
Submitted:
30 May 2023
Posted:
31 May 2023
You are already at the latest version
Abstract
Bread wheat (Triticum aestivum L.) is one of the world's most important staple food crop providing 20% global energy and dietary proteins. It is widely grown in sub-tropical and tropical areas and as such exposed to heat-stress especially at grain filling period (GFP). Global warming has further affected its production and productivity in these heat-stressed environments. We examined the effect of heat-stress on 18 morpho-physiological and yield-related traits in 96 bread wheat accessions. Heat susceptibility index (HSI <0.60) and yield stability (i <0.55) used as criteria for selecting the tolerant accessions. Heat-stress, imposed by delayed sowing, decreased crop growth and GFP, and as a result reduced morphological and yield-related traits, namely days to 50% anthesis, plant height, peduncle length, flag-leaf area, spike length, spikelets per spike, grain length and width, thousand grain weight (TGW), harvest index and yield. The reduction in the trait values was severe in susceptible accessions (48.2% yield reduction in IC277741) than the tolerant. Physiological traits like chlorophyll content, canopy temperature depression (CTD), normalized difference vegetation index (NDVI), plant waxiness and leaf rolling showed higher expression in the tolerant accessions under heat-stress. Scanning electron microscopy of matured wheat grains revealed ultrastructural changes in endosperm and aleurone cells caused due to heat-stress. The reduction of size and density of large starch granules is the major cause of yield and TGW decrease in the heat-stress susceptible lines. The most stable and high-yielding accessions namely IC566223, IC128454, IC335792, EC576707, IC535176, IC529207, IC446713 and IC416019 were identified as the climate-smart germplasm lines. Germplasm lines possessing desirable traits were selected as po-tential parents for the development of bi-parental and multi-parental populations.
Keywords:
Bread wheat
; climate-smart germplasm
; adaptation
; global warming
; heat tolerance
; yield stability
1. Introduction
Bread wheat (Triticum aestivum L., 2n = 6X = 42, BBAADD), an important cereal crop, is a staple food for 40% of the world's population [1]. It provides 20% of the total dietary calories consumed globally [2], and contributes proteins, vitamins, dietary fiber and phytochemicals to human diet and for health benefits [3]. Wheat was cultivated on 219.0 million ha land with a global production of 760.9 million tonnes during 2020 and it contributed 8% to the world's food basket [4]. The demand of wheat is expected to rise by 60% from today's level by 2050 and its production is expected to decrease by ~30% during this period due to extreme weather events [5]. Climate changes have impacted the agriculture production and productivity globally during the past decades and seriously threatened the food supply [6,7]. Therefore, major advances in global food systems are required to ensure food security to burgeoning human population, which is expected to reach 10 billion by 2050 and for sustainable development [8]. With increasing climate instability due to global warming, agricultural productivity will continue to be negatively impacted [9,10]. Inter-Governmental Panel on Climatic Change (IPCC) 2018 report has predicted an increase in the global mean surface temperature between 3-4°C by 2100 relative to 1986-2005 if carbon emissions continue at the current rate [11]. Studies using global models suggest that every 1°C rise in average global surface temperature will lead to decline in wheat yields from 4.1 to 6.4% worldwide and 8.0% in India [12].
The narrow genetic base of elite cultivars further emphasizes the need to utilize the available germplasm in genebanks to enhance resilience to abiotic and biotic stresses [13]. Efficient utilization of germplasm resources is crucial for enhancing the genetic gains to address the challenges posed by global warming [14]. Genetic diversity is paramount for genetic improvement of cultivated crops, and for bread wheat this resides in three genomes, which were constituted by polyploidization of ancestral diploid species [15]. The bread wheat was originated in Fertile Crescent after few events of initial allopolyploidization and its cultivation spread to all continents except Antarctica [16]. During the course of evolution and later on its world-wide cultivation, bread wheat has adapted to various agro-ecologies from temperate (cold) to sub-tropical (hot and dry) and tropical (hot and humid) environments. The widely adapted germplasm with tolerance to abiotic and biotic stresses is being conserved in genebanks of national and international research institutes (e.g. World Wheat Collection housed at CIMMYT, Mexico), which needs to be utilized in breeding programmes for further enhancing the genetic gains in wheat [17,18].
Late sowing of bread wheat is common in western part of Indo-Gangetic Plain (IGP) region of India, where popular rice-wheat, potato-wheat and sugarcane-wheat cropping systems are followed, which delays sowing of bread wheat exposing it to terminal-heat stress. A large area in eastern and central part of India is also under late-sown condition due to delayed harvesting of Kharif crops and in such areas the wheat crop is exposed to terminal-heat stress resulting in significant yield losses [19,20]. IGP contributes about 15% of global wheat production, however, ~51% of its area might be re-classified as a heat-stressed (HS), short season production mega- environment by 2050 [21]. The significant part of eastern IGP, peninsular and central part of India are under severe heat-stress, but it is moderate in north-western parts of IGP [22,23]. Globally, high temperatures during grain filling period (GFP) is a major yield reducing factor in many wheat growing areas [24]. High temperature-stress during GFP, referred to as terminal-heat stress, adversely affects plant growth and grain yield. Every 1°C rise in temperature above the optimum of 28°C during GFP has resulted in yield losses of 3-17% in Great Plains of USA and the eastern Gangetic Plains of India [25,26]. Hence, study on impact of heat-stress on productivity of wheat in these regions has emerged as top priority in the climate change scenario [22,27] and as a novel strategy in wheat breeding for HS environment should focus on the ideotype development [28].
Several adaptive morpho-physiological traits like early ground cover, epicuticular wax, leaf rolling, stay-green, biomass, and flag-leaf area contribute toward heat-stress tolerance ability of wheat plant [29,30]. The adaptive physiological traits such CTD, cell membrane thermo-stability, NDVI, chlorophyll content and fluorescence are associated with heat-stress tolerance and significantly contribute to the performance of the tolerant wheat lines in HS environment during GFP [31,32,33]. Heat-stress during crop growth and grain development stage significantly reduces morphological and yield-realated traits viz., plant height, tillers′ number, peduncle length, spike length, spikelets and grains per spike, TGW and yield in wheat [34,35]. Endosperm shrinkage in the wheat grains of heat-stressed plants is a major cause of yield and TGW reduction [36,37]. Heat-stress causes damage to cellular structure and affects various metabolic pathways, mainly those related to membrane thermostability, photosynthesis and starch synthesis [38,39,40]. Each genotype responds to changed environment differently due to its genetic makeup and interaction with the environment [19,41]. The genotypes, which maintain high TGW and yield under HS environment, seem to possess higher tolerance to hot environments [42,43]. Exposure of wheat plants at anthesis and during GFP to higher than the optimal temperature affects the seed development and as a result reduces productivity [44,45]. The knowledge of source-sink relationships during grain development is critical for the selection of germplasm that can produce better yield and quality under global warming [46]. Thus, understanding of sink-source relationship and plant metabolism under heat- stress in a wide range of wheat germplasm could be useful for selecting tolerant lines.
Genotype-Environment (G × E) interactions are important factors in the expression of quantitative traits such as yield and its component traits, and are major bottlenecks in selection of superior breeding lines. Stability analysis has been used to identify the varieties with superior performance and yield stability under different environments [47]. Germplasm offers the best opportunity to develop varieties which show small G × E interaction. Climate-smart varieties tolerate negative effects of climate change better and produce higher yield and better quality in unfavourable environments such as heat, cold, salinity, drought etc. Therefore, selection of climate-smart germplasm by screening a large genebank collection is a right approach to develop climate-resilience [13]. The evaluation of diverse germplasm under HS environment would be useful in understanding the complexity of plant responses to heat-stress and identification of superior lines for the development of mapping populations, which might be utilized further in development of heat-stress tolerant cultivars [48]. The morpho-physiological traits have been used to screen germplasm for heat stress tolerance in numerous studies world-wide [24,49]. Thus, aims of the study were to identify superior germplasm based on morpho-physiological traits and yield stability, and to select desirable parents for the creation of bi-parental and MAGIC (Multi-parent Advanced Generation Inter-Cross) populations.
2. Materials and Methods
2.1. Plant Materials
Plant materials for the present study consisted of a subset of 96 bread wheat accessions (79 indigenous and 17 exotic collections), selected from a large genebank collection of wheat [50], which included some of released varieties for late sown condition, trait-specific germplasm lines and genetic stocks. Seeds of each accession were obtained from the working collection of National Gene Bank (NGB) at ICAR-National Bureau of Plant Genetic Resources (NBPGR), New Delhi. Passport data of wheat accessions used for this study are provided in Supplementary Table S1. Geo-referencing of bread wheat accessions on the world map was done using software ‘DIVA-GIS’ [51]. The germplasm accessions chosen for the present study were genetically diverse and represented all wheat growing zones of India (Supplementary Figure S1).
2.2. Experimental Site, Design and Weather Conditions
Field experiments were conducted at Experimental Farm, ICAR-NBPGR, New Delhi, situated at latitude 28.649°N, longitude 77.152°E and altitude 220 m, during two successive years 2018-2019 and 2019-2020. The farm area is in north western plain zone (NWPZ) of wheat production with semi-arid and sub-tropical climate and sandy-loam alluvial soil, slightly alkaline in pH and low in organic matter content. In each crop season, two sowing dates were i) Normal (sown in 1st week of Dec.; NS environment) and ii) late sown wheat (sown in 1st week of Jan.; HS environment). Thus, late sown wheat accessions were exposed to high temperature stress during GFP. Combination of year and sowing dates made our field experiments with four environments, that included two NS and two HS environments for testing the stability of germplasm for heat-stress tolerance. The field trial was laid out in augmented block design (ABD) with five blocks, where four checks namely Raj3765, HD2932, WR544 and HD2967 were randomized and replicated in each block. Each experimental plot consisted of three rows of 2 m length with 25 cm spacing between rows (1.5 m2). The standard crop management practices for irrigated ecology followed for raising the healthy wheat crop. Weather parameters namely temperature and rainfall recorded during entire crop period from sowing to maturity for two crop seasons during 2018-19 and 2019-20. Crop duration expressed in standard meteorological weeks (SMW).
2.3. Field Phenotyping and Data Recording
The morpho-physiological and yield related traits for terminal heat-stress tolerance were recorded as per Manual on Physiological Breeding II: A field guide to wheat phenotyping [52]. Phenological parameters were recorded using Zadoks scale observing the phenotype of whole plot [53]. Eighteen traits were recorded during different growth phases of the bread wheat crop trials (Table 1).
2.4. Scanning Electron Microscopy
Mature grains from heat-stress tolerant and susceptible accessions of bread wheat were transverse sectioned into three small pieces. The middle sections were mounted on an aluminum stub using double side adhesive carbon tape. The specimens were uniformly coated with thin layer of gold-palladium using Emitech SC7620 sputter coater. Specimens were examined under Scanning Electron Microscope (Model Tescan Vega3, Tescan Analytics, France) operated at 10.0 kV using secondary electron detector. Aleurone layer, endosperm cells and starch granules were observed and their images captured.
2.5. Statistical Analysis
Adjusted mean values were used for the statistical analysis of ABD trial data for 18 morpho-physiological and yield related traits of both NS and HS environments over two years. Combined analysis of variance (ANOVA) was performed following a test of homogeneity of variances and applying Aitkin's transformation using SAS software version 9.4 [58]. Pearson's correlation coefficients (r) were derived using IBM SPSS statistics software version 20.0 [59] for both NS and HS environments. Cluster analysis was performed with Ward's minimum variance using Euclidean distance matrices and dendrogram was constructed using unweighted paired group method of arithmetic averages (UPGMA) algorithm with SAS software. We performed boxplot analysis for 18 quntitative traits in tolerant and susceptible accessions. The bread wheat accessions were categorized as heat-stress tolerant or susceptible based on heat susceptibility index (HSI) values calculated on grain yield (GY) data using following formula [60].
HSI = (1 - Xa/Xb)/(1 - Ya/Yb) ,
Where, Xa and Xb are mean value of GY of individual accession under HS and NS environment, respectively and Ya and Yb are mean value of GY of all accessions under HS and NS environment, respectively. HSI values were used to categorize bread wheat accessions as highly tolerant (< 0.50), tolerant (0.51-1.0), susceptible (1.0–1.50) and highly susceptible (>1.50). Phenotypic variance (σ2ph), genotypic variance (σ2g), phenotypic coefficient of variation (PCV), genotypic coefficient of variation (GCV), heritability (H′) in broad sense and genetic advance (GA) were calculated on mean data of four checks using analyzed using SAS software version 9.4.
Heritability (H) % = (σ2g/σ2ph) × 100 , and Genetic advance (GA) = H × k × σph
Where, σph is phenotypic standard deviation and k is constant, 2.06 at 5% selection intensity. The diversity estimates based on quantitative traits were derived utilizing Shannon-Weiner index [61]. The formula for calculating Shannon-Weiner index is as follows.
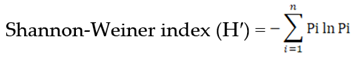
Where n is the number of phenotypic classes for a character, Pi is the relative frequency in the ith class of the jth trait and ln is the natural logarithm of Pi. The extent of diversity was interpreted as <0.50 value of H′: low diversity, 0.50 to 1.00 value of H′: high and >1.00 value of H′: very high diversity. The traits, which showed H′ >1.00, revealed the existence of great genetic diversity among the accessions.
