
Submitted:
25 May 2023
Posted:
26 May 2023
You are already at the latest version
Abstract
High-quality rice (Oryza sativa L.) is increasingly widely planted in China with the improvement of people's living standards and the achievement of rice breeding efforts in recent years. However, the agronomic and physiological performances of high-quality indica rice (HQIR) under different nitrogen(N) application conditions in southern China are little known. Two-year consecutive field experiments were conducted with two HQIR and two ordinary-quality indica rice (OQIR) varieties under moderate and high N application rates, with yield and yield components, biomass, N up-take, and their related traits investigated. We found that grain yields of HQIR were slightly de-creased, but grain yields of OQIR were significantly increased by 6.60%-8.88% under high N rate compared with moderate N rate in both years. Thereby, OQIR produced 8.34%-11.87% and 22.00%-22.50% higher grain yield than HQIR under moderate and high N rates, respectively. The different responses of grain yield to N application rates were mainly due to decreased grain set-ting rate in HQIR and increased spikelets m-2 in OQIR under high N rate. Furthermore, high N rate significantly reduced pre-anthesis AE and improved grain-leaf area ratio, while did not in-crease post-anthesis DM compared with moderate N rate in HQIR, which might result in carbon metabolic deterioration and imbalance of source-sink relationship and subsequently lower supply of carbohydrate to panicle. Our results suggest that a moderate N rate (165 kg N ha-1) is beneficial for HQIR varieties to balance the maximum grain yield and high quality in southern China
Keywords:
grain yield
; biomass
; N application rate
; high-quality rice
1. Introduction
Rice (Oryza sativa L.) is a vital important staple food for more than 60% of the population and contributes nearly 40% of the people’s total calorie intake in China [1]. Over the past several decades, the high-yielding traits were always regarded as the principal targets for rice breeding and selecting to meet the population growth of China [2]. For instance, with the development of hybrid rice in 1976 and “super” rice in 1996 in China, the newly released rice varieties which have higher canopy photosynthesis and larger sink size showed about 10% higher yield potential than their check varieties [1,2,3]. Correspondingly, the national total rice yield and average rice yield have increased 3.09 Mt year−1 and 0.127 t ha−1 year−1 from 1976–1995, and 1.26 Mt year−1 and 0.039 t ha−1 year−1 from 1996–2020, respectively, in despite of the national total rice production area showing a decreasing trend (FAOSTAT, 2020) [4]. These achievements have contributed much to national food self-sufficiency and food security.
Presently, with the development of social economy and the continuous improvement of living standards, the consumer demand for rice food has shifted from quantity to high-quality in China [5,6]. However, there has long been a contradiction in achieving high yield and superior quality simultaneously for rice breeders [7,8]. Most high-yielding hybrid indica rice varieties generally show poor rice quality, like higher chalkiness degree, lower cooking and eating quality [9,10]. The reasons for this outcome are that most rice yield- or quality-related traits are quantitative, and yield and quality are generally negatively correlated with each other [11,12,13]. Hence, it is difficult to develop new elite rice varieties with both high yield and superior quality using traditional breeding approaches [13,14]. Fortunately, based on technological innovation of rice breeding (i.e., molecular marker-assisted breeding, molecular design breeding), the improvement of quality of high-yielding rice varieties has been increasingly highlighted in breeding efforts and more and more attentions have been paid on high-quality rice breeding [9,13,14]. So far, over 50% varieties released by province and state have reached the national and ministerial standards of high-quality rice since 2017, which greatly accelerates the popularizing rate of high-quality varieties in rice production in China [15]. Particularly in late-season rice (grown from July to November) of double-rice cropping system of southern China, more than 80% of the late rice varieties planted by farmers have high quality as the optimal temperature in the late-season is conducive to the formation of rice quality.
In comparison with the ordinary-quality rice, the high-quality rice varieties usually have lower yield potential but higher price per unit yield and overall benefits [6,8]. As we all known, nitrogen (N) input plays a vital role in rice production and has significant effects on rice yield and quality. To get more returns, however, rice farmer unrealistically applied a large amount of nitrogen (N) fertilizer to realize higher yield of high-quality rice varieties [16]. Many studies indicated that the overuse of N fertilizer was prevalent in rice production of China, subsequently leading to rice lodging, increased pests and diseases, low N use efficiency, and higher environmental costs [17,18]. In fact, potential yields of most high-yielding rice varieties do not depend on greater N fertilizer input under moderate and high soil fertility conditions [19,20]. On the other hand, excessive N fertilizer input may increase grain protein content and alter starch properties, resulting in remarkable decline in cooking and eating quality for high-quality rice [21,22,23]. Previous studies showed that optimizing N management, such as reducing total N and/or late-stage N application rate, is beneficial to balance rice yield and quality [16,22]. Most results indicated that the rational N application rate of simultaneously obtaining high yield and superior quality varied with varieties and was 180–270 kg N ha−1 for high-quality japonica rice (HQJR) [21,22,23], but such rational N application rate was 135–165 kg N ha−1 for high-quality indica rice (HQIR) [24,25,26], which is close to the N application rate of producing maximum yield in ordinary-quality indica rice (OQIR) [20,27].
However, the abovementioned studies focused more on the effect of N management on balancing rice yield and quality. The agronomic and physiological responses of yield formation to N application rates are not detailed in HQIR varieties. In this study, we hypothesized that HQIR and OQIR had similar responses in terms of yield formation under different N application rates. Therefore, to verify the hypothesis, two-year field experiments were carried out with two HQIR and two OQIR varieties under moderate and high N rates. The objectives of this study were to explore yield performance of two types of rice varieties with contrasting quality under moderate and high N conditions, and to further reveal the relevant physiological mechanism related to yield formation. The results may provide an insight into the fundamental of balancing high yield and superior quality of HQIR by adopting a reasonable N management.
2. Materials and Methods
2.1. Plant materials
The tested rice varieties included Yexiangyoulisi (YXY), Wanxiangyou-982 (WXY), Jiyou T025 (JY) and Keyou-5 (KY), which are all hybrid late indica rice. The YXY and WXY are high-quality indica rice (HQIR) varieties, while JY and KY are ordinary-quality indica rice (OQIR) varieties, the related key quality properties were showed in Table 1. In addition, the YXY and JY were first released in 2017, and WXY and KY were first released in 2019.
2.2. Experimental design
Field experiments were conducted in Jiangxi Shanggao Rice Science and Technology Backyard in 2020 and 2021, which located in Sixi Township, Shanggao County, Jiangxi Province, China (115° 06′ E, 28° 20′ N, 38 m altitude). The soil during the two experimental years were clay loam. The upper 20 cm of soil properties in 2020 were: pH 5.49, 29.51 g kg−1 organic C, 18.73 mg kg−1 available P, 65.13 mg kg−1 available K, and total 1.98 g kg−1 N content. In 2021, the soil properties were pH 5.62, 34.95 g kg−1 organic C, 19.29 mg kg−1 available P, 76.31 mg kg−1 available K, and 2.09 g kg−1 total N content. Climate parameters including daily average temperature and solar radiation were collected during the growth period from sowing to maturity from an automatic weather station (Vantage Pro 2, Davis instruments Corp., Hayward, CA, USA) located near the experimental site (Figure 1).
The experiments were all arranged in a split-plot design with N application rates as main plots and rice varieties as subplots. The experiments were replicated three times with a main plot size of 120 m2 and subplot size of 30 m2. Each main plot separated by a ridge with plastic film inserted into the soil to a depth of 0.3 m to minimize leakage and nutrient loss. N application rates were 165 kg N ha−1 (moderate N rate) and 225 kg N ha−1 (high N rate). For both moderate and high N rates, urea was used as the N fertilizer and was split-applied with 50% as basal (1 day before transplanting), 30% at early-tillering (7 days after transplanting), and 20% at panicle initiation.
Pre-germinated seeds were sown on June 27th in 2020 and June 24th in 2021. The 25-day-old seedlings were transplanted at a hill spacing of 25 cm × 14 cm with two seedlings per hill. The heading date of four rice varieties was on September 8th to 11th in both years, and physiological maturity date was October 26th in 2020 and October 16th in 2021. The amount and method of P and K fertilizer were consistent during the two years; rice plants received 105 kg ha−1 P2O5 and 180 kg ha−1 K2O. The P was applied as basal, while the K was split equally at basal and panicle initiation. The regime for water management was in the sequence of flooding, midseason drainage, reflooding, and moist intermittent irrigation. Weeds, pests, and diseases were intensively controlled using chemical treatments.
