
Submitted:
25 May 2023
Posted:
26 May 2023
You are already at the latest version
Abstract
Euphorbia ingens is traditionally used to treat and manage cancer in Ambeere community of Embu County in Kenya. Whilst research has demonstrated the bioactivities of E. ingens including antimicrobial, antitubercular and antifungal activities, scientific validation of its anticancer properties is limited. This study evaluated the antiproliferative potentials of E. ingens on human prostate cancer cell line (DU-145). The 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide assay was used to assess the antiproliferative activity, chemical constituents were analysed by qualitative colour method and Gas Chromatography-Mass Spectrometry analysis. At the same time, the investigation of putative molecular targets and mechanisms of action of E. ingens was done through network pharmacological analysis. Reverse transcription-quantitative polymerase chain reaction (RT-qPCR) was carried out to validate the network predictions of putative targets. Our result showed E. ingens ethyl acetate inhibited DU-145 growth (IC50 of 9.71 ± 0.4 µg/ml) with a high selectivity index of 8.26. There was the presence of phenols, terpenoids, flavonoids, tannins, sterols, and saponins; additional 18 compounds were identified by the GC-MS approach. ESR1, IL6, MMP9, CDK2, MAP2K1, AR, PRKCD, CDK1, CDC25B, and JAK2 were indicated as key targets of E. ingens against prostate cancer with the PI3K-AKT, MAPK, and p53 signalling pathways identified as the most probable mechanisms of action. There was significant downregulation of AR and BCL2, and upregulation of p53 and caspase-3 in E. ingens-treated DU-145 cells compared to 0.2 % DMSO negative control. Our results suggest that E. ingens has phytochemical compounds efficacious at inhibiting the proliferation of DU-145 cells; therefore, the plant can be considered a potential source of compounds that may be used to manage and treat prostate cancer; however, further in vivo evaluations are needed.
Keywords:
Euphorbia ingens
; Anticancer
; Antiproliferative
; Prostate Cancer
; Network Pharmacology
; Phyto-chemicals
; Mechanism of action
; Molecular targets
1. Introduction
Cancer is one of the major cause of death globally, and a significant contributor to decreased life expectancy [1]. Cancer of the prostate is an adenocarcinoma that often forms in the glandular prostate and can be identified by its characteristic glandular patterns under the microscope [2]. This cancer can advance by spreading outside the prostate or to remain contained in the prostate for a very long time. The lymph nodes and bone are common sites where prostate cancer metastasizes; it is hypothesized that the prostatic venous plexus's connection to the spinal veins contributes to the latter [3]. According to the World Health Organisation (WHO), there are 1,414,259 new cases of prostate cancer (PCa) each year, making it the second most common cancer in men and the fifth greatest cause of cancer-related mortality globally [4]. In Africa, the rate of prostate cancer is 26.6% per 100,000 people, and in sub-Saharan Africa, there are an estimated 70,000 new cases per year [5]. Countries with low socio-demographic indexes such as Nigeria and Kenya are affected the most by prostate cancer, and this has been attributed to the lack of effective preventive and treatment strategies in the countries [6].
Prostatectomy and local radiation are used to treat localised PCa, which accounts for 90 % of PCa cases. However, PCa is diagnosed in an advanced stage in around 90 % of men with the disease, when androgen deprivation therapy (ADT) and chemotherapy are common treatment options. [5,7]. ADT treatment leads to recurrent androgen-independent prostate cancer within 2–3 years, with frequent metastases to regional lymph nodes or the pelvis, making the disease advanced [8]. Although there are several prostate cancer chemotherapy treatments, there is still a gap in the therapeutic options for advanced prostate cancer. Drugs that target rapidly proliferating cancer cells such as docetaxel and paclitaxel can also damage healthy cells, causing fatigue, hypertension, hot flushes, arthralgia, fractures, peripheral oedema and rash [9]. Therefore, there is a serious need to prospect better therapeutic interventions against PCa and herbal plant sources provide the greatest promise, and the “lowest hanging fruits”.
The use of herbal plants in cancer has gained substantial attention, and recently, research is ongoing, with the US National Cancer Institute (NCI) playing a pivotal role in the research of traditional medicine to treat cancer [10]. Herbal plants have various advantages over chemical products due to their lower chances of inducing adverse effects, cost-effectiveness, their tolerability, and their reduced chances of developing resistance [11]. Findings have shown that herbal plants exercise their anticancer properties owing to the existence of phytochemicals which act by creating antioxidant effects, repairing damaged DNA, boosting the immune system, inducing apoptosis and suppressing cell cycle [12,13].
Euphorbia ingens E.Mey. ex Boiss belongs to the plant family of Euphorbiaceae [14], it is known to contain latex, which is applied in traditional medicine for the treatment of cancer and other abnormalities including swellings, fistula, lesions, wounds, abscesses and burns [15,16]. In East Africa, E. ingens is also used for the treatment of snakebites suggesting that the plant may possess an anti-venom for snakes[17]. Additionally, there are scientific data demonstrating ichthyoidal, antitubercular, antimicrobial and antifungal activities of E. ingens [18,19,20]. We have previously shown that E. ingens extract contain phytochemicals including phenols, tannins, terpenoids, flavonoids and saponins, which are generally associated with anticancer activity [21]. However, despite the aforementioned roles of E. ingens as a viable target for the management and treatment of diseases, scientific validation of its ethnobotanical use in cancer management and treatment is lacking. Therefore, we hypothesised that E. ingens has the ability to selectively stop the proliferation of prostate cancer cells while not harming normal cells.
To test this hypothesis, we first applied network pharmacology approaches to determine whether compounds in E. ingens extract have molecular targets associated with PCa and further investigated the antiproliferative activity of the extract so as to validate the putative activity and targets. We showed that E. ingens has selective antiproliferative effect against prostate cancer cell line, with no toxicity towards the non-cancerous Vero cell line.
2. Results
2.1. Plant extract preparation
The percentage yield of the dichloromethane:methanol root extract of E. ingens was 36.25%. Of the four fractions (crude dichloromethane:methanol, hexane, ethyl acetate and water) obtained after solvents partitioning, the crude and hexane fractions dissolved sparingly in dimethyl sulfoxide (DMSO), and so, they were not considered for further bioassays as account for their concentrations would not be possible.
2.2. Antiproliferative & cytotoxic activity
The ethyl acetate and water fractions were evaluated in a blind screening for their potential antiproliferative activity against DU-145 at a fixed concentration of 200µg/ml (Figure 1). Only the ethyl acetate fraction displayed antiproliferative activity of <50% at 200 μg/ml, and therefore, it was prioritized for concentration-dependent testing through 2-fold serial dilution.
The inhibition of cell proliferation by the ethyl acetate fraction in a concentration-response dependent manner against DU-145 is shown in (Figure 2); whereby the IC50 of the fraction was calculated at 9.71 ± 0.4 µg/ml while the standard reference drug, doxorubicin had a IC50 of 5.30 ± 0.11 µg/ml (Supplementary Figure S1). There was significant (p = 0.0115) difference when the two treatments were compared (Table 1).
The fraction in vitro safety was determined using non-cancerous Vero E6 cells (Figure 3). The cytotoxic concetration killing 50% of treated cells (CC50) of the E. ingens ethyl acetate was 80.19 ± 6.12, which is significantly (p ˂ 0.05) different as shown in Table 1 from the doxorubicin drug (176.10 ± 8.09) (Supplementary Figure S2).
2.3. Selectivity index(SI)
The IC50 and CC50 values for the tested ethyl acetate fraction were determined from the data extracted in Figure 2 and Figure 3 respectively; while for doxorubicin from Supplementary Figures S1 and S2 respectively, and the SI calculated. Table 1 shows the summary of all the values, whereby, the IC50 of 9.71 µg/ml and CC50 80.19 µg/ml of the ethyl acetate fraction of E. ingens gave an SI of 8.26; the SI calculated for doxorubicinfor an IC50 of 5.30 µg/ml and CC50 µg/ml of 176.10 is 33.23.
2.4. Identification and analysis of E. ingens bioactive compounds
2.4.1. Qualitative colour method
The results of the qualitative phytochemical screening to identify the class of compounds that to some extent could be responsible for the observed antiproliferative activity of the ethyl acetate fraction is shown in Table 2, whereby, tannins, terpenoids, flavonoids, saponins and sterols were found to be abundant with phenol moderately present. Alkaloid and quinones were not detected.
2.4.2. In-depth compounds characterisation by Gas chromatography–mass spectrometry (GC-MS)
To further identify the specific compounds that were present in the ethyl acetate fraction, GC-MS was used. The chromatogram of GC-MS spectra of compounds is shown in Figure 4. The individual compound’s identification was established on the basis of the peak area, and retention time and the details are presented in Table 3. Twenty two peaks were observed in the chromatogram (Figure 4) comprising 18 compounds (Table 3), majorly terpenoids. The most abundant compounds include 6-pentylidene-4,5-secoandrostane-4,17.beta.-diol (55.39 %), 2-bornanol (16.75 %) and 1-octadecene appearing two times to make up 8.1 %.
2.5. In silico work
2.5.1. Screening for drug-like compounds in E. ingens ethyl acetate fraction
Out of the 18 E. ingens ethyl acetate compounds that were identified through GC-MS analysis, only 7 were considered ideal drug-like candidate using the Lipinski rule of 5; which includes the compounds inability to penetrate the blood brain barrier, as a non-inhibitors of cytochrome P450 enzymes and having topological polar surface area that were less than 140 square angstrom (Å2). The prioritized 7 compounds (1-dodecene, 1-heneicosanol, 1-octadecene, octadecyl trifluoroacetate, andrographolide and squalene) are presented in Table 4 while detailed results on all 18 compounds are shown in Supplementary Table S1.
2.5.2. The targets of E. ingens prioritized drug-like compounds and therapeutic targets for prostate cancer
From the SWISS TargetPrediction (STP) and BindingDB (BDB) databases, a total of 87 potential targets were identified as targets for the 7 prioritized E. ingens compounds (Supplementary Table S2). 76 total targets were retained after removing duplicated targets. Additionally, we retrieved a total of 12,674, and 4,389 target genes that are closely related to PCa from GeneCards and DisGenet, respectively. A total of 12,889 genes were identified after duplicates were eliminated (Supplementary Table S3). Gene datasets obtained from the obtained 76 E. ingens compounds-related targets and the 12,889 PCa-related targets were imported into an online Venn diagram, overlapped and a total of 65 intersecting targets were obtained. The analysed relationship of gene targets between E. ingens(drug) and disease resulted into 65 intersection genes (key targets) shown in Figure 5, and further details are presented in Supplementary Table S4.
2.5.3. Compound-disease target protein–protein interaction (PPI) network
A total of 65 key genes were obtained by mapping the E. ingens drug-like compounds targets to the PCa disease targets. The 65 targets were imported into the STRING database and imported into Cytoscape for visualization and analysis, and a network consisting of 65 nodes and 195 edges was obtained (Figure 6). The average node degree is 6 and average local clustering coefficient of 0.48; the PPI enrichment p-value was < 1.0e-16; thus, proteins have more interactions among themselves than would be expected for a random set of proteins of similar size drawn from the genome. Such a significant enrichment indicated that the proteins are at least partially biologically connected as a group. TTL, ACVRL1, PDE10A and SQLE were not analysed in the PPI network, as they do not interact with other proteins. In the network, the top ten targets with the highest the Maximal Clique Centrality (MCC) scores were estrogen receptor alpha (ESR1), interleukin-6 (IL6), matrix metalloproteinase 9 (MMP9), cyclin-dependent kinase 2 (CDK2), mitogen-activated protein kinase kinase 1 (MAP2K1), androgen receptor (AR), protein kinase C delta (PRKCD), cyclin-dependent kinase 1 (CDK1), Cell Division Cycle 25B (CDC25B) and tyrosine-protein kinase JAK2 (Figure 7).
2.5.4. GO and KEGG pathway enrichment
The 65 key targets of E. ingens against PCa were enriched in 1,418 GO (gene ontology) terms, including 1000 for biological processes (BP) terms, 107 for cellular component (CC) terms, and 311 for molecular functions (MF) terms (p < 0.05). The top 20 significant GO terms for each category are shown in the dot plot chart (Figure 8A-C). The y-axis represents the enriched categories and the x-axis represents the number of enrichment and the order of importance was ranked from the top to bottom by −Log10 (p value). GO analysis showed that the key targets mainly play a role in the biological processes of responding to oxygen-containing compound, lipid and organic cyclic compound. CC was mainly enriched in the membrane raft, membrane microdomain and cyclin-dependent protein kinase holoenzyme complex. MF analysis associated the key targets to protein kinase, phosphotransferase and nuclear receptor activities. KEGG (Kyoto encyclopedia of genes and genomes) signalling pathway analysis indicated that the key targets were significantly enriched in 170 pathways, the top 20 signalling pathways were screened by - LogP values (Figure 9A), and the prostate cancer pathway was selected for analysis as the relevant molecular pathways affected by E. ingens drug-like compounds treatment on the prostate cancer cell line (Figure 9B).