The stability analysis for grain yield and 1000-grain weight was carried out using Eberhart and Russell (1966) model [62]. We performed statistical analyses using Windostat software version 9.2 (IndoStat Services, Hyderabad, India). The stability model used is as follows: Yij = µi + βiIj + δij
Where, Yij is accession mean of ith accession in jth environment, µi is ith accession mean over all environments, βi is regression coefficient of ith accession to varying environments, Ij is environmental index, and δij is deviation from regression of ith accession at jth environment. According to this model, an ideal (stable) accession has higher mean than the population mean, regression coefficient (βi = 1) and deviations from the regression (S2di) as small as possible. These accessions are well adapted to all environments. The accessions having higher mean value than the population mean, βi >1 and S2di as small as possible, are specifically adapted to favorable (NS) environment. The accessions exhibiting higher mean than the population mean, βi <1 and S2di as small as possible, are specially adapted to unfavourable (HS) environment. Bread wheat accessions classified into four categories namely highly tolerant, tolerant, susceptible and highly susceptible on the basis of mean performance, stability and HSI.
3. Results
3.1. Weather Conditions during Crop Seasons
The maximum and minimum temperatures and rainfall occurred during GFP of 2018-19 and 2019-20 crop growing seasons are presented in Figure 1.
Grain filling initiated from 8 SMW (18-24 Feb.) and completed during 14 SMW (1-7 Apr.) in normal sown crop (NS environment), while the GFP was from 12 SMW (18-24 Mar.) to 17 SMW (22-28 Apr.) for the late sown crop (HS environment) of both the seasons. Light to moderate rainfall occurred during GFP under NS environment in 2018-19, but it was moderate to heavy rainfall during 2019-20. The light rainfall during GFP slightly pulled down the rising day and night temperatures under HS environment in both the crop seasons. The HS environment (sown in January) was exposed to day temperatures of 35°C in 2018-19 and 33ºC in 2019-20 during the grain filling stage. Similarly, night temperature was also higher during GFP under the HS environment than the NS environment in both the crop seasons. On average, the maximum temperature was 7.7°C higher in 2018-19 and 5.7°C in 2019-20 under HS environment. Thus, the prevailing air temperature during GFP in late sown crop was significantly higher than the temperature needed for the normal grain filling (22-25°C). Hence, the late sown crops were exposed to the heat-stress during the grain filling stage.
3.2. Crop Growth and Genetic Variability
The differences in the climatic conditions in the environments were reflected by the variations observed for the phenological and agronomic traits as presented in Table 2. Significant differences observed for DA, PH, PL, FLA, SL, NSS and GFP between environments. Bread wheat accessions showed significant differences for all the traits except CTD, under both NS and HS environments (Supplementary Table S2). CTD showed large variation due to year effect and hence exhibited the non-significant variation for the genotype. The other physiological traits like NDVI and MSI also showed greater variation due to year than the genotype. Thus, the variation due to year was significant for almost all traits except CC, LR, NSS, HI and YPP, under both NS and HS environments. Genotype-year (G×Y) interaction showed non-significant differences in most of the traits.
NDVI had significant G×Y interaction in both NS and HS environments, while TGW showed significant G×Y interaction in NS environment. Morphological traits, namely PH, PL, SL and GL showed significant G×Y interaction under heat-stress. Thus the morpho- morphological and yield-related traits showed a wide-range of phenotypic variability under both NS and HS environments. Check variety WR544 (IC296383) had earliest anthesis on 83.4 and 66.6 days in normal and late-sown trials, respectively. Accessions 89 (IC393878) and 7 (IC252725) showed the highest GFP of 39.5 and 32.0 days under NS and HS environments, respectively. Accession 34 (EC577013) was the tallest (150.9 cm) under NS, while accession 84 (EC576585) was the tallest (127.0 cm) under HS environment. However, accession 38 (IC335792 produced the shortest plant (84.9 cm) under NS and accession 64 (IC443653) developed the shortest plants (73.2 cm) under heat-stress. Accession 9 (IC277741) produced the highest grain yield (802.5 g) in NS environment, while accession 91 (IC566223) produced the highest yield (598.1g) under HS environment. The lowest grain yield (300.1 g) produced in accession 26 (IC542509) under NS, whereas accession 15 (IC539287) produced the lowest yield (176.7 g) under HS environment. Accessions 58 (IC573461) produced bold grains (TGW = 52.2 g) under NS, while accession 14 (IC539221) produced bold grains (TGW=46.6 g) under HS environment. Accession 64 (IC443653) showed highest harvest index (48.0%) under HS environment.
Bread wheat accessions exhibited significant variation for most of the physiological traits. They showed high variability in physiological traits such as plant waxiness and leaf rolling (Figure 2). Accessions 43 (IC528965) and 60 (IC529207) developed the highest plant waxiness (score 10) under both the environments. Accession 22 (IC416019) showed the highest leaf rolling (score 9.5) in NS environment, whereas accessions 22 (IC416019) and 30 (IC416055) expressed the highest leaf rolling trait (score 10) under HS condition.
For chlorophyll content, accession 58 (IC573461) showed the highest value (38.5) under NS, while accession 59 (IC252444) had the highest CC (37.2) under HS condition. Accession 80 (IC542547) showed the lowest CC in both the environments. Accessions 26 (IC542509) and 91 (IC566223) showed the highest value of NDVI in both NS (0.72) and HS (0.64) environments. CTD was higher in HS than the NS environment. Accession 81 (EC190962) showed highest value of CTD (9.7°C) under NS environment, while accession 26 (IC542509) exhibited the highest CTD (12.0°C) in HS environment. The accessions 9 (IC277741) and 83 (EC573527) exhibited the highest value for MSI under NS (77.5%) environment, while accession 90 (IC542544) showed the highest value of MSI (72.6%) under HS environment. Detailed data for the morpho-physiological and yield-related traits of all accessions are shown in Supplementary Tables S3 and S4.
A wide-range of genetic variability was recorded for most of the traits (Table 2). PCV was higher than GCV for all the quantitative traits studied. The highest PCV was observed for CTD (26.1%) under HS environment followed by plant waxiness (23.8%) under NS environment. The highest GCV recorded for plant waxiness (19.6%) followed by leaf rolling (17.5 %) under NS environment. The estimates of high heritability (>70%,) were recorded for days to 50% anthesis (84.9% under NS and 85.4% under HS), grain length (77.9% under NS and 89.3% under HS) in both the environments, CC (75.9%) and PL (79.4%) under HS environment, and LR (71.4%) under NS environment. Genetic advance was the highest for PW (33.2 %) followed by LR (30.5%) under the NS environment. Shannon-Weiner diversity index (H′) revealed a high morpho- physiological diversity among studied bread wheat accessions considering all traits under NS (H′=1.56) and HS environments (H′=1.47). The most polymorphic traits were PW (H′=1.90), CTD (H′=1.88), NSS (H′=1.79) and NDVI (H′=1.78) under NS environment, while under the HS environment, the traits displaying high diversity were NSS (H′=1.78), PW (H′= 1.77), MSI (H′=1.70) and CTD (H′ =1.69). Grain length showed the least variability under both the NS (H′=1.19) and HS (H′= 1.14) environments. Frequency distribution of wheat accessions for 12 important phenological, physiological and yield-related traits, namely days to 50% anthesis, GFP, chlorophyll content, CTD, NDVI, MSI, plant waxiness, leaf rolling, plant height, peduncle length, TGW and grain yield also supported the existence of wide-range of genetic variability in the germplasm (Supplementary Figure S2).
3.3. Genetic Relationship in Wheat Accessions
UPGMA dendrogram grouped all the 96 bread wheat accessions into 6 major clusters using the data of 18 morpho-physiological and yield-related traits in HS environment (Figure 3). Most of the accessions (80 acc.) included in first three clusters with cluster-I, -II and -III consisted of 21, 42 and 17 accessions, respectively. Remaining three clusters grouped 16 accessions with cluster-IV, -V and -VI comprised of 10, 5 and 1, accessions, respectively. Most of the accessions adapted to heat-stress viz., accession number 91 (IC566223), 60 (IC529207), 12 (IC535176), 18 (EC534487), 27 (IC252348), 13 (IC401976), 25 (IC075240), 16 (IC539531) and 5 (EC574731) grouped in cluster-III along with two national check varieties, namely 2 (HD2932) and 4 (HD2967). Accession 26 (IC542509) took the longest time to 50% anthesis (119 days in NS and 94 days in HS environment), produced the lowest yield in both the NS and HS environments and formed a solitary accession in cluster VI. The accessions 84 (EC576585), 68 (IC536162), 70 (IC252999), 85 (EC190899), 56 (IC252867), 69 (IC536050), 20 (IC416075), 55 (IC572925), 64 (IC443653) and WR544 flowered early (<69 days) in HS environment and were grouped in cluster-I.
3.4. Correlations with Grain Yield
Grain yield associated positively with plant waxiness, leaf rolling, GFP, GW and HI under HS environment (Figure 4a). However, under NS environment, yield associated positively with CTD, TGW, GFP, GW and HI, but negatively with NDVI and DA (Figure 4b). TGW showed positive association with GFP, GL, GW and HI under both the environments, whereas under the heat-stress, TGW also associated positively with PH, PL and FLA, but negatively with MSI, LR and DA. The strong positive correlations recorded between PH and PL, and SL and NSS under both the environments. GFP associated positively with GW, TGW and HI, but negatively with DA under both the environments. Similarly, DA also associated negatively with HI, TGW and GW under both environments.
Among physiological traits, positive association was observed between plant waxiness and leaf rolling under both the environments. NDVI associated positively with both chlorophyll content and CTD under heat-stress. The cooler canopy (CTD) showed positive correlation with DA, PH, PL, FLA, SL and NSS under the HS environment. However, CTD associated positively with MSI under NS environment and negatively under the HS environment.
3.5. Impact of Heat-stress on Yield and Morpho-Physiological Traits
Late showing of bread wheat reduced the optimal period for crop growth and grain development. The unfavorable weather conditions under HS environment had an adverse effect on yield and its component traits, which showed reduced expression (Table 2). However, physiological adaptive traits namely CTD, plant waxiness and leaf rolling showed increased expression in HS environment. Phenological trait like days to 50% anthesis was reduced by 17 days under HS environment. Plant height, peduncle length and GFP traits were decreased in HS environment by 9.7 cm, 4.1 cm and 6 days, respectively. Likewise, heat-stress reduced flag leaf area by 13.6 cm².
High-temperature stress prevailed at the onset of reproductive phase and during grain development stage decreased spike length, number of spikelets per spike, and grain length and width. Furthermore, the heat-stress condition during GFP had adverse effect on grain yield and TGW, and consequently reduced these traits by 138.6 g and 6.0 g, respectively, under HS environment. The harvest index was decreased from 39.7 to 34.4 under HS environment. Among the physiological traits, chlorophyll content, NDVI and MSI were reduced by 4.2, 0.14 and 7.2%, respectively, under the HS environment. However, CTD was slightly increased from 6.2 to 6.9 °C under HS environment to maintain optimal sub-cellular temperature during GFP. Plant waxiness and leaf rolling were also increased from 6.2 to 6.9 and 5.8 to 6.5, respectively, under HS environment.
Boxplot analysis clearly showed differential effect of heat-stress on the morpho- physiological traits in tolerant and susceptible accessions (Figure 5). All the traits were reduced under the HS environment in both tolerant and susceptible accessions except CTD, PW and LR. However, mean values of CTD, PW and LR were higher under the HS environment in tolerant accessions. The heat-stress tolerant accessions showed higher mean values for all traits as compared to heat susceptible ones under HS environment except MSI. The higher MSI was observed in the susceptible accessions because of more leakage of cell contents at 100 °C and showed higher electrical conductivity as compared to the tolerant bread wheat accessions.
The effect of heat-stress on important morpho-physiological and yield- related traits was also investigated in five classes of accessions, namely highly tolerant, tolerant, susceptible and highly susceptible accessions along with check varieties (Figure 6). Under heat-stress, reduction of chlorophyll and NDVI was more evident in susceptible accessions as compared to the tolerant. However, under the HS environment, MSI showed relatively higher values in susceptible accessions than the tolerant accessions. CTD, PW and LR increased in the tolerant accessions as compared to the susceptible ones under HS environment. DA, PH, PL, FLA, GFP, SL, TGW, HI and GY all decreased in both tolerant and susceptible accessions under the heat-stress. The tolerant accessions produced higher yield and bold grains than the susceptible under the heat-stress mainly due to reduced number of grains per spike and faster grain developmentin under the HS environment.
3.6. Impact of Heat-stress on Wheat Grains
Heat-stress reduced grain width, length and TGW in all the accessions. However, the effect of the heat-stress during GFP on wheat grain traits (GL, GW and TKW) was more evident in susceptible than the tolerant accessions (Figure 5 and Figure 7). The reduction in GW was significantly high in the susceptible than the tolerant accessions in heat-stress (Figure 7a). Scanning electron microscopy of aleurone layer and endosperm was carried out in mature grains of heat tolerant and susceptible accessions of bread wheat grown under NS and HS environments (Figure 7b). Heat-stress during GFP adversely affected aleurone layer and endosperm of wheat grains.
The concentration and morphology of starch granules changed under heat stress. The size, shape and structure of aleurone layer and starch granules of heat-stress tolerant grains were quite different from the heat-stress susceptible genotypes. The aleurone layer was de-structured in susceptible accessions as compared to tolerant ones (Figure 7b, upper panel). Heat-stress showed adverse effects on structure and packing of starch granules. Robust, bold and well-structured large starch granules (LG) were observed in both heat-stress tolerant and susceptible accessions in NS environment, whereas un-structured, shrivelled LG with prominence of small starch granules (SG) were present in the wheat grains developed under HS environment. Density of LG (A-type; 15-35 µm) was higher in both heat-stress tolerant and susceptible accessions in NS environment, whereas, in HS environment the density of LG slightly reduced in tolerant accessions while it was considerably reduced in the susceptible accessions. The density of SG (B-type; 2-8 µm) was higher in both tolerant and susceptible accessions in the HS environment as compared to NS environment.
