
Submitted:
29 May 2023
Posted:
31 May 2023
You are already at the latest version
Abstract
Oxidative stress (OS) is a condition in which there is an excess of reactive oxygen species (ROS) in the body, which can lead to cell and tissue damage. This occurs when there is an overproduction of ROS or when the body's antioxidant defense systems are overwhelmed. Quercetin is part of a group of compounds called flavonoids. It is found in high concentrations of vegetables, fruits, and other foods. Over the past decade, a growing number of studies have highlighted the therapeutic potential of flavonoids to modulate neuronal function and prevent age-related neurodegeneration. Therefore, quercetin has been shown to have antioxidant, anticancer, and anti-inflammatory properties, both in vitro and in vivo. Due to its antioxidant character, quercetin alleviates oxidative stress, thus improving cognitive function, reducing the risk of neurodegenerative diseases. On the other hand, quercetin can also help support the body's natural antioxidant defense systems, thus being a potentially practical supplement for managing oxidative stress. This review focuses on experimental studies supporting the neuroprotective effects of quercetin in Alzheimer’s disease (AD), Parkinson’s disease (PD), Huntington's disease (HD) and epilepsy.
Keywords:
oxidative stress
; neurodegenerative diseases
; quercetin
; neuroprotective effects
1. Introduction
Excessive oxidative stress is the result of disturbing the balance between oxidation and antioxidant systems, with a tendency to oxidize. Oxidative stress can cause many reactions, such as protease stimulation, neutrophil infiltration, and the explosion of oxidative intermediates [1]. In addition, OS is thought to play a key role in progressive degeneration and/or death of nerve cells, especially in neurodegenerative diseases, where it acts as a mediator of the side effects of several neurotoxic substances and as a mechanism of age-related degenerative processes [2]. The ROS scavenger is often used to counteract the effects of oxidative stress in neurons [3,4]. Numerous studies have shown that quercetin, by eliminating oxygen radicals and by metal chelating operations, attenuates neuronal damage mediated by oxidative stress [5,6].
To survive, aerobic multicellular organisms need molecular oxygen (O2), to the detriment of oxygen, which is susceptible to radical formation due to its electronic structure. Reactive oxygen species are the natural by-product of normal oxygen metabolism and play significant roles in homeostasis and cellular signaling. Oxidative stress increases in the cellular environment when oxygen homeostasis is not maintained. ROS are oxygen free radicals or small molecules derived from oxygen, such as peroxyl radical (ROO•), hydroxyl radical (⋅OH), superoxide anion (•O2–), and alkoxyl (RO-). ROS could also come from non-radicals such as ozone (O3), hypochlorous acid (HOCl), singlet oxygen (1O2), and hydrogen peroxide (H2O2). These non-radicals are oxidizing agents or are easily converted to radicals [7,8].
Neurological diseases are a consequence of genetics, environmental factors, and even age [9]. During periods of environmental stress, such as ultraviolet A (UVA) and ultraviolet B (UVB) radiation, exposure to heat, and ionizing radiation, their levels could increase dramatically. A study by Erden et al., 2001 [10] showed that exposure to UVA radiation can induce ROS production leading to damage to cellular elements, but also the benefits that antioxidant quercetin brings to cells, protecting them from the harmful effects of radiation.
Neurodegeneration is characterized by progressive deterioration of the structure and function of neurons and is accompanied by severe cognitive deficits. Aging is the main risk factor for neurodegenerative disorders such as Alzheimer’s disease, Parkinson’s disease, and Huntington’s disease. Mitochondrial dysfunction and oxidative stress also trigger neurodegeneration. Recent studies have supported the mechanisms by which quercetin supports brain health [11].
First studies involving flavonoids to reduce oxidative stress have been performed since the end of the twentieth century [12]. Numerous in vitro and in vivo studies have reported the neuroprotective properties of quercetin [11,13,14]. Thus, it has been observed to protect neurons from oxidative damage and reduces lipid peroxidation as well (see Figure 1). On the other hand, besides its antioxidant properties, quercetin can inhibit the formation of amyloid-β proteins in the fibrils, counteracting cell lysis, and inflammatory cascade [13]. Flavonoids, but also foods containing flavonoids can have multiple beneficial effects in the treatment of conditions involving oxidative stress, such as Alzheimer’s disease, Parkinson’s disease, aging itself, atherosclerosis, and ischemia [12,15,16].
Quercetin or 2-(3,4-dihydroxy phenyl)-3,5,7- trihydroxychromen-4-one is commonly found in our diet and is found in abundance as a secondary metabolite in vegetables and fruits. According to the USDA (United States Department of Agriculture) database on flavonoid concentration in foods [18] and Table 1, the highest quercetin concentrations are in capers, dill weed, oregano, onions, cranberries, cherries, and red fruits; in addition to fruits and vegetables, they are also found in beverages such as red wine and black tea. It has also been isolated and marketed as a dietary supplement, in the form of free aglycone, used in doses of 1000 mg per day, exceeding the usual levels of food intake, which is 200-500 mg per day in individuals who consume large amounts of vegetables and fruits [18,19].
Quercetin is a more potent antioxidant than other antioxidant nutrients, such as vitamin C, vitamin E, and β-carotene [20]. Due to the five hydroxyl groups present in its structure that can bind to ROS, quercetin has a higher antioxidant potential than many other flavonoids [21,22]. In addition to its antioxidant activity, quercetin has effects anti-cancer [6,7,8,9,10], and anti-inflammatory [21,28,29,30], antiviral [31,32,33], antibacterial properties [34,35,36,37,38], cardioprotective effects [39], neuroprotective effects vs. brain ischemia [1,40].
In many studies, quercetin is also reported to have adverse effects, such as induction of mutations, chromosomal aberrations, and single-stranded deoxyribonucleic acid (DNA) ruptures in various eukaryotic cell systems in vitro [41].
Quercetin is less toxic than curcumin or gallic acid, due to the LD50 value of 484 µg mL−1, while the LD50 values for curcumin it is 135 µg mL−1 and for gallic acid it is 304 µg mL−1 [42].
This review focuses on the preventive and therapeutic capacity of quercetin in neurological and neurodegenerative diseases along with its potential mechanisms of action. Furthermore, we also summarized the biological sources and other pharmacological activities of this antioxidant compound.
Quercetin acts as a protector of neurons against severe oxidative stress, but also against free radical attack by easily intercalating its molecules in DNA, thus forming a protective barrier against stronger intercalators and/or ROS attack [43].
2. Methodology
This updated review covers the neuroprotective effects and potential alternative therapeutic options of quercetin in neurological disorders. The literature databases searched for information used in the present review until inception (May 2023) were: PubMed (https://pubmed.ncbi.nlm.nih.gov/), ScienceDirect (https://www.sciencedirect.com/), Scopus (https://www.scopus.com/), and Google Scholar (https://scholar.google.com/).
We used “quercetin”, “neurological disorders”, “Alzheimer’s disease”, “Parkinson’s disease”, “neurodegenerative diseases”, “oxidative stress”, “neuroinflammation” and “amyloid beta” as the keywords for the literature search.
The chemical structure was revised by consulting the open PubChem database (https://pubchem.ncbi.nlm.nih.gov/) and the scientific names of the plants were revised according to PlantList (http://www.theplantlist.org/). The purpose is to include in vitro and in vivo experimental studies that highlighted the main neuroprotective effects of quercetin. To this review, only full-text articles in English from the years 2013–2023 were chosen, with some important publications from earlier years. Finally, 115 articles are referred to in this review.
3. Oxidative stress
OS is a condition in which there is an imbalance between production of ROS and the body’s ability to neutralize them, causing damage to lipids, proteins, and DNA. ROS are naturally produced by the body as a by-product of normal metabolism but can also be produced by environmental factors such as exposure to pollution, radiation, and certain chemicals. In the context of neurological disease, oxidative stress has been shown to play a role in the development and progression of several conditions.
Recent studies have shown that quercetin can protect against oxidative stress-induced cell death by inhibiting the activity of caspase-3 [44] and increasing the body’s antioxidant capacity by regulating glutathione (GSH) levels [45]. Quercetin has also been shown to reduce inflammation and oxidative stress and improve wound healing in animal models of Alzheimer’s disease [43] and reduce inflammatory pain by inhibiting oxidative stress pathways [46] (see Figure 2). These findings suggest that quercetin may be beneficial in reducing oxidative stress and inflammation in humans.
In AD, OS has been shown to accelerate the formation of amyloid plaques in the brain, which are a hallmark of the disease. Also, in PD, OS can cause the death of dopaminergic neurons, leading to loss of motor function. And in HD, stress can increase levels of the glutamate neurotransmitter in the brain, which can lead to neuron death and worsening symptoms. Thus, Figure 2, we highlighted the main enzymes that have the role of protecting the body from oxidative stress and have neuroprotective effects in the case of neurodegenerative diseases.
3.1. Alzheimer’s disease
Alzheimer’s disease is the most common neurodegenerative disease, accounting for about two-thirds (60-80%) of all cases of dementia, and it affects mainly the elderly (aged 65 or older) [48]. The pathogenesis of AD is commonly associated with extracellular accumulation of amyloid-β (1-40, 1-42) aggregates and hyperphosphorylation of tau proteins, leading to neurofibrillary tangles (NFT) and synaptic dysfunction [13,49,50,51]. An estimated 44 million people worldwide are affected by AD or a related form of dementia, with a prevalence rate of 4.6 million new cases each year. The prevalence rate of AD increases with age: the rate doubles every 5 years from the age of 60 [52,53].
Oxidative stress plays an important role in AD, which, through ROS generation, can amplify or initiate the disease. The reduction reaction of hydrogen peroxide results in the production of reactive oxygen species, thus damaging brain tissue and disrupting brain cell repair [54,55]. Thus, the administration (ad) of quercetin before the treatment decreases the damage to the cell membrane induced by the oxidative stress caused by H2O2 [55].
AD is characterized by neuronal loss, which is preceded by extracellular accumulation of Aβ1–40, and Aβ1–42. Antioxidants such as quercetin increase the resistance of neurons to oxidative stress by modulating cell death mechanisms. Thus, quercetin protects the mouse hippocampal cell line HT-22 from glutamate-induced oxidative toxicity and lipid peroxidation, by blocking the production of free radicals [54]. Also, pretreatment of primary hippocampal cultures with quercetin significantly attenuated Aβ1–42 induced cytotoxicity, protein oxidation (protein carbonyl, 3-nitrotyrosine), lipid peroxidation (protein-bound 4-hydroxy-2-nonenal), and apoptosis. There were also observed protective effects against Aβ1–42 toxicity by modulating oxidative stress at lower concentrations (5 and 10 μM), while in the cases of higher concentrations (20 and 40 μM), effects were not only non-neuroprotective but toxic [13].
