
Submitted:
26 May 2023
Posted:
29 May 2023
You are already at the latest version
Abstract
Abstract: In Australia adoption of reduced tillage and stubble retention cropping systems to conserve soil moisture by growers has seen an increase in the prevalence of the disease Fusarium crown rot (FCR) caused by the stubble-borne fungal pathogen Fusarium pseudograminearum. Durum wheat is particularly susceptible to FCR expressing significant yield and quality losses in the presence of infection. Increasing rates of nitrogen (N) application at sowing exacerbates FCR. However, to achieve the desired grain protein and quality suited to pasta manufacture, N application is necessary and this creates a dilemma for growers. The purpose of this study was to investigate the effects of FCR infection in the presence of different N fertiliser application rates in durum wheat varieties on yield and technological quality. Two durum varieties were evaluated at the same location over two seasons (2020 and 2021). These seasons were characterised by being wetter than normal and showed different responses to FCR and N application. In general, the 2021 season showed better responses to applied N with no impact of FCR. The FCR inoculation while it resulted in significant infection, it had no impacts on yield or grain quality in 2021 while in 2020 yield was reduced and only gluten index was reduced. Jandaroi was found to hold its kernel vitreosity at all protein levels obtaining the premium grade while DBA Lillaroi did not and would suffer downgrade if N application was insufficient. However, higher N application rates needed to achieve 12% + protein saw a reduction in dough strength with Jandaroi holding its dough strength much better than DBA Lillaroi. This was related to the lower glutenin/gliadin (Gli/Glu) ratio in response to applied N at sowing in Jandaroi which helped retain kernels with high vitreousness. This suggests genetics plays an important role in a genotype response to N fertilisation and should be considered when selecting a variety where higher premium grades are desirable.
Keywords:
Fusarium crown rot
; durum wheat
; agronomy
; glutenin
; gliadin
; semolina quality
; grain quality
1. Introduction
Durum wheat (Triticum durum L. ssp. Durum Desf.) is an important crop for the human diet grown in many parts of the world predominantly for semolina and resulting pasta production. Durum is a high yielding wheat typically grown under rainfed conditions in semiarid regions, that are characterised by unpredictable and highly variable seasonal rainfall affecting yield and quality [1]. In the Australian environment, most durum wheat is grown in Northern New South Wales (NNSW) and South Australia. In NNSW durum wheat is grown as a dryland crop on fertile cracking clay soils that can store water in the profile from the typical summer rains and this enables the crop to better tolerate heat and moisture stress during ear emergence, anthesis and grain filling. Durum attracts a premium price in the market but this depends on meeting grading standards in some countries [2] and in Australia the key quality attributes to ensure the highest grade (ADR1) are grain protein ≥13% (11% mb) (GP), test weight ≥76 kg/hl (TW), screenings <5%, hard vitreous kernels ≥80% (HVK) and other factors related to seed contamination and staining [3]. To achieve high grain protein usually requires high levels of nitrogen (N) input, reliant on the application of artificial N, commonly urea, which represents around 30-40% of input costs when targeting these higher quality cereal options [4]. Decisions on how much N to apply, when and choice of variety together with climatic and soil factors can all affect yield and market grading achieved for durum wheat in the NNSW environment. Understanding these interactions is important to achieve the most profitable outcome for the grower.
In Australia adoption of reduced tillage and stubble retention cropping systems to conserve soil moisture by growers revolutionised crop production in this region but has seen an increase in the prevalence of the disease Fusarium crown rot (FCR) caused by the stubble-borne fungal pathogen Fusarium pseudograminearum [5,6]. FCR is a basal infection in winter cereal plants that restricts water flow from the roots to the heads under moisture and/or temperature stress during grain-fill. This results in the formation of distinctive ‘whiteheads’ which are associated with yield loss along with reduced grain size that increases levels of screenings at harvest with potential to downgrade the grain [7]. Increasing rates of N application at sowing exacerbates the incidence and increases yield and grain quality (screenings) losses associated with FCR [8,9]. Consequently, producing ADR1 durum wheat in NNSW under dryland conditions has an inherently elevated risk of FCR given the level of climatic variability within and between seasons possibly becoming worse with more extreme and frequent drought and high temperature stress during grain-fill [10]. This balance between risk vs. reward is particularly frustrating for growers when harvested grain fails to meet quality specifications in terms of protein achievement, screenings and HVK. Although quantitative data on the proportion of grain receivals meeting ADR in NNSW is not freely available, analysis of National Variety Trials from 2014-2018 indicates that only 49% of the durum crop achieves the top ADR1 grade, which is supported by estimates from GrainCorp Pty. Ltd. (personal communication). This indicates that the majority of growers are not getting this balance right, where maximum yield may not necessarily equal optimum nitrogen use efficiency, water-use efficiency or profitability.
Despite the large number of studies that have examined the effect of N fertiliser applications on grain protein and quality in bread wheat (for a review see 11), there have been fewer studies on durum wheat and indeed no studies to the authors knowledge on the effect of N fertilisation in the presence of FCR on durum quality. The objectives of this study were to investigate the effects of FCR infection in the presence of different N fertiliser application rates in two commercial durum wheat varieties on grain yield, grain and semolina quality and relate the changes obtained to the mature grain gluten composition. Understanding these interrelationships will help determine the optimal management strategy for durum wheat production in the NNSW environment where FCR is an endemic constraint to production.
2. Materials and Methods
2.1. Plant material and field trials
2.1. Location and Soil Characteristics
Field experiments were conducted at the Tamworth Agricultural Institute (S 31.1468; E 150.9823) in northern NSW, Australia. The experiments were repeated in sequential years across the 2020 and 2021 winter growing seasons. Soil nitrogen (N) levels were determined from segmented soil cores (0-30 cm, 30-60 cm, 60-90 cm and 90-120 cm) collected from each site at sowing. In 2020, the field experiment was conducted on a red chromosol soil whilst in 2021 the experiment was located on a grey vertosol. There was around 250 kg/ha of nitrate N in 2020 and 64 kg/ha in 2021 within the soil profile (0-120 cm) at sowing.
2.2. Experimental Treatments and Growing Conditions
Two durum wheat varieties, DBA Lillaroi and Jandaroi were grown in each of the two experimental years across each site at a target plant population of 120 plants/m2 based on seed germination levels and 1000 grain weights.
Three levels of FCR infection were created using inoculum of Fusarium pseudograminearum prepared as outlined in Forknall et al. [12]. Inoculation rates in 2020 were nil, 0.5 g/m row (low infection level) or 2.0 g/m row (high infection level) of FCR inoculum applied at sowing. Inoculum rates were adjusted in 2021 to provide more intermediate levels of FCR infection of nil, 0.75 g/m row (low-medium) or 1.5 g/m row (medium-high). Five rates of N application of 0, 50, 100, 150 or 200 units N/ha were applied as urea at sowing in 2020. Sorghum was grown across the site in the summer of 2020/21 prior to establishing the durum experiment in 2021 to lower residual soil nitrogen levels. Consequently, N application rates were varied to 0, 20, 40, 80 and 160 units N/ha applied as urea at sowing in 2021 to provide increased ability to determine responses and interactions at lower levels of N nutrition. In both years the field experiments were sown using a small plot seeder using tynes with a split boot arrangement that physically separate the urea from the seed at sowing by offsetting the fertilizer to the side (~2 cm) and below (~2 cm) seed within the planting furrow. This avoided the experiments being compromised by any potential damage to emerging seedlings from fertiliser burn, especially at the higher rates of N application. Plots in both seasons consisted of five plant rows on 33 cm spacings (1.65 m wide) with a 5.0 m plot length. There were four replicate plots of each treatment in a fully randomised complete block design. Grain yield was determined at physiological maturity using a small plot header and grain retained for quality assessment. The incidence of FCR infection was determined at harvest from 25 random primary tillers collected from each plot. Crown segments from each tiller (2 cm length) were trimmed, surface sterilised and cultured on 1/4 strength Potato Dextrose Agar (PDA) + novobiocin (10 g PDA, 15 g technical agar plus 0.1 g novobiocin/L water) and incubated under alternating white and near ultraviolet lights for a 12 h photoperiod of 66.6% alternating fluorescent (FL36W/865, Sylvania, East Sussex, United Kingdom) and 33.3% blacklight blue (F36T8 BLB, Crompton lighting, Bradford, United Kingdom) light for 7 days at 25°C. Following incubation, distinctive colonies of Fusarium pseudograminaerum arising from surface sterilised crowns were counted to determine the incidence of FCR infection within each plot. Climatic conditions of rainfall and temperature were recorded from the nearest Bureau of Meteorology site in each season (Table 1).
2.2. Grain and semolina analyses
Harvested grain was assessed for screenings defined as the percentage by weight of the total sample passing through a 2-mm slotted screen after 40 shakes of a sieve. Following harvest, grain samples were cleaned with a Carter-Day dockage tester and individual field replicates analysed for grain protein (GP%, 11%mb), moisture and hardness (SKHI) by near infrared reflectance spectroscopy. The test weight (TW), thousand grain weight (TGW) and falling number were determined on clean grain which was then milled (3 kg) into semolina using a Buhler laboratory mill as described elsewhere [13]. Milling performance assessed as milling yield (MY = Total Flour Weight/Total Products Weight *100), semolina yield (SY = semolina weight (three break fractions + semolina)/(total products weight-bran, pollard, break, reduction and semolina fractions) × 100); semolina only yield (Semo Only = semolina weight/total products weight × 100). The percentage of hard vitreous kernels (HVK%) was determined using a Pohl Farinator device (https://www.bastak.com/eng/products/pohl-farinator-for-cutting-kernels). HVK% = ([300-number of mottled kernels]/300) X 100. Semolina colour was evaluated by measuring L* (brightness, 100 = white, 0 = black), a* (+ve is redness. –ve is greenness), b* (+ve is yellow, –ve is blue) and whiteness index parameters by means of a Minolta Chroma Meter CR-410 (Biolab Australia, Sydney) in triplicate. Dough properties were assessed using a Mixograph and Glutomatic with the mean of duplicate analyses presented for each field replicate [13]. Key measures captured were mixograph dough development time (MPT) and resistance breakdown (RBD) [100x((width of the curve at peak mixing time – width of the curve after 8 min mixing)/width of the curve at peak mixing time)], wet gluten (WG) and gluten index (GI).
2.3. Gluten protein composition analysis of harvested grain from 2021 trial
Mature, harvested grain was milled into wholemeal using a Perten falling number mill and the glutenin and gliadin proteins extracted and separated on a BioSep size-exclusion column using a high performance liquid chromatograph (HPLC) apparatus described elsewhere [14]. Briefly, the extraction produces two protein fractions, one soluble in a phosphate buffer containing a detergent (SDS) and the other fraction protein that can only be solubilised using a sonication device to disrupt the polymer. Each flour sample was extracted in duplicate and each protein fraction (designated soluble and insoluble) injected onto the HPLC column and protein constituents separated. The soluble fraction separates into three fractions, glutenin polymers, gliadins and albumin/globulins. While the insoluble fraction separates into three polymeric glutenin fractions. Key measures from the chromatograms are area under a peak as percentage of total area (soluble A%, soluble B%, soluble C%, insoluble A%, insoluble B% and insoluble C%) and derived terms such as polymeric/monomeric ratio (P/M=[SolA area+InsolA area]/[SolB area+SolC area+InsolB area+InsolC area]), Gliadin/Glutenin ratio (Gli/Glu=[SolB area+InsolB area]/[SolA area+InsolA area]) and the percentage of unextractable polymeric protein (UPP=InsolA area/[InsolA area+SolA area] x 100).
2.4. Statistical analysis
Data were analysed using the statistical programme GenStat version 17.1.0.14713 with a generalised linear model and the means were tested for significant differences by the least significant difference statistic (LSD), P < 0.05. Data were checked for normality.
3. Results
3.1. Results from the 2020 season
High background soil nitrogen (N) levels at sowing (~250 kg/ha nitrate N in soil between 0-120 cm) resulted in this site being unresponsive to N application in terms of yield in both varieties (Figure 1). Except for DBA Lillaroi at the 0 and 100 Nrates, FCR infection resulted in significant yield loss compared with uninoculated (nil) plots at both infection levels. Yield loss under moderate FCR infection (0.5 g/m of row) ranged from 15 to 27% in Jandaroi and from not significant to 19% in DBA Lillaroi. At the higher level of FCR infection (2.0 g/m of row) yield loss ranged from 19 to 32% in Jandaroi and 18 to 38% in DBA Lillaroi. There was no interaction between yield loss from FCR and rate of N application rate at sowing at this site in 2020. Baseline grain protein (GP) with no applied N was high (13.7 to 13.9%) with a blunted but significant increase in GP in response to applied N rates (0-200 kgN/ha) increasing in Jandaroi by 7.1% and for DBA Lillaroi by 8.7% (Figure 2). Jandaroi had a higher GP and WG than DBA Lillaroi. Starting soil N levels at this site in 2020 appeared to be too high which limited responses to additional N inputs. Inoculation with Fusarium pseudograminearum (FCR treatment) had no significant effect on GP (data not shown). The functional component of the grain protein, the wet gluten content follows a similar trend to GP because the majority of protein in the kernel is gluten, so a high correlation (r = 0.75, n = 120, p<0.05) was observed, although only small increases in wet gluten were obtained (Table S1).
Grain size expressed as 1000 grain weight (TGW), packing density as test weight (TW) and percentage of small kernels, as screenings (SCR) were affected differently by the three factors (variety, Nrate and FCR). For TGW only variety and Nrate were significant. DBA Lillaroi has a larger grain size at all N rates than Jandaroi with no effect of Nrate on TGW in Jandaroi while it tended to decrease in DBA Lillaroi which was not consistent at 150N (Figure 3). TW was not affected by Nrate but variety and disease were significant, with no difference in DBA Lillaroi with FCR level but decreased in Jandaroi (Figure 4). Screenings (SCR) was significantly affected by variety and FCR with Jandaroi having higher and more variable SCR than DBA Lillaroi, sometimes exceeding the market cut-off of 5% for a downgrade to feed grade. FCR increased SCR in Jandaroi only with no difference at the two rates of inoculation (Table S2). All grain samples showed very high vitreousness (HVK) averaging 99.8% (data not shown).
Kernel characteristics can impact the milling potential of the grain which is important for economic returns of high yielding low speck and acceptable ash content semolina. Milling quality was assessed by measuring laboratory milling yields. The semolina yield (SY) was significantly affected by variety and disease but not Nrate (Figure 5, Table S3). DBA Lillaroi had significantly higher SY under all treatments than Jandaroi with a tendency to increase with FCR inoculation being highest at 2 g/m rate while Jandaroi SY was not significantly changed. A more sensitive indicator is the measure, Semolina Only yield (Semo only) but this was significant for variety and Nrate not FCR. Again, DBA Lillaroi had a Semo only yield higher at all Nrates than Jandaroi (Figure 6). There was a tendency for Semo only to decline in both varieties with Nrate but more so for DBA Lillaroi being lowest at 200 kg/ha compared to no additional applied N at sowing.
Semolina quality was also assessed by its colour parameters, brightness (L*), redness (a*) and yellowness (b*). The colour of the semolina is important as this determines the colour of the pasta. Brightness L* achieved high values ~80 with minimal differences between samples (data not shown) while b* mean varied from 27.9-31.5 with DBA Lillaroi having higher yellowness at all treatments than Jandaroi (Figure 7). Yellowness tended to decline slightly with increased Nrate (Figure 7) and FCR inoculum rate (data not shown) but not always consistently.
