
Submitted:
25 May 2023
Posted:
29 May 2023
You are already at the latest version
Abstract
5-Hydroxytryptamine (5-HT) or serotonin plays a crucial role as a neuromodulator and/or neurotransmitter of several nervous system functions. Its actions are complex and depend on multiple factors, including the type of effector or receptor activated. Briefly, 5-HT can activate: (i) metabotropic (G-protein-coupled) receptors to promote inhibition (5-HT1, 5-HT5) or activation (5-HT4, 5-HT6, 5 HT7) of adenylate cyclase, as well as activation (5-HT2) of phospholipase C; and (ii) ionotropic receptors (5-HT3), which form a ligand-gated Na+/K+ channel. Regarding blood pressure regulation (and beyond the intricacy of central 5-HT effects), this monoamine also exerts direct postjunctional (on vascular smooth muscle and endothelium) or indirect prejunctional (on autonomic and sensory perivascular nerves) effects. At the prejunctional level, 5-HT can facilitate or preclude the release of autonomic (e.g., noradrenaline and acetylcholine) or sensory (e.g., calcitonin gene-related peptide) neurotransmitters facilitating hypertensive or hypotensive effects. Hence, we cannot formulate a specific impact of 5-HT on blood pressure level, since an increase or decrease in neurotransmitter release would be favoured depending on the type of prejunctional receptor involved. This review summarizes and discusses the current knowledge on the prejunctional mechanisms involved in blood pressure regulation by 5-HT and its impact on some vascular-related diseases.
Keywords:
Serotonin
; CGRP
; blood pressure
; migraine
; hypertension
1. Introduction
Among all biogenic monoamines, serotonin (5-hydroxytryptamine; 5-HT) stands out for its complex effects, the participation of a wide variety of receptors (which include the 5-HT1, 5-HT2, 5-HT3, 5-HT4, 5-HT5, 5-HT6 and 5-HT7 receptors), and its extensive distribution in vertebrates and invertebrates [1]. In mammals, 5-HT is mainly synthesised in enterochromaffin cells (~90 %) and in serotonergic neurons of the brain (1-2 %) [2]. Indeed, this monoamine is predominantly found in platelets, enterochromaffin cells and in the central nervous system (CNS), but in many cases, its physiological role remains elusive [1,3,4,5]. Fortunately, with the progressive development of agonists and antagonists that act selectively on 5-HT receptors, many functions of 5-HT in the CNS and in the periphery have been discovered [1,3,4,5].
1.1. A summary on 5-HT receptors
This review will not document historical aspects of 5-HT research, discovery or 5-HT receptors. However, published research on the mechanisms involved in the effects of 5-HT (even long before its identification as 5-HT) has accumulated over 130 years [1,3,4,5,6,7,8,9,10,11].
As summarized in Table 1, with the conjunction of structural, transductional and operational (pharmacological) criteria, 5-HT receptors have classified into seven receptor types (5-HT1-5-HT7) that can be grouped into: (i) six metabotropic (G-protein-coupled) receptors, namely: the 5-HT1 (further subdivided into the 5-HT1A, 5-HT1B, 5-HT1D, 5-ht1e and 5-HT1F subtypes), 5-HT2 (further subdivided into the 5-HT2A, 5-HT2B and 5-HT2C subtypes), 5-HT4, 5-HT5 (further subdivided into the 5-HT5A and 5-ht5B subtypes), 5-HT6 and 5-HT7 receptor types; and (ii) one ligand-gated ion channel represented by the ionotropic 5- HT3 receptor type [1,3,6,7,8]. The corresponding subtypes of the 5-HT1, 5-HT2 and 5-HT5 receptor types share similar structural and transductional properties, but display very different pharmacological profiles.
Some agonists and antagonists employed to identify the pharmacological profile of each 5-HT receptor type are shown in Table 1. As previously established [1,3,6,7,8], the pharmacological identification of a specific 5-HT receptor type is based on the application of: (i) inclusion criteria (i.e., selective agonists for this receptor mimic the effects of 5-HT, while selective antagonists for this receptor produce a blockade of the effects of 5-HT and the corresponding agonist); and (ii) exclusion criteria (i.e., agonists and antagonists for the other 5-HT receptors -and sometimes even for receptors unrelated to 5-HT- are inactive) (see Table 1).
This knowledge has helped to establish the role of 5-HT receptors in a number of diseases, including anxiety, depression, schizophrenia, drug addiction, cardiovascular pathologies (e.g., systemic, pulmonary and portal hypertension), cardiac disorders, migraine, etc., and has also led to the development of agonists and antagonists at 5-HT receptors for the therapeutic treatment of these -and other- diseases [1,3,4,5,6,7,8,9,10,11,12,13,14].
1.2. An overview of the effects of 5-HT in the cardiovascular system
As previously described in other reviews dealing with 5-HT and the cardiovascular system [3,4,5,8,9,10,11,12,13,14,15], the cardiovascular effects of 5-HT are complex and include bradycardia/tachycardia, hypotension/hypertension and vasodilatation/vasoconstriction. This complexity of effects is due to: (i) the capability of 5-HT to interact at various levels, including the heart and blood vessels, as well as the central and peripheral (autonomic and sensory) nervous systems; and (ii) the involvement of serotonin 5-HT1, 5-HT2, 5-HT3, 5-HT4, 5-HT5A and 5-HT7 receptors, as well as a tyramine-like action or unidentified mechanisms, depending on the species and the experimental conditions [3,4,5,8,9,10,11,12,13,14,15]. Interestingly, the 5-HT6 receptor is not involved in the cardiovascular effects of 5-HT [14,16].
1.3. The specific interactions of 5-HT at peripheral and central levels to induce cardiovascular effects
1.3.1. Sensory afferents
Overall, an intravenous (i.v.) bolus injection of 5-HT in anaesthetised animals results in a reflex bradycardia and hypotension by stimulating 5-HT3 receptors on vagal sensory afferents [3,4,5,9]. These neuronal 5-HT3 receptors were identified by using selective agonists and antagonists (see Table 1).
1.3.2. Sympathetic ganglia
I.v. administration of 5-HT may induce stimulation and/or inhibition at sympathetic ganglia which, in turn, may result in sympatho-excitation and/or sympatho-inhibition and, consequently, in vasopressor, vasodepressor, tachycardic and/or bradycardic responses [3,4,5,9]. Furthermore, 5-HT-induced hyperpolarization of sympathetic ganglia activates 5-HT1A receptors in rats; these 5-HT1A receptors were identified by using selective agonists and antagonists (see Table 1).
1.3.3. Cardiac effects of 5-HT
Central or i.v. administration of 5-HT may produce bradycardia and/or tachycardia, and the 5-HT receptors involved in these effects have been identified by using some of the agonists and antagonists shown in Table 1 [3,4,5,9,12].
In general, central 5-HT pathways regulating the cardiovascular system involve two main receptors, namely: 5-HT1A receptors (which mostly mediate sympatho-inhibition) and 5-HT2 receptors (which typically induce sympatho-excitation) [3,17,18]; some of the agonists and antagonists used to identify these receptors (with the inclusion and exclusion criteria described in section 1.1.) are shown in Table 1. Admittedly, central administration of 5-HT elicits complex and contradictory cardiac effects which depend, among other factors, on the species, the exact site of central application, the drug used and the dose employed [3,17,18]. In contrast, the bradycardia or tachycardia produced by i.v. administration of 5-HT is more controllable and consistent (see below) in view of the implied simplicity of the procedure.
1.3.3.1. Bradycardia
I.v. administration of 5-HT in intact animals results in a pronounced and transient bradycardia that is abolished after ganglion blockade, vagotomy, atropine, spinal section or 5-HT3 receptor antagonists [3,4,5,9,12]. This response involves the von Bezold-Jarisch reflex originating from the depolarization of afferent cardiac sensory neurons via activation of 5-HT3 receptors [3,4,5,9,12]. Furthermore, 5-HT can also produce bradycardia by: (i) a cardiac sympatho-inhibition via activation of prejunctional 5-HT1B, 5-HT1D and 5-HT5A receptors in pithed rats [19,20,21]; or (ii) a cardiac vagal stimulation via activation of 5-HT3 receptors on parasympathetic ganglia and postganglionic vagal nerves in rabbits [3,4,5,9,12] (see Table 1 for pharmacological tools).
1.3.3.2. Tachycardia
I.v. administration of 5-HT in vagotomized animals induces a tachycardic effect that may be mediated by a wide variety of receptors/mechanisms depending on the species and the experimental conditions [3,4,5,9,12]. These receptors/mechanisms include: (i) a tyramine-like action in spinal guinea-pigs; (ii) direct stimulation of 5-HT2A receptors on the cardiac pacemaker in reserpinized pithed rats; (iii) activation of 5-HT3 receptors on cardiac sympathetic neurons in the rabbit perfused heart, resulting in noradrenaline release and cardiac stimulation; (iv) activation of 5-HT3 receptors on calcitonin gene-related peptide (CGRP)-containing sensory neurons in isolated guinea-pig atrium, resulting in CGRP release and cardiac stimulation; (v) direct stimulation of 5-HT3 receptors on the cardiac pacemaker in conscious dogs; (vi) direct stimulation of 5-HT4 receptors on the cardiac pacemaker in healthy anaesthetized pigs (which is also involved in the positive inotropic effects of 5-HT in isolated human atria and in rats with chronic heart failure); (vii) direct stimulation of 5-HT7 receptors on the cardiac pacemaker in spinal cats; and (viii) unidentified mechanisms in the isolated hearts of certain lamellibranch and gastropod species (including Mercenaria mercenaria, Patella vulgata, Tapes watlingi, Helix aspersa, Aplysia, etc.). These receptors were pharmacologically identified by using selective agonists and antagonists for each type and the inclusion and exclusion criteria explained in section 1.1. (see Table 1).
1.3.4. Vascular and blood pressure effects of 5-HT
As explained in other reviews [3,4,5,13,14], i.v. administration of 5-HT results in a triphasic effect on arterial blood pressure, consisting of an initial transient vasodepressor effect followed by a vasopressor effect, and then a late long-lasting vasodepressor effect.
1.3.4.1. Initial transient vasodepressor effect
This response results from an abrupt bradycardia (and the consequent decrease in cardiac output) following stimulation of 5-HT3 receptors on afferent cardiac vagal afferents (i.e., the von Bezold-Jarisch reflex; see above and Table 1).
1.3.4.2. Vasopressor effect
This effect (which varies quantitatively depending on the species and the experimental conditions) involves the activation of vascular 5-HT2 receptors in resistance blood vessels (resulting in peripheral vasoconstriction). It is worthy of note that a release of catecholamines by adrenomedullary 5-HT2 receptors is also involved in dogs, while 5-HT1B receptors mediate vasoconstriction in cranial blood vessels of humans, pigs and dogs, as well as in the saphenous vein and external/internal carotid arterial beds of dogs [3,4,5]. Moreover, both 5-HT1B and 5-HT2 receptors induce vasoconstriction in the canine internal carotid arterial bed, while 5-HT may act directly on α-adrenoceptors in isolated rabbit ear and external carotid arteries [3,4,5]. Some of the agonists and antagonists used to identify these receptors (applying the inclusion and exclusion criteria defined in section 1.1.) are shown in Table 1.
1.3.4.3. Late long-lasting vasodepressor effect
This effect is predominantly, but not exclusively (see below), mediated by musculotropic 5-HT7 receptors [3,4,5,12,13,14]. Notwithstanding, several vascular mechanisms may contribute to different degrees in different experimental conditions and species. These mechanisms contributing to the late long-lasting vasodepressor effect of 5-HT may include:
(i) Direct vasodilatation. The direct vasodilatation to 5-HT is mediated by 5-HT7 receptors in a wide variety of blood vessels in different species under different experimental conditions [3,4,12,13,14]. Some of the agonists and antagonists used to identify these receptors (applying the aforementioned inclusion and exclusion criteria) are shown in Table 1. Moreover, in the blood vessels where 5-HT7 receptors produce vasodilatation and 5-HT2/5-HT1B receptors produce vasoconstriction, the final effect of 5-HT would depend on the pre-existing vascular tone, the dose employed, and the proportions in which these receptors are distributed [3,4].
