
Submitted:
25 May 2023
Posted:
26 May 2023
You are already at the latest version
Abstract
Lead (Pb) is a non-essential, highly toxic, and persistent element widely recognised as one of the most concerning pollutants, and it is listed in the Priority List of Hazardous Substances. Widespread environmental contamination from Pb is a serious issue for human health and wildlife. The liver, a key compartment for metal detoxification and excretion, is also the organ in which Pb mainly accumulates in fish. Herein, we investigated for the first time the morphological and functional injuries induced in zebrafish (Danio rerio) liver by two very low and environmentally relevant concentrations of Pb (2.5 and 5 μg/L) after 48, 96, and 192 hours of exposure. We showed significant histological alterations in all the exposed samples, also demonstrating that the extent of injuries increased with dose and exposure time. The most common modifications observed were congestion of blood vessels and sinusoids, cytoplasmic vacuolizations, parenchyma dyschromia, and macrophage proliferation. Pb administration also resulted in a significant increase in lipid content and the upregulation of key genes involved in metal detoxification (mtf1) and the defensive response against oxidative stress (sod1 and cat). We showed that even very low doses of Pb can disrupt liver morphology and function.
Keywords:
lead
; liver
; molecular biomarkers
; morphological biomarkers
; MTs
; SOD
; CAT
1. Introduction
Heavy metals are a major concern for terrestrial and aquatic ecosystems because of their negative effects even in very low concentrations, their nondegradable nature, their high capacity for bioaccumulating, and their long-term persistence making them one of the main environmental problems of the 21st century worldwide [1,2,3]. Some heavy metals have been classified in terms of biological function as beneficial or essential for living organisms based on their biological functions while others, such as lead (Pb), cadmium (Cd), mercury (Hg), and arsenic (As), are considered to be non-threshold micropollutants able to induce toxic effects in living organisms [4,5,6,7].
Pb is a highly toxic and non-biodegradable heavy metal listed in the Priority List of Hazardous Substances released by the Agency for Toxic Substances and Disease Registry (ATSDR) [8,9]. Although Pb is naturally occurring in the environment, its unregulated use in a number of human activities (e.g., mining and agriculture activities, paint pigments production) has resulted in an increase in its levels in all environmental comparts, raising great concern for both human and wildlife [10]. Lead pollution of the aquatic environment occurs through agricultural, domestic, and industrial wastewater discharges, exerting a wide range of toxic effects on aquatic biota, including fish [11]. Fish play a prominent role in the functioning and balance of aquatic ecosystems [12] and are particularly sensitive to environmental pollutants [13,14,15], thus representing good indicators of environmental water quality.
Fish assimilate Pb by direct ingestion (i.e., food and water), ion exchange across lipophilic membranes (e.g., the gills), or adsorption across specific tissue and membrane surfaces [3]. In fish, numerous detrimental effects induced by Pb have been reported, among which are genotoxicity [16], oxidative stress induction [17,18,19,20] change in the activities of immune-related enzymes and genes [8,21,22] and histological changes of some tissues and organs [16,23,24,25].
Given its role in metal detoxification and excretion, the liver is a target organ for Pb [21,26]. In freshwater species, waterborne Pb mainly enters through the gills reaching the liver via the circulatory system [26]. Therefore, the liver has been claimed as the main target for Pb accumulation in fish [23,27,28], and the noxious effect induced in this complex organ have been investigated in several species. A systematic review of the literature provides plenty of evidence of adverse effects induced in the liver, including a modification of liver enzyme activity [29], the occurrence of metabolic disorders [30], alteration of liver morphology [31,32,33,34]. However, all these studies refer to high Pb concentrations, and there is a lack of knowledge on the effects of Pb at environmentally relevant concentrations [35,36].
It must be emphasized that fish exposed to very low concentrations of Pb may not show obvious signs of pathology, but the subtle morphological and functional alterations induced by the metal can reduce the health of individuals with important and dramatic repercussions at the population level.
Since the 1980s, zebrafish (Danio rerio) has been used in a broad spectrum of research fields due to its small body size, short reproductive cycle, easy husbandry, and high homology to the human genome. All these studies provided a powerful basis for using zebrafish as a model organism for aquatic ecotoxicology [37,38]. It is surprising that a very limited number of researches investigated the hepatotoxic effects of Pb in zebrafish liver, and so far, only three studies are available focusing on the induction of metabolic disorder and oxidative stress [39,40] and morphological alterations following chronic exposure to high Pb concentrations (60 mg/L) [41].
To fill the knowledge gap on Pb hepatotoxicity in fish, here we evaluated, for the first time, the effect induced in zebrafish liver by two very low and environmentally relevant concentrations of Pb (2.5 and 5 μg/L) after 48, 96, and 192 hours of exposure. The tested doses have been selected based on Pb concentrations found in aquatic environments worldwide and, particularly, in the concentrations range of Pb in surface waters; therefore, our results would also support the implementation of risk assessment protocols.
Given the general paucity of information about morphological and functional injuries induced by Pb in the fish liver, we first assessed the histological alterations, which are widely recognised as the best tool for assessing the effects of chemical contaminants, including heavy metals [42]. Moreover, to allow a more reliable and objective comparison among experimental groups, we applied a semi-quantitative method for evaluating the severity of histological changes.
Since histopathological lesions represent an integration of the effects of prior biochemical and physiological perturbations [43], in a second step, we analyzed the modulations of some genes involved in i) metal detoxification (metallothionein, mtf1) and ii) oxidative stress defense (i.e., superoxide dismutase-sod1 and catalase-cat) to better clarify the molecular mechanisms underlying Pb hepatotoxicity.
Metallothionein (MTs) are low-molecular-weight proteins responsible for metal binding, which are effective in non-essential metal detoxification and protection from oxidative stress [44]. The biosynthesis of MTs in fish is induced by a variety of metals, including Pb, providing an excellent biomarker of exposure to metals [45,46].
Pb toxicity in the liver could be mediated through different mechanisms, but the most common response in both fish and mammalian models is the imbalance between reactive oxygen species (ROS) production and the removal of such molecules [47]. The cell counteracts ROS overproduction by induction of antioxidant molecules, which may be either enzymatic (e.g., catalase and superoxide dismutase) or non-enzymatic (e.g., glutathione). In fish exposed to Pb, the antioxidant responses have been demonstrated in several species, thus supporting the role of such molecules as biomarkers of oxidative stress induced by heavy metals [21,48,49,50].
To the best of our knowledge, this is the first study documenting the morphological, morphometric, and functional alterations of low Pb concentrations in fish liver. Our results, providing new insights into lead-induced hepatotoxicity, will also contribute to a better understanding of the risk posed by heavy metals to wildlife species under a realistic exposure scenario.
2. Materials and Methods
2.1. Fish maintenance
A total of 70 individuals of both sexes (length, 3.5±0.5 cm, and weight, 0.43±0.06 g) were obtained from a local fish retailer. For two weeks, fish were acclimatized under controlled conditions in aquaria filled with dechlorinated tap water (temperature=26±0.5°C, pH=7.3, conductivity=300 μs/cm, dissolved oxygen=8±1 mg/L, hardness=100 mg/L CaCO3, 14:10 light regime). During the acclimatization period, half of the water was renewed daily, and fish were fed daily with commercial fish food.
2.2. Exposure Conditions
To obtain the two nominal concentrations of 2.5 µg/L (low concentration) and 5 µg/L (high concentration), a stock solution (1000 µg/L) of lead acetate was prepared in distilled water [Pb(CH3CO2)2 3H2O, Sigma-Aldrich Chemical Co., St. Louis, MO, USA]; then an adequate amount was diluted in dechlorinated water.
To determine Pb in water samples, an Elan DRC-e inductively Coupled Plasma-Mass Spectrometry (ICP-MS) (PerkinElmer SCIEX, Woodbridge, ON, Canada) was used. Samples were diluted in ultrapure nitric acid (500 µL) and then introduced into the instrument system using a PerkinElmer AS-93 plus autosampler and a cross-flow nebulizer with a Scott-type spray chamber. Quantitative analysis was performed by constructing the calibration curve for the lead on five different Plasma-Mass (calibration range of 0.1-50 g/L). The analytical verification of the actual concentrations was performed throughout the experiment (starting from time 0 every 24 hours) (Table S1). No evident variation was recorded in agreement with previous literature data [25 and references therein].
The two selected concentrations corresponded to 0.00146% and 0.00292% of the median lethal concentration at 96 hours (LC5096h), respectively for adult zebrafish [39]. Moreover, both doses were selected considering pre-existing data on the worldwide concentration of Pb in surface water and can be considered very low and environmentally realistic [3,35].
Fish were exposed to the low or the high Pb dose for 48, 96, and 192 hours, resulting in 6 experimental groups (n=10). The control group (n=10) was maintained in aquaria filled with dechlorinated tap water.
During the experiment, temperature, pH, conductivity, dissolved oxygen, hardness, and photoperiod were monitored daily and kept constant, as described for the acclimatization period. Fish were fed on alternate days, and food waste and debris were removed daily using a fine mesh.
After 48, 96, and 192 hours, fish were immersed in an anesthetic water bath containing ethyl 3-aminobenzoate methanesulfonate (20 mg/L MS 222, Sandoz, Sigma-Aldrich, St. Louis, MO, USA), and the liver was rapidly dissected and processed for subsequent analyses as reported below. For each experimental unit, including the control, two replicates were conducted. The use of animals in this study was approved by the Institutional Animal Care and Use Committee at the National University of Entre Rios and the Italian University Institute of Rosario (Rosario, Argentina; protocol N°028/12).
2.3. Histology and Histopathological assessment
Excised liver samples (n=4) were immediately fixed in 4% glutaraldehyde (Electron Microscopy Sciences, Hatfield, PA, USA) in phosphate-buffered saline solution (PBS 0.1 M, pH 7.2, 4°C) and post-fixed in osmium tetroxide (1% in PBS) for 2 hours. Samples were dehydrated through a graded ethanol series, placed in propylene oxide, and embedded in Epon-Araldite (Araldite 502/Embed 812, Electron Microscopy Sciences). Longitudinal serial semithin sections of 1 μm, obtained using a Leica UltraCut UCT (Leica Microsystems, Wetzlar, Germany), were mounted on glass slides, stained with toluidine blue, and observed under an LM Leitz Dialux 20 E.B. (Leica Microsystems, Wetzlar, Germany) equipped with a digital camera.
