
Submitted:
30 May 2023
Posted:
30 May 2023
You are already at the latest version
Abstract
Cancer cells cannot proliferate and survive unless they obtain sufficient levels of the 20 proteinogenic amino acids (AAs). Unlike normal cells, cancer cells have genetic and metabolic alterations that may limit their capacity to obtain adequate levels of the 20 AAs under challenging metabolic environments. However, since normal diets provide all AAs at relatively constant levels and ratios, these potentially lethal genetic and metabolic defects are eventually harmless to cancer cells. If we temporarily replace the normal diet of cancer patients with artificial diets in which the levels of specific AAs are manipulated, cancer cells may be unable to proliferate and survive. This article reviews in vivo studies that have evaluated the antitumor activity of diets restricted or supplemented with the 20 proteinogenic AAs, individually and in combinations. It also reviews our recent studies that show that manipulating the levels of several AAs simultaneously can lead to marked survival improvements in mice with metastatic cancers.
Keywords:
cancer metabolism
; anticancer activity
; artificial diets
; in vivo
; mice
; essential amino acids
; non-essential amino acids
; restriction
; leucine
; methionine
; cysteine
; arginine
; serine
; glutamine
; asparagine
1. Introduction
The first metabolic alteration of cancer cells was discovered almost one century ago by the German biochemist Otto Warburg. He observed that, unlike normal cells, cancer cells converted high amounts of glucose into lactate in the presence of normal oxygen levels [1]. This alteration in glucose metabolism, known as aerobic glycolysis or Warburg effect, is now widely used in diagnostic imaging to trace cancers and evaluate cancer treatment response [2,3]. Clinical use of FDG-based PET imaging continually shows that most primary and metastatic cancers have a significant increase in glucose uptake when compared to normal tissues [2,3].
During many decades, the Warburg effect was considered an irrelevant oddity of cancer cells, probably because it was unknown why cancer cells use this primitive form of energy production when the availability of oxygen allows for a much more effective way of producing energy: oxidative phosphorylation. The explanation of the Warburg effect is simple when one realizes that glycolysis not only serves to produce energy, but also to generate building blocks to generate new cells [4]. Cancer cells have high glycolytic rates because the breakdown of glucose molecules generates building blocks needed to produce many cellular components for the new cancer cells created during cell division. One cell cannot divide to produce two cells unless glucose is broken down into these building blocks. Since both glucose and oxygen are supplied together through the blood, cancer cells have no choice but to activate glycolysis in the presence of oxygen in order to proliferate. Since oxygen inhibits glycolysis (Pasteur Effect) indirectly via ATP generation, cancer cells partially uncouple oxygen utilization from ATP production to activate glycolysis in the presence of normal oxygen levels. By deviating oxygen metabolism from the route that generate ATP to the route that produces reactive oxygen species (ROS), cancer cells manage to keep sustained glycolytic rates under aerobic conditions [4,5,6]. The subsequent increased production of ROS, such as superoxide anion and hydrogen peroxide, leads to a state of increased basal oxidative stress, which represents another metabolic hallmark of cancer cells [5,7,8,9,10].
Targeting the Warburg effect for cancer therapy is difficult because cells from different normal tissues also need glucose for their survival and proliferation. However, understanding the Warburg effect is important to realize that the genetic alterations of cancer cells are insufficient for cancer cell proliferation and survival. Cancer cells also need to take glucose and other nutrients, such as amino acids (AAs), from the extracellular environment to proliferate and survive under conditions of elevated oxidative stress. Although the metabolic changes of cancer cells play an important role in carcinogenesis and cancer progression, these changes can also be exploited to develop new cancer therapies [10,11,12,13,14,15].
The altered AA metabolism of cancer cells is one of most therapeutically relevant metabolic features of cancer. Several excellent reviews have summarized the role of AA metabolism in cancer development and the potential of targeting AA metabolism for therapeutic intervention [16,17,18,19,20,21]. Briefly, cancer cells have elevated requirements of some AAs to maintain the high biosynthetic and bioenergetic demands of cell proliferation [18]. In addition, many cancer cells are unable to synthesize sufficient levels of certain non-essential AAs (NEAAs) [21] and depend on their external supply to maintain their cellular functions. Several dietary and pharmacological interventions have been developed to target the altered AA metabolism of cancer cells [22]. For example, L-asparaginase (ASNase) is a clinically useful anticancer drug that depletes the NEAA asparagine (Asn) from the blood and selectively kills leukemia cells that cannot biosynthesize this AA [23]. Several AA-depleting enzymes, and many small-molecule drugs targeting AA uptake or their metabolic pathways, are currently under preclinical and clinical development [13,21]. The altered AA metabolism of cancer cells can also be targeted without drugs, through dietary manipulation of the levels of certain AAs [24,25,26,27].
In this work, we review studies that have evaluated the cancer therapeutic potential of dietary AA manipulation in vivo. Since dietary proteins are the primary source of AAs for cancer cells, we first briefly review key studies showing that protein restriction can inhibit tumor growth. Then, we review in vivo studies assessing the antitumor activity of dietary strategies based on restricting or increasing the levels of each of the 20 proteinogenic AAs, beginning with the 9 EAAs and ending with the 11 NEAAs (Figure 1). Finally, we discus recent studies showing that manipulating the levels of several AAs simultaneously can lead to marked survival improvements in mice with different types of metastatic cancers.
2. Protein restriction
Dietary protein restriction can increase life expectancy [28] and reduce the incidence of age-related diseases such as cancer [29]. It is well known that proliferating cancer cells must produce new proteins for the new cells created during cell division. Since dietary proteins provide the AAs needed to generate the proteins of the new cancer cells, it is not surprising that low-protein diets can restrict tumor growth in animal models [30]. Table 1 summarizes several studies that evaluated the effect of changing the amount and type of protein in the diet on cancer progression in mice [31,32,33,34,35,36].
A reduction in IGF-1 levels has been proposed as a key mechanism by which low-protein diets induce anticancer activity. Murine models of melanoma and breast cancer revealed that mice fed a low protein diet (4% kcal protein) had reduced IFG-1 levels and reduced tumor progression compared to those fed a high protein diet (18% kcal protein) [31]. Weight loss was observed in elderly mice but not in young mice. Similarly, a low protein diet reduced IGF-1 levels in patients aged 50-65 years and reduced the risk of cancer death, while a low-protein diet increased mortality among elderly patients (+65 years old) [31]. This suggests that low protein diets might induce anticancer activity in middle-aged adults but not in elderly patients. Low protein diets are not active in all cancer types. For example, experiments in a syngeneic glioma model showed no reductions in tumor growth in animals fed a low protein diet (4% kcal) when compared to animals fed a high protein diet (18% kcal protein) [32].
Enhanced cancer immunosurveillance is another possible mechanism by which low protein diets induce anticancer activity. A reduction in dietary proteins (17-15% protein vs 19% protein) induced IRE1α-dependent endoplasmic reticulum (ER) stress in cancer cells, which resulted in cytokine production and improved anticancer immune response [33]. Lower protein intake (12%) reversed the anticancer effect, which suggested that a certain level of protein intake was needed for activity [33]. However, other studies showed that diets with lower protein intake (7% protein vs. 21% protein) inhibited cancer progression and induced a synergistic effect when combined with anti-PD-1 immunotherapy [34]. Low protein diets also induced anticancer activity in immunosuppressed mice, therefore suggesting that the anticancer activity of protein restriction is not necessarily mediated by the immune system [35]. For example, a low protein diet (7% vs 21% protein diets) reduced cancer progression in immunodeficient mice implanted with human breast and prostate cancer cells [35].
The type of protein can also modulate cancer progression. Mice fed a 20% plant-based protein diet showed a reduced tumor growth in xenograft and syngeneic cancer models when compared to mice fed a 20% animal-based protein diet [35,36]. The anticancer activity of diets based on plant proteins was explained by declines in the plasma levels of IFG -1 and insulin, which decreased the activity of the IGF/AKT/mTOR pathway and led to epigenetic modifications that restricted tumor growth [35,36]. Since animal and plant proteins have different AA levels, it is important to identify which individual AAs are involved in the anticancer activity of diets based on plant proteins. Understanding the anticancer effect induced by restriction of each AA may be useful to develop more effective diets for cancer therapy.
3. Essential Amino Acids
3.1. Leucine
Leucine (Leu) is one of the nine EAAs for humans; this means that we cannot biosynthesize it from other nutrients and that we must take it from the diet. Like all 20 proteinogenic AAs, Leu is necessary for protein synthesis. Leu is also important for other cellular functions. For example, Leu is a key intracellular sensor of AAs under starvation conditions, and it regulates protein turnover through mTORC1 signaling [37]. Like isoleucine (Ile) and valine (Val), Leu is a branched-chain amino acid (BCAA); these AAs can regulate lipid metabolism in cancer cells by providing carbon skeletons for fatty-acid biosynthesis [38].
Dietary restriction of Leu can induce in vivo anticancer effects. In 1956, Sugimura et al. [39] found that dietary deprivation of Leu for 5 days reduced the growth rate of Walker tumors in rats by 24%; however, it also induced body weight loss. In 1971, reducing dietary levels of Leu from 0.8% to 0.1% for 3 weeks significantly reduced tumor growth in mice with breast adenocarcinoma; tumor weights were 32 ± 10 g in mice fed a standard AA-based diet (0.80% Leu), 38 ± 4 g for a 0.50% Leu diet, 32 ± 4 g for a 0.25% Leu diet, and 16 ± 6 g for a 0.10% Leu diet [40]. Body weight loss was observed in mice fed the 0.10% Leu diet [40]. More recent studies have shown that 14 days on a Leu-free diet combined with an autophagy inhibitor induced anticancer activity in mice with melanoma xenografts without causing significant toxicity [41]. Only 4 days of a Leu-free diet was sufficient to induce anticancer activity in mice with triple negative breast cancer xenografts [42].
Mechanistically, Leu limitation restricts protein synthesis, cell division and tumor growth. In addition, Leu restriction can reduce Leu catabolism and limit fatty acid biosynthesis and lipogenesis in cancer cells. BCAAs catabolism plays an important role in pancreatic cancer growth by regulating lipogenesis [38]. BCAT2 and BCKDHA knockdown impaired pancreatic cancer cell proliferation in vitro and in vivo by inhibiting fatty acid synthesis [38]. Furthermore, inhibition of BCAT1, the first enzyme in the catabolism of BCAAs, induced anticancer activity in vitro and in vivo [43,44,45,46,47,48]. Leu restriction also decreased the expression of the enzyme fatty acid synthase (FASN) [42]; FASN overexpression or palmitic acid supplementation (the product of FASN) blocked the anticancer activity of Leu restriction [42].
Since Leu restriction can reduce tumor growth, it makes sense to think that Leu supplementation may facilitate cancer progression. A study showed that 5% Leu supplementation increased cancer growth in a syngeneic model of pancreatic cancer [49]. However, our recent studies in mice with different types of metastatic cancers indicate that supplementing Leu can increase the anticancer activity of diets deficient in other AAs [26,27]. Supplementing 2.5% Leu to several casein-based artificial diets markedly improved their anticancer activity [26,27]. Importantly, the activity of these artificial diets in mice with metastatic cancers were higher than the observed in mice treated with the standard therapies used in cancer patients [26,27].
Maintaining high Leu levels may be important to prevent proteolysis, which could be beneficial in certain circumstances. Leu is a critical intracellular sensor of AAs under starvation conditions. This AA activates mTORC1 signaling and inhibits autophagy and proteasome-mediated proteolysis. Supplementing Leu may therefore prevent intracellular and extracellular proteolysis [50,51,52,53]. If muscle and liver proteolysis is not prevented, the lysis of proteins in these organs would supply any AA restricted in the diet [26,27,54]. Inhibition of proteolysis is also important to avoid weight loss and cachexia. Cachexia is a syndrome of progressive body weight loss with reduction in skeletal muscle and fat mass [55]. Ultimately, cachexia reduces the tolerability of anticancer treatments and leads to a reduced life expectancy and quality of life [55,56]. Leu supplementation can alleviate cancer cachexia by activating mTORC1 and decreasing protein degradation [51,55,57,58]. Several preclinical studies have shown that 3% Leu supplementation can ameliorate cancer cachexia in the Walker-256 rat model [59,60,61,62,63,64,65,66] and in the C26 murine model [67]. The anti-cachectic effect of Leu supplementation can be improved with fish oil supplementation [68], glutamine (Gln) supplementation [64], and aerobic physical exercise [63,64]. Evidence suggests that supplementation of 3% Leu is sufficient to improve cachexia [55,59,60,61,62,63,64,65,66,67,68].
3.2. Isoleucine
Like all proteinogenic AAs, the BCAA Ile is necessary for protein synthesis. Ile also participates in other biological processes, including lipogenesis and immune function regulation [37,38,69]. Experiments conducted several decades ago revealed that complete dietary Ile restriction for 5 days inhibited tumor growth by 40% in Walker tumor bearing rats; however, this force-fed intervention caused the animals to lose 1-2 g per day [39]. Dietary Ile restriction (from 0.5% to 0.05%) also resulted in tumor growth inhibition in C57BL/6 mice with BW10232 mammary carcinomas [40]. Tumor weights were 32 ± 10 g in mice fed a standard AA-based diet (0.50% Ile), 31 ± 5 g for a 0.30% Ile diet, 17 ± 7 g for a 0.15% Ile diet, and 7 ± 3 g for a 0.05% Ile diet [40]. The tumors of mice fed the 0.15% Ile diet were significantly smaller, whereas the final tumor-free weight of the mice was relatively unaffected. Both tumor weight and final tumor-free weight were significantly reduced in mice fed the 0.05% Ile diet. This means that moderate Ile restriction was sufficient to reduce tumor growth without significantly decreasing mice body weight [40]. Mechanistically, Ile limitation restricts protein synthesis, cell division and tumor growth. Ile restriction can also reduce Ile catabolism and limit fatty acid biosynthesis and lipogenesis in cancer cells [38,43,44,45,46,47,48].
3.3. Valine
Like Leu and Ile, Val is an essential and proteinogenic BCAA. It is also involved in other cellular functions, including regulation of lipid and glucose metabolism [37,38,70]. Dietary depletion of Val for 5 days reduced tumor growth by 41% in Walker tumor bearing rats [39]. However, all the animals on this Val-free diet rapidly sickened and failed to survive beyond 9 days on this diet. Dietary limitation of Val (from 0.7% to 0.1%) significantly decreased tumor growth in mice with breast adenocarcinoma, but also induced body weight loss [40]. Tumor weights were 32 ± 10 g in mice fed a standard AA-based diet (0.70% Val), 36 ± 5 g for a 0.40% Val diet, 25 ± 8 g for a 0.20% Val diet, and 16 ± 6 g for a 0.10% Val diet [40]. With the 0.20% Val diet, a reduction in tumor weight was achieved with little reduction in the final tumor-free weight of the mice. Reducing the Val levels further, to 0.10 %, significantly reduced tumor weight as well as mice tumor-free weight [40]. Mechanistically, Val limitation restricts protein synthesis, cell division and tumor growth. Like the other two BCAAs, Val restriction can reduce Val catabolism and therefore limit the production of carbon skeletons for fatty acid biosynthesis in cancer cells [38,43,44,45,46,47,48].
3.4. Threonine
Threonine (Thr) is an essential and proteinogenic AA. Like other AAs, Thr catabolism can also provide amino groups for the synthesis of NEAAs and carbon skeletons for biosynthesis and energy production [37]. Force feeding of a diet lacking Thr for 5 days reduced tumor growth by 28% in Walker tumor-bearing rats [39]. This diet caused the animals to lose between 0.2 and 1.0 g./day over an 11-day period [39]. Another study revealed that dietary limitation of Thr for 3 weeks significantly decreased cancer growth in mice with breast adenocarcinoma [40], but also caused weight loss. Tumor weights were 32 ± 10 g in mice fed a standard AA-based diet (0.50% Thr), 37 ± 6 g for a 0.30% Thr diet, 30 ± 7 g for a 0.15% Thr diet, and 15 ± 5 g for a 0.05% Thr diet [40]. In a group of mice not inoculated with the breast adenocarcinoma cells, feeding the 0.05% Thr diet for three weeks produced a 31% weight loss [40]. Thr limitation restricts protein synthesis, cell division and tumor growth.
3.5. Lysine
Lys is an essential and proteinogenic AA, whose deficiency can trigger severe malnutrition [37,71]. Lys is also used for carnitine production and participates in protein methylation, acetylation, ubiquitination and glycosylation [37]. The anticancer activity of Lys restriction was evaluated 80 years ago in mice with spontaneous breast cancer [72]. The author of this research first devised a Lys-deficient diet suitable for human consumption (palatable, adequate in calories, minerals and vitamins, and sufficient for keeping nitrogen balance). After observing in two healthy humans that nitrogen equilibrium could be maintained with this diet, he obtained and reproduced a strain of mice characterized by a high incidence of spontaneous mammary carcinomas. The mice that developed tumors were fed with the Lys-deficient diet. The diet inhibited the growth rate of the tumors, but also the rate of normal growth in mice. These inhibitory effects were abolished upon the addition of Lys, therefore indicating that Lys was essential for both normal and malignant growth. When the Lys-deficient diet was fed for several weeks, the antitumor effect wore off and the tumors resumed rapid growth. In addition, the inhibitory effect was either not apparent or very short when the Lys-deficient diet was started in mice with tumors that had reached an advanced stage of growth. The author concluded that the therapeutic potential of the Lys-deficient diet was low [72]. In 1959, force feeding of a diet lacking Lys for 5 days did not reduce tumor growth in Walker tumor-bearing rats [39]. In 1971, Lys limitation (from 0.6% to 0.15%) did not significantly reduce tumor growth in C57BL mice with BW10232 mammary carcinomas [40]. Tumor weights were 36 ± 13 g in mice fed a standard AA-based diet (0.90% Lys), 36 ± 6 g for a 0.60% Lys diet, 36 ± 6 g for a 0.30% Lys diet, and 31 ± 8 g for a 0.15% Lys diet [40].
3.6. Phenylalanine
Phenylalanine (Phe) is an essential and proteinogenic AA with an aromatic group in its structure. Phe can be used to synthesize tyrosine (Tyr), a proteinogenic NEAA that produces important molecules such as catecholamines (dopamine, epinephrine, and norepinephrine) and melanin [37]. Dietary Phe limitation is used in people with phenylketonuria, an inborn disease caused by inactivity of the enzyme phenylalanine hydroxylase, which converts Phe into Tyr; the accumulation of Phe can lead to seizures and intellectual disability [73].
In 1959, force feeding of a diet lacking Phe for 5 days was found to reduce tumor growth by 15% in Walker tumor-bearing rats [39]. Several years later, a Phe-deficient diet (0.12% Phe) was reported to reduced tumor growth by 23% in C57L/J mice with BW7756 hepatoma and 32% in C3H/HeJ mice with C3HBA mammary adenocarcinoma [74]. In combination with ρ-fluorophenylalanine (a metabolic analog of Phe), the Phe-deficient diet reduced tumor growth by 94% in BW7756 hepatoma and 42% in C3HBA mammary adenocarcinoma [74].
Since Phe is a precursor of Tyr, dual restriction of Phe and Tyr was evaluated in several studies conducted between the 1960s and the early 2000s. In 1966, a diet with 0.12% Phe and 0.06% Tyr was found to reduce the growth of melanoma (but not of sarcoma) in mice [75]. In 1971, dietary restriction of Phe and Tyr reduced tumor growth in C57BL mice with BW10232 mammary carcinomas [40]. Tumor weights were 41 ± 9 g in mice fed a standard AA-based diet (0.60% Phe + 0.30% Tyr), 35 ± 10 g for a 0.40% Phe + 0.20% Tyr diet, 29 ± 6 g for a 0.20% Phe + 0.10% Tyr diet, and 11 ± 5 g for a 0.10% Phe + 0.05% Tyr diet [40]. In a group of mice not inoculated with the breast cancer cells, feeding the 0.10% Phe + 0.05% Tyr diet for three weeks produced a 21% weight loss [40]. Dietary limitation of Phe and Tyr also showed anticancer activity in mice with breast cancer but not fibrosarcoma [76]. A diet with 0.08% Phe and 0.04% Tyr reduced the metastatic potential of cancer cells in several in vivo models, including melanoma, lung, and hepatocarcinoma [77,78]. In vitro experiments using different types of cancer cells support the in vivo anticancer activity of Phe and Tyr limitation [79,80,81,82,83,84,85,86].
In humans, several case reports have shown reductions in tumor bulk and regression of lymph nodes in patients with malignant melanoma, Hodgkins lymphoma and cancer of the uterus [87,88]. Stabilization of choroidal malignant melanoma has also been reported [89]. In 1985, no tumor responses were observed in three patients with disseminated malignant melanoma who received a low Phe/Tyr diet for two months [90]. In 2002, 3 patients with metastatic melanoma and 3 patients with metastatic breast cancer agreed to consume a low protein diet providing approximately 10 mg/kg Phe/Tyr per day; the diet was based on several fixed products complemented with different foods [91]. A possible decline in the rate of disease progression was observed in one patient with metastatic melanoma; this patient had a prognosis of 8 weeks at recruitment, but survived a further 7 months after stopping the low Phe/Tyr diet [91]. All the patients of this pilot study experienced side effects such as increases in anxiety and depression [91].
3.7. Histidine
Histidine (His) is an aromatic EAA required for protein synthesis. This AA is involved in other cellular functions, including the synthesis of histamine and carnitine [37,92]. Force feeding of a diet lacking His for 5 days reduced tumor growth by 19% in the Walker rat model [39]. More recently, dietary limitation of His was found to selectively limit the growth of MYC-dependent neural tumors in a Drosophila model [93]. In contrast, supplementing His can activate His catabolism, which consumes tetrahydrofolate and increase the anticancer activity of methotrexate by reducing the tetrahydrofolate cellular pool [94]. This study found that administration of His (injection of 18.4 mg His) significantly increased the anticancer activity of methotrexate in mice xenografted with human leukemia cells [94].
3.8. Tryptophan
Although tryptophan (Trp) is the least abundant EAA in the diet, it is necessary for protein synthesis and for the production of a variety of biologically active compounds, including serotonin, melatonin, and niacin (a component of NAD and NADP) [37,95]. In addition, Trp and its catabolic derivatives modulate the immune function and play a key role in autoimmune diseases and antitumor immunity [95,96].
In 1959, total Trp restriction for 5 days inhibited tumor growth by 19% in Walker tumor bearing rats, with moderate weight loss in the animals [39]. In 1971, Trp limitation (from 0.10% to 0.02%) reduced tumor growth in C57BL/6 mice with BW10232 mammary carcinomas [40]. Tumor weights were 33 ± 6 g in mice fed a standard AA-based diet (0.15% Trp), 33 ± 13 g for a 0.10% Trp diet, 31 ± 10 g for a 0.05% Trp diet, and 16 ± 8 g for a 0.02% Trp diet [40]. Mice fed 0.02% Trp diet lost 28% of their weight in 3 weeks [40]. Moderate dietary limitation of Trp (0.05%) did not show anticancer activity in C3H mice bearing mammary adenocarcinomas [97].
Recent research on Trp and cancer therapy has focused on a catabolic pathway known as the kynurenine (Kyn) pathway, in which Trp is catabolized into Kyn by the enzymes indoleamine-2,3-dioxygenase 1 and 2 (IDO1/2) and tryptophan-2,3-dioxygenase (TDO2) [96]. Tumor and myeloid cells in the tumor microenvironment are known to metabolize Trp to Kyn [95]. The drop in Trp levels and the increase in the levels of metabolites of the Kyn pathway can lead to an immunosuppressive state that supports cancer survival [95,96,98,99]. For example, the antitumor activity of anti-PD1 immunotherapy was reduced in mice fed a low Trp diet [100]. Trp limitation may therefore facilitate cancer progression by impairing cancer immunosurveillance. Several IDO1 and TDO2 inhibitors enhanced the anticancer activity of checkpoint inhibitors in preclinical studies, and some of them have entered clinical trials, including epacadostat (phase I to III), BMS-986205 (phase I-II), indoximod (phase II) and navoximod (phase I) [95,96,101]. Unfortunately, epacadostat and other compounds have yielded disappointing clinical results. Epacadostat plus pembrolizumab did not improve progression-free survival and overall survival compared to pembrolizumab alone in a phase III clinical trial with 706 melanoma patients [102]. More research is needed to fully understand the relevance of Trp metabolism in cancer progression and immunity.
3.9. Methionine
Methionine (Met) is an essential and proteinogenic AA that contains a sulfur atom in its structure. Met is the precursor of S-adenosyl methionine (SAM), which is a methyl donor involved in DNA methylation and epigenetics. Met also produces Cys through the irreversible transsulfuration pathway, which in turn produces several sulfur-containing molecules with important cellular roles, including glutathione (GSH), hydrogen sulfide (H2S), and taurine (Tau) [24,37,103,104].
Dietary Met restriction has shown anticancer activity in numerous preclinical studies [26,27,34,39,40,105,106,107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125]. Dietary Met depletion (0%) induced anticancer activity in rats [39,121,122,123,124] and mice [105,106,107,108,109,110,111,125] with different types of cancer. Force feeding of a diet lacking Met for 5 days reduced tumor growth by 39% in the Walker rat model, but this diet caused the animals to lose l-2 g weight/day [39]. Met restriction (from 0.60% to 0.10%) reduced tumor growth in C57BL mice with BW10232 mammary carcinomas [40]. Tumor weights were 36 ± 13 g in mice fed a standard AA-based diet (0.90% Met; 0.2% Cys), 33 ± 11 g for a 0.60% Met diet, 30 ± 6 g for a 0.40% Met diet, 29 ± 7 g for a 0.20% Met diet, and 16 ± 7 g for a 0.10% Met diet [40]. Mice fed the 0.10% Met diet lost 10% of their initial weight in 3 weeks [40]. Several studies suggest that Met intake can be reduced to 0.12% without causing significant protein loss or noticeable toxicities in healthy animals [126,127,128]. Limiting Met intake to 0.17-0.12% did not significantly decrease body weight of mice with different types of cancer [112,113,114,115,116]. Since Met is necessary to biosynthesize Cys, the dietary levels of Cys can condition the dietary requirements of Met. Dietary restriction of Met can increase the antitumor effect of a variety of drugs, including 5-fluorouracil [106,114,117,121,124], anti-PD-1 immunotherapy [34,118], vincristine [122], cisplatin [105], lexatumimab (TNFα receptor agonist) [109], auranofin (TXNRDs inhibitor) [110], ethionine (Met analog) [123], and radiotherapy [114]. Table 3 shows representative studies assessing the in vivo anticancer activity of Met restriction.
Mechanistically, Met limitation restricts protein synthesis, cell division and tumor growth. Met restriction can also reduce the cellular levels of the methyl donor SAM, which may alter DNA methylation and epigenetics in dividing cancer cell. Met restriction can also compromise the biosynthesis of polyamines, which are involved in several key processes of cell growth and survival, including maintenance of protein and nucleic acid synthesis, stabilization of chromatin structure, and protection from oxidative damage [130]. Since Met is necessary to synthesize Cys, Met restriction can lead to Cys restriction when the dietary intake of Cys is low. In this case, Met restriction can reduce Cys and GSH (Glu-Cys-Gly) levels, which may lead to the accumulation of cytotoxic concentrations of ROS in cancer cells. Accordingly, evidence suggests that Cys supplementation can reduce the anticancer activity of Met restriction [131,132], and many studies restrict or eliminate Cys in the diet to increase the anticancer activity of Met restriction [26,27,34,40,111,113,120,121,122,133]. It has been proposed that cancer cells are more vulnerable than normal cells to Met restriction because cancer cells may be unable to recycle Met from homocysteine (HCys) or SAM through the Met salvage pathways [24]. Normal cells can proliferate under Met restricted conditions if supplied with HCys, while cancer cells cannot obtain sufficient Met from HCys [134,135,136]. This vulnerability may be explained by defects in enzymes involved in Met salvage pathways [137,138]. Furthermore, some cancer cells use HCys to synthesize Cys, therefore limiting their ability to recycle Met [139].
The role of Met in antitumor immunity is complex. Met supplementation can improve immune function because this AA is highly consumed by immune T-cells. Accordingly, intratumorally/intraperitoneally Met administration improved antitumor immunity and increased the anticancer activity of checkpoint inhibitors in several syngeneic cancer models [140]. On the other hand, some studies showed that dietary Met limitation can improve antitumor immunity and increase the activity of anti-PD-1 immunotherapies [34,118].
Dietary Met restriction has been evaluated in cancer patients in phase I clinical trials. In 1995, 14 patients with preoperative gastric cancer were randomly divided into two groups: 5-fluorouracil plus total parenteral nutrition lacking Met, or 5-fluorouracil plus total parenteral nutrition with Met [141]. The combination of 5-fluoruracil plus Met-restriction showed a lower tumor burden and thymidylate synthase activity compared to 5-fluoruracil alone [141]. In 2002, 8 patients with solid cancers received a Met-restricted diet for approximately 17 weeks (range 8-39 weeks) [142]. This Met-restricted diet was safe and tolerable for patients [142]. In the late 2000s, two small clinical trials were conducted in a total of 29 patients with melanoma and 3 patients with glioma [143,144], who received an intermittent Met-free diet plus cystemustine [144] or nitrosourea [143]. Treatment was well tolerated, but little benefit was observed in patient survival. In 2010, a phase I trial was conducted in 11 colon cancer patients who received 3 cycles of Met-free diet for 3 consecutive days plus FOLFOX6 chemotherapy [145]. Plasma Met levels were reduced by 58% on the first day and treatment was well tolerated, but little benefit was observed [145]. Recently, after observing that Met restriction produced therapeutic responses in patient-derived xenograft models of chemotherapy-resistant RAS-driven colorectal cancer in mice, the authors conducted a feeding study in humans that revealed that Met restriction induced changes in systemic metabolism that were similar to those obtained in mice [114].
Pharmacological approaches based on enzymatic depletion of Met support the idea that Met restriction has potential for cancer therapy. Methioninase (METase) was the first enzyme developed to deplete Met. The injection of this recombinant bacterial enzyme can deplete Met from plasma and induce anticancer activity [146]. In mice, each administration completely depletes Met for 8 hours, showing anticancer activity against xenograft and syngeneic cancer models [147,148,149,150,151,152]. METase can also deplete Met for 8 hours in primates [153]. However, since METase had a short half-life (immune clearance) and its continuous administration could trigger anaphylactic reactions [153], the enzyme was pegylated to minimize its immunogenicity. PEGylated-METase extended its half-life and prevented anaphylactic reactions in primates [154]. Surprisingly, oral administration of METase, which removes Met from the gastrointestinal tract, was as effective as intraperitoneal administration in lowering serum Met levels and inducing anticancer activity in several murine cancer models [155,156,157,158,159,160,161,162]. Other human recombinant enzymes have recently been developed [163,164]. These human recombinant enzymes eluded the immunogenic problems of the original METase and demonstrated in vivo anticancer activity against neuroblastoma [163] and prostate cancer [164]. Met enzymatic depletion has been combined with several anticancer drugs. Enzymatic Met depletion showed synergistic effects with 5-fluorouracil [149], cisplatin [148], temozolomide [150,151], nitrosourea [150] and doxorubicin [152]. Oral METase also showed a synergistic effect in combination with various anticancer drugs [155,156,157,158,159,160,161,162]. The pharmacokinetics and safety of METase has also been evaluated in patients. Two small phase I clinical trials conducted during the late 1990s showed that METase reduced the plasma levels of Met without causing evident toxicity [165,166]. Recently, two case reports have shown that oral administration of METase plus a low Met diet achieved stable long-term disease in a patient with locally recurrent rectal cancer [167] and in a patient with stage IV pancreatic cancer treated with FOLFIRINOX [168].