Dough strength (gluten index, GI) showed significant effects by genotype and disease. Jandaroi is known for its high GI averaging 93 compared to DBA Lillaroi of 68 in the absence of FCR inoculation. Values above 90 are considered to have very strong dough properties. Inoculation had a significant effect in reducing GI in both varieties with a larger decline at the highest FCR inoculation rate (Figure S1). This was a bigger issue for DBA Lillaroi declining by up to 21% compared to Jandaroi 10%. While Jandaroi also showed reduced GI with inoculation, it still achieved high GI values dropping only to 83, which is still high but the decline in DBA Lillaroi would cause processing issues. In 2020 there was minimal change in dough strength with N rate (Figure 8). The mixograph is another instrument to assess the dough properties of semolina and two keys measures, MPT and RBD both showed significant genotype and disease effects (Figure 9, Table S4). Jandaroi MPT declined slightly with low and high inoculation while DBA Lillaroi MPT did not change. However, RBD is a better indicator of dough strength correlating with GI (r = 0.49, p<0.05) with no change in Jandaroi but an increase in DBA Lillaroi with moderate-high FCR infection consistent with the large effect on GI from inoculation. Nrate had no effect on RBD in this season. Overall, GI was the most responsive trait to FCR infection (Table S4).
3.2. Results from the 2021 season and comparison to 2020 season
The mild weather conditions, wet and cool during grain filling in 2020 reduced the expression of FCR and the high pre-sowing soil N level also restricted the grain protein response. To reduce the likelihood of this happening in the 2021 season, the field trial was established following a commercial sorghum crop grown across the site in the summer prior to sowing the durum wheat experiment and this significantly reduced starting soil N levels (~64 kg/ha nitrate N in soil between 0-120 cm).
Grain yield in both durum varieties increased in response to applied N (20 to 160 kg/ha) at sowing when compared to the nil N treatment by up to 29% and 36% for Jandaroi and DBA Lillaroi, respectively in the absence of FCR inoculation (Figure 10). There was also a significant decrease in yield due to FCR infection which were highest at the 1.5 g/m row inoculation rate but no significant effect from FCR inoculation in 2021 except at the highest rate for Jandaroi 20N and DBA Lillaroi 80N compared to nil FCR. In 2020 Nrate had no effect on GY compared to a large positive effect in 2021 because of the high starting soil N and so considerably higher yields obtained (overall yield 2020, ~2 t/ha vs. 2021, 4.2 - 5.5 t/ha in absence of inoculum). There were no significant interactions between FCR infection and N rate.
There were good GP responses in 2021 (range 2.4%) compared to 2020 (range 1.04%) from 10.1% at 0 N up to 12.8% at 160 kgN/ha increasing on average by 23% and 24% for Jandaroi and DBA Lillaroi, respectively (Figure 2). There was a bigger increase in GP between 40 to 160 kgN/ha application rates that were significant compared to between 20 to 40 kgN/ha that were not significant. Both varieties achieved similar protein levels with Jandaroi slightly higher than DBA Lillaroi over the range of N levels. There was no effect of FCR infection on GP as was the case in 2020. The application rates selected differed from those in 2020 due to more refinement of the treatments to obtain a better spread in FCR infection levels. Much higher responses in GP were achieved in 2021 using the strategy to lower starting soil N rather than the blunted and narrow range obtained over a larger range of applied N in 2020. This aligned with the GY response in 2021 although FCR inoculation did not significantly affect GP due to prolonged wet and mild temperature conditions during grain-fill limiting the expression of FCR. Periods of heavy rain around harvest impacted grain falling number averaging 250 sec ranging from 185-290 sec (data not shown). Screenings level was very low ranging 0.7-1.2% well below the market cut-off of 5% in 2021 compared to some plots experiencing over 5% in 2020 season (Table S2). Grain hardness measured by SKHI showed a narrow range (79.6-99.8), indicative of very hard grain typical of durum wheat. The low SCR observed in all plots is very much related to the sound TW (78-79.2 kg/hl) and high TGW achieved (43.7-49 g) in 2021. TW was not influenced by either Nrate or FCR whereas in 2020, FCR reduced TW significantly although only marginally (in Jandaroi only), still meeting the minimum level of 76 kg/hl (Figure 4). Higher TGW was obtained for both varieties in 2021 compared to 2020 (Figure 3) with no effect of Nrate in Jandaroi and a slight increase for DBA Lillaroi at the highest Nrate compared to a decline in 2020 except for the blip at 150 kg/ha. DBA Lillaroi had larger grain than Jandaroi as noted in the 2020 season.
While grain dimensions and low SCR level in 2021 indicate good grain quality, another important trait for durum and market grading is the percentage of hard vitreous kernels (HVK). The HVK was quite variable ranging from 53.8-98.2 with both varieties displaying higher HVK with increased applied N and not affected by FCR infection. Low N <20 kg/ha reduced HVK in DBA Lillaroi to a low level well below the market cut-off of 70% but never in Jandaroi even with nil N (Figure 11A). At 160N both varieties achieved similar HVK but at all other N rates, DBA Lillaroi had lower HVK than Jandaroi. Grain TGW, TW, SCR, SKHI and HVK can all impact milling potential yields. The SY% values for DBA Lillaroi were all higher than for Jandaroi at all N application rates (Figure 5), but this was not so for Semo only at all Nrates where values between varieties were closer in 2021 only (Figure 6). SY values were higher in 2020 than in 2021. In 2020, increasing N had no impact on SY but increased SY in 2021 (Figure 5). The highest SY was at 40N with no further increase as Nrate increased to 160N for both varieties.
For semolina colour there was a tendency for semolina L* to decrease with Nrate while genotype and disease were not significant (data not shown). However, only up to 1.3 units were lost which is not considered important enough for pasta production, the true indicator of brightness not the semolina which is more predictive having different particle size and shape. More important is semolina b* with significant genotype and Nrate effects. DBA Lillaroi achieved a higher semolina b* than Jandaroi at all Nrates as was found in 2020 (Figure 7). In 2021 there was a significant increase in semo-b* in DBA Lillaroi with increasing rates of applied N from as low as 20 kgN/ha and continued to increase reaching highest levels at 160 kgN/ha N. For Jandaroi 20N increased semo-b* with no further increases with more applied N (Figure 7). This is a desirable trait but needs to be balanced by the slight decrease in semo-L*.
As observed in 2020, Jandaroi showed much stronger dough properties than DBA Lillaroi (longer MPT, lower RBD, higher GI). In 2020 there was minimal change in dough strength with N rate (RBD and GI). However, in 2021 Nrate led to a decline in GI in DBA Lillaroi but not Jandaroi (Figure 8) and an increase in RBD for both varieties (Figure 9). Jandaroi typically had a very high GI staying above 90 at all N rates and was unaffected by FCR infection (Figure S1). However, in 2020 both varieties were impacted by FCR with reduced GI not seen in 2021 likely because there was little to no FCR expression in this season (Figure S1). The much larger decline in GI in DBA Lillaroi was evident in 2021 with a more restrained fall in GI in the 2020 season with Nrate. DBA Lillaroi, while having high GI 90+ at 0 kgN/ha and up to 40 kgN/ha application rate, its GI tended to decline with increasing rates of applied N reaching lowest levels at 160 kgN/ha application rate with a GI ~70-80, which is still an acceptable level. Mixograph RBD hardly changed with Nrate in 2020 for both varieties but in 2021, both showed large increases in RBD (indicative of less stable dough) in line with the declining GI with increasing Nrate (Figure 9) although there was a larger decline in GI for DBA Lillaroi but little difference in RBD between the varieties with increasing N rate. While FCR had a significant effect on RBD only in 2020 and especially for DBA Lillaroi (data not shown). Lack of FCR impact in 2021 is consistent with lack of expression not infection in 2021 (Table S5). The season was too wet and temperatures mild during grain filling which limited the expression of whiteheads and hence yield loss and many quality traits were unaffected from FCR infection due to these climatic conditions. While in 2020 there was infection and expression with significant effects of variety, Nrate and a Disease x Nrate interaction from the ANOVA (Table S6).
3.3. Protein compositional changes (2021 season)
The protein composition of the mature grain samples can provide some explanation for observed effects of the three treatments based on understanding of the role of gluten proteins in grain quality. Only data from the 2021 season was examined by HPLC. As the protein content of the sample increased, the total area of each peak also increased (Table S7). Looking for trends in the data with variety, Nrate and FCR infection reveals significant variety effects for all six HPLC peak areas and for three derived parameters except SolC, while for Nrate the effect was only significant for SolB, InsolA, InsolA, InsolB and Gli/Glu with no significant effect of FCR inoculation and no interactions (Table S7). Higher SolB mean values were found in DBA Lillaroi at the same corresponding Nrate than Jandaroi (Figure 12A). Only the higher N application led to a significant increase in the gliadin to glutenin ratio changing the protein composition of the grain. For insolA while some significant differences were apparent with Nrate, there was no consistent trend (Table S7). Overall mean Gli/Glu for DBA Lillaroi (0.806) was significantly higher than Jandaroi (0.777) but Jandaroi showed more response to Nrate plateauing at 80N increasing from 0-160N by 8.1% compared to DBA Lillaroi of 5.1% (Figure 12B). A correlation analysis shows significant (p <0.05) and strong correlations (defined as r ~ 0.7-0.9) between Gli/Glu and L* (-0.72), MPT (-0.74), RBD (0.78) and GI (-0.70) and between SolA% and L* (-0.78), MPT (-0.88), RBD (0.87) and GI (-0.80) and for InsolA and TGW (-0.82), SY (-0.94), b* (-0.92), GI (0.72). Low correlations (r ~ 0.3-0.7) were obtained between HVK and all these three HPLC parameters.
Principal component analysis (PCA) enables insight into the presence of patterns and relationships in available data by providing information of defined variables, which behave similarly to each other. Traits showing simple correlations were analysed and separated by Nrate (Figure 15 A, C) or variety (Figure 13 B, D). The first two principal components accounted for 90.73% or 98.79% of the variability of the data, respectively. For example, in Figure 15A, MPT was negatively correlated to RBD while GP, Yield and WG were strongly positively correlated and higher values for GP, Yield, WG, HVK were associated with higher Nrates. For varieties these same traits (Figure 15B) show that Jandaroi data points cluster to higher values for HVK, GI. For milling traits, SY, Semo only and TGW were all correlated (Figure 15 C) and for variety split, higher milling and grain weight values were evident for DBA Lillaroi over Jandaroi (Figure 13 D).
4. Discussion
Increased nitrogen (N) inputs are required to achieve higher grain protein levels desirable for obtaining premium durum wheat grades in Australia. Unfortunately, higher N inputs can exacerbate infection and yield loss from FCR [8,9], which is prevalent across the northern grain region of Northern New South Wales [6]. The two seasons were unseasonably wet compared to the long term average rainfall (Table 1) but with important differences between the seasons during the critical grain filling period, September-November in this environment. Total rainfall in 2020 during this period was 103.4 mm with a higher average maximum temperature of 27.4°C showing that 2020 was a drier and hotter season compared to 2021 with continued rainfall during grain-filling and mild temperatures (rainfall 279.8 mm and 24.5 °C). This would help to explain why there was more impact on grain yield and some quality measures from FCR inoculation in 2020 and not in 2021. In 2020, high residual soil N levels reduced N responsiveness with limited impacts on durum quality. Potentially background soil N levels following extended mineralisation during drought years (2019 and earlier) limited the impact of N treatments in 2020, as seen with yield and GP, which consequently limited impact on grain protein synthesis (lay down of glutenin and gliadin) that impacts grain structure and hence vitreosity (no change, very high). In 2021, the field experimental site was preceded by a sorghum crop which reduced soil N levels and resulted in significant GP and yield responses to varying N application rates and consequent effects on grain quality and gluten protein composition. Over the two years of this project we did not have the combination of low starting soil N levels to drive strong responsiveness to N application rates and a drier/hotter conditions during grain-fill to favour FCR expression following inoculation. A third season (2022) would have been desirable but as it turned out, due to a persistent La Niña, wet and mild conditions continued in 2022 (September-November 389.8 mm rainfall and average maximum temperature of 23.3°C) that would have also limited FCR expression at this location.
4.1. Comparison of seasonal impact on yield and quality
Grain yield in 2020 showed no significant difference between variety and Nrate but FCR infection impacted GY in both varieties reducing it with the higher dose of inoculum having more impact in most situations. In the 2021 season higher yields were obtained (overall mean 2020, 2.0 t/ha; 2021, 4.8 t/ha) with much more responsiveness to applied N in both varieties but no significant effect from FCR inoculation. Yield increases were similar between varieties in 2021. Grain yield is responsive to applied N fertiliser in durum wheat as N fertilization is largely considered to be the main factor affecting yield and protein content [15] where the N is converted into protein and biomass. This was reflected in the GP responses being blunted in 2020 (range 1.04%) compared to more responsive at all N rates in 2021 (range 2.4%), especially between 40-160N. These increases had expected effects on technological quality traits. Nitrogen fertilization contributes to increases in protein content when fertilizer rates satisfy the requirements of both yield and protein synthesis [16,17]. Indeed, in 2021 traits significantly affected by Nrate were GY, GP, WG, TW, TGW, HVK, SKHI, SY, Semo only, L*, b*, MPT, RBD, MPH and GI compared to 2020 where only GP, WG, TGW, b* and Semo only were significantly affected by Nrate. In 2021 both varieties responded similarly to applied N with DBA Lillaroi starting GP lower than Jandaroi. Response was larger at 80N and above compared to lower rates. If the soil N at sowing is high then the plant relies less on the applied N to fill the grain with protein. GP content obtained in the mature grain depends on other factors such as water and N availability, length of grain filling, weather conditions prevailing during grain filling (like higher rainfall, lower temperatures prolonging flowering and grain filling period), partitioning of N and dry matter to the grain and source-sink relations with leaves and stems [18]. An inverse relationship between grain yield and grain N concentration has been reported in durum wheat [19,20,21,22,23]. Against this trend was a strong positive correlation between GY and GP in 2021 (r = 0.84, p<0.05) but not in 2020. This relationship can be affected by soil fertility, water availability etc. [18]. The combination of above average rainfall and high rates of N application satisfying the requirements for both yield and protein synthesis could explain this positive correlation in 2021 [16]. Based on the GP responses, both varieties appear to have similar abilities to transfer the applied N to more grain protein. Grains were smaller in 2020 than in 2021 (mean TGW 2020, 41.8 g compared to 2021, 46.4 g) and this could also result in the higher protein in that season (mean 0N GP 13.8% in 2020 vs. 12.7% at 160N in 2021).
Genetic differences in grain weight were evident between the two varieties and DBA Lillaroi was selected during its breeding to achieve high TGW and milling potential and the data supports this as do other studies [24,25]. The overall TGW achieved are typical for durum wheat grown in the NNSW region under non-drought situations as was the case in the two seasons of this study. Jandaroi TGW did not respond to Nrate while DBA Lillaroi tended to decrease but was not consistent in 2020 and hardly changed in 2021. FCR had no impact on TW in both varieties in 2021 because there was very little FCR expression despite the infection from the inoculation but reduced TW for Jandaroi in 2020 significantly from 78.7 kg/hl to 77.6 kg/hl. This is not a large change and of no commercial significance as these levels are above the trading discount of 76 kg/hl. There was some evidence of weather damage in the visual appearance of the kernels especially in 2021 where much lower HVK was found. Rainfall at or near harvest (1st December 2021) of 191 mm in November (Table 1) could have been responsible for the low falling numbers from 185-290 sec in 2021 that was not found in the 2020 season with only 3 mm of rain near harvest, 24th November, 2020 (mean FN 571 sec). While this is below the minimum GTA trading value of 300 sec required for acceptable grain [3], evidence from the literature suggests only if weather damage is severe in durum wheat (FN < 100 sec or thereabouts), is there any influence on pasta colour and cooking quality [27].