3.7. Selection of Heat-stress Adapted Germplasm
Grain yield and TGW were used for stability analysis using four environment datasets. ANOVA showed significant differences (p <0.01) among accessions (= Genotype, G) and environments (E) for both the traits (Table 3). The yield exhibited significant interactions for G × E, G × E (Linear), E + (G × E) and E (Linear). However, TGW revealed significant interactions for E + (G × E) and E (Linear), but non- significant for G × E and G × E (Linear). On the basis of mean performance, regression coefficient (βi) and HSI, wheat accessions were classified as adapted to unfavorable (HS) environment, favorable (NS) environment or both environments (Table 4). Accession number 91 (IC566223), 60 (IC529207), 38 (IC335792), 12 (IC535176), 6 (EC576707), 29 (IC128454), 22 (IC416019), 23 (IC446713), 41 (IC265318), 27 (IC252348), 13 (IC401976), 25 (IC075240) and 16 (IC539531) performed well under HS environment and were identified as highly tolerant genotypes. Accessions 65 (IC252431), 9 (IC277741), 85 (EC190899), 96 (CUO/79/Pru 11A), 95 (EC277134), 57 (IC524299), 94 (IC553599), 84 (EC576585), 58 (IC573461), 82 (EC576066) performed well in NS environment but poor in HS environment, were referred as highly susceptible ones. Accessions 89 (IC393878), 19 (IC416018), 14 (IC539221) and 17 (IC443661) performed well under both NS and the HS environments were called as general adapters.
Likewise, stability for TGW revealed that some accessions were well adapted to HS, NS or both environments (Table 4, Figure 8). The detail of mean performance and stability parameters of 96 bread wheat accessions for TGW and grain yield are presented in Supplementary Table S5. The accessions showing higher mean than the population mean, regression coefficient (βi <1) and HSI < 0.5 and producing more than 500 g grain yield were considered as highly tolerant genotypes (Supplementary Table S6). The top 10 accessions were screened out for various morphological, physiological, yield and its contributing traits under both the NS and HS environments (Supplementary Table S7).
We selected germplasm lines based on their performance under HS environment as potential parents for the development of bi-parent and MAGIC populations (Table 5). The germplasm accessions, which showed extreme phenotype under HS environment, were chosen as the potential parents for the creation of trait-specific bi-parental mapping populations. However, for the selection of promising parents for 4-parental and 8-parental MAGIC populations, accessions showing the highest expression of yield and its associated morpho-physiological traits were considered.
4. Discussion
The production and productivity of bread wheat is adversely affected due to terminal-heat stress as a result of delayed sowing in many parts of India [23]. In the present study, we have evaluated a diverse set of 96 accessions of bread wheat to analyse plant responses in terms of effect of heat-stress on morpho-physiological and yield-related traits. Proper endosperm development under heat-stress is the key to maintain TGW, yield level, grain quality in wheat, and to facilitate the selection of superior and stable genotypes under HS environment [42,63]. The heat-stress imposed by sowing the wheat trial in a very late condition during first week of January in both years, created a unique environment to test germplasm for heat-stress tolerance. The delayed sown wheat is exposed to higher temperatures at reproductive and grain filling stages, and is a widely used strategy to screen germplasm for yield and other traits under heat-stress [28,64]. Yield, a complex quantitative trait, is the end-product of many interactions between genes for physiological and yield component traits. High temperature stress has a wide range of effects on plants in terms of physiological, biochemical and gene regulation pathways [65].
4.1. Trait Variability and Impact of Heat-stress
Bread wheat accessions showed varied response to heat-stress and provided ample scope for further selection of trait-specific accessions for terminal heat-tolerance. The high genotypic variability obtained in present study is due to selection of diverse germplasm accessions for different traits related to heat-stress tolerance. Non-significant differences observed for genotype × year (G × Y) interaction for most of the traits reveal that accessions had similar expressions in both the years for these traits. Significant G × Y interactions for NDVI in both environments, TGW under NS, and plant height, peduncle length, spike length and grain length under HS environment indicate that accessions performed differently over the years for these traits. Our results corroborate with earlier studies [41,66], which also reported varied responses of different traits in different genotypes and their significant interactions with environment under the heat-stress. However, minor differences in the range of quantitative traits are expected in these studies due to differences in the genetic make-up accessions used and test environments. The heat-adapted genotypes with best yielding ability also possessed high early biomass, high grain filling rates and low canopy temperatures [67].
The response to heat stress involves physiological adaptations that are required to protect the cellular functions including photosynthesis, assimilate-partitioning and accumulation of secondary metabolites [65,68]. Heat-stress showed an adverse effect on crop growth and development, and consequently it negatively affected morphological and yield-related traits such as days to 50% anthesis, plant height, peduncle length, flag-leaf area, GFP, spike length, spikelets per spike, grain length and width, TGW, harvest index and yield. The physiological traits, namely chlorophyll content, NDVI and MSI were reduced, whereas CTD, PW and LR showed enhanced expressions under the HS environment. The reduction in the expression of traits was more prominent in susceptible accessions as compared to the tolerant ones. Under heat-stress, reduced expression of morphological and yield-related traits was also reported in several studies conducted across the globe [28, 34, 42, 43). CTD found to be a reliable and non-invasive method for selecting heat-stress tolerant lines and it showed the best results for evaluation of wheat genotypes under terminal heat stress condition [69].
Significantly reductions in the days to heading, GFP, TGW, harvest index and yield in late sown wheat were observed earlier [42] and well-supported our results in these traits. Similarly, reduction in days to heading, GFP, yield, plant height, harvest index and TGW under heat-stress was also reported in a previous study [34]. The extent of reduction of the traits varied in these studies due to use of different set of genotypes and test conditions. The yield and its component traits are more severely affected with the increase of heat-stress [43]. High temperature during post-anthesis stage reduced grain filling duration, yield and TGW [70]. The yield parameters were also reduced in plants exposed to high temperatures in wheat landraces [71]. Recently in India, the reduction of days to anthesis, MSI and yield traits was reported in late sown wheat compared to normal [72]. They also observed that HSI was the lowest in the heat tolerant germplasm lines and supports our observations on HSI. The heat-stress reduced plant height, days to anthesis and maturity, grain weight and yield between 4-7% with every 1°C rise in mean maximum temperature above the optimum of 25°C [28]. The higher leaf waxiness under HS condition reported earlier [30] supports our results of increased waxiness under heat-stress, which protects plant against excess radiation and water loss through reflection of visible and infrared wavelengths [73]. Heritability of waxiness was low because of significant environ ment interactions as also reported in previous study [74]. Genotypes with stay green traits performed better under heat stress and donor genotypes were identified for utilization in heat-stress tolerance breeding [75].
4.2. Association of Grain Yield with Other Traits
Yield, being a complex quantitative character, is determined by interaction between morphological, physiological and other related parameters. Under HS environment, yield showed positive correlations with GFP, HI, GW, PW and LR. The positive correlation of yield with GFP reveals that the longer grain development period is an important contributing factor for improving yield under HS environment. In a previous study [42], grain yield and HI showed positive correlation with TGW under both normal and terminal-heat stress conditions, whereas GFP positively associated with TGW only under heat-stress. This study advocates that the selection for low TGW reduction as an indirect criterion to identify high yielding lines under terminal heat-stress. In the present study, TGW was associated with yield only under the NS environment. However, another study [76] reported the association of TGW with yield under both optimal and HS environments. The positive association of yield with GFP and TGW was also reported under the heat-stress [77]. There was non-significant association of grain yield with days to 50% anthesis, plant height and TGW in present study under HS environment, which confirms the results of an earlier study [28]. Hence, to increase yield in wheat under HS environment, the focus should be given on traits which have high and significant association with yield for the selection of heat tolerant lines.
In our study, there was non-significant correlation between chlorophyll content and grain yield under both NS and HS environments, whereas, CTD was significant associated with grain yield only under NS environment. Similarly, Elbasyoni [41] also observed no correlation between chlorophyll content and grain yield. However, a high correlation of both CTD and chlorophyll content with yield under heat-stress environment was reported earlier [31,78]. The high genetic variation for chlorophyll retention and seed weight along with positive association between chlorophyll content and seed weight was found under heat stress condition in diverse elite winter wheat lines [79]. Similarly, association of canopy temperature with grain yield was also observed under both NS and HS environments [26]. Under heat-stress, the grain yield showed a strong positive correlation with both CTD and days to heading [80,81,82]. It was suggested that the longer time before heading enables the development of larger spikes and more numbers of spikelets producing enhanced grain numbers per plant [82]. The present and earlier association studies revealed that grain yield in bread wheat correlates positively with its contributing and physiological traits under HS environment. Therefore, selection of these traits could be valuable in wheat breeding programme designed for heat-stress tolerance.
4.3. Grain Development under Heat Stress
Heat stress during GFP adversely affects the grain size in bread wheat. In our study, the effect of heat-stress on grain width was more severe as compared to length, which resulted into shrivelled grains in susceptible accessions. The development of shrivelled grains in the susceptible accessions was due to changes in ultrastructure of aleurone cells and starch granules in endosperm. Our findings on grain development under heat-stress corroborate with a recent study [83], wherein they observed severe effect of heat stress on grain characters like grain length, width and area along with starch synthesis. The effect of heat-stress shows a significantly reduced grain width and perimeter. The endosperm of mature wheat grain contains two types of starch granules: large (10–35 µm) A-type and small (1–10 µm) B-type [84,85]. The density and size of large type of starch granules were slightly reduced in heat-stress tolerant accessions while it was considerably reduced in heat susceptible ones under HS environment. The concentration of small type of starch granules was higher in both heat tolerant and susceptible accession under HS environment as compared to NS environment. Our findings on the size and density of starch granules are well supported by previous study [86], where it reported differences in size, shape and structure of starch granules in wheat grain after heat-shock. They also observed that the ratio of large and small type of starch granules decreases significantly under heat-stress and this limits the potential sink size for dry matter deposition in grain. Dias et al. [37] also reported that the heat-stress during GFP triggers ultrastructural changes in the aleurone layer and endosperm cells and causes disordered cells, grain shrinkage and reduced weight in heat-stress susceptible genotypes.
Grain filling is influenced by various metabolic processes occurring in leaves, mainly the production and translocation of photoassimilates and importing of precursors for biosynthesis of grain reserves, minerals and other functional constituents [87]. It is, therefore, important to know the physiological, biochemical and genetic mechanisms, which govern the grain filling events under heat-stress to devise strategies for yield enhancement in wheat. Identification and selection of germplasm lines with higher yield and grain weight along with early maturity, semi-tall plant height, higher NDVI during grain filling and higher content of chlorophyll at milky stage should be the hallmark of breeding strategies for heat-stress tolerance [28,76]. Harnessing the genetic variability for these traits in the germplasm of bread wheat is vital for breeding of heat-stress tolerant cultivars [14].
4. Yield Stability and Selection of Accessions Adapted to Heat-stress
Heat-stress adaptation is a complex phenomenon and is influenced by several factors such as genotype and its interaction with environment over a period of time from decades to centuries. Wheat plant achieves adaptation through variation in phenology and related traits determining plant architecture [49]. Hence, it is crucial to understand genes that underpin the variations in plant phenology and their interactions with other genes, morpho-physiological traits and the environment. The yield stability across the environments is a reliable criterion for the selection of heat-stress adapted germplasm [47,88]. Climate change associated global warming has severely affected yield stability in cereal crops [89]. Genotype – environment interactions are of major importance to the plant breeder for developing improved varieties. We used stability analysis [62] to identify stable and better performing accessions under HS environment. The excellent performing accessions in HS environment may be used as donor parents for heat-stress adaptation breeding programme. Similar study was also carried [41] to select superior yielding lines under heat-stress. However, G × E interaction biplots for yield were used to select genotypes with stable performance across all environments [19]. In a similar study [35], genotype by environment interaction was used to identify superior yielding genotypes with heat stress tolerance. A selection strategy was also suggested to improve adaptation to heat stress in the bread wheat [28]. Heat stress affects significantly all the yield contributing traits and TGW an important trait for selection of tolerant lines. The heat tolerant lines with high grain yield could be selected using heat tolerance index and both AMMI and GGE biplot stability analyses [90]. The stable and higher yielding accessions identified under HS environment could be utilized in the breeding programme for the development of terminal heat-stressed tolerant cultivars.
5. Conclusions
Wheat genetic resources used in the present study represent a part of the reference set for heat-stress tolerance and hence exhibited high extent of genetic variability for morpho-physiological and yield-related traits. The terminal heat-stress during late sown bread wheat crop negatively affected yield and its contributing traits. The higher than the optimum temperature during GFP reduced the grain size, and eventually decreased TGW and yield under heat-stressed environment. Ultrastructural analysis of wheat grains showed that the decrease in size and density of large starch granules in the endosperm is the main cause of yield and TGW reduction in the heat-stress susceptible germplasm. The better yielding accessions possessing desirable morpho-physiological traits and adaptation to heat-stress identified, could be utilized for the development of mapping populations and genetic improvement of bread wheat.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Figure S1 to S2: Supplementary Table S1 to S7.
