
Submitted:
20 May 2023
Posted:
23 May 2023
You are already at the latest version
Abstract
The importance of PAs for CNS functions are well known. Less clear is, however, where PAs in the brain are derived from. Principally, there are three possibilities: (i) intake by nutrition, release into the bloodstream, and subsequent uptake from CNS capillaries, (ii) products of parenchymatous organs as the liver, and again uptake from CNS capillaries, and (iii) uptake of precursors such as arginine from the blood and subsequent local biosynthesis of polyamines within the CNS.The present investigation aimed to unequivocally answer the question, whether PAs, especially the higher ones like SPD and SPM can or cannot be taken up into the brain from the bloodstream. For this purpose a biotin-labelled analogue of SPM was synthesized, characterized, and used to visualize its uptake into brain cells subsequent to application to surviving brain slices, to the intraventricular space, or to the bloodstream.In surviving slices uptake of B-X-SPM is massive into protoplasmic and absent in fibrous type astrocytes. It is also taken up by neurons but to a lesser degree. Under in-vivo conditions astrocyte uptake of biotinylated SPM also is massive from the brain interstitial fluid subsequent to intraventricular application. In contrast, following intracardial injection there is no uptake from the bloodstream, indicating that the brain is completely dependent on the local synthesis of polyamines.
Keywords:
Polyamines
; blood brain barrier
; local biosynthesis
; metabolism
; protoplasmic astrocytes
; fibrous astrocytes
Introduction
Polyamines (PAs) may be derived from alimentary sources or by local biosynthesis [1]. In the mammalian gut lumen, the predominant share of PAs stems from food intake, while a variable quantity may be produced by large intestine microbiota. PAs in the gut lumen are mainly absorbed by the duodenal and jejunal mucosa [2,3] and subsequently transferred [4] into the bloodstream ([1], Figure 1). PAs in the gut lumen may reach almost millimolar concentrations after a meal and disappear rapid and completely [3], whereby the luminal PA content returns to the fasting level in about 120 minutes [5]. Plasma levels of SPD and SPM show only mild (up to 20 µM) increases after a meal [5]. Most likely PA uptake in peripheral tissues keeps the plasma concentrations low ([1], Table 2).
While the importance of the biological functions of PAs in the CNS is well known [6], it is not clear, whether the brain retrieves its PAs from the bloodstream or whether it solely depends on local synthesis. While a transport system for PUT had been demonstrated in brain slices [7], it was hardly detectable in vivo [8]. Subsequent data indicated that the brain is unable to take up PAs from the bloodstream [9].
The present investigation now aimed to unequivocally answer the question, whether PAs, especially the higher ones like SPD and SPM, can or cannot be taken up from the bloodstream into the brain. For this purpose we synthesized a biotin-labelled analogue of SPM and used it as a tool to visualize this PA analogue in brain sections subsequent to its application to surviving brain slices, to the bloodstream, or to the intraventricular space.
Materials and Methods
Materials
All chemicals and other materials were obtained from Sigma-Aldrich Chemie GmbH (82024 Taufkirchen, Germany) unless indicated otherwise.
Synthesis of B-X-SPM
Biotinylated spacer-extended spermine (B-X-SPM) was synthesized following established methods to conjugate N-hydroxy-succinimide esters of haptens with amino group containing molecules [10,11]. In short, 17.4 mg of spermine tetrahydrochloride and 27 μl triethanolamine (to deprotonize SPM) were dissolved in 1 ml of dimethylformamide (50 mM SPM, solution A). Furthermore, 6.8 mg of 6-biotinamido-hexanoic acid N-hydroxysuccinimide ester (Molecular Probes) were dissolved in another 1 ml DMFA (20 mM; solution B). For conjugation, 100 μl of solution A was combined with 150 μl of DMFA and supplemented with 250 μl of solution B. After 2 hours at room temperature the reaction was stopped by the addition of 4.5 ml of pure ethanol, resulting in a 1 mM final concentration of biotinylated spermine.
Characterization of B-X-SPM
Analysis and characterization of the reaction products were analyzed commercially (WITA GmbH, Berlin, Germany). Components of the reaction the mixture were separated using reverse phase high pressure liquid chromatography, yielding two major peaks. The structure of the compound producing the first peak turned out to be N1-X-biotinyl-SPM (Figure 1) as confirmed by mass spectroscopy. The second peak most likely corresponds to N1,N12-doubly biotinylated SPM as the presence of acylated secondary amino groups was ruled out by mass spectroscopy. This compound was not characterized in further detail.
Solutions and Drugs
The saline solution (ACSF) contained (in mM): 125 NaCl, 25 NaHCO3, 2.5 KCl, 1.25 NaH2PO4, 2 CaCl2, 2 MgCl2, and 25 D-glucose, pH 7.4 maintained by saturation with carbogen (95% O2/5% CO2). In the sucrose-ACSF used for brain preparation and during slicing, 50 mM sucrose was substituted for NaCl (final 75 mM). Spermine, biotinylated spermine (B-X-SPM, and betaine and carnitine as organic cation transporter agonists were dissolved as stock solutions and added to final concentrations in ACSF prior to application.
Preparation of Brain Slices
Brain slices were derived from adult male Wistar rats from an institutional breeder (Forschungseinrichtungen für Experimentelle Medizin, Charité-Universitätsmedizin Berlin, Germany). All experiments were approved by the Regional Berlin Animals Ethics Committee (T0127/02) and performed in strict accordance with the European Communities Council directive regarding care and use of animals for experimental procedures. All efforts were made to minimize the number of specimens and animal suffering.
Animals were deeply anesthetized with diethylether and decapitated. Brains were quickly dissected and placed into ice-cold sucrose-substituted artificial cerebrospinal fluid (ACSF). Coronal slices (300 µm) containing the habenular complex were cut using a vibrating microtome (VT 1000 S; Leica Instruments, Nussloch, Germany). Slices were placed in a holding vial, incubated in oxygenated ACSF (recording solution, see below) at 32-35°C and continuously perfused with recording solution at a rate of 2.5-5.0 ml/min.
Superfusion of Brain Slices
For superfusion with selected drugs the aCSF solution was complemented with SPM (10 µM) or with B-X-SPM (5 to 20 µM). For competition experiments in addition to B-X-SPM the aCSF contained ethylenediamine, carnithine, betain, or SPM, all at concentrations of 20 µM. Subsequent to superfusion slices were shortly rinsed in aCSF and immediately fixed for 2 h in PGPic (see below). Thereafter they were cryoprotected (see below), mounted on a flat block of prefrozen Tissue Tek, shock-frozen in hexane at -70 °C, and stored at -80 °C until resectioning in a cryostat.