2.3. Sampling and measurements
2.3.1. Yield and yield components
At maturity, ten hills on a diagonal from the centre of each subplot were sampled to determine the aboveground total biomass and yield components. The sampled plants were separated into straw, filled grains, unfilled grains, and rachis. Panicle number was recorded from the 10 hills and the panicles were hand threshed. The filled grains were separated from unfilled grains using a seed winnowing machine (SXJ-80A, Hangzhou, China) and then were oven-dried at 70 °C to a constant weight. The number of filled and unfilled grains were calculated using an automatic seed counter (DC-3, Zhengzhou, China). Grain yield was determined from 5 m2 in each plot, and yields were then adjusted to the standard moisture content of 0.135 g H2O g–1 fresh weight.
2.3.2. Biomass and related properties
At heading stage, ten hills, excluding two borders, were sampled from each subplot. The rice plants were separated to leaves, stems and panicles, and then were oven-dried at 70 °C to a constant weight to determine aboveground total dry weight. At maturity, the total biomass including straw, filled grains, unfilled grains, and rachis were determined from abovementioned 2.3.1. Apparent exportation of pre-anthesis stem and leave dry matter (pre-anthesis AE), contribution of pre-anthesis AE, post-anthesis dry matter (pre-anthesis DM), contribution of post-anthesis DM, and harvest index were calculated by the following formulas.
Pre-anthesis AE = stem and leaf dry weight at heading – stem and leaf dry weight at maturity.
Contribution of pre-anthesis AE = (pre-anthesis AE/grain yield) × 100
Pre-anthesis DM = biomass at maturity – biomass at heading
contribution of post-anthesis DM = (Pre-anthesis DM/grain yield) × 100
Harvest index = filled spikelet weight/aboveground total biomass) × 100
2.3.3. Leaf area index (LAI) and grain-leaf ratio
Green leaves from six hills at heading stages was measured with a leaf area meter (LI-3000C, LI-COR, Lincoln, NE, USA). LAI was calculated as leaf area/the specific land area. Grain-leaf ratio was calculated as spikelets m−2 (filled grains m−2 or grain weight m−2)/leaf area m−2.
2.3.4. Radiation use efficiency (RUE) and net photosynthetic rate (Pn)
Canopy light interception was measured between 11:00 h and 13:00 h at middle-tillering, panicle initiation, booting, heading, 15 days after heading (HD15), and maturity using the SunScan Canopy Analysis System (Delta-T Devices Ltd., Burwell, Cambridge, UK). The measuring methods and computing methods referenced to Zhang et al. [28]. Briefly intercepted radiation during the whole growing season was the summation of intercepted radiation during each growth period. RUE (aboveground total biomass/intercepted radiation during the whole growing season) was calculated.
The Pn was measured on five flag leaves for each subplot at heading, 15 days after heading (HD15) and 30 days after heading (HD30) using a portable photosynthesis system (LI-6400, Li-Cor, Lincoln, NE, USA). The measurements were collected between 9:00 and 11:00 when photosynthetic active radiation above the canopy was 1000–1200 μmol m−2 s−1. Meanwhile, a light intensity of 1200 μmol m–2 s–1, a leaf temperature of 30 °C, a constant CO2 concentration of 380 μmol mol–1 and a relative humidity of 70% were set up in the sample chamber. Moreover, a chlorophyll meter (SPAD-502, Minolta Camera Co. Ltd., Japan) was used to measure the SPAD value for the corresponding leaves.
2.3.5. Plant N uptake and N use efficiency
The aboveground plants from heading and maturity stages were ground to a fine powder to determine plant N concentration using a fully automatic Kjeldahl apparatus (Kjeltec 8400, FOSS Analytical A/S, Hilleroed, Denmark). N uptake, post-anthesis N uptake, pre-anthesis N exportation and N use efficiency for grain production (NUEg) were calculated by the following formulas.
N uptake = N concentration × dry matter weight.
Post-anthesis N uptake = N uptake at maturity – N uptake at heading
Pre-anthesis N exportation = N uptake of stem and leaf at heading – N uptake of stem and leaf at maturity.
NUEg = grain yield/N uptake at maturity.
2.4. Statistical Analysis
Crop data were analysed using analysis of variance (ANOVA) in Statistix 8.0 (Analytical software, Tallahassee, FL, USA), and the means of the treatments were examined with the least significant (LSD) test at the 5% probability level. Graphs were drawn using Origin 2018 (OriginLab Corp, Northampton, MA, USA).
3. Results
3.1. Grain yield and its components
N application rate and variety had significant effects on grain yield and its components, except for the effect of N application rate on grain weight in 2020 and 2021; Interaction effects between N application rate and variety were observed on spikelets m−2, grain setting rate, grain yield in both years and grain weight in 2021(Table 2). Compared to moderate N rate, grain yields of HQIR were slightly decreased, while grain yields of OQIR were significantly increased by 6.60%–8.88% under high N rate in both years. High N rate significantly increased panicles m−2 of HQIR and OQIR, but decreased spikelets per panicle of HQIR, which resulted in significantly increased in spikelets m−2 of OQIR. In addition, high N rate also led to a marked decline in grain setting rate of HQIR, while had no effect for OQIR.
OQIR produced higher grain yield of 8.34%–11.87% under moderate N rate and 22.00%–22.50% under high N rate than HQIR in both years. Although panicles m−2 of OQIR were significantly lower than HQIR, OQIR had significantly higher spikelets per panicle and spikelets m−2 than HQIR under two N rates. However, significant higher grain setting rate was only observed in OQIR than that in HQIR under high N rate. Moreover, OQIR had higher grain weight than YXY (one of the varieties of HQIR) but lower grain weight than WXY.
3.2. Biomass production
Significant effects on biomass at heading and maturity stages, pre-anthesis AE and post-anthesis DM and its contribution to grain yield, as well as harvest index were observed among N application rate, variety, and their interaction in two years, except for interaction effect on the contribution of post-anthesis DM to grain yield (Table 3 and Figure 2). Compared to moderate N rate, high N rate significantly increased biomass of HQIR and OQIR at heading and maturity stages, post-anthesis DM of OQIR, and contribution of post-anthesis DM to grain yield of HQIR and OQIR (excluding 2020). On the other hand, high N rate markedly reduced pre-anthesis AE and harvest index of HQIR, and the contribution of pre-anthesis AE to grain of HQIR and OQIR.
At heading stage, HQIR produced more biomass than OQIR under moderate and high N rates. On the contrary, OQIR had more biomass at maturity stage and subsequently showed significantly higher post-anthesis DM and its contribution to grain yield than OQIR. However, in comparison with OQIR, higher pre-anthesis AE (excluding high N rate in 2020) and its contribution to grain yield were observed in HQIR under two N rates. Moreover, OQIR showed higher harvest index than HQIR under two N application rates in both years.
3.3. Net photosynthetic rate (Pn), RUE, and sink-source relationship
Compared to moderate N rate, high N rate improved SPAD values of HQIR and OQIR at HD, HD15d and HD30d, excluding SPAD values of HQIR at HD and of OQIR at HD15d in 2020 (Table 4). Meanwhile, high N rate significantly enhanced Pn of HQIR and OQIR at HD15d in 2020 and HD30d in 2021, and of OQIR at HD30d in 2020 and HD in 2021. At HD and HD15d, significant higher SPAD values and Pn of OQIR were found than those of HQIR, while OQIR showed lower SPAD values and Pn than HQIR at HD30d in both years.
High N rate significantly increased intercepted radiation and RUE of HQIR and OQIR, compared to moderate N rate in 2020 and 2021. OQIR showed markedly higher intercepted radiation but lower RUE than HQIR under two N rates (Figure 3).
In comparison with moderate N rate, high N rate significantly increased LAI of HQIR and OQIR at heading, but significantly decreased spikelets-leaf ratio, filled grains-leaf ratio and grain weight-leaf ratio of HQIR in 2020 and 2021(Table 5). Furthermore, HQIR had higher LAI and lower spikelets-leaf ratio, filled grains-leaf ratio and grain weight-leaf ratio than OQIR under two N rates.
