
Submitted:
26 May 2023
Posted:
29 May 2023
You are already at the latest version
Abstract
Many studies link changes in avian phenology in Europe to the North Atlantic Oscillation (NAO), as a proxy of conditions at western Europe. But effects of climate variation in other regions of Eu-rope on phenology of short-distance migrants with wide non-breeding grounds remain unclear. We determined the combined influence of large-scale climate indices, NAO, the Mediterranean Oscillation Index (MOI), and the Scandinavian Pattern (SCAND), during the preceding year on spring migration timing of European Wren at the southern Baltic coast during 1982–2021. We modelled the effects of these climate variables on the entire passage and subsequent percentiles of Wren’s passage at Bukowo-Kopań and Hel ringing stations. The start and median of migration shifted earlier at Hel, but the end of passage shifted later at both stations over 1982–2021. In the effect, the duration of passage at Hel extended by 7.6 days. Early passage at Hel was related with high MOI1 in spring and in preceding autumn. Spring passage at Bukowo-Kopań was late after high NAO in the previous breeding season, and high winter and spring NAO. At both stations late spring passage occurred after high SCAND in previous summer. Early beginning or median of passage at our stations followed high local temperatures. We conclude that phenology of Wren’s spring migration at the Baltic coast was shaped by conditions that different populations encounter at wintering quarters in western Europe, where NAO operates, and in the south-eastern Europe, where the MOI1 operates, in combination with conditions in Scandinavia during previous breed-ing season. We showed that climate variability in different parts of the migrants’ range has com-bined carry-over effects on in migrant’s phenology in Europe.
Keywords:
climate change
; spring phenology
; short–distance migrants
; European Wren
; Troglodytes troglodytes
; MOI
; NAO
; SCAND
1. Introduction
Average global temperature has risen by more than 1°C since the beginning of the XXth century and continues to increase [1], affecting life cycles of the world’s flora and fauna at multiple levels. Birds are excellent indicators of the impact of climate change on animals [2] because they occur in all climates, and many species undertake seasonal migrations, and show plasticity in response to environmental conditions [3]. Changing climate has been shown to cause advancement of spring migration in many birds, in response to increase of temperatures on their breeding grounds [4], along migratory routes [5,6,7,8,9] or at wintering sites [10,11,12]. Numerous studies link such shifts in migrants’ phenology with climatic variability in the northern hemisphere, reflected by such large-scale climate indices as the North Atlantic Oscillation Index (NAO) [13,14,15,16]. NAO has been used as a predictor of spring conditions in Europe, because it reflects temperature and precipitation [17], which are main drivers of advancement of the breeding season in Europe [4,18]. Also Scandinavian Pattern (SCAND), which is a proxy of weather conditions over the Scandinavia, has been related to migrants’ timing [19,20,21]. Large-scale indices of climate in Africa, such as the Sahel Precipitation Index (SPI), Indian Ocean Dipole (IOD) or Southern Oscillation Index (SOI), were also shown to influence birds’ phenology, especially in long-distance migrants that winter in areas where these indices operate [21,22,23,24,25]. But the Mediterranean Oscillation Index (MOI), the predictor of climatic conditions in the Mediterranean region, where many European birds overwinter or stopover, has rarely been considered in the context of bird migrations. This is so probably because this climate index has been proposed only in 1989, and its influence on climate in southern Europe intertwines with that of NAO [26]. These large-scale indices reflect ecological conditions that migrants encounter at their breeding grounds, migration routes, or wintering areas, which may have carry-over effects on their subsequent life-stages [21,27,28]. Many passerines have advanced the beginning and middle of their spring passage across Europe over the recent decades [9,20,29,30]. In combination with a delayed autumn migration, the residence period on the breeding grounds has extended in some European species [28,29,31], which may favour increased number of broods [28,32,33]. Shifts to earlier spring arrivals over the last decades have been most pronounced for short-distance migrants [15,16,17,34]. In addition, Forchhammer et al. [35] showed that conditions related with NAO at the wintering grounds had greater influence on arrival in Norway for short-distance migrants than for long-distance ones. Conditions at the wintering grounds provide short-distance migrants with a cue to adjust their departure to conditions on the breeding grounds, which are relatively close [17]. For this reason, larger fluctuations in migration phenology in Europe have been observed in this group, than in long-distance migrants, for which conditions at their distant wintering grounds in Africa are poor predictors of the weather on their spring arrival in Europe [17,36,37]. Many studies have focused on the effect of climatic variability on phenology of long-distance passerine migrants [4,6,21,38,39,40]. But fewer studies attempted to identify carry-over effects of conditions at breeding and non-breeding areas on migration timing in species with shorter migrations but wide wintering quarters in Europe [12,30,41]. To fill in this gap, we set out to identify annual variation and any long-term trends in spring migration phenology over 40 years in a regular short-distance migrant, the European Wren Troglodytes troglodytes, at the southern coast of Baltic, Poland. We also aimed to determine any relationships between this species’ spring phenology and three large-scale indices, which reflect climate across Europe, at breeding and non-breeding grounds of this species.
2. Materials and Methods
2.1. Study Species
The European Wren (henceforth Wren), is a small insectivorous passerine common in deciduous woodlands throughout Europe [42]. Populations breeding at the north and east of the species’ range are migratory, those from western Europe are partially sedentary, and populations of the southern part of the range and endemic subspecies of Atlantic Islands are sedentary [43]. Our study focuses on the northern populations, which usually migrate for about 1000–1500 km, exceptionally as far as about 2500 km, which we chose as an example of a regular short-distance migrant (Figure 1) [44]. Scandinavian breeding populations winter mostly in southern Sweden, central and western Europe, and rarely on the Iberian Peninsula and in Great Britain (Figure 1) [44,45,46,47]. Wrens that migrate in spring and autumn through the southern coast of the Baltic Sea, originate from Fennoscandia, and the Baltic countries, with most ringing recoveries from Sweden. They overwinter mostly in western and central Europe, and in the Apennine Peninsula, but some ringing recoveries suggest they also winter in the Balkans [44,48,49]. In Poland, including the Baltic coast, spring passage of Wrens occurs in March–April [50]. At the breeding grounds in southern Sweden, the first individuals appear in the second decade of April, and in northern Sweden from mid-May [45]. The breeding season in Scandinavia begins in May and lasts until July (Figure 2) [45,46]. Wrens usually raise one clutch, but sometimes have two to three broods [43,46]. They depart from the northern breeding grounds in August–September, reach non-breeding grounds by the end of October, and stay there until February–early March [43,45,46]. Thus, for the Wren, we considered „spring migration” as March–April, „breeding” as May–July, „autumn migration” as August–October, and „non–breeding period” as November–February (Figure 2).
2.2. Study Sitesand Material
We set out to analyse the timing of spring migration of the Wren at the southern coast of the Baltic Sea in 1982–2021. We used data from two ringing stations at the Polish coast (Figure 1): Bukowo–Kopań (54°20′13″N, 16°14′36″E) and Hel (54°44′25″N, 18°33′49″E). At both stations birds were caught during spring migration (26 March–15 May), according to standardised monitoring protocol [52]. This study period covered the entire migration of Wrens through the Polish coast. Migrants were captured daily, from dusk to dawn, using 8m-long mist-nets. The number of mist-nets at each station was stable through each season, but varied from 35 to 58 between the years. After ringing, each bird was aged, measured and weighed, using the established fieldwork protocol [52]. Sexing Wrens was not possible in field, because they show no sexual dimorphism in size or plumage [53,54]. In spring, most adult and immature Wrens look the same, only some immatures are detectable by plumage [53,54], thus we treated all age groups jointly in this study.
2.3. Climate Indices
We used 11 large-scale climate indices, reflecting weather conditions on summer breeding grounds (SCAND, NAO), spring migration routes (SCAND, NAO, MOI1) and on the wintering areas (NAO, MOI1) of Wren, and the local spring temperature at each ringing station over 1981-2021 (Table 1). We averaged values of these indices for periods of months corresponding to subsequent stages in the Wren’s year (Figure 2), and we used these averaged indices relevant to the regions where Wrens should stay in each period. Thus, for example we selected the Scandinavian Pattern (SCAND) for March–April, May–July, and August–October (Figure 2), i.e. the periods when Wrens are likely to stay within the influence of this climate pattern, but we excluded SCAND for November–December, when most Wrens stay at their wintering grounds in southern Europe (Figure 1), away of the effect of SCAND. The large-scale climate indices use differences in the sea level pressure between two geographically distant weather stations to reflect weather patterns, which manifest in temperature, precipitation, wind direction and cloudiness over large areas [55]. The positive phase of SCAND in spring (March–May) and summer (July–August) corresponds with high average temperature and dry conditions over Scandinavia, and low temperature with rainy conditions over central and western Europe and central Russia [19,55,56]. SCAND in March–April and May–June showed a significant decreasing trend over our study period, but no long-time trend occurred for SCAND in July–August (Figure A1). Positive North Atlantic Oscillation Index (NAO) is related with warm winter (November–February) and spring (March–May) over western Europe, moist conditions between Iceland and Scandinavia, including western Europe, and with dry conditions over Greenland, central and southern Europe [26]. In summer (June–August), the positive NAO is correlated with dry and warm weather over northern Europe [57]. NAO in July–August showed a significant decreasing trend over 1981-2021, but no clear trends for others months (Figure A1). Remisiewicz & Underhill [21] provided more details on the effects of these large-scale on long-distance migrant birds. The Mediterranean Oscillation Index (MOI1) we used as a proxy for conditions of the eastern Mediterranearn region was calculated as the difference in the normalised air pressure at the sea level between Algiers and Cairo [58,59,60]. Positive values of MOI1 in November–February and March–May are related with low rainfall, high temperature and consequently mild winters and springs in the eastern Mediterranean area [26,60]. MOI1 showed no long-term trends for any of the periods of months we distinguished (Figure A1).
Additionally, as a proxy for local spring conditions at the ringing stations, we used the mean daily temperatures averaged for March–April from the nearest weather stations (Table 1). We used these temperatures as the monthly anomalies in relation to the 1982–2020 baseline for March and April, respectively, which we then averaged for both months. For Bukowo–Kopań (54°20′13″N, 16°14′36″E) we used temperature anomalies averaged for two nearby weather stations in Koszalin (54°12′15″N,16°09′19″) and Łeba (54°45′12″N, 17°32′04″) (TBK, Table 1). For the Hel ringing station, we used temperatures from the weather station in the Hel Harbour (54°36′12″N, 18°48′42″E) (THL, Table 1). We used the monthly anomalies averaged for these weather stations provided by Climate Explorer facility (https://climexp.knmi.nl/), based on the European Climate Assessment and Dataset (http://www.ecad.eu). These spring temperatures for Bukowo-Kopań and Hel increased by 1.2°C and 1.1°C, respectively, over 1982–2021 (Figure A1).
2.4. Statistical Analysis
We used the daily numbers and spring totals of Wrens caught at each station, including only the first capture of an individual in a season (Table A1). For single days when mistnetting was suspended because of storms or other obstacles, the missing daily totals were imputed with the numbers estimated based on the numbers of Wrens caught at this date during six earlier and six later years at that station, as in other studies [20,21,61,62]. Imputations considered only 1–4 days in 9 seasons at Hel and in 13 seasons at Bukowo-Kopań. The analysis did not cover springs of 2011 and 2020 at Bukowo-Kopań, when this station did not operate, because of logistics or COVID–19 constraints. Years when less than 10 birds were caught during spring at a station were excluded from the study. Thus, we analysed data from 38 years at Bukowo-Kopań and 32 years at Hel, within the period 1982–2021 (Table A1).
For each station, based on daily catching totals, we calculated the Annual Anomaly (AA), which indicates the departure of the cumulative curve of the spring migration in one year from the many-year average curve (1982–2021), as in Remisiewicz and Underhill [20]. We calculated AA for each spring, and compiled them in two time-series, one for each station, which reflected variation in overall spring migration timing over 1982–2021. To investigate the timing of subsequent phases of passage in more detail, we calculated the dates (Julian day) when the first 5% (beginning of passage), 50% (middle) and 95% (end of passage) of Wrens occurred every spring at each station, as in other studies [8,9,29]. In this way, for each station we obtained time-series over 1982–2021 for four measures of migration timing: AA, and the dates of 5%, 50% and 95% of passage. We also calculated the duration of migration at each station, as the difference between the dates for 5% and 95% of passage in each year. We compared these parameters of passage between stations over 1982–2021 using Mann-Whitney test. We also used simple linear regression models to check for any significant trends in all the four measures of migration timing, and for migration duration, at each station over 1982–2021.
To explore the effects of climate indices on Wren’s passage at each station, we used all four measures as response variables for Bukowo–Kopań and Hel separately in multiple regression models against „Year” and 12 climate factors, which were the explanatory variables. These climate variables included 11 large-scale climate indices and one index of local temperature relevant for each station, TBK or THL, respectively (Table 1). The indices of climate in spring came from the same year as the analysed spring migration. But for the climate indices that operate at wintering grounds, autumn migration routes and the breeding grounds, where Wrens occur three to ten months prior to the spring migration in question, we used the data from the previous year (Table 1). All explanatory and response variables, which were in different scales, were standardised to have the mean of 0 and standard deviation of 1, except for the „Year”, which was used as the year number since 1982. All climate variables were used in regression models in linear terms, as in the previous papers [20,21,63].
Before running any multiple regression models we used Pearson correlation coefficient to check for any strong correlations between climate variables (|r| ≥ 0.7), which could cause multicollinearity [64]. For most climate variables correlation coefficients, if ever significant, were below this threshold (Table A2), but MOI1 and NAO in November–February were correlated at r = 0.74 (Table A2). However, these two indices did not occur together in any variant of the resulting models, so we decided to include them both into the initial full models to allow the model selection procedure to indicate which of these variables had a stronger relation to Wren’s passage.
We selected the best model based on „all subsets regression” and ranking the models by the Akaike Information Criteria (AICc). For the best models, with the lowest AICc, we calculated partial correlation coefficients (pR) to check the direction and strength of the correlation between the response variable and each explanatory variable, while excluding the effects of the other variables. The proportion of variation explained by each best model was reflected by the adjusted coefficient of determination (AdjR2). We also calculated Variance Inflation Factors (VIFs) for explanatory variables in each best model, to monitor for any effects of multicollinearity [64]. Statistical analyses were conducted in R 4.2.2 [65] and Statistica 13.3 [66].
3. Results
3.1. Long-Term Trends in Timing of Wren Spring Passage at the southern Baltic Sea Coast
The overall spring migration (AA) of Wren showed a tendency to shift earlier by 1.7 days on average at Bukowo–Kopań (Figure 3A) and 2.7 days at Hel (Figure 3B) over the 40 years we studied, but these trends were not significant (Table A3). The timing of passage showed large year-to-year variation (Figure 3A,B), which was correlated between the stations (rS =0.42, p =0.02). At Bukowo-Kopań the start of passage (5%) was delayed by 0.02 days, the middle (50%) advanced on average by 5.3 days, but the end (95%) shifted later by 2.3 days (Figure 3C) over 1982–2021, thus the duration of passage extended also by 2.3 days (Figure A2), but none of these trends was significant (Table A3). At Hel, the start of Wren’s passage shifted earlier by 3.6 days, the middle by 5.0 days, but the end of passage shifted later by 3.9 days over the 40 years of our study. These trends were not significant (Figure 3D, Table A3), but the duration of spring passage at Hel extended significantly (t = 2.20, p = 0.04) on average by 7.6 days, in the effect of advance of the initial phase of passage and the delay of its final phase (Table A3, Figure A2). The dates of 5%, 50% and 95% of passage were on average 2–4 days later at Hel than at Bukowo–Kopań (Table A3, A4), and the difference was the largest for the start (5%) of passage (Mann–Whitney test, Z =6.24, p<0.01).