The percentage of kernels that are vitreous is an important grade determinant for durum wheat because of its impact on semolina milling yield and end-product quality. Durum wheat with high HVK (>80) and test weight will generally give high semolina yield with a minimum number of white starchy particles [27]. According to the grading standards in Australia [3], durum is graded as, ADRI HVK >80%; ADR2 HVK 70-80%; ADR3 <70% which means that despite what the grain protein is, grain could be downgraded at grain receival centres from low HVK with consequent financial penalties for growers. Grain vitreousness was affected in 2021 but not in 2020 where HVK average values >95% (data not shown) probably because protein was >13% and unresponsive to applied N. In 2021, the increase in N fertilisation led to an increase in HVK which is mostly likely due to the increase in GP that has been found by others [28,29]. High grain protein content allows the formation of a compact grain structure due to the starch granules in the amyloplasts surrounded by the protein matrix and this creates reduced air spaces in the grain structure resulting in a vitreous or “glassy” appearance, i.e. high HVK score. In contrast, a less compact grain structure has many open spaces, with a lower density endosperm [28] and the protein matrix is discontinuous creating opaque appearance due to light diffraction and diffusion at the void spaces [28]. There is generally a strong correlation between GP and HVK as observed in the 2021 data (GP vs. HVK, r = 0.81, P<0.001, Fig 11B) in line with other reports [2,26]. This relationship differed between Jandaroi and DBA Lillaroi with the former variety showing a more gradual change and holding the HVK mostly above 75% even at 9-10% GP, whereas DBA Lillaroi slope was larger with HVK<80% as GP declined below 11.5% reaching values as low as 42%. Given that HVK>80 ensures top grade (ADR1) in Australia, Jandaroi for most field plots met this requirement whereas DBA Lillaroi did not especially at Nrates below 80 kg/ha. Even with no applied N, many Jandaroi samples achieved HVK>80% whereas for DBA Lillaroi, the applied N was found to be critical in achieving the highest grade and needed 80N minimum, with downgrading to the lowest grade at ≤20N. This would strongly impact the price a grower would receive. Jandaroi has more potential to achieve a higher GP than DBA Lillaroi in the NNSW environment although quite often lower in yield [25]. So despite the known relationship between HVK and GP, this was not observed for Jandaroi since even at very low GP 9-11%, the HVK was mostly >75%. It has been noted by others that this strong association between protein and HVK is inconsistent [2] and Sieber et al. [30] found large variation in grain protein in non-vitreous kernels. A possible explanation is related to the protein composition and how that influences grain structure. At the same Gli/Glu ratio range of 0.65-0.79, Jandaroi had mean HVK 89 ± 8.3% and GP 11.4 ± 1% while DBA Lillaroi had mean HVK 70 ± 14.6% with a similar GP 10.8 ± 1.1%, showing that despite very similar GP and Gli/Glu between the varieties, the grain structure in Jandaroi must be quite different to DBA Lillaroi to account for this large difference in HVK, so other factors are operating. The question then is why is Jandaroi able to hold its HVK so high at low protein? Samson et al. [28] found variety Néodur was able to synthesise protein more efficiently from applied N than other varieties examined (like the case for Jandaroi vs. DBA Lillaroi in this study) and showed the highest density kernels, consistent with high HVK. We found that as HVK increased the gliadin to glutenin ratio increased in both durum varieties (Figure 14). This result is consistent with other studies showing preferential accumulation of gliadin in vitreous kernels [2,26]. Samson et al. [28] suggested that gliadin proteins facilitate the formation of vitreous endosperm by providing better adhesion of the protein matrix to starch granules during kernel desiccation. Our data suggests this would at least have some dependence on the genotype but none of the above studies examined the relationship between HVK, GP and Glu/Gli in multiple genotypes. Further analysis of the glutenin proteins during grain maturation should confirm if Jandaroi does synthesise gliadins faster during grain development than DBA Lillaroi and this may assist the formation of a “tighter grain structure” which translates into higher mature HVK levels in this variety irrespective of grain protein content making Jandaroi a better variety when soil N levels could be low but more studies are needed to confirm this.
Grain weight, TW and vitreousness can contribute to milling yield and purity (ash content and bran specks in semolina) that can impact pasta appearance and colour [31]. For the trait Semo only in 2020 there was a tendency for this to decline with increased N but only significantly for DBA Lillaroi (Figure 6). However, in 2021 there was a much greater change (increase) in Semo only which was very clear for DBA Lillaroi, increasing significantly from 20N peaking at 80N while Jandaroi showed little response to applied N (Figure 6). This was less apparent for trait SY% with smaller increases with applied N, also noting no change in SY% for both varieties in 2020 (Figure 5). The increase in Semo only in DBA Lillaroi in 2021 could be due to a good response from applied N for HVK and less so for TGW that was not the case for Jandaroi. It seems that DBA Lillaroi has more potential to improve its milling potential than Jandaroi possibly by translating the N into more starch synthesis through higher starch synthase activity or improving the milling potential of this variety by changes in endosperm to bran ratio or the separability of the bran or other factors [32]. However, further testing across more environments is needed to confirm this response which is very interesting showing potential to manipulate milling potential with applied N that is genotype dependent. Interestingly, despite DBA Lillaroi at Nrates <80 kg/ha (2021 data only) having HVK<75, the Semo only and SY% were still superior to Jandaroi and even at 0N DBA Lillaroi has higher milling yield than Jandaroi receiving 160N (for SY% only). It was noted by Fu et al., [2] that low protein durum can have acceptable milling yield even with HVK<70%. Other factors come into play in particular genetic factors [25]. DBA Lillaroi genetic tendency to produce high TGW would contribute to higher semolina yield than Jandaroi and not so much, if at all, from HVK contributions. In particular the N response seen in TGW in DBA Lillaroi and not in Jandaroi would help in achieving a higher semolina yield. Fu et al. [2] found using composite durum wheat samples varying in GP from 14.5 to 9.9% that providing HVK>70%, the semolina yield did not change with GP content but semolina yield would decline significantly if low HVK coincided with low grain protein. This could explain why semolina yield in Jandaroi did not respond to N fertilisation (even at GP ~10.5% with 0N) because mean HVK>80%. Whereas for DBA Lillaroi at N<80 kg/ha where HVK was <75% and GP<11.5% that this combination resulted in the grain producing significantly less semolina on milling, as observed. In 2020 season both GP >13% and HVK >96% were high which could explain the lack of SY response to applied N in that season. High HVK confers superior milling performance and high wheat protein can mitigate the negative impact of low HVK on durum milling by reducing the size of the starchy areas in the piebald kernels [2].
Semolina colour is an important characteristic in durum because high semolina yellowness tends to translate to high pasta yellowness although during milling and pasta processing loss of colour can occur [33]. Higher protein semolina is known to be duller (lower L*) as protein affects brightness/redness of semolina [34] and the semolina L* decreased with increasing N rate (data not shown). High HVK grain can negate the effect of duller semolina due to high grain protein resulting in the final pasta still remaining a bright yellow colour [2]. DBA Lillaroi has a higher semolina b* than Jandaroi because this variety has a higher natural yellow pigment level in the endosperm, as a result of years of breeding and selection to enhance semolina b* levels [25]. While there was no significant changes in semolina b* in 2020, in 2021 there was a significant increase for DBA Lillaroi but not Jandaroi with increasing N. This is probably related to the increase in GP, as Dalla Marta et al. [35] reported an association between protein content increase and increase in yellow pigment content.
Jandaroi was selected in its breeding for its high dough strength reflected in the MPT (3-5 min), low RBD (<50) and very high GI (>90) superior to DBA Lillaroi [25]. While there was little effect of Nrate on GI in the 2020 season for both varieties, in 2021 increasing N application reduced GI slightly in Jandaroi (95 to 85) but to a much larger extent for DBA Lillaroi (92 to 76). The FCR inoculum however significantly reduced GI in both varieties in 2020 but had no effect in 2021, (Figure S1). The season was too wet and temperatures mild during grain filling which limited the expression of whiteheads and hence yield loss and many quality traits being unaffected from FCR infection in 2021. The RBD in both varieties in 2021 increased with increasing N in both varieties (Jandaroi from 27 to 46; DBA Lillaroi from 27 to 53) but not in 2020. These data indicate a weaking of dough strength with increasing N application consistent with the decrease in GI. The reason for the seasonal differences and effect of N is complex and may depend on other conditions like water availability and other environmental changes as a genotype’s dough properties are affected by both genotypic and environmental factors [24,36]. Giuliani et al., [18] found in their N application study in durum wheat that while GI was affected by N in one year, there was no change in another season. Other studies in wheat have shown a decrease in gluten strength with N fertilization in hard white winter wheats due to a preferential increase of gliadins over glutenins [37]. The negative influence of a higher Gli/Glu ratio on common wheat dough rheological properties is well known [38,39]. Increasing N led to an increase in the Gli/Glu ratio in both varieties with different responses between varieties (Figure 12B). The regression of Gli/Glu on RBD and GI (Figure 15) show that as gliadin percentage increases the dough becomes weaker. In Jandaroi the increase in Gli/Glu did not result in much change in GI whereas in DBA Lillaroi there was a much stronger relationship suggesting that changes in Jandaroi Gli/Glu are much less likely to result in large shifts in GI compared to DBA Lillaroi supporting the findings of Saint Pierre et al. [37] but with qualification. This relationship is more complex and related to genetic differences in varieties ability to retain gluten strength despite changing protein composition. However, both varieties showed a similar change in RBD, increasing with an increase in Gli/Glu but that did not translate to much reduction in GI for Jandaroi. While mixograph parameters and the gluten index tests are correlated, they measure different aspects of dough properties because they are fundamentally different methods to evaluate dough [40]. Horvat et al., [41] showed in common wheat that increasing nitrogen level led to higher grain protein content with changes in the gliadin to glutenin ratio to a lesser extent which was variety specific.
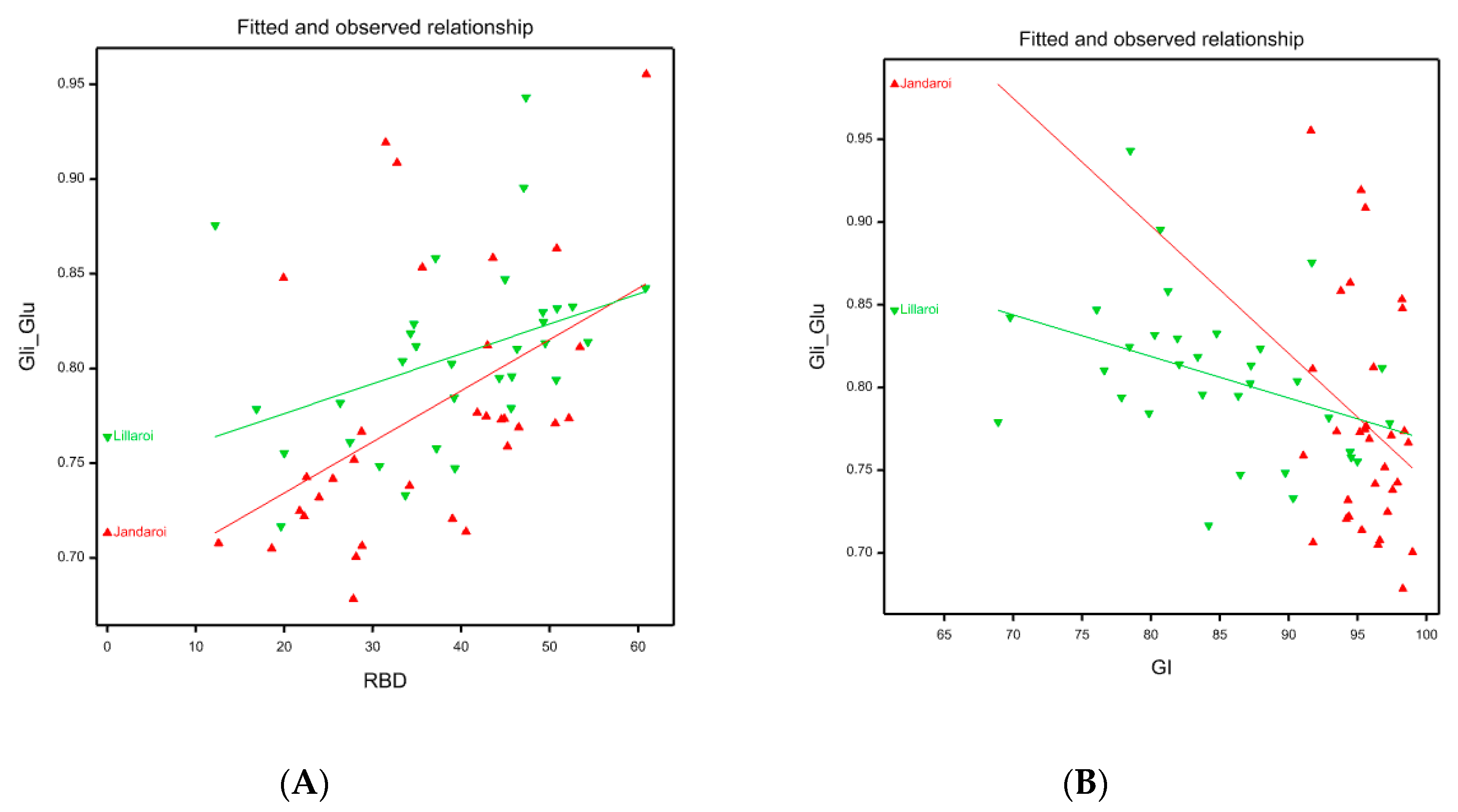
Figure 15.
Linear models showing fitted values (line) and observed (triangles) between Gli/Glu ratio and RBD (A) and between Gli/Glu ratio and GI(B) according to variety.
Figure 15.
Linear models showing fitted values (line) and observed (triangles) between Gli/Glu ratio and RBD (A) and between Gli/Glu ratio and GI(B) according to variety.
5. Conclusions
This study was performed to illustrate the impact of fusarium crown rot (FCR) infection and its interaction with nitrogen fertiliser application levels at sowing on grain yield and quality. However, cooler and wetter conditions in 2021 prevented the expression of FCR despite inoculation of plants and due to soil management in the summer of 2021 prior to sowing, good nitrogen responses in terms of grain yield and grain protein content were obtained with consequent effects on grain and semolina traits. Jandaroi was found to hold its kernel vitreosity at all protein levels allowing a good chance of a grower obtaining the premium grade while DBA Lillaroi did not and would suffer downgrade if nitrogen application was insufficient. However, higher nitrogen application needed to achieve 12% + protein can reduce dough strength under some conditions, as in 2021 but Jandaroi held its dough strength much better than DBA Lillaroi. While dough strength is not reflected in grain trading standards, it can impact pasta processing and pasta quality so a variety that can hold this trait is important to pasta processors. The protein composition of the two varieties response to nitrogen fertilisation differed and it was proposed that Jandaroi tendency to a lower Gli/Glu ratio in response to nitrogen helps to hold a high grain vitreousness and high dough strength. This suggests genetics plays an important role in a genotype response to nitrogen fertilisation and should be considered when selecting a variety where high grade is desirable. The potential of nitrogen application to increase the milling yield found in the 2021 season especially in DBA Lillaroi should be explored and the mechanism for this effect determined as potential to increase value to the miller. DBA Lillaroi genetic potential for higher grain weight through breeding is an important factor. Further validation of these findings is needed over drier and warmer grain filling periods which favours the expression of FCR to better understand the relationships between genotype, Nrate and FCR.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1. Means and LSD for Wet Gluten for both varieties at different N application rates at sowing for both seasons. Table S2. Means and LSD for screenings for both varieties at different N application rates at sowing for both seasons. Table S3. Means and LSD for SY for both varieties at different disease application rates - Tamworth 2020. Table S4. Means and LSD for GI, MPT and RBD for both varieties at different inoculation application . Table S5. Mean SE-HLPC data for two durum wheat varieties at 4N rates and 2 CR inoculations in mature grain from 2021 season. Figure S1. Boxplot showing GI for both varieties at different CR inoculation rates (g/m) for 2020 and 2021 seasons. LSD statistic shown for each year.