In-Vivo Injections
Animals were kept deeply anesthetized with isoflurane (T0127/02, see preceeding paragraph). Rats were placed into a stereotaxic frame (David Kopf Instruments, Tujinga, CA) and holes were drilled through the skull at selected positions for intracerebroventricular injections of B-X-SPM. For intracardial injections of B-X-SPM deeply anesthetized animals underwent thoracotomy under artificial respiration to expose the heart. Subsequent to injections, animals were fixed via transcardial perfusion with PGPic (4 % paraformaldehyde, 0.05 % glutaraldehyde, and 0.2 % picric acid in 0.1 M phosphate buffer, pH 7.4; [12]). Brains, livers, and kidneys were removed, cryoprotected in 0.4 M sucrose for about 4 h and in 0.8 M sucrose overnight, cut into blocks, shock-frozen in hexane at -70 °C, and stored at -80 °C until use.
Immunocytochemistry
Freely floating cryostat sections (25 µm) were subjected to immunocytochemistry as described earlier [13]. In short, sections were rinsed in PBS (phosphate buffered saline; 150 mM sodium chloride in 10 mM phosphate buffer, pH 7.4), treated for 15 min with 1% sodium borohydride in PBS to remove residual aldehyde groups from the fixative, and again thoroughly washed in PBS. Sections were pretreated for 30 minutes in a blocking and permeabilizing solution (10% normal goat serum in 0.3% Triton X-100 and 0.05% phenylhydrazine in PBS) at room temperature (RT).
Anti-SPD/SPM antibodies [14] were applied for 36 hours at appropriate dilutions in PBS containing 10% NGS, 0.3% Triton X-100, 0.1% sodium azide, and 0.01% thimerosal at 4° C. Sections were thoroughly rinsed in PBS, pretreated for 1 hour with PBS-A (0.2% bovine serum albumine in PBS), and exposed for another 24 hours at RT to the biotinylated goat anti-rabbit antibody (Vector Laboratories, Burlingame, USA), diluted 1:2.000 in PBS-A containing 0.1% Triton X-100 and 0.1% sodium azide. After repeated washings in PBS and preincubation for one hour in PBS-A, the Elite avidin-biotin-peroxidase-complex (1:200 dilution in PBS-A; Vector Labofatories) was attached to biotinylated secondary antibodies by incubation for another 12 hours at RT.
Next, sections were preincubated for 15 minutes in a solution of 0.5 mg/ml diaminobenzidine, 3 mg/ml ammonium nickel sulfate, and 10 mM imidazole in 50 mM Tris buffer, pH 7.6. Visualization of the antigen-antibody complexes was started by the addition of 0.0015% hydrogen peroxide and stopped after 15 minutes at RT by repeated washings with PBS. Sections were mounted onto gelatine-coated slides, air-dried not longer than 30 minutes, dehydrated through a graded series of ethanol, transferred into xylene, and coverslipped with entellan.
Selective Visualization of Biotinylated SPM in Tissues
The B-X-SPM molecule contains amino groups, which guarantee that it is retained in the tissue subsequent to aldehyde fixation. For visualization sections were pretreated as above. Subsequently they were directly exposed to the Elite avidin-biotin-peroxidase-complex (1:200 dilution in PBS-A; Vector Laboratories) and the B-X-SPM visualized with diaminobenzidine as describe above [15].
Results
The present investigation aimed to understand, where polyamines in the mammalian CNS are derived from. Principally, there are three possibilities: (i) intake by nutrition including the potential contribution from gut bacteria [1], release into the bloodstream from the gut, and subsequent uptake from CNS capillaries, (ii) products of general metabolism, release into the bloodstream from parenchymatous organs as the liver, and again uptake from CNS capillaries, and (iii) uptake of precursors such as arginine [1] from brain capillaries and subsequent local biosynthesis of polyamines within the CNS.
In the present investigation we focused on the most characteristic PA, spermine. Simulating uptake with natural SPM cannot produce unequivocal results, as the separate detection of endogenous and exogenously added PAs is hardly possible. Consequently, we first prepared and characterized a biotinylated and extended analogue of SPM, called B-X-SPM here. Subsequently, this SPM-analogue was used in brain slices and in-vivo experiments to understand, which parameters are important for SPM uptake in the mammalian brain.
B-SPM was synthesized by directly coupling an extended and activated derivative of biotin (NHS-X-biotin; shown in green) to SPM (shown in red) as described in detail in the Methods section. Formation of B-SPM results in the release of N-hydroxy-succinimide.
Synthesis and Characterization of Biotinylated and Extended Spermine (B-X-SPM)
B-X-SPM (Figure 1) was synthesized by direct coupling an extended and activated derivative of biotin (NHS-X-biotin) to SPM as described in detail in the Methods section. Subsequently, the components of the reaction the mixture were separated using reverse phase high pressure liquid chromatography, yielding two major peaks. The structure of the compound producing the first peak turned out to be N1-X-biotinyl-SPM (Figure 1) as confirmed by mass spectroscopy. This will be the active compound in our subsequent physiological experiments. The second peak most likely corresponds to N1,N12-doubly biotinylated SPM as the presence of acylated secondary amino groups was ruled out by mass spectroscopy. This compound was not characterized in further detail.
Three distinct procedures were used to investigate, whether B-X-SPM is taken up by astrocytes of other brain cells from superfusing aCSF (procedure 1), from the brain extracellular space subsequent to intracerebroven-tricaular injection (procedure 2) or from the blood stream (procedure 3).
In-Vitro Uptake of Native or Biotinylated SPM in Brain Slices
Brain slices provide a simple tool to simulate the uptake of native or biotinylated PAs from the intracellular fluid. To verify this idea, in a first step we tested, whether this system can be used to detect the uptake of native SPM (Figure 2; Procedure 1). SPM-containing artificial cerebrospinal fluid (CSF) provides a constant source or PAs for all types of brain cells, when it is used to superfuse surviving slices.
Uptake of Native SPM in Brain Slices
In a first set of experiments we asked, whether intracellular SPM might be lost during pretreatment of surviving slices.
When slices are fixed immediately after sectioning astrocytes display prominent SPD/SPM-immunoreactivity, Neurons are positive also, but stained less intensely (Figure 3A). After 90 minutes of equilibration, astrocytes retain strong immunoreactivity, while neurons now appear as translucent spots largely (Figure 3B), suggesting the PAs have been lost from these cells during habituation.
Tiny dots detectable already in survey micrographs (Figure 3C) indicate that astrocytes retain SPM-immunoreactivity even after superfusion with SPM-free aCSF. They are most prominent (Figure 3B, right upper corner) in the area of the Corpus callosum (labeled cc in Figure 3A). Staining becomes more intense, when slices are superfused with SPM-containing artificial CSF (compare Figure 3C and Figure 3D). This higher magnification also discloses that the tiny dots (Figure 3D) mostly represent astrocytes.