Author Contributions
Mahesh C. Yadav conceptualized and validated the study. Anil Patidar performed the experiments and recorded field and lab data. Jyoti Kumari provided germplasm lines and analysed quantitative data. Shailesh Tiwari performed data recording and crop management. Gautam Chawla performed ultrastructural analysis of wheat grains. MCY and AP wrote and prepared the original draft and finalized the manuscript. JK and Vijay Paul reviewed and edited the manuscript. MCY supervised the study, was in charge of the project administration, and acquired the funding for the study. All authors contributed to the research article and approved the submitted version.
Data availability statement
The original contributions presented in the study are included in the article/ Supplementary Materials. Further queries can be directed to the corresponding author.
Acknowledgments and Fundings
First author is grateful to the Director, ICAR-IARI, New Delhi for granting senior research fellowship during Ph.D. study. This work was funded by ICAR Network Project on National Innovations in Climate Resilient Agriculture (NICRA) entitled "Focused collection of climate-smart germplasm of rice and wheat, their valuation and genetic enhancement through pre-breeding for abiotic stress tolerance" with scheme code 13921 and project number 1006607.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Acevedo, M.; Zurn, J.D.; Molero, G.; Singh, P.; He, X.; Aoun, M.; McCandless, L. The role of wheat in global food security. In Agricultural Development and Sustainable Intensification: Technology and Policy Challenges in the Face of Climate Change, 1st Ed. New York, Routledge, 2018; pp. 81-110. [CrossRef]
- Shiferaw, B.; Smale, M.; Braun, H.J.; Duveiller, E.; Reynolds, M.; Muricho, G. Crops that feed the world 10. Past successes and future challenges to the role played by wheat in global food security. Food Sec. 2013, 5, 291–317. [Google Scholar] [CrossRef]
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef]
- FAO. World Food and Agriculture – Statistical Yearbook. 2022, Rome. [CrossRef]
- CIMMYT. FFAR grant develops climate-resilient wheat. CIMMYT press release 11 January, 2021. Available at https://www. cimmyt.org/news/ffar-grant-develops-climate-resilient-wheat/.
- Lobell, D.B.; Schlenker, W.; Costa-Roberts, J. Climate trends and global crop production since 1980. Science 2011, 333, 616–620. [Google Scholar] [CrossRef]
- Wheeler, T.; von Braun, J. Climate change impacts on global food security. Science 2013, 341, 508–513. [Google Scholar] [CrossRef]
- FAO. The future of Food and Agriculture – Trends and challenges. Food and Agriculture Organization of the United Nations, Rome, 2017; pp. 151.
- Wang, P.; Deng, X.; Jiang, S. Global warming, grain production and its efficiency: Case study of major grain production region. Ecol. Indic. 2018, 105, 563–570. [Google Scholar] [CrossRef]
- Massel, K.; Lam, Y.; Wong, A.C.S.; Hickey, L.T.; Borrell, A.K.; Godwin, I.D. Hotter, drier, CRISPR: the latest edit on climate change. Theor. Appl. Genet. 2021, 134, 1691–1709. [Google Scholar] [CrossRef]
- IPCC. Summary for Policymakers. In Global warming of 1.5°C - An IPCC Special Report on the impacts of global warming of 1.5°C. World Meteorological Organization, Geneva, Switzerland, 2018; pp. 1–32.
- Liu, B.; Asseng, S.; Müller, C.; Ewert, F.; Elliott, J.; Lobell, D.B.; et al. Similar estimates of temperature impacts on global wheat yield by three independent methods. Nat. Clim. Change 2016, 6, 1130–1136. [Google Scholar] [CrossRef]
- McCouch, S.R.; Navabi, Z.K.; Abberton, M.; Anglin, N.L.; Barbieri, R.L.; Baum, M.; et al. Mobilizing crop biodiversity. Mol. Plant 2020, 13, 1341–1344. [Google Scholar] [CrossRef]
- Reynolds, M.; Tattaris, M.; Cossani, C.M.; Ellis, M.; Yamaguchi-Shinozaki, K.; Pierre, C.S. Exploring genetic resources to increase adaptation of wheat to climate change. In Advances in Wheat Genetics: From Genome to Field. Ogihara, Y., Takumi, S., Handa, H., Eds.; Springer, Tokyo, 2015; pp. 355–368. [CrossRef]
- Mujeeb-Kazi, A.; Kazi, A.G.; Dundas, I.; Rasheed, A.; Ogbonnaya, F.; Kishii, M.; Bonnett, D.; et al. Genetic diversity for wheat improvement as a conduit to food security. Adv. Agron. 2013, 122, 179–257. [Google Scholar] [CrossRef]
- Matsuoka, Y. Evolution of polyploid Triticum wheats under cultivation: the role of domestication, natural hybridization and allopolyploid speciation in their diversification. Plant Cell Physiol. 2011, 52, 750–764. [Google Scholar] [CrossRef]
- Tadesse, W.; Sanchez-Garcia, M.; Assefa, S.G.; Amri, A.; Bishaw, Z.; Ogbonnaya, F.C.; Baum, M. Genetic gains in wheat breeding and its role in feeding the world. Crop Breed. Genet. Genom. 2019, 1, e190005. [Google Scholar] [CrossRef]
- Singh, S.; Vikram, P.; Sehgal, D.; Burgueño, J.; Sharma, A.; Singh, S.K.; Sansaloni, C.P.; Joynson, R.; Brabbs, T.; Ortiz, C.; et al. Harnessing genetic potential of wheat germplasm banks through impact-oriented-prebreeding for future food and nutritional security. Sci. Rep. 2018, 8, 12527. [Google Scholar] [CrossRef] [PubMed]
- Rane, J.; Pannu, R.K.; Sohu, V.S.; Saini, R.S.; Mishra, B.; Shoran, J.; Crossa, J.; Vargas, M.; Joshi, A.K. Performance of yield and stability of advanced wheat genotypes under heat stress environments of the Indo-Gangetic plains. Crop Sci. 2007, 47, 1561–1573. [Google Scholar] [CrossRef]
- Kumar, S.N.; Aggarwal, P.K.; Rani, D.N.S.; Saxena, R.; Chauhan, N.; Jain, S. Vulnerability of wheat production to climate change in India. Clim. Res. 2014, 59, 173–187. [Google Scholar] [CrossRef]
- Ortiz, R.; Sayre, K.D.; Govaerts, B.; Gupta, R.; Subbarao, G.V.; Ban, T.; et al. Climate change: can wheat beat the heat? Agric. Ecosyst. Environ. 2008, 126, 46–58. [Google Scholar] [CrossRef]
- Joshi, A.K.; Mishra, B.; Chatrath, R.; Ortiz -Ferrara, G.; Singh, R.P. Wheat improvement in India: present status, emerging challenges and future prospects. Euphytica 2007, 157, 431–446. [Google Scholar] [CrossRef]
- Joshi, A.K.; Ortiz-Ferrara, G.; Crossa, J.; Singh, G.; Sharma, R.C.; Chand, R.; Parsad, R. Combining superior agronomic performance and terminal heat tolerance with resistance to spot blotch (Bipolaris sorokiniana) of wheat in the warm humid Gangetic Plains of South Asia. Field Crops Res. 2007, 103, 53–61. [Google Scholar] [CrossRef]
- Farooq, M.; Bramley, H.; Palta, J.A.; Siddique, K.H. Heat stress in wheat during reproductive and grain-filling phases. Crit. Rev. Plant Sci. 2011, 30, 491–507. [Google Scholar] [CrossRef]
- Lobell, D.B.; Burke, M.B.; Tebaldi, C.; Mastrandrea, M.D.; Falcon, W.P.; Naylor, R.L. Prioritizing climate change adaptation needs for food security in 2030. Science 2008, 319, 607–610. [Google Scholar] [CrossRef]
- Mondal, S.; Singh, R.P.; Crossa, J.; Huerta-Espino, J.; Sharma, I.; Chatrath, R.; et al. Earliness in wheat: a key to adaptation under terminal and continual high temperature stress in South Asia. Field Crops Res. 2013, 151, 19–26. [Google Scholar] [CrossRef]
- Joshi, A.K.; Chand, R.; Arun, B.; Singh, R.P.; Ortiz, R. Breeding crops for reduced-tillage management in the intensive, rice–wheat systems of South Asia. Euphytica 2007, 153, 135–151. [Google Scholar] [CrossRef]
- Ullah, S.; Bramley, H.; Mahmood, T.; Trethowan, R. A strategy of ideotype development for heat-tolerant wheat. J. Agro. Crop Sci. 2019, 206, 229–241. [Google Scholar] [CrossRef]
- Reynolds, M.P.; Ortiz-Monasterio, J.I.; McNab, A. Application of Physiology in Wheat Breeding. Mexico, D.F. CIMMYT, 2001.
- Huggins, T.D.; Mohammed, S.; Sengodon, P.; Ibrahim, A.M.H.; Tilley, M.; Hays, D.B. Changes in leaf epicuticular wax load and its effect on leaf temperature and physiological traits in wheat cultivars (Triticum aestivum L.) exposed to high temperatures during anthesis. J. Agro. Crop Sci. 2017, 204, 49–61. [Google Scholar] [CrossRef]
- Reynolds, M.P.; Singh, R.P.; Ibrahim, A.; Ageeb, O.A.A.; Larque-Saavedra, A.; Quick, J.S. Evaluating physiological traits to complement empirical selection for wheat in warm environments. Euphytica 1998, 100, 85–94. [Google Scholar] [CrossRef]
- Fokar, M.; Nguyen, H.T.; Blum, A. Heat tolerance in spring wheat. I. Estimating cellular thermotolerance and its heritability. Euphytica 1998, 104, 1–8. [Google Scholar] [CrossRef]
- Zhao, H.; Dai, T.; Jing, Q.; Jiang, D.; Cao, W. Leaf senescence and grain filling affected by post-anthesis high temperatures in two different wheat cultivars. Plant Growth Regul. 2007, 51, 149–158. [Google Scholar] [CrossRef]
- Mondal, S.; Singh, R.P.; Huerta-Espino, J.; Kehel, Z.; Autrique, E. Characterization of heat- and drought-stress tolerance in high-yielding spring wheat. Crop Sci. 2015, 55, 1–11. [Google Scholar] [CrossRef]
- Telfer, P.; Edwards, J.; Bennett, D.; Ganesalingam, D.; Able, J.; Kuchel, H. A field and controlled environment evaluation of wheat (Triticum aestivum) adaptation to heat stress. Field Crops Res. 2018, 229, 55–65. [Google Scholar] [CrossRef]
- Jenner, C.F. Starch synthesis in the kernel of wheat under high temperature conditions. Aust. J. Plant Physiol. 1994, 21, 791–806. [Google Scholar] [CrossRef]
- Dias, A.S.; Bagulho, A.S.; Lidon, F.C. Ultrastructure and biochemical traits of bread and durum wheat grains under heat stress. Braz. J. Plant Physiol. 2008, 20, 323–333. [Google Scholar] [CrossRef]
- Keeling, P.L.; Bacon, P.J.; Holt, D.C. Elevated temperature reduces starch deposition in wheat endosperm by reducing the activity of soluble starch synthase. Planta 1993, 191, 342–348. [Google Scholar] [CrossRef]
- Shah, N.H.; Paulsen, G.M. Interaction of drought and high temperature on photosynthesis and grain-filling of wheat. Plant Soil 2003, 257, 219–226. [Google Scholar] [CrossRef]
- Cossani, C.M.; Reynolds, M.P. Physiological traits for improving heat tolerance in wheat. Plant physiol. 2012, 160, 1710–1718. [Google Scholar] [CrossRef]
- Elbasyoni, I.S. Performance and stability of commercial wheat cultivars under terminal heat stress. Agronomy 2018, 8, 37. [Google Scholar] [CrossRef]
- Sharma, R.C.; Tiwary, A.K.; Ortiz-Ferrara, G. Reduction in kernel weight as a potential indirect selection criterion for wheat grain yield under terminal heat stress. Plant Breed. 2008, 127, 241–248. [Google Scholar] [CrossRef]
- Fleitas, M.C.; Mondal, S.; Gerard, G.S.; Hernández-Espinosa, N.; Singh, R.P.; Crossa, J.; Guzmán, C. Identification of CIMMYT spring bread wheat germplasm maintaining superior grain yield and quality under heat-stress. J. Cereal Sci. 2020, 93, 102981. [Google Scholar] [CrossRef]
- Asseng, S.; Ewert, F.; Martre, P.; Rötter, R.P.; Lobell, D. B.; Cammarano, D.; et al. Rising temperatures reduce global wheat production. Nat. Clim. Chang. 2015, 5, 143–147. [Google Scholar] [CrossRef]
- Ray, D.K.; Gerber, J.S.; MacDonald, G.K.; West, P.C. Climate variation explains a third of global crop yield variability. Nat. Comm. 2015, 6, 1–9. [Google Scholar] [CrossRef]
- Abdelrahman, M.; Burritt, D.J.; Gupta, A.; Tsujimoto, H.; Tran, L.S.P. Heat stress effects on source–sink relationships and metabolome dynamics in wheat. J. Exp. Bot. 2020, 71, 543–554. [Google Scholar] [CrossRef]
- Kang, M.S.; Prabhakaran, V.T.; Mehra, R.B. Genotype-by-environment interaction in crop improvement. In Plant Breeding - Mendelian to Molecular Approaches, Jain H.K.; Kharkwal, M.C., Eds., Narosa Publishing House, New Delhi, India, 2004; pp. 535–572.