3.4. Plant N uptake and N use efficiency
Compared with moderate N rate, high N rate significantly increased N uptake of HQIR by 25.38%–25.39% at heading and 14.50%–17.40% at maturity, and of OQIR by12.96%–17.90% at heading and 2.75%–18.81% at maturity (Table 6). In contrast, high N rate significantly decreased the post-anthesis N uptake of HQIR by 28.72%–33.06% in two years and of OQIR by 8.98% in 2020 (but increased by 55.73% in 2021). However, high N rate significantly increased pre-anthesis N exportation of HQIR and OQIR by 26.84%–37.23% and 17.06%–22.51%, respectively. Finally, the significant declines in NUEg of HQIR by 14.72%–17.90% and of OQIR by 5.42%–8.40% were found under high N rate compared to moderate N rate.
Compared with HQIR, OQIR showed significantly higher N uptake at heading under moderate N rate and N uptake at maturity under two N rates. Similarly, significant higher post-anthesis N uptake under high N rate and pre-anthesis N expropriation under two N rates were observed in OQIR than in HQIR. Accordingly, OQIR also had higher NUEg under two N rates than HQIR.
4. Discussion
4.1. Yield responses of HQIR to N application rates
N is an essential nutrient for rice growth, development, and yield formation in almost all environments. Over the past several decades, most previous studies have demonstrated that the continuously increase of rice yield in China and the world is mainly attributed to genetic improvement and N fertilizer input [17,19,29,30]. On further improving rice yield potential, China had made two breakthroughs in breeding for hybrid and “super” rice varieties since the semidwarf rice varieties were successfully bred in 1956 [1,2]. In addition, many modern high-yielding rice varieties usually have great N responsiveness and higher lodging resistance [19]. On this account, rice farmers often apply a large amount of N fertilizers to obtain the highest grain yield of high-yielding rice varieties [16,17,20]. Ultimately, a synchronous increased tendency is observed between total rice grain yields and total N fertilizers consumption in China during the past several decades [4]. In fact, numerous studies have shown that there is a quadratic function relationship between grain yield (per unit area yield) and N application rates in most high-yielding rice varieties [20,27,30,31]. These results suggest that higher yields of high-yielding rice varieties do not depend on much more N fertilizer inputs.
In this study, we find that different yield responses of HQIR and OQIR to moderate and high N application rates were mainly explained by the difference in spikelets m−2 and/or grain setting rate. High N rate significantly increased OQIR’s yield by 6.60%–8.88% but had no or slightly adverse effect on HQIR’s yield in comparison with moderate N rate (Table 2). The results indicate that HQIR can obtain the highest grain yield under the moderate N rate (165 kg N ha−1), which is in accord with the recommended optimum N application rates (135–165 kg N ha−1) from previous reports for HQIR grown in late-season of double-rice cropping system [24,25,26]. Meanwhile, our results also suggest that further increase in N fertilizer input is beneficial to gain the maximum grain yield for OQIR compared with the moderate N rate. However, previous studies revealed that OQIR could produce the maximum grain yield under moderate N rates (120–190 kg N ha−1) in the single- and double-rice cropping systems in southern China [27,32]. The difference in OQIR maybe because the site-specific N management technology was adopted in previous studies, which would significantly improve rice grain yield and N use efficiency as well as reduce N loss [17,27]. To the end, OQIR produced 8.34%–11.87% and 22.00%–22.50% higher grain yield than did HQIR under moderate and high N rates, respectively.
4.2. Agronomical and physiological responses of HQIR to N application rates
In the present study and previous studies, the high N rate did not increase spikelets m−2 but led to a significant decline in grain setting rate of HQIR [24,25,26]. On the contrary, significant increases in spikelets m−2 and no effect on grain setting rate of OQIR were observed under high N rate (Table 2). Fu et al. [31] have found that high N rate could reduce the grain setting rate of spikelets at the panicle base of “super” rice without the increased spikelets per panicle. In general, “super” rice varieties have numerous spikelets per panicle, but they often fail to achieve their high yield potential because of poor grain-filling of inferior spikelets, particularly under high N rates [33]. The main reason may be due to lower partitioning of assimilates in developing inferior spikelets resulted from low activity of enzymes involved in carbohydrate metabolism [33,34]. On the other hand, high N rate could significantly increase plant (panicles, stems, and leaves) N concentration, which might result in enhancement of plant N metabolism and overconsumption of carbohydrate, thus decreasing carbohydrate supply to panicles [35]. However, the spikelets per panicle of HQIR (140–150 spikelets panicle−1) and OQIR (160–170 spikelets panicle−1) in our study are less than that of “super” rice (more than 180 spikelets panicle−1). The decreased grain setting rate of HQIR under high N rate are probably related to the above-mentioned interpretations.
In addition, HQIR had 8.99%–13.59% higher LAI than OQIR, and the LAI of HQIR and OQIR were increased by 6.82%–6.97% and 2.64%–3.58%, respectively, under high N rate compared with moderate N rate (Table 5). Meanwhile, we also found that high N rate significantly decreased pre-anthesis AE in HQIR but increased post-anthesis DM in OQIR compared with moderate N rate (Table 3); HQIR had obviously higher canopy intercepted radiation but lower RUE than OQIR under moderate and high N application rates. These results confirm that higher LAI of HQIR might lead to canopy closure and result in further aggravated carbohydrate metabolism under high N rate [23,31]. Generally, grain-leaf area ratio, including spikelets-leaf area ratio, filled grains-leaf area ratio, and grain weight-leaf area ratio, is an important parameter to evaluate the relationship between source and sink, and is taken as a comprehensive index to breed and select high-yielding rice varieties [36]. Thereby, it is an efficient approach to improving rice grain yield by increasing the grain-leaf area ratio [37]. In our study, grain-leaf ratios were significantly decreased in HQIR, but there was no effect in OQIR under high N rate compared with moderate N rate (Table 4). The results indicate that high N rate may result in an imbalance of source and sink relationship in HQIR. On the other hand, HQIR had higher or close N uptake at heading but lower pre-anthesis N exportation, as well as higher Pn at the late grain filling stage than OQIR under high N rate (Table 4 and Table 6). The results suggest that the carbon and nitrogen metabolisms of HQIR could be disturbed by excessive N fertilizer input, resulting in lower supply of carbohydrate for panicle [38]. In comparision with 2021, grain setting rate and grain yield of HQIR and OQIR were significantly decreased in 2020, which resulted from lower daily average temperature (4.17 °C) and daily average solar radiation (3.81 MJ m−2) in 2020. Nevertheless, the agronomical and physiological mechanism underlying lower grain setting rate of HQIR under high N rate need to be further studied in future.
5. Conclusions
The present study showed that the yield responses of HQIR and OQIR to moderate and high N application rates were different, which mainly explained by the difference in spikelets m−2 and/or grain setting rate. Compared with moderate N rate, slightly decreased grain yield of HQIR was due to decreased grain setting rate, whereas significant increased grain yield of OQIR was attributed to increased spikelets m−2 under high N rate. High N rate reduced pre-anthesis AE and its contribution, as well as grain-leaf area ratio of HQIR, but did not increase its post-anthesis DM compared with moderate N rate. Therefore, the mechanism underlying lower grain setting rate of HQIR under high N rate might be aggravated carbohydrate metabolism and imbalance source-sink relathionship. These results suggest that mederate N rate is beneficial for HQIR to balance the maximum grain yield and higher quality.
Author Contributions
Conceptualization, X.X., Y.Z. (Yongjun Zeng) and X.P.; Investigation, G.D., J.W. and R.Q.; Data curation, G.D.; Writing—original draft preparation, G.D., J.W. R.Q; Writing—review and editing, X.X., J.W.; Funding acquisition, X.X., Y.Z. (Yoanhua Zeng), Y.Z. (Yongjun Zeng) and X.P. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Earmarked Fund for Jiangxi Agricultural Research System (JXARS-04); Youthful Innovation Research Team of Jiangxi Agricultural University (JXAUCXTD004).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data reported in this study is contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cheng, S.; Cao, L.; Zhuang, J.; Chen, S.; Zhan, X.; Fan, Y.; Zhu, D.; Min, S. Super hybrid rice breeding in China: achievements and prospects. J. Integr. Plant Biol. 2007, 49, 805–810. [Google Scholar] [CrossRef]
- Peng, S.; Khush, G.; Virk, P.; Tang, Q.; Zou, Y. Progress in ideotype breeding to increase rice yield potential. Field Crops Res. 2008, 108, 32–38. [Google Scholar] [CrossRef]
- Yuan, L. Increasing yield potential in rice by exploitation of heterosis. In: Virmani SS (ed) Hybrid Rice Technology: New Development and Future Prospects. International Rice Research Institute, Los Baños, Philippines 1994, pp 1-6. [Google Scholar]
- FAOSTAT. https://www.fao.org/faostat/zh/#data.