Surprisingly, separate types of neurons or neurons in distinct areas may accumulate PAs to a different level. Thus, GABAergic interneurons in the hippocampus and especially in the hilus of the dentate gyrus are largely devoid of SPD/SPM-immunoreactivity (Figure 3E,F). In contrast, inhibitory neurons in the reticular thalamic nucleus are positive without and even more when superfused with SPM (Rt, Figure 3G,H). Neurons in the ventral posterior thalamic nucleus display also but weaker SPD/SPM-immunoreactivity (VP, Figure 3G,H), while the posterior thalamic nucleus is largely devoid of positive neurons (Po, Figure 3G,H), independently whether the superfusion solution does or does not contain SPM. Astrocytes, however, are strongly positive even in the Po, while immunoreactivity of the neurpil is increased after superfusion with SPM (compare Figure 3G–H).
When slices are fixed immediately after sectioning astrocytes display prominent SPD/SPM-immunoreactivity (A). Neurons are positive also, but stained less intensely (A). After equilibration, astrocytes retain strong immunoreactivity, while neurons now are largely devoid of staining (B). Note that neuronal cell bodies stand out as translucent circles in the positive neuropil.
To identify the most important hippocampal areas in immunostained sections they are represented here after staining with cresyl violet (Nissl stain). Tiny dots detectable already in survey micrographs (D) indicate that astrocytes retain SPM-immunoreactivity even after superfusion with SPM-free aCSF. They are most prominent (D, right upper corner) in the area of the Corpus callosum (labeled cc in C). Staining becomes more intense, when slices are Superfusion with SPM-containing aCSF increases glial staining. Note that when superfused with SPM-free aCSF the upper leaflet of the dentate gyrus contains many weakly stained astrocytes (E). After superfusion with SPM-containing aCSF most of them display strong immunoreactivity (F).
Neurons in distinct areas accumulate PAs to a different level. Thus, GABAergic interneurons in the hippocampus and especially in the hilus of the dentate gyrus are largely devoid of SPD/SPM-immunoreactivity (E, F). In contrast, inhibitory neurons in the reticular thalamic nucleus are positive without and even more when superfused with SPM (G, H). Neurons in the ventral posterior thalamic nucleus display also but weaker SPD/SPM-immunoreactivity (G, H), while the posterior thalamic nucleus is largely devoid of positive neurons (G, H), independently whether the superfusion solution does (H) or does not (G)contain SPM. Astrocytes, however, are strongly positive (small arrows) even in the Po, while immunoreactivity of the neurpil is increased after superfusion with SPM (compare G to H). Rt: reticular thalamic nucleus; VP: ventral posterior thalamic nucleus; Po: posterior thalamic nucleus. Bar in (H) indicates 540 µm in (C), 300 µM in (D), (G), (H), and 70 µM in (A), (B), (E), and (F).
Uptake of Biotinylated SPM (B-X-SPM) in Brain Slices
The use of biotinylated SPM (B-X-SPM; see Figure 1) enabled us to visualize SPM freshly taken up in the presence of SPM, which had been in the cell before. For this purpose surviving slices were superfused with B-X-SPM, subsequently resectioned, and used for immunocytochemistry (see Figure 2). Total SPM was demonstrated with our anti-SPM antibody [14], while staining directly with the ABC complex selectively visualizes the freshly internalized biotinylated SPM.
At the first glance both stainings appear rather similar (compare Figure 4A,B). Closer inspection however, falsely suggests that considerably more SPM-positive cell bodies are detected by the anti-SPM antibodies (Figure 4A) as compared to the ABC complex alone (Figure 4B), which might indicate that not all astrocytes are able to take up B-X-SPM. This, however, is not the case. When focusing on the stratum radiatum (SR) of the hippocampal CA3, this area again appears to contain more astrocytes labeled for total SPM as for B-X-SPM (compare Figure 4C,D). Rough counting, however, indicates that the total number
The use of biotinylated SPM allows to visualize SPM freshly taken up in the presence of SPM, which had been in the cell before. For this purpose surviving slices were superfused with B-X-SPM, subsequently resectioned, and used for immunocytochemistry. Total SPM was demonstrated with our anti-SPM antibody, while staining directly with the ABC complex selectively visualizes the freshly internalized biotinylated SPM.
At the first glance both stainings appear rather similar (A and B). Closer inspection however, falsely suggests that considerably more SPM-positive cell bodies are detected by the anti-SPM antibodies (A, C) as compared to the ABC complex alone (B, D), which might indicate that not all astrocytes are able to take up B-X-SPM. This, however, is not the case. When focusing on the stratum radiatum (SR) of the CA3, this hippocampal area again appears to contain more astrocytes positive for total SPM as for B-X-SPM (compare C and D). Rough counting, however, indicates that the total number of astrocytes is quite similar. Instead, the cells contain much more total SPM as evident by the prominently stained astrocyte processes with the anti-SPM antibody (C), which are less easily detectable, when only the newly incorporated B-X-SPM is visualized (D). In fact, however, not all astrocytes are able to take up B-X-SPM. Thus, the fibrous-type astrocytes in the Corpus callosum do present SPM-like immunoreactivity (E), but remain negative for newly uptaken B-X-SPM (F). Bar in (F) indicates 300 µM in (A), (B), 230 µM in (C), (D), and 70 µM in E), and (F) of astrocytes is quite similar. Instead, the cells contain much more total SPM as evident by the prominently stained astrocyte processes with the anti-SPM antibody (Figure 4C), which are less easily detectable, when only the newly incorporated B-X-SPM is visualized (Figure 4D).
In fact, however, not all astrocytes are able to take up B-X-SPM. Thus, the fibrous-type astrocytes in the Corpus callosum do present SPM-like immunoreactivity (Figure 4E), but they remain negative for newly uptaked B-X-SPM (Figuer 4F). Based on this novel information, fibrous-type and protoplasmic-type astrocytes apparently are quite distinct in their ability to take up PAs from the surrounding intercellular fluid.
Incorporation of B-X- SPM into Astrocytes Is Based on a PA-Specific Uptake System
Uptake of B-X-SPM as surrogate for SPM as visualized above might be due to any more or less specific cation transporter. When this uptake, however, was due to a PA-specific system, it is expected that added PAs, but not other cations, would act as competitors, thereby reducing the uptake of B-X-SPM. Exactly this competition was confirmed here. When betain, carnitine, or ethylenediamine were used as competitors, B-X-SPM uptake was good visualized as if no competitor was present. In contrast, when the slice was superfused with additional 20 mM SPM, uptake of B-X-SPM could not be detected any longer (Supplemental Figure 1).
