
Submitted:
30 May 2023
Posted:
31 May 2023
You are already at the latest version
Abstract
Neurodegenerative disorders are characterized by a progressive process of degeneration and neuronal death, where oxidative stress and neuroinflammation are key factors that contribute to the progression of these diseases. Therefore, two major pathways involved in these pathologies have been proposed as relevant therapeutic targets: The nuclear transcription factor erythroid 2 (Nrf2), which responds to oxidative stress with cytoprotecting activity and the nuclear factor NF-κB pathway, which is highly related to the neuroinflammatory process by promoting cytokine expression. Caffeic acid phenethyl ester (CAPE) is a phenylpropanoid naturally found in propolis that shows important biological activities, including neuroprotective activity by modulating the Nrf2 and NF-kB pathways, promoting antioxidant enzyme expression and inhibition of proinflammatory cytokine expression. Its simple chemical structure has inspired the synthesis of many derivatives, with aliphatic and/or aromatic moieties, some of which have improved the biological properties. Moreover, new drug delivery systems increase the bioavailability of these compounds in vivo, allowing its transcytosis through the blood-brain barrier, thus protecting brain cells from the increased inflammatory status associated to neurodegenerative and psychiatric disorders. This review summarizes the biosynthesis and chemical synthesis of CAPE derivatives, their miscellaneous activities, and relevant studies (from 2010 to 2023) addressing their neuroprotective activity in vitro and in vivo.
Keywords:
caffeic acid phenethyl ester
; CAPE derivatives
; anti-inflammatory
; NF-kB
; antioxidant
; Nrf2
; neuroprotection
1. Introduction
The current demographic shift of modern societies has led to a surge in the prevalence of diseases affecting older adults, including neurodegenerative disorders. Among them, Alzheimer's disease (AD) is the first cause of dementia and a significant global public health problem, estimated to affect about 131.5 million people worldwide, with an annual incidence of 4 to 6 million new cases (Dubois, Villain et al. 2021). On the other hand, the second neurodegenerative disorder is Parkinson's disease (PD), affecting one in every hundred people over 60 years of age. It is estimated that by 2030 there will be 9 million patients with idiopathic PD worldwide (Erkkinen, Kim et al. 2018). Moreover, psychiatric disorders with increasing worldwide prevalence, such as major depressive disorder (MDD), are relevant and potentially modifiable risk factors for dementia. Neurodegenerative disorders are characterized by excessive damage of key brain structures, which is followed by neuronal function loss, structural alterations and decrease of cellular survival (Heppner, Ransohoff et al. 2015). However, despite our increasing knowledge about their pathogenic mechanisms, successful therapeutic approaches to tackle neurodegeneration have remained highly elusive.
A wealth of evidence has supported that oxidative stress and neuroinflammation may have a crucial role in the pathogenesis of neurodegenerative disorders, as they are associated with complex systems that activate programmed cell death cascades (Ebadi, Srinivasan et al. 1996, Markesbery and Carney 1999, Barber and Shaw 2010, Leiva, Martínez-Sanguinetti et al. 2019). Activation of the Nrf2 and NF-kB pathways are expected to have a prominent role in the control of oxidative stress and inflammation. Nuclear erythroid transcription factor 2 (Nrf2) responds to oxidative stress, mediating cytoprotection in mammalian cells by the production of a set of antioxidant genes called phase II genes, which produce phase II proteins associated to ROS stabilization (Cullinan, Gordan et al. 2004, Kobayashi, Kang et al. 2004, Sykiotis and Bohmann 2010). In turn, NF-kB controls pro-inflammatory gene expression he synthesis of cytokines such as tumor necrosis factor alpha (TNF-α), interleukin (IL)-1β, IL-6, and IL-8 are directly mediated by NF-kB, as well as the expression of cyclooxygenase-2 (COX2) (Almowallad, Alqahtani et al. 2022).
To date, conventional single drug-based therapeutic approaches for addressing neurodegenerative disorders have not been entirely satisfactory. For instance, in the pharmacological treatment of AD, since the 1990s, only tree cholinesterase inhibitors including galantamine, rivastigmine, and donepezil together with memantine a glutamatergic antagonist are available. Therefore, targeting multiple pharmacological pathways, such as inflammation and oxidative stress, holds great promise for increasing the likelihood of obtaining potent neuroprotective effects. Among the novel and emerging therapeutic approaches, consuming functional foods enriched with natural bioactive molecules has shown great potential as a palliative or preventative measure for the middle-aged population. For instance, propolis is considered a food supplement with relevant therapeutic properties including antidiabetic, anticancer or antibacterial activities. Propolis is a large mixture of polyphenols, essential oils, and waxes produced by plants as resinous secretions. This is collected by honeybees, which use it as a waterproof substance to seal hive cracks or holes (Borba, Wilson et al. 2017). Furthermore, bees use propolis as an antiseptic material to prevent infections due to its powerful antibacterial (Przybyłek and Karpiński 2019) and antiparasitic properties. (Drescher, Klein et al. 2017) The large molecular variety of propolis depends on many factors, such as location, the season, and plants distributed around the hive. (Machado, Silva et al. 2016) Despite this, propolis always shows antioxidant, anti-inflammatory, bactericidal, and antifungal properties, that could be explained due to the high content and variety of flavonoids with a particular chemical composition. (Freires, Queiroz et al. 2016; Pazin, Monaco et al. 2017; and Tiveron, Rosalen et al. 2016)
Caffeic acid phenethyl ester (CAPE) is found mainly in propolis. It is a natural polyphenol from the phenylpropanoid family, formed by the esterification of caffeic acid and phenethyl alcohol (Figure 1) (Metzner, Bekemeier et al. 1979, Romero, Freire et al. 2019) (Kurek-Górecka, Rzepecka-Stojko et al. 2013). This polyphenol is not the major component of propolis, but is one of the most potent antioxidants, given by the caffeic acid moiety which has a higher antioxidant capacity than related phenylpropanoids such as ferulic acid, and p-coumaric acid (Rice-Evans, Miller et al. 1996). The concentration of CAPE in propolis has a large difference, from 0 to 11 mg/g of propolis, quantification by high performance liquid chromatography (HPLC) (Gómez-Caravaca, Gómez-Romero et al. 2006) showed, for example, that propolis collected in Mexico (Ures) has a CAPE concentration of 11.4 mg/g (Hernandez, Goycoolea et al. 2007), followed by propolis from Viveros and Valencia (Spain), with 10,047 and 10,399 mg/g respectively (García Argente 2018). On the other hand, the product from Chile showed 5,798 mg/g, while propolis from China had 4.9 mg/g, in contrast to the propolis from Honduras that did not present CAPE (Argente 2019). The same results were obtained from propolis collected in different regions of Turkey (Ozdal, Ceylan et al. 2019).
CAPE displays a broad spectrum of beneficial activities, including antitumor activity by inducing apoptosis in many cancer cell lines (Chen, Shiao et al. 2001, Watabe, Hishikawa et al. 2004, Xiang, Wang et al. 2006, Hernandez, Goycoolea et al. 2007). For example, on human leukemia HL-60 cells CAPE inhibits DNA, RNA, and protein synthesis with an IC50 of 1.0 μM, 5.0 μM, and 1.5 μM, respectively (Chen, Shao et al. 1996). Moreover, CAPE induces anti-inflammatory and immunomodulatory responses. These activities have been explained by the activation of the erythroid nuclear transcription factor 2 (Nrf2) and the inhibition of the nuclear transcription factor (NF-kB). Thus, CAPE it is available as NF-kB inhibitor for in vitro and in vivo studies. (Murtaza, Sajjad et al. 2015)
This review provides an in-depth examination of the biosynthesis and chemical synthesis of CAPE derivatives, as well as their various activities, with a particular focus on their effects on the Nrf2 and NF-kB signaling pathways implicated in oxidative and inflammatory damage underlying neurodegeneration.
2. Biosynthesis of CAPE
CAPE is synthesized by the phenylpropanoid pathway, beginning with the amino acids phenylalanine or tyrosine, producing 4-coumaric acid. The biosynthesis of CAPE was reviewed by Song, 2019, and is summarized in Figure 2.
4-coumaric acid is bound to coenzyme A by 4-coumaric acid CoA-ligase (4CL) enzymatic activity, producing 4-coumaroyl CoA with an activated carbonyl. This product is further meta hydroxylated by p-coumarate 3-hydroxylase giving the intermediate caffeoyl CoA, which is the precursor of caffeic acid by hydrolysis of CoA, but in the case of biosynthesis of CAPE, the CoA is substituted in one-step with phenethyl alcohol by ester linkage. Phenethyl alcohol is synthesized by two enzymes, an aromatic amino acid decarboxylase such as phenylalanine decarboxylase (PDC) followed by a monoamine oxidase such as phenethylamine oxidase (PEO) (Song, Cho et al. 2019).
The absorption and metabolism of CAPE have not yet been well studied, but it should be similar to caffeic acid or related caffeic acid esters. The absorption and metabolism of the radiolabel [3-14C] caffeic acid have been studied in rats, showing that absorption starts in the stomach after 1 h post-ingestion and is quickly absorbed in the first portion of the intestine, obtaining high concentrations in plasma after 2h, then is excreted mainly by urine as nine derivatives of type sulfonated and glucuronide, with no accumulation in tissues (Omar, Mullen et al. 2012). Caffeic acid esters, such as chlorogenic acid and rosmarinic acid, have shown a similar pattern, being excreted by urine after 4 - 8 h post-ingestion, remaining only 3.3 to 4.0% of total polyphenol content (Omar, Mullen et al. 2012, de Oliveira, Sampaio et al. 2017). Anyway, the pharmacodynamic and pharmacokinetics of murine and human organisms are different. For instance, CAPE shows more stability in human than in rat plasma, where its concentration decreases quickly, producing caffeic acid and by-products. Still, CAPE showed more stability in human plasma, suggesting that CAPE is hydrolyzed by enzymes present in rat plasma but not in human (Celli, Dragani et al. 2007).
3. Biological Properties of CAPE
Numerous studies have evidenced the medicinal properties of propolis, positioning it as a superfood. Many of these beneficial effects are attributable to CAPE, Table 1.
4. CAPE Derivatives as Novel Bioactive Compounds
The structure of CAPE has been the inspiration to organic chemists for the development of chemical analogs that improve the activities of the original molecule. The synthesis of CAPE derivatives has been improved over the years, from the initial Fisher esterification catalyzed with acids, giving low yields, to the use of coupling reagents as DCC or by the condensation of acid chlorine derivative, or using microwave assistance. Here we resume some methods that produce acceptable yields and are easy to use (Figure 3).
Choi et al 2019 produced ester derivatives with the starting alcohol and 1.2 equivalents of Meldrum's acid, by reflux overnight, getting the corresponding malonic acid monoester, then 1 eq. of an aldehyde produces the corresponding CAPE derivative by Knoevenagel condensation, as shown in the Figure 3 entry 1 to 3 (Choi, Villegas et al. 2019). An alternative pathway involves microwave assistance, considerably reducing reaction times but giving moderate yields (entries 4 to 6).
The N,N′-dicyclohexylcarbodiimide (DCC) coupling method was used by Chen et al. in the synthesis of CAPE. Starting with caffeic acid and 5 equivalents of phenethyl alcohol, produced 38% yield of CAPE after purification (Figure 4, entry 1). Nagaoka synthetized twenty CAPE analogues by acyl chlorine condensation, starting with caffeic acid and chlorine thionyl in a dry atmosphere, then the corresponding alcohol was added, producing the corresponding CAPE derivative (Figure 4, entry 2a to 2c). The product 4-phenylbutyl caffeate showed antiproliferative activity with EC50 of 20 μM against murine colon 26-L5 carcinoma cells (Nagaoka, Banskota et al. 2002).
Different methods have achieved the synthesis of CAPE and its derivatives. However, some routes were not shown here because they present low yields, inconvenient reagents, or burdensome purification procedures, in contrast to the methods summarized in the Figure 3 and Figure 4. CAPE derivatives were studied in vitro and in vivo due to antiviral, and anticancer activities showed. In Figure 5 some of its esters are shown.
Compound (E)-phenyl 3-(3,4-dihydroxyphenyl) acrylate is the best result of a screening of 19 CAPE-derivatives, searching for inhibitors of the enzyme xanthine oxidase (XO). XO inhibition is a therapeutic strategy for treating hyperuricemia, which results in uric acid crystals accumulation in joints, producing gout or inflammatory arthritis. This CAPE-derivative inhibits XO more efficiently than CAPE by π-π interactions with the enzyme. This enzymatic activity, together with the anti-inflammatory capacities of the related molecules, suggests its use in the treatment of hyperuricemia (Choi, Villegas et al. 2019). Moreover, n-Alkyl esters of CAPE have shown antiviral activity against the hepatitis C virus (HCV), with actions depending on and length of the aliphatic chain and substituents in the catechol moiety. Among derivatives with 1, 4, 6, 8, and 10 carbons, octyl caffeate (CAOE, Figure 5) exhibited the highest activity against HCV with an EC50 value of 2.7 mM, inhibiting genotype 1b and 2a of the virus, independent of interferon signaling. Moreover, CAOE showed synergistic activity with antivirals such as interferon-alpha 2b, daclatasvir, and VX-222, enhancing their activities. Shen and collaborators evidenced that CAOE is protective against hepato-toxic stimuli and mitochondrial dysfunction induced by tert-butyl hydroperoxide in HepG2 Cells. CAOE showed the best results compared to ten different compounds, including amides and esters of caffeic acid (Shen, Yamashita et al. 2013, Tsai, Yu et al. 2017).
A more extended side chain derivative, with 12 carbons, decyl caffeate (DC, Figure 5) showed inhibition of human colorectal cancer cells HT-29 and HCT-116 cell proliferation, in vivo and in vitro, on xenograft mouse model, enhancing cell-cycle arrest at the S phase, via blockade of the STAT3 and Akt signaling pathways. Therefore, DC inhibits cell proliferation by inducing autophagy in HCT-116 colon cells (Chen, Kuo et al. 2020).
The replacement of the ester bonding by amide bonding could increase the stability of the molecules regarding acidic pH and esterase activity, increasing their bioavailability. Thus, many caffeoyl amides have been synthesized (Figure 6), and some of them showed neuroprotective and photoprotective effects in cell and animal models.
