate was completely dried, the optical density absorbance was measured at 570nm on the surface of the biofilm formed at 225 points of each well. For each strain, two replicates were done to detect the biofilm formation ability.
Using the i-control software, the mean optical density (OD) and standard deviation (SD) values were calculated for each test isolate and all replicates. The cut-off point (ODc) was calculated using the following formula: ODc = mean OD of the negative control + (3 x standard deviation (SD) of the negative control). The averaged OD value of the tested isolates was reduced by the ODc value. The ODc value was calculated for each 12-well plate separately.
The strains were categorized according by Stepanović [30] into four categories: 0: non-biofilm producers (OD variable below cut-off), 1: Poor biofilm producers (OD variable ≤ 2 x cut-off), 2: Moderate biofilm producers (OD variable) from 2 x to 4 x cut-off), 3: Strong biofilm producers (>4 x cut-off).
2.4.2. Detection of biofilm-associated genes
Genomic DNA was extracted as described above.
Detection of biofilm-associated genes was was carried out in PCR. The genes bap, csuE, ompA, bfmS, espA were identified [Table 2]. PCR amplification was performed using Color OptiTaq PCR Master Mix (EURx Ltd., Poland) in a final volume of 25 μl with a final primer concentration of 0.1 μM for each primer. Bacterial DNA functioned as a template. For genes ompA, bfmS, and espA PCR was performed with an initial denaturation step of 5 minutes at 94°C, followed by 35 cycles of 1 minute at 94°C, 1 minute at 55°C and 45 seconds at 72°C for amplification and a final extension step of 5 minutes at 72°C. PCR products were analyzed by gel electrophoresis. For Bap and csuE PCR was performed with an initial denaturation step of 5 minutes at 96°C, followed by 35 cycles of 1 minute at 96°C, 1 minute at 56.5°C for bap and 57°C for csuE and 1 minute at 72°C for amplification and a final extension step of 10 minutes at 72°C. PCR products were analyzed by gel electrophoresis.
2.5. Statistical analysis
In statistical analyses, the determination of significant differences between the groups of isolates with low and moderate biofilm production and the group with intensive biofilm production and the clinical types of infection or colonization (BSI, PNEU, SSTI, colonization) was demonstrated in the cross-analysis in the Fisher-Freeman-Halton exact test with two-sided exact significance p ≤ 0.05.The differences between the prevalence of resistant isolates and the ability to form a biofilm were analysed based on the Pearson Chi-square test. The Fisher-Freeman-Halton test was used to show the variability between resistant strains isolated from different clinical forms of infection. The univariate analysis of variance (ANOVA) was used to find significance between association the different combination of the biofilm genes and the ODs and significance between the MDR or XDR and the type of the produced biofilm. The results were considered significant at p ≤ 0.05.
3. Results
3.1. Bacterial isolates
A total of 100 Acinetobacter baumannii isolates were tested following the principle of one strain per patient, including 25 from PNEU, 25 from SSTI, 25 from BSI and 25 from colonization. Fifty three percent of the strains came from ICU patients, including all colonization strains from ICU, 56% from PNEU infections and 44% from BSI infections. In contrast, only 12% of SSTIs were from ICU. Distribution of Acinetobacter baumannii in ICU and non-ICU isolates was showed in Table 3.
3.2. Antimicrobial Susceptibility Testing
The highest rate of resistance was for cefoperazone/sulbactam (95%) and the lowest was for colistin (8%). Strains resistant to both carbapenems (imipenem and meropenem) were nearly before 70%, and 45% were resistant to both carbapenems, fluoroquinolones and aminoglycosides tested. Significant difference was found for resistance to imipenem (p=0.023) and meropenem (p=0.023) between strains from the colonization and SSTI groups and gentamicin between the colonization and SSTI; BSI and PNEU groups (p=0.024, p=0.001, p=0.048 respectively), no significant differences were observed in case of other antimicrobials and the other groups of strains (Table 4).
The most common pattern of resistance among all AB strains was resistance to all tested antibiotics except colistin (37%; n=37) (Table 5). Among the tested isolates, 75% were classified as XDR, and 2% as E-XDR. While 9.4% of the ICU strains were not multidrug resistant, nearly 87% of the strains classified as to XDR and E-XDR (Table 6).
3.3. Selected carbapenemases genes
The most frequently detected gene among all AB strains was blaOXA-40 (42%); considering the clinical form of infection, it was detected in 72% of strains from SSTI and 56% of strains from colonization. In turn, in BSI strains, the blaOXA-23 gene was found more often (44%). Three strains isolated from patient colonization had the blaNDM gene (Table 7). None of the isolates carried the blaOXA-58 gene.
3.4. Quantitative biofilm formation assessment
Quantification of biofilm production showed that all tested isolates (n=100) produce biofilm: 3% were classified as weak producer of biofilm, 25% as moderate and 72% as strong (Figure 1). Among the ICU strains, 77% (n=41) were classified as strong biofilm producers, and 66% (n=31) from non-ICU (Table 8).
3.4. Correlation between the ability to biofilm formation and resistance among strains with different clinical forms of infection or colonization.
A group of strains characterized by weak and moderate biofilm production was combined for statistical analyses due to the small number of strains of the group of weak biofilm producers (n=3 and n=25).
Most of the strains strongly producing biofilm were in the group of colonization and PNEU. Significant differences (two-sided exact significance p<0.001) were found between the type of infection and the ability to form a biofilm among the isolates from the colonization group in relation to BSI and SSTI. No differences were found between isolates from different clinical forms of infection (Figure 2).
The correlation between biofilm formation and resistance to specific antibiotics was also analyzed. All colistin-resistant strains (n=8) were strong biofilm producers. A statistically significant difference (p=0.012) was found between the number of isolates resistant to gentamicin in the group of strong biofilm producers (47.2%; n=34) and in the group of weak and moderate biofilm producers (75%; n=21).
The distribution of resistance by resistance groups (nMDR, MDR, XDR and E-XDR) and groups of different biofilm producers (strong, moderate, weak) in different groups of origin of the strains (BSI, PNEU, SSTI, colonization) is shown in the radar chart (Figure 3). There was no significant correlation between the resistance group and the type of biofilm producer.
AB strains, regardless of the origin of the strains were characterized by a similar distribution of XDR resistance. Among SSTI isolates it was 68% (n=17), BSI 76% (n=19) and PNEU 76% (n=19); colonization 80% (n=20).
All weak biofilm-producing strains (n=3) were isolated from SSTI, while moderate biofilm-producing strains were present in all other groups, and most of them (56 %; n=14) were isolated from BSI.
3.5. Selected biofilm associated genes.
Among the tested isolates, the most frequently detected gene was ompA 99%, while the least frequently detected was epsA gene 26%.
The most common characteristic genotype was bap/bfmS/csuE/ompA, observed in 57% of isolates. Among them, 66.67% (n=38) belonged to the group of strong biofilm producers.
The set of all five tested genes bap/bfmS/csuE/ompA/epsA was less frequent (19%) but 78.9% (n=15) of isolates with this set were strong producers of biofilm. The third most common set of genes bfmS/csuE/ompA, present in 12% of isolates, concerned 74% (n=9) of strong biofilm producers.
In the group of the most characteristic genotype bap/bfmS/csuE/ompA, more than half of isolates were isolates from the colonization group (52.63%; n=20). The same genotype was the least frequent in the group of isolates from BSI (10.53%; n=4). Moreover, the same set of genes was found among all E-XDR strains (5.26%; n=2) and in 89.47% (n=34) XDR strains (Figure 4a and b).
No significant association was found between different combinations of biofilm genes and ODs biofilm in the biofilm assay for strong biofilm formers.
4. Discussion
In our study, the vast majority of A. baumannii strains (72%) strongly produced biofilm and were characterized by the presence of four genes associated with biofilm formation (bap, bfmS, csuE, ompA). Also, the most belonged to the XDR group (75%) and were resistant to imipenem and meropenem (69%), noting that only half of the isolates were from ICU. It is alarming to note 8 colistin-resistant strains that were isolated during screening tests from asymptomatic patients.
The WHO has listed carbapenem-resistant A. baumannii as a critical priority pathogen among those bacteria that require research and development of new drugs [1]. The situation in Europe for resistant ABs is not uniform as it is the pathogen with the greatest cross-country distribution. By far the highest prevalence of strains resistant to carbapenems as well as to three groups of drugs combined is found in southern and eastern Europe and it reaches over 90% in Greece, Romania and the Balkan countries (the current European average is 34.1% and it has increased by 1.8% compared to the average from 2016).