However, recent research shows that the more hydroxyl groups the structure of the molecule contains, the stronger the anti-amyloidogenic activity. Therefore, one of the potential benefits of quercetin is its ability to act as an anti-amyloidogenic agent due to its five hydroxyl groups (see Figure 2), which means it can prevent the formation of amyloid plaques in the brain [56,57].
Numerous studies demonstrate anticancer and apoptosis-inducing effects in vitro on a variety of cancer cell lines, including murine neuroblastoma HT-22 cells [54,58,59]. Quercetin was found to be non-cytotoxic and strongly protected HT-22 cells from fibril formation [59]. In addition, Ishige et al., 2001, [54] using the HT-22 mouse hippocampal cell line found three distinct mechanisms of flavonoid protection in cell death, including increased intracellular GSH, direct decrease in ROS levels, and prevention of Ca2+ influx.
In a study on the stable cell line of the Swedish mutant of amyloid precursor protein (APP695 transfected SH-SY5Y) no effects of quercetin were observed in the middle-late stage of AD; instead, effects were observed in the mid-early stage, when the reduction of β-amyloid converting enzyme 1 (BACE1) activity was recorded [57].
The optimal concentration of quercetin required for effective destabilization of Aβ fibrils has been found to be in the range of 0.1–1 μM [58,60]. Thus, in a study using neurons of the rat hippocampal region, optimal doses of quercetin administration were beneficial for protecting against Aβ25-35-induced amnesic injury by reducing lipid peroxidase, ROS, and GPx [60]. In another study on pheochromocytoma (PC12) cells, quercetin was found to increase the survival rate of H2O2-damaged cells, decrease lipid peroxidation and GSH level, and provide mitochondrial protection mechanisms [55]. On the other hand, Yu et al., 2020, [61] have shown that quercetin has beneficial effects, so it can increase PC12 cell survival damaged by Aβ25-35, antagonize toxicity Aβ, promote cell proliferation, and provide some neuroprotective effects.
Quercetin-3-glucuronide (Q3G), a glucuronide conjugate of quercetin, has been identified as a potential intervention for AD due to its ability to target the brain. Thus, several studies have shown that Q3G may be able to alleviate neuroinflammation and reduce oxidative stress in nerve cells, both of which being associated with AD [62,63]. Additionally, Q3G has been found to specifically localize in human brain tissue, suggesting that it may be able to cross the blood-brain barrier and reach areas of the brain affected by AD [62]. Thus, Ho et al., 2013, [64] observed that quercetin-3-O-glucuronide significantly reduced the generation of β-amyloid peptides by cultures of primary neurons generated by the mouse model Tg2576 AD.
A high concentration of quercetin was found in Ginkgo biloba, thus showing that Ginkgo biloba extract (EGb761) and its constituents, quercetin and ginkgolide B, which by intraperitoneal administration (i.p.) have protective effects against the cytotoxic action of Aβ1–42, thereby ameliorating oxidative phosphorylation deficits, and mitochondrial dysfunction in AD [65]. Another plant used in pharmacotherapy and in which quercetin is found is Acanthopanax henryi, that it can potentiate cholinergic activity by inhibiting acetylcholinesterase (AchE) [66].

Table 2.
Protective effects against oxidative stress, neuroinflammation and Aβ accumulation, induced by quercetin in vitro.
Table 2.
Protective effects against oxidative stress, neuroinflammation and Aβ accumulation, induced by quercetin in vitro.
| Types of quercetin | Concentration of quercetin | Model | Exposure | Effects | Ref. |
|---|---|---|---|---|---|
| Quercetin | Dosage: 2.2 μM Duration: 24 h. |
HT-22 mouse hippocampal cell | H2O2 | ↓lipid peroxidation, ↑intracellular GSH, ↓ROS | [54] |
| Dosage: 10-100 µmol L−1 Duration: 10 min |
PC12 cells | H2O2 | ↓lipid peroxidation, ↓GSH, mitochondrial protection mechanisms. | [55] | |
| Dosage: 10 μM | APP695-transfected SH-SY5Y cells | Aβ25–35 | ↓ROS, ↓BACE, ↓Aβ, ↓GSH, ↓lipid peroxidation | [57] | |
| Dosage: 10 and 50 μM Duration: 7 days |
HT-22 mouse hippocampal cells | Aβ1–42 or Aβ1–40 | ↓Aβ peptides, ↓the performed mature fibrils | [58] | |
| Dosage: 2.4 µg mL−1 | HT-22 murine neuroblastoma cells | Aβ25–35 | ↓amyloidogenic Aβ peptides, inhibited Aβ fibril formation. |
[59] | |
| Quercetin-3′-glucoside | Dosage: 10, 20, 40, and 80 μmol L−1 Duration: 24 h, 48 h, and 72 h |
PC12 cells | Aβ25–35 | ↑ the survival rate of PC12 injured by Aβ25-35, promote cell proliferation, and antagonize the toxicity of Aβ, ↑ CREB/BDNF signaling pathway, ↓ROS | [61] |
| Q3G | Dosage: 50 mg kg−1 Ad: gavage Duration: 24 h, 48 h, and 72 h |
APP695-transfected SH-SY5Y cells | Aβ1–42 | ↓Aβ peptides, ↑CREB signaling, ↓Aβ aggregation, ↑mitogen-activated protein | [64] |
| Dosage: 50 mg kg −1 Ad: gavage Duration: 4 months |
Tg2576 AD primary neuron cultures | ↑neuronal survival, ↑c-Jun N-terminal kinases, ↓stress-induced impairments | |||
| Quercetin/Ginkgo biloba | Dosage: 1.5-6 μg mL−1 Ad: i.p. |
SHSY5Y human neuroblastoma cells | Aβ1–42 | ↓ Akt signaling pathways, ↓Aβ toxicity, ↓platelet-activating factor | [65] |
| Quercetin/ Acanthopanax henryi |
Dosage: 2.5, 5, 10, 20, and 40 μg mL−1 | Cell free system | ↓ AchE activity, ↑antioxidant activity |
[66] |
In mitochondria, the first free radical to form is the superoxide radical, which is catalyzed by superoxide dismutase (SOD) and can cause irreversible damage to nucleic acids, proteins, phospholipids and / or signaling pathways, thus contributing to apoptosis and intoxication [67].
Thus, it has been observed that the administration of quercetin can attenuate the levels of the SOD enzyme, reduce the amount of lactate dehydrogenase (LDH) release and protect human brain microvascular endothelial cells (hBMECs) from fibrillar β-amyloid1–40 (fAβ1–40)-induced toxicity [68]. In vivo studies in triple transgenic mice model of AD (3xTg-AD) have shown that quercetin can disaggregate amyloid fibrils, such as extracellular amyloid β-peptide, tauopathy astrogliosis, and microgliosis in the hippocampus and amygdala, and improves their spatial memory and learning [69,70]. Additionally, the results showed that quercetin tended to improve active behaviors of 3xTG-AD mice and decreased neurodegeneration markers in mice [70]. Additionally, in the case of APPswe/PS1dE9 transgenic mice it was observed that long-term quercetin consumption prevents memory loss, Aβ-induced neurotoxicity, and mitochondrial dysfunctions [71].
Furthermore, Hayakawa et al., 2015, [72] have shown that quercetin has memory enhancing effects in older mice and delays the deterioration of memory in the early stages of Alzheimer’s, since it reduces eIF2a and ATF4 expression by inducing GADD34 in the brain. Also, quercetin can partially block the effect of other genes that play an important role in Alzheimer’s disease, such as tumor necrosis factor-alfa (TNF-α), IL-1β, IL-6 [73].
Quercetin has the effect of reducing anion superoxide levels that increased with H2O2 and Aβ treatment in hippocampal neurons [74,75]. On the other hand, oral (p.o.) treatment with 500 mg kg−1 b.w. quercetin for 10 days can significantly increase brain apoE levels and reduce insoluble Aβ levels in the cortex of 5xFAD amyloid model mice [76].
Interestingly, this memory impairment was markedly ameliorated by oral treatment with quercetin nanoencapsulated in zein nanoparticles (25 mg kg−1 every 48 h for 2 months), while the administration of free quercetin was not able to reverse the faulty behavior, despite a higher administration frequency [77].
In a study involving homozygotic transgenic mouse line B6.129S7-Sod2tm1Leb/J, where quercetin was administered orally at a dose of 50 mg kg−1 body weight (b.w.) twice a week for four weeks, the results showed that quercetin had a protective effect against hydrogen peroxide- and paraquat-induced oxidative stress in the mice [74].
Scopolamine administration causes short-term and long-term memory loss because it blocks muscarinic cholinergic receptors in the brain and interferes with learning and memory [78,79]. There are studies that have found that quercetin alleviates scopolamine-induced memory deficits by protecting against neuroinflammation and neurodegeneration by inhibiting oxidative stress and acetylcholinesterase activity, reverses synaptic loss in the cortex and hippocampus of the brain of adult mice, and suppresses memory impairment [78,79].
Aluminum is a toxic metal that has neurological effects, including Alzheimer’s disease, by generating ROS [80]. Increased production of reactive oxygen species leads to the disruption of cellular antioxidant defense systems and to the release of cytochrome c from mitochondria into the cytosol, resulting in apoptotic cell death [80,81]. Thus, the administration of 10 mg kg−1 b.w quercetin reduces the effects induced by aluminum, thus reducing oxidative stress, in addition, it prevents cytochrome c translocation [80].
In addition, Hou et al., 2010 [82] have shown that flavonols can antagonize A toxicity Aβ and improve the expression of brain-derived neurotrophic factor (BDNF) in the hippocampus of double transgenic mice.
Table 3.
Protective effects against oxidative stress, neuroinflammation, and Aβ accumulation, induced by quercetin in vivo.
Table 3.