The prevalence of each histological alteration was obtained by doing the ratio between the number of fish affected by a specific alteration and the total number of fish. We also determined the histological changes' severity using a semi-quantitative method according to previous literature data [51,52]. Briefly, the alterations were attributed to a specific reaction pattern (circulatory disturbances, regressive changes, progressive changes, and inflammation). Then an importance factor was assigned to each observed alteration following the relevance of the change and its pathological importance (from 1, minimal pathological importance, to 3, marked pathological importance). A score value was then assigned based on the degree and extent of each lesion, as follows: 0 (unchanged), 2 (mild occurrence), 4 (moderate occurrence), and 6 (severe occurrence). (Table S2). The organ index (Iorg) representing the degree of organ damage was calculated using the importance factor and the score value according to the following formula:
where: org=organ, rp=reaction pattern, alt=alteration; a=score value; w=importance factor.
Iorg = Σrp Σalt(aorg rp alt× worg rp alt)
2.4. Lipid droplets content
Lipid droplets analysis has been performed on semithin sections (toluidine blue-stained). Four liver sections were photographed (100×) for each animal of both the control and Pb exposed groups (n=4) and evaluated for the percentage of area occupied by lipid droplets. The lipid granules were isolated in each micrograph using the free and open-source ImageJ software (NIH, developed at the National Institutes of Health, a part of the U.S. Department of Health and Human Services), and the total area occupied by the granules was quantified.
The results, expressed as the percentage of area occupied by the lipid granules in each section, were statistically compared using the two-way ANOVA followed by Tukey's multiple comparisons tests (at a significance level of 0.05). Data were checked for normality (Shapiro-Wilk test) and presented as mean±standard deviation.
2.5. Quantitative Real-Time PCR
Excised liver samples of animals of both treated and control groups (n=6) were promptly stored at -80°C for subsequent real-time PCR analyses. Total RNA was extracted using the PureLink RNA Mini Kit and the PureLink™ DNase Set (Thermo Fisher Scientific, Waltham, MA, USA) following the manufacturer's protocol. The quantity and quality of RNA were verified using a NanoDrop One spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) and 1.5% agarose gel electrophoresis, respectively. 2 μg of total RNA was utilized for first-strand cDNA synthesis using the high capacity RNA to cDNA kit (Applied Biosystems, Foster City, CA, USA); the resulting cDNA was kept at -20°C. cDNA was used as a template for quantitative reverse transcription-polymerase chain reaction (RT-qPCR) analysis to quantify the expression of the metal-regulatory transcription factor 1 (mtf1, NCBI Reference Sequence NM_152981.1), the superoxide dismutase 1 (sod1, NCBI Reference Sequence NM_131294.1) and the catalase (cat, NCBI Reference Sequence NM_130912.2). RT-qPCR was performed in triplicate in a Light Cycler (Applied Biosystems StepOne, Real-Time PCR System, Foster City, CA, USA) using the TaqMan Gene Expression Assays (Thermo Fisher Scientific, Waltham, MA, USA). Each reaction contained 2 μL of cDNA, 10 μL of master mix (TaqMan Universal Master Mix II, Applied Biosystems), 1 μL of assay mix (TaqMan Gene Expression Assay), and 7 μL of RNase- and DNase-free water and was run according to the manufacturer's instructions: one cycle at 50°C for 2 minutes, 95°C for 10 minutes, 40 cycles at 95°C for 15 seconds, and 60°C for 1 minute.
The glyceraldehyde-3-phosphate dehydrogenase (gapdh, NCBI Reference Sequence: NM_001115114.1) and the actin beta 1 (actb1, NCBI Reference Sequence: NM_131031.2) were used as internal reference genes. The relative copy number of each analyzed gene was calculated according to the 2−ΔΔCT comparative CT method.
3. Results
No mortality occurred during the whole experimental period, neither in the control nor in exposed groups.
3.1. Control group
The morphology of the Danio rerio liver is similar to that of other freshwater Teleosts [53,54]; only a brief description will be furnished in the present study. The parenchymal mass, even and compact, was crossed by a network of sinusoids surrounded by cords of hepatocytes (Figure 1a,b).
The space of Disse is recognizable between the sinusoidal endothelium and the hepatocytes (Figure 1b). Veins bordered by a continuous endothelium are scattered within the parenchyma; erythrocytes and a few macrophages can be detected in their lumen (Figure 1b). The bile ducts, lined by cuboidal epithelium, can be seen in the parenchyma (Figure 1a). Hepatocytes exhibit a polygonal shape with central spherical nuclei, numerous glycogen granules, and a few lipid droplets in their cytoplasm (Figure 1b).
3.2. Exposed group
3.2.1. Low Pb concentration
After 48 hours of exposure to the low Pb concentration, the overall morphological organization of the liver parenchyma was maintained (Class I normal organ structure, Table 1) (Figure 2a). However, the frequency of bile duct degeneration and the congestion of blood vessels and sinusoids significantly increased compared to the control (Figure 2b,c) (Figure 3a,b). An increase of lipid droplets in the hepatocyte cytoplasm was also recognizable (Figure 2c).
The extent and intensity of histological alterations increased after 96 hours of exposure, becoming significantly higher compared to both control and 48 hours-exposed groups (Table 1), and their severity degree ranged from normal to slightly altered (Class II). The typical architecture of the bile ducts was no longer recognizable, and cuboidal cells often showed signs of degeneration (Figure 2d). Moreover, detachment of the duct epithelium from connective tissue was frequently observed (Figure 2d) (Figure 3c). The lumen of some vessels and sinusoids was filled by a large number of blood cells and proliferating macrophages, which sporadically also migrated to the liver parenchyma (Figure 2e,f). Commonly observed hepatocyte abnormalities included cytoplasmic vacuolization, increased lipid droplets, and the emergence of the distinctive signs of apoptosis, such as deeply stained cytoplasm and degenerated nuclei (Figure 2e,f) (Figure 3d-g). It was also observed that the occurrence of lysed areas was significantly higher than in control and the 48 hours exposed groups (Figure 2g) (Figure 3j).
After 192 hours of exposure, the architecture of the liver parenchyma was markedly compromised. The degrees of histopathological changes significantly increased compared to the control and 96 hours-exposed groups (Class IV pronounced histological alterations, Table 1). Hepatocyte cytoplasmic vacuolization, enhancement of lipid droplets, the appearance of extensive lysed areas, and congestion of vessels and sinusoids were frequently detected. Still, their frequency did not differ from the 96 hours exposed group (Figure 3). In contrast, detachment of the bile duct epithelium dramatically increased and macrophage proliferation, bile duct degeneration, pale necrotic hepatocytes, and dark-colored apoptotic hepatocytes were observed in all samples (Figure 2h-j) (Figure 3a,c,f,g,k).
3.2.2. High Pb concentration
After 48 hours of exposure to the high Pb concentration, the extent and intensity of histological alterations significantly increased compared to the control group (Class II slight histological alterations, Table 1). All morphological alterations observed in low-Pb exposed groups precociously appeared (Figure 3a-k) (Figure 4a-c). Degeneration of the bile ducts, bile duct epithelial detachment, an increase of lipid droplets in hepatocytes cytoplasm, and the appearance of lysed areas were observed at a significantly higher frequency than in control (Figure 3a,c,e,j) (Figure 4a,c).
The incidence of cytoplasmic vacuolization, blood vessel congestion, and apoptotic and necrotic hepatocytes was significantly increased compared to the control and the low Pb concentration group (Figure 3b,d,f,g) (Figure 4a,b). Moreover, macrophage proliferation was detected in 100% of the samples (Figure 3k) (Figure 4b).
The tissue degeneration significantly increased as the exposure proceeded, and after 96 hours of exposure, the extent and intensity of histological alterations were higher compared to the control and low Pb concentration group (Class III severe histological modifications, Table 1). The frequency of cytoplasmic vacuolization, lysed areas, bile duct degeneration, and bile duct epithelial detachment was significantly higher compared to the control and 48 hours exposed group (Figure 3a,c,d,j) (Figure 4d,e). Numerous alterations, such as vessels and sinusoid congestion, macrophage proliferation, and apoptotic hepatocytes, were detected in 100% of samples (Figure 3b,f,k) (Figure 4f). Moreover, an increase in lipid droplet contents was evident (Figure 3e) (Figure 3f).
The liver structure markedly changed after 192 hours of exposure. The extent and intensity of histological alterations reached a peak and were statistically significant compared to the control and all treated groups (Class V severe histological alterations, Table 1). All the considered alterations were detected with an incidence of 100% (Figure 3a-k). Bile ducts displayed a severely modified architecture (Figure 4g,h). The lumen of vessels and sinusoids was filled with macrophages frequently migrated in the liver parenchyma (Figure 4i). Hepatic dyschromia was more evident due to the increase in the numbers of both necrotic and apoptotic hepatocytes (Figure 4i). Cytoplasmic vacuolization and growth of lipid droplets were prominent (Figure 4i).
3.3. Lipid droplets content
Lipid droplet amount, expressed as the percentage of the section area occupied by lipid droplets, showed a statistically significant difference in Pb-exposed groups compared to the control. The lipid droplet content significantly increased after exposure to both tested concentrations at all time points (Figure 5). More in detail, after 48 hours, the increase was significant and became highly significant as the exposure proceeded when also a significant difference could be noted between low- and high-exposed groups (Figure 5).
3.4. Gene expression
Metallothioneins (mtf1) – Exposure to the low Pb concentration induced a significant upregulation of mtf1 compared to the control, starting from 96 hours of exposure and peaking after 192 hours (Figure 6a). Highly significant upregulation was observed at 48 and 96 hours when the high Pb dose was administered. A significant difference was detected between low- and high-concentration groups at these time points. The expression level decreased after 192 hours, remaining significantly higher than the control (Figure 6a).
Catalase (cat) – After exposure to the low Pb concentration, a significant increase in cat expression was detected at all time points compared to the control, and the maximum level was reached after 192 hours (Figure 6b). A similar transcriptional response was detected after exposure to the high Pb concentration. The highest expression level was noted after 192 hours when the upregulation was significantly higher also compared to the low-Pb exposed groups (Figure 6b).
Superoxide dismutase (sod1) – Exposure to the low Pb concentration induced a significant upregulation of sod1 at all time points compared to the control, peaking after 192 hours. A similar pattern was detected when the high Pb dose was administered (Figure 6c).
4. Discussion
Extensive literature data demonstrate that heavy metals must be recognized as priority pollutants due to their pervasive and persistent distribution in all environmental compartments. Pb is highly toxic and non-biodegradable, and it is widely acknowledged as one of the most dangerous heavy metals for living organisms [8,9,55].
Information on the effects of naturally occurring Pb concentrations is mandatory for environmental and public safety in order to effectively assess the presumed harmful outcomes. However, most of the information about Pb toxicity in fish comes from studies on high Pb concentrations which are not representative of a naturally occurring event of contamination whereas the number of studies explicitly assessing the impact of low Pb concentrations on fish species is extremely low.
Since the liver is responsible for many vital functions, including the accumulation and detoxification of pollutants [56], pathological alterations of this organ may easily interfere with the functioning of all physiological processes. Moreover, the liver of teleost fish is an organ widely used as a biomarker for fish health assessment because the effects of pollutant exposure can be evident at its cellular and tissue level [57,58].
