
Submitted:
30 May 2023
Posted:
01 June 2023
You are already at the latest version
Abstract
The melanoma differentiation-associated protein 5 (MDA5; encoded by the IFIH1 gene) mediates the activation of the interferon pathway in response to viral infection. This protein is also upregulated in autoimmune diseases and psoriasis skin lesions. IFIH1 gene variants that increased the MDA5 activity have been associated with increased risk for immune mediated diseases, including psoriasis. Our aim was to determine the association between three IFIH1 variants (rs35337543, intron8 +1G>C; rs35744605, Glu627Stop; and rs1990760, Ala946Thr) and the main clinical findings in a cohort of Spanish patients with psoriatic disease (N=572; 77% early-onset). Early-onset psoriasis (EOPs) had a significant higher frequency of severe disease and Cw6+. Carriers of the 946Thr variant were more common in EOPs (p<0.001), and the effect was more pronounced among Cw6-negatives. This variant was also associated with an increased risk of psoriatic arthritis (PsA) independently of other factors (OR=1.62, 95%CI=1.11-2.37). The rs3533754 and rs35744605 have been reported as risk factors for viral infection and protective for autoimmune diseases, but we did not find significant differences between the two onset age or PsA groups. However, due to the reduced frequency of the two variants (<0.02) the size of our cohort was too low to conclude a significant effect.
In conclusion, the common IFIH1 rs1990760 T allele that has been linked to increased gene expression was significantly more frequent in EOPs patients. This variant was also an independent risk factor for PsA in our cohort. This risk allele was in linkage disequilibrium with other variants previously associated with the risk of psoriasis and PsA. Our study reinforces the widely reported role of IFIH1 gene variants on psoriatic disease and other immune mediated diseases.
Keywords:
psoriasis
; psoriatic arthritis
; IFIH1
; MDA5
; polymorphism
; genetic association
1. Introduction
Psoriasis is a chronic inflammatory skin disease in which acquired and inherited risk factors contribute to define the individual’s risk [1]. Both rare and common gene variants have been associated with the risk of developing psoriasis as well with the onset age, disease severity, or the clinical phenotype, including the risk for psoriatic arthritis (PsA) [2,3]. In addition to the HLA-Cw6*0602 allele, other genes from immunological pathways have been linked to the risk of developing psoriasis [4,5,6,7,8,9].
The role of innate immunity genes in psoriatic disease is well stablished, including components of the interferon and anti-viral response pathways such as the interferon-induced helicase C domain-containing protein 1 (IFIH1), also known as melanoma differentiation-associated protein 5 (MDA5; encoded by the IFIH1 gene). IFIH1/MDA5 contains two N-terminal CARD (caspase activation recruitment) domains followed by a helicase ATP-binding domain and a C-terminal RIG-1 like domain. Upon recognition of double stranded RNAs (dsRNAs) by the helicase domains, MDA5 undergoes conformational changes that promote assembly into filaments and association of the CARD-domain with anti-viral proteins, leading to the transcription of early innate type-I interferon response genes [10,11,12,13,14,15]. Type-I interferons and their downstream products are also increased in psoriatic lesions [16,17,18].
Functional variants in the IFIH1 that decrease the protein expression and MDA5 activity have been associated with increased risk of viral infections and the extent of viral disease, including COVID-19 (caused by SARS-CoV-2) [19,20,21,22,23]. While low-activity variants would predispose to infection, IFIH1 variants that result in a MDA5 gain of function might result in the over activation of inflammatory pathways and increased risk of inflammatory and autoimmune diseases [24,25,26]. Several studies have investigated IFIH1/MDA5 in psoriasis. Among others, cytokines such as TNFα and IFNγ can increase the expression of MDA5 in keratinocytes, and MDA5 is increased in psoriatic skin lesions [27,28,29]. Moreover, improvement of the skin lesions after treatment with ultraviolet would be correlated with the reduction of MDA5 levels [29]. In agreement with this functional role, IFIH1 gene variants have been associated with the risk of developing psoriatic disease [30,31].