4. Non-Essential Amino Acids
4.1. Cysteine
Cys is a sulfur-containing NEAA with multiple cellular roles [169]. Humans can biosynthesize Cys from the EAA Met through the transsulfuration pathway [24,169,170] (Figure 2). In addition to being necessary for protein synthesis, Cys is essential for the production of a variety of sulfur-containing molecules with important biological roles [170]. These include iron-sulfur clusters found in enzymes of the electron transport chain (ETC) [171], coenzyme A and thioredoxins [172]. Cys also produces taurine [37], hydrogen sulfide (H2S) [37], and is the rate-limiting AA for the production of the tripeptide GSH (Glu-Cys-Gly) [169,170]. GSH is essential for protecting cells against the toxic effects ROS [169].
The importance of Cys in tumor growth was first reported in 1936 [133]. In this study, Voegtlin et al. observed that a diet deficient in Cys/Met reduced tumor growth in mice with spontaneous breast cancer, and the addition of Cys abruptly stimulated tumor growth [133]. Since Met can produce Cys, Met is usually restricted in may studies evaluating the anticancer activity of Cys depletion/restriction [26,27,34,40,111,113,120,121,122,133]. Intravenous parenteral nutrition with double Cys/Met restriction showed anticancer activity in rats with sarcoma [121,122], and inhibited cancer proliferation in mice xenografted with human glioma cells [111]. Dietary restriction of Cys and Met also showed anticancer activity in a spontaneous mouse model of breast cancer [40] and in a transgenic prostate cancer model [34]. We recently showed that an artificial diet restricted in Cys and Met (formulated with 6% casein, 5% Gln, and 2.5% Leu) induced a marked anticancer activity in two metastatic colon cancer models; Cys supplementation blocked its anticancer activity [26]. Although limiting Met levels can increase the effect of Cys restriction, anticancer activity has been observed with Cys-restricted diets with normal levels of Met, for example in animal models of colon cancer [174,175] and glioma [176]. We recently found that a diet lacking 6 NEAAs (including Cys), with normal Met levels (0.6%), showed a marked anticancer activity in mice with disseminated renal cell carcinoma; the anticancer activity of this diet was reduced by Cys supplementation [27]. However, supplementing Cys under certain conditions may be important for the anticancer activity of an anticancer diet; we recently observed that supplementing 0.2% Cys to an inactive casein-based diet (restricted in Cys) markedly improved the survival of mice with disseminated renal cell carcinoma [27]. Cys restriction may therefore have a positive or negative effect on the anticancer activity of a diet depending on the levels of other dietary components [27]. Table 4 shows representative studies assessing the in vivo anticancer activity of Cys restriction.
Mechanistically, Cys restriction may induce anticancer activity by reducing the capacity of cancer cells to eliminate ROS. Cancer cells produce high levels of ROS, which may accumulate and produce cell death [8]. Cancer cells rely on GSH to reduce ROS levels [24]. Dietary Cys restriction can decrease Cys plasma levels [176], reduce GSH biosynthesis [175,176] and increase ROS levels in cancer cells [174,175,176]. Cys restriction may also lead to the accumulation of cytotoxic levels of ROS in cancer cells by interfering with the activity of the polyamine pathway, which cancer cells use for ROS protection [174].
Since Cys is necessary for immune cells, Cys restriction may reduce the ability of the immune system to eliminate cancer cells. Cys is essential for T-cell activation and function [177]. High CysS plasma levels have been associated with a higher probability of response to immune checkpoint inhibitors in patients with lung cancer [178,179]. However, the negative effect of Cys restriction on the immune antitumor response is controversial, because other studies suggest that Cys restriction can increase the antitumor immune response [34,180].
Pharmacological approaches based on enzymatic depletion of Cys and inhibition of Cys transporters support the idea that Cys restriction has potential for cancer therapy. These pharmacological interventions have been useful to understand possible mechanisms by which Cys restriction induces in vivo anticancer effects. In 2017, an optimized human cyst(e)inase enzyme was able to reduce Cys and CysS plasma levels in mice and primates without causing toxicity [181]. Cyst(e)inase showed anticancer activity in mouse models of a variety of cancers including prostate, breast, chronic lymphocytic leukemia, pancreas, lung, renal, melanoma and ovarian cancer [180,181,182,183,184,185,186]. Cyst(e)inase administration increased ROS levels, depleted intracellular levels of GSH, and triggered ferroptosis in cancer cells [181,182,183,184,185,186]. Ferroptosis is a form of iron-dependent cell death triggered by lipid peroxidation. GPX4, which prevents lipid peroxidation, needs GSH as a cofactor for its activity [170,187,188]. Cys depletion can lead to lipid peroxidation and trigger ferroptotic cancer cell death.
Pharmacological inhibition of the xCT antiporter (SLC7A11), which imports exogenous CysS and exports glutamate (Glu), suggest that cancer cells depend on external Cys supply for their survival. Due to the oxidizing conditions of the extracellular environment, most of the extracellular Cys is in the form of CysS (the oxidized dimer of Cys) [189]. Inhibition of xCT antiporter decreased intracellular Cys levels, caused loss of antioxidant protection and induced ferroptosis [189]. Several drugs are known to inhibit xCT antiporter activity, including sulfasalazine, sorafenib, erastin, imidazole ketone erastin (IKE) and HG106 [189]. These drugs, some of them approved for clinical use, induce anticancer activity in a variety of cancer models [190,191,192,193,194,195,196,197,198,199,200]. Sulfasalazine, which is used for the treatment of rheumatic arthritis, has been evaluated in several clinical studies. Sulfasalazine, at a maximum dose of 6 g/day, did not induce clinical response in 10 patients with gliomas [201]. In a dose-escalation study, sulfasalazine reduced intratumoral GSH levels and CD44 positive cancer stem cells in patients with gastric cancer [202]. In a clinical trial conducted in 8 patients with CD44 positive gastric cancer who received sulfasalazine plus cisplatin, only 1 patient achieved stable disease for 4 months [203]. A phase I/II clinical trial in patients with glioblastoma revealed that the addition of sulfasalazine to a temozolomide plus radiotherapy intervention did not increase overall survival and progression-free survival [204].
4.2. Serine
Serine (Ser) is synthesized from 3-phosphoglycerate (glucose metabolite) and Glu (nitrogen donor) through de novo Ser synthesis pathway [205]. In addition to being a proteinogenic AA, Ser plays an important role in one-carbon metabolism [205,206,207]. Ser is the main source of carbon units in the folate cycle, which is mainly used for the synthesis of purines and pyrimidines, and for the conversion of HCys into Met. Ser is also used to produce Gly and provides the carbon skeleton for the synthesis of Cys through the transsulfuration pathway. It also has other important functions, such as production of certain lipids, including ceramide and phosphatidylserine [205,206,207].
Ser and Gly are easily interconverted by the SHMT1/2 enzymes [207]. Therefore, Gly is usually restricted in most dietary studies evaluating the anticancer activity of Ser limitation. Dietary Ser/Gly can reduce Ser and Gly levels in plasma [208] and tumors [209]. Although both AAs can be synthesized by human cells, cancer cells may depend on external supply of these AAs to keep their high proliferative demands. Cancer cells may also have mutations (e.g., in p53) that increase their dependency on these AAs. Dietary double restriction of Ser/Gly have shown anticancer activity against a variety of cancers in mice [208,209,210,211,212,213,214,215,216,217,218,219,220,221]. These studies are summarized in Table 5.
Mechanistically, dietary Ser/Gly restriction can induce anticancer activity by restricting two important building blocks for biosynthesis. The new cancer cells created during tumor growth need new proteins, nucleic acids and specific lipids; these processes require the synthesis or acquisition of sufficient levels of these two AAs. For example, dietary Ser/Gly restriction induced anticancer activity in colon cancer xenograft model by altering the biosynthesis of sphingolipids [212]. Dietary Ser/Gly restriction may also induce anticancer activity by increasing the cellular levels of ROS in cancer cells. Ser is needed to synthesize Gly and Cys, which are necessary to produce the antioxidant tripeptide GSH (Glu-Cys-Gly). Ser restriction caused oxidative stress in p53-deficient cancer cells, and dietary Ser/Gly restriction induced anticancer activity in mice [214]. The combination of Ser-Gly restricted diets with other pro-oxidant treatments showed a synergistic anticancer response in murine cancer models [213,218].
Some mutations found in cancer cells can increase their susceptibility to dietary Ser/Gly restriction, including mutations involved in the synthesis of Ser/Gly, or some mutations in p53. However, overexpression of enzymes involved in in the biosynthesis of these AAs may compromise the anticancer activity of dietary Ser/Gly restriction. For example, many cancer cells overexpress PHGDH, the first enzyme involved in the synthesis of Ser (Figure 2) [206]. Certain p53 mutations and the activation of KRAS, MYCN, NRF2 and MDM2 can induce the overexpression of enzymes involved in Ser biosynthesis, conferring resistance to dietary Ser/Gly restriction [24,208,211,215,222]. As expected, reduced expression of PHGDH and PSAT1 sensitized cancer cells to Ser-Gly restricted diets [211,221]. Low expression of PHGDH has been observed in platinum-resistant ovarian cancer cells, making this subtype of ovarian cancer vulnerable to Ser/Gly dietary restriction [210]. Combination of dietary Ser/Gly restriction with an inhibitor of PHGDH (PH755) showed anticancer activity in colon cancer [217].
The importance of Ser and Gly for cancer cell proliferation and survival is supported by studies showing that pharmacological inhibition of enzymes involved in the Ser/Gly biosynthesis pathway induce anticancer effects. The PHGDH inhibitors NTC-503 [219,223] and CBR-5884 [224] induced antiproliferative effects, but lacked selectivity for PHGDH. The PHGDH inhibitor PH755 showed higher selectivity and induced anticancer activity in vitro and in vivo [212,217,225]. Small-molecule dual SHMT1/2 inhibitors also showed anticancer activity against B-cell and T-cell lymphoblastic leukemia [226,227].
In immunogenic tumors, however, dietary Ser/Gly restriction may reduce the ability of immune cells to eliminate cancer cells. For example, Ser restriction can impair expansion of T cells in vivo, probably because Ser supplies Gly and one-carbon units for de novo nucleotide biosynthesis in proliferating T cells [228]. We recently observed in mice with disseminated renal cell carcinoma that a diet with both Ser and Gly was better than the same diet without Ser and Gly; untreated mice lived 30.3 ± 1.3 days, mice fed the diet without Ser/Gly lived 40.3 ± 2.0 days and mice fed the diet with Ser/Gly lived 54.7 ± 7.8 days [27].
4.3. Glycine
Gly is a NEAA that can be synthetized from Ser. Gly is essential for protein synthesis. Collagen, which is the most abundant protein in the human body (30-40% of total body protein), contains approximately a 33% of Gly [229]. This AA also acts as an inhibitory neurotransmitter [37]. Gly can also be used for the synthesis of the antioxidant tripeptide GSH, Ser, purines, creatine, and heme group [37]. Evidence suggest that rapidly growing cancer cells have a high Gly dependency [230].
As discussed in the previous section, Ser and Gly are easily interconverted by the enzymes SHMT1/2 [207]. Therefore, in most studies evaluating the anticancer activity of dietary Gly restriction, Ser was also restricted. Gly/Ser restriction has shown anticancer activity in different types of cancer [208,209,210,211,212,213,214,215,216,217,218,219,220,221,222]. Table 5 shows representative studies assessing the in vivo anticancer activity of Ser/Gly restriction. We recently observed that an artificial diet lacking several NEAAs (Ser, Cys, Tyr, Pro, Asn and Glu) markedly improved the survival of mice with disseminated renal cell carcinoma; Gly elimination, however, reduced the activity of this diet. These studies suggest that restricting Gly can have a positive or negative effect on the anticancer activity of a diet depending on the levels of other dietary components [27].
4.4. Arginine
Arginine (Arg) is a NEAA used for protein synthesis. It also participates in many other biological processes, including synthesis of nitric oxide, creatinine, ornithine, agmatine and polyamines [37,231]. It also plays a key role in the urea cycle [37]. Normal cells can synthesize Arg from citrulline and aspartate (Asp) through ASS1 (argininosuccinate synthase 1) and ASL (argininosuccinate lyase) in the urea cycle.
The anticancer potential of Arg restriction was evaluated in 1959 by Sugimura et al. [39]. In this study, 5 days of an Arg-free diet reduced the growth of Walker tumors in rats, and no weight loss was observed after 11-days on the Arg-free diet [39]. In the early 1990s, an Arg-free diet showed anticancer effects in mouse models of skin carcinogenesis [232] and colon cancer [233]; the Arg-free diet inhibited tumor growth and Arg supplementation stimulated tumor growth [233]. More recently, dietary Arg restriction reduced cancer growth in a xenograft model of ASS1-deficient breast cancer [234]. Arg restricted diets also suppressed cancer growth in colon cancer [235], prostate cancer [236] and liver cancer [237] xenografts. On the other hand, since adequate levels of Arg are important for T-cell proliferation, dietary supplementation of Arg has been found to improve T-cell function and induce anticancer immunity in murine cancer models alone and in combination with other anticancer drugs [238,239,240,241,242]. Table 6 summarizes studies assessing the in vivo anticancer activity of dietary Arg restriction and supplementation.
Arg-free diets can decrease plasma levels of Arg in healthy volunteers. An Arg-free diet taken for 6 days reduced Arg plasma levels by approximately 20-40% [243]. In another study, 4 weeks of dietary restriction of Arg and other precursors of Arg (Asp, Pro, and Glu) significantly decreased Arg plasma levels without causing side effects [244].
Mechanistically, dietary Arg deprivation may induce selective anticancer activity because many cancer cells express low levels of ASS1, which is involved in the synthesis of Arg. Downregulation of ASS1 facilitates cancer cell proliferation by increasing aspartate availability for pyrimidine biosynthesis [245]. In addition, downregulating the expression of ASS1 under acidic or hypoxic environments provides cancer cells with a survival advantage [246]. However, ASS1-deficient cancer cells rely on the external supply of Arg for their survival, which may explain why Arg deprivation induces anticancer activity [246].
The importance of Arg for cancer cell proliferation and survival is supported by numerous studies that have shown that pharmacological depletion of Arg levels with Arg-depleting enzymes induces anticancer activity. Two different enzymes are currently under clinical development: ADI-PEG20 (pegylated arginine deiminase) and PEG-BCT-100 (pegylated recombinant human arginase 1). These enzymes have shown anticancer activity in a wide variety of cancers, including melanoma, hepatocarcinoma, and glioblastoma [231,234,247,248,249]. Eight phase I-II clinical trials are completed or ongoing for PEG-BCT-100 [250]. In patients with hepatocellular carcinoma, this drug was well tolerated and showed anticancer activity alone [251,252,253] and in combination with chemotherapy (capecitabine plus oxaliplatin) [254]. PEG-BCT-100 also showed anticancer activity in melanoma and prostate cancer patients [255], and induced a complete remission in an immunotherapy-resistant melanoma patient with absent expression of enzymes involved in the synthesis of Arg [256]. ADI-PEG20 has received more clinical attention, with 30 completed or ongoing clinical trials, three of them in phase III [257]. ADI-PEG20 monotherapy was well tolerated and safe in most clinical trials [258,259,260,261,262,263,264,265,266]. Combinations of ADI-PEG20 with other anticancer drugs are also being studied in phase I-II clinical trials [267,268,269,270,271,272,273]. In a phase III clinical trials with more than 600 patients with hepatocellular carcinoma, ADI-PEG20 monotherapy did not show significant improvements in overall survival and progression-free survival [265]. More research is needed to elucidate possible mechanisms of resistance, as well as the potential benefit of the combination of this enzyme with other anticancer therapies.
As occurs with other AAs, Arg restriction may have a negative impact on immunogenic cancers. Some cancer cells create an immunosuppressive microenvironment by converting myeloid cells into M2 macrophages or myeloid-derived suppressive cells [274]. These immunosuppressive cells express arginase, which hydrolyzes Arg to ornithine and urea, therefore reducing Arg levels in the tumor microenvironment [274]. Arg is essential for T-cell proliferation, and the expression of arginase can disrupt antitumor immunity [241,274,275]. Accordingly, arginase inhibitors (which increase Arg levels) have shown anticancer activity [276,277,278,279] and have reached phase I-II clinical trials [280,281]. Dietary supplementation of Arg can improve the immune function and induce anticancer activity in murine cancer models alone and in combination with other anticancer drugs [238,239,240,241,242]. However, in a recent double-blind clinical trial with 65 colorectal cancer patients, 10 g/day of Arg supplementation did not prevent immunosuppression compared to placebo [282]. In summary, since Arg in important for both cancer cells and immune cells, patients with Arg-auxotrophic tumors may benefit from Arg restriction therapies, while patients with immunogenic cancers may benefit from Arg supplementation.
4.5. Glutamine
Gln is a non-essential proteinogenic AA that can be considered essential under certain conditions [283]. It is the most abundant AA in human plasma and tissues and is involved in many biological processes [284]. It participates in the transport and detoxification of ammonia in the urea cycle, helping to maintain the pH balance [285,286]. Gln is the main source of nitrogen atoms for the biosynthesis of nucleotides (pyrimidines and purines) and NEAAs (Glu, Asn, Ala, Asp, Ser, Pro and citrulline) [285]. Gln also mediates the cellular uptake of certain EAAs; for example, LAT1 imports the EAA Leu while simultaneously exports Gln [287]. Importantly, Gln catabolism is used as a key energy source in highly proliferating cells, such as intestinal cells, immune cells and cancer cells [286]. Gln is deaminated in two steps by GLS and glutamate dehydrogenase (GDH), yielding Glu and α-ketoglutarate (αKG); the latter can enter the tricarboxylic acid (TCA) cycle and eventually produces ATP [286]. Gln indirectly participates in maintaining cellular redox balance. Gln catabolism generates Glu, which is used for the synthesis of GSH and for the uptake of CysS through the antiporter xCT [288]. Gln is a versatile biosynthetic substrate to supply carbon and nitrogen atoms for the generation of key precursors for biosynthesis and cell proliferation [289].
Proliferating cancer cells have a high Gln demand. Cancer cells obtain high Gln levels by increasing their biosynthesis or by obtaining it from the extracellular environment [285]. The increased Gln uptake of cancer cells has been associated with lower plasma levels of Gln in patients with several types of cancer [290,291]. The increased Gln uptake by tumors is actually being studied for diagnostic purposes with PET imaging using 18F-(2S,4R)-4-fluoroglutamine [292,293,294,295]. The increased Gln uptake of cancer cells is related with their high expression of ASCT2 (SLC1A5) [296,297,298,299,300]; this chief Gln transporter is upregulated by the oncogenes MYC and KRAS [301,302]. Once Gln is inside the cell, Gln catabolism serves to supply the TCA cycle, to support lipogenesis, to biosynthesize NEAAs and nucleotides, and to help to maintain high GSH levels [285,303]. The GLS1 isoform, which catalyzes Gln deamination to generate Glu, is upregulated by the oncogene MYC [301]. GLS1 is upregulated in many cancer types and its overexpression is associated with a poor prognosis [304,305,306,307,308,309]. The conversion of Glu into αKG is also enhanced in many cancer cells, probably because MYC, KRAS and PI3KCA expression upregulate the enzymes GLUD1, GOT1, GOT2 and GPT2 [310,311,312]. Gln may become an essential AA for cancer cells driven by oncogenic MYC, KRAS and PIK3CA [310].
Limiting Gln levels and targeting Gln acquisition and utilization have been studied as possible anticancer strategies. Few studies have evaluated the in vivo anticancer activity of diets deficient in Gln. In 2017, dietary restriction of Gln was found to induce anticancer activity in vitro and in vivo in a p73-expressing medulloblastoma xenograft model [313]. The Gln restricted diet increased mice survival and also showed a synergistic effect with cisplatin. Although the only difference between the control and experimental diets was the presence/restriction of Gln, both diets also lacked Glu, Ala, Asn, Asp, and Pro. This diet reduced Gln and Glu levels in the cerebellum and cerebrospinal fluid of mice [313]. In another study, a Gln and Glu free diet significantly decreased Gln plasma levels and impaired disease progression in mice with Notch1-expressing leukemia [314].
Most anticancer strategies targeting the altered Gln metabolism of cancer cells have focused on pharmacological inhibition of Gln acquisition and utilization [285]. These include inhibition of GLS1, with inhibitors such as CB-839 (telaglenastat) [315], BPTES [316] and C.968 [317,318]. CB-839, which is orally bioavailable, has been tested in clinical trials. There are at least 21 completed or ongoing phase I-II clinical trials, of which 8 have been completed [319]. In general, CB-839 was safe and well tolerated by cancer patients [320,321,322,323,324,325,326]. In most of the completed clinical trials, CB-839 was combined with other anticancer drugs [285]. The benefit on cancer progression has been modest so far [285]. There are other experimental anticancer drugs targeting Gln metabolism. Inhibition of Gln uptake by V-9302, an inhibitor of the ASCT2 transporter, induced anticancer activity in murine cancer models [327]. JHU083, which is a prodrug of the Gln antagonist DON [328], is selectively activated in the tumor microenvironment and disrupts cancer cell metabolism while improving T-cell anticancer responses. This compound induced marked anticancer activity alone and in combination with immunotherapies in several murine cancer models [328,329,330,331,332,333]. The off-target effects of some anticancer drugs can also impact Gln metabolism. For example, the Asn-depleting enzyme L-asparaginase (ASNase), which is used against leukemia that rely on external supply of Asn, can also deplete Gln; this effect may explain its anticancer activity in a murine leukemia model resistant to Asn depletion [334,335].
Gln restriction may cause toxicity, because this AA is necessary for non-malignant proliferating cells [336,337]. Dietary restriction of Gln induced small intestine mucosal atrophy and muscle weight loss in rats [338]. Oral Gln supplementation has been shown to ameliorate mucosal damage (mucositis, stomatitis, pharyngitis, esophagitis, and enteritis) induced by chemotherapy and radiotherapy in randomized clinical trials with cancer patients [339,340,341]. Gln plays a key role in the cellular uptake of Leu [287], and its supplementation could enhance the beneficial effects of Leu on cancer cachexia [55].
A recent report have shown that Gln supplementation induced in vivo anticancer activity in a transgenic melanoma model and sensitized tumors to a BRAF inhibitor via epigenetic reprogramming [342]. The authors observed that a diet containing very high levels of Gln (20%) increased the concentration of Gln and αKG in tumors, without increasing other biosynthetic intermediates necessary for cell proliferation. The increase in αKG concentration led to hypomethylation of H3K4me3, thereby suppressing epigenetically-activated oncogenic pathways in melanoma [342]. Our recent investigation has revealed that supplementing specific artificial diets with Gln can increase their anticancer activity in mice with metastatic cancers; most of our active diets contained 5-6% Gln in their composition [26,27,343]. Therefore, although Gln plays a key role in cancer cell metabolism, anticancer activity can be achieved by both restricting and supplementing this AA.
4.6. Glutamate
Glu is a NEAA closely related to Gln. This AA is used in protein synthesis and has many other cellular functions. Glu is as a nitrogen donor for transaminases [37]. It is used in the synthesis of many NEAAs, including Ala, Asp, Ser, Pro and Gln (Figure 2) [54]. Transaminases and glutamic dehydrogenase (GDH) can convert Glu into αKG, which can be used to fuel the TCA cycle for energy production. In the brain, Glu is an excitatory neurotransmitter and can also be used for the synthesis of the inhibitory neurotransmitter γ-aminobutyric acid (GABA) [37]. Glu participates in ROS protection by allowing CysS uptake by the xCT antiporter, and by directly participating in the synthesis of the tripeptide GSH [288].
Although Glu supports cancer cell proliferation and survival, the anticancer activity of dietary Glu restriction has not been extensively studied, probably because Glu can be easily obtained from Gln, Asp and Ala, and is also produced in the degradation pathways of many AAs, including Leu, Ile, Val, Lys, Phe, His, Tyr and Pro. We recently observed that a diet lacking 6 NEAAs (including Glu) induced marked anticancer activity in mice with disseminated renal cell carcinoma; supplementing Glu to this diet markedly reduced its anticancer activity [27]. Glu is particularly abundant in the brain. Glioblastomas and brain tumors use this AA for energy production [344] and nucleotide biosynthesis [345]. As mentioned previously, dietary Gln restriction showed anticancer activity in a medulloblastoma xenograft model, and a significant drop in the levels of both Gln and Glu was observed in the cerebellum and cerebrospinal fluid of mice treated with this diet [313]. The activity of this diet may therefore be mediated, at least in part, by a reduction in the levels of Glu. Similarly, Gln and Glu restricted diet achieved anticancer response in mice with Notch1-expressing leukemia [314]. Accordingly, as mentioned previously, the enzyme GLS produces Glu from Gln, and several GLS inhibitors have shown in vivo anticancer activity, including CB-839 (telaglenastat) [315], BPTES [316] and C.968 [317,318]. CB-839 has been evaluated in clinical trials [319,320,321,322,323,324,325,326]; however, its clinical benefit has been moderate [285].
4.7. Asparagine
Asn is a NEAA that can be synthesized from Asp by the enzyme ASNase. Asn is needed for protein synthesis, but the importance of Asn in other cellular processes is less understood [20]. Asn can modulate mTORC1 activity, can serve as an exchange molecule for the uptake of other AAs (e.g., Ser, Arg, and His), and maintenance of intracellular Asn levels seems to be critical for cancer cell growth [346].
Asn is commonly used to exemplify the relevance of NEAA restriction in cancer therapy, because the Asn-depleting enzyme ASNase is a useful drug for patients with acute lymphoblastic leukemia (ALL) and acute lymphoblastic lymphoma (ALLy). ASNase is an enzyme from E. coli that deaminates Asn to Asp and ammonium; its intravenous administration quickly depletes Asn from serum and cells [347]. ALL cells usually rely on external Asn for their survival, and the depletion of Asn with ASNase leads to apoptosis in leukemia cells [348]. ASNase is pegylated (PEG-ASNase) to extend the half-life and reduce the immunogenicity of the enzyme. Nowadays, ASNase is included in most chemotherapy regimens for pediatric ALL and ALLy, achieving high survival rates [23]. The efficacy of ASNase is generally correlated with the expression of ASNS in leukemia cells [349,350]; this enzyme allows the synthesis of Asn from Asp. However, in some cases, ASNS expression after ASNase was not associated with resistance to treatment [351]. ASNase also has Gln-depleting activity, which may participate in the anticancer activity of this enzyme [334,335]. ASNase treatment can produce adverse effects attributed to Gln-depleting activity, to ammonia production, and to the development of immunogenicity against the enzyme [352]. Antibody neutralization and subsequent inactivation of ASNase is a key cause of treatment failure [23]. As an alternative for patients with hypersensitivity reactions and silent neutralization to E. coli ASNase, Erwinia ASNase can be used as a second-line treatment [353]. Although ASNase was considered inactive against most solid tumors [354], growing evidence suggests that ASNase has anticancer activity in several types of solid tumors in preclinical animal models [216,355,356,357,358,359,360].
Dietary Asn restriction induces in vivo anticancer activity [216,357,360], and may serve as an alternative to treatment with ASNase. Limiting Asn availability by ASNS knockdown, treatment with ASNase, or dietary Asn restriction reduced the number of lung metastases in an orthotopic triple negative breast cancer model, whereas increased dietary Asn or enforced ASNS expression promoted metastatic progression [357]. Serum Asn levels were proportional to the Asn content in each diet (0% Asn in the restricted diet, 0.6% in the standard diet and 4% in the supplemented diet) [357]. Another study revealed that dietary Asn restriction reduced tumor growth in KEAP1 mutant cancer cells in vivo [216]. Cancer cells with KEAP1/NRF1-mutations display a high endogenous oxidative stress response dependent on the external supply of several AAs including Asn [216]. ASNase showed more anticancer activity than dietary Asn restriction, and the combination of both enzymatic and dietary restriction showed the same activity as ASNase alone [216]. Recently, the combination of dietary Asn restriction with the electron transport chain (ETC) inhibitors metformin or IACS-010759 induced anticancer activity in xenograft and transgenic lung cancer models [360]. Similar results were obtained with ASNase in combination with metformin [360]. Supplementing 0.6% or 4% Asn restored tumor growth [360]. Mechanistically, inhibition of ETC with metformin limits Asp availability for Asn synthesis during Asn-restricted conditions [360].
4.8. Aspartate
In addition to its role in protein synthesis, Asp participates in the synthesis of purines, pyrimidines, Asn and Arg [37]. It also plays a role in the urea cycle, the malate-Asp shuttle, and in transamination reactions [37]. Due to its role in the synthesis of nucleotides, Asp is crucial for proliferating cancer cells.
Although Asp can become a limiting factor for tumor growth, the antitumor activity of dietary Asp deprivation has not been evaluated individually, probably because this AA can be easily obtained from Glu and OAA through GOT1/2 (AST) transaminases (Figure 2). Since these enzymes are expressed in many tissues, including the liver, dietary Asp restriction would not result in systemic Asp restriction.
Recent evidence indicates that Asp is an endogenous metabolic limitation for tumor growth. Asp has poor cell permeability, which prevents its environmental acquisition by tumor cells [361]. Cancer cells synthetize Asp from the OAA originated in the TCA cycle; this process requires ETC activity to consume NADH and allow OAA synthesis from malate [362,363]. Therefore, cancer cells rely on the TCA cycle and ETC to obtain Asp for proliferation and for other processes such as Asn biosynthesis. As mentioned before, this has been exploited to increase the anticancer activity of dietary Asn restriction. The combination of an Asn-restricted diet with an ETC inhibitor suppressed tumor progression in transgenic and xenograft lung cancer models [360]. Inhibition of ETC (metformin or IACS-010759) reduced the intracellular Asp available for Asn synthesis under Asn restricted conditions [360]. Some cancer cells overcome the dependence on Asp production from the TCA cycle by expressing the Asp/Glu carrier 1 (AGC1 or SLC1A3), allowing Asp uptake into cells [364]. The expression of SLC1A3 in cancer cells provided resistance to ETC inhibition in xenograft lung cancer models [364]. Gln catabolism can feed the TCA cycle and therefore Asp synthesis; the deletion of SLC1A3 synergized with CB-839 (GLS inhibitor) in a syngeneic model of lung cancer [365].
4.9. Tyrosine
Tyr is an aromatic NEAA that can be obtained from the EAA Phe. In addition to its role in protein synthesis, Tyr is necessary to produce catecholamines (dopamine, epinephrine, and norepinephrine) and melanin [37].
