Submitted:
24 May 2023
Posted:
26 May 2023
You are already at the latest version
Abstract
MicroRNAs (MiRNAs) are a class of non-coding single-stranded RNA molecules of approximately 20-24 nucleotides in plants that play an important regulatory role in a variety of biological processes such as plant growth and development and response to various abiotic stresses. For example Drought, Salt, Cold, High temperature, Heavy metals and Nutrition. MiRNAs affect gene expression by manipulating the cleavage, translational expression or DNA methylation of target mRNAs. This review describes the current progress made on the way miRNAs are produced and regulated and the way miRNA/target gene is used in plant responses to abiotic stresses. Studying the molecular mechanism of action of miRNAs downstream target genes can optimize the genetic manipulation of crop growth and development conditions that provide a more theoretical basis for improving crop production.
Keywords:
microRNA
; target gene
; plant growth and development
; abiotic stress response
1. Introduction
MicroRNAs, a class of plant non-coding single-stranded RNA molecules of approximately 20-24 nucleotides in length encoded by endogenous genes, have a variety of important regulatory roles in cells, participating in the regulation of plant growth and development, stress responses and hormone signalling through the negative regulation of plant gene expression, and post-transcriptional regulation of gene expression in plants [1]. microRNAs can complementarily bind to the 3’UTR region of the target mRNA, thus achieving negative regulation of gene expression. Several miRNAs can also regulate the same gene and be regulated by a combination of several miRNAs. It has been shown that miRNAs are not only conserved in gene location, but also exhibit a high degree of sequence homology, that most mammalian miRNAs are located in transcriptionunits (TUs), and most of them are located in intronic regions [2,3,4]. The high degree of conservation is strongly related to its functional importance, potentially demonstrating that homologous miRNAs serve similar functions across different species. The purpose of this review is to provide a deeper understanding of molecular mechanisms of miRNA involvement in response to abiotic stresses, For example Drought, Salt, Cold, High temperature, Heavy metals and Nutrition. Since the first discovery of microRNAs in animals in 1993, more and more researchers have become interested in this non-coding RNA [3]. Furthermore, the aim of this paper is to provide a functional perspective on the role of miRNAs in plant adaptation to agronomic abiotic stress conditions, as well as to provide a theoretical basis for miRNA improving plant resistance in abiotic stress with a view to increasing crop yields.
2. Production and Mechanism of miRNAs
2.1. Production of miRNAs
Gene transcription processing: RNA polymerase II in most cases and RNA polymerase III in a few cases transcribe from genomic sequences to form the precursor pri-miRNA. pri-miRNA is very long, ranging from a few hundred to several thousand nucleotides. pri-miRNA maintains its non-activated state by cap structure and polyadenylation and coiling of the spatial structure. Intranuclear processing: pri-miRNA is edited by the adenylate deaminase of RNA and forms pre-miRNA hairpin sequences after being sheared by the microprocessor formed by the enzyme.
Nuclear export processing: The nucleoplasmic transporter protein recognizes the two nucleotides protruding from the 3′ ends of the pre-miRNA hairpin sequence and transports the pre-miRNA from the nucleus to the cytoplasm. Nucleus processing: The nuclease Dicer begins to shear the stem-loop structure in the hairpin sequence, after which it is able to produce miRNA double strands. Pre-miRNA precursors are processed into miRNA/miRNA double-stranded bodies via the DICER-LIKE 1 (DCL1) protein, nucleocapsid binding complex (CBC), mushroom (DDL) and sawtooth (SE), then the double-stranded body is translocated into the cytoplasm.The pre-miRNA is then processed into a mature double-stranded miRNA/miRNA complex with the help of the HUA ENHANCER 1 (HEN1) protein helper complex. When the mature miRNA is embedded in an RNA-induced silencing complex, the antisense miRNA is degraded in the cytoplasm. The RNA-induced silencing complex (RISC), known as the miRNA-ribonucleoprotein complex (miRNP), contains mature miRNAs and proteins. The ribonucleoprotein complex miRNP interacts with target mRNAs through its cognate mature miRNAs, negatively regulates the expression of target genes at the post-transcriptional level, leading to mRNA degradation or translational repression under specific mechanisms of action [5,6,7](Figure 1).
2.2. Mechanism of miRNAs
Since most plant miRNAs are derived from the reverse copy of the target gene, the bases of the miRNA are complementary to those of the target mRNA. after the plant miRNA recognizes and binds to the target mRNA, the AGO will shear the target mRNA at the 10th and 11th nucleotides of the miRNA binding site. the AGO is the protein for the miRNA to carry out its function, miRNAs must bind to AGO to function. Since most plant miRNAs are derived from the reverse replication of target genes, the bases of miRNAs are complementary to those of target mRNAs, and when plant miRNAs bind to target mRNAs, the AGO will shear the target mRNA of the miRNA binding site [8,9]. Ten AGOs have been identified in Arabidopsis, which AGO1 has four structural domains, PAZ, Mid, PIWI and the N-terminal domain, respectively. The PIWI domain has nucleic acid endonuclease activity and functions as a shear for its target mRNA. Complete base complementarity between bases near the miRNA shear site and the target mRNA is important for AGO shear [8] (Figure 2A).
Another mechanism of plant miRNAs is through translational repression. The miRNA does not bind to the 3′ UTR region of the target mRNA in its entirety and the RNA-induced silencing complex inhibits the initiation of translation or the specific degradation of the synthetic ribosome to achieve translation inhibition. Aukerman found that overexpression of miR172 did not reduce the expression abundance of target mRNAs, but the corresponding levels of proteins encoded by target mRNAs were significantly reduced [10]. they therefore proposed for the first time that plant miRNAs could also repress translation of target genes. Subsequently, several studies have shown that translational repression of target genes by plant miRNAs is also a common phenomenon, and even the same miRNA may regulate target genes in both a shearing and translational repression manner. This is one of the reasons why plant miRNAs are not fully complementary to the expression of their target genes [11,12] (Figure 2B). In plants, miRNAs are an important modality for mRNA regulation through translational repression.
3. For miRNA and Drought Stress
Water is a vital resource for the survival of all life and has played an important role in the evolution of life. The most abundant substance in plant cells is water, which is an essential component of the plant body. With sufficient water, the stalks and branches of plants can stand up and stretch in the air, and the flowers can bloom better and facilitate the completion of pollination. Water is also one of the raw materials for photosynthesis in green plants and if there is a lack of water, the plant’s photosynthesis will be weakened. Leaves will wilt and in severe cases can lead to the death of the plant [13,14,15]. MiRNA156 was one of the first miRNAs identified in plants, and numerous studies have linked miRNA156 to drought stress. anthocyanins act as a secondary metabolite by scavenging ROS to protect plants from stress. In Arabidopsis, rice, alfalfa and poplar, miR156/SPL is present in Arabidopsis, rice, alfalfa and poplar by regulating anthocyanin accumulation levels in response to plant drought stress. mechanism in response to plant drought stress by regulating the level of anthocyanin accumulation [16,17]. López-Galiano et al. showed that drought conditions lead to down-regulation of miRNA159 and up-regulation of its target gene transcription factor MYB33 in tomato [18]. Reyeset al showed that during Arabidopsis seed germination, ABA induces the accumulation of microRNA 159 (miR159) in an ABI3-dependent manner, and miRNA159 mediates the cleavage of MYB101 and MYB33 transcripts in vitro and in vivo [19]. Zhang et al. showed that fine localization and functional analysis identified the candidate gene ZmLRT of qLRT5-1 as expressing the major transcript of microRNA miR166a, and that knockdown of ZmLRT enhanced drought tolerance in maize seedlings [20].
Stomata play a central role in the exchange of gases between plants and their environment, and their opening and closing is influenced by environmental signals, as well as being regulated by endogenous hormones, which in turn affect the plant’s response and tolerance to drought stress. ABA is the most critical hormone in drought stress, it regulating water loss, stomatal opening and closing [21,22,23,24]. MiR393 positively regulated stomatal density and negatively regulated guard cell length, while overexpressing plants had the opposite phenotype to the deletion mutant, possibly due to miR393 regulating the expression of growth hormone response factor 5 (ARF5) and three stomatal development-related genes, epidermal pattern factor 1 (EPF1), SPEECHLESS (SPCH) and MUTE. The miR393 overexpression strain was more sensitive to drought treatment, accumulating more malondialdehyde (MDA) and hydrogen peroxide (H2O2) compared to the wild type, and also inhibiting the accumulation of ABA in leaves. These results also demonstrate that miR393 responds to plant drought stress by interacting with ABA and regulating stomatal density [25]. Zhao et al. showed that plants overexpressing miR393a exhibited enhanced drought stress tolerance associated with stomatal density and epidermal densification. MiR393 regulates the expression of Auxin signalling F-BOX 2 (AsAFB2) and TRANSPORTINHIBITOR RESPONSE 1 (AsTIR1) [26]. To adapt to drought stress plants require a hormone monocrotaline lactone, and exogenous monocrotaline lactone applied to tomato induces the accumulation of miR156. miR156-OE and monocrotaline lactone treatments both result in reduced stomatal conductance and increased ABA sensitivity in plants [27]. MiR398c was able to negatively regulate drought resistance in soybean. Overexpression of miR398c in Arabidopsis reduced the expression of GmCSD1a/b, GmCSD2a/b/c and GmCCS, impaired the ability to scavenge active oxygen, and increased relative electrolyte leakage and stomatal opening. This resulted in reduced germination, increased water loss from leaves, reduced survival under water deficit and demonstrated sensitivity to drought during seed germination and seedling growth [28].