Author Contributions
Conceptualization, M.S. and S.S.; methodology, M.S., N.E. and S.S.; formal analysis, M.S. ; investigation, M.S. N.E.; resources, M.S. and S.S; writing—original draft preparation, M.S.; writing—review and editing, N.E. and S.S.; supervision, M.S. and S.S.; project administration, S.S.; funding acquisition, M.S. and S.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded under the Grains Agronomy and Pathology Partnership between New South Wales Department of Primary.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Please contact the corresponding author with requests for data.
Acknowledgments
Would like to acknowledge the contributions of technical staff at New South Wales Department of Primary Industries for their technical support for the project.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Richards, R.A.; Rebetzke, G.J.; Condon, A.G.; van Herwaarden, A.F. Breeding opportunities for increasing the efficiency of water use and crop yield in temperate cereals. Crop Science 2002, 42, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Fu, B.X.; Wang, K.; Dupuis, B.; Taylor, D.; Nam, S. Kernel vitreousness and protein content: Relationship, interaction and synergistic effects on durum wheat quality. Journal of Cereal Science 2018, 79, 210–217. [Google Scholar] [CrossRef]
- Anonymous. (2020). Grain Trade Australia Wheat Standards 2020/21. http://www.graintrade.org.au/sites/default/files/WHEAT%20STANDARDS%202020-2021%20All%20Grades%20updated %20061120.pdf. Accessed 22 November 2022. 22 November.
- Herbert, A. An international Benchmarking Comparison of Australian Crop production and Profitability, in GRDC Updates, GRDC, Editor. 2017: Perth.
- Chakraborty, S.; Liu, C.L.; Mitter, V.; Scott, J.B.; Akinsanmi, O.A.; Ali, S.; Dill-Macky, R.; Backhouse, D.; Simpfendorfer, S. Pathogen population structure and epidemiology are keys to wheat crown rot and Fusarium head blight management. Australasian Plant Pathology 2006, 35, 643–655. [Google Scholar] [CrossRef]
- Simpfendorfer S, McKay A, Ophel-Keller K (2019). New approaches to crop disease management in conservation agriculture. In (Eds J Pratley and J Kirkegaard) “Australian Agriculture in 2020: From Conservation to Automation” pp 173-188 (Agronomy Australia and Charles Sturt University: Wagga Wagga).
- Alahmad, S.; Simpfendorfer, S.; Bentley, A.R.; Hickey, L.T. Crown rot of wheat in Australia: Fusarium pseudograminearum taxonomy, population biology and disease management. Australas. Plant Pathol. 2018, 47, 285–299. [Google Scholar] [CrossRef]
- Davis, R.A.; Huggins, D.R.; Cook, J.R.; Paulitz, T.C. Nitrogen and crop rotation effects on fusarium crown rot in no-till spring wheat. Canadian Journal of Plant Pathology, 2009, 31, 456–467. [Google Scholar] [CrossRef]
- Buster, M.; Simpfendorfer, S.; Guppy, C.; Sissons, M.; Flavel, R.J. Interactions of Fusarium Crown Rot of Wheat with Nitrogen. Plants 2023, 12, 533. [Google Scholar] [CrossRef]
- Hughes, N.; Galeano, D.; Hatfield-Dodds, S. The effects of drought and climate variability on Australian farms, ABARES insights report 2019.
- Zörb, C.; Ludewig, U.; Hawkesford, M.J. Perspective on Wheat Yield and Quality with Reduced Nitrogen Supply. Trends in Plant Science 2018, 23, 1029–1037. [Google Scholar] [CrossRef]
- Forknall, C.R.; Simpfendorfer, S.; Kelly, A.M. Using yield response curves to measure variation in the tolerance and resistance of wheat cultivars to Fusarium crown rot. Phytopathology 2019, 109, 932–941. [Google Scholar] [CrossRef]
- Sissons, M.; Ovenden, B.; Adorada, D.; Milgate, A. Durum wheat quality in high input irrigation systems in south-eastern Australia. Crop & Pasture Science 2014, 65, 411–422. [Google Scholar]
- Sissons, M.J.; Batey, I.L. Protein and Starch Properties of some Tetraploid Wheats. Cereal Chemistry 2003, 80, 468–475. [Google Scholar] [CrossRef]
- Fuertes-Mendizabal, T.A.; Aizpurua, M.B.; Gonzalez-Moro, J.M.; Estavillo, J.M. Improving wheat breadmaking quality by splitting the N fertilizer rate. Eur. J. Agron. 2010, 33, 52–61. [Google Scholar] [CrossRef]
- Johansson, V.A.; Mattern, P.J. Wheat, rye, and triticale. In Nutritional quality of cereal grains Olson, R.A.; Frey, K.J. Eds.; Agron. Monogr. 1987 28. ASA, CSSA, and SSSA, Madison, WI; pp. 133–182.
- Reznick, J.P.K.; Barth, G.; Kaschuk, G.; Pauletti, V. Nitrogen and cultivars as field strategies to improve the nutritional status of wheat grain and flour. Journal of Cereal Science 2021, 102, 103290. [Google Scholar] [CrossRef]
- Giuliani, M.M.; Giuzio, L.; De Caro, A.; Flagella, Z. Relationships between nitrogen utilization and grain technological quality in durum wheat: II. Grain yield and quality. Agronomy Journal 2011, 103, 1668–1675. [Google Scholar]
- Ehdaie, B.; Waines, J.G. Sowing date and nitrogen rate effects on dry matter and nitrogen partitioning in bread and durum wheat. Field Crops Res. 2001, 73, 47–61. [Google Scholar] [CrossRef]
- Martre, P.; Porter, J.R.; Jamieson, P.D.; Triboi, E. Modelling grain nitrogen accumulation and protein composition to understand the sink/source regulation of nitrogen remobilization for wheat. Plant Physiol. 2003, 133, 1959–1967. [Google Scholar] [CrossRef] [PubMed]
- Clarke, J.M.; Clarke, F.R.; Pozniak, C.J. Forty-six years of genetic improvement in Canadian durum wheat cultivars. Canadian Journal of Plant Science 2010, 90, 791–801. [Google Scholar] [CrossRef]
- DePauw, R.M.; Knox, R.E.; Clarke, F.R.; Wang, H.; Fernandez, M.R.; Clarke, J.M.; McCaig, T.N. Shifting undesirable correlations. Euphytica 2007, 157, 409–415. [Google Scholar] [CrossRef]
- Motzo, R.; Fois, S.; Giunta, F. Relationship between grain yield and quality of durum wheats from different eras of breeding. Euphytica 2004, 140, 147–154. [Google Scholar] [CrossRef]
- Sissons, M.; Kadkol, G.; Taylor, J. Genotype by Environment effects on Durum Wheat Quality and Yield -Implications for Breeding. Crop Breed Genet Genom. 2020, 2, e2000XX. [Google Scholar] [CrossRef]
- Kadkol, G.; Sissons, M.; Lambert, N.; Lisle, C. Genetic improvement in grain yield and quality of Australian durum wheat over six decades of breeding. Cereal Chemistry 2022, 1–22. [Google Scholar] [CrossRef]
- Dexter, J.E.; D’Egidio, M.G. Grading factors impacting durum wheat processing quality. In Durum Wheat Chemistry and Technology, Sissons, M.S.; Abecassis, J.; Marchylo, B; Carcea, M. Eds.; 2nd edition, AACC International Press. 2012; pp. 235-250.
- Dexter, J.E.; Williams, P.C.; Edwards, N.M.; Martin, D.G. The relationships between durum wheat vitreousness, kernel hardness and processing quality. J. Cereal Sci. 1988, 7, 169–181. [Google Scholar] [CrossRef]
- Samson, M.-F.; Mabille, F.; Cheret, R.; Abecassis, J.; Morel, M.H. Mechanical and physicochemical characterization of vitreous and mealy durum wheat endosperm. Cereal Chemistry 2005, 82, 81–87. [Google Scholar] [CrossRef]
- Boukef, S.; Karmous, C.; Trifa, Y.; Rezgui, S. Durum Wheat Grain Quality Traits as Affected by Nitrogen Fertilization Sources under Mediterranean Rainfed Conditions. Journal of Agriculture and Sustainability 2013, 4, 99–114. [Google Scholar]
- Sieber, A.N; Würschum, T.; Longin, C.G.H. Vitreosity, its stability and relationship to protein content in durum wheat. Journal of Cereal Science 61 2015, 71–77. [Google Scholar] [CrossRef]
- Wang, K.; Taylor, D.; Chen, Y.; Suchy, J.; Fu, B.X. Effect of Kernel Size and Its Potential Interaction with Genotype on Key Quality Traits of Durum Wheat. Foods 2021, 10, 2992. [Google Scholar] [CrossRef] [PubMed]
- Sissons, M.; Abecassis, J.; Cubadda, R.; Marchylo, B. Methods used to assess and predict quality of durum wheat, semolina and pasta In: Durum wheat chemistry and technology, Sissons, M.S.; Abecassis, J.; Marchylo, B; Carcea, M. Eds.; 2nd edition, AACC International Press. 2012; pp. 213-234.
- Ficco, D.B.M.; Mastrangelo, A.M.; Trono, D.; Borrelli, G.M.; Vita, P.D.; Fares, C.; Beleggia, R.; Platani, C.; Papa, R. The colours of durum wheat: a review. Crop & Pasture Science, 2014, 65, 1–15. [Google Scholar] [CrossRef]
- Feillet, P.; Autran, J.C.; Icard-Verniere, C. Pasta brownness: an assessment. J. Cereal Sci. 2000, 32, 215–233. [Google Scholar] [CrossRef]
- Dalla Marta, A.D.; Grifoni, M.; Mancini, G.; Zipoli, S.; Orlandini, S. The influence of climate on durum wheat quality in Tuscany, central Italy. Int. J. Biometeorol. 2011, 55, 87–96. [Google Scholar] [CrossRef]
- Fois, S.; Schlichting, L.; Marchylo, B.; Dexter, J.; Motzoa, R.; Giuntaa, F. Environmental conditions affect semolina quality in durum wheat (Triticum turgidum ssp. durum L.) cultivars with different gluten strength and gluten protein composition. J. Sci. Food Agric. 2011, 91, 2664–73. [Google Scholar] [CrossRef]
- Saint Pierre, C.; Peterson, C.J.; Ross, A.S.; Ohm, J.B.; Verhoeven, M.C.; Larson, M.; Hoefer, B. Winter wheat cultivars under different levels of nitrogen and water stress: Changes in grain protein composition. J. Cereal Sci. 2008, 47, 407–416. [Google Scholar] [CrossRef]
- Barak, S.; Mudgil, D.; Khatkar, B.S. Influence of gliadin and glutenin fractions on rheological, pasting, and textural properties of dough. Int. J. Food. Prop. 2014, 17, 1428–1438. [Google Scholar] [CrossRef]
- Tang, J.W.; Liu, J.J.; Zhang, P.P.; Zhang, Y.; Xiao, Y.G.; Qu, Y.Y.; Zhang, Y.; He, Z.H. Effects of gluten protein fractions on dough property and products quality in common wheat. Sci. Agric. Sin. Sci. 2008, 41, 2937–2946. [Google Scholar]
- AbuHammad, W.A.; Elias, E.M.; Manthey, F.A.; Alamri, M.S.; Mergoum, M. A comparison of methods for assessing dough and gluten strength of durum wheat and their relationship to pasta cooking quality. International Journal Food Science Technology, 2012, 47, 2561–2673. [Google Scholar] [CrossRef]
- Horvat, D.; Šimi´c, G.; Dvojkovi´c, K.; Ivi´c, M.; Plavšin, I.; Novoselovi´c, D. Gluten Protein Compositional Changes in Response to Nitrogen Application Rate. Agronomy 2021, 11, 325. [Google Scholar] [CrossRef]
Figure 1.
Boxplot showing grain yield for both varieties at different N application (kg/ha) and CR inoculation (g/m) rates from 2020 season. Boxplots show median and interquartile range of trial data. LSD statistic shown as bar with value, p<0.05.
Figure 1.
Boxplot showing grain yield for both varieties at different N application (kg/ha) and CR inoculation (g/m) rates from 2020 season. Boxplots show median and interquartile range of trial data. LSD statistic shown as bar with value, p<0.05.
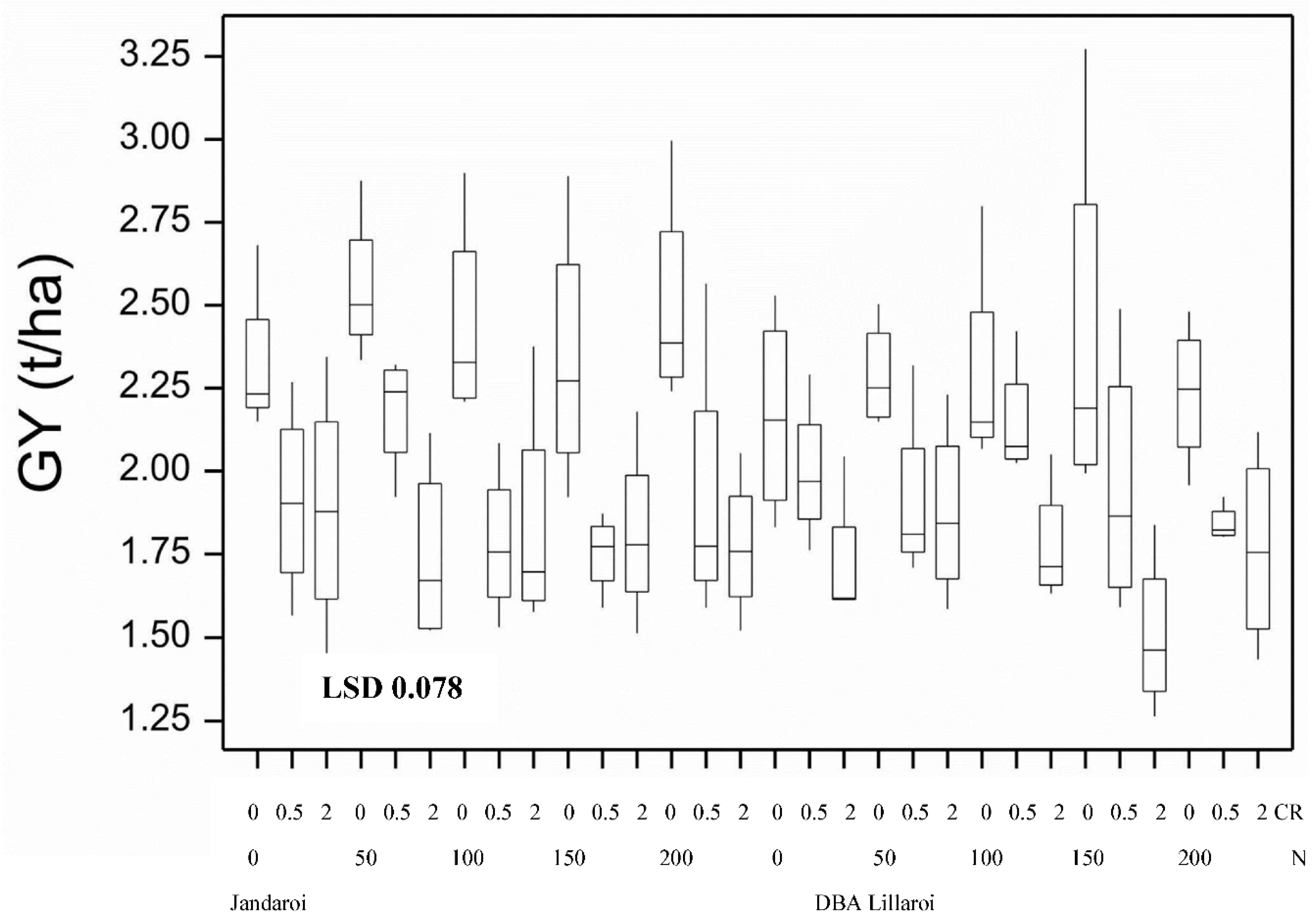
Figure 2.