- Akter, N.; Islam, M.R. Heat stress effects and management in wheat - A review. Agron. Sustain. Dev. 2017, 37, 1–17. [Google Scholar] [CrossRef]
- Hyles, J.; Bloomfield, M.T.; Hunt, J.R.; Trethowan, R.M.; Trevaskis, B. Phenology and related traits for wheat adaptation. Heredity 2020, 125, 417–430. [Google Scholar] [CrossRef] [PubMed]
- Phogat, B.S.; Kumar, S.; Kumari, J.; Kumar, N.; Pandey, A.C.; Singh, T.P.; et al. Characterization of wheat germplasm conserved in the Indian National Genebank and establishment of a composite core collection. Crop Sci. 2021, 61, 604–620. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Guarino, L.; Cruz, M.; Rojas, E. Computer tools for spatial analysis of plant genetic resources data: 1. DIVA-GIS. Plant Genet. Resour. Newslett. 2001, 127, 15–19. [Google Scholar]
- Pask, A.J.D.; Pietragalla, J.; Mullan, D.M.; Reynolds, M.P. Physiological Breeding II: A Field Guide to Wheat Phenotyping. Mexico, D.F. CIMMYT, 2012.
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Reynolds, M.P.; Nagarajan, S.; Razzaque, M.A.; Ageeb, O.A.A. Using Canopy Temperature Depression to Select for Yield Potential of Wheat in Heat-Stressed Environments. Wheat Special Report. No. 42. Mexico, D.F.: CIMMYT, 1997.
- Babar, M.A.; Reynolds, M.P.; Van Ginkel, M.; Klatt, A.R.; Raun, W.R.; Stone, M.L. Spectral reflectance to estimate genetic variation for in-season biomass, leaf chlorophyll and canopy temperature in wheat. Crop Sci. 2006, 46, 1046–1057. [Google Scholar] [CrossRef]
- Sairam, R.K.; Deshmukh, P.S.; Shukla, D.S. Tolerance of drought and temperature stress in relation to increased antioxidant enzyme activity in wheat. J. Agron. Crop Sci. 1997, 178, 171–178. [Google Scholar] [CrossRef]
- Aldesuquy, H.; Baka, Z.; Mickky, B. Kinetin and spermine mediated induction of salt tolerance in wheat plants: Leaf area, photosynthesis and chloroplast ultrastructure of flag leaf at ear emergence. Egypt. J. Basic Appl. Sci. 2014, 1, 77–87. [Google Scholar] [CrossRef]
- SAS Institute. Statistical analysis system for windows version 9.4, SAS Institute Inc., Cary, NC, USA, 2013.
- IBM SPSS. IBM SPSS (Statistical Package for the Social Sciences) Statistics Software for Windows, Version 20.0. Armonk, NY, IBM Corp. 2011. Available at: https://hadoop.apache.org.
- Fischer, R.A.; Maurer, R. Drought resistance in spring wheat cultivars. I. Grain yield responses. Aust. J. Agr. Res. 1978, 29, 897–912. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The mathematical theory of communication. University of Illinois Press, Urbana, 1949.
- Eberhart, S.A.; Russell, W.A. Stability parameters for comparing varieties. Crop Sci. 1966, 6, 36–40. [Google Scholar] [CrossRef]
- Mondal, S.; Rutkoski, J.E.; Velu, G.; Singh, P.K.; Crespo-Herrera, L.A.; Guzman, C.; Bhavani, S.; Lan, C.; He, X.; and Singh, R.P. Harnessing diversity in wheat to enhance grain yield, climate resilience, disease and insect pest resistance and nutrition through conventional and modern breeding approaches. Front. Plant Sci. 2016, 7, 991. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, S.K.; Basu, S.; Kumar, S.; Kumar, G.; Prakash, V.; Kumar, S.; et al. Heat stress induced impairment of starch mobilisation regulates pollen viability and grain yield in wheat: Study in eastern Indo-Gangetic Plains. Field Crops Res. 2017, 206, 106–114. [Google Scholar] [CrossRef]
- Bita, C.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 273. [Google Scholar] [CrossRef] [PubMed]
- Gowda, D.S.S.; Singh, G.P.; Singh, A.M. Relationship between canopy temperature depression, membrane stability, relative water content and grain yield in bread wheat (Triticum aestivum) under heat-stress environments. Indian J. Agric. Sci. 2011, 81, 197–202. [Google Scholar]
- Pinto, R.S.; Molero, G.; Reynolds, M.P. Identification of heat tolerant wheat lines showing genetic variation in leaf respiration and other physiological traits. Euphytica 2017, 213, 76. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Lepekhov, S.B. Canopy temperature depression for drought- and heat stress tolerance in wheat breeding. Vavilov J. Genet. Breed. 2022, 26, 196–201. [Google Scholar] [CrossRef]
- Kaur, V.; Behl, R.K. Grain yield in wheat as affected by short periods of high temperature, drought and their interaction during pre-and post-anthesis stages. Cereal Res. Commun. 2010, 38, 514–520. [Google Scholar] [CrossRef]
- Tomás, D.; Coelho, L.P.; Rodrigues, J.C.; Viegas, W.; Silva, M. Assessment of four Portuguese wheat landrace diversity to cope with global warming. Front. Plant Sci. 2020, 11, 594977. [Google Scholar] [CrossRef]
- Agarwal, V.P.; Gupta, N.K.; Gupta, S.; Singh, G. Screening of wheat germplasm for terminal heat tolerance under hyper-arid conditions. Cereal Res. Commun. 2021, 49, 375–383. [Google Scholar] [CrossRef]
- Shepherd, T.; Griffiths, D.W. The effects of stress on plant cuticular waxes. New Phytol. 2006, 171, 469–499. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, S.; Huggins, T.D.; Beecher, F.; Chick, C.; Sengodon, P.; Mondal, S.; et al. The role of leaf epicuticular wax in the adaptation of wheat (Triticum aestivum L.) to high temperatures and moisture deficit conditions. Crop Sci. 2018, 58, 679–689. [Google Scholar] [CrossRef]
- Soni, A.; Munjal, R. Characterisation and evaluation of wheat genetic resources for heat stress tolerance using stay-green traits. Crop Pasture Sci. 2023. [Google Scholar] [CrossRef]
- Mondal, S.; Dutta, S.; Crespo-Herrera, L.; Huerta-Espino, J.; Braun, H.J.; Singh, R.P. Fifty years of semi-dwarf spring wheat breeding at CIMMYT: Grain yield progress in optimum, drought and heat stress environments. Field Crops Res. 2020, 250, 107757. [Google Scholar] [CrossRef]
- Al-Ashkar, I.; Alotaibi, M.; Refay, Y.; Ghazy, A.; Zakri, A.; Al-Doss, A. Selection criteria for high-yielding and early-flowering bread wheat hybrids under heat stress. PLoS ONE 2020, 15, e0236351. [Google Scholar] [CrossRef]
- Reynolds, M.P.; Pierre, C.S.; Saad, A.S.I.; Vargas, M.; Condon, A.G. Evaluating potential genetic gains in wheat associated with stress-adaptive trait expression in elite genetic resources under drought and heat stress. Crop Sci. 2007, 47, S172–S189. [Google Scholar] [CrossRef]
- Fu, J.; Bowden, R.L.; Jagadish, S.V.K.; Prasad, P.V.V. Genetic variation for terminal heat stress tolerance in winter wheat. Front. Plant Sci. 2023, 14, 1132108. [Google Scholar] [CrossRef]
- Ayeneh, A.; van Ginkel, M.; Reynolds, M.P.; Ammar, K. Comparison of leaf, spike, peduncle and canopy temperature depression in wheat under heat stress. Field Crops Res. 2002, 79, 173–184. [Google Scholar] [CrossRef]
- Bahar, B.; Yildirim, M.; Yucel, C. Heat and drought resistance criteria in spring bread wheat (Triticum aestivum L.): morpho- physiological parameters for heat tolerance. Sci. Res. Essays. 2011, 6, 2212–2220. [Google Scholar] [CrossRef]
- Lordkaew, S.; Yimyam, N.; Wongtamee, A.; Jamjod, S.; Rerkasem, B. Evaluating a heat-tolerant wheat germplasm in a heat stress environment. Plant Genet. Resour. 2019, 17, 339–345. [Google Scholar] [CrossRef]
- Chaubey, R.K.; Bhutia, D.D.; Navathe, S.; Mishra, V.K.; Singh, A.K.; Chand, R. Interrelationships among different grain characteristics of wheat grown under optimum and late sowing date conditions in the Eastern Indo-Gangetic plains of India. Cereal Res. Commun. 2021, 49, 449–455. [Google Scholar] [CrossRef]
- Chiotelli, E.; Le Meste, M. Effect of small and large wheat starch granules on thermomechanical behavior of starch. Cereal Chem. 2002, 79, 286–293. [Google Scholar] [CrossRef]
- Uthayakumaran, S.; Wrigley, C. Wheat: Grain-quality Characteristics and Management of Quality Requirements. In Cereal Grains-Assessing and Managing Quality, Batey, C.W.I., Miskelly, D.,Eds.; Cambridge, England, Woodhead Publishing, 2017; pp. 91–134. [CrossRef]
- Liu, P.; Guo, W.; Jiang, Z.; Pu, H.; Feng, C.; Zhu, X.; Peng, Y.; Kuang, A.; Little, C.R. Effects of high temperature after anthesis on starch granules in grains of wheat (Triticum aestivum L.). J. Agric. Sci. 2011, 149, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, A.; Sita, K.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; et al. Drought or/and heat-stress effects on seed filling in food crops: Impacts on functional biochemistry, seed yields, and nutritional quality. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- Braun, H. J.; Atlin, G.; Payne, T. Multi-location testing as a tool to identify plant response to global climate change. In Climate Change and Crop Production, Reynolds, M.P., Ed., CABI, USA, 2010; pp. 115–138.
- Powell, N.; Ji, X.; Ravash, R.; Edlinton, J.; Dolferus, R. Yield stability for cereals in a changing climate. Funct. Plant Biol. 2012, 39, 539–552. [Google Scholar] [CrossRef]
- Gupta, V.; Mehta, G.; Kumar, S.; Ramdas, S.; Tiwari, R.; Singh, G.P.; Sharma, P. AMMI and GGE biplot analysis of yield under terminal heat tolerance in wheat. Mol. Biol. Rep. 2023, 50, 3459–3467. [Google Scholar] [CrossRef]
Figure 1.
Temperature and rainfall variation during grain filling period (GFP; from anthesis to physiological maturity) in both non-stressed (NS) and heat-stressed (HS) environments in crop seasons of 2018-19 and 2019-20. RF-rainfall, Tmax-maximum temp., Tmin - minimum temp.
Figure 1.
Temperature and rainfall variation during grain filling period (GFP; from anthesis to physiological maturity) in both non-stressed (NS) and heat-stressed (HS) environments in crop seasons of 2018-19 and 2019-20. RF-rainfall, Tmax-maximum temp., Tmin - minimum temp.
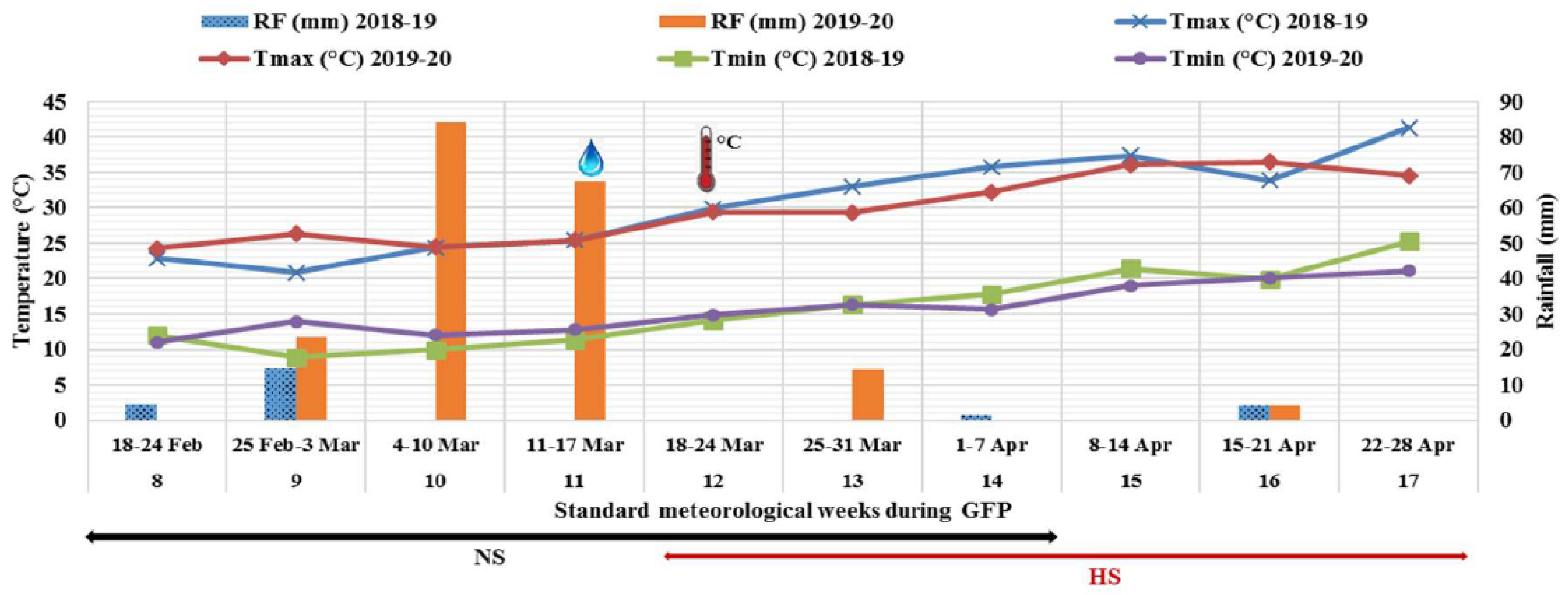
Figure 2.