In-Vivo Uptake of Native or Biotinylated SPM into the brain from intraventricular CSF or from the blood stream
Our data so far demonstrate that B-X-SPM is a good surrogate to visualize the uptake of polyamines like SPM from the surrounding brain intercellular space into the brain. Slices represent an interesting model, but cannot provide realistic information, whether the system works in the intact animal.
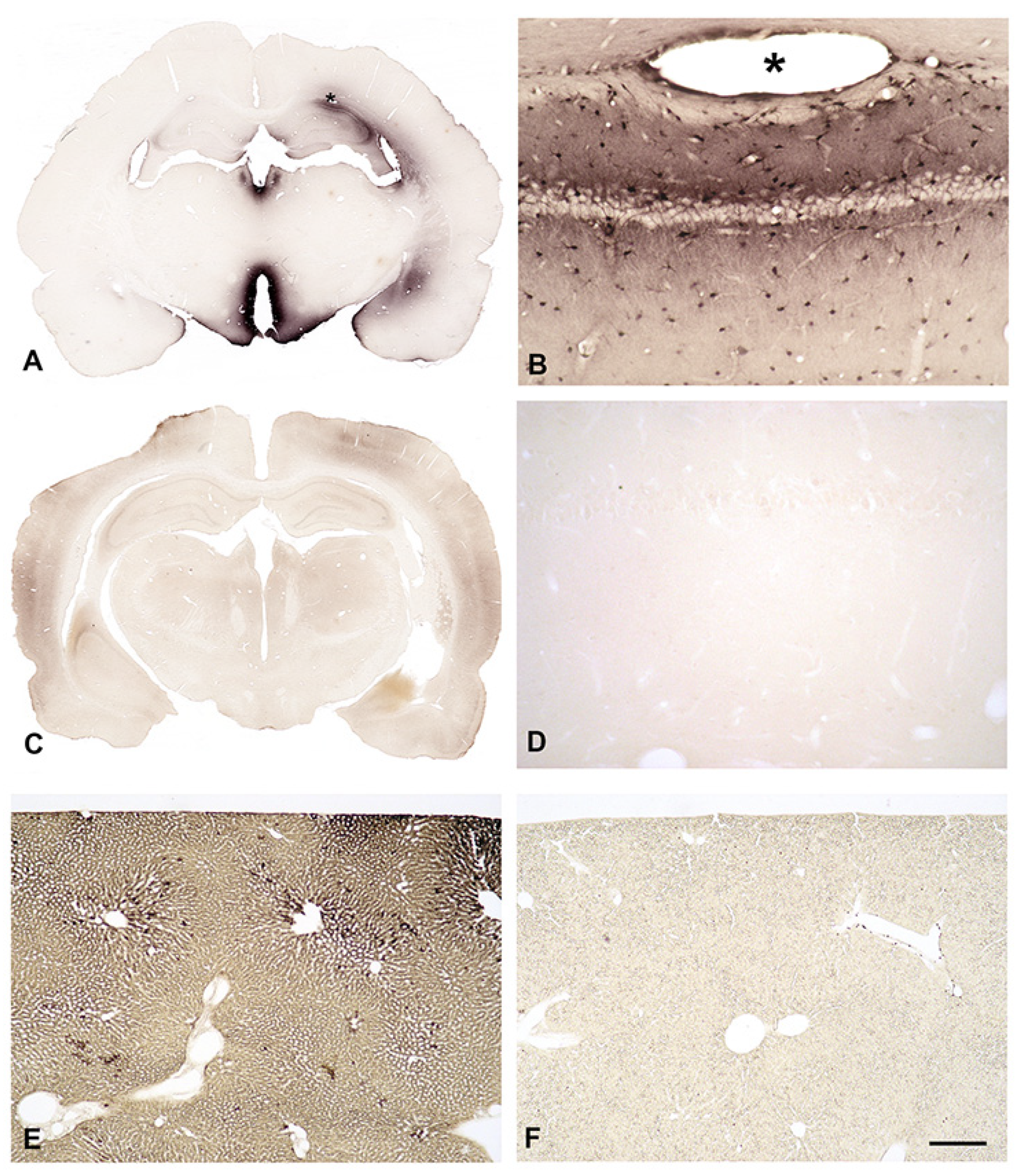
When B-X-SPM is injected into the right lateral cerebral ventricle of deeply anesthesized rats it is easily detected in the corresponding brains (A, B). Ventricles, especially the third one, are surrounded by strong staining for B-X-SPM (A). Higher magnification of the CA1 hippocampal area displays positive neuropil and strongly stained astrocytes (B). In contrast, when B-X-SPM in injected intracardially, no uptake of B-X-SPM into the CNS is observed (C). Even at high magnification absolutely no immunoreactivity can be detected (D). However, B-X-SPM is easily taken up in liver (E) indicating that in fact it can be taken up from the blood stream. The liver of an control animal, which did not receive B-X-SPM, is largely negative (F). Bar in (F) indicates 1700 µM in (A), (C), 260 µM in (E), (F), and 100 µM in (B), and (D).
In-Vivo Uptake of Biotinylated SPM into the Brain from Intraventricular CSF
To learn, whether astrocytes do collect PAs like SPM from the intercellular fluid, we injected B-X-SPM into the right lateral cerebral ventricle of deeply anesthesized rats. After transcardial perfusion fixation and adequate histological treatment, B-X-SPM is easily detected in the corresponding brains (Figure 5A,B). Ventricles, especially the third one, are surrounded by strong staining for B-X-SPM (Figure 5A). Higher magnification of the CA1 hippocampal area displays positive neurpil and strongly stained astrocytes (Figure 5B).
In-Vivo Uptake of Biotinylated SPM into the Brain from the Blood Stream
To learn, whether astrocytes do collect PAs like SPM from the blood stream, we injected single doses of B-X-SPM intracardially in deeply anesthesized rats. After 30 minutes animals were euthanasized by transcardial fixation and brain, liver and kidneys were subjected to immunocytochemistry.
Surprisingly, no uptake of B-X-SPM into the CNS could be observed (Figure 5C). Even at higher magnification absolutely no immunoreactivity can be detected (Figure 5D). However, B-X-SPM is easily taken up in liver (Figure 5E) and kidneys (not shown), indicating that indeed in peripheral organs polyamines can be taken from the blood stream.
Discussion
Our newly synthesized B-X-SPM already had been used in a previous investigation [15]. Here it is characterized in more detail and examined as invaluable tool to follow the uptake of PAs, especially SPM, by distinct cell types and organs using in-vitro and in-vivo conditions.
Technical Considerations
For coupling biotin to SPM we used a N-hydroxysuccinimide activated and spacer extended biotin derivative and native SPM (Figure 1). As SPM contains two primary and two secondary amino groups it appeared unclear, which of these two groups would be attacked by the activated SPM derivative. Reversed phase chromatography (see methods section) of the reaction products resulted in two major peaks (not shown). The first one represented the desired compound (Figure 1), as characterized by mass spectroscopy. The second one most likely contained a doubly coupled SPM and was not investigated further by economic reasons. Using either the material in the first peak or the complete mixture after reaction in pilot experiments for our biological studies revealed no major differences. Consequently, the complete mixture was used in all subsequent attempts.