Since Phe is a precursor of Tyr, both AAs are usually restricted simultaneously in most cancer studies. As shown in the Phenylalanine section (section 3.6.), dual restriction of Phe and Tyr has been evaluated in animal studies and cancer patients with several positive results. We recently found that a diet lacking 6 NEAAs (Tyr, Cys, Ser, Pro, Asn, and Glu) markedly improved the survival of mice with disseminated renal cell carcinoma. Adding Tyr to this AA-manipulated diet did not decrease its anticancer activity, suggesting that restricting Tyr was not essential to achieve anticancer activity in this cancer model [27].
4.10. Alanine
Ala is a proteinogenic NEAA with other important metabolic functions. It is involved in transamination reactions and in the glucose-alanine cycle (Cahill cycle). Ala can be easily converted into pyruvate by GPT1/2 transaminases [37]; pyruvate is a carbon source for energy production, fatty acid biosynthesis and gluconeogenesis [37,366].
The antitumor activity of dietary Ala deprivation has not been evaluated independently of other AAs, probably because Ala can be easily obtained from Glu and pyruvate through GPT1/2 transaminases (Figure 2). Since these enzymes are expressed in many tissues, including the liver, dietary Ala restriction would not result in systemic Ala restriction.
Recent evidence suggest that Ala is a critical substrate for pancreatic cancer cells. Pancreatic cancer cells obtain Ala from stroma-associated pancreatic stellate cells by upregulating the SLC38A2 transporter. Pancreatic cancer cells then deaminate Ala to obtain pyruvate and support the TCA cycle and the biosynthesis of NEAAs and lipids. Pyruvate derived from Ala can actually outcompete glucose and Gln-derived carbon skeletons for these processes [367,368].
4.11. Proline
Pro is a proteinogenic NEAA that can be synthesized from Glu or ornithine [369] (Figure 2). Pro can be used for the synthesis of Arg, Glu and polyamines, and participates in wound healing and the immune response [37,370]. Like Gly, Pro is a major building block for the synthesis of collagen [37]. Collagen is the main Pro storage in the human body [369], and some cancer cells, such as pancreatic cancer cells, can use extracellular collagen to obtain Pro under conditions of nutrient deprivation [371].
Dietary Pro restriction inhibited tumor growth in mice xenografted with PC-9 lung cancer cells, but not in mice with PaTu-8902 pancreatic cancer cells [372]. Mechanistically, Pro starvation induced endoplasmic reticulum stress in cancer cells with hyperactivation of mTORC1-mediated 4EBP1 signaling [372]. A diet restricted in several NEAAs (including Pro) showed anticancer activity in mice inoculated with B16F10 melanoma cells; the diet was initiated one week before the inoculation of the cancer cells [373]. We recently observed that the addition of Pro to a diet lacking 6 NEAAs (Tyr, Cys, Ser, Pro, Asn, and Glu) did not block its anticancer activity in mice with disseminated renal cell carcinoma [27].
Pharmacological treatment of enzymes involved in Pro metabolism suggests that this AA plays an important role in the metabolism of cancer cells. Pro catabolism is used by cancer cells to generate Glu and αKG, which can fuel the TCA cycle [369]. Since PRODH is a key enzyme in Pro catabolism, several PRODH inhibitors have been developed to target cancer cells. N-propargyl-glycine is a suicide inhibitor of PRODH that showed anticancer activity in breast cancer xenografts [374]. Inhibition of PRODH with L-tetrahydro-2-furoic acid (L-THFA) decreased growth and metastases in breast and lung cancer models [375,376]. On the other hand, overexpression of enzymes involved in Pro biosynthesis, such as PYCR1 and OAT (Figure 2) can facilitate cancer progression, and their pharmacological inhibition can suppress cancer growth in vivo [377,378,379,380,381].
5. Manipulation of Multiple Amino Acids Simultaneously
Since the metabolic routes of many AAs are interconnected (Figure 2), the cellular requirements of specific AAs are probably influenced by the levels of other AAs. Manipulating several AAs simultaneously may therefore be more therapeutically useful than restricting AAs individually. As discussed in the previous sections, several pairs of AAs have usually been restricted together. Phe is the precursor of Tyr, and several studies showed in vivo anticancer activity when both AAs were restricted together [40,75,76,77,78]. Similarly, since Met can be used to synthesize Cys, dietary restriction of both AAs induced in vivo anticancer activity in different cancer types [26,27,34,40,111,113,120,121,122,133]. The NEAAs Ser and Gly can be easily interconverted by SHMT1/2 enzymes (Figure 2), and the simultaneous restriction of both AAs induced anticancer activity in several murine cancer models [208,209,210,211,212,213,214,215,216,217,218,219,220,221,222].
We have recently reported that the manipulation of multiple AAs simultaneously induced marked anticancer activities in mice with different types of metastatic cancers [26,27,119,343]. We screened 18 artificial diets for anticancer activity in mice with disseminated renal cell carcinoma and observed that mice fed a diet lacking six AAs (Ser, Cys, Tyr, Pro, Asn and Glu) lived longer than mice treated with sunitinib or anti-PD-1 immunotherapy (which are standard therapies for patients with metastatic renal cell carcinoma). Controlling the levels of several AAs (e.g., Cys, Met, and Leu) and lipids was important for the anticancer activity of the diets [27]. We have also tested several artificial diets in mice with metastatic colon cancer and compared their activity with that of capecitabine, which is a first-line treatment for patients with this disease. Mice fed a diet lacking 10 AAs (all NEEAAs except Gln), or a diet with 6% casein, 5% Gln, and 2.5% Leu, lived longer than untreated mice; several mice survived the treatment. The casein-based diet was better than several cycles of capecitabine in two animal model; the models were established by injecting CT26.WT murine colon cancer cells in the peritoneum (peritoneal dissemination) or in the tail vein (pulmonary metastases) of immunocompetent BALB/cAnNRj mice. We found that Cys supplementation blocked the activity of both diets, but Cys restriction was insufficient for activity [26]. We have also reported that the survival of mice with metastatic triple negative breast cancer (TNBC) could be markedly increased by replacing their normal diet with artificial diets in which the levels of AAs and lipids were strongly manipulated. AA manipulation led to modest improvements in mice survival when the levels or lipids were normal. Reducing lipid levels to 1% markedly improved the activity of several diets with different AA content. Mice fed the artificial diets as monotherapy lived longer than mice treated with first-line drugs doxorubicin and capecitabine. An artificial diet without 10 NEAAs, with reduced levels of EAAs, and with 1% lipids improved the survival not only of mice with TNBC but also of mice with other types of metastatic cancers [343]. Two additional diets, with altered levels of sulfur AAs also improved the survival of mice with metastatic colon cancer, ovarian cancer and renal cell carcinoma [119]. These data suggest that dietary manipulation of multiple AAs simultaneously has therapeutic potential for patients with metastatic cancers. Based on these results, we are currently testing the safety and efficacy of one of our artificial diets as monotherapy in patients with different types of metastatic cancers.
6. Discussion
Two decades ago, few cancer scientists considered metabolism as a relevant area of cancer research, probably because it was assumed that the accumulation of mutations in a cell was sufficient for malignant transformation, cell division and tumor growth. The explanation of the Warburg effect revealed that the genetic alterations of cancer cells are insufficient for cell proliferation and tumor growth. Cell proliferation requires that the dividing cell takes nutrients, such as glucose and certain AAs, from the extracellular environment. If you culture any type of cancer cell in PBS (which lacks glucose and AAs), the cell will not proliferate no matter what type of mutations it has. Cells must uptake glucose and AAs to produce building blocks to create new cells. Widespread clinical use of FDG-based PET imaging remind us that most cancers have increased glucose uptake when compared to normal tissues [2,3]. Unfortunately, glucose deprivation is not a feasible therapeutic strategy for cancer patients, because normal tissues also need glucose for their survival and proliferation. Severe hypoglycemia, which may occur in patients with type I diabetes receiving insulin, can actually be lethal if untreated. Depriving cells of certain AAs, however, can selectively kill cancer cells [[384] and can also be useful in patients with specific cancers [23]. Today, AA metabolism is considered a therapeutically relevant area of cancer research.
It is important to stress that selectivity is the key feature of an effective anticancer treatment. Cancer patients need therapies able to eliminate their malignant cells without significantly affecting their normal cells. The available anticancer drugs can kill cancer cells through a variety of mechanisms of action, but they also target normal cells at similar concentrations. The consequence of this limited selectivity is that patients cannot be treated with the drug doses needed to eliminate their malignant cells, because these doses would also kill their normal cells and would be fatal. Cancer patients receive tolerable doses instead of effective doses, which are generally insufficient to eradicate the cancer cells and cure the disease. This means that, when searching for clinically relevant therapies based on AA manipulation, the key is not to find the most toxic strategies for cancer cells. The key is to find strategies able to eliminate cancer cells without significantly affecting normal cells.
Restricting NEAAs may be more clinically relevant than restricting EAAs. Complete deprivation of EAAs would be toxic to both cancer cells and normal cells, because human cells cannot synthesize these AAs. However, normal cells can synthesize NEAAs, while cancer cells may be unable to obtain all of them because of their mutated genomes. This difference may confer selectivity. In vitro experiments have revealed that restricting NEAAs, individually and simultaneously, can induce selective anticancer effects [26,27,382] However, these experiments should be interpreted cautiously, because the metabolic environment of cells growing in vitro and in vivo is extremely different. For example, the low AA concentrations tested in most in vitro experiments are difficult to achieve in the systemic circulation, because the liver and muscles provide AAs to ensure sufficient plasma levels. Since in vitro data are difficult to extrapolate to an in vivo situation, this review article is focused on in vivo studies that have evaluated the antitumor activity of diets restricted or supplemented with the 20 proteinogenic AAs, individually and in combinations.
Restricting protein levels is the simplest strategy to reduce all AAs simultaneously. Several studies showed that low-protein diets reduced tumor growth in animal models [30,31,32,33,34,35]. Reducing the levels of plant proteins induced higher antitumor activities than reducing the levels of animal proteins, probably because each type of protein contains different ratios of AAs [34,35]. Reducing protein levels, however, does not exploit the full therapeutic potential of AA manipulation. Since each protein has a constant AA ratio, changing protein levels does not allow to alter the levels of specific AAs while keeping normal levels of others. Using mixtures of AAs, alone or in combinations with proteins, can drastically change AA ratios and may increase the potential of dietary AA manipulation for cancer therapy.
Diets lacking any EAA are lethal if taken continuously. However, these diets may induce anticancer activity if taken temporarily. Reducing EAAs to certain levels, rather than eliminating them, may also be therapeutically useful. Several decades ago, researchers started to evaluate in rodents the antitumor activity of diets lacking each EAA and diets with reduced levels of each EAA [39,40,72]. For most EAAs, the antitumor activity of the diets was linked to marked reductions in body weights; depriving EAAs was toxic for both cancer tissues and normal tissues. However, reducing some EAAs to certain levels (e.g., Ile, Met, Phe) inhibited tumor growth without significantly affecting body weights. For example, moderate restriction of Ile in the diet (0.15%) was sufficient to reduce tumor growth without significantly decreasing body weights [40]. More recent experiments revealed that limiting Met intake to 0.17-0.12% induced anticancer activity without causing significant weight loss in mice [112,113,114,115,116]. Many cancer cells have defects in the Met salvage pathways [134,135,136,137,138,139], which may limit their ability to recycle Met under low Met conditions [24]. In cancer patients, several case-reports and pilot clinical studies showed that dietary restriction of Phe/Tyr or Met only induced mild anticancer effects (see sections 3.6 and 3.9).
Diets lacking NEAAs are not lethal because our cells can synthesize these AAs. Some cancer cells, however, may have lost the ability to synthesize particular NEAAs due to mutations or low expression of enzymes involved in NEAA biosynthesis (Figure 2). For example, some cancer cells are known to depend on extracellular Asn or Arg for their survival. The restriction of Asn with the anticancer drug ASNase has been used in patients with leukemia for decades [23,349,350]. Dietary Asn restriction have also shown anticancer activity in preclinical models of solid tumors [216,357,360]. Similarly, some cancer cells lack the Arg-producing enzyme ASS1 and depend on exogenous Arg for their survival [231]. Dietary Arg restriction induces anticancer activity in preclinical models [232,233,234,235,236,237]. Enzymatic depletion of Arg has also shown preclinical anticancer activity and has reached clinical trials [250,258,259,260,261,262,263,264,265,266]. In a phase III clinical trial with more than 600 patients with hepatocellular carcinoma, the Arg-depleting enzyme ADI-PEG20 did not improve progression-free survival, although a mild overall survival benefit associated with prolonged Arg depletion was noted [265].
Limiting individual AAs in the diet has shown anticancer activity in animal models. However, the available clinical data show that this strategy may be insufficient to improve the survival rates achieved with the standard pharmacological treatments. To date, Asn is the only AA whose restriction is approved for patients with specific cancers. Since the cellular requirements of specific AAs are influenced by the levels of others, manipulating several AAs simultaneously may be more clinically useful than restricting AAs individually. In addition, it is important to realize that increasing, rather than reducing, the levels of particular AAs may improve the efficacy of diets based on AA manipulation. For example, if we restrict or eliminate some AAs in a diet, we may need to keep sufficient levels of Leu to avoid muscle and liver proteolysis; otherwise, the lysis of proteins in these organs would supply the AAs restricted in the diet. Supplementing a diet with particular AAs may also be important to create a permissive environment for antitumor immunity. Cells of the immune system must proliferate to mount an efficient immune response, and immune cells need to acquire sufficient levels of certain AAs (e.g., Trp, Met, Cys, Ser, Gly, Arg, Gln, and Asn) to proliferate. For example, Arg supplementation improved antitumor immunity alone and in combination with immunotherapies [238,239,240,241,242]. Limiting AAs may therefore be a double-edged sword in cancer therapy. Reducing AA levels may inhibit cancer cell proliferation, but may also inhibit the capacity of the immune system to eliminate cancer cells. Increasing AA levels may facilitate cancer cell eradication by the immune system, but may also promote cancer growth.
Several years ago, we proposed a new approach to manipulate AA levels for cancer therapy. Instead of reducing the levels of a particular AA, we proposed to use artificial diets to create massive changes in AA levels and ratios to generate toxic metabolic environments for cancer cells. The therapy would consist of replacing during several weeks the normal diet of cancer patients with an artificial diet prepared in the laboratory from scratch [54]. The rationale for this approach is as follows. Although cancer cells have mutations (and other DNA changes) that provide them with a survival advantage under a standard physiological environment, these same mutations may cause their death in a different environment. It is well known that evolution and survival depend not only on the acquisition of beneficial mutations, but also on favorable environments for these mutations. A mutation that facilitates survival in a specific environment can be lethal in a different environment. All cancer cells have originated and evolved under metabolic environments shaped by normal diets, which provide the 20 proteinogenic AAs at relatively constant levels and ratios. Creating massive changes in AA levels and ratios with artificial diets may therefore create new and unfavorable metabolic environments for cancer cells. Under these new environments, the DNA aberrations of cancer cells may cause their death [54]. Based on this idea, we have evaluated the therapeutic potential of numerous artificial diets in mice with different types of metastatic cancers [26,27,119,343]. As discussed in the previous section, some of the diets induced higher survival rates in mice with metastatic cancer than the achieved with the standard pharmacological treatments, and one of the diets is currently being tested (in monotherapy) in patients with different types of metastatic cancers.
Our in vivo studies in mice with disseminate cancers revealed important conclusions that may be useful to further develop this anticancer strategy. It is generally assumed that the presence or absence of a particular AA determines the anticancer activity. However, the screening of numerous artificial diets under similar experimental conditions revealed that the restriction of a particular AA could have a positive, negative or neutral effect on the activity of a diet depending on the levels of other dietary components [27]. For example, in mice with disseminated renal cell carcinoma, Cys supplementation reduced the high survival rates achieved with the most active diet formulated with free AAs (diet T2), therefore suggesting that Cys restriction was important for the activity. However, supplementing Cys to an inactive casein-based diet deficient in Cys markedly improved the survival rates of mice with the same cancer type under the same experimental conditions [27]. In mice with metastatic colon cancer, Cys supplementation markedly reduced the activity of two active diets [26]; however, in mice with peritoneally disseminated ovarian cancer, Cys supplementation did not have a significant impact on the anticancer activity of the diets [119]. We observed a similar complexity for Met. Several diets formulated with free AAs that contained relatively high amounts of Met induced a marked antitumor effect in mice with renal cell carcinoma; however, supplementing similar levels of Met to a different set of casein-based diets completely abolished their anticancer activity in the same cancer model [27]. In mice with metastatic colon cancer, Met supplementation strongly reduced the anticancer activity of two diets (diets B1 and B2B) [119]. In contrast, Met supplementation did not significantly reduce the activity of diet B2B in mice with ovarian cancer [119]. Altogether, our results strongly suggest that changing the levels of a particular AA can have a positive, negative or neutral effect on the anticancer activity of a diet depending not only on the type of cancer but also on levels of other AAs and nutrients. The whole composition is what determines the activity.
The mechanisms by which dietary manipulation of AAs induce selective anticancer activity are not completely understood. Unlike normal cells, cancer cells may have mutations or other DNA defects in the metabolic pathways involved in the synthesis of NEAAs, and may be unable to obtain sufficient levels if these AAs are removed from the diet [54]. In addition, since cancer cells produce higher levels of ROS than normal cells, they may have a higher dependency on Cys (and its precursor Met) to maintain sufficient GSH levels and prevent ROS-induced cell death. Another possible reason is that cancer cells proliferate faster than most normal cells and therefore need higher levels of AAs to produce building blocks for the dividing cells. Dietary AA restriction would restrict biomass production for cancer cell division and tumor growth, while non-dividing normal cells would be unaffected. In addition, normal cells have functional checkpoints and may exit the cell cycle into a quiescent state under conditions of AA deprivation, while cancer cells with DNA defects in the cell cycle checkpoint machinery may fail to enter quiescence. If a cell cannot stop dividing under conditions of nutrient deprivation, the lack of specific nutrients may trigger cell death [382]. Finally, as discussed previously, the DNA aberrations that provide cancer cells with a survival advantage under normal metabolic environments may become lethal under unfavorable metabolic environments created through dietary manipulation of AAs. Normal cells have a functional DNA and would resist these temporal AA imbalances. Importantly, these unfavorable environments can be created with different types of diets, and a new metabolic environment may be toxic to cancer cells with different sets of mutations [54]. This would explain why different diets induce antitumor activity in the same cancer type, and why the same diet can be active in different types of cancer [343].
Although our review is focused on dietary manipulation of AAs, it is important to note that the anticancer efficacy of this therapeutic strategy can be increased by simultaneously manipulating other dietary constituents. In fact, many of our active diets in mice with metastatic cancers were formulated with very low levels of lipids (1%) [26,27,119,343]. Currently, we are also manipulating other dietary constituents, such as vitamins and minerals. Since artificial diets can be prepared from scratch, any dietary component can be completely eliminated. In addition, although dietary manipulation of AAs and other nutrients may be clinically useful as monotherapy, future studies should evaluate artificial diets in combination with the standard therapies used in cancer patients. Although a diet can have a major impact on cancer progression and response to treatment, cancer patients are currently allowed to choose any type of diet. Future research will probably show that patient survival can be improved by matching specific therapies with specific and controlled diets.
7. Conclusions
Cancer cells reprogram their metabolism to produce the large amounts of building blocks required for biosynthesis and proliferation, to fulfill their high energy demands, and to survive under conditions of elevated oxidative stress. The altered AA metabolism of cancer cells is one of most therapeutically relevant metabolic features of cancer. In this work, we have reviewed studies that have evaluated the cancer therapeutic potential of dietary AA manipulation in vivo. These studies suggest that dietary manipulation of AAs with artificial diets is a feasible strategy to create harmful metabolic environments for cancer cells and achieve anticancer activity in vivo.
8. Patents
J.J. Jiménez-Alonso and M. López-Lázaro are inventors of a patent related to this work licensed to AMINOVITA, S.L., and the University of Seville.
Author Contributions
Conceptualization, M.L.-L.; investigation, J.J.J.-A. and M.L.-L.; writing—original draft preparation, J.J.J.-A.; writing—review and editing, J.J.J.-A and M.L.-L.; supervision, M.L.-L.; funding acquisition, M.L.-L. All authors have read and agreed to the published version of the manuscript.