Compounds K36 and K36H are closely related to CAPE. They change the ester bonding by amide linkage, showing antiphotodamage properties, induced by ultraviolet A (UVA), which is a preponderant factor in skin damage and aging. They were synthesized by condensation of caffeic acid with 2-phenylethanamine or 2-(4-bromophenyl)ethanamine producing K36 or K36H, respectively. K36 reduces ROS levels via nuclear factor erythroid 2-related factor 2 (Nrf2), with the consequent antioxidant response mechanisms, such as the expression of the antioxidant enzyme heme oxygenase-1 (HO-1), together with the inhibition of metalloproteinase (MMP)-1, MMP-2, IL-6, and PGE2. K36, as well as CAPE, reduces iNOS and COX-2 expression by inhibition of NF-κB (Kuo, Chen et al. 2015). Moreover, K36 reduces the activation of extracellular-signal-regulated kinase (ERK) and c-Jun N-terminal kinases (JNK), as well as 8-hydroxy-20-deoxyguanosine (8-OHdG) levels (Chu, Wu et al. 2019). The p-bromine derivative K36H displays similar behavior as K36, reducing ROS generation, PGE2, NO formation, DNA damage, overexpression of matrix metalloproteinase (MMP)-1 and MMP-2, as well as the expression of Bax and caspase-3, in line with photoprotective properties, and suggesting that K36 and K36H could be used as active components of new photoprotector products (Kuo, Chiang et al. 2020).
The long-chain amides, CAPE dodecyl amide (CAF12) together with CAPE hexyl amide (CAF6), showed neuroprotective effects in an animal model of Diabetic Retinopathy by ameliorating oxidative stress via increasing antioxidant enzyme retinal superoxide dismutase (SOD). Moreover, treatment with CAF12 or CAF6 decreased iPF2α levels and AKT phosphorylation in diabetic rats, with the promotion of survival and growth of retinal neurons. This resulted in the prevention of negative morphological changes in the retina of diabetic rat models treated with streptozotocin (a diabetic inductor drug) compared with rats treated with CAF12 or CAF6 intravitreally for two weeks (Fathalipour, Eghtedari et al. 2019). Moreover, prior studies with aliphatic amides of caffeic acid with C3, C4, C6, C8, and C12 were evaluated on AD models, showing that CAF12 increases PC12 pheochromocytoma cell survival in serum-deprived conditions, enhancing the nerve growth factor (NGF) effect, and inducing neurite outgrowth via the ERK1/2 and AKT pathways, by activation of PI3K. Moosavi and colleagues suggested that these amides could have neuroprotective properties (Moosavi, Hosseini et al. 2017). A CAF12 and CAF6-related amide, denominated compound 13, with a p-methoxy group incorporated into the phenyl moiety, revealed a potent antioxidant and neuroprotection activity with better results compared to other 20 related compounds evaluated (Chai, Zhao et al. 2018). Compound 13 induced memory loss impairment in a transgenic Drosophila model, as well as inhibitory or disaggregation activity on Aβ peptides in Alzheimer’s disease (AD) models (Chai, Zhao et al. 2018). Although these amides are structurally related to the esters shown in Figure 3, and possess antiviral and anticancer activities, they have yet not been studied adequately in these disease models. Only a molecule with a shorter side chain, the amide PT-93, was studied in glioblastoma multiforme (GBM) such as T98G, U87, U251 and HT22 cell lines. The compound showed antiproliferative activities in these models by MMP-2 and MMP-9 inhibition (Li, Tu et al. 2017).
Amides and esters presented here are the result of inducing chemical variations in the phenethyl moiety of CAPE, producing a variety of bioactive compounds. Moreover, Wan, et al 2019 improved the solubility and absorption of CAPE with the synthesis pf the 4-O-glucoside of CAPE, called FA-97, by coupling of CAPE to an acetyl protected brominated D-glucose, according to the Figure 7 (Wan, Wang et al. 2019).
FA-97 enhances antioxidant and scavenger properties in neuronal cell cultures by directly inhibiting ROS production while also activating Nrf2, which produces an antioxidant cell response. In SH-SY5Y human neuroblastoma and PC12 rat pheochromocytoma cells, FA-97 attenuated H2O2-induced apoptosis by decreasing ROS and malondialdehyde (MDA) levels, also induces cellular superoxide (SOD) and glutathione (GSH) dismutase, quinone oxidoreductase 1(NQO1) and HO-1, by activation of the Nrf2 pathway. On AD models, FA-97 reduced learning and memory impairments in mice via reducing neuronal apoptosis (Wan, Wang et al. 2019). On another hand, FA-97 showed to be a potential drug against inflammatory bowel disease (IBD), which is an inflammatory intestinal disorder. In fact, on a mouse model of colitis employing dextran sodium sulfate (DSS) to induce epithelial damage, FA-97 reduced body weight loss, as well as colon damage and length shortening, via activation of HO-1/Nrf2, showing the excellent properties of this compound in gastrointestinal affections (Mei, Wang et al. 2019).
5. CAPE Inhibits Oxidative Stress by Modulation of the Nrf2 Pathway
The astonishing metabolic activity of the brain renders it particularly susceptible to oxidative damage. Indeed, despite the brain accounts for only 2% of our body weight, it consumes 20% of the whole body's oxygen supply. This puts neuronal cells at risk of generating an excess of free radicals that can overwhelm the available antioxidant systems, leading to oxidative stress (OS). Furthermore, OS is associated with the development and progression of different neurodegenerative disorders, including Alzheimer's disease (AD), Parkinson's disease (PD), and Huntington's disease (HD) (Sies 1986, Halliwell and Gutteridge 2015). Interestingly, transcription factors such as Nrf2 govern the homeostatic oxidative responses, highlighting its potential as a therapeutic target for counterbalancing oxidative damage during neurodegenerative disorders. Indeed, Nrf2 has been shown to upregulate the expression of various antioxidant enzymes, which can be further stimulated with CAPE. Next, we will briefly provide mechanistic insights regarding the effects of chemical modifications during OS associated with neurodegeneration. Further, we will examine evidence supporting the potential of CAPE for positively impacting Nrf2 signaling during neurodegeneration (Dawson and Dawson 1996, Halliwell 2001).
Metabolic activity is a major source of OS in the cells. As such, mitochondria are the major sources of cellular ATP and the main generators of Reactive Oxygen Species (ROS) and OS inside the cells. Indeed around 2% of the oxygen consumed by cells is converted into ROS during oxidative phosphorylation in the mitochondria. Different mechanisms may account for the tight association of OS with neurodegeneration. For instance, mitochondrial exposure to ROS may cause damage of its DNA (DNAmit), including DNA fragmentation, chromatid exchange, translocations and consequent mutations. Altered DNAmit impacts on the expression and abnormal activity of proteins involved in ATP synthesis by uncoupling the electronic transport chain from oxidative phosphorylation (Dawson and Dawson 1996, Halliwell 2001), decreasing ATP production and therefore the activity of the plasma membrane ATPase. Decreased activity of the sodium/potassium transporter (Na/K-ATPase) leads to neuronal membrane depolarization, thus generating an increase in cellular excitation, and facilitating the opening of voltage-sensitive Ca2+ channels. In turn, increased glutamate release (and decreased uptake by the glutamate transporter) leads to activation of the N-methyl-D-aspartate (NMDA)- glutamate receptor, thus contributing to increased Ca2+ influx (Hammer, Parker et al. 1993, Halliwell 2001). Moreover, calcium as activates several Ca2+-dependent signaling pathways, such as nitric oxide synthases (NOS) leading to an excess in the production of nitric oxide (NO) which also generates free radicals causing oxidative damage (Sapolsky 1996, Egan, Kojima et al. 2003, Ceccatelli, Tamm et al. 2007, Liu, Lee et al. 2009).
Oxidative damage is manifested as covalent modifications of different subcellular structures and compartments. Accordingly, ROS can oxidate macromolecules of various nature, including lipids, proteins and nucleic acids, modifying their structure and function (Anand, Kunnumakkara et al. 2007; Tonda-Turo, Origlia et al. 2018). As such, lipid damage by peroxidation is associated to early and late markers, including as hexanonyl-lysine (HEL), acrolein-lysine (ACR), malondialdehyde, F2-isoprostanes and 4-hydroxynonenal (4-HNE) adducts. In turn, oxidative damage of DNA and RNA is reflected in markers such as 8-hydroxy-2-deoxyguanosine (8-OHdG) and 8-hydroxyguanine (8-oxo-dGo) respectively, which have a high mutagenic power (Hayashi 2009, Halliwell and Gutteridge 2015). In the case of AD and HD, an increase of oxidized molecules such as proteins, lipids and nucleic acids is generated by the presence of 3-nitrotyrosine and 3,3'-dithyrosine in the hippocampus, neocortex and cerebrospinal ventricular fluid of patients with the condition (Marlatt, Lucassen et al. 2008). In addition, nuclear and mitochondrial DNA modification and lipoperoxidation are associated with increased 8-OHdG, acrolein, malondialdehyde, 4-HNE and F2-isoprostanes (Kamat, Gadal et al. 2008). It has been shown that there is a relationship between increased 4-HNE in cultured hippocampal cells and altered Na+/K+ ATPase function (Tabrizi, Cleeter et al. 1999, Sonnen, Breitner et al. 2008, Zhou, Huang et al. 2008). PD patients show a reduced activity of antioxidant enzymes such as catalase and glutathione peroxidase and the most affected dopaminergic brain region presents a decrease in reduced glutathione (GSH) levels, so the ratio between GSH/GSSG (oxidized glutathione) decreases during neuronal degeneration, which favors the formation of free radicals (Borrás, Sastre et al. 2003, Zhou, Huang et al. 2008). Also, as in amyotrophic lateral sclerosis (ALS), there is a decrease in the activity of Complex I of the mitochondrial electron transport chain accompanied by up to 7-fold increase in O2, H2O2 and NO production (Barber and Shaw 2010).
The transcription factor Nrf2 is the intrinsic controller of cellular redox homeostasis and regulates the expression of a set of antioxidant genes called phase II genes, which produce phase II proteins associated to ROS stabilization. Under normal redox conditions, Nrf2 associates with the Keap1 protein, forming the Keap1-Nrf2 complex that is retained in the cytosol (cytoskeleton) (Cullinan, Gordan et al. 2004, Kobayashi, Kang et al. 2004, Sykiotis and Bohmann 2010). On the other hand, in an oxidizing environment, phosphatidylinositol 3-kinase (PI3K) produces depolymerization of actin microfilaments, realizing the Nrf2 from the cytoskeleton (Lee and Johnson 2004), facilitating its nuclear translocation and allowing its binding to a specific DNA sequence known as ARE (Antioxidant Response Element), producing enzymes of phase II as NQO1 (NADPH Quinone Dehydrogenase 1), glutathione S-transferase (GST) that conjugates ROS and hydrophobic electrophiles to glutathione, UDP-glucuronosyl transferases that conjugate xenobiotics to glucuronic acid for excretion, ferritin for iron storage, epoxide hydrolase that inactivates epoxides, and γ-glutamyl cysteine synthetase (γ-GCS) involved in GSH synthesis, heme oxygenase 1 (HO-1) that catabolizes heme groups, as well as exerting antioxidant and anti-inflammatory actions (Itoh, Tong et al. 2004, Loboda, Damulewicz et al. 2016).
CAPE shows potent antioxidant activity, reducing the formation of the superoxide anion produced during the autoxidation of β-mercaptoethanol and quenching the 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical. It also inhibited the activity of xanthine oxidase (XO). In addition, CAPE exhibits potent cytoprotective and antigenotoxic antilipoperoxidative potential against oxidative damage (Wang, Chen et al. 2008, Chen, Huang et al. 2009, Wang, Stavchansky et al. 2010). CAPE activates the expression of the antioxidant HO-1 and protects the kidneys against aging-related oxidative injury in rats, by reducing malondialdehyde (MDA) levels with a simultaneous elevation of superoxide dismutase, catalase, glutathione peroxidase and reduced glutathione in the renal tissues of old rats (Chen, Tran et al. 2012, Eşrefoğlu, Iraz et al. 2012). CAPE possesses a marked iron-binding capacity, therefore, interferes with the formation of the ferrous-ferrozin complex (Göçer and Gülçin 2011). Sun et al. demonstrated that CAPE at micromolar concentrations in a concentration-dependent manner is capable of increasing Nrf2 by inhibiting ubiquitination and promoting nuclear translocation of Nrf2 with subsequent HO-1 expression (Kim, Kim et al. 2013). Furthermore, they demonstrated that CAPE regulates NF-κB signaling and the inflammatory response through the Nrf2/HO-1 signaling pathway in IL-1β-stimulated chondrocytes (Sun, Xie et al. 2022).
CAPE attenuates oxidative stress and neurotoxicity, by modulating MAPK and Akt/glycogen synthase kinase 3β (GSK3β) signaling pathways, reducing hippocampal cells dead in an AD mouse model (Huang Y), thus is particularly beneficial in AD treatment due to Alzheimer's disease is closely related to toxicity in hippocampus. Moreover, it was found to protect transient ischemic injury and prevent neonatal encephalopathy, by blocking NF-κB-mediated neuronal inflammation, evoking antioxidant effects and mitochondrial apoptosis (Khan, Elango et al. 2007).
CAPE may be able to ameliorate Parkinson's disease, in PD murine models, attenuating dopaminergic loss and decreasing behavioral abnormality through oxidative stress and reducing neuroinflammation, causing a protective effect on nigral dopaminergic neurons by HO-1-dependent MAPK signaling (Fontanilla, Ma et al. 2011, Kurauchi, Hisatsune et al. 2012).
In an acute brain damage model of stroke, CAPE decreased MDA levels, prevented lipid peroxidation due to ROS scavenging, and increased GSH levels and NO in brain tissue (Aladag, Turkoz et al. 2006). When CAPE was compared with alpha-tocopherol, a potent free radical scavenger molecule (vitamin e) on ischemia-reperfusion brain injury, CAPE proved to be better than vitamin e in reducing the brain level of MDA, a marker of oxidative stress (Irmak, Fadillioglu et al. 2003). Moreover, CAPE is highly effective in scavenging free radicals, even more than their structurally related ferulic acid and ethyl ferulate, as well as in blocking the enhanced release of cardiolipin and cytochrome c, therefore it protects from functional alterations in isolated mouse brain mitochondria challenged by anoxia/reoxygenation (Feng, Lu et al. 2008). This compound further exhibits an effect on isolated brain mitochondria by directly inhibiting Ca2+-induced cytochrome c release, caspase-3, caspase-1, expression of inducible nitric oxide synthase by hypoxia-mediated, in vivo. Moreover, CAPE potently blocked neurotoxicity in vitro due to nitric oxide-induced (Wei, Zhao et al. 2004). The mechanisms of action of CAPE on oxidative stress and activation of the Nrf2 pathway are summarized in Figure 8.