Several IFIH1 common variants have been associated with the risk of immune mediated diseases, including the single nucleotide polymorphism (SNPs) rs1990760 C>T. This results in a missense change (p.Ala946Thr) that would affect the ATPase activity due to a conformational protein change and has been associated with type 1 diabetes and other autoimmune diseases [32,33,34,35,36,37,38,39,40]. This SNP could be also linked to IFIH1 expression, since the autoimmune risk allele T (946Thr) was associated with increased IFIH1 transcription in interferon-β stimulated peripheral blood mononuclear cells [39]. Human peripheral blood mononuclear cells with the rs1990760 T allele expressed higher basal type I interferons, while IFIH1946T knock-in mice displayed enhanced basal expression of type I interferons and survived a lethal viral challenge, but also exhibited increased risk for autoimmune traits [34].
In addition to common functional polymorphisms, IFIH1 has several rare deleterious variants. SNP rs35744605 (c.1879 G>T) results in a premature stop-codon (p.Glu627*) with the loss of the 399 C-terminal amino acids of MDA5. This variant would be in approximately 1% of the Europeans and was associated with increased risk for recurrent viral infection but decreased risk of type 1 diabetes (T1D) [20,39,40]. SNP rs35337543 (c.1641 +1G>C) is a putative splicing change that would skips exon 8 of the mRNA, removing 39 amino acids of the MDA5 helicase domain [20,41]. This variant would be a risk factor for recurrent viral infection while protective for T1D [20,42,43]. These rare T1D-protective alleles were linked to reduced expression of IFIH1 among heterozygous individuals [40].
The association between IFIH1 variants and psoriasis has been supported by some authors [44]. Among others, rs1990760 was associated with psoriasis by Genome Wide Association Studies (GWAS) with OR=1.22 [45,46]. Other studies reported significant association with SNP rs17715343 and rs17716942, in strong linkage disequilibrium with rs1990760 [47,48].
Our aim was to investigate the association of the IFIH1 SNPs rs1990760, rs35744605, and rs35337543 and the risk of developing psoriasis, and whether these variants have a significant effect on onset age, disease severity and the risk of PsA.
2. Methods
2.1. Study Population and Data Collection
The study was approved by the Ethical Committee of the Principality of Asturias and involved a total of 572 psoriatic disease patients aged ≥18 years. They were of Caucasian ancestry and from the region of Asturias (Northern Spain, total population 1 million). The patient´s cohort was registered as a Biobank Collection by the Spanish Instituto de Salud Carlos III (reference C.0003441).
The patients had been diagnosed with chronic plaque psoriasis and recruited through the Departments of Dermatology and Rheumatology of Hospital Universitario Central de Asturias (HUCA) and Hospital Universitario Alvarez Buylla (HUAB) between January 2007 and August 2017. The main demographic and clinical characteristics of the patients are summarized in Table 1. Psoriasis was diagnosed based on clinical findings, and disease severity was defined according to the Psoriasis Area and Severity Index (PASI; severe, PASI ≥10). All the patients were genotyped for the HLA-Cw6 (PSORS1) [49]. Early-onset psoriasis (EOPs) was defined as a disease beginning at age ≤40 years. The presence of arthritis was assessed by a rheumatologist according to CASPAR criteria. Patients who had not developed arthritis 10 or more years after psoriasis onset were categorized as having pure cutaneous psoriasis (PCP). This is because 10 years is the average time between the onset of skin lesions and the onset of arthritis in most studies, making it unlikely that these patients with PCP would develop arthritis over time. This form of categorization has been shown to be reliable in other studies [50].
2.2. IFIH1 Variants Genotyping
The DNA was obtained from whole blood leukocytes and all the individuals were genotyped for the IFIH1 polymorphisms with real time PCR Taqman assays (Fisher scientific): rs35337543, assay id C_25985625_10; rs35744605, assay id C_25982959_10; rs1990760, C_2780299_30. The quality of the genotyping method was determined by sequencing PCR fragments with different genotypes.
2.3. Statistical Analysis
Patient’s data were recorded in an Excel file (Microsoft office). The statistical analysis was performed with the R-software (http://www.r-project.org). Allele and genotype frequencies between the groups were compared with the Chi2 and Fisher’s exact tests. A p< 0.05 value was considered as statistically significant.