Boxplot showing grain protein % (GP%) for both varieties at different N application rates at sowing for 2020 and 2021 seasons. J=Jandaroi; L=DBA Lillaroi; numbers are N kg/ha. Boxplots show median and interquartile range of trial data. LSD statistic shown for each year.
Figure 2.
Boxplot showing grain protein % (GP%) for both varieties at different N application rates at sowing for 2020 and 2021 seasons. J=Jandaroi; L=DBA Lillaroi; numbers are N kg/ha. Boxplots show median and interquartile range of trial data. LSD statistic shown for each year.
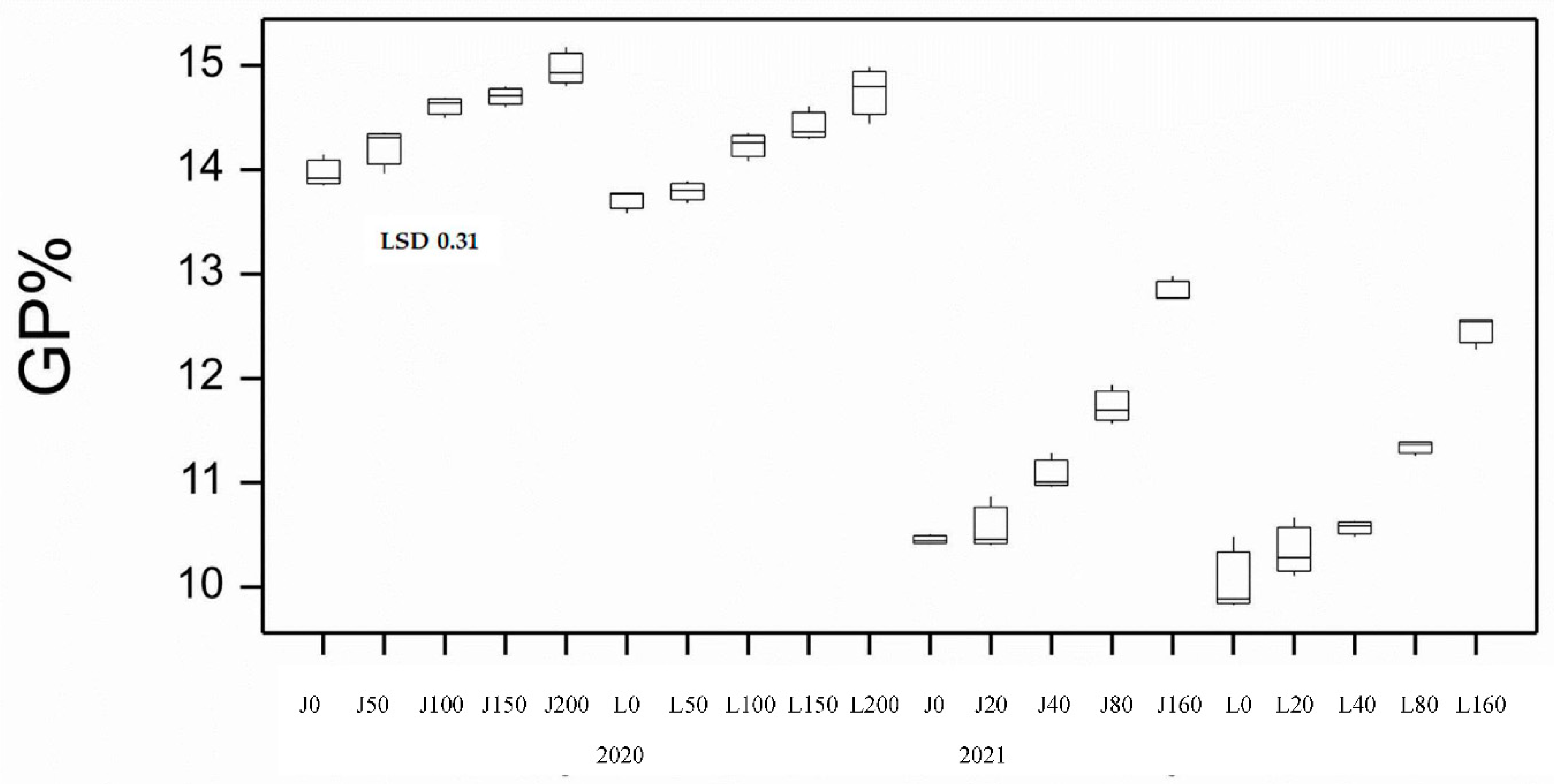
Figure 3.
Boxplot showing 1000 grain weight (TGW) for both varieties at different N application rates at sowing for 2020 and 2021 seasons. LSD statistic shown for each year.
Figure 3.
Boxplot showing 1000 grain weight (TGW) for both varieties at different N application rates at sowing for 2020 and 2021 seasons. LSD statistic shown for each year.
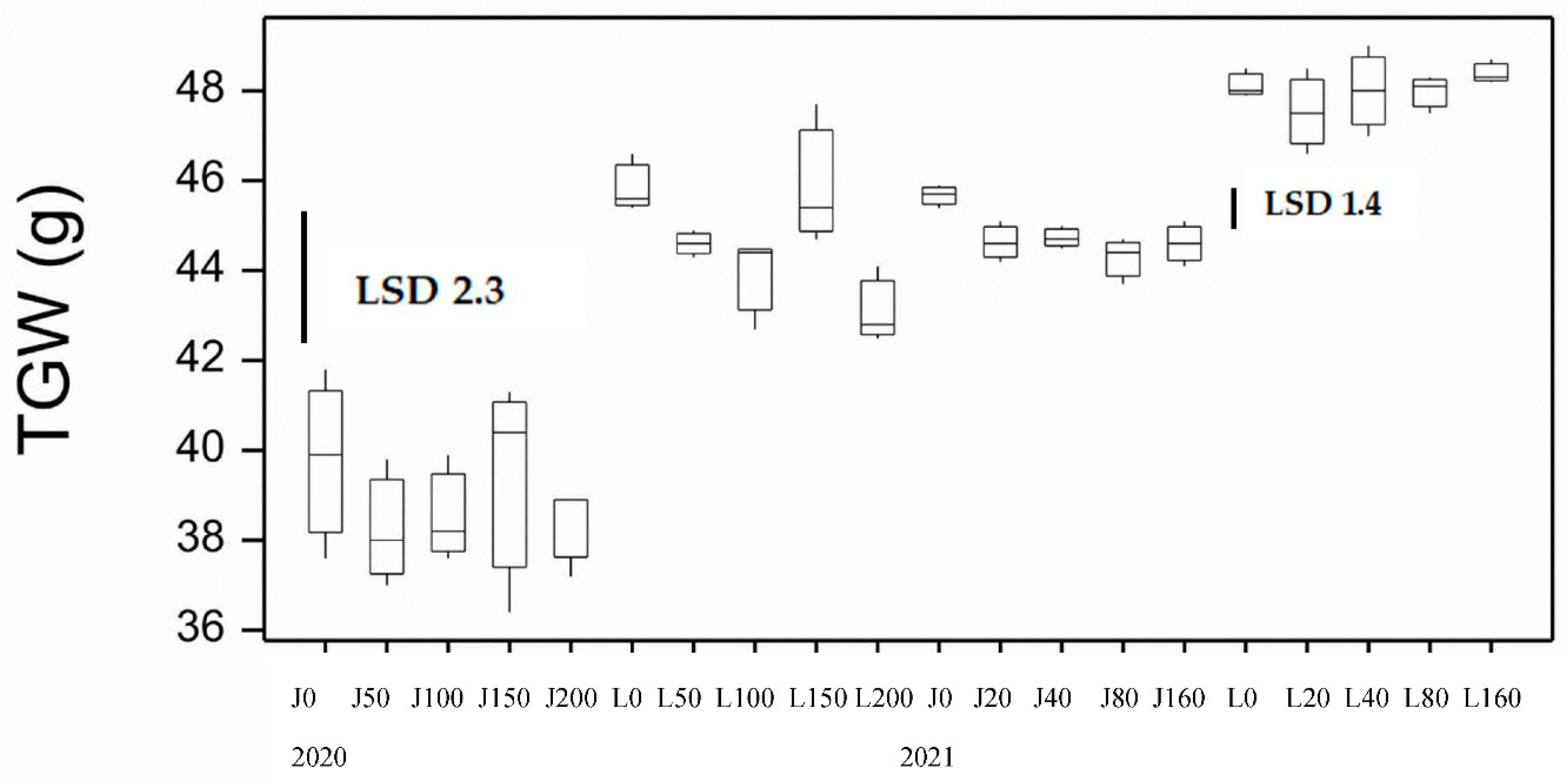
Figure 4.
Boxplot showing test weight (TW) for both varieties at different CR inoculation rates (g/m) for 2020 and 2021 seasons. LSD statistic shown for each year.
Figure 4.
Boxplot showing test weight (TW) for both varieties at different CR inoculation rates (g/m) for 2020 and 2021 seasons. LSD statistic shown for each year.
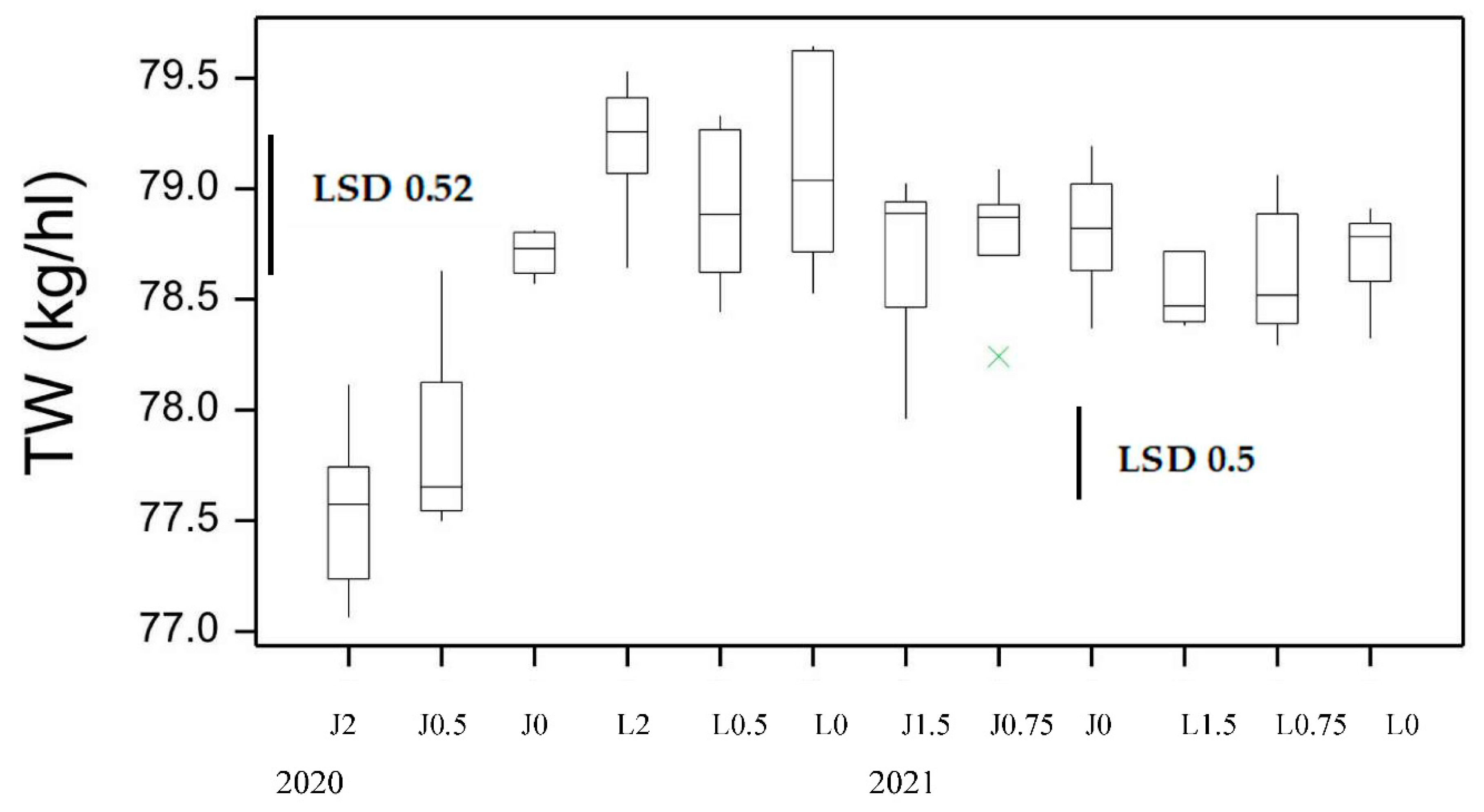
Figure 5.
Boxplot showing semolina yield % (SY) for both varieties at different N application rates at sowing for 2020 and 2021 seasons. LSD statistic shown for each year.
Figure 5.
Boxplot showing semolina yield % (SY) for both varieties at different N application rates at sowing for 2020 and 2021 seasons. LSD statistic shown for each year.