Plants can also improve their drought stress tolerance by altering root conformation and adjusting leaf size and curl to reduce water evaporation. Hang et al. showed that the increased drought tolerance in OsmiR408 transgenic plants may be due to changes in leaf morphology that facilitate the maintenance of water status, as well as increased antioxidant capacity to protect against damage from reactive oxygen specie (ROS) under stress [29]. Wang et al. showed that miR9674a showed progressive up-regulation in response to drought stress treatment. Overexpressing miR9674a lines exhibited different growth characteristics under drought and salt treatment in tobacco, with significant improvements in plant biomass, leaf area and root length, while its knockout expression line showed significant alleviation in the above growth traits compared to the wild type [30].
4. For miRNA and Salt Stress
Soil salinity affects around 6% of the world’s land and 23% of arable land, causing considerable economic losses through crop stress and reduced yields. Because salinity plays a vital role in plant growth, above a certain limit, excess soluble salts will have a toxic effect on plants, affecting the levels of a wide range of endogenous plant signalling molecules such as ABA, ethylene, gibberellin (GA), ROS, and Nitric oxide (NO), These hormones can greatly affect the growth and development of plants, ultimately resulting in reduced yields [31,32]. In recent years, a number of miRNAs have been identified through miRNA studies in response to salt stress. The increased abundance of miR399 under salt stress, and therefore the altered expression of PHO2 target genes, resulted in significant changes in the expression levels of two PO4 transporter genes, PHOSPHATE TRANSPORTER1;4 (PHT1;4) and PHT1;9. In salt-stressed Arabidopsis would enhance PO4 transport from roots to shoot tissues, and these aerial tissues could use these resources to maintain essential biological processes or to generate adaptive responses to salt stress [33]. The mRNA for PpDCL1a encodes an essential Dicer protein for microRNA (miRNA) biogenesis and contains an intron miRNA (miR1047). Precise deletion of the intron containing MIR1047 to abrogate PpDCL1a autoregulatory feedback control revealed a hypersensitive response to salt stress and an insensitive response to the phytohormone ABA, and the physiological importance of feedback control of miR1047 on the abundance of PpDCL1a transcripts, which controls miRNA expression and its homologous target RNAs during salt stress adaptation[34]. Overexpression of sly-miR398b inhibited plant growth under salinity conditions in tomato, including less above-ground and root biomass and shorter plant height. Further analysis showed that overexpression of sly-miR398b down-regulated the expression of Cu/Zn superoxide dismutase (CSD) [35]. Liu et al. identified that two contrasting F. velutina cuttings clones, salt-tolerant (R7) and salt-sensitive (S4), and found to exhibit higher salt tolerance in R7 than in S4. In R7 leaves, miR164d, miR171b/c, miR396a and miR160g targeting NAC1, SCL22, GRF1 and ARF18, respectively, were involved in salt tolerance. In R7 roots, miR396a, miR156a/b, miR8175, miR319a/d and miR393a targeting TGA2.3, SBP14, GR-RBP, TCP2/4 and TIR1, respectively. That were involved in salt stress response [36]. Yuan et al. found that Osa-miR396c transgenic plants exhibited reduced biomass, shorter internodes, reduced leaf area and reduced leaf size compared with wild-type, while The transgenic plants showed increased water retention under high salt stress [37].
5. For miRNA and Temperature Stress
5.1. For miRNA and Cold Stress
Temperature is the main environmental factor affecting plant growth and development and the quality of life of the fruit after harvest. Low temperature can inhibit plant growth and is a very important abiotic stressor. A variety of miRNAs can be involved in the low temperature stress response of plants by affecting the IAA or ABA signalling pathway [38]. Wang et al. found that miR319 has been shown to target the TEOSINTE BRANCHED/CYCLOIDEA/PCF (TCP) transcription factors, which are involved in regulating multiple processes in plant growth and development by controlling cell proliferation. miR319 expression is down-regulated by low temperature induction, while its target genes, OsPCF6 and OsTCP21 are reversed. Osa-miRNA319 overexpression enhanced tolerance to low temperature stress [39]. Overexpression of miRNA156 resulted in increased cell viability and growth rate under cold stress in Arabidopsis, pine and rice. OsmiR156 increased plant cold tolerance by targeting OsSPL3, which positively regulates the expression of OsWRKY71, a negative regulator of the transcription factors OsMYB2 and OsMYB3R-2 [40]. Dong et al. found that SlNAM3 enhances cold tolerance and Sl-miR164a/b-5p plays a negative role in cold tolerance by repressing the expression upstream of SlNAM3. miR164a-NAM3 module induces ethylene synthesis by directly regulating the expression of SlACS1A, SlACS1B, SlACO1 and SlACO4, thereby conferring cold tolerance in tomato [41]. The APETALA2/ethylene response factor (ERF) transcription factor OsERF096 was identified as a target of miR1320, which negatively regulates cold stress tolerance. Overexpression miR1320 leads to increased cold tolerance, while miR1320 knockdown lines decreases cold tolerance. The MiR1320-OsERF096 module regulates cold tolerance by inhibiting the JA-mediated cold signalling pathway [42].
5.2. For miRNA and High Temperature Stress
The response of plants to temperature stress is a complex process involving a variety of metabolic and biochemical processes. Not only low temperatures affect plant growth and development, but also high temperatures negatively affect processes such as growth, development and reproduction [43,44,45,46,47,48]. Wang et al. found that SRL10, a double-stranded RNA-binding protein, regulates leaf morphology and heat tolerance in rice by altering microRNA biogenesis. The srl10 mutant has a semi-curled leaf phenotype and increased heat sensitivity. SRL10 interacts directly with catalase isozyme B (CATB) to enhance hydrogen peroxide (H2O2) scavenging, thereby promoting heat tolerance [49]. Li et al. found that Overexpression of cucumber miR9748 in Arabidopsis thaliana increased high temperature tolerance. Transcriptome analysis suggests that miR9748 may mediate high temperature tolerance through the phytohormone signalling pathway. The target gene of miR9748 is CsNPF4.4, which negatively regulates high temperature stress tolerance by repressing the JA signalling pathway [50]. Ahmed et al. found that Novel and conserved heat responsive miRNAs were identified in Chinese cabbage using a high-throughput sequencing approach using heat stress treatment at 38°C . This analysis identified 41 conserved miRNAs from 19 families, with miRNA156, miRNA159, miRNA168, miRNA171 and miRNA1885 having the most abundant molecules [51].
6. For miRNA and Heavy Metals Stress
The excessive accumulation of heavy metals in plants can poison plants, affect their growth and productivity, affect humans and animals through enrichment in the food chain, cause disease and induce cell damage. Metal elements include essential and non-essential elements. Essential metals are required for many physiological processes in living organisms, such as zinc, manganese and copper, non-essential metals are cadmium, lead or mercury [52,53,54,55,56,57,58,59,60,61,62]. Zhang et al. found that overexpressing miR156 accumulated significantly less Cd in their branches and showed enhanced tolerance to Cd stress in plants. The reason for this is that miR156 positively regulates Cd stress tolerance by regulating ROS levels and Cd uptake/transport gene expression [63]. Plants overexpressing miR408 showed severe susceptibility to low sulphur (LS), arsenite As(III) and LS+As(III) stresses due to altered and miR408 knockout mutants showed tolerance that regulated expression of genes involved in the sulphur reduction pathway and affecting the accumulation of sulphate and glutathione [64]. Nie et al. found miRNA167a, novel_miRNA15, novel_miRNA22 and their targets may be involved in Cr transport and chelation. In addition, miRNA156a, miRNA164, miRNA396d and novel_miRNA155 were identified as being involved in the detoxification of plant Cr [65]. Zhou et al. by comparing miRNAs and transcriptome analysis, a total of three known and 19 new differentially expressed microRNAs (DEMs) and 1561 differentially expressed genes (DEGs) were identified following Cd treatment, mainly because miRNAs play an important role in Cd-stressed wheat by regulating targets such as TaHMA2;1 [66]. Overexpression of miR393 abolished the inhibition of root elongation by aluminium ions. In addition, overexpression miR393 attenuated the effect of exogenous growth hormone on aluminium-induced root growth inhibition and down-regulated the expression of growth hormone-responsive genes under aluminium stress [67].
7. For miRNA and Nutrient Stress
The nutrients of plants include nitrogen, phosphorus and potassium, which play a very important role in the growth and development of plants [68,69,70,71]. Nitrogen is a major component of many important compounds in plants, participating in a range of biochemical reactions and playing an important role in crop biomass accumulation and yield enhancement [72]. Phosphorus is involved in photosynthesis, respiration, energy storage and transfer, cell division, cell enlargement and a number of other processes in the plant [73]. Potassium is involved in osmoregulation, material transport and other processes, and can improve plant stress tolerance [74]. It has been found in studies that nutrient deficiencies in plants cause plants to exhibit reduced dry weight of tissues in the above and below ground parts, reduced root length, root surface area, root volume, root vigour and reduced root respiration. Therefore, a deficiency of elements will greatly affect plant growth and in severe cases cause plant death [75].
Nitrogen stress: In past studies miRNA functions were identified in response to nitrate and N deficiency. MiR167 is able to limit root growth, mainly because it controls the response of adventitious plants to N and even controls N metabolizing enzymes produced downstream of nitrification and uptake, thereby affecting plant growth through N [76]. miR393 is activated by N signalling transmitted during nitrification and uptake. Nitrate had no effect on primary root development in overexpressing miR393 plants or afb3-1 mutants, but it controlled horizontal root development in response to nitrate treatment [77,78].