ECDC and WHO data for Poland indicate that the number of ABs resistant to fluoroquinolones, aminoglycosides and carbapenems together increased from 59.3% in 2016 to 64.2% (2020) [5]. AB also accounted for 55% of all bacterial isolates from ICU [5]. Our current study indicates that the situation seems to be worse than in our previous studies, where we recorded 80 and 86% of XDR, but nearly 80% of isolates came from ICU (now we tested only 53% ICU isolates) [7,35]. Other authors from Poland reported 76.5% XDR among ICU isolates [36], our current study shows 86.8% XDR (including E-XDR) while we consider only ICU strains. Additionally, we found two strains resistant to all antibiotics tested in this work (E-XDR). So far, no data have been published in Poland indicating the presence of A. baumannii isolates PDR or E-XDR. These strains originated from the colonization of patient but unfortunately, we do not have data on whether they later contributed to infection in these patients. Our strains were overwhelmingly sensitive to colistin 92%, but also to ampicillin-sulbactam 69%. We did not test new drugs such as cefiderocol and ervacycline, and we reported only 19% susceptibility to tigecycline In recent years, isolates resistant to all drugs (PDR) or to the vast majority of subjects, including colistin (E-XDR), have appeared mainly in Asian countries [37,38].
The predominant carbapenemase genes in studied AB strains were blaOXA-40 (42%) and blaOXA-23 (26%), which confirms our previous studies as well as other reports from Poland [7,39,40]. blaNDM gene was also detected in three strains from colonized patients. Resistance in AB results mainly from the production of carbapenem-hydrolyzing class D β-lactamases (CHLD) and also from non-enzymatic mechanisms of resistance, e.g., activity of efflux pumps. One family of efflux pumps is the RND-family. This efflux pump is also involved in biofilm formation and maturation. Yoon et al. [41] showed that in mutants in RND pump genes is significantly reduced biofilm formation compared to wild-type strains. This could explain the association between multidrug resistance and strong biofilm production. The antibiotic resistance of bacteria growing in the biofilm will be higher even when the strains growing in the form of planktonic cells do not have the acquired resistance mechanisms [23,42]. Kim et al. [43] indicates, however, that correlations between efflux pump genes and biofilm formation and resistance are not always clear-cut, in his studies increased efflux activity occurred among poor biofilm producers, although it also correlated with resistance to tigecycline and cefotaxim.
The vast majority of studied strains tested in this publication were biofilm-producing strains, including strongly producers which accounted for 72%. In studies, where like in ours, the association between biofilm formation and antimicrobial resistance was checked, regardless of the mechanism, a positive correlation was found much more often, strains strongly producing biofilm were characterized by higher resistance, primarily to antibiotics from the group of B-lactams and aminoglycosides, or the XDR phenotype in general [10,44,45,46,47].
In our earlier research [48], the vast majority of strains produced biofilm nearly 82%, but most were included in the moderate biofilm producers group. In this study, we observed that a large number of biofilm-producing strains were susceptible to amikacin or tobramycin and these were strains isolated from ICU patients [48]. These findings showed how important it is to take into account other factors, such as the types of hospital units, when describing the relationship between biofilm and resistance.
In our current study, most of the strains were classified as strong biofilm producers. We introduced - in relation to previous experiments - a modification of the study and assessment of biofilm intensity, namely we read the absorbance of crystal violet on the surface of the created biofilm instead of in the solution, and additionally the measurement was made at 225 points. This approach allows the classification of strains into a given group of biofilm producers with a high accuracy. We showed a significant relationship in strains isolated from patients without clinical symptoms of infection and strains isolated from patient with pneumonia and between AB strong biofilm producers. Most of these patients were hospitalized in ICUs. The vast majority of strains strongly producing biofilm were XDR type, including all of them resistant to colistin. The only antibiotic to which those isolates were more likely to produce less biofilm were resistant was gentamicin.
AB survival on often touched surfaces may have an impact on the spread of AB strains in hospital environment. It is extremely important to understand the impact of biofilm formation and antibiotic resistance on AB survival in the hospital environment. Greene et al. [18] indicate interesting differences between clinical and environmental strains, in the case of the latter, the ability to form a biofilm is critical for the survival of the strains, while in the case of clinical strains, the MDR phenotype is more important. This study demonstrates a trade-off between antibiotic resistance and desiccation tolerance in hospital strains [18]. In his research, Qi et al. showed that a strong ability to form biofilm can be a mechanism that allows bacteria to survive better, especially in the case of isolates with a sufficiently high level of resistance [49]. Our research, in turn, seems to confirm the thesis that the AB-HAIs in Polish ICUs are dominated by strains that are characterized by both high enzymatic drug resistance and high virulence (including biofilm formation). Ababneh et al. showed that in hospital environment frequently touched surfaces especially in ICUs for adults and children are contaminated with A. baumannii strains with the XDR phenotype. The source of these strains may be patients even with a distant history of infections with multidrug-resistant AB strains [50].
In Poland, for many years there have been difficulties in the eradication of multidrug-resistant AB strains, especially in ICUs [7]. It is of great concern that our tested AB isolates originating from patient colonization are highly biofilm-forming and multi-drug resistant; it may stem from the fact that these strains are likely to be present in the hospital environment for a long time and are difficult to eliminate and may contribute to later infections. Unfortunately, we had no information on possible AB infections or lack of them in these patients.
The presence of genes that are mainly associated with biofilm production was confirmed in both strong and moderate biofilm producers. The strong biofilm-producing A. baumannii represents 70 % of the most common set of genes (bap, bfmS, csuE and ompA), from which 87. 2% are XDR. Other studies report high frequency of csuE, bap and ompA genes [10,17,51]. The Csu and Bap systems significantly increase adherence to the cell line, and Bap is also involved in the formation and maintenance of mature biofilm. The least common gene in our research was epsA, which codes for extracellular exopolysaccharide, which is consistent with the Thummeepak et al. reports and inconsistent with the Zeighami et al. [10,17]. It is also believed that ompA and bap gene products may contribute to the drug-resistant AB phenotype, especially OmpA, an outer membrane porin.
The resistance of AB to most antibiotics and the fact that it persists the hospital environment for a long time causes a high risk of transmission of resistant (E-XDR and XDR) and highly biofilm-forming strains. The resistance of AB to most antibiotics and the fact that it persists in the hospital environment for a long time complicates the treatment of infections caused by biofilm-forming E-XDR and XDR AB [52,53]. These strains pose a serious threat to patients and a challenge for physicians in treatment [54,55]. In the used combination therapy, some combinations of drugs also showed significant inhibition of Acinetobacter biofilm, which may be an advantage of using the combination therapy. Such activities are demonstrated by, among others, imipenem-rifampicin, colistin-rifampicin, meropenem-sulbactam, tigecycline-sulbactam [56,57]. Biofilm inhibitors in combination with antibiotics such as zinc lactate or furanone with carbapenems, tigecycline or polymyxin B are also being tested. Such combinations work synergistically in vitro studies [58].
New strategies are also being sought to combat biofilm-forming and multidrug-resistant strains, such as new antibiotics e.g., synthetic lipopeptides [59], natural products, e.g., myrtenol, which also suppresses biofilm-forming genes [60], therapy with bacteriophages alone or in combination with an antibiotic [61,62]. High hopes are associated with cefiderocol, a new synthetic, siderophore cephalosporin. In a study by Kazmierczak et al., cefiderocol showed strong in vitro activity against most meropenem-resistant Enterobacteriaceae, P. aeruginosa and A. baumannii strains (96.7%, 100% and 96.9%, respectively, inhibited at cefiderocol MIC ≤4 µg/ml) [63]. Bassetii et al. [64] systematically reviewed the papers and concluded that cefiderocol is a promising and safe antibiotic option for the treatment of patients with carbapenem-resistant A. baumannii infections. Cefiderocol was approved for the treatment of infections caused by aerobic Gram-negative bacteria in adults with limited treatment options by the European Medicines Agency (EMA) in November 2019, and in Poland in March 2021 [65,66].
Regardless of the search for new solutions in the fight against multidrug-resistant A. baumannii, infection prevention and control is important in reducing of A. baumannii infections. These include hand hygiene, environmental cleaning, provision and appropriate use of personal protective equipment, appropriate training of healthcare staff, and promotion of antimicrobial stewardship programmes [5,6,67]. In the case of carbapenem-resistant AB strains, it is difficult to introduce surveillance of HAIs similar to the surveillance of carbapenem-resistant Enterobacterales (CRE), because there screening tests for Enterobacterales-specific carbapenemases are based on rapid cassette tests detecting KPC, NDM, VIM, IMP and OXA-48 carbapenemases. The variety of types of carbapenemases in AB means that there are no similar tests for AB.
Current research on biofilm-forming Acinetobacter baumannii, including ours, focuses mainly on the assessment of biofilm formation and its impact on bacterial resistance and survival in the environment, however, research on agents that destroy biofilm and interact with antibiotic therapy is also needed. Therefore, in our future studies, we plan to research among others, the influence of bacteriophages on biofilm.