6. CAPE Inhibits Neuroinflammation by Modulation of the NF-κB Pathway
Cumulating evidence supports that the transcription factor NF-κB plays a critical role in regulating the immune response and inflammation implicated in the pathogenesis of neurodegenerative disorders. The inflammatory response involves the activation of the innate and adaptive immune system, leading to the release of proinflammatory and anti-inflammatory cytokines, which play a crucial role in inflammation resolution. These systemic inflammatory responses are also manifested inside the brain's parenchyma, which is known as neuroinflammation. Furthermore, there is a particular inflammatory signature in the bloodstream and brain of patients with different neurodegenerative disorders, including AD, PD, ALS, and FTD. Interestingly, CAPE may be a relevant therapeutic agent for controlling NF-κB signaling and reducing neuroinflammation in these disorders.
NF-kB activation can be mediated through three pathways: canonical (or classical), the non-canonical (or alternative) and a non-enzymatic regulatory component. In the canonical pathway, NF-kB is activated in response to viral and microbial infections or exposure to pro-inflammatory cytokines. This occurs with the participation of RelA, and c-Rel dimers together with P50. After activation, the IKKB subunit phosphorylates IkB at serine residues, causing its ubiquitin-dependent degradation via proteasome, translocating NF-kB (p65/p50) to the nucleus to act as a transcription factor for target gene (Mincheva, Garcera et al. 2011). On the other hand, the alternative pathway is activated via members of the TNF-α family, such as lymphotoxin B and BAFF; CK2, independent of the IKK complex (IKKα and IKKβ). The pathway mainly activates IKKa, which is responsible for the phosphorylation of p100, causing proteolysis to give an active form, which is p52. The dimers can then be translocated into the nucleus, acting as transcription factors (Demchenko, Glebov et al. 2010).
Most of the genes under NF-κB transcriptional control are involved in immune system signaling and inflammatory responses. In fact, transcriptional control of cytokine expression by NF-κB is probably one of the most important factors when assessing the pathogenic role of NF-κB in some diseases. Some of these cytokines include TNFα, IL-1α/β, IL-2, 3, 6, 12, GM-CSF, M-CSF and G-CSF, the chemokines MCP-1, KC, MIP-1 and CCL together with adhesion molecules ICAM-1, E-selectin and VCAM-1, which enable the recruitment and binding of immune system cells to sites where the inflammatory process develops (Lai, Liu et al. 2017). NF-κB prevents apoptosis in several inflammatory cells, including macrophages, dendritic cells, T and B-lymphocytes, and neutrophils, and promotes survival of several malignant tumors, especially of lymphomas. In contrast, the inflammatory response can induce apoptotic cell death. This inflammatory cell death response is initiated by the production of cell death receptors (Fas and FasL) and intracellular apoptosis-inducing proteins. This apoptotic death is further assisted by activated immune cells, which secrete granzyme, perforin and nitric oxide, all apoptosis-inducing factors regulated by NF-κB (Lai, Liu et al. 2017).
The mammalian NF-κB family consists of five subfamilies: NF-κB1 (p105 and p50), NF-κB2 (p100 and p52), RelA (p65), RelB and c-Rel. In most cell types, NF-kB dimers are predominantly cytoplasmic because they are in constant interaction with NF-kB inhibitors (IkBα, IkBβ, IkBε and Bcl-3) remaining transcriptionally inactive (Karin and Ben-Neriah 2000). IkBα, IkBβ, IkBε interact with NF-kB dimers and are responsible for their retention in the cytoplasm. These NF-kB inhibitors contain conserved serine residues that can be phosphorylated by IkB kinases (or IKKs), to be subsequently degraded by the proteasome and finally generate the translocation of NF-kB dimers to the nucleus (Oeckinghaus and Ghosh 2009, Sun, Xie et al. 2022).
In 1996 for the first time, the molecular mechanisms by which caffeic acid derivatives prevent NF-kB activation were analyzed. Initially, it was shown that activation of NF-kB by tumor necrosis factor (TNF) is completely blocked by CAPE in a dose- and time-dependent manner. In addition to TNF, CAPE also inhibited NF-kB activation induced by other inflammatory agents, including phorbol ester, ceramide, hydrogen peroxide, and okadaic acid. However, reducing agents reverse inhibitory effects exerted by CAPE, suggesting a role for critical sulfhydryl groups in NF-kB activation (Natarajan, Singh et al. 1996). CAPE prevents translocation of the p65 subunit of NF-kB to the nucleus but has no significant effect on TNF-induced degradation of IκBα; however, it delays IκBα resynthesis. Therefore, these data suggest that CAPE prevented NF-κB activation by inhibiting p65 translocation and IκBα degradation in human chondrocytes (Sun, Xie et al. 2022).
The effect of CAPE on the inhibition of NF-kB binding to is target-DNA sites, as well as the binding to other transcription factors, including AP-1, Oct-1, and TFIID, has been found to be unaffected by CAPE (Natarajan, Singh et al. 1996). When several synthetic structural analogs of CAPE were tested, it was found that a bicyclic, rotationally restricted, 5,6-dihydroxy form exhibits high inhibitory activity, whereas the 6,7-dihydroxy variant is less active. In this sense, other investigations in which caffeic acid phenethyl ester (CAPE) and 3,4-dihydroxybenzalacetone (DBL), both phenylpropanoid derivatives containing catechol with and presenting diverse bioactivities, have been evaluated. It was determined that the proinflammatory effects on NO synthase expression was strongly suppressed by CAPE and, to a lesser extent, by DBL and caffeic acid ethyl ester. Thus, the induction of genes downstream of LPS-activated NF-κB, such as NO synthase, IL-1β and IL6, and translocation of NF-κB p65 to the nucleus were reduced, to a greater extent by CAPE than by DBL. Interestingly, phosphorylation of p65 was reduced by both compounds, especially by CAPE, even when the levels of IκB were not altered. One explanation for the action of these compounds is related to the effect of CAPE and DBL on the thiol groups of p65 which were modified by these molecules, however the inhibitory effects on p65 phosphorylation and nitrite production were reversed by pretreatment with thiol-containing reagents, suggesting that CAPE has strong inhibitory effects on NF-κB activation due to modification of thiol groups and phosphorylation of p65 (Natarajan, Singh et al. 1996).
In activated astroglia cells CAPE pretreatment abrogated TNF-α-induced expression of chemokine (CC motif) ligand 2 (CCL-2) and intercellular adhesion molecule 1 (ICAM-1) through inhibition of NF-κB activation in a cell-specific manner (Choi and Choi 2008). In another study CAPE was found to mediate up-regulation of Abcd2 expression and peroxisomal β-oxidation, this causes a decrease in very long chain fatty acid (VLCFA) levels in ABCD1-deficient U87 cells; VLCFA accumulation is characteristic of a neuroinflammatory disease associated with demyelination of the cerebral white matter known as: X-linked adrenoleukodystrophy (X-ALD). CAPE reduced NF-κB activation, iNOS expression and inflammatory cytokines in primary cultures of astrocytes derived from ABCD1 / ABCD2 silenced mice (Singh, Khan et al. 2013).
Microglial activation promotes to mediate inflammatory processes which are crucial in the development of neurodegenerative disorders. CAPE showed to inhibit cyclooxygenase-2 (COX-2) and subsequent NO production in both in vitro and in vivo models of microglial activation, mediated by α-adenosine monophosphate-activated protein kinase 5′ (AMPK), erythropoietin (EPO) and HO-1 (Tsai, Yu et al. 2017). It is known that in the case of Amyotrophic Lateral Sclerosis the mutation of superoxide dismutase (SOD) is highly correlated with its pathogenesis. A study using murine model showed that CAPE prolongs the survival of mice expressing mutant SOD1 (G93A) by attenuating neuroinflammation and motor neuron cell death as consequence of p38 phosphorylation reduction (Fontanilla, Wei et al. 2012). CAPE may exert its anti-inflammatory effects by inhibiting ROS production at the transcriptional level, through suppressing NF-κB activation and by directly inhibiting the catalytic activity of iNOS (Ilhan et al. 2004). Supposed mechanisms of CAPE actions through the NF-kB pathway are summarized in Figure 9.
7. Delivery Systems for Novel CAPE Formulations
The low solubility of CAPE and therefore its low bioavailability, limits its therapeutical use. These problems can be overcome by packing the drug in nanoparticles o vesicles, which provide better therapeutic effects at lower dose. Table 3 summarizes the main delivery systems that have been used with the aim of enhancing the pharmacological effects of CAPE.
8. Conclusions
Nowadays neurodegenerative diseases are a global concern with dramatically fast increasing and treatments still been palliatives. In this context functional food could be an alternative in the prevention or deceleration of neurodegenerative problems. It is interesting for the future treatment of AD that propolis has shown to potentiate the neuroprotective effect of memantine in preclinical models of AD (Moriguchi et al. 2022). This evidence supports a new potential avenue of investigation of a combination therapy of CAPE with other drugs to modify the evolution of AD. CAPE has a wide range of pharmacological activities, including neuroprotective effects by inhibition of NF-κB pathways and evoking the intrinsic cell protection by increasing the Nrf-2 pathway. The main objective of this review is to provide a comprehensive overview of the therapeutical potential of CAPE in the treatment of neurodegenerative disorders reviewing the published data about its neuroprotective effects in the last decade. We also shed light on its molecular mechanisms of action and summarize the structure-activity relationship of synthetic derivatives of CAPE, which will be useful for the further design and development of better derivatives.
Author Contributions
manuscript writing, R.P., V.B., V.M., J.O. L.O.; manuscript editing, A.C., H.U., I.G.-C., U.W., C.P.
Funding
Authors thank the financial support given by the support from ANID FONDECYT 1220831 and FAPESP 2018/07366-4.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Aladag, M.A.; Turkoz, Y.; Ozcan, C.; Sahna, E.; Parlakpinar, H.; Akpolat, N.; Cigremis, Y. Caffeic acid phenethyl ester (CAPE) attenuates cerebral vasospasm after experimental subarachnoidal haemorrhage by increasing brain nitric oxide levels. Int J Dev Neurosci 2006, 24, 9–14. [Google Scholar] [CrossRef]
- Almowallad, S.; Alqahtani, L.S.; Mobashir, M. NF-kB in Signaling Patterns and Its Temporal Dynamics Encode/Decode Human Diseases. Life (Basel) 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Ang, E.S.; Pavlos, N.J.; Chai, L.Y.; Qi, M.; Cheng, T.S.; Steer, J.H.; Joyce, D.A.; Zheng, M.H.; Xu, J. Caffeic acid phenethyl ester, an active component of honeybee propolis attenuates osteoclastogenesis and bone resorption via the suppression of RANKL-induced NF-kappaB and NFAT activity. J Cell Physiol 2009, 221, 642–649. [Google Scholar] [CrossRef]
- Argente, P.G. Phenolic profile of propolis from diferent geographical origins. Final masters thesis in food safety and quality management polytechnic. University of Valencia, 2019.