- Zhou, H.; Xia, D.; He, Y. Rice grain quality-traditional traits for high quality rice and health-plus substances. Mol Breeding 2020, 40, 1–17. [Google Scholar] [CrossRef]
- Liu, X.; Liu, C.; Wang, Y.; Ning, M.; Jing, Q.; Zhang, C. Current situation and suggestions on the development of high-quality rice branding in China. China rice 2022, 28, 12–15. (In Chinese) [Google Scholar] [CrossRef]
- Liao, F.; Zhou, K.; Yang, H.; Xu, Q. Comparison of grain quality between F1 hybrids and their parents in indica hybrid rice. Chinese J. Rice Sci 2003, 17, 134–140. (In Chinese) [Google Scholar]
- Peng, S.; Tang, Q.; Zou, Y. Current status and challenges of rice production in China. Plant Prod Sci. 2009, 12, 3–8. [Google Scholar] [CrossRef]
- Liu, Q.; Li, Q.; Cai, X.; Wang, H.; Tang, S.; Yu, H.; Wang, Z.; Gu, M. Molecular marker-assisted selection for improved cooking and eating quality of two elite parents of hybrid rice. Crop Sci. 2006, 46, 2354–2360. [Google Scholar] [CrossRef]
- Feng, F.; Li, Y.; Qin, X.; Liao, Y.; Siddique, K. Changes in rice grain quality of indica and japonica type varieties released in China from 2000 to 2014. Front in Plant Sci. 2017, 8, 01863. [Google Scholar] [CrossRef]
- Wang, Y.; Zheng, Y.; Cai, Q.; Liao, C.; Mao, X.; Xie, H.; Zhu, Y.; Lian, L.; Luo, X.; Xie, H.; Zhang, J. Population structure and association analysis of yield and grain quality traits in hybrid rice primal parental lines. Euphytica 2016, 212, 261–273. [Google Scholar] [CrossRef]
- Kinoshita, N.; Kato, M.; Koyasaki, K.; Kawashima, T.; Nishimura, T.; Hirayama, Y.; Takamure, I.; Sato, T.; Kato, K. Identification of quantitative trait loci for rice grain quality and yield-related traits in two closely related Oryza sativa L. subsp. japonica cultivars grown near the northernmost limit for rice paddy cultivation. Breeding Sci. 2017, 67, 191–206. [Google Scholar] [CrossRef]
- Zeng, D.; Tian, Z.; Rao, Y.; Dong, G.; Yang, Y.; Huang, L.; Leng, Y.; Xu, J.; Sun, C.; Zhang, G.; Hu, J.; Zhu, L.; Gao, Z.; Hu, X.; Guo, L.; Xiong, G.; Wang, Y.; Li, J.; Qian, Q. Rational design of high-yield and superior-quality rice. Nat Plants 2017, 17031. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wu, K.; Yuan, Q.; Liu, X.; Liu, Z.; Lin, X.; Zeng, R.; Zhu, H.; Dong, G.; Qian, Q.; Zhang, G.; Fu, X. Control of grain size, shape and quality by OsSPL16 in rice. Nat Genet. 2012, 44, 950–954. [Google Scholar] [CrossRef]
- Lin, H.; Li, H.; E, Z.; Pang, Q. Analysis on characteristics of rice varieties registered in China in 2020. China rice 2021, 27, 6–11. (In Chinese) [Google Scholar] [CrossRef]
- Cheng, B.; Jiang, Y.; Cao, C. Balance rice yield and eating quality by changing the traditional nitrogen management for sustainable production in China. J. Clean. Prod. 2021, 312, 127793. [Google Scholar] [CrossRef]
- Peng, S.; Buresh, R.; Huang, J.; Yang, J.; Zou, Y.; Zhong, X.; Wang, G.; Zhang, F. Strategies for overcoming low agronomic nitrogen use efficiency in irrigated rice systems in China. Field Crops Res. 2006, 96, 37–47. [Google Scholar] [CrossRef]
- Chen, X.; Cui, Z.; Fan, M.; Vitousek, P.; Zhao, M.; Ma, W.; Wang, Z.; Zhang, W.; Yan, X.; Yang, J.; Deng, X.; Gao, Q.; Zhang, Q.; Guo, S.; Ren, Jun. ; Li, S.; Ye, Y.; Wang, Z.; Huang, J.; Tang, Q.; Sun, Y.; Peng, X.; Zhang, J.; He, M.; Zhu, Y.; Xue, J.; Wang, G.; Wu, L.; An, N.; Wu,L.; Ma, L.; Zhang, W.; Zhang, F. Producing more grain with lower environmental costs. Nature 2014, 514, 486–489. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Peng, S. Yield potential and nitrogen use efficiency of China’s super rice. J. Integr. Agr. 2017, 16, 1000–1008. [Google Scholar] [CrossRef]
- Huang, M.; Jiang, P.; Shan, S.; Gao, W.; Ma, G.; Zou, Y.; Uphoff, N.; Yuan, L. Higher yields of hybrid rice do not depend on nitrogen fertilization under moderate to high soil fertility conditions. Rice 2017, 10, 43. [Google Scholar] [CrossRef]
- Gu, J.; Chen, J.; Chen, L.; Wang, Z.; Zhang, H.; Yang, J. Grain quality changes and responses to nitrogen fertilizer of japonica rice cultivars released in the Yangtze River Basin from the 1950s to 2000s. Crop Jour. 2015, 3, 285–297. [Google Scholar] [CrossRef]
- Zhu, D.; Zhang, H.; Guo, Ba.; Xu, K.; Dai, Q.; Wei, H.; Gao, H.; Hu, Y.; Cui, P.; Huo, Z. Effects of nitrogen level on yield and quality of japonica soft super rice. J. Integr. Agr. 2017, 16, 1018–1027. [Google Scholar] [CrossRef]