4.1. Morphological Modifications
In the present study, we demonstrated for the first time that exposure to two very low Pb concentrations induces severe histopathological and functional changes in zebrafish liver. As revealed by our semi-quantitative analyses, the severity of injuries was time and dose-dependent. Indeed, an evident pathological progression could be seen as the experiment proceeded in both experimental groups, but alterations precociously arose in the group exposed to the high Pb dose, also showing a higher severity at all exposure times.
According to previous reports on Oreochromis niloticus chronically exposed to high Pb concentrations [33,59], the first and most frequent alteration observed in D. rerio liver was the congestion of blood vessels and sinusoids. Such circulatory alterations have been regarded as reversible modifications that do not alter the normal function of the tissue [57], leading us to suppose that a recovery of the health status would be possible if the input of the toxicant ceased. In contrast, inflammation, regressive, and progressive changes that appeared or worsened starting from 96 hours of exposure are non-reversible modifications resulting in the complete loss of the liver parenchyma arrangement.
In our experiment, we also frequently observed the appearance of cytoplasmic vacuolizations, as previously reported, under both laboratory and field conditions, in other freshwater species after exposure to Pb [60,61]. It has been suggested that hepatic vacuolations following exposure to heavy metals would be due to lipid and/or glycogen deposition, which are indicative of metabolic disorders [62]. Interestingly a significant increase in lipid content and cytoplasmic vacuolizations were synchronously detected in all D. rerio samples exposed to Pb, thus suggesting that the appearance of vacuole structures would be related to lipid depositions. Our findings are in general agreement with literature data on rare minnows (Gobiocypris rarus) following acute waterborne cadmium exposure [63].
Another commonly observed phenomenon induced in zebrafish liver was the emergence of lysed areas, in agreement with available reports on zebrafish liver after exposure to other heavy metals [54,64].
D. rerio also responded to Pb exposure by the proliferation of macrophages, which play a crucial role in regulating immune response, host protection, and tissue homeostasis [65]. Macrophage activation in fish is considered a bio-indicator of exposition to chemical contaminants, especially those associated with oxidative stress and lipid peroxidation [66,67].
Exposure to Pb affects ROS and reactive nitrogen species production through different mechanisms [21,68]. Whatever the prooxidant pathways, Pb triggers a cascade of oxidative reactions leading to protein unfolding, DNA/RNA damage, and peroxidation of unsaturated lipids in cell membranes.
Among other heavy metals, Pb especially encourages iron-initiated membrane lipid oxidation in fish [21], which is considered an essential mediator of ferroptosis, a lytic form of regulated cell death (LRCD) [69]. Although nonmammalian vertebrates, including fish, exhibit such an oxidation pathway, the extent to which ferroptosis per se is involved it is still poorly understood [70]. Metal-induced ferroptosis has been recently suggested in Japanese flounder exposed to nickel and cobalt [71], as demonstrated by the enhancement of ferroptosis-related pathways, simultaneous cell swelling, and cytoplasmic depletion in hepatocytes.
Different types of cell death may coexist in the Pb-induced pathological context, including lytic forms of hepatocellular death, such as ferroptosis, which share morphologic features with passive necrosis [72]. In our study, we clearly showed the concurrent presence of dark-stained hepatocytes, representing the apoptotic cell population, and pale and swollen hepatocytes which might belong to both necrotic or ferroptotic cells. Therefore, based on the histological results and the significant upregulation of antioxidant enzymes (see below), we speculated that ferroptosis may be a key regulator of Pb-induced liver injury.
4.2. Gene-Expression
Superoxide dismutase (SOD) and Catalase (CAT) - In fish, the first antioxidant response against elevated ROS levels is the modulation of key enzymes, including superoxide dismutase (SOD) and catalase (CAT) which are two major antioxidant enzymes and good indicators of oxidative stress [73,74].
Here we clearly showed a significant upregulation of such enzymes after exposure to both Pb concentrations at all time points. SOD and CAT modulation induced by Pb in fish liver has been investigated in several species with contradictory results, as both the upregulation and downregulation of these enzymes have been reported [17,21,41,75,76]. In zebrafish liver, Wang and colleagues [41] recently demonstrated that chronic exposure to high Pb concentration resulted in an initial increased activity of SOD and CAT (45 days), followed by a reduction when the experiment was prolonged (90 days). They suggested that the excess ROS may exhaust the antioxidant system giving the reason for the trend reversal in SOD and CAT activity. It must be emphasized that a direct comparison of our results with literature data is difficult since available information is limited to the effects induced by high Pb concentrations and/or deals with chronic exposure assays. More studies are needed to understand the physiological and molecular mechanisms underlying the modulation of antioxidant enzymes after Pb exposure to low and realistic environmental concentrations.
Metallothioneins (MTs) - MTs are recognized as sensitive biomarkers of heavy metal exposure in aquatic organisms. They play an important role in maintaining redox potentials, essential metals' homeostasis, and detoxifying non-essential metals [45]. Under the basal condition, MTs are expressed in the fish's liver, but their increase is strictly related to metal exposure [49].
An increase in MTs expression has been demonstrated in the liver of both marine and freshwater fish after dietary exposure to Pb and in fish coming from Pb-contaminated areas [49,77]. Furthermore, our recent study reported an increase in the expression of mtf1 in zebrafish gills after Pb exposure [25]. Although MTs induction has been demonstrated in several fish organs after exposure to heavy metals, the liver is one of the organs that first responds to the toxic input [77]. Accordingly, here we confirm the use of the liver as a sensitive organ to investigate early exposure to Pb and the importance of using MTs as valuable biomarkers. We demonstrated an upregulation of metallothionein starting from 96 hours of exposure to the low concentration of Pb, while the overexpression is precocious (from 48 hours) when the high dose of the contaminant is administrated.
5. Conclusions
Overall, data presented here clearly show that short-term exposure to two very low and naturally found concentrations of Pb is associated with significant histological alterations in the Danio rerio liver. Our semi-quantitative morphological evaluation shows that the severity and extent of injuries increase with dose and exposure time, resulting in irreversible histological changes at the end of exposure to the high tested concentration. We also demonstrated that Pb administration induces metabolic disorders, evident in the significant increase in lipid content in all exposed groups.
Our results confirm that the production of reactive oxygen species is an essential mechanism of Pb toxicity in the liver leading to an upregulation of the antioxidant enzymes (sod and cat). MTs upregulation is confirmed by results presented here as a powerful marker of lead contamination.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Table S1: Detected Pb concentrations in the control group and exposure solutions.; Table S2: Histopathological changes observed in Danio rerio liver.
Author Contributions
Methodology, R.M., V.C., E.P., L.O.; investigation, R.M., V.C., E.B.; formal analysis, A.I.M.A., F.T., L.O., M.M.; data curation, F.T., S.S.; software, A.I.M.A., F.T. S.S.; writing—original draft preparation, R.M., V.C., E.B.; writing—review and editing, E.B.; project administration, E.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Institutional Review Board Statement
The use of animals in this study was approved by the Institutional Animal Care and Use Committee at the National University of Entre Rios and the Italian University Institute of Rosario (Rosario, Argentina; protocol N. 028/12).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kortei, N.K.; Heymann, M.E.; Essuman, E.K.; Kpodo, F.M.; Akonor, P.T.; Lokpo, S.Y.; Boadi, N.O.; Ayim-Akonor, M.; Tettey, C. Health risk assessment and levels of toxic metals in fishes (Oreochromis noliticus and Clarias anguillaris) from Ankobrah and Pra basins: Impact of illegal mining activities on food safety. Toxicol. Rep. 2020, 7, 360–369. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.W.; Lee, D.Y.; Chen, P.J. Use of embedded Chelex chelating resin and sediment toxicity bioassays with medaka embryos to determine the bioavailability and toxicity of lead-contaminated sediment. Sci. Total Environ. 2020, 745, 140794. [Google Scholar] [CrossRef]
- Ezemonye, L.I.; Adebayo, P.O.; Enuneku, A.A.; Tongo, I.; Ogbomida, E. Potential health risk consequences of heavy metal concentrations in surface water, shrimp (Macrobrachium macrobrachion) and fish (Brycinus longipinnis) from Benin River, Nigeria. Toxicol. Rep. 2019, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bansod, B.; Kumar, T.; Thakur, R.; Rana, S.; Singh, I. A review on various electrochemical techniques for heavy metal ions detection with different sensing platforms. Biosens. Bioelectron. 2017, 94, 443–455. [Google Scholar] [CrossRef]
- Brunelli, E.; Mauceri, A.; Maisano, M.; Bernabò, I.; Giannetto, A.; de Domenico, E.; Corapi, B.; Tripepi, S.; Fasulo, S. Ultrastructural and immunohistochemical investigation on the gills of the teleost, Thalassoma pavo L., exposed to cadmium. Acta Histochem. 2011, 113, 201–213. [Google Scholar] [CrossRef]
- Cai, L.M.; Wang, Q.S.; Luo, J.; Chen, L.G.; Zhu, R.L.; Wang, S.; Tang, C.H. Heavy metal contamination and health risk assessment for children near a large Cu-smelter in central China. Sci. Total Environ. 2019, 650, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Cheema, A.I.; Liu, G.; Yousaf, B.; Abbas, Q.; Zhou, H. A comprehensive review of biogeochemical distribution and fractionation of lead isotopes for source tracing in distinct interactive environmental compartments. Sci. Total Environ. 2020, 719, 135658. [Google Scholar] [CrossRef]
- Shi, L.; Wang, N.; Hu, X.; Yin, D.; Wu, C.; Liang, H.; Cao, W.; Cao, H. Acute toxic effects of lead (Pb2+) exposure to rare minnow (Gobiocypris rarus) revealed by histopathological examination and transcriptome analysis. Environ. Toxicol. Pharmacol. 2020, 78, 103385. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, A.; MMS, C.P.; Chaturvedi, A.K.; Shabnam, A.A.; Subrahmanyam, G.; Mondal, R.; Gupta, D.K.; Malyan, S.K.; Kumar, S.S.; Khan, S.A.; Yadav, K.K. Lead toxicity: Health hazards, influence on food chain, and sustainable remediation approaches. Int. J. Environ. Res. Public Health 2020, 17, 2179. [Google Scholar] [CrossRef] [PubMed]
- United States Environmental Protection Agency, "Learn About Lead". EPA, 2022. https://www.epa.gov/lead/learn-about-lead.