B-X-SPM certainly only represents a surrogate for native SPM or other polyamines. Our competition studies, however, indicate that its uptake is based on the same system as that of natural SPM and, most likely, that of other PAs like SPD also.
Native and Biotinylated SPM Are Taken Up by Astrocytes and Neurons in Brain Slices
Astrocytic cells are not able to synthesize PAs on their own [16] but accumulate them much more effectively than neurons [1,14,17,18,19]). In contrast, neurons do synthesize PAs, most likely from arginine taken up from the bloodstream [1]. They express ornithine decarboxylase [20,21] and Spd/SPM synthase [16] and also display SPD/SPM immunoreactivity [22].
Interestingly, neuronal SPD/SPM immunoreactivity largely differs between neuronal areas and cell types. Here we find that neuronal staining is weak in the cerebral cortex and in the hippocampus (see Figure 3). Even interneurons in dentate hilus largely are devoid of immunostaining, while inhibitory neurons in the reticular thalamic nucleus are clearly SPD/SPM-positive, even increased after preincubation with native SPM. In contrast, in the same section neurons in the ventral posterior thalamic nucleus display weak staining only, and in the posterior thalamic nucleus no staining is evident at all (see Figure 3G,H). At the first glance such differences are surprising, but have also been describe earlier [22].
Uptake of Biotinylated SPM (B-X-SPM) by Brain Slices Is Different in Protoplasmic and Fibrous Astrocytes
Uptake of B-X-SPM into brain slices is due to a specific cation transporter system [15] as also supported here. Competing uptake by native SPM completely abolished the appearance of B-X-SPM in brain slices, while betain, carnitine, or ethylenediamine produced no effects (Supplemental Figure S1).
Most interesting, not all types of astrocytes can easily take up B-X-SPM. Astrocytes in the Corpus callosum, generally known to belong to the fibrous type, apparently are not able to take up B-X-SPM easily (see Figure 4E,F). The importance of this old division of astrocytes into protoplasmic and fibrous types has recently been supported by novel data [23]. Experiments with transgenic mice indicate that the Hedgehog pathway, well known for its role in the developing CNS, is active in astrocytes in the mature mouse forebrain. Not all astrocytes, however, respond to Sonic hedgehog (Shh), which in responsive cells activates Gli1 to achieve its effects. Gli1-expressing astrocytes display a morphology corresponding to the protoplasmic type of astrocytes, while fibrous-type astrocytes are not affected by Shh [23]. The importance of this fact is evident when one knows that neurons are the source of Shh. Apparently, only protoplasmic type astrocytes can respond to this neuronal communication and fibrous type cannot [23]. This is not suprising when one considers that fibrous astrocytes mostly reside in the white matter, where neurons are extremely rare. The mouse data above in combination with ours give further support to the idea that protoplasmic and fibrous type astrocytes belong to different subfamilies with potentially distinct biological functions.
Native or Biotinylated SPM Are not Taken Up into the Brain from the Blood Stream
Subsequent to its intraventricular injection B-X-SPM is easily detected in the surrounding brain areas (Figure 5A,B), indicating an unhindered uptake of PAs from the CSF. In contrast, following intracardial injection no B-X-SPM could be detected in the parenchyma of the brain. Consequently, from our data it is now completely clear that in line with earlier indirect evidences [7,8,9] the brain is not able to take up higher PAs from the bloodstream.
Conclusions
A biotinylated SPM analogue (B-X-SPM) was found to be an invaluable tool to investigate the uptake of PAs, especially SPM, by distinct cell types and organs using in-vitro and in-vivo conditions. In surviving slices uptake of B-X-SPM is massive into protoplasmic and absent in fibrous type astrocytes. It is also taken up by neurons but to a lesser degree. Under in-vivo conditions astrocyte uptake of biotinylated SPM also is massive from the brain interstitial fluid subsequent to intraventricular application. In contrast, following intracardial injection there is no uptake from the bloodstream, indicating that the brain is completely dependent on the local synthesis of polyamines.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
References
- Rieck, J.; Skatchkov, S.N.; Derst, C.; Eaton, M.J.; Veh, R.W. Unique Chemistry, Intake, and Metabolism of Polyamines in the Central Nervous System (CNS) and Its Body. Biomolecules 2022, 12, 501.
- Matsumoto, M.; Kakizoe, K.; Benno, Y. Comparison of Fecal Microbiota and Polyamine Concentration in Adult Patients with Intractable Atopic Dermatitis and Healthy Adults. Microbiology and Immunology 2007, 51, 37-46, . [CrossRef]
- Madeo, F.; Hofer, S.J.; Pendl, T.; Bauer, M.A.; Eisenberg, T.; Carmona-Gutierrez, D.; Kroemer, G. Nutritional Aspects of Spermidine. Annual Review of Nutrition 2020, 40, 135-159. [CrossRef]
- Kibe, R.; Kurihara, S.; Sakai, Y.; Suzuki, H.; Ooga, T.; Sawaki, E.; Muramatsu, K.; Nakamura, A.; Yamashita, A.; Kitada, Y.; et al. Upregulation of colonic luminal polyamines produced by intestinal microbiota delays senescence in mice. Scientific Reports 2014, 4, 4548. [CrossRef]
- Milovic, V. Polyamines in the gut lumen: bioavailability and biodistribution. European Journal of Gastroenterology & Hepatology 2001, 13, 1021-1025.
- Skatchkov, S.N.; Antonov, S.M.; Eaton, M.J. Glia and glial polyamines. Role in brain function in health and disease. Biochemistry (Moscow) Supplement Series A: Membrane and Cell Biology 2016, 10, 73-98. [CrossRef]
- Lajtha, A.; Sershen, H. Substrate specificity of uptake of diamines in mouse brain slices. Archives of Biochemistry and Biophysics 1974, 165, 539-547, . [CrossRef]
- Sershen, H.; Lajtha, A. Perinatal changes of transport systems for amino acids in slices of mouse brain. Neurochemical Research 1976, 1, 417-428. [CrossRef]
- Shin, W.-W.; Fong, W.-F.; Pang, S.-F.; Wong, P.C.-L. Limited Blood-Brain Barrier Transport of Polyamines. Journal of Neurochemistry 1985, 44, 1056-1059, . [CrossRef]
- Hopman, A.H.; Ramaekers, F.C.; Speel, E.J. Rapid synthesis of biotin-, digoxigenin-, trinitrophenyl-, and fluorochrome-labeled tyramides and their application for In situ hybridization using CARD amplification. Journal of Histochemistry & Cytochemistry 1998, 46, 771-777.