Adaptation to terminal heat-stress by highly tolerant accessions of bread wheat germplasm during grain filling period. Comparisons of accession IC529207 (highly tolerant) with highest plant waxiness (score 10) (a) vs. accession IC252431 (highly susceptible) showing the least plant waxiness (score 1) (b), and accession IC416019 (highly tolerant) showing the highest leaf rolling (score 10) (c) vs. IC553599 (highly susceptible) showing the least leaf rolling (score 4) (d).
Figure 2.
Adaptation to terminal heat-stress by highly tolerant accessions of bread wheat germplasm during grain filling period. Comparisons of accession IC529207 (highly tolerant) with highest plant waxiness (score 10) (a) vs. accession IC252431 (highly susceptible) showing the least plant waxiness (score 1) (b), and accession IC416019 (highly tolerant) showing the highest leaf rolling (score 10) (c) vs. IC553599 (highly susceptible) showing the least leaf rolling (score 4) (d).

Figure 3.
UPGMA dendrogram constructed with Ward's minimum variance method for 96 bread wheat accessions using data of 18 morpho-physiological and yield-related traits recorded under heat-stress environment. Six clusters are marked on the left side of the dendrogram.
Figure 3.
UPGMA dendrogram constructed with Ward's minimum variance method for 96 bread wheat accessions using data of 18 morpho-physiological and yield-related traits recorded under heat-stress environment. Six clusters are marked on the left side of the dendrogram.
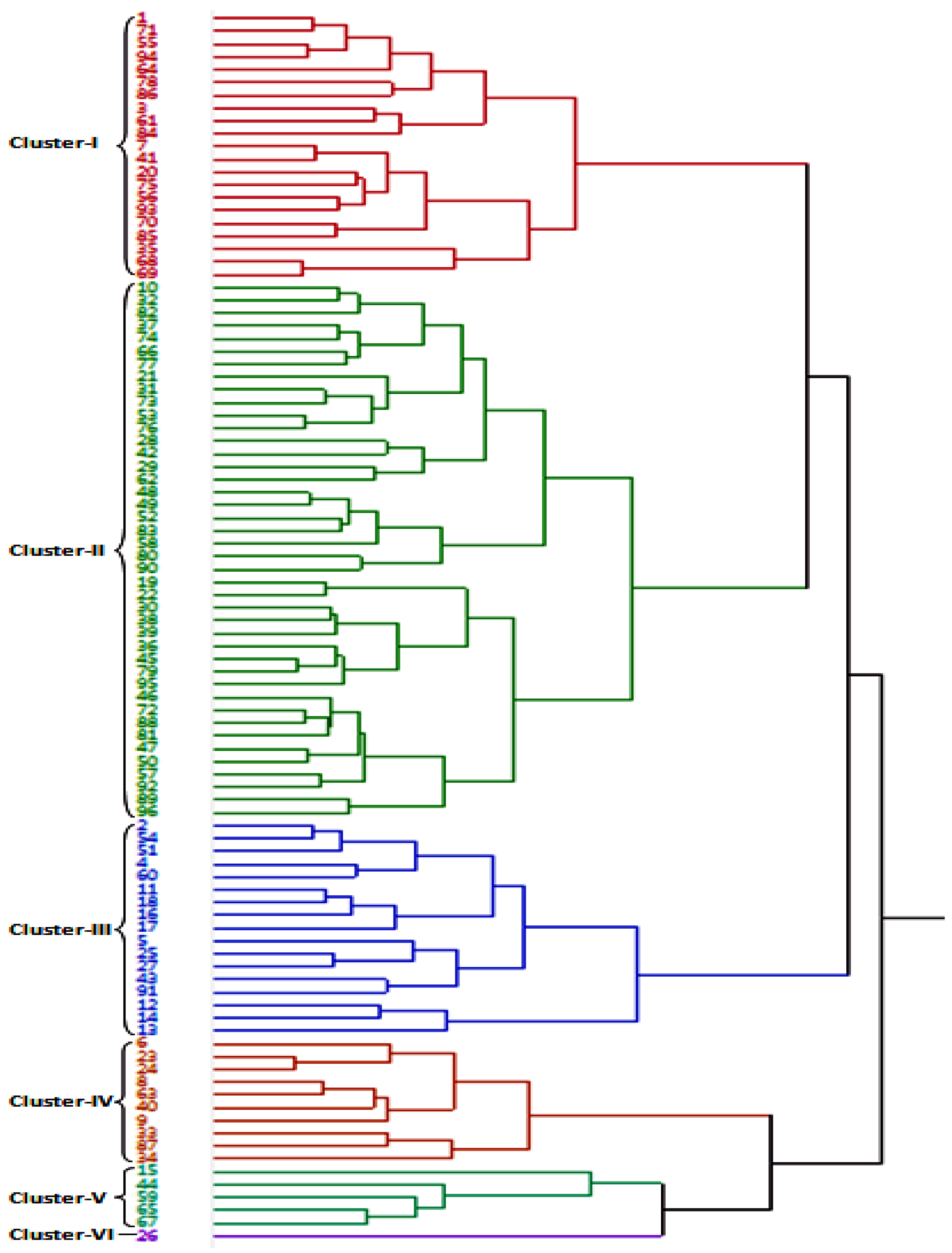
Figure 4.
Pearson's correlations coefficients derived between 18 morpho-physiological and yield traits under non-stressed (a) and heat-stressed (b) environments in 96 bread wheat accessions.
Figure 4.
Pearson's correlations coefficients derived between 18 morpho-physiological and yield traits under non-stressed (a) and heat-stressed (b) environments in 96 bread wheat accessions.
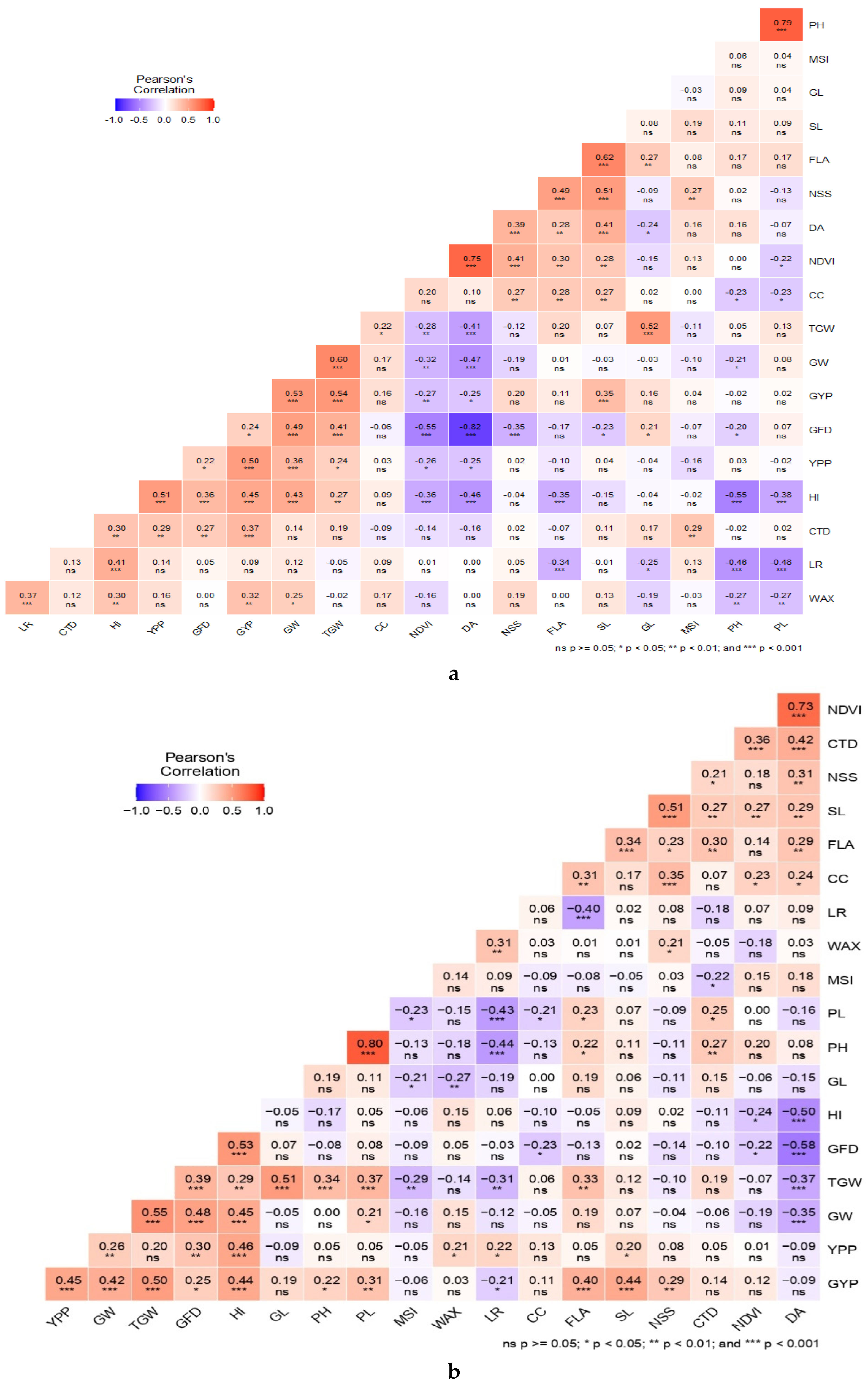
Figure 5.
Boxplot distribution of variability assessed for 18 morpho-physiological and yield contributing traits under non-stressed (NS) and heat-stressed (HS) environments in 96 bread wheat accessions categorized based on HSI values as tolerant (HSI <1.0).
Figure 5.
Boxplot distribution of variability assessed for 18 morpho-physiological and yield contributing traits under non-stressed (NS) and heat-stressed (HS) environments in 96 bread wheat accessions categorized based on HSI values as tolerant (HSI <1.0).
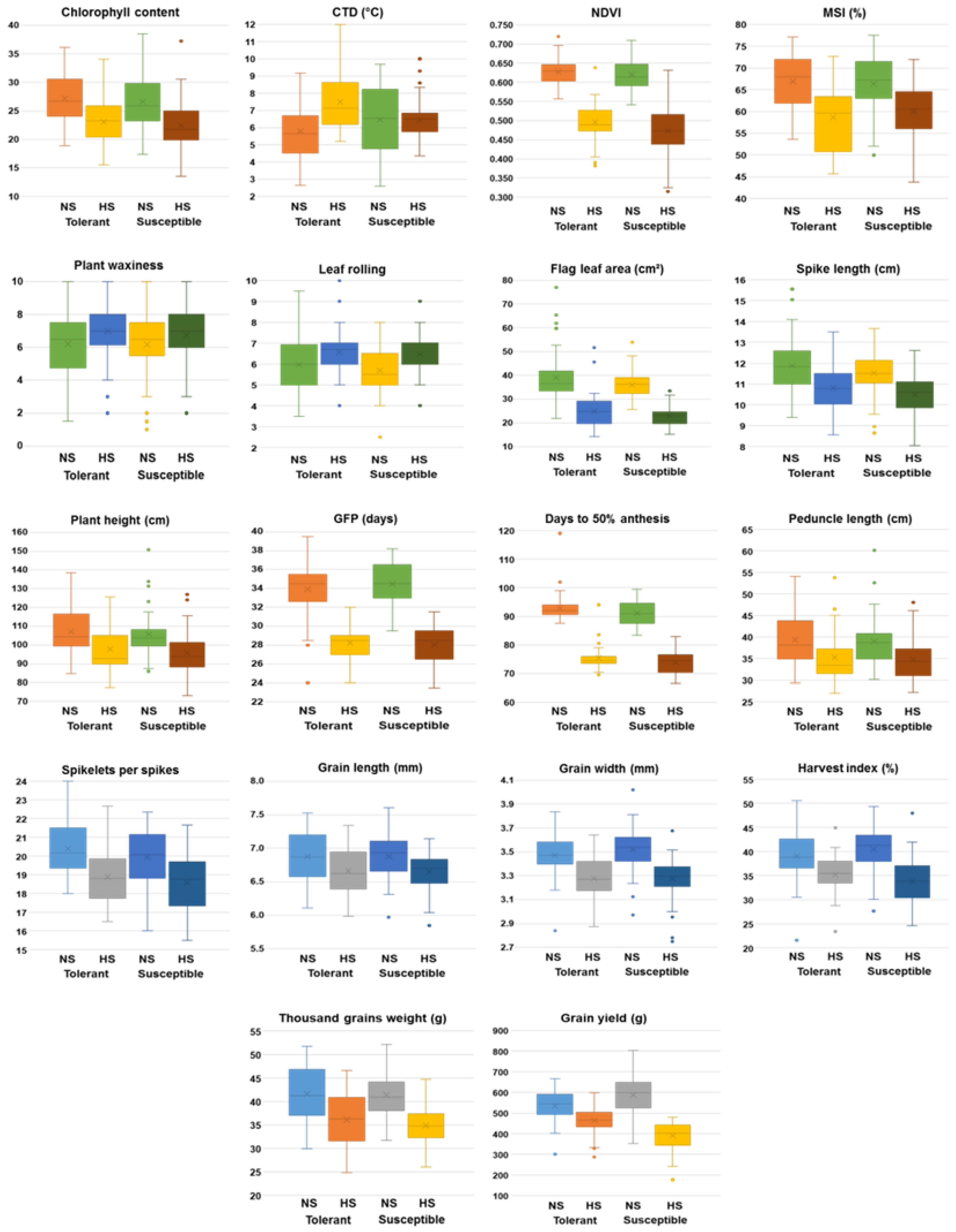
Figure 6.