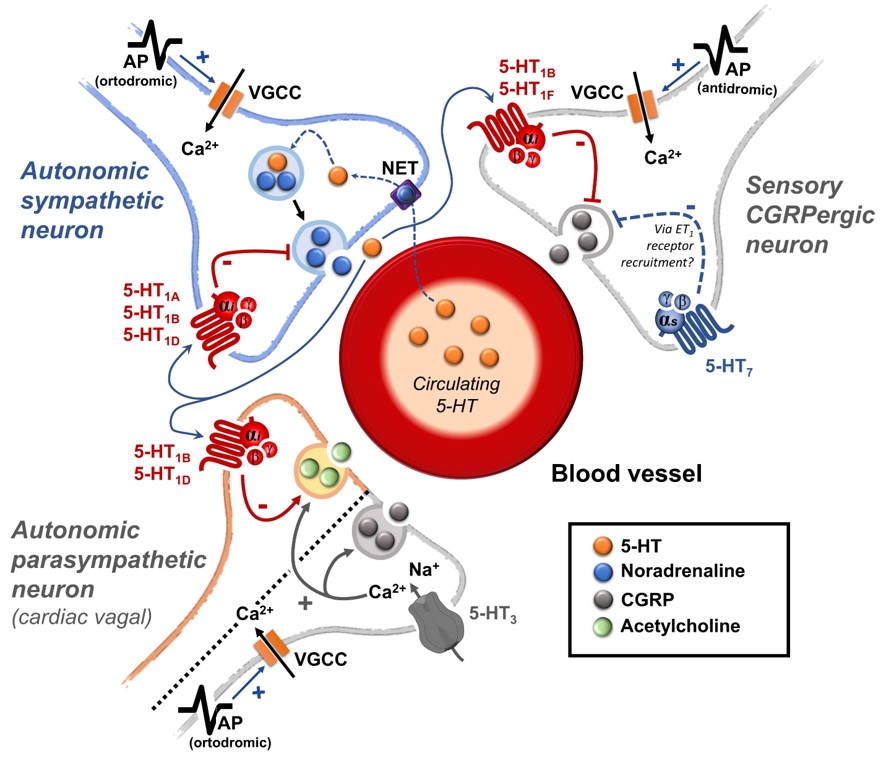
(ii) Prejunctional inhibition of perivascular sympathetic neurons. The prejunctional inhibition induced by 5-HT and related agonists on noradrenaline release from perivascular sympathetic neurons has been confirmed in vitro and in vivo in many blood vessels [3,4]. This vascular sympatho-inhibition, generally mediated by 5-HT1 receptors, may involve the 5-HT1A, 5-HT1B and/or 5-HT1D receptor subtypes depending on the vascular bed under study, the species and the experimental conditions [3,4]. Interestingly, sympatho-inhibitory 5-HT7 receptors could also be involved when rats are chronically pretreated with the 5-HT2 receptor antagonist sarpogrelate [3,22]. These receptors were pharmacologically identified by applying the inclusion and exclusion criteria explained in section 1.1. (see Table 1 and Figure 1).
(iii) Endothelium-dependent vasodilatation. In isolated blood vessels of several species without a functional endothelium, the vasodilatation to 5-HT is attenuated while the vasoconstriction is augmented [3,4,5]. This vasodilator effect of 5-HT, involving endothelial release of nitric oxide, is predominantly mediated by 5-HT1 receptors [3,4]. Interestingly, in porcine blood vessels, the 5-HT-induced endothelium-dependent vasodilatation involves: (i) 5-HT1B/1D receptors in coronary arteries; or (ii) 5-HT2B receptors in pulmonary arteries (see Table 1).
(iv) Actions in the CNS. Central administration of 5-HT may produce vasodepressor, vasopressor or biphasic effects depending on the exact site of application, dose employed, depth of anaesthesia, the species used, etc. [3,4,5]. As previously reported [3,16,18], the cardiovascular regulation by central 5-HT neurons involves: (i) 5-HT1A receptors (associated with sympatho-inhibition, hypotension and bradycardia); and (ii) 5-HT2 receptors (associated with sympatho-excitation and hypertension). Indeed, when directly applied in the CNS, 5-HT may produce both sympatho-inhibition and cardiac-vagal stimulation via 5-HT1A receptors [17,23]. In fact, psychiatric conditions that involve alterations in the serotoninergic limbic components are usually accompanied by an autonomic imbalance; for example, the posttraumatic stress disorder includes clinical manifestations such as cardiac arrhythmia, tachycardia, high blood pressure, etc. [24,25]. Moreover, anxiety correlates strongly with adrenaline levels in a positive direction [26], while aberrations in the autonomic nervous system (ANS) have been reported in patients with depression or other mood alterations [27]. Hence, central 5-HT is a powerful modulator of the ANS whose complex mechanisms fall beyond the scope of the present review. Interestingly, cerebral 5-HT can cross the blood-brain barrier via the 5-HT transporter (SERT) in endothelial cells and reach systemic circulation [28].
1.3.5. Receptor-independent actions of 5-HT
Apart from the above cardiovascular effects of 5-HT mediated by 5-HT receptors, other studies suggest that 5-HT can also play cardiovascular (patho)physiological roles independent of 5-HT receptor activation [3]. For example: (i) rats pretreated with fluoxetine were protected from monocrotaline-induced pulmonary hypertension [29]; and (ii) 5-HT uptake can “serotonylate” proteins by transglutaminase-2 [30], a mechanism involved in the mitogenic and profibrotic effects of 5-HT without receptor activation [31].
2. Peripheral autonomic nervous system and prejunctional 5-HT receptors
2.1. An overview of the peripheral actions of 5-HT regulating the vascular function
Although 5-HT modulates the ANS at the central level [16,17,18], presynaptic and pre-junctional mechanisms by which 5-HT controls perivascular cholinergic and adrenergic outflows is relevant. Indeed, mutant mice lacking the SERT gene showed increased noradrenaline levels in plasm after mechanic immobilization [32], suggesting that peripheral 5-HT reuptake may be an essential mechanism involved in the systemic catecholaminergic modulation by 5-HT during stressful situations. Nevertheless, acute and systemic administration of selective SERT inhibitors may produce sympathetic inhibition (mainly via a central mechanism) [33].
Certainly, both SERT and 5-HT receptors are expressed in rodent adrenal glands, particularly in chromaffin cells [34], and 5-HT is involved in the development of the adrenal medulla [35]. Moreover, the number of adrenal chromaffin cells in mice embryos seems to be controlled by 5-HT3 receptors expressed in their Schwann cell precursors [36]. Hence, 5-HT modulates adrenal chromaffin cells since its development and, probably, during the rest of the lifetime.
Interestingly, when considering the distribution of SERTs in the adrenal chromaffin cells population, 5-HT seems to be strategically taken up by cells that exert an autocrine/paracrine modulation on the rest of chromaffin cells that release several vasocontractile mediators to the systemic circulation; these include adrenaline (~79%), noradrenaline (~18%) and other mediators (~1-3%) during a sympathetic fight/flight situation induced by fear, stress, exercise, or conflict [34,37]. In this manner, the adrenal chromaffin release is controlled both neurogenically (by the ANS) and non-neurogenically (by several mediators, including 5-HT) [34,37].
On the other hand, it is noteworthy that adrenal chromaffin cells do not synthesize 5-HT by themselves [38,39], but they can take up 5-HT via the high expression of SERTs [34,39]. Moreover, activation of 5 HT1A receptors decreased adrenal chromaffin release [27,38]. Hence, 5-HT may act as a neuroendocrine tool to modulate (negatively) catecholamines release after stressful events via 5-HT1 receptors. In addition, the autonomic control of the sympathetic nervous system at the vascular level is strategically organized to exert a local modulation of blood vessel sections or even complete vascular beds [40], forming a complex varicose network that surrounds the blood vessels at the level of the adventitia layer in close proximity with the smooth muscle cells. However, neurotransmitters can diffuse and reach the endothelium [41]; this opens the possibility for a highly specific modulation by 5-HT of each blood vessel layer, namely, tunica intima, tunica media, and tunica externa (also called tunica adventitia).
The parasympathetic branch of the ANS innervates only cerebral vascular beds, whereas it does not innervate peripheral resistance blood vessels [42,43]. Particularly, intracerebral-posterior blood vessels are richly innervated by parasympathetic fibres that seem to exert an essential control of blood flow in the polygon of Willis [44]. In peripheral blood vessels, vagal parasympathetic molecules (mainly acetylcholine) may be released systemically and reach the endothelium exerting vasorelaxant neuroendocrine actions [41]. In short, both sympathetic and vagal parasympathetic varicosities express 5-HT receptors [45,46]. Thus, 5-HT may modulate sympathetic and parasympathetic perivascular nerves and exert direct vascular actions [47,48], as described in section 1.
2.2. The role of prejunctional 5-HT receptors
There are several sources of 5-HT that may contribute to the modulation of perivascular autonomic and sensory nerve terminals; these include: (i) the systemic circulation, where 5-HT is transported via blood platelets and released upon activation [49,50]; (ii) chromaffin cells of the adrenal medulla [34,38]; (iii) enterochromaffin gastrointestinal cells [51]; (iv) a subgroup of trigeminal C-fibres which store 5-HT [52]; and (v) cortical terminals from raphe neurons [28].
In the parasympathetic branch, the sphenopalatine ganglion (SPG) positively regulates cerebral blood flow; interestingly, more than 96% of the SPG body cells express 5-HT receptors [53]. Hence, 5-HT may be seen as a ubiquitous autonomic modulator.
5-HT1/2/3 receptors are highly active during motor, sensory and autonomic neuron development [54]. Thus, it is logical to suppose those receptors keep homeostatic functions on the developed organism’s motor, sensory, and autonomic neurons. According to their transduction systems, 5-HT1 serotonin receptors are mainly involved in sympathetic inhibition, whereas 5-HT2 and 5-HT3 serotonin receptors may facilitate parasympathetic outflow (see below).
2.2.1. The 5-HT receptors inhibiting the autonomic outflow
On the sidelines of its central sites of action, 5-HT can inhibit the tachycardia induced by sympathetic electrical stimulation but not the one induced by exogenous noradrenaline [55]. This finding revealed the existence of a 5-HT-induced cardiac sympatho-inhibition at the prejunctional level (Figure 1).
5-HT1 receptors are widely expressed in sympathetic perivascular and cardiac terminals; its activation is linked to cardiovascular sympathetic inhibition [56,57,58,59]. Specifically, selective stimulation of 5-HT1A, 5-HT1B, and 5-HT1D serotonin receptors produced inhibition of the sympathetic vasopressor outflow in pithed rats [46,60]; this is a useful experimental model to pharmacologically study the modulation of the sympathetic prejunctional terminals [61]. Furthermore, in vitro experiments in the human atrium have shown that noradrenergic terminals express the 5-HT1D receptors, which mediate sympathetic inhibition [62]. Similarly, in pithed rats, the sympathetic cardioaccelerator outflow is inhibited by 5-HT1B/1D receptor activation [19].
2.2.2. The 5-HT receptors as facilitators of the autonomic outflow
In pithed rats, bradycardia induced by vagal electric stimulation may be increased by 5-HT during the blockade of 5-HT1/2 receptors and by selective 5-HT3 receptor agonists [63]. In contrast, activation of 5-HT2 receptors inhibited this bradycardia induced by vagal electrical stimulation [63]. These findings suggest a dual role for 5-HT receptors in the cardiac parasympathetic outflow. Interestingly, in cerebral blood vessels, most SPG parasympathetic neurons: (i) highly express 5-HT3A > 5-HT3B serotonin receptors; (ii) slightly express 5-HT2B > 5-HT2A > 5-HT1B receptors; and (iii) practically lack the expression of 5-HT1A, 5-HT1D, 5-HT1F, 5-HT2C, 5-HT4, 5-HT5A, 5-HT5B, 5-HT6 and 5-HT7 serotonin receptors [53]. In view that the 5-HT3 receptor forms a ligand-gated Na+/K+ channel, its parasympathetic expression in SPG neurons leads to acetylcholine, nitric oxide (NO), and vasoactive intestinal polypeptide (VIP) release, which, in turn, results in vasodilatation [64]. On the other hand, it remains unclear whether major cerebral blood vessels are rich in 5-HT receptors [65].