- Madai, V.I.; Poller, W.C.; Peters, D.; Berger, J.; Paliege, K.; Bernard, R.; Veh, R.W.; Laube, G. Synaptic localisation of agmatinase in rat cerebral cortex revealed by virtual pre-embedding. Amino Acids 2012, 43, 1399-1403. [CrossRef]
- Somogyi, P.; Takagi, H. A note on the use of picric acid-paraformaldehyde-glutaraldehyde fixative for correlated light and electron microscopic immunocytochemistry. Neuroscience 1982, 7, 1779-1783, . [CrossRef]
- Bernard, R.; Veh, R.W. Individual neurons in the rat lateral habenular complex project mostly to the dopaminergic ventral tegmental area or to the serotonergic raphe nuclei. Journal of Comparative Neurology 2012, 520, 2545-2558, . [CrossRef]
- Laube, G.; Veh, R.W. Astrocytes, not neurons, show most prominent staining for spermidine/spermine-like immunoreactivity in adult rat brain. Glia 1997, 19, 171-179, . [CrossRef]
- Malpica-Nieves, C.J.; Rivera, Y.; Rivera-Aponte, D.E.; Phanstiel, O.; Veh, R.W.; Eaton, M.J.; Skatchkov, S.N. Uptake of Biotinylated Spermine in Astrocytes: Effect of Cx43 siRNA, HIV-Tat Protein and Polyamine Transport Inhibitor on Polyamine Uptake. Biomolecules 2021, 11, 1187.
- Krauss, M.; Langnaese, K.; Richter, K.; Brunk, I.; Wieske, M.; Ahnert-Hilger, G.; Veh, R.W.; Laube, G. Spermidine synthase is prominently expressed in the striatal patch compartment and in putative interneurones of the matrix compartment. Journal of Neurochemistry 2006, 97, 174-189. [CrossRef]
- Biedermann, B.; Skatchkov, S.N.; Brunk, I.; Bringmann, A.; Pannicke, T.; Bernstein, H.-G.; Faude, F.; Germer, A.; Veh, R.; Reichenbach, A. Spermine/spermidine is expressed by retinal glial (müller) cells and controls distinct K+ channels of their membrane. Glia 1998, 23, 209-220. [CrossRef]
- Skatchkov, S.N.; Eaton, M.J.; Krušek, J.; Veh, R.W.; Biedermann, B.; Bringmann, A.; Pannicke, T.; Orkand, R.K.; Reichenbach, A. Spatial distribution of spermine/spermidine content and K+-current rectification in frog retinal glial (Müller) cells. Glia 2000, 31, 84-90. [CrossRef]
- Kovács, Z.; Skatchkov, S.N.; Veh, R.W.; Szabó, Z.; Németh, K.; Szabó, P.T.; Kardos, J.; Héja, L. Critical Role of Astrocytic Polyamine and GABA Metabolism in Epileptogenesis. Frontiers in Cellular Neuroscience 2022, 15, doi:10.3389/fncel.2021.787319.
- Bernstein, H.-G.; Müller, M. The cellular localization of the l-ornithine decarboxylase/polyamine system in normal and diseased central nervous systems. Progress in Neurobiology 1999, 57, 485-505. [CrossRef]
- Kilpeläinen, P.; Rybnikova, E.; Hietala, O.; Pelto-Huikko, M. Expression of ODC and its regulatory protein antizyme in the adult rat brain. Journal of Neuroscience Research 2000, 62, 675-685. [CrossRef]
- Laube, G.; Bernstein, H.-G.; Wolf, G.; Veh, R.W. Differential distribution of spermidine/spermine-like immunoreactivity in neurons of the adult rat brain. Journal of Comparative Neurology 2002, 444, 369-386. [CrossRef]
- Garcia, A.D.R.; Petrova, R.; Eng, L.; Joyner, A.L. Sonic Hedgehog Regulates Discrete Populations of Astrocytes in the Adult Mouse Forebrain. The Journal of Neuroscience 2010, 30, 13597-13608. [CrossRef]
Figure 1.
Synthesis of biotinylated and spacer-extended spermine (B-X-SPM).
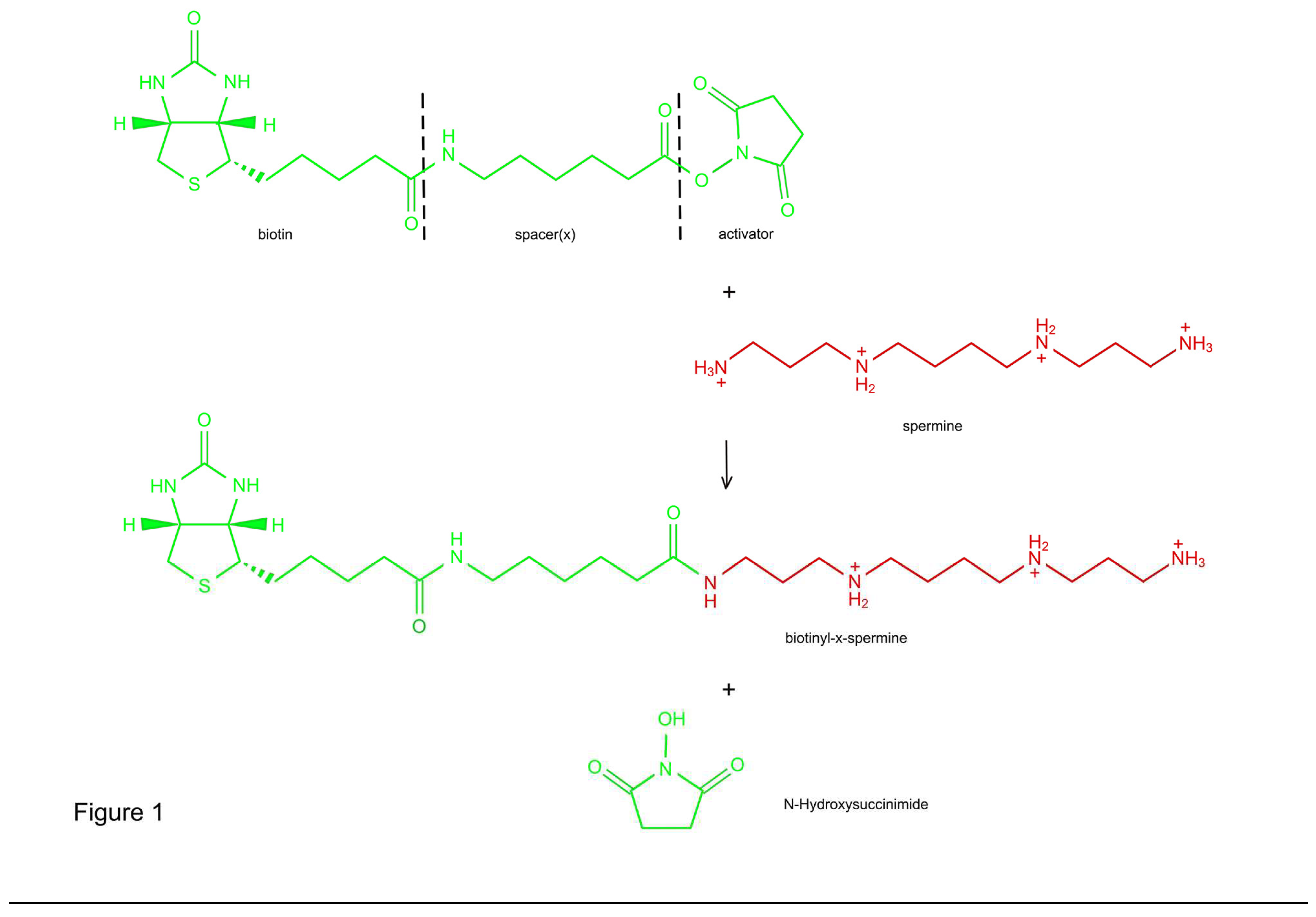
Figure 2.