3. Results
A total of 440 patients had an onset age ≤40 years (EOPs) compared to 132 cases >40 years (Table 1). Severe disease (PASI ≥10) and the presence of Cw6 was significantly more frequent in EOPs patients (p<0.001). Arthritis was also more common in this group (32% vs 24%) without significant difference with patients >40 years. In reference to the IFIH1 variants, rs1990760 T (946Thr) that has been linked to increased gene expression and interferon response was significantly more frequent in the ≤40 years group (0.66 vs 0.52), with higher frequencies for the CC and CT genotypes (Table 1). Thus, this allele was associated with a significantly increased risk for EOPs (CC+TC vs CC, p<0.001) with an OR=3.65 (95%IC=2.21-6.02). Allele frequencies for the rs1990760 SNP were T=0.38 and C=0.62 in the whole cohort, close to the reported frequencies among Europeans (T=0.36-0.40).
The two rare rs35744605 T (627Stop) and rs35337543 C (splicing variant) were found in our patients, all heterozygotes, without significant difference between the two age groups. There were no homozygotes for the two rare alleles, and the reduced sample size did not allow to determine the putative protective effect of the two rare homozygous genotypes (Table 1).
The multiple logistic regression analysis showed that severe psoriasis, Cw6 positivity, and carrying the rs1990760 T allele were independent associated factors for EOPs (Table 1). Psoriasis has been associated with the presence of the Cw6 variant, that would distinguish early versus late-onset disease. In agreement with this, a total of 48% and 23% had an onset age ≤40 and >40 years, respectively (p<0.001). We determined the distribution of the onset-age according to the Cw6 + rs1990760 status. Among Cw6+ there were higher frequencies of rs1990760 TT among EOPs (45% vs 27%, p=0.06), while among Cw6- the TT and TC were increased among EOPs patients (93% vs 72%, p<0.001) (Figure 1). This suggested that the high expression 946Thr might increase disease risk more pronouncedly among Cw6 negative patients than among Cw6+ ones.
We also determined the association between the IFIH1 variants and PsA (Table 2). The presence of arthritis was significantly associated with a severe psoriasis, with a trend toward increased risk among early-onset patients. We found a significantly increased frequency of rs1990760 TT (946Thr homozygotes) in the arthritis group (47% vs 35%, p=0.006). There were no significant differences between the rs35744605 T and rs35337543 C variants and PsA, although the number of the rare allele carriers was too low to reach a minimum statistical power (Table 2). We performed multiple logistic regression analyses and the presence of PsA was significantly associated with a severe PASI (OR=2.14, 95%CI=1.46-3.16), female sex (OR=1.63, 1.12-2.38), and the rs1990760 TT genotype (OR=1.62, 1.11-2.37), while the Cw6 positivity was protective (OR=0.65, 0.44-0.95) and there was a trend toward increased risk for patients with EOPs (OR=1.29, 0.81-2.09).
4. Discussion
The population genetics of the functional IFIH1 variants would be shaped by their effect on the interferon response pathways. Accordingly, variants associated with higher expression/function might be associated with increased protection against viral infection [51,52,53,54]. Recently, IFIH1 variants have been also associated with increased risk for SARS-CoV-2 severe disease (COVID-19) [23,55,56,57]. In opposition to its antiviral protective effect these higher expression variants might also increase the risk for autoimmune diseases [32,33,34,35,36,37,38,39,40,58]. IFIH1 encodes the MDA5 protein, a RIG-I-like receptor dsRNA helicase enzyme which activation induces the transcription of type 1 interferon genes. The type 1 IFN pathway has been implicated in the pathogenesis of psoriatic disease. Thus, type 1 IFN pathway genes were found upregulated in psoriatic skin lesions [17,59]. In mice, the type 1 IFN pathway would play an important role in the T cell-mediated skin inflammation, and mice treated with IFNα or IFNβ neutralizing antibodies showed an attenuated Th17 mediated skin inflammation [60,61].