2.3. Clinical relevance and therapeutic potential
The serotoninergic negative modulation of sympathetic cardiovascular activity by 5-HT1A/1B/1D receptors may be achieved endogenously through platelet activation by catecholamines [50]. Indeed, a recent clinical study exposed 79 healthy male and female volunteers to tryptophan enhancement and 85 others to tryptophan depletion conditions to analyse adrenaline and noradrenaline plasma levels [66]. Participants from the tryptophan enhancement condition showed a clear increment in plasma adrenaline, while noradrenaline decreased. Interestingly, this depletion condition slightly increased adrenaline and noradrenaline levels compared with baseline [66], suggesting that some preclinical findings are also observed clinically. Hence, 5-HT1 serotonin receptors on perivascular fibres represent therapeutic targets to decrease sympathetic noradrenaline release.
On the other hand, a metanalysis with cancer and cancer-depressed patients concluded that management of stress-linked-emotions (which include serotoninergic alterations) is a crucial element in preventing comorbidities related to disruption of endocrine and autonomic (sympathetic) nervous system homeostasis [67] and in improving survival time in these patients. Hence, stabilizing 5-HT levels may be a strategy to prevent autonomic disorders as comorbidities in diseases with associated high emotional stress.
It is important to keep in mind that some blood vessels (e.g., those from the coronary and carotid vascular beds) express the 5-HT1 receptor in the smooth muscle layer, whose activation produces vasoconstriction [68,69]. These receptors are directly and/or indirectly involved in the physiopathology of migraine, and some of the prophylactic (e.g., methysergide) and acute (e.g., triptans and ergots) antimigraine drugs interact with these receptors [69,70,71]. Hence, as a well-founded concern, direct vascular effects should be considered in any strategy that modifies 5-HT levels.
3. Sensory CGRPergic perivascular nerves and prejunctional 5-HT receptors
3.1. The sensory perivascular CGRPergic neurons as an intrinsic modulator of vascular tone
In general, CGRP is a potent vasodilator that can be released by capsaicin [72]; hence, CGRP release is associated with the activation of TRPV1 receptors on sensory nerves [73,74]. Nevertheless, the role of other TRP ion channels (e.g., TRPA1, TRPM3) located on nociceptors inducing the release of CGRP has also been documented [75]. It is noteworthy that sensory nerves, which originate from the spinal cord [76], can exert: (i) afferent actions [76]; and (ii) efferent actions via local (axonal) or central (dorsal root) reflexes [15]. In contrast to the efferent autonomic perivascular innervation from the spinal ventral horn, the sensory-afferent fibres arrive at the spinal dorsal horn conveying information from the periphery to the spinal cord [15].
The relevance of the sensory nervous system (particularly CGRP) as an intrinsic modulator of vascular tone was elegantly demonstrated by a series of in situ and in vivo experiments led by the group of Kawasaki in the early 90s. Indeed, they showed that, after pharmacological blockade of autonomic function, electrical stimulation of perivascular sensory nerves resulted in a vasodilator action mediated by CGRP release (blocked by CGRP(8-37), a CGRP receptor antagonist), which was insensitive to blockade of β-adrenergic, muscarinic and histaminergic receptors [77,78,79,80]. More recently, our group has shown in pithed rats that after CGRP receptor blockade with olcegepant, not only are the neurogenic and non-neurogenic vasodepressor responses to CGRP precluded, but a potentiation of the noradrenergic vasopressor responses is also unmasked [81]. Together, these data demonstrate that selective stimulation of perivascular sensory nerves results in CGRP release at the prejunctional level, activating CGRP receptors and evoking vasodilation. Current data strongly support the notion that CGRPergic sensory transmission modulates vascular tone via smooth muscle or endothelial mechanisms [82,83].
At the prejunctional level, several mechanisms have been reported to impact the sensory release of CGRP. One of the first lines of evidence suggesting that prejunctional heteroreceptors in sensory nerves modulate CGRP release was observed in experiments performed in the mesenteric vascular beds [78]. Briefly, Kawasaki et al. [78] showed that the vasodilation induced by periarterial nerve stimulation is smaller in vascular beds precontracted with noradrenaline (the endogenous ligand; non-selective α1/2- and β-adrenergic agonist) than in those precontracted with methoxamine (a selective α1-adrenoceptor agonist); this finding correlated with activation of α2-adrenoceptor activation [77]. These data suggest that the sympathetic perivascular outflow induces a direct vasoconstrictor effect mediated by vascular activation of α1/2-adrenoceptors and an indirect action by inhibiting the vasodilator function of sensory perivascular fibres. Furthermore, since α2-adrenoceptors are divided into three functional subtypes (α2A/2B/2C-), further pharmacological analysis in pithed rats showed that a fine-tuning of the perivascular sensory release of CGRP at the systemic level exists by selective activation of α2A/2C-adrenoceptors [84]. In this regard, several other prejunctional heteroreceptors facilitating (e.g., TRPV1) or inhibiting (e.g., µ-opioid, D2-like, CB1, H3, P2Y1/13, and 5-HT1 receptors) CGRPergic neurovascular transmission have been described (for references see [15]).
It is worthy of note that the potential relevance of serotonergic transmission modulating the perivascular sensory CGRPergic outflow has been established in the last 15 years [85,86]. In the case of 5-HT receptors modulating perivascular CGRPergic transmission, special attention has been paid in the context of migraine pathophysiology and pharmacotherapy. Indeed, triptans like sumatriptan, which is a 5-HT1B/1D/1F receptor agonist considered the gold standard in acute migraine treatment ([11] relieves migraine attacks by producing: (i) direct vasoconstriction of intracranial and extracranial arteries; (ii) and inhibition of CGRP release at the trigeminal level and on perivascular sensory nerves [70,87].
3.2. Prejunctional 5-HT receptors are mainly inhibitors of the perivascular sensory CGRPergic outflow
As mentioned above, triptans and ergots (both agonists at 5-HT1 receptors) can prejunctionally inhibit CGRP release at the trigeminovascular level [88,89]. Indeed, the first evidence about the role of 5-HT1 receptors as inhibitors of CGRPergic transmission derived from pharmacological research on the mechanisms involved in the therapeutic effects of acute antimigraine drugs [90,91,92,93]. Admittedly, the discussion on the relevance of serotonergic mechanisms modulating CGRPergic outflow in the context of migraine (i.e., at trigeminovascular level) falls beyond the scope of the present review since several excellent reviews have been published elsewhere (see refs. [11,70,88,94,95,96,97,98]).
Nevertheless, considering that triptans and ergots are associated with cardiovascular side effects [11,70], a study in pithed rats demonstrated that acute (rather than prophylactic) antimigraine drugs are capable of inhibiting the perivascular sensory CGRPergic outflow at the systemic level, via prejunctional mechanisms [99]. Specifically, the pithed rat model was used to analyse vascular and prejunctional mechanisms excluding the influence of any central compensatory reflex mechanisms. Under these experimental conditions, in animals infused with hexamethonium (a sympathetic ganglionic blocker) and methoxamine (an α1-adrenoceptor agonist to induce a sustained systemic vasoconstriction), the treatment with sumatriptan, ergotamine, or dihydroergotamine inhibited the vasodepressor responses elicited by electrical stimulation of the T9-T12 spinal cord segments (an effect associated with inhibition of CGRP release from perivascular sensory nerves; [99]).
The above data strongly support the hypothesis that 5-HT receptors located on perivascular sensory nerve terminals modulate CGRP release in the vascular system (such as at the trigeminovascular level) (Figure 1). Indeed, molecular evidence at the dorsal root ganglion level has suggested that mRNA expression correlates with 5-HT1B and 5-HT1F, but not with 5-HT1A or 5-HT1D, receptors [100]. In this regard, further functional pharmacological experiments using the pithed rat model showed that the selective 5-HT1B receptor agonist, CP-93,129, selectively inhibits the neurogenic CGRPergic vasodepressor responses via prejunctional sensory mechanisms [86]. Likewise, some data suggest that trigeminal activation of prejunctional 5-HT1B receptors (by sumatriptan or donitriptan) inhibits the external carotid vasodilation induced by capsaicin [101,102], highlighting the relevance of this receptor subtype in the modulation of CGRP release. Furthermore, as discussed by Rubio-Beltrán et al. [103], since 5-HT1F receptors have been found on sensory nerves, the role of these receptors in the modulation of CGRP release is suggested. Indeed, lasmiditan (a selective 5-HT1F receptor agonist) can prejunctionally inhibit CGRP release not only at the central (trigeminal) level, but also at the peripheral (meninges) level [89].
Considering that sumatriptan is a non-selective 5-HT1A/1B/1D/1F receptor agonist, the role of these receptor subtypes was also analysed in the inhibition of the vasodepressor sensory CGRPergic outflow in pithed rats [85]. The data using selective agonists and antagonists for each 5-HT1 receptor subtype (see Table 1): (i) corroborated the relevance of 5-HT1B receptors, and further showed that activation of prejunctional 5-HT1F receptors inhibited CGRP release; and (ii) excluded the role of 5-HT1A and 5-HT1D receptors [85]. It is noteworthy that the role of prejunctional 5-HT1D receptors inhibiting CGRP release has also been suggested [104]; however, it must be emphasised that the development of a selective agonist for this receptor (i.e., PNU-142633) to treat migraine, a disorder where trigeminal release of CGRP plays a key role, was not effective [105].
In the case of the ergots, ergotamine and dihydroergotamine can also inhibit the perivascular sensory CGRPergic outflow. Nevertheless, their pharmacology is much more complex since these compounds display affinity for all 5-HT (except 5-HT3) receptors and also interact with dopaminergic and noradrenergic receptors. Indeed, a detailed pharmacological analysis showed that, apart from prejunctional 5-HT1B/1F receptors, prejunctional D2-like and α2-adrenergic receptors also inhibit the vasodepressor responses elicited by spinal electrical stimulation of the vasodepressor sensory CGRPergic outflow [70,106].
It is interesting to note that, mechanistically, the 5-HT1 receptor family is canonically coupled to Gi/o proteins [1,4] which, in turn: (i) via the Gα subunit reduces the activity of adenylate cyclase, diminishing intracellular cAMP levels and consequently inhibiting the activity of protein kinase A; and (ii) via the Gβ/γ subunits increases the activity of K+ channels. Both mechanisms are intrinsically associated with the inhibition of neurotransmitter release [107].
From this point of view, the finding that prejunctional 5-HT7 receptor activation with AS-19 inhibited the vasodepressor sensory CGRPergic outflow in pithed rats was surprising and counterintuitive [108], particularly if we consider that this receptor is positively coupled to Gsproteins [1,4]. Hence, one would have expected facilitation rather than inhibition of the rat vasodepressor sensory CGRPergic outflow. Nonetheless, the possibility exists that this 5-HT7 receptor-induced sensory inhibition may involve: (i) an ATP-dependent K+ channel-mediated hyperpolarization sensitive to glibenclamide [108], as previously reported for the 5-HT-induced inhibition of the contractile and electrical activities in the guinea-pig mesenteric bed [109]; and (ii) the endothelin pathway, as this response was blocked by sulfisoxazole [108], an endothelin ETA receptor antagonist [110]. Indeed, it has been shown that endothelin-1 inhibits the neuroeffector transmission in smooth muscle [111], and Filipelli et al. [112] demonstrated that endothelin-1 inhibits the capsaicin-induced CGRP release. Hence, the prejunctional 5-HT7 receptor seems to promote endothelin-1 secretion, which inhibits CGRP release.