Procedures to visualize the potential uptake of SPM or B-X-SPM.
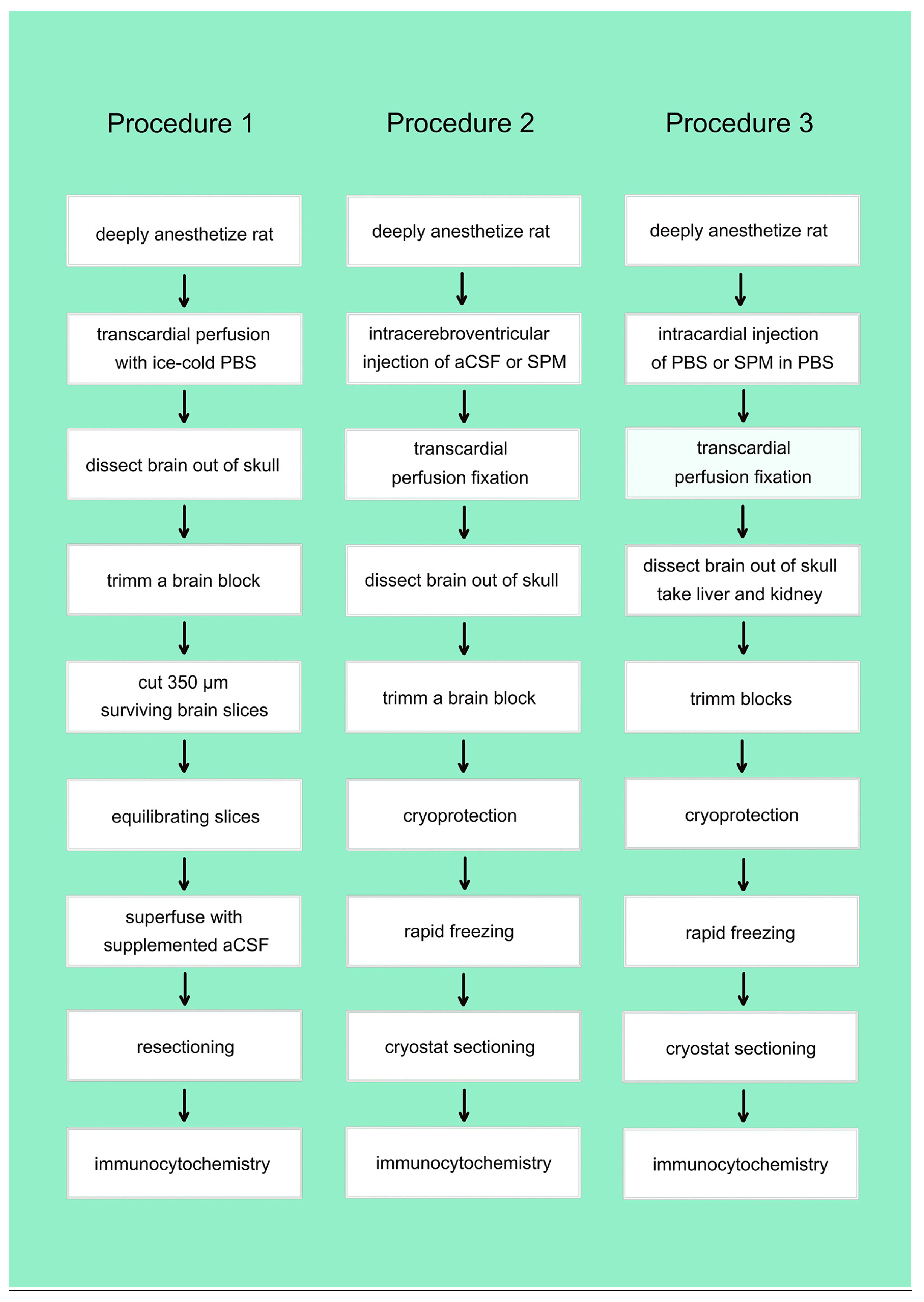
Figure 3.
SPM-immunoreactivity in astrocytes and neurons after superfusion with aICF devoid of or complemented with SPM.
Figure 3.
SPM-immunoreactivity in astrocytes and neurons after superfusion with aICF devoid of or complemented with SPM.