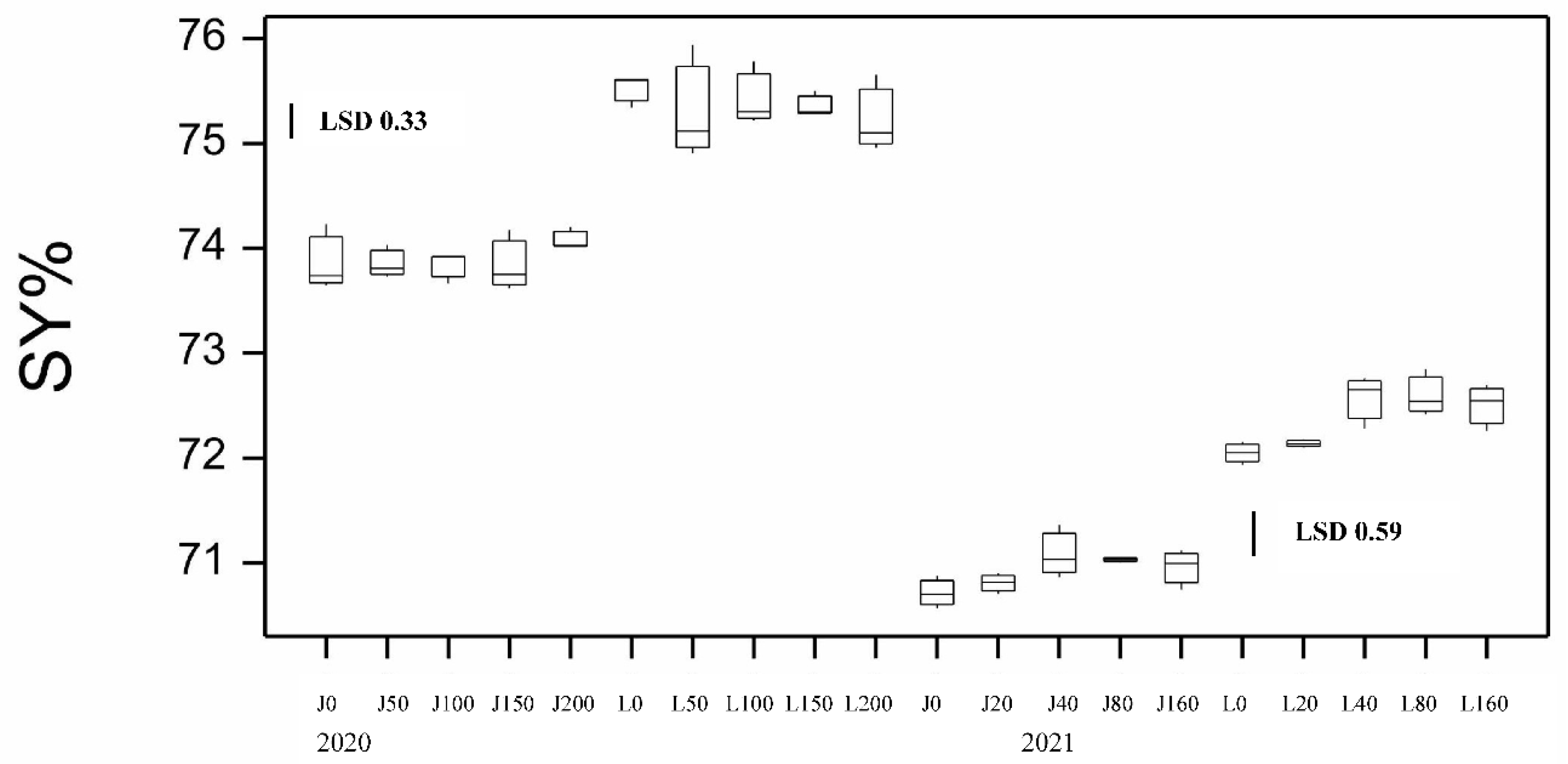
Figure 6.
Boxplot showing semolina only yield for both varieties at different N application rates at sowing for 2020 and 2021 seasons. LSD statistic shown for each year.
Figure 6.
Boxplot showing semolina only yield for both varieties at different N application rates at sowing for 2020 and 2021 seasons. LSD statistic shown for each year.
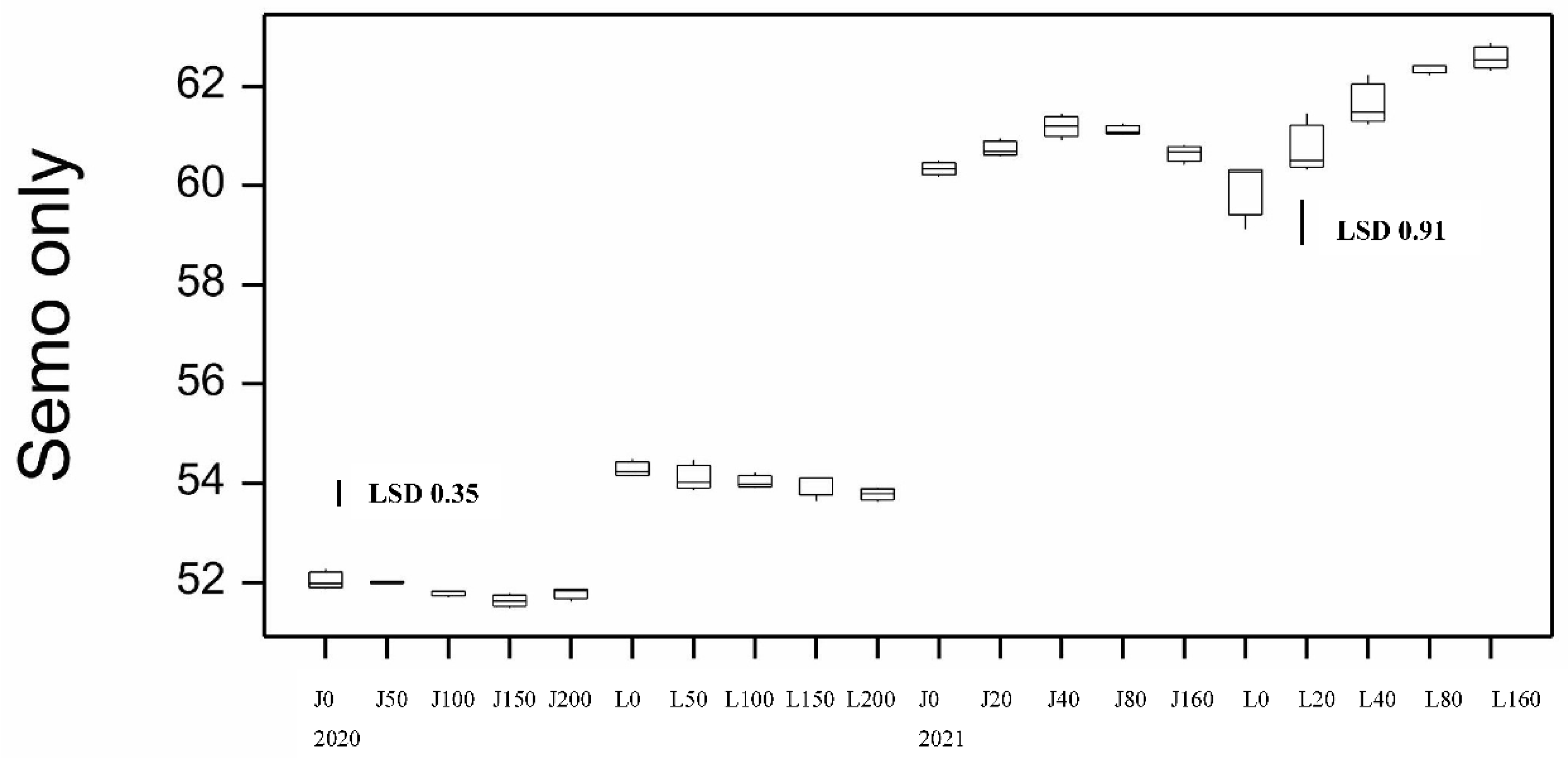
Figure 7.
Boxplot showing semolina yellowness (b*) for both varieties at different N application rates at sowing for 2020 and 2021 seasons. LSD statistic shown for each year.
Figure 7.
Boxplot showing semolina yellowness (b*) for both varieties at different N application rates at sowing for 2020 and 2021 seasons. LSD statistic shown for each year.
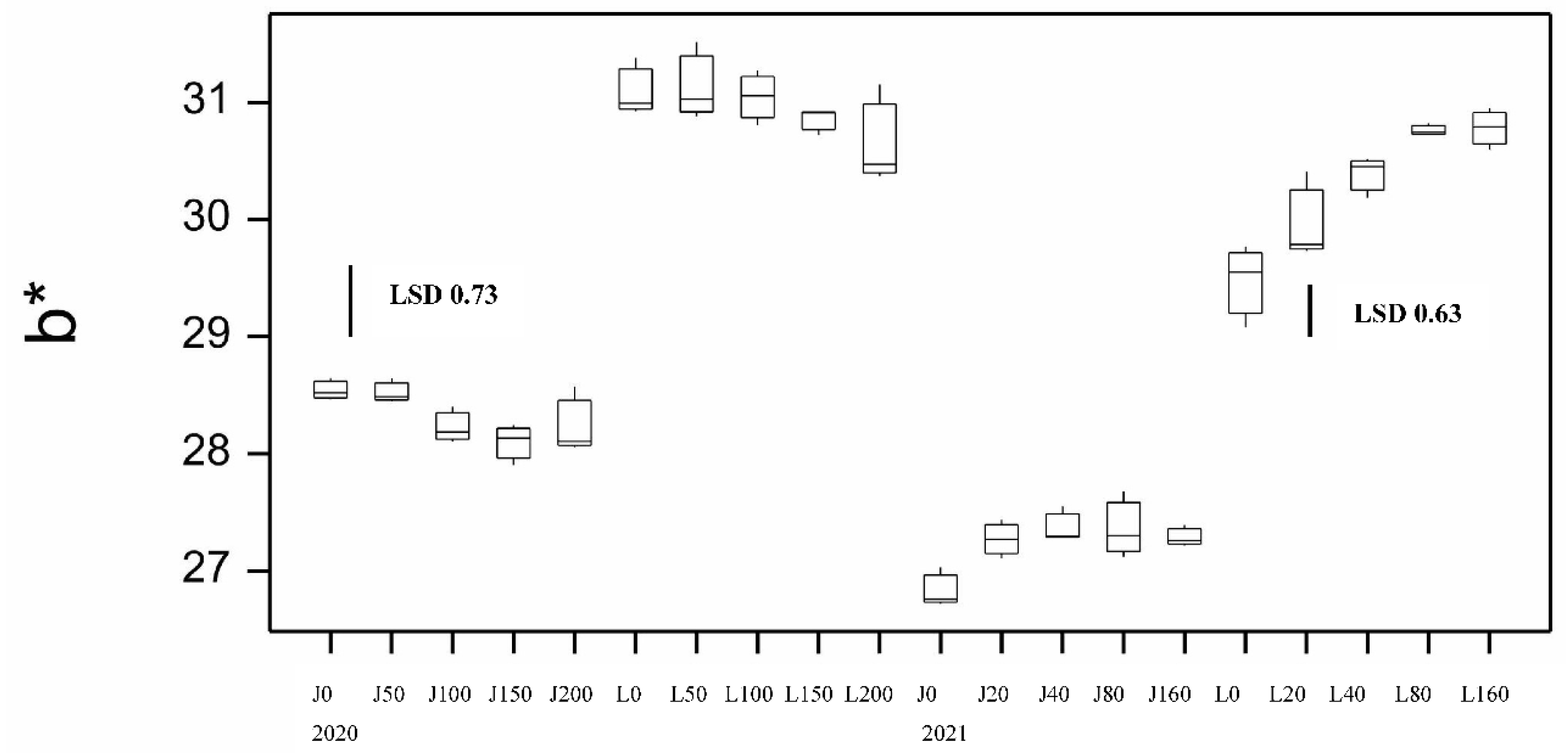
Figure 8.
Boxplot showing gluten index (GI) for both varieties at different N application rates at sowing for 2020 and 2021 seasons. LSD statistic shown for each year.
Figure 8.
Boxplot showing gluten index (GI) for both varieties at different N application rates at sowing for 2020 and 2021 seasons. LSD statistic shown for each year.
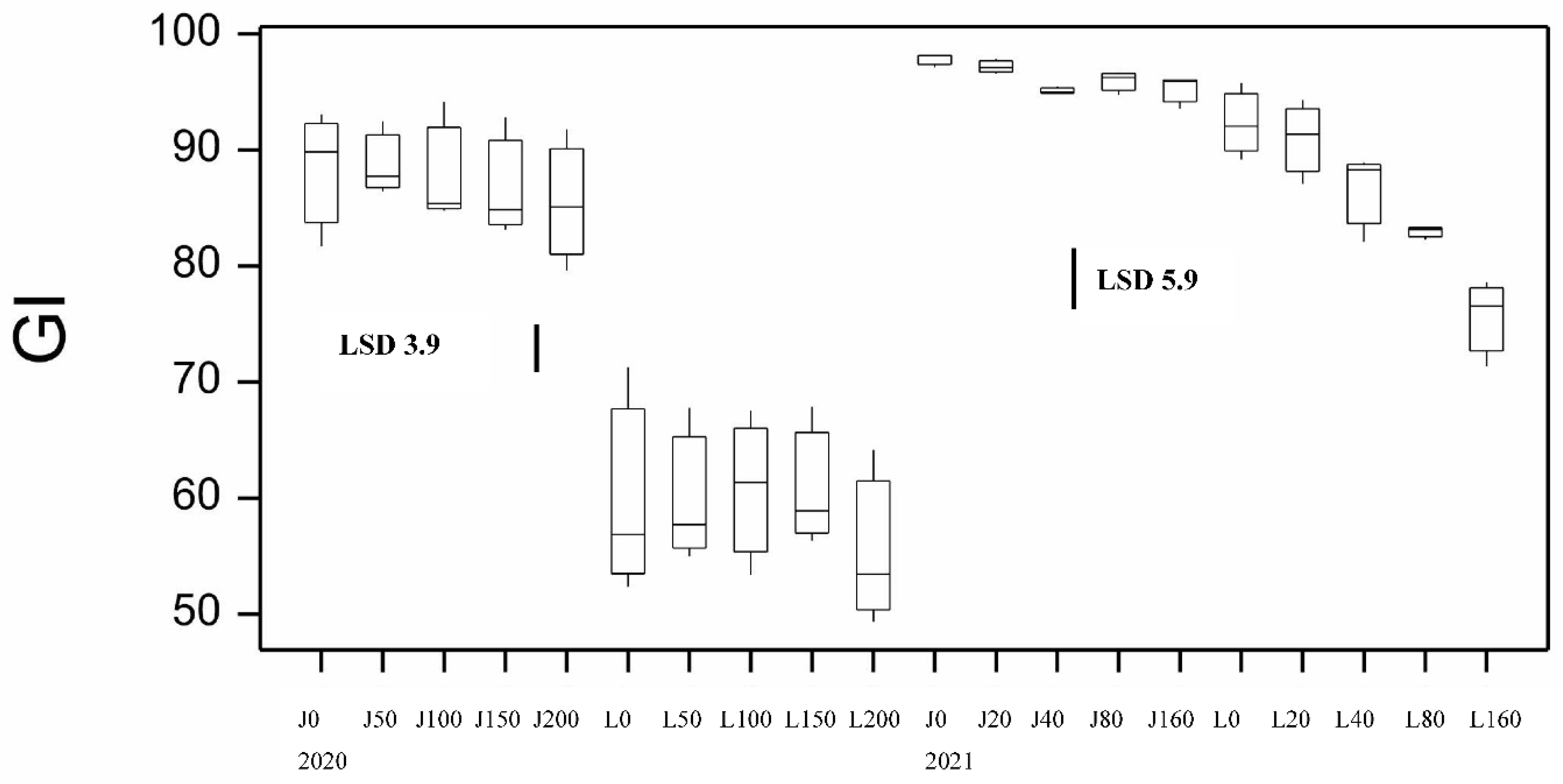
Figure 9.
Boxplot showing mixograph resistance breakdown (RBD) for both varieties at different N application rates at sowing for 2020 and 2021 seasons. LSD statistic shown for each year.
Figure 9.
Boxplot showing mixograph resistance breakdown (RBD) for both varieties at different N application rates at sowing for 2020 and 2021 seasons. LSD statistic shown for each year.