3.2. Climate Indices and the Timing of Spring Passage
At Bukowo-Kopań the overall spring passage (AA) was later with the higher values of NAO in July–August of the preceding breeding period, which explained AdjR2 = 15% of variation (Figure 4A, Table 2). But MOI1 was not selected for any best model, in contrast to Hel (Table 2). At Bukowo-Kopań the timing of the first phase of passage (5%), analogously as the whole passage, was later with the higher NAO and SCAND in July–August of the previous year, which jointly explained AdjR2 = 28% of its variation (Table 2). The median date of passage (50%) was later with higher SCAND in May–June and NAO in July–August of the previous year, and NAO in November–February and March–April, but earlier with higher MOI1, SCAND and local temperature during spring migration in March–April (Table 2, Figure 4A). The best model including all these climate variables explained AdjR2 =40% of the variation (Table 2). The end (95%) of passage was later with high NAO in July–August and with low SCAND in July–August of the previous year; this model explained only AdjR2 =10% variation (Table 2, Figure 4A).
At Hel the overall timing of passage was earlier with higher spring MOI1 in March–April, and with lower SCAND in July–August of the previous year; this best model explained AdjR2 = 16% in variation of the AA, but did not include NAO (Table 2, Figure 4B). The initial phase of passage (5%) was earlier with higher spring local temperatures at Hel (in March–April), and the previous last years’ autumn MOI1 (September–October), and with lower SCAND in May–July the previous year; these factors jointly explained AdjR2 =39% of the variation. The median date of passage (50%) was the later the higher was last years’ the previous summer’s SCAND (July–August), which explained AdjR2 =14% of the variation. The end of passage (95%) was earlier with high spring MOI1 (March–April), but this model explained only AdjR2 = 1% of the variation (Table 2, Figure 4B). All VIFs <10 in all the best models (Table 2) indicated no collinearity between the selected variables.
4. Discussion
We revealed large year-to-year variation of Wren’s spring migration at the southern Baltic coast, but no many-year trends for any advance or delay of passage. Spring migration at the more western Bukowo-Kopań station was shaped mainly by NAO during the previous breeding period, and by winter and spring NAO. In contrast, the passage of Wrens at Hel responded mostly to spring and autumnMOI1, but not to NAO. We will suggest an explanation for these differences in responses of Wren’s passage to these climate factors at two coastal stations located only 160 km apart, in the context of results from other ringing stations in northern Europe. At both stations late spring passage occurred after high SCAND during breeding season, and the beginning of passage at Hel, and the median at Bukowo-Kopań were early with high local spring temperatures. We will discuss how these climate factors, operating mostly at breeding and wintering grounds, might have a carry-over effect on Wren’s spring migration timing at the Baltic coast three to ten months later.
4.1. Multi–year Trends in the Timing of Wren Spring Migration
A number of European passerines [3,16,29,68,69], as well as North American species [9,70,71], including House Wren Troglodytes aedon [72], advanced their spring migration timing. Analogous shifts to earlier spring arrivals have occurred also in the European Wren, based on data from the other parts of the Baltic region [8,73], and from the Helgoland Island in the south-eastern part of the North Sea [14]. At Helgoland, the mean date of Wren’s spring passage advanced significantly over 1960–2000 [14], but we found only small advances in the median date of passage over 1982–2021 at both our stations, located 550–700 km to the east (Figure 1). A slight advance of the passage of Wren on the southern Baltic coast we found is similar to that in Ottenby station at the Baltic island of Öland (Figure 1) in 1971–2002 [69]. However, our results differ from those at Christiansø Island (Figure 1), where both beginning (5%) and end (95%) of Wren’s passage advanced significantly over 1976–1997 [8], but in our study the end of passage was slightly delayed. In our study the most pronounced advance was that of the median (50%) of migration, but at Christiansø the shift of the median was the least pronounced shift of all the percentiles. These discrepancies can be explained in several ways. Firstly, the study period at our stations extends over more recent years than the periods analysed for the other stations [8,14,69,73]. Many-year trends may differ depending on the range of years selected for analyses, which would include various combinations of early and late migrations, as demonstrated for the Song Thrush Turdus philomelos migration in the Baltic region [30,74]. Considering large inter-annual variation of passage in Wren (Figure 3), late passage in several springs within 2011–2021 likely caused a lack of clear trends for earlier arrivals, in contrast to trends from earlier periods at other stations. Secondly, proportions of various migratory populations, each having its own many-year trends and fluctuations, likely differ between distant stopover locations, and our two stations are located more to the east than Christiansø [8] and the other compared sites (Figure 1).Thirdly, before crossing ecological barriers, such as open sea, birds intensively refuel. This can cause mixing of different populations at stopover sites, and may affect the timing of their further migration, which might cause discrepancies between coastal and island sites [75,76,77]. Also, adults and immatures differ in their propensity to cross ecological barriers, which leads to the „coastal effect”, i.e. greater proportion of immatures observed at the coast than in land [78]. Thus, we suspect that different proportions of age groups might cause differences in the timing of passage between the coastal (Bukowo-Kopań, Hel) and the island (Christiansø, Ottenby) stations in the Baltic region. However, we have no clear evidence to confirm this assumption, considering that ageing of Wrens is problematic in spring [53,54].
The few days later occurrence of the corresponding phases of passage at Hel than at Bukowo–Kopań, might reflect the time that Wrens need to cross the extra ca 160 km to the west to reach Hel (Figure 1), considering they migrate slowly [76]. An analogous time difference in spring migration was found for the European Robin Erithacus rubecula at these two stations [79,80]. But this shift in migration timing might also be a result of different proportions of various migratory populations of Wrens passing through both stations, which we will discuss later.
The delayed end of migration on Hel significantly extended the duration of passage, which did not occur at Bukowo-Kopań. A tendency towards extended duration of spring passage is relatively rare among European migratory birds. But a tendency to advanced beginning and delayed end of spring migration, and thus to prolonged passage, occurred in some long-distance migrants, such as Pied Flycatcher Ficedula hypoleuca and Red–backed Shrike Lanius collurio at Christiansø [8]. For the Song Thrush Turdus philomelos, which has a similar migration distance and routes as Wren [74], the end of spring passage at the Baltic coast of Poland was delayed after warm December–February at their wintering grounds in south–western Europe [30]. We suspect a similar mechanism of delaying the end of passage in Wrens, in response to conditions at their wintering grounds.
4.2. Effects of Conditions on Spring Migration Routes on the Timing of Spring Passage at the Baltic Coast
The earlier migration of Wrens with higher local spring temperatures we found at both stations (Table2, Figure 4A,B), is consistent with the overall trend of migrants to shift spring migration earlier in response to increasing temperatures in Europe [6,9,14,15,17]. This trend mainly considers short-distance migrants [15,17,23], including our study species. With high local temperatures vegetation is early, allowing for efficient foraging, which in turn may shorten stopover duration [35,81], thus the passage through subsequent stations would also occur earlier and quicker. With warm conditions and insects abundance, birds mobility will also be greater than on cold and rainy days, increasing the likelihood of catching them in mistnets [82]. Local temperatures at Bukowo-Kopań (TBK) and Hel (THL) are related also to the conditions birds meet earlier on the spring migration routes, as temperatures are correlated over larger regions of Europe [83,84]. The timing of arrivals of the first 5% of Wrens at Hel (Figure 4B) and the 50% at Bukowo-Kopań(Figure 4A) were related with local temperatures, as in other studies [8,23]. But the overall migration (AA) at both stations was related rather with the indices of climate at earlier stages of Wren’s spring migration, and in the previous breeding season, which suggests the local spring temperatures only fine-tune the timing of passage, as it was described in other species [17,21].
Spring passage of Wrens at the southern coast of Baltic was earlier with higher spring MOI1 (March–April), which reflects with warm and dry weather in the eastern Mediterranean area [26,59]. Such conditions would promote early and fast passage farther north, and thus early arrivals at the Baltic coast. The overall migration of Wren at Hel, and in particular the end of their passage, were early with high spring MOI (Figure 4A), and thus warm spring on route from the eastern Mediterranean. Such conditions would create favourable feeding conditions on stopovers, which would enable even immatures and weakest individuals, which migrate as last to arrive relatively early at the Baltic coast [77,85]. Analogously, warm and dry conditions in the eastern Mediterranean would explain the early median of passage through Bukowo–Kopań with high spring MOI1 (Figure 4B).
Spring MOI1 reflects climate over the eastern Mediterranean area, but less so in western Europe [26], which in the spring is rather under climatic influence of NAO. Both indices were not correlated in spring (Table A2). Stronger influence of spring MOI1 on overall passage at Hel than at Bukowo-Kopań corresponds with the pattern of ringing recoveries of Wren, which suggests that Hel supports larger proportion of individuals wintering in the eastern Mediterranean region, than Bukowo-Kopań, where the populations wintering in south-western Europe seem to prevail [44,48,49]. This pattern is also in line with the relationship of spring NAO (March–April) with the median date of passage at Bukowo-Kopań, but not with timing of migration at Hel (Figure 4). Surprisingly, high spring NAO (March–April) was related with late median of passage at Bukowo–Kopań (Figure 4). Positive phase of NAO brings warm and wet conditions over most of continental Europe [86]. Some nocturnal species, like thrushes, avoid departing stopover sites during rainfall [87], thus such conditions on migration route may also delay Wren migration.
The positive phase of spring SCAND (March–April) was related with the late beginning of passage at Hel (Figure 4B). In winter and early spring positive SCAND results in low temperatures in western Europe and high precipitation over central and southern Europe [19]. These conditions might be unfavourable for passage [88], and delay Wrens departures from these regions, and thus their arrival at Hel. As spring progresses, the centre of influence of the positive phase of SCAND shifts southwards,which brings warm and dry conditions to western Europe [89]. This change might explain early median of Wren migration at Bukowo-Kopań with high SCAND in March–April (Figure 4A).
4.3. Effects of Conditions on Wintering Grounds on the Timing of Spring Passage
Winter NAO has been shown to influence spring migration of many European short-distance migrants, like Goldcrest Regulus regulus, Song Trush Turdus philomelos or Skylark Alauda arvensis, as well as long-distance migrants, such as Willow Warbler Phylloscopus trochilus or Lesser Whitethroat Curruca curruca [10,35,62]. With positive winter NAO, which indicates mild and wet winters and early spring in western and central Europe [14,69,90], migration of majority of these passerines in spring at various locations in Europe was also early [14,21,69]. Favourable conditions on wintering grounds improve fitness of migrants, which enables them to depart early and reach the breeding grounds quickly in spring [14,91]. Mild winters may also cause shortening of migration distance, and thus early arrivals at the north, as suitable wintering conditions might occur closer to the breeding grounds than during cold winters [10,92]. In contrast to these studies, which emphasize dearly spring migration of Wren after positive winter NAO [14,69], we found that the median date (50%) of passage at Bukowo-Kopań was late with high NAO in November–February. The explanation might be that mild winter conditions, related with positive winter NAO, may favour survival of unexperienced and weak individuals, especially from late broods [43]. The increased proportion of immatures and weak individuals which survived mild winter well and arrive at Bukowo-Kopań later than the fittest birds, may delay the bulk of passage and thus move the median late, as in the Song Thrush at the Baltic coast [30]. This influence of winter NAO suggests that the wintering grounds of Wren populations migrating through Bukowo-Kopań are located mainly in south-western Europe. We found no analogous influence of winter NAO on Wren’s passage at Hel, which implies that populations that pass through that station are likely to winter mainly outside the influence of NAO [14,69], likely in the more south-easterly part of Europe, where the climate is shaped by MOI1 [26,60].
4.4. Carry-over Effects of Conditions at the Breeding Grounds on the Following Spring Passage
We found that with positive values of indices of climate at the breeding grounds in Scandinavia, such as NAO and SCAND in summer (May–June and July–August), the overall migration, and most phases of Wren’s passage were late in spring next year at both our stations. The only exception was an early end (95%) of passage at Hel with positive SCAND in July–August. SCAND shows strong association with conditions in early summer [19], which is the time of the first and second broods in Wren [43]. NAO has the most pronounced effect on conditions in late summer (July–August) in north-western and central Europe, which is the period of second and third broods [43], but has the weakest influence on weather in June [57]. Our results are consistent with these patterns, as the SCAND in May–June and July–August was related with timing of passage the next spring at both our stations (Figure 4A,B).
High pressure in the continental Europe and the Azores correlated with positive NAO, and the center of air circulation over Scandinavia [21], in summer are both related with high temperatures and low precipitation on Wrens’ breeding grounds in the Baltic region and further east. High SCAND and NAO in summer would thus favour early breeding and provide suitable conditions for Wrens to undertake additional broods, which would increase the numbers of juveniles [18,93], and thereby the abundance of their migration next spring [94]. Youngs from early broods have more time to mature and prepare for autumn migration, than those from later broods, and thus greater chances to take a winter territory, which would further increase their survival rate [95,96]. This might result in numerous arrivals of these individuals in springs after summers with high NAO and SCAND (Figure 4A,B). Immatures usually migrate in spring later than more experienced adults [88]. Thus, numerous spring returns of youngs after warm summers, would result in later timing of overall migration, and of its subsequent phases, than in springs after cold summers and fewer immatures, as in the Song Thrush [30]. Favourable conditions on the breeding grounds may improve fitness of immatures and adults [4], which would enable some individuals to reach farther wintering grounds, from where they will arrive later, than after cold and wet summers. At Hel, the effect of summer SCAND (July–August) on spring passage was well pronounced, but we found no influence of summer NAO (Figure 4B). In contrast, the late summer NAO (July–August) had the strongest influence of all climate variables on spring passage at Bukowo-Kopań (Figure 4A), but some effect of SCAND in May–June and July–August was also pronounced for subsequent phases of passage at that station (Figure 4B). The influence of summer NAO on migration through Bukowo-Kopań but not at Hel, suggest that populations passing through these stations encounter different conditions on breeding grounds and thus they breeding range partly differ. This corresponds with the distribution of ringing recoveries, which suggests that at Bukowo-Kopań a greater proportion of Wrens originate from north-western breeding grounds, as Sweden and Norway (Figure 1), where NAO operates, than at Hel, which supports more birds breeding to the east, under greater climatic influence of SCAND [44,48,49].
The late median of passage at Bukowo-Kopań with positive of SCAND in previous May–June (Figure 4A), can be explained by a prolonged residence on the breeding grounds of youngs from early broods enabled by early and warm summer. Such good conditions may allow them to prepare well for autumn migration. As a consequence, some immatures might be able to migrate farther, and thus return in spring later than after unfavourable summers. The late end of passage at Bukowo–Kopań after positive NAO in July–August (Figure 4A) might be the result of the increased proportion of youngs from second broods arriving at the Baltic coast late next spring [96].
On the other hand, the end of migration at Bukowo-Kopań was early after positive SCAND in July–August. We consider that the last cohorts of migrants may include some proportion of a population originating from eastern Europe, where positive summer SCAND means low temperature and high precipitation [19,55,56]. It could have the opposite effect to July-August NAO influence, causing fewer broods, and low fitness and survival rate of immatures. However, given that conditions at the breeding grounds explained only 10% of variation in the dates of 95% at Bukowo-Kopań (Table 2, Figure 4A), we approach these results with caution.