In the case of the 5-HT7 receptor and nociceptive sensory transmission, activation of this receptor at the spinal cord exerts an antinociceptive action, whereas at the peripheral level enhances the peripheral capsaicin-induced sensitization [113]. Certainly, as previously mentioned, the effect of 5-HT is complex and depends not only on the 5-HT receptor subtype involved but also on the location of the receptor.
Finally, it is interesting to note that molecular expression analysis of 5-HT receptor expression in dorsal root ganglion neurons showed that, apart from 5-HT1B, 5-HT1F, and 5-HT7 receptors, also 5-HT2A, 5-HT2C, 5-HT3, 5-HT5A, 5-HT5B, and 5-HT6 receptors can be found in this type of cells [100,114,115]. Although the functional role of the later receptor (sub)types in sensory vascular neurotransmission has not yet been reported, experiments exploring nociception showed that activation of 5-HT2A or 5-HT4 receptors seems to enhance CGRPergic transmission [116,117], whereas 5-HT5A receptors have the opposite effect [118]. Furthermore, in guinea-pig isolated cardiac atria, 5-HT favours CGRP release via sensory 5-HT3 receptor activation, leading to a positive inotropic response [119]. However, it must be highlighted that these findings do not necessarily imply that similar results can be obtained at the vascular sensory neuroeffector level, as illustrated with the case of the 5-HT7 receptor.
3.3. Clinical relevance
Apart from the well-established therapeutic relevance of acute antimigraine serotonergic drugs inhibiting trigeminal CGRPergic transmission by 5-HT1B/1D/1F receptor activation [11,95,103], little attention has been paid to the interaction between 5-HT and the perivascular sensory nerves modulating systemic vascular responses (i.e., changes in arterial blood pressure). Admittedly, this is partly because there is no consensus on the pivotal role of CGRP in maintaining blood pressure [82,120]. In addition, since the pharmacology of serotonergic transmission is complex at the peripheral and central levels, the (cardio)vascular effects resulting from activation of the different 5-HT receptors at both levels are hard to explain [4,13,14,16].
Regarding perivascular CGRPergic transmission on the systemic vasculature, some findings seem to exclude the relevance of this neuropeptide in regulating blood pressure since acute CGRP receptor blockade in anaesthetized rats does not significantly impact blood pressure levels [83,120,121]. Accordingly, resting blood pressure is not affected in transgenic mice lacking the CGRP receptor [122]; conversely, continuous recording of blood pressure (in CGRP receptor KO mice) showed that this parameter is globally increased by an enhancement of the sympathetic autonomic function [123]. Indeed, in pithed rats (where central compensatory cardiovascular reflexes are excluded since the CNS in not functional), acute pharmacological blockade of the CGRP receptor with olcegepant not only inhibits the vasodepressor sensory CGRPergic outflow elicited electrically, but also enhanced the sympathetic vasopressor responses [81]. These data may imply that continuous blockade of CGRPergic vascular transmission with olcegepant (or any other CGRP antagonist) could favour a hypertensive state [81]. Indeed, although this seemed absent in clinical trials [124], real-world studies now suggest that the use of CGRP (receptor) blocking medications may increase blood pressure [125]. As elsewhere discussed [82,126,127], CGRP may play a physiological protective role in the cardiovascular system, but the relevance of CGRPergic transmission in blood pressure regulation is only unmasked under pathological cardiovascular alterations [128,129].
In this regard, a decrease in CGRP levels has been observed in spontaneously hypertensive rats and humans with essential hypertension [130,131], and it has been suggested that a diminution of the perivascular CGRPergic innervation may play a role in the development of this pathology [132]. Beyond the use of selective ligands to activate or antagonize the different 5-HT receptor subtypes favouring vasodilatory or vasoconstrictor effects, we need to keep in mind that, globally, 5-HT produces vasopressor responses by activation of vascular 5-HT2A receptors [4,14,16]. Moreover, under vascular damage conditions (e.g., hypertension), the vasculature is more sensitive to 5-HT to cause contraction [14]. Thus, apart from an enhanced 5-HT-induced vasoconstriction in hypertensive subjects [14], we are tempted to suggest that the release of CGRP in these subjects may be diminished by activation of prejunctional 5-HT1B/1F and 5-HT7 receptors, favouring a pro-hypertensive state.
4. Future directions
Physiologically, blood pressure is regulated by changes in peripheral vascular tone (caused by resistance blood vessels) and cardiac output, and these parameters are homeostatically maintained by neuronal, humoral, and local mechanisms [133]. When considering the neurovascular junction, it is well known that vascular tone is modulated by: (i) autonomic sympathetic nerves, which produce vasoconstriction by noradrenaline release [133,134]; and (ii) primary sensory nerves, which produce vasodilatation by neuropeptides release, mainly CGRP [78,79,135,136,137,138]. Several mechanisms exist at the neurovascular junction to modulate the neuronal outflow to the blood vessels; one of these mechanisms is the serotonergic transmission. Certainly, 5-HT can generally modulate the autonomic and sensory outflows via prejunctional receptors (see Figure 1) to regulate blood pressure.
From a global perspective, the actions of 5-HT on haemodynamic parameters are complex and sometimes opposite, depending on the experimental conditions [13]. This may be explained in terms of the numerous sites of action to 5-HT, which include: (i) the CNS; (ii) autonomic ganglia; (iii) perivascular nerve terminals; (iv) endothelial cells; (v) smooth muscle cells; and (vi) the heart [4,11,45,46,54,139,140,141]. Consequently, rather than assigning a single cardiovascular function to 5-HT, it is clear that 5-HT exerts multiple cardiovascular actions [4].
As evident from the present review, 5-HT induces a plethora of complex, and sometimes opposing, actions in the cardiovascular system, which may be even further complicated by pathophysiological states, such as hypertension [13] and pain [142]. While this may seem an impediment for the exploration of the therapeutic potential of 5-HT ligands, this simultaneously also provides options (e.g., in hypertension, as reviewed by Watts and Davis [13]). Future research should focus on the significance of species differences, the (patho)physiological conditions that may affect the function of 5-HT and its receptors, as well as interindividual differences caused by gender, ethnic background, or age. Research on human differentiated tissues obtained from induced pluripotent stem cells (iPSCs) may be a valuable tool for the study of rare diseases and the influence of different (culture) conditions. Because of the many modulating roles of 5-HT in other systems, targeting specific 5-HT receptors may provide valuable novel therapeutic avenues, besides its currently known therapeutic applications.
Author Contributions
Conceptualization, A.G-H., B.A.M-C., A.MVDB., C.M.V.; investigation, A.G-H., B.A.M-C., A.MVDB., C.M.V.; writing-original draft preparation, A.G-H., B.A.M-C., A.MVDB., C.M.V.; writing-review and editing, A.G-H., B.A.M-C., A.MVDB., C.M.V. All authors have read and agreed to the final version of this work.
Funding
This work was supported by Consejo Nacional de Ciencia y Tecnología (CONACyT Mexico) to: (i) A.G-H. (Fondo Sectorial de Investigación para la Educación, Grant No. A1-S-23631); and (ii) C.M.V. (Grant No. 219707). Furthermore, C.M.V acknowledges the SEP-Cinvestav Research Support Fund (Grant No. 50) and B.A.M.C. appreciates the “Dirección General de Investigación y Posgrado” of the Autonomous University of Aguascalientes (Grant No. PIFF21-1).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.References
Data Availability Statement
The articles cited in this paper are available on PubMed®, UptoDate®, and Cochrane®.
Conflicts of interest
The authors have no relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the present manuscript.
References
- Barnes, N.M.; Ahern, G.P.; Becamel, C.; Bockaert, J.; Camilleri, M.; Chaumont-Dubel, S.; Claeysen, S.; Cunningham, K.A.; Fone, K.C.; Gershon, M.; et al. International Union of Basic and Clinical Pharmacology. CX. Classification of Receptors for 5-hydroxytryptamine; Pharmacology and Function. Pharmacol Rev 2021, 73, 310–520. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Liu, F. The Role of Serotonin beyond the Central Nervous System during Embryogenesis. Front Cell Neurosci 2017, 11, 74. [Google Scholar] [CrossRef] [PubMed]
- Villalón, C.M. Chapter Three - The role of serotonin receptors in the control of cardiovascular function. In The Serotonin System, Tricklebank, M.D., Daly, E., Eds.; Academic Press: 2019; pp. 45-61.
- Villalón, C.M.; Centurión, D. Cardiovascular responses produced by 5-hydroxytriptamine:a pharmacological update on the receptors/mechanisms involved and therapeutic implications. Naunyn Schmiedebergs Arch Pharmacol 2007, 376, 45–63. [Google Scholar] [CrossRef] [PubMed]
- Saxena, P.R.; Villalón, C.M. Cardiovascular effects of serotonin agonists and antagonists. Journal of cardiovascular pharmacology 1990, 15 Suppl 7, S17–34. [Google Scholar] [CrossRef]
- Hoyer, D.; Clarke, D.E.; Fozard, J.R.; Hartig, P.R.; Martin, G.R.; Mylecharane, E.J.; Saxena, P.R.; Humphrey, P.P. International Union of Pharmacology classification of receptors for 5-hydroxytryptamine (Serotonin). Pharmacol Rev 1994, 46, 157–203. [Google Scholar] [PubMed]
- Hoyer, D.; Hannon, J.P.; Martin, G.R. Molecular, pharmacological and functional diversity of 5-HT receptors. Pharmacol Biochem Behav 2002, 71, 533–554. [Google Scholar] [CrossRef]
- Saxena, P.R.; De Vries, P.; Villalón, C.M. 5-HT1-like receptors: a time to bid goodbye. Trends Pharmacol Sci 1998, 19, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Saxena, P.R.; Villalón, C.M. 5-Hydroxytryptamine: a chameleon in the heart. Trends Pharmacol Sci 1991, 12, 223–227. [Google Scholar] [CrossRef]
- Villalón, C.M.; Centurión, D.; Valdivia, L.F.; de Vries, P.; Saxena, P.R. Migraine: pathophysiology, pharmacology, treatment and future trends. Curr Vasc Pharmacol 2003, 1, 71–84. [Google Scholar] [CrossRef]