Phosphorus stress: In past studies miRNA functions were identified in response to phosphorus. MiR399 is an important component of the phosphorus starvation signalling pathway. The function of miR399 in phosphorus starvation signalling was first elucidated in Arabidopsis. miR399 expression was increased under phosphorus starvation conditions, increasing the uptake and translocation of inorganic phosphorus by plants in response to phosphorus deficiency [79]. Hu et al. showed that up-regulation of genes in response to phosphorus starvation, many genes involved in iron, potassium, sodium and calcium uptake were also significantly up-regulated in overexpression miR399 strains, with increased concentrations of iron, potassium, sodium and calcium. In addition, function of Ospho2 mutant also resulted in increased concentrations of these nutrients as well as upregulation of related genes. This demonstrates that miRNA399 influences plant responses to nutrient stress by regulating PHO2 expression [80].
Potassium stress: In past studies miRNA functions were identified in response to potassium. Researchers have demonstrated that miRNA expression in cotton and wheat is altered by low dietary potassium utilisation. K deficiency treatment resulted in altered expression of 16 of the 20 miRNAs. As a response to K deficiency, wheat increases root growth and nutrient uptake through molecular mechanisms. In peanut plants, root development is influenced by miRNAs, which play a key role in K deficiency conditions. miR156 and miR390, together with miR160, miR164 and miR393, are proposed to be up-regulated in response to potassium deficiency [81,82]. Under low K stress in barley, many miRNAs appear to be differentially expressed including Hvu-miR160a, Hvu-miR169h and Hvu-miR396c. Due to the induction of Hvu-miR319 under low K, it is able to repress the expression of growth response factor (HvGRF) and thus promote Hvu-miR396 transcription in barley [83]. The dormancy-associated MADS-box (OsMADS23) target gene is significantly up-regulated in response to potassium deficiency, while Osa-miR444a clearly regulates N and P accumulation [84].
Except nitrogen, phosphorus and potassium, there are other elements in plants that play a very important role in plant growth, such as magnesium (Mg), Iron (Fe), sulfate (S), manganese (Mn), copperand (Cu), and boron (B). Mg is one of the main components of chlorophyll and promotes the activation of phosphatase and glucose convertase, facilitating the conversion of monosaccharides. Fe is an essential element for chlorophyll formation and is directly or indirectly involved in the formation of chloroplast proteins. S is protein, amino acid, vitamin and enzyme component. Promotes redox, growth regulation and is involved in chlorophyll formation and sugar metabolism. Cu is a core element in the activation groups of various oxidative enzymes in the crop and plays an important role in catalysing redox reactions in the crop. Mn is an activator of enzymes and a component of chloroplasts. B is involved in water, sugar and nitrogen metabolism and cell membrane pectin formation, it is involved in promoting the differentiation of meristematic tissues, the development of flowering organs and seed formation [85,86,87,88,89]. During sulfate limitation, miR395 expression is significantly upregulated. miR395 targets two genes capable of participating in the sulfate metabolism pathway, ATP sulfatase (encoded by the APS gene) and sulfate transporter protein 2;1 (SULTR2;1, also known as AST68) [90]. Valdés et al. found Discovery of novel common bean stress response miRNAs for manganese toxicity [91]. Kayihan et al. found expression levels of miRNAs for transcription factors related to JA and ethylene metabolism were significantly induced in moderate B toxicity but not in severe B toxicity, with the most significant regulation obtained in Arabidopsis by miR172 and miR319 [92]. Ozhuner et al. found a total of 31 known miRNAs and 3 new miRNAs were identified in barley, 25 of these were found to be responsive to boron treatment [93]. Thus, miRNAs may therefore plant regulate the expression of downstream genes to help plants to resist the stress.
8. Conclusion and Prospects
Environmental stresses, such as drought, salt, temperature, heavy metals and nutrient stress, affect the metabolic processes of plants, which in turn regulate the expression of secondary metabolites, the synthesis of which reduces the toxic effects of reactive oxygen groups through signal transduction, redox and other mechanisms to ensure the continued survival of the plant (Table 1). Much research have shown that differential expression of miRNAs is induced in plants in response to different environmental stresses. MiRNAs are important regulators in the gene regulatory network and have various functions in regulating the growth, development, programmed cell death and metabolism of organisms [120,121]. MiRNAs can cause changes in the expression of various genes in plants, and therefore their study can help improve the resistance of plants to abiotic stresses. Although miRNAs have been studied for a long time, there is little data available on the link between secondary metabolites and abiotic stresses, and there are still many plant miRNA functions that have not yet been verified. Most articles focus on the role of miRNAs and their target genes in biological processes, while the molecular mechanisms of how miRNAs receive upstream signals and influence various downstream regulatory pathways through cascade responses are still unclear.
With in-depth research on the formation, function and mechanism of action of plant miRNAs and continuous improvement and innovation in miRNA research method, more and more miRNAs will be validated to play a critical role in plant resistance to abiotic stresses, laying the foundation for a more systematic miRNA regulatory network. In summary, miRNAs are essential for plants to regulate mRNA translation in plants, and research to explore the mechanisms of miRNA downstream target gene action can provide a more theoretical basis for improving food production. In the future researchers focus on miRNAs, where genomic information is scarce, will be of great significance in broadening the scope of species and research areas.
Author Contributions
All the authors contributed to the present form of the manuscript. Z.M collected the data, drafted the manuscript; Z.M. and L.J. edited; Z.M. and L.J. created figures and tables; Z.M. and L.J. supervised; Z.M. and L.J. finalized and approved the final version of manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by National Natural Science Foundation of China (32201695) and Scientific Research Project of Education Department of Jilin Province of China (JJKH20211130KJ).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no competing interests.
References
- Laubinger S, Sachsenberg T, Zeller G, Busch W, Lohmann JU, Rätsch G, Weigel D. Dual roles of the nuclear cap-binding complex and SERRATE in pre-mRNA splicing and microRNA processing in Arabidopsis thaliana. Proc Natl Acad Sci USA 2008, 105, 8795–800. [CrossRef] [PubMed]
- Rodriguez, A.; Griffiths-Jones, S.; Ashurst, J.L.; Bradley, A. Identification of Mammalian microRNA Host Genes and Transcription Units. Genome Res. 2004, 14, 1902–1910. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Kim VN, Nam JW. Genomics of microRNA. Trends Genet. 2006, 22, 165–73.
- Iki, T.; Yoshikawa, M.; Meshi, T.; Ishikawa, M. Cyclophilin 40 facilitates HSP90-mediated RISC assembly in plants. EMBO J. 2011, 31, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Park, M.Y.; Wu, G.; Gonzalez-Sulser, A.; Vaucheret, H.; Poethig, R.S. Nuclear processing and export of microRNAs in Arabidopsis. Proc. Natl. Acad. Sci. 2005, 102, 3691–3696. [Google Scholar] [CrossRef] [PubMed]
- Thieme CJ, Schudoma C, May P, Walther D. Give It AGO: The Search for miRNA-Argonaute Sorting Signals in Arabidopsis thaliana Indicates a Relevance of Sequence Positions Other than the 5’-Position Alone. Front Plant Sci. 2012, 3, 272.
- Liu, Q.; Wang, F.; Axtell, M.J. Analysis of Complementarity Requirements for Plant MicroRNA Targeting Using a Nicotiana benthamiana Quantitative Transient Assay. Plant Cell 2014, 26, 741–753. [Google Scholar] [CrossRef]
- Franco-Zorrilla, J.M.; Valli, A.; Todesco, M.; Mateos, I.; Puga, M.I.; Rubio-Somoza, I.; Leyva, A.; Weigel, D.; García, J.A.; Paz-Ares, J. Target mimicry provides a new mechanism for regulation of microRNA activity. Nat. Genet. 2007, 39, 1033–1037. [Google Scholar] [CrossRef]
- Aukerman, M.J.; Sakai, H. Regulation of flowering time and floral organ identity by a microRNA and its APETALA2-like target genes. Plant Cell 2003, 15, 2730–2741. [Google Scholar] [CrossRef]
- Brodersen, P.; Sakvarelidze-Achard, L.; Bruun-Rasmussen, M.; Dunoyer, P.; Yamamoto, Y.Y.; Sieburth, L.; Voinnet, O. Widespread Translational Inhibition by Plant miRNAs and siRNAs. Science 2008, 320, 1185–1190. [Google Scholar] [CrossRef] [PubMed]
- Lanet, E.; Delannoy, E.; Sormani, R.; Floris, M.; Brodersen, P.; Crété, P.; Voinnet, O.; Robaglia, C. Biochemical Evidence for Translational Repression by Arabidopsis MicroRNAs. Plant Cell 2009, 21, 1762–1768. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Ma, Z.; Hu, L.; Huang, K.; Zhang, M.; Zhang, S.; Jiang, W.; Wu, T.; Du, X. Involvement of rice transcription factor OsERF19 in response to ABA and salt stress responses. Plant Physiol. Biochem. 2021, 167, 22–30. [Google Scholar] [CrossRef]
- Ma Z, Jin YM, Wu T, Hu L, Zhang Y, Jiang W, Du X. OsDREB2B, an AP2/ERF transcription factor, negatively regulates plant height by conferring GA metabolism in rice. Front Plant Sci. 2022, 13, 1007811.
- Singh, A.; Jain, D.; Pandey, J.; Yadav, M.; Bansal, K.C.; Singh, I.K. Deciphering the role of miRNA in reprogramming plant responses to drought stress. Crit. Rev. Biotechnol. 2022, 43, 613–627. [Google Scholar] [CrossRef] [PubMed]
- Arshad, M.; Gruber, M.Y.; Hannoufa, A. Transcriptome analysis of microRNA156 overexpression alfalfa roots under drought stress. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Feyissa BA, Arshad M, Gruber MY, Kohalmi SE, Hannoufa A. The interplay between miR156/SPL13 and DFR/WD40-1 regulate drought tolerance in alfalfa. BMC Plant Biol. 2019, 19, 434.