- Almowallad, S.; Alqahtani, L.S.; Mobashir, M. NF-kB in Signaling Patterns and Its Temporal Dynamics Encode/Decode Human Diseases. Life (Basel) 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.; Kunnumakkara, A.B.; Newman, R.A.; Aggarwal, B.B. Bioavailability of curcumin: problems and promises. Mol Pharm 2007, 4, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Borba, R.S.; Wilson, M.B.; Spivak, M. Hidden benefits of honeybee propolis in hives. Beekeeping–from science to practice 2017, 17–38. [CrossRef]
- Barber, S.C.; Shaw, P.J. Oxidative stress in ALS: key role in motor neuron injury and therapeutic target. Free Radic Biol Med 2010, 48, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Borrás, C.; JSastre; García-Sala, D. ; Lloret, A.; Pallardó, F.V.; Viña, J. Mitochondria from females exhibit higher antioxidant gene expression and lower oxidative damage than males. Free Radic Biol Med 2003, 34, 546–552. [Google Scholar] [CrossRef] [PubMed]
- Ceccatelli, S.; Tamm, C.; Zhang, Q.; Chen, M. Mechanisms and modulation of neural cell damage induced by oxidative stress. Physiol Behav 2007, 92, 87–92. [Google Scholar] [CrossRef]
- Celli, N.; Dragani, L.K.; Murzilli, S.; Pagliani, T.; Poggi, A. In vitro and in vivo stability of caffeic acid phenethyl ester, a bioactive compound of propolis. J Agric Food Chem 2007, 55, 3398–3407. [Google Scholar] [CrossRef] [PubMed]
- Chai, T.; Zhao, X.B.; Wang, W.F.; Qiang, Y.; Zhang, X.Y.; Yang, J.L. Design, Synthesis of N-phenethyl Cinnamide Derivatives and Their Biological Activities for the Treatment of Alzheimer's Disease: Antioxidant, Beta-amyloid Disaggregating and Rescue Effects on Memory Loss. Molecules 2018, 23. [Google Scholar] [CrossRef]
- Chan, G.C.; Cheung, K.W.; Sze, D.M. The immunomodulatory and anticancer properties of propolis. Clin Rev Allergy Immunol 2013, 44, 262–273. [Google Scholar] [CrossRef]
- Chen, C.; Kuo, Y.H.; Lin, C.C.; Chao, C.Y.; Pai, M.H.; Chiang, E.I.; Tang, F.Y. Decyl caffeic acid inhibits the proliferation of colorectal cancer cells in an autophagy-dependent manner in vitro and in vivo. PLoS One 2020, 15, e0232832. [Google Scholar] [CrossRef]
- Chen, H.; Tran, J.T.; Anderson, R.E.; Mandal, M.N. Caffeic acid phenethyl ester protects 661W cells from H2O2-mediated cell death and enhances electroretinography response in dim-reared albino rats. Mol Vis 2012, 18, 1325–1338. [Google Scholar] [PubMed]
- Chen, J.H.; Shao, Y.; Huang, M.T.; Chin, C.K.; Ho, C.T. Inhibitory effect of caffeic acid phenethyl ester on human leukemia HL-60 cells. Cancer Lett 1996, 108, 211–214. [Google Scholar] [CrossRef]
- Chen, M.J.; Chang, W.H.; Lin, C.C.; Liu, C.Y.; Wang, T.E.; Chu, C.H.; Shih, S.C.; Chen, Y.J. Caffeic acid phenethyl ester induces apoptosis of human pancreatic cancer cells involving caspase and mitochondrial dysfunction. Pancreatology 2008, 8, 566–576. [Google Scholar] [CrossRef]
- Chen, T.G.; Lee, J.J.; Lin, K.H.; Shen, C.H.; Chou, D.S.; Sheu, J.R. Antiplatelet activity of caffeic acid phenethyl ester is mediated through a cyclic GMP-dependent pathway in human platelets. Chin J Physiol 2007, 50, 121–126. [Google Scholar]
- Chen, Y.J.; Shiao, M.S.; Hsu, M.L.; Tsai, T.H.; Wang, S.Y. Effect of caffeic acid phenethyl ester, an antioxidant from propolis, on inducing apoptosis in human leukemic HL-60 cells. Journal of agricultural and food chemistry 2001, 49, 5615–5619. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Huang, A.C.; Chang, H.H.; Liao, H.F.; Jiang, C.M.; Lai, L.Y.; Chan, J.T.; Chen, Y.Y.; Chiang, J. Caffeic acid phenethyl ester, an antioxidant from propolis, protects peripheral blood mononuclear cells of competitive cyclists against hyperthermal stress. J Food Sci 2009, 74, H162–H167. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.C.; Chi, P.L.; Shen, M.C.; Shu, C.W.; Wann, S.R.; Liu, C.P.; Tseng, C.J.; Huang, W.C. Caffeic Acid Phenethyl Ester Rescues Pulmonary Arterial Hypertension through the Inhibition of AKT/ERK-Dependent PDGF/HIF-1α In Vitro and In Vivo. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Choi, S.S.; Kim, E.S.; Jedrychowski, M.P.; Yang, Y.R.; Jang, H.J.; Suh, P.G.; Banks, A.S.; Gygi, S.P.; Spiegelman, B.M. Thrap3 docks on phosphoserine 273 of PPARγ and controls diabetic gene programming. Genes Dev 2014, 28, 2361–2369. [Google Scholar] [CrossRef]
- Choi, K.; Choi, C. Differential regulation of c-Jun N-terminal kinase and NF-kappaB pathway by caffeic acid phenethyl ester in astroglial and monocytic cells. J Neurochem 2008, 105, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.H.; Lee, D.Y.; Kang, S.; Lee, M.K.; Lee, J.H.; Lee, S.H.; Lee, H.L.; Lee, H.Y.; Jeong, Y.I. Caffeic Acid Phenethyl Ester-Incorporated Radio-Sensitive Nanoparticles of Phenylboronic Acid Pinacol Ester-Conjugated Hyaluronic Acid for Application in Radioprotection. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.; Villegas, V.; Istre, H.; Heppler, B.; Gonzalez, N.; Brusman, N.; Snider, L.; Hogle, E.; Tucker, J.; Oñate, A.; Oñate, S.; Ma, L.; Paula, S. Synthesis and characterization of CAPE derivatives as xanthine oxidase inhibitors with radical scavenging properties. Bioorg Chem 2019, 86, 686–695. [Google Scholar] [CrossRef]
- Chu, Y.; Wu, P.Y.; Chen, C.W.; Lyu, J.L.; Liu, Y.J.; Wen, K.C.; Lin, C.Y.; Kuo, Y.H.; Chiang, H.M. Protective Effects and Mechanisms of N-Phenethyl Caffeamide from UVA-Induced Skin Damage in Human Epidermal Keratinocytes through Nrf2/HO-1 Regulation. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Cullinan, S.B.; Gordan, J.D.; Jin, J.; Harper, J.W.; Diehl, J.A. The Keap1-BTB protein is an adaptor that bridges Nrf2 to a Cul3-based E3 ligase: oxidative stress sensing by a Cul3-Keap1 ligase. Mol Cell Biol 2004, 24, 8477–8486. [Google Scholar] [CrossRef]
- Dawson, T.M.; Dawson, V.L. Nitric oxide synthase: role as a transmitter/mediator in the brain and endocrine system. Annu Rev Med 1996, 47, 219–227. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, D.M.; Sampaio, G.R.; Pinto, C.B.; Catharino, R.R.; Bastos, D.H.M. Bioavailability of chlorogenic acids in rats after acute ingestion of maté tea (Ilex paraguariensis) or 5-caffeoylquinic acid. Eur J Nutr 2017, 56, 2541–2556. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, J.S.; Monte-Alto-Costa, A. Caffeic acid phenethyl ester improves burn healing in rats through anti-inflammatory and antioxidant effects. J Burn Care Res 2013, 34, 682–688. [Google Scholar] [CrossRef]
- Dubois, B.; Villain, N.; Frisoni, G.B.; Rabinovici, G.D.; Sabbagh, M.; Cappa, S.; Bejanin, A.; Bombois, S.; Epelbaum, S.; Teichmann, M.; Habert, M.O.; Nordberg, A.; Blennow, K.; Galasko, D.; Stern, Y.; Rowe, C.C.; Salloway, S.; Schneider, L.S.; Cummings, J.L.; Feldman, H.H. Clinical diagnosis of Alzheimer's disease: recommendations of the International Working Group. Lancet Neurol 2021, 20, 484–496. [Google Scholar] [CrossRef] [PubMed]
- Dawson, T.M.; Dawson, V.L. Nitric oxide synthase: role as a transmitter/mediator in the brain and endocrine system. Annu Rev Med 1996, 47, 219–227. [Google Scholar] [CrossRef]
- Demchenko, Y.N.; Glebov, O.K.; Zingone, A.; Keats, J.J.; Bergsagel, P.L.; Kuehl, W.M. Classical and/or alternative NF-kappaB pathway activation in multiple myeloma. Blood 2010, 115, 3541–3552. [Google Scholar] [CrossRef] [PubMed]
- Drescher, N.; Klein, A.M.; Neumann, P.; Yañez, O.; Leonhardt, S.D. Inside Honeybee Hives: Impact of Natural Propolis on the Ectoparasitic Mite Varroa destructor and Viruses. Insects 2017, 8. [Google Scholar] [CrossRef]
- Erkkinen, M.G.; Kim, M.O.; Geschwind, M.D. Clinical Neurology and Epidemiology of the Major Neurodegenerative Diseases. Cold Spring Harb Perspect Biol 2018, 10. [Google Scholar]
- Ebadi, M.; Srinivasan, S.K.; Baxi, M.D. Oxidative stress and antioxidant therapy in Parkinson's disease. Prog Neurobiol 1996, 48, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Egan, M.F.; Kojima, M.; Callicott, J.H.; Goldberg, T.E.; Kolachana, B.S.; Bertolino, A.; Zaitsev, E.; Gold, B.; Goldman, D.; Dean, M.; Lu, B.; Weinberger, D.R. The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell 2003, 112, 257–269. [Google Scholar] [CrossRef]
- Eşrefoğlu, M.; Iraz, M.; Ateş, B.; Gül, M. Not only melatonin but also caffeic acid phenethyl ester protects kidneys against aging-related oxidative damage in Sprague Dawley rats. Ultrastruct Pathol 2012, 36, 244–251. [Google Scholar]
- Fathalipour, M.; Eghtedari, M.; Borges, F.; Silva, T.; Moosavi, F.; Firuzi, O.; Mirkhani, H. Caffeic Acid Alkyl Amide Derivatives Ameliorate Oxidative Stress and Modulate ERK1/2 and AKT Signaling Pathways in a Rat Model of Diabetic Retinopathy. Chem Biodivers 2019, 16, e1900405. [Google Scholar] [CrossRef]
- Feng, Y.; Lu, Y.W.; Xu, P.H.; Long, Y.; Wu, W.M.; Li, W.; Wang, R. Caffeic acid phenethyl ester and its related compounds limit the functional alterations of the isolated mouse brain and liver mitochondria submitted to in vitro anoxia-reoxygenation: relationship to their antioxidant activities. Biochim Biophys Acta 2008, 1780, 659–672. [Google Scholar] [CrossRef]
- Fontanilla, C.V.; Ma, Z.; Wei, X.; Klotsche, J.; Zhao, L.; Wisniowski, P.; Dodel, R.C.; Farlow, M.R.; Oertel, W.H.; Du, Y. Caffeic acid phenethyl ester prevents 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced neurodegeneration. Neuroscience 2011, 188, 135–141. [Google Scholar] [CrossRef]
- Fontanilla, C.V.; Wei, X.; Zhao, L.; Johnstone, B.; Pascuzzi, R.M.; Farlow, M.R.; Du, Y. Caffeic acid phenethyl ester extends survival of a mouse model of amyotrophic lateral sclerosis. Neuroscience 2012, 205, 185–193. [Google Scholar] [CrossRef]
- García Argente, P. (2018). "Perfil fenólico de propóleos de diferentes orígenes geográficos.".
- Göçer, H.; Gülçin, I. Caffeic acid phenethyl ester (CAPE): correlation of structure and antioxidant properties. Int J Food Sci Nutr 2011, 62, 821–825. [Google Scholar] [CrossRef]
- Gómez-Caravaca, A.M.; Gómez-Romero, M.; Arráez-Román, D.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Advances in the analysis of phenolic compounds in products derived from bees. Journal of pharmaceutical and biomedical analysis 2006, 41, 1220–1234. [Google Scholar]
- Freires, I.A.; Queiroz, V.; Furletti, V.F.; Ikegaki, M.; de Alencar, S.M.; Duarte, M.C.T.; Rosalen, P.L. Chemical composition and antifungal potential of Brazilian propolis against Candida spp. J Mycol Med 2016, 26, 122–132. [Google Scholar] [CrossRef]
- Gupta, V.K.; Fakhri, A.; Agarwal, S.; Ahmadi, E.; Nejad, P.A. Synthesis and characterization of MnO(2)/NiO nanocomposites for photocatalysis of tetracycline antibiotic and modification with guanidine for carriers of Caffeic acid phenethyl ester-an anticancer drug. J Photochem Photobiol B 2017, 174, 235–242. [Google Scholar] [CrossRef]
- Ha, J.; Choi, H.S.; Lee, Y.; Lee, Z.H.; Kim, H.H. Caffeic acid phenethyl ester inhibits osteoclastogenesis by suppressing NF kappaB and downregulating NFATc1 and c-Fos. Int Immunopharmacol 2009, 9, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Role of free radicals in the neurodegenerative diseases: therapeutic implications for antioxidant treatment. Drugs Aging 2001, 18, 685–716. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine; Oxford University Press: 2015.
- Hammer, B.; Parker, W.D., Jr.; Bennett, J.P., Jr. NMDA receptors increase OH radicals in vivo by using nitric oxide synthase and protein kinase C. Neuroreport 1993, 5, 72–74. [Google Scholar] [CrossRef] [PubMed]