2.6. RT-qPCR results
The mRNA expression of AR, CDK1, p53, BCL-2 and Caspase-3 was measured by RT-qPCR to validate the genes associated with the prostate cancer pathway (Figure 9B) as demonstrated by network pharmacology. AR and CDK1 were among the ten hub genes. Further, we picked p53, BCL-2 and caspase-3 genes due to their involment in tumorigenesis and cancer chemotherapy through deregulation of cell survival and apoptosis. The quantification cycle, also known as the threshold cycle (Ct), was calculated, and relative mRNA expression levels of target genes were normalized to β -Actin using the 2-ΔΔCt method [22]. The results are presented in Figure 10.
There was significant downregulation of AR (p = 0.0129) and BCL2 (p = 0.0002, relative fold change recorded was 5.0). Furthermore, while significant upregulation was also observed in the expression level of p53 (p ˂ 0.0001, relative fold change of 2.5), there was no significant difference in the upregulation of caspase 3 (p = 0.1004). Though, CDK1 was observed to be upregulated, the RT-qPCR results still allude that drug-like compounds in E. ingens ethyl acetate fraction interfere with prostate cancer cells proliferation by inducing apoptosis.
3. Discussion
Active surveillance, surgery, radiotherapy, hormonal therapy, immunotherapy and chemotherapy are treatment options currently available for prostate cancer. These options have major limitations, particularly the accompanying adverse side effects; hence, prostate cancer remains to be incurable. Nevertheless, natural products provide an outlet for identification of promising new anticancer agents that are highly efficient with low toxicity [23]. Cancer cells exhibit sustained proliferation and resistance to cell death; and thus, compounds that can stop or slow down cell proliferation in rapidly dividing cells hold great promise as anticancer therapeutics [24]. Based on the cell-based MTT assay, our findings demonstrated a significant selective antiproliferative effects of extracts from E. ingens roots towards the DU-145 prostate cancer cell lines without affecting the non-cancerous ones (Vero E6). An IC50 < 30 μg/ml is one of the established criteria by the US National Cancer Institute (NCI) to consider crude extracts as being cytotoxic. Interestingly, we observed that the ethyl acetate fractions of E. ingens caused cytotoxicity in DU-145 with IC50 value that was less than 10 µg/mL (Table 1). Therefore, E. ingens has significant potential for further studies as a potential chemotherapeutic agent. The inhibitory effects of E. ingens ethyl acetate on DU-145 is better when compared to that reported for longifolene isolated from Chrysopogon zizanioides (IC50 of 78.64 µg/mL) on the same DU-145 cells [25]. Meanwhile, El-Hawary et al.[26] found E. ingens to be inactive when tested against human colon adenocarcinoma (CACO2), human hepatoma (HepG2) and human breast adenocarcinoma (MCF-7) cell lines. This might be related to the difference in the solvents extraction, hence a likely different set of chemical compounds; while they used the methanolic extract of the plant, we test the ethyl acetate fraction of crude (dichloromethane:methanol) extract. A future study to investigate the cytotoxic effect of ethyl acetate fraction of E. ingens on the CACO2 and HepG2 cell lines is encouraged. Meanwhile, a drug with a measure of the safety margin (selectivity index) of ≥2 is considered highly selective [27]. The ethyl acetate fraction of E. ingens had an SI greater than 2, an indicator of high selectivity for the cancer cells. Previous studies have also shown that E. ingens bioactivities are not a result of general toxicity [19].
The findings of this study depict a high abundance of terpenoids which act on different stages of tumour development and exhibit their anticancer properties by inducing autophagy in cancer cells via a complex signalling pathway [28,29]. In recent study, sesquiterpenoids have been shown to restrict cell cycle in prostate cancer through induction of apoptosis [30]. Flavonoids are known antioxidants under normal conditions and potent pro-oxidants in cancer cells, triggering the apoptotic pathways and downregulating pro-inflammatory signalling pathways [31,32]. Other present phytoconstituents, phenols, tannins, saponins, and sterols, were previously shown to possess anticancer properties. Phenols and tannins have been demonstrated to participate in cell cycle arrest, induce apoptosis, and suppress cancer cell proliferation and invasiveness [32,33]. Similar mechanisms as well as regulation of angiogenesis have been attributed to saponins and sterols [13,34]. Some of the GC-MS-identified compounds in this study have been isolated and reported to elicit anticancer activity while others have been identified to synergistically act with other compounds in many medicinal plants. For example, andrographolide, diterpenoid, that was isolated from Andrographis paniculata Nees induced cell cycle arrest and apoptosis in HT-29 human colon cancer cells [35]. By increasing intracellular reactive oxygen species, squalene induced anti-proliferative activity against ovarian, breast, lung, and colon cancers [36]. Similarly, diterpene phenol, ferruginol has pharmacological properties such as inhibition of the growth rate of cancer cells [37]. Other compounds which have also been reported to have anticancer activity include 1-octadecene, 1-heneicosanol, and 2,4-di-tert-butylphenol [38,39,40]. Given that these compounds are found in the ethyl acetate fraction of E. ingens, we suggest that they may be responsible for the demonstrated antiproliferative effects of the plant on DU-145 cells. However, further research is required to determine the antiproliferative activity of these isolated compounds, particularly on prostate cell lines.
Network pharmacology is currently used in cancer therapy to develop new drugs [41]. The multi-target pathways application of network pharmacology is widely adopted to study the mechanism of action of traditional medicine; it identifies the active ingredients of plants, predict their targets, and subsequently combine them with disease targets to generate a presentable drug-target-disease relationship [42]. We selected 7 GC-MS-identified compounds with good absorption, distribution, metabolism, and excretion (ADME) activity using the RO5, as well as blood–brain barrier, total polar surface area, CYP2D6 and CYP3A4. Drug screening and development relies heavily on pharmacokinetic characteristics. Without appropriate pharmacokinetic qualities, drugs will fail to attain requisite concentration in the target organs where they are needed to produce therapeutic effects [43]. The candidate targets of the selected E. ingens bioactive compounds for prostate cancer treatment were obtained; of which ESR1, IL6, MMP9, CDK2, MAP2K1, AR, PRKCD, CDK1, CDC25B and JAK2 were at the core position in the PPI network. These targets are considered the possible molecular targets of the drug-like compounds in E. ingens ethyl acetate fraction against prostate cancer cells.
The MMPs family is known to have proteolytic effect on the cell membrane; member proteins such as MMP9 release proangiogenic factors which acts on endothelial cells to induce cell migration and proliferation [44,45]. An increased level of MMP9 was reported to cause metastasis in androgen-independent prostate cancer [46]. There have been reports on the role of ESR1, PRKCD, and IL6 in the proliferation and migration of PCa cells, and inhibitors of JAK2 have be suggested to be important in the treatment of advanced PCa [47,48,49,50]. Targeted inhibition of MAP2K1 expression has been shown to elicit cell apoptosis and weakening of cell proliferation in DU-145 and PC-3 prostate cancer cells [51]. The cell cycle protein-dependent kinase (CDK) and cell cycle proteins, as well as CDK inhibitors are essential in the regulation of cell cycle, hence, impairment in the activities of these cell cycle mediators is observed in many types of cancer. CDK2 is a core regulator of cell cycle through late G1-phase and S-phase. CDK2 is thought to be strongly linked to development of cancer, and accumulating evidence shows that inhibition of CDK2 induces cancer cell apoptosis without cellular damage [52]. CDC25B is a cell cycle transitions regulatory enzyme; it is an important target of the checkpoint machinery in maintaining genome stability during DNA damage [53]. Overexpression of CDC25B has been reported in many types of human cancers, and targeted cellular depletion of the enzyme in DU-145 facilitated rapamycin anticancer effects [54]. Given the clear evidence that these genes are involved in the development and progression of prostate cancer, the ability to modulate them would have contributed to the observed antiproliferative property of the ethyl acetate fraction of E. ingens.
The KEGG enrichment results showed that many disease pathways that were not relevant to this study were enriched, probably because the same molecular targets exist in the development of different diseases. The prostate cancer pathway was selected for analysis, the pathway showed a sets of multiple pathways, through which the therapeutic effect of E. ingens on prostate cancer may be produced; these include the PI3K/Akt, MAPK and p53 signalling pathways. The phosphatidylinositol-3 kinase (PI3K)/protein kinase B (Akt) signalling pathway is an important tumour cell pathway which participates in the occurrence, invasion, and distant metastasis of prostate cancer [55]. Long-term ADT can abnormally activate the PI3K/Akt pathway thereby enhancing the antiapoptotic ability of tumour [56]. Therefore, the PI3K/Akt pathway is an important potential target of non-AR pathway in the treatment of PCa. Mitogen-activated protein kinases (MAPKs) are serine-threonine kinases that could link extracellular signals to fundamental cellular processes such as cell growth, proliferation, differentiation, migration, and apoptosis. The p53 pathway is one of the crucial signalling pathways for cancer cell apoptosis [57], as a cancer suppressor gene, p53 regulate the downstream genes and take a part in DNA repair and regulation of the cell cycle and apoptosis. Hence, reactivation and restoration of p53 function holds great potential for the treatment of PCa.
Multiple lines of evidence in the field of cancer research have demonstrated the role of secondary metabolites in natural products is to affect gene expression by influencing significant transcription factors involved in carcinogenesis [58]. We used RT-qPCR to validate the predicted molecular targets of E. ingens in prostate cancer. Overexpression of CDK1 promotes progression of PCa, and when this kinase is hyperactivated, it mediates the phosphorylation that activates AR when there are no ligands [59,60]. AR signalling plays a crucial role in the growth of normal prostate tissue and PCa pathogenesis and progression. In advanced PCa stages, AR mutations and overexpression contribute to sustained proliferation [61]. The treatment of DU-145 prostate cancer cells with E. ingens resulted into a significant downregulation in AR when compared with the untreated control. Similar results were reported on the modulation of AR expression by the flavonoid genistein. [62] However, E. ingens showed upregulation in the expression of CDK1. This unusual finding was also observed by Mustafa et al.[63]. CDK1 has been said to be capable of avoiding an accumulation of oncogenic mutations during cell division, that might explain the result [64]. In cancer cells, the evasion of cell death is often achieved by either upregulating anti-apoptotic proteins like BCL-2 or by impairing the function of pro-apoptotic proteins such as caspase-3. BCL-2 controls the mitochondrial apoptotic pathway by binding to pro-apoptotic proteins and preventing pore formation and cytochrome c release. Apoptosis is executed by caspases and various upstream regulatory factors, including p53. We observed an upregulation in the expression of caspase-3 and p53 with concomitant downregulation of BCL-2 when E. ingens-treated prostate cancer cells were compared with the untreated prostate cancer cells; this pattern was also reported by Eltamany et al.[65]. Moreover, recent computational study revealed that flavonoids, found to be abundant in our study, have strong interaction with caspase-3, BCL-2 and p53 [66]. The difference in the expression level of AR, p53, BCL-2 and caspase-3 between the E. ingens-treated human prostate cancer cells and the untreated cells justifies the network pharmacology predictions; as these genes are distributed in the PI3K/Akt, MAPK and p53 signalling pathways, the results further suggest that E. ingens’ antiprofelirative activity is likely by modulating the pathways to cause induction of apoptosis as well as suppression of cell cycle.
4. Materials and Methods
4.1. Plant collection and extract preparation
Fresh root samples of E. ingens were obtained on 26th March, 2022 from Embu County, Kenya (0° 46' 27.0" South, 37° 40' 54.9" East) where it grows naturally. Plant identification and authentication was carried out at Egerton University, Kenya by a plant taxonomist, and a voucher specimen number NSN9 was subsequently deposited in the same place. The dried roots of the plant were extracted using dichloromethane:methanol as previously described [21]. The obtained extract of 149 g and the yield percentage was calculated using the following equation [67]:
Percentage yield of extracts = [(Weight of the obtained extract material/Weight of original fine plant powder used)* 100]
We further partitioned the crude (dichloromethane:methanol) extract into hexane, ethyl acetate and water fractions. 20 g of dried crude extract were weighed and dissolved in 50 ml of a 1:1 solution of dichloromethane and methanol. In a separating funnel, 300 ml of hexane was mixed with the solution that was made. The mixture was then left undisturbed for 30 minutes to ensure proper separation. The top hexane part was then put in a beaker. The lower part was washed over and over with hexane, and the different fractions of hexane were collected and concentrated. This process was repeated using 300 ml of ethyl acetate and distilled water (1:1). At the end, the ethyl acetate fraction was collected and concentrated, while the bottom distilled water part was taken for freeze drying. Following partitioning, the crude, and obtained hexane, ethyl acetate, and aqueous fractions were stored at -20 ˚C until further analysis.
4.2. Cell culture
A human prostate carcinoma cell line (DU-145) and a kidney epithelial cell line derived from African green monkeys (Vero E6) were purchased from the American Type Culture Collection (ATCC) and cultured at the Kenya Medical Research Institute's Centre for Traditional Medicine and Drug Research. The cells were grown in Modified Eagle's Medium (MEM) supplemented with 1% L-glutamine (200 mM), 10 % fetal bovine serum (FBS), 1.5 % sodium bicarbonate, 1 % HEPES (1 M), 1% penicillin/streptomycin, and 0.4 % phenol red at 37 °C and 5 % CO2. The logarithmic growth phase of the cells was used in all studies.