Protective effects against oxidative stress, neuroinflammation, and Aβ accumulation, induced by quercetin in vivo.
| Types of quercetin | Concentration | Model | Exposure | Effects | Ref. | |
|---|---|---|---|---|---|---|
| Quercetin | Dosage: 5 or 10 mg kg−1 b.w.; Ad: p.o.; Duration: once daily; |
hBMECs | fAβ1–40 | ↓SOD, ↓LDH | [68] | |
| Dosage: 25 mg kg−1 b.w.; Ad: i.p.; Duration: every 2 days for 3 months; |
3xTg-AD mice | ↓tauopathy, ↓β-amyloidosis, ↑memory, ↑learning↓ microgliosis, ↓astrogliosis | [69] | |||
| Dosage: 100 mg kg−1 b.w.; Ad: gavage; Duration: every 48 h for 12 months; |
3xTg-AD mice | ↓neurodegeneration, ↓β-amyloidosis | [70] | |||
| Dosage: 20 and 40 mg kg−1 b.w.; Ad: p.o.; Duration: 16 weeks; |
adult male C57BL mice | ↑ MMP, ↑ATP levels, ↓ROS | [71] | |||
| Dosage: 20 mg; Ad: p.o.; Duration: 5 weeks; |
APP23 AD mice model | Aβ | ↓eIF2α, ↓ATF4, ↓GADD34, ↑memory in aged mice, ↓memory deterioration in the early stage of AD, ↓memory dysfunction, ↓OS | [72] | ||
| Dosage: 1% in mouse chow; Ad: p.o.; Duration: from 3 to 13 months; |
double transgenic female mice | ↓neuroinflammation, ↓neurodegeneration, ↓ IL-1β |
[73] | |||
| Dosage: 50 mg kg−1 b.w.; Duration: 2 times a week for 4 weeks; |
homozygotic transgenic mouse line B6.129S7-Sod2tm1Leb/J | H2O2 and Aβ | ↓ROS levels, improved the typical morphology of mitochondria, prevented mitochondrial dysfunction | [74] | ||
| Dosage: 100 mg kg−1 b.w.; Ad: p.o.; Duration: 22 days; |
adult male Sprague-Dawley rats | Aβ1–42 | ↑expression of Nrf2/HO-1 in rat brain, ↓Aβ1-42 level, ↓antioxidant activity | [75] | ||
| Dosage: 25 mg kg−1; Ad: p.o.; Duration: 2 times a week for 2 months; |
SAMP8 mice | ↑ the cognition and memory impairments, ↓ astrogliosis | [77] | |||
| Dosage: 12.5 and 25 mg kg−1; | mice | Scopolamine | ↓OS, ↓AchE activity | [78] | ||
| Dosage: 30 mg kg−1 b.w.; Ad: i.p.; Duration: every day for 8 days; |
male albino Wistar rats | Scopolamine | abridged transfer latency, ↓avoidance response, ↓3,4-methylenedioxyamphetamine, acetylcholinesterase levels, ↑CAT, ↑ GSH levels | [79] | ||
| Dosage: 10 mg kg −1 b.w.; Ad: p.o.; Duration: every day for 12 weeks; |
male albino Wistar rats | aluminum | ↓ROS production, ↑mitochondrial superoxide dismutase activity |
[80] | ||
| Quercetin/ ginkgo flavonols | Dosage: 4.8% in extract, all based on weight; |
Double Transgenic (TgAPP/PS1) mice | - | Reversed the spatial learning deficit | [82] | |
Several studies have shown that type 2 diabetes is a risk factor for the onset of AD [83,84]. Moreover, changes in glucose regulation that accompany type 2 diabetes appear to affect regions of the hippocampus involved in age-related memory loss [84].
The link between AD and type 2 diabetes is not fully understood. However, uncontrolled blood sugar can increase the risk of AD [85]. Some researchers have referred to AD as “diabetes of the brain” or “type 3 diabetes” [86,87,88]. Environmental factors and comorbidities, as well as other diseases that a certain person suffers from, can make them more susceptible to the onset of AD, but all studies so far point to the fact that the accumulation of amyloid is the fundamental cause of dementia.
3.2. Parkinson’s disease
Parkinson’s disease is the second most common neurodegenerative disorder worldwide, affecting 1% of the global population aged 65 years and older; it has significant morbidity and mortality [89]. An increasing percentage of research indicates the association of Parkinson’s disease with microglial activation, resulting in an increase in various inflammatory mediators and neuroinflammation [90,91]. 1-Methyl-4-phenylpyridinium (MPP+) is the ultimate toxic agent formed by metabolism of MPTP and can activate glial cells to induce neuroinflammation [92]. Research has shown that MPP+ induces microglial activation and degeneration of dopaminergic neurons, as well as generation of ROS in dopaminergic neurons [93]. On the other hand, quercetin administration protects microglia cells against MPP+- induced increases in mRNA protein level of IL-1, IL-6 and TNF-α, due to its antioxidant action [92]. In addition to the loss of dopaminergic neurons in the substantia nigra pars compacta, PD is also characterized by the abnormal accumulation and aggregation of α-synuclein (α-Syn) in the form of Lewy bodies [94,95]. Thus, the formation of α-Syn fibrillations can be inhibited by quercetin and oxidized quercetin through their 1:1 covalent binding [95].
Another neurotoxic synthetic organic compound used by researchers to selectively destroy dopaminergic and noradrenergic neurons is 6-hydroxydopamine (6-OHDA) [96,97]. It is a hydroxylated analogue of dopamine and is a benzenetriol with hydrogens on the phenyl ring at positions 2, 4 and 5. Isoquercetin, a flavonol derived from quercetin, has also been found to have protective effects against 6-OHDA-induced oxidative damage in a rat model of Parkinson’s disease, they observed that antioxidant enzymes, catalase (CAT), SOD, GPX and GSH levels, which were previously attenuated by 6-OHDA, increased significantly [96,97].
Table 4.
Protective effects against oxidative stress and neuroinflammation, induced by quercetin in vitro, in the case of Parkinson’s disease.
Table 4.
Protective effects against oxidative stress and neuroinflammation, induced by quercetin in vitro, in the case of Parkinson’s disease.
| Types of quercetin | Concentration | Model | Exposure | Effects | Ref. |
|---|---|---|---|---|---|
| Quercetin | Dosage: 0.1 μM |
Microglial (N9)-neuronal (PC12) cells | MPP | ↓iNOS gene expression, ↓ROS, ↓ cellular death, ↓DNA fragmentation, ↑apoptosis, ↓nuclear translocation of apoptosis-inducing factor, ↓caspase-3activation | [92] |
| Dosage:10 mM |
PC12 cells | α-Synuclein | ↓Aβ fibrillation | [95] | |
| Isoquercetin | Dosage: 10, 50, and 100 μM |
PC12 cells | 6-OHDA | ↓ROS, ↑SOD, ↑GSH, ↑CAT, ↑GPx | [96] |
| Quercetin glycoside | Dosage: 10, 50, and 100 μM | PC12 cells | 6-OHDA | ↑antioxidant activity, ↑GSH, ↑GPx | [97] |
In vitro studies have shown that quercetin can improve mitochondrial quality control, reduce oxidative stress, and increase levels of antioxidant enzymes [92,95,96,97]. Instead, in vivo studies in mice and 6-OHDA-induced PD rat models demonstrated that quercetin can improve locomotor and muscle activity, increase striatal dopamine levels, and protect neurons from mitochondrial dysfunction [98,99,100,101,102].
Studies have shown that quercetin has neuroprotective effects against MPTP-induced neurotoxicity in Wistar rats and adult male C57BL/6 mice [98,99,100]. Quercetin was found to reduce oxidative stress and neuroinflammatory cytokines in rats [98,99], as well as restores motor and non-motor symptoms (depression and cognitive impairment) of PD in rats injected with rotenone [103,104]. Additionally, quercetin supplementation was found to improve striatal cholinergic function and reduce rotenone-induced oxidative stress in rats [104].
On the other hand, administration of fish oil can attenuate rotenone-induced oxidative impairments and mitochondrial dysfunctions in rat brain [105]. Combined oral supplementation with fish oil and quercetin has been found to enhance neuroprotection in a chronic rotenone rat model, suggesting potential relevance for Parkinson’s disease [105].
Table 5.
Protective effects against oxidative stress and neuroinflammation of quercetin in vivo in the case of Parkinson’s disease.
Table 5.
Protective effects against oxidative stress and neuroinflammation of quercetin in vivo in the case of Parkinson’s disease.
| Types of quercetin | Concentration | Model | Exposure | Effects | Ref. |
|---|---|---|---|---|---|
| Quercetin | Dosage: 25 mg kg−1 Ad: p.o. |
Wistar rats | Haloperidol MPTP | ↓cataleptic score, ↑actophotometer activity score, ↑GSH, ↓lipid peroxidation, ↓ROS | [98] |
| Dosage: 25 and 50 mg kg−1, Ad: intragastrically Duration: 14 days |
Wistar rats | MPTP | ↓TNF-α, ↓IL-1β and ↓IL-6, ↓glutamate level, | [99] | |
| Dosage: 50, 100 and 200 mg kg−1 Ad: p.o. Duration: 14 days. |
adult male C57BL/6 mice | MPTP | ↓striatal dopamine depletion, ↓level of acetylcholine, ↑AchE activity, ↑motor deficits, ↑GPx, ↑SOD | [100] | |
| Dosage: 100, 200 and 300 mg kg −1 Duration: 14 days |
Wistar rats | 6-OHDA | ↑spatial memory, ↓OS, ↓AchE activity, ↑antioxidant activity, ↓neuronal damage | [101] | |
| Dosage: 20 mg kg−1 Ad: i.p. Duration: 1 month. |
Wistar rats | 6-OHDA | ↓neuroplastic changes in neural circuits, ↓excitability in neurons involved in epilepsy, ↓NMDA receptor functionality | [102] | |
| Dosage: 25-75 mg kg−1 Duration: 12 h intervals for 4 days |
Wistar rats | Rotenone | ↓nigral GSH depletion, ↓ROS, ↓striatal DA loss, ↑mitochondrial complex, ↓neuronal death | [103] | |
| Dosage: 50 mg kg−1, Ad: p.o. Duration: 14 days |
Wistar rats | Rotenone | ↑AchE activity, ↑SOD, ↓GPx, ↓CAT | [104] | |
| Quercetin + fish oil | Dosage: 25 mg kg−1 Ad: p.o. Duration: 28 days |
Wistar rats | Rotenone | ↑mitochondrial functions, ↑GSH, ↑antioxidant defenses | [105] |
3.3. Huntington’s disease
Along with Alzheimer’s and Parkinson’s, Huntington’s disease (HD) is a major health problem worldwide, with a major financial impact [106]. Huntington’s disease is an autosomal dominant, inherited disorder, treatment of which is clinically available but provides only symptomatic relief. These drugs are available by prescription and have side effects such as anxiety and depression.
For an experimental model of Huntington’s disease, 3-nitropropionic acid (3-NPA) is administered, which alters the mitochondrial metabolism, decreases cellular ATP level, and includes the nerve cell death, by increasing OS.