4.5. Carry-over Effects of Conditions on Autumn Migration on the Following Spring Passage
We found that with high MOI1 in September–October, thus with high temperatures and low precipitation in the eastern Mediterranean region [60,97], the beginning of arrivals at Hel was early the following spring (Figure 4B). With recently observed changes of migratory behavior and great plasticity of short-distance European migrants [7,98,99], we consider shortening of migration distance as a plausible explanation. Mild autumn conditions may cause some individuals to overwinter closer to the breeding grounds [92,100,101], or to skip migration and stay for winter in the breeding area [102]. With favourable conditions along the autumn migration route, Wrens may winter closer to breeding grounds, e.g. in the Apennine Peninsula, without crossing the Mediterranean Sea, which would result in their early return in spring to the north. Shorter distance between the wintering and breeding sites would benefit Wrens in several ways. Birds staying at closer wintering quarters would be able to predict conditions on the breeding grounds with greater precision than those staying farther away [3]. Wren’s energy demands during migratory flight are high, due to their short and rounded wings and low rate of fat deposition [76], shorter distance to cross would improve their survival of both autumn and spring migration. Closer wintering grounds might promote early occupation of breeding territories in males [81,103,104]. According to the dominance theory and given the greater physiological constraints [105], females and juveniles will be more likely to migrate farther south to overwinter in conditions that will ensure their survival [106,107]. Such patterns have been shown in the European Robin, which is territorial on the wintering grounds as Wren [102], and in the close North American relative of the Wren, Marsh Wren Cistothorus palustris [105]. Shortening of migration distance has been observed among passerines in response to climate change [88,101], in e.g. Blackcap Sylvia atricapilla [100] or Blue Tit Cyanistes caeruleus [108].
We suspect also another explanation of early arrivals of Wren at Hel after warm autumns in the eastern Mediterranean region. Conditions at autumn stopover sites are crucial for the rate of migration, as birds spend up to seven-fold more time on foraging and resting, than on actual flight [109]. Efficiency of energy accumulation depends indirectly on conditions at stopover sites [88,110]. Warm autumn favours robust vegetation and the availability of insects for birds on migration [20,83]. Wrens’ diet is composed exclusively of invertebrates [111]. Increased availability of such prey in autumn might have a positive effects on their condition, and enable their early arrivals the following spring at the Baltic coast. An analogous positive carry-over effect of favourable conditions on autumn stopover sites in the Mediterranean region on survival and subsequent early arrivals at the breeding grounds has been shown in the Great Reed Warblers Acrocephalus arundinaceus [33]. A lack of analogous effect of autumn MOI1on spring migration at Bukowo–Kopań, might be related with likely lower proportion of Wrens arriving at that station from the eastern Mediterranean region than at Hel, which we discussed earlier [44,48].
4.6. Long-term Irends and Inter-annual Variation in Wren’s Spring Migration in Response to Rrends and Variability in the climate indices
Our results showed significant negative trends over 1982-2021 for SCAND in March–April and in May–June, and NAO in July–August (Figure A1), indices related with spring and summer conditions in northern Europe [19,57]. As SCAND (May–June) and NAO (July–August) were positively associated with the passage at Bukowo-Kopań (Figure 4A), their combined linear multi-year trends might explain the slight shift to later end (95%) of passage at this station (Table 2, Figure 4A, 4B). Analogously, the small advance in the beggining of passage at Hel (Figure 3D) might be be an effect of the positively correlated SCAND March–April (Figure 4B), which showed long-term trends. The influence of local spring temperature at Hel (THL), which increased during our study period (Figure A1), might contribute to that small advance in the start of passage at Hel (Figure 3D), as in other passerines [21]. Local spring temperatures near Bukowo–Kopań (TBK), which also increased over 1981-2021 (Figure A1), might also contribute to the slight advance of the median date at this station (Figure 3C). These advances of the early phases of Wren’s passage at our stations correspond with the overall trend in short- and medium-distance migrants in Europe to shift beginning of passage earlier with high local spring temperatures [14,15,17].
The delay of the end of migration, which was the main cause of the extension of the spring passage over the 40 studied years at Hel, cannot be attributed to the effect of spring MOI1 (Figure 4B), which had no trend over study period (Figure A1). Hence, other factors, likely those related with changes in the number of broods and population productivity over the study period, which we discussed earlier, provide a better explanation for prolonged passage at Hel (Figure A2, Table A3). An increased number of inexperienced and late migrating immatures after warm summers allowing for two or three broods, would delay the end of migration.
The insignificant trends, with a large year-to-year variation in timing of spring passage at both stations (Figure 3A,B) might be explained by a large interannual variation in the remaining climate indices related with the Wren’s passage. Since the 1980s NAO and MOI1 have shown large year–to-year variablity (Figure A1) [26,67,89]. Summer SCAND has shown mostly negative linear trend since mid-XXth century [112], but its inter-anual variation increased from 1990s [113], so this trend in SCAND (July–August) is no longer valid (Figure A1). A lack of significant long-term trend in overall passage (AA) at Hel (Figure 3A), which was influenced by MOI1 in March–April and SCAND in July–August (Figure 4B), is consistent with no long-term trends in both these climate indices (Figure A1). Analogously, for Hel, large variation in the overall passage (AA) (Figure 3B), and in the dates of beginning and the end of passage (Figure 3D) is in line with large multi-annual variation of spring MOI1 in March–April and September–October of the previous year (Figure A1). NAO in spring (March–April) and winter (November–February), which showed large annual changes but no many-year trends (Figure A1), might contribute to the large variation and thus a weak trend in the median date of passage at Bukowo-Kopań.
5. Conclusions
Our results provide new insight into drivers of spring migration phenology of short-distance migrants with wide non-breeding grounds, such as Wren by considering a wide set of climate indices that cover most of Europe. The timing of Wren’s spring passage at the Baltic coast was shaped by a combination of carry-over effects of climate factors the birds encountered during the year prior to spring migration in different parts of their range. High spring MOI1, SCAND promoted early spring arrivals of Wren’s at the Baltic coast. Favourable conditions during autumn migration to the eastern Mediterranean, reflected by high MOI1 in September–October, advanced arrivals at Hel the next spring, probably by enabling Wrens to overwinter closer to the breeding grounds. Warm summers on breeding grounds, indicated by high summer SCAND and NAO, and mild winters in south-western Europe related with high winter NAO, delayed next years’ spring migration, probably by the positive effect on the numbers and survival rate of immatures. These large-scale indices of climate in regions where Wrens stayed during non-breeding and breeding periods had greater effects on their spring phenology than local spring temperatures, which only fine-tuned their migration timing. We attribute the weak multi-year trends in timing of spring passage of Wren at both our stations, different to advanced arrivals at other sites in Europe, to the effects of climate indices with large year-to year variation, such as MOI1, which likely override the influence of the climate indices with long-term trends. Strong effects of MOI1 on Wren’s migration at more eastern station Hel, emphasize that other large-scale climate indices than just the most commonly analysed NAO, shape spring migration phenology, especially in central and eastern Europe. The effect of MOI1 on Wren’s spring migration highlights the relevance of this climate index, so far rather neglected, in understanding of the effects of climate change on phenology of bird migration in Europe.
Author Contributions
Conceptualization, I.G. and M.R.; methodology, M.R.; software, M.R.; formal analysis, I.G.; data collecting, M.R. and I.G.; writing—original draft preparation, I.G.; writing—review and editing, M.R.; visualization, I.G.; supervision, M.R.; funding acquisition, M.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research has been supported over the years by the Special Research Facility grants (SPUB) from the Polish Ministry of Education and Science to the Bird Migration Research Station, University of Gdańsk.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data supporting reported results can be found at the Global Biodiversity Information Facility database at: Ringing Data from the Bird Migration Research Station, University of Gdańsk (gbif.org) [145].
Acknowledgments
Thousands of citizen scientists collected the data we used at Operation Baltic’s Bukowo ringing station over four decades. The staff of the Bird Migration Research Station, especially Jarosław K. Nowakowski, Krzysztof Stępniewski, Wioletta Wójcik and Justyna Szulc, compiled the databases on birds. We acknowledge the climate data providers in the US National Oceanic and Atmospheric Administration, National Weather Service, Climate Prediction Center (http://www.cpc.ncep.noaa.gov/,accessed on 22 May 2023), the European Climate Assessment and Dataset (http://www.ecad.eu, accessed on 22 May 2023), the Climatic Research Unit at the University of East Anglia, UK (https://crudata.uea.ac.uk/cru/data/moi/,accessed on 22 May 2023), and KNMI Climate Explorer (http://climexp.knmi.nl, accessed on 22 May 2023).
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A. Detailed results of the models presented in the study: Table A1, Table A2, Table A3, Table A4 and Table A5, Figure A1 and Figure A2

Table A1.
Numbers of European Wrens caught at Bukowo–Kopań and Hel stations in 1982–2021. “–” indicate seasons in which the stations did not operate, grey indicates years excluded from the analysis, when < 10 individuals were caught at a station. Total = the total number of Wrens ringed at each station in1982–2021.
Table A1.
Numbers of European Wrens caught at Bukowo–Kopań and Hel stations in 1982–2021. “–” indicate seasons in which the stations did not operate, grey indicates years excluded from the analysis, when < 10 individuals were caught at a station. Total = the total number of Wrens ringed at each station in1982–2021.
| Year | Station | Year | Station | |||
| Hel | Bukowo -Kopań | Hel | Bukowo -Kopań | |||
| 1982 | 36 | 12 | 2002 | 30 | 36 | |
| 1983 | 49 | 46 | 2003 | 25 | 35 | |
| 1984 | 49 | 54 | 2004 | 24 | 16 | |
| 1985 | 7 | 18 | 2005 | 32 | 30 | |
| 1986 | 13 | 24 | 2006 | 36 | 33 | |
| 1987 | 16 | 37 | 2007 | 57 | 49 | |
| 1988 | 7 | 31 | 2008 | 61 | 69 | |
| 1989 | 7 | 36 | 2009 | 52 | 43 | |
| 1990 | 6 | 25 | 2010 | 18 | 26 | |
| 1991 | 7 | 55 | 2011 | 67 | – | |
| 1992 | 2 | 24 | 2012 | 33 | 49 | |
| 1993 | 8 | 21 | 2013 | 24 | 32 | |
| 1994 | 4 | 32 | 2014 | 30 | 19 | |
| 1995 | 29 | 32 | 2015 | 18 | 26 | |
| 1996 | 99 | 42 | 2016 | 41 | 31 | |
| 1997 | 19 | 13 | 2017 | 23 | 28 | |
| 1998 | 30 | 54 | 2018 | 17 | 26 | |
| 1999 | 30 | 41 | 2019 | 32 | 20 | |
| 2000 | 29 | 60 | 2020 | 40 | – | |
| 2001 | 11 | 56 | 2021 | 22 | 36 | |
| Total | 1140 | 1317 | ||||
Figure A1.
Trends and variation for the 11 climate indices used in the study over 1981–2021. *=statistical significance at p<0.05.
Figure A1.
Trends and variation for the 11 climate indices used in the study over 1981–2021. *=statistical significance at p<0.05.
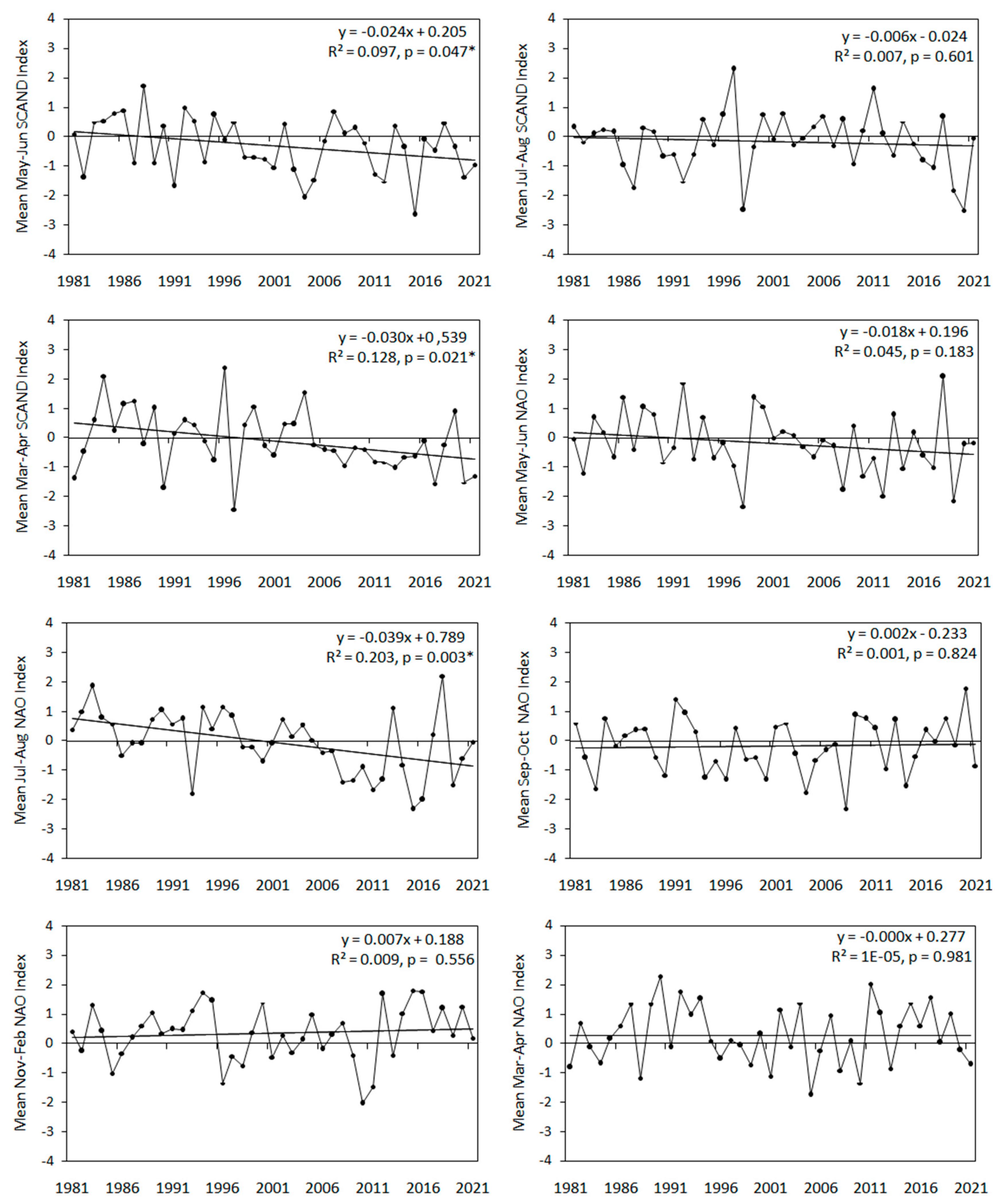

Figure A2.
Duration of spring migration on Bukowo–Kopań (A) and Hel (B) stations based on Annual Annomaly (AA) in 1982–2021. „Year” is used in the equations as the explanatory variable, * = statistical significance at p < 0.05.
Figure A2.
Duration of spring migration on Bukowo–Kopań (A) and Hel (B) stations based on Annual Annomaly (AA) in 1982–2021. „Year” is used in the equations as the explanatory variable, * = statistical significance at p < 0.05.