Effects of heat-stress on important morpho-physiological and yield-traits on highly tolerant, tolerant, susceptible and highly susceptible accessions with national checks. Reduction of CC and NDVI was more evident in highly susceptible and susceptible accessions compared to highly tolerant and tolerant accessions under HS (a-b). MSI was more in highly susceptible and susceptible accessions than the tolerant accessions in HS environment (c). CTD, plant waxiness and leaf rolling increased in the tolerant accessions as compared to the susceptible ones under HS environment (d-f). Under heat-stress, days to 50% anthesis, plant height and peduncle length were reduced in both the tolerant and susceptible accessions (g-i). GFP, flag-leaf area and spike length all decreased in both tolerant and susceptible accessions under heat-stress (j-l). TGW, HI and grain yield were higher in the tolerant accessions than the susceptible ones in HS environment (m-o).
Figure 6.
Effects of heat-stress on important morpho-physiological and yield-traits on highly tolerant, tolerant, susceptible and highly susceptible accessions with national checks. Reduction of CC and NDVI was more evident in highly susceptible and susceptible accessions compared to highly tolerant and tolerant accessions under HS (a-b). MSI was more in highly susceptible and susceptible accessions than the tolerant accessions in HS environment (c). CTD, plant waxiness and leaf rolling increased in the tolerant accessions as compared to the susceptible ones under HS environment (d-f). Under heat-stress, days to 50% anthesis, plant height and peduncle length were reduced in both the tolerant and susceptible accessions (g-i). GFP, flag-leaf area and spike length all decreased in both tolerant and susceptible accessions under heat-stress (j-l). TGW, HI and grain yield were higher in the tolerant accessions than the susceptible ones in HS environment (m-o).
Figure 7.
Impact of heat-stress on grain morphology and ultrastructure as visualized by scanning electron microscopy (SEM) in tolerant and susceptible accessions of bread wheat. Reduction of grain width due to heat-stress was more evident in susceptible accession IC573461 as compared to three tolerant accessions IC535176, IC443661 and IC539221(a). Ultrastructural changes caused by heat-stress in endosperm and aleurone layer of grains in very late sown bread wheat accessions (b). SEM revealed ultrastructure of matured wheat grains showing aleurone layer and endosperm (b, upper panel; low magnification) and packing of starch granules (structure and density as seen in close-up view) in the endosperm (b, Lower panel; high magnification) in tolerant (IC443661) and susceptible (IC573461) accessions in non-stressed (NS) and heat-stressed (HS) environments.
Figure 7.
Impact of heat-stress on grain morphology and ultrastructure as visualized by scanning electron microscopy (SEM) in tolerant and susceptible accessions of bread wheat. Reduction of grain width due to heat-stress was more evident in susceptible accession IC573461 as compared to three tolerant accessions IC535176, IC443661 and IC539221(a). Ultrastructural changes caused by heat-stress in endosperm and aleurone layer of grains in very late sown bread wheat accessions (b). SEM revealed ultrastructure of matured wheat grains showing aleurone layer and endosperm (b, upper panel; low magnification) and packing of starch granules (structure and density as seen in close-up view) in the endosperm (b, Lower panel; high magnification) in tolerant (IC443661) and susceptible (IC573461) accessions in non-stressed (NS) and heat-stressed (HS) environments.
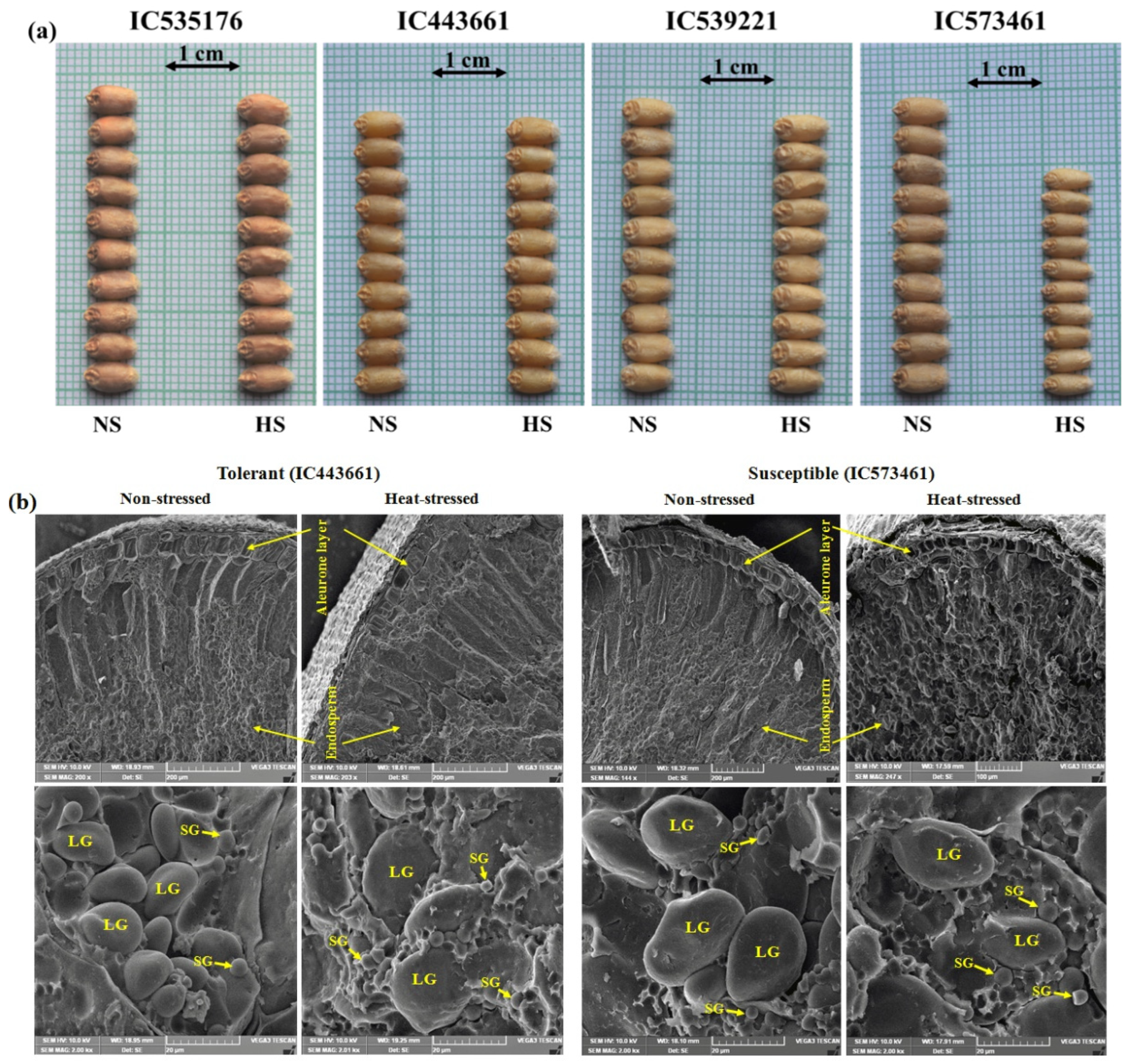
Figure 8.
Stability parameters based on Eberhart and Russell model. Frequency distribution of accessions for grain yield per plot and thousand grain weight (a-b). Mean performance of accessions and their regression value (βi) for grain yield and thousand grain weight (c-d). Stability of accessions based on deviation from linearity (S2Di) for grain yield and TGW (e-f).
Figure 8.
Stability parameters based on Eberhart and Russell model. Frequency distribution of accessions for grain yield per plot and thousand grain weight (a-b). Mean performance of accessions and their regression value (βi) for grain yield and thousand grain weight (c-d). Stability of accessions based on deviation from linearity (S2Di) for grain yield and TGW (e-f).
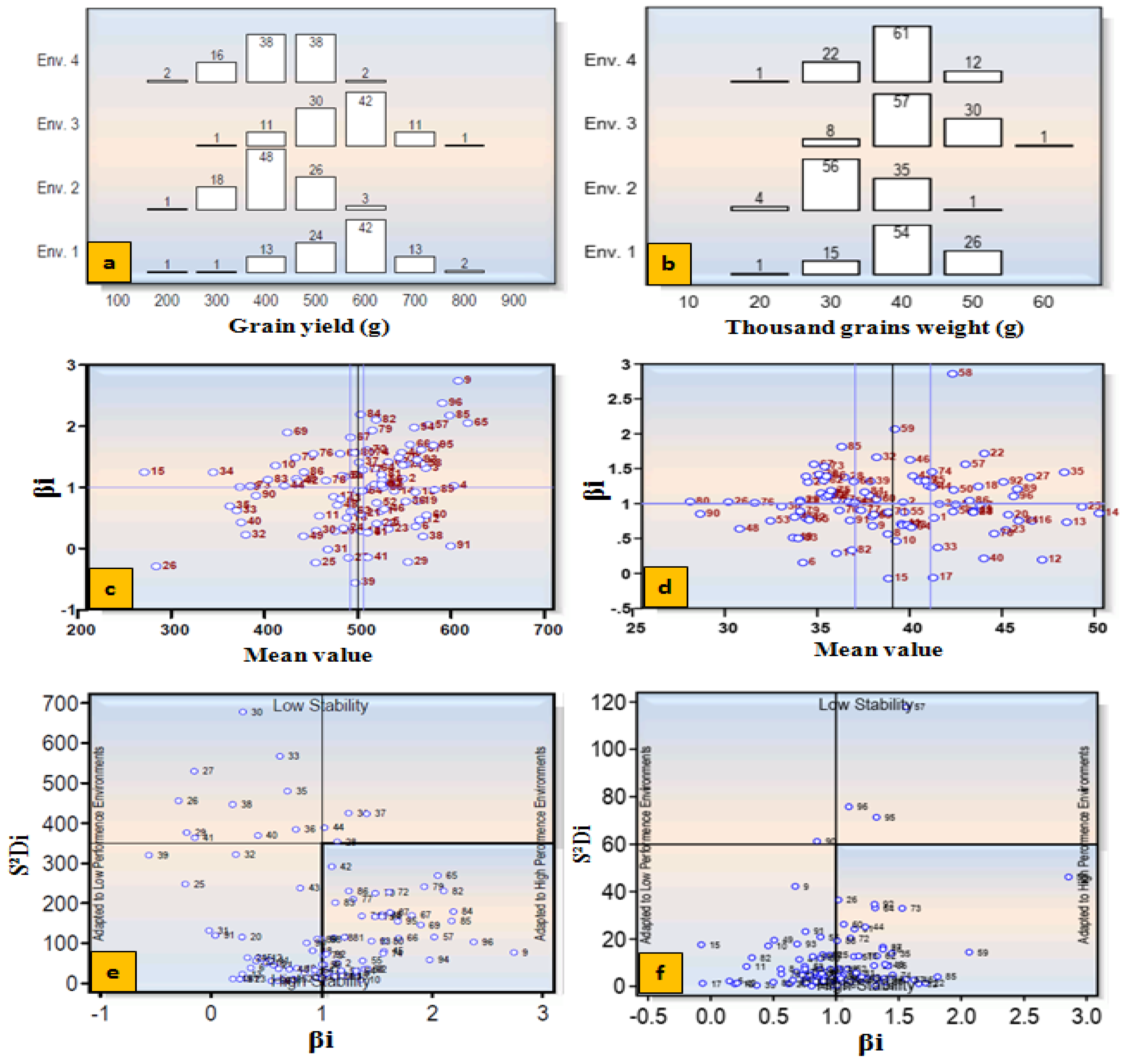

Table 1.
Morpho-physiological and yield-related traits studied in 96 bread wheat accessions under non-stressed and heat-stressed environments during two crop seasons of 2018-19 and 2019-20.
Table 1.