Limitations
In the study, we did not detect the presence of efflux pumps from the RND family in the tested strains. Also, the effectiveness of cefiderocol on the tested strains has not been tested.
Author Contributions
TK-planned the experiments, performed all microbiological analyses, carried out the biofilm experiments, analysed data, performed the statistical analyses, prepared tables and figure, drafted the manuscript, DR- curated the strains collection, MPW and EJM – collected and identificated data, JWM-participated in the study design, participated in reviewed manuscript, ACH-participated in study design, drafted the manuscript, research supervision .
Funding
This research was funded by National Science Centre, grant number 2020/39/O/NZ7/00145 and by Jagiellonian University Collegium Medicum, grant number DBS/000698.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by Bioethics Committee of the Jagiellonian University (opinion number KBET 1072.6120.274.2021 and KBET 1072.6120.2.2021)
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- WHO Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed. Available online: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 7 February 2023).
- Nguyen, M.; Joshi, S.G. Carbapenem Resistance in Acinetobacter Baumannii, and Their Importance in Hospital-Acquired Infections: A Scientific Review. J. Appl. Microbiol. 2021, 131, 2715–2738. [Google Scholar] [CrossRef] [PubMed]
- Høiby, N.; Bjarnsholt, T.; Moser, C.; Bassi, G.L.; Coenye, T.; Donelli, G.; Hall-Stoodley, L.; Holá, V.; Imbert, C.; Kirketerp-Møller, K.; Lebeaux, D.; Oliver, A.; Ullmann, A.J.; Williams, C. ; ESCMID Study Group for Biofilms (ESGB); Consulting External Expert Werner Zimmerli ESCMID Guideline for the Diagnosis and Treatment of Biofilm Infections 2014. Clin. Microbiol. Infect. 2015, 21 Suppl 1, S1–S25. [Google Scholar] [CrossRef]
- Gedefie, A.; Demsis, W.; Ashagrie, M.; Kassa, Y.; Tesfaye, M.; Tilahun, M.; Bisetegn, H.; Sahle, Z. Acinetobacter Baumannii Biofilm Formation and Its Role in Disease Pathogenesis: A Review. Infect. Drug Resist. 2021, 14, 3711–3719. [Google Scholar] [CrossRef] [PubMed]
- Antimicrobial Resistance Surveillance in Europe 2022 - 2020 Data. Available online: https://www.ecdc.europa.eu/en/publications-data/antimicrobial-resistance-surveillance-europe-2022-2020-data (accessed on 7 February 2023).
- Antimicrobial Resistance in the EU/EEA (EARS-Net) - Annual Epidemiological Report for 2020. Available online: https://www.ecdc.europa.eu/en/publications-data/antimicrobial-resistance-eueea-ears-net-annual-epidemiological-report-2020 (accessed on 7 February 2023).
- Chmielarczyk, A.; Pilarczyk-Zurek, M.; Kamińska, W.; Pobiega, M.; Romaniszyn, D.; Ziółkowski, G.; Wójkowska-Mach, J.; Bulanda, M. Molecular Epidemiology and Drug Resistance of Acinetobacter Baumannii Isolated from Hospitals in Southern Poland: ICU as a Risk Factor for XDR Strains. Microb. Drug Resist. 2016, 22, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Wałaszek, M.; Różańska, A.; Bulanda, M.; Wojkowska-Mach, J. ; Polish Society of Hospital Infections Team Alarming Results of Nosocomial Bloodstream Infections Surveillance in Polish Intensive Care Units. Przegl. Epidemiol. 2018, 72, 33–44. [Google Scholar]
- Azimi, L.; Fallah, F.; Karimi, A.; Shirdoust, M.; Azimi, T.; Sedighi, I.; Rahbar, M.; Armin, S. Survey of Various Carbapenem-Resistant Mechanisms of Acinetobacter Baumannii and Pseudomonas Aeruginosa Isolated from Clinical Samples in Iran. Iran. J. Basic Med. Sci. 2020, 23, 1396–1400. [Google Scholar] [CrossRef]
- Zeighami, H.; Valadkhani, F.; Shapouri, R.; Samadi, E.; Haghi, F. Virulence Characteristics of Multidrug Resistant Biofilm Forming Acinetobacter Baumannii Isolated from Intensive Care Unit Patients. BMC Infect. Dis. 2019, 19. [Google Scholar] [CrossRef]
- Adams-Haduch, J.M.; Paterson, D.L.; Sidjabat, H.E.; Pasculle, A.W.; Potoski, B.A.; Muto, C.A.; Harrison, L.H.; Doi, Y. Genetic Basis of Multidrug Resistance in Acinetobacter Baumannii Clinical Isolates at a Tertiary Medical Center in Pennsylvania. Antimicrob. Agents Chemother. 2008, 52, 3837–3843. [Google Scholar] [CrossRef]
- Falagas, M.E.; Karveli, E.A.; Kelesidis, I.; Kelesidis, T. Community-Acquired Acinetobacter Infections. Eur. J. Clin. Microbiol. Infect. Dis. 2007, 26, 857–868. [Google Scholar] [CrossRef]
- Ababneh, Q.; Al-Rousan, E.; Jaradat, Z. Fresh Produce as a Potential Vehicle for Transmission of Acinetobacter Baumannii. Int. J. Food Contam. 2022, 9, 1–9. [Google Scholar] [CrossRef]
- Carvalheira, A.; Silva, J.; Teixeira, P. Lettuce and Fruits as a Source of Multidrug Resistant Acinetobacter Spp. Food Microbiol. 2017, 64, 119–125. [Google Scholar] [CrossRef]
- Rafei, R.; Hamze, M.; Pailhoriès, H.; Eveillard, M.; Marsollier, L.; Joly-Guillou, M.L.; Dabboussi, F.; Kempf, M. Extrahuman Epidemiology of Acinetobacter Baumannii in Lebanon. Appl. Environ. Microbiol. 2015, 81, 2359–2367. [Google Scholar] [CrossRef]
- Ababneh, Q.; Abu Laila, S.; Jaradat, Z. Prevalence, Genetic Diversity, Antibiotic Resistance and Biofilm Formation of Acinetobacter Baumannii Isolated from Urban Environments. J. Appl. Microbiol. 2022, 133, 3617–3633. [Google Scholar] [CrossRef]
- Thummeepak, R.; Kongthai, P.; Leungtongkam, U.; Sitthisak, S. Distribution of Virulence Genes Involved in Biofilm Formation in Multi-Drug Resistant Acinetobacter Baumannii Clinical Isolates. Int. Microbiol. 2016, 19, 121–129. [Google Scholar] [CrossRef]
- Greene, C.; Vadlamudi, G.; Newton, D.; Foxman, B.; Xi, C. The Influence of Biofilm Formation and Multidrug Resistance on Environmental Survival of Clinical and Environmental Isolates of Acinetobacter Baumannii. Am. J. Infect. Control 2016, 44, e65–e71. [Google Scholar] [CrossRef]
- Lin, M.-F.; Lan, C.-Y. Antimicrobial Resistance in Acinetobacter Baumannii: From Bench to Bedside. World J. Clin. Cases WJCC 2014, 2, 787. [Google Scholar] [CrossRef] [PubMed]
- Harding, C.M.; Hennon, S.W.; Feldman, M.F. Uncovering the Mechanisms of Acinetobacter Baumannii Virulence. Nat. Rev. Microbiol. 2018, 16, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Badave, G.K.; Dhananjay, K. Biofilm Producing Multidrug Resistant Acinetobacter Baumannii: An Emerging Challenge. J. Clin. Diagn. Res. 2015, 9, DC08. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.Y.; Baek, W.K.; Kim, H.A. Association of Biofilm Production with Colonization among Clinical Isolates of Acinetobacter Baumannii. Korean J. Intern. Med. 2017, 32, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Rumbo-Feal, S.; Gómez, M.J.; Gayoso, C.; Álvarez-Fraga, L.; Cabral, M.P.; Aransay, A.M.; Rodríguez-Ezpeleta, N.; Fullaondo, A.; Valle, J.; Tomás, M.; Bou, G.; Poza, M. Whole Transcriptome Analysis of Acinetobacter Baumannii Assessed by RNA-Sequencing Reveals Different MRNA Expression Profiles in Biofilm Compared to Planktonic Cells. PLoS One 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Szczypta, A.; Talaga-ćwiertnia, K.; Kielar, M.; Krzyściak, P.; Gajewska, A.; Szura, M.; Bulanda, M.; Chmielarczyk, A. Investigation of Acinetobacter Baumannii Activity in Vascular Surgery Units through Epidemiological Management Based on the Analysis of Antimicrobial Resistance, Biofilm Formation and Genotyping. Int. J. Environ. Res. Public Health 2021, 18, 1–15. [Google Scholar] [CrossRef]
- Eucast: The 13.0 Versions of Breakpoints, Dosing and QC (2023) Published. Available online: https://www.eucast.org/eucast_news/news_singleview?tx_ttnews%5Btt_news%5D=518&cHash=2509b0db92646dffba041406dcc9f20c (accessed on 23 April 2023).