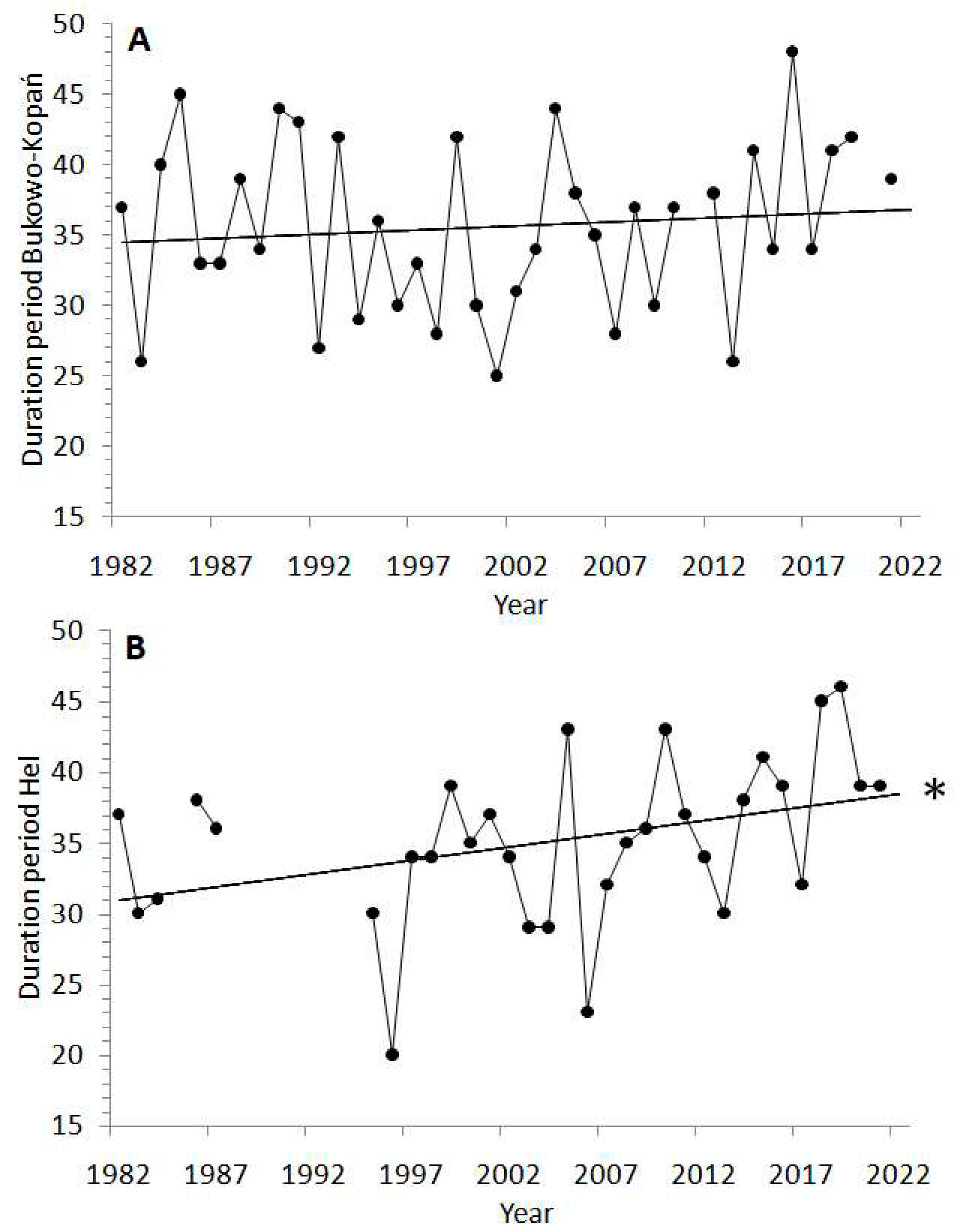
Table A2.
Pearson’s correlation coefficients of the climate indices in 1982–2021 analysed in multiple regression models. Abbreviations of the climate variables as in Table 1. “_1Y” = climate indices from the year preceding analysed spring. Significant correlations after Benjamini-Hochberg correction for multiple comparisons with an assumed 5% false positive rate highlighted in bold face.
Table A2.
Pearson’s correlation coefficients of the climate indices in 1982–2021 analysed in multiple regression models. Abbreviations of the climate variables as in Table 1. “_1Y” = climate indices from the year preceding analysed spring. Significant correlations after Benjamini-Hochberg correction for multiple comparisons with an assumed 5% false positive rate highlighted in bold face.
| Climate indices | NAO_ MAR_APR |
NAO_ SEP_OCT_1Y |
NAO_ MAY_JUN_1Y |
NAO_ JUL_AUG_1Y |
MOI1_ NOV_FEB |
MOI1_ MAR_APR |
MOI1_ SEP_OCT_1Y |
SCAND_ MAR_APR |
SCAND_ MAY_JUN_1Y |
SCAND_ JUL_AUG_1Y |
TBK_ MAR_APR |
THL_ MAR_APR |
| NAO_NOV_FEB | 0.16 | 0.11 | 0.06 | -0.05 | 0.74 | -0.10 | 0.09 | -0.15 | -0.43 | -0.11 | 0.42 | 0.43 |
| NAO_MAR_APR | 0.00 | 0.12 | -0.11 | 0.20 | 0.30 | -0.09 | 0.01 | 0.13 | 0.20 | 0.30 | 0.31 | |
| NAO_SEP_OCT_1Y | 0.26 | -0.06 | -0.12 | 0.01 | 0.13 | 0.11 | 0.11 | -0.03 | 0.05 | 0.07 | ||
| NAO_MAY_JUN_1Y | 0.39 | 0.21 | 0.32 | -0.01 | 0.19 | 0.19 | 0.18 | 0.01 | 0.01 | |||
| NAO_JUL_AUG_1Y | 0.12 | 0.19 | 0.05 | 0.41 | 0.34 | 0.23 | -0.08 | -0.09 | ||||
| MOI1_NOV_FEB | -0.07 | -0.10 | -0.05 | -0.41 | 0.06 | 0.43 | 0.43 | |||||
| MOI1_MAR_APR | -0.07 | -0.23 | 0.37 | 0.29 | 0.14 | 0.13 | ||||||
| MOI1_SEP_OCT_1Y | 0.26 | 0.22 | -0.28 | -0.29 | -0.24 | |||||||
| SCAND_MAR_APR | 0.26 | 0.01 | -0.24 | -0.22 | ||||||||
| SCAND_MAY_JUN_1Y | 0.11 | -0.07 | -0.07 | |||||||||
| SCAND_JUL_AUG_1Y | -0.01 | -0.03 | ||||||||||
| TBK_MAR_APR | 1.00 |
Table A3.
Summary statistics for linear regressions over the year for the metrics of spring migration phenology of the European Wren during 1982–2021. 5%–95% = subsequent percentiles of spring migrants, AA = the Annual Anomaly, Duration of passage = the difference between the dates when 5% and 95% of migrants passed through each station. Mean = many-year mean date, “–” = mean date not applicable, ß slope = regression coefficient; SE = its Standard Error; R2 = determination coefficient; t, p = results of t-test, p<0.05 marked in bold face, 40 x ß = estimated change in days of timing over 1982–2021, negative values reflect advance of migration.
Table A3.
Summary statistics for linear regressions over the year for the metrics of spring migration phenology of the European Wren during 1982–2021. 5%–95% = subsequent percentiles of spring migrants, AA = the Annual Anomaly, Duration of passage = the difference between the dates when 5% and 95% of migrants passed through each station. Mean = many-year mean date, “–” = mean date not applicable, ß slope = regression coefficient; SE = its Standard Error; R2 = determination coefficient; t, p = results of t-test, p<0.05 marked in bold face, 40 x ß = estimated change in days of timing over 1982–2021, negative values reflect advance of migration.
| Parameter/Station | Mean | SD | ß slope | SE | R2 | t30 | p | 40 x ß (days) |
| Bukowo–Kopań | ||||||||
| BK_AA | – | – | -0.0432 | 0.06 | 0.02 | –0.75 | 0.46 | -1.73 |
| BK_5% | 26 Mar | 1.12 | 0.0004 | 0.02 | 0.00 | 0.03 | 0.98 | 0.02 |
| BK_50% (median) | 13 Apr | 6.07 | -0.1323 | 0.09 | 0.06 | –1.52 | 0.14 | -5.29 |
| BK_95% | 03 May | 6.02 | 0.0577 | 0.09 | 0.01 | 0.67 | 0.51 | 2.31 |
| BK_Duration of passage (5%–95%) | – | – | 0.0573 | 0.09 | 0.01 | 0.66 | 0.52 | 2.29 |
| Hel | ||||||||
| HL_AA | – | – | -0.0662 | 0.06 | 0.03 | –1.04 | 0.31 | -2.65 |
| HL_5% | 30 Mar | 3.49 | -0.0911 | 0.06 | 0.08 | –1.65 | 0.11 | -3,64 |
| HL_50% (median) | 15 Apr | 6.30 | -0.1260 | 0.10 | 0.05 | –1.27 | 0.21 | -5.04 |
| HL_95% | 07 May | 4.87 | 0.0985 | 0.08 | 0.05 | 1.28 | 0.21 | 3.94 |
| HL_Duration of passage (5%–95%) | – | – | 0.19 | 0.09 | 0.14 | 2.20 | 0.04 | 7.6 |
Table A4.
Results of the Mann–Whitney test for differences in the dates of subsequent stages of the spring passage of Wren through Bukowo-Kopań (N = 38 years) and Hel (N = 32 years) stations in 1982–2021. 5%, 50%, 95% = dates of these selected percentiles of spring migration, Z, p = results of the Mann–Whitney test, p =statistical significance of differences, p<0.05 marked in bold face.
Table A4.
Results of the Mann–Whitney test for differences in the dates of subsequent stages of the spring passage of Wren through Bukowo-Kopań (N = 38 years) and Hel (N = 32 years) stations in 1982–2021. 5%, 50%, 95% = dates of these selected percentiles of spring migration, Z, p = results of the Mann–Whitney test, p =statistical significance of differences, p<0.05 marked in bold face.
| Parameter | Mean date Bukowo-Kopań | Mean date Hel | Z | p |
| 5% | 26 Mar | 30 Mar | –6.33 | <0.01 |
| 50% | 13 Apr | 15 Apr | –3.60 | <0.01 |
| 95% | 03 May | 07 May | –2.97 | <0.01 |
Table A5.
Model selection procedure by “all subsets”, according to AICc, from the full model. The models represents the relationship between 12 climate variables and Year, used as explanatory variables, and Annual Anomaly (AA) or subsequent percentiles (5%, 50%, 95%) of spring passage used as response variables, of Wren Troglodytes troglodytes ringed at Bukowo–Kopań station in 1982–2021. The table presents models with ΔAICc <2. The models are ranked according to Akaike’s information criterion adjusted for small samples (AICc), k is the number of parameters estimated in the model, ∆AICc is the difference between AICc from the model and lowest AICc value, wi is the Akaike weight. The best model, discussed in the text, in bold face.
Table A5.
Model selection procedure by “all subsets”, according to AICc, from the full model. The models represents the relationship between 12 climate variables and Year, used as explanatory variables, and Annual Anomaly (AA) or subsequent percentiles (5%, 50%, 95%) of spring passage used as response variables, of Wren Troglodytes troglodytes ringed at Bukowo–Kopań station in 1982–2021. The table presents models with ΔAICc <2. The models are ranked according to Akaike’s information criterion adjusted for small samples (AICc), k is the number of parameters estimated in the model, ∆AICc is the difference between AICc from the model and lowest AICc value, wi is the Akaike weight. The best model, discussed in the text, in bold face.
| Response variable | Explanatory variables in models | k | AICc | ΔAICc | Wi |
|---|---|---|---|---|---|
| BK_AA | NAO_JUL_AUG_1Y | 2 | 103.92 | 0 | 0.21 |
| MOI1_SEP_OCT_1Y+NAO_JUL_AUG_1Y | 3 | 104.84 | 0.92 | 0.13 | |
| MOI1_NOV_FEB+NAO_JUL_AUG_1Y | 3 | 104.87 | 0.95 | 0.13 | |
| NAO_JUL_AUG_1Y+SCAND_MAY_JUN_1Y | 3 | 104.94 | 1.02 | 0.13 | |
| NAO_JUL_AUG_1Y+NAO_SEP_OCT_1Y | 3 | 105.11 | 1.18 | 0.12 | |
| MOI1_NOV_FEB+NAO_JUL_AUG_1Y+NAO_NOV_FEB | 4 | 105.47 | 1.55 | 0.1 | |
| NAO_JUL_AUG_1Y+TBK_MAR_APR | 3 | 105.68 | 1.75 | 0.09 | |
| MOI1_MAR_APR+NAO_JUL_AUG_1Y | 3 | 105.78 | 1.86 | 0.08 | |
| BK_5%~ | NAO_JUL_AUG_1Y+SCAND_JUL_AUG_1Y | 3 | 99.3 | 0 | 0.2 |
| MOI1_MAR_APR+NAO_JUL_AUG_1Y+SCAND_JUL_AUG_1Y | 4 | 99.38 | 0.08 | 0.19 | |
| NAO_JUL_AUG_1Y+NAO_MAR_APR+SCAND_JUL_AUG_1Y | 4 | 99.61 | 0.31 | 0.17 | |
| NAO_JUL_AUG_1Y+SCAND_JUL_AUG_1Y+TBK_MAR_APR | 4 | 99.79 | 0.5 | 0.16 | |
| MOI1_MAR_APR+NAO_JUL_AUG_1Y+SCAND_JUL_AUG_1Y+TBK_MAR_APR | 5 | 100.58 | 1.29 | 0.1 | |
| NAO_JUL_AUG_1Y+NAO_MAY_JUN_1Y+SCAND_JUL_AUG_1Y | 4 | 100.77 | 1.47 | 0.1 | |
| MOI1_MAR_APR+NAO_JUL_AUG_1Y+NAO_MAR_APR | 5 | 100.98 | 1.68 | 0.09 | |
| BK_50%~ | MOI1_MAR_APR+NAO_JUL_AUG_1Y+NAO_MAR_APR+NAO_NOV_FEB+SCAND_MAR_APR +SCAND_MAY_JUN_1Y+TBK_MAR_APR | 8 | 100.34 | 0 | 0.08 |
| MOI1_MAR_APR+NAO_JUL_AUG_1Y+NAO_MAR_APR+SCAND_MAR_APR+SCAND_MAY_JUN_1Y | 6 | 100.72 | 0.38 | 0.06 | |
| MOI1_MAR_APR+NAO_JUL_AUG_1Y+NAO_MAR_APR+SCAND_MAR_APR+SCAND_MAY_JUN_1Y+TBK_MAR_APR | 7 | 100.76 | 0.42 | 0.06 | |
| BK_50%~ | NAO_JUL_AUG_1Y+NAO_NOV_FEB+SCAND_MAY_JUN_1Y | 4 | 100.81 | 0.47 | 0.06 |
| MOI1_MAR_APR+NAO_JUL_AUG_1Y+NAO_NOV_FEB+SCAND_MAR_APR+SCAND_MAY_JUN_1Y | 6 | 100.91 | 0.57 | 0.06 | |
| NAO_JUL_AUG_1Y+SCAND_MAY_JUN_1Y | 3 | 100.98 | 0.65 | 0.05 | |
| MOI1_MAR_APR+MOI1_NOV_FEB+NAO_JUL_AUG_1Y+NAO_MAR_APR+NAO_NOV_FEB+SCAND_MAR_APR +SCAND_MAY_JUN_1Y+TBK_MAR_APR |
9 | 101.01 | 0.67 | 0.05 | |
| MOI1_MAR_APR+NAO_JUL_AUG_1Y+NAO_NOV_FEB+SCAND_MAR_APR+SCAND_MAY_JUN_1Y+TBK_MAR_APR | 7 | 101.02 | 0.68 | 0.05 | |
| NAO_JUL_AUG_1Y+NAO_NOV_FEB+SCAND_MAY_JUN_1Y+TBK_MAR_APR | 5 | 101.03 | 0.69 | 0.05 | |
| MOI1_MAR_APR+NAO_JUL_AUG_1Y+NAO_NOV_FEB+SCAND_MAY_JUN_1Y | 5 | 101.14 | 0.8 | 0.05 | |
| NAO_JUL_AUG_1Y | 2 | 101.27 | 0.93 | 0.05 | |
| MOI1_MAR_APR+NAO_JUL_AUG_1Y+SCAND_MAR_APR+SCAND_MAY_JUN_1Y | 5 | 101.28 | 0.94 | 0.05 | |
| MOI1_MAR_APR+MOI1_NOV_FEB+NAO_JUL_AUG_1Y+NAO_NOV_FEB+SCAND_MAY_JUN_1Y | 6 | 101.38 | 1.04 | 0.04 | |
| MOI1_NOV_FEB+NAO_JUL_AUG_1Y+NAO_NOV_FEB+SCAND_MAY_JUN_1Y | 5 | 101.49 | 1.15 | 0.04 | |
| MOI1_MAR_APR+NAO_JUL_AUG_1Y+NAO_MAR_APR+NAO_NOV_FEB+SCAND_MAR_APR+SCAND_MAY_JUN_1Y+TBK_MAR_APR | 8 | 100.34 | 0 | 0.08 | |
| MOI1_MAR_APR+NAO_JUL_AUG_1Y+NAO_MAR_APR+SCAND_MAR_APR+SCAND_MAY_JUN_1Y | 6 | 100.72 | 0.38 | 0.06 | |
| MOI1_MAR_APR+NAO_JUL_AUG_1Y+NAO_MAR_APR+SCAND_MAR_APR+SCAND_MAY_JUN_1Y+TBK_MAR_APR | 7 | 100.76 | 0.42 | 0.06 | |
| MOI1_NOV_FEB+NAO_JUL_AUG_1Y+NAO_NOV_FEB | 4 | 101.89 | 1.56 | 0.03 | |
| MOI1_MAR_APR+NAO_JUL_AUG_1Y+NAO_MAR_APR+SCAND_MAY_JUN_1Y | 5 | 102.1 | 1.76 | 0.03 | |
| MOI1_MAR_APR+NAO_JUL_AUG_1Y+NAO_NOV_FEB+SCAND_MAY_JUN_1Y+TBK_MAR_APR | 6 | 102.23 | 1.9 | 0.03 | |
| NAO_JUL_AUG_1Y+NAO_NOV_FEB+SCAND_MAR_APR+SCAND_MAY_JUN_1Y+TBK_MAR_APR | 6 | 102.26 | 1.92 | 0.03 | |
| BK_95%~ | NAO_JUL_AUG_1Y+SCAND_JUL_AUG_1Y | 3 | 107.41 | 0 | 0.12 |
| NAO_MAY_JUN_1Y+SCAND_JUL_AUG_1Y | 3 | 108.23 | 0.82 | 0.08 | |
| MOI1_SEP_OCT_1Y+NAO_JUL_AUG_1Y+SCAND_JUL_AUG_1Y | 4 | 108.41 | 1 | 0.07 | |
| NAO_JUL_AUG_1Y+NAO_MAY_JUN_1Y+SCAND_JUL_AUG_1Y | 4 | 108.56 | 1.15 | 0.07 | |
| NAO_JUL_AUG_1Y | 2 | 108.62 | 1.21 | 0.07 | |
| SCAND_JUL_AUG_1Y | 2 | 108.72 | 1.31 | 0.06 | |
| MOI1_SEP_OCT_1Y+NAO_MAY_JUN_1Y+SCAND_JUL_AUG_1Y | 4 | 108.76 | 1.34 | 0.06 | |
| NAO_JUL_AUG_1Y+NAO_NOV_FEB+SCAND_JUL_AUG_1Y | 4 | 108.81 | 1.39 | 0.06 |
Table A6.