4.3. Cellular proliferation assay
The antiproliferative activity of tested fractions was assessed using the 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) assay, as previously described [68]. The DU-145 (1 × 104 cells/well) were seeded in 96-well plates and grown for 24 h. The seeding medium was aspirated from the plates, and 100 µl of a 200 µg/ml working concentration of plant fractions were added for 48 h as screening treatments. The same concentration of doxorubicin was used as a positive control, while 0.2 % dimethyl sulfoxide (DMSO) was used as a negative control. 10 μl of freshly prepared MTT (5 mg/ml) was added to each well. After 4 h incubation, MTT was aspirated out and 100 μl of 100% DMSO was added to solubilize the formazan crystals. Plates were then read on an Infinite M1000 by Tecan (a plate reader) using the absorbance at 570 nm. Experiments were carried out in triplicates, and the percentage cell survival was calculated using the formula below [24]:
Percentage cell viability = [(Absorbance of treated cells –Absorbance of culture medium/Absorbance of untreated cells –Absorbance of culture medium) * 100]
The extracts with equal or less than 50% cell viability after 48 hours of treatments were considered active [69]. After the initial screening at a single concentration (200 µg/ml), only the active fraction was selected for further anti-proliferative studies using a range of concentrations (6.25-200 µg/ml) and the 50% inhibition concentration (IC50) was determined. The cytotoxicity of E. ingens fraction was further assessed on Vero E6 cells using the MTT cell proliferation assay previously described, and the 50% cytotoxicity concentration (CC50) was calculated.
4.4. Selectivity index
The selectivity index (SI) for each of the tested plant fraction was determined by dividing the IC50 by the corresponding CC50.
4.5. Identification of plant compounds
The plant fraction that showed antiproliferative activity was subjected to the colour change qualitative phytochemical screening and GC-MS analysis as previously described [21]. This was to identify the compounds responsible for the activity as well as the class of the bioactive compounds.
4.6. In silico work
4.6.1. Drug candidate screening test
The Canonical SMILES (Simplified Molecular Input Line Entry System) of the GC-MS identified compounds were retrieved from PubChem (https://pubchem.ncbi.nlm.nih.gov/). The SMILES were then submitted to the Swiss ADME (absorption, distribution, metabolism, and excretion) tool (http://www.swissadme.ch/index.php) to predict the drug likeness and physicochemical properties of the compounds [70]. The prediction was based on the parameters of the blood–brain barrier, total polar surface area, cytochrome P450s (CYP2D6 and CYP3A4) and Lipinski’s rule of five (RO5) (a drug candidate should have a molecular weight of fewer than 500 daltons, a number of rotatable bonds of less than 10, a number of hydrogen bond donors of less than 5, a number of hydrogen bond acceptors of less than 10, and a lipophilicity (log P) value of less than 5). It was predicted that a molecule would be a non-orally accessible medicine if two or more of the RO5 were not met [71].
The polar atoms of a molecule add up to the topological polar surface area (TPSA), which can be used to predict how the drug will be transported. In most cases, the amount of TPSA in an approved medicine was below 140 Å2 [70]. Substances that are able to breach the blood-brain barrier (BBB) can be hazardous to the nervous system because they can pass from the more hydrophilic blood to the more lipophilic brain. A drug candidate should not block cytochrome P450 enzymes (such as CYP2D6 and CYP3A4) because they are essential for drug metabolism [70].
4.6.2. Identification of candidate targets in Euphorbia ingens against Pca
Compounds targets were predicted using BindingDB (https://bindingdb.org/rwd/bind/chemsearch/marvin/FMCT.jsp) correspondence to the known ligand molecules having minimum similarity of >0.7 and their Gene IDs were retrieved from UniProtKB (https://www.uniprot.org) database [72]. Similarly, the SMILES of the compounds were uploaded to the Swiss TargetPrediction (http://www.swisstargetprediction.ch/)database, with “humans” (Homo sapiens) as the study species, and the probability of each potential target was determined to be >0 [73]. The retrieved targets were converted into standardized abbreviations by UniProt. The resulting predicted compounds targets from the two databases were pooled together and duplicates were removed [74].
Disease targets of PCa were collected from the GeneCards (https://www.genecards.org/) and the DisGeNET (https://www.disgenet.org/) databases [75]. The targets from the databases were searched using the keyword “prostate cancer”; retrieved results were merged, and ensured there was no repeats. Finally, the targets of active compounds of E. ingens and disease targets of PCa were intersected using the bioinformatics and evolutionary genomics platform (https://bioinformatics.psb.ugent.be/webtools/Venn/).
4.6.3. Construction of the protein-protein interaction (PPI) network
The intersection obtained, which are considered the common potential targets were up loaded to the STRING 11.5 database (https://string-db.org/), the species was set as “Homo sapiens”, and the minimum interaction threshold was set to 0.4. The cytohubba plug-in in Cytoscape software (version 3.9.1) was used to to analyse the topology of the network and the Maximal Clique Centrality (MCC) algorithm was used to filter out the top 10 key targets [76].
4.6.4. Gene Ontology (GO) & Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis
The GO and KEGG enrichment analyses for the targets of E. ingens for PCa treatment were conducted in the enrichment tool ShinyGO version 0.76 (http://ge-lab.org/go/). The following parameters were considered for the analyses: species = Human, false discovery rate (FDR) cut-off = 0.05, and number of pathways to show = 20. The biological process (BP), the cellular component (CC), and the molecular function (MF) categories were considered for the gene ontology analysis.
4.7. Quantitative real-time polymerase chain reaction (RT-qPCR)
Eighty (80) % confluent DU-145 in T-25 flask were treated with E. ingen fraction at concentration equivalent to the calculated IC50. Negative control cells were exposed to fresh growth media with 0.2 percent DMSO. The cells were incubated for 48 h. Cells were collected for the extraction of total RNA and the reverse transcription was conducted using FIRE Script RT cDNA synthesis kit (Solis BioDyne, Estonia). Luna Universal qPCR Master Mix (New England Biolabs) was used for RT-qPCR) detection. The primer sequences (Macrogen Europe BV, Netherlands) are listed as follows:
4.8. Statistical analysis
GraphPad Prism version 8.4.0 software (San Diego, CA, USA) was used for statistical analysis. Data was presented as the mean ± SEM. An independent-sample T test was used for the comparison between two groups and one-way ANOVA was used for the comparison between multiple groups, and p < 0.05 was considered as statistically significant.
5. Conclusions
Our findings are the first to demonstrate that E. ingens has a selective cytotoxicity effect against prostate cancer cell line. Network pharmacology pointed to ESR1, IL6, MMP9, CDK2, MAP2K1, AR, PRKCD, CDK1, CDC25B and JAK2 as likely molecular targets of E. ingens against prostate cancer, with the regulation of the PI3K/Akt, MAPK and p53 signalling pathways as the plant’s molecular mechanism of action. Interestingly, the expression level of AR, p53, caspase-3 and BCL-2 was reversed in E. ingens-treated prostate cancer cells when they were compared with the untreated cancer cells. Additional use of other prostate cancer cell lines (PC3, and LNCaP) might have further added to the robustness of our data; this we considered the limitation of this study. Nonetheless, ethyl acetate fraction of E. ingens could be a source of new effective and safe alternative to current prostate cancer treatments. However, additional in vivo research is necessary to better enhance the potential clinical use of the plant.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
I.O.: Conceptualization; Methodology; Data curation; Writing—Original draft preparation. F.N.: Conceptualization; Supervision; Reviewing and Editing. M.K.: Conceptualization, Supervision; Reviewing and Editing. J.O.: Conceptualization, Reviewing and Editing. S.N.: Conceptualization; Methodology; Supervision; Reviewing and Editing. All authors have read and agreed to the published version of the manuscript.
Funding
The African Union scholarship program funded the research through the Pan African University Institute for Basic Sciences, Technology, and Innovation (PAUSTI), REF: PAU/ADM/PAUSTI/2020/8; and the APC was supported by F.N.; M.K.; S.N.; J.O.; and I.O.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We acknowledge PAUSTI, the Centre for Traditional Medicine and Drug Research, Kenya Medical Research Institute (KEMRI), and Jomo Kenyatta University of Agriculture and Technology for provision of laboratories and resources.
Conflicts of Interest
The authors declare no conflict of interest.
Sample Availability
Samples of the compounds are not available from the authors.
References
- Bray, F.; Laversanne, M.; Weiderpass, E.; Soerjomataram, I. The Ever-Increasing Importance of Cancer as a Leading Cause of Premature Death Worldwide. Cancer 2021, 127, 3029–3030. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Shen, M.M. Cell Types of Origin for Prostate Cancer. Curr. Opin. Cell Biol. 2015, 37, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Thorpe, C.S.; Bhangoo, R.S.; Anderson, J.D.; Jason Shen, J.; Vargas, C.E. Prostate Cancer. Princ. Pract. Part. Ther. 2023, 383–410. [Google Scholar] [CrossRef]
- Belkahla, S.; Nahvi, I.; Biswas, S.; Nahvi, I.; Ben Amor, N. Advances and Development of Prostate Cancer, Treatment, and Strategies: A Systemic Review. Front. Cell Dev. Biol. 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Ekwan, R.; Bua, E.; Nantale, R.; Opito, R.; Abingwa, P.; Sserwanja, Q.; Kuteesa, J.; Mukunya, D. Uptake of Prostate Cancer Screening and Associated Factors among Men Aged 50 Years and above in Lira City, Uganda: A Cross-Sectional Study. BMC Public Health 2023, 23, 432. [Google Scholar] [CrossRef]
- Nowroozi, A.; Roshani, S.; Ghamari, S.-H.; Shobeiri, P.; Abbasi-Kangevari, M.; Ebrahimi, N.; Rezaei, N.; Yoosefi, M.; Malekpour, M.-R.; Rashidi, M.-M.; et al. Global and Regional Quality of Care Index for Prostate Cancer: An Analysis from the Global Burden of Disease Study 1990–2019. Arch. Public Heal. 2023, 81, 70. [Google Scholar] [CrossRef] [PubMed]
- Bosland, M.C.; Shittu, O.B.; Ikpi, E.E.; Akinloye, O. Potential New Approaches for Prostate Cancer Management in Resource-Limited Countries in Africa. Ann. Glob. Heal. 2023. [Google Scholar] [CrossRef]
- Thomas, T.S.; Pachynski, R.K. Treatment of Advanced Prostate Cancer. Mo. Med. 2018, 115, 156. [Google Scholar] [CrossRef]
- Shore, N.; Garcia-Horton, V.; Terasawa, E.; Ayyagari, R.; Grossman, J.P.; Adrianus, &; Waldeck, R. Safety Differences across Androgen Receptor Inhibitors in Nonmetastatic Castration-Resistant Prostate Cancer. https://doi.org/10.2217/fon-2022-1123 2023. [CrossRef]
- Okaiyeto, K.; Oguntibeju, O.O. African Herbal Medicines: Adverse Effects and Cytotoxic Potentials with Different Therapeutic Applications. Int. J. Environ. Res. Public Health 2021, 18. [Google Scholar] [CrossRef]
- Salehi, B.; Fokou, P.V.T.; Yamthe, L.R.T.; Tali, B.T.; Adetunji, C.O.; Rahavian, A.; Mudau, F.N.; Martorell, M.; Setzer, W.N.; Rodrigues, C.F.; et al. Phytochemicals in Prostate Cancer: From Bioactive Molecules to Upcoming Therapeutic Agents. Nutrients 2019, 11. [Google Scholar] [CrossRef]
- Khan, T.; Ali, M.; Khan, A.; Nisar, P.; Jan, S.A.; Afridi, S.; Shinwari, Z.K. Anticancer Plants: A Review of the Active Phytochemicals, Applications in Animal Models, and Regulatory Aspects. Biomolecules 2019, 10, 47. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Tan, L.; Chen, M.; He, C. Pharmacological Activities and Molecular Mechanisms of Pulsatilla Saponins. Chinese Med. (United Kingdom) 2022, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Simpson, M.G. Diversity and Classification of Flowering Plants: Eudicots. Plant Syst. 2010, 275–448. [Google Scholar] [CrossRef]
- Jain, V.; Dudhal, A.; Chitale, R.; Bankar, P. A Survey of Some Medicinally Important Plants of the Family Euphorbiaceae from Baramati Area of Pune District of Maharashtra, India. Prasad Bank. 2021, 6, 456–462. [Google Scholar]
- Koduru, S.; Grierson, D.S.; Afolayan, A.J. Ethnobotanical Information of Medicinal Plants Used for Treatment of Cancer in the Eastern Cape Province, South Africa. Curr. Sci. 2017, 92, 906–908. [Google Scholar]