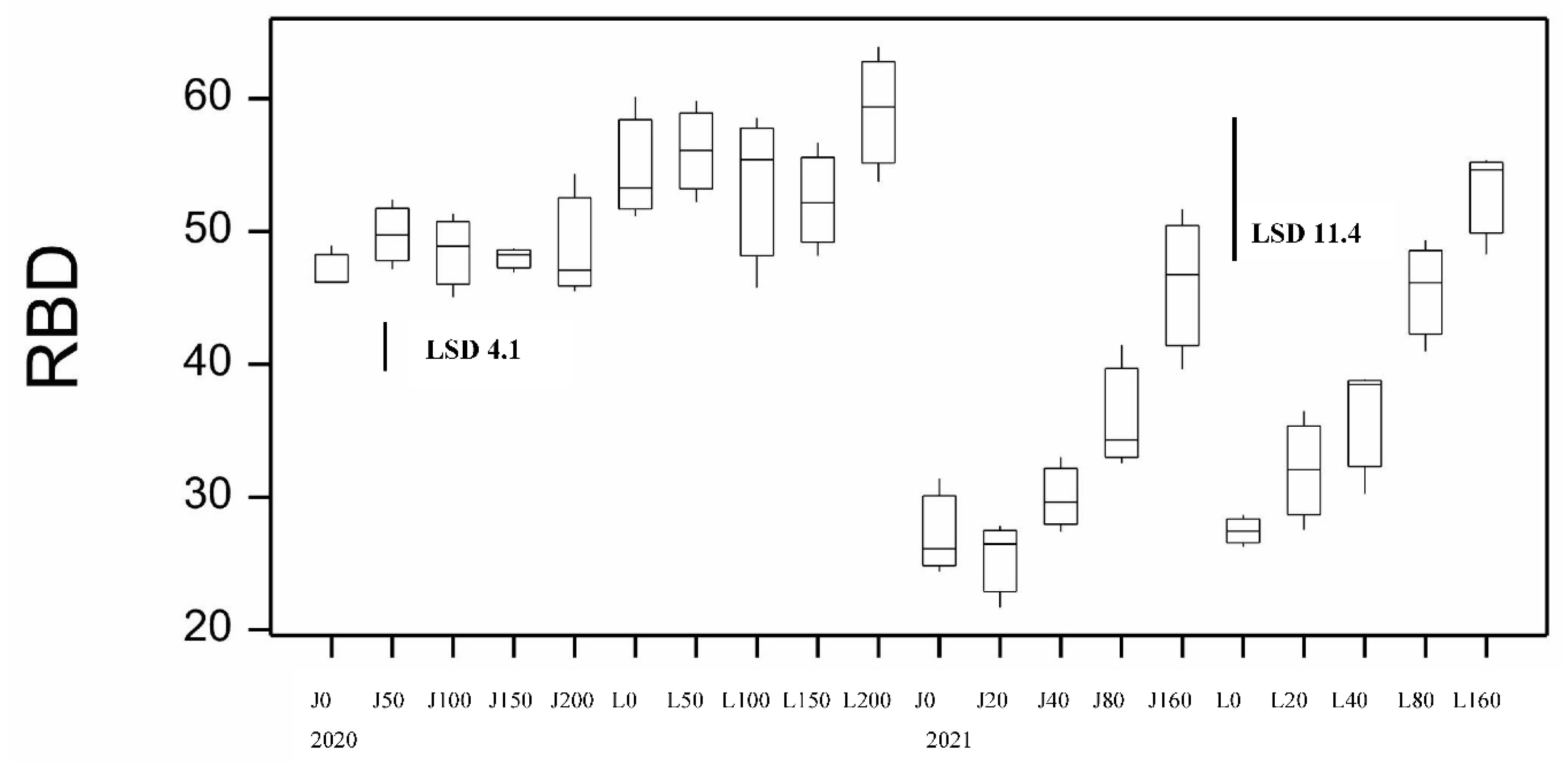
Figure 10.
Boxplot showing grain yield for both varieties at different N application and CR inoculation rates from 2021 season.
Figure 10.
Boxplot showing grain yield for both varieties at different N application and CR inoculation rates from 2021 season.
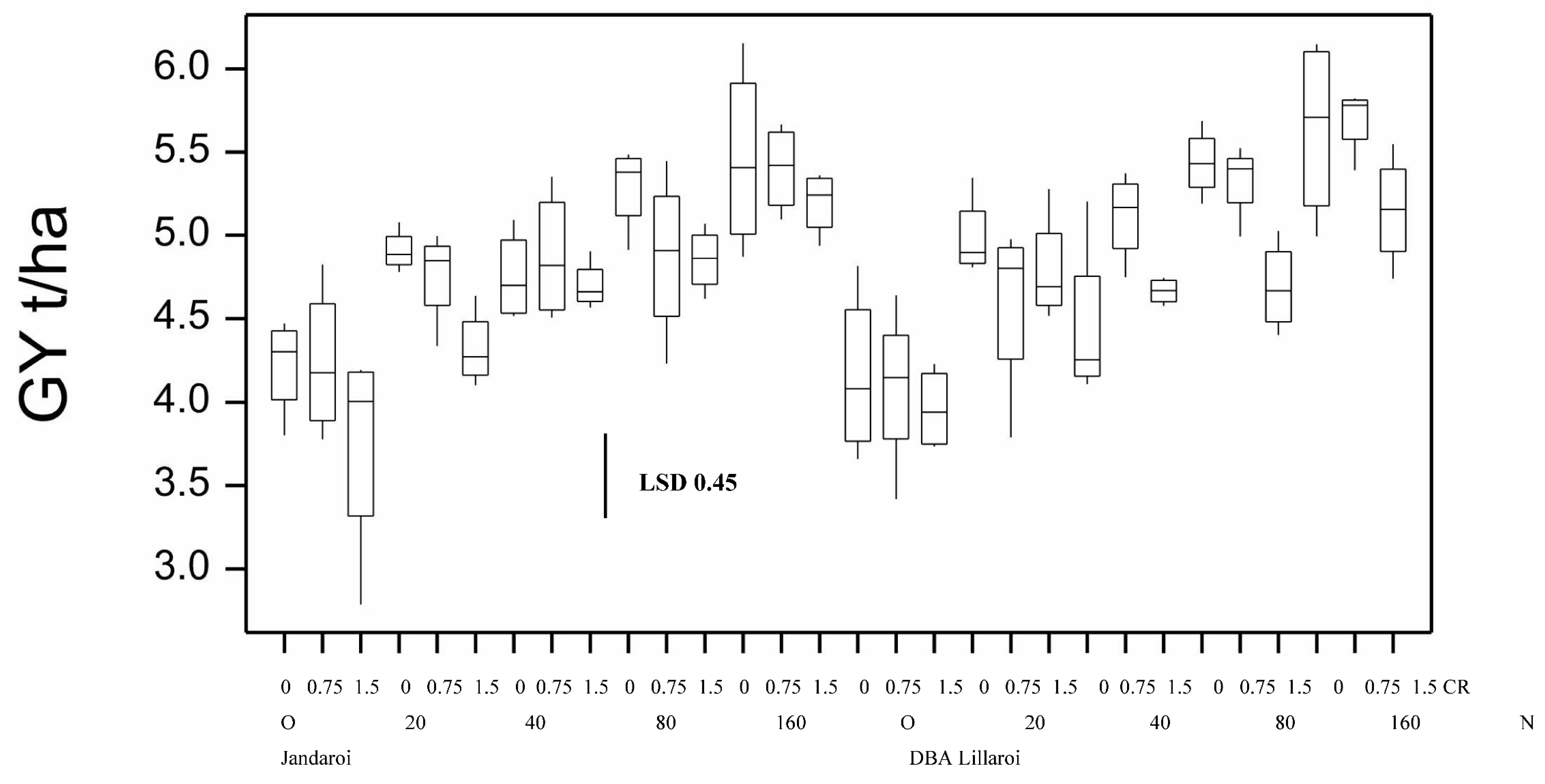
Figure 11.
A: Boxplot showing HVK% for both varieties at different N application rates at sowing for 2021 season. LSD statistic shown. B: Linear regression plots for each variety, lines are best fit, r2 = 0.76, p<0.001.
Figure 11.
A: Boxplot showing HVK% for both varieties at different N application rates at sowing for 2021 season. LSD statistic shown. B: Linear regression plots for each variety, lines are best fit, r2 = 0.76, p<0.001.
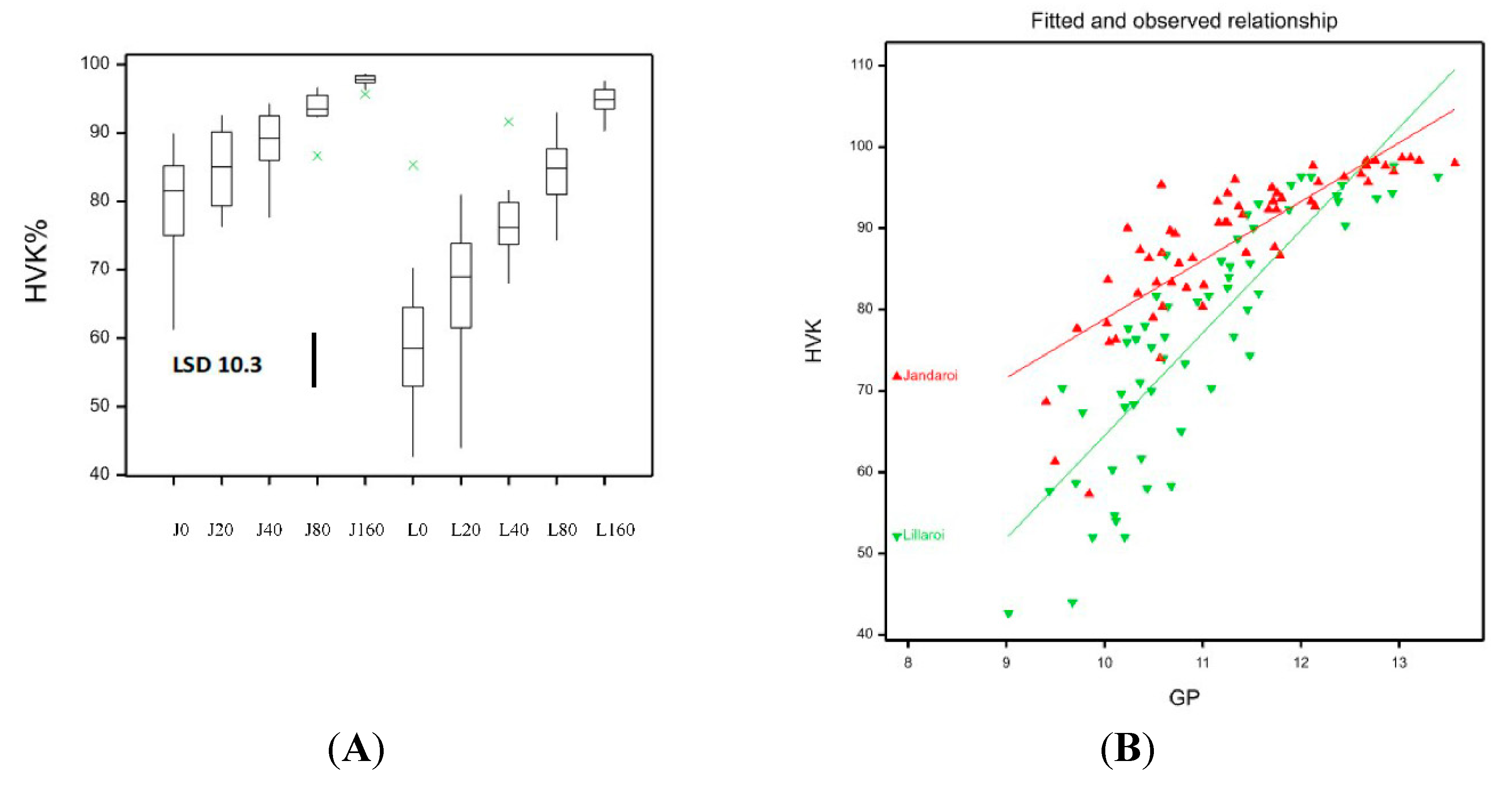
Figure 12.
A: Boxplot showing SE-HPLC soluble B (SolB%) and B:gliadin:glutenin ratio (Gli/Glu) for both varieties at different N application rates at sowing for 2021 season. LSD statistic shown.
Figure 12.
A: Boxplot showing SE-HPLC soluble B (SolB%) and B:gliadin:glutenin ratio (Gli/Glu) for both varieties at different N application rates at sowing for 2021 season. LSD statistic shown.
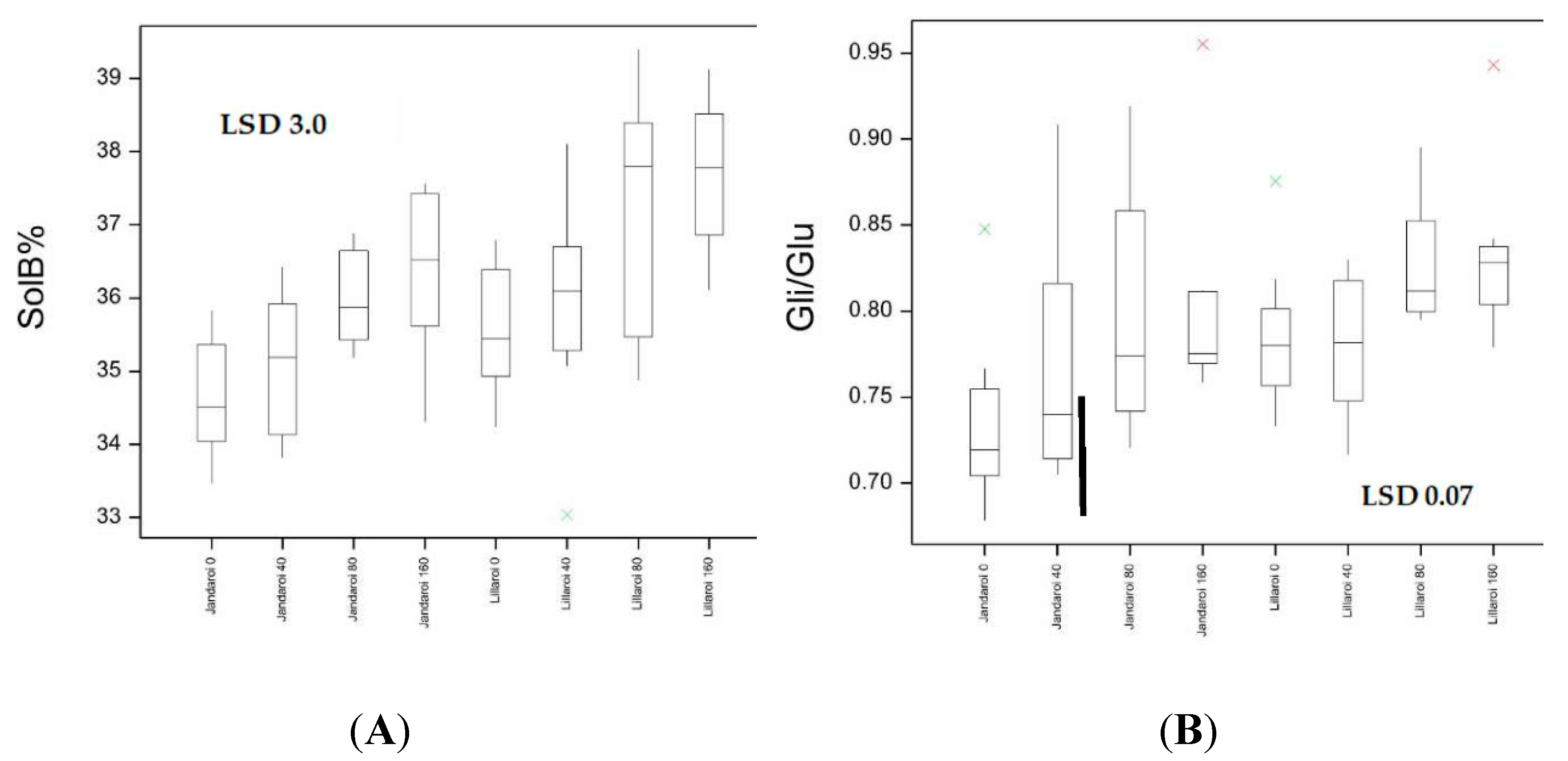
Figure 13.