In a study by Sandhir and Mehrotra [107], in which female Wistar rats were used as model organisms, in which quercetin was orally administered at a dose of 25 mg kg−1 for 21 days, for 17 of these 21 days concomitantly with 3-NPA, was observed an attenuation of motor deficits which were assessed using the narrow-beam walking test and fingerprint analysis. Furthermore, molecular changes induced by 3-NPA acid were observed, which were reversed, thus increasing the level of oxidative stress, and lowering the ATP concentration [107].
On the other hand, a study by Chakraborty [108] failed to confirm the beneficial effect of quercetin on the 3-NP-induced striatal neuronal lesion. However, the conditions of the two studies varied little, Chakraborty used male rats as model organisms, and the duration of administration of 3-NP and quercetin was 4 days, and the concentration was higher (25-50 mg kg−1), than that which was administered by Sandhir and Mehrotra, [107] where the dose was administered subchronically (25 mg kg−1). Although quercetin had no effects on 3-NP-induced striatal neuronal injury, it significantly attenuated neurotoxin-induced anxiety, decreased microglial proliferation, and increased the number of astrocytes in the lesion core [108].
In addition, quercetin in combination with other antioxidants, such as lycopene, decreases anxiety and depression [109]. Furthermore, the use of dietary antioxidants as adjuvants with n-3 fatty acids is increasingly being used, as they offer a higher degree of protection. Thus, the efficacy of quercetin in combination with fish oil was observed in a rat model previously treated with 3-NPA, where it decreased OS, and improved motor function [110].
Quinolinic acid (QA) is also a paradigm of Huntington’s disease, and co-administration of antioxidants such as quercetin with sesamol minimizes neurochemical, behavioral, and biochemical alterations in rat brains [111]. However, these data appear inconsistent and unequivocal conclusions cannot be drawn.
Table 6.
Protective effects against oxidative stress and neuroinflammation, induced by quercetin in vitro in the case of Huntington’s disease.
Table 6.
Protective effects against oxidative stress and neuroinflammation, induced by quercetin in vitro in the case of Huntington’s disease.
| Types of quercetin | Concentration | Model | Exposure | Effects | Ref. |
|---|---|---|---|---|---|
| Quercetin | Dosage: 25 mg kg−1 Ad: p.o. Duration: 21 days; |
Wistar rats | 3-NPA | ↑ATP, ↑activity of complex II and V enzyme of respiratory chain complex, ↓ROS, ↑SOD, ↑CAT, ↓lipid peroxidation | [107] |
| Dosage: 25–50 mg kg−1 Ap: i.p.; Duration: 4 days |
Sprague Dawley rats |
3-NPA | ↓gait despair, ↓microglial proliferation, ↓anxiety, ↑astrocyte numbers in the lesion core, ↓motor coordination deficits, ↓serotonin metabolism | [108] | |
| Quercetin + lycopene | Dosage: 50 mg kg−1, Duration: 14 days |
Wistar rats | 3-NPA |
↓anxiety, ↓depression | [109] |
| Quercetin + fish oil | Dosage: 25 mg kg−1 | Wistar rats | 3-NPA | ↓OS, ↑motor function | [110] |
| Quercetin + sesamol | Dosage: 25, 50, and 100 mg kg−1, Ad: i.p. Duration: 14 days before and 14 days after quinolinic acid administration |
Wistar rats | QA | ↓behavioral, biochemical, and neurochemical alterations in the rat brain, ↑antioxidant effects, ↑anti-inflammatory activity | [111] |
3.4. Epilepsy
Epilepsy is a neurological disorder characterized by recurrent spontaneous seizures, being caused by an imbalance in excitatory and inhibitory neurotransmission [112]. Glutamate and γ-amino butyric acid (GABA) are the major excitatory and inhibitory neurotransmitters in the CNS [113]. A GABA receptor antagonist is pentylenetetrazol (PTZ), which is used to create a chemically induced seizure model in animals [113].
In a study using PTZ-induced seizure model rats, quercetin administration at 10 mg kg−1 intraperitoneally 30 min before PTZ injection significantly prolonged the onset and reduced the severity of the seizure, but, at an increased concentration of 40 mg kg−1, quercetin failed to prevent the effects of PTZ [113]. Also, Nassiri-Asl et al., [111] showed that administration of 35 mg kg-1 PTZ after 50 mg kg-1 quercetin reduces seizure severity during kindling and improves performance in a passive avoidance task in kindled rats. In addition, Choudhary et al., 2011, [114] isolated and evaluated the antiepileptic potential of the flavonoid fractions of Anisomeles malabarica leaves, both acute and chronic. In the acute treatment (25 and 50 mg kg−1, i.p.) with toxic effects observed, respectively chronic treatment for one week (6.25 and 12.5 mg kg−1, i.p.) a significant antiepileptic effect was observed without causing side effects neurotoxic [114].
Table 7.
Protective effects against oxidative stress and neuroinflammation, induced by quercetin in the case of epilepsy.
Table 7.
Protective effects against oxidative stress and neuroinflammation, induced by quercetin in the case of epilepsy.
| Types of quercetin | Concentration | Model | Type of test | Exposure | Effects | Ref. |
|---|---|---|---|---|---|---|
| Quercetin | Dosage: 5, 10, 20, 40 mg kg−1 | Albino rats | in vivo | PTZ | ↑antiseizure effect, ↑anticonvulsant effect | [113] |
| Dosage: 25, 50, and 100. mg kg−1 Ad: i.p. |
Wistar rats | in vivo | PTZ | ↑anticonvulsant effects, ↓seizure severity, ↓lipid peroxidation, ↑antioxidant effect, ↑memory retrieval in the passive avoidance task | [115] | |
| Quercetin/ Anisomelesma labarica |
Dosage: 25 and 50 mg kg−1 Ad: i.p. |
Wistar rat | in vivo | PTZ |
↓ locomotor activity and motor activity performance |
[114] |
| Dosage: 6.25 and 12.5 mg kg−1 Ad: i.p. Duration: 1 week |
Wistar rat | in vivo | PTZ |
potentiating the GABAergic system, inhibition of the NMDA receptor and Na+ channels. |
4. Conclusions
Research on quercetin’s neuroprotective effects suggests that it can be used to protect against various neurodegenerative diseases. Quercetin has been shown to reduce OS and inflammation, which are both associated with neurodegeneration. It has also been found to reduce hippocampal tau phosphorylation, which is a marker of AD.
In addition, quercetin has been found to act through numerous mechanistic targets to provide neuroprotection, including the modulation of receptor pathways. It has also been found to have protective effects when combined with vitamin C, lycopene, fish oil, or sesamol.
Overall, research suggests that quercetin may be an effective agent for the prevention of progressive age-related neurodegenerative diseases such as AD, PD, HD, or epilepsy, respectively. However, more research is needed to draw more concrete conclusions about the efficacy of quercetin in these disorders.
Author Contributions
Conceptualization, methodology, analysis, writing—original draft preparation, V.R. and A.C.; writing—review and editing, supervision, validation, M.N. and D.U. All authors have read and agreed to the published version of the manuscript.
Funding
The authors are thankful to the Romanian Ministry of Research, Innovation and Digitization, within Program 1—Development of the national RD system, Subprogram 1.2—Institutional Performance—RDI excellence funding projects, Contract no. 11PFE/30.12.2021, for financial support.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets used and analyzed during the current study are available from the corresponding author upon request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhang, Y.M.; Zhang, Z.Y.; Wang, R.X. Protective Mechanisms of Quercetin Against Myocardial Ischemia Reperfusion Injury. Front Physiol 2020. [CrossRef] [PubMed]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24. [Google Scholar] [CrossRef] [PubMed]
- Shukla, V.; Mishra, S.K.; Pant, H.C. Oxidative Stress in Neurodegeneration. Adv Pharmacol Sci 2011, 1–13. [Google Scholar] [CrossRef]
- Gkekas, I.; Gioran, A.; Boziki, M.K.; Grigoriadis, N.; Chondrogianni, N.; Petrakis, S. Oxidative Stress and Neurodegeneration: Interconnected Processes in Polyq Diseases. Antioxidants 2021, 1450. [Google Scholar] [CrossRef] [PubMed]
- Bandiwadekar, A.; Jose, J.; Khayatkashani, M.; Habtemariam, S.; Khayat Kashani, H.R.; Nabavi, S.M. Emerging Novel Approaches for the Enhanced Delivery of Natural Products for the Management of Neurodegenerative Diseases. Journal of Molecular Neuroscience 2021, 72, 653–676. [Google Scholar] [CrossRef] [PubMed]
- Chapman, J.M.; Muday, G.K. Flavonols Modulate Lateral Root Emergence by Scavenging Reactive Oxygen Species in Arabidopsis Thaliana. Journal of Biological Chemistry 2021, 296. [Google Scholar] [CrossRef] [PubMed]
- Ozcan, A.; Ogun, M. Biochemistry of Reactive Oxygen and Nitrogen Species. In Basic Principles and Clinical Significance of Oxidative Stress; 2015.
- Klebanoff, S.J. Oxygen Metabolism and the Toxic Properties of Phagocytes. Ann Intern Med 1980, 93, 391–398. [Google Scholar] [CrossRef]
- Sheikh, S. ; Safia; Haque, E.; Mir, S.S. Neurodegenerative Diseases: Multifactorial Conformational Diseases and Their Therapeutic Interventions. J Neurodegener Dis, 2013; 1–8. [Google Scholar] [CrossRef]
- Erden Inal, M.; Kahraman, A.; Köken, T. Beneficial Effects of Quercetin on Oxidative Stress Induced by Ultraviolet A. Clin Exp Dermatol 2001, 26, 536–539. [Google Scholar] [CrossRef]
- Elumalai, P.; Lakshmi, S. Role of Quercetin Benefits in Neurodegeneration. In Advances in Neurobiology; 2016; Vol. 12, pp. 229–245.