- Bashir, I.; Lone, F.A.; Bhat, R.A.; Mir, S.A.; Dar, Z.A.; Dar, S.A. Concerns and threats of contamination on aquatic ecosystems. In Bioremediation and Biotechnology; Hakeem, K., Bhat, R., Qadri, H., Eds.; Springer: Cham, Switzerland, 2020; pp. 1–26. [Google Scholar] [CrossRef]
- Su, G.; Logez, M.; Xu, J.; Tao, S.; Villéger, S.; Brosse, S. Human impacts on global freshwater fish biodiversity. Science 2021, 371, 835–838. [Google Scholar] [CrossRef]
- Keke, U.N.; Mgbemena, A.S.; Arimoro, F.O.; Omalu, I.C. Biomonitoring of effects and accumulations of heavy metals insults using some helminth parasites of fish as bio-indicators in an Afrotropical stream. Front. Environ. Sci. 2020, 8, 576080. [Google Scholar] [CrossRef]
- Łuczyńska, J.; Paszczyk, B.; Łuczyński, M.J. Fish as a bioindicator of heavy metals pollution in aquatic ecosystem of Pluszne Lake, Poland, and risk assessment for consumer's health. Ecotoxicol. Environ. Saf. 2018, 153, 60–67. [Google Scholar] [CrossRef]
- Macirella, R.; Brunelli, E. Morphofunctional alterations in zebrafish (Danio rerio) gills after exposure to mercury chloride. Int. J. Mol. Med. Sci. 2017, 18, 824. [Google Scholar] [CrossRef]
- Paul, S.; Mandal, A.; Bhattacharjee, P.; Chakraborty, S.; Paul, R.; Mukhopadhyay, B.K. Evaluation of water quality and toxicity after exposure of lead nitrate in fresh water fish, major source of water pollution. Egypt. J. Aquat. Res. 2019, 45, 345–351. [Google Scholar] [CrossRef]
- Dai, J.; Zhang, L.; Du, X.; Zhang, P.; Li, W.; Guo, X.; Li, Y. Effect of lead on antioxidant ability and immune responses of Crucian carp. Biol. Trace Elem. Res. 2018, 186, 546–553. [Google Scholar] [CrossRef]
- Zhao, L.; Zheng, Y.G.; Feng, Y.H.; Li, M.Y.; Wang, G.Q.; Ma, Y.F. Toxic effects of waterborne lead (Pb) on bioaccumulation, serum biochemistry, oxidative stress and heat shock protein-related genes expression in Channa argus. Chemosphere 2020, 261, 127714. [Google Scholar] [CrossRef] [PubMed]
- Ishaque, A.; Ishaque, S.; Arif, A.; Abbas, H.G. Toxic effects of lead on fish and human. Biol. Clin. Sci. Res. J. 2020, 2020, e045. [Google Scholar] [CrossRef]
- Jing, H.; Zhang, Q.; Li, S.; Gao, X.J. Pb exposure triggers MAPK-dependent inflammation by activating oxidative stress and miRNA-155 expression in carp head kidney. Fish Shellfish Immunol. 2020, 106, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Choi, H.; Hwang, U.K.; Kang, J.C.; Kang, Y.J.; Kim, K.I.; Kim, J.H. Toxic effects of lead exposure on bioaccumulation, oxidative stress, neurotoxicity, and immune responses in fish: A review. Environ. Toxicol. Pharmacol. 2019, 68, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Paul, N.; Chakraborty, S.; Sengupta, M. Lead toxicity on non-specific immune mechanisms of freshwater fish Channa punctatus. Aquat. Toxicol. 2014, 152, 105–112. [Google Scholar] [CrossRef]
- Al-Balawi, H.F.A.; Al-Akel, A.S.; Al-Misned, F.; Suliman, E.A.M.; Al-Ghanim, K.A.; Mahboob, S.; Ahmad, Z. Effects of sub-lethal exposure of lead acetate on histopathology of gills, liver, kidney and muscle and its accumulation in these organs of Clarias gariepinus. Braz. Arch. Biol. Technol. 2013, 56, 293–302. [Google Scholar] [CrossRef]
- Macirella, R.; Sesti, S.; Bernabò, I.; Tripepi, M.; Godbert, N.; Brunelli, E. Lead toxicity in seawater teleosts: A morphofunctional and ultrastructural study on the gills of the Ornate wrasse (Thalassoma pavo L.). Aquat. Toxicol. 2019, 211, 193–201. [Google Scholar] [CrossRef]
- Curcio, V.; Macirella, R.; Sesti, S.; Ahmed, A. I.; Talarico, F.; Tagarelli, A.; Mezzasalma, M.; Brunelli, E. Morphological and functional alterations induced by two ecologically relevant concentrations of Lead on Danio rerio gills. Int. J. Mol. Sci. 23, 9165. [CrossRef] [PubMed]
- Zhai, Q.; Wang, H.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W. Dietary Lactobacillus plantarum supplementation decreases tissue lead accumulation and alleviates lead toxicity in Nile tilapia (Oreochromis niloticus). Aquacult. Res. 2017, 48, 5094–5103. [Google Scholar] [CrossRef]
- Bawuro, A.A.; Voegborlo, R.B.; Adimado, A.A. Bioaccumulation of heavy metals in some tissues of fish in Lake Geriyo, Adamawa State, Nigeria. J. Environ. Public Health 2018, 2018. [Google Scholar] [CrossRef]
- Tanhan, P.; Imsilp, K.; Lansubsakul, N.; Thong-asa, W. Oxidative response to Cd and Pb accumulation in coastal fishes of Pattani Bay. Ital. J. Anim. Sci. 2023, 22, 148–156. [Google Scholar] [CrossRef]
- Kumar, E.K.; Midhun, S.J.; Vysakh, A.; James, T.J. Antagonistic effects of dietary Moringa oleifera on hemato-biochemical and oxidative stress of lead nitrate intoxicated Nile tilapia, Oreochromis niloticus. Aquacult. Res. 2021, 52, 6164–6178. [Google Scholar] [CrossRef]
- Eroglu, A.; Dogan, Z.; Kanak, E.G.; Atli, G.; Canli, M. Effects of heavy metals (Cd, Cu, Cr, Pb, Zn) on fish glutathione metabolism. Environ. Sci. Pollut. Res. 2015, 22, 3229–3237. [Google Scholar] [CrossRef]
- Rajamanickam, D.; Devadason, C.G. Histopathological alterations in gill, liver, and brain of Nile Tilapia (Oreochromis Niloticus), exposed to Lead Nitrate (Pb [NO3]2). Int. J. Sci. Technol. Res. 2021, 3, 149–153. [Google Scholar]
- Mustafa, S.A.; Al-Faragi, J.K.; Salman, N.M.; Al-Rudainy, A.J. Histopathological alterations in gills, liver and kidney of common carp, Cyprinus carpio exposed to lead Acetate. Adv. Anim. Vet. Sci. 2017, 5, 371–376. [Google Scholar] [CrossRef]
- Doaa, M.M.; Hanan, H.A. Histological changes in selected organs of Oreochromis niloticus exposed to doses of lead acetate. J. Life Sci. Biomed. 2013, 3, 256–263. [Google Scholar]
- Abdel-Warith, A.W.A.; Younis, E.S.M.; Al-Asgah, N.A.; Rady, A.M.; Allam, H.Y. Bioaccumulation of lead nitrate in tissues and its effects on hematological and biochemical parameters of Clarias gariepinus. Saudi J. Biol. Sci. 2020, 27, 840–845. [Google Scholar] [CrossRef]
- Li, X.; Zhang, B.; Li, N.; Ji, X.; Liu, K.; Jin, M. Zebrafish neurobehavioral phenomics applied as the behavioral warning methods for fingerprinting endocrine disrupting effect by lead exposure at environmentally relevant level. Chemosphere 2019, 231, 315–325. [Google Scholar] [CrossRef]
- Curcio, V.; Macirella, R.; Sesti, S.; Ahmed, A.I.; Talarico, F.; Pizzolotto, R.; Tagarelli, A.; Mezzasalma, M.; Brunelli, E. The role of exposure window and dose in determining lead toxicity in developing Zebrafish. Chemosphere 307, 136095. [CrossRef]
- Lei, P.; Zhang, W.; Ma, J.; Xia, Y.; Yu, H.; Du, J.; Fang, Y.; Wang, L.; Zhang, K.; Jin, L.; Sun, D.; Zhong, J. Advances in the utilization of Zebrafish for assessing and understanding the mechanisms of Nano-/Microparticles toxicity in water. Toxics 2023, 11, 380. [Google Scholar] [CrossRef] [PubMed]
- Magyary, I. Recent advances and future trends in zebrafish bioassays for aquatic ecotoxicology. Ecocycles 2018, 4, 12–18. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, Y.; Liu, R.; Liu, C.; Chen, Y. Molecular mechanism of lead-induced superoxide dismutase inactivation in zebrafish livers. J. Phys. Chem. B 2014, 118, 14820–14826. [Google Scholar] [CrossRef]
- Xia, J.; Lu, L.; Jin, C.; Wang, S.; Zhou, J.; Ni, Y.; Zhengwei, F.; Jin, Y. Effects of short-term lead exposure on gut microbiota and hepatic metabolism in adult zebrafish. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2018, 209, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wang, T.; Zhang, X.; Chen, J.; Feng, C.; Yun, S.; Cheng, Y.; Cheng, F.; Cao, J. Sex-specific effects of fluoride and lead exposures on histology, antioxidant physiology, and immune system in the liver of zebrafish (Danio rerio). Ecotoxicology 2022, 31, 396–414. [Google Scholar] [CrossRef]
- Shah, Z.U.; Parveen, S. Oxidative, biochemical and histopathological alterations in fishes from pesticide contaminated river Ganga, India. Sci. Rep. 2022, 12, 3628. [Google Scholar] [CrossRef]
- Opute, P.A.; Oboh, I.P. Hepatotoxic effects of atrazine on Clarias gariepinus (Burchell, 1822): biochemical and histopathological studies. Arch. Environ. Contam. Toxicol. 2021, 80, 414–425. [Google Scholar] [CrossRef]
- Bakiu, R.; Pacchini, S.; Piva, E.; Schumann, S.; Tolomeo, A.M. , Ferro, D.; Irato, P.; Santovito, G. Metallothionein expression as a physiological response against metal toxicity in the striped rockcod Trematomus hansoni. Int. J. Mol. Sci. 2022, 23, 12799. [Google Scholar] [CrossRef]
- Wang, W.C.; Mao, H.; Ma, D.D.; Yang, W.X. Characteristics, functions, and applications of metallothionein in aquatic vertebrates. Front. Mar. Sci. 2014, 1, 34. [Google Scholar] [CrossRef]