Morpho-physiological and yield-related traits studied in 96 bread wheat accessions under non-stressed and heat-stressed environments during two crop seasons of 2018-19 and 2019-20.
| Sl.No. | Traits studied | Code | How was the trait measured? |
|---|---|---|---|
| 1. | Chlorophyll Content |
CC | Estimated on five flag leaves of main tillers in each accession by non-destructive method using hand-held Chlorophyll Content Meter (Model-CCM-200 plus, Opti- Science, USA) and expressed as chlorophyll content index (CCI) [52]. |
| 2. | Canopy Temp. Depression (°C) | CTD | Measured on warm, sunny and cloudless day using portable Infrared Thermometer (Fisher Scientific, England) [54]. |
| 3. | Normalized Difference Vegetation Index | NDVI | NDVI was recorded using hand-held crop sensor (Green Seeker®, Trimble, USA) covering entire plot. The value for crop canopy ranged from 0 to 1; where 0 represents no green area and 1 represent maximum greenness [55]. |
| 4. | Membrane Stability Index (%) | MSI | MSI estimated as per procedure of Sairam et al. [56]. Leaf samples (0.1 g) from each plot were taken and cut into uniform small discs. MSI was calculated using formula: MSI = [1 - (C1/C2)] × 100, where C1 and C2 represent reading of EC recorded using digital conductivity meter at 45 ºC and 100 ºC, respectively. |
| 5. | Plant Waxiness (0-10 scale) | PW | Plant waxiness measured by visual observations of whole plot during mid of GFP and scored using the scale from 0 (0%) to 10 (100%) in increment of 10 %. |
| 6. | Leaf Rolling (0-10 scale) | LR | Leaf rolling measured at mid grain filling period by visual observation of whole plot and rated the proportion of the leaves showing rolling effect using a scale from 0 (0 %) to 10 (100 %) in increments of 10% [52]. |
| 7. | Days to 50 % Anthesis | DA | Recorded as the period between date of sowing and the date at which 50% of spikes start to extrude their anthers [53]. |
| 8. | Grain Filling Period (days) | GFP | GFP calculated as the difference between days to 50% anthesis and days to physiological maturity. |
| 9. | Plant Height (cm) | PH | Plant height was measured from base of the plant to top of the spike excluding awns of the main tiller at maturity. |
| 10. | Peduncle Length (cm) | PL | Measured from uppermost node to the spike collar of the main tiller at maturity in three plants per accession. |
| 11. | Flag Leaf Area (cm2) | FLA | Derived on five randomly selected plant's flag leaves using following equation [57]: Leaf area = Length × Breadth × 0.75. |
| 12. | Spike Length (cm) | SL | Measured from the spike collar to tip of spike excluding awns of the main tiller in three plants per accession. |
| 13. | Number of Spikelets per Spike | NSS | Spikelets per spike were counted on the main tiller spike of three plants per accession. |
| 14. | Grain Length (mm) | GL | Measured on five grains per accession by Digimatic Caliper (Model-CD-6''ASX, Mitutoyo Corporation, Japan). |
| 15. | Grain Width (mm) | GW | Grain width was measured from randomly selected five grains per accession by Digimatic Caliper. |
| 16. | 1000-Grain Weight (g) | TGW | TGW recorded on randomly selected 1000 grains from plot yield and weighted using sensitive electronic balance (d = 0.1 mg, Sartorius, model CPA64, Germany). |
| 17. | Harvest Index (%) | HI | HI calculated using the following formula: HI = (Grain yield per plant/ Biological yield per plant) × 100. |
| 18. | Grain Yield (g/m2) | GY | Plot yield of individual accession harvested at maturity and threshed manually. Weight of grains recorded using electronic balance and expressed as grain yield per unit area. |
Table 2.
Descriptive statistics of pooled data of 18 morpho-physiological and yield traits recorded in 96 accessions of bread wheat in NS and HS environments during two crop seasons of 2018-2020.
Table 2.
Descriptive statistics of pooled data of 18 morpho-physiological and yield traits recorded in 96 accessions of bread wheat in NS and HS environments during two crop seasons of 2018-2020.
| Trait | Environment | Range | Mean ± S.E. | SD | CV (%) |
PCV (%) |
GCV (%) | H (%) |
GA (%) |
|
|---|---|---|---|---|---|---|---|---|---|---|
| Min. | Maxi. | |||||||||
| CCI | NS | 17.4 | 38.5 | 26.9±0.43 | 4.21 | 15.69 | 18.1 | 15.0 | 68.3 | 25.5 |
| HS | 13.5 | 37.2 | 22.7±0.49 | 4.10 | 18.10 | 18.6 | 16.2 | 75.9 | 29.0 | |
| CTD (°C) | NS | 2.6 | 9.7 | 6.2±0.18 | 1.80 | 29.19 | 23.3 | 10.5 | 20.3 | 9.7 |
| HS | 4.4 | 12.0 | 6.9±0.15 | 1.47 | 21.24 | 26.1 | 11.0 | 17.8 | 9.6 | |
| NDVI (0 - 1) | NS | 0.54 | 0.72 | 0.62±0.01 | 0.04 | 6.03 | 7.5 | 5.1 | 42.9 | 6.6 |
| HS | 0.32 | 0.64 | 0.48±0.01 | 0.06 | 12.61 | 10.9 | 8.5 | 63.8 | 14.3 | |
| MSI (%) | NS | 50.0 | 77.5 | 66.6±0.70 | 6.89 | 10.35 | 10.5 | 6.0 | 33.0 | 7.2 |
| HS | 43.7 | 72.6 | 59.4±0.68 | 6.70 | 11.29 | 8.5 | 2.5 | 8.7 | 1.5 | |
| PW (0 - 10) | NS | 1.0 | 10.0 | 6.2±0.20 | 1.94 | 31.35 | 23.8 | 19.6 | 67.6 | 33.2 |
| HS | 2.0 | 10.0 | 6.9±0.17 | 1.68 | 24.47 | 21.4 | 17.2 | 64.6 | 28.5 | |
| LR (0 - 10) | NS | 2.5 | 9.5 | 5.8±0.13 | 1.28 | 22.04 | 20.8 | 17.5 | 71.4 | 30.5 |
| HS | 4.0 | 10.0 | 6.5±0.13 | 1.26 | 19.29 | 19.1 | 15.0 | 61.4 | 24.1 | |
| Days to 50% anthesis | NS | 83.4 | 119.0 | 91.9±0.48 | 4.72 | 5.13 | 6.1 | 5.6 | 84.9 | 10.6 |
| HS | 66.6 | 94.0 | 74.6±0.40 | 3.88 | 5.20 | 6.8 | 6.3 | 85.4 | 11.9 | |
| GFP (days) | NS | 24.0 | 39.5 | 34.2±0.25 | 2.45 | 7.16 | 19.9 | 10.7 | 29.0 | 11.9 |
| HS | 23.5 | 32.0 | 28.1±0.20 | 1.94 | 6.91 | 8.7 | 5.7 | 43.0 | 7.7 | |
| Plant height (cm) | NS | 84.9 | 150.9 | 106.6±1.31 | 12.83 | 12.03 | 5.9 | 2.5 | 17.8 | 2.2 |
| HS | 73.2 | 127.0 | 96.9±1.25 | 12.23 | 12.63 | 7.1 | 4.4 | 39.1 | 5.7 | |
| Peduncle length (cm) | NS | 29.4 | 60.1 | 39.2±0.61 | 6.00 | 15.33 | 14.7 | 12.1 | 67.7 | 20.6 |
| HS | 27.0 | 53.8 | 35.1±0.54 | 5.27 | 15.04 | 14.5 | 12.9 | 79.4 | 23.7 | |
| Flag leaf area (cm²) | NS | 21.9 | 77.0 | 37.4±0.88 | 8.61 | 23.01 | 20.4 | 16.1 | 62.4 | 26.2 |
| HS | 14.2 | 51.7 | 23.8±0.61 | 5.95 | 24.96 | 17.8 | 14.5 | 66.3 | 24.3 | |
| Spike length (cm) | NS | 8.7 | 15.6 | 11.7±0.12 | 1.18 | 10.14 | 6.8 | 3.0 | 19.4 | 2.7 |
| HS | 8.1 | 13.5 | 10.6±0.11 | 1.10 | 10.38 | 4.8 | 1.3 | 7.1 | 0.7 | |
| Spikelets per spike | NS | 16.0 | 24.0 | 20.2±0.15 | 1.46 | 7.23 | 5.9 | 2.1 | 12.7 | 1.6 |
| HS | 15.5 | 22.7 | 18.7±0.15 | 1.49 | 7.98 | 5.9 | 2.4 | 16.3 | 2.0 | |
| Grain length (mm) | NS | 5.97 | 8.76 | 6.90±0.04 | 0.39 | 5.59 | 5.6 | 4.9 | 77.9 | 9.0 |
| HS | 5.85 | 8.62 | 6.68±0.04 | 0.38 | 5.63 | 6.1 | 5.8 | 89.3 | 11.3 | |
| Grain width (mm) | NS | 2.84 | 4.02 | 3.49±0.02 | 0.19 | 5.31 | 3.5 | 2.0 | 33.0 | 2.4 |
| HS | 2.64 | 3.68 | 3.27±0.02 | 0.18 | 5.52 | 3.3 | 1.4 | 17.9 | 1.2 | |
| 1000-grain weight (g) | NS | 30.0 | 52.2 | 41.5±0.53 | 5.19 | 12.51 | 6.0 | 3.0 | 24.1 | 3.0 |
| HS | 24.9 | 46.6 | 35.5±0.48 | 4.73 | 13.34 | 8.0 | 3.3 | 16.7 | 2.8 | |
| Harvest index (%) | NS | 21.6 | 50.6 | 39.7±0.49 | 4.84 | 12.19 | 9.4 | 1.7 | 13.3 | 0.6 |
| HS | 23.4 | 48.0 | 34.4±0.45 | 4.38 | 12.76 | 9.3 | 3.2 | 11.9 | 2.3 | |
| Grain yield (g/m2) | NS | 300.0 | 802.5 | 562.2±8.77 | 86.00 | 15.30 | 11.3 | 7.3 | 42.1 | 9.8 |
| HS | 176.7 | 598.1 | 423.6±7.34 | 71.95 | 16.97 | 10.5 | 5.8 | 30.2 | 6.6 | |
Table 3.
ANOVA for TGW and grain yield over four environments for stability analysis using Eberhart and Russell model.
Table 3.
ANOVA for TGW and grain yield over four environments for stability analysis using Eberhart and Russell model.
| Sources of variation | D.F. | TGW (MSS) | Grain Yield (MSS) |
|---|---|---|---|
| Genotypes | 95 | 89.54** | 19206.70** |
| Environment + (G × E) | 288 | 28.00** | 9164.46** |
| Environment | 3 | 1589.07** | 614125.02** |
| Environment G × E | 285 | 11.57 | 2796.45** |
| Environment (Linear) | 1 | 4767.22** | 1842375.06** |
| G × E (Linear) | 95 | 9.53 | 18197.54** |
| Pooled deviation | 192 | 12.46 | 148.88 |
| Total | 383 | 43.27 | 11655.34 |
** significant at 0.01 level of probability; TGW: 1000-grain weight, MSS: Mean sum of squares.
Table 4.
Grouping of different accessions based on Eberhart and Russell stability model, their mean performance under NS and HS environments along with heat susceptibility index. .
Table 4.
Grouping of different accessions based on Eberhart and Russell stability model, their mean performance under NS and HS environments along with heat susceptibility index. .
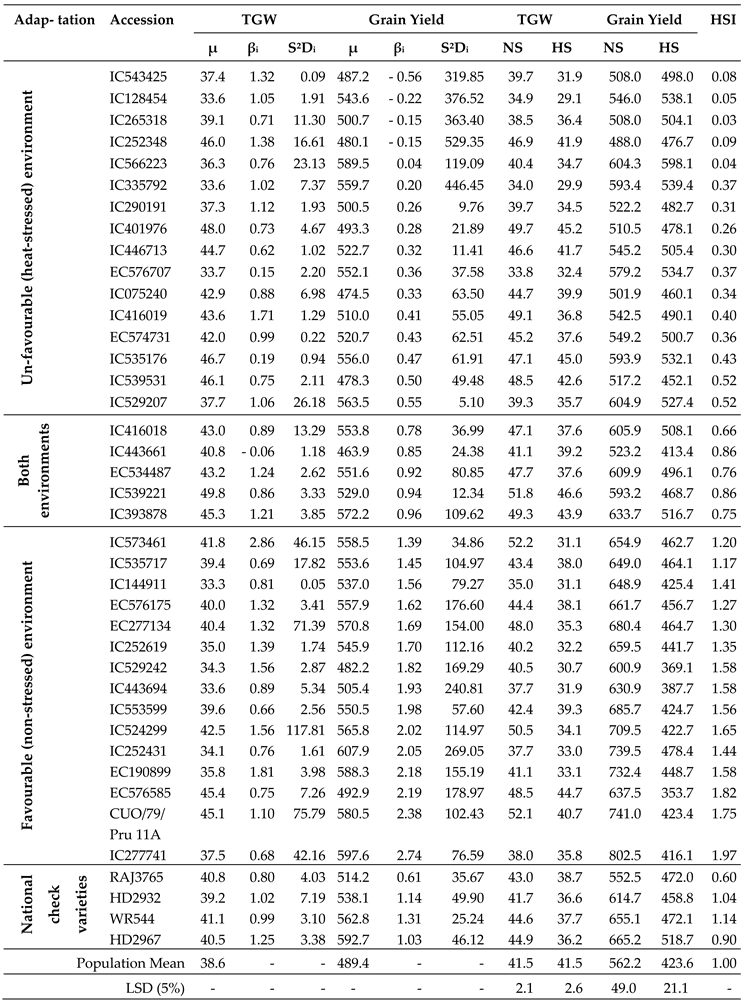 |
Abbreviations: TGW: 1000-grain weight, HSI: Heat susceptibility index, NS: Non-stressed environment, HS: Heat-stressed environment, μ: Mean, βi: Regression coefficient, S2di: Deviations from the regression.
Table 5.
Identification and selection of parents for the development of mapping populations for heat-stress tolerance in bread wheat.
Table 5.
Identification and selection of parents for the development of mapping populations for heat-stress tolerance in bread wheat.
| Sl. No. | Traits for mapping population | Parents with desirable traits for heat-stress tolerance | |
|---|---|---|---|
| a). | Bi-parental population | Parent(Higher value) | Parent(Lower value) |
| 1. | Plant waxiness | IC529207, IC528965 | IC252431, IC252444 |
| 2. | Leaf rolling | IC416019, IC416055 | IC553599, IC252816 |
| 3. | Earliness | IC296383 | IC542509 |
| 4. | Grain filling period | IC252725 | EC577013 |
| 5. | Grain width | IC401976 | IC112258 |
| 6. | 1000-grain weight | IC539221 | IC542544 |
| 7. | Harvest index | IC443653 | IC542509 |
| 8. | Grain yield | IC566223 | EC577013 |
| b) | MAGIC Population | Parents with desirable traits | |
| 1. | 4-parent MAGIC | IC566223, IC529207, IC416019, IC296383 | |
| 2. | 8-parent MAGIC |
IC128454, IC519900, IC528965, IC416055, IC539221, IC401976, IC535176, IC566223 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