Model selection procedure by “all subsets”, according to AICc, from the full model. The model represents the relationship between 12 climate variables and Year, used as explanatory variables, and Annual Anomaly (AA) or subsequent percentiles (5%, 50%, 95%) of spring passage used as response variables, of Wren Troglodytes troglodytes ringed at Hel station in 1982–2021. The table presents models with ΔAICc <2. The models are ranked according to Akaike’s adjusted information criterion for small samples (AICc), k is the number of parameters estimated in the model, ∆AICc is the difference between AICc from the model and lowest AICc value, wi is the Akaike weight.The best model, discussed in the text, in bold face.
Table A6.
Model selection procedure by “all subsets”, according to AICc, from the full model. The model represents the relationship between 12 climate variables and Year, used as explanatory variables, and Annual Anomaly (AA) or subsequent percentiles (5%, 50%, 95%) of spring passage used as response variables, of Wren Troglodytes troglodytes ringed at Hel station in 1982–2021. The table presents models with ΔAICc <2. The models are ranked according to Akaike’s adjusted information criterion for small samples (AICc), k is the number of parameters estimated in the model, ∆AICc is the difference between AICc from the model and lowest AICc value, wi is the Akaike weight.The best model, discussed in the text, in bold face.
| Response variable | Explanatory variables in models | k | AICc | ΔAICc | Wi |
|---|---|---|---|---|---|
| HL_AA | MOI1_MAR_APR+SCAND_JUL_AUG_1Y | 3 | 89.05 | 0 | 0.33 |
| MOI1_MAR_APR+NAO_MAY_JUN_1Y+SCAND_JUL_AUG_1Y | 4 | 89.84 | 0.79 | 0.22 | |
| MOI1_MAR_APR+NAO_JUL_AUG_1Y+SCAND_JUL_AUG_1Y | 4 | 90.19 | 1.14 | 0.18 | |
| MOI1_MAR_APR+MOI1_SEP_OCT_1Y+SCAND_JUL_AUG_1Y | 4 | 90.74 | 1.69 | 0.14 | |
| SCAND_JUL_AUG_1Y | 2 | 90.91 | 1.86 | 0.13 | |
| HL_5%~ | MOI1_SEP_OCT_1Y+SCAND_MAR_APR+THL_MAR_APR | 4 | 83.23 | 0 | 0.17 |
| MOI1_MAR_APR+MOI1_SEP_OCT_1Y+NAO_MAR_APR+THL_MAR_APR | 5 | 83.67 | 0.44 | 0.13 | |
| MOI1_MAR_APR+MOI1_SEP_OCT_1Y+THL_MAR_APR | 4 | 83.79 | 0.56 | 0.13 | |
| MOI1_MAR_APR+MOI1_SEP_OCT_1Y+SCAND_MAR_APR+THL_MAR_APR | 5 | 84.05 | 0.82 | 0.11 | |
| MOI1_SEP_OCT_1Y+THL_MAR_APR | 3 | 84.07 | 0.84 | 0.11 | |
| MOI1_MAR_APR+MOI1_SEP_OCT_1Y+NAO_MAR_APR+SCAND_MAR_APR+ THL_MAR_APR | 6 | 84.72 | 1.49 | 0.08 | |
| MOI1_SEP_OCT_1Y+NAO_SEP_OCT_1Y+SCAND_MAR_APR+THL_MAR_APR | 5 | 84.82 | 1.59 | 0.08 | |
| MOI1_MAR_APR+MOI1_SEP_OCT_1Y+SCAND_JUL_AUG_1Y+THL_MAR_APR | 5 | 84.84 | 1.61 | 0.07 | |
| MOI1_SEP_OCT_1Y+NAO_MAR_APR+SCAND_MAR_APR+THL_MAR_APR | 5 | 85.08 | 1.85 | 0.07 | |
| MOI1_MAR_APR+MOI1_SEP_OCT_1Y+SCAND_MAY_JUN_1Y+THL_MAR_APR | 5 | 85.21 | 1.98 | 0.06 | |
| HL_50%~ | SCAND_JUL_AUG_1Y | 2 | 88.38 | 0 | 0.16 |
| MOI1_MAR_APR+SCAND_JUL_AUG_1Y | 3 | 89.2 | 0.81 | 0.11 | |
| SCAND_JUL_AUG_1Y+SCAND_MAR_APR | 3 | 89.59 | 1.2 | 0.09 | |
| NAO_SEP_OCT_1Y+SCAND_JUL_AUG_1Y | 3 | 89.74 | 1.36 | 0.08 | |
| NAO_JUL_AUG_1Y+SCAND_JUL_AUG_1Y | 3 | 89.86 | 1.47 | 0.08 | |
| NAO_MAR_APR+SCAND_JUL_AUG_1Y | 3 | 89.91 | 1.53 | 0.08 | |
| MOI1_MAR_APR+NAO_MAR_APR+SCAND_JUL_AUG_1Y | 4 | 89.93 | 1.55 | 0.08 | |
| MOI1_SEP_OCT_1Y+SCAND_JUL_AUG_1Y | 3 | 90.08 | 1.69 | 0.07 | |
| NAO_NOV_FEB+SCAND_JUL_AUG_1Y | 3 | 90.17 | 1.79 | 0.07 | |
| MOI1_NOV_FEB+SCAND_JUL_AUG_1Y | 3 | 90.25 | 1.86 | 0.06 | |
| MOI1_MAR_APR+NAO_JUL_AUG_1Y+NAO_MAR_APR+SCAND_JUL_AUG_1Y | 5 | 90.29 | 1.9 | 0.06 | |
| NAO_MAY_JUN_1Y+SCAND_JUL_AUG_1Y | 3 | 90.3 | 1.92 | 0.06 | |
| HL_95%~ | MOI1_MAR_APR | 1 | 91.93 | 0 | 0.28 |
| SCAND_JUL_AUG_1Y | 2 | 92.87 | 0.94 | 0.17 | |
| MOI1_SEP_OCT_1Y | 2 | 93.64 | 1.71 | 0.12 | |
| NAO_MAY_JUN_1Y | 2 | 93.71 | 1.78 | 0.11 | |
| NAO_SEP_OCT_1Y | 2 | 93.8 | 1.87 | 0.11 | |
| YearN | 2 | 93.84 | 1.91 | 0.11 |
References
- Allen, M.R.; Dube, O.P; Solecki, W.; Aragón–Durand, F.; Cramer W, Humphreys S, Kainuma M, Kala J, Mahowald N, Mulugetta Y, Perez R, Wairiu M, Zickfeld K. Framing and Context. In: Global Warming of 1.5°C. An IPCC Special Report on the impacts of global warming of 1.5°C above pre–industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty [Masson–Delmotte V, Zhai P, Pörtner HO, Roberts D, Skea J, Shukla PR, Pirani A, Moufouma–Okia W, Péan C, Pidcock R, Connors S, Matthews JBR, Chen Y, Zhou X, Gomis MI, Lonnoy E, Maycock T, Tignor M, Waterfield T.]. Cambridge University Press, Cambridge, UK and New York, NY, USA, 2018, 49–92. [CrossRef]
- Newson, E.S.; Mendes, S.; Crick, H.Q.P.; Dulvy, N.K.; Houghton, J.D.R.; Hays, G.C.; Hutson, A.M.; MacLeod, C.D.; Pierce, G.J.; Robinson, R.; A. Indicators of the impact of climate change on migratory species. Endang. Species 2009, 7, 101–113. [Google Scholar] [CrossRef]
- Visser, M.E.; Both, C. ; Shifts in phenology due to global climate change: The need for a yardstick. Proceedings of the Royal Society B: Biological Sciences 2005, 272, 2561–2569. [Google Scholar] [CrossRef] [PubMed]
- Ockendon, N.; Leech, D.; Pearce–Higgins, J.W. Climatic effects on breeding grounds are more important drivers of breeding phenology in migrant birds than carryover effects from wintering grounds. Biology Letters 2013, 9, 20130669. [Google Scholar] [CrossRef] [PubMed]
- Crick, H.Q.P.; Dudley, C.; Glue, D.E.; Thomson, D.L. UK birds are laying eggs earlier. Nature 1997, 388, 526–527. [Google Scholar] [CrossRef]
- Ahola, M.; Laaksonen, T.; Sippola, K.; Eeva, T.; Rainio, K.; Lehikoinen, E. Variation in Climate Warming along the Migration Route Uncouples Arrival and Breeding Dates. Glob. Chang. Biol. 2004, 10, 1610–1617. [Google Scholar] [CrossRef]
- Carey, C. ; The impacts of climate change on the annual cycles of birds. Philosophical Transactions of the Royal Society B: Biological Sciences 2009, 364, 3321–3330. [Google Scholar] [CrossRef] [PubMed]
- Tøttrup, A.P.; Thorup, K.; Rahbek, C. Patterns of change in timing of springmigration in North European songbird populations. Journal of Avian Biology 2006, 37, 84–92. [Google Scholar] [CrossRef]
- Lehikoinen, A.; Lindén, A.; Karlsson, M.; Andersson, A.; Crewe, T.L.; Dunn, E.H.; Gregory, G.; Karlsson, L.; Kristiansen, V.; Mackenzie, S.; Newman, S.; Røer, J.E.; Sharpe, C.; Sokolov, L V.; Steinholtz, Å.; Stervander, M.; Tirri, IS.; Tjørnløv, RS.Phenology of the avian spring migratory passage in Europe and North America: Asymmetric advancement in time and increase in duration. Ecological Indicators 2019, 101, 985–991. [CrossRef]
- Vähätalo, A.V.; Rainio, K.; Lehikoinen, A.; Lehikoinen, E. Spring Arrival of Birds Depends on the North Atlantic Oscillation. J. Avian Biol 2004, 35, 210–216. [Google Scholar] [CrossRef]
- Rainio, K.; Laaksonen, T.; Ahola, M.; Vähätalo, A.V.; Lehikoinen, E. Climatic Responses in Spring Migration of Boreal and Arctic Birds in Relation to Wintering Area and Taxonomy. J. Avian Biol. 2006, 37, 507–515. [Google Scholar] [CrossRef]
- Ambrosini, R.; Cuervo, JJ.; du Feu, C.; Fiedler, W.; Musitelli, F.; Rubolini, D.; Sicurella, B.; Spina, F.; Saino, N.; Møller, AP. Migratory connectivity and effects of winter temperatures on migratory behaviour of the European robin Erithacus rubecula: A continent–wide analysis. Journal of Animal Ecology 2016, 85, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Møller, A.P. North Atlantic Oscillation (NAO) effects of climate on the relative importance of first and second clutches in a migratory passerine bird. Journal of Animal Ecology 2002, 71, 201–210. [Google Scholar] [CrossRef]
- Hüppop, O.; Hüppop, K. North Atlantic Oscillation and Timing of Spring Migration in Birds. Proc. R. Soc. B Biol. Sci. 2003, 270, 233–240. [Google Scholar] [CrossRef]
- Palm, V.; Leito, A.; Truu, J.; Tomingas, O. The Spring Timing of Arrival of Migratory Birds: Dependence on Climate Variables and Migration Route. Ornis Fenn. 2009, 86, 97–108. [Google Scholar]
- Maggini, I.; Cardinale, M.; Sundberg, J.H.; Spina, F.; Fusani, L. Recent phenological shifts of migratory birds at a Mediterranean spring stopover site: Species wintering in the Sahel advance passage more than tropical winterers. PLoS One 2020, 15(9), e0239489. [Google Scholar] [CrossRef] [PubMed]
- Tøttrup, A.P.; Rainio, K.; Coppack, T.; Lehikoinen, E.; Rahbek, C.; Thorup, K. Local Temperature Fine–Tunes the Timing of Spring Migration in Birds. Integr. Comp. Biol. 2010, 50, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Crick, H.Q.P.; Sparks, T.H. Climate change related to egg–laying trends. Nature 1999, 399, 423–424. [Google Scholar] [CrossRef]
- Bueh, C.; Nakamura, H. Scandinavian Pattern and Its Climatic Impact. Q. J. R. Meteorol. Soc. 2007, 133, 2117–2131. [Google Scholar] [CrossRef]
- Remisiewicz, M.; Underhill, L.G. Climatic Variation in Africa and Europe Has Combined Effects on Timing of Spring Migration in a Long–Distance Migrant Willow Warbler Phylloscopus trochilus. PeerJ 2020, 8, 1–30. [Google Scholar] [CrossRef]
- Remisiewicz, M.; Underhill, L.G. Large–scale climatic patterns have stronger carry–over effects than local temperatures on spring phenology of long–distance passerine migrants between Europe and Africa. Animals 2022, 12, 1–57. [Google Scholar] [CrossRef]
- Ranasinghe, R.; McLoughlin, R.; Short, A.; Symonds, G. The Southern Oscillation Index, wave climate, and beach rotation. Marine Geology 2004, 204, 273–287. [Google Scholar] [CrossRef]
- Tryjanowski, P.; Stenseth, N.C.; Matysioková, B. The Indian Ocean Dipole as an Indicator of Climatic Conditions Affecting European Birds. Clim. Res. 2013, 57, 45–49. [Google Scholar] [CrossRef]
- Okonkwo, C. An Advanced Review of the Relationships between Sahel Precipitation and Climate Indices: A Wavelet Approach. International Journal of Atmospheric Sciences 2014, 1–11. [Google Scholar] [CrossRef]
- Tobolka, M.; Dylewski, L.; Wozna, J.T.; Zolnierowicz, K.M. How Weather Conditions in Non–Breeding and Breeding Grounds Affect the Phenology and Breeding Abilities of White Storks. Sci. Total Environ. 2018, 636, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Criado–Aldeanueva, F.; Soto–Navarro, J. Climatic indices over the Mediterranean sea: A review. Applied Sciences (Switzerland) 2020, 10, 5790. [Google Scholar] [CrossRef]
- Tomotani, B.M.; van der Jeugd, H.; Gienapp, P.; de la Hera, I.; Pilzecker, J.; Teichmann, C.; Visser, M.E. Climate Change Leads to Differential Shifts in the Timing of Annual Cycle Stages in a Migratory Bird. Glob. Chang. Biol. 2018, 24, 823–835. [Google Scholar] [CrossRef]
- Pinszke, A.; Remisiewicz, M. Long–term changes in autumn migration timing of Garden Warblers Sylvia borin at southern Baltic coast in response to spring, summer and autumn temperatures. European Zoological Journal 2023, 90(1), 283–295. [Google Scholar] [CrossRef]
- Miles, W.T.S.; Bolton, M.; Davis, P.; Dennis, R.; Broad, R.; Robertson, I.; Riddiford, N.J.; Harvey, P.V.; Riddington, R.; Shaw, D.N.; et al. Quantifying Full Phenological Event Distributions Reveals Simultaneous Advances, Temporal Stability and Delays in Spring and Autumn Migration Timing in Long–Distance Migratory Birds. Glob. Chang. Biol. 2017, 23, 1400–1414. [Google Scholar] [CrossRef]