- CLSI Publishes M100—Performance Standards for Antimicrobial Susceptibility Testing, 31st Edition. Available online: https://clsi.org/about/press-releases/clsi-publishes-m100-performance-standards-for-antimicrobial-susceptibility-testing-31st-edition/ (accessed on 7 February 2023).
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; Paterson, D.L.; Rice, L.B.; Stelling, J.; Struelens, M.J.; Vatopoulos, A.; Weber, J.T.; Monnet, D.L. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Mohamed, H.M.A.; Abd-Elhafeez, H.H.; Al-Jabr, O.A.; El-Zamkan, M.A. Characterization of Acinetobacter Baumannii Isolated from Raw Milk. Biology (Basel). 2022, 11, 1845. [Google Scholar] [CrossRef]
- Cerezales, M.; Biniossek, L.; Gerson, S.; Xanthopoulou, K.; Wille, J.; Wohlfarth, E.; Kaase, M.; Seifert, H.; Higgins, P.G. Novel Multiplex PCRs for Detection of the Most Prevalent Carbapenemase Genes in Gram-Negative Bacteria within Germany. J. Med. Microbiol. 2021, 70. [Google Scholar] [CrossRef] [PubMed]
- Stepanović, S.; Vuković, D.; Hola, V.; Di Bonaventura, G.; Djukić, S.; Ćirković, I.; Ruzicka, F. Quantification of Biofilm in Microtiter Plates: Overview of Testing Conditions and Practical Recommendations for Assessment of Biofilm Production by Staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Sharon Goh, H.M.; Beatson, S.A.; Totsika, M.; Moriel, D.G.; Phan, M.D.; Szubert, J.; Runnegar, N.; Sidjabat, H.E.; Paterson, D.L.; Nimmo, G.R.; Lipman, J.; Schembri, M.A. Molecular Analysis of the Acinetobacter Baumannii Biofilm-Associated Protein. Appl. Environ. Microbiol. 2013, 79, 6535. [Google Scholar] [CrossRef]
- Turton, J.F.; Gabriel, S.N.; Valderrey, C.; Kaufmann, M.E.; Pitt, T.L. Use of Sequence-Based Typing and Multiplex PCR to Identify Clonal Lineages of Outbreak Strains of Acinetobacter Baumannii. Clin. Microbiol. Infect. 2007, 13, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Tayabali, A.F.; Nguyen, K.C.; Shwed, P.S.; Crosthwait, J.; Coleman, G.; Seligy, V.L. Comparison of the Virulence Potential of Acinetobacter Strains from Clinical and Environmental Sources. PLoS One 2012, 7, e37024. [Google Scholar] [CrossRef] [PubMed]
- Liou, M.L.; Soo, P.C.; Ling, S.R.; Kuo, H.Y.; Tang, C.Y.; Chang, K.C. The Sensor Kinase BfmS Mediates Virulence in Acinetobacter Baumannii. J. Microbiol. Immunol. Infect. 2014, 47, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Chmielarczyk, A.; Pobiega, M.; Romaniszyn, D.; Wójkowska-Mach, J. Multi-Locus Sequence Typing (MLST) of Non-Fermentative Gram-Negative Bacilli Isolated from Bloodstream Infections in Southern Poland. Folia Microbiol. (Praha). 2018, 63, 191. [Google Scholar] [CrossRef]
- Ziółkowski, G.; Pawłowska, I.; Krawczyk, L.; Wojkowska-Mach, J. Antibiotic Consumption versus the Prevalence of Multidrug-Resistant Acinetobacter Baumannii and Clostridium Difficile Infections at an ICU from 2014-2015. J. Infect. Public Health 2018, 11, 626–630. [Google Scholar] [CrossRef]
- Inchai, J.; Liwsrisakun, C.; Theerakittikul, T.; Chaiwarith, R.; Khositsakulchai, W.; Pothirat, C. Risk Factors of Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Acinetobacter Baumannii Ventilator-Associated Pneumonia in a Medical Intensive Care Unit of University Hospital in Thailand. J. Infect. Chemother. 2015, 21, 570–574. [Google Scholar] [CrossRef]
- Kim, Y.J.; Yoon, J.H.; Kim, S.I.; Hong, K.W.; Kim, J.I.; Choi, J.Y.; Yoon, S.K.; You, Y.K.; Lee, M.D.; Moon, I.S.; Kim, D.G.; Kang, M.W. High Mortality Associated with Acinetobacter Species Infection in Liver Transplant Patients. Transplant. Proc. 2011, 43, 2397–2399. [Google Scholar] [CrossRef]
- Depka, D.; Mikucka, A.; Bogiel, T.; Rzepka, M.; Zawadka, P.; Gospodarek-Komkowska, E. Conventional and Real-Time PCR Targeting BlaOXA Genes as Reliable Methods for a Rapid Detection of Carbapenem-Resistant Acinetobacter Baumannii Clinical Strains. Antibiotics 2022, 11. [Google Scholar] [CrossRef]
- Słoczyńska, A.; Wand, M.E.; Tyski, S.; Laudy, A.E. Analysis of BlaCHDL Genes and Insertion Sequences Related to Carbapenem Resistance in Acinetobacter Baumannii Clinical Strains Isolated in Warsaw, Poland. Int. J. Mol. Sci. 2021, Vol. 22, Page 2486 2021, 22, 2486. [Google Scholar] [CrossRef]
- Yoon, E.J.; Balloy, V.; Fiette, L.; Chignard, M.; Courvalin, P.; Grillot-Courvalin, C. Contribution of the Ade Resistance-Nodulation-Cell Division-Type Efflux Pumps to Fitness and Pathogenesis of Acinetobacter Baumannii. MBio 2016, 7. [Google Scholar] [CrossRef]
- Alav, I.; Sutton, J.M.; Rahman, K.M. Role of Bacterial Efflux Pumps in Biofilm Formation. J. Antimicrob. Chemother. 2018, 73, 2003–2020. [Google Scholar] [CrossRef]
- Kim, C.M.; Park, G.; Ko, Y.J.; Kang, S.H.; Jang, S.J. Relationships between Relative Expression of RND Efflux Pump Genes, H33342 Efflux Activity, Biofilm-Forming Activity, and Antimicrobial Resistance in Acinetobacter Baumannii Clinical Isolates. Jpn. J. Infect. Dis. 2021, 74, 499–506. [Google Scholar] [CrossRef]
- Yang, C.H.; Su, P.W.; Moi, S.H.; Chuang, L.Y. Biofilm Formation in Acinetobacter Baumannii: Genotype-Phenotype Correlation. Molecules 2019, 24. [Google Scholar] [CrossRef]
- Ranjbar, R.; Farahani, A. Study of Genetic Diversity, Biofilm Formation, and Detection of Carbapenemase, MBL, ESBL, and Tetracycline Resistance Genes in Multidrug-Resistant Acinetobacter Baumannii Isolated from Burn Wound Infections in Iran. Antimicrob. Resist. Infect. Control 2019, 8. [Google Scholar] [CrossRef]
- Celik, B. Evaluation of the Correlation between Biofilm Formation and Drug Resistance in Clinical Isolates of Acinetobacter Baumannii. Int. J. Pathog. Res. 2020, 5, 16–27. [Google Scholar] [CrossRef]
- Asaad, A.M.; Soma, S.A.; Ajlan, E.; Awad, S.M. Epidemiology of Biofilm Producing Acinetobacter Baumannii Nosocomial Isolates from a Tertiary Care Hospital in Egypt: A Cross-Sectional Study. Infect. Drug Resist. 2021, 14, 709–717. [Google Scholar] [CrossRef]