- Ernst, M.; Grace, O.M.; Saslis-Lagoudakis, C.H.; Nilsson, N.; Simonsen, H.T.; Rønsted, N. Global Medicinal Uses of Euphorbia L.(Euphorbiaceae). J. Ethnopharmacol. 2015, 176, 90–101. [Google Scholar] [CrossRef]
- Kena, M.A. Antifungal Activities of Monsonia Burkeana and Euphorbia Ingens Extracts against Penicillium Digitatum, the Causal Agent of Citrus Green Mould. African Plant Prot. 2016, 19, 1–3. [Google Scholar] [CrossRef]
- Njeru, S.N.; Meshack, A.O. Potency of Extracts of Selected Plant Species from Mbeere, Embu County-Kenya against Mycobacterium Tuberculosis. J. Med. Plants Res. 2016, 10, 149–157. [Google Scholar] [CrossRef]
- Ross, M.J.; Steyn, G.J. Preliminary Results on the Ichthyocidal Properties of Euphorbia Ingens (Euphorbiaceae). African J. Aquat. Sci. 2004, 29, 265–269. [Google Scholar] [CrossRef]
- Okpako, I.O.; Ng’ong’a, F.A.; Kyama, M.C.; Njeru, S.N. Phytochemical Screening and Gas Chromatography- Mass Spectrometry Analysis of Euphorbia Ingens Organic Root Extract. J. Med. Plants Res. 2023, 17, 100–105. [Google Scholar] [CrossRef]
- Lalremruati, M.; Lalmuansangi, C.; Zosangzuali, M.; Tochhawng, L.; Trivedi, A.K.; Kumar, N.S.; Siama, Z. Mussaenda Macrophylla Wall. Exhibit Anticancer Activity against Dalton’s Lymphoma Ascites (DLA) Bearing Mice via Alterations of Redox-Homeostasis and Apoptotic Genes Expression. J. Basic Appl. Zool. 2022 831 2022, 83, 1–14. [Google Scholar] [CrossRef]
- Al Kury, L.T.; Taha, Z.; Mahmod, A.I.; Talib, W.H. Xanthium Spinosum L. Extracts Inhibit Breast Cancer in Mice by Apoptosis Induction and Immune System Modulation. Pharmaceuticals 2022, 15, 1504. [Google Scholar] [CrossRef]
- Eghianruwa, Q.; Osoniyi, O.; Wachira, S.; Maina, N.; Mbugua, R.; Imbuga, M. In Vitro Antiproliferative Studies of Extracts of the Marine Molluscs: Tympanatonus Fuscatus Var Radula (Linnaeus) and Pachymelania Aurita (Muller). Int. J. Biochem. Mol. Biol. 2019, 10, 1–8. [Google Scholar] [PubMed]
- Grover, M.; Behl, T.; Virmani, T.; Sanduja, M.; Makeen, H.A.; Albratty, M.; Alhazmi, H.A.; Meraya, A.M.; Bungau, S.G. Exploration of Cytotoxic Potential of Longifolene/Junipene Isolated from Chrysopogon Zizanioides. Molecules 2022, 27. [Google Scholar] [CrossRef] [PubMed]
- El-Hawary, S.S.; Mohammed, R.; Tawfike, A.F.; Lithy, N.M.; Abouzid, S.F.; Amin, M.N.; Abdelmohsen, U.R.; Amin, E. Cytotoxic Activity and Metabolic Profiling of Fifteen Euphorbia Species. Metabolites 2020, 11, 15. [Google Scholar] [CrossRef] [PubMed]
- Chikwana, N.; Maina, E.N.; Gavamukulya, Y.; Bulimo, W.; Wamunyokoli, F. Antiproliferative Activity, c-Myc and FGFR1 Genes Expression Profiles and Safety of Annona Muricata Fruit Extract on Rhabdomyosarcoma and BALB/c Mice. J. Complement. Altern. Med. Res. 2021, 14, 30–46. [Google Scholar] [CrossRef]
- El-Baba, C.; Baassiri, A.; Kiriako, G.; Dia, B.; Fadlallah, S.; Moodad, S.; Darwiche, N. Terpenoids’ Anti-Cancer Effects: Focus on Autophagy. Apoptosis 2021, 26, 491–511. [Google Scholar] [CrossRef]
- Ansari, I.A.; Akhtar, M.S. Current Insights on the Role of Terpenoids as Anticancer Agents: A Perspective on Cancer Prevention and Treatment. Nat. Bio-active Compd. Chem. Pharmacol. Heal. Care Pract. 2019, 2, 53–80. [Google Scholar] [CrossRef]
- Dhyani, P.; Sati, P.; Sharma, E.; Attri, D.C.; Bahukhandi, A.; Tynybekov, B.; Szopa, A.; Sharifi-Rad, J.; Calina, D.; Suleria, H.A.R.; et al. Sesquiterpenoid Lactones as Potential Anti-Cancer Agents: An Update on Molecular Mechanisms and Recent Studies. Cancer Cell Int. 2022, 22, 1–18. [Google Scholar] [CrossRef]
- Ansari, I.A.; Akhtar, M.S. Recent Insights on the Anticancer Properties of Flavonoids: Prospective Candidates for Cancer Chemoprevention and Therapy. Nat. Bio-active Compd. Prod. Appl. 2019, 1, 425–448. [Google Scholar] [CrossRef]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as Anticancer Agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Zhang, J.; Chen, N.G.; Shi, Z.; Qiu, J.; He, C.; Chen, M. Recent Advances in Anticancer Activities and Drug Delivery Systems of Tannins. Med. Res. Rev. 2017, 37, 665–701. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Quispe, C.; Sharifi-Rad, J.; Cruz-Martins, N.; Nigam, M.; Mishra, A.P.; Konovalov, D.A.; Orobinskaya, V.; Abu-Reidah, I.M.; Zam, W. Phytosterols: From Preclinical Evidence to Potential Clinical Applications. Front. Pharmacol. 2021, 11, 1819. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Khan, F.; Farooqui, A.; Ansari, I.A. Andrographolide Exhibits Anticancer Potential Against Human Colon Cancer Cells by Inducing Cell Cycle Arrest and Programmed Cell Death via Augmentation of Intracellular Reactive Oxygen Species Level. Nutr. Cancer 2018, 70, 787–803. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Grande, M.A.; Gorinstein, S.; Espitia-Rangel, E.; Dávila-Ortiz, G.; Martínez-Ayala, A.L. Plant Sources, Extraction Methods, and Uses of Squalene. Int. J. Agron. 2018, 2018, 1–13. [Google Scholar] [CrossRef]
- Xiong, W.D.; Gong, J.; Xing, C. Ferruginol Exhibits Anticancer Effects in OVCAR-3 Human Ovary Cancer Cells by Inducing Apoptosis, Inhibition of Cancer Cell Migration and G2/M Phase Cell Cycle Arrest. Mol. Med. Rep. 2017, 16, 7013–7017. [Google Scholar] [CrossRef]
- Seenivasan, A.; Manikkam, R.; Kaari, M.; Sahu, A.K.; Said, M.; Dastager, S.G. 2,4-Di-Tert-Butylphenol (2,4-DTBP) Purified from Streptomyces Sp. KCA1 from Phyllanthus Niruri: Isolation, Characterization, Antibacterial and Anticancer Properties. J. King Saud Univ. - Sci. 2022, 34, 102088. [Google Scholar] [CrossRef]
- Tonisi, S.; Okaiyeto, K.; Hoppe, H.; Mabinya, L. V; Nwodo, U.U.; Okoh, A.I. Chemical Constituents, Antioxidant and Cytotoxicity Properties of Leonotis Leonurus Used in the Folklore Management of Neurological Disorders in the Eastern Cape, South Africa. 3 Biotech 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Thekkangil, A.; George, B.; Prakash, S.M.U.; Suchithra, T. V Mechanism of Streptomyces Albidoflavus STV1572a Derived 1-Heneicosanol as an Inhibitor against Squalene Epoxidase of Trichophyton Mentagrophytes. Microb. Pathog. 2021, 154, 104853. [Google Scholar] [CrossRef]
- Zheng, J.; Wu, M.; Wang, H.; Li, S.; Wang, X.; Li, Y.; Wang, D.; Li, S. Network Pharmacology to Unveil the Biological Basis of Health-Strengthening Herbal Medicine in Cancer Treatment. Cancers (Basel). 2018, 10, 461. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Fu, L.; Xu, P.; Wang, T.; Li, P. Network Pharmacology and Experimental Validation to Explore the Effect and Mechanism of Kanglaite Injection Against Triple-Negative Breast Cancer. Drug Des. Devel. Ther. 2023, 17, 901–917. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Wang, J.; Li, Y.; Li, D.; Xu, L.; Hou, T. The Application of in Silico Drug-Likeness Predictions in Pharmaceutical Research. Adv. Drug Deliv. Rev. 2015, 86, 2–10. [Google Scholar] [CrossRef]
- Conway, E.M.; Collen, D.; Carmeliet, P. Molecular Mechanisms of Blood Vessel Growth. Cardiovasc. Res. 2001, 49, 507–521. [Google Scholar] [CrossRef] [PubMed]
- Libra, M.; Scalisi, A.; Vella, N.; Clementi, S.; Sorio, R.; Stivala, F.; Spandidos, D.A.; Mazzarino, C. Uterine Cervical Carcinoma: Role of Matrix Metalloproteinases (Review). Int. J. Oncol. 2009, 34, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.C.; Peng, C.C.; Peng, C.H.; Hsieh, C.L.; Peng, R.Y. The Aqueous Soluble Polyphenolic Fraction of Psidium Guajava Leaves Exhibits Potent Anti-Angiogenesis and Anti-Migration Actions on DU145 Cells. Evidence-based Complement. Altern. Med. 2011, 2011. [Google Scholar] [CrossRef]
- Beinhoff, P.; Sabharwal, L.; Udhane, V.; Maranto, C.; LaViolette, P.S.; Jacobsohn, K.M.; Tsai, S.; Iczkowski, K.A.; Wang, L.; Hall, W.A.; et al. Second-Generation Jak2 Inhibitors for Advanced Prostate Cancer: Are We Ready for Clinical Development? Cancers (Basel). 2021, 13. [Google Scholar] [CrossRef]
- Jariwala, U.; Prescott, J.; Jia, L.; Barski, A.; Pregizer, S.; Cogan, J.P.; Arasheben, A.; Tilley, W.D.; Scher, H.I.; Gerald, W.L.; et al. Identification of Novel Androgen Receptor Target Genes in Prostate Cancer. Mol. Cancer 2007, 6, 39. [Google Scholar] [CrossRef]
- Chen, S.; Lu, K.; Hou, Y.; You, Z.; Shu, C.; Wei, X.; Wu, T.; Shi, N.; Zhang, G.; Wu, J.; et al. YY1 Complex in M2 Macrophage Promotes Prostate Cancer Progression by Upregulating IL-6. J. Immunother. Cancer 2023, 11, e006020. [Google Scholar] [CrossRef]
- Palicelli, A.; Croci, S.; Bisagni, A.; Zanetti, E.; De Biase, D.; Melli, B.; Sanguedolce, F.; Ragazzi, M.; Zanelli, M.; Chaux, A. What Do We Have to Know about PD-L1 Expression in Prostate Cancer? A Systematic Literature Review (Part 6): Correlation of PD-L1 Expression with the Status of Mismatch Repair System, BRCA, PTEN, and Other Genes. Biomedicines 2022, 10, 236. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Li, Y.N.; Mu, X.; Pan, Z.L.; Liu, W.B. Targeted Regulation of MiR-195 on MAP2K1 for Suppressing ADM Drug Resistance in Prostate Cancer Cells. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 8599–8608. [Google Scholar] [CrossRef] [PubMed]
- Tadesse, S.; Anshabo, A.T.; Portman, N.; Lim, E.; Tilley, W.; Caldon, C.E.; Wang, S. Targeting CDK2 in Cancer: Challenges and Opportunities for Therapy. Drug Discov. Today 2020, 25, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Boutros, R.; Lobjois, V.; Ducommun, B. CDC25 Phosphatases in Cancer Cells: Key Players? Good Targets? Nat. Rev. Cancer 2007, 7, 495–507. [Google Scholar] [CrossRef]
- Chen, R.Q.; Yang, Q.K.; Lu, B.W.; Yi, W.; Cantin, G.; Chen, Y.L.; Fearns, C.; Yates, J.R.; Lee, J.D. CDC25B Mediates Rapamycin-Lnduced Oncogenic Responses in Cancer Cells. Cancer Res. 2009, 69, 2663–2668. [Google Scholar] [CrossRef] [PubMed]
- Makwana, V.; Rudrawar, S.; Anoopkumar-Dukie, S. Signalling Transduction of O-GlcNAcylation and PI3K/AKT/MTOR-Axis in Prostate Cancer. Biochim. Biophys. acta. Mol. basis Dis. 2021, 1867. [Google Scholar] [CrossRef] [PubMed]
- Pisano, C.; Tucci, M.; Di Stefano, R.F.; Turco, F.; Scagliotti, G.V.; Di Maio, M.; Buttigliero, C. Interactions between Androgen Receptor Signaling and Other Molecular Pathways in Prostate Cancer Progression: Current and Future Clinical Implications. Crit. Rev. Oncol. Hematol. 2021, 157, 103185. [Google Scholar] [CrossRef]
- He, R.; Ou, S.; Chen, S.; Ding, S. Network Pharmacology-Based Study on the Molecular Biological Mechanism of Action for Compound Kushen Injection in Anti-Cancer Effect. Med. Sci. Monit. 2020, 26, 1–15. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Lee, J.H.; Chai, E.Z.P.; Kanchi, M.M.; Kar, S.; Arfuso, F.; Dharmarajan, A.; Kumar, A.P.; Ramar, P.S.; Looi, C.Y.; et al. Cancer Prevention and Therapy through the Modulation of Transcription Factors by Bioactive Natural Compounds. Semin. Cancer Biol. 2016, 40–41, 35–47. [Google Scholar] [CrossRef]
- Huang, Y.; Lu, S.; Chen, Y.; Qing, Y.; Wu, R.; Ma, T.; Zhang, Z.; Wang, Y.; Li, K. Verification of Cell Cycle-Associated Cyclin-Dependent Kinases Facilitated Prostate Cancer Progression by Integrated Bioinformatic Analysis and Experimental Validation. Heliyon 2022, 8, e10081. [Google Scholar] [CrossRef]
- Gao, X.T.; Liang, J.; Wang, L.Y.; Zhang, Z.; Yuan, P.; Wang, J.; Gao, Y.; Ma, F.; Calagua, C.; Ye, H.; et al. Phosphorylation of the Androgen Receptor at Ser81 Is Co-Sustained by CDK1 and CDK9 and Leads to AR-Mediated Transactivation in Prostate Cancer. Mol. Oncol. 2021, 15, 1901–1920. [Google Scholar] [CrossRef]
- Hatano, K.; Nonomura, N. Systemic Therapies for Metastatic Castration-Resistant Prostate Cancer: An Updated Review. World J. Mens. Health 2023, 41. [Google Scholar] [CrossRef] [PubMed]
- Lazarevic, B.; Karlsen, S.J.; Saatcioglu, F. Genistein Differentially Modulates Androgen-Responsive Gene Expression and Activates JNK in LNCaP Cells. Oncol. Rep. 2008, 19, 1231–1235. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, K.; Mohamed, H.; Shah, A.M.; Yu, S.; Akhlaq, M.; Xiao, H.; Li, S.; Naz, T.; Nosheen, S.; Bai, X. In Vitro Anticancer Potential of Berberis Lycium Royle Extracts against Human Hepatocarcinoma (HepG2) Cells. Biomed Res. Int. 2020, 2020. [Google Scholar] [CrossRef] [PubMed]
- Malumbres, M.; Barbacid, M. Cell Cycle, CDKs and Cancer: A Changing Paradigm. Nat. Rev. cancer 2009, 9, 153–166. [Google Scholar] [CrossRef]
- Eltamany, E.E.; Goda, M.S.; Nafie, M.S.; Abu-Elsaoud, A.M.; Hareeri, R.H.; Aldurdunji, M.M.; Elhady, S.S.; Badr, J.M.; Eltahawy, N.A. Comparative Assessment of the Antioxidant and Anticancer Activities of Plicosepalus Acacia and Plicosepalus Curviflorus: Metabolomic Profiling and In Silico Studies. Antioxidants 2022, 11, 1249. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.C.; Kamarudin, M.N.A.; Naidu, R. Anticancer Mechanism of Flavonoids on High-Grade Adult-Type Diffuse Gliomas. Nutrients 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Evans, W.C. Trease and Evans’ Pharmacognosy: Sixteenth Edition; Elsevier Inc., 2009; ISBN 9780702029332.