- Redlisiak, M.; Remisiewicz, M.; Mazur, A. Sex–Specific Differences in Spring Migration Timing of Song Thrush Turdusphilomelos at the Baltic Coast in Relation to Temperatures on the Wintering Grounds. European Zoological Journal 2021, 88, 191–203. [Google Scholar] [CrossRef]
- Jenni, L.; Kéry, M. Timing of autumn bird migration under climate change: advances in long–distance migrants, delays in short–distance migrants. Proceedings of the Royal Society of London. Series B: Biological Sciences 2003, 270, 1467–1471. [Google Scholar] [CrossRef]
- Najmanova, L.; Adamik, P. Effect of climatic change on the duration of the breeding season in three European thrushes. Bird Study 2009, 56, 349–365. [Google Scholar] [CrossRef]
- Halupka, L.; Wierucka, K.; Sztwiertnia, H.; Klimczuk, E. Conditions at autumn stopover sites affect survival of a migratory passerine. Journal of Ornithology 2017, 158, 979–988. [Google Scholar] [CrossRef]
- Tryjanowski, P.; Kuźniak, S.; Sparks, T. Earlier arrival of some farmland migrants in western Poland. Ibis 2002, 144, 62–68. [Google Scholar] [CrossRef]
- Forchhammer, M.C.; Post, E.; Stenseth, N.C. North Atlantic Oscillation Timing of Long– and Short–Distance Migration. J. Anim. Ecol. 2002, 71, 1002–1014. [Google Scholar] [CrossRef]
- Usui, T.; Butchart, S.H.M.; Phillimore, A.B. Temporal Shifts and Temperature Sensitivity of Avian Spring Migratory Phenology: A Phylogenetic Meta–Analysis. J. Anim. Ecol. 2017, 86, 250–261. [Google Scholar] [CrossRef] [PubMed]
- Dinsbergs, I.; Keišs, O.; Briedis, M. Spring phenology is advancing at a faster rate than arrival times of Common Starling. Journal of Ornithology 2023, 164, 367–375. [Google Scholar] [CrossRef]
- Saino, N.; Ambrosini, R. Climatic Connectivity between Africa and Europe May Serve as a Basis for Phenotypic Adjustment of Migration Schedules of Trans–Saharan Migratory Birds. Glob. Chang. Biol. 2008, 14, 250–263. [Google Scholar] [CrossRef]
- Visser, M.E.; Gienapp, P.; Husby, A.; Morrisey, M.; de la Hera, I.; Pulido, F.; Both, C. ; Effects of Spring Temperatures on the Strength of Selection on Timing of Reproduction in a Long–Distance Migratory Bird. PLoS Biology 2015, 13, 1002120. [Google Scholar] [CrossRef]
- Haest, B.; Hüppop, O.; Bairlein, F. Weather at the Winter and Stopover Areas Determines Spring Migration Onset, Progress, and Advancements in Afro–Palearctic Migrant Birds. PNAS 2020, 117, 17056–17062. [Google Scholar] [CrossRef]
- Rolshausen, G.; Segelbacher, G.; Hobson, K.A.; Schaefer, H.M. Contemporary Evolution of Reproductive Isolation and Phenotypic Divergence in Sympatry along a Migratory Divide. Current Biology 2009, 19, 2097–2101. [Google Scholar] [CrossRef]
- Clements, J.F.; Schulenberg, T.S.; Iliff, M.J.; Fredericks, T.A.; Gerbracht, J.A.; Lepage, D.; Billerman, S.M.; Sullivan, B.L.; Wood, C.L. The eBird/Clements checklist of Birds of the World: 2022. Available online: https://www.birds.cornell.edu/clementschecklist/download/ (accessed on 2 January 2023).
- Armstrong, E.A.; Whitehouse, H.L.K. Behavioural adaptaptions of the Wren (Troglodytes troglodytes). Biol. Rev. 1977, 52, 235–294. [Google Scholar] [CrossRef]
- Spina, F.; Baillie, S.R.; Bairlein, F.; Fiedler, W.; Thorup, K. The Eurasian African Bird Migration Atlas. Available online: https://migrationatlas.org (accessed on 31 March 2023).
- Cramp, S. The Birds of the Western Palearctic. Handbook of the Birds of Europe, the Middle East and North Africa. Vol. VI: Warblers; Oxford University Press: Oxford, UK, 1992. [Google Scholar]
- Hoyo, J.; Elliott, A.; Christie, D. Handbook of the Birds of the World. Vol. 10. Cuckoo–shrikes to Thrushes; Lynx Edicions, Barcelona, 2005.
- BirdLife International and Handbook of the Birds of the World. Bird species distribution maps of the world. Version 2019.1. Available online: http://datazone.birdlife.org/species/requestdis (accessed on 1 January 2023).
- Maciąg, T.; Remisiewicz, M.; Nowakowski, J.K.; Redlisiak, M.; Rosińska, K.; Stępniewski, K.; Stępniewska, K.; Szulc, J. Website of the Bird Migration Research Station. Available online: https://en–sbwp.ug.edu.pl/badania/monitoringwyniki/maps–of–ringing–recoveries/ (accessed on 2 June 2023).
- Nowakowski, J.K. Ringing Data from the Bird Migration Research Station, University of Gdańsk. Available online: https://www.gbif.org/dataset/8186b0c0−925e−11da−8900-b8a03c50a862 (accessed on 26 May 2023).
- Tomiałojć, L.; Stawarczyk, T. Awifauna Polski: Rozmieszczenie, Liczebność i Zmiany; PTPP “pro Natura”: Wrocław, Poland, 2003. [Google Scholar]
- Trepte, A. Avi–Fauna – Vögel in Deutschland. Available online: https://www.avi–fauna.info/ (accessed on 2 January 2023).
- Busse, P.; Meissner, W. Bird Ringing Station Manual; De Gruyter Open Ltd.: Warsaw, Poland; Berlin, Germany, 2015. [Google Scholar]
- Identification Guide to European Passerines; Svensson, L., Ed.; British Trust for Ornithology: Thetford, Norfolk, UK, 1992. [Google Scholar]
- Demongin, L. Identification Guide to Birds in the Hand; Laurent Demongin: Beauregard–Vendon, France, 2016. [Google Scholar]
- National Oceanic and Atmospheric Administration US Department of Commerce National Weather Service.Climate 1130 Prediction Center. Climate & Weather Linkage. Available online: https://www.cpc.ncep.noaa.gov/ (accessed on 31 March 2022).
- Barstone, A.G.; Livezey, R.E. Classification, Seasonality and Persistence of Low–Frequency Atmospheric Circulation Patterns. Monthly Weather Review 1987, 115, 1083–1126. [Google Scholar] [CrossRef]
- Linderholm, H.W.; Folland, C.; Knight, J.; Fereday, D.; Ineson, S.; Hurrell, J.W. Influences of the Summer North Atlantic Oscillation (SNAO) on regional climate –from the past to the future. IOP Conference Series: Earth and Environmental Science 2009, 6, 072039. [Google Scholar] [CrossRef]
- Conte, M.; Giuffrida, A.; Tedesco, S. The Mediterranean Oscillation. Impact on precipitation and hydrology in Italy Climate Water; Publications of the Academy of Finland: Helsinki, 1989. [Google Scholar]
- Palutikof, J.P.; Conte, M.; Casimiro Mendes, J.; Goodess, C.M.; Espirito Santo, F. Climate and climate change. In: Brandt CJ, Thornes JB. (eds): Mediterranean desertification and land use; John Wiley and Sons: London, 1996. [Google Scholar]
- Palutikof, J.P.Analysis of Mediterranean climate data: measured and modelled. In: Bolle HJ. (eds): Mediterranean climate: Variability and trends; Springer–Verlag, Berlin, 2003.
- Nowakowski, J.K. Terms of autumn migration of the genus Sylvia in Central Poland. The Ring 1999, 21, 3–13. [Google Scholar]
- Redlisiak, M.; Remisiewicz, M.; Nowakowski, J.K. Long–term changes in migration timing of Song Thrush Turdus philomelos at the southern Baltic coast in response to temperatures on route and at breeding grounds. International Journal of Biometeorology 2018, 62, 1595–1605. [Google Scholar] [CrossRef] [PubMed]
- Remisiewicz, M.; Underhill, L.G. Climate in Africa Sequentially Shapes Spring Passage of Willow Warbler Phylloscopus trochilus across the Baltic Coast. PeerJ 2022, 10, e8770. [Google Scholar] [CrossRef] [PubMed]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; Münkemüller, T.; Mcclean, C.; Osborne, P.E.; Reineking, B.; Schroder, B.; Skidmore, A.K.; Zurell, D.; Lautenbach, S. Collinearity: A Review of Methods to Deal with It and a Simulation Study Evaluating Their Performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing. Available online: https://www.r–project.org (accessed on 11 February 2023).
- TIBCO Software Inc. 2017. Statistica (data analysis software system), version 13.3. Available online: http://statistica.io (accessed on 11 February 2023).
- Martin–Vide, J.; Lopez–Bustins, J. The western Mediteranean oscillation and rainfall in the Iberian Peninsula. Int. J. Climatol. 2006, 26, 1455–1475. [Google Scholar] [CrossRef]
- Cotton, P.A. Avian migration phenology and global climate change. Proceedings of the National Academy of Sciences of the United States of America 2003, 100, 12219–12222. [Google Scholar] [CrossRef]
- Stervander, M.; Lindstrom, A.; Jonzen, N.; Andersson, A. Timing of spring migration in birds: long–term trends, North Atlantic Oscillation and the significance of different migration routes. Journal of Avian Biology 2005, 36, 210–221. [Google Scholar] [CrossRef]
- Mills, A.M. Changes in the timing of spring and autumn migration in North American migrant passerines during a period of global warming. Ibis 2005, 147(2), 259–269. [Google Scholar] [CrossRef]
- Miller–Rushing, A.J.; Lloyd–Evans, T.L.; Primack, R.B.; Satzinger, P. Bird Migration Times, Climate Change, and Changing Population Sizes. Glob. Chang. Biol. 2008, 14, 1959–1972. [Google Scholar] [CrossRef]
- Butler, C. J. The disproportionate effect of global warming on the arrival dates of short–distance migratory birds in North America. Ibis 2003, 145, 484–495. [Google Scholar] [CrossRef]
- Sokolov, L.V.; Markovets, M.Y.; Shapoval, A.P.; Morozov, Y.G. Long–term trends in the timing of spring migration of passerines on the CourishSpit of the Baltic Sea. Avian Ecology and Behaviour 1998, 1, 1–21. [Google Scholar]
- Redlisiak, M.; Remisiewicz, M.; Nowakowski, J.K. Long–term changes in migration timing of Song Thrush Turdus philomelos at the southern Baltic coast in response to temperatures on route and at breeding grounds. International Journal of Biometeorology 2018, 62, 1595–1605. [Google Scholar] [CrossRef] [PubMed]
- Ehnbom, S.; Karlsson, L. , Ylvén, R., Åkesson, S. A. comparison of autumn migration strategies in Robins Erithacus rubecula at a coastal and an inland site in southern Sweden. Ring. & Migr. 1993, 14, 84–93. [Google Scholar]
- Chernetsov, N. Migratory stopovers of Wrens Troglodytes troglodytes on the south–eastern Baltic coast. Avian Ecology Behaviour 2010, 17, 13–22. [Google Scholar]
- Kristensen, M.W.; Tøttrup, A.P.; Thorup, K. Migration of the Common Redstart (Phoenicurus phoenicurus). The Auk 2013, 130(2), 258–264. [Google Scholar] [CrossRef]
- Payevsky, V. Age structure of passerine migrants at the eastern Baltic coast: The analysis of the ‘coastal effect’. Ornis Svecica 1998, 8(4), 171–178. [Google Scholar] [CrossRef]
- Koprowska, D. Wpływ temperatur na lęgowiskach i trasach migracji na terminy wiosennej i jesiennej wędrówki rudzika (Erithacus rubecula) przez Hel i Mierzeję Wiślaną. MSc thesis.Bird Migration Research Station, Faculty of Biology, University of Gdańsk, Poland, 2021.
- Rybant, M. Wpływ temperatur na lęgowiskach, trasach migracji i zimowiskach na terminy wiosennej i jesiennej wędrówki rudzików Erithacus rubecula przez stację Bukowo–Kopań. MSc thesis. Bird Migration Research Station, Facultyof Biology, University of Gdańsk, Poland, 2022.
- Newton, I. The Migration Ecology of Birds; Academic Press, London, 2008.