- López-Galiano, M.J.; García-Robles, I.; González-Hernández, A.I.; Camañes, G.; Vicedo, B.; Real, M.D.; Rausell, C. Expression of miR159 Is Altered in Tomato Plants Undergoing Drought Stress. Plants 2019, 8, 201. [Google Scholar] [CrossRef]
- Reyes, J.L.; Chua, N. ABA induction of miR159 controls transcript levels of two MYB factors during Arabidopsis seed germination. Plant J. 2007, 49, 592–606. [Google Scholar] [CrossRef]
- Zhang, M.; Chen, Y.; Xing, H.; Ke, W.; Shi, Y.; Sui, Z.; Xu, R.; Gao, L.; Guo, G.; Li, J.; et al. Positional cloning and characterization reveal the role of a miRNA precursor gene ZmLRT in the regulation of lateral root number and drought tolerance in maize. J. Integr. Plant Biol. 2022, 65, 772–790. [Google Scholar] [CrossRef]
- Kaushal, M. Microbes in Cahoots with Plants: MIST to Hit the Jackpot of Agricultural Productivity during Drought. Int. J. Mol. Sci. 2019, 20, 1769. [Google Scholar] [CrossRef] [PubMed]
- Romero-Romero JL, Inostroza-Blancheteau C, Orellana D, Aquea F, Reyes-Díaz M, Gil PM, Matte JP, Arce-Johnson P. Stomata regulation by tissue-specific expression of the Citrus sinensis MYB61 transcription factor improves water-use efficiency in Arabidopsis. Plant Physiol Biochem. 2018, 130, 54–60.
- Hoshika, Y.; Fares, S.; Pellegrini, E.; Conte, A.; Paoletti, E. Water use strategy affects avoidance of ozone stress by stomatal closure in Mediterranean trees—A modelling analysis. Plant, Cell Environ. 2019, 43, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Lertngim, N.; Ruangsiri, M.; Klinsawang, S.; Raksatikan, P.; Thunnom, B.; Siangliw, M.; Toojinda, T.; Siangliw, J.L. Photosynthetic Plasticity and Stomata Adjustment in Chromosome Segment Substitution Lines of Rice Cultivar KDML105 under Drought Stress. Plants 2022, 12, 94. [Google Scholar] [CrossRef]
- Yuan, W.; Suo, J.; Shi, B.; Zhou, C.; Bai, B.; Bian, H.; Zhu, M.; Han, N. The barley miR393 has multiple roles in regulation of seedling growth, stomatal density, and drought stress tolerance. Plant Physiol. Biochem. 2019, 142, 303–311. [Google Scholar] [CrossRef]
- Zhao J, Yuan S, Zhou M, Yuan N, Li Z, Hu Q, Bethea FG Jr, Liu H, Li S, Luo H. Transgenic creeping bentgrass overexpressing Osa-miR393a exhibits altered plant development and improved multiple stress tolerance. Plant Biotechnol J. 2019, 17, 233–251.
- Visentin I, Pagliarani C, Deva E, Caracci A, Turečková V, Novák O, Lovisolo C, Schubert A, Cardinale F. A novel strigolactone-miR156 module controls stomatal behaviour during drought recovery. Plant Cell Environ. 2020, 43, 1613–1624.
- Zhou Y, Liu W, Li X, Sun D, Xu K, Feng C, Kue Foka IC, Ketehouli T, Gao H, Wang N, Dong Y, Wang F, Li H. Integration of sRNA, degradome, transcriptome analysis and functional investigation reveals gma-miR398c negatively regulates drought tolerance via GmCSDs and GmCCS in transgenic Arabidopsis and soybean. BMC Plant Biol. 2020, 20, 190.
- Hang N, Shi T, Liu Y, Ye W, Taier G, Sun Y, Wang K, Zhang W. Overexpression of Os-microRNA408 enhances drought tolerance in perennial ryegrass. Physiol Plant. 2021, 172, 733–747.
- Wang, L. , Bai X. , Qiao Y., Si L., Yu Z., Ni C., Li T., Guo C., Xiao K. Tae-MiR9674a, a MicroRNA Member of Wheat, Confers Plant Drought and Salt Tolerance through Modulating the Stomata Movement and ROS Homeostasis. Plant Biotechnol Rep. 2022, 1, 3. [Google Scholar]
- Mir, I.R.; Rather, B.A.; Sehar, Z.; Masood, A.; Khan, N.A. Nitric oxide in co-ordination with nitrogen reverses cadmium-inhibited photosynthetic activity by interacting with ethylene synthesis, strengthening the antioxidant system, and nitrogen and sulfur assimilation in mustard (Brassica juncea L.). Sci. Hortic. 2023, 314. [Google Scholar] [CrossRef]
- Ma, Z.; Wu, T.; Huang, K.; Jin, Y.-M.; Li, Z.; Chen, M.; Yun, S.; Zhang, H.; Yang, X.; Chen, H.; et al. A Novel AP2/ERF Transcription Factor, OsRPH1, Negatively Regulates Plant Height in Rice. Front. Plant Sci. 2020, 11, 709. [Google Scholar] [CrossRef] [PubMed]
- Pegler JL, Oultram JMJ, Grof CPL, Eamens AL. Molecular Manipulation of the miR399/PHO2 Expression Module Alters the Salt Stress Response of Arabidopsis thaliana. Plants (Basel). 2020, 10, 73.
- Arif, M.A.; Top, O.; Csicsely, E.; Lichtenstern, M.; Beheshti, H.; Adjabi, K.; Frank, W. DICER-LIKE1a autoregulation based on intronic microRNA processing is required for stress adaptation in Physcomitrium patens. Plant J. 2021, 109, 227–240. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhou, J.X.; Hu, Y.F.; Fang, C.Y.; Yu, Y.J.; Yang, J.; Zhu, B.; Ruan, Y.L.; Zhu, Z.J. Overexpression of sly-miR398b increased salt sensitivity likely via regulating antioxidant system and photosynthesis in tomato. Environ. Exp. Bot. 2021, 181, 104273. [Google Scholar] [CrossRef]
- Liu, J.N.; Ma, X.; Yan, L.; Liang, Q.; Fang, H.; Wang, C.; Dong, Y.; Chai, Z.; Zhou, R.; Bao, Y.; et al. MicroRNA and Degradome Profiling Uncover Defense Response of Fraxinus velutina Torr. to Salt Stress. Front. Plant Sci. 2022, 13, 847853. [Google Scholar] [CrossRef]
- Yuan, S.; Zhao, J.; Li, Z.; Hu, Q.; Yuan, N.; Zhou, M.; Xia, X.; Noorai, R.; Saski, C.; Li, S.; et al. MicroRNA396-mediated alteration in plant development and salinity stress response in creeping bentgrass. Hortic. Res. 2019, 6, 1–13. [Google Scholar] [CrossRef]
- Abla, M.; Sun, H.; Li, Z.; Wei, C.; Gao, F.; Zhou, Y.; Feng, J. Identification of miRNAs and Their Response to Cold Stress in Astragalus Membranaceus. Biomolecules 2019, 9, 182. [Google Scholar] [CrossRef]
- Wang, S.-T.; Sun, X.-L.; Hoshino, Y.; Yu, Y.; Jia, B.; Sun, Z.-W.; Sun, M.-Z.; Duan, X.-B.; Zhu, Y.-M. MicroRNA319 Positively Regulates Cold Tolerance by Targeting OsPCF6 and OsTCP21 in Rice (Oryza sativa L.). PLOS ONE 2014, 9, e91357. [Google Scholar] [CrossRef]
- Zhou, M.; Tang, W. MicroRNA156 amplifies transcription factor-associated cold stress tolerance in plant cells. Mol. Genet. Genom. 2018, 294, 379–393. [Google Scholar] [CrossRef]
- Dong, Y.; Tang, M.; Huang, Z.; Song, J.; Xu, J.; Ahammed, G.J.; Yu, J.; Zhou, Y. The miR164a-NAM3 module confers cold tolerance by inducing ethylene production in tomato. Plant J. 2022, 111, 440–456. [Google Scholar] [CrossRef] [PubMed]
- Sun M, Shen Y, Chen Y, Wang Y, Cai X, Yang J, Jia B, Dong W, Chen X, Sun X. Osa-miR1320 targets the ERF transcription factor OsERF096 to regulate cold tolerance via JA-mediated signaling. Plant Physiol. 2022, 189, 2500–2516.