- Sergazy, S.; Vetrova, A.; Orhan, I.E.; Senol Deniz, F.S.; Kahraman, A.; Zhang, J.Y.; Aljofan, M. Antiproliferative and Cytotoxic Activity of Geraniaceae Plant Extracts against Five Tumor Cell Lines. Futur. Sci. OA 2022, 8. [Google Scholar] [CrossRef]
- Fadeyi, S.A.; Fadeyi, O.O.; Adejumo, A.A.; Okoro, C.; Myles, E.L. In Vitro Anticancer Screening of 24 Locally Used Nigerian Medicinal Plants. BMC Complement. Altern. Med. 2013, 13, 79. [Google Scholar] [CrossRef]
- Navid, M.; Rad, S.; Behrouz, S.; Aghajani, S.; Behrouz, M.; Zarenezhad, E.; Ghanbariasad, A. Design, Synthesis, Anticancer and in Silico Assessment of 8-Caffeinyl-Triazolylmethoxy Hybrid Conjugates. RSC Adv. 2023, 13, 3056–3070. [Google Scholar] [CrossRef]
- Patel, N.; Patel SmtSSPatel, L.N.; Patel, N.B.; Patel, L.N.; Patel, K.D.; Patel, M. V; Kalasariya, H.S. ADMET & Cytotoxicity Prediction of Red Seaweed Gracillaria Dura: An in Silico Approach. Artic. World J. Pharm. Res. 2020, 9, 991–1004. [Google Scholar] [CrossRef]
- Borse, S.; Joshi, M.; Saggam, A.; Bhat, V.; Walia, S.; Marathe, A.; Sagar, S.; Chavan-Gautam, P.; Girme, A.; Hingorani, L.; et al. Ayurveda Botanicals in COVID-19 Management: An in Silico Multi-Target Approach. PLoS One 2021, 16, e0248479. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A.; Wu, Y.; Li, A.S.M.; Fu, X.Q.; Yu, Z.L. Network Pharmacology and Molecular Docking-Based Prediction of Active Compounds and Mechanisms of Action of Cnidii Fructus in Treating Atopic Dermatitis. BMC Complement. Med. Ther. 2022, 22, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Tashrifi, Z.; Mohammadi-Khanaposhtani, M.; Hamedifar, H.; Larijani, B.; Ansari, S.; Mahdavi, M. Synthesis and Pharmacological Properties of Polysubstituted 2-Amino-4H-Pyran-3-Carbonitrile Derivatives. Mol. Divers. 2020, 24, 1385–1431. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Lin, J.; Zhang, N.; Wang, Q.; Li, N.; Yao, K. Effect of Xuefu Zhuyu Capsule on Myocardial Infarction: Network Pharmacology and Experimental Verification. Evidence-based Complement. Altern. Med. 2023, 2023. [Google Scholar] [CrossRef]
- Zhu, B.-J.; Nai, G.-Y.; Pan, T.-X.; Ma, Z.-F.; Huang, Z.-D.; Shi, Z.-Z.; Pang, Y.-H.; Li, N.; Lin, J.-X.; Ling, G.-M. To Explore the Active Constituents of Sedum Aizoon L in the Treatment of Coronary Heart Disease Based on Network Pharmacology and Molecular Docking Methodology. Ann. Transl. Med. 2022, 10, 1327–1327. [Google Scholar] [CrossRef]
Figure 1.
Screening for cellular proliferation inhibition of E. ingens fractions: Inhibition of cellular proliferation following 48 h treatment at 200 µg/ml concentration of the water and ethyl acetate fractions of E. ingens on DU-145. Doxorubicin at 200 µg/ml was used as positive control, and 0.2% DMSO as negative control. Values are expressed as Mean ± SEM. All treatments were done in triplicates (n = 3).
Figure 1.
Screening for cellular proliferation inhibition of E. ingens fractions: Inhibition of cellular proliferation following 48 h treatment at 200 µg/ml concentration of the water and ethyl acetate fractions of E. ingens on DU-145. Doxorubicin at 200 µg/ml was used as positive control, and 0.2% DMSO as negative control. Values are expressed as Mean ± SEM. All treatments were done in triplicates (n = 3).
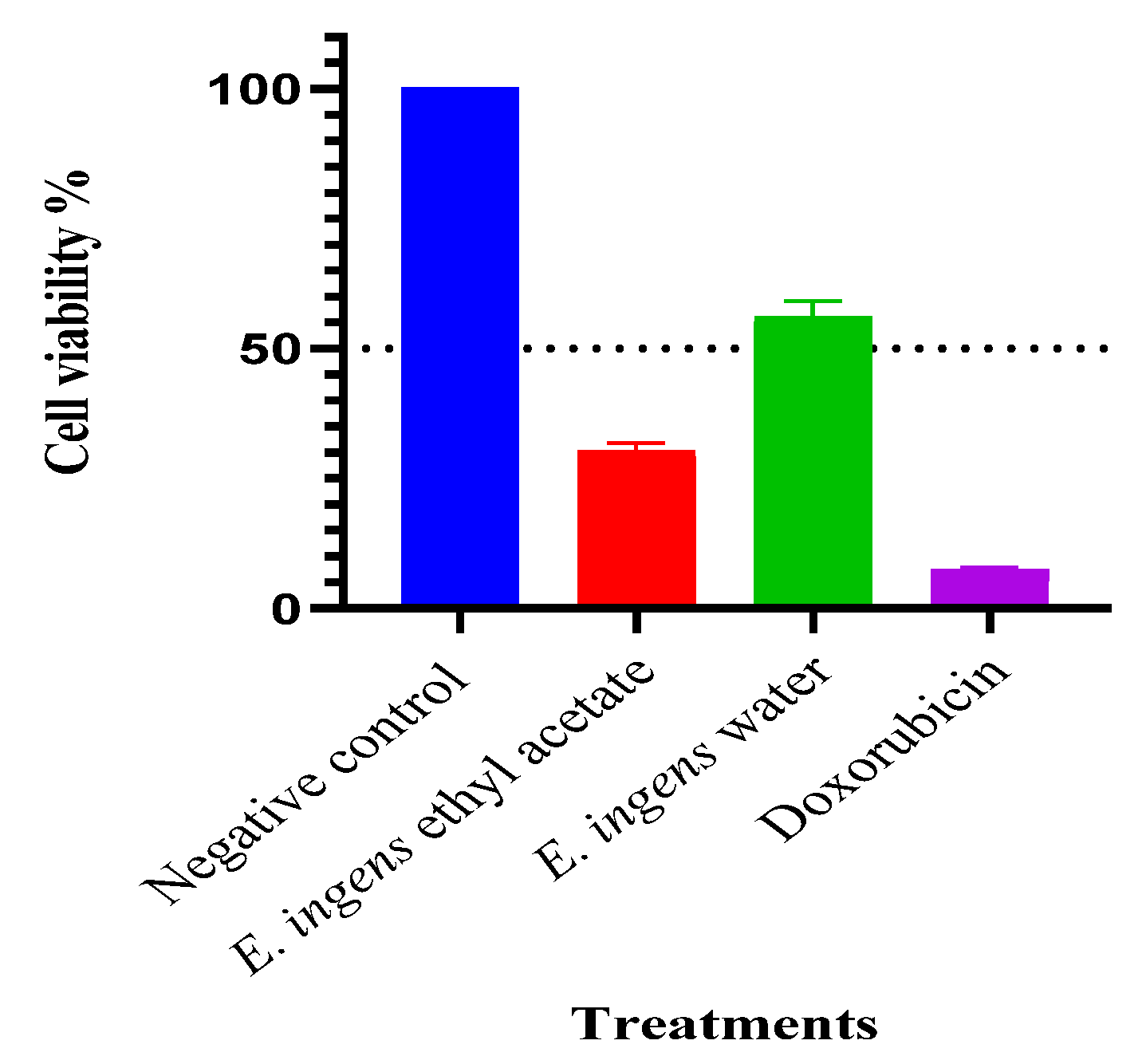
Figure 2.
Inhibition of cellular proliferation by E. ingens ethyl acetate fraction: Inhibition of cellular proliferation following 48 hours treatment with serial concentrations of E. ingens ethyl acetate fraction on DU-145 so as to determine IC50. 0.2% DMSO acted as negative control. Values are expressed as Mean ± SEM. All treatments were done in triplicates (n = 3).
Figure 2.
Inhibition of cellular proliferation by E. ingens ethyl acetate fraction: Inhibition of cellular proliferation following 48 hours treatment with serial concentrations of E. ingens ethyl acetate fraction on DU-145 so as to determine IC50. 0.2% DMSO acted as negative control. Values are expressed as Mean ± SEM. All treatments were done in triplicates (n = 3).