- Villalón, C.M.; VanDenBrink, A.M. The Role of 5-Hydroxytryptamine in the Pathophysiology of Migraine and its Relevance to the Design of Novel Treatments. Mini Rev Med Chem 2017, 17, 928–938. [Google Scholar] [CrossRef]
- Kaumann, A.J.; Levy, F.O. 5-hydroxytryptamine receptors in the human cardiovascular system. Pharmacology & therapeutics 2006, 111, 674–706. [Google Scholar] [CrossRef]
- Watts, S.W.; Davis, R.P. 5-hydroxtryptamine receptors in systemic hypertension: an arterial focus. Cardiovasc Ther 2011, 29, 54–67. [Google Scholar] [CrossRef] [PubMed]
- Watts, S.W.; Morrison, S.F.; Davis, R.P.; Barman, S.M. Serotonin and blood pressure regulation. Pharmacol Rev 2012, 64, 359–388. [Google Scholar] [CrossRef] [PubMed]
- González-Hernández, A.; Marichal-Cancino, B.A.; Lozano-Cuenca, J.; López-Canales, J.S.; Muñoz-Islas, E.; Ramírez-Rosas, M.B.; Villalón, C.M. Heteroreceptors Modulating CGRP Release at Neurovascular Junction: Potential Therapeutic Implications on Some Vascular-Related Diseases. Biomed Res Int 2016, 2016, 2056786. [Google Scholar] [CrossRef]
- Ramage, A.G.; Villalón, C.M. 5-hydroxytryptamine and cardiovascular regulation. Trends Pharmacol Sci 2008, 29, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Ramage, A.G. Influence of 5-HT1A receptor agonists on sympathetic and parasympathetic nerve activity. Journal of cardiovascular pharmacology 1990, 15 Suppl 7, S75–85. [Google Scholar] [CrossRef]
- Ramage, A.G. Central cardiovascular regulation and 5-hydroxytryptamine receptors. Brain Res Bull 2001, 56, 425–439. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Lopez, A.; Centurión, D.; Vázquez, E.; Arulmani, U.; Saxena, P.R.; Villalón, C.M. Pharmacological profile of the 5-HT-induced inhibition of cardioaccelerator sympathetic outflow in pithed rats: correlation with 5-HT1 and putative 5-ht5A/5B receptors. Br J Pharmacol 2003, 140, 725–735. [Google Scholar] [CrossRef]
- García-Pedraza, J.; Hernández-Abreu, O.; García, M.; Morán, A.; Villalón, C.M. Chronic 5-HT(2) receptor blockade unmasks the role of 5-HT(1F) receptors in the inhibition of rat cardioaccelerator sympathetic outflow. Can J Physiol Pharmacol 2018, 96, 328–336. [Google Scholar] [CrossRef]
- Sánchez-López, A.; Centurión, D.; Vázquez, E.; Arulmani, U.; Saxena, P.R.; Villalón, C.M. Further characterization of the 5-HT1 receptors mediating cardiac sympatho-inhibition in pithed rats: pharmacological correlation with the 5-HT1B and 5-HT1D subtypes. Naunyn Schmiedebergs Arch Pharmacol 2004, 369, 220–227. [Google Scholar] [CrossRef]
- García-Pedraza, J.; García, M.; Martín, M.L.; Gómez-Escudero, J.; Rodríguez-Barbero, A.; Román, L.S.; Morán, A. Peripheral 5-HT₁D and 5-HT₇ serotonergic receptors modulate sympathetic neurotransmission in chronic sarpogrelate treated rats. Eur J Pharmacol 2013, 714, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Dabiré, H. Central 5-hydroxytryptamine (5-HT) receptors in blood pressure regulation. Therapie 1991, 46, 421–429. [Google Scholar] [PubMed]
- Bedi, U.S.; Arora, R. Cardiovascular manifestations of posttraumatic stress disorder. J Natl Med Assoc 2007, 99, 642–649. [Google Scholar]
- Tania, V.; Catherine, V. Roles of the Serotoninergic System in Coping with Traumatic Stress. In Serotonin and the CNS, Berend, O., Ed.; IntechOpen: Rijeka, 2021. [Google Scholar]
- Paine, N.J.; Watkins, L.L.; Blumenthal, J.A.; Kuhn, C.M.; Sherwood, A. Association of depressive and anxiety symptoms with 24-hour urinary catecholamines in individuals with untreated high blood pressure. Psychosom Med 2015, 77, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Brindley, R.L.; Bauer, M.B.; Blakely, R.D.; Currie, K.P.M. An interplay between the serotonin transporter (SERT) and 5-HT receptors controls stimulus-secretion coupling in sympathoadrenal chromaffin cells. Neuropharmacology 2016, 110, 438–448. [Google Scholar] [CrossRef]
- Nakatani, Y.; Sato-Suzuki, I.; Tsujino, N.; Nakasato, A.; Seki, Y.; Fumoto, M.; Arita, H. Augmented brain 5-HT crosses the blood-brain barrier through the 5-HT transporter in rat. Eur J Neurosci 2008, 27, 2466–2472. [Google Scholar] [CrossRef]
- Wang, H.M.; Wang, Y.; Liu, M.; Bai, Y.; Zhang, X.H.; Sun, Y.X.; Wang, H.L. Fluoxetine inhibits monocrotaline-induced pulmonary arterial remodeling involved in inhibition of RhoA-Rho kinase and Akt signalling pathways in rats. Can J Physiol Pharmacol 2012, 90, 1506–1515. [Google Scholar] [CrossRef]
- Lin, J.C.; Chou, C.C.; Tu, Z.; Yeh, L.F.; Wu, S.C.; Khoo, K.H.; Lin, C.H. Characterization of protein serotonylation via bioorthogonal labeling and enrichment. J Proteome Res 2014, 13, 3523–3529. [Google Scholar] [CrossRef]
- Penumatsa, K.C.; Fanburg, B.L. Transglutaminase 2-mediated serotonylation in pulmonary hypertension. American journal of physiology. Lung cellular and molecular physiology 2014, 306, L309–315. [Google Scholar] [CrossRef]
- Tjurmina, O.A.; Armando, I.; Saavedra, J.M.; Goldstein, D.S.; Murphy, D.L. Exaggerated adrenomedullary response to immobilization in mice with targeted disruption of the serotonin transporter gene. Endocrinology 2002, 143, 4520–4526. [Google Scholar] [CrossRef]
- Tiradentes, R.V.; Pires, J.G.; Silva, N.F.; Ramage, A.G.; Santuzzi, C.H.; Futuro Neto, H.A. Effects of acute administration of selective serotonin reuptake inhibitors on sympathetic nerve activity. Braz J Med Biol Res 2014, 47, 554–559. [Google Scholar] [CrossRef] [PubMed]
- Schroeter, S.; Levey, A.I.; Blakely, R.D. Polarized expression of the antidepressant-sensitive serotonin transporter in epinephrine-synthesizing chromaffin cells of the rat adrenal gland. Mol Cell Neurosci 1997, 9, 170–184. [Google Scholar] [CrossRef] [PubMed]
- Furlan, A.; Dyachuk, V.; Kastriti, M.E.; Calvo-Enrique, L.; Abdo, H.; Hadjab, S.; Chontorotzea, T.; Akkuratova, N.; Usoskin, D.; Kamenev, D.; et al. Multipotent peripheral glial cells generate neuroendocrine cells of the adrenal medulla. Science 2017, 357. [Google Scholar] [CrossRef] [PubMed]
- Kameneva, P.; Melnikova, V.I.; Kastriti, M.E.; Kurtova, A.; Kryukov, E.; Murtazina, A.; Faure, L.; Poverennaya, I.; Artemov, A.V.; Kalinina, T.S.; et al. Serotonin limits generation of chromaffin cells during adrenal organ development. Nature Communications 2022, 13, 2901. [Google Scholar] [CrossRef] [PubMed]
- Carbone, E.; Borges, R.; Eiden, L.E.; García, A.G.; Hernández-Cruz, A. Chromaffin Cells of the Adrenal Medulla: Physiology, Pharmacology, and Disease. Compr Physiol 2019, 9, 1443–1502. [Google Scholar] [CrossRef]
- Brindley, R.L.; Bauer, M.B.; Blakely, R.D.; Currie, K.P.M. Serotonin and Serotonin Transporters in the Adrenal Medulla: A Potential Hub for Modulation of the Sympathetic Stress Response. ACS Chem Neurosci 2017, 8, 943–954. [Google Scholar] [CrossRef]
- Linder, A.E.; Beggs, K.M.; Burnett, R.J.; Watts, S.W. Body distribution of infused serotonin in rats. Clin Exp Pharmacol Physiol 2009, 36, 599–601. [Google Scholar] [CrossRef]
- Kobayashi, S.; Tsukahara, S.; Sugita, K.; Nagata, T. Adrenergic and cholinergic innervation of rat cerebral arteries. Consecutive demonstration on whole mount preparations. Histochemistry 1981, 70, 129–138. [Google Scholar] [CrossRef]
- Sheng, Y.; Zhu, L. The crosstalk between autonomic nervous system and blood vessels. Int J Physiol Pathophysiol Pharmacol 2018, 10, 17–28. [Google Scholar]
- Koep, J.L.; Taylor, C.E.; Coombes, J.S.; Bond, B.; Ainslie, P.N.; Bailey, T.G. Autonomic control of cerebral blood flow: fundamental comparisons between peripheral and cerebrovascular circulations in humans. The Journal of physiology 2022, 600, 15–39. [Google Scholar] [CrossRef]
- Suzuki, N.; Hardebo, J.E. The cerebrovascular parasympathetic innervation. Cerebrovasc Brain Metab Rev 1993, 5, 33–46. [Google Scholar] [PubMed]
- Roloff, E.v.L.; Tomiak-Baquero, A.M.; Kasparov, S.; Paton, J.F.R. Parasympathetic innervation of vertebrobasilar arteries: is this a potential clinical target? The Journal of physiology 2016, 594, 6463–6485. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.E.; Salvatierra, A.T. Apposition of enkephalin- and neurotensin-immunoreactive neurons by serotonin-immunoreactive varicosities in the rat spinal cord. Neuroscience 1998, 85, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Villalón, C.M.; Centurión, D.; Rabelo, G.; de Vries, P.; Saxena, P.R.; Sánchez-López, A. The 5-HT1-like receptors mediating inhibition of sympathetic vasopressor outflow in the pithed rat: operational correlation with the 5-HT1A, 5-HT1B and 5-HT1D subtypes. Br J Pharmacol 1998, 124, 1001–1011. [Google Scholar] [CrossRef] [PubMed]
- Rapport, M.M.; Green, A.A.; Page, I.H. Serum vasoconstrictor, serotonin; isolation and characterization. J Biol Chem 1948, 176, 1243–1251. [Google Scholar] [CrossRef] [PubMed]
- Rapport, M.M.; Green, A.A.; Page, I.H. Partial purification of the vasoconstrictor in beef serum. J Biol Chem 1948, 174, 735–741. [Google Scholar] [CrossRef] [PubMed]
- Herr, N.; Bode, C.; Duerschmied, D. The Effects of Serotonin in Immune Cells. Front Cardiovasc Med 2017, 4, 48. [Google Scholar] [CrossRef]
- Palermo, A.; del Rosso, G.; Costantini, C.; Bertalero, P.; Rizzi, S.; Libretti, A. Platelet content of serotonin and response to stress. J Hypertens Suppl 1986, 4, S43–45. [Google Scholar]
- Teff, K.L.; Young, S.N. Effects of carbohydrate and protein administration on rat tryptophan and 5-hydroxytryptamine: differential effects on the brain, intestine, pineal, and pancreas. Can J Physiol Pharmacol 1988, 66, 683–688. [Google Scholar] [CrossRef]
- Edvinsson, J.C.A.; Maddahi, A.; Christiansen, I.M.; Reducha, P.V.; Warfvinge, K.; Sheykhzade, M.; Edvinsson, L.; Haanes, K.A. Lasmiditan and 5-Hydroxytryptamine in the rat trigeminal system; expression, release and interactions with 5-HT1 receptors. The Journal of Headache and Pain 2022, 23, 26. [Google Scholar] [CrossRef]
- Ishida, Y.; Sugiura, Y.; Magome, T.; Kamakura, T.; Takimoto, Y.; Hanada, Y.; Kitayama, K.; Nakamura, Y.; Shimada, S.; Ohta, N.; et al. Expression Analysis of Serotonin Receptors, Serotonin Transporter and l-Amino Acid Decarboxylase in the Mouse Sphenopalatine Ganglion by RT-PCR, Northern Blot Analysis and In Situ Hybridization. Neuroscience 2019, 411, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Punda, H.; Mardesic, S.; Filipovic, N.; Kosovic, I.; Benzon, B.; Ogorevc, M.; Bocina, I.; Kolic, K.; Vukojevic, K.; Saraga-Babic, M. Expression Pattern of 5-HT (Serotonin) Receptors during Normal Development of the Human Spinal Cord and Ganglia and in Fetus with Cervical Spina Bifida. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Satoh, S. Presynaptic inhibition by serotonin of cardiac sympathetic transmission in dogs. Clin Exp Pharmacol Physiol 1983, 10, 535–542. [Google Scholar] [CrossRef] [PubMed]