PCA plots of mean values of samples for selected traits from the 2021 season. Colour coding for Nrate (A and C) and variety (C and D) are shown in legends.
Figure 13.
PCA plots of mean values of samples for selected traits from the 2021 season. Colour coding for Nrate (A and C) and variety (C and D) are shown in legends.
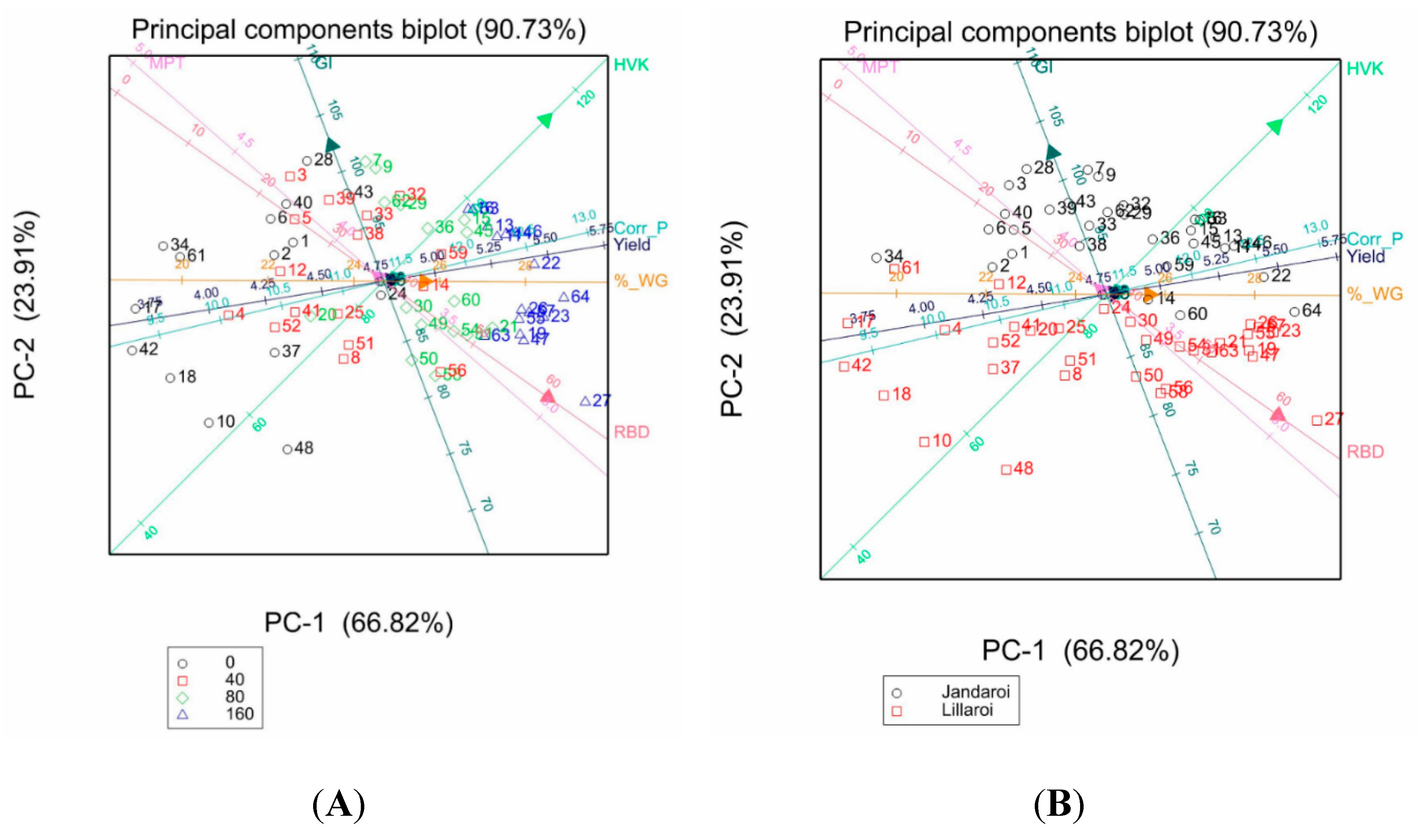
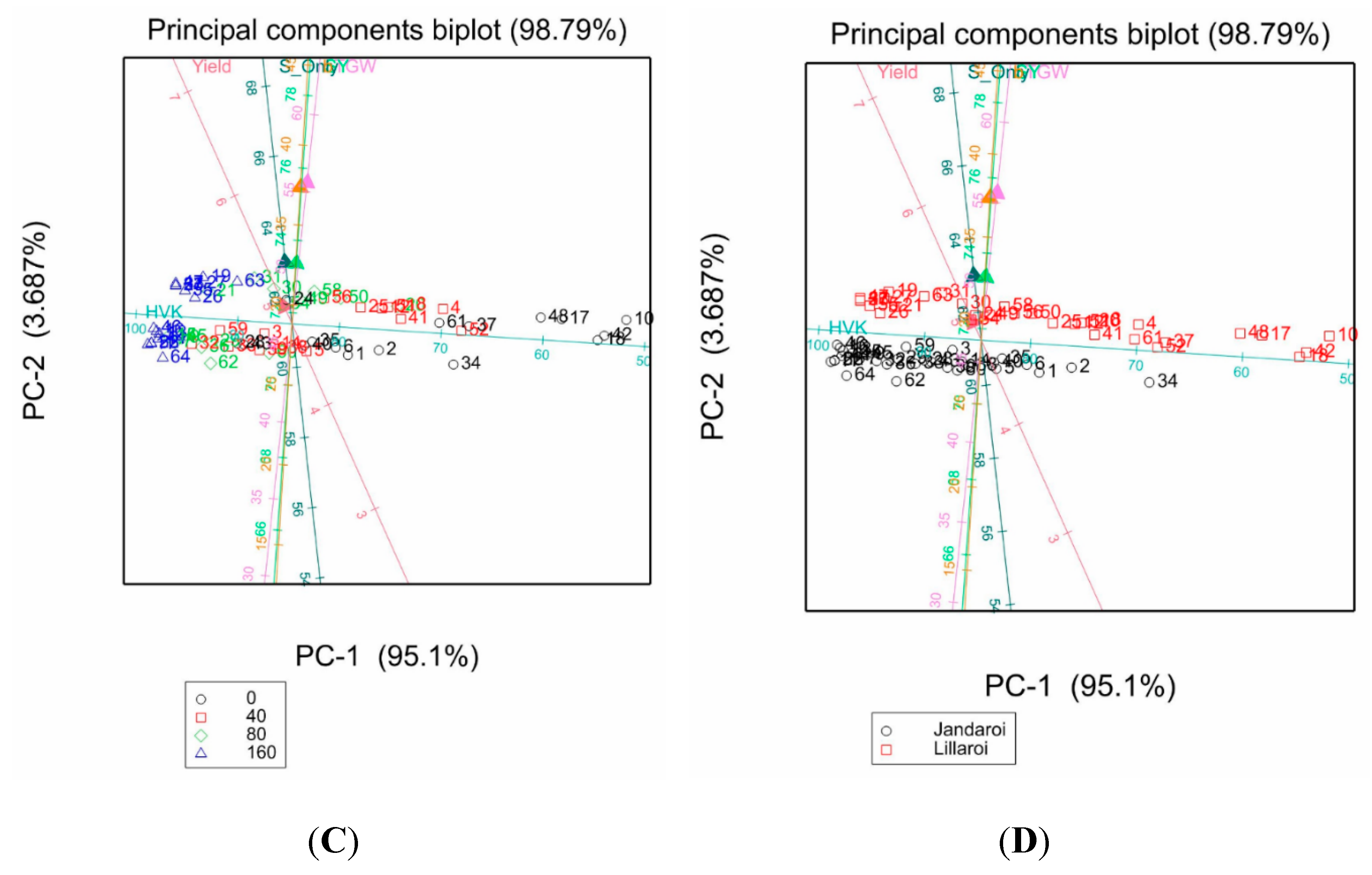
Figure 14.
Linear model showing fitted values (line) and observed (triangles) between HVK and Gli/Glu ratio by variety.
Figure 14.
Linear model showing fitted values (line) and observed (triangles) between HVK and Gli/Glu ratio by variety.
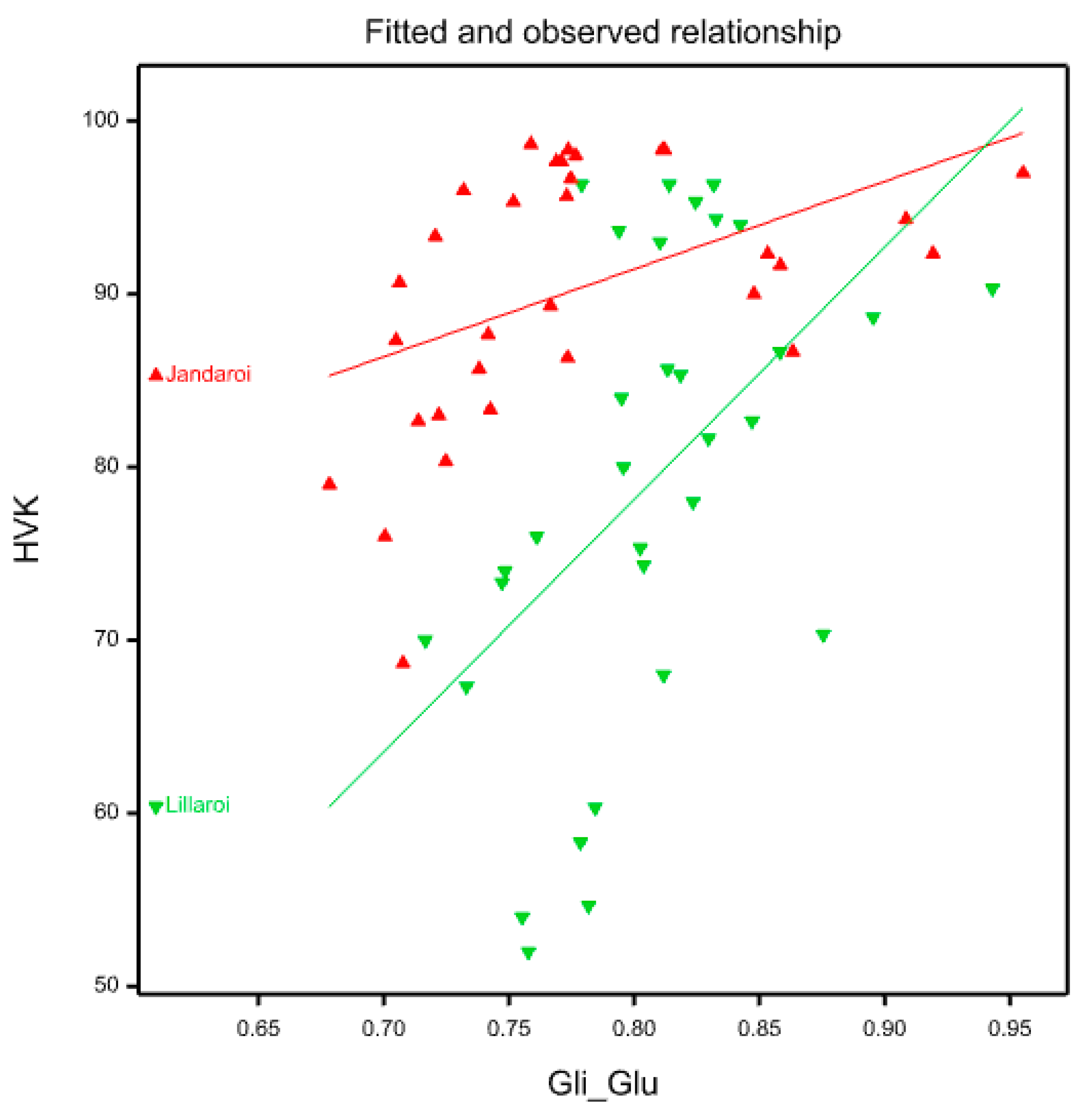

Table 1.
Seasonal rainfall (mm), minimum temperature and maximum temperature (°C) at Tamworth in 2020, 2021 and LTA average data (1993-2021).
Table 1.
Seasonal rainfall (mm), minimum temperature and maximum temperature (°C) at Tamworth in 2020, 2021 and LTA average data (1993-2021).
| Metric | Jan | Feb | March | April | May | June | July | Aug | Sept | Oct | Nov | Dec | Annual |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rainfall 2020 | 97.4 | 202.2 | 83.4 | 49.0 | 39.2 | 29.4 | 37.0 | 29.2 | 26.8 | 73.6 | 3.0 | 223.6 | 893.8 |
| Rainfall 2021 | 23.6 | 108.2 | 162.0 | 22.6 | 24.6 | 125.0 | 65.4 | 48.4 | 36.8 | 52.0 | 191.0 | 70.6 | 930.2 |
| Rainfall LTA | 61.2 | 73.9 | 57.2 | 25.3 | 30.4 | 53.4 | 40.9 | 39.5 | 46.3 | 58.3 | 82.3 | 80.4 | 638.5 |
| Min T 2020 | 21.4 | 19.0 | 14.4 | 9.8 | 5.1 | 3.7 | 3.5 | 2.9 | 6.6 | 10.3 | 13.3 | 17.0 | |
| Min T 2021 | 15.3 | 15.7 | 14.4 | 7.0 | 4.1 | 3.7 | 2.9 | 3.4 | 4.1 | 8.7 | 14.0 | 14.7 | |
| Min T LTA | 17.7 | 16.9 | 14.5 | 10.0 | 6.0 | 3.6 | 2.3 | 2.8 | 5.8 | 9.7 | 13.3 | 15.7 | |
| Max T 2020 | 36.3 | 29.7 | 27.5 | 24.1 | 18.9 | 17.5 | 16.9 | 17.6 | 23.3 | 27.1 | 31.8 | 30.4 | |
| Max T 2021 | 30.8 | 29.9 | 27.4 | 23.9 | 20.5 | 16.8 | 16.6 | 19.9 | 22.1 | 25.4 | 25.9 | 29.9 | |
| Max T LTA | 33.0 | 31.5 | 29.2 | 25.5 | 20.8 | 17.0 | 16.5 | 18.5 | 22.0 | 25.6 | 28.6 | 30.8 |
LTA=long term average from Australian Bureau of Meteorology data http://www.bom.gov.au/jsp/ncc/cdio/weatherData/av?p_nccObsCode=139&p_display_type=dataFile&p_startYear=&p_c=&p_stn_num=055325.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