- Halliwell, B. Oxidants and Human Disease: Some New Concepts 1. The FASEB Journal 1987, 1, 358–364. [Google Scholar] [CrossRef]
- Ansari, M.A.; Abdul, H.M.; Joshi, G.; Opii, W.O.; Butterfield, D.A. Protective Effect of Quercetin in Primary Neurons against Aβ(1-42): Relevance to Alzheimer’s Disease. Journal of Nutritional Biochemistry 2009, 20, 269–275. [Google Scholar] [CrossRef]
- Islam, M.S.; Quispe, C.; Hossain, R.; Torequl Islam, M.; Al-Harrasi, A.; Al-Rawahi, A.; Martorell, M.; Mamurova, A.; Seilkhan, A.; Altybaeva, N.; et al. Neuropharmacological Effects of Quercetin: A Literature-Based Review. [CrossRef]
- Ames, B.N.; Shigenaga, M.K.; Hagen, T.M. Oxidants, Antioxidants, and the Degenerative Diseases of Aging. Proc Natl Acad Sci U S A 1993, 90, 7915–7922. [Google Scholar] [CrossRef] [PubMed]
- Zalpoor, H.; Nabi-Afjadi, M.; Forghaniesfidvajani, R.; Tavakol, C.; Farahighasreaboonasr, F.; Pakizeh, F.; Dana, V.G.; Seif, F. Quercetin as a JAK–STAT Inhibitor: A Potential Role in Solid Tumors and Neurodegenerative Diseases. Cell Mol Biol Lett 2022, 27. [Google Scholar] [CrossRef] [PubMed]
- Shen, P.; Lin, W.; Deng, X.; Ba, X.; Han, L.; Chen, Z.; Qin, K.; Huang, Y.; Tu, S. Potential Implications of Quercetin in Autoimmune Diseases. Front Immunol 2021, 12. [Google Scholar] [CrossRef]
- Haytowitz, D.; Wu, X.; Bhagwat, S. USDA Database for the Flavonoid Content of Selected Foods, Release 3.3. USDA Agricultureal Research Service 2018. [Google Scholar]
- Harwood, M.; Danielewska-Nikiel, B.; Borzelleca, J.F.; Flamm, G.W.; Williams, G.M.; Lines, T.C. A Critical Review of the Data Related to the Safety of Quercetin and Lack of Evidence of in Vivo Toxicity, Including Lack of Genotoxic/Carcinogenic Properties. Food and Chemical Toxicology 2007, 45, 2179–2205. [Google Scholar] [CrossRef]
- Mishra, D.; Flora, S.J.S. Quercetin Administration during Chelation Therapy Protects Arsenic-Induced Oxidative Stress in Mice. Biol Trace Elem Res 2008, 122, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Lesjak, M.; Beara, I.; Simin, N.; Pintać, D.; Majkić, T.; Bekvalac, K.; Orčić, D.; Mimica-Dukić, N. Antioxidant and Anti-Inflammatory Activities of Quercetin and Its Derivatives. J Funct Foods 2018, 40, 68–75. [Google Scholar] [CrossRef]
- Dhaouadi, Z.; Nsangou, M.; Garrab, N.; Anouar, E.H.; Marakchi, K.; Lahmar, S. DFT Study of the Reaction of Quercetin with · O2- and · OH Radicals. Journal of Molecular Structure: THEOCHEM, 2009; 35–42. [Google Scholar] [CrossRef]
- Reyes-Farias, M.; Carrasco-Pozo, C. The Anti-Cancer Effect of Quercetin: Molecular Implications in Cancer Metabolism. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Vafadar, A.; Shabaninejad, Z.; Movahedpour, A.; Fallahi, F.; Taghavipour, M.; Ghasemi, Y.; Akbari, M.; Shafiee, A.; Hajighadimi, S.; Moradizarmehri, S.; et al. Quercetin and Cancer: New Insights into Its Therapeutic Effects on Ovarian Cancer Cells. Cell Biosci 2020, 10. [Google Scholar] [CrossRef]
- Shafabakhsh, R.; Asemi, Z. Quercetin: A Natural Compound for Ovarian Cancer Treatment. J Ovarian Res 2019, 12. [Google Scholar] [CrossRef]
- Tang, S.M.; Deng, X.T.; Zhou, J.; Li, Q.P.; Ge, X.X.; Miao, L. Pharmacological Basis and New Insights of Quercetin Action in Respect to Its Anti-Cancer Effects. Biomedicine and Pharmacotherapy 2020, 121. [Google Scholar] [CrossRef] [PubMed]
- Ezzati, M.; Yousefi, B.; Velaei, K.; Safa, A. A Review on Anti-Cancer Properties of Quercetin in Breast Cancer. Life Sci 2020. [CrossRef] [PubMed]
- Hou, D.D.; Zhang, W.; Gao, Y.L.; Sun, Y. zhe; Wang, H.X.; Qi, R.Q.; Chen, H.D.; Gao, X.H. Anti-Inflammatory Effects of Quercetin in a Mouse Model of MC903-Induced Atopic Dermatitis. Int Immunopharmacol 2019, 74. [Google Scholar] [CrossRef]
- Kawabata, K.; Baba, N.; Sakano, T.; Hamano, Y.; Taira, S.; Tamura, A.; Baba, S.; Natsume, M.; Ishii, T.; Murakami, S.; et al. Functional Properties of Anti-Inflammatory Substances from Quercetin-Treated Bifidobacterium Adolescentis. Biosci Biotechnol Biochem 2018, 82, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Saeedi-Boroujeni, A.; Mahmoudian-Sani, M.R. Anti-Inflammatory Potential of Quercetin in COVID-19 Treatment. Journal of Inflammation (United Kingdom) 2021, 18. [Google Scholar] [CrossRef]
- Liu, M.; Yu, Q.; Xiao, H.; Li, M.; Huang, Y.; Zhang, Q.; Li, P. The Inhibitory Activities and Antiviral Mechanism of Medicinal Plant Ingredient Quercetin Against Grouper Iridovirus Infection. Front Microbiol 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Di Petrillo, A.; Orrù, G.; Fais, A.; Fantini, M.C. Quercetin and Its Derivates as Antiviral Potentials: A Comprehensive Review. Phytotherapy Research 2022, 36, 266–278. [Google Scholar] [CrossRef]
- Kim, C.H.; Kim, J.E.; Song, Y.J. Antiviral Activities of Quercetin and Isoquercitrin against Human Herpesviruses. Molecules 2020, 25. [Google Scholar] [CrossRef]
- Wang, S.; Yao, J.; Zhou, B.; Yang, J.; Chaudry, M.T.; Wang, M.; Xiao, F.; Li, Y.; Yin, W. Bacteriostatic Effect of Quercetin as an Antibiotic Alternative in Vivo and Its Antibacterial Mechanism in Vitro. J Food Prot 2018, 81, 69–78. [Google Scholar] [CrossRef]
- Olewnik-Kruszkowska, E.; Gierszewska, M.; Richert, A.; Grabska-Zielińska, S.; Rudawska, A.; Bouaziz, M. Antibacterial Films Based on Polylactide with the Addition of Quercetin and Poly(Ethylene Glycol). Materials 2021, 14. [Google Scholar] [CrossRef]
- Júnior, S.D. da C.; Santos, J.V. de O.; Campos, L.A. de A.; Pereira, M.A.; Magalhães, N.S.S.; Cavalcanti, I.M.F. Antibacterial and Antibiofilm Activities of Quercetin against Clinical Isolates of Staphyloccocus Aureus and Staphylococcus Saprophyticus with Resistance Profile. International Journal of Environment, Agriculture and Biotechnology 2018, 3. [Google Scholar] [CrossRef]
- Jaisinghani, R.N. Antibacterial Properties of Quercetin. Microbiol Res (Pavia) 2017, 8. [Google Scholar] [CrossRef]
- Aljadaan, S.A.N.; Elias, R.S.; Al-Anssari, R.A. Investigation of the Antioxidant and Antibacterial Activity of Novel Quercetin Derivatives. Biointerface Res Appl Chem 2020, 10. [Google Scholar] [CrossRef]
- Patel, R. V.; Mistry, B.M.; Shinde, S.K.; Syed, R.; Singh, V.; Shin, H.S. Therapeutic Potential of Quercetin as a Cardiovascular Agent. Eur J Med Chem 2018, 115, 889–904. [Google Scholar] [CrossRef]
- Lei, X.; Chao, H.; Zhang, Z.; Lv, J.; Li, S.; Wei, H.; Xue, R.; Li, F.; Li, Z. Neuroprotective Effects of Quercetin in a Mouse Model of Brain Ischemic/Reperfusion Injury via Anti-Apoptotic Mechanisms Based on the Akt Pathway. Mol Med Rep 2015, 12, 3688–3696. [Google Scholar] [CrossRef]
- Andres, S.; Pevny, S.; Ziegenhagen, R.; Bakhiya, N.; Schäfer, B.; Hirsch-Ernst, K.I.; Lampen, A. Safety Aspects of the Use of Quercetin as a Dietary Supplement; 2018; Vol. 62; ISBN 4930184126.