- Krzyściak, P.; Chmielarczyk, A.; Pobiega, M.; Romaniszyn, D.; Wójkowska-Mach, J. Acinetobacter Baumannii Isolated from Hospital-Acquired Infection: Biofilm Production and Drug Susceptibility. APMIS 2017, 125, 1017–1026. [Google Scholar] [CrossRef]
- Qi, L.; Li, H.; Zhang, C.; Liang, B.; Li, J.; Wang, L.; Du, X.; Liu, X.; Qiu, S.; Song, H. Relationship between Antibiotic Resistance, Biofilm Formation, and Biofilm-Specific Resistance in Acinetobacter Baumannii. Front. Microbiol. 2016, 7, 483. [Google Scholar] [CrossRef]
- Ababneh, Q.; Abulaila, S.; Jaradat, Z. Isolation of Extensively Drug Resistant Acinetobacter Baumannii from Environmental Surfaces inside Intensive Care Units. Am. J. Infect. Control 2022, 50, 159–165. [Google Scholar] [CrossRef]
- Ghasemi, E.; Ghalavand, Z.; Goudarzi, H.; Yeganeh, F.; Hashemi, A.; Dabiri, H.; Mirsamadi, E.S.; Foroumand, M. Phenotypic and Genotypic Investigation of Biofilm Formation in Clinical and Environmental Isolates of Acinetobacter Baumannii. Arch. Clin. Infect. Dis. 2018 134 2018, 13, 12914. [Google Scholar] [CrossRef]
- Poulikakos, P.; Tansarli, G.S.; Falagas, M.E. Combination Antibiotic Treatment versus Monotherapy for Multidrug-Resistant, Extensively Drug-Resistant, and Pandrug-Resistant Acinetobacter Infections: A Systematic Review. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1675–1685. [Google Scholar] [CrossRef]
- Papathanakos, G.; Andrianopoulos, I.; Papathanasiou, A.; Koulenti, D.; Gartzonika, K.; Koulouras, V. Pandrug-Resistant Acinetobacter Baumannii Treatment: Still a Debatable Topic with No Definite Solutions. 2020, 75. [Google Scholar] [CrossRef]
- Karakonstantis, S.; Kritsotakis, E.I.; Gikas, A. Pandrug-Resistant Gram-Negative Bacteria: A Systematic Review of Current Epidemiology, Prognosis and Treatment Options. J. Antimicrob. Chemother. 2020, 75, 271–282. [Google Scholar] [CrossRef]
- Sobouti, B.; Mirshekar, M.; Fallah, S.; Tabaei, A.; Mehrabadi, J.F.; Darbandi, A. Pan Drug-Resistant Acinetobacter Baumannii Causing Nosocomial Infections among Burnt Children. Med. J. Islam. Repub. Iran 2020, 34, 24. [Google Scholar] [CrossRef]
- Wang, Y.C.; Kuo, S.C.; Yang, Y.S.; Lee, Y.T.; Chiu, C.H.; Chuang, M.F.; Lin, J.C.; Chang, F.Y.; Chen, T.L. Individual or Combined Effects of Meropenem, Imipenem, Sulbactam, Colistin, and Tigecycline on Biofilm-Embedded Acinetobacter Baumannii and Biofilm Architecture. Antimicrob. Agents Chemother. 2016, 60, 4670–4676. [Google Scholar] [CrossRef]
- Song, J.Y.; Cheong, H.J.; Noh, J.Y.; Kim, W.J. In Vitro Comparison of Anti-Biofilm Effects against Carbapenem-Resistant Acinetobacter Baumannii: Imipenem, Colistin, Tigecycline, Rifampicin and Combinations. Infect. Chemother. 2015, 47, 27. [Google Scholar] [CrossRef]
- Peng, Q.; Lin, F.; Ling, B. In Vitro Activity of Biofilm Inhibitors in Combination with Antibacterial Drugs against Extensively Drug-Resistant Acinetobacter Baumannii. Sci. Reports 2020 101 2020, 10, 1–8. [Google Scholar] [CrossRef]
- Roberts, K.D.; Zhu, Y.; Azad, M.A.K.; Han, M.L.; Wang, J.; Wang, L.; Yu, H.H.; Horne, A.S.; Pinson, J.A.; Rudd, D.; Voelcker, N.H.; Patil, N.A.; Zhao, J.; Jiang, X.; Lu, J.; Chen, K.; Lomovskaya, O.; Hecker, S.J.; Thompson, P.E.; et al. A Synthetic Lipopeptide Targeting Top-Priority Multidrug-Resistant Gram-Negative Pathogens. Nat. Commun. 2022, 13, 1625. [Google Scholar] [CrossRef]
- Selvaraj, A.; Valliammai, A.; Sivasankar, C.; Suba, M.; Sakthivel, G.; Pandian, S.K. Antibiofilm and Antivirulence Efficacy of Myrtenol Enhances the Antibiotic Susceptibility of Acinetobacter Baumannii. Sci. Reports 2020 101 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Vukotic, G.; Obradovic, M.; Novovic, K.; Di Luca, M.; Jovcic, B.; Fira, D.; Neve, H.; Kojic, M.; McAuliffe, O. Characterization, Antibiofilm, and Depolymerizing Activity of Two Phages Active on Carbapenem-Resistant Acinetobacter Baumannii. Front. Med. 2020, 7, 426. [Google Scholar] [CrossRef]
- Grygorcewicz, B.; Wojciuk, B.; Roszak, M.; Łubowska, N.; Blstrokaejczak, P.; Jursa-Kulesza, J.; Rakoczy, R.; Masiuk, H.; Dogowska, B. Environmental Phage-Based Cocktail and Antibiotic Combination Effects on Acinetobacter Baumannii Biofilm in a Human Urine Model. Microb. Drug Resist. 2021, 27, 25–35. [Google Scholar] [CrossRef]
- Kazmierczak, K.M.; Tsuji, M.; Wise, M.G.; Hackel, M.; Yamano, Y.; Echols, R.; Sahm, D.F. In Vitro Activity of Cefiderocol, a Siderophore Cephalosporin, against a Recent Collection of Clinically Relevant Carbapenem-Non-Susceptible Gram-Negative Bacilli, Including Serine Carbapenemase- and Metallo-β-Lactamase-Producing Isolates (SIDERO-WT-2014 Study). Int. J. Antimicrob. Agents 2019, 53, 177–184. [Google Scholar] [CrossRef]
- Bassetti, M.; Vena, A.; Castaldo, N.; Giacobbe, D.R.; Peghin, M.; Grossi, P.A. Clinical Evidence Supporting Cefiderol for Serious Acinetobacter Baumannii Infections. Curr. Opin. Infect. Dis. 2022, 35, 545–551. [Google Scholar] [CrossRef]
- European Medicines Agency Fetcroja | European Medicines Agency. Available online: https://www.ema.europa.eu/en/medicines/human/EPAR/fetcroja (accessed on 16 March 2023).
- RPL. Available online: https://rejestrymedyczne.ezdrowie.gov.pl/rpl/search/public (accessed on 16 March 2023).
- Kinross, P.; Gagliotti, C.; Merk, H.; Plachouras, D.; Monnet, D.L.; Högberg, L.D. ; EARS-Net Study Group; EARS-Net Study Group participants Large Increase in Bloodstream Infections with Carbapenem-Resistant Acinetobacter Species during the First 2 Years of the COVID-19 Pandemic, EU/EEA, 2020 and 2021. Euro Surveill. 2022, 27, 2200845. [Google Scholar] [CrossRef]
Figure 1.
Ability to form a biofilm among A. baumannii isolates. OD absorbance distribution and upper and lower quartiles.
Figure 1.
Ability to form a biofilm among A. baumannii isolates. OD absorbance distribution and upper and lower quartiles.
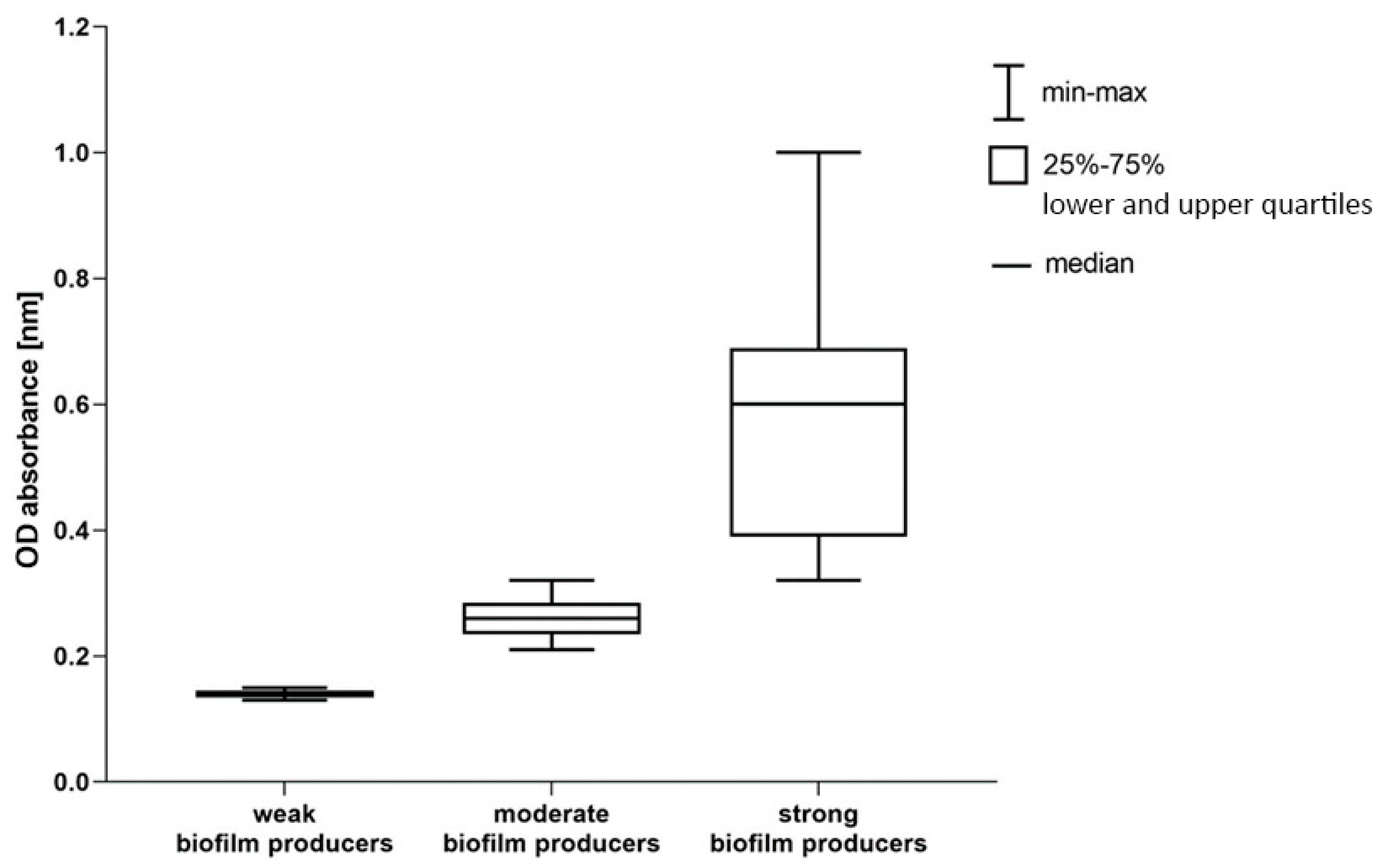
Figure 2.