- Zhou, C.; Huang, Y.; Jia, B.; Wang, Y.; Wang, Y.; Xu, Q.; Li, R.; Wang, S.; Dou, F. Effects of cultivar, nitrogen rate, and planting density on rice-grain quality. Agronomy 2018, 8, 246. [Google Scholar] [CrossRef]
- Zhang, G.; Zhao, R.; Liu, Y.; Yao, B.; Tang, W. Effect of different amount of nitrogen on the yield and the quality of high-quality rice and its nitrogen utilization efficiency. J. Hunan Agr. Univ. (Nat. Sci.) 2019, 45, 231–236. (In Chinese) [Google Scholar]
- Wu, J.; Que, R.; Qi, W.; Duan, G.; Wu, J.; Zeng, Y.; Pan, X.; Xie, X. Varietal variances of grain nitrogen content and its relations to nitrogen accumulation and yield of high-quality rice under different nitrogen rates. Agronomy 2022, 12, 2719. [Google Scholar] [CrossRef]
- Zhou, M.; Wang, X.; Wang, S.; Hao, R.; Dang, C.; Hu, Y.; Liu, Y.; Mu, Q.; Tian, X. Effect of nitrogen application rate on yield and quality of high⁃quality rice in Jianghan Plain. J. Henan Agr. Sci. 2022, 51, 25–34. (In Chinese) [Google Scholar] [CrossRef]
- Huang, J.; He, F.; Cui, K.; Buresh, R. J.; Xu, B.; Gong, W.; Peng, S. Determination of optimal nitrogen rate for rice varieties using a chlorophyll meter. Field Crops Res. 2008, 105, 70–80. [Google Scholar] [CrossRef]
- Zhang, Y.; Tang, Q.; Zou, Y.; Li, D.; Qin, J.; Yang, S.; Chen, Li.; Xia, B.; Peng, S. Yield potential and radiation use efficiency of “super” hybrid rice grown under subtropical conditions. Field Crops Res. 2009, 114, 91–98. [Google Scholar] [CrossRef]
- Ladha, J.; Tirol-Padre, A.; Reddy, C.; Verma, S.; Powlson, D. S.; van Kessel, C; Richter, D. B.; Chakraborty, D.; Pathak, H. Global nitrogen budgets in cereals: A 50-year assessment for maize, rice and wheat production systems. Sci Rep 2016, 6, 19355. [Google Scholar] [CrossRef]
- Huang, M.; Lei, T.; Cao, F.; Chen, J.; Shan, S.; Zou, Y. Grain yield responses to nitrogen rate in two elite double-cropped inbred rice cultivars released 41 years apart. Field Crops Res. 2020, 259, 107970. [Google Scholar] [CrossRef]
- Fu, P.; Wang, J.; Zhang, T.; Huang, J; Peng, S. High nitrogen input causes poor grain filling of spikelets at the panicle base of super hybrid rice. Field Crops Res. 2019, 244, 107635. [Google Scholar] [CrossRef]
- Jiang, P.; Xie, X.; Huang, M.; Zhou, X.; Zhang, R.; Chen, J.; Wu, D.; Xia, B.; Xiong, H.; Xu, F.; Zou, Y. Potential yield increase of hybrid rice at five locations in southern China. Rice 2016, 9, 11. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, J. Grain-filling problem in “super” rice. J. Exp. Bot. 2010, 61, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, P.; Panigrahi, R. Ethylene control of grain development in the inferior spikelets of rice panicle. Adv. Plant Physiol. 2011, 12, 79–89. [Google Scholar]
- Liang, Z.; Bao, A.; Li, H.; Cai, H. The effect of nitrogen level on rice growth, carbon-nitrogen metabolism and gene expression. Biologia 2015, 70, 1340–1350. [Google Scholar] [CrossRef]
- Ling, Q.; Yang, J. Studies on “grain-leaf ratio” of population and cultural approaches of high yield in rice plants. Sci Agric Sin 1986, 19, 1–8. (In Chinese) [Google Scholar]
- Zhang, H.; Chen, T.; Liu, L.; Wang, Z.; Yang, J.; Zhang, J. Performance in grain yield and physiological traits of rice in the Yangtze River Basin of China during the last 60 yr. J. Integr. Agr. 2013, 12, 57–66. [Google Scholar] [CrossRef]
- Wei, H.; Meng, T.; Li, X.; Dai, Q.; Zhang, H.; Yin, X. Sink-source relationship during rice grain filling is associated with grain nitrogen concentration. Field Crops Res. 2018, 215, 23–38. [Google Scholar] [CrossRef]
Figure 1.
Daily average temperature and solar radiation during the late rice growing seasons in Shanggao County, Jiangxi Province, China in 2020 and 2021.
Figure 1.
Daily average temperature and solar radiation during the late rice growing seasons in Shanggao County, Jiangxi Province, China in 2020 and 2021.
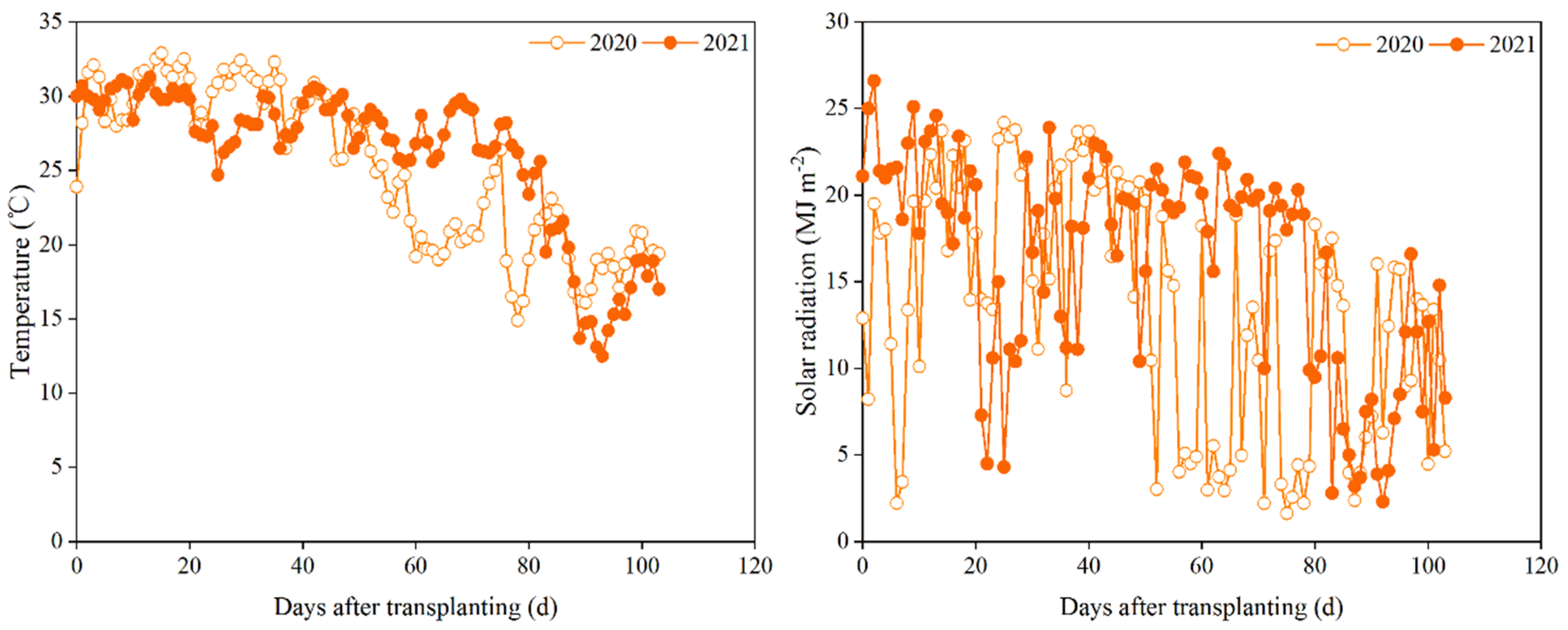
Figure 2.
Harvest index of HQR and OQR varieties under moderate and high N rates. Note: Different lowercase letters between varieties in the same N rate indicate significant differences at the p < 0.05 level.
Figure 2.
Harvest index of HQR and OQR varieties under moderate and high N rates. Note: Different lowercase letters between varieties in the same N rate indicate significant differences at the p < 0.05 level.