- Quinlan, S.E.; BoydR.L., *!!! REPLACE !!!*. Mist netting success in relation to weather. North American Bird Bander 1976, 1(4), 168–170. [Google Scholar]
- Gordo, O. Why Are Bird Migration Dates Shifting? A Review of Weather and Climate Effects on Avian Migratory Phenology. Clim. Res. 2007, 35, 37–58. [Google Scholar] [CrossRef]
- Koenig, W.D. Spatial Autocorrelation of Ecological Phenomena. Trends Ecol. Evol. 1999, 14, 22–26. [Google Scholar] [CrossRef]
- Patchett, R.; Kirschel, A.N.G.; Robins King, J.; Styles, P.; Cresswell, W. Age–related changes in migratory behaviour within the first annual cycle of a passerine bird. PLoS ONE 2022, 17(10), e0273686. [Google Scholar] [CrossRef]
- Kożuchowski, K.; Wibig, J. Zmienność oscylacji północnoatlantyckiej na podstawie wskaźnika NAO DJFM Hurrella (1864–2019). [Variation of the North Atlantic Oscillation According to Hurrell’s NAO DJFM Index (1864–2019)]. Przegląd Geofizyczny 2021, 66, 3–4. [Google Scholar] [CrossRef]
- Smith, A.D.; McWilliams, S.R. What to do when stopping over: behavioral decisions of a migrating songbird during stopover are dictated by initial change in their body condition and mediated by key environmental conditions. Behavioral Ecology 2014, 25(6), 1423–1435. [Google Scholar] [CrossRef]
- Newton, I. Can conditions experienced during migration limit the population levels of birds? J. Ornithol 2006, 147, 146–166. [Google Scholar] [CrossRef]
- World Climate Service. What is the Scandinavian Pattern Climate Index. Available online: https://www.worldclimateservice.com (accessed on 14 March 2023).
- Post, E.; Forchhammer, M.C.; Stenseth, N.C.; Callaghan, T.V. The timing of life–history events in a changing climate. Proc. R. Soc. Lond. 2001, 268, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Dossman, B.C.; Rodewald, A.D.; Studds, C.E.; Marra, P.P. Migratory birds with delayed spring departure migrate faster but pay the costs. Ecology 2023, 104, e3938. [Google Scholar] [CrossRef]
- Visser, M.E.; Perdeck, A.C.; van Balen, J.H.; Both, C. Climate change leads to decreasing bird migration distances. Global Change Biology 2009, 15(8), 1859–1865. [Google Scholar] [CrossRef]
- Vengerov, P.D. Effect of rise in spring air temperature on the arrival dates and reproductive success of the Song Thrush, Turdus philomelos (C.L. Brehm, 1831) in the forest–steppe of the Russian Plain. Russian Journal of Ecology 2017, 48, 178–184. [Google Scholar] [CrossRef]
- Newton, I. Advances in the study of irruptive migration. Ardea 2006, 94, 433–460. [Google Scholar]
- Schwabl, H. Winter and Breeding Territorial Behaviour and Levels of Reproductive Hormones of Migratory European Robins. Ornis Scandinavica 1992, 23, 271–276. [Google Scholar] [CrossRef]
- Armstrong, E.A. Territory in the Wren Troglodytes troglodytes. Ibis 1955, 98, 430–437. [Google Scholar] [CrossRef]
- Milošević, DD.; Savić, S.M.; Pantelić, M.; Stankov, U.; Žiberna, I.; Dolinaj, D.; Leščešen, I. Variability of seasonal and annual precipitation in Slovenia and its correlation with large–scale atmospheric circulation. Open Geosciences 2016, 8(1), 593–605. [Google Scholar] [CrossRef]
- Van Buskirk, J.; Mulvihill, R.S.; Leberman, R.C. Phenotypic plasticity alone cannot explain climate–induced change in avian migration timing. Ecology and Evolution 2012, 2(10), 2430–2437. [Google Scholar] [CrossRef]
- Davidson, L.J. The Impact of Climate Change on the Phenology of Short– and Long–Distance Migratory Birds. Undergraduate Thesis, Lakehead University, London, Thunder Bay, April 2021. [Google Scholar]
- Pulido, F.; Berthold, P. Current selection for lower migratory activity willdrive the evolution of residency in a migratory bird population. PNAS 2010, 107(16), 7341–7346. [Google Scholar] [CrossRef]
- Morganti, M. Birds facing climate change: a qualitative model for the adaptive potential of migratory behaviour. Rivista Italiana Di Ornitologia 2015, 85(1), 3–13. [Google Scholar] [CrossRef]
- Catry, P.; Campos, A.; Almada, V.; Cresswell, W. Winter segregation of migrant European robins Erithacus rubecula inrelation to sex, age and size. J. Avian Biol. 2004, 35, 204–209. [Google Scholar] [CrossRef]
- Prescott, D.R.; Middleton, A.L.A. Age and sex differences in winter distribution of American Goldfinches in eastern North America. Ornis Scandinavica 1990, 21, 99–104. [Google Scholar] [CrossRef]
- Adriaensen, F.; Dhondt, A.A. Population dynamics and partial migration of the European Robin (Erithacus rubecula) in different habitats. Journal of Animal Ecology 1990, 59. [Google Scholar] [CrossRef]
- Cristol, D. A.; Baker, M. B.; Carbone, C. Differential migration revisited: Latitudunal segergation by age and sex class. In book: Current Ornithology, Volume 15, Springer, New York, 1999.
- Catry, P.; Lecoq, M.; Conway, G.; Felgueiras, M.; King, J. M. B.; Hamidi, S. Are blackcaps Sylvia atricapilla differential distance migrants by sex? Ardeola 2006, 53, 31–38. [Google Scholar]
- Arizaga, J.; Alonso, D.; Barba, M. Patterns of migration and wintering of Robins Erithacus rubecula in northern Iberia. Ringing & Migration 2010, 25, 7–14. [Google Scholar] [CrossRef]
- Smallegange, I.M.; Fiedler, W.; Köppen, U.; Geiter, O.; Bairlein, F. Tits on the move: exploring the impact of environmental change on blue tit and great tit migration distance. Journal of Animal Ecology 2010, 79(2), 350–357. [Google Scholar] [CrossRef] [PubMed]
- Hedenström, A.; Alerstam, T. Optimum Fuel Loads in Migratory Birds: Distinguishing Between Time and Energy Minimization. J. theor. Biol. 1997, 189, 227–234. [Google Scholar] [CrossRef]
- Rogues, S.; Henry, P.Y.; Guyot, G.; Bargain, B.; Cam, E.; Pradel, R. When to depart from a stopover site? Time since arrival matters more than current weather conditions. Ornithology 2021, 139(1), 1–13. [Google Scholar] [CrossRef]
- Parrish, J.D. Patterns of frugivory and energetic conditio in nearctic landbirds during autumn migration. The Condor 1997, 99, 681–697. [Google Scholar] [CrossRef]
- Vyshkvarkova, E.; Sukhonos, O. Compound Extremes of Air Temperature and Precipitation in Eastern Europe. Climate 2022, 10, 133. [Google Scholar] [CrossRef]
- Nakbin, C.; Myong–In, L.; Dong–Hyun, C.; Young–Kwon, L.; Kyu–Myong, K. Decadal Changes in the Inter–anual Variability of Heatwaves in East Asia Caused by Atmospheric Teleconnection Changes. Journal of Climate 2020, 33, 1505–1522. [Google Scholar] [CrossRef]
Figure 1.
Geographical range of the European Wren, two ringing stations at the southern Baltic coast in Poland, where the analysed data was collected, and other ringing stations mentioned in the text (map after Trepte 2022 [51], modified).
Figure 1.
Geographical range of the European Wren, two ringing stations at the southern Baltic coast in Poland, where the analysed data was collected, and other ringing stations mentioned in the text (map after Trepte 2022 [51], modified).
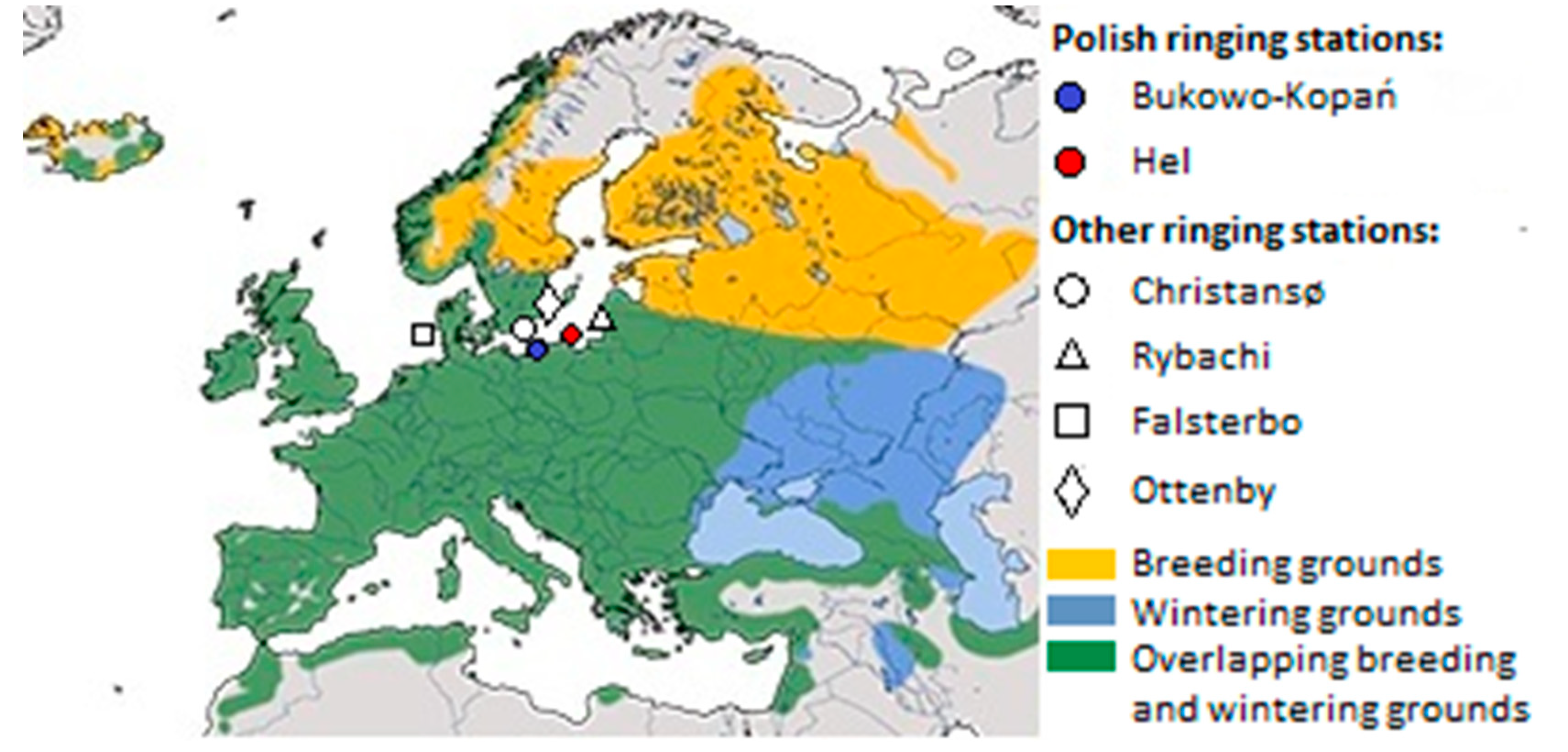
Figure 2.
Four main periods of the European Wren life cycle we used in analyses. Areas marked with transition between two colours indicate possible overlapping of the life stages.
Figure 2.
Four main periods of the European Wren life cycle we used in analyses. Areas marked with transition between two colours indicate possible overlapping of the life stages.
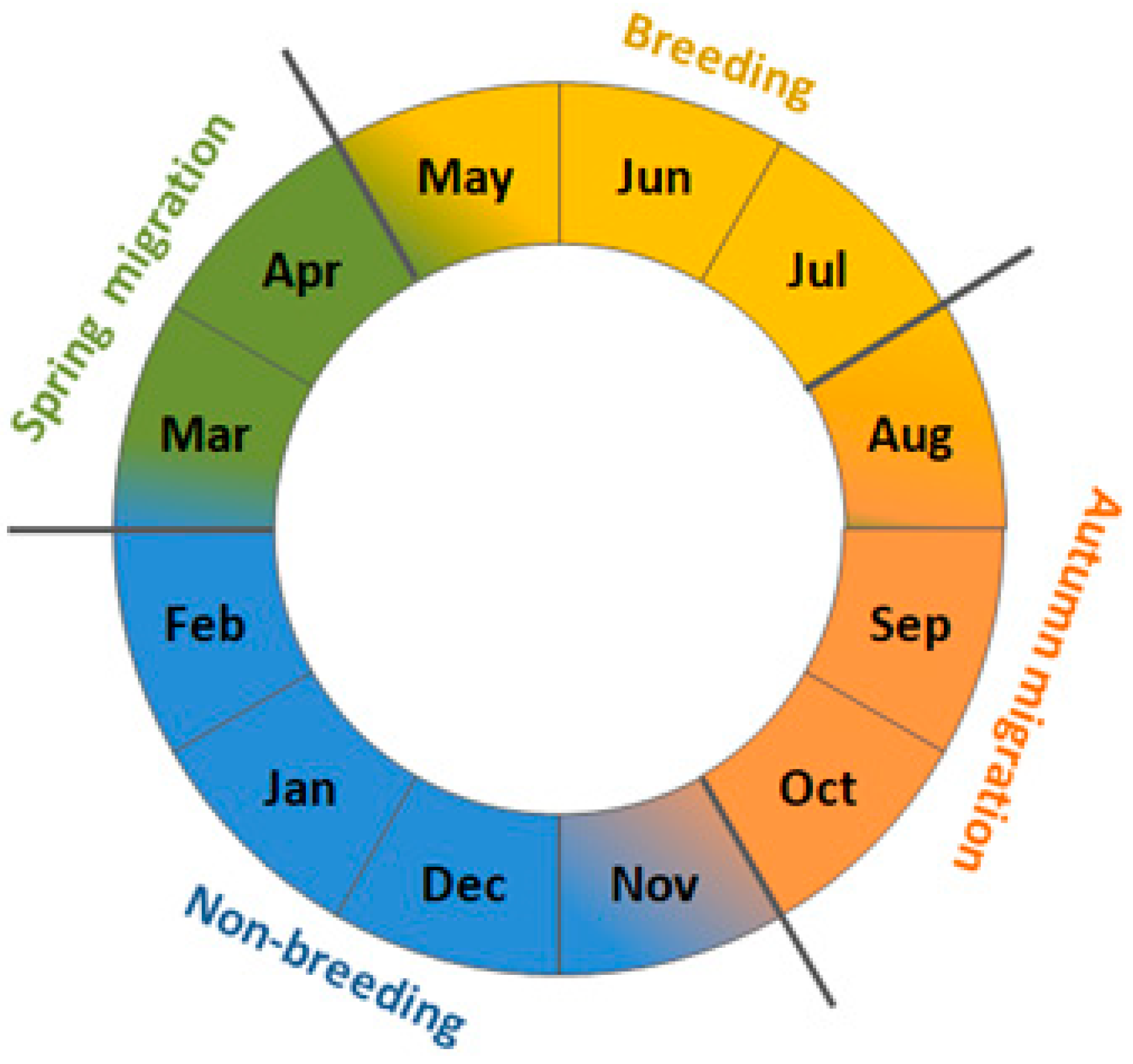
Figure 3.
Many-year trends in the spring migration ofthe European Wrenover 1982–2021, based on Annual Annomaly (AA) of passage at (A) Bukowo–Kopań and (B) Hel, and based on 5%, 50%, and 95% percentiles of passageat (C) Bukowo–Kopań and (D) Hel.Statistics of the regression equations in Table A3.
Figure 3.