Distribution of strong biofilm producing Acinetobacter baumannii isolates from various clinical forms of infection and colonization. Legend: BSI-bloodstream infection, PNEU-pneumonia, SSTI- skin and soft tissue infection, colonization; significance is marked with an asterisk * p<0.001.
Figure 2.
Distribution of strong biofilm producing Acinetobacter baumannii isolates from various clinical forms of infection and colonization. Legend: BSI-bloodstream infection, PNEU-pneumonia, SSTI- skin and soft tissue infection, colonization; significance is marked with an asterisk * p<0.001.
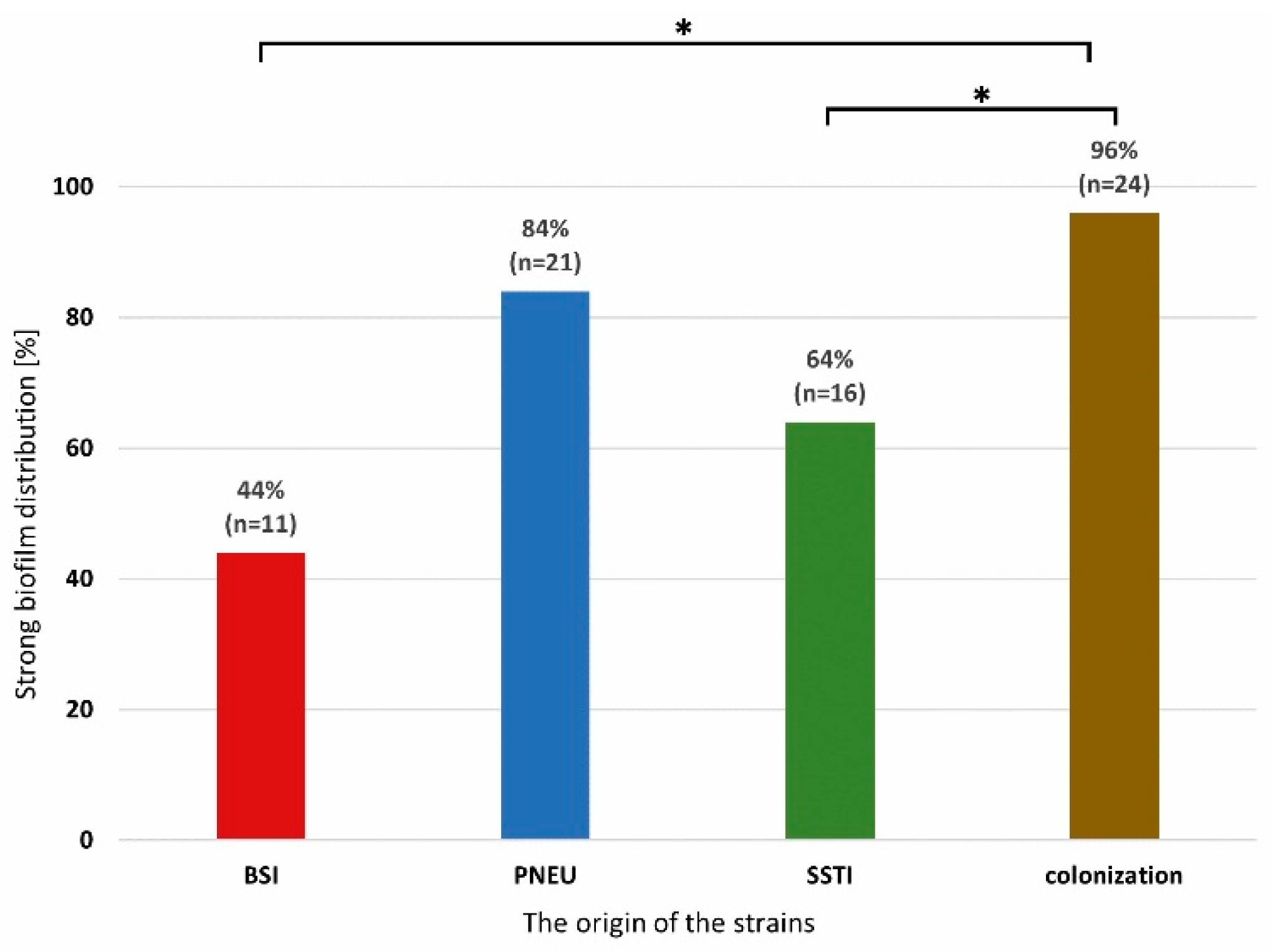
Figure 3.
Distribution of XDR, E-XDR and moderate and strong biofilm producing Acinetobacter baumannii among clinical forms of infection. Legend: BSI-bloodstream infection, PNEU-pneumonia, SSTI- skin and soft tissue infection, nMDR- no multidrug resistant, MDR-multidrug resistant, XDR-extensively drug resistant, E-XDR-extra extensively drug resistant.
Figure 3.
Distribution of XDR, E-XDR and moderate and strong biofilm producing Acinetobacter baumannii among clinical forms of infection. Legend: BSI-bloodstream infection, PNEU-pneumonia, SSTI- skin and soft tissue infection, nMDR- no multidrug resistant, MDR-multidrug resistant, XDR-extensively drug resistant, E-XDR-extra extensively drug resistant.