- Harishkumar, R.; Reddy, L.P.K.; Karadkar, S.H.; Murad, M. Al; Karthik, S.S.; Manigandan, S.; Selvaraj, C.I.; Christopher, J.S.G. Toxicity and Selective Biochemical Assessment of Quercetin, Gallic Acid, and Curcumin in Zebrafish. Biol Pharm Bull 2019, 42, 1969–1976. [Google Scholar] [CrossRef]
- Simunkova, M.; Alwasel, S.H.; Alhazza, I.M.; Jomova, K.; Kollar, V.; Rusko, M.; Valko, M. Management of Oxidative Stress and Other Pathologies in Alzheimer’s Disease. Arch Toxicol 2019, 93, 2491–2513. [Google Scholar] [CrossRef]
- Kook, D.; Wolf, A.H.; Yu, A.L.; Neubauer, A.S.; Priglinger, S.G.; Kampik, A.; Welge-Lüssen, U.C. The Protective Effect of Quercetin against Oxidative Stress in the Human RPE in Vitro. Invest Ophthalmol Vis Sci 2008, 49, 1712–1720. [Google Scholar] [CrossRef]
- Xu, D.; Hu, M.J.; Wang, Y.Q.; Cui, Y.L. Antioxidant Activities of Quercetin and Its Complexes for Medicinal Application. Molecules 2019, 24. [Google Scholar] [CrossRef]
- Valério, D.A.; Georgetti, S.R.; Magro, D.A.; Casagrande, R.; Cunha, T.M.; Vicentini, F.T.M.C.; Vieira, S.M.; Fonseca, M.J.V.; Ferreira, S.H.; Cunha, F.Q.; et al. Quercetin Reduces Inflammatory Pain: Inhibition of Oxidative Stress and Cytokine Production. J Nat Prod 2009, 72. [Google Scholar] [CrossRef]
- Grewal, A.K.; Singh, T.G.; Sharma, D.; Sharma, V.; Singh, M.; Rahman, M.H.; Najda, A.; Walasek-Janusz, M.; Kamel, M.; Albadrani, G.M.; et al. Mechanistic Insights and Perspectives Involved in Nfeuroprotective Action of Quercetin. Biomedicine and Pharmacotherapy 2021, 140, 111729. [Google Scholar] [CrossRef] [PubMed]
- Deture, M.A.; Dickson, D.W. The Neuropathological Diagnosis of Alzheimer’s Disease. Mol Neurodegener 2019, 14, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Kommaddi, R.P.; Das, D.; Karunakaran, S.; Nanguneri, S.; Bapat, D.; Ray, A.; Shaw, E.; Bennett, D.A.; Nair, D.; Ravindranath, V. Aβ Mediates F-Actin Disassembly in Dendritic Spines Leading to Cognitive Deficits in Alzheimer’s Disease. Journal of Neuroscience 2018, 38, 1085–1099. [Google Scholar] [CrossRef] [PubMed]
- Parent, M.J.; Zimmer, E.R.; Shin, M.; Kang, M.S.; Fonov, V.S.; Mathieu, A.; Aliaga, A.; Kostikov, A.; do Carmo, S.; Dea, D.; et al. Multimodal Imaging in Rat Model Recapitulates Alzheimer’s Disease Biomarkers Abnormalities. Journal of Neuroscience 2017, 37, 12263–12271. [Google Scholar] [CrossRef]
- Wallace, R.A.; Dalton, A.J. What Can We Learn from Study of Alzheimer’s Disease in Patients with Down Syndrome for Early-Onset Alzheimer’s Disease in the General Population? Alzheimers Res Ther 2011, 3. [Google Scholar] [CrossRef]
- Hollingworth, P.; Harold, D.; Jones, L.; Owen, M.J.; Williams, J. Alzheimer’s Disease Genetics: Current Knowledge and Future Challenges. Int J Geriatr Psychiatry 2011, 26, 793–802. [Google Scholar] [CrossRef]
- Mayeux, R.; Stern, Y. Epidemiology of Alzheimer Disease. Cold Spring Harb Perspect Med 2012, 2. [Google Scholar] [CrossRef]
- Ishige, K.; Schubert, D.; Sagara, Y. Flavonoids Protect Neuronal Cells from Oxidative Stress by Three Distinct Mechanisms. Free Radic Biol Med 2001, 30, 433–446. [Google Scholar] [CrossRef]
- Ho, J.H.; Chang, Y.L. Protective Effects of Quercetin and Vitamin C against Oxidative Stress-Induced Neurodegeneration. J Agric Food Chem 2004, 52, 7514–7517. [Google Scholar] [CrossRef]
- Porat, Y.; Abramowitz, A.; Gazit, E. Inhibition of Amyloid Fibril Formation by Polyphenols: Structural Similarity and Aromatic Interactions as a Common Inhibition Mechanism. Chemical Biology Drug Design 2006, 67, 27–37. [Google Scholar] [CrossRef]
- Jiménez-Aliaga, K.; Bermejo-Bescós, P.; Benedí, J.; Martín-Aragón, S. Quercetin and Rutin Exhibit Antiamyloidogenic and Fibril-Disaggregating Effects in Vitro and Potent Antioxidant Activity in APPswe Cells. Life Sci 2011, 89, 939–945. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Yoshiike, Y.; Takashima, A.; Hasegawa, K.; Naiki, H.; Yamada, M. Potent Anti-Amyloidogenic and Fibril-Destabilizing Effects of Polyphenols in Vitro: Implications for the Prevention and Therapeutics of Alzheimer’s Disease. J Neurochem 2003, 87, 172–181. [Google Scholar] [CrossRef]
- Kim, H.; Park, B.S.; Lee, K.G.; Cheol, Y.C.; Sung, S.J.; Kim, Y.H.; Lee, S.E. Effects of Naturally Occurring Compounds on Fibril Formation and Oxidative Stress of β-Amyloid. J Agric Food Chem 2005, 53, 8537–8541. [Google Scholar] [CrossRef]
- Rattanajarasroj, S.; Unchern, S. Comparable Attenuation of Aβ25-35-Induced Neurotoxicity by Quercitrin and 17β-Estradiol in Cultured Rat Hippocampal Neurons. Neurochem Res 2010, 35, 1196–1205. [Google Scholar] [CrossRef]
- Yu, X.; Li, Y.; Mu, X. Effect of Quercetin on PC12 Alzheimer’s Disease Cell Model Induced by Aβ 25-35 and Its Mechanism Based on Sirtuin1/Nrf2/HO-1 Pathway. Biomed Res Int 2020, 1–10. [Google Scholar] [CrossRef]
- Ishisaka, A.; Mukai, R.; Terao, J.; Shibata, N.; Kawai, Y. Specific Localization of Quercetin-3-O-Glucuronide in Human Brain. Arch Biochem Biophys 2014, 557, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.; Ullah, H.; Aschner, M.; Cheang, W.S.; Akkol, E.K. Neuroprotective Effects of Quercetin in Alzheimer’s Disease. Biomolecules 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Ho, L.; Ferruzzi, M.G.; Janle, E.M.; Wang, J.; Gong, B.; Chen, T.Y.; Lobo, J.; Cooper, B.; Wu, Q.L.; Talcott, S.T.; et al. Identification of Brain-Targeted Bioactive Dietary Quercetin-3-O- Glucuronide as a Novel Intervention for Alzheimer’s Disease. FASEB Journal 2013, 27, 769–781. [Google Scholar] [CrossRef]
- Shi, C.; Zhao, L.; Zhu, B.; Li, Q.; Yew, D.T.; Yao, Z.; Xu, J. Protective Effects of Ginkgo Biloba Extract (EGb761) and Its Constituents Quercetin and Ginkgolide B against β-Amyloid Peptide-Induced Toxicity in SH-SY5Y Cells. Chem Biol Interact 2009, 181, 115–123. [Google Scholar] [CrossRef]
- Zhang, X.D.; Liu, X.Q.; Kim, Y.H.; Whang, W.K. Chemical Constituents and Their Acetyl Cholinesterase Inhibitory and Antioxidant Activities from Leaves of Acanthopanax Henryi: Potential Complementary Source against Alzheimer’s Disease. Arch Pharm Res 2014, 37, 606–616. [Google Scholar] [CrossRef]
- Miriyala, S.; K. Holley, A.; St Clair, D.K. Mitochondrial Superoxide Dismutase—Signals of Distinction. Anticancer Agents Med Chem 2012, 11, 181–190. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, S.; Li, J.; Sun, Y.; Hasimu, H.; Liu, R.; Zhang, T. Quercetin Protects Human Brain Microvascular Endothelial Cells from Fibrillar β-Amyloid1-40-Induced Toxicity. Acta Pharm Sin B 2015, 5, 47–54. [Google Scholar] [CrossRef]