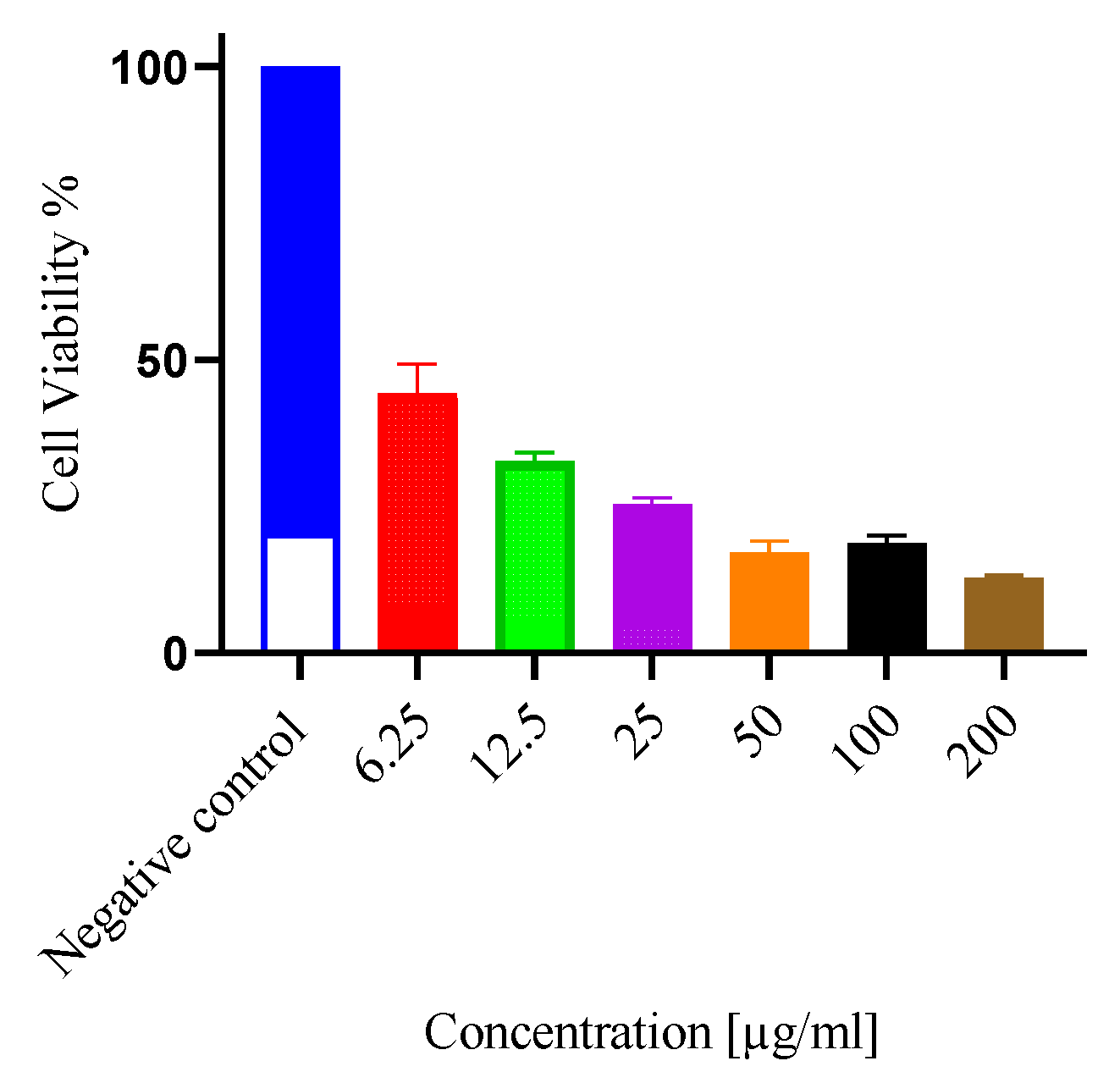
Figure 3.
Cellular safety: The potential cellular safety of E. ingens ethyl acetate was measured using noncancerous Vero E6. Cells were treated with serial concentrations of the E. ingens extract fraction for 48 hours. Values were expressed as Mean ± SEM, all treatments were done in triplicates (n = 3).
Figure 3.
Cellular safety: The potential cellular safety of E. ingens ethyl acetate was measured using noncancerous Vero E6. Cells were treated with serial concentrations of the E. ingens extract fraction for 48 hours. Values were expressed as Mean ± SEM, all treatments were done in triplicates (n = 3).
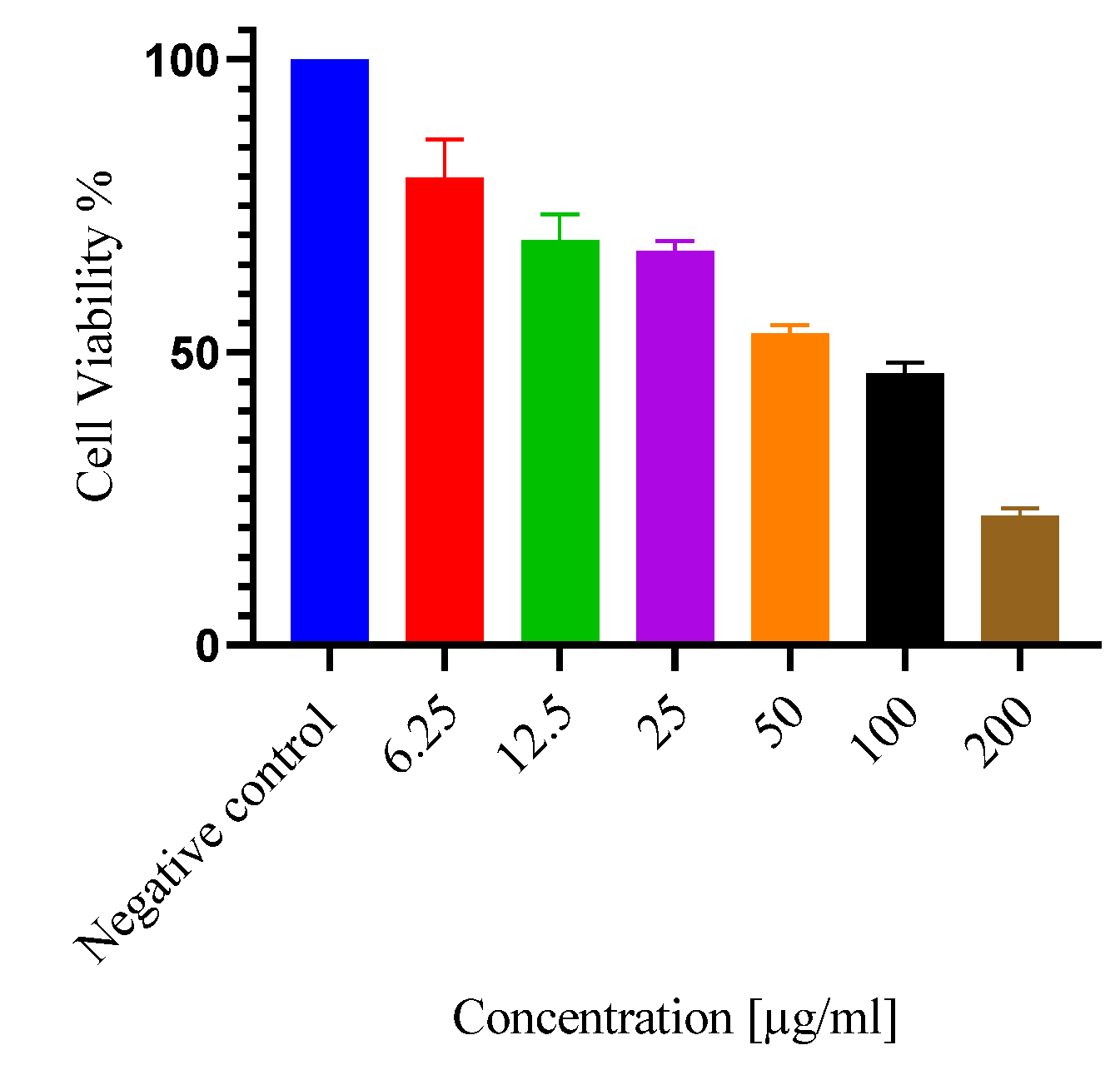
Figure 4.
GC-MS chromatogram of the chemical constituents present in E. ingens ethyl acetate fraction.
Figure 4.
GC-MS chromatogram of the chemical constituents present in E. ingens ethyl acetate fraction.
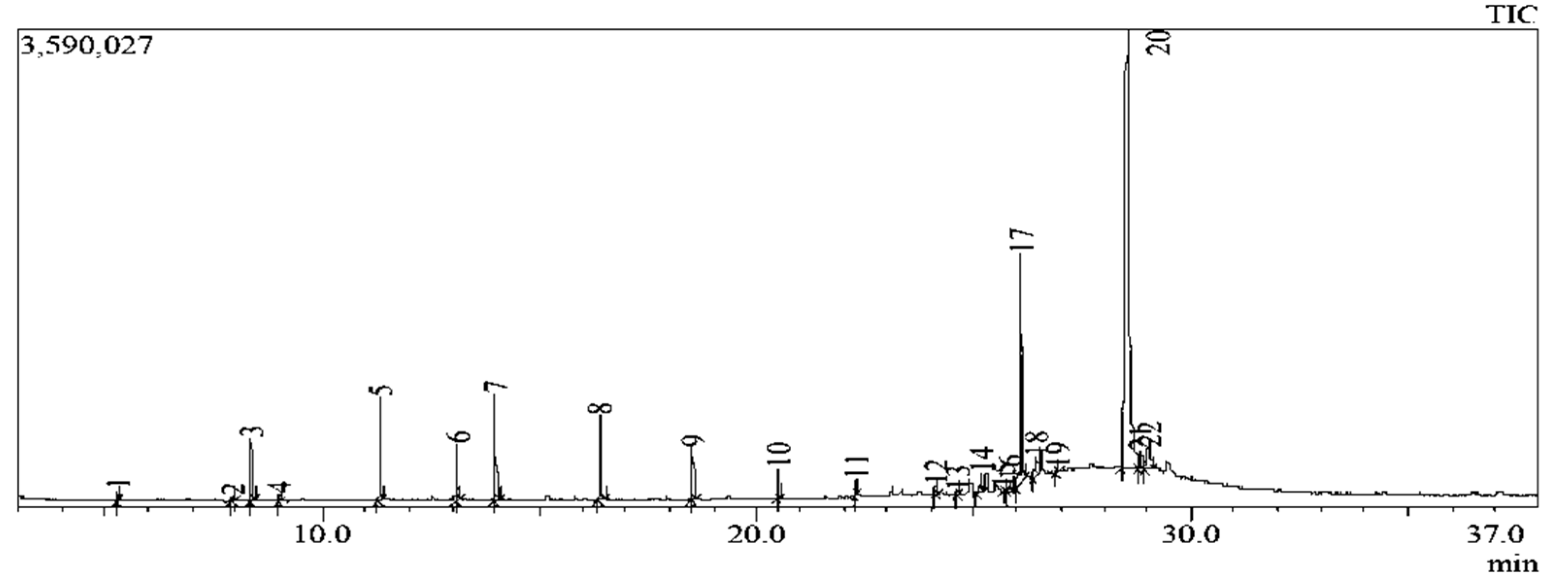
Figure 5.
Compound-disease targets Venny intersection diagram, with the compound targets on the left, the disease (PCa) targets on the right, and the two intersection targets in the middle.
Figure 5.
Compound-disease targets Venny intersection diagram, with the compound targets on the left, the disease (PCa) targets on the right, and the two intersection targets in the middle.
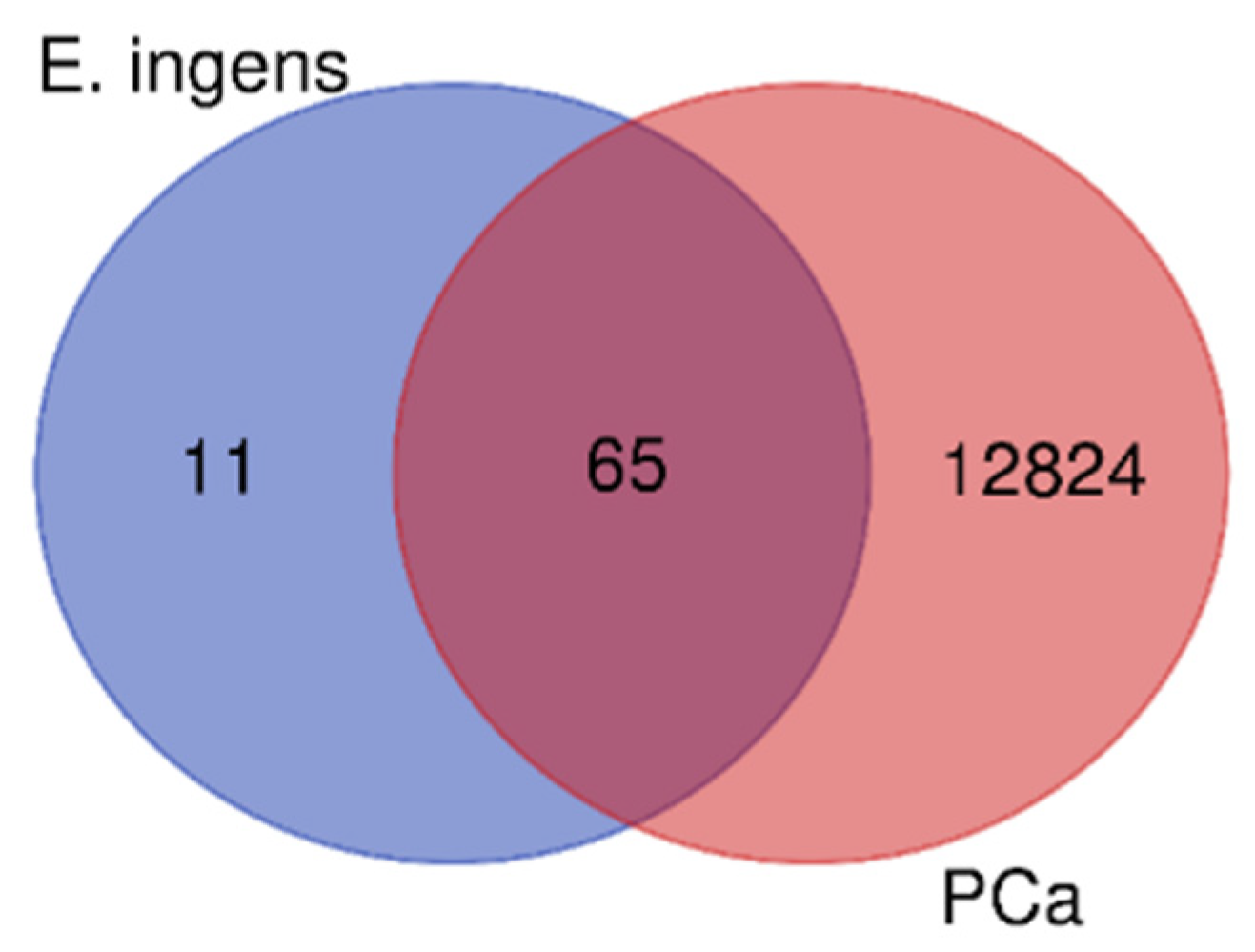
Figure 6.