Funding
Part of the experimental research reviewed in this work was funded by AMINOVITA, S.L. (PRJ201803388). Additional support was given by Junta de Andalucía (grant numbers 2017/CTS-657; 2019/CTS-657; 2021/CTS-657).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
J.J.J.-A thanks Ministerio de Universidades (Spain) for a predoctoral FPU fellowship (FPU17/02185).
Conflicts of Interest
M. López-Lázaro has shares in AMINOVITA, S.L. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Warburg, O. On the Origin of Cancer Cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef]
- Kostakoglu, L.; Agress, H.; Goldsmith, S.J. Clinical Role of FDG PET in Evaluation of Cancer Patients. Radiographics 2003, 23, 315–340. [Google Scholar] [CrossRef] [PubMed]
- Ben-Haim, S.; Ell, P. 18F-FDG PET and PET/CT in the Evaluation of Cancer Treatment Response. J. Nucl. Med. 2009, 50, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Lazaro, M. The Warburg Effect: Why and How Do Cancer Cells Activate Glycolysis in the Presence of Oxygen? Anticancer. Agents Med. Chem. 2008, 8, 305–312. [Google Scholar] [CrossRef] [PubMed]
- López-Lázaro, M. Does Hypoxia Really Control Tumor Growth? Anal. Cell. Pathol. 2006, 28, 327–329. [Google Scholar] [CrossRef]
- Lopez-Lazaro, M. A New View of Carcinogenesis and an Alternative Approach to Cancer Therapy. Mol. Med. 2010, 16, 144–153. [Google Scholar] [CrossRef]
- López-Lázaro, M. Excessive Superoxide Anion Generation Plays a Key Role in Carcinogenesis. Int. J. Cancer 2007, 120, 1378–1380. [Google Scholar] [CrossRef]
- López-Lázaro, M. Dual Role of Hydrogen Peroxide in Cancer: Possible Relevance to Cancer Chemoprevention and Therapy. Cancer Lett. 2007, 252, 1–8. [Google Scholar] [CrossRef]
- Hayes, J.D.; Dinkova-Kostova, A.T.; Tew, K.D. Oxidative Stress in Cancer. Cancer Cell 2020, 38, 167–197. [Google Scholar] [CrossRef]
- Pavlova, N.N.; Zhu, J.; Thompson, C.B. The Hallmarks of Cancer Metabolism: Still Emerging. Cell Metab. 2022, 34, 355–377. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Pavlova, N.N.; Thompson, C.B. The Emerging Hallmarks of Cancer Metabolism. Cell Metab. 2016, 23, 27–47. [Google Scholar] [CrossRef] [PubMed]
- Stine, Z.E.; Schug, Z.T.; Salvino, J.M.; Dang, C.V. Targeting Cancer Metabolism in the Era of Precision Oncology. Nat. Rev. Drug Discov. 2022, 21, 141–162. [Google Scholar] [CrossRef]
- Pranzini, E.; Pardella, E.; Paoli, P.; Fendt, S.-M.; Taddei, M.L. Metabolic Reprogramming in Anticancer Drug Resistance: A Focus on Amino Acids. Trends in Cancer 2021, 7, 682–699. [Google Scholar] [CrossRef] [PubMed]
- Bergers, G.; Fendt, S.-M. The Metabolism of Cancer Cells during Metastasis. Nat. Rev. Cancer 2021, 21, 162–180. [Google Scholar] [CrossRef]
- Wei, Z.; Liu, X.; Cheng, C.; Yu, W.; Yi, P. Metabolism of Amino Acids in Cancer. Front. Cell Dev. Biol. 2021, 8, 1628. [Google Scholar] [CrossRef]
- Endicott, M.; Jones, M.; Hull, J. Amino Acid Metabolism as a Therapeutic Target in Cancer: A Review. Amino Acids 2021, 53, 1169–1179. [Google Scholar] [CrossRef] [PubMed]
- Lukey, M.J.; Katt, W.P.; Cerione, R.A. Targeting Amino Acid Metabolism for Cancer Therapy. Drug Discov. Today 2017, 22, 796–804. [Google Scholar] [CrossRef]
- Safrhansova, L.; Hlozkova, K.; Starkova, J. Targeting Amino Acid Metabolism in Cancer. Int. Rev. Cell Mol. Biol. 2022, 373, 37–79. [Google Scholar] [CrossRef]
- Choi, B.-H.; Coloff, J.L. The Diverse Functions of Non-Essential Amino Acids in Cancer. Cancers (Basel). 2019, 11, 675. [Google Scholar] [CrossRef]
- Butler, M.; van der Meer, L.T.; van Leeuwen, F.N. Amino Acid Depletion Therapies: Starving Cancer Cells to Death. Trends Endocrinol. Metab. 2021, 32, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Vernieri, C.; Casola, S.; Foiani, M.; Pietrantonio, F.; de Braud, F.; Longo, V. Targeting Cancer Metabolism: Dietary and Pharmacologic Interventions. Cancer Discov. 2016, 6, 1315–1333. [Google Scholar] [CrossRef]
- Pui, C.H.; Mullighan, C.G.; Evans, W.E.; Relling, M.V. Pediatric Acute Lymphoblastic Leukemia: Where Are We Going and How Do We Get There? Blood 2012, 120, 1165–1174. [Google Scholar] [CrossRef] [PubMed]
- Tajan, M.; Vousden, K.H. Dietary Approaches to Cancer Therapy. Cancer Cell 2020, 37, 767–785. [Google Scholar] [CrossRef]
- Connolly-Schoonen, J.; Biamonte, S.F.; Danowski, L.; Montrose, D.C. Modifying Dietary Amino Acids in Cancer Patients. Int. Rev. Cell Mol. Biol. 2022, 373, 1–36. [Google Scholar] [CrossRef]
- Jiménez-Alonso, J.J.; Guillén-Mancina, E.; Calderón-Montaño, J.M.; Jiménez-González, V.; Díaz-Ortega, P.; Burgos-Morón, E.; López-Lázaro, M. Artificial Diets Based on Selective Amino Acid Restriction versus Capecitabine in Mice with Metastatic Colon Cancer. Nutrients 2022, 14, 3378. [Google Scholar] [CrossRef] [PubMed]
- Calderón-Montaño, J.M.; Guillén-Mancina, E.; Jiménez-Alonso, J.J.; Jiménez-González, V.; Burgos-Morón, E.; Mate, A.; Pérez-Guerrero, M.C.; López-Lázaro, M. Manipulation of Amino Acid Levels with Artificial Diets Induces a Marked Anticancer Activity in Mice with Renal Cell Carcinoma. Int. J. Mol. Sci 2022, 2022, 16132. [Google Scholar] [CrossRef]
- Kitada, M.; Ogura, Y.; Monno, I.; Koya, D. The Impact of Dietary Protein Intake on Longevity and Metabolic Health. EBioMedicine 2019, 43, 632–640. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, H.; Suarez, J.A.; Longo, V.D. Protein and Amino Acid Restriction, Aging and Disease: From Yeast to Humans. Trends Endocrinol. Metab. 2014, 25, 558–566. [Google Scholar] [CrossRef]
- Yin, J.; Ren, W.; Huang, X.; Li, T.; Yin, Y. Protein Restriction and Cancer. Biochim. Biophys. Acta - Rev. Cancer 2018, 1869, 256–262. [Google Scholar] [CrossRef]
- Levine, M.E.; Suarez, J.A.; Brandhorst, S.; Balasubramanian, P.; Cheng, C.-W.; Madia, F.; Fontana, L.; Mirisola, M.G.; Guevara-Aguirre, J.; Wan, J.; et al. Low Protein Intake Is Associated with a Major Reduction in IGF-1, Cancer, and Overall Mortality in the 65 and Younger but Not Older Population. Cell Metab. 2014, 19, 407–417. [Google Scholar] [CrossRef]
- Brandhorst, S.; Wei, M.; Hwang, S.; Morgan, T.E.; Longo, V.D. Short-Term Calorie and Protein Restriction Provide Partial Protection from Chemotoxicity but Do Not Delay Glioma Progression. Exp. Gerontol. 2013, 48, 1120–1128. [Google Scholar] [CrossRef]
- Rubio-Patiño, C.; Bossowski, J.P.; De Donatis, G.M.; Mondragón, L.; Villa, E.; Aira, L.E.; Chiche, J.; Mhaidly, R.; Lebeaupin, C.; Marchetti, S.; et al. Low-Protein Diet Induces IRE1α-Dependent Anticancer Immunosurveillance. Cell Metab. 2018, 27, 828–842. [Google Scholar] [CrossRef]
- Orillion, A.; Damayanti, N.P.; Shen, L.; Adelaiye-Ogala, R.; Affronti, H.; Elbanna, M.; Chintala, S.; Ciesielski, M.; Fontana, L.; Kao, C.; et al. Dietary Protein Restriction Reprograms Tumor-Associated Macrophages and Enhances Immunotherapy. Clin. Cancer Res. 2018, 24, 6383–6395. [Google Scholar] [CrossRef]
- Fontana, L.; Adelaiye, R.M.; Rastelli, A.L.; Miles, K.M.; Ciamporcero, E.; Longo, V.D.; Nguyen, H.; Vessella, R.; Pili, R. Dietary Protein Restriction Inhibits Tumor Growth in Human Xenograft Models of Prostate and Breast Cancer. Oncotarget 2013, 4, 2451–2461. [Google Scholar] [CrossRef]
- Taha, A.A.A.; Koshiyama, M.; Matsumura, N.; Abiko, K.; Yamaguchi, K.; Hamanishi, J.; Baba, T.; Kharma, B.; Mohamed, I.H.; Ameen, M.M.; et al. The Effect of the Type of Dietary Protein on the Development of Ovarian Cancer. Oncotarget 2018, 9, 23987–23999. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Amino Acids: Metabolism, Functions, and Nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef]
- Lee, J.H.; Cho, Y. ra; Kim, J.H.; Kim, J.; Nam, H.Y.; Kim, S.W.; Son, J. Branched-Chain Amino Acids Sustain Pancreatic Cancer Growth by Regulating Lipid Metabolism. Exp. Mol. Med. 2019, 51, 1–11. [Google Scholar] [CrossRef]
- Sugimura, T.; Birnbaum, S.M.; Winitz, M.; Greenstein, J.P. Quantitative Nutritional Studies with Water-Soluble, Chemically Defined Diets. VIII. The Forced Feeding of Diets Each Lacking in One Essential Amino Acid. Arch. Biochem. Biophys. 1959, 81, 448–455. [Google Scholar] [CrossRef]
- Theuer, R.C. Effect of Essential Amino Acid Restriction on the Growth of Female C57BL Mice and Their Implanted BW10232 Adenocarcinomas. J. Nutr. 1971, 101, 223–232. [Google Scholar] [CrossRef]
- Sheen, J.-H.; Zoncu, R.; Kim, D.; Sabatini, D.M. Defective Regulation of Autophagy upon Leucine Deprivation Reveals a Targetable Liability of Human Melanoma Cells in Vitro and in Vivo. Cancer Cell 2011, 19, 613–628. [Google Scholar] [CrossRef]
- Xiao, F.; Wang, C.; Yin, H.; Yu, J.; Chen, S.; Fang, J.; Guo, F. Leucine Deprivation Inhibits Proliferation and Induces Apoptosis of Human Breast Cancer Cells via Fatty Acid Synthase. Oncotarget 2016, 7, 63679–63689. [Google Scholar] [CrossRef]
- Tönjes, M.; Barbus, S.; Park, Y.J.; Wang, W.; Schlotter, M.; Lindroth, A.M.; Pleier, S.V.; Bai, A.H.C.; Karra, D.; Piro, R.M.; et al. BCAT1 Promotes Cell Proliferation through Amino Acid Catabolism in Gliomas Carrying Wild-Type IDH1. Nat. Med. 2013, 19, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Q.; Faddaoui, A.; Bachvarova, M.; Plante, M.; Gregoire, J.; Renaud, M.C.; Sebastianelli, A.; Guillemette, C.; Gobeil, S.; Macdonald, E.; et al. BCAT1 Expression Associates with Ovarian Cancer Progression: Possible Implications in Altered Disease Metabolism. Oncotarget 2015, 6, 31522–31543. [Google Scholar] [CrossRef]
- Zheng, Y.H.; Hu, W.J.; Chen, B.C.; Grahn, T.H.M.; Zhao, Y.R.; Bao, H.L.; Zhu, Y.F.; Zhang, Q.Y. BCAT1, a Key Prognostic Predictor of Hepatocellular Carcinoma, Promotes Cell Proliferation and Induces Chemoresistance to Cisplatin. Liver Int. 2016, 36, 1836–1847. [Google Scholar] [CrossRef]
- Hattori, A.; Tsunoda, M.; Konuma, T.; Kobayashi, M.; Nagy, T.; Glushka, J.; Tayyari, F.; McSkimming, D.; Kannan, N.; Tojo, A.; et al. Cancer Progression by Reprogrammed BCAA Metabolism in Myeloid Leukaemia. Nature 2017, 545, 500–504. [Google Scholar] [CrossRef]
- Zhu, Z.; Achreja, A.; Meurs, N.; Animasahun, O.; Owen, S.; Mittal, A.; Parikh, P.; Lo, T.W.; Franco-Barraza, J.; Shi, J.; et al. Tumour-Reprogrammed Stromal BCAT1 Fuels Branched-Chain Ketoacid Dependency in Stromal-Rich PDAC Tumours. Nat. Metab. 2020, 2, 775–792. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Han, J. Branched-Chain Amino Acid Transaminase 1 (BCAT1) Promotes the Growth of Breast Cancer Cells through Improving MTOR-Mediated Mitochondrial Biogenesis and Function. Biochem. Biophys. Res. Commun. 2017, 486, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.A.; Lashinger, L.M.; Rasmussen, A.J.; Hursting, S.D. Leucine Supplementation Differentially Enhances Pancreatic Cancer Growth in Lean and Overweight Mice. Cancer Metab. 2014, 2. [Google Scholar] [CrossRef]
- Atherton, P.J.; Smith, K.; Etheridge, T.; Rankin, D.; Rennie, M.J. Distinct Anabolic Signalling Responses to Amino Acids in C2C12 Skeletal Muscle Cells. Amino Acids 2010, 38, 1533–1539. [Google Scholar] [CrossRef] [PubMed]
- Han, J.M.M.; Jeong, S.J.J.; Park, M.C.C.; Kim, G.; Kwon, N.H.H.; Kim, H.K.K.; Ha, S.H.H.; Ryu, S.H.H.; Kim, S. Leucyl-TRNA Synthetase Is an Intracellular Leucine Sensor for the MTORC1-Signaling Pathway. Cell 2012, 149, 410–424. [Google Scholar] [CrossRef]
- Takahara, T.; Amemiya, Y.; Sugiyama, R.; Maki, M.; Shibata, H. Amino Acid-Dependent Control of MTORC1 Signaling: A Variety of Regulatory Modes. J. Biomed. Sci. 2020, 27, 87. [Google Scholar] [CrossRef] [PubMed]
- Kamei, Y.; Hatazawa, Y.; Uchitomi, R.; Yoshimura, R.; Miura, S. Regulation of Skeletal Muscle Function by Amino Acids. Nutrients 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- López-Lázaro, M. Selective Amino Acid Restriction Therapy (SAART): A Non- Pharmacological Strategy against All Types of Cancer Cells. Oncoscience 2015, 2, 857. [Google Scholar] [CrossRef] [PubMed]
- Beaudry, A.G.; Law, M.L. Leucine Supplementation in Cancer Cachexia: Mechanisms and a Review of the Pre-Clinical Literature. Nutrients 2022, 14, 2824. [Google Scholar] [CrossRef]
- Baracos, V.E.; Martin, L.; Korc, M.; Guttridge, D.C.; Fearon, K.C.H. Cancer-Associated Cachexia. Nat. Rev. Dis. Prim. 2018, 4, 17105. [Google Scholar] [CrossRef] [PubMed]
- Saxton, R.A.; Sabatini, D.M. MTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef]
- Osburn, S.C.; Vann, C.G.; Church, D.D.; Ferrando, A.A.; Roberts, M.D. Proteasome- and Calpain-Mediated Proteolysis, but Not Autophagy, Is Required for Leucine-Induced Protein Synthesis in C2C12 Myotubes. Physiologia 2021, 1, 22–33. [Google Scholar] [CrossRef]
- Viana, L.R.; Chiocchetti, G. de M. e; Oroy, L.; Vieira, W.F.; Busanello, E.N.B.; Marques, A.C.; Salgado, C. de M.; de Oliveira, A.L.R.; Vieira, A.S.; Suarez, P.S.; et al. Leucine-Rich Diet Improved Muscle Function in Cachectic Walker 256 Tumour-Bearing Wistar Rats. Cells 2021, 10, 3272. [Google Scholar] [CrossRef]
- Cruz, B.; Oliveira, A.; Viana, L.R.; Lopes-Aguiar, L.; Canevarolo, R.; Colombera, M.C.; Valentim, R.R.; Garcia-Fóssa, F.; de Sousa, L.M.; Castelucci, B.G.; et al. Leucine-Rich Diet Modulates the Metabolomic and Proteomic Profile of Skeletal Muscle during Cancer Cachexia. Cancers (Basel). 2020, 12, 1880. [Google Scholar] [CrossRef] [PubMed]
- Toneto, A.T.; Ferreira Ramos, L.A.; Salomão, E.M.; Tomasin, R.; Aereas, M.A.; Gomes-Marcondes, M.C.C. Nutritional Leucine Supplementation Attenuates Cardiac Failure in Tumour-Bearing Cachectic Animals. J. Cachexia. Sarcopenia Muscle 2016, 7, 577–586. [Google Scholar] [CrossRef]
- Ventrucci, G.; Mello, M.A.R.; Gomes-Marcondes, M.C.C. Proteasome Activity Is Altered in Skeletal Muscle Tissue of Tumour-Bearing Rats Fed a Leucine-Rich Diet. Endocr. Relat. Cancer 2004, 11, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Salomão, E.M.; Toneto, A.T.; Silva, G.O.; Gomes-Marcondes, M.C.C. Physical Exercise and a Leucine-Rich Diet Modulate the Muscle Protein Metabolism in Walker Tumor-Bearing Rats. Nutr. Cancer 2010, 62, 1095–1104. [Google Scholar] [CrossRef] [PubMed]
- Salomão, E.M.; Gomes-Marcondes, M.C.C. Light Aerobic Physical Exercise in Combination with Leucine and/or Glutamine-Rich Diet Can Improve the Body Composition and Muscle Protein Metabolism in Young Tumor-Bearing Rats. J. Physiol. Biochem. 2012, 68, 493–501. [Google Scholar] [CrossRef]
- Gomes-Marcondes, M.C.C.; Ventrucci, G.; Toledo, M.T.; Cury, L.; Cooper, J.C. A Leucine-Supplemented Diet Improved Protein Content of Skeletal Muscle in Young Tumor-Bearing Rats. Brazilian J. Med. Biol. Res. 2003, 36, 1589–1594. [Google Scholar] [CrossRef] [PubMed]
- Cruz, B.; Oliveira, A.; Gomes-Marcondes, M.C.C. L-Leucine Dietary Supplementation Modulates Muscle Protein Degradation and Increases pro-Inflammatory Cytokines in Tumour-Bearing Rats. Cytokine 2017, 96, 253–260. [Google Scholar] [CrossRef]
- Peters, S.J.; Van Helvoort, A.; Kegler, D.; Argilès, J.M.; Luiking, Y.C.; Laviano, A.; Van Bergenhenegouwen, J.; Deutz, N.E.P.; Haagsman, H.P.; Gorselink, M.; et al. Dose-Dependent Effects of Leucine Supplementation on Preservation of Muscle Mass in Cancer Cachectic Mice. Oncol. Rep. 2011, 26, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Van Norren, K.; Kegler, D.; Argilés, J.M.; Luiking, Y.; Gorselink, M.; Laviano, A.; Arts, K.; Faber, J.; Jansen, H.; Van Der Beek, E.M.; et al. Dietary Supplementation with a Specific Combination of High Protein, Leucine, and Fish Oil Improves Muscle Function and Daily Activity in Tumour-Bearing Cachectic Mice. Br. J. Cancer 2009, 100, 713–722. [Google Scholar] [CrossRef]
- Gu, C.; Mao, X.; Chen, D.; Yu, B.; Yang, Q. Isoleucine Plays an Important Role for Maintaining Immune Function. Curr. Protein Pept. Sci. 2019, 20, 644–651. [Google Scholar] [CrossRef]
- Xiao, F.; Yu, J.; Guo, Y.; Deng, J.; Li, K.; Du, Y.; Chen, S.; Zhu, J.; Sheng, H.; Guo, F. Effects of Individual Branched-Chain Amino Acids Deprivation on Insulin Sensitivity and Glucose Metabolism in Mice. Metabolism. 2014, 63, 841–850. [Google Scholar] [CrossRef]
- Williams, C.D. A Nutritional Disease of Childhood Associated with a Maize Diet. Arch. Dis. Child. 1983, 58, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Kocher, R.A. Effects of a Low Lysine Diet on the Growth of Spontaneous Mammary Tumors in Mice and on the N2 Balance in Man. Cancer Res. 1944, 4, 251–256. [Google Scholar]
- van Spronsen, F.J.; Blau, N.; Harding, C.; Burlina, A.; Longo, N.; Bosch, A.M. Phenylketonuria. Nat. Rev. Dis. Prim. 2021, 7, 36. [Google Scholar] [CrossRef] [PubMed]
- Ryan, W.L.; Elliott, J.A. Fluorophenylalanine Inhibition of Tumors in Mice on a Phenylalanine-Deficient Diet. Arch. Biochem. Biophys. 1968, 125, 797–801. [Google Scholar] [CrossRef]
- Demopoulos, H.B. Effects of Low Phenylalanine-Tyrosine Diets on S91 Mouse Melanomas. J. Natl. Cancer Inst. 1966, 37, 185–190. [Google Scholar] [CrossRef]
- Bounous, G.; Kongshavn, P.A.L. The Effect of Dietary Amino Acids on the Growth of Tumors. Experientia 1981, 37, 271–273. [Google Scholar] [CrossRef]
- Abdallah, R.M.; Starkey, J.R.; Meadows, G.G. Dietary Restriction of Tyrosine and Phenylalanine: Inhibition of Metastasis of Three Rodent Tumors. J. Natl. Cancer Inst. 1987, 78, 759–769. [Google Scholar] [CrossRef]
- Elstad, C.A.; Meadows, G.G.; Abdallah, R.M. Specificity of the Suppression of Metastatic Phenotype by Tyrosine and Phenylalanine Restriction. Clin. Exp. Metastasis 1990, 8, 393–416. [Google Scholar] [CrossRef]
- Elstad, C.A.; Thrall, B.D.; Raha, G.; Meadows, G.G. Tyrosine and Phenylalanine Restriction Sensitizes Adriamycin-Resistant P388 Leukemia Cells to Adriamycin. Nutr. Cancer 1996, 25, 47–60. [Google Scholar] [CrossRef]
- Uhlenkott, C.E.; Huijzer, J.C.; Cardeiro, D.J.; Elstad, C.A.; Meadows, G.G. Attachment, Invasion, Chemotaxis, and Proteinase Expression of B16-BL6 Melanoma Cells Exhibiting a Low Metastatic Phenotype after Exposure to Dietary Restriction of Tyrosine and Phenylalanine. Clin. Exp. Metastasis 1996, 14, 125–137. [Google Scholar] [CrossRef]
- Pelayo, B.A.; Fu, Y.M.; Meadows, G.G. Inhibition of B16BL6 Melanoma Invasion by Tyrosine and Phenylalanine Deprivation Is Associated with Decreased Secretion of Plasminogen Activators and Increased Plasminogen Activator Inhibitors. Clin. Exp. Metastasis 1999, 17, 841–848. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.M.; Yu, Z.X.; Li, Y.Q.; Ge, X.; Sanchez, P.J.; Fu, X.; Meadows, G.G. Specific Amino Acid Dependency Regulates Invasiveness and Viability of Androgen-Independent Prostate Cancer Cells. Nutr. Cancer 2003, 45, 60–73. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.M.; Zhang, H.; Ding, M.; Li, Y.Q.; Fu, X.; Yu, Z.X.; Meadows, G.G. Specific Amino Acid Restriction Inhibits Attachment and Spreading of Human Melanoma via Modulation of the Integrin/Focal Adhesion Kinase Pathway and Actin Cytoskeleton Remodeling. Clin. Exp. Metastasis 2005, 21, 587–598. [Google Scholar] [CrossRef]
- Fu, Y.M.; Zhang, H.; Ding, M.; Li, Y.Q.; Fu, X.; Yu, Z.X.; Meadows, G.G. Selective Amino Acid Restriction Targets Mitochondria to Induce Apoptosis of Androgen-Independent Prostate Cancer Cells. J. Cell. Physiol. 2006, 209, 522–534. [Google Scholar] [CrossRef] [PubMed]
- Núñez, N.P.; Liu, H.; Meadows, G.G. PPAR-γ Ligands and Amino Acid Deprivation Promote Apoptosis of Melanoma, Prostate, and Breast Cancer Cells. Cancer Lett. 2006, 236, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Fu, Y.-M.; Meadows, G.G. Differential Effects of Specific Amino Acid Restriction on Glucose Metabolism, Reduction/Oxidation Status and Mitochondrial Damage in DU145 and PC3 Prostate Cancer Cells. Oncol. Lett. 2011, 2, 349–355. [Google Scholar] [CrossRef]
- Demopoulos, H.B. Effects of Reducing the Phenylalanine-Tyrosine Intake of Patients with Advanced Malignant Melanoma. Cancer 1966, 19, 657–664. [Google Scholar] [CrossRef]
- Lorincz, A.B.; Kuttner, R.E. Response of Malignancy to Phenylalanine Restriction. A Preliminary Report on a New Concept of Managing Malignant Disease. Nebr. State Med. J. 1965, 50, 609–617. [Google Scholar]
- Edmund, J.; Jensen, O.A.; Egeberg, J. Reduced Intake of Phenylalanine and Tyrosine as Treatment of Choroidal Malignant Melanoma. Mod. Probl. Ophthalmol. 1974, 12, 504–509. [Google Scholar]
- Lawson, D.H.; Stockton, L.H.; Bleier, J.C.; Acosta, P.B.; Heymsfield, S.B.; Nixon, D.W. The Effect of a Phenylalanine and Tyrosine Restricted Diet on Elemental Balance Studies and Plasma Aminograms of Patients with Disseminated Malignant Melanoma. Am. J. Clin. Nutr. 1985, 41, 73–84. [Google Scholar] [CrossRef]
- Harvie, M.N.; Campbell, I.T.; Howell, A.; Thatcher, N. Acceptability and Tolerance of a Low Tyrosine and Phenylalanine Diet in Patients with Advanced Cancer - a Pilot Study. J. Hum. Nutr. Diet. 2002, 15, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, M.E.; Brosnan, J.T. Histidine Metabolism and Function. J. Nutr. 2020, 150, 2570S–2575S. [Google Scholar] [CrossRef] [PubMed]
- Froldi, F.; Pachnis, P.; Szuperák, M.; Costas, O.; Fernando, T.; Gould, A.P.; Cheng, L.Y. Histidine Is Selectively Required for the Growth of Myc-dependent Dedifferentiation Tumours in the Drosophila CNS. EMBO J. 2019, 38, e99895. [Google Scholar] [CrossRef] [PubMed]
- Kanarek, N.; Keys, H.R.; Cantor, J.R.; Lewis, C.A.; Chan, S.H.; Kunchok, T.; Abu-Remaileh, M.; Freinkman, E.; Schweitzer, L.D.; Sabatini, D.M. Histidine Catabolism Is a Major Determinant of Methotrexate Sensitivity. Nature 2018, 559, 632–636. [Google Scholar] [CrossRef] [PubMed]
- Opitz, C.A.; Somarribas Patterson, L.F.; Mohapatra, S.R.; Dewi, D.L.; Sadik, A.; Platten, M.; Trump, S. The Therapeutic Potential of Targeting Tryptophan Catabolism in Cancer. Br. J. Cancer 2020, 122, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Tomek, P. Tryptophan: A Rheostat of Cancer Immune Escape Mediated by Immunosuppressive Enzymes IDO1 and TDO. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Kamath, S.K.; Conrad, N.C.; Olson, R.E.; Kohrs, M.B.; Ghosh, L. Amino Acid-Restricted Diets in the Treatment of Mammary Adenocarcinoma in Mice. J. Nutr. 1988, 118, 1137–1142. [Google Scholar] [CrossRef]
- Brenk, M.; Scheler, M.; Koch, S.; Neumann, J.; Takikawa, O.; Häcker, G.; Bieber, T.; von Bubnoff, D. Tryptophan Deprivation Induces Inhibitory Receptors ILT3 and ILT4 on Dendritic Cells Favoring the Induction of Human CD4 + CD25 + Foxp3 + T Regulatory Cells. J. Immunol. 2009, 183, 145–154. [Google Scholar] [CrossRef]
- Fallarino, F.; Grohmann, U.; You, S.; McGrath, B.C.; Cavener, D.R.; Vacca, C.; Orabona, C.; Bianchi, R.; Belladonna, M.L.; Volpi, C.; et al. The Combined Effects of Tryptophan Starvation and Tryptophan Catabolites Down-Regulate T Cell Receptor ζ-Chain and Induce a Regulatory Phenotype in Naive T Cells. J. Immunol. 2006, 176, 6752–6761. [Google Scholar] [CrossRef]
- Schramme, F.; Crosignani, S.; Frederix, K.; Hoffmann, D.; Pilotte, L.; Stroobant, V.; Preillon, J.; Driessens, G.; van den Eynde, B.J. Inhibition of Tryptophan-Dioxygenase Activity Increases the Antitumor Efficacy of Immune Checkpoint Inhibitors. Cancer Immunol. Res. 2020, 8, 32–45. [Google Scholar] [CrossRef]
- Günther, J.; Däbritz, J.; Wirthgen, E. Limitations and Off-Target Effects of Tryptophan-Related IDO Inhibitors in Cancer Treatment. Front. Immunol. 2019, 10, 1801. [Google Scholar] [CrossRef] [PubMed]
- Long, G.V.; Dummer, R.; Hamid, O.; Gajewski, T.F.; Caglevic, C.; Dalle, S.; Arance, A.; Carlino, M.S.; Grob, J.J.; Kim, T.M.; et al. Epacadostat plus Pembrolizumab versus Placebo plus Pembrolizumab in Patients with Unresectable or Metastatic Melanoma (ECHO-301/KEYNOTE-252): A Phase 3, Randomised, Double-Blind Study. Lancet Oncol. 2019, 20, 1083–1097. [Google Scholar] [CrossRef]
- Mato, J.M.; Martínez-Chantar, M.L.; Lu, S.C. S-Adenosylmethionine Metabolism and Liver Disease. Ann. Hepatol. 2013, 12, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Weber, R.; Birsoy, K. The Transsulfuration Pathway Makes, the Tumor Takes. Cell Metab. 2019, 30, 845–846. [Google Scholar] [CrossRef] [PubMed]
- Hoshiya, Y.; Kubota, T.; Matsuzaki, S.W.; Kitajima, M.; Hoffman, R.M. Methionine Starvation Modulates the Efficacy of Cisplatin on Human Breast Cancer in Nude Mice. Anticancer Res. 1996, 16, 3515–3518. [Google Scholar] [PubMed]
- Hoshiya, Y.; Kubota, T.; Inada, T.; Kitajima, M.; Hoffman, R.M. Methionine-Depletion Modulates the Efficacy of 5-Fluorouracil in Human Gastric Cancer in Nude Mice. Anticancer Res. 1997, 17, 4371–4375. [Google Scholar]
- Guo, H.; Lishko, V.K.; Herrera, H.; Groce, A.; Kubota, T.; Hoffman, R.M. Therapeutic Tumor-Specific Cell Cycle Block Induced by Methionine Starvation in Vivo. Cancer Res. 1993, 53, 5676–5679. [Google Scholar] [PubMed]