- de Lima, C.F.F.; Kleine-Vehn, J.; De Smet, I.; Feraru, E. Getting to the root of belowground high temperature responses in plants. J. Exp. Bot. 2021, 72, 7404–7413. [Google Scholar] [CrossRef]
- Cohen, S.P.; E Leach, J. High temperature-induced plant disease susceptibility: more than the sum of its parts. Curr. Opin. Plant Biol. 2020, 56, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Posch, B.C.; Kariyawasam, B.C.; Bramley, H.; Coast, O.; Richards, R.A.; Reynolds, M.P.; Trethowan, R.; Atkin, O.K. Exploring high temperature responses of photosynthesis and respiration to improve heat tolerance in wheat. J. Exp. Bot. 2019, 70, 5051–5069. [Google Scholar] [CrossRef]
- Sadok W, Lopez JR, Smith KP. Transpiration increases under high-temperature stress: Potential mechanisms, trade-offs and prospects for crop resilience in a warming world. Plant Cell Environ. 2021, 44, 2102–2116. [Google Scholar] [CrossRef]
- Sadura, I.; Janeczko, A. Brassinosteroids and the Tolerance of Cereals to Low and High Temperature Stress: Photosynthesis and the Physicochemical Properties of Cell Membranes. Int. J. Mol. Sci. 2021, 23, 342. [Google Scholar] [CrossRef]
- Singh, R.K.; Prasad, A.; Maurya, J.; Prasad, M. Regulation of small RNA-mediated high temperature stress responses in crop plants. Plant Cell Rep. 2021, 41, 765–773. [Google Scholar] [CrossRef]
- Wang, J.; Xu, J.; Wang, L.; Zhou, M.; Nian, J.; Chen, M.; Lu, X.; Liu, X.; Wang, Z.; Cen, J.; et al. SEMI-ROLLED LEAF 10 stabilizes catalase isozyme B to regulate leaf morphology and thermotolerance in rice (Oryza sativa L.). Plant Biotechnol. J. 2023, 21, 819–838. [Google Scholar] [CrossRef]
- Li, L.; Chen, G.; Yuan, M.; Guo, S.; Wang, Y.; Sun, J. CsbZIP2-miR9748-CsNPF4.4 Module Mediates High Temperature Tolerance of Cucumber Through Jasmonic Acid Pathway. Front. Plant Sci. 2022, 13, 883876. [Google Scholar] [CrossRef]
- Ahmed, W.; Xia, Y.; Zhang, H.; Li, R.; Bai, G.; Siddique, K.H.M.; Guo, P. Identification of conserved and novel miRNAs responsive to heat stress in flowering Chinese cabbage using high-throughput sequencing. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Matthews, C.; Arshad, M.; Hannoufa, A. Alfalfa response to heat stress is modulated by microRNA156. Physiol. Plant. 2018, 165, 830–842. [Google Scholar] [CrossRef]
- Arshad, M.; Puri, A.; Simkovich, A.J.; Renaud, J.; Gruber, M.Y.; Marsolais, F.; Hannoufa, A. Label-free quantitative proteomic analysis of alfalfa in response to microRNA156 under high temperature. BMC Genom. 2020, 21, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Arshad, M.; Hannoufa, A. Alfalfa transcriptome profiling provides insight into miR156-mediated molecular mechanisms of heat stress tolerance. Genome 2022, 65, 315–330. [Google Scholar] [CrossRef]
- Pandey, A.K.; Zorić, L.; Sun, T.; Karanović, D.; Fang, P.; Borišev, M.; Wu, X.; Luković, J.; Xu, P. The Anatomical Basis of Heavy Metal Responses in Legumes and Their Impact on Plant–Rhizosphere Interactions. Plants 2022, 11, 2554. [Google Scholar] [CrossRef] [PubMed]
- Gavrilescu, M. Enhancing phytoremediation of soils polluted with heavy metals. Curr. Opin. Biotechnol. 2021, 74, 21–31. [Google Scholar] [CrossRef]
- Chot, E.; Reddy, M.S. Role of Ectomycorrhizal Symbiosis Behind the Host Plants Ameliorated Tolerance Against Heavy Metal Stress. Front. Microbiol. 2022, 13, 855473. [Google Scholar] [CrossRef]
- Tighe-Neira, R.; Gonzalez-Villagra, J.; Nunes-Nesi, A.; Inostroza-Blancheteau, C. Impact of nanoparticles and their ionic counterparts derived from heavy metals on the physiology of food crops. Plant Physiol. Biochem. 2022, 172, 14–23. [Google Scholar] [CrossRef]
- Sharma, A.; Kapoor, D.; Gautam, S.; Landi, M.; Kandhol, N.; Araniti, F.; Ramakrishnan, M.; Satish, L.; Singh, V.P.; Sharma, P.; et al. Heavy metal induced regulation of plant biology: Recent insights. Physiol. Plant. 2022, 174, e13688. [Google Scholar] [CrossRef]
- Velusamy, K.; Periyasamy, S.; Kumar, P.S.; Rangasamy, G.; Pauline, J.M.N.; Ramaraju, P.; Mohanasundaram, S.; Vo, D.-V.N. Biosensor for heavy metals detection in wastewater: A review. Food Chem. Toxicol. 2022, 168, 113307. [Google Scholar] [CrossRef]
- Vaid, N.; Sudan, J.; Dave, S.; Mangla, H.; Pathak, H. Insight Into Microbes and Plants Ability for Bioremediation of Heavy Metals. Curr. Microbiol. 2022, 79, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Vega, A.; Delgado, N.; Handford, M. Increasing Heavy Metal Tolerance by the Exogenous Application of Organic Acids. Int. J. Mol. Sci. 2022, 23, 5438. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ding, H.; Jiang, H.; Wang, H.; Chen, K.; Duan, J.; Feng, S.; Wu, G. Regulation of cadmium tolerance and accumulation by miR156 in Arabidopsis. Chemosphere 2019, 242, 125168. [Google Scholar] [CrossRef]
- Kumar, R.S.; Sinha, H.; Datta, T.; Asif, M.H.; Trivedi, P.K. microRNA408 and its encoded peptide regulate sulfur assimilation and arsenic stress response in Arabidopsis. Plant Physiol. 2023, 192, 837–856. [Google Scholar] [CrossRef] [PubMed]
- Nie G, Liao Z, Zhong M, Zhou J, Cai J, Liu A, Wang X, Zhang X. MicroRNA-Mediated Responses to Chromium Stress Provide Insight Into Tolerance Characteristics of Miscanthus sinensis. Front Plant Sci. 2021, 12, 666117.
- Zhou, M.; Zheng, S.; Liu, R.; Lu, L.; Zhang, C.; Zhang, L.; Yant, L.; Wu, Y. The genome-wide impact of cadmium on microRNA and mRNA expression in contrasting Cd responsive wheat genotypes. BMC Genom. 2019, 20, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Bai, B.; Bian, H.; Zeng, Z.; Hou, N.; Shi, B.; Wang, J.; Zhu, M.; Han, N. miR393-Mediated Auxin Signaling Regulation is Involved in Root Elongation Inhibition in Response to Toxic Aluminum Stress in Barley. Plant Cell Physiol. 2017, 58, 426–439. [Google Scholar] [CrossRef]
- Zinta, R.; Tiwari, J.K.; Buckseth, T.; Thakur, K.; Goutam, U.; Kumar, D.; Challam, C.; Bhatia, N.; Poonia, A.K.; Naik, S.; et al. Root system architecture for abiotic stress tolerance in potato: Lessons from plants. Front. Plant Sci. 2022, 13, 926214. [Google Scholar] [CrossRef]
- Phour, M.; Sindhu, S.S. Mitigating abiotic stress: microbiome engineering for improving agricultural production and environmental sustainability. Planta 2022, 256, 1–34. [Google Scholar] [CrossRef]
- Prasad, R. Cytokinin and Its Key Role to Enrich the Plant Nutrients and Growth Under Adverse Conditions-An Update. Front. Genet. 2022, 13, 883924. [Google Scholar] [CrossRef]
- Swain, R.; Sahoo, S.; Behera, M.; Rout, G.R. Instigating prevalent abiotic stress resilience in crop by exogenous application of phytohormones and nutrient. Front. Plant Sci. 2023, 14, 1104874. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Singh, R.K.; Li, H.-B.; Guo, D.-J.; Sharma, A.; Verma, K.K.; Solanki, M.K.; Upadhyay, S.K.; Lakshmanan, P.; Yang, L.-T.; et al. Nitrogen fixation and phytohormone stimulation of sugarcane plant through plant growth promoting diazotrophic Pseudomonas. Biotechnol. Genet. Eng. Rev. 2023, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Jezek M, Allan AC, Jones JJ, Geilfus CM. New Phytol. 2023.
- Johnson, R.; Vishwakarma, K.; Hossen, S.; Kumar, V.; Shackira, A.; Puthur, J.T.; Abdi, G.; Sarraf, M.; Hasanuzzaman, M. Potassium in plants: Growth regulation, signaling, and environmental stress tolerance. Plant Physiol. Biochem. 2022, 172, 56–69. [Google Scholar] [CrossRef]
- Lyzenga, W.J.; Liu, Z.; Olukayode, T.; Zhao, Y.; Kochian, L.V.; Ham, B.-K. Getting to the roots of N, P, and K uptake. J. Exp. Bot. 2023, 74, 1784–1805. [Google Scholar] [CrossRef]
- Yousuf, P.Y.; Shabir, P.A.; Hakeem, K.R. miRNAomic Approach to Plant Nitrogen Starvation. Int. J. Genom. 2021, 2021, 1–14. [Google Scholar] [CrossRef]
- Vega, A.; O’brien, J.A.; A Gutiérrez, R. Nitrate and hormonal signaling crosstalk for plant growth and development. Curr. Opin. Plant Biol. 2019, 52, 155–163. [Google Scholar] [CrossRef]
- Islam, W.; Tauqeer, A.; Waheed, A.; Zeng, F. MicroRNA Mediated Plant Responses to Nutrient Stress. Int. J. Mol. Sci. 2022, 23, 2562. [Google Scholar] [CrossRef]
- Du, Q.; Wang, K.; Zou, C.; Xu, C.; Li, W.-X. The PILNCR1-miR399 Regulatory Module Is Important for Low Phosphate Tolerance in Maize. Plant Physiol. 2018, 177, 1743–1753. [Google Scholar] [CrossRef]
- Hu, B.; Wang, W.; Deng, K.; Li, H.; Zhang, Z.; Zhang, L.; Chu, C. MicroRNA399 is involved in multiple nutrient starvation responses in rice. Front. Plant Sci. 2015, 6, 188. [Google Scholar] [CrossRef]
- Thornburg TE, Liu J, Li Q, Xue H, Wang G, Li L, Fontana JE, Davis KE, Liu W, Zhang B, Zhang Z, Liu M, Pan X. Potassium Deficiency Significantly Affected Plant Growth and Development as Well as microRNA-Mediated Mechanism in Wheat (Triticum aestivum L.). Front Plant Sci. 2020, 11, 1219.