- García-Pedraza, J.; Hernández-Abreu, O.; Morán, A.; Carretero, J.; García-Domingo, M.; Villalón, C.M. Role of peripheral 5-HT(5A) receptors in 5-HT-induced cardiac sympatho-inhibition in type 1 diabetic rats. Sci Rep 2020, 10, 19358. [Google Scholar] [CrossRef] [PubMed]
- Morán, A.; Fernández, M.M.; Velasco, C.; Martín, M.L.; San Román, L. Characterization of prejunctional 5-HT1 receptors that mediate the inhibition of pressor effects elicited by sympathetic stimulation in the pithed rat. Br J Pharmacol 1998, 123, 1205–1213. [Google Scholar] [CrossRef] [PubMed]
- Pilowsky, P.M. Chapter 16 - Serotonin in Central Cardiovascular Regulation: Ex Uno Plura (From One Comes Many). In Serotonin, Pilowsky, P.M., Ed.; Academic Press: Boston, 2019; pp. 335–347. [Google Scholar]
- Morán, A.; Velasco, C.; Salvador, T.; Martín, M.L.; San Román, L. Inhibitory 5-hydroxytryptamine receptors involved in pressor effects obtained by stimulation of sympathetic outflow from spinal cord in pithed rats. Br J Pharmacol 1994, 113, 1358–1362. [Google Scholar] [CrossRef] [PubMed]
- Villamil-Hernández, M.T.; Alcántara-Vázquez, O.; Sánchez-López, A.; Gutiérrez-Lara, E.J.; Centurión, D. Pharmacological evidence that 5-HT1A/1B/1D, α2-adrenoceptors and D2-like receptors mediate ergotamine-induced inhibition of the vasopressor sympathetic outflow in pithed rats. Eur J Pharmacol 2014, 740, 512–521. [Google Scholar] [CrossRef]
- Marichal-Cancino, B.A.; González-Hernández, A.; Muñoz-Islas, E.; Villalón, C.M. Monoaminergic Receptors as Modulators of the Perivascular Sympathetic and Sensory CGRPergic Outflows. Curr Neuropharmacol 2020, 18, 790–808. [Google Scholar] [CrossRef]
- Molderings, G.J.; Frölich, D.; Likungu, J.; Göthert, M. Inhibition of noradrenaline release via presynaptic 5-HT1D alpha receptors in human atrium. Naunyn Schmiedebergs Arch Pharmacol 1996, 353, 272–280. [Google Scholar] [CrossRef]
- Morán, A.; Velasco, C.; Martín, M.L.; San Román, L. Pharmacological characterization of 5-HT receptors in parasympathetic innervation of rat heart. Eur J Pharmacol 1994, 252, 161–166. [Google Scholar] [CrossRef]
- Lee, T.J.; Liu, J.; Evans, M.S. Cholinergic-nitrergic transmitter mechanisms in the cerebral circulation. Microsc Res Tech 2001, 53, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Jackowski, A.; Crockard, A.; Burnstock, G. 5-Hydroxytryptamine demonstrated immunohistochemically in rat cerebrovascular nerves largely represents 5-hydroxytryptamine uptake into sympathetic nerve fibres. Neuroscience 1989, 29, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Boyle, S.H.; Brummett, B.H.; Kuhn, C.M.; Barefoot, J.C.; Siegler, I.C.; Williams, R.B.; Georgiades, A. The Effects of Tryptophan Enhancement and Depletion on Plasma Catecholamine Levels in Healthy Individuals. Psychosom Med 2019, 81, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Švec, J.; Švec, P.; Bencová, V.; Krčméry, V. [Anxio-depressive Syndrome - Biopsychosocial Model of Supportive Care]. Klin Onkol 2015, 28, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Razzaque, Z.; Pickard, J.D.; Ma, Q.P.; Shaw, D.; Morrison, K.; Wang, T.; Longmore, J. 5-HT1B-receptors and vascular reactivity in human isolated blood vessels: assessment of the potential craniovascular selectivity of sumatriptan. Br J Clin Pharmacol 2002, 53, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Villalón, C.M.; Sánchez-López, A.; Centurión, D. Operational characteristics of the 5-HT1-like receptors mediating external carotid vasoconstriction in vagosympathectomized dogs. Naunyn-Schmiedeberg's Archives of Pharmacology 1996, 354, 550–556. [Google Scholar] [CrossRef]
- Gonzalez-Hernandez, A.; Marichal-Cancino, B.A.; MaassenVanDenBrink, A.; Villalon, C.M. Side effects associated with current and prospective antimigraine pharmacotherapies. Expert Opin Drug Metab Toxicol 2018, 14, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Marichal-Cancino, B.A.; González-Hernández, A.; Guerrero-Alba, R.; Medina-Santillán, R.; Villalón, C.M. A critical review of the neurovascular nature of migraine and the main mechanisms of action of prophylactic antimigraine medications. Expert Rev Neurother 2021, 1–16. [Google Scholar] [CrossRef]
- Zaidi, M.; Bevis, P.J.; Girgis, S.I.; Lynch, C.; Stevenson, J.C.; MacIntyre, I. Circulating CGRP comes from the perivascular nerves. Eur J Pharmacol 1985, 117, 283–284. [Google Scholar] [CrossRef]
- Escott, K.J.; Connor, H.E.; Brain, S.D.; Beattie, D.T. The involvement of calcitonin gene-related peptide (CGRP) and substance P in feline pial artery diameter responses evoked by capsaicin. Neuropeptides 1995, 29, 129–135. [Google Scholar] [CrossRef]
- Van der Schueren, B.J.; de Hoon, J.N.; Vanmolkot, F.H.; Van Hecken, A.; Depre, M.; Kane, S.A.; De Lepeleire, I.; Sinclair, S.R. Reproducibility of the capsaicin-induced dermal blood flow response as assessed by laser Doppler perfusion imaging. Br J Clin Pharmacol 2007, 64, 580–590. [Google Scholar] [CrossRef] [PubMed]
- Dux, M.; Rosta, J.; Messlinger, K. TRP Channels in the Focus of Trigeminal Nociceptor Sensitization Contributing to Primary Headaches. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Julius, D.; Basbaum, A.I. Molecular mechanisms of nociception. Nature 2001, 413, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, H.; Nuki, C.; Saito, A.; Takasaki, K. Adrenergic modulation of calcitonin gene-related peptide (CGRP)-containing nerve-mediated vasodilation in the rat mesenteric resistance vessel. Brain research 1990, 506, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, H.; Takasaki, K.; Saito, A.; Goto, K. Calcitonin gene-related peptide acts as a novel vasodilator neurotransmitter in mesenteric resistance vessels of the rat. Nature 1988, 335, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, T.; Kawasaki, H.; Imamura, T.; Takasaki, K. Endogenous calcitonin gene-related peptide mediates nonadrenergic noncholinergic depressor response to spinal cord stimulation in the pithed rat. Circulation research 1992, 71, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, H.; Saito, A.; Takasaki, K. Age-related decrease of calcitonin gene-related peptide-containing vasodilator innervation in the mesenteric resistance vessel of the spontaneously hypertensive rat. Circulation research 1990, 67, 733–743. [Google Scholar] [CrossRef]
- Avilés-Rosas, V.H.; Rivera-Mancilla, E.; Marichal-Cancino, B.A.; Manrique-Maldonado, G.; Altamirano-Espinoza, A.H.; Maassen Van Den Brink, A.; Villalón, C.M. Olcegepant blocks neurogenic and non-neurogenic CGRPergic vasodepressor responses and facilitates noradrenergic vasopressor responses in pithed rats. Br J Pharmacol 2017, 174, 2001–2014. [Google Scholar] [CrossRef]
- Russell, F.A.; King, R.; Smillie, S.J.; Kodji, X.; Brain, S.D. Calcitonin gene-related peptide: physiology and pathophysiology. Physiological reviews 2014, 94, 1099–1142. [Google Scholar] [CrossRef]
- Russo, A.F.; Hay, D.L. CGRP physiology, pharmacology, and therapeutic targets: migraine and beyond. Physiological reviews 2023, 103, 1565–1644. [Google Scholar] [CrossRef]
- Villalón, C.M.; Albarrán-Juárez, J.A.; Lozano-Cuenca, J.; Pertz, H.H.; Görnemann, T.; Centurión, D. Pharmacological profile of the clonidine-induced inhibition of vasodepressor sensory outflow in pithed rats: correlation with alpha(2A/2C)-adrenoceptors. Br J Pharmacol 2008, 154, 51–59. [Google Scholar] [CrossRef]
- González-Hernández, A.; Manrique-Maldonado, G.; Lozano-Cuenca, J.; Muñoz-Islas, E.; Centurión, D.; Maassen VanDenBrink, A.; Villalón, C.M. The 5-HT(1) receptors inhibiting the rat vasodepressor sensory CGRPergic outflow: further involvement of 5-HT(1F), but not 5-HT(1A) or 5-HT(1D), subtypes. Eur J Pharmacol 2011, 659, 233–243. [Google Scholar] [CrossRef] [PubMed]
- González-Hernández, A.; Muñoz-Islas, E.; Lozano-Cuenca, J.; Ramírez-Rosas, M.B.; Sánchez-López, A.; Centurión, D.; Ramírez-San Juan, E.; Villalón, C.M. Activation of 5-HT1B receptors inhibits the vasodepressor sensory CGRPergic outflow in pithed rats. Eur J Pharmacol 2010, 637, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Ibrahimi, K.; Danser, A.; Terwindt, G.M.; van den Meiracker, A.H.; MaassenVanDenBrink, A. A human trigeminovascular biomarker for antimigraine drugs: A randomised, double-blind, placebo-controlled, crossover trial with sumatriptan. Cephalalgia 2017, 37, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Benemei, S.; Cortese, F.; Labastida-Ramírez, A.; Marchese, F.; Pellesi, L.; Romoli, M.; Vollesen, A.L.; Lampl, C.; Ashina, M. Triptans and CGRP blockade - impact on the cranial vasculature. J Headache Pain 2017, 18, 103. [Google Scholar] [CrossRef] [PubMed]
- Labastida-Ramírez, A.; Rubio-Beltrán, E.; Haanes, K.A.; Chan, K.Y.; Garrelds, I.M.; Johnson, K.W.; Danser, A.H.J.; Villalón, C.M.; MaassenVanDenBrink, A. Lasmiditan inhibits calcitonin gene-related peptide release in the rodent trigeminovascular system. Pain 2020, 161, 1092–1099. [Google Scholar] [CrossRef] [PubMed]
- Buzzi, M.G.; Carter, W.B.; Shimizu, T.; Heath, H., 3rd; Moskowitz, M.A. Dihydroergotamine and sumatriptan attenuate levels of CGRP in plasma in rat superior sagittal sinus during electrical stimulation of the trigeminal ganglion. Neuropharmacology 1991, 30, 1193–1200. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Akerman, S.; van den Maagdenberg, A.M.; Saxena, P.R.; Goadsby, P.J.; van den Brink, A.M. Intravital microscopy on a closed cranial window in mice: a model to study trigeminovascular mechanisms involved in migraine. Cephalalgia 2006, 26, 1294–1303. [Google Scholar] [CrossRef]
- Limmroth, V.; Katsarava, Z.; Liedert, B.; Guehring, H.; Schmitz, K.; Diener, H.C.; Michel, M.C. An in vivo rat model to study calcitonin gene related peptide release following activation of the trigeminal vascular system. Pain 2001, 92, 101–106. [Google Scholar] [CrossRef]
- Williamson, D.J.; Hargreaves, R.J.; Hill, R.G.; Shepheard, S.L. Sumatriptan inhibits neurogenic vasodilation of dural blood vessels in the anaesthetized rat--intravital microscope studies. Cephalalgia 1997, 17, 525–531. [Google Scholar] [CrossRef]