- Jeon, H.; Kim, J.H.; Lee, E.; Jang, Y.J.; Son, J.E.; Kwon, J.Y.; Lim, T.; Kim, S.; Park, J.H.Y.; Kim, J.-E.; et al. Methionine Deprivation Suppresses Triple-Negative Breast Cancer Metastasis in Vitro and in Vivo. Oncotarget 2016, 7, 67223–67234. [Google Scholar] [CrossRef]
- Strekalova, E.; Malin, D.; Good, D.M.; Cryns, V.L. Methionine Deprivation Induces a Targetable Vulnerability in Triple-Negative Breast Cancer Cells by Enhancing TRAIL Receptor-2 Expression. Clin. Cancer Res. 2015, 21, 2780–2791. [Google Scholar] [CrossRef]
- Malin, D.; Lee, Y.; Chepikova, O.; Strekalova, E.; Carlson, A.; Cryns, V.L. Methionine Restriction Exposes a Targetable Redox Vulnerability of Triple-Negative Breast Cancer Cells by Inducing Thioredoxin Reductase. Breast Cancer Res. Treat. 2021, 190, 373–387. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, W.; Wang, K.; Wang, X.; Yin, F.; Li, C.; Wang, C.; Zhao, B.; Zhong, C.; Zhang, J.; et al. Methionine and Cystine Double Deprivation Stress Suppresses Glioma Proliferation via Inducing ROS/Autophagy. Toxicol. Lett. 2015, 232, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.; Cooper, T.K.; Rogers, C.J.; Sinha, I.; Turbitt, W.J.; Calcagnotto, A.; Perrone, C.E.; Richie, J.P. Dietary Methionine Restriction Inhibits Prostatic Intraepithelial Neoplasia in TRAMP Mice. Prostate 2014, 74, 1663–1673. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Li, Y.; Gao, X.; Kang, K.; Williams, J.G.; Tong, L.; Liu, J.; Ji, M.; Deterding, L.J.; Tong, X.; et al. HNF4α Regulates Sulfur Amino Acid Metabolism and Confers Sensitivity to Methionine Restriction in Liver Cancer. Nat. Commun. 2020, 11, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Sanderson, S.M.; Dai, Z.; Reid, M.A.; Cooper, D.E.; Lu, M.; Richie, J.P.; Ciccarella, A.; Calcagnotto, A.; Mikhael, P.G.; et al. Dietary Methionine Influences Therapy in Mouse Cancer Models and Alters Human Metabolism. Nature 2019, 572, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Hens, J.R.; Sinha, I.; Perodin, F.; Cooper, T.; Sinha, R.; Plummer, J.; Perrone, C.E.; Orentreich, D. Methionine-Restricted Diet Inhibits Growth of MCF10AT1-Derived Mammary Tumors by Increasing Cell Cycle Inhibitors in Athymic Nude Mice. BMC Cancer 2016, 16, 349. [Google Scholar] [CrossRef]
- Komninou, D.; Leutzinger, Y.; Reddy, B.S.; Richie, J.P. Methionine Restriction Inhibits Colon Carcinogenesis. Nutr. Cancer 2006, 54, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wang, J.L.; Wu, D.Z.; Yuan, Y.W.; Xin, L. Methionine Restriction Enhances the Chemotherapeutic Sensitivity of Colorectal Cancer Stem Cells by MiR-320d/c-Myc Axis. Mol. Cell. Biochem. 2022, 477, 2001–2013. [Google Scholar] [CrossRef]
- Li, T.; Tan, Y.-T.; Chen, Y.-X.; Zheng, X.-J.; Wang, W.; Liao, K.; Mo, H.-Y.; Lin, J.; Yang, W.; Piao, H.-L.; et al. Methionine Deficiency Facilitates Antitumour Immunity by Altering m 6 A Methylation of Immune Checkpoint Transcripts. Gut 2023, 72, 501–511. [Google Scholar] [CrossRef]
- Jiménez-Alonso, J.J.; Guillén-Mancina, E.; Calderón-Montaño, J.M.; Jiménez-González, V.; Díaz-Ortega, P.; Burgos-Morón, E.; López-Lázaro, M. Artificial Diets with Altered Levels of Sulfur Amino Acids Induce Anticancer Activity in Mice with Metastatic Colon Cancer, Ovarian Cancer and Renal Cell Carcinoma. Int. J. Mol. Sci. 2023, 24, 4587. [Google Scholar] [CrossRef]
- Upadhyayula, P.S.; Higgins, D.M.; Mela, A.; Banu, M.; Dovas, A.; Zandkarimi, F.; Patel, P.; Mahajan, A.; Humala, N.; Nguyen, T.T.T.; et al. Dietary Restriction of Cysteine and Methionine Sensitizes Gliomas to Ferroptosis and Induces Alterations in Energetic Metabolism. Nat. Commun. 2023, 14, 1187. [Google Scholar] [CrossRef]
- Goseki, N.; Endo, M.; Onodera, T.; Kosaki, G. Anti-Tumor Effect of L-Methionine-Deprived Total Parenteral Nutrition with 5-Fluorouracil Administration on Yoshida Sarcoma-Bearing Rats. Ann. Surg. 1991, 214, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Goseki, N.; Nagahama, T.; Maruyama, M.; Endo, M. Enhanced Anticancer Effect of Vincristine with Methionine Infusion after Methionine-Depleting Total Parenteral Nutrition in Tumor-Bearing Rats. Japanese J. Cancer Res. 1996, 87, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Tan, Y.; Kubota, T.; Moossa, A.R.; Hoffman, R.M. Methionine Depletion Modulates the Antitumor and Antimetastatic Efficacy of Ethionine. Anticancer Res. 1996, 16, 2719–2723. [Google Scholar] [PubMed]
- Xiao, H.B.; Cao, W.X.; Yin, H.R.; Lin, Y.Z.; Ye, S.H. Influence of L-Methionine-Deprived Total Parenteral Nutrition with 5-Fluorouracil on Gastric Cancer and Host Metabolism. World J. Gastroenterol. 2001, 7, 698–701. [Google Scholar] [CrossRef]
- Hoshiya, Y.; Guo, H.; Kubota, T.; Inada, T.; Asanuma, F.; Yamada, Y.; Koh, J.I.; Kitajima, M.; Hoffman, R.M. Human Tumors Are Methionine Dependent in Vivo. Anticancer Res. 1995, 15, 717–718. [Google Scholar] [PubMed]
- Miller, R.A.; Buehner, G.; Chang, Y.; Harper, J.M.; Sigler, R.; Smith-Wheelock, M. Methionine-Deficient Diet Extends Mouse Lifespan, Slows Immune and Lens Aging, Alters Glucose, T4, IGF-I and Insulin Levels, and Increases Hepatocyte MIF Levels and Stress Resistance. Aging Cell 2005, 4, 119–125. [Google Scholar] [CrossRef]
- Jonsson, W.O.; Margolies, N.S.; Anthony, T.G. Dietary Sulfur Amino Acid Restriction and the Integrated Stress Response: Mechanistic Insights. Nutrients 2019, 11, 1349. [Google Scholar] [CrossRef]
- Forney, L.A.; Wanders, D.; Stone, K.P.; Pierse, A.; Gettys, T.W. Concentration-Dependent Linkage of Dietary Methionine Restriction to the Components of Its Metabolic Phenotype. Obesity 2017, 25, 730–738. [Google Scholar] [CrossRef]
- Breillout, F.; Hadida, F.; Echinard-Garin, P.; Lascaux, V.; Poupon, M.F. Decreased Rat Rhabdomyosarcoma Pulmonary Metastases in Response to a Low Methionine Diet. Anticancer Res. 1987, 7, 861–868. [Google Scholar]
- Casero, R.A.; Murray Stewart, T.; Pegg, A.E. Polyamine Metabolism and Cancer: Treatments, Challenges and Opportunities. Nat. Rev. Cancer 2018, 18, 681–695. [Google Scholar] [CrossRef]
- Gomez, A.; Gomez, J.; Torres, M.L.; Naudi, A.; Mota-Martorell, N.; Pamplona, R.; Barja, G. Cysteine Dietary Supplementation Reverses the Decrease in Mitochondrial ROS Production at Complex I Induced by Methionine Restriction. J. Bioenerg. Biomembr. 2015, 47, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Elshorbagy, A.K.; Valdivia-Garcia, M.; Mattocks, D.A.L.; Plummer, J.D.; Smith, A.D.; Drevon, C.A.; Refsum, H.; Perrone, C.E. Cysteine Supplementation Reverses Methionine Restriction Effects on Rat Adiposity: Significance of Stearoyl-Coenzyme a Desaturase. J. Lipid Res. 2011, 52, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Voegtlin, C.; Johnson, J.M.; Thompson, J.W. Glutathione and Malignant Growth. Public Heal. Reports 1936, 51, 1689. [Google Scholar] [CrossRef]
- Cavuoto, P.; Fenech, M.F. A Review of Methionine Dependency and the Role of Methionine Restriction in Cancer Growth Control and Life-Span Extension. Cancer Treat. Rev. 2012, 38, 726–736. [Google Scholar] [CrossRef] [PubMed]
- Halpern, B.C.; Clark, B.R.; Hardy, D.N.; Halpern, R.M.; Smith, R.A. The Effect of Replacement of Methionine by Homocystine on Survival of Malignant and Normal Adult Mammalian Cells in Culture. Proc. Natl. Acad. Sci. U. S. A. 1974, 71, 1133–1136. [Google Scholar] [CrossRef]
- Kreis, W.; Goodenow, M. Methionine Requirement and Replacement by Homocysteine in Tissue Cultures of Selected Rodent and Human Malignant and Normal Cells. Cancer Res. 1978, 38, 2259–2262. [Google Scholar]
- Bertino, J.R.; Waud, W.R.; Parker, W.B.; Lubin, M. Targeting Tumors That Lack Methylthioadenosine Phosphorylase (MTAP) Activity. Cancer Biol. Ther. 2011, 11, 627–632. [Google Scholar] [CrossRef]
- Chaturvedi, S.; Hoffman, R.M.; Bertino, J.R. Exploiting Methionine Restriction for Cancer Treatment. Biochem. Pharmacol. 2018, 154, 170–173. [Google Scholar] [CrossRef]
- Lien, E.C.; Ghisolfi, L.; Geck, R.C.; Asara, J.M.; Toker, A. Oncogenic PI3K Promotes Methionine Dependency in Breast Cancer Cells through the Cystine-Glutamate Antiporter XCT. Sci. Signal. 2017, 10. [Google Scholar] [CrossRef]
- Bian, Y.; Li, W.; Kremer, D.M.; Sajjakulnukit, P.; Li, S.; Crespo, J.; Nwosu, Z.C.; Zhang, L.; Czerwonka, A.; Pawłowska, A.; et al. Cancer SLC43A2 Alters T Cell Methionine Metabolism and Histone Methylation. Nature 2020, 585, 277–282. [Google Scholar] [CrossRef]
- Goseki, N.; Yamazaki, S.; Shimojyu, K.; Kando, F.; Maruyama, M.; Endo, M.; Koike, M.; Takahashi, H. Synergistic Effect of Methionine-Depleting Total Parenteral Nutrition with 5-Fluorouracil on Human Gastric Cancer: A Randomized, Prospective Clinical Trial. Japanese J. Cancer Res. 1995, 86, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Epner, D.E.; Morrow, S.; Wilcox, M.; Houghton, J.L. Nutrient Intake and Nutritional Indexes in Adults with Metastatic Cancer on a Phase I Clinical Trial of Dietary Methionine Restriction. Nutr. Cancer 2002, 42, 158–166. [Google Scholar] [CrossRef]
- Thivat, E.; Durando, X.; Demidem, A.; Farges, M.C.; Rapp, M.; Cellarier, E.; Guenin, S.; D’Incan, M.; Vasson, M.P.; Chollet, P. A Methionine-Free Diet Associated with Nitrosourea Treatment down-Regulates Methylguanine-DNA Methyl Transferase Activity in Patients with Metastatic Cancer. Anticancer Res. 2007, 27, 2779–2783. [Google Scholar] [PubMed]
- Thivat, E.; Farges, M.-C.; Bacin, F.; D’Incan, M.; Mouret-Reynier, M.-A.; Cellarier, E.; Madelmont, J.-C.; Vasson, M.-P.; Chollet, P.; Durando, X. Phase II Trial of the Association of a Methionine-Free Diet with Cystemustine Therapy in Melanoma and Glioma. Anticancer Res. 2009, 29, 5235–5240. [Google Scholar] [PubMed]
- Durando, X.; Farges, M.-C.; Buc, E.; Abrial, C.; Petorin-Lesens, C.; Gillet, B.; Vasson, M.-P.; Pezet, D.; Chollet, P.; Thivat, E. Dietary Methionine Restriction with FOLFOX Regimen as First Line Therapy of Metastatic Colorectal Cancer: A Feasibility Study. Oncology 2010, 78, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Xu, M.; Tan, X.; Tan, X.; Wang, X.; Saikawa, Y.; Nagahama, T.; Sun, X.; Lenz, M.; Hoffman, R.M. Overexpression and Large-Scale Production of Recombinant L-Methionine-α-Deamino-γ-Mercaptomethane-Lyase for Novel Anticancer Therapy. Protein Expr. Purif. 1997, 9, 233–245. [Google Scholar] [CrossRef]
- Tan, Y.; Mingxu, X.U.; Guo, H.; Sun, X.; Kubota, T.; Hoffman, R.M. Anticancer Efficacy of Methioninase in Vivo. Anticancer Res. 1996, 16, 3931–3936. [Google Scholar]
- Tan, Y.; Sun, X.; Xu, M.; Tan, X.; Sasson, A.; Rashidi, B.; Han, Q.; Tan, X.; Wang, X.; An, Z.; et al. Efficacy of Recombinant Methioninase in Combination with Cisplatin on Human Colon Tumors in Nude Mice. Clin. Cancer Res. 1999, 5, 2157–2163. [Google Scholar]
- Yoshioka, T.; Wada, T.; Uchida, N.; Maki, H.; Yoshida, H.; Ide, N.; Kasai, H.; Hojo, K.; Shono, K.; Maekawa, R.; et al. Anticancer Efficacy in Vivo and in Vitro, Synergy with 5-Fluorouracil, and Safety of Recombinant Methioninase. Cancer Res. 1998, 58, 2583–2587. [Google Scholar]
- Kokkinakis, D.M.; Hoffman, R.M.; Frenkel, E.P.; Wick, J.B.; Han, Q.; Xu, M.; Tan, Y.; Schold, S.C. Synergy between Methionine Stress and Chemotherapy in the Treatment of Brain Tumor Xenografts in Athymic Mice. Cancer Res. 2001, 61, 4017–4023. [Google Scholar]
- Kawaguchi, K.; Igarashi, K.; Li, S.; Han, Q.; Tan, Y.; Kiyuna, T.; Miyake, K.; Murakami, T.; Chmielowski, B.; Nelson, S.D.; et al. Combination Treatment with Recombinant Methioninase Enables Temozolomide to Arrest a BRAF V600E Melanoma in a Patientderived Orthotopic Xenograft (PDOX) Mouse Model. Oncotarget 2017, 8, 85516–85525. [Google Scholar] [CrossRef]
- Igarashi, K.; Kawaguchi, K.; Li, S.; Han, Q.; Tan, Y.; Murakami, T.; Kiyuna, T.; Miyake, K.; Miyake, M.; Singh, A.S.; et al. Recombinant Methioninase in Combination with Doxorubicin (DOX) Overcomes First-Line DOX Resistance in a Patient-Derived Orthotopic Xenograft Nude-Mouse Model of Undifferentiated Spindle-Cell Sarcoma. Cancer Lett. 2018, 417, 168–173. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, J.; Yoshioka, T.; Li, B.; Lu, Q.; Li, S.; Sun, X.; Tan, Y.; Yagi, S.; Frenkel, E.P.; et al. Pharmacokinetics, Methionine Depletion, and Antigenicity of Recombinant Methioninase in Primates. Clin. Cancer Res. 2004, 10, 2131–2138. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, J.; Lu, Q.; Xu, J.; Kobayashi, Y.; Takakura, T.; Takimoto, A.; Yoshioka, T.; Lian, C.; Chen, C.; et al. PEGylation Confers Greatly Extended Half-Life and Attenuated Immunogenicity to Recombinant Methioninase in Primates. Cancer Res. 2004, 64, 6673–6678. [Google Scholar] [CrossRef]
- Higuchi, T.; Kawaguchi, K.; Miyake, K.; Han, Q.; Tan, Y.; Oshiro, H.; Sugisawa, N.; Zhang, Z.; Razmjooei, S.; Yamamoto, N.; et al. Oral Recombinant Methioninase Combined with Caffeine and Doxorubicin Induced Regression of a Doxorubicin-Resistant Synovial Sarcoma in a PDOX Mouse Model. Anticancer Res. 2018, 38, 5639–5644. [Google Scholar] [CrossRef]
- Kawaguchi, K.; Miyake, K.; Han, Q.; Li, S.; Tan, Y.; Igarashi, K.; Kiyuna, T.; Miyake, M.; Higuchi, T.; Oshiro, H.; et al. Oral Recombinant Methioninase (o-RMETase) Is Superior to Injectable RMETase and Overcomes Acquired Gemcitabine Resistance in Pancreatic Cancer. Cancer Lett. 2018, 432, 251–259. [Google Scholar] [CrossRef]
- Higuchi, T.; Sugisawa, N.; Yamamoto, J.; Oshiro, H.; Han, Q.; Yamamoto, N.; Hayashi, K.; Kimura, H.; Miwa, S.; Igarashi, K.; et al. The Combination of Oral-Recombinant Methioninase and Azacitidine Arrests a Chemotherapy-Resistant Osteosarcoma Patient-Derived Orthotopic Xenograft Mouse Model. Cancer Chemother. Pharmacol. 2020, 85, 285–291. [Google Scholar] [CrossRef]
- Yamamoto, J.; Miyake, K.; Han, Q.; Tan, Y.; Inubushi, S.; Sugisawa, N.; Higuchi, T.; Tashiro, Y.; Nishino, H.; Homma, Y.; et al. Oral Recombinant Methioninase Increases TRAIL Receptor-2 Expression to Regress Pancreatic Cancer in Combination with Agonist Tigatuzumab in an Orthotopic Mouse Model. Cancer Lett. 2020, 492, 174–184. [Google Scholar] [CrossRef]
- Igarashi, K.; Kawaguchi, K.; Kiyuna, T.; Miyake, K.; Miyaki, M.; Yamamoto, N.; Hayashi, K.; Kimura, H.; Miwa, S.; Higuchi, T.; et al. Metabolic Targeting with Recombinant Methioninase Combined with Palbociclib Regresses a Doxorubicin-Resistant Dedifferentiated Liposarcoma. Biochem. Biophys. Res. Commun. 2018, 506, 912–917. [Google Scholar] [CrossRef] [PubMed]
- Sugisawa, N.; Hamada, K.; Qinghong, H.A.N.; Yamamoto, J.; Yu, S.U.N.; Nishino, H.; Kawaguchi, K.; Bouvet, M.; Unno, M.; Hoffman, R.M. Adjuvant Oral Recombinant Methioninase Inhibits Lung Metastasis in a Surgical Breast-Cancer Orthotopic Syngeneic Model. Anticancer Res. 2020, 40, 4869–4874. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, T.; Han, Q.; Miyake, K.; Oshiro, H.; Sugisawa, N.; Tan, Y.; Yamamoto, N.; Hayashi, K.; Kimura, H.; Miwa, S.; et al. Combination of Oral Recombinant Methioninase and Decitabine Arrests a Chemotherapy-Resistant Undifferentiated Soft-Tissue Sarcoma Patient-Derived Orthotopic Xenograft Mouse Model. Biochem. Biophys. Res. Commun. 2020, 523, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Aoki, Y.; Tome, Y.; Qinghong, H.A.N.; Yamamoto, J.; Hamada, K.; Masaki, N.; Kubota, Y.; Bouvet, M.; Nishida, K.; Hoffman, R.M. Oral-Recombinant Methioninase Converts an Osteosarcoma from Methotrexate-Resistant to -Sensitive in a Patient-Derived Orthotopic-Xenograft (PDOX) Mouse Model. Anticancer Res. 2022, 42, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Stone, E.; Paley, O.; Hu, J.; Ekerdt, B.; Cheung, N.K.; Georgiou, G. De Novo Engineering of a Human Cystathionine-γ-Lyase for Systemic l-Methionine Depletion Cancer Therapy. ACS Chem. Biol. 2012, 7, 1822–1829. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.C.; Saha, A.; Yan, W.; Garrison, K.; Lamb, C.; Pandey, R.; Irani, S.; Lodi, A.; Lu, X.; Tiziani, S.; et al. Enzyme-Mediated Depletion of Serum L-Met Abrogates Prostate Cancer Growth via Multiple Mechanisms without Evidence of Systemic Toxicity. Proc. Natl. Acad. Sci. U. S. A. 2020, 117, 13000–13011. [Google Scholar] [CrossRef]
- Tan, Y.; Zavala, J.; Mingxu, X.U.; Zavala, J.; Hoffman, R.M. Serum Methionine Depletion without Side Effects by Methioninase in Metastatic Breast Cancer Patients. Anticancer Res. 1996, 16, 3937–3942. [Google Scholar]
- Tan, Y.; Zavala, J.; Han, Q.; Xu, M.; Sun, X.; Tan, X.; Tan, X.; Magana, R.; Geller, J.; Hoffman, R.M. Recombinant Methioninase Infusion Reduces the Biochemical Endpoint of Serum Methionine with Minimal Toxicity in High-Stage Cancer Patients. Anticancer Res. 1997, 17, 3857–3860. [Google Scholar]
- Kubota, Y.; Han, Q.; Hamada, K.; Aoki, Y.; Masaki, N.; Obara, K.; Tsunoda, T.; Hoffman, R.M. Long-Term Stable Disease in a Rectal-Cancer Patient Treated by Methionine Restriction With Oral Recombinant Methioninase and a Low-Methionine Diet. Anticancer Res. 2022, 42, 3857–3861. [Google Scholar] [CrossRef]
- Kubota, Y.; Han, Q.; Hozumi, C.; Masaki, N.; Yamamoto, J.; Aoki, Y.; Tsunoda, T.; Hoffman, R.M. Stage IV Pancreatic Cancer Patient Treated With FOLFIRINOX Combined With Oral Methioninase: A Highly-Rare Case With Long-Term Stable Disease. Anticancer Res. 2022, 42, 2567–2572. [Google Scholar] [CrossRef]
- Combs, J.A.; DeNicola, G.M. The Non-Essential Amino Acid Cysteine Becomes Essential for Tumor Proliferation and Survival. Cancers (Basel). 2019, 11, 678. [Google Scholar] [CrossRef]
- Zhang, H.-F.; Klein Geltink, R.I.; Parker, S.J.; Sorensen, P.H. Transsulfuration, Minor Player or Crucial for Cysteine Homeostasis in Cancer. Trends Cell Biol. 2022, 32, 800–814. [Google Scholar] [CrossRef]
- Poltorack, C.D.; Dixon, S.J. Understanding the Role of Cysteine in Ferroptosis: Progress & Paradoxes. FEBS J. 2022, 289, 374–385. [Google Scholar] [CrossRef]
- Harris, I.S.; Treloar, A.E.; Inoue, S.; Sasaki, M.; Gorrini, C.; Lee, K.C.; Yung, K.Y.; Brenner, D.; Knobbe-Thomsen, C.B.; Cox, M.A.; et al. Glutathione and Thioredoxin Antioxidant Pathways Synergize to Drive Cancer Initiation and Progression. Cancer Cell 2015, 27, 211–222. [Google Scholar] [CrossRef]
- HumanCyc HumanCyc: Encyclopedia of Human Genes and Metabolism. Available online: https://humancyc.org/ (accessed on 29 May 2023).
- Zhang, T.; Bauer, C.; Newman, A.C.; Uribe, A.H.; Athineos, D.; Blyth, K.; Maddocks, O.D.K. Polyamine Pathway Activity Promotes Cysteine Essentiality in Cancer Cells. Nat. Metab. 2020, 2, 1062–1076. [Google Scholar] [CrossRef]
- Wu, J.; Yeung, S.-C.J.; Liu, S.; Qdaisat, A.; Jiang, D.; Liu, W.; Cheng, Z.; Liu, W.; Wang, H.; Li, L.; et al. Cyst(e)Ine in Nutrition Formulation Promotes Colon Cancer Growth and Chemoresistance by Activating MTORC1 and Scavenging ROS. Signal Transduct. Target. Ther. 2021, 6, 188. [Google Scholar] [CrossRef]
- Ruiz-Rodado, V.; Dowdy, T.; Lita, A.; Kramp, T.; Zhang, M.; Jung, J.; Dios-Esponera, A.; Zhang, L.; Herold-Mende, C.C.; Camphausen, K.; et al. Cysteine Is a Limiting Factor for Glioma Proliferation and Survival. Mol. Oncol. 2022, 16, 1777–1794. [Google Scholar] [CrossRef]
- Srivastava, M.K.; Sinha, P.; Clements, V.K.; Rodriguez, P.; Ostrand-Rosenberg, S. Myeloid-Derived Suppressor Cells Inhibit T Cell Activation by Depleting Cystine and Cysteine. Cancer Res. 2010, 70, 68. [Google Scholar] [CrossRef]
- Bonifácio, V.D.B.; Pereira, S.A.; Serpa, J.; Vicente, J.B. Cysteine Metabolic Circuitries: Druggable Targets in Cancer. Br. J. Cancer 2021, 124, 862–879. [Google Scholar] [CrossRef]
- Hatae, R.; Chamoto, K.; Kim, Y.H.; Sonomura, K.; Taneishi, K.; Kawaguchi, S.; Yoshida, H.; Ozasa, H.; Sakamori, Y.; Akrami, M.; et al. Combination of Host Immune Metabolic Biomarkers for the PD-1 Blockade Cancer Immunotherapy. JCI Insight 2020, 5. [Google Scholar] [CrossRef]
- Wang, W.; Green, M.; Choi, J.E.; Gijón, M.; Kennedy, P.D.; Johnson, J.K.; Liao, P.; Lang, X.; Kryczek, I.; Sell, A.; et al. CD8+ T Cells Regulate Tumour Ferroptosis during Cancer Immunotherapy. Nature 2019, 569, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Cramer, S.L.; Saha, A.; Liu, J.; Tadi, S.; Tiziani, S.; Yan, W.; Triplett, K.; Lamb, C.; Alters, S.E.; Rowlinson, S.; et al. Systemic Depletion of L-Cyst(e)Ine with Cyst(e)Inase Increases Reactive Oxygen Species and Suppresses Tumor Growth. Nat. Med. 2017, 23, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Kshattry, S.; Saha, A.; Gries, P.; Tiziani, S.; Stone, E.; Georgiou, G.; DiGiovanni, J. Enzyme-Mediated Depletion of l-Cyst(e)Ine Synergizes with Thioredoxin Reductase Inhibition for Suppression of Pancreatic Tumor Growth. npj Precis. Oncol. 2019, 3. [Google Scholar] [CrossRef] [PubMed]
- Saha, A.; Zhao, S.; Chen, Z.; Georgiou, G.; Stone, E.; Kidane, D.; DiGiovanni, J. Combinatorial Approaches to Enhance DNA Damage Following Enzyme-Mediated Depletion of L-Cys for Treatment of Pancreatic Cancer. Mol. Ther. 2021, 29, 775–787. [Google Scholar] [CrossRef] [PubMed]
- Badgley, M.A.; Kremer, D.M.; Maurer, H.C.; DelGiorno, K.E.; Lee, H.-J.; Purohit, V.; Sagalovskiy, I.R.; Ma, A.; Kapilian, J.; Firl, C.E.M.; et al. Cysteine Depletion Induces Pancreatic Tumor Ferroptosis in Mice. Science 2020, 368, 85–89. [Google Scholar] [CrossRef]
- Poursaitidis, I.; Wang, X.; Crighton, T.; Labuschagne, C.; Mason, D.; Cramer, S.L.; Triplett, K.; Roy, R.; Pardo, O.E.; Seckl, M.J.; et al. Oncogene-Selective Sensitivity to Synchronous Cell Death Following Modulation of the Amino Acid Nutrient Cystine. Cell Rep. 2017, 18, 2547–2556. [Google Scholar] [CrossRef]
- Kerimoglu, B.; Lamb, C.; McPherson, R.D.; Ergen, E.; Stone, E.M.; Ooi, A. Cyst(e)Inase-Rapamycin Combination Induces Ferroptosis in Both In Vitro and In Vivo Models of Hereditary Leiomyomatosis and Renal Cell Cancer. Mol. Cancer Ther. 2022, 21, 419–426. [Google Scholar] [CrossRef]
- Yang, W.S.; Sriramaratnam, R.; Welsch, M.E.; Shimada, K.; Skouta, R.; Viswanathan, V.S.; Cheah, J.H.; Clemons, P.A.; Shamji, A.F.; Clish, C.B.; et al. Regulation of Ferroptotic Cancer Cell Death by GPX4. Cell 2014, 156, 317–331. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, X.; Jin, S.; Chen, Y.; Guo, R. Ferroptosis in Cancer Therapy: A Novel Approach to Reversing Drug Resistance. Mol. Cancer 2022, 21, 47. [Google Scholar] [CrossRef]
- Koppula, P.; Zhuang, L.; Gan, B. Cystine Transporter SLC7A11/XCT in Cancer: Ferroptosis, Nutrient Dependency, and Cancer Therapy. Protein Cell 2021, 12, 599–620. [Google Scholar] [CrossRef]
- Lanzardo, S.; Conti, L.; Rooke, R.; Ruiu, R.; Accart, N.; Bolli, E.; Arigoni, M.; Macagno, M.; Barrera, G.; Pizzimenti, S.; et al. Immunotargeting of Antigen XCT Attenuates Stem-like Cell Behavior and Metastatic Progression in Breast Cancer. Cancer Res. 2016, 76, 62–72. [Google Scholar] [CrossRef]
- Zheng, Z.; Luo, G.; Shi, X.; Long, Y.; Shen, W.; Li, Z.; Zhang, X. The Xc− Inhibitor Sulfasalazine Improves the Anti-Cancer Effect of Pharmacological Vitamin C in Prostate Cancer Cells via a Glutathione-Dependent Mechanism. Cell. Oncol. 2020, 43, 95–106. [Google Scholar] [CrossRef]
- Hu, K.; Li, K.; Lv, J.; Feng, J.; Chen, J.; Wu, H.; Cheng, F.; Jiang, W.; Wang, J.; Pei, H.; et al. Suppression of the SLC7A11/Glutathione Axis Causes Synthetic Lethality in KRAS-Mutant Lung Adenocarcinoma. J. Clin. Invest. 2020, 130, 1752–1766. [Google Scholar] [CrossRef]
- Li, Y.; Yan, H.; Xu, X.; Liu, H.; Wu, C.; Zhao, L. Erastin/Sorafenib Induces Cisplatin-Resistant Non-Small Cell Lung Cancer Cell Ferroptosis through Inhibition of the Nrf2/XCT Pathway. Oncol. Lett. 2020, 19, 323–333. [Google Scholar] [CrossRef]
- Miess, H.; Dankworth, B.; Gouw, A.M.; Rosenfeldt, M.; Schmitz, W.; Jiang, M.; Saunders, B.; Howell, M.; Downward, J.; Felsher, D.W.; et al. The Glutathione Redox System Is Essential to Prevent Ferroptosis Caused by Impaired Lipid Metabolism in Clear Cell Renal Cell Carcinoma. Oncogene 2018, 37, 5435–5450. [Google Scholar] [CrossRef]
- Kim, E.H.; Shin, D.; Lee, J.; Jung, A.R.; Roh, J.L. CISD2 Inhibition Overcomes Resistance to Sulfasalazine-Induced Ferroptotic Cell Death in Head and Neck Cancer. Cancer Lett. 2018, 432, 180–190. [Google Scholar] [CrossRef]
- Byun, J.K.; Lee, S.; Kang, G.W.; Lee, Y.R.; Park, S.Y.; Song, I.S.; Yun, J.W.; Lee, J.; Choi, Y.K.; Park, K.G. Macropinocytosis Is an Alternative Pathway of Cysteine Acquisition and Mitigates Sorafenib-Induced Ferroptosis in Hepatocellular Carcinoma. J. Exp. Clin. Cancer Res. 2022, 41, 98. [Google Scholar] [CrossRef]
- Feng, H.; Stockwell, B.R. Unsolved Mysteries: How Does Lipid Peroxidation Cause Ferroptosis? PLoS Biol. 2018, 16. [Google Scholar] [CrossRef]