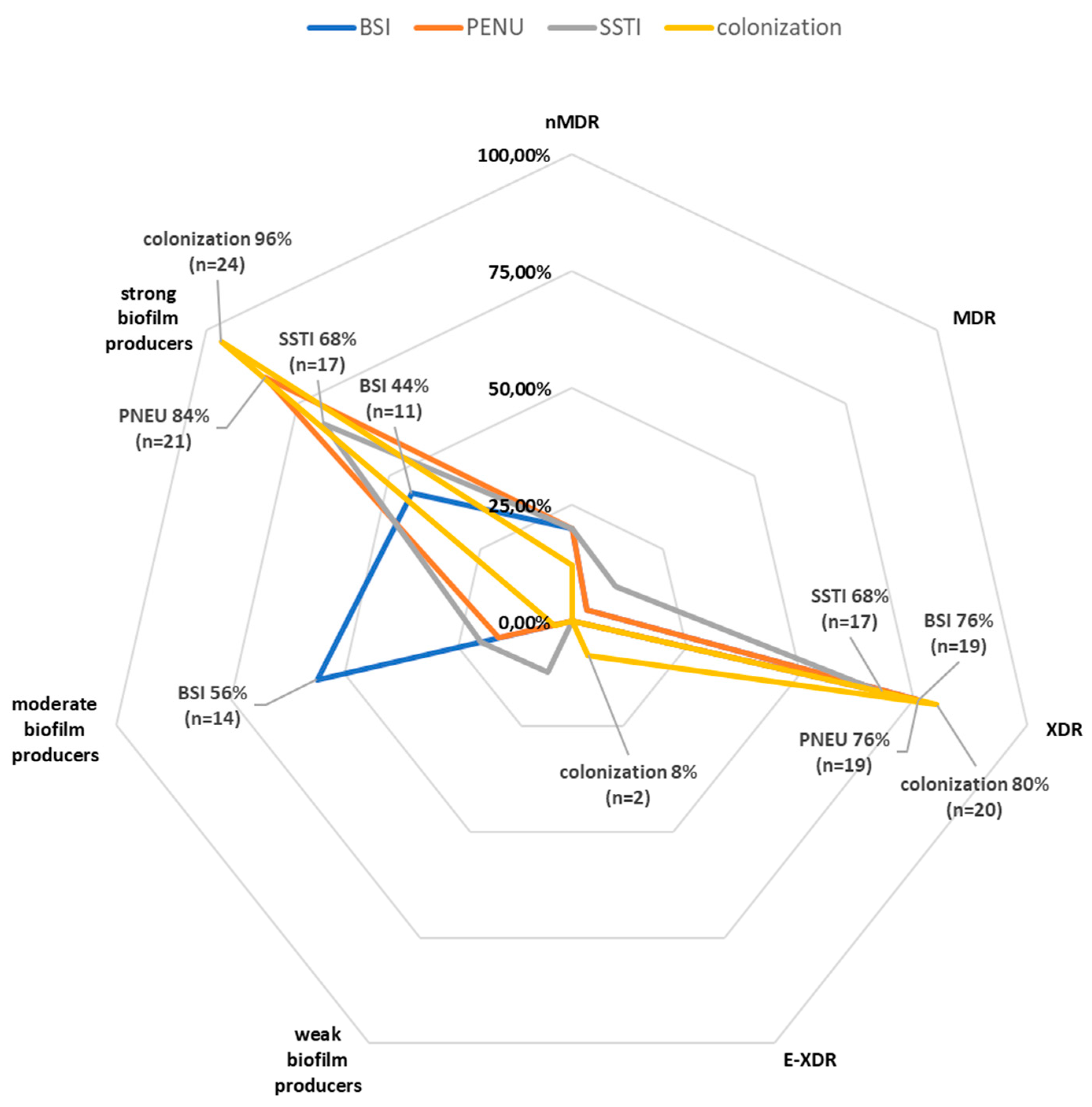
Figure 4.
Occurrence of set of genes associated with biofilm formation among isolates showing differences in biofilm production, resistance (b) and from different forms of clinical infection and colonization (a). Legend: BSI-bloodstream infection, PNEU-pneumonia, SSTI- skin and soft tissue infection, nMDR- no multidrug resistant, MDR-multidrug resistant, XDR-extensively drug resistant, E-XDR-extra extensively drug resistant.
Figure 4.
Occurrence of set of genes associated with biofilm formation among isolates showing differences in biofilm production, resistance (b) and from different forms of clinical infection and colonization (a). Legend: BSI-bloodstream infection, PNEU-pneumonia, SSTI- skin and soft tissue infection, nMDR- no multidrug resistant, MDR-multidrug resistant, XDR-extensively drug resistant, E-XDR-extra extensively drug resistant.
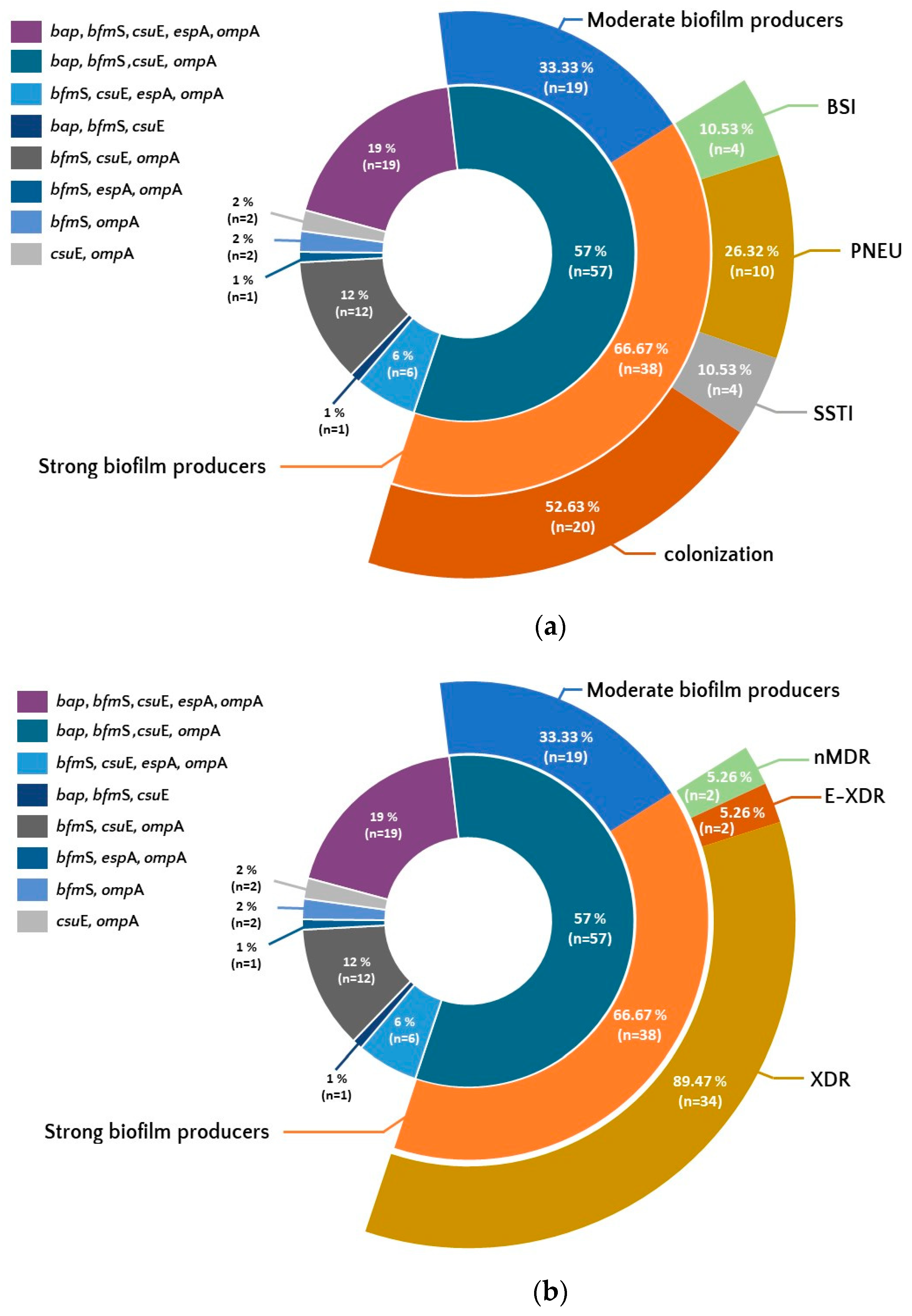

Table 1.
Primers used in detection of the carbapenemases genes in Acinetobacter baumannii.
| Detected genes | Primer sequences (5’-3’)1 | Product size (bp) | Annealing temperature | Reference |
|---|---|---|---|---|
| blaOXA-23 | F: TCTGGTTGTACGGTTCAGCA | 718 | 58 °C | [29] |
| R: GCATTTCTGACCGCATTTCC | ||||
| blaOXA-40 | F: GCATTGTCAGCAGTTCCAGT | 402 | 58 °C | [29] |
| R: AGAACCAGACATTCCTTCTTTCA | ||||
| blaNDM | F: GTTTGATCGTCAGGGATGGC | 517 | 58 °C | [29] |
| R: CTCATCACGATCATGCTGGC | ||||
| blaOXA-58 | F: ATCAAGAATTGGCACGTCGT | 303 | 58 °C | [29] |
| R: CCACATACCAACCCACTTGC |
Table 2.
Primers used in detection of genes associated with the biofilm formation in Acinetobacter baumannii.
Table 2.
Primers used in detection of genes associated with the biofilm formation in Acinetobacter baumannii.
| Detected genes | Primer sequences (5’-3’)1 | Product size (bp) | Annealing temperature | Reference |
|---|---|---|---|---|
| bap | F: TACTTCCAATCCAATGCTAGGGAGGGTACCAATGCAG | 1225 | 56.5 °C | [31] |
| R: TTATCCACTTCCAATGATCAGCAACCAAACCGCTAC | ||||
| csuE | F: ATGCATGTTCTCTGGACTGATGTTGAC | 976 | 57 °C | [32] |
| R: CGACTTGTACCGTGACCGTATCTTGATAAG | ||||
| ompA | F: CGCTTCTGCTGGTGCTGAAT | 531 | 55 °C | [33] |
| R: CGTGCAGTAGCGTTAGGGTA | ||||
| bfmS | F: TTGCTCGAACTTCCAATTTATTATAC | 1368 | 55 °C | [34] |
| R: TTATGCAGGTGCTTTTTTATTGGTC | ||||
| espA | F: AGCAAGTGGTTATCCAATCG | 451 | 55 °C | [33] |
| R: ACCAGACTCACCCATTACAT |
1 F – forward; R- reverse.
Table 3.