- Hassan, N.A.; El-Bassossy, H.M.; Mahmoud, M.F.; Fahmy, A. Caffeic acid phenethyl ester, a 5-lipoxygenase enzyme inhibitor, alleviates diabetic atherosclerotic manifestations: effect on vascular reactivity and stiffness. Chem Biol Interact 2014, 213, 28–36. [Google Scholar] [CrossRef]
- Hayashi, M. Oxidative stress in developmental brain disorders. Neuropathology 2009, 29, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Heppner, F.L.; Ransohoff, R.M.; Becher, B. Immune attack: the role of inflammation in Alzheimer disease. Nat Rev Neurosci 2015, 16, 358–372. [Google Scholar] [CrossRef]
- Hernandez, J.; Goycoolea, F.M.; Quintero, J.; Acosta, A.; Castañeda, M.; Dominguez, Z.; Robles, R.; Vazquez-Moreno, L.; Velazquez, E.F.; Astiazaran, H.; Lugo, E. Sonoran propolis: chemical composition and antiproliferative activity on cancer cell lines. Planta medica 2007, 73, 1469–1474. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Jin, M.; Pi, R.; Zhang, J.; Chen, M.; Ouyang, Y.; Liu, A.; Chao, X.; Liu, P.; Liu, J.; Ramassamy, C. Protective effects of caffeic acid and caffeic acid phenethyl ester against acrolein-induced neurotoxicity in HT22 mouse hippocampal cells. Neurosci Lett 2013, 535, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Ilhan, A.; Akyol, O.; Gurel, A.; Armutcu, F.; Iraz, M.; Oztas, E. Protective effects of caffeic acid phenethyl ester against oxidative stress induced by experimental allergic encephalomyelitis in rats. Free Rad. Biol. Medicine 2004, 37, 386–394. [Google Scholar] [CrossRef]
- Irmak, M.K.; Fadillioglu, E.; Sogut, S.; Erdogan, H.; Gulec, M.; Ozer, M.; Yagmurca, M.; Gozukara, M.E. Effects of caffeic acid phenethyl ester and alpha-tocopherol on reperfusion injury in rat brain. Cell Biochem Funct 2003, 21, 283–289. [Google Scholar] [CrossRef]
- Itoh, K.; Tong, K.I.; Yamamoto, M. Molecular mechanism activating Nrf2-Keap1 pathway in regulation of adaptive response to electrophiles. Free Radic Biol Med 2004, 36, 1208–1213. [Google Scholar] [CrossRef] [PubMed]
- Jung, B.I.; Kim, M.S.; Kim, H.A.; Kim, D.; Yang, J.; Her, S.; Song, Y.S. Caffeic acid phenethyl ester, a component of beehive propolis, is a novel selective estrogen receptor modulator. Phytother Res 2010, 24, 295–300. [Google Scholar] [CrossRef]
- Kamat, C.D.; Gadal, S.; Mhatre, M.; Williamson, K.S.; Pye, Q.N.; Hensley, K. Antioxidants in central nervous system diseases: preclinical promise and translational challenges. J Alzheimers Dis 2008, 15, 473–493. [Google Scholar] [CrossRef]
- Kapare, H.S.; Lohidasan, S.; Sinnathambi, A.; Mahadik, K. Formulation Development of Folic Acid Conjugated PLGA Nanoparticles for Improved Cytotoxicity of Caffeic Acid Phenethyl Ester. Pharm Nanotechnol 2021, 9, 111–119. [Google Scholar] [CrossRef]
- Kazancioglu, H.O.; Aksakalli, S.; Ezirganli, S.; Birlik, M.; Esrefoglu, M.; Acar, A.H. Effect of caffeic acid phenethyl ester on bone formation in the expanded inter-premaxillary suture. Drug Des Devel Ther 2015, 9, 6483–6488. [Google Scholar] [CrossRef]
- Kazancioglu, H.O.; Bereket, M.C.; Ezirganli, S.; Aydin, M.S.; Aksakalli, S. Effects of caffeic acid phenethyl ester on wound healing in calvarial defects. Acta Odontol Scand 2015, 73, 21–27. [Google Scholar] [CrossRef]
- Khan, M.; Elango, C.; Ansari, M.A.; Singh, I.; Singh, A.K. Caffeic acid phenethyl ester reduces neurovascular inflammation and protects rat brain following transient focal cerebral ischemia. J Neurochem 2007, 102, 365–377. [Google Scholar] [CrossRef]
- Kim, H.; Kim, W.; Yum, S.; Hong, S.; Oh, J.-E.; Lee, J.-W.; Kwak, M.-K.; Park, E.J.; Na, D.H.; Jung, Y. Caffeic acid phenethyl ester activation of Nrf2 pathway is enhanced under oxidative state: Structural analysis and potential as a pathologically targeted therapeutic agent in treatment of colonic inflammation. Free Radical Biology and Medicine 2013, 65, 552–562. [Google Scholar]
- Kızıldağ, A.; Arabacı, T.; Albayrak, M.; Taşdemir, U.; Şenel, E.; Dalyanoglu, M.; Demirci, E. Therapeutic effects of caffeic acid phenethyl ester on alveolar bone loss in rats with endotoxin-induced periodontitis. J Dent Sci 2019, 14, 339–345. [Google Scholar] [CrossRef]
- Kobayashi, A.; Kang, M.I.; Okawa, H.; Ohtsuji, M.; Zenke, Y.; Chiba, T.; Igarashi, K.; Yamamoto, M. Oxidative stress sensor Keap1 functions as an adaptor for Cul3-based E3 ligase to regulate proteasomal degradation of Nrf2. Mol Cell Biol 2004, 24, 7130–7139. [Google Scholar] [CrossRef] [PubMed]
- Krol, W.; Scheller, S.; Shani, J.; Pietsz, G.; Czuba, Z. Synergistic effect of ethanolic extract of propolis and antibiotics on the growth of staphylococcus aureus. Arzneimittelforschung 1993, 43, 607–609. [Google Scholar] [PubMed]
- Kuo, Y.H.; Chen, C.W.; Chu, Y.; Lin, P.; Chiang, H.M. In Vitro and In Vivo Studies on Protective Action of N-Phenethyl Caffeamide against Photodamage of Skin. PLoS One 2015, 10, e0136777. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.H.; Chiang, H.L.; Wu, P.Y.; Chu, Y.; Chang, Q.X.; Wen, K.C.; Lin, C.Y.; Chiang, H.M. Protection against Ultraviolet A-Induced Skin Apoptosis and Carcinogenesis through the Oxidative Stress Reduction Effects of N-(4-bromophenethyl) Caffeamide, A Propolis Derivative. Antioxidants (Basel) 2020, 9. [Google Scholar] [CrossRef]
- Kurauchi, Y.; Hisatsune, A.; Isohama, Y.; Mishima, S.; Katsuki, H. Caffeic acid phenethyl ester protects nigral dopaminergic neurons via dual mechanisms involving haem oxygenase-1 and brain-derived neurotrophic factor. Br J Pharmacol 2012, 166, 1151–1168. [Google Scholar] [CrossRef]
- Kurek-Górecka, A.; Rzepecka-Stojko, A.; Górecki, M.; Stojko, J.; Sosada, M.; Świerczek-Zięba, G.J.M. Structure and antioxidant activity of polyphenols derived from propolis. Molecules 2013, 19, 78–101. [Google Scholar] [CrossRef]
- Karin, M.; Ben-Neriah, Y. Phosphorylation meets ubiquitination: the control of NF-[kappa]B activity. Annu Rev Immunol 2000, 18, 621–663. [Google Scholar] [CrossRef]
- Lai, J.L.; Liu, Y.H.; Liu, C.; Qi, M.P.; Liu, R.N.; Zhu, X.F.; Zhou, Q.G.; Chen, Y.Y.; Guo, A.Z.; Hu, C.M. Indirubin Inhibits LPS-Induced Inflammation via TLR4 Abrogation Mediated by the NF-kB and MAPK Signaling Pathways. Inflammation 2017, 40, 1–12. [Google Scholar] [CrossRef]
- Lee, H.Y.; Jeong, Y.I.; Kim, E.J.; Lee, K.D.; Choi, S.H.; Kim, Y.J.; Kim, D.H.; Choi, K.C. Preparation of caffeic acid phenethyl ester-incorporated nanoparticles and their biological activity. J Pharm Sci 2015, 104, 144–154. [Google Scholar] [CrossRef]
- Lee, J.M.; Johnson, J.A. An important role of Nrf2-ARE pathway in the cellular defense mechanism. J Biochem Mol Biol 2004, 37, 139–143. [Google Scholar] [CrossRef]
- Leiva, A.M.; Martínez-Sanguinetti, M.A.; Troncoso-Pantoja, C.; Nazar, G.; Petermann-Rocha, F.; Celis-Morales, C. [Parkinson's Disease in Chile: Highest Prevalence in Latin America]. Rev Med Chil 2019, 147, 535–536. [Google Scholar] [CrossRef]
- Li, K.; Tu, Y.; Liu, Q.; Ouyang, Y.; He, M.; Luo, M.; Chen, J.; Pi, R.; Liu, A. PT93, a novel caffeic acid amide derivative, suppresses glioblastoma cells migration, proliferation and MMP-2/-9 expression. Oncol Lett 2017, 13, 1990–1996. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Y.; Lee, C.F.; Wei, Y.H. Role of reactive oxygen species-elicited apoptosis in the pathophysiology of mitochondrial and neurodegenerative diseases associated with mitochondrial DNA mutations. J Formos Med Assoc 2009, 108, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: an evolutionarily conserved mechanism. Cell Mol Life Sci 2016, 73, 3221–3247. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Li, S.; Dai, D.; Zhang, Q.; Min, Z.; Yang, C.; Sun, S.; Ye, L.; Teng, C.; Cao, X.; Yin, H.; Lv, L.; Lv, W.; Xin, H. Enhanced treatment of cerebral ischemia-Reperfusion injury by intelligent nanocarriers through the regulation of neurovascular units. Acta Biomater 2022, 147, 314–326. [Google Scholar] [CrossRef]
- Markesbery, W.R.; Carney, J.M. Oxidative alterations in Alzheimer's disease. Brain Pathol 1999, 9, 133–146. [Google Scholar] [CrossRef]
- Marlatt, M.W.; Lucassen, P.J.; Perry, G.; Smith, M.A.; Zhu, X. Alzheimer's disease: cerebrovascular dysfunction, oxidative stress, and advanced clinical therapies. J Alzheimers Dis 2008, 15, 199–210. [Google Scholar] [CrossRef]
- Matsunaga, T.; Tsuchimura, S.; Azuma, N.; Endo, S.; Ichihara, K.; Ikari, A. Caffeic acid phenethyl ester potentiates gastric cancer cell sensitivity to doxorubicin and cisplatin by decreasing proteasome function. Anticancer Drugs 2019, 30, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Mei, Y.; Wang, Z.; Zhang, Y.; Wan, T.; Xue, J.; He, W.; Luo, Y.; Xu, Y.; Bai, X.; Wang, Q.; Huang, Y. FA-97, a New Synthetic Caffeic Acid Phenethyl Ester Derivative, Ameliorates DSS-Induced Colitis Against Oxidative Stress by Activating Nrf2/HO-1 Pathway. Front Immunol 2019, 10, 2969. [Google Scholar] [CrossRef]
- Metzner, J.; Bekemeier, H.; Paintz, M.; Schneidewind, E. [On the antimicrobial activity of propolis and propolis constituents (author's transl)]. Pharmazie 1979, 34, 97–102. [Google Scholar]
- Moosavi, F.; Hosseini, R.; Rajaian, H.; Silva, T.; Magalhães, E.S.D.; Saso, L.; Edraki, N.; Miri, R.; Borges, F.; Firuzi, O. Derivatives of caffeic acid, a natural antioxidant, as the basis for the discovery of novel nonpeptidic neurotrophic agents. Bioorg Med Chem 2017, 25, 3235–3246. [Google Scholar] [CrossRef] [PubMed]
- Machado, B.A.; Silva, R.P.; Gde, A.B.; Costa, S.S.; Silva, D.F.; Brandão, H.N.; Rocha, J.L.; Dellagostin, O.A.; Henriques, J.A.; Umsza-Guez, M.A.; Padilha, F.F. Chemical Composition and Biological Activity of Extracts Obtained by Supercritical Extraction and Ethanolic Extraction of Brown, Green and Red Propolis Derived from Different Geographic Regions in Brazil. PLoS One 2016, 11, e0145954. [Google Scholar] [CrossRef] [PubMed]
- Mincheva, S.; Garcera, A.; Gou-Fabregas, M.; Encinas, M.; Dolcet, X.; Soler, R.M. The canonical nuclear factor-κB pathway regulates cell survival in a developmental model of spinal cord motoneurons. J Neurosci 2011, 31, 6493–6503. [Google Scholar] [CrossRef] [PubMed]
- Moriguchi, S.; Inagaki, R.; Saito, T.; Saido, T.C.; Fukunaga, K. Propolis Promotes Memantine-Dependent Rescue of Cognitive Deficits in APP-KI Mice. Mol Neurobiol 2022, 59, 4630–4646. [Google Scholar] [CrossRef]
- Morroni, F.; Sita, G.; Graziosi, A.; Turrini, E.; Fimognari, C.; Tarozzi, A.; Hrelia, P. Neuroprotective Effect of Caffeic Acid Phenethyl Ester in A Mouse Model of Alzheimer's Disease Involves Nrf2/HO-1 Pathway. Aging Dis 2018, 9, 605–622. [Google Scholar] [CrossRef]
- Murtaza, G.; Sajjad, A.; Mehmood, Z.; Shah, S.H.; Siddiqi, A.R. Possible molecular targets for therapeutic applications of caffeic acid phenethyl ester in inflammation and cancer. J Food Drug Anal 2015, 23, 11–18. [Google Scholar] [CrossRef]
- Natarajan, K.; Singh, S.; Burke, T.R., Jr.; Grunberger, D.; Aggarwal, B.B. Caffeic acid phenethyl ester is a potent and specific inhibitor of activation of nuclear transcription factor NF-kappa B. Proc Natl Acad Sci USA 1996, 93, 9090–9095. [Google Scholar] [CrossRef]