- Zhang, Y.; Tan, H.; Daniels, J.D.; Zandkarimi, F.; Liu, H.; Brown, L.M.; Uchida, K.; O’Connor, O.A.; Stockwell, B.R. Imidazole Ketone Erastin Induces Ferroptosis and Slows Tumor Growth in a Mouse Lymphoma Model. Cell Chem. Biol. 2019, 26, 623–633. [Google Scholar] [CrossRef]
- Zhang, Y.; Swanda, R.V.; Nie, L.; Liu, X.; Wang, C.; Lee, H.; Lei, G.; Mao, C.; Koppula, P.; Cheng, W.; et al. MTORC1 Couples Cyst(e)Ine Availability with GPX4 Protein Synthesis and Ferroptosis Regulation. Nat. Commun. 2021, 12. [Google Scholar] [CrossRef]
- Alborzinia, H.; Flórez, A.F.; Kreth, S.; Brückner, L.M.; Yildiz, U.; Gartlgruber, M.; Odoni, D.I.; Poschet, G.; Garbowicz, K.; Shao, C.; et al. MYCN Mediates Cysteine Addiction and Sensitizes Neuroblastoma to Ferroptosis. Nat. Cancer 2022, 3, 471–485. [Google Scholar] [CrossRef]
- Robe, P.A.; Martin, D.H.; Nguyen-Khac, M.T.; Artesi, M.; Deprez, M.; Albert, A.; Vanbelle, S.; Califice, S.; Bredel, M.; Bours, V. Early Termination of ISRCTN45828668, a Phase 1/2 Prospective, Randomized Study of Sulfasalazine for the Treatment of Progressing Malignant Gliomas in Adults. BMC Cancer 2009, 9, 372. [Google Scholar] [CrossRef] [PubMed]
- Shitara, K.; Doi, T.; Nagano, O.; Imamura, C.K.; Ozeki, T.; Ishii, Y.; Tsuchihashi, K.; Takahashi, S.; Nakajima, T.E.; Hironaka, S.; et al. Dose-Escalation Study for the Targeting of CD44v+ Cancer Stem Cells by Sulfasalazine in Patients with Advanced Gastric Cancer (EPOC1205). Gastric Cancer 2017, 20, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Shitara, K.; Doi, T.; Nagano, O.; Fukutani, M.; Hasegawa, H.; Nomura, S.; Sato, A.; Kuwata, T.; Asai, K.; Einaga, Y.; et al. Phase 1 Study of Sulfasalazine and Cisplatin for Patients with CD44v-Positive Gastric Cancer Refractory to Cisplatin (EPOC1407). Gastric Cancer 2017, 20, 1004–1009. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, S.; Wada, K.; Nagatani, K.; Otani, N.; Osada, H.; Nawashiro, H. Sulfasalazine and Temozolomide with Radiation Therapy for Newly Diagnosed Glioblastoma. Neurol. India 2014, 62, 42. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Vousden, K.H. Serine and One-Carbon Metabolism in Cancer. Nat. Rev. Cancer 2016, 16, 650–662. [Google Scholar] [CrossRef] [PubMed]
- Possemato, R.; Marks, K.M.; Shaul, Y.D.; Pacold, M.E.; Kim, D.; Birsoy, K.K.; Sethumadhavan, S.; Woo, H.-K.K.; Jang, H.G.; Jha, A.K.; et al. Functional Genomics Reveal That the Serine Synthesis Pathway Is Essential in Breast Cancer. Nature 2011, 476, 346–350. [Google Scholar] [CrossRef] [PubMed]
- Labuschagne, C.F.; van den Broek, N.J.F.; Mackay, G.M.; Vousden, K.H.; Maddocks, O.D.K. Serine, but Not Glycine, Supports One-Carbon Metabolism and Proliferation of Cancer Cells. Cell Rep. 2014, 7, 1248–1258. [Google Scholar] [CrossRef] [PubMed]
- Maddocks, O.D.K.K.; Athineos, D.; Cheung, E.C.; Lee, P.; Zhang, T.; Van Den Broek, N.J.F.F.; Mackay, G.M.; Labuschagne, C.F.; Gay, D.; Kruiswijk, F.; et al. Modulating the Therapeutic Response of Tumours to Dietary Serine and Glycine Starvation. Nature 2017, 544, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Gravel, S.-P.; Hulea, L.; Toban, N.; Birman, E.; Blouin, M.-J.; Zakikhani, M.; Zhao, Y.; Topisirovic, I.; St-Pierre, J.; Pollak, M. Serine Deprivation Enhances Antineoplastic Activity of Biguanides. Cancer Res. 2014, 74, 7521–7533. [Google Scholar] [CrossRef]
- Van Nyen, T.; Planque, M.; van Wagensveld, L.; Duarte, J.A.G.; Zaal, E.A.; Talebi, A.; Rossi, M.; Körner, P.R.; Rizzotto, L.; Moens, S.; et al. Serine Metabolism Remodeling after Platinum-Based Chemotherapy Identifies Vulnerabilities in a Subgroup of Resistant Ovarian Cancers. Nat. Commun. 2022, 13, 25. [Google Scholar] [CrossRef]
- Sullivan, M.R.; Mattaini, K.R.; Dennstedt, E.A.; Nguyen, A.A.; Sivanand, S.; Reilly, M.F.; Meeth, K.; Muir, A.; Darnell, A.M.; Bosenberg, M.W.; et al. Increased Serine Synthesis Provides an Advantage for Tumors Arising in Tissues Where Serine Levels Are Limiting. Cell Metab. 2019, 29, 1410–1421.e4. [Google Scholar] [CrossRef]
- Muthusamy, T.; Cordes, T.; Handzlik, M.K.; You, L.; Lim, E.W.; Gengatharan, J.; Pinto, A.F.M.; Badur, M.G.; Kolar, M.J.; Wallace, M.; et al. Serine Restriction Alters Sphingolipid Diversity to Constrain Tumour Growth. Nature 2020, 586, 790–795. [Google Scholar] [CrossRef]
- Fujihara, K.M.; Zhang, B.Z.; Jackson, T.D.; Ogunkola, M.O.; Nijagal, B.; Milne, J.V.; Sallman, D.A.; Ang, C.-S.; Nikolic, I.; Kearney, C.J.; et al. Eprenetapopt Triggers Ferroptosis, Inhibits NFS1 Cysteine Desulfurase, and Synergizes with Serine and Glycine Dietary Restriction. Sci. Adv. 2022, 8, eabm9427. [Google Scholar] [CrossRef] [PubMed]
- Maddocks, O.D.K.K.; Berkers, C.R.; Mason, S.M.; Zheng, L.; Blyth, K.; Gottlieb, E.; Vousden, K.H. Serine Starvation Induces Stress and P53-Dependent Metabolic Remodelling in Cancer Cells. Nature 2013, 493, 542–546. [Google Scholar] [CrossRef] [PubMed]
- Humpton, T.J.; Hock, A.K.; Maddocks, O.D.K.; Vousden, K.H. P53-Mediated Adaptation to Serine Starvation Is Retained by a Common Tumour-Derived Mutant. Cancer Metab. 2018, 6, 18. [Google Scholar] [CrossRef] [PubMed]
- LeBoeuf, S.E.; Wu, W.L.; Karakousi, T.R.; Karadal, B.; Jackson, S.R.E.; Davidson, S.M.; Wong, K.K.; Koralov, S.B.; Sayin, V.I.; Papagiannakopoulos, T. Activation of Oxidative Stress Response in Cancer Generates a Druggable Dependency on Exogenous Non-Essential Amino Acids. Cell Metab. 2020, 31, 339–350.e4. [Google Scholar] [CrossRef] [PubMed]
- Tajan, M.; Hennequart, M.; Cheung, E.C.; Zani, F.; Hock, A.K.; Legrave, N.; Maddocks, O.D.K.; Ridgway, R.A.; Athineos, D.; Suárez-Bonnet, A.; et al. Serine Synthesis Pathway Inhibition Cooperates with Dietary Serine and Glycine Limitation for Cancer Therapy. Nat. Commun. 2021, 12. [Google Scholar] [CrossRef]
- Falcone, M.; Uribe, A.H.; Papalazarou, V.; Newman, A.C.; Athineos, D.; Stevenson, K.; Sauvé, C.E.G.; Gao, Y.; Kim, J.K.; Del Latto, M.; et al. Sensitisation of Cancer Cells to Radiotherapy by Serine and Glycine Starvation. Br. J. Cancer 2022, 1–14. [Google Scholar] [CrossRef]
- Pranzini, E.; Pardella, E.; Muccillo, L.; Leo, A.; Nesi, I.; Santi, A.; Parri, M.; Zhang, T.; Uribe, A.H.; Lottini, T.; et al. SHMT2-Mediated Mitochondrial Serine Metabolism Drives 5-FU Resistance by Fueling Nucleotide Biosynthesis. Cell Rep. 2022, 40, 111233. [Google Scholar] [CrossRef]
- Polet, F.; Corbet, C.; Pinto, A.; Rubio, L.I.; Martherus, R.; Bol, V.; Drozak, X.; Grégoire, V.; Riant, O.; Feron, O. Reducing the Serine Availability Complements the Inhibition of the Glutamine Metabolism to Block Leukemia Cell Growth. Oncotarget 2016, 7, 1765–1776. [Google Scholar] [CrossRef]
- Méndez-Lucas, A.; Lin, W.; Driscoll, P.C.; Legrave, N.; Novellasdemunt, L.; Xie, C.; Charles, M.; Wilson, Z.; Jones, N.P.; Rayport, S.; et al. Identifying Strategies to Target the Metabolic Flexibility of Tumours. Nat. Metab. 2020, 2, 335–350. [Google Scholar] [CrossRef]
- Hamanaka, R.B.; Nigdelioglu, R.; Meliton, A.Y.; Tian, Y.; Witt, L.J.; O’Leary, E.; Sun, K.A.; Woods, P.S.; Wu, D.; Ansbro, B.; et al. Inhibition of Phosphoglycerate Dehydrogenase Attenuates Bleomycin-Induced Pulmonary Fibrosis. Am. J. Respir. Cell Mol. Biol. 2018, 58, 585–593. [Google Scholar] [CrossRef]
- Pacold, M.E.; Brimacombe, K.R.; Chan, S.H.; Rohde, J.M.; Lewis, C.A.; Swier, L.J.Y.M.; Possemato, R.; Chen, W.W.; Sullivan, L.B.; Fiske, B.P.; et al. A PHGDH Inhibitor Reveals Coordination of Serine Synthesis and One-Carbon Unit Fate. Nat. Chem. Biol. 2016, 12, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Mullarky, E.; Lucki, N.C.; Zavareh, R.B.; Anglin, J.L.; Gomes, A.P.; Nicolay, B.N.; Wong, J.C.Y.; Christen, S.; Takahashi, H.; Singh, P.K.; et al. Identification of a Small Molecule Inhibitor of 3-Phosphoglycerate Dehydrogenase to Target Serine Biosynthesis in Cancers. Proc. Natl. Acad. Sci. U. S. A. 2016, 113, 1778–1783. [Google Scholar] [CrossRef] [PubMed]
- Ngo, B.; Kim, E.; Osorio-Vasquez, V.; Doll, S.; Bustraan, S.; Liang, R.J.; Luengo, A.; Davidson, S.M.; Ali, A.; Ferraro, G.B.; et al. Limited Environmental Serine and Glycine Confer Brain Metastasis Sensitivity to PHGDH Inhibition. Cancer Discov. 2020, 10, 1352–1373. [Google Scholar] [CrossRef]
- Ducker, G.S.; Ghergurovich, J.M.; Mainolfi, N.; Suri, V.; Jeong, S.K.; Li, S.H.J.; Friedman, A.; Manfredi, M.G.; Gitai, Z.; Kim, H.; et al. Human SHMT Inhibitors Reveal Defective Glycine Import as a Targetable Metabolic Vulnerability of Diffuse Large B-Cell Lymphoma. Proc. Natl. Acad. Sci. U. S. A. 2017, 114, 11404–11409. [Google Scholar] [CrossRef]
- García-Cañaveras, J.C.; Lancho, O.; Ducker, G.S.; Ghergurovich, J.M.; Xu, X.; da Silva-Diz, V.; Minuzzo, S.; Indraccolo, S.; Kim, H.; Herranz, D.; et al. SHMT Inhibition Is Effective and Synergizes with Methotrexate in T-Cell Acute Lymphoblastic Leukemia. Leukemia 2021, 35, 377–388. [Google Scholar] [CrossRef]
- Ma, E.H.; Bantug, G.; Griss, T.; Condotta, S.; Johnson, R.M.; Samborska, B.; Mainolfi, N.; Suri, V.; Guak, H.; Balmer, M.L.; et al. Serine Is an Essential Metabolite for Effector T Cell Expansion. Cell Metab. 2017, 25, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Wu, G. Roles of Dietary Glycine, Proline, and Hydroxyproline in Collagen Synthesis and Animal Growth. Amino Acids 2018, 50, 29–38. [Google Scholar] [CrossRef]
- Jain, M.; Nilsson, R.; Sharma, S.; Madhusudhan, N.; Kitami, T.; Souza, A.L.; Kafri, R.; Kirschner, M.W.; Clish, C.B.; Mootha, V.K. Metabolite Profiling Identifies a Key Role for Glycine in Rapid Cancer Cell Proliferation. Science 2012, 336, 1040–1044. [Google Scholar] [CrossRef]
- Chen, C.-L.; Hsu, S.-C.; Ann, D.K.; Yen, Y.; Kung, H.-J. Arginine Signaling and Cancer Metabolism. Cancers (Basel). 2021, 13, 3541. [Google Scholar] [CrossRef]
- Gonzalez, G.G.; Byus, C.V. Effect of Dietary Arginine Restriction upon Ornithine and Polyamine Metabolism during Two-Stage Epidermal Carcinogenesis in the Mouse. Cancer Res. 1991, 51, 2932–2939. [Google Scholar] [PubMed]
- Yeatman, T.J.; Risley, G.L.; Brunson, M.E. Depletion of Dietary Arginine Inhibits Growth of Metastatic Tumor. Arch. Surg. 1991, 126, 1376–1382. [Google Scholar] [CrossRef]
- Cheng, C.-T.T.; Qi, Y.; Wang, Y.-C.C.; Chi, K.K.; Chung, Y.; Ouyang, C.; Chen, Y.-R.Y.-H.Y.H.R.; Oh, M.E.; Sheng, X.; Tang, Y.; et al. Arginine Starvation Kills Tumor Cells through Aspartate Exhaustion and Mitochondrial Dysfunction. Commun. Biol. 2018, 1, 178. [Google Scholar] [CrossRef]
- Alexandrou, C.; Al-Aqbi, S.S.; Higgins, J.A.; Boyle, W.; Karmokar, A.; Andreadi, C.; Luo, J.-L.; Moore, D.A.; Viskaduraki, M.; Blades, M.; et al. Sensitivity of Colorectal Cancer to Arginine Deprivation Therapy Is Shaped by Differential Expression of Urea Cycle Enzymes. Sci. Rep. 2018, 8, 12096. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.C.; Chen, C.L.; Cheng, M.L.; Chu, C.Y.; Changou, C.A.; Yu, Y.L.; Yeh, S. Der; Kuo, T.C.; Kuo, C.C.; Chuu, C.P.; et al. Arginine Starvation Elicits Chromatin Leakage and CGAS-STING Activation via Epigenetic Silencing of Metabolic and DNA-Repair Genes. Theranostics 2021, 11, 7527–7545. [Google Scholar] [CrossRef] [PubMed]
- Missiaen, R.; Anderson, N.M.; Kim, L.C.; Nance, B.; Burrows, M.; Skuli, N.; Carens, M.; Riscal, R.; Steensels, A.; Li, F.; et al. GCN2 Inhibition Sensitizes Arginine-Deprived Hepatocellular Carcinoma Cells to Senolytic Treatment. Cell Metab. 2022, 34, 1151–1167.e7. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Feng, Y.; Zhang, Y.; Zhu, X.; Jin, F. L-Arginine Supplementation Inhibits the Growth of Breast Cancer by Enhancing Innate and Adaptive Immune Responses Mediated by Suppression of MDSCs in Vivo. BMC Cancer 2016, 16. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, Q.; Du, Y.; Liu, F.; Zhang, Y.; Feng, Y.; Jin, F. L-Arginine and Docetaxel Synergistically Enhance Anti-Tumor Immunity by Modifying the Immune Status of Tumor-Bearing Mice. Int. Immunopharmacol. 2016, 35, 7–14. [Google Scholar] [CrossRef]
- Geiger, R.; Rieckmann, J.C.; Wolf, T.; Basso, C.; Feng, Y.; Fuhrer, T.; Kogadeeva, M.; Picotti, P.; Meissner, F.; Mann, M.; et al. L-Arginine Modulates T Cell Metabolism and Enhances Survival and Anti-Tumor Activity. Cell 2016, 167, 829–842.e13. [Google Scholar] [CrossRef]
- Satoh, Y.; Kotani, H.; Iida, Y.; Taniura, T.; Notsu, Y.; Harada, M. Supplementation of L-Arginine Boosts the Therapeutic Efficacy of Anticancer Chemoimmunotherapy. Cancer Sci. 2020, 111, 2248–2258. [Google Scholar] [CrossRef]
- He, X.; Lin, H.; Yuan, L.; Li, B. Combination Therapy with L-Arginine and α-PD-L1 Antibody Boosts Immune Response against Osteosarcoma in Immunocompetent Mice. Cancer Biol. Ther. 2017, 18, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Castillo, L.; Chapman, T.E.; Sanchez, M.; Yu, Y.M.; Burke, J.F.; Ajami, A.M.; Vogt, J.; Young, V.R. Plasma Arginine and Citrulline Kinetics in Adults given Adequate and Arginine-Free Diets. Proc. Natl. Acad. Sci. U. S. A. 1993, 90, 7749–7753. [Google Scholar] [CrossRef] [PubMed]
- Tharakan, J.F.; Yu, Y.M.; Zurakowski, D.; Roth, R.M.; Young, V.R.; Castillo, L. Adaptation to a Long Term (4 Weeks) Arginine- and Precursor (Glutamate, Proline and Aspartate)-Free Diet. Clin. Nutr. 2008, 27, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Rabinovich, S.; Adler, L.; Yizhak, K.; Sarver, A.; Silberman, A.; Agron, S.; Stettner, N.; Sun, Q.; Brandis, A.; Helbling, D.; et al. Diversion of Aspartate in ASS1-Deficient Tumours Fosters de Novo Pyrimidine Synthesis. Nature 2015, 527, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Silberman, A.; Goldman, O.; Assayag, O.B.; Jacob, A.; Rabinovich, S.; Adler, L.; Lee, J.S.; Keshet, R.; Sarver, A.; Frug, J.; et al. Acid-Induced Downregulation of ASS1 Contributes to the Maintenance of Intracellular PH in Cancer. Cancer Res. 2019, 79, 518–533. [Google Scholar] [CrossRef]
- Agrawal, V.; Woo, J.H.; Mauldin, J.P.; Jo, C.; Stone, E.M.; Georgiou, G.; Frankel, A.E. Cytotoxicity of Human Recombinant Arginase I (Co)-PEG5000 in the Presence of Supplemental L-Citrulline Is Dependent on Decreased Argininosuccinate Synthetase Expression in Human Cells. Anticancer. Drugs 2012, 23, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Bowles, T.L.; Kim, R.; Galante, J.; Parsons, C.M.; Virudachalam, S.; Kung, H.J.; Bold, R.J. Pancreatic Cancer Cell Lines Deficient in Argininosuccinate Synthetase Are Sensitive to Arginine Deprivation by Arginine Deiminase. Int. J. Cancer 2008, 123, 1950–1955. [Google Scholar] [CrossRef]
- Hajji, N.; Garcia-Revilla, J.; Soto, M.S.; Perryman, R.; Symington, J.; Quarles, C.C.; Healey, D.R.; Guo, Y.; Orta-Vázquez, M.L.; Mateos-Cordero, S.; et al. Arginine Deprivation Alters Microglial Polarity and Synergizes with Radiation to Eradicate Non-Arginine-Auxotrophic Glioblastoma Tumors. J. Clin. Invest. 2022, 132. [Google Scholar] [CrossRef]
- Clinical Trials Using PEG-BCT-100 in Cancer | List Results - ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/results?cond=Cancer&term=PEG-BCT-100&cntry=&state=&city=&dist= (accessed on 29 May 2023).
- Yau, T.; Cheng, P.N.; Chan, P.; Chan, W.; Chen, L.; Yuen, J.; Pang, R.; Fan, S.T.; Poon, R.T. A Phase 1 Dose-Escalating Study of Pegylated Recombinant Human Arginase 1 (Peg-RhArg1) in Patients with Advanced Hepatocellular Carcinoma. Invest. New Drugs 2013, 31, 99–107. [Google Scholar] [CrossRef]
- Yau, T.; Cheng, P.N.; Chan, P.; Chen, L.; Yuen, J.; Pang, R.; Fan, S.T.; Wheatley, D.N.; Poon, R.T. Preliminary Efficacy, Safety, Pharmacokinetics, Pharmacodynamics and Quality of Life Study of Pegylated Recombinant Human Arginase 1 in Patients with Advanced Hepatocellular Carcinoma. Invest. New Drugs 2015, 33, 496–504. [Google Scholar] [CrossRef]
- Chan, S.L.; Cheng, P.N.M.; Liu, A.M.; Chan, L.L.; Li, L.; Chu, C.M.; Chong, C.C.N.; Lau, Y.M.; Yeo, W.; Ng, K.K.C.; et al. A Phase II Clinical Study on the Efficacy and Predictive Biomarker of Pegylated Recombinant Arginase on Hepatocellular Carcinoma. Invest. New Drugs 2021, 39, 1375–1382. [Google Scholar] [CrossRef]
- Yau, T.; Cheng, P.N.M.; Chiu, J.; Kwok, G.G.W.; Leung, R.; Liu, A.M.; Cheung, T.T.; Ng, C.T. A Phase 1 Study of Pegylated Recombinant Arginase (PEG-BCT-100) in Combination with Systemic Chemotherapy (Capecitabine and Oxaliplatin)[PACOX] in Advanced Hepatocellular Carcinoma Patients. Invest. New Drugs 2022, 40, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.N.M.; Liu, A.M.; Bessudo, A.; Mussai, F. Safety, PK/PD and Preliminary Anti-Tumor Activities of Pegylated Recombinant Human Arginase 1 (BCT-100) in Patients with Advanced Arginine Auxotrophic Tumors. Invest. New Drugs 2021, 39, 1633–1640. [Google Scholar] [CrossRef] [PubMed]
- De Santo, C.; Cheng, P.; Beggs, A.; Egan, S.; Bessudo, A.; Mussai, F. Metabolic Therapy with PEG-Arginase Induces a Sustained Complete Remission in Immunotherapy-Resistant Melanoma. J. Hematol. Oncol. 2018, 11, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Clinical Trials Using ADI-PEG20 in Cancer | List Results - ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/results?cond=Cancer&term=ADI-PEG20&cntry=&state=&city=&dist= (accessed on 29 May 2023).
- Delman, K.A.; Brown, T.D.; Thomas, M.; Ensor, C.M.; Holtsberg, F.W.; Bomalaski, J.S.; Clark, M.A.; Curley, S.A. Phase I/II Trial of Pegylated Arginine Deiminase (ADI-PEG20) in Unresectable Hepatocellular Carcinoma. J. Clin. Oncol. 2005, 23, 4139–4139. [Google Scholar] [CrossRef]
- Feun, L.G.; You, M.; Wu, C.; Wangpaichitr, M.; Kuo, M.T.; Marini, A.; Jungbluth, A.; Savaraj, N. Final Results of Phase II Trial of Pegylated Arginine Deiminase (ADI-PEG20) in Metastatic Melanoma (MM). J. Clin. Oncol. 2010, 28, 8528–8528. [Google Scholar] [CrossRef]
- Glazer, E.S.; Piccirillo, M.; Albino, V.; Di Giacomo, R.; Palaia, R.; Mastro, A.A.; Beneduce, G.; Castello, G.; De Rosa, V.; Petrillo, A.; et al. Phase II Study of Pegylated Arginine Deiminase for Nonresectable and Metastatic Hepatocellular Carcinoma. J. Clin. Oncol. 2010, 28, 2220–2226. [Google Scholar] [CrossRef]
- Feun, L.G.; Marini, A.; Walker, G.; Elgart, G.; Moffat, F.; Rodgers, S.E.; Wu, C.J.; You, M.; Wangpaichitr, M.; Kuo, M.T.; et al. Negative Argininosuccinate Synthetase Expression in Melanoma Tumours May Predict Clinical Benefit from Arginine-Depleting Therapy with Pegylated Arginine Deiminase. Br. J. Cancer 2012, 106, 1481–1485. [Google Scholar] [CrossRef]
- Ott, P.A.; Carvajal, R.D.; Pandit-Taskar, N.; Jungbluth, A.A.; Hoffman, E.W.; Wu, B.-W.W.; Bomalaski, J.S.; Venhaus, R.; Pan, L.; Old, L.J.; et al. Phase I/II Study of Pegylated Arginine Deiminase (ADI-PEG 20) in Patients with Advanced Melanoma. Invest. New Drugs 2013, 31, 425–434. [Google Scholar] [CrossRef]
- Szlosarek, P.W.; Steele, J.P.; Nolan, L.; Gilligan, D.; Taylor, P.; Spicer, J.; Lind, M.; Mitra, S.; Shamash, J.; Phillips, M.M.; et al. Arginine Deprivation With Pegylated Arginine Deiminase in Patients With Argininosuccinate Synthetase 1-Deficient Malignant Pleural Mesothelioma: A Randomized Clinical Trial. JAMA Oncol. 2017, 3, 58–66. [Google Scholar] [CrossRef]
- Tsai, H.J.; Jiang, S.S.; Hung, W.C.; Borthakur, G.; Lin, S.F.; Pemmaraju, N.; Jabbour, E.; Bomalaski, J.S.; Chen, Y.P.; Hsiao, H.H.; et al. A Phase II Study of Arginine Deiminase (ADI-PEG20) in Relapsed/Refractory or Poor-Risk Acute Myeloid Leukemia Patients. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Abou-Alfa, G.K.; Qin, S.; Ryoo, B.Y.; Lu, S.N.; Yen, C.J.; Feng, Y.H.; Lim, H.Y.; Izzo, F.; Colombo, M.; Sarker, D.; et al. Phase III Randomized Study of Second Line ADI-PEG 20 plus Best Supportive Care versus Placebo plus Best Supportive Care in Patients with Advanced Hepatocellular Carcinoma. Ann. Oncol. 2018, 29, 1402–1408. [Google Scholar] [CrossRef] [PubMed]
- Hall, P.E.; Ready, N.; Johnston, A.; Bomalaski, J.S.; Venhaus, R.R.; Sheaff, M.; Krug, L.; Szlosarek, P.W. Phase II Study of Arginine Deprivation Therapy With Pegargiminase in Patients With Relapsed Sensitive or Refractory Small-Cell Lung Cancer. Clin. Lung Cancer 2020, 21, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.J.; Hsiao, H.H.; Hsu, Y.T.; Liu, Y.C.; Kao, H.W.; Liu, T.C.; Cho, S.F.; Feng, X.; Johnston, A.; Bomalaski, J.S.; et al. Phase I Study of ADI-PEG20 plus Low-Dose Cytarabine for the Treatment of Acute Myeloid Leukemia. Cancer Med. 2021, 10, 2946–2955. [Google Scholar] [CrossRef]
- Lowery, M.A.; Yu, K.H.; Kelsen, D.P.; Harding, J.J.; Bomalaski, J.S.; Glassman, D.C.; Covington, C.M.; Brenner, R.; Hollywood, E.; Barba, A.; et al. A Phase 1/1B Trial of ADI-PEG 20 plus Nab-Paclitaxel and Gemcitabine in Patients with Advanced Pancreatic Adenocarcinoma. Cancer 2017, 123, 4556–4565. [Google Scholar] [CrossRef]
- Harding, J.J.; Do, R.K.; Dika, I. El; Hollywood, E.; Uhlitskykh, K.; Valentino, E.; Wan, P.; Hamilton, C.; Feng, X.; Johnston, A.; et al. A Phase 1 Study of ADI-PEG 20 and Modified FOLFOX6 in Patients with Advanced Hepatocellular Carcinoma and Other Gastrointestinal Malignancies. Cancer Chemother. Pharmacol. 2018, 82, 429–440. [Google Scholar] [CrossRef]
- Hall, P.E.; Lewis, R.; Syed, N.; Shaffer, R.; Evanson, J.; Ellis, S.; Williams, M.; Feng, X.; Johnston, A.; Thomson, J.A.; et al. A Phase I Study of Pegylated Arginine Deiminase (Pegargiminase), Cisplatin, and Pemetrexed in Argininosuccinate Synthetase 1-Deficient Recurrent High-Grade Glioma. Clin. Cancer Res. 2019, 25, 2708–2716. [Google Scholar] [CrossRef]
- Yao, S.; Janku, F.; Subbiah, V.; Stewart, J.; Patel, S.P.; Kaseb, A.; Westin, S.N.; Naing, A.; Tsimberidou, A.M.; Hong, D.; et al. Phase 1 Trial of ADI-PEG20 plus Cisplatin in Patients with Pretreated Metastatic Melanoma or Other Advanced Solid Malignancies. Br. J. Cancer 2021, 124, 1533–1539. [Google Scholar] [CrossRef]
- Tomlinson, B.K.; Thomson, J.A.; Bomalaski, J.S.; Diaz, M.; Akande, T.; Mahaffey, N.; Li, T.; Dutia, M.P.; Kelly, K.; Gong, I.Y.; et al. Phase i Trial of Arginine Deprivation Therapy with ADI-PEG 20 plus Docetaxel in Patients with Advanced Malignant Solid Tumors. Clin. Cancer Res. 2015, 21, 2480–2486. [Google Scholar] [CrossRef]
- Bomalaski, J.S.; Chen, K.-T.; Chuang, M.-J.; Liau, C.-T.; Peng, M.-T.; Chen, P.-Y.; Lee, C.-C.; Johnston, A.; Liu, H.-F.; Huang, Y.-L.S.; et al. Phase IB Trial of Pegylated Arginine Deiminase (ADI-PEG 20) plus Radiotherapy and Temozolomide in Patients with Newly Diagnosed Glioblastoma. J. Clin. Oncol. 2022, 40, 2057–2057. [Google Scholar] [CrossRef]
- Rath, M.; Müller, I.; Kropf, P.; Closs, E.I.; Munder, M. Metabolism via Arginase or Nitric Oxide Synthase: Two Competing Arginine Pathways in Macrophages. Front. Immunol. 2014, 5, 532. [Google Scholar] [CrossRef]
- Fletcher, M.; Ramirez, M.E.; Sierra, R.A.; Raber, P.; Thevenot, P.; Al-Khami, A.A.; Sanchez-Pino, D.; Hernandez, C.; Wyczechowska, D.D.; Ochoa, A.C.; et al. L-Arginine Depletion Blunts Antitumor T-Cell Responses by Inducing Myeloid-Derived Suppressor Cells. Cancer Res. 2015, 75, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Baggio, R.; Elbaum, D.; Kanyo, Z.F.; Carroll, P.J.; Cavalli, R.C.; Ash, D.E.; Christianson, D.W. Inhibition of Mn2+2-Arginase by Borate Leads to the Design of a Transition State Analogue Inhibitor, 2(S)-Amino-6-Boronohexanoic Acid. J. Am. Chem. Soc. 1997, 119, 8107–8108. [Google Scholar] [CrossRef]
- Steggerda, S.M.; Bennett, M.K.; Chen, J.; Emberley, E.; Huang, T.; Janes, J.R.; Li, W.; MacKinnon, A.L.; Makkouk, A.; Marguier, G.; et al. Inhibition of Arginase by CB-1158 Blocks Myeloid Cell-Mediated Immune Suppression in the Tumor Microenvironment. J. Immunother. Cancer 2017, 5, 101. [Google Scholar] [CrossRef] [PubMed]
- Grzybowski, M.M.; Stańczak, P.S.; Pomper, P.; Błaszczyk, R.; Borek, B.; Gzik, A.; Nowicka, J.; Jędrzejczak, K.; Brzezińska, J.; Rejczak, T.; et al. OATD-02 Validates the Benefits of Pharmacological Inhibition of Arginase 1 and 2 in Cancer. Cancers (Basel). 2022, 14, 3967. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, M.A.; Ugel, S.; Hübbe, M.L.; Carretta, M.; Perez-Penco, M.; Weis-Banke, S.E.; Martinenaite, E.; Kopp, K.; Chapellier, M.; Adamo, A.; et al. Arginase 1–Based Immune Modulatory Vaccines Induce Anticancer Immunity and Synergize with Anti–PD-1 Checkpoint Blockade. Cancer Immunol. Res. 2021, 9, 1316–1326. [Google Scholar] [CrossRef]
- Clinical Trials Using INCB001158 in Cancer | List Results - ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/results?cond=cancer&term=incb001158&cntry=&state=&city=&dist= (accessed on 29 May 2023).
- Clinical Trials Using Arginase Vaccine in Cancer | List Results - ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/results?cond=cancer&term=arginase+vaccine&cntry=&state=&city=&dist= (accessed on 29 May 2023).