- Fontana JE, Wang G, Sun R, Xue H, Li Q, Liu J, Davis KE, Thornburg TE, Zhang B, Zhang Z, Pan X. Impact of potassium deficiency on cotton growth, development and potential microRNA-mediated mechanism. Plant Physiol Biochem. 2020, 153, 72–80.
- Ye, Z.; Zeng, J.; Long, L.; Ye, L.; Zhang, G. Identification of microRNAs in response to low potassium stress in the shoots of Tibetan wild barley and cultivated. Curr. Plant Biol. 2020, 25, 100193. [Google Scholar] [CrossRef]
- Yan, Y.; Wang, H.; Hamera, S.; Chen, X.; Fang, R. miR444a has multiple functions in the rice nitrate-signaling pathway. Plant J. 2014, 78, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Prusty, S.; Sahoo, R.K.; Nayak, S.; Poosapati, S.; Swain, D.M. Proteomic and Genomic Studies of Micronutrient Deficiency and Toxicity in Plants. Plants 2022, 11, 2424. [Google Scholar] [CrossRef] [PubMed]
- Ninkuu, V.; Liu, Z.; Sun, X. Genetic regulation of nitrogen use efficiency in Gossypium spp. Plant, Cell Environ. 2023, 46, 1749–1773. [Google Scholar] [CrossRef]
- Huang, S.; Wang, P.; Yamaji, N.; Ma, J.F. Plant Nutrition for Human Nutrition: Hints from Rice Research and Future Perspectives. Mol. Plant 2020, 13, 825–835. [Google Scholar] [CrossRef] [PubMed]
- Robinson, R.S.; Smart, S.M.; Cybulski, J.D.; McMahon, K.W.; Marcks, B.; Nowakowski, C. Insights from Fossil-Bound Nitrogen Isotopes in Diatoms, Foraminifera, and Corals. Annu. Rev. Mar. Sci. 2023, 15, 407–430. [Google Scholar] [CrossRef]
- Helliwell, K.E. Emerging trends in nitrogen and phosphorus signalling in photosynthetic eukaryotes. Trends Plant Sci. 2022, 28, 344–358. [Google Scholar] [CrossRef]
- Kong, W.W.; Yang, Z.M. Identification of iron-deficiency responsive microRNA genes and cis-elements in Arabidopsis. Plant Physiol. Biochem. 2010, 48, 153–159. [Google Scholar] [CrossRef]
- Valdés-López, O.; Yang, S.S.; Aparicio-Fabre, R.; Graham, P.H.; Reyes, J.L.; Vance, C.P.; Hernández, G. MicroRNA expression profile in common bean (Phaseolus vulgaris) under nutrient deficiency stresses and manganese toxicity. New Phytol. 2010, 187, 805–818. [Google Scholar] [CrossRef] [PubMed]
- Kayihan, D.S.; Kayihan, C.; Özden Çiftçi, Y. Moderate level of toxic boron causes differential regulation of micrornas related to jasmonate and ethylene metabolisms in arabidopsis thaliana. Turk. J. Botany. 2019, 43, 167–172. [Google Scholar] [CrossRef]
- Ozhuner, E.; Eldem, V.; Ipek, A.; Okay, S.; Sakcali, S.; Zhang, B.; Boke, H.; Unver, T. Boron Stress Responsive MicroRNAs and Their Targets in Barley. PLOS ONE 2013, 8, e59543. [Google Scholar] [CrossRef]
- Giacomelli, J.I.; Weigel, D.; Chan, R.L.; Manavella, P.A. Role of recently evolved miRNA regulation of sunflower HaWRKY6 in response to temperature damage. New Phytol. 2012, 195, 766–773. [Google Scholar] [CrossRef]
- Zhang, N.; Yang, J.; Wang, Z.; Wen, Y.; Wang, J.; He, W.; Liu, B.; Si, H.; Wang, D. Identification of Novel and Conserved MicroRNAs Related to Drought Stress in Potato by Deep Sequencing. PLOS ONE 2014, 9, e95489. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.-P.; Montgomery, T.A.; Fahlgren, N.; Kasschau, K.D.; Nonogaki, H.; Carrington, J.C. Repression of AUXIN RESPONSE FACTOR10 by microRNA160 is critical for seed germination and post-germination stages. Plant J. 2007, 52, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Boualem, A.; Laporte, P.; Jovanovic, M.; Laffont, C.; Plet, J.; Combier, J.; Niebel, A.; Crespi, M.; Frugier, F. MicroRNA166 controls root and nodule development in Medicago truncatula. Plant J. 2008, 54, 876–887. [Google Scholar] [CrossRef]
- Trindade, I.; Capitão, C.; Dalmay, T.; Fevereiro, M.P.; dos Santos, D.M. miR398 and miR408 are up-regulated in response to water deficit in Medicago truncatula. Planta 2009, 231, 705–716. [Google Scholar] [CrossRef]
- Li, W.-X.; Oono, Y.; Zhu, J.; He, X.-J.; Wu, J.-M.; Iida, K.; Lu, X.-Y.; Cui, X.; Jin, H.; Zhu, J.-K. The Arabidopsis NFYA5 Transcription Factor Is Regulated Transcriptionally and Posttranscriptionally to Promote Drought Resistance. Plant Cell 2008, 20, 2238–2251. [Google Scholar] [CrossRef]
- Jagadeeswaran, G.; Li, Y.; Sunkar, R. Redox signaling mediates the expression of a sulfate-deprivation-inducible microRNA395 in Arabidopsis. Plant J. 2013, 77, 85–96. [Google Scholar] [CrossRef]
- E Barrera-Figueroa, B.; Gao, L.; Diop, N.N.; Wu, Z.; Ehlers, J.D.; A Roberts, P.; Close, T.J.; Zhu, J.-K.; Liu, R. Identification and comparative analysis of drought-associated microRNAs in two cowpea genotypes. BMC Plant Biol. 2011, 11, 127–127. [Google Scholar] [CrossRef] [PubMed]
- Sunkar, R.; Zhu, J.-K. Novel and Stress-Regulated MicroRNAs and Other Small RNAs from Arabidopsis[W]. Plant Cell 2004, 16, 2001–2019. [Google Scholar] [CrossRef]
- Liu, H.-H.; Tian, X.; Li, Y.-J.; Wu, C.-A.; Zheng, C.-C. Microarray-based analysis of stress-regulated microRNAs in Arabidopsis thaliana. RNA 2008, 14, 836–843. [Google Scholar] [CrossRef]
- Shamimuzzaman; Vodkin, L. Identification of soybean seed developmental stage-specific and tissue-specific miRNA targets by degradome sequencing. BMC Genom. 2012, 13, 310–310. [Google Scholar] [CrossRef] [PubMed]
- Szabados, L.; Savouré, A. Proline: a multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Tao, Y.; Zhu, C. Emerging roles of microRNAs in the mediation of drought stress response in plants. J. Exp. Bot. 2013, 64, 3077–3086. [Google Scholar] [CrossRef]
- Liu, Y.; Li, D.; Yan, J.; Wang, K.; Luo, H.; Zhang, W. MiR319 mediated salt tolerance by ethylene. Plant Biotechnol. J. 2019, 17, 2370–2383. [Google Scholar] [CrossRef]
- He F, Xu C, Fu X, Shen Y, Guo L, Leng M, Luo K. The MicroRNA390/TRANS-ACTING SHORT INTERFERING RNA3 Module Mediates Lateral Root Growth under Salt Stress via the Auxin Pathway. Plant Physiol. 2018, 177, 775–791.
- Bai, Q.; Wang, X.; Chen, X.; Shi, G.; Liu, Z.; Guo, C.; Xiao, K. Wheat miRNA TaemiR408 Acts as an Essential Mediator in Plant Tolerance to Pi Deprivation and Salt Stress via Modulating Stress-Associated Physiological Processes. Front. Plant Sci. 2018, 9, 499. [Google Scholar] [CrossRef]
- Guo, X.; Niu, J.; Cao, X. Heterologous Expression of Salvia miltiorrhiza MicroRNA408 Enhances Tolerance to Salt Stress in Nicotiana benthamiana. Int. J. Mol. Sci. 2018, 19, 3985. [Google Scholar] [CrossRef]
- Wang, W.; Liu, D.; Chen, D.; Cheng, Y.; Zhang, X.; Song, L.; Hu, M.; Dong, J.; Shen, F. MicroRNA414c affects salt tolerance of cotton by regulating reactive oxygen species metabolism under salinity stress. RNA Biol. 2019, 16, 362–375. [Google Scholar] [CrossRef] [PubMed]
- Aslam M, Sugita K, Qin Y, Rahman A. Aux/IAA14 Regulates microRNA-Mediated Cold Stress Response in Arabidopsis Roots. Int J Mol Sci. 2020, 21, 8441.