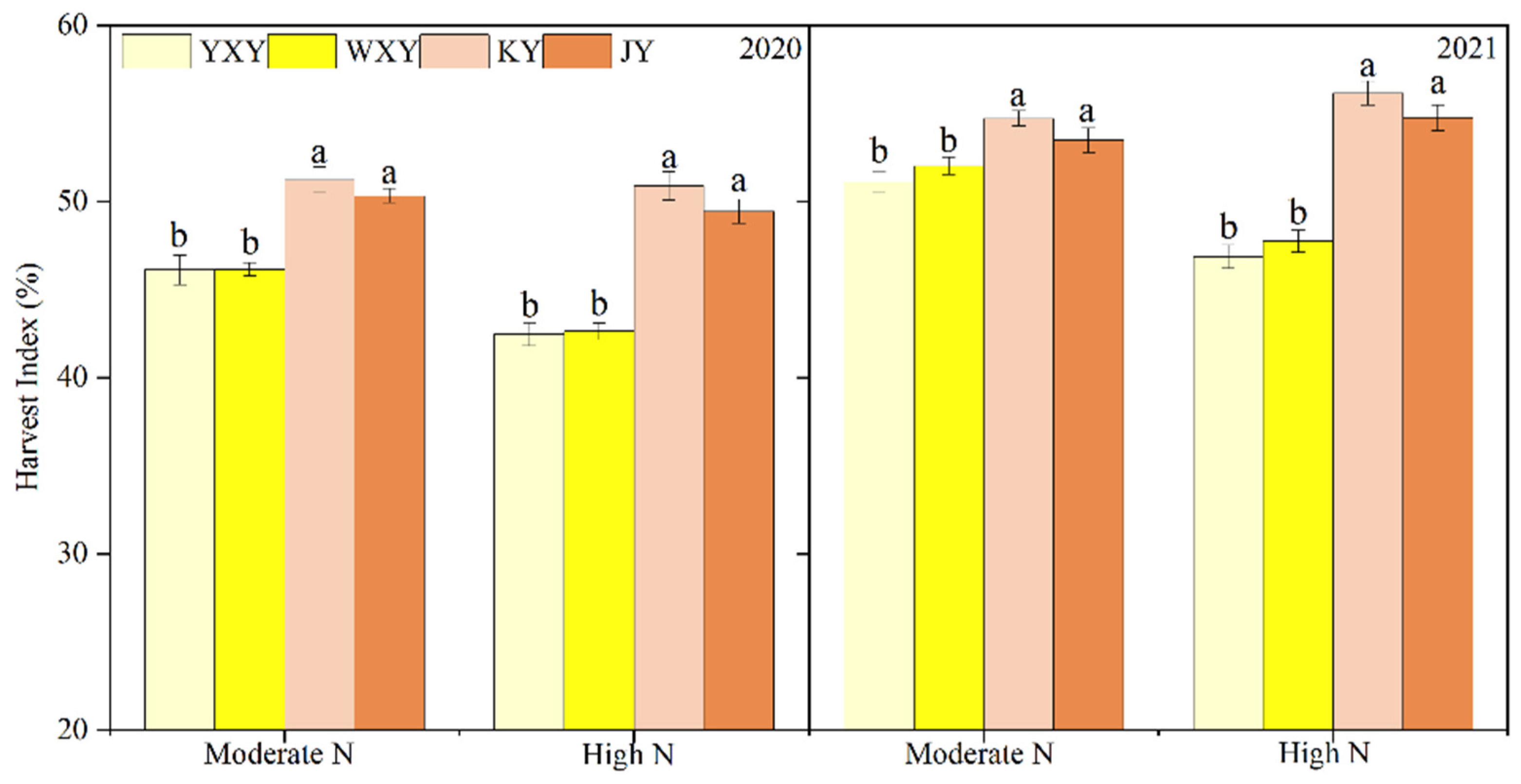
Figure 3.
Intercepted radiation and RUE of HQR and OQR varieties under moderate and high N rates. Note: Different lowercase letters between varieties in the same N rate indicate significant differences at the p <0.05 level.
Figure 3.
Intercepted radiation and RUE of HQR and OQR varieties under moderate and high N rates. Note: Different lowercase letters between varieties in the same N rate indicate significant differences at the p <0.05 level.
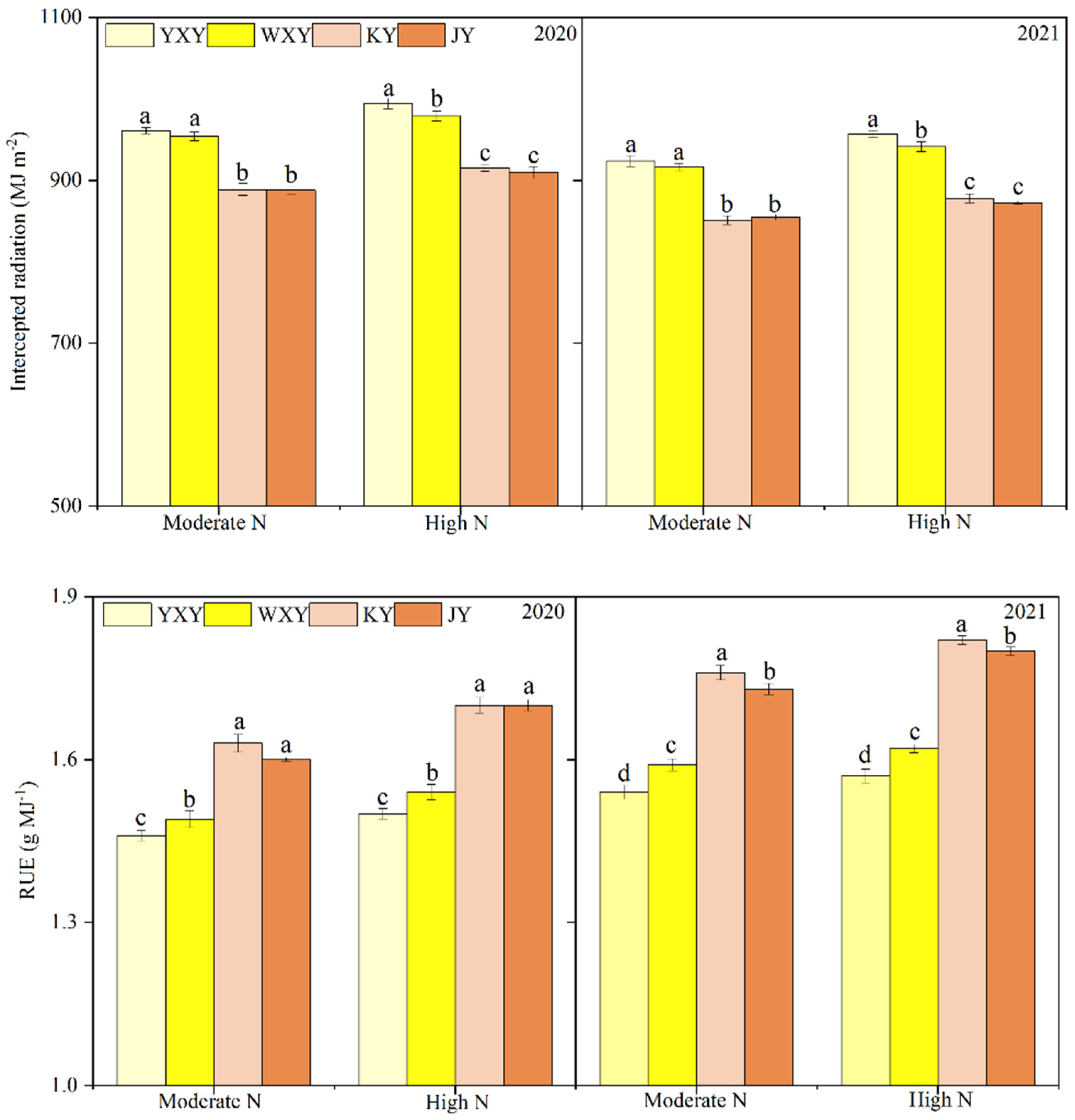

Table 1.
The related key quality properties of tested varieties.
| Type | Variety | Length-width ratio | Percentage of chalky grains (%) | Chalkiness degree (%) | Gel consistency (mm) | Amylose content (%) |
|---|---|---|---|---|---|---|
| HQIR | YXY | 4.0 | 5.0 | 0.9 | 69 | 13.9 |
| WXY | 3.9 | 9.0 | 1.8 | 73 | 15.0 | |
| OQIR | JY | 3.3 | 23.0 | 4.8 | 30 | 25.9 |
| KY | 2.9 | 33.0 | 8.2 | 36 | 18.5 |
Note: Data obtained from China rice data center, https://www.ricedata.cn/variety/index.htm.
Table 2.
Grain yield and its components of HQR and OQR varieties under moderate and high N rates.