- Nagaoka, T.; Banskota, A.H.; Tezuka, Y.; Saiki, I.; Kadota, S. Selective antiproliferative activity of caffeic acid phenethyl ester analogues on highly liver-metastatic murine colon 26-L5 carcinoma cell line. Bioorg Med Chem 2002, 10, 3351–3359. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, K.; Singh, S.; Burke, T.R., Jr.; Grunberger, D.; Aggarwal, B.B. Caffeic acid phenethyl ester is a potent and specific inhibitor of activation of nuclear transcription factor NF-kappa B. Proc Natl Acad Sci USA 1996, 93, 9090–9095. [Google Scholar] [CrossRef]
- Niu, Y.; Wang, K.; Zheng, S.; Wang, Y.; Ren, Q.; Li, H.; Ding, L.; Li, W.; Zhang, L. Antibacterial Effect of Caffeic Acid Phenethyl Ester on Cariogenic Bacteria and Streptococcus mutans Biofilms. Antimicrob Agents Chemother 2020, 64. [Google Scholar] [CrossRef]
- Omar, M.H.; Mullen, W.; Stalmach, A.; Auger, C.; Rouanet, J.M.; Teissedre, P.L.; Caldwell, S.T.; Hartley, R.C.; Crozier, A. Absorption, disposition, metabolism, and excretion of [3-(14)C]caffeic acid in rats. J Agric Food Chem 2012, 60, 5205–5214. [Google Scholar] [CrossRef] [PubMed]
- Ozdal, T.; Ceylan, F.D.; Eroglu, N.; Kaplan, M.; Olgun, E.O.; Capanoglu, E.J.F.R.I. Investigation of antioxidant capacity, bioaccessibility and LC-MS/MS phenolic profile of Turkish propolis. Food Research International 2019, 122, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Oeckinghaus, A.; Ghosh, S. The NF-kappaB family of transcription factors and its regulation. Cold Spring Harb Perspect Biol 2009, 1, a000034. [Google Scholar] [CrossRef]
- Pazin, W.M.; Monaco, L.D.M.; Soares, A.E.E.; Miguel, F.G.; Berretta, A.A.; Ito, A.S. Antioxidant activities of three stingless bee propolis and green propolis types. Journal of Apicultural Research 2017, 56, 40–49. [Google Scholar] [CrossRef]
- Przybyłek, I.; Karpiński, T.M. Antibacterial Properties of Propolis. Molecules 2019, 24. [Google Scholar] [CrossRef]
- Sies, H. Biochemistry of oxidative stress. Angewandte Chemie International Edition in English 1986, 25, 1058–1071. [Google Scholar] [CrossRef]
- Pittalà, V.; Salerno, L.; Romeo, G.; Acquaviva, R.; Di Giacomo, C.; Sorrenti, V. Therapeutic Potential of Caffeic Acid Phenethyl Ester (CAPE) in Diabetes. Curr Med Chem 2018, 25, 4827–4836. [Google Scholar] [CrossRef]
- Colpan, R.D.; Erdemir, A. Co-delivery of quercetin and caffeic acid phenethyl ester by polymeric nanoparticles to enhance antitumor efficacy in colon cancer cells. Journal of Microencapsulation 2021, 38, 381–393. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free radical biology and medicine 1996, 20, 933–956. [Google Scholar] [CrossRef] [PubMed]
- Romero, M.; Freire, J.; Pastene, E.; García, A.; Aranda, M.; González, C. Propolis polyphenolic compounds affect the viability and structure of Helicobacter pylori in vitro. Revista Brasileira de Farmacognosia 2019, 29, 325–332. [Google Scholar] [CrossRef]
- Roos, T.U.; Heiss, E.H.; Schwaiberger, A.V.; Schachner, D.; Sroka, I.M.; Oberan, T.; Vollmar, A.M.; Dirsch, V.M. Caffeic acid phenethyl ester inhibits PDGF-induced proliferation of vascular smooth muscle cells via activation of p38 MAPK, HIF-1α, and heme oxygenase-1. J Nat Prod 2011, 74, 352–356. [Google Scholar] [CrossRef]
- Sapolsky, R.M. Why stress is bad for your brain. Science 1996, 273, 749–750. [Google Scholar] [CrossRef]
- Scheller, S.; Dworniczak, S.; Waldemar-Klimmek, K.; Rajca, M.; Tomczyk, A.; Shani, J. Synergism between ethanolic extract of propolis (EEP) and anti-tuberculosis drugs on growth of mycobacteria. Z Naturforsch C J Biosci 1999, 54, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Serarslan, G.; Altuğ, E.; Kontas, T.; Atik, E.; Avci, G. Caffeic acid phenethyl ester accelerates cutaneous wound healing in a rat model and decreases oxidative stress. Clin Exp Dermatol 2007, 32, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Yamashita, A.; Nakakoshi, M.; Yokoe, H.; Sudo, M.; Kasai, H.; Tanaka, T.; Fujimoto, Y.; Ikeda, M.; Kato, N.; Sakamoto, N.; Shindo, H.; Maekawa, S.; Enomoto, N.; Tsubuki, M.; Moriishi, K. Inhibitory effects of caffeic acid phenethyl ester derivatives on replication of hepatitis C virus. PLoS One 2013, 8, e82299. [Google Scholar] [CrossRef]
- Singh, J.; Khan, M.; Singh, I. Caffeic acid phenethyl ester induces adrenoleukodystrophy (Abcd2) gene in human X-ALD fibroblasts and inhibits the proinflammatory response in Abcd1/2 silenced mouse primary astrocytes. Biochim Biophys Acta 2013, 1831, 747–758. [Google Scholar] [CrossRef]
- Song, M.K.; Cho, A.R.; Sim, G.; Ahn, J.H. Synthesis of Diverse Hydroxycinnamoyl Phenylethanoid Esters Using Escherichia coli. J Agric Food Chem 2019, 67, 2028–2035. [Google Scholar] [CrossRef]
- Sonnen, J.A.; Breitner, J.C.; Lovell, M.A.; Markesbery, W.R.; Quinn, J.F.; Montine, T.J. Free radical-mediated damage to brain in Alzheimer's disease and its transgenic mouse models. Free Radic Biol Med 2008, 45, 219–230. [Google Scholar] [CrossRef]
- Sorrenti, V.; Raffaele, M.; Vanella, L.; Acquaviva, R.; Salerno, L.; Pittalà, V.; Intagliata, S.; Di Giacomo, C. Protective Effects of Caffeic Acid Phenethyl Ester (CAPE) and Novel Cape Analogue as Inducers of Heme Oxygenase-1 in Streptozotocin-Induced Type 1 Diabetic Rats. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Sun, W.; Xie, W.; Huang, D.; Cui, Y.; Yue, J.; He, Q.; Jiang, L.; Xiong, J.; Sun, W.; Yi, Q. Caffeic acid phenethyl ester attenuates osteoarthritis progression by activating NRF2/HO-1 and inhibiting the NF-κB signaling pathway. Int J Mol Med 2022, 50. [Google Scholar] [CrossRef] [PubMed]
- Sykiotis, G.P.; Bohmann, D. Stress-activated cap'n'collar transcription factors in aging and human disease. Sci Signal 2010, 3, re3. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Xie, W.; Huang, D.; Cui, Y.; Yue, J.; He, Q.; Jiang, L.; Xiong, J.; Sun, W.; Yi, Q. Caffeic acid phenethyl ester attenuates osteoarthritis progression by activating NRF2/HO-1 and inhibiting the NF-κB signaling pathway. Int J Mol Med 2022, 50. [Google Scholar] [CrossRef] [PubMed]
- Tiveron, A.P.; Rosalen, P.L.; Franchin, M.; Lacerda, R.C.; Bueno-Silva, B.; Benso, B.; Denny, C.; Ikegaki, M.; Alencar, S.M. Chemical Characterization and Antioxidant, Antimicrobial, and Anti-Inflammatory Activities of South Brazilian Organic Propolis. PLoS One 2016, 11, e0165588. [Google Scholar] [CrossRef]
- Tonda-Turo, C.; Origlia, N.; Mattu, C.; Accorroni, A.; Chiono, V. Current Limitations in the Treatment of Parkinson's and Alzheimer's Diseases: State-of-the-Art and Future Perspective of Polymeric Carriers. Curr Med Chem 2018, 25, 5755–5771. [Google Scholar] [CrossRef] [PubMed]
- Tabrizi, S.J.; Cleeter, M.W.; Xuereb, J.; Taanman, J.W.; Cooper, J.M.; Schapira, A.H. Biochemical abnormalities and excitotoxicity in Huntington's disease brain. Ann Neurol 1999, 45, 25–32. [Google Scholar] [CrossRef]
- Tolba, M.F.; El-Serafi, A.T.; Omar, H.A. Caffeic acid phenethyl ester protects against glucocorticoid-induced osteoporosis in vivo: Impact on oxidative stress and RANKL/OPG signals. Toxicol Appl Pharmacol 2017, 324, 26–35. [Google Scholar] [CrossRef]
- Tomur, A.; Kanter, M.; Gurel, A.; Erboga, M. The efficiency of CAPE on retardation of hepatic fibrosis in biliary obstructed rats. J Mol Histol 2011, 42, 451–458. [Google Scholar] [CrossRef]
- Tsai, T.H.; Yu, C.H.; Chang, Y.P.; Lin, Y.T.; Huang, C.J.; Kuo, Y.H.; Tsai, P.J. Protective Effect of Caffeic Acid Derivatives on tert-Butyl Hydroperoxide-Induced Oxidative Hepato-Toxicity and Mitochondrial Dysfunction in HepG2 Cells. Molecules 2017, 22. [Google Scholar] [CrossRef] [PubMed]
- Veloz, J.J.; Alvear, M.; Salazar, L.A. Antimicrobial and Antibiofilm Activity against Streptococcus mutans of Individual and Mixtures of the Main Polyphenolic Compounds Found in Chilean Propolis. Biomed Res Int 2019, 2019, 7602343. [Google Scholar] [CrossRef]
- Wan, T.; Wang, Z.; Luo, Y.; Zhang, Y.; He, W.; Mei, Y.; Xue, J.; Li, M.; Pan, H.; Li, W.; Wang, Q.; Huang, Y. FA-97, a New Synthetic Caffeic Acid Phenethyl Ester Derivative, Protects against Oxidative Stress-Mediated Neuronal Cell Apoptosis and Scopolamine-Induced Cognitive Impairment by Activating Nrf2/HO-1 Signaling. Oxid Med Cell Longev 2019, 2019, 8239642. [Google Scholar] [CrossRef]
- Wang, J.; Bhargava, P.; Yu, Y.; Sari, A.N.; Zhang, H.; Ishii, N.; Yan, K.; Zhang, Z.; Ishida, Y.; Terao, K.; Kaul, S.C.; Miyako, E.; Wadhwa, R. Novel Caffeic Acid Phenethyl Ester-Mortalin Antibody Nanoparticles Offer Enhanced Selective Cytotoxicity to Cancer Cells. Cancers (Basel) 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Chen, L.; Wu, W.; Long, Y.; Wang, R. Potential cytoprotection: antioxidant defence by caffeic acid phenethyl ester against free radical-induced damage of lipids, DNA, and proteins. Can J Physiol Pharmacol 2008, 86, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Stavchansky, S.; Kerwin, S.M.; Bowman, P.D. Structure-activity relationships in the cytoprotective effect of caffeic acid phenethyl ester (CAPE) and fluorinated derivatives: effects on heme oxygenase-1 induction and antioxidant activities. Eur J Pharmacol 2010, 635, 16–22. [Google Scholar] [CrossRef]
- Watabe, M.; Hishikawa, K.; Takayanagi, A.; Shimizu, N.; Nakaki, T.J.J.O.B.C. Caffeic acid phenethyl ester induces apoptosis by inhibition of NFκB and activation of Fas in human breast cancer MCF-7 cells. Journal of Biological Chemistry 2004, 279, 6017–6026. [Google Scholar] [CrossRef]
- Wei, X.; Zhao, L.; Ma, Z.; Holtzman, D.M.; Yan, C.; Dodel, R.C.; Hampel, H.; Oertel, W.; Farlow, M.R.; Du, Y. Caffeic acid phenethyl ester prevents neonatal hypoxic-ischaemic brain injury. Brain 2004, 127 Pt 12, 2629–2635. [Google Scholar] [CrossRef]
- Xiang, D.; Wang, D.; He, Y.; Xie, J.; Zhong, Z.; Li, Z.; Xie, J. Caffeic acid phenethyl ester induces growth arrest and apoptosis of colon cancer cells via the β-catenin/T-cell factor signaling. Anti-cancer drugs 2006, 17, 753–762. [Google Scholar] [CrossRef]
- Yasui, N.; Nishiyama, E.; Juman, S.; Negishi, H.; Miki, T.; Yamori, Y.; Ikeda, K. Caffeic acid phenethyl ester suppresses oxidative stress in 3T3-L1 adipocytes. J Asian Nat Prod Res 2013, 15, 1189–1196. [Google Scholar] [CrossRef]
- Yordanov, Y.; Aluani, D.; Tzankova, V.; Rangelov, S.; Odzhakov, F.; Apostolov, A.; Yoncheva, K. Safety assessment of a newly synthesized copolymer for micellar delivery of hydrophobic caffeic acid phenethyl ester. Pharmaceutical Development and Technology 2020, 25, 1271–1280. [Google Scholar] [CrossRef] [PubMed]
- Zawawi, M.S.; Perilli, E.; Stansborough, R.L.; Marino, V.; Cantley, M.D.; Xu, J.; Dharmapatni, A.A.; Haynes, D.R.; Gibson, R.J.; Crotti, T.N. Caffeic acid phenethyl ester abrogates bone resorption in a murine calvarial model of polyethylene particle-induced osteolysis. Calcif Tissue Int 2015, 96, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Huang, Y.; Przedborski, S. Oxidative stress in Parkinson's disease: a mechanism of pathogenic and therapeutic significance. Ann N Y Acad Sci 2008, 1147, 93–104. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chemical structure of caffeic acid phenethyl ester (CAPE).
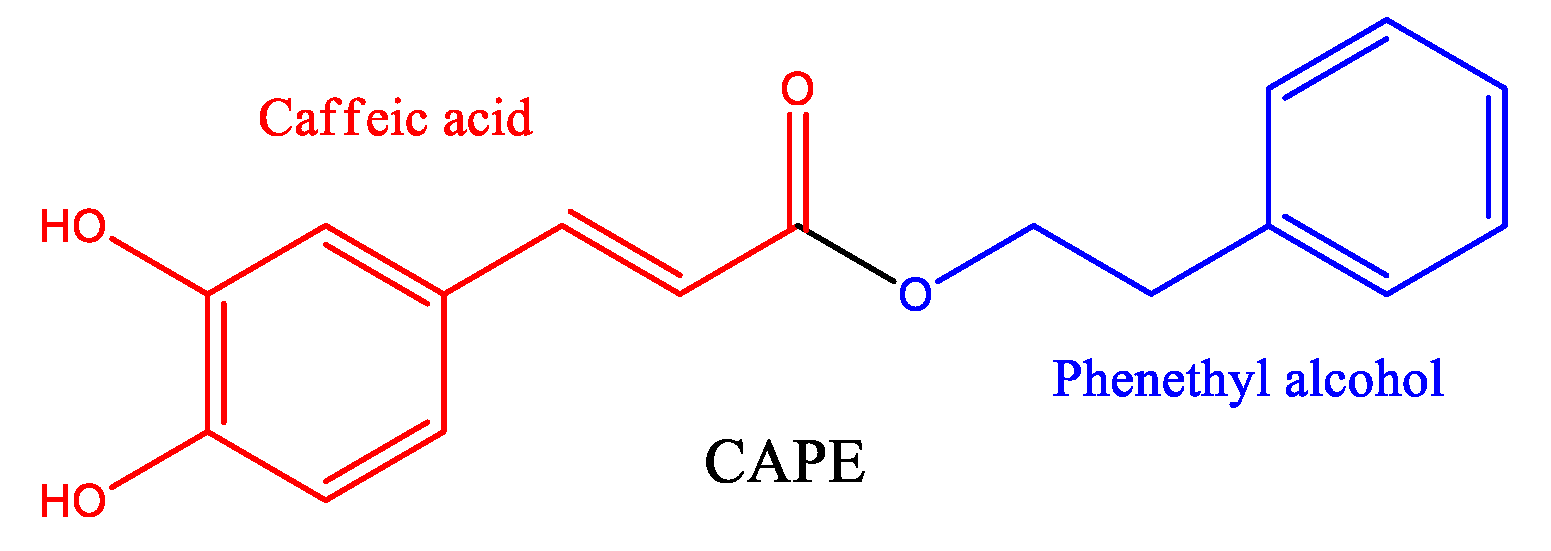
Figure 2.
Biosynthetic pathways for the formation of CAPE. The involved enzymes are abbreviated: PAL = phenylalanine ammonia lyase; C4H = cinnamate 4-hydroxilase; 4CL = 4-coumaric acid CoA-ligase; C3H = p-coumarate 3-hydroxylase.
Figure 2.
Biosynthetic pathways for the formation of CAPE. The involved enzymes are abbreviated: PAL = phenylalanine ammonia lyase; C4H = cinnamate 4-hydroxilase; 4CL = 4-coumaric acid CoA-ligase; C3H = p-coumarate 3-hydroxylase.
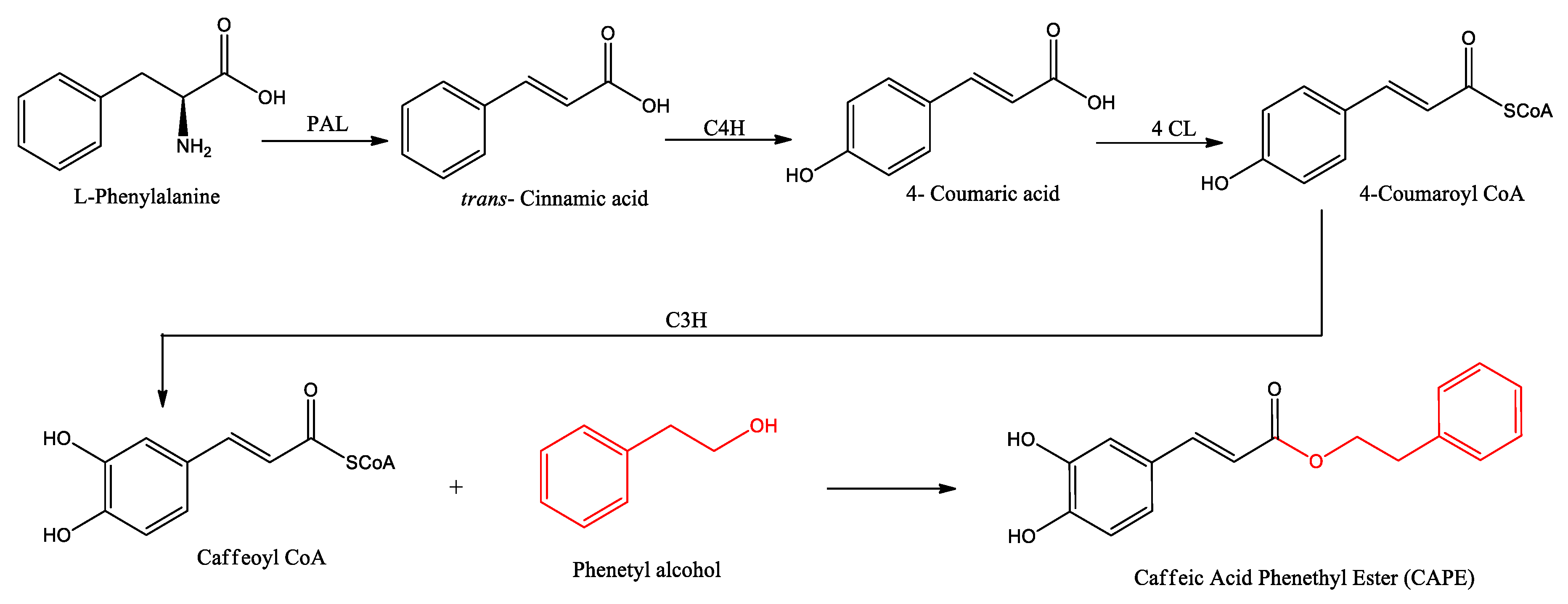
Figure 3.