Distribution of Acinetobacter baumannii in ICU and non-ICU isolates.
| The origin of the strains |
ICU |
non-ICU |
||
|---|---|---|---|---|
| n (%) | % of total N=100 |
n (%) | % of total N=100 |
|
| BSI | 11 (20.7) | 44% | 14 (29.8) | 56% |
| PNEU | 14 (26.4) | 56% | 11 (23.4) | 44% |
| SSTI | 3 (5.7) | 12% | 22 (46.8) | 88% |
| colonization | 25 (47.2) | 100% | 0 (0) | 0% |
| Total | 53 (100) | 53% | 47 (100) | 47% |
Legend: ICU-intensive care unit, BSI-bloodstream infection, PNEU-pneumonia, SSTI- skin and soft tissue infection.
Table 4.
Distribution of resistance of Acinetobacter baumannii isolates according to clinical form of infections and colonization.
Table 4.
Distribution of resistance of Acinetobacter baumannii isolates according to clinical form of infections and colonization.
| Antibiotic Classes | Antimicrobial | Resistant Isolate; number (%) | ||||
|---|---|---|---|---|---|---|
| Total | The origin of the strains | |||||
| BSI | PNEU | SSTI | colonization | |||
| Penicillin | ampicillin/sulbactam piperacillin/tazobactam |
65 (65%) 86 (86%) |
16 (64%) 21 (84%) |
17 (68%) 21 (84%) |
12 (48%) 22 (88%) |
20 (80%) 22 (88%) |
| Cephalosporins | cefoperazone/sulbactam | 95 (95%) | 23 (92%) | 24 (96%) | 23 (92%) | 25 (100%) |
| Carbapenems | Imipenem* meropenem* |
69 (69%) 69 (69%) |
18 (72%) 18 (72%) |
17 (68%) 17 (68%) |
12 (48%) 12 (48%) |
22 (88%) 22 (88%) |
| Fluoroquinolones | ciprofloxacin levofloxacin |
87 (87%) 80 (80%) |
23 (92%) 19 (76%) |
23 (92%) 21 (82%) |
18 (68%) 18 (72%) |
22 (88%) 22 (88%) |
| Aminoglycosides | amikacin gentamycin** tobramycin |
69 (69%) 55 (55%) 70 (70%) |
19 (76%) 19 (76%) 17 (68%) |
18 (72%) 14 (56%) 19 (76%) |
14 (56%) 15 (60%) 16 (64%) |
18 (72%) 7 (28%) 18 (72%) |
| Tetracyclines | tigecycline | 71 (71%) | 18 (72%) | 19 (76%) | 15 (60%) | 19 (76%) |
| Miscellaneous agents | colistin trimethoprim/sulfamethoxazole |
8 (8%) 77 (77%) |
0 (0%) 20 (80%) |
0 (0%) 19 (76%) |
0 (0%) 16 (64%) |
8 (32%) 22 (88%) |
Legend: BSI-bloodstream infection, PNEU-pneumonia, SSTI- skin and soft tissue infection, *A significant difference (p=0.023) was found for resistance to imipenem and meropenem between strains from the colonization and SSTI (p=0.023) groups. ** A significant difference was found for resistance to gentamicin (p=0.007) between the colonization and SSTI (p=0.024), colonization and BSI (p=0.001) and colonization and PNEU (p=0.048) groups.
Table 5.
Antimicrobial resistance patterns of Acinetobacter baumannii isolates found in two or more strains.
Table 5.
Antimicrobial resistance patterns of Acinetobacter baumannii isolates found in two or more strains.
| Antibiotic Patterns* | No. of Isolates | MAR Index |
|---|---|---|
| SAM, TZP, SCF, IMP, MEM, CIP, LEV, AMI, GEN, TN, TIG, SXT, | 37 | 0,92 |
| SAM, TZP, SCF, IMP, MEM, CIP, LEV, AMI, TN, TIG, SXT, | 10 | 0,85 |
| TZP, SCF, IMP, MEM, CIP, LEV, AMI, GEN, TN, TIG, SXT, | 5 | 0,85 |
| SCF, | 4 | 0,08 |
| SCF, CIP, | 4 | 0,15 |
| SAM, TZP, SCF, IMP, MEM, CIP, LEV, AMI, TN, TIG, SXT, CL | 3 | 0,92 |
| CIP, | 2 | 0,08 |
| SAM, TZP, SCF, CIP, LEV, AMI, TN, TIG, SXT, | 2 | 0,69 |
| SAM, TZP, SCF, CIP, LEV, GEN, TIG, SXT, | 2 | 0,62 |
| SAM, TZP, SCF, IMP, MEM, CIP, LEV, AMI, GEN, TN, TIG, SXT, CL | 2 | 1,00 |
| SAM, TZP, SCF, IMP, MEM, CIP, LEV, AMI, TN, SXT, | 2 | 0,77 |
| SAM, TZP, SCF, IMP, MEM, CIP, LEV, TIG, SXT, CL | 2 | 0,77 |
| TZP, | 2 | 0,08 |
| TZP, SCF, | 2 | 0,15 |
Legend: SAM—ampililin/sulbactam; TZP—piperacilin/tazobactam; SCF—cefoperazone/sulbactam; IMP—imipenem; MEM—meropenem; CIP—ciprofloxacin; LEV—levofloxacin; AMI—amikacin; GEN—gentamicin; TN—tobramycin; TIG—tigecycline; SXT- trimethoprim/sulfamethoxazole; CL—colistin.
Table 6.
Antimicrobial resistance groups of Acinetobacter baumannii in ICU and non- ICU isolates.
| Group of resistance | ICU | non-ICU | ||
|---|---|---|---|---|
| n (%) | % of total N=100 |
n (%) | % of total N=100 |
|
| nMDR | 5 (9.4) | 27.8% | 13 (27.6) | 72.2% |
| MDR | 2 (3.8) | 40% | 3 (6.4) | 60% |
| XDR | 44 (83) | 58.7% | 31 (66) | 42.3% |
| E-XDR | 2 (3.8) | 100% | 0 (0) | 0% |
| Total | 53 (100) | 53% | 47 (100) | 47% |
Table 7.
Presence of selected carbapenemase genes among the Acinetobacter baumannii isolates according to type of unit, clinical form of infection and group of resistance.
Table 7.
Presence of selected carbapenemase genes among the Acinetobacter baumannii isolates according to type of unit, clinical form of infection and group of resistance.
|
blaOXA-23 N (%) |
blaOXA-40 N (%) |
blaNDM N (%) |
None of the tested genes N (%) |
|
|---|---|---|---|---|
| Type of unit | ||||
| Non-ICU | 13 (27.6) | 23 (48,9) | 0 | 13 (27.6) |
| ICU | 13 (24.5) | 19 (35.8) | 3 (5.7) | 19 (35.8) |
| The origin of the strains | ||||
| BSI | 11 (44) | 5 (20) | 0 | 9 (36) |
| PNEU | 7 (28) | 5 (20) | 0 | 13 (52) |
| SSTI | 3 (12) | 18 (72) | 0 | 4 (16) |
| colonization | 5 (20) | 14 (56) | 3 (12) | 6 (24) |
| Group of resistance | ||||
| nMDR | 2 (11.1) | 7 (38.9) | 0 | 9 (50) |
| MDR | 0 | 2 (40) | 0 | 3 (60) |
| XDR | 24 (32) | 31 (41.3) | 3 (4) | 20 (26.7) |
| E-XDR | 0 | 2 (100) | 0 | 0 |
| Both carbanenems resistant strains | 21 (30.4) | 31 (44.9) | 3 (4.3) | 7 (10.1) |
| Total | 26 (26) | 42 (42) | 3 (3) | 32 (32) |
Legend: ICU-intensive care unit, BSI-bloodstream infection, PNEU-pneumonia, SSTI- skin and soft tissue infection, nMDR- no multidrug resistant, MDR-multidrug resistant, XDR-extensively drug resistant, E-XDR-extra extensively drug resistant.
Table 8.
Biofilm production Acinetobacter baumannii in ICU and non- ICU isolates.
| Group of biofilm producers | ICU | non-ICU | ||
|---|---|---|---|---|
| n(%) | % of total N=100 |
n (%) | % of total N=100 |
|
| Weak+moderate biofilm producers | 12 (22.6) | 42.8% | 16 (34) | 57.2% |
| Strong biofilm producers | 41 (77) | 57% | 31 (66) | 43% |
| Total | 53 (100) | 53% | 47 (100) | 47% |
Legend: ICU-intensive care unit, nMDR- no multidrug resistant, MDR-multidrug resistant, XDR-extensively drug resistant, E-XDR-extra extensively drug resistant; weak biofilm producers (n=3) and moderate biofilm producer (n=25).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