- Huo, C.; Zhang, B.; Wang, R. Research progress on plant noncoding RNAs in response to low-temperature stress. Plant Signal. Behav. 2021, 17, 2004035. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Zhang, N.; Wang, Q.; Fu, Y.; Wang, F.; Su, Y.; Xue, B.; Zhou, L.; Liao, H. The Effect of Low Temperature Stress on the Leaves and MicroRNA Expression of Potato Seedlings. Front. Ecol. Evol. 2021, 9. [Google Scholar] [CrossRef]
- Stief, A.; Altmann, S.; Hoffmann, K.; Pant, B.D.; Scheible, W.-R.; Bäurle, I. Arabidopsis miR156 Regulates Tolerance to Recurring Environmental Stress through SPL Transcription Factors. Plant Cell 2014, 26, 1792–1807. [Google Scholar] [CrossRef] [PubMed]
- Zhang M, An P, Li H, Wang X, Zhou J, Dong P, Zhao Y, Wang Q, Li C. The miRNA-Mediated Post-Transcriptional Regulation of Maize in Response to High Temperature. Int J Mol Sci. 2019, 20, 1754.
- Gong, J.; Li, D.; Li, H.; Zhou, H.; Xu, J. Identification of manganese-responsive microRNAs in Arabidopsis by small RNA sequencing. Czech J. Genet. Plant Breed. 2019, 55, 76–82. [Google Scholar] [CrossRef]
- da Silva, R.G.; Rosa-Santos, T.M.; França, S.d.C.; Kottapalli, P.; Kottapalli, K.R.; Zingaretti, S.M. Microtranscriptome analysis of sugarcane cultivars in response to aluminum stress. PLOS ONE 2019, 14, e0217806. [Google Scholar] [CrossRef]
- Shi, D.-Q.; Zhang, Y.; Ma, J.-H.; Li, Y.-L.; Xu, J. Identification of Zinc Deficiency-Responsive MicroRNAs in Brassica juncea Roots by Small RNA Sequencing. J. Integr. Agric. 2013, 12, 2036–2044. [Google Scholar] [CrossRef]
- Halder, K.; Chaudhuri, A.; Abdin, M.Z.; Datta, A. Tweaking the Small Non-Coding RNAs to Improve Desirable Traits in Plant. Int. J. Mol. Sci. 2023, 24, 3143. [Google Scholar] [CrossRef]
- Yan, G.; Hua, Y.; Jin, H.; Huang, Q.; Zhou, G.; Xu, Y.; He, Y.; Zhu, Z. Sly-miR398 Participates in Cadmium Stress Acclimation by Regulating Antioxidant System and Cadmium Transport in Tomato (Solanum lycopersicum). Int J Mol Sci. 2023, 24, 1953. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The origin, biogenesis of miRNAs in the plant: (a) RNA polymerase II in most cases and RNA polymerase III in a few cases transcribe from genomic sequences to form the precursor pri-miRNA. In the nuclear pri-miRNA is edited by the adenylate deaminase of RNA and forms pre-miRNA hairpin sequences after being sheared by the microprocessor formed by the enzyme. (b) The nucleoplasmic transporter protein recognises the two nucleotides protruding from the 3’ end of the pre-miRNA hairpin sequence and transports the pre-miRNA from the nucleus to the cytoplasm. The nuclease begins to shear the stem-loop structure in the hairpin sequence, after which it is able to produce the miRNA duplex. The pre-miRNA precursor is processed into a mature double-stranded miRNA/miRNA complex by processing the miRNA/miRNA double-stranded body and then the double-stranded body translocates with the help of a protein helper complex. When the mature miRNA is embedded in the RNA-induced silencing complex, the antisense miRNA is degraded in the cytoplasm. The ribonucleoprotein complex miRNP negatively regulates the expression of target genes at the post-transcriptional level through the interaction of its cognate mature miRNA with the target mRNA.
Figure 1.
The origin, biogenesis of miRNAs in the plant: (a) RNA polymerase II in most cases and RNA polymerase III in a few cases transcribe from genomic sequences to form the precursor pri-miRNA. In the nuclear pri-miRNA is edited by the adenylate deaminase of RNA and forms pre-miRNA hairpin sequences after being sheared by the microprocessor formed by the enzyme. (b) The nucleoplasmic transporter protein recognises the two nucleotides protruding from the 3’ end of the pre-miRNA hairpin sequence and transports the pre-miRNA from the nucleus to the cytoplasm. The nuclease begins to shear the stem-loop structure in the hairpin sequence, after which it is able to produce the miRNA duplex. The pre-miRNA precursor is processed into a mature double-stranded miRNA/miRNA complex by processing the miRNA/miRNA double-stranded body and then the double-stranded body translocates with the help of a protein helper complex. When the mature miRNA is embedded in the RNA-induced silencing complex, the antisense miRNA is degraded in the cytoplasm. The ribonucleoprotein complex miRNP negatively regulates the expression of target genes at the post-transcriptional level through the interaction of its cognate mature miRNA with the target mRNA.
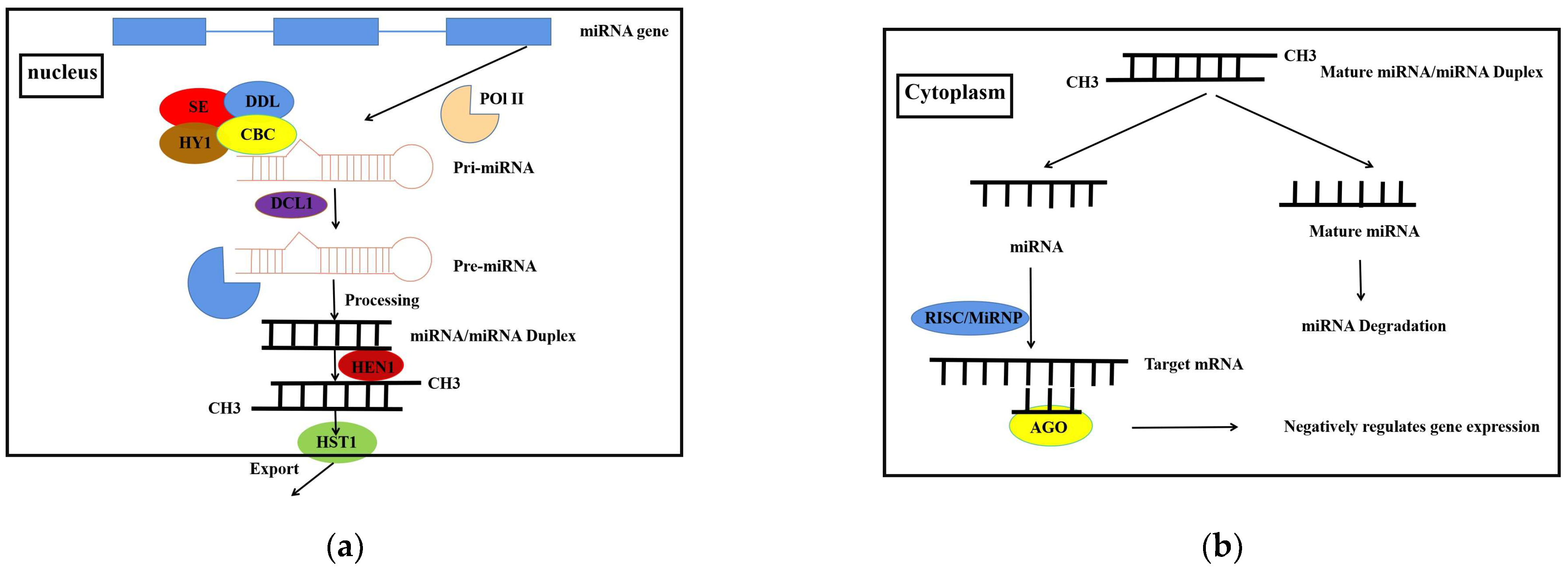
Figure 2.
The mechanism of miRNAs in the plant: (a) After the plant miRNA recognizes and binds to the target mRNA, the AGO will shear the target mRNA at the 10th and 11th nucleotides of the miRNA binding site. When plant miRNAs bind to target mRNAs, the AGO will shear the target mRNA of the miRNA binding site. (b) Another mechanism of plant miRNAs is through translational repression. The miRNA does not bind to the 3′ UTR region of the target mRNA in its entirety and the RNA-induced silencing complex inhibits the initiation of translation or the specific degradation of the synthetic ribosome to achieve translation inhibition.
Figure 2.
The mechanism of miRNAs in the plant: (a) After the plant miRNA recognizes and binds to the target mRNA, the AGO will shear the target mRNA at the 10th and 11th nucleotides of the miRNA binding site. When plant miRNAs bind to target mRNAs, the AGO will shear the target mRNA of the miRNA binding site. (b) Another mechanism of plant miRNAs is through translational repression. The miRNA does not bind to the 3′ UTR region of the target mRNA in its entirety and the RNA-induced silencing complex inhibits the initiation of translation or the specific degradation of the synthetic ribosome to achieve translation inhibition.