- Sabogal-Guáqueta, A.M.; Muñoz-Manco, J.I.; Ramírez-Pineda, J.R.; Lamprea-Rodriguez, M.; Osorio, E.; Cardona-Gómez, G.P. The Flavonoid Quercetin Ameliorates Alzheimer’s Disease Pathology and Protects Cognitive and Emotional Function in Aged Triple Transgenic Alzheimer’s Disease Model Mice. Neuropharmacology 2015, 93, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Paula, P.C.; Maria, S.G.A.; Luis, C.H.; Patricia, C.G.G. Preventive Effect of Quercetin in a Triple Transgenic Alzheimer’s Disease Mice Model. Molecules 2019, 24. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.M.; Li, S.Q.; Wu, W.L.; Zhu, X.Y.; Wang, Y.; Yuan, H.Y. Effects of Long-Term Treatment with Quercetin on Cognition and Mitochondrial Function in a Mouse Model of Alzheimer’s Disease. Neurochem Res 2014, 39, 1533–1543. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, M.; Itoh, M.; Ohta, K.; Li, S.; Ueda, M.; Wang, M.X.; Nishida, E.; Islam, S.; Suzuki, C.; Ohzawa, K.; et al. Quercetin Reduces EIF2α Phosphorylation by GADD34 Induction. Neurobiol Aging 2015, 36, 2509–2518. [Google Scholar] [CrossRef]
- JUNG, S.H.; Murphy, E.A.; McClellan, J.L.; Carmichael, M.D.; Davis, J.M. The Dietary Flavonoid Quercetin Decreases Neuroinflammation in a Mouse Model of Alzheimer’s Disease. The FASEB Journal 2010, 24. [Google Scholar] [CrossRef]
- Godoy, J.A.; Lindsay, C.B.; Quintanilla, R.A.; Carvajal, F.J.; Cerpa, W.; Inestrosa, N.C. Quercetin Exerts Differential Neuroprotective Effects Against H2O2 and Aβ Aggregates in Hippocampal Neurons: The Role of Mitochondria. Mol Neurobiol 2017, 54, 7116–7128. [Google Scholar] [CrossRef]
- Li, Y.; Tian, Q.; Li, Z.; Dang, M.; Lin, Y.; Hou, X. Activation of Nrf2 Signaling by Sitagliptin and Quercetin Combination against β-Amyloid Induced Alzheimer’s Disease in Rats. Drug Dev Res 2019, 80, 837–845. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, J.; Zhong, L.; Wang, N.; Yang, L.; Liu, C.C.; Li, H.; Wang, X.; Zhou, Y.; Zhang, Y.; et al. Quercetin Stabilizes Apolipoprotein e and Reduces Brain Aβ Levels in Amyloid Model Mice. Neuropharmacology 2016, 179–192. [Google Scholar] [CrossRef]
- Moreno, L.C.G. e. I.; Puerta, E.; Suárez-Santiago, J.E.; Santos-Magalhães, N.S.; Ramirez, M.J.; Irache, J.M. Effect of the Oral Administration of Nanoencapsulated Quercetin on a Mouse Model of Alzheimer’s Disease. Int J Pharm 2017, 14, 927–937. [Google Scholar] [CrossRef] [PubMed]
- Olayinka, J.N.; Eduviere, A.; Adeoluwa, O.; Akinluyi, E.; Obisesan, A.; Akawa, O.; Adebanjo, A. Quercetin Mitigates Scopolamine-Induced Memory Dysfunction: Impact on Oxidative Stress and Cholinergic Mechanisms. Metab Brain Dis 2022, 37, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Palle, S.; Neerati, P. Quercetin Nanoparticles Attenuates Scopolamine Induced Spatial Memory Deficits and Pathological Damages in Rats. Bulletin of Faculty of Pharmacy, Cairo University 2017, 55, 101–106. [Google Scholar] [CrossRef]
- Sharma, D.R.; Wani, W.Y.; Sunkaria, A.; Kandimalla, R.J.; Sharma, R.K.; Verma, D.; Bal, A.; Gill, K.D. Quercetin Attenuates Neuronal Death against Aluminum-Induced Neurodegeneration in the Rat Hippocampus. Neuroscience 2016, 324, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Wani, W.Y.; Gudup, S.; Sunkaria, A.; Bal, A.; Singh, P.P.; Kandimalla, R.J.L.; Sharma, D.R.; Gill, K.D. Protective Efficacy of Mitochondrial Targeted Antioxidant MitoQ against Dichlorvos Induced Oxidative Stress and Cell Death in Rat Brain. Neuropharmacology 2011, 61, 1193–1201. [Google Scholar] [CrossRef]
- Hou, Y.; Aboukhatwa, M.A.; Lei, D.L.; Manaye, K.; Khan, I.; Luo, Y. Anti-Depressant Natural Flavonols Modulate BDNF and Beta Amyloid in Neurons and Hippocampus of Double TgAD Mice. Neuropharmacology 2010, 58, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Barbagallo, M. Type 2 Diabetes Mellitus and Alzheimer’s Disease. World J Diabetes 2014, 5, 889–893. [Google Scholar] [CrossRef]
- Tumminia, A.; Vinciguerra, F.; Parisi, M.; Frittitta, L. Type 2 Diabetes Mellitus and Alzheimer’s Disease: Role of Insulin Signalling and Therapeutic Implications. Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef]
- Xu, W.L.; Von Strauss, E.; Qiu, C.X.; Winblad, B.; Fratiglioni, L. Uncontrolled Diabetes Increases the Risk of Alzheimer’s Disease: A Population-Based Cohort Study. Diabetologia 2009. [Google Scholar] [CrossRef]
- Michailidis, M.; Moraitou, D.; Tata, D.A.; Kalinderi, K.; Papamitsou, T.; Papaliagkas, V. Alzheimer’s Disease as Type 3 Diabetes: Common Pathophysiological Mechanisms between Alzheimer’s Disease and Type 2 Diabetes. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Kandimalla, R.; Thirumala, V.; Reddy, P.H. Is Alzheimer’s Disease a Type 3 Diabetes? A Critical Appraisal. Biochim Biophys Acta Mol Basis Dis 2017, 1863, 1078–1089. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Ta, Q.T.H.; Nguyen, T.K.O.; Nguyen, T.T.D.; Giau, V. Van Type 3 Diabetes and Its Role Implications in Alzheimer’s Disease. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Halli-Tierney, A.D.; Luker, J.; Carroll, D.G. Parkinson Disease. Am Fam Physician 2020. [Google Scholar] [CrossRef]
- Vargas-Restrepo, F.; Sabogal-Guáqueta, A.M.; Cardona-Gómez, G.P. Quercetin Ameliorates Inflammation in CA1 Hippocampal Region in Aged Triple Transgenic Alzheimer’s Disease Mice Model. Biomedica 2018, 38, 1–23. [Google Scholar] [CrossRef]
- Benameur, T.; Soleti, R.; Porro, C. The Potential Neuroprotective Role of Free and Encapsulated Quercetin Mediated by Mirna against Neurological Diseases. Nutrients 2021, 13. [Google Scholar] [CrossRef]
- Bournival, J.; Plouffe, M.; Renaud, J.; Provencher, C.; Martinoli, M.G. Quercetin and Sesamin Protect Dopaminergic Cells from MPP +-Induced Neuroinflammation in a Microglial (N9)-Neuronal (PC12) Coculture System. Oxid Med Cell Longev 2012. [Google Scholar] [CrossRef]
- Bureau, G.; Longpré, F.; Martinoli, M.G. Resveratrol and Quercetin, Two Natural Polyphenols, Reduce Apoptotic Neuronal Cell Death Induced by Neuroinflammation. J Neurosci Res 2008, 86, 403–410. [Google Scholar] [CrossRef]
- Amanzadeh, E.; Esmaeili, A.; Rahgozar, S.; Nourbakhshnia, M. Application of Quercetin in Neurological Disorders: From Nutrition to Nanomedicine. Rev Neurosci 2019, 30, 555–572. [Google Scholar] [CrossRef]
- Zhu, M.; Han, S.; Fink, A.L. Oxidized Quercetin Inhibits α-Synuclein Fibrillization. Biochim Biophys Acta Gen Subj 2013, 1830, 2872–2881. [Google Scholar] [CrossRef]
- Magalingam, K.B.; Radhakrishnan, A.; Haleagrahara, N. Protective Effects of Flavonol Isoquercitrin, against 6-Hydroxy Dopamine (6-OHDA)—Induced Toxicity in PC12 Cells. BMC Res Notes 2014, 7. [Google Scholar] [CrossRef]