- Hauser-Davis, R.A. The current knowledge gap on metallothionein mediated metal-detoxification in Elasmobranchs. PeerJ 2020, 8, e10293. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Wang, A.P.; Li, W.F.; Shi, R.; Jin, H.T.; Wei, J.F. Sensitive biomarkers identification for differentiating Cd and Pb induced toxicity on zebrafish embryos. Environ. Toxicol. Pharm. 2017, 56, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Maiti, A.K.; Saha, N.C.; Paul, G. Effect of lead on oxidative stress, Na+K+ATPase activity and mitochondrial election transport chain activity of the brain of Clarias batrachus L. Bull. Environ. Contam. Toxicol. 2010, 84, 672–676. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, J.C. Effects of sub-chronic exposure to lead (Pb) and ascorbic acid in juvenile rockfish: antioxidant responses, MT gene expression, and neurotransmitters. Chemosphere 2017, 171, 520–527. [Google Scholar] [CrossRef] [PubMed]
- Morcillo, P.; Esteban, M.Á.; Cuesta, A. Heavy metals produce toxicity, oxidative stress and apoptosis in the marine teleost fish SAF-1 cell line. Chemosphere 2016, 144, 225–233. [Google Scholar] [CrossRef]
- Bernet, D.; Schmidt, H.; Meier, W.; Burkhardt-Holm, P.; Wahli, T. Histopathology in fish: proposal for a protocol to assess aquatic pollution. J. Fish Dis. 1999, 22, 25–34. [Google Scholar] [CrossRef]
- Santos, R.M.B.; Monteiro, S.M.V.; Cortes, R.M.V.; Pacheco, F.A.L.; Fernandes, L.F.S. Seasonal differences in water pollution and liver histopathology of Iberian barbel (Luciobarbus bocagei) and Douro nase (Pseudochondrostoma duriense) in an agricultural watershed. Water 2022, 14, 444. [Google Scholar] [CrossRef]
- Macirella, R.; Curcio, V.; Ahmed, A.I.M.; Pellegrino, D.; Brunelli, E. Effect of short-term exposure to low concentration of tebuconazole: Morphological, histometric and functional modifications in Danio rerio liver. Eur. Zool. J. 2022, 89, 331–345. [Google Scholar] [CrossRef]
- Macirella, R.; Guardia, A.; Pellegrino, D.; Bernabò, I.; Tronci, V.; Ebbesson, L.O.; Sesti, S.; Tripepi, S.; Brunelli, E. Effects of two sublethal concentrations of mercury chloride on the morphology and metallothionein activity in the liver of zebrafish (Danio rerio). Int. J. Mol. Sci. 2016, 17, 361. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Ilahi, I. Environmental chemistry and ecotoxicology of hazardous heavy metals: Environmental persistence, toxicity, and bioaccumulation. J. Chem. 2019, 2019, 6730305. [Google Scholar] [CrossRef]
- Agbohessi, P.; Olowo, L.; Degila, B.; Houedjissi, G.; Imorou Toko, I.; Mandiki, S.N.; Kestemont, P. Comparative assessment of acute toxicity and histological changes in liver of African catfish Clarias gariepinus exposed to cotton insecticides. J. Environ. Sci. Health. Part B 2023, 58, 31–44. [Google Scholar] [CrossRef]
- Agamy, E. Histopathological changes in the livers of rabbit fish (Siganus canaliculatus) following exposure to crude oil and dispersed oil. Toxicol Pathol. 2012, 40, 1128–1140. [Google Scholar] [CrossRef] [PubMed]
- Popović, N.T.; Čižmek, L.; Babić, S.; Strunjak-Perović, I.; Čož-Rakovac, R. Fish liver damage related to the wastewater treatment plant effluents. Environ. Sci. Pollut. Res. 2023, 30, 48739–48768. [Google Scholar] [CrossRef]
- Khidr, B.M.; Mekkawy, I.A.; Harabawy, A.S.; Ohaida, A.S. Effect of lead nitrate on the liver of the cichlid fish (Oreochromis niloticus): a light microscope study. PJBS 2012, 15, 854–862. [Google Scholar] [CrossRef] [PubMed]
- Rajeshkumar, S.; Liu, Y.; Ma, J.; Duan, H.Y.; Li, X. Effects of exposure to multiple heavy metals on biochemical and histopathological alterations in common carp, Cyprinus carpio L. Fish Shellfish Immunol. 2017, 70, 461–472. [Google Scholar] [CrossRef]
- Shahid, S.; Sultana, T.; Sultana, S.; Hussain, B.; Irfan, M.; Al-Ghanim, K.A.; A-Misnedb, F.; Mahboob, S. Histopathological alterations in gills, liver, kidney and muscles of Ictalurus punctatus collected from pollutes areas of River. Braz. J. Biol. 2020, 81, 814–821. [Google Scholar] [CrossRef]
- Abalaka, S.E. Heavy metals bioaccumulation and histopathological changes in Auchenoglanis occidentalis fish from Tiga dam, Nigeria. J. Environ. Health Sci. Eng. 2015, 13, 1–8. [Google Scholar] [CrossRef]
- Liu, X.H.; Pang, X.; Jin, L.; Pu, D.Y.; Wang, Z.J.; Zhang, Y.G. Exposure to acute waterborne cadmium caused severe damage on lipid metabolism of freshwater fish, revealed by nuclear lipid droplet deposition in hepatocytes of rare minnow. Aquat. Toxicol. 2023, 257, 106433. [Google Scholar] [CrossRef]
- Paris-Palacios, S.; Biagianti-Risbourg, S.; Vernet, G. Biochemical and (ultra) structural hepatic perturbations of Brachydanio rerio (Teleostei, Cyprinidae) exposed to two sublethal concentrations of copper sulfate. Aquat. Toxicol. 2000, 50, 109–124. [Google Scholar] [CrossRef]
- Broeg, K. Acid phosphatase activity in liver macrophage aggregates as a marker for pollution-induced immunomodulation of the non-specific immune response in fish. Helgol. Mar. Res. 2003, 57, 166–175. [Google Scholar] [CrossRef]
- Valdez Domingos, F.X.; Assis, H.C.S.; Silva, M.D.; Damian, R.C.; Almeida, A.I.M.; Cestari, M.M.; Randi, M.A.F.; Oliveira Ribeiro, C.A. Anthropic Impact Evaluation of Two Brazilian Estuaries Trough Biomarkers in Fish. J. Braz. Soc. Ecotoxicol. 2009, 4, 21–30. [Google Scholar] [CrossRef]
- Sinha, R. Macrophage: a key player of teleost immune system. In Macrophages-140 Years of Their Discovery. IntechOpen 2022. [CrossRef]
- Curcio, V.; Macirella, R.; Sesti, S.; Pellegrino, D.; Ahmed, A.I.; Brunelli, E. Morphological and molecular alterations induced by lead in embryos and larvae of Danio rerio. Appl. Sci. 2021, 11, 7464. [Google Scholar] [CrossRef]
- Xia, X.; He, B.; Zhang, X.; Cheng, Z.; Liu, M.; Wei, X.; Jiang, J.; Hu, J. Lytic regulated cell death in aquaculture fish. Rev. Aquacult. 2021, 13, 1549–1564. [Google Scholar] [CrossRef]
- Conrad, M.; Kagan, V.E.; Bayir, H.; Pagnussat, G.C.; Head, B.; Traber, M.G.; Stockwell, B.R. Regulation of lipid peroxidation and ferroptosis in diverse species. Genes Dev. 2018, 32, 602–619. [Google Scholar] [CrossRef]
- Sun, Z.; Gong, C.; Ren, J.; Zhang, X.; Wang, G.; Liu, Y.; Ren, Y.; Zhao, Y.; Yu, Q.; Wang, Y.; Hou, J. Toxicity of nickel and cobalt in Japanese flounder. Environ. Poll. 2020, 263, 114516. [Google Scholar] [CrossRef] [PubMed]
- Gautheron, J.; Gores, G.J.; Rodrigues, C.M. Lytic cell death in metabolic liver disease. J. Hepatol. 2020, 73, 394–408. [Google Scholar] [CrossRef]
- Stephenie, S.; Chang, Y.P.; Gnanasekaran, A.; Esa, N.M.; Gnanaraj, C. An insight on superoxide dismutase (SOD) from plants for mammalian health enhancement. J. Funct. Foods 2020, 68, 103917. [Google Scholar] [CrossRef]
- Shi, Q.; Xiong, X.; Wen, Z.; Qin, C.; Li, R.; Zhang, Z.; Gong, Q.; Wu, X. Cu/Zn Superoxide dismutase and catalase of Yangtze Sturgeon, Acipenser dabryanus: molecular cloning, tissue distribution and response to fasting and refeeding. Fishes, 2022, 7, 35. [Google Scholar] [CrossRef]
- Alak, G.; Atamanalp, M.; Topal, A.; Arslan, H.; Kocaman, E.M.; Oruc, E. Effect of sub-lethal lead toxicity on the histopathological and antioxidant enzyme activity of rainbow trout (Oncorhynchus mykiss). Fresenius Environ. Bull. 2013, 22, 733–738. [Google Scholar]
- Jing, D.; Li, M.; Zhang, Y.; Yuan, L.; Wang, R.; Gong, Y. Differential induction of enzymes and genes involved in oxidative stress in gill and liver tissues of mudskipper Boleophthalmus pectinirostris exposed to lead. Turk. J. Fish. Aquat. Sci. 2017, 17, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Girgis, S.M.; Mabrouk, D.M.; Hanna, M.I.; Abd ElRaouf, A. Seasonal assessment of some heavy metal pollution and Metallothionein gene expression in cultured Oreochromis niloticus. Bull. Natl. Res. Cent. 2019, 43, 1–8. [Google Scholar] [CrossRef]
Figure 1.
Light micrographs of Danio rerio liver under basal condition. (a) General organization of the liver parenchyma; note the bile ducts enclosed by cuboidal epithelium. (b) High magnification showing the space of Disse between the hepatocytes and the sinusoid wall. Note erythrocytes and a few macrophages in the lumen of the veins. bd=bile duct, s=sinusoid, v=vein, n=nucleus, arrow=glycogen granules, arrowhead=lipid droplets, sd=space of Disse.
Figure 1.
Light micrographs of Danio rerio liver under basal condition. (a) General organization of the liver parenchyma; note the bile ducts enclosed by cuboidal epithelium. (b) High magnification showing the space of Disse between the hepatocytes and the sinusoid wall. Note erythrocytes and a few macrophages in the lumen of the veins. bd=bile duct, s=sinusoid, v=vein, n=nucleus, arrow=glycogen granules, arrowhead=lipid droplets, sd=space of Disse.