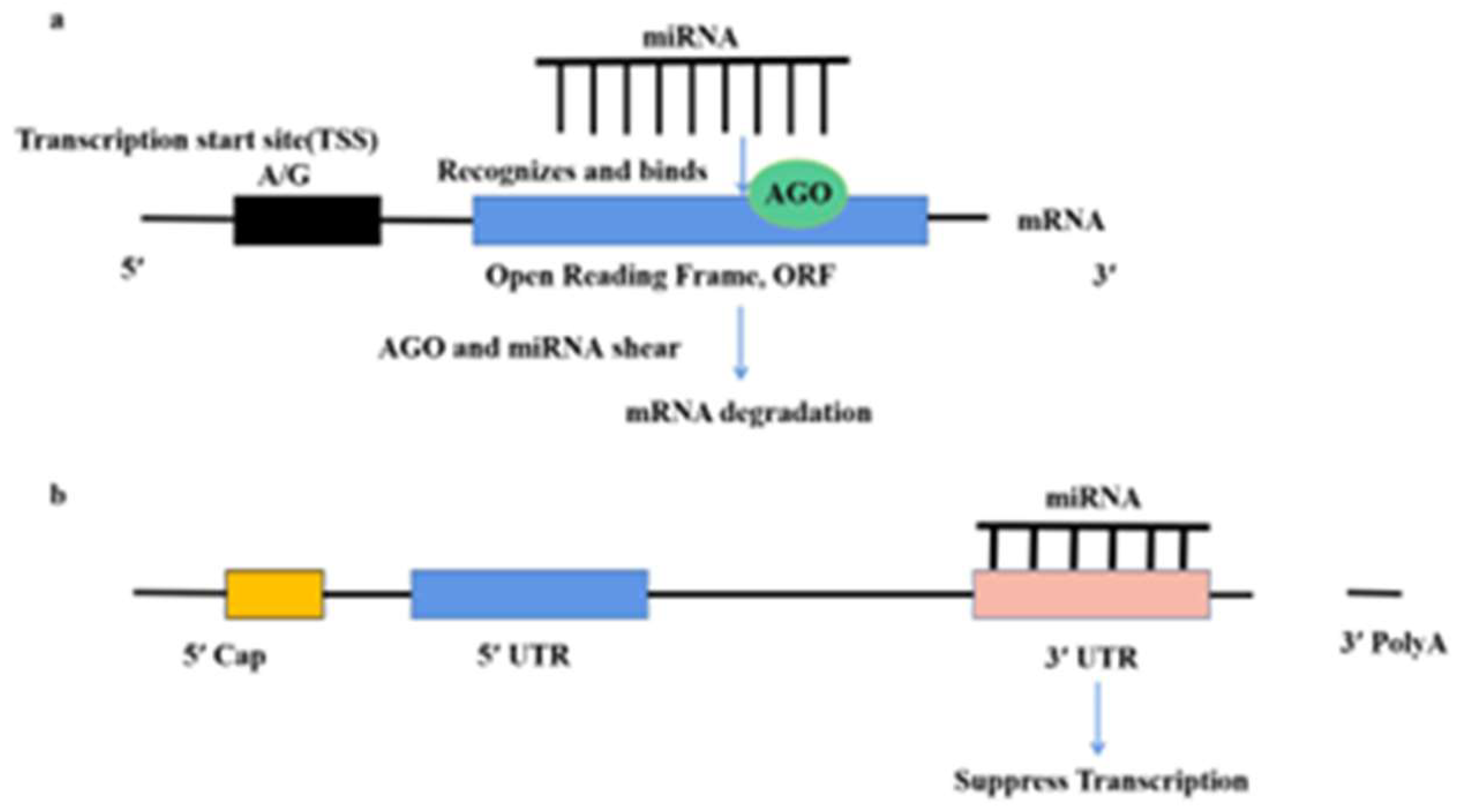

Table 1.
Abiotic stress responsive miRNAs: their regulations and target genes in plants.
| Abiotic Stress Type | miRNA | Expression | Species | Target Genes | Reference |
|---|---|---|---|---|---|
| Drought | MicroRNA-157 | Upregulated | Arabidopsis thaliana | SPB Transcription factor | [94] |
| Drought | MicroRNA-159 | Upregulated | Arabidopsis thaliana | MYB and TCP Transcription factors | [95] |
| Drought | MicroRNA-160 | Downregulated | Arabidopsis thaliana | ARF10, ARF16, ARF17 | [96] |
| Drought | MicroRNA-166 | Upregulated | Medicago truncatula | HD-ZIPIII Transcription factors | [97,98] |
| Drought | MicroRNA-167 | Upregulated | Arabidopsis thaliana | ARF6,ARF8 | [94] |
| Drought | MicroRNA-168 | Upregulated | Arabidopsis thaliana | ARGONAUTE, MAPK | [94] |
| Drought | MicroRNA-169 | Downregulated | Arabidopsis thaliana | NF-YA transcription factor, SIMRP1 | [99] |
| Drought | MicroRNA-171 | Upregulated | Arabidopsis thaliana | GRAS transcription factor | [94] |
| Drought | MicroRNA-319 | Upregulated | Arabidopsis thaliana | TCP Family | [100] |
| Drought | MicroRNA-390 | Upregulated | Vigna unguiculata | ARF Family | [101] |
| Drought | MicroRNA-393 | Upregulated | Arabidopsis thaliana | (TIR1, AFB2, AFB3) (ARF5, EPF1, SPCH) | [102,103] |
| Drought | MicroRNA-396 | Upregulated | Arabidopsis thaliana | GRL transcription factor | [94] |
| Drought | MicroRNA-397 | Downregulated | Oryza sativa | Laccase genes | [104] |
| Drought | MicroRNA-398 | Upregulated | Medicago truncatula | Superoxide dismutase | [98] |
| Drought | MicroRNA-398c | Downregulated | Soybean | GmCSD1a/b,GmCSD2a/b/c,GmCCS | [28] |
| Drought | MicroRNA-408 | Upregulated | Arabidopsis thaliana | Chemocyanin precursor, kinases | [94] |
| Drought | MicroRNA-474 | Upregulated | Zea mays | PDH, PPR | [105] |
| Drought | MicroRNA-528 | Downregulated | Zea mays | POD | [105] |
| Drought | MicroRNA-811 | Downregulated | Catharanthus roseus | MYB transcription factor | [106] |
| Drought | MicroRNA-814 | Downregulated | Phaseolus vulgaris | Hydroxyproline-rich glycoprotein | [106] |
| Drought | MicroRNA-835 | Downregulated | Ricinus communis | Aquaporin | [106] |
| Drought | MicroRNA-4398 | Downregulated | Solanum tuberosum | WRKY transcription factor | [106] |
| Salt | MicroRNA-319b | Upregulated | Switchgrass | PvPCF5 | [107] |
| Salt | MicroRNA-390 | Downregulated | Poplar | ARF3.1, ARF3.2,ARF4 | [108] |
| Salt | MicroRNA-390a | Downregulated | Creeping bentgrass | AsTIR1, AsAFB2 | [26] |
| Salt | MicroRNA-396c | Upregulated | Creeping bentgrass | GRF | [37] |
| Salt | MicroRNA-408 | Upregulated | Wheat | TaCP,TaMP,TaBCP,TaFP,TaKRP,TaABP | [109] |
| Salt | MicroRNA-408 | Upregulated | Salvia miltiorrhiza | NbSOD, NbPOD, NbCAT | [110] |
| Salt | MicroRNA-414c | Downregulated | Cotton | GhFSD1 | [111] |
| Cold | MicroRNA-160 | Downregulated | Maize | [112] | |
| Cold | MicroRNA-319 | Downregulated | Rice | PCF6/TCP21 | [113] |
| Cold | MicroRNA-319 | Downregulated | Maize | [112] | |
| Cold | MicroRNA-408a | Upregulated | Maize | [112] | |
| Cold | MicroRNA-528 | Upregulated | Maize | [112] | |
| Cold | MicroRNA-5125 | Upregulated | Potato | ABF8011 | [114] |
| Cold | MicroRNA-10881 | Upregulated | Potato | GA3ox123158 | [114] |
| High temperature | MicroRNA-156 | Downregulated | Arabidopsis thaliana | SPL transcription factor | [115] |
| High temperature | MicroRNA-159 | Downregulated | Maize | MYB transcription factor | [116] |
| High temperature | MicroRNA-164 | Downregulated | Maize | NAC transcription factor | [116] |
| High temperature | MicroRNA-166 | Downregulated | Maize | HD zip | [116] |
| High temperature | MicroRNA-169 | Downregulated | Maize | SBP | [116] |
| High temperature | MicroRNA-172 | Downregulated | Maize | AP2/ERF | [116] |
| High temperature | MicroRNA-396 | Downregulated | Maize | GRF, | [116] |
| High temperature | MicroRNA-5381 | Downregulated | Maize | SAC2 | [116] |
| Heavy metals-Cd | MicroRNA-167 | Zea mays | [117] | ||
| Heavy metals-Cd | MicroRNA-393 | Zea mays | [117] | ||
| Heavy metals-Cu | MicroRNA-398 | Grape | VvCSD1 and VvCSD2 | [63] | |
| Heavy metals-Al | MicroRNA-160 | Sugarcance | [117] | ||
| Heavy metals-Al | MicroRNA-162 | Sugarcance | [117] | ||
| Heavy metals-Al | MicroRNA-164 | Sugarcance | [117] | ||
| Heavy metals-Al | MicroRNA-166 | Sugarcance | [117] | ||
| Heavy metals-Al | MicroRNA-167 | Sugarcance | [117] | ||
| Nutrients-Zn | MicroRNA-158 | Upregulated | Brassica juncea | FUT1 | [118] |
| Nutrients-K | MicroRNA-169 | Triticum aestivum | Pentose pathway | [119] | |
| Nutrients-N | MicroRNA-169 | Downregulated | Arabidopsis thaliana | HAP2 | [81] |
| Nutrients-B | MicroRNA-319 | Upregulated | Riticum aestivum | MYB transcription factor | [92] |
| Nutrients-K | MicroRNA-319 | Downregulated | Hordeum vulgare | TCP | [92] |
| Nutrients-K | MicroRNA-396 | Downregulated | Hordeum vulgare | GRF | [83] |
| Nutrients-P | MicroRNA-399 | Downregulated | Arabidopsis thaliana | Ubiquitin conjugase E2 | [117] |
| Nutrients-Mn | MicroRNA-781 | Upregulated | Arabidopsis thaliana | MCM2 | [117] |
| Nutrients-Mn | MicroRNA-826 | Upregulated | Arabidopsis thaliana | Alkenylhydroxalkylproducing 2 | [117] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