PPI network of the key targets in E. ingens’ action against human prostate cancer.
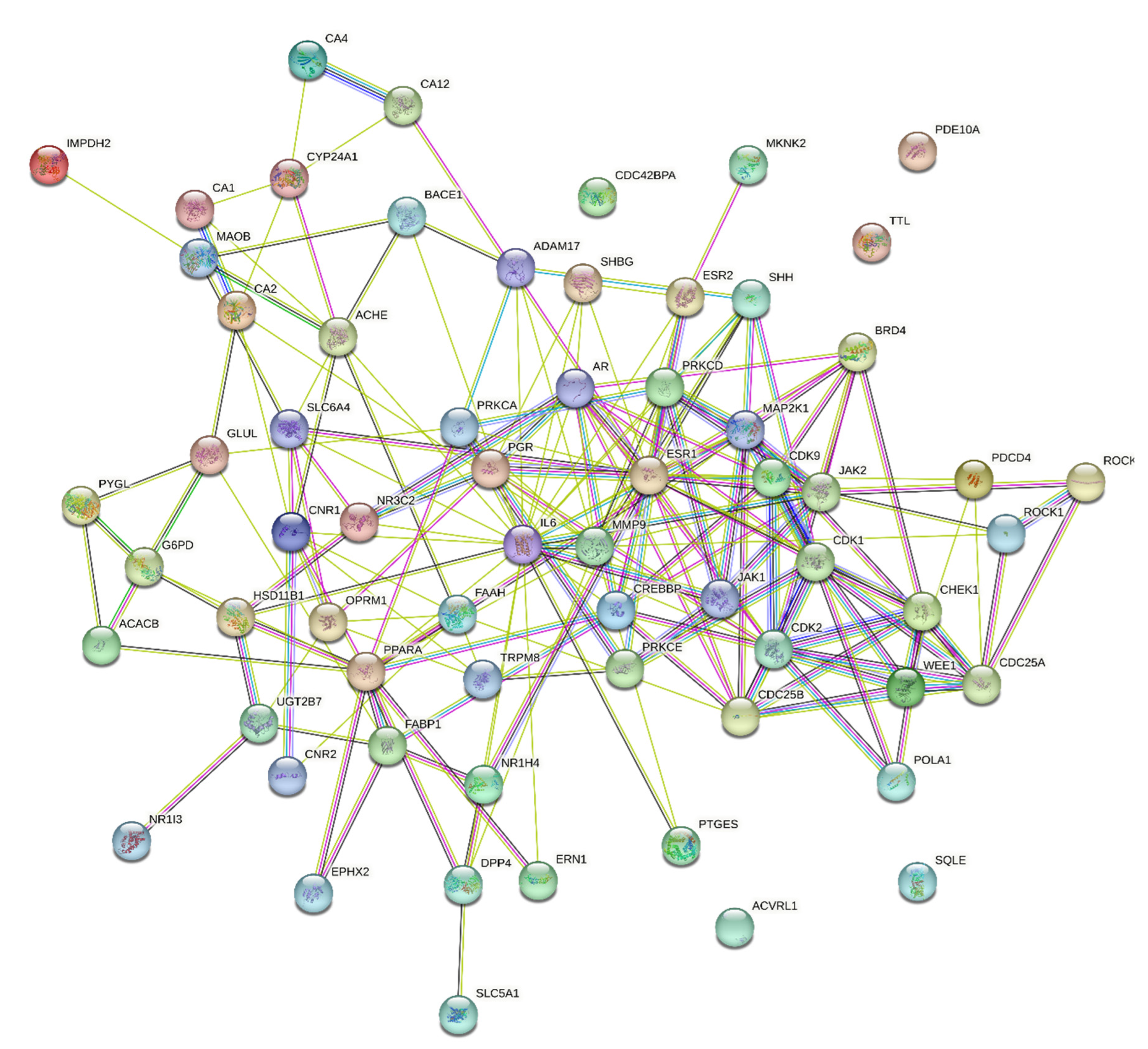
Figure 7.
Obtained ten hub genes. The larger the node is, the more important the target in the network.
Figure 7.
Obtained ten hub genes. The larger the node is, the more important the target in the network.
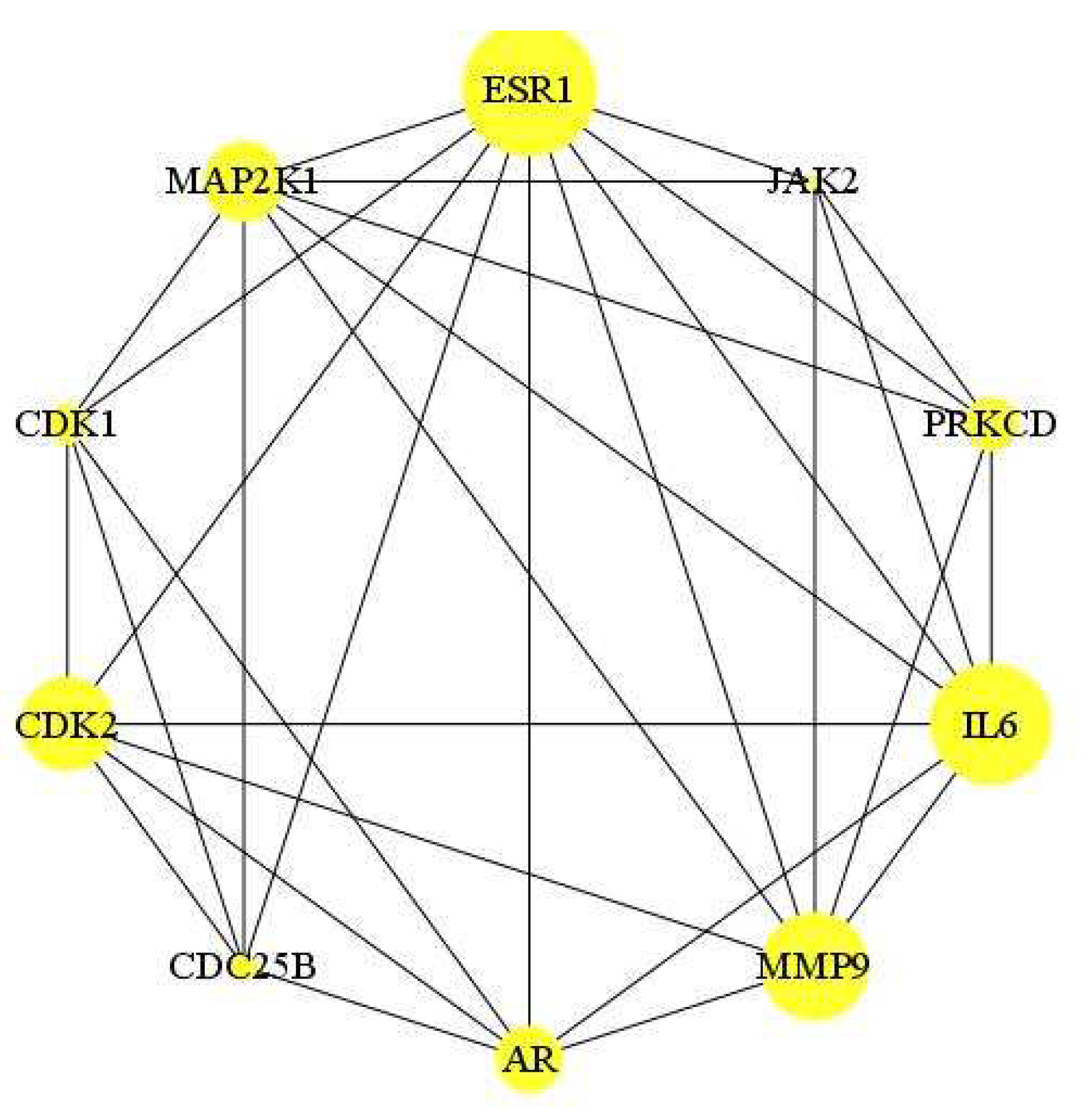
Figure 8.
Euphorbia ingens GO enrichment analysis (A) BP terms (B) CC terms (C) MF terms.
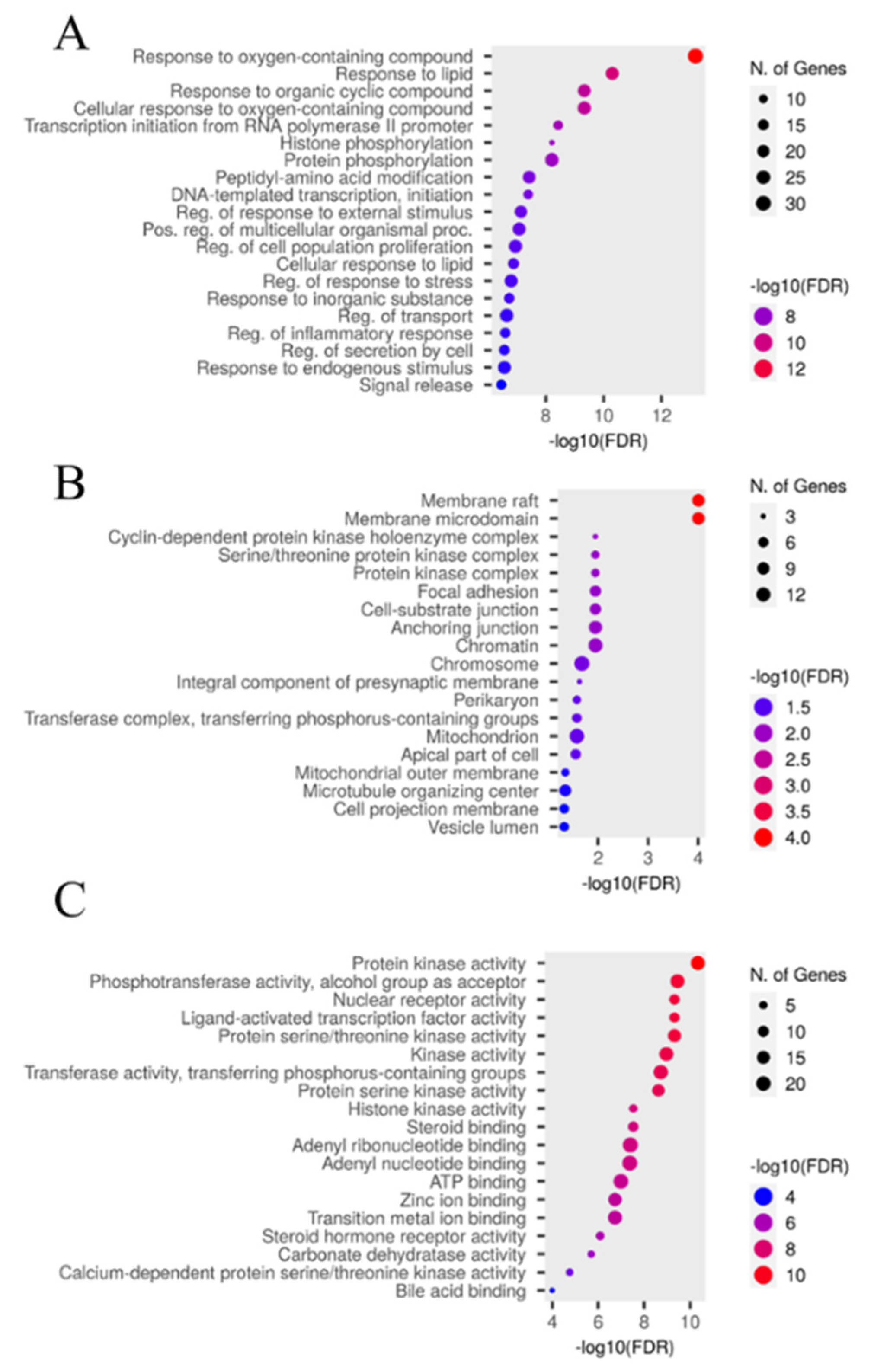
Figure 9.
KEGG signaling pathway analysis and prostate cancer pathway of E. ingens-PCa. (A) KEGG signaling pathway analysis. Each bubble represents a KEGG pathway on the vertical axis. The - logP values are shown on the horizontal axis. The size of each bubble indicates the number of genes enriched in each KEGG pathway. Larger bubbles indicate more genes involved in the pathway. The colour of each bubble represents the adjusted P-value of each KEGG pathway, with redder color indicating smaller adjusted P-value (B) Prostate cancer pathway (red marks represent potential targets for E. ingens intervention).
Figure 9.