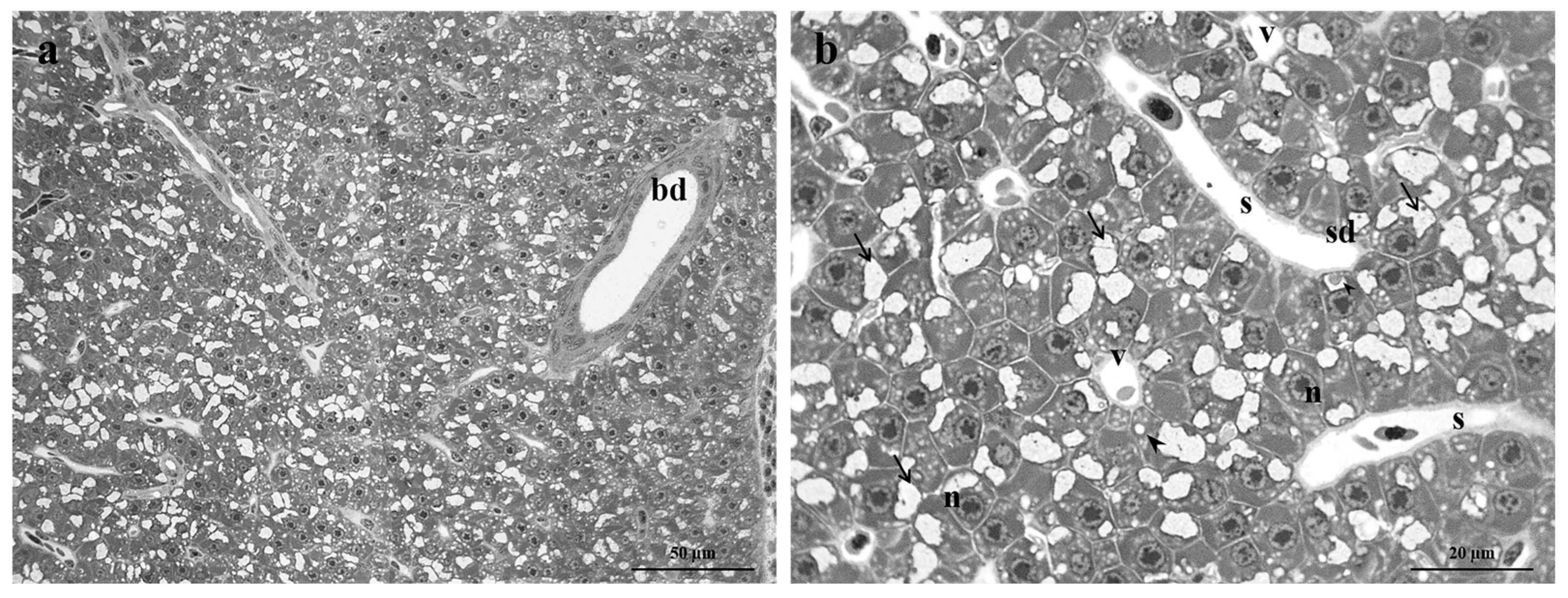
Figure 2.
Light micrographs of Danio rerio liver after exposure to 2.5 µg/L of Pb. (a-c) After 48 hours of exposure, bile duct degeneration and the congestion of blood vessels and sinusoids were observed. Note the increase in lipid droplet content. (d-g) After 96 hours of exposure, the cuboidal epithelium lining the bile ducts was modified, and the detachment of the duct epithelium was evident. Note the congestion of blood vessels and sinusoids, macrophage proliferation, cytoplasmic vacuolization, the increase in lipid droplet content, and the appearance of both apoptotic and necrotic hepatocytes. Also, lysed areas were frequently observed. (h-j) After 192 hours of exposure, cytoplasm vacuolization, the congestion of vessels and sinusoids, and numerous lipid droplets were frequently detected. Note the detachment of bile duct epithelium, macrophage proliferation, and both apoptotic and necrotic hepatocytes. bdd=bile duct degeneration, vc=vessel congestion, sc=sinusoids congestion, black arrow= bile duct epithelial detachment, black arrowhead=lipid droplets, white arrowhead=cytoplasmic vacuolization, white arrow=macrophages proliferation, white star=apoptotic cell, black star=necrotic cell, la=lysed area.
Figure 2.
Light micrographs of Danio rerio liver after exposure to 2.5 µg/L of Pb. (a-c) After 48 hours of exposure, bile duct degeneration and the congestion of blood vessels and sinusoids were observed. Note the increase in lipid droplet content. (d-g) After 96 hours of exposure, the cuboidal epithelium lining the bile ducts was modified, and the detachment of the duct epithelium was evident. Note the congestion of blood vessels and sinusoids, macrophage proliferation, cytoplasmic vacuolization, the increase in lipid droplet content, and the appearance of both apoptotic and necrotic hepatocytes. Also, lysed areas were frequently observed. (h-j) After 192 hours of exposure, cytoplasm vacuolization, the congestion of vessels and sinusoids, and numerous lipid droplets were frequently detected. Note the detachment of bile duct epithelium, macrophage proliferation, and both apoptotic and necrotic hepatocytes. bdd=bile duct degeneration, vc=vessel congestion, sc=sinusoids congestion, black arrow= bile duct epithelial detachment, black arrowhead=lipid droplets, white arrowhead=cytoplasmic vacuolization, white arrow=macrophages proliferation, white star=apoptotic cell, black star=necrotic cell, la=lysed area.
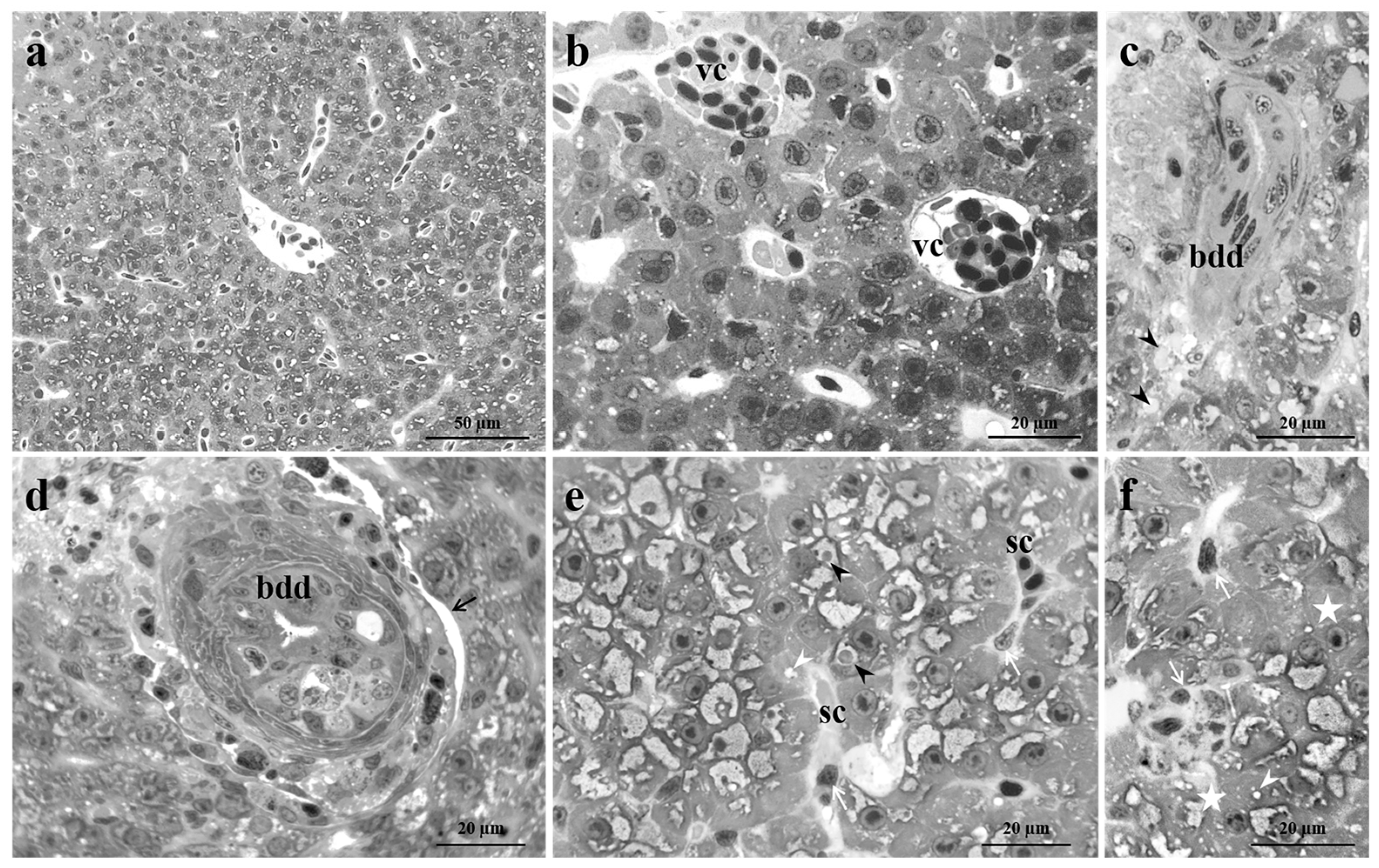
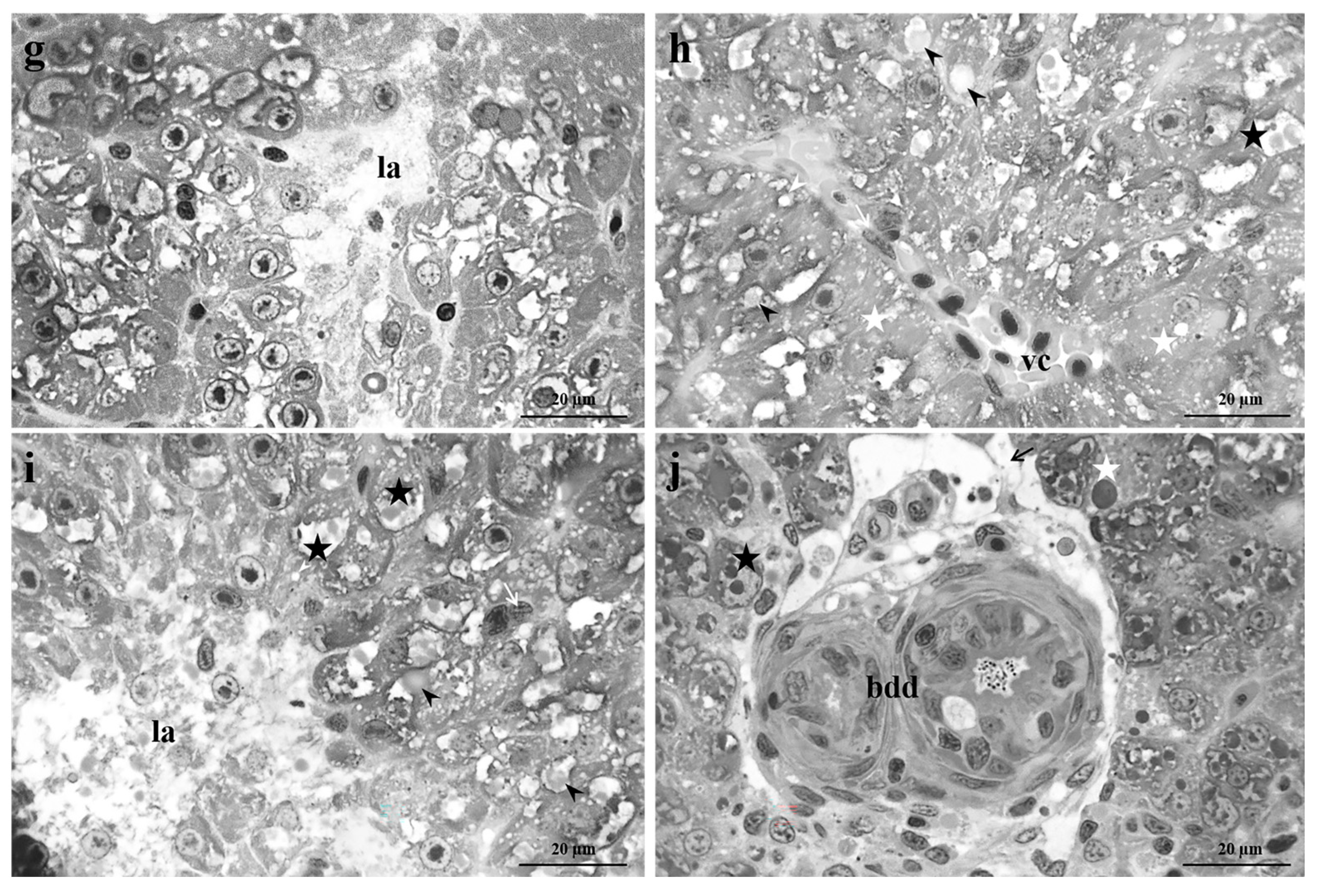
Figure 3.