Synthesis of CAPE and derivatives by Knoevenagel condensation using thermal or microwave methods.
Figure 3.
Synthesis of CAPE and derivatives by Knoevenagel condensation using thermal or microwave methods.
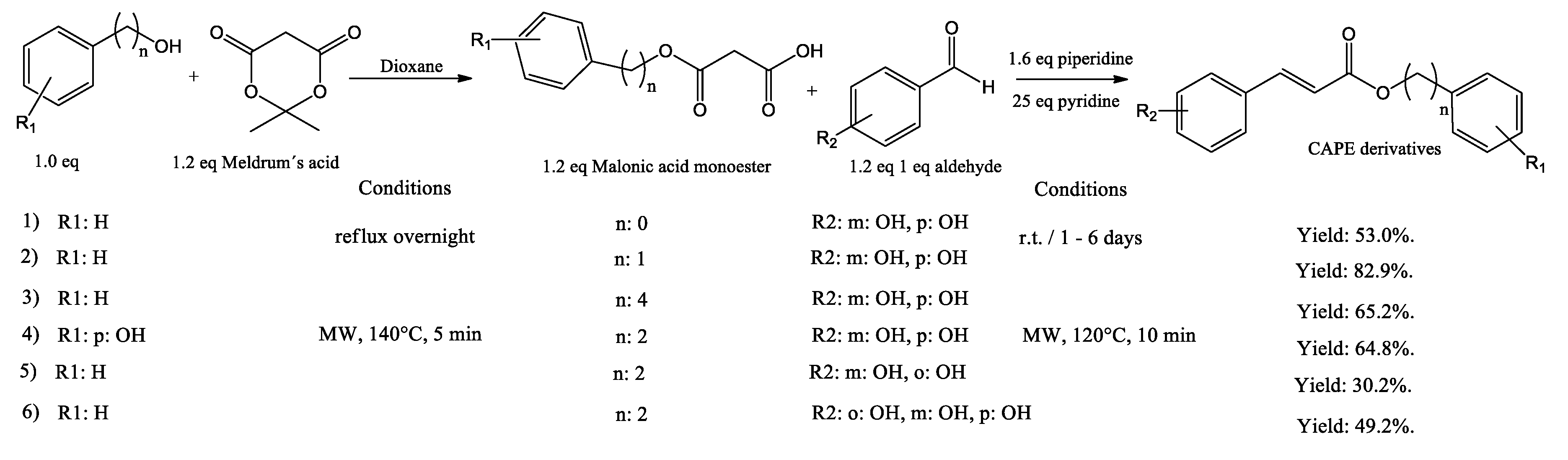
Figure 4.
Synthesis of CAPE derivatives by DCC and acyl chlorine condensation.
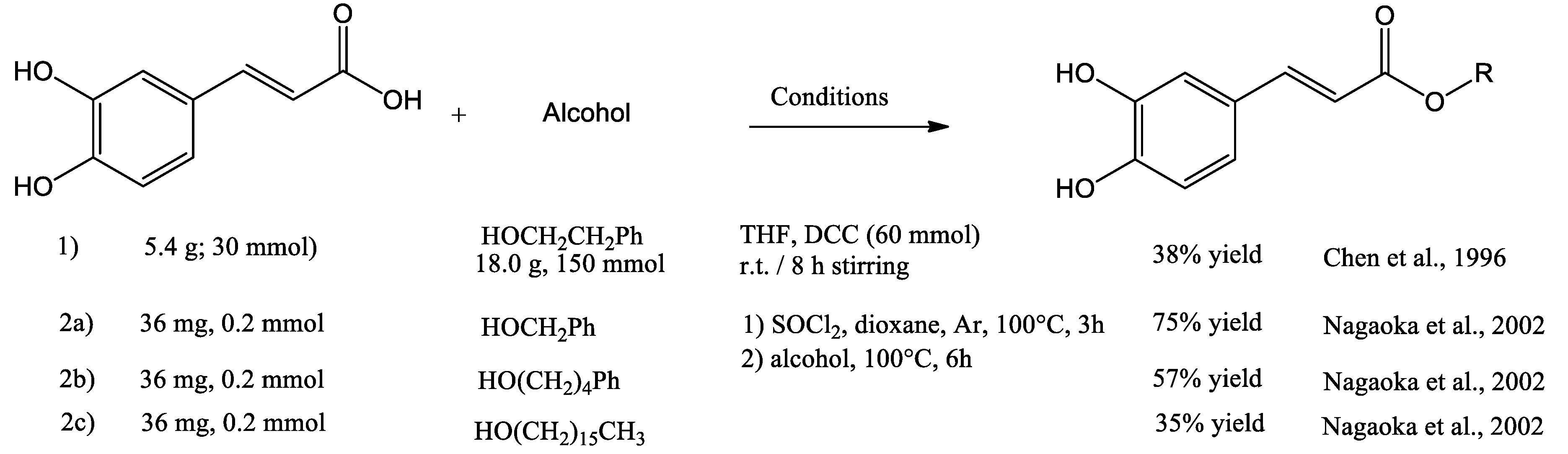
Figure 5.
chemical structure of caffeoyl esters derivatives.
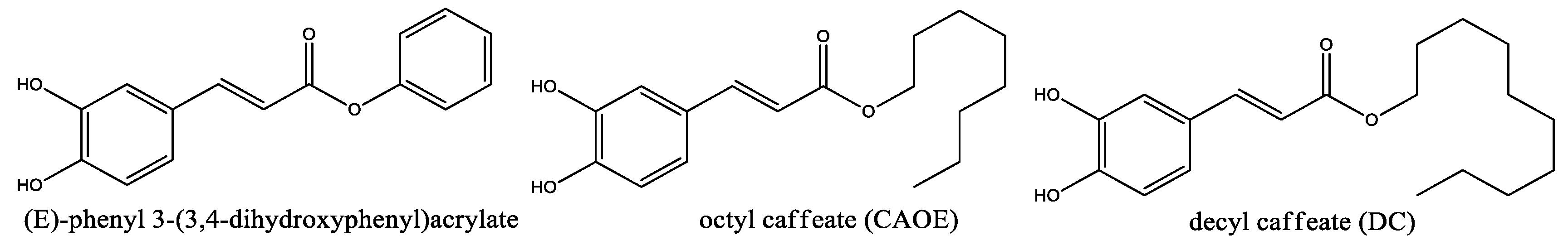
Figure 6.
Chemical structure of caffeoyl amides derivatives.
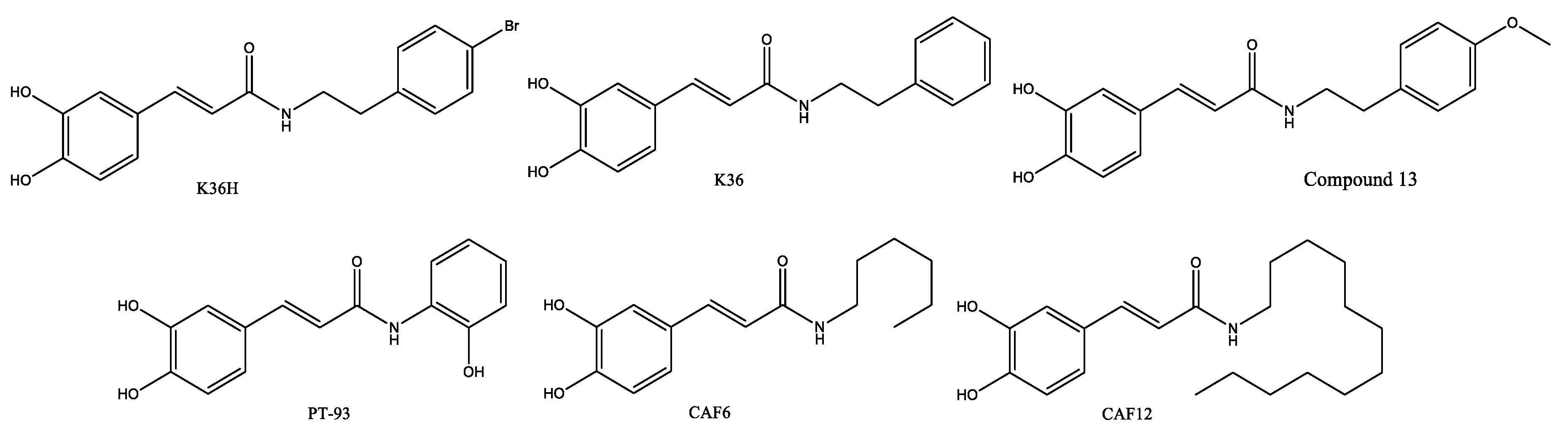
Figure 7.
Synthesis of caffeic acid phenethyl ester 4-O-glucoside, or FA-97 (Wan, Wang et al. 2019).
Figure 7.
Synthesis of caffeic acid phenethyl ester 4-O-glucoside, or FA-97 (Wan, Wang et al. 2019).
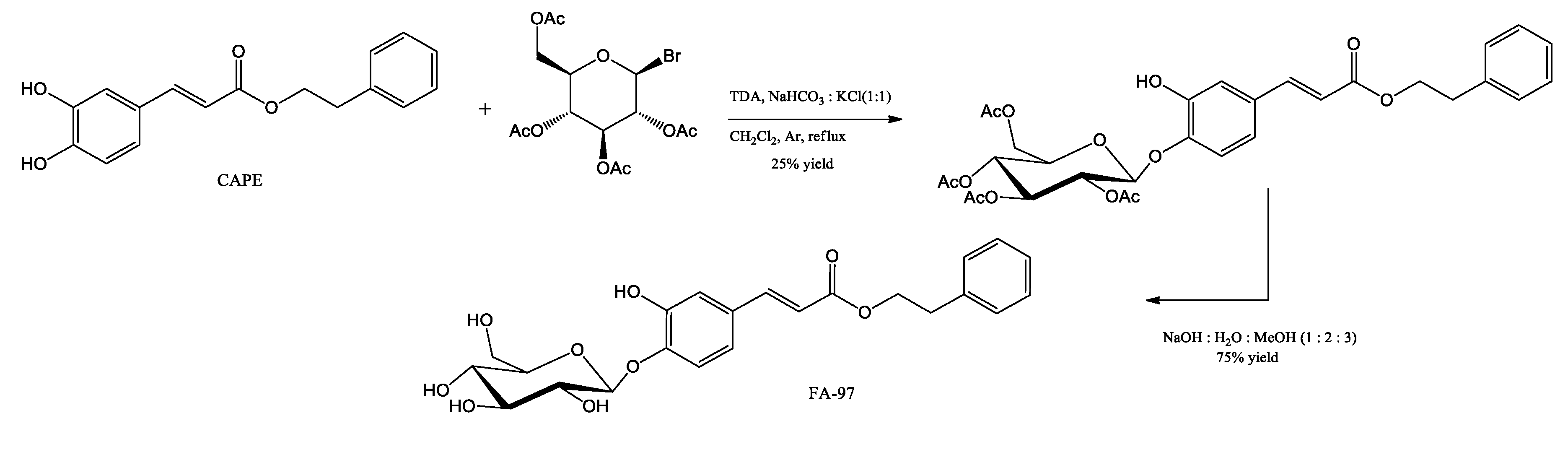
Figure 8.
CAPE action mechanisms on oxidative stress and activation of the Nrf2 pathway. (-) routes that inactivate. (+) activating routes.
Figure 8.
CAPE action mechanisms on oxidative stress and activation of the Nrf2 pathway. (-) routes that inactivate. (+) activating routes.