Many-year trends in the spring migration ofthe European Wrenover 1982–2021, based on Annual Annomaly (AA) of passage at (A) Bukowo–Kopań and (B) Hel, and based on 5%, 50%, and 95% percentiles of passageat (C) Bukowo–Kopań and (D) Hel.Statistics of the regression equations in Table A3.
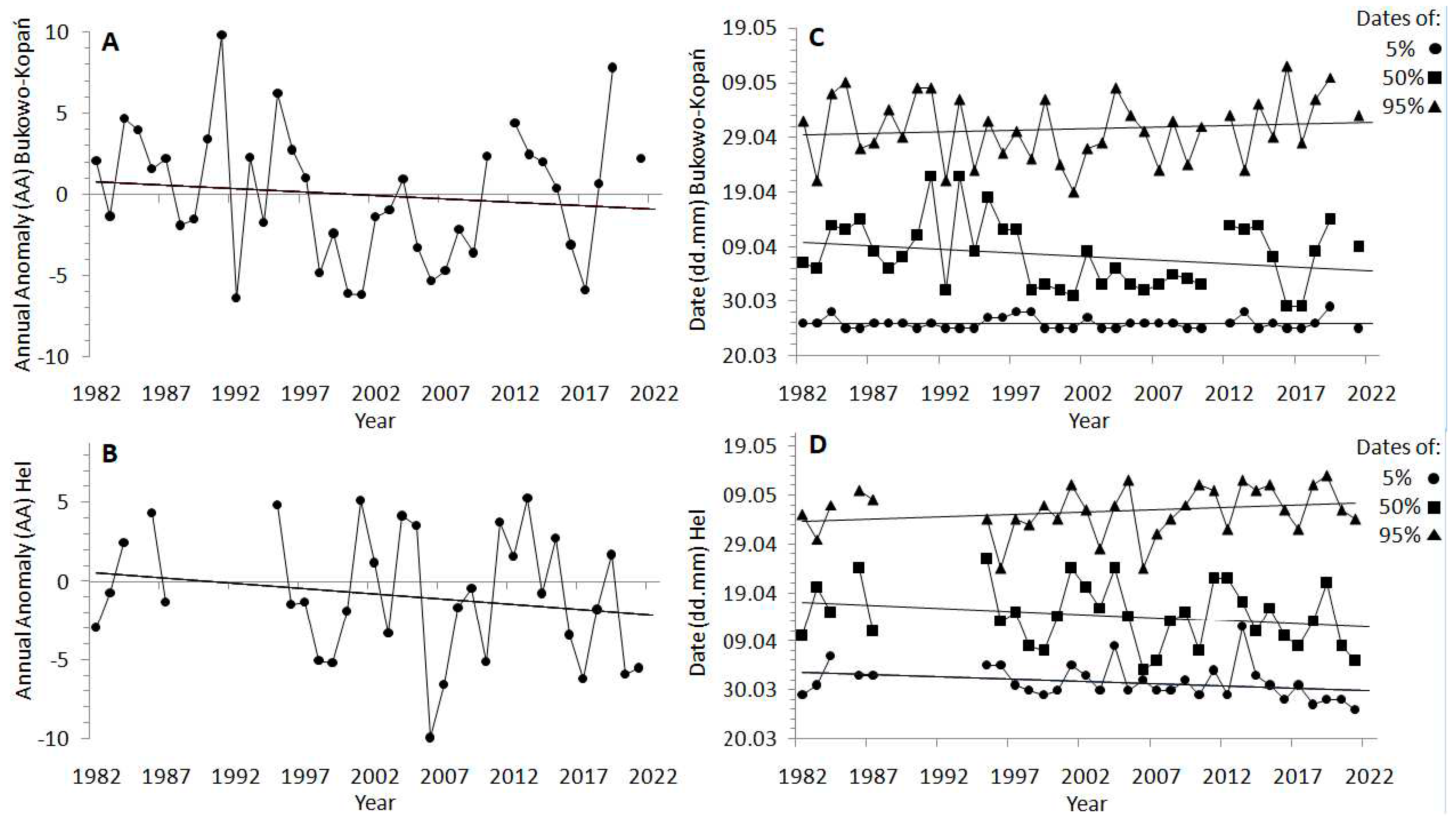
Figure 4.
Partial correlation coefficients for the Annual Anomaly (AA) and selected percentiles (5%, 50%,95%) of spring passage of the European Wren ringed in 1982–2021in N Poland, (A )at Bukowo–Kopań, and (B) at Hel, against the climate indices selected in the best multiple regression models. The details of these models are presented in Table 2.
Figure 4.
Partial correlation coefficients for the Annual Anomaly (AA) and selected percentiles (5%, 50%,95%) of spring passage of the European Wren ringed in 1982–2021in N Poland, (A )at Bukowo–Kopań, and (B) at Hel, against the climate indices selected in the best multiple regression models. The details of these models are presented in Table 2.
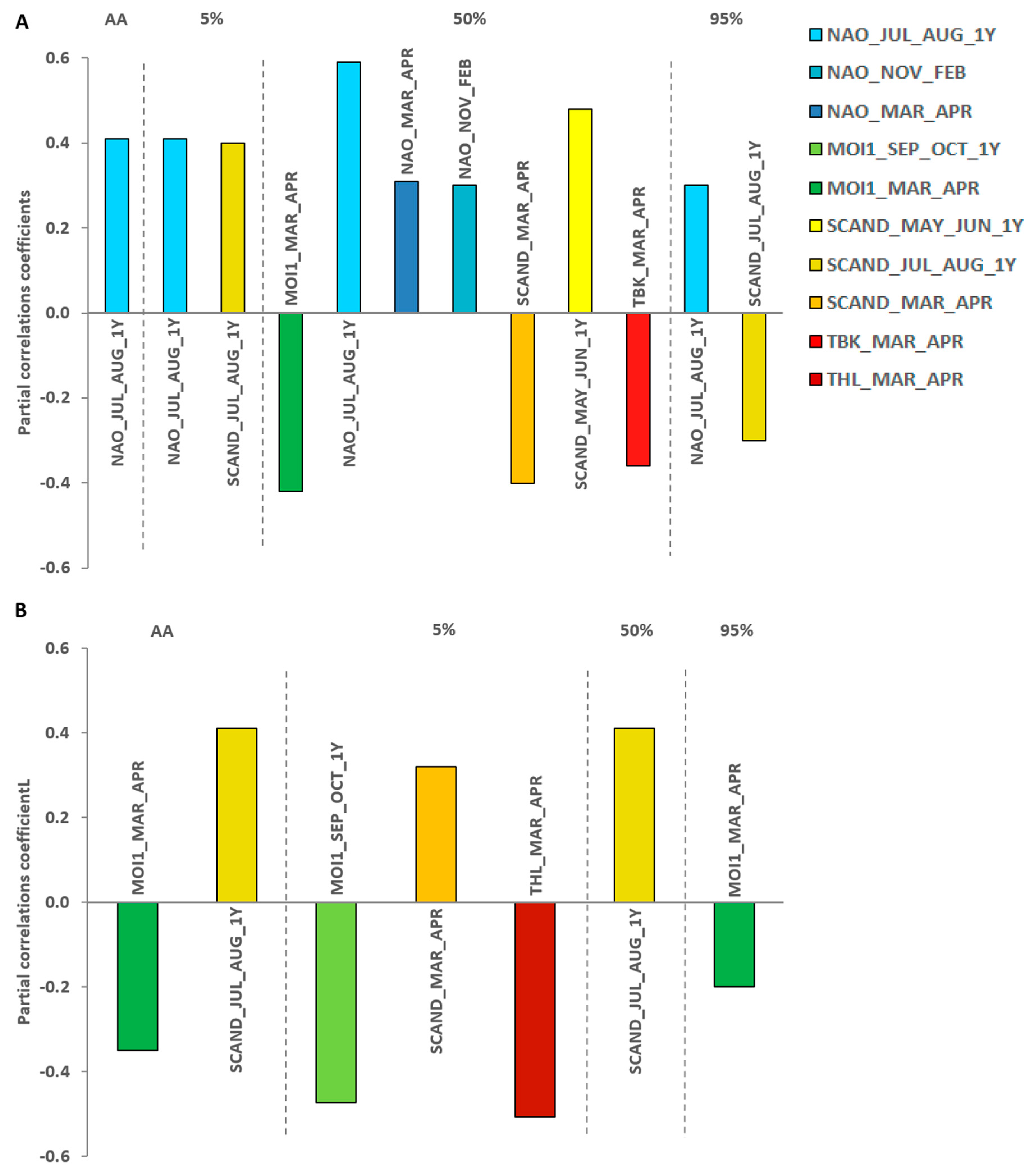
Table 1.
Climate variables and the “Year” used as explanatory variables in modelling the European Wren spring migration (26 March–15 May) over 1982–2021 at Bukowo-Kopań and Hel ringing stations, N Poland. Months = months corresponding with a life stage of the Wren (Figure 2), for which the climate index was averaged, “_1Y” = months of the year prior to the analysed spring migration.
Table 1.
Climate variables and the “Year” used as explanatory variables in modelling the European Wren spring migration (26 March–15 May) over 1982–2021 at Bukowo-Kopań and Hel ringing stations, N Poland. Months = months corresponding with a life stage of the Wren (Figure 2), for which the climate index was averaged, “_1Y” = months of the year prior to the analysed spring migration.
| No | Symbol | Climate Index | Months | Source of Data | |
|---|---|---|---|---|---|
| 1 | NAO_MAY_JUN_1Y | North Atlantic Oscillation Index |
May–Jun_1Y | https://www.cpc.ncep.noaa.gov/ products/precip/CWlink/pna/nao.shtml |
|
| 2 | NAO_JUL_AUG_1Y | Jul–Aug_1Y | |||
| 3 | NAO_SEP_OCT_1Y | Sep–Oct_1Y | |||
| 4 | NAO_NOV_FEB | Nov_1Y–Feb | |||
| 5 | NAO_MAR_APR | Mar–Apr | |||
| 6 | MOI1_SEP_OCT_1Y | Mediterranean Oscillation Index |
Sep–Oct_1Y | https://crudata.uea.ac.uk/cru/data/moi/ | |
| 7 | MOI1_NOV_FEB | Nov_1Y–Feb | |||
| 8 | MOI1_MAR_APR | Mar–Apr | |||
| 9 | SCAND_MAY_JUN_1Y | Scandinavian Pattern Index | May–Jun_1Y | https://ftp.cpc.ncep.noaa.gov/wd52dg/ data/indices/scand_index.tim | |
| 10 | SCAND_JUL_AUG_1Y | Jul–Aug_1Y | |||
| 11 | SCAND_MAR_APR | Mar–Apr | |||
| 12 | TBK_MAR_APR | Mean temperaturę anomaly for Koszalin and Łeba(near Bukowo-Kopań) | Mar–Apr | http://www.ecad.eu/ https://climexp.knmi.nl/ |
|
| 13 | THL_MAR_APR | Meant emperaturę anomaly at Hel | |||
| 14 | Year | 1982 = year 1 | Our data |
Table 2.
Relationships, as shown by the best models, between the climate variables and Annual Anomaly (AA), and selected percentiles (5%, 50%, 95%) of spring passage of the European Wren ringed at Bukowo–Kopań (BK) and Hel, N Poland, in 1982–2021. Estimate = coefficients from multiple regression, SE = standard error of the estimates; t, p = t–test and significance of each estimate, p<0.05 marked in bold face, VIF = variance inflation factor. R2= partial determination coefficients, pR =Pearson’s partial correlation coefficient (visualised at Figure 4A for Bukowo-Kopań and at Figure 4B for Hel).
Table 2.
Relationships, as shown by the best models, between the climate variables and Annual Anomaly (AA), and selected percentiles (5%, 50%, 95%) of spring passage of the European Wren ringed at Bukowo–Kopań (BK) and Hel, N Poland, in 1982–2021. Estimate = coefficients from multiple regression, SE = standard error of the estimates; t, p = t–test and significance of each estimate, p<0.05 marked in bold face, VIF = variance inflation factor. R2= partial determination coefficients, pR =Pearson’s partial correlation coefficient (visualised at Figure 4A for Bukowo-Kopań and at Figure 4B for Hel).
| Station/Response variable | Model statistics | Explanatory variable | Estimate | SE | t | p | VIF | R2 | pR | |
|---|---|---|---|---|---|---|---|---|---|---|
| Bukowo-Kopań | ||||||||||
| BK_AA | AdjR2=15%, F1,37= 7.79 | NAO_JUL_AUG_1Y | 0.42 | 0.15 | 2.79 | 0.01 | 1.00 | 0.17 | 0.41 | |
| BK_5% | AdjR2=28%, F2,36= 8.23 | NAO_JUL_AUG_1Y | 0.38 | 0.14 | 2.69 | 0.01 | 1.02 | 0.17 | 0.41 | |
| SCAND_JUL_AUG_1Y | 0.37 | 0.14 | 2.63 | 0.01 | 1.02 | 0.16 | 0.40 | |||
| BK_50% | AdjR2=40%, F7,31= 4.69 | MOI1_MAR_APR | –0.41 | 0.16 | –2.58 | 0.01 | 1.61 | 0.18 | –0.42 | |
| NAO_JUL_AUG_1Y | 0.56 | 0.14 | 4.04 | <0.00 | 1.21 | 0.34 | 0.59 | |||
| NAO_MAR_APR | 0.29 | 0.16 | 1.83 | 0.08 | 1.63 | 0.10 | 0.31 | |||
| NAO_NOV_FEB | 0.27 | 0.15 | 1.77 | 0.09 | 1.48 | 0.09 | 0.30 | |||
| SCAND_MAR_APR | –0.38 | 0.16 | –2.41 | 0.02 | 1.62 | 0.16 | –0.40 | |||
| SCAND_MAY_JUN_1Y | 0.46 | 0.15 | 3.03 | <0.00 | 1.44 | 0.23 | 0.48 | |||
| TBK_MAR_APR | –0.33 | 0.15 | –2.15 | 0.04 | 1.49 | 0.13 | –0.36 | |||
| BK_95% | AdjR2 = 10%, F2,36= 3.16 | NAO_JUL_AUG_1Y | 0.30 | 0.16 | 1.91 | 0.06 | 1.02 | 0.09 | 0.30 | |
| SCAND_JUL_AUG_1Y | –0.29 | 0.16 | –1.88 | 0.07 | 1.02 | 0.09 | –0.30 | |||
| Hel | ||||||||||
| HL_AA | AdjR2 = 16%, F2,30= 4.02 | MOI1_MAR_APR | –0.35 | 0.17 | –2.08 | 0.05 | 1.09 | 0.13 | –0.35 | |
| SCAND_JUL_AUG_1Y | 0.41 | 0.17 | 2.45 | 0.02 | 1.09 | 0.17 | 0.41 | |||
| HL_5% | AdjR2 = 39%, F3,29 = 6.30 | MOI1_SEP_OCT_1Y | –0.44 | 0.15 | –2.90 | 0.01 | 1.11 | 0.22 | –0.47 | |
| SCAND_MAR_APR | 0.28 | 0.15 | 1.82 | 0.08 | 1.10 | 0.10 | 0.32 | |||
| THL_MAR_APR | –0.48 | 0.15 | –3.17 | <0.00 | 1.09 | 0.26 | –0.51 | |||
| HL_50% | AdjR2 = 14%, F1,31 = 6.19 | SCAND_JUL_AUG_1Y | 0.41 | 0.16 | 2.49 | 0.02 | 1.00 | 0.17 | 0.41 | |
| HL_95% | AdjR2 = 1%, F1,31 = 1.33 | MOI1_MAR_APR | –0.20 | 0.18 | –1.15 | 0.26 | 1.00 | 0.04 | –0.20 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