Prevalence of histological alterations in Danio rerio liver after exposure to 2.5 and 5 µg/L of Pb for 48, 96, and 192 hours. a significance of low concentration group with respect to the control group, b significance of the high concentration group with respect to the control group, c significance of the high concentration group with respect to the low concentration group. *p≤0.05; **p≤0.005; ****p≤0.0001.
Figure 3.
Prevalence of histological alterations in Danio rerio liver after exposure to 2.5 and 5 µg/L of Pb for 48, 96, and 192 hours. a significance of low concentration group with respect to the control group, b significance of the high concentration group with respect to the control group, c significance of the high concentration group with respect to the low concentration group. *p≤0.05; **p≤0.005; ****p≤0.0001.
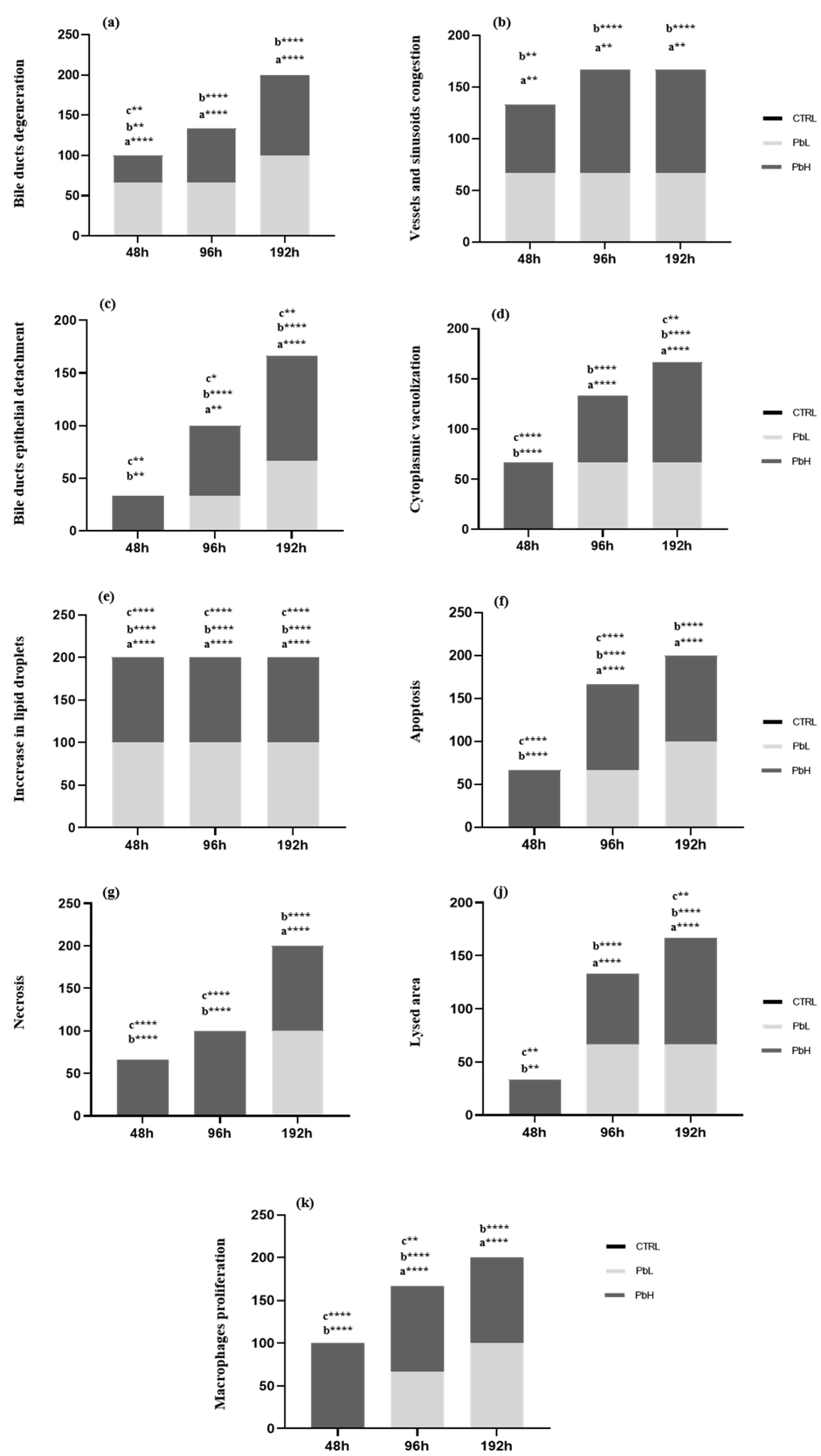
Figure 4.
Light micrographs of Danio rerio liver after exposure to 5 µg/L of Pb. (a-c) After 48 hours of exposure, bile duct degeneration, bile duct epithelial detachment, wide lysed areas and numerous lipid droplets could be detected. Moreover, the vacuolization of hepatocytes cytoplasm, the congestion of blood vessels, and both apoptotic and necrotic were frequently visible. Note the proliferation of macrophages. (d-f) After 96 hours of exposure, numerous degenerations such as vessels and sinusoid congestion, macrophage proliferation, and apoptotic hepatocytes, were visible in all samples. Also, the increase in lipid contents, cytoplasmic vacuolization, bile duct degeneration, detachment of the bile duct epithelium and lysed areas were frequently detected. (g-i) After 192 hours of exposure, all the considered alterations were detected in all samples. bdd=bile duct degeneration, vc=vessel congestion, sc=sinusoids congestion, black arrow= bile duct epithelial detachment, black arrowhead=lipid droplets, white arrowhead=cytoplasmic vacuolization, white arrow=macrophages proliferation, white star=apoptotic cell, black star=necrotic cell, la=lysed area.
Figure 4.
Light micrographs of Danio rerio liver after exposure to 5 µg/L of Pb. (a-c) After 48 hours of exposure, bile duct degeneration, bile duct epithelial detachment, wide lysed areas and numerous lipid droplets could be detected. Moreover, the vacuolization of hepatocytes cytoplasm, the congestion of blood vessels, and both apoptotic and necrotic were frequently visible. Note the proliferation of macrophages. (d-f) After 96 hours of exposure, numerous degenerations such as vessels and sinusoid congestion, macrophage proliferation, and apoptotic hepatocytes, were visible in all samples. Also, the increase in lipid contents, cytoplasmic vacuolization, bile duct degeneration, detachment of the bile duct epithelium and lysed areas were frequently detected. (g-i) After 192 hours of exposure, all the considered alterations were detected in all samples. bdd=bile duct degeneration, vc=vessel congestion, sc=sinusoids congestion, black arrow= bile duct epithelial detachment, black arrowhead=lipid droplets, white arrowhead=cytoplasmic vacuolization, white arrow=macrophages proliferation, white star=apoptotic cell, black star=necrotic cell, la=lysed area.
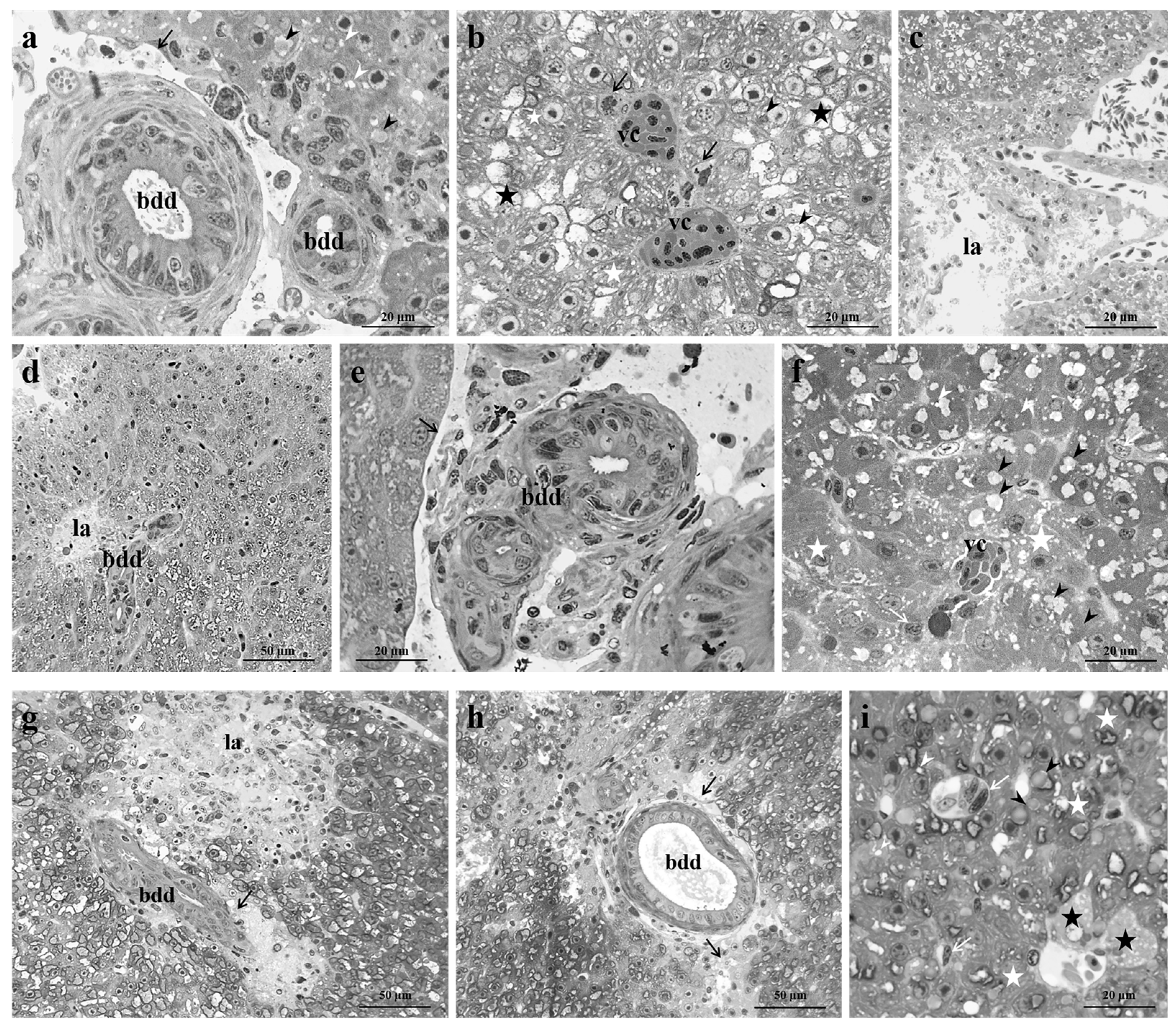
Figure 5.