- Ashina, M.; Hansen, J.M.; Do, T.P.; Melo-Carrillo, A.; Burstein, R.; Moskowitz, M.A. Migraine and the trigeminovascular system-40 years and counting. Lancet Neurol 2019, 18, 795–804. [Google Scholar] [CrossRef] [PubMed]
- de Vries, T.; Villalón, C.M.; MaassenVanDenBrink, A. Pharmacological treatment of migraine: CGRP and 5-HT beyond the triptans. Pharmacology & therapeutics 2020, 211, 107528. [Google Scholar] [CrossRef]
- Goadsby, P.J.; Holland, P.R.; Martins-Oliveira, M.; Hoffmann, J.; Schankin, C.; Akerman, S. Pathophysiology of Migraine: A Disorder of Sensory Processing. Physiological reviews 2017, 97, 553–622. [Google Scholar] [CrossRef]
- Gupta, S.; Villalón, C.M. The relevance of preclinical research models for the development of antimigraine drugs: focus on 5-HT(1B/1D) and CGRP receptors. Pharmacology & therapeutics 2010, 128, 170–190. [Google Scholar] [CrossRef]
- Moreno-Ajona, D.; Chan, C.; Villar-Martínez, M.D.; Goadsby, P.J. Targeting CGRP and 5-HT(1F) Receptors for the Acute Therapy of Migraine: A Literature Review. Headache 2019, 59 Suppl 2, 3–19. [Google Scholar] [CrossRef]
- Lozano-Cuenca, J.; González-Hernández, A.; Muñoz-Islas, E.; Sánchez-López, A.; Centurión, D.; Cobos-Puc, L.E.; Villalón, C.M. Effect of some acute and prophylactic antimigraine drugs on the vasodepressor sensory CGRPergic outflow in pithed rats. Life sciences 2009, 84, 125–131. [Google Scholar] [CrossRef]
- Nicholson, R.; Small, J.; Dixon, A.K.; Spanswick, D.; Lee, K. Serotonin receptor mRNA expression in rat dorsal root ganglion neurons. Neurosci Lett 2003, 337, 119–122. [Google Scholar] [CrossRef]
- Muñoz-Islas, E.; Gupta, S.; Jiménez-Mena, L.R.; Lozano-Cuenca, J.; Sánchez-López, A.; Centurión, D.; Mehrotra, S.; MaassenVanDenBrink, A.; Villalón, C.M. Donitriptan, but not sumatriptan, inhibits capsaicin-induced canine external carotid vasodilatation via 5-HT1B rather than 5-HT1D receptors. Br J Pharmacol 2006, 149, 82–91. [Google Scholar] [CrossRef]
- Muñoz-Islas, E.; Lozano-Cuenca, J.; González-Hernández, A.; Ramírez-Rosas, M.B.; Sánchez-López, A.; Centurión, D.; Maassenvandenbrink, A.; Villalón, C.M. Spinal sumatriptan inhibits capsaicin-induced canine external carotid vasodilatation via 5-HT1B rather than 5-HT1D receptors. Eur J Pharmacol 2009, 615, 133–138. [Google Scholar] [CrossRef]
- Rubio-Beltrán, E.; Labastida-Ramírez, A.; Villalón, C.M.; MaassenVanDenBrink, A. Is selective 5-HT(1F) receptor agonism an entity apart from that of the triptans in antimigraine therapy? Pharmacology & therapeutics 2018, 186, 88–97. [Google Scholar] [CrossRef]
- Fujii, H.; Takatori, S.; Zamami, Y.; Hashikawa-Hobara, N.; Miyake, N.; Tangsucharit, P.; Mio, M.; Kawasaki, H. Adrenergic stimulation-released 5-HT stored in adrenergic nerves inhibits CGRPergic nerve-mediated vasodilatation in rat mesenteric resistance arteries. Br J Pharmacol 2012, 166, 2084–2094. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Mancilla, B.; Cutler, N.R.; Leibowitz, M.T.; Spierings, E.L.; Klapper, J.A.; Diamond, S.; Goldstein, J.; Smith, T.; Couch, J.R.; Fleishaker, J.; et al. Safety and efficacy of PNU-142633, a selective 5-HT1D agonist, in patients with acute migraine. Cephalalgia 2001, 21, 727–732. [Google Scholar] [CrossRef] [PubMed]
- González-Hernández, A.; Marichal-Cancino, B.A.; Lozano-Cuenca, J.; MaassenVanDenBrink, A.; Villalón, C.M. Functional Characterization of the Prejunctional Receptors Mediating the Inhibition by Ergotamine of the Rat Perivascular Sensory Peptidergic Drive. ACS Chem Neurosci 2019, 10, 3173–3182. [Google Scholar] [CrossRef] [PubMed]
- de Jong, A.P.; Verhage, M. Presynaptic signal transduction pathways that modulate synaptic transmission. Current opinion in neurobiology 2009, 19, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Cuesta, C.; García-Pedraza, J.; García, M.; Villalón, C.M.; Morán, A. Role of 5-HT7 receptors in the inhibition of the vasodepressor sensory CGRPergic outflow in pithed rats. Vascul Pharmacol 2014, 63, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.K.; von der Weid, P.Y. 5-HT decreases contractile and electrical activities in lymphatic vessels of the guinea-pig mesentery: role of 5-HT 7-receptors. Br J Pharmacol 2003, 139, 243–254. [Google Scholar] [CrossRef]
- Chan, M.F.; Okun, I.; Stavros, F.L.; Hwang, E.; Wolff, M.E.; Balaji, V.N. Identification of a new class of ETA selective endothelin antagonists by pharmacophore directed screening. Biochem Biophys Res Commun 1994, 201, 228–234. [Google Scholar] [CrossRef]
- Wiklund, N.P.; Wiklund, C.U.; Cederqvist, B.; Ohlén, A.; Hedqvist, P.; Gustafsson, L.E. Endothelin modulation of neuroeffector transmission in smooth muscle. Journal of cardiovascular pharmacology 1991, 17 Suppl 7, S335–339. [Google Scholar] [CrossRef]
- Filippelli, A.; Falciani, M.; Piucci, B.; D'Amico, M.; D'Agostino, B.; Filippelli, W.; Rossi, F. Endothelin-1 affects capsaicin-evoked release of neuropeptides from rat vas deferens. Eur J Pharmacol 1999, 364, 183–191. [Google Scholar] [CrossRef]
- Brenchat, A.; Zamanillo, D.; Hamon, M.; Romero, L.; Vela, J.M. Role of peripheral versus spinal 5-HT(7) receptors in the modulation of pain undersensitizing conditions. Eur J Pain 2012, 16, 72–81. [Google Scholar] [CrossRef]
- Chen, J.J.; Vasko, M.R.; Wu, X.; Staeva, T.P.; Baez, M.; Zgombick, J.M.; Nelson, D.L. Multiple subtypes of serotonin receptors are expressed in rat sensory neurons in culture. J Pharmacol Exp Ther 1998, 287, 1119–1127. [Google Scholar] [PubMed]
- Pierce, P.A.; Xie, G.X.; Meuser, T.; Peroutka, S.J. 5-Hydroxytryptamine receptor subtype messenger RNAs in human dorsal root ganglia: a polymerase chain reaction study. Neuroscience 1997, 81, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Sugiuar, T.; Bielefeldt, K.; Gebhart, G.F. TRPV1 function in mouse colon sensory neurons is enhanced by metabotropic 5-hydroxytryptamine receptor activation. J Neurosci 2004, 24, 9521–9530. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Chen, T.; Gao, Y.; Quirion, R.; Hong, Y. Inhibition of SNL-induced upregulation of CGRP and NPY in the spinal cord and dorsal root ganglia by the 5-HT(2A) receptor antagonist ketanserin in rats. Pharmacol Biochem Behav 2012, 101, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Islas, E.; Vidal-Cantú, G.C.; Bravo-Hernández, M.; Cervantes-Durán, C.; Quiñonez-Bastidas, G.N.; Pineda-Farias, J.B.; Barragán-Iglesias, P.; Granados-Soto, V. Spinal 5-HT₅A receptors mediate 5-HT-induced antinociception in several pain models in rats. Pharmacol Biochem Behav 2014, 120, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Tramontana, M.; Giuliani, S.; Del Bianco, E.; Lecci, A.; Maggi, C.A.; Evangelista, S.; Geppetti, P. Effects of capsaicin and 5-HT3 antagonists on 5-hydroxytryptamine-evoked release of calcitonin gene-related peptide in the guinea-pig heart. Br J Pharmacol 1993, 108, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Smillie, S.J.; Brain, S.D. Calcitonin gene-related peptide (CGRP) and its role in hypertension. Neuropeptides 2011, 45, 93–104. [Google Scholar] [CrossRef]
- Arulmani, U.; Schuijt, M.P.; Heiligers, J.P.; Willems, E.W.; Villalón, C.M.; Saxena, P.R. Effects of the calcitonin gene-related peptide (CGRP) receptor antagonist BIBN4096BS on alpha-CGRP-induced regional haemodynamic changes in anaesthetised rats. Basic Clin Pharmacol Toxicol 2004, 94, 291–297. [Google Scholar] [CrossRef]
- Lu, J.T.; Son, Y.J.; Lee, J.; Jetton, T.L.; Shiota, M.; Moscoso, L.; Niswender, K.D.; Loewy, A.D.; Magnuson, M.A.; Sanes, J.R.; et al. Mice lacking alpha-calcitonin gene-related peptide exhibit normal cardiovascular regulation and neuromuscular development. Mol Cell Neurosci 1999, 14, 99–120. [Google Scholar] [CrossRef]
- Mai, T.H.; Wu, J.; Diedrich, A.; Garland, E.M.; Robertson, D. Calcitonin gene-related peptide (CGRP) in autonomic cardiovascular regulation and vascular structure. Journal of the American Society of Hypertension : JASH 2014, 8, 286–296. [Google Scholar] [CrossRef]
- Kudrow, D.; Pascual, J.; Winner, P.K.; Dodick, D.W.; Tepper, S.J.; Reuter, U.; Hong, F.; Klatt, J.; Zhang, F.; Cheng, S.; et al. Vascular safety of erenumab for migraine prevention. Neurology 2020, 94, e497–e510. [Google Scholar] [CrossRef] [PubMed]
- de Vries Lentsch, S.; van der Arend, B.W.H.; Maassen VanDenBrink, A.; Terwindt, G.M. Blood Pressure in Patients With Migraine Treated With Monoclonal Anti-CGRP (Receptor) Antibodies: A Prospective Follow-up Study. Neurology 2022, 99, e1897–e1904. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Potts, J.D.; DiPette, D.J. Protective Role of α-Calcitonin Gene-Related Peptide in Cardiovascular Diseases. Frontiers in physiology 2019, 10, 821. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Williamson, M.; Hess, A.; DiPette, D.J.; Potts, J.D. Alpha-Calcitonin Gene Related Peptide: New Therapeutic Strategies for the Treatment and Prevention of Cardiovascular Disease and Migraine. Frontiers in physiology 2022, 13, 826122. [Google Scholar] [CrossRef] [PubMed]
- MaassenVanDenBrink, A.; Meijer, J.; Villalón, C.M.; Ferrari, M.D. Wiping Out CGRP: Potential Cardiovascular Risks. Trends Pharmacol Sci 2016, 37, 779–788. [Google Scholar] [CrossRef] [PubMed]
- MaassenVanDenBrink, A.; Terwindt, G.M.; van den Maagdenberg, A. Calcitonin gene-related peptide (receptor) antibodies: an exciting avenue for migraine treatment. Genome Med 2018, 10, 10. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.Y.; Yang, Y.; Zhao, Y.T. [Plasma calcitonin gene-related peptide (CGRP) level in patients with essential hypertension]. Zhonghua Nei Ke Za Zhi 1990, 29, 616–618. [Google Scholar] [PubMed]
- Xu, D.; Wang, X.A.; Wang, J.P. [Calcitonin gene-related peptide in normotensive and spontaneously hypertensive rats]. Zhonghua Xin Xue Guan Bing Za Zhi 1989, 17, 174–177. [Google Scholar] [CrossRef]
- Watson, R.E.; Supowit, S.C.; Zhao, H.; Katki, K.A.; Dipette, D.J. Role of sensory nervous system vasoactive peptides in hypertension. Braz J Med Biol Res 2002, 35, 1033–1045. [Google Scholar] [CrossRef]
- Guyton, A.C.; Coleman, T.G.; Cowley, A.V., Jr.; Scheel, K.W.; Manning, R.D., Jr.; Norman, R.A., Jr. Arterial pressure regulation. Overriding dominance of the kidneys in long-term regulation and in hypertension. Am J Med 1972, 52, 584–594. [Google Scholar] [CrossRef]
- Hoffman, B.B. Catecholamines, sympathomimetic drugs, and adrenergic receptor antagonists. In Goodman & Gilman´s The Pharmacological Basis of Therapeutics (10th Edition). Hardman JG, L.L., Goodman Gilman A (eds), Ed.; McGraw-Hill: New York, 2001; pp. pp. 215-268.