| Year | N rate | Variety | Panicles (m−2) |
Spikelets (panicle−1) |
Spikelets (×103 m−2) |
Grain setting rate (%) |
Grain weight (mg) |
Grain yield (t hm−2) |
|---|---|---|---|---|---|---|---|---|
| 2020 | Moderate | YXY | 311.19 a | 146.71 b | 45.64 c | 73.89 a | 19.81 d | 7.51 c |
| N | WXY | 298.76 b | 144.99 b | 43.31 d | 72.74 a | 23.38 a | 7.64 c | |
| MeanHQR | 304.97 B | 145.85 B | 44.48 C | 73.32 A | 21.60 B | 7.58 C | ||
| KY | 283.00 c | 172.82 a | 48.90 a | 74.43 a | 21.54 c | 8.63 a | ||
| JY | 279.62 c | 170.19 a | 47.58 b | 70.16 b | 22.53 b | 8.33 b | ||
| MeanOQR | 281.31 D | 171.51 A | 48.24 B | 72.29 AB | 22.03 A | 8.48 B | ||
| High N | YXY | 327.69 a | 140.33 b | 45.97 c | 69.95 b | 19.72 d | 7.35 c | |
| WXY | 315.90 b | 137.70 b | 43.50 d | 67.63 c | 23.13 a | 7.46 c | ||
| MeanHQR | 321.80 A | 139.02 C | 44.74 C | 68.79 C | 21.42 B | 7.41 C | ||
| KY | 300.14 c | 170.09 a | 51.04 a | 73.67 a | 21.73 c | 9.19 a | ||
| JY | 293.48 c | 169.24 a | 49.66 b | 69.67 b | 22.62 b | 8.90 b | ||
| MeanOQR | 296.81 C | 169.66 A | 50.35 A | 71.67 B | 22.17 A | 9.04 A | ||
| 2021 | Moderate | YXY | 301.02 a | 146.17 b | 43.00 b | 81.95 a | 20.71 c | 8.47 b |
| N | WXY | 286.38 b | 142.67 b | 40.85 c | 81.58 a | 23.68 a | 8.79 b | |
| MeanHQR | 293.70 B | 144.42 B | 42.42 C | 81.76 A | 22.19 B | 8.63 C | ||
| KY | 281.67 b | 164.66 a | 46.37 a | 82.15 a | 23.24 b | 9.37 a | ||
| JY | 280.29 b | 163.18 a | 45.73 a | 82.61 a | 23.51 a | 9.34 a | ||
| MeanOQR | 280.98 C | 163.92 A | 46.05 B | 82.35 A | 23.38 A | 9.35 B | ||
| High N | YXY | 325.49 a | 136.08 b | 44.27 c | 77.39 b | 20.57 c | 8.18 b | |
| WXY | 305.99 b | 133.88 b | 40.96 d | 76.99 b | 23.43 b | 8.44 b | ||
| MeanHQR | 315.74 A | 134.98 C | 42.62 C | 77.19 B | 21.00 B | 8.31 C | ||
| KY | 300.95 bc | 163.12 a | 49.09 a | 82.09 a | 23.39 b | 10.29 a | ||
| JY | 293.61 c | 160.18 a | 47.02 b | 81.31 a | 23.74 a | 10.08 a | ||
| MeanOQR | 297.28 B | 161.65 A | 48.06 A | 81.73 A | 23.56 A | 10.18 A | ||
| Analysis of variance | ||||||||
| 2020 | N rate | ** | * | ** | ** | ns | * | |
| Variety | *** | *** | *** | *** | *** | *** | ||
| N rate ×Variety | ns | ns | *** | ** | ns | ** | ||
| 2021 | N rate | * | * | * | * | ns | * | |
| Variety | *** | *** | *** | ** | *** | *** | ||
| N rate ×Variety | ns | ns | *** | * | ** | *** | ||
Note: Within a column, different lowercase letters between varieties in the same N rate indicate significant differences at the p < 0.05 level; different uppercase letters between varietal types across two N rates indicate significant differences at the p < 0.05 level. * , ** and *** represent significant differences at the p < 0.05, p < 0.01 and p < 0.001 probability levels, respectively; ns means no significance.
Table 3.
Biomass production and its contribution to grain yield of HQR and OQR varieties under moderate and high N rates.
Table 3.
Biomass production and its contribution to grain yield of HQR and OQR varieties under moderate and high N rates.
| Year | N rate | Variety | Biomass (g m−2) | Pre-anthesis AE (g m−2) |
Contribution of pre-anthesis AE (%) |
Post-anthesis DM (g m−2) |
Contribution of post-anthesis DM (%) | |
|---|---|---|---|---|---|---|---|---|
| Heading | Maturity | |||||||
| 2020 | Moderate | YXY | 1020.44 a | 1401.26 c | 261.17 a | 34.75 a | 380.82 d | 50.70 c |
| N | WXY | 1017.14 a | 1423.75 b | 248.60 b | 32.55 b | 406.61 c | 53.22 c | |
| MeanHQR | 1018.79 B | 1412.50 D | 254.89 A | 33.65 A | 393.72 C | 51.96 C | ||
| KY | 926.18 c | 1449.41 b | 216.34 d | 25.07 d | 523.23 a | 60.61 a | ||
| JY | 943.90 b | 1424.06 a | 234.17 c | 28.10 c | 480.16 b | 57.62 b | ||
| MeanOQR | 935.04 D | 1436.73 C | 225.26 B | 26.58 C | 501.70 B | 59.12 A | ||
| High N | YXY | 1097.13 a | 1488.93 c | 223.21 ab | 30.37 a | 391.80 d | 53.28 d | |
| WXY | 1090.88 a | 1504.64 b | 208.60 c | 27.96 b | 413.76 c | 55.48 c | ||
| MeanHQR | 1094.00 A | 1496.78 B | 215.91 B | 29.17 B | 402.78 C | 54.38 B | ||
| KY | 980.86 c | 1553.18 a | 215.52 bc | 23.45 d | 572.32 a | 62.29 a | ||
| JY | 1018.90 b | 1547.35 a | 229.42 a | 25.79 c | 528.45 b | 59.39 b | ||
| MeanOQR | 999.88 C | 1550.26 A | 222.47 B | 24.62 D | 550.39 A | 60.84 A | ||
| 2021 | Moderate | YXY | 1019.01 a | 1424.81 d | 309.24 a | 36.52 a | 405.80 d | 47.93 d |
| N | WXY | 1012.80 a | 1452.75 c | 277.29 b | 31.56 b | 439.95 c | 50.05 c | |
| MeanHQR | 1015.90 B | 1438.78 D | 293.27 A | 34.04 A | 422.88 C | 48.99 D | ||
| KY | 903.15 c | 1473.05 b | 230.22 d | 24.57 c | 569.90 a | 60.81 a | ||
| JY | 949.54 b | 1501.22 a | 243.39 c | 26.07 c | 551.68 b | 59.10 b | ||
| MeanOQR | 926.35 D | 1487.13 C | 236.81 C | 25.32 C | 560.79 B | 59.95 B | ||
| High N | YXY | 1089.88 a | 1501.06 c | 272.30 a | 33.28 a | 411.18 d | 50.27 b | |
| WXY | 1072.48 b | 1520.58 b | 253.93 b | 30.07 b | 448.10 c | 53.08 b | ||
| MeanHQR | 1081.18 A | 1510.82 B | 263.12 B | 31.68 B | 429.64 C | 51.67 C | ||
| KY | 929.89 d | 1575.81 a | 222.42 d | 21.62 d | 645.92 a | 62.80 a | ||
| JY | 961.56 c | 1583.48 a | 235.86 c | 23.39 c | 621.92 b | 61.70 a | ||
| MeanOQR | 945.73 C | 1579.64 A | 229.14 C | 22.51 D | 633.92 A | 62.25 A | ||
| Analysis of variance | ||||||||
| 2020 | N rate | *** | *** | ** | ** | ** | * | |
| Variety | *** | *** | ** | *** | *** | *** | ||
| N rate ×Variety | *** | *** | *** | * | *** | ns | ||
| 2021 | N rate | *** | *** | ** | ** | * | ||
| Variety | *** | *** | *** | ** | *** | *** | ||
| N rate ×Variety | *** | *** | *** | *** | *** | ns | ||
Note: Within a column, different lowercase letters between varieties in the same N rate indicate significant differences at the p < 0.05 level; different uppercase letters between varietal types across two N rates indicate significant differences at the p < 0.05 level. * , ** and *** represent significant differences at the p < 0.05, p < 0.01 and p < 0.001 probability levels, respectively; ns means no significance.
Table 4.
SPAD value and net photosynthetic rate (Pn) of HQR and OQR varieties under moderate and high N rates.
Table 4.