Percentage of the area occupied by lipid droplets in Danio rerio liver after exposure to 2.5 and 5 µg/L of Pb for 48, 96, and 192 hours. Graphs indicate the mean ± S.D. Asterisks indicate significant differences between the treated and control groups. Hashtags indicate significant differences between the high Pb concentration group and the low Pb concentration group. ***p≤0.001; ****p≤0.0001; ##p≤0.005; ###p≤0.001.
Figure 5.
Percentage of the area occupied by lipid droplets in Danio rerio liver after exposure to 2.5 and 5 µg/L of Pb for 48, 96, and 192 hours. Graphs indicate the mean ± S.D. Asterisks indicate significant differences between the treated and control groups. Hashtags indicate significant differences between the high Pb concentration group and the low Pb concentration group. ***p≤0.001; ****p≤0.0001; ##p≤0.005; ###p≤0.001.
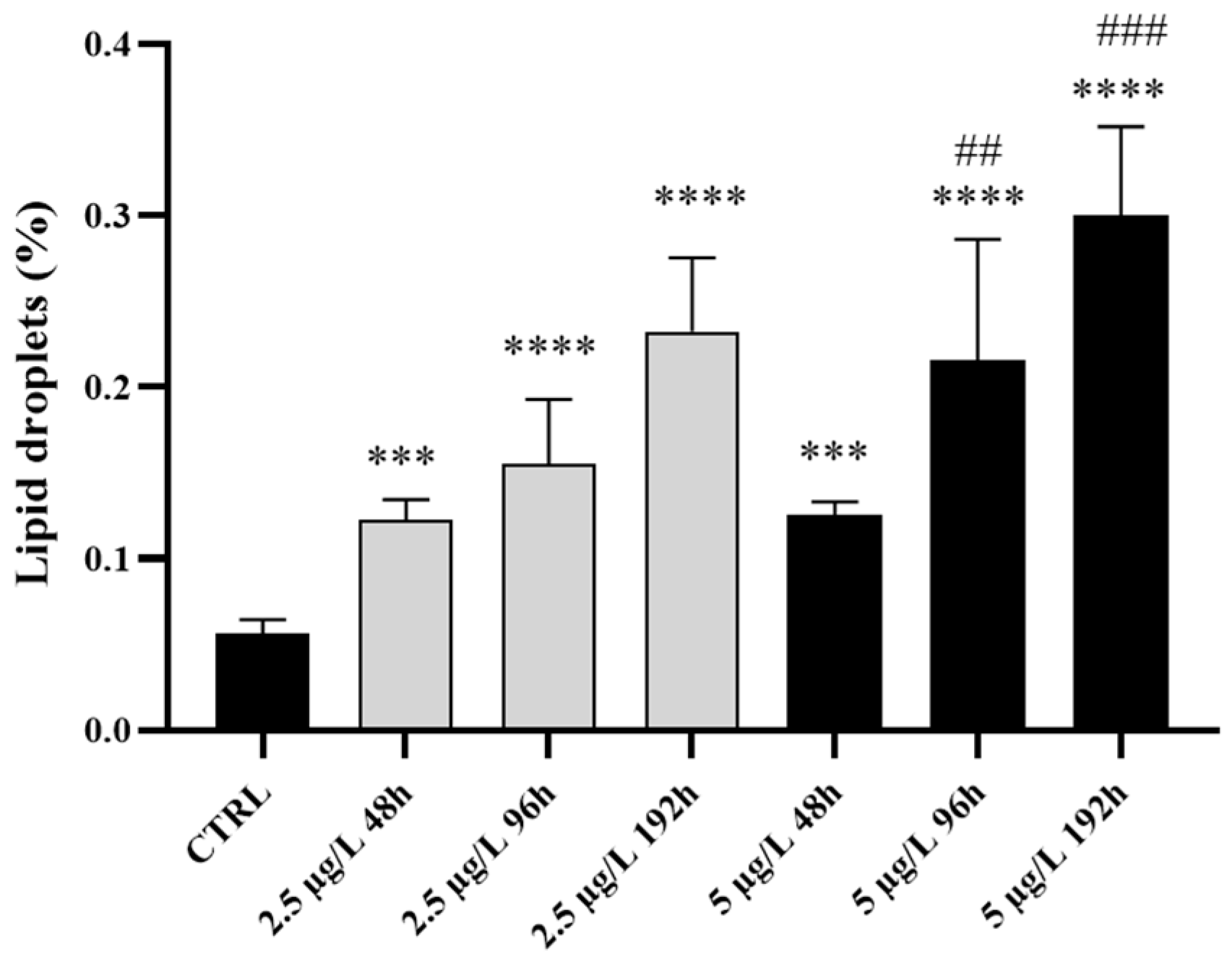
Figure 6.
Gene expression in Danio rerio liver after exposure to 2.5 and 5 µg/L of Pb for 48, 96, and 192 hours. Graphs indicate the mean±S.D. (a) Metallothioneins (mtf1), (b) Catalase (cat), (c) Superoxide dismutase (sod1). Asterisks indicate significant differences between treated and control groups. Hashtags indicate significant differences between the high concentration and the low concentration groups. *p≤0.05; **p≤0.005; ***p≤0.001; ****p≤0.0001; ####p≤0.0001.
Figure 6.
Gene expression in Danio rerio liver after exposure to 2.5 and 5 µg/L of Pb for 48, 96, and 192 hours. Graphs indicate the mean±S.D. (a) Metallothioneins (mtf1), (b) Catalase (cat), (c) Superoxide dismutase (sod1). Asterisks indicate significant differences between treated and control groups. Hashtags indicate significant differences between the high concentration and the low concentration groups. *p≤0.05; **p≤0.005; ***p≤0.001; ****p≤0.0001; ####p≤0.0001.
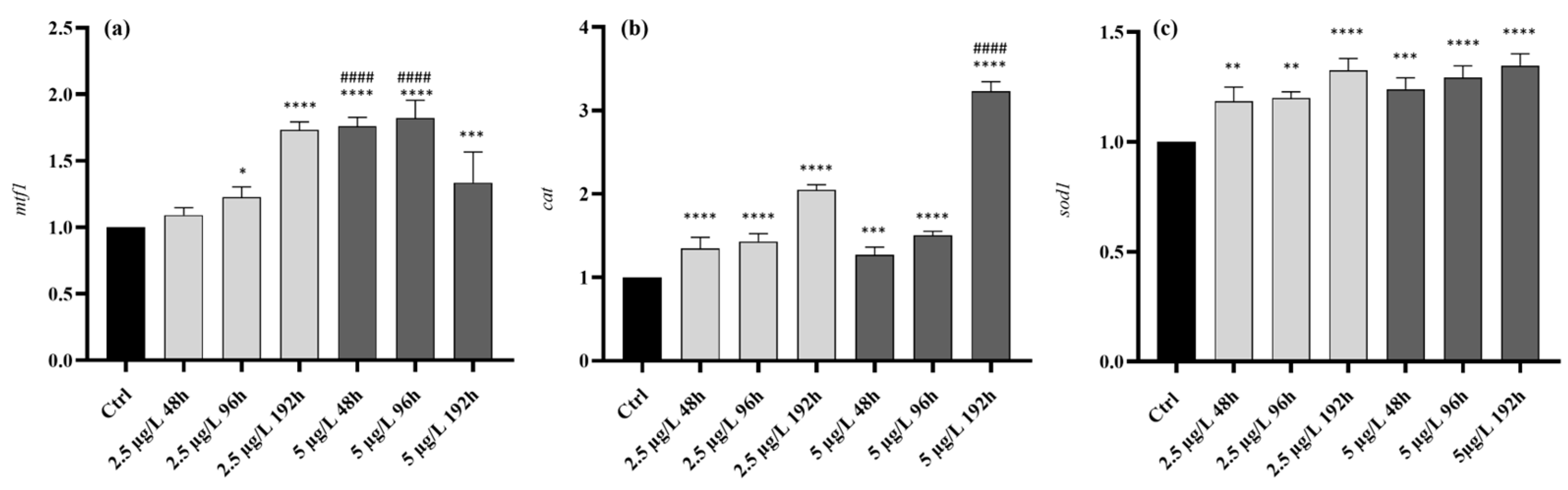

Table 1.
Comparison in the organ index (mean ± SD) between control and Pb exposed groups.
| 2.5 µg/L Pb | 5 µg/L Pb | |
|---|---|---|
| CTRL | 0.00±0.00 | 0.00±0.00 |
| 48 hours | 2.66±1.15 | 16.00±5.29(a***)(b**) |
| 96 hours | 14.66±3.05(a**)(c*) | 42.00±2.40(a****)(b****)(c****) |
| 192 hours | 35.33±4.61(a****)(c****) | 53.33±3.05(a****)(b****)(c*) |
Class I (index ≤ 10) normal organ structure; Class II (index 11–20) slight histological alterations; Class III (index 21–30) moderate histological alterations; Class IV (index 31-40) pronounced histological alterations of the organ; Class V (index > 40) severe histological alterations. a = significance of low or high concentration group with respect to control group; b = significance of high concentration group with respect to low concentration group; c = significance of 96 h treatment with respect to 48 h treatment or 192 h treatment with respect to 96 h treatment. *p≤0.05; **p≤0.01; ***p=0.001; ****p≤0.0001.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