- Gardiner, S.M.; Compton, A.M.; Bennett, T. Regional hemodynamic effects of calcitonin gene-related peptide. Am J Physiol 1989, 256, R332–338. [Google Scholar] [CrossRef] [PubMed]
- Han, S.P.; Naes, L.; Westfall, T.C. Calcitonin gene-related peptide is the endogenous mediator of nonadrenergic-noncholinergic vasodilation in rat mesentery. J Pharmacol Exp Ther 1990, 255, 423–428. [Google Scholar] [PubMed]
- Han, S.P.; Naes, L.; Westfall, T.C. Inhibition of periarterial nerve stimulation-induced vasodilation of the mesenteric arterial bed by CGRP (8-37) and CGRP receptor desensitization. Biochem Biophys Res Commun 1990, 168, 786–791. [Google Scholar] [CrossRef] [PubMed]
- Holzer, P. Local effector functions of capsaicin-sensitive sensory nerve endings: involvement of tachykinins, calcitonin gene-related peptide and other neuropeptides. Neuroscience 1988, 24, 739–768. [Google Scholar] [CrossRef] [PubMed]
- Machida, T.; Iizuka, K.; Hirafuji, M. 5-hydroxytryptamine and its receptors in systemic vascular walls. Biol Pharm Bull 2013, 36, 1416–1419. [Google Scholar] [CrossRef]
- Marichal-Cancino, B.A.; Gonzalez-Hernandez, A.; Manrique-Maldonado, G.; Ruiz, S., II; Altamirano-Espinoza, A.H.; MaassenVanDenBrink, A.; Villalon, C.M. Intrathecal dihydroergotamine inhibits capsaicin-induced vasodilatation in the canine external carotid circulation via GR127935- and rauwolscine-sensitive receptors. Eur J Pharmacol 2012, 692, 69–77. [Google Scholar] [CrossRef]
- Sumner, M.J. Characterization of the 5-HT receptor mediating endothelium-dependent relaxation in porcine vena cava. Br J Pharmacol 1991, 102, 938–942. [Google Scholar] [CrossRef]
- Rivasi, G.; Menale, S.; Turrin, G.; Coscarelli, A.; Giordano, A.; Ungar, A. The Effects of Pain and Analgesic Medications on Blood Pressure. Curr Hypertens Rep 2022, 24, 385–394. [Google Scholar] [CrossRef]
Figure 1.
Prejunctional 5-HT receptors are involved in the inhibition of postganglionic autonomic and sensory CGRPergic function at the vascular level. Generally, 5-HT can inhibit the release of noradrenaline, acetylcholine, and CGRP via activation of the 5-HT1 receptor family (coupled to Gi/o proteins; this figure shows the corresponding Gα/β/γ subunits). In the case of the parasympathetic outflow, activation of 5-HT3 (ligand-gated ion channel) receptors favours the release of acetylcholine. Furthermore, in sensory CGRPergic neurones, prejunctional activation of 5-HT7 receptors seems to recruit the endothelin system (via an unknown pathway), favouring the activation of the ET1 receptor and promoting inhibition of CGRP release. Interestingly: (i) in some isolated cases, activation of prejunctional 5-HT3 receptors on parasympathetic fibres facilitates the release of CGRP; and (ii) circulating 5-HT can be recaptured via NET, and subsequently vesiculated and released upon electrical stimulation of the sympathetic outflow. See text for details. AP: action potential; NET: noradrenaline transporters; VGCC: voltage-gated ion channels.
Figure 1.
Prejunctional 5-HT receptors are involved in the inhibition of postganglionic autonomic and sensory CGRPergic function at the vascular level. Generally, 5-HT can inhibit the release of noradrenaline, acetylcholine, and CGRP via activation of the 5-HT1 receptor family (coupled to Gi/o proteins; this figure shows the corresponding Gα/β/γ subunits). In the case of the parasympathetic outflow, activation of 5-HT3 (ligand-gated ion channel) receptors favours the release of acetylcholine. Furthermore, in sensory CGRPergic neurones, prejunctional activation of 5-HT7 receptors seems to recruit the endothelin system (via an unknown pathway), favouring the activation of the ET1 receptor and promoting inhibition of CGRP release. Interestingly: (i) in some isolated cases, activation of prejunctional 5-HT3 receptors on parasympathetic fibres facilitates the release of CGRP; and (ii) circulating 5-HT can be recaptured via NET, and subsequently vesiculated and released upon electrical stimulation of the sympathetic outflow. See text for details. AP: action potential; NET: noradrenaline transporters; VGCC: voltage-gated ion channels.
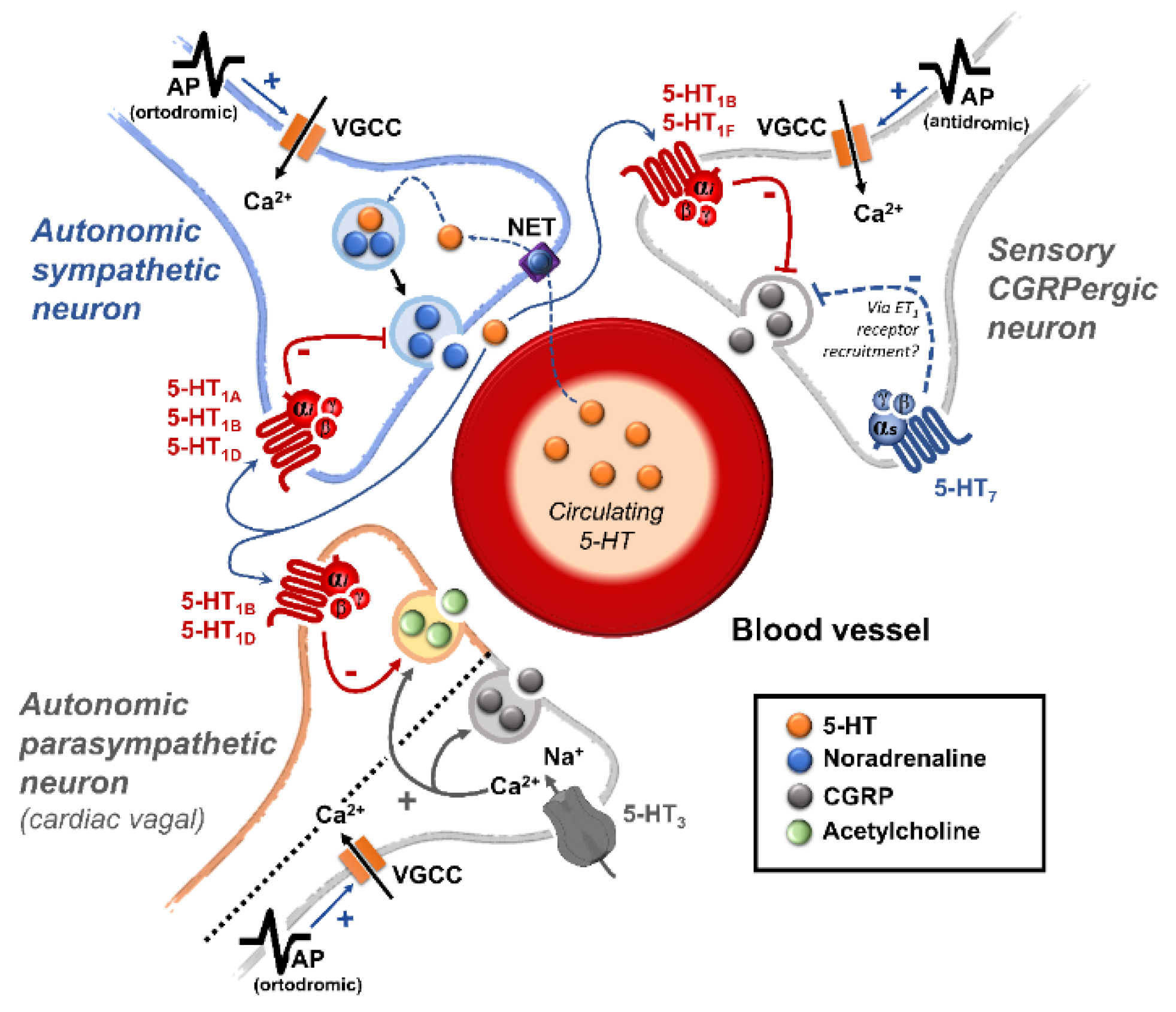

Table 1.
Classification of 5-HT receptors a.
| 5-HT receptor | Receptor subtype | Agonists | Antagonists | Some functions | Canonical transduction system |
|---|---|---|---|---|---|
| 5-HT1 | 5-HT1A | 8-OH-DPAT | WAY 100635 | Central hypotension |
G-protein coupled receptor (Gi)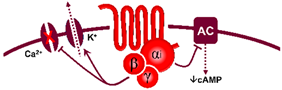
|
| 5-HT1B | Sumatriptan CP-93,129 (rodents) |
SB224289 | Vasoconstriction, sympatho-inhibition | ||
| 5-HT1D | PNU-109291 PNU-142633 |
BRL15572 | Autoreceptor, sympatho-inhibition | ||
| 5-HT1e* | 5-HT >> 5-CT LY334370 | Methiothepin (non-selective) | Unknown | ||
| 5-HTF | LY344864 lasmiditan, LY334370 | Methysergide (non-selective) | (-) Trigeminal system | ||
| 5-HT5 | 5-HT5A | 5-HT, ergotamine | SB699551 | Cardiac sympatho-inhibition in rats | |
| 5-HT5b* | 5-CT (non-selective) | Unknown | Unknown | ||
| 5-HT4 | - | Renzapride BIMU8, ML10302 SC53116 | GR 113808 SB204070 | (+) Neuronal activity, vasodilatation, tachycardia in pigs and humans |
G-protein coupled receptor (Gs)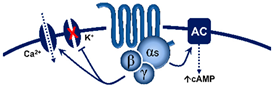
|
| 5-HT6 | - | 5-MeO-T ≥ 5-HTSB357134 SB271046 | Ro 630563 | Memory, not involved in cardiovascular regulation by 5-HT | |
| 5-HT7 | - | 5-CT>>5-HT AS-19 |
SB269970 SB258719 | Circadian rhythm, vasodilatation, tachycardia in cats | |
| 5-HT2 | 5-HT2A | DOI, DOB α-methyl-5-HT |
MDL100907 ketanserin | Vasoconstriction, platelet aggregation |
G-protein coupled receptor (Gq)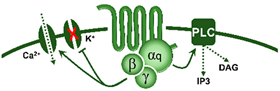
|
| 5-HT2B | DOI, BW723C86α-methyl-5-HT | SB204741 RS-127445 |
Vasoconstriction, release of NO | ||
| 5-HT2C | DOI, Ro 60-0175α-methyl-5-HT | SB242084 RS-102221 |
CSF production | ||
| 5-HT3 | Pentameric ion channel** | Phenylbiguanideα-methyl-5-HT | Tropisetron Granisetron MDL-72222 | (+) Neuronal activity, reflex bradycardia |
Ligand-gated ion channel
|
Modified from Villalón and Centurión (2007) and Villalón (2019). AS-19, (2S)-(+)-5-(1,3,5-Trimethylpyrazol-4-yl)-2-(dimethylamino)tetralin; CNS, central nervous system; CSF, cerebrospinal fluid; LSD, lysergic acid diethylamide; 5-MeOT, 5-methoxytryptamine; 5-CT, 5-carboxamidotryptamine; DOI, 1-(2,5-dimethoxy-4-iodophenyl)-2-aminopropane; NO, nitric oxide; (-), inhibits; (+), stimulates. * Lowercase used by convention to denote a receptor whose functional role (i.e., in native cells or tissues) has not been elucidated. ** Five known subunits have been described (5-HT3A – 5-HT3E) forming homomeric or heteromeric complexes. At least two subunits of 5-HT3A type are required to form a functional ion channel. a The pharmacological profile of each 5-HT receptor type is identified with the application of inclusion and exclusion criteria, as explained in section 1.1.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