SPAD value and net photosynthetic rate (Pn) of HQR and OQR varieties under moderate and high N rates.
| Year | N rate | Variety | SPAD value | Pn (μmol CO2 m−2 s−1) | |||||
|---|---|---|---|---|---|---|---|---|---|
| HD | HD15d | HD30d | HD | HD15d | HD30d | ||||
| 2020 | Moderate | YXY | 34.89 d | 30.89 d | 21.01 b | 17.96 c | 15.43 b | 12.21 a | |
| N | WXY | 35.99 c | 32.10 c | 21.89 a | 18.10 c | 16.02 b | 11.46 b | ||
| MeanHQR | 35.44 C | 31.50 C | 21.45 B | 18.03 B | 15.73 C | 11.84 A | |||
| KY | 39.06 a | 33.00 b | 16.55 d | 23.25 a | 16.84 a | 10.34 c | |||
| JY | 37.98 b | 34.21 a | 18.77 c | 22.68 b | 16.99 a | 10.59 c | |||
| MeanOQR | 38.52 B | 33.61A | 17.66 D | 22.96 A | 16.92 B | 10.47 C | |||
| High N | YXY | 35.62 d | 31.89 c | 23.41 b | 18.39 c | 17.56 b | 12.37 a | ||
| WXY | 37.89 c | 33.78 b | 26.99 a | 18.84 c | 18.00 b | 12.00 b | |||
| MeanHQR | 36.76 C | 32.84 B | 25.20 A | 18.62 B | 17.78 B | 12.19 A | |||
| KY | 42.00 a | 34.88 a | 17.56 d | 25.51 a | 20.45 a | 11.00 d | |||
| JY | 39.44 b | 33.83 b | 19.83 c | 21.78 b | 18.00 b | 11.34 c | |||
| MeanOQR | 40.72 A | 34.36 A | 18.69 C | 23.65 A | 19.23 A | 11.17 B | |||
| 2021 | Moderate | YXY | 33.84 c | 29.50 c | 21.37 b | 17.80 b | 15.12 c | 11.99 a | |
| N | WXY | 36.41 b | 33.57 a | 24.46 a | 18.26 b | 15.93 b | 11.54 b | ||
| MeanHQR | 35.13 D | 31.54 C | 22.92 B | 18.03C | 15.53C | 11.77B | |||
| KY | 39.32 a | 32.19 b | 15.96 d | 23.02 a | 16.03 b | 10.01 c | |||
| JY | 38.57 a | 34.29 a | 16.65 c | 22.88 a | 16.94 a | 9.99 c | |||
| MeanOQR | 38.95 B | 33.24 B | 16.30 D | 22.95 B | 16.49 AB | 10.00 D | |||
| High N | YXY | 35.90 c | 32.78 c | 22.63 b | 18.19 d | 16.31 c | 12.32 a | ||
| WXY | 38.33 b | 34.23 b | 27.36 a | 18.73 c | 16.00 c | 11.94 b | |||
| MeanHQR | 37.11 C | 33.50 B | 25.00 A | 18.46 C | 16.16 BC | 12.13 A | |||
| KY | 41.78 a | 33.84 b | 16.96 d | 25.28 a | 17.34 a | 10.62 d | |||
| JY | 42.24 a | 36.55 a | 18.93 c | 24.41 b | 16.98 b | 11.05 c | |||
| MeanOQR | 42.01 A | 35.20 A | 17.94 C | 24.85 A | 17.16 A | 10.84 C | |||
Note: Within a column, different lowercase letters between varieties in the same N rate indicate significant differences at the p < 0.05 level; different uppercase letters between varietal types across two N rates indicate significant differences at the p < 0.05 level.
Table 5.
LAI and grain-leaf area ratio of HQR and CMR varieties under moderate and high N rates.
| N rate | Variety | 2020 | 2021 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| LAI | Spikelets-leaf area ratio (cm−2) |
Filled grains-leaf area ratio (cm−2) |
Grain weight- leaf area ratio (mg cm−2) |
LAI | Spikelets-leaf area ratio (cm−2) |
Filled grains-leaf area ratio (cm−2) |
Grain weight- leaf area ratio (mg cm−2) |
||
| Moderate | YXY | 7.08 a | 0.64b | 0.48c | 9.44c | 7.13 a | 0.62b | 0.51b | 10.48c |
| N | WXY | 6.99 a | 0.62c | 0.45d | 10.54b | 6.94 a | 0.59b | 0.48c | 11.26b |
| MeanHQR | 7.04 B | 0.63B | 0.46B | 9.99B | 7.03 B | 0.60B | 0.49B | 10.87B | |
| KY | 6.51 b | 0.75a | 0.56a | 12.05a | 6.51 b | 0.73a | 0.60a | 14.07a | |
| JY | 6.36 b | 0.75a | 0.52b | 11.82a | 6.39 b | 0.72a | 0.59a | 13.90a | |
| MeanOQR | 6.43 D | 0.75A | 0.54A | 11.94A | 6.45 D | 0.72A | 0.60A | 13.98A | |
| High N | YXY | 7.68 a | 0.60b | 0.42c | 8.24c | 7.60 a | 0.58b | 0.45c | 9.27c |
| WXY | 7.36 b | 0.59b | 0.40d | 9.25b | 7.44 b | 0.55b | 0.42d | 10.03b | |
| MeanHQR | 7.52 A | 0.59C | 0.41C | 8.74C | 7.52 A | 0.57C | 0.44C | 9.65C | |
| KY | 6.76 c | 0.76a | 0.56a | 12.09a | 6.72 c | 0.73a | 0.61a | 14.19a | |
| JY | 6.57 d | 0.76a | 0.53b | 11.91a | 6.52 d | 0.72a | 0.59b | 14.04a | |
| MeanOQR | 6.66 C | 0.76A | 0.54A | 12.00A | 6.62 C | 0.73A | 0.60A | 14.11A | |
Note: Within a column, different lowercase letters between varieties in the same N rate indicate significant differences at the p < 0.05 level; different uppercase letters between varietal types across two N rates indicate significant differences at the p < 0.05 level.
Table 6.
Aboveground plant N uptake and NUEg of HQR and OQR varieties under moderate and high N rates.
Table 6.
Aboveground plant N uptake and NUEg of HQR and OQR varieties under moderate and high N rates.
| Year | N rate | Variety | N uptake at heading (kg ha−2) |
N uptake at maturity (kg ha−2) |
Post-anthesis N uptake (kg ha−2) |
Pre- anthesis N exportation (kg ha−2) |
NUEg (kg kg−1) |
|---|---|---|---|---|---|---|---|
| 2020 | Moderate | YXY | 123.47 c | 149.87c | 26.40b | 44.55b | 50.22bc |
| N | WXY | 122.38 c | 157.87b | 35.49a | 41.78b | 48.41c | |
| MeanHQR | 122.92 C | 153.87D | 30.95AB | 43.16C | 49.31B | ||
| KY | 129.81 b | 164.59a | 34.78a | 55.29a | 52.45a | ||
| JY | 134.72 a | 162.73a | 28.01b | 59.37a | 51.22ab | ||
| MeanOQR | 132.26 B | 163.66C | 31.40A | 57.33B | 51.84A | ||
| High N | YXY | 156.24 ab | 177.80b | 21.56b | 62.18b | 41.37b | |
| WXY | 152.01 bc | 174.57b | 22.56b | 56.29c | 42.76b | ||
| MeanHQR | 154.13 A | 176.18B | 22.06C | 59.23B | 42.06C | ||
| KY | 151.50 c | 184.54a | 33.04a | 68.41a | 49.83a | ||
| JY | 160.38 a | 184.49a | 24.11b | 69.89a | 48.23a | ||
| MeanOQR | 155.94 A | 184.52A | 28.58B | 69.15A | 49.03B | ||
| 2021 | Moderate | YXY | 125.62 bc | 142.64d | 17.02c | 50.92c | 59.36b |
| N | WXY | 124.42 c | 146.90c | 22.49ab | 47.23d | 59.85ab | |
| MeanHQR | 125.02 D | 144.77D | 19.75B | 49.07C | 59.60A | ||
| KY | 128.88 ab | 152.56b | 23.68a | 56.89b | 61.45a | ||
| JY | 136.72 a | 155.11a | 18.38bc | 61.22a | 60.24ab | ||
| MeanOQR | 132.80 C | 153.84C | 21.03B | 59.05B | 60.84A | ||
| High N | YXY | 159.63 a | 172.13b | 12.50c | 64.66bc | 47.54c | |
| WXY | 153.87 b | 167.80c | 13.94c | 59.81c | 50.32b | ||
| MeanHQR | 156.75 A | 169.97B | 13.22C | 62.24B | 48.93C | ||
| KY | 145.35 c | 181.80a | 36.45a | 68.64b | 56.59a | ||
| JY | 154.68 b | 183.73a | 29.05b | 76.04a | 54.87a | ||
| MeanOQR | 150.01 B | 182.77A | 32.75A | 72.34A | 55.73B |
Note: Within a column, different lowercase letters between varieties in the same N rate indicate significant differences at the p < 0.05 level; different uppercase letters between varietal types across two N rates indicate significant differences at the p < 0.05 level.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