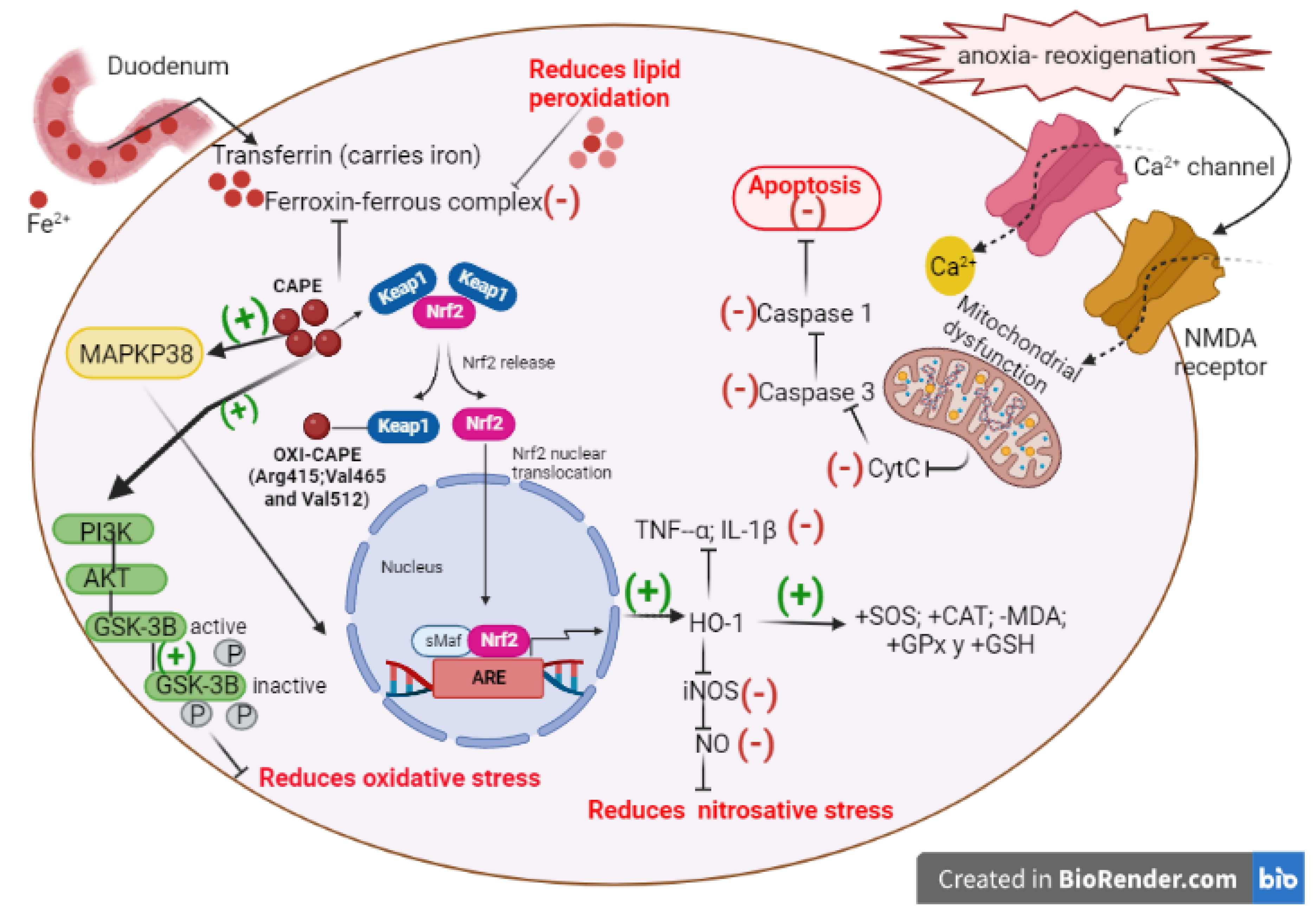
Figure 9.
Mechanism of action of CAPE against neuroinflammation and NF-κB pathway activation. (-) routes that inactivate. (+) activating routes.
Figure 9.
Mechanism of action of CAPE against neuroinflammation and NF-κB pathway activation. (-) routes that inactivate. (+) activating routes.
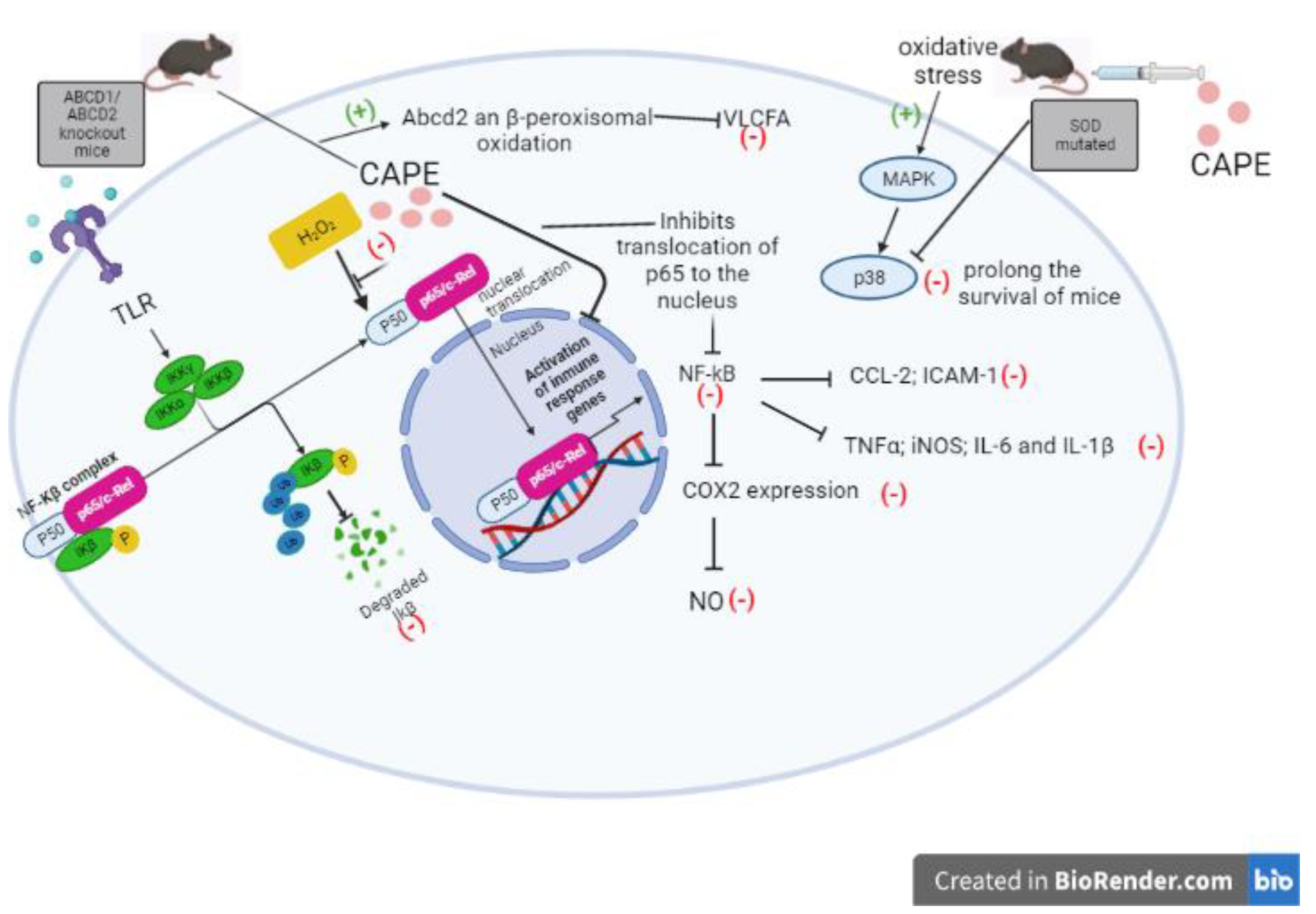

Table 1.
Biological properties of CAPE.
| Pharmacological effect | Molecular mechanism | Reference |
|---|---|---|
| Wound repair | CAPE promotes early inflammatory response (increased NOS2, TNF-α, and NF-κB) associated with a short-term event, leading to fast skin wound healing and inhibition of inflammation. Significant increase in the glutathione (GSH) level, an endogenous antioxidant that plays a key role in cellular defense against oxidative stress. Considerable decrease in malondialdehyde (MDA) and superoxide dismutase activity. | (Serarslan, Altuğ et al. 2007, dos Santos and Monte-Alto-Costa 2013) |
|
Antidiabetic Properties |
CAPE is a heme oxygenase 1 (HO-1) inducer, combating the increase of reactive oxygen species (ROS), induced by hyperglycemia. It inhibits the 5-lipoxygenase, alleviating diabetic atherosclerotic symptoms (an important macrovascular complication of diabetes). It restores adipocyte function by increasing adiponectin and PPARγ (their activation leads to improved insulin sensitivity), leading the reduction of proinflammatory factors. | (Choi, Choi et al. 2014, Hassan, El-Bassossy et al. 2014, Pittalà, Salerno et al. 2018, Sorrenti, Raffaele et al. 2019) |
| Anticancer properties | Inhibition of DNA synthesis, disruption of growth signal transmission, induction of apoptosis through an internal apoptotic pathway and promotion of anti-angiogenic effects. It enhances the anti-cancer effect of first-line chemotherapy drugs, an example being the drug paclitaxel in a rat model of DMBA-induced breast cancer, resulting in lower tumor weights compared to those with paclitaxel alone. CAPE protects normal cells against the effects of anticancer drugs by acting as a chemopreventive agent, in the case of the drug irinotecan it protects normal blood, liver and kidney cells without affecting the cytotoxicity of irinotecan in the in vivo model of Ehrlich ascites tumor cells. |
(Natarajan, Singh et al. 1996, Chen, Chang et al. 2008, Roos, Heiss et al. 2011, Chan, Cheung et al. 2013, Yasui, Nishiyama et al. 2013, Matsunaga, Tsuchimura et al. 2019) |
| Periodontitis treatment | It improves bone healing, preventing RANKL-induced osteoclastogenesis (causes follicular bone destruction), suggesting its use as a regenerative agent in therapy of bone resorption. | (Kazancioglu, Aksakalli et al. 2015, Kazancioglu, Bereket et al. 2015, Kızıldağ, Arabacı et al. 2019) |
|
Antibacterial activity |
High activity against Mycobactrium tuberculosis, Mycobacterium avium, Streptococcus piogenes Klebsiella pneumoniae and Staphylococcus epidermidis. It presents synergistic activity with many antituberculosis drugs, including rifamycin, streptomycin, isoniazid and antibiotics as gentamicin, tetracycline, chloramphenicol, vancomycin, clindamycin, netilmicin. CAPE Inhibits biofilm formation and lactic acid and extracellular polysaccharide production in S. mutans. | (Krol, Scheller et al. 1993, Scheller, Dworniczak et al. 1999, Veloz, Alvear et al. 2019, Niu, Wang et al. 2020) |
|
Antiatherogenic effects |
In human platelets CAPE (15 and 25 µM) markedly inhibited platelet aggregation stimulated by collagen (2 µg/ml). |
(Chen, Lee et al. 2007) |
|
Estrogenic effects |
CAPE is a selective agonist of ER-β estrogen receptors (present in the lungs, blood vessels, brain and bones). | (Jung, Kim et al. 2010) |
| Hepatoprotective effects | In liver CAPE elevates tissue catalase (CAT) activity and ameliorates ultrastructural changes associated with aging, also gives protection against necrosis, lipid peroxidation, aberrant cell proliferation and p65 activation (decreased number of preneoplastic nodules and reduced incidence of liver tumors). Reduced malondialdehyde (MDA) levels and increased activities of glutathione peroxidase (GPx) together with superoxide dismutase (SOD) in liver tissue. | (Tomur, Kanter et al. 2011, Eşrefoğlu, Iraz et al. 2012) |
| Lung protection | Effects on pulmonary arterial hypertension and fibrosis by HIF-1α/platelet-derived growth factor (PDGF)-dependent Akt/ERK pathways. | (Cheng, Chi et al. 2019) |
|
Protection of the bone diseases |
CAPE induce osteoclast apoptosis, antioxidant effects and modulation of osteoprotegrin signaling pathways, by suppressing NF-κB activity, CAPE significantly inhibits osteoclastogenesis and osteoclast differentiation. |
(Ang, Pavlos et al. 2009, Ha, Choi et al. 2009, Zawawi, Perilli et al. 2015, Tolba, El-Serafi et al. 2017) |
| Alzheimer’s disease | CAPE significantly inhibits neuronal apoptosis and neuroinflammation, induces Nrf2 activation, inhibits glycogen synthase kinase 3β in the hippocampus, and improves learning and memory and cognition. | (Morroni, Sita et al. 2018) |
Table 3.
Bioactive formulation of CAPE.
| Formulation | Target | Results | Reference |
|---|---|---|---|
| Poly(lactic-co-glycolic acid) (PLGA) co-loaded nanoparticles (QuCaNP) quercetin and CAPE. | Improving anticancer efficacy in HT-29 human colorectal carcinoma. | Increased caspase-3 (2.38-fold) and caspase-9 (2-fold) mRNA levels and expressions of key proteins in the intrinsic apoptosis pathway in HT-29 cells. | (Dilsu and Erdemir 2021) |
| Copolymer: polyglycerol and poly(allyl glycidyl ether) (C12-PAGE-PG), loaded with CAPE | To evaluate the in vitro and in vivo safety of a CAPE-loaded micellar system as a drug delivery platform on HepG2 cells. | Empty micelles loaded with CAPE showed no cytotoxic effects and retained the cytotoxic activity of CAPE loaded in the micelles, making it a good strategy to use this hydrophobic compound and improve the effectiveness of the treatments. | (Yordanov et al. 2020) |
| NiO nanoparticles and MnO2 /NiO nanocomposites with guanidine and CAPE as a carrier | Evaluate its capacity as an anchoring method for drug carriers. | The drug loading time was 100 min and drug release in 1-10 h with 20-80 % drug release. | (Gupta, Fakhri et al. 2017) |
| Hyaluronic acid (HA) conjugated with phenylboronic acid pinacol ester (PBPE) with radiosensitive delivery of CAPE | Manufacture radiosensitive delivery of CAPE for application in radioprotection. | Prevention of radiation-induced apoptosis and intracellular ROS accumulation, with increased survivability of mice against radiation-induced death. | (Choi, Lee et al. 2021) |
| Methoxy poly(ethylene glycol)-b -poly(ε-caprolactone) copolymer nanoparticles (CE) with CAPE. | Study antitumor activity against lung metastasis by CT26 colon carcinoma cells. | Superior anti-metastatic efficacy against the tumor than CAPE itself. | (Lee, Jeong et al. 2015) |
|
Caffeic acid phenethyl ester-morphthalin antibody nanoparticles |
Generate a potent anticancer drug by recruiting an anti-mortalin antibody (hsp70 chaperone that is enriched on the cancer cell surface). | Enhanced growth arrest/apoptosis of cancer cells through down-regulation of cyclin D1-CDK4, phospho-Rb, PARP-1 and the anti-apoptotic protein Bcl2. Significantly increased expression of p53, p21 WAF1, and caspase cleavage. Significantly improved down-regulation of proteins involved in cell migration |
(Wang, Bhargava et al. 2020) |
| Folic acid-conjugated PLGA nanoparticles. | Improve cytotoxicity, solubility, and achieve sustained release of CAPE. | It showed enhanced cytotoxicity in vivo and in vitro, causing a decrease in cell proliferation by 46%. | (Kapare, Lohidasan et al. 2021) |
| Stimuli-responsive liposomal nanocarrier loaded with CAPE modified with ac peptide (RGDyK) | Attack ischemic lesions and remodel neurovascular units (NVU) to reduce the progression of brain injury. | Drugs release in response to pathological signaling stimuli, localization of cerebral ischemia-reperfusion injury and remodeling of neurovascular units by reducing neuronal apoptosis, regulating microglia polarization and repair of vascular endothelial cells. | (Lu, Li et al. 2022) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