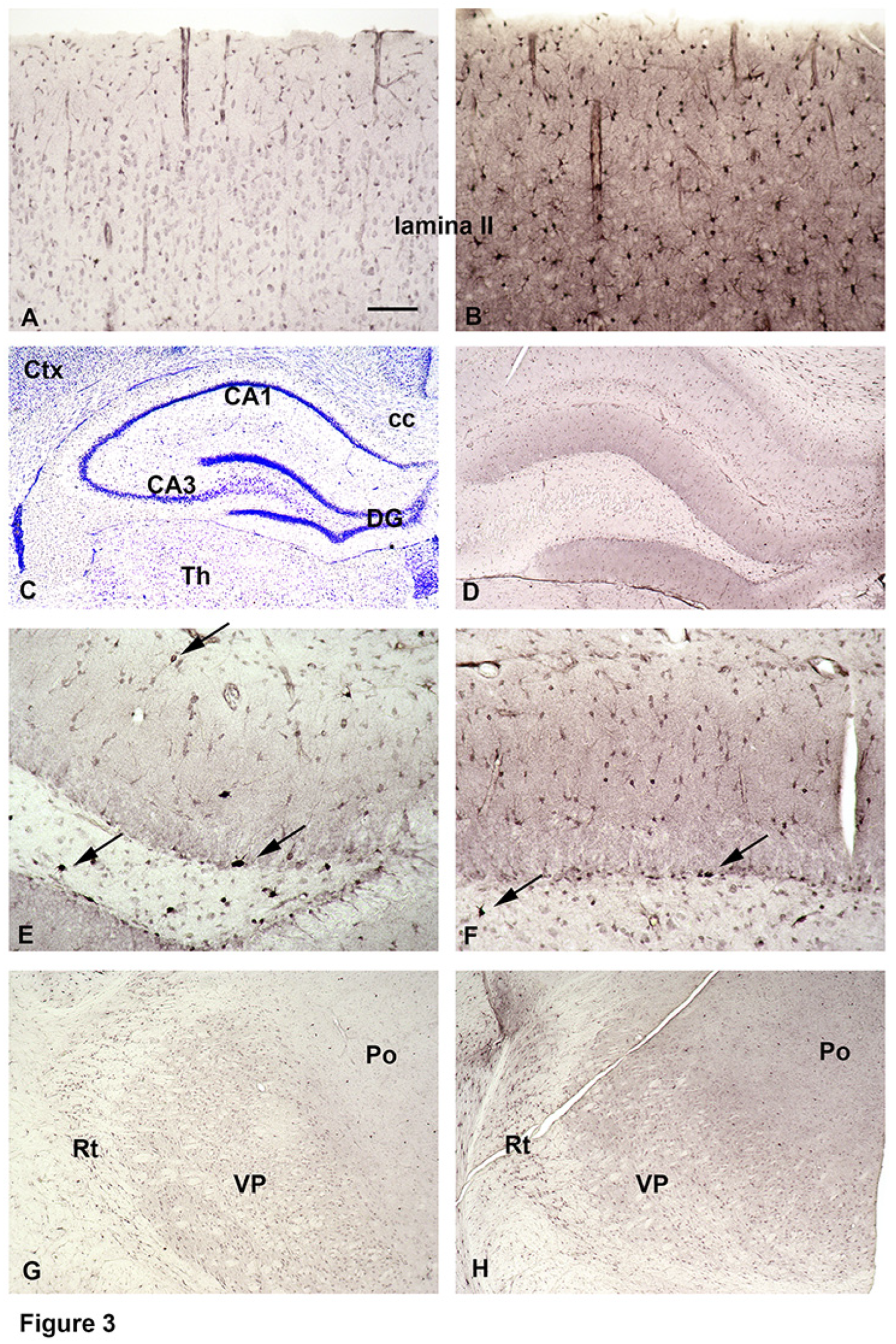
Figure 4.
SPM, which has been freshly taken by astrocytes, is visualized in the presence of SPM, which had been in the cell before.
Figure 4.
SPM, which has been freshly taken by astrocytes, is visualized in the presence of SPM, which had been in the cell before.
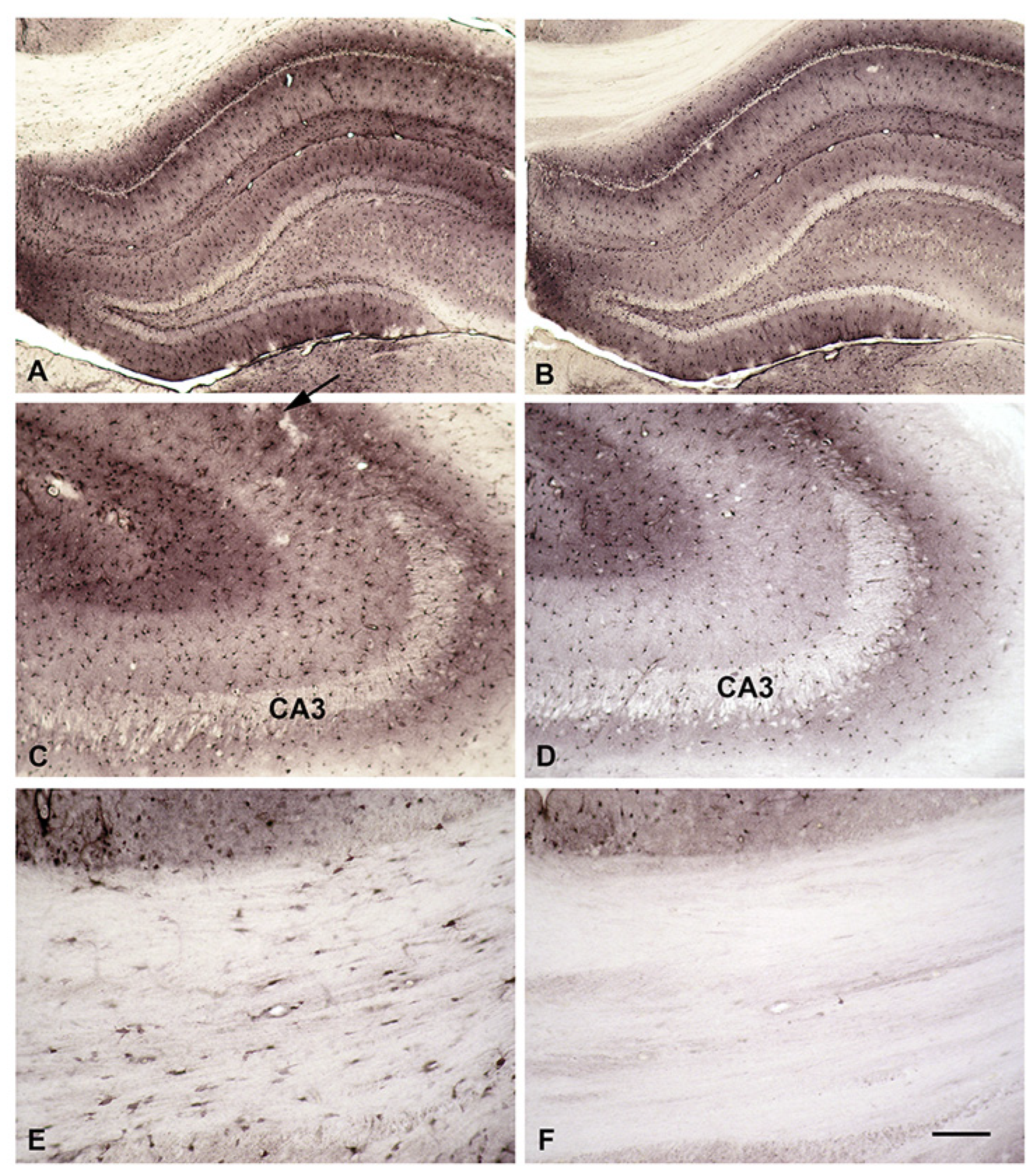
Figure 5.
In-vivo uptake of biotinylated SPM from intraventricular CSF or from the bloodstream.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