- Szefel, J.; Ślebioda, T.; Walczak, J.; Kruszewski, W.J.; Szajewski, M.; Ciesielski, M.; Stanisławowski, M.; Buczek, T.; Małgorzewicz, S.; Owczarzak, A.; et al. The Effect of L-Arginine Supplementation and Surgical Trauma on the Frequency of Myeloid-Derived Suppressor Cells and T Lymphocytes in Tumour and Blood of Colorectal Cancer Patients. Adv. Med. Sci. 2022, 67, 66–78. [Google Scholar] [CrossRef]
- Lacey, J.M.; Wilmore, D.W. Is Glutamine a Conditionally Essential Amino Acid? Nutr. Rev. 1990, 48, 297–309. [Google Scholar] [CrossRef]
- Soeters, P.B.; Grecu, I. Have We Enough Glutamine and How Does It Work? A Clinician’s View. Ann. Nutr. Metab. 2012, 60, 17–26. [Google Scholar] [CrossRef]
- Halama, A.; Suhre, K. Advancing Cancer Treatment by Targeting Glutamine Metabolism—A Roadmap. Cancers (Basel). 2022, 14, 553. [Google Scholar] [CrossRef]
- Cruzat, V.; Macedo Rogero, M.; Noel Keane, K.; Curi, R.; Newsholme, P. Glutamine: Metabolism and Immune Function, Supplementation and Clinical Translation. Nutrients 2018, 10, 1564. [Google Scholar] [CrossRef] [PubMed]
- Hodson, N.; Brown, T.; Joanisse, S.; Aguirre, N.; West, D.; Moore, D.; Baar, K.; Breen, L.; Philp, A. Characterisation of L-Type Amino Acid Transporter 1 (LAT1) Expression in Human Skeletal Muscle by Immunofluorescent Microscopy. Nutrients 2017, 10, 23. [Google Scholar] [CrossRef] [PubMed]
- Daemen, A.; Liu, B.; Song, K.; Kwong, M.; Gao, M.; Hong, R.; Nannini, M.; Peterson, D.; Liederer, B.M.; de la Cruz, C.; et al. Pan-Cancer Metabolic Signature Predicts Co-Dependency on Glutaminase and De Novo Glutathione Synthesis Linked to a High-Mesenchymal Cell State. Cell Metab. 2018, 28, 383–399.e9. [Google Scholar] [CrossRef]
- Jiang, J.; Srivastava, S.; Zhang, J. Starve Cancer Cells of Glutamine: Break the Spell or Make a Hungry Monster? Cancers (Basel). 2019, 11, 804. [Google Scholar] [CrossRef]
- Kubota, A.; Meguid, M.M.; Hitch, D.C. Amino Acid Profiles Correlate Diagnostically with Organ Site in Three Kinds of Malignant Tumors. Cancer 1992, 69, 2343–2348. [Google Scholar] [CrossRef]
- Miyagi, Y.; Higashiyama, M.; Gochi, A.; Akaike, M.; Ishikawa, T.; Miura, T.; Saruki, N.; Bando, E.; Kimura, H.; Imamura, F.; et al. Plasma Free Amino Acid Profiling of Five Types of Cancer Patients and Its Application for Early Detection. PLoS One 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Pollard, A.C.; Paolillo, V.; Radaram, B.; Qureshy, S.; Li, L.; Maity, T.; Wang, L.; Uddin, M.N.; Wood, C.G.; Karam, J.A.; et al. PET/MR Imaging of a Lung Metastasis Model of Clear Cell Renal Cell Carcinoma with (2S,4R)-4-[18F]Fluoroglutamine. Mol. imaging Biol. 2022. [Google Scholar] [CrossRef]
- Dunphy, M.P.S.; Harding, J.J.; Venneti, S.; Zhang, H.; Burnazi, E.M.; Bromberg, J.; Omuro, A.M.; Hsieh, J.J.; Mellinghoff, I.K.; Staton, K.; et al. In Vivo PET Assay of Tumor Glutamine Flux and Metabolism: In-Human Trial of 18f-(2S,4R)-4-Fluoroglutamine. Radiology 2018, 287, 667–675. [Google Scholar] [CrossRef]
- Xu, X.; Zhu, H.; Liu, F.; Zhang, Y.; Yang, J.; Zhang, L.; Zhu, L.; Li, N.; Kung, H.F.; Yang, Z. Imaging Brain Metastasis Patients with 18F-(2S,4 R)-4-Fluoroglutamine. Clin. Nucl. Med. 2018, 43, e392–e399. [Google Scholar] [CrossRef]
- Grkovski, M.; Goel, R.; Krebs, S.; Staton, K.D.; Harding, J.J.; Mellinghoff, I.K.; Humm, J.L.; Dunphy, M.P.S. Pharmacokinetic Assessment of 18F-(2S,4R)-4-Fluoroglutamine in Patients with Cancer. J. Nucl. Med. 2020, 61, 357–366. [Google Scholar] [CrossRef]
- van Geldermalsen, M.; Wang, Q.; Nagarajah, R.; Marshall, A.D.; Thoeng, A.; Gao, D.; Ritchie, W.; Feng, Y.; Bailey, C.G.; Deng, N.; et al. ASCT2/SLC1A5 Controls Glutamine Uptake and Tumour Growth in Triple-Negative Basal-like Breast Cancer. Oncogene 2016, 35, 3201–3208. [Google Scholar] [CrossRef]
- Bhutia, Y.D.; Ganapathy, V. Glutamine Transporters in Mammalian Cells and Their Functions in Physiology and Cancer. Biochim. Biophys. Acta - Mol. Cell Res. 2016, 1863, 2531–2539. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Xiang, W.; Xu, Y.; Han, L.; Li, Q.; Dai, W.; Cai, G. Enhanced Glutamine Utilization Mediated by SLC1A5 and GPT2 Is an Essential Metabolic Feature of Colorectal Signet Ring Cell Carcinoma with Therapeutic Potential. Ann. Transl. Med. 2020, 8, 302–302. [Google Scholar] [CrossRef]
- Cormerais, Y.; Massard, P.A.; Vucetic, M.; Giuliano, S.; Tambutté, E.; Durivault, J.; Vial, V.; Endou, H.; Wempe, M.F.; Parks, S.K.; et al. The Glutamine Transporter ASCT2 (SLC1A5) Promotes Tumor Growth Independently of the Amino Acid Transporter LAT1 (SLC7A5). J. Biol. Chem. 2018, 293, 2877–2887. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, R.; Shuai, Y.; Huang, Y.; Jin, R.; Wang, X.; Luo, J. ASCT2 (SLC1A5)-Dependent Glutamine Uptake Is Involved in the Progression of Head and Neck Squamous Cell Carcinoma. Br. J. Cancer 2020, 122, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Wise, D.R.; Deberardinis, R.J.; Mancuso, A.; Sayed, N.; Zhang, X.Y.; Pfeiffer, H.K.; Nissim, I.; Daikhin, E.; Yudkoff, M.; McMahon, S.B.; et al. Myc Regulates a Transcriptional Program That Stimulates Mitochondrial Glutaminolysis and Leads to Glutamine Addiction. Proc. Natl. Acad. Sci. U. S. A. 2008, 105, 18782–18787. [Google Scholar] [CrossRef]
- Toda, K.; Nishikawa, G.; Iwamoto, M.; Itatani, Y.; Takahashi, R.; Sakai, Y.; Kawada, K. Clinical Role of ASCT2 (SLC1A5) in KRAS-Mutated Colorectal Cancer. Int. J. Mol. Sci. 2017, 18, 1632. [Google Scholar] [CrossRef]
- Zhang, J.; Pavlova, N.N.; Thompson, C.B. Cancer Cell Metabolism: The Essential Role of the Nonessential Amino Acid, Glutamine. EMBO J. 2017, 36, 1302–1315. [Google Scholar] [CrossRef]
- Lee, J.S.; Kang, J.H.; Lee, S.H.; Hong, D.; Son, J.; Hong, K.M.; Song, J.; Kim, S.Y. Dual Targeting of Glutaminase 1 and Thymidylate Synthase Elicits Death Synergistically in NSCLC. Cell Death Dis. 2016, 7, e2511–e2511. [Google Scholar] [CrossRef]
- Xiang, L.; Mou, J.; Shao, B.; Wei, Y.; Liang, H.; Takano, N.; Semenza, G.L.; Xie, G. Glutaminase 1 Expression in Colorectal Cancer Cells Is Induced by Hypoxia and Required for Tumor Growth, Invasion, and Metastatic Colonization. Cell Death Dis. 2019, 10, 40. [Google Scholar] [CrossRef]
- Dorai, T.; Dorai, B.; Pinto, J.T.; Grasso, M.; Cooper, A.J.L. High Levels of Glutaminase II Pathway Enzymes in Normal and Cancerous Prostate Suggest a Role in ‘Glutamine Addiction’. Biomolecules 2019, 10, 2. [Google Scholar] [CrossRef] [PubMed]
- Pan, T.; Gao, L.; Wu, G.; Shen, G.; Xie, S.; Wen, H.; Yang, J.; Zhou, Y.; Tu, Z.; Qian, W. Elevated Expression of Glutaminase Confers Glucose Utilization via Glutaminolysis in Prostate Cancer. Biochem. Biophys. Res. Commun. 2015, 456, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Myint, Z.W.; Sun, R.C.; Hensley, P.J.; James, A.C.; Wang, P.; Strup, S.E.; McDonald, R.J.; Yan, D.; St. Clair, W.H.; Allison, D.B. Evaluation of Glutaminase Expression in Prostate Adenocarcinoma and Correlation with Clinicopathologic Parameters. Cancers (Basel). 2021, 13, 2157. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, D.H.; Jung, W.H.; Koo, J.S. Expression of Glutamine Metabolism-Related Proteins According to Molecular Subtype of Breast Cancer. Endocr. Relat. Cancer 2013, 20, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.C.; Yu, Y.C.; Sung, Y.; Han, J.M. Glutamine Reliance in Cell Metabolism. Exp. Mol. Med. 2020, 52, 1496–1516. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Samuels, Y.; Li, Q.; Krokowski, D.; Guan, B.J.; Wang, C.; Jin, Z.; Dong, B.; Cao, B.; Feng, X.; et al. Oncogenic PIK3CA Mutations Reprogram Glutamine Metabolism in Colorectal Cancer. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef]
- Bernfeld, E.; Foster, D.A. Glutamine as an Essential Amino Acid for KRas-Driven Cancer Cells. Trends Endocrinol. Metab. 2019, 30, 357–368. [Google Scholar] [CrossRef]
- Niklison-Chirou, M.V.; Erngren, I.; Engskog, M.; Haglöf, J.; Picard, D.; Remke, M.; McPolin, P.H.R.; Selby, M.; Williamson, D.; Clifford, S.C.; et al. TAp73 Is a Marker of Glutamine Addiction in Medulloblastoma. Genes Dev. 2017, 31, 1738–1753. [Google Scholar] [CrossRef]
- Nguyen, T.L.; Nokin, M.; Terés, S.; Tomé, M.; Bodineau, C.; Galmar, O.; Pasquet, J.; Rousseau, B.; Liempd, S.; Falcon-Perez, J.M.; et al. Downregulation of Glutamine Synthetase, Not Glutaminolysis, Is Responsible for Glutamine Addiction in Notch1-driven Acute Lymphoblastic Leukemia. Mol. Oncol. 2021, 15, 1412–1431. [Google Scholar] [CrossRef]
- Gross, M.I.; Demo, S.D.; Dennison, J.B.; Chen, L.; Chernov-Rogan, T.; Goyal, B.; Janes, J.R.; Laidig, G.J.; Lewis, E.R.; Li, J.; et al. Antitumor Activity of the Glutaminase Inhibitor CB-839 in Triple-Negative Breast Cancer. Mol. Cancer Ther. 2014, 13, 890–901. [Google Scholar] [CrossRef]
- Robinson, M.M.; McBryant, S.J.; Tsukamoto, T.; Rojas, C.; Ferraris, D.V.; Hamilton, S.K.; Hansen, J.C.; Curthoys, N.P. Novel Mechanism of Inhibition of Rat Kidney-Type Glutaminase by Bis-2-(5-Phenylacetamido-1,2,4-Thiadiazol-2-Yl)Ethyl Sulfide (BPTES). Biochem. J. 2007, 406, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Wang, J. Bin; Erickson, J.W.; Fuji, R.; Ramachandran, S.; Gao, P.; Dinavahi, R.; Wilson, K.F.; Ambrosio, A.L.B.; Dias, S.M.G.; Dang, C.V.; et al. Targeting Mitochondrial Glutaminase Activity Inhibits Oncogenic Transformation. Cancer Cell 2010, 18, 207–219. [Google Scholar] [CrossRef]
- Hoerner, C.R.; Chen, V.J.; Fan, A.C. The ‘Achilles Heel’ of Metabolism in Renal Cell Carcinoma: Glutaminase Inhibition as a Rational Treatment Strategy. Kidney Cancer 2019, 3, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Clinical Trials Using CB839 in Cancer| List Results - ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/results?cond=Cancer&term=CB839&cntry=&state=&city=&dist= (accessed on 29 May 2023).
- Kalinsky, K.; Harding, J.; DeMichele, A.; Infante, J.; Gogineni, K.; Owonikoko, T.; Isakoff, S.; Iliopoulos, O.; Patel, M.; Munster, P.; et al. Abstract PD3-13: Phase 1 Study of CB-839, a First-in-Class Oral Inhibitor of Glutaminase, in Combination with Paclitaxel in Patients with Advanced Triple Negative Breast Cancer. Cancer Res. 2018, 78, PD3-13. [Google Scholar] [CrossRef]
- Calithera Biosciences, I. Initial Results from Phase 2 Study of CB-839 in Combination with Opdivo® (Nivolumab) to Be Presented at the Society for Immunotherapy of Cancer Meeting. Available online: https://www.globenewswire.com/en/news-release/2017/11/07/1176454/32478/en/Initial-Results-from-Phase-2-Study-of-CB-839-in-Combination-with-Opdivo-nivolumab-to-be-Presented-at-the-Society-for-Immunotherapy-of-Cancer-Meeting.html (accessed on 29 September 2023).
- Harding, J.J.; Telli, M.L.; Munster, P.N.; Le, M.H.; Molineaux, C.; Bennett, M.K.; Mittra, E.; Burris, H.A.; Clark, A.S.; Dunphy, M.; et al. Safety and Tolerability of Increasing Doses of CB-839, a First-in-Class, Orally Administered Small Molecule Inhibitor of Glutaminase, in Solid Tumors. J. Clin. Oncol. 2015, 33, 2512–2512. [Google Scholar] [CrossRef]
- Lee, C.H.; Motzer, R.; Emamekhoo, H.; Matrana, M.; Percent, I.; Hsieh, J.J.; Hussain, A.; Vaishampayan, U.; Liu, S.; McCune, S.; et al. Telaglenastat plus Everolimus in Advanced Renal Cell Carcinoma: A Randomized, Double-Blinded, Placebo-Controlled, Phase II ENTRATA Trial. Clin. Cancer Res. 2022, 28, 3248–3255. [Google Scholar] [CrossRef]
- Tannir, N.M.; Agarwal, N.; Porta, C.; Lawrence, N.J.; Motzer, R.; McGregor, B.; Lee, R.J.; Jain, R.K.; Davis, N.; Appleman, L.J.; et al. Efficacy and Safety of Telaglenastat Plus Cabozantinib vs Placebo Plus Cabozantinib in Patients With Advanced Renal Cell Carcinoma. JAMA Oncol. 2022. [Google Scholar] [CrossRef]
- Wang, E.S.; Frankfurt, O.; Orford, K.W.; Bennett, M.; Flinn, I.W.; Maris, M.; Konopleva, M. Phase 1 Study of CB-839, a First-in-Class, Orally Administered Small Molecule Inhibitor of Glutaminase in Patients with Relapsed/Refractory Leukemia. Blood 2015, 126, 2566–2566. [Google Scholar] [CrossRef]
- Motzer, R.J.; Lee, C.-H.; Emamekhoo, H.; Matrana, M.; Percent, I.; Hsieh, J.J.; Hussain, A.; Vaishampayan, U.N.; Graham, R.; Liu, S.; et al. ENTRATA: Randomized, Double-Blind, Phase II Study of Telaglenastat (Tela; CB-839) + Everolimus (E) vs Placebo (Pbo) + E in Patients (Pts) with Advanced/Metastatic Renal Cell Carcinoma (MRCC). Ann. Oncol. 2019, 30, v889–v890. [Google Scholar] [CrossRef]
- Schulte, M.L.; Fu, A.; Zhao, P.; Li, J.; Geng, L.; Smith, S.T.; Kondo, J.; Coffey, R.J.; Johnson, M.O.; Rathmell, J.C.; et al. Pharmacological Blockade of ASCT2-Dependent Glutamine Transport Leads to Antitumor Efficacy in Preclinical Models. Nat. Med. 2018, 24, 194–202. [Google Scholar] [CrossRef]
- Leone, R.D.; Zhao, L.; Englert, J.M.; Sun, I.-M.; Oh, M.-H.; Sun, I.-H.; Arwood, M.L.; Bettencourt, I.A.; Patel, C.H.; Wen, J.; et al. Glutamine Blockade Induces Divergent Metabolic Programs to Overcome Tumor Immune Evasion. Science 2019, 366, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- Hanaford, A.R.; Alt, J.; Rais, R.; Wang, S.Z.; Kaur, H.; Thorek, D.L.J.; Eberhart, C.G.; Slusher, B.S.; Martin, A.M.; Raabe, E.H. Orally Bioavailable Glutamine Antagonist Prodrug JHU-083 Penetrates Mouse Brain and Suppresses the Growth of MYC-Driven Medulloblastoma. Transl. Oncol. 2019, 12, 1314–1322. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, A.S.; da Costa Rosa, M.; Stumpo, V.; Rais, R.; Slusher, B.S.; Riggins, G.J. The Glutamine Antagonist Prodrug JHU-083 Slows Malignant Glioma Growth and Disrupts MTOR Signaling. Neuro-Oncology Adv. 2021, 3, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Suru, A.; Islam, M.; Tam, A.; Gross, J.; Llosa, N. 580 Glutamine Blockade in Combination with Immune Checkpoint Blockade Remodels the Myeloid Landscape in Mouse Models of Soft Tissue Sarcomas. J. Immunother. Cancer 2021, 9, A609–A610. [Google Scholar] [CrossRef]
- Oh, M.-H.; Travers, M.; Brown, S.; Zhao, L.; Sun, I.-M.; Sun, I.-H.; Arwood, M.; Xu, W.; Collins, S.; Leone, R.; et al. Abstract LB-022: Targeting Glutamine Metabolism as a Mean of Treating a Murine Model of Ovarian Cancer and Ascites Development. Cancer Res. 2019, 79, LB-022. [Google Scholar] [CrossRef]
- Oh, M.H.; Sun, I.H.; Zhao, L.; Leone, R.D.; Sun, I.M.; Xu, W.; Collins, S.L.; Tam, A.J.; Blosser, R.L.; Patel, C.H.; et al. Targeting Glutamine Metabolism Enhances Tumor-Specific Immunity by Modulating Suppressive Myeloid Cells. J. Clin. Invest. 2020, 130, 3865–3884. [Google Scholar] [CrossRef]
- Chan, W.K.; Horvath, T.D.; Tan, L.; Link, T.; Harutyunyan, K.G.; Pontikos, M.A.; Anishkin, A.; Du, D.; Martin, L.A.; Yin, E.; et al. Glutaminase Activity of L-Asparaginase Contributes to Durable Preclinical Activity against Acute Lymphoblastic Leukemia. Mol. Cancer Ther. 2019, 18, 1587–1592. [Google Scholar] [CrossRef] [PubMed]
- Offman, M.N.; Krol, M.; Patel, N.; Krishnan, S.; Liu, J.Z.; Saha, V.; Bates, P.A. Rational Engineering of L-Asparaginase Reveals Importance of Dual Activity for Cancer Cell Toxicity. Blood 2011, 117, 1614–1621. [Google Scholar] [CrossRef]
- Reinfeld, B.I.; Madden, M.Z.; Wolf, M.M.; Chytil, A.; Bader, J.E.; Patterson, A.R.; Sugiura, A.; Cohen, A.S.; Ali, A.; Do, B.T.; et al. Cell-Programmed Nutrient Partitioning in the Tumour Microenvironment. Nature 2021, 593, 282–288. [Google Scholar] [CrossRef]
- Pallett, L.J.; Dimeloe, S.; Sinclair, L. V.; Byrne, A.J.; Schurich, A. A Glutamine ‘Tug-of-War’: Targets to Manipulate Glutamine Metabolism for Cancer Immunotherapy. Immunother. Adv. 2021, 1. [Google Scholar] [CrossRef]
- Horvath, K.; Jami, M.; Hill, I.D.; Papadimitriou, J.C.; Magder, L.S.; Chanasongcram, S. Isocaloric Glutamine-Free Diet and the Morphology and Function of Rat Small Intestine. J. Parenter. Enter. Nutr. 1996, 20, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, S.; Saha, A.; Azam, M.; Mukherjee, A.; Sur, P. Role of Oral Glutamine in Alleviation and Prevention of Radiation-Induced Oral Mucositis: A Prospective Randomized Study. South Asian J. Cancer 2014, 3, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Tsujimoto, T.; Yamamoto, Y.; Wasa, M.; Takenaka, Y.; Nakahara, S.; Takagi, T.; Tsugane, M.; Hayashi, N.; Maeda, K.; Inohara, H.; et al. L-Glutamine Decreases the Severity of Mucositis Induced by Chemoradiotherapy in Patients with Locally Advanced Head and Neck Cancer: A Double-Blind, Randomized, Placebo-Controlled Trial. Oncol. Rep. 2015, 33, 33–39. [Google Scholar] [CrossRef]
- Anderson, P.M.; Lalla, R.V. Glutamine for Amelioration of Radiation and Chemotherapy Associated Mucositis during Cancer Therapy. Nutrients 2020, 12, 1675. [Google Scholar] [CrossRef] [PubMed]
- Ishak Gabra, M.B.; Yang, Y.; Li, H.; Senapati, P.; Hanse, E.A.; Lowman, X.H.; Tran, T.Q.; Zhang, L.; Doan, L.T.; Xu, X.; et al. Dietary Glutamine Supplementation Suppresses Epigenetically-Activated Oncogenic Pathways to Inhibit Melanoma Tumour Growth. Nat. Commun. 2020, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Guillén-Mancina, E.; Jiménez-Alonso, J.J.; Calderón-Montaño, J.M.; Jiménez-González, V.; Díaz-Ortega, P.; Burgos-Morón, E.; López-Lázaro, M. Artificial Diets with Selective Restriction of Amino Acids and Very Low Levels of Lipids Induce Anticancer Activity in Mice with Metastatic Triple-Negative Breast Cancer. Cancers (Basel). 2023, 15, 1540. [Google Scholar] [CrossRef] [PubMed]
- van Lith, S.A.M.; Navis, A.C.; Verrijp, K.; Niclou, S.P.; Bjerkvig, R.; Wesseling, P.; Tops, B.; Molenaar, R.; van Noorden, C.J.F.; Leenders, W.P.J. Glutamate as Chemotactic Fuel for Diffuse Glioma Cells: Are They Glutamate Suckers? Biochim. Biophys. Acta - Rev. Cancer 2014, 1846, 66–74. [Google Scholar] [CrossRef]
- Tardito, S.; Oudin, A.; Ahmed, S.U.; Fack, F.; Keunen, O.; Zheng, L.; Miletic, H.; Sakariassen, P.Ø.; Weinstock, A.; Wagner, A.; et al. Glutamine Synthetase Activity Fuels Nucleotide Biosynthesis and Supports Growth of Glutamine-Restricted Glioblastoma. Nat. Cell Biol. 2015, 17, 1556–1568. [Google Scholar] [CrossRef]
- Krall, A.S.; Xu, S.; Graeber, T.G.; Braas, D.; Christofk, H.R. Asparagine Promotes Cancer Cell Proliferation through Use as an Amino Acid Exchange Factor. Nat. Commun. 2016, 7, 1–13. [Google Scholar] [CrossRef]
- Appel, I.M.; Kazemier, K.M.; Boos, J.; Lanvers, C.; Huijmans, J.; Veerman, A.J.P.; van Wering, E.; den Boer, M.L.; Pieters, R. Pharmacokinetic, Pharmacodynamic and Intracellular Effects of PEG-Asparaginase in Newly Diagnosed Childhood Acute Lymphoblastic Leukemia: Results from a Single Agent Window Study. Leukemia 2008, 22, 1665–1679. [Google Scholar] [CrossRef]
- Story, M.D.; Voehringer, D.W.; Stephens, L.C.; Meyn, R.E. L-Asparaginase Kills Lymphoma Cells by Apoptosis. Cancer Chemother. Pharmacol. 1993, 32, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, C.; Weinert, B.T.; Nishida, Y.; Verdin, E.; Mann, M. The Growing Landscape of Lysine Acetylation Links Metabolism and Cell Signalling. Nat. Rev. Mol. Cell Biol. 2014, 15, 536–550. [Google Scholar] [CrossRef] [PubMed]
- Aslanian, A.M.; Kilberg, M.S. Multiple Adaptive Mechanisms Affect Asparagine Synthetase Substrate Availability in Asparaginase-Resistant MOLT-4 Human Leukaemia Cells. Biochem. J. 2001, 358, 59–67. [Google Scholar] [CrossRef]
- Fine, B.M.; Kaspers, G.J.L.; Ho, M.; Loonen, A.H.; Boxer, L.M. A Genome-Wide View of the in Vitro Response to L-Asparaginase in Acute Lymphoblastic Leukemia. Cancer Res. 2005, 65, 291–299. [Google Scholar] [CrossRef]
- Brumano, L.P.; da Silva, F.V.S.; Costa-Silva, T.A.; Apolinário, A.C.; Santos, J.H.P.M.; Kleingesinds, E.K.; Monteiro, G.; Rangel-Yagui, C. de O.; Benyahia, B.; Junior, A.P. Development of L-Asparaginase Biobetters: Current Research Status and Review of the Desirable Quality Profiles. Front. Bioeng. Biotechnol. 2019, 6, 212. [Google Scholar] [CrossRef]
- Pieters, R.; Hunger, S.P.; Boos, J.; Rizzari, C.; Silverman, L.; Baruchel, A.; Goekbuget, N.; Schrappe, M.; Pui, C.-H. L-Asparaginase Treatment in Acute Lymphoblastic Leukemia. Cancer 2011, 117, 238–249. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Batra, S.; Zhang, J. Asparagine: A Metabolite to Be Targeted in Cancers. Metabolites 2021, 11, 402. [Google Scholar] [CrossRef] [PubMed]
- Pavlova, N.N.; Hui, S.; Ghergurovich, J.M.; Fan, J.; Intlekofer, A.M.; White, R.M.; Rabinowitz, J.D.; Thompson, C.B.; Zhang, J. As Extracellular Glutamine Levels Decline, Asparagine Becomes an Essential Amino Acid. Cell Metab. 2018, 27, 428–438.e5. [Google Scholar] [CrossRef]
- Gwinn, D.M.; Lee, A.G.; Briones-Martin-del-Campo, M.; Conn, C.S.; Simpson, D.R.; Scott, A.I.; Le, A.; Cowan, T.M.; Ruggero, D.; Sweet-Cordero, E.A. Oncogenic KRAS Regulates Amino Acid Homeostasis and Asparagine Biosynthesis via ATF4 and Alters Sensitivity to L-Asparaginase. Cancer Cell 2018, 33, 91–107.e6. [Google Scholar] [CrossRef]
- Knott, S.R.V.; Wagenblast, E.; Khan, S.; Kim, S.Y.; Soto, M.; Wagner, M.; Turgeon, M.O.; Fish, L.; Erard, N.; Gable, A.L.; et al. Asparagine Bioavailability Governs Metastasis in a Model of Breast Cancer. Nature 2018, 554, 378–381. [Google Scholar] [CrossRef]
- Halbrook, C.J.; Thurston, G.; Mccarthy, A.; Nelson, B.S.; Sajjakulnukit, P.; Krall, A.S.; Mullen, P.J.; Zhang, L.; Batra, S.; Viale, A.; et al. Clonal Heterogeneity Supports Mitochondrial Metabolism in Pancreatic Cancer. bioRxiv, 0983. [Google Scholar] [CrossRef]
- Hinze, L.; Labrosse, R.; Degar, J.; Han, T.; Schatoff, E.M.; Schreek, S.; Karim, S.; McGuckin, C.; Sacher, J.R.; Wagner, F.; et al. Exploiting the Therapeutic Interaction of Wnt Pathway Activation and Asparaginase for Colorectal Cancer Therapy. Cancer Discov. 2020, 10, 1690–1705. [Google Scholar] [CrossRef] [PubMed]
- Krall, A.S.; Mullen, P.J.; Surjono, F.; Momcilovic, M.; Schmid, E.W.; Halbrook, C.J.; Thambundit, A.; Mittelman, S.D.; Lyssiotis, C.A.; Shackelford, D.B.; et al. Asparagine Couples Mitochondrial Respiration to ATF4 Activity and Tumor Growth. Cell Metab. 2021, 33, 1013–1026.e6. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, L.B.; Luengo, A.; Danai, L.V.; Bush, L.N.; Diehl, F.F.; Hosios, A.M.; Lau, A.N.; Elmiligy, S.; Malstrom, S.; Lewis, C.A.; et al. Aspartate Is an Endogenous Metabolic Limitation for Tumour Growth. Nat. Cell Biol. 2018, 20, 782–788. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, L.B.; Gui, D.Y.; Hosios, A.M.; Bush, L.N.; Freinkman, E.; Vander Heiden, M.G. Supporting Aspartate Biosynthesis Is an Essential Function of Respiration in Proliferating Cells. Cell 2015, 162, 552–563. [Google Scholar] [CrossRef] [PubMed]
- Birsoy, K.; Wang, T.; Chen, W.W.; Freinkman, E.; Abu-Remaileh, M.; Sabatini, D.M. An Essential Role of the Mitochondrial Electron Transport Chain in Cell Proliferation Is to Enable Aspartate Synthesis. Cell 2015, 162, 540–551. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Bermudez, J.; Baudrier, L.; La, K.; Zhu, X.G.; Fidelin, J.; Sviderskiy, V.O.; Papagiannakopoulos, T.; Molina, H.; Snuderl, M.; Lewis, C.A.; et al. Aspartate Is a Limiting Metabolite for Cancer Cell Proliferation under Hypoxia and in Tumours. Nat. Cell Biol. 2018, 20, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Alkan, H.F.; Walter, K.E.; Luengo, A.; Madreiter-Sokolowski, C.T.; Stryeck, S.; Lau, A.N.; Al-Zoughbi, W.; Lewis, C.A.; Thomas, C.J.; Hoefler, G.; et al. Cytosolic Aspartate Availability Determines Cell Survival When Glutamine Is Limiting. Cell Metab. 2018, 28, 706–720.e6. [Google Scholar] [CrossRef]
- Sarabhai, T.; Roden, M. Hungry for Your Alanine: When Liver Depends on Muscle Proteolysis. J. Clin. Invest. 2019, 129, 4563–4566. [Google Scholar] [CrossRef]
- Sousa, C.M.; Biancur, D.E.; Wang, X.; Halbrook, C.J.; Sherman, M.H.; Zhang, L.; Kremer, D.; Hwang, R.F.; Witkiewicz, A.K.; Ying, H.; et al. Pancreatic Stellate Cells Support Tumour Metabolism through Autophagic Alanine Secretion. Nature 2016, 536, 479–483. [Google Scholar] [CrossRef]
- Parker, S.J.; Amendola, C.R.; Hollinshead, K.E.R.; Yu, Q.; Yamamoto, K.; Encarnación-Rosado, J.; Rose, R.E.; LaRue, M.M.; Sohn, A.S.W.; Biancur, D.E.; et al. Selective Alanine Transporter Utilization Creates a Targetable Metabolic Niche in Pancreatic Cancer. Cancer Discov. 2020, 10, 1018–1037. [Google Scholar] [CrossRef]
- Geng, P.; Qin, W.; Xu, G. Proline Metabolism in Cancer. Amino Acids 2021, 53, 1769–1777. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Burghardt, R.C.; Johnson, G.A.; Kim, S.W.; Knabe, D.A.; Li, P.; Li, X.; McKnight, J.R.; Satterfield, M.C.; et al. Proline and Hydroxyproline Metabolism: Implications for Animal and Human Nutrition. Amino Acids 2011, 40, 1053–1063. [Google Scholar] [CrossRef]
- Olivares, O.; Mayers, J.R.; Gouirand, V.; Torrence, M.E.; Gicquel, T.; Borge, L.; Lac, S.; Roques, J.; Lavaut, M.N.; Berthezène, P.; et al. Collagen-Derived Proline Promotes Pancreatic Ductal Adenocarcinoma Cell Survival under Nutrient Limited Conditions. Nat. Commun. 2017, 8, 1–14. [Google Scholar] [CrossRef]
- Sahu, N.; Dela Cruz, D.; Gao, M.; Sandoval, W.; Haverty, P.M.; Liu, J.; Stephan, J.-P.; Haley, B.; Classon, M.; Hatzivassiliou, G.; et al. Proline Starvation Induces Unresolved ER Stress and Hinders MTORC1-Dependent Tumorigenesis. Cell Metab. 2016, 24, 753–761. [Google Scholar] [CrossRef] [PubMed]
- Ragni, M.; Ruocco, C.; Tedesco, L.; Carruba, M.O.; Valerio, A.; Nisoli, E. An Amino Acid-Defined Diet Impairs Tumour Growth in Mice by Promoting Endoplasmic Reticulum Stress and MTOR Inhibition. Mol. Metab. 2022, 60, 101478. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.K.; Yau, C.; Becker, B.C.; Khateeb, S.; Mahoney, S.; Jensen, M.B.; Hann, B.; Cowen, B.J.; Pegan, S.D.; Benz, C.C. Targeting Mitochondrial Proline Dehydrogenase with a Suicide Inhibitor to Exploit Synthetic Lethal Interactions with P53 Upregulation and Glutaminase Inhibition. Mol. Cancer Ther. 2019, 18, 1374–1385. [Google Scholar] [CrossRef] [PubMed]
- Elia, I.; Broekaert, D.; Christen, S.; Boon, R.; Radaelli, E.; Orth, M.F.; Verfaillie, C.; Grünewald, T.G.P.; Fendt, S.-M. Proline Metabolism Supports Metastasis Formation and Could Be Inhibited to Selectively Target Metastasizing Cancer Cells. Nat. Commun. 2017, 8, 15267. [Google Scholar] [CrossRef]
- Liu, Y.; Mao, C.; Wang, M.; Liu, N.; Ouyang, L.; Liu, S.; Tang, H.; Cao, Y.; Liu, S.; Wang, X.; et al. Cancer Progression Is Mediated by Proline Catabolism in Non-Small Cell Lung Cancer. Oncogene 2020, 39, 2358–2376. [Google Scholar] [CrossRef]
- Loayza-Puch, F.; Rooijers, K.; Buil, L.C.M.; Zijlstra, J.; Oude Vrielink, J.F.; Lopes, R.; Ugalde, A.P.; Van Breugel, P.; Hofland, I.; Wesseling, J.; et al. Tumour-Specific Proline Vulnerability Uncovered by Differential Ribosome Codon Reading. Nature 2016, 530, 490–494. [Google Scholar] [CrossRef]
- Ding, Z.; Ericksen, R.E.; Escande-Beillard, N.; Lee, Q.Y.; Loh, A.; Denil, S.; Steckel, M.; Haegebarth, A.; Wai Ho, T.S.; Chow, P.; et al. Metabolic Pathway Analyses Identify Proline Biosynthesis Pathway as a Promoter of Liver Tumorigenesis. J. Hepatol. 2020, 72, 725–735. [Google Scholar] [CrossRef]
- Ding, J.; Kuo, M.-L.; Su, L.; Xue, L.; Luh, F.; Zhang, H.; Wang, J.; Lin, T.G.; Zhang, K.; Chu, P.; et al. Human Mitochondrial Pyrroline-5-Carboxylate Reductase 1 Promotes Invasiveness and Impacts Survival in Breast Cancers. Carcinogenesis 2017, 38, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wu, L.; Li, K.; Liu, F.; Wang, L.; Zhang, D.; Zhou, J.; Ma, X.; Wang, S.; Yang, S. Ornithine Aminotransferase Promoted the Proliferation and Metastasis of Non-small Cell Lung Cancer via Upregulation of MiR-21. J. Cell. Physiol. 2019, 234, 12828–12838. [Google Scholar] [CrossRef] [PubMed]
- Zigmond, E.; Ya’acov, A. Ben; Lee, H.; Lichtenstein, Y.; Shalev, Z.; Smith, Y.; Zolotarov, L.; Ziv, E.; Kalman, R.; Le, H.V.; et al. Suppression of Hepatocellular Carcinoma by Inhibition of Overexpressed Ornithine Aminotransferase. ACS Med. Chem. Lett. 2015, 6, 840–844. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.; Lamb, J.; Smith, S.; Wheatley, D.N. Single Amino Acid (Arginine) Deprivation: Rapid and Selective Death of Cultured Transformed and Malignant Cells. Br. J. Cancer 2000, 83, 800–810. [Google Scholar] [CrossRef]
Figure 1.