KEGG signaling pathway analysis and prostate cancer pathway of E. ingens-PCa. (A) KEGG signaling pathway analysis. Each bubble represents a KEGG pathway on the vertical axis. The - logP values are shown on the horizontal axis. The size of each bubble indicates the number of genes enriched in each KEGG pathway. Larger bubbles indicate more genes involved in the pathway. The colour of each bubble represents the adjusted P-value of each KEGG pathway, with redder color indicating smaller adjusted P-value (B) Prostate cancer pathway (red marks represent potential targets for E. ingens intervention).
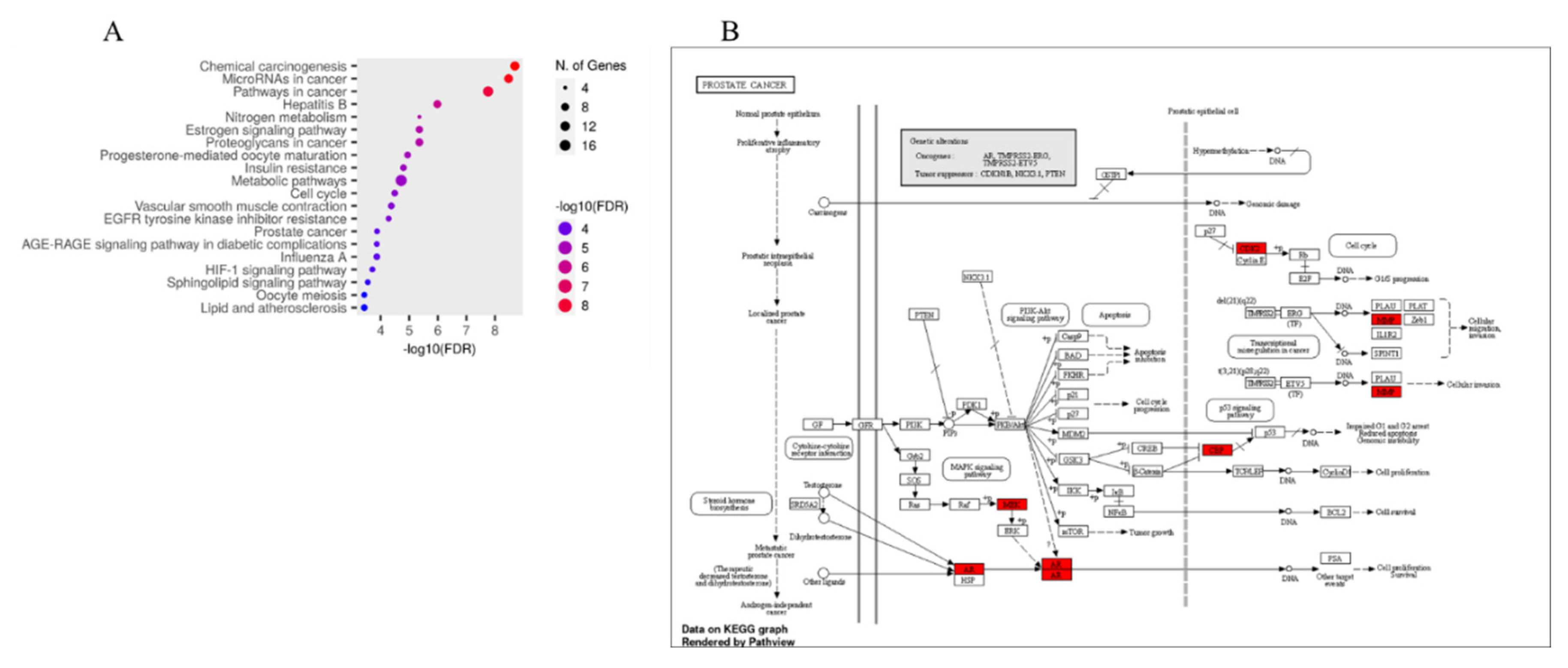
Figure 10.
Relative gene expression analysis of E. ingens treated and untreated DU-145 cells (A) AR, (B) BCL-2, (C) CDK1, (D) caspase-3 and (E) p53. ns p ˃ 0.05, * p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001, **** p ≤ 0.0001 as compared to untreated control.
Figure 10.
Relative gene expression analysis of E. ingens treated and untreated DU-145 cells (A) AR, (B) BCL-2, (C) CDK1, (D) caspase-3 and (E) p53. ns p ˃ 0.05, * p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001, **** p ≤ 0.0001 as compared to untreated control.
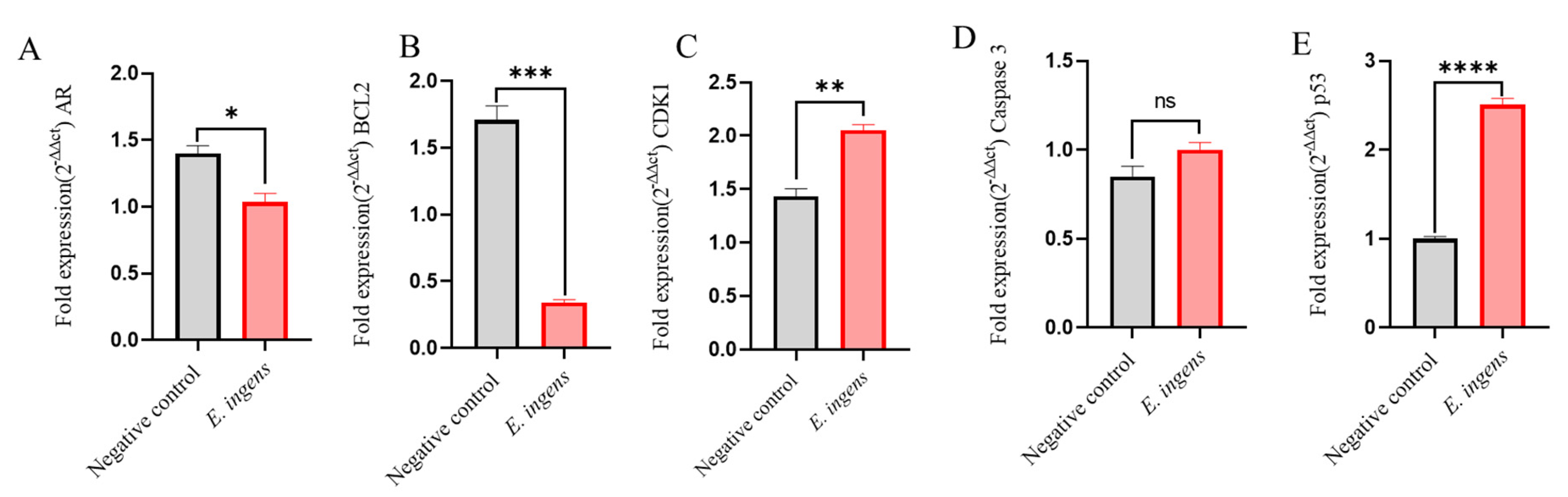

Table 1.
Summary of IC50, CC50 and selectivity index values.
| Extract | IC50 (µg/ml) | CC50 (µg/ml) | SI |
|---|---|---|---|
| E. ingens ethyl acetate | 9.71 ± 0.40a | 80.19 ± 6.12a | 8.26 |
| Doxorubicin | 5.30 ± 0.11b | 176.10 ± 8.09b | 33.23 |
Values that bear another superscript different from that of doxorubicin in a column differ significantly (p<0.05) from doxorubicin. Values are expressed as Mean ± SEM.
Table 2.
Phytochemical profile of E. ingens ethyl acetate fraction.
| Phytoconstituent | E. ingens ethyl acetate |
|---|---|
| Alkaloids | - |
| Phenols | ++ |
| Tannins | +++ |
| Terpenoids | +++ |
| Flavonoids | +++ |
| Saponins | +++ |
| Quinones | - |
| Sterols | +++ |
+++= highly abundant, ++= moderately abundant, += less abundant, - = absent.
Table 3.
The GC-MS identified compounds from E. ingens ethyl acetate fraction.
| Peak$Nr. | Rt (min) | Compound identified | Peak$Area % | MW$(g/mol) | MF | Structure Type |
|---|---|---|---|---|---|---|
| 5.302 | 1-decene | 0.29 | 140 | C10H20 | Alkene | |
| 7.952 | Bicyclo[3.1.1]heptan-3-ol | 0.23 | 152 | C10H16O | Terpenoid | |
| 8.381 | 1-dodecene | 2.11 | 168 | C12H24 | Alkene | |
| 9.034 | Bicyclo[3.1.1]hept-3-en-2-one | 0.25 | 150 | C10H14O | Terpenoid | |
| 11.339 | 1-tridecene | 3.64 | 182 | C13H26 | Alkene | |
| 13.123 | 2,4-di-tert-butylphenol | 2.03 | 206 | C14H22O | Phenol | |
| 14.003 | 1-octadecene | 4.05 | 252 | C18H36 | Alkene | |
| 14.003 | 1-octadecene | 4.05 | 252 | C18H36 | Alkene | |
| 18.543 | 1-heneicosanol | 1.86 | 312 | C21H44O | Fatty alcohol | |
| 18.543 | 1-heneicosanol | 1.86 | 312 | C21H44O | Fatty alcohol | |
| 18.543 | 1-heneicosanol | 1.86 | 312 | C21H44O | Fatty alcohol | |
| 24.128 | Octadecyl trifluoroacetate | 0.37 | 366 | C20H37F3O2 | Fatty Acid | |
| 24.639 | Prasterone | 0.14 | 288 | C19H28O2 | Sterol | |
| 25.177 | Andrographolide | 1.22 | 350 | C20H30O5 | Diterpenoid | |
| 25.695 | Ferruginol | 0.13 | 286 | C20H30O | Diterpenoid | |
| 25.834 | (1R,7S,E)-7-isopropyl-4,10-dimethylenecyclodec-5-enol | 0.63 | 220 | C15H24O | Sesquiterpenoid | |
| 26.079 | 2-bornanol | 16.75 | 348 | C16H20N4O5 | Terpenoid | |
| 26.435 | 11-oxoandrosterone | 1.53 | 376 | C22H36O3Si | Sterol | |
| 26.933 | Squalene | 0.20 | 410 | C30H50 | Triterpenoid | |
| 28.535 | 6-pentylidene-4,5-secoandrostane-4,17.beta.-diol | 55.39 | 362 | C24H42O2 | Sterol | |
| 28.827 | 17.beta.-hydroxy-6.alpha.-pentyl-4-nor-3,5-secoandrostan-3-oic acid, methyl ester | 1.78 | 378 | C24H42O3 | Fatty acid methyl ester | |
| 28.827 | 17.beta.-hydroxy-6.alpha.-pentyl-4-nor-3,5-secoandrostan-3-oic acid, methyl ester | 1.78 | 378 | C24H42O3 |
Key: Rt, retention time; MW, Molecular Weight; MF, Molecular Formular.
Table 4.
E. ingens ethyl acetate compounds with ideal drug candidate qualities.
| S/N | Compounds | MW $(g/mol) | HD | HA | Mol$LogP | Lipinski’s $Rule $(Violation) | BBB | CYP2D6 | CYP3A4 | TPSA |
|---|---|---|---|---|---|---|---|---|---|---|
| 1-dodecene | 168.32 | 0 | 0 | 5.25 | Yes; 1 | No | No | No | 0 | |
| 1-heneicosanol | 312.6 | 1 | 1 | 5.62 | Yes; 1 | No | No | No | 20.23 | |
| 1-octadecene | 252.5 | 0 | 0 | 6.77 | Yes; 1 | No | No | No | 0 | |
| 1-tridecene | 182.35 | 0 | 0 | 5.52 | Yes; 1 | No | No | No | 0 | |
| Octadecyl trifluoroacetate | 366.5 | 0 | 5 | 5.47 | Yes; 1 | No | No | No | 26.30 | |
| Andrographolide | 350.4 | 3 | 5 | 1.98 | Yes; 0 | No | No | No | 86.99 | |
| Squalene | 410.7 | 0 | 0 | 3.59 | Yes; 0 | No | No | No | 0 |
Keys: MW, molecular weight; HD, number of hydrogen bond donors; HA, number of hydrogen bond acceptors; MolLogP lipophilicity; BBB, blood brain barrier; CYP2D6, Cytochrome P450 2D6; CYP3A4 Cytochrome P450 3A4 and TPSA, total polar surface area.
Table 5.
Primers sequences.
| Genes | Primers |
|---|---|
| AR | Forward- GCTTTATCAGGGAGAACAGCCT$Reverse- TGCAGCTCTCTCGCAATCTG |
| BCL2 | Forward- GGCCTCAGGGAACAGAATGAT$Reverse- TCCTGTTGCTTTCGTTTCTTTC |
| CDK1 | Forward- GAACACCACTTGTCCCTCTAAGAT$Reverse- CTGCTTAGTTCAGAGAAAAGTGC |
| Caspase-3 | Forward- CAAAGAGGAAGCACCAGAACCC$Reverse- GGACTTGGGAAGCATAAGCGA |
| P53 | Forward- CTTCGAGATGTTCCGAGAGC$Reverse- GACCATGAAGGCAGGATGAG |
| β –Actin | Forward- GCCAACTTGTCCTTACCCAGA$Reverse- AGGAACAGAGACCTGACCCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