- Magalingam, K.B.; Radhakrishnan, A.; Haleagrahara, N. Protective Effects of Quercetin Glycosides, Rutin, and Isoquercetrin against 6-Hydroxydopamine (6-OHDA)-Induced Neurotoxicity in Rat Pheochromocytoma (PC-12) Cells. Int J Immunopathol Pharmacol 2016, 29, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Pany, S.; Pal, A.; Sahu, P.K. Neuroprotective Effect of Quercetin in Neurotoxicity Induced Rats: Role of Neuroinflammation in Neurodegeneration. Asian Journal of Pharmaceutical and Clinical Research 2014, 7, 152–156. [Google Scholar]
- Singh, S.; Jamwal, S.; Kumar, P. Neuroprotective Potential of Quercetin in Combination with Piperine against 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine-Induced Neurotoxicity. Neural Regen Res 2017, 12, 1137–1144. [Google Scholar] [CrossRef]
- Lv, C.; Hong, T.; Yang, Z.; Zhang, Y.; Wang, L.; Dong, M.; Zhao, J.; Mu, J.; Meng, Y. Effect of Quercetin in the 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine- Induced Mouse Model of Parkinson’s Disease. Evidence-based Complementary and Alternative Medicine, 2016; 1–6. [Google Scholar] [CrossRef]
- Wattanathorn, J.; Sriraksa, N.; Muchimapura, S.; Tiamkao, S.; Brown, K.; Chaisiwamongkol, K. Cognitive-Enhancing Effect of Quercetin in a Rat Model of Parkinson’s Disease Induced by 6-Hydroxydopamine. Evidence-based Complementary and Alternative Medicine, 2016. [Google Scholar] [CrossRef]
- Mehdizadeh, M.; Joghataei, M.T.; Nobakht, M.; Aryanpour, R. Neuroprotective Effect of Quercetin in a Model of Parkinson’s Disease in Rat: A Histochemical Analysis. Basic Clin Neurosci 2009, 1, 3–6. [Google Scholar]
- Karuppagounder, S.S.; Madathil, S.K.; Pandey, M.; Haobam, R.; Rajamma, U.; Mohanakumar, K.P. Quercetin Up-Regulates Mitochondrial Complex-I Activity to Protect against Programmed Cell Death in Rotenone Model of Parkinson’s Disease in Rats. Neuroscience 2013, 236, 136–148. [Google Scholar] [CrossRef] [PubMed]
- Madiha, S.; Batool, Z.; Tabassum, S.; Liaquat, L.; Sadir, S.; Shahzad, S.; Naqvi, F.; Saleem, S.; Yousuf, S.; Nawaz, A.; et al. Quercetin Exhibits Potent Antioxidant Activity, Restores Motor and Non-Motor Deficits Induced by Rotenone Toxicity. PLoS One 2021, 16. [Google Scholar] [CrossRef]
- Denny Joseph, K.M. ; Muralidhara Combined Oral Supplementation of Fish Oil and Quercetin Enhances Neuroprotection in a Chronic Rotenone Rat Model: Relevance to Parkinson’s Disease. Neurochem Res 2015, 40, 894–905. [Google Scholar] [CrossRef]
- Bombardi Duarte, A.C.; Santana, M.G.; di Camilo Orfali, G.; de Oliveira, C.T.P.; Priolli, D.G. Literature Evidence and ARRIVE Assessment on Neuroprotective Effects of Flavonols in Neurodegenerative Diseases’ Models. CNS Neurol Disord Drug Targets 2017, 17, 34–42. [Google Scholar] [CrossRef]
- Sandhir, R.; Mehrotra, A. Quercetin Supplementation Is Effective in Improving Mitochondrial Dysfunctions Induced by 3-Nitropropionic Acid: Implications in Huntington’s Disease. Biochim Biophys Acta Mol Basis Dis 2013, 1832, 421–430. [Google Scholar] [CrossRef]
- Chakraborty, J.; Singh, R.; Dutta, D.; Naskar, A.; Rajamma, U.; Mohanakumar, K.P. Quercetin Improves Behavioral Deficiencies, Restores Astrocytes and Microglia, and Reduces Serotonin Metabolism in 3-Nitropropionic Acid-Induced Rat Model of Huntington’s Disease. CNS Neurosci Ther 2014, 20, 10–19. [Google Scholar] [CrossRef]
- Jain, D.; Gangshettiwar, A. Combination of Lycopene, Quercetin and Poloxamer188 Alleviates Anxiety and Depression in 3-Nitropropionic Acid-Induced Huntingtons Disease in Rats. J Intercult Ethnopharmacol 2014. [Google Scholar] [CrossRef] [PubMed]
- Denny Joseph, K.M. ; Muralidhara Enhanced Neuroprotective Effect of Fish Oil in Combination with Quercetin against 3-Nitropropionic Acid Induced Oxidative Stress in Rat Brain. Prog Neuropsychopharmacol Biol Psychiatry 2013. [Google Scholar] [CrossRef] [PubMed]
- Kuhad, A.; Singla, S.; Arora, V.; Chopra, K. Neuroprotective Effect of Sesamol and Quercetin against QA Induced Neurotoxicity: An Experimental Paradigm of Huntington’s Disease. J Neurol Sci 2013, 333. [Google Scholar] [CrossRef]
- Patel, D.C.; Tewari, B.P.; Chaunsali, L.; Sontheimer, H. Neuron–Glia Interactions in the Pathophysiology of Epilepsy. Nat Rev Neurosci 2019, 20, 282–297. [Google Scholar] [CrossRef]
- Sefil, F.; Kahraman, I.; Dokuyucu, R.; Gokce, H.; Ozturk, A.; Tutuk, O.; Aydin, M.; Ozkan, U.; Pinar, N. Ameliorating Effect of Quercetin on Acute Pentylenetetrazole Induced Seizures in Rats. Int J Clin Exp Med 2014, 7, 2471–2477. [Google Scholar]
- Choudhary, N.; Bijjem, K.R. V.; Kalia, A.N. Antiepileptic Potential of Flavonoids Fraction from the Leaves of Anisomeles Malabarica. J Ethnopharmacol 2011, 135, 238–242. [Google Scholar] [CrossRef]
- Nassiri-Asl, M.; Moghbelinejad, S.; Abbasi, E.; Yonesi, F.; Haghighi, M.R.; Lotfizadeh, M.; Bazahang, P. Effects of Quercetin on Oxidative Stress and Memory Retrieval in Kindled Rats. Epilepsy and Behavior 2013, 28, 151–155. [Google Scholar] [CrossRef]
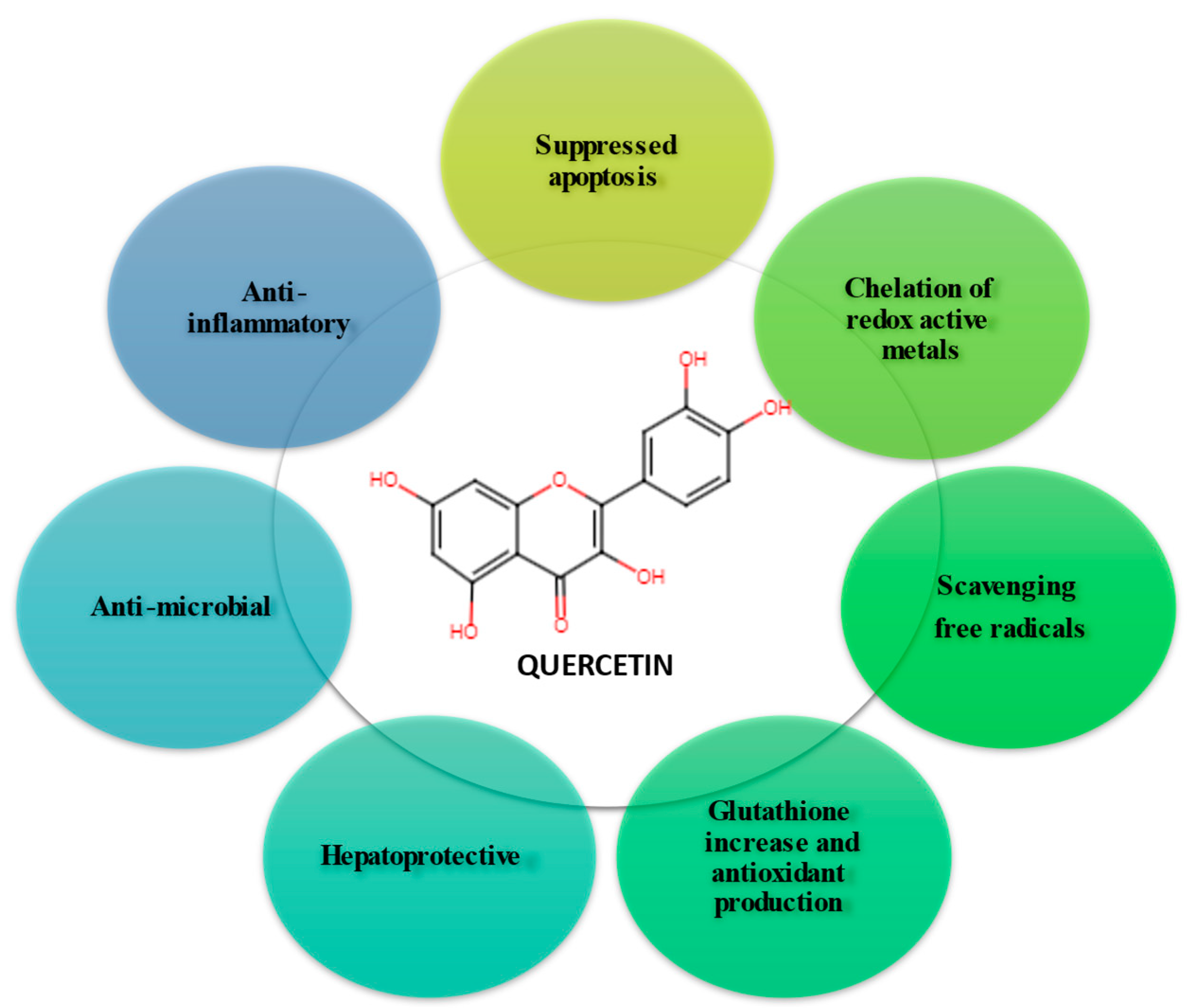
Figure 2.
Diagram with possible neuroprotective effects of quercetin. Abbreviations: ↑, increase; ↓, decrease; Aβ, amyloid beta-peptide; CAT, catalase; GPx, superoxide dismutase; GSH, glutathione; OS, oxidative stress; ROS, reactive oxygen species; SOD, superoxide dismutase [14,16,47].
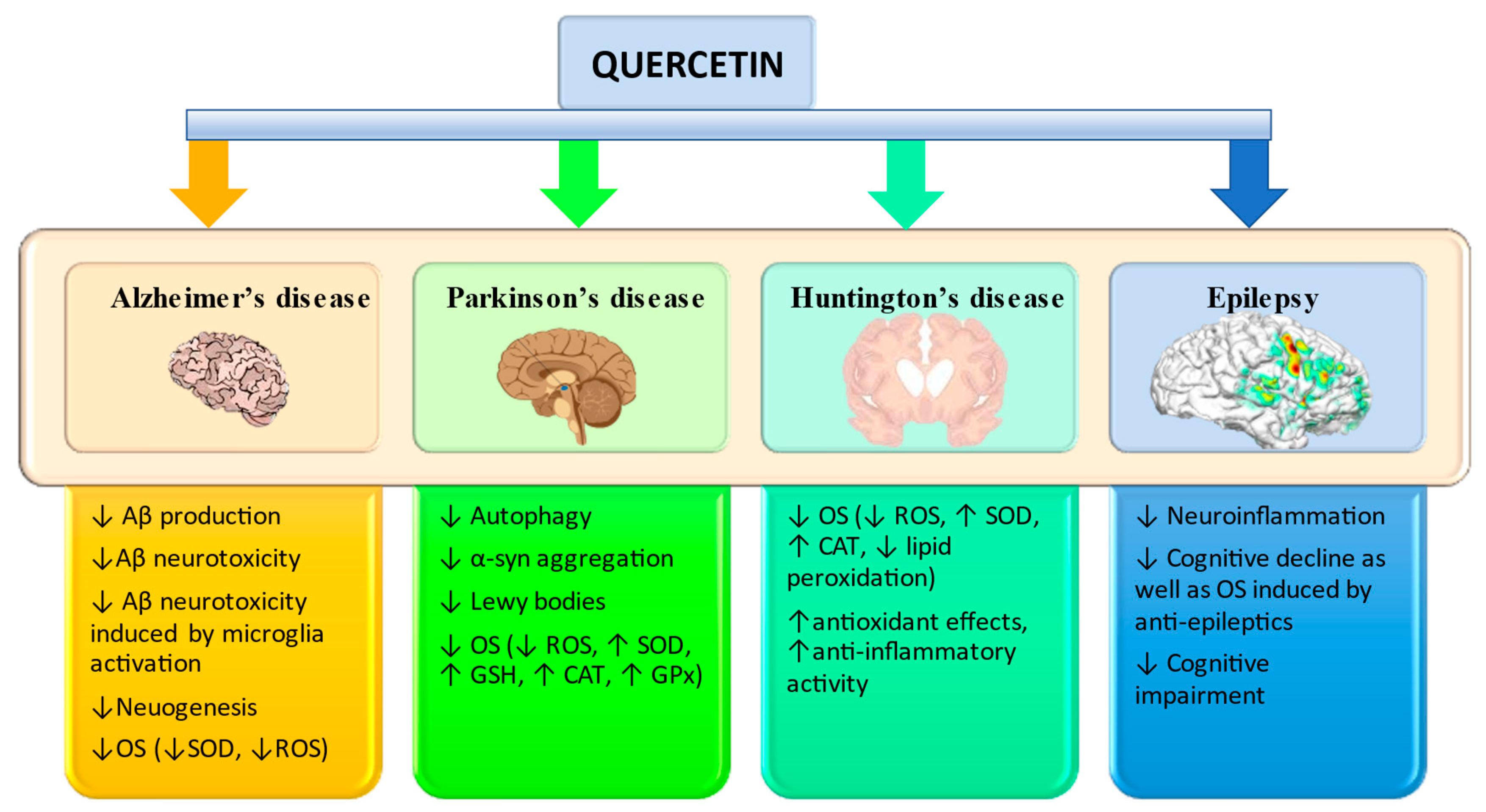
Table 1.
Quercetin content (mg 100 g−1or mg 100 mL−1) in selected foods and beverages [18].
Table 1.
Quercetin content (mg 100 g−1or mg 100 mL−1) in selected foods and beverages [18].
| Source | Quercetin | |||
|---|---|---|---|---|
| Food | Common Name | Scientific Name | Active Portions | mg 100 g−1 weight |
| Fruits | Acerola | Malpighia emarginata | Fruits | 4.74 |
| Apple | Malus domestica | Fruits | 19.36 | |
| Cranberry | Vaccinium oxycoccus | Fruits | 25.0 | |
| Apricots | Prunus armeniaca | Fruits | 1.63 | |
| Blackberries | Rubus spp. | Fruits | 3.58 | |
| Blueberries | Vaccinium spp. | Fruits | 7.67 | |
| Cherries | Prunus avium | Fruits | 17.44 | |
| Cranberries | Vaccinium macrocarpon | Fruits | 14.84 | |
| Grapefruit | Citrus paradisi | Fruits | 0.50 | |
| Grapes | Vitis vinifera | Fruits | 1.04 | |
| Vegetables | Capers, raw | Capparis spinosa | Flower buds | 233.84 |
| Onions, raw | Allium cepa | Bulbs | 20.30 | |
| Dill weed, fresh | Anethum graveolens | Leaves | 55.15 | |
| Oregano | Origanum vulgare | Leaves | 42.00 | |
| Tarragon, fresh | Artemisia dracunculus | Leaves | 10.00 | |
| Chicory | Cichorium intybus | Leaves | 6.49 | |
| Beverage | mg 100 mL−1 | |||
| Black tea | 2.50 | |||
| Red wine | 3.16 |
Source: Phenol Explorer and USDA Database for the Flavonoid Content of Selected Foods.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