Chemical structure of proteinogenic AAs: EAAs (a) and NEAAs (b).
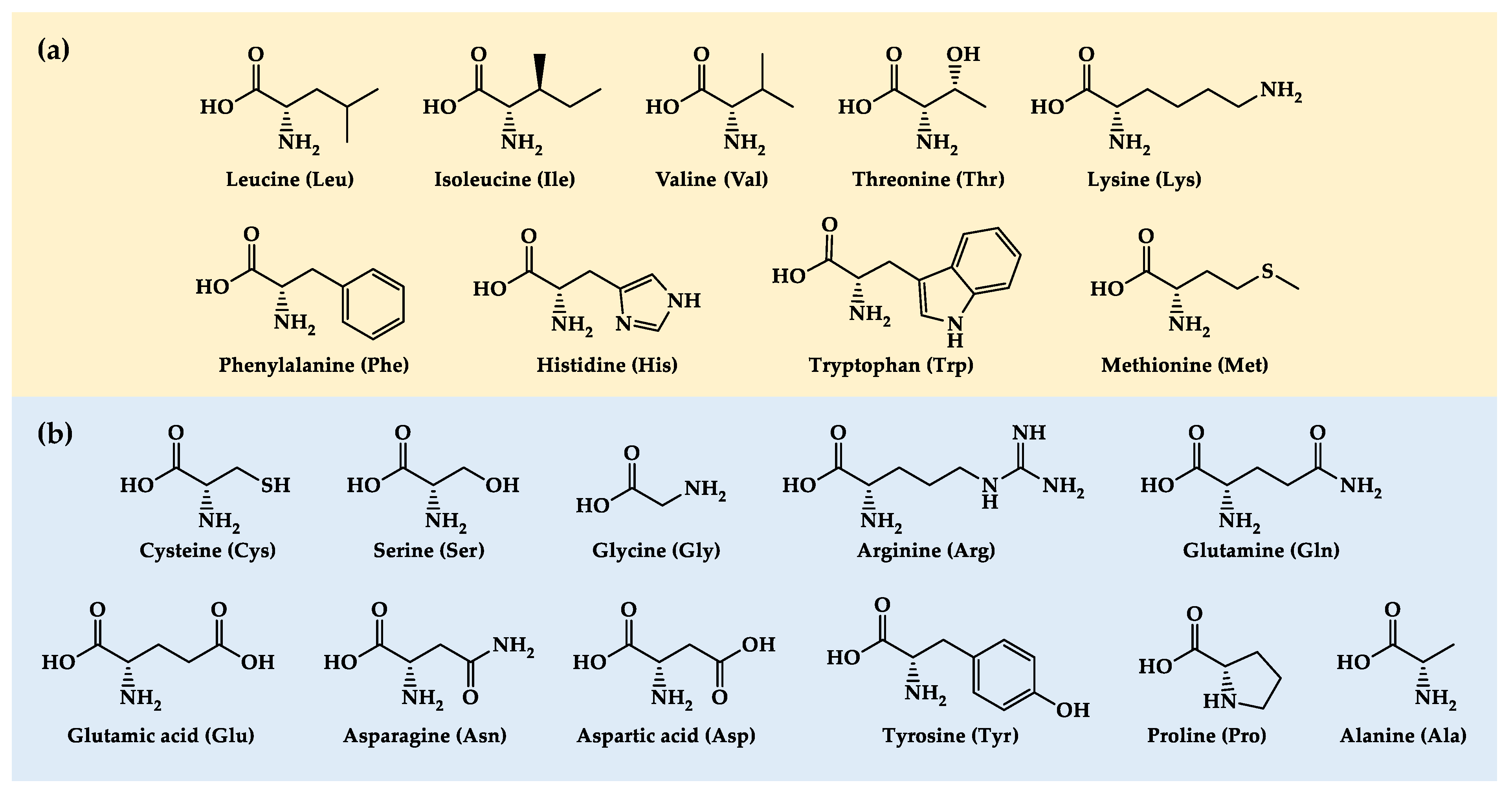
Figure 2.
Schematic representation of key metabolic pathways for the biosynthesis of NEAAs, the enzymes in each pathway, and the main functions of each AA [54,173]. NEAAs are represented in blue and EAAs in red. Leucine (Leu), isoleucine (Ile), histidine (His), valine (Val), phenylalanine (Phe), threonine (Thr), methionine (Met), lysine (Lys), tryptophan (Trp), glutamine (Gln), alanine (Ala), aspartate (Asp), asparagine (Asn), arginine (Arg), tyrosine (Tyr), glutamate (Glu), cysteine (Cys), glycine (Gly), proline (Pro), and serine (Ser). 3-phospho-D-glycerate (3-PG), S-adenosylmethionine (SAM), homocysteine (HCys), glutathione (GSH), α-ketoglutarate (α-KGlu), tricarboxylic acid cycle (TCA), oxaloacetate (OAA), reactive oxygen species (ROS). D-3-phosphoglycerate dehydrogenase (PHGDH), phosphoserine aminotransferase-1 (PSAT1), vacuolar protein sorting-associated protein-29 (VPS29), phosphoserine phosphatase (PSPH), serine hydroxymethyltransferase-1 (SHMT1), serine hydroxymethyltransferase-2 (SHMT2), S-adenosylmethionine synthase isoform type-1 (MAT1A), S-adenosylmethionine synthetase isoform type-2 (MAT2A), methionine adenosyltransferase 2 subunit beta (MAT2B), adenosylhomocysteinase (AHCY), cystathionine β-synthase (CBS), cystathionine γ-lyase (CTH), methionine synthase (MTR), betaine-homocysteine methyltransferase (BHMT), betaine-homocysteine methyltransferase-2 (BHMT2), S-adenosylmethionine decarboxylase (AMD1), spermidine synthase (SRM), 5’-methylthioadenosine phosphorylase (MTAP), methylthioribose-1-phosphate isomerase (MRI1), methylthioribulose 1-phosphate dehydratase (APIP), enolase-phosphatase (ENOPH1), 1,2-dihydroxy-3-keto-5-methylthiopentene dioxygenase (ADI1), 2-oxo-4-methylthiobutanoate aminotransferase (KYAT1), phenylalanine hydroxylase (PAH), alanine aminotransferase-1 (GPT), alanine aminotransferase-2 (GPT2), glutaminase-1 (GLS1), glutaminase-2 (GLS2), glutamine synthetase (GLUL), glutamate dehydrogenase-1 (GLUD1), glutamate dehydrogenase-2 (GLUD2), aspartate aminotransferase-1 (GOT1), aspartate aminotransferase-2 (GOT2), asparagine synthetase (ASNS), asparaginase (ASRGL1), aspartylglucosaminidase (AGA), argininosuccinate synthase (ASS1), argininosuccinate lyase (ASL), ornithine aminotransferase (OAT), pyrroline-5-carboxylate reductase-1 (PYCR1), pyrroline-5-carboxylate reductase-2 (PYCR2), δ-1-pyrroline-5-carboxylate synthase (ALDH18A1). Enzymes that participate in consecutive steps in a metabolic pathway are separated by “-”, enzymes that catalyze the same step in a metabolic pathway are separated by “/“.
Figure 2.
Schematic representation of key metabolic pathways for the biosynthesis of NEAAs, the enzymes in each pathway, and the main functions of each AA [54,173]. NEAAs are represented in blue and EAAs in red. Leucine (Leu), isoleucine (Ile), histidine (His), valine (Val), phenylalanine (Phe), threonine (Thr), methionine (Met), lysine (Lys), tryptophan (Trp), glutamine (Gln), alanine (Ala), aspartate (Asp), asparagine (Asn), arginine (Arg), tyrosine (Tyr), glutamate (Glu), cysteine (Cys), glycine (Gly), proline (Pro), and serine (Ser). 3-phospho-D-glycerate (3-PG), S-adenosylmethionine (SAM), homocysteine (HCys), glutathione (GSH), α-ketoglutarate (α-KGlu), tricarboxylic acid cycle (TCA), oxaloacetate (OAA), reactive oxygen species (ROS). D-3-phosphoglycerate dehydrogenase (PHGDH), phosphoserine aminotransferase-1 (PSAT1), vacuolar protein sorting-associated protein-29 (VPS29), phosphoserine phosphatase (PSPH), serine hydroxymethyltransferase-1 (SHMT1), serine hydroxymethyltransferase-2 (SHMT2), S-adenosylmethionine synthase isoform type-1 (MAT1A), S-adenosylmethionine synthetase isoform type-2 (MAT2A), methionine adenosyltransferase 2 subunit beta (MAT2B), adenosylhomocysteinase (AHCY), cystathionine β-synthase (CBS), cystathionine γ-lyase (CTH), methionine synthase (MTR), betaine-homocysteine methyltransferase (BHMT), betaine-homocysteine methyltransferase-2 (BHMT2), S-adenosylmethionine decarboxylase (AMD1), spermidine synthase (SRM), 5’-methylthioadenosine phosphorylase (MTAP), methylthioribose-1-phosphate isomerase (MRI1), methylthioribulose 1-phosphate dehydratase (APIP), enolase-phosphatase (ENOPH1), 1,2-dihydroxy-3-keto-5-methylthiopentene dioxygenase (ADI1), 2-oxo-4-methylthiobutanoate aminotransferase (KYAT1), phenylalanine hydroxylase (PAH), alanine aminotransferase-1 (GPT), alanine aminotransferase-2 (GPT2), glutaminase-1 (GLS1), glutaminase-2 (GLS2), glutamine synthetase (GLUL), glutamate dehydrogenase-1 (GLUD1), glutamate dehydrogenase-2 (GLUD2), aspartate aminotransferase-1 (GOT1), aspartate aminotransferase-2 (GOT2), asparagine synthetase (ASNS), asparaginase (ASRGL1), aspartylglucosaminidase (AGA), argininosuccinate synthase (ASS1), argininosuccinate lyase (ASL), ornithine aminotransferase (OAT), pyrroline-5-carboxylate reductase-1 (PYCR1), pyrroline-5-carboxylate reductase-2 (PYCR2), δ-1-pyrroline-5-carboxylate synthase (ALDH18A1). Enzymes that participate in consecutive steps in a metabolic pathway are separated by “-”, enzymes that catalyze the same step in a metabolic pathway are separated by “/“.
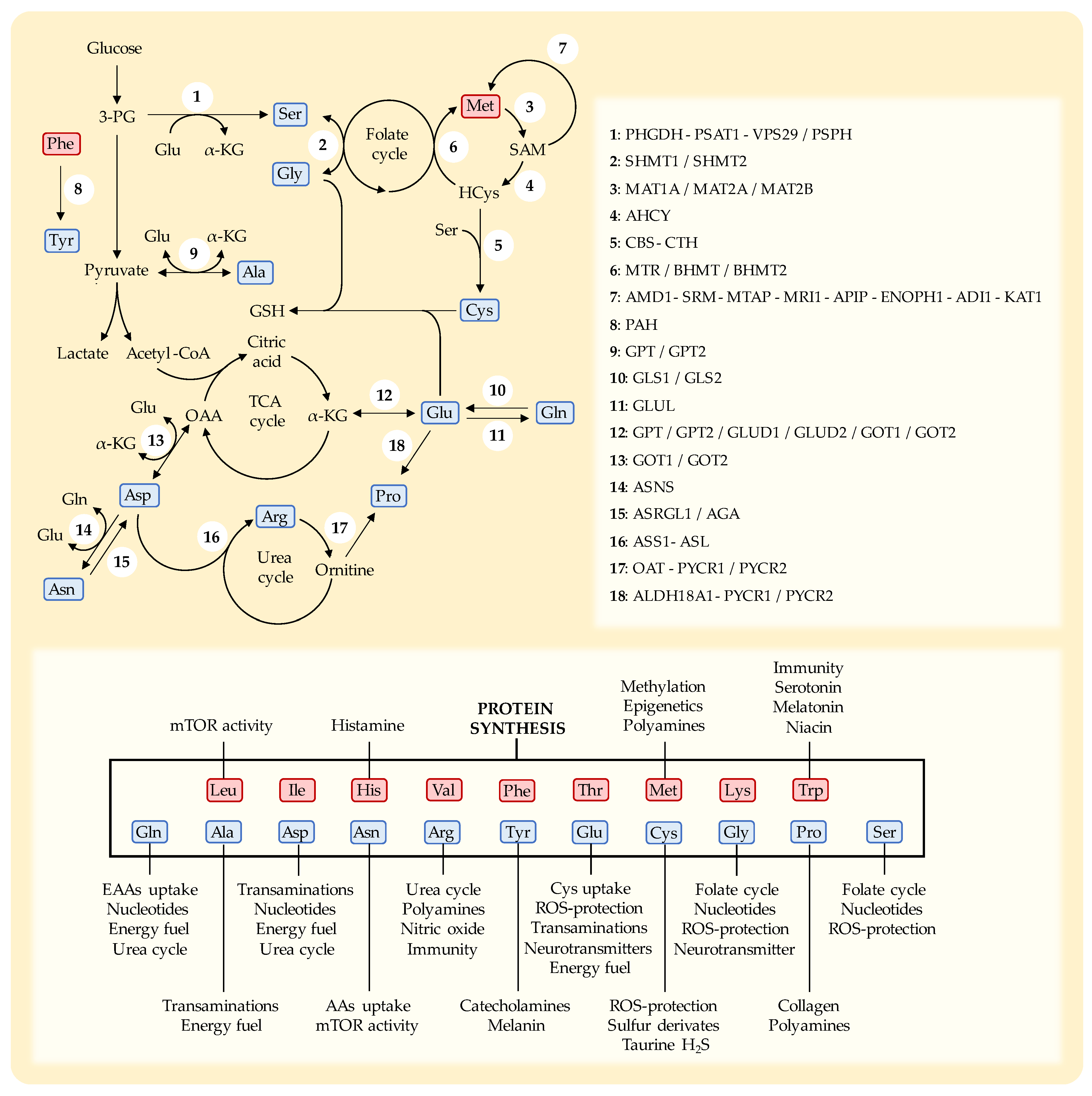

Table 1.
Effect of manipulating dietary protein intake in mice with cancer.
| Articles | Relevant results in preclinical in vivo cancer models |
|---|---|
| Levine et al. 2014 [31] | Low protein diet (4% vs. 18% kcal protein) reduced IFG-1 levels and decreased tumor growth in syngeneic models of melanoma and breast cancer. Weight loss was observed in older mice. |
| Brandhorst et al. 2013 [32] | Low protein diet (4% vs. 19% kcal protein) did not reduce cancer progression in a syngeneic glioma murine model. |
| Rubio-Patiño et al. 2018 [33] | Low protein diet (15-17% vs 19.5% protein) reduced cancer progression in syngeneic models of lymphoma and colon cancer. Low protein diet enhanced anticancer immunity. |
| Orillion et al. 2018 [34] | Low protein diet (7% vs 21% protein) reduced tumor growth in syngeneic models of prostate and renal cancer by increasing antitumor immunity. Synergistic effect with immunotherapies. |
| Fontana et al. 2013 [35] | Low protein diet (7% vs. 20% protein) reduced tumor growth in mice xenografts of prostate and breast cancer. Diet with a 20% plant protein showed lower tumor growth than diet with 20% animal protein. |
| Taha et al. 2018 [36] | Plant-based protein diet induced tumor growth inhibition compared to animal-based protein diet in two syngeneic models of ovarian cancer (20% protein in both diets). |
Table 2.
Results of Leu restriction / supplementation in cancer therapy in vivo.
| Articles | Relevant results in preclinical in vivo cancer models |
|---|---|
| Sugimura et al. 1959 [39] | Leu restricted diet for 5 days reduced tumor growth in Walker cancer bearing rats. Approximately 1-2 g/day body weight loss was observed in rats fed the Leu restricted diet. |
| Theuer 1971 [40] | Dietary limitation of Leu (from 0.8% to 0.1%) for 3 weeks reduced tumor growth in mice with breast adenocarcinoma. Significant body weight loss was also observed. |
| Sheen et al. 2011 [41] | Leu restricted diet for 14 days plus autophagy inhibitor (chloroquine) synergistically suppressed cancer growth in a xenograft melanoma model. |
| Xiao et al. 2016 [42] | Leu restricted diet for 4 days reduced tumor growth in mice xenografted with human triple-negative breast cancer cells. Supplementation with palmitic acid and overexpression of FASN enzyme blocked the anticancer effect of Leu restriction. |
| Liu et al. 2014 [49] | Diet supplemented with 5% Leu enhanced tumor growth in a syngeneic pancreatic cancer model. This effect was observed in lean and overweight mice. |
| Jiménez-Alonso et al. 2022 [26] | Supplementation of 2.5% Leu to casein-based diets increased their anticancer activity in two syngeneic murine models of metastatic colon cancer. |
| Calderón-Montaño et al. 2022 [27] | Supplementation of 2.5% Leu to casein-based diets increased their anticancer activity in mice with disseminated renal cell carcinoma. |
Table 3.
In vivo anticancer effects of Met restriction.
| Articles | Relevant results in preclinical in vivo cancer models |
|---|---|
| Sugimura et al. 1959 [39] | Dietary Met depletion (0%) for 11 days suppressed tumor growth in Walker cancer bearing rats, causing the animals to lose l-2 g weight/day. |
| Goseki et al. 1991 [121] | Restriction of Met and Cys in total parenteral diet inhibited tumor growth, reduced the number of metastases and improved survival in a sarcoma rat model. Synergistic effect with 5-fluorouracil. |
| Goseki et al. 1996 [122] | Restriction of Met and Cys in total parenteral diet followed by vincristine improved survival of rats with sarcoma. |
| Guo et al. 1996 [123] | Dietary Met depletion (0%) plus ethionine (Met analogue and antagonist) showed synergic anticancer activity in a sarcoma rat cancer model. |
| Xiao et al. 2001 [124] | Restriction of Met in total parenteral diet for 7 days suppressed cancer growth and prolonged survival of rats with gastric cancer. Synergistic effect with 5-fluorouracil. |
| Hoshiya et al. 1995 [125] | Met-free diet inhibited growth of human cancer xenografts in nude mice. |
| Hoshiya et al. 1996 [105] | Dietary Met depletion (0%) induced anticancer activity in mice xenografted with human breast cancer cells and increased the antitumor activity of cisplatin. |
| Hoshiya et al. 1997 [106] | Dietary Met depletion (0%) induced anticancer activity in mice xenografted with human gastric cancer cells and increased the antitumor activity of 5-fluorouracil. |
| Guo et al. 1993 [107] | Met-free diet extended mice survival in a xenograft sarcoma model, with eventual cancer regression. |
| Jeon et al. 2016 [108] | Met-free diet for 10 days decreased the number of tumors in the lungs in a syngeneic triple-negative breast cancer model in mice. This diet induced significant weight loss. Mice survival was not evaluated. |
| Strekalova et al. 2015 [109] | Met-free diet for 5 weeks inhibited tumor growth in mice xenografted with human triple-negative breast cancer cells. Synergistic effect with lexatumumab (TNF receptor agonist). |
| Malin et al. 2021 [110] | Met-free diet for 4 weeks showed anticancer activity against triple-negative breast cancer xenograft and PDX models. Synergistic effect with auranofin (TXNRDs inhibitor). |
| Lui et al. 2015 [111] | Double Cys and Met deprivation inhibited tumor growth and triggered autophagy in a xenograft glioma model in mice. |
| Breillout et al. 1987 [129] | Met-restricted diet supplemented with homocysteine reduced the metastatic dissemination of cancer cells in a rhabdomyosarcoma rat model. |
| Orillion et al. 2018 [34] | Dietary Met restriction (0.092%) induced anticancer activity in models of prostate (RP-B6Myc) and renal (RENCA) cell carcinoma. Synergistic effect with immunotherapies. |
| Theuer 1971 [40] | Dietary Met restriction (0.10%) for 3 weeks showed anticancer activity in a spontaneous breast adenocarcinoma model. Diets with higher Met levels (0.20-0.60%) showed no anticancer activity. All diets were also restricted in Cys. |
| Sinha et al. 2014 [112] | Dietary Met restriction (0.12%) for 11 weeks reduced the development and severity of prostate cancer in a transgenic murine model of prostate adenocarcinoma. |
| Hens et al. 2016 [115] | Dietary Met restriction (0.12%) for 12 weeks induced anticancer activity in mice xenografted with human breast cancer cells. Reduced plasma levels of Met, Cys and Tau were reported. |
| Gao et al. 2019 [114] | Dietary Met restriction (0.12%) induced anticancer activity in a transgenic sarcoma and patient-derived xenograft colorectal cancer models. The diet increased the anticancer effect of radiotherapy and 5-fluorouracil. |
| Liu et al. 2022 [117] | Dietary Met restriction (0.12%) induced anticancer activity in mice xenografted with human colorectal cancer cells. Synergistic effect with 5-fluorouracil. |
| Li et al. 2023 [118] | Dietary Met restriction (0.12%) improved antitumor immunity and showed a synergistic effect with anti-PD-1 immunotherapy in two syngeneic models of colorectal cancer. |
| Upadhyayula et al. 2023 [120] | Dietary restriction of Cys and Met (0.0% Cys 0.15% Met vs. 0.40% Cys 0.43% Met) for 7 days induced anticancer activity in a murine glioma model. Synergistic effect with GPX4 inhibitor. |
| Xu et al. 2020 [113] | Dietary Met restriction (0.17%) inhibited HNF4α-positive liver cancer growth in mice |
| Komninou et al. 2006 [116] | Met limited diet (0.17%) inhibited the development and proliferation of colonic tumors in an induced colon cancer rat model. |
| Calderón-Montaño et al. 2022 [27] | Artificial diets with manipulated levels of AAs markedly improved survival of mice with disseminated renal cell carcinoma. Several active diets formulated with free AAs contained 0.60% Met. However, the anticancer activity of casein-based diets (0.17% Met) was completely blocked by adding 0.5% Met supplement. |
| Jiménez-Alonso et al. 2023 [119] | Artificial diets restricted in Met (0.17%) showed anticancer activity in mice with metastatic colon cancer, ovarian cancer and renal cell carcinoma. Met supplementation blocked the anticancer activity in mice with colon cancer. |
Table 4.
In vivo anticancer effects of Cys restriction.
| Articles | Relevant results in preclinical cancer in vivo models |
|---|---|
| Voegtlin et al. 1936 [133] | A diet deficient in Cys and Met (approximately 0.06% Cys and 0.17% Met) reduced tumor growth in mice with spontaneous breast adenocarcinomas. Addition of 0.6% CysS stimulated tumor growth abruptly. |
| Theuer 1971 [40] | 3 weeks of treatment with a diet restricted in Cys and limited in Met (0.10%) showed anticancer activity in a spontaneous breast adenocarcinoma model. Diets restricted in Cys but with higher Met levels (0.20-0.60%) showed no anticancer activity. |
| Zhang et al. 2020 [174] | Dietary Cys restriction reduced tumor growth in mice xenografted with human colon cancer cells. Loss/inhibition of MTAP upregulated polyamine metabolism and increased the activity of Cys restriction. |
| Wu et al. 2021 [175] | Dietary Cys restriction suppressed cancer growth in mice xenografted with human colon cancer cells without causing weight loss. Synergistic effect with oxaliplatin. |
| Ruiz-Rodado et al. 2022 [176] | Dietary Cys restriction reduced plasma levels of Cys and GSH and increased mice survival in a xenograft orthotopic glioma model. |
| Goseki et al. 1991 [121] | Total parenteral diet without Cys and Met induced anticancer activity in a sarcoma rat model. Synergistic effect with 5-fluorouracil. |
| Goseki et al. 1996 [122] | Total parenteral diet without Cys and Met followed by vincristine improved survival in a sarcoma rat cancer model. |
| Lui et al. 2015 [111] | Dietary depletion of Cys and Met inhibited tumor growth and triggered autophagy in a xenograft glioma model in mice. |
| Orillion et al. 2018 [34] | Diet restricted in Cys and limited in Met (0.092%) showed anticancer activity in a transgenic prostate cancer model in mice. Synergistic effect with immunotherapies. |
| Upadhyayula et al. 2023 [120] | Dietary restriction of Cys and Met (0.0% Cys 0.15% Met vs. 0.40% Cys 0.43% Met) for 7 days induced anticancer activity in a murine glioma model. Synergistic effect with GPX4 inhibitor. |
| Jiménez-Alonso et al. 2022 [26] | Two artificial diets deficient in Cys/Met improved survival of mice with metastatic colon cancer. The addition of 0.2% Cys blocked the anticancer activity of both diets. |
| Calderón-Montaño et al. 2022 [27] | Artificial diet lacking 6 NEAAs (including Cys) and with normal Met levels (0.6%) showed marked anticancer activity in mice with disseminated renal cell carcinoma; the anticancer activity of this diet was reduced by supplementing Cys. However, supplementing 0.2% Cys to an inactive casein-based diet markedly improved its anticancer activity in mice with disseminated renal cell carcinoma. |
Table 5.
In vivo anticancer effects of Ser and Gly restriction.
| Articles | Relevant results in preclinical cancer in vivo models |
|---|---|
| Maddocks et al. 2013 [214] 1 | Dietary Ser/Gly restriction induced anticancer activity in mice xenografted with p53-defective colon cancer cells. |
| Maddocks et al. 2017 [208] 1 | Dietary Ser/Gly restriction induced in vivo anticancer activity that could be improved by antagonizing the anti-oxidant response |
| Humpton et al. 2018 [215] 1 | A commonly occurring p53 mutant, R248W, retains wild-type ability to support survival under serine starvation. The growth of R248W-expressing tumors was resistant to dietary Ser/Gly restriction. |
| LeBoeuf et al. 2020 [216] 1 | Dietary Ser/Gly restriction inhibited tumor growth in mice with mutated KEAP1. Synergistic effect with a GLS inhibitor (CB-839) even in non-mutated cancers. |
| Tajan et al. 2021 [217] 1 | Dietary Ser/Gly restriction plus PH755 (PHGDH inhibitor) synergistically improved the anticancer activity in colon cancer xenografts |
| Falcone et al. 2022 [218] 1 | Dietary Ser/Gly restriction improved the anticancer effect of radiotherapy in syngeneic models of pancreatic cancer and triple-negative breast cancer. |
| Pranzini et al. 2022 [219] 1 | Dietary Ser/Gly restriction was inactive in syngeneic and xenograft colon cancer models. However, the combination with 5-fluorouracil showed a synergistic anticancer effect. |
| Gravel et al. 2014 [209] | Dietary Ser/Gly restriction reduced Ser levels in plasma and tumors. The combination of Ser-Gly restricted diet plus phenformin reduced tumor growth in a syngeneic colon cancer model. |
| Polet et al. 2016 [220] | Dietary Ser/Gly restriction improved survival in a murine syngeneic model of leukemia. Combination with a GLS inhibitor (BPTES) synergistically improved mice survival. |
| Méndez-Lucas et al. 2020 [221] | Dietary Ser/Gly restriction plus inhibition of Ser biosynthesis (PSAT1 knockdown) showed anticancer activity in a murine model of c-MYC-induced liver cancer. Each intervention alone did not show activity. |
| Van Nyen et al. 2022 [210] | Dietary Ser/Gly restriction reduced tumor growth in a platinum-resistant ovarian cancer model in mice. Mice with platinum-sensitive ovarian cancer cells were insensitive to the diet. |
| Sullivan et al. 2019 [211] | Dietary Ser/Gly restriction plus PHGDH knockdown significantly reduced tumor growth in triple-negative breast cancer xenografts. PHGDH overexpression reduced the effect of this restriction. |
| Muthusamy et al. 2020 [212] | Dietary Ser/Gly restriction altered the biosynthesis of sphingolipids and decreased tumor growth in a xenograft model of colon cancer. |
| Fujihara et al. 2022 [213] | Dietary Ser/Gly restriction induced anticancer activity in a xenograft model of esophageal cancer. Combination with a ferroptosis inducer (eprenetapopt) synergistically improved mice survival. |
| Calderón-Montaño et al. 2022 [27] | An artificial diet lacking Ser and other 5 NEAAs (Ser, Cys, Tyr, Pro, Asn and Glu) markedly improved the survival of mice with disseminated renal cell carcinoma. Ser supplementation did not decrease the activity of the diet, and Gly elimination did not improve the activity of the diet. |
1 The control diet and the Ser-Gly restricted diet both lacked the NEAAs Ala, Pro, Glu, Asn and Ast (the only difference was the addition / restriction of Ser and Gly).
Table 6.
In vivo anticancer effect of Arg dietary restriction or supplementation.
| Articles | Relevant results in preclinical cancer in vivo models |
|---|---|
| Gonzalez and Byus 1991 [232] | Dietary Arg restriction reduced the ornithine available for polyamine biosynthesis and reduced the incidence and multiplicity of papillomas in a mouse model of skin cancer. |
| Yeatman et al. 1991 [233] | Dietary Arg restriction inhibited tumor growth in a syngeneic model of colon cancer in mice. Arg supplementation stimulated tumor growth. |
| Alexandrou et al. 2018 [235] | Dietary Arg restriction reduced tumor growth in mice xenografted with human colorectal cancer cells deficient in ASS1 and OTC (ornithine transcarbamylase). |
| Cheng et al. 2018 [234] | Dietary Arg restriction reduced tumor size in two xenograft breast cancer models of ASS1 deficient cells. No weight loss was observed in the animals. |
| Hsu et al. 2021 [236] | Dietary Arg restriction suppressed prostate cancer growth in xenograft models. The Arg-free diet reduced cancer cell proliferation and enhanced inflammatory response. |
| Missiaen et al. 2022 [237] | Dietary Arg restriction induced anticancer activity in xenograft model of hepatic cancer. Combination with GNC2 and BCL2 inhibitors synergistically enhanced the anticancer response. |
| Cao, Feng, et al. 2016 [238] | Arg supplementation (1.5 g/kg) reduced tumor growth in a syngeneic orthotopic breast cancer model. Reduction of myeloid-derived suppressor cells (MDSCs) and enhanced antitumor immune responses were observed. |
| Cao, Wang et al. 2016 [239] | Arg supplementation (1.5 g/kg) plus docetaxel synergistically inhibited tumor growth in a syngeneic breast cancer model. Reduction of MDSCs and enhanced antitumor immune responses were observed. |
| Geiger et al 2016 [240] | Arg supplementation (1.5 g/kg) suppressed tumor growth and enhanced survival of mice with an immunogenic melanoma. |
| Satoh et al. 2020 [241] | Arg supplementation (1.5 g/kg) combined with cyclophosphamide and anti-PD-1 significantly increased the number of cured mice in a syngeneic colon cancer model. |
| He et al. 2017 [242] | Arg supplementation (2.0 g/kg) combined with anti-PD-1 immunotherapy synergistically increased the survival of mice with osteosarcoma. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