Our main finding was the significant association of the rs1990760 T (946Thr) and EOPs. Interestingly, in agreement with our results, the alleles associated with increased risk of psoriasis were also associated with younger age of psoriasis onset by other authors [62]. Differences in the genetic association between early and late onset disease have been described for other immune pathway polymorphisms [63]. The association between psoriasis and IFIH1 variants, either isolated or interacting with other gene variants, has been previously reported [64]. The risk alleles of these variants were in strong linkage disequilibrium (Figure 2). Among others, the rare rs35667974 (p.Ile923Val) was found to be protective for PsA [30,65]. The risk rs35667974 T allele was in strong linkage disequilibrium with the risk rs1990760 T allele. Another IFIH1 SNP previously associated with psoriasis is rs984971 A/G, with the risk A allele defining haplotype with rs1990760 T [66]. A study in Spanish patients found a significant increased risk for the rs17716942 T allele carriers, with a more pronounced effect for PsA (OR=1.36) [67]. Notably, the T-risk allele is in strong linkage disequilibrium with rs1990760 T.
Although highly conserved among vertebrates the rs1990760 missense change was predicted not to affect the protein function by Shigemoto et al. These authors determined the functional effect of several IFIH1 variants and found that the Ala946Thr influences neither dsRNA binding nor IFN gene activation [68]. In contrast, the Ile923Val exhibited loss of function without affecting the dsRNA binding activity, a fact that suggested a novel role for this residue in the IFN-signaling. The lack of functional effect for rs1990760 would reinforce the hypothesis that linkage disequilibrium with other IFIH1 variants might explain the reported association with psoriasis and autoimmune diseases. In contrast, Gorman et al. provided strong evidence of a direct functional effect for the Ala946Thr variant, since peripheral blood mononuclear cells with the risk rs1990760 T allele expressed higher basal IFN-1, and IFIH1946T knock-in mice displayed enhanced basal expression of type I interferons and increased risk for autoimmune traits such as type 1 diabetes and systemic lupus [34].
We did not find significant differences between the two psoriasis age groups for the rare rs35744605 and rs35337543 polymorphisms. The two were likely pathogenic variants causing a truncating protein and an alternative transcript, and were previously associated with increased risk for recurrent viral infections [20]. The rare alleles at the two variants could be also protective against autoimmune diseases [42]. The rare rs35337543 C allele causes skipping of exon 8 (IFIH1-Δ8) without frameshifting, but removes 39 amino acids of the helicase 1 and the linker part between helicase 1 and helicase 2 domains [20]. Cells transfected with IFIH1-Δ8 plasmids showed an attenuated IFN-β induction compared to IFIH1-wt transfected cells, and cotransfection of IFIH1-wt with the mutant isoform showed interference with IFN-β production [20]. Moreover, the ATPase activity after dsRNA stimulation was undetectable in cells transfected with the mutant compared to IFIH1-wt isoform. The deleterious effect of the IFIH1-Δ8 and other mutations might be explained by a reduction in the protein stability and interference with the wild-type protein [20]. The rare rs35744605 allele results in a premature stop codon and a truncated protein lacking the C-terminal RNA-helicase domain. The functional effect of this variant was measured in peripheral blood monocytes transfected with compounds that mimic viral dsRNA, and cells from wild-type individuals showed increased IFN-β secretion compared to cells from mutation carriers [39]. The lack of association between these variants and Psor onset-age might be explained by the reduced size of our study cohort. The rare rs35744605 and rs35337543 alleles showed a frequency of 0.01 in the whole patient cohort, close to the one reported among Caucasians (0.01-0.02). Due to the very low frequencies, to conclude a protective effect on the risk for psoriatic disease would require larger cohort of patients.
We also confirmed the widely reported increased frequency of Cw6 among EOPs compared to late-onset cases. Interestingly, the effect was more pronounced among Cw6 negative patients with rs1990760 TT+TC that was significantly increased among early-onset patients.
The rs1990760 TT genotype was significantly increased also among patients who developed PsA. However, because the arthritis status was stablished taking into account the 10 years period after the initial psoriasis diagnosis, we cannot exclude that this association with PsA might be biased by not including patients who would develop PsA later. Nonetheless, the ten- year period is the average time between the onset of skin lesions and joint complaints, making very unlikely that most of PCP patients will develop arthritis [50]. Moreover, the association between IFIH1 variants and PsA has been previously reported. For instance, a large-scale study by Stuart et al described a significant association between the rs1990760 T allele and psoriatic disease with higher odds ratio for PsA (OR=1.585; p=1x10-14) than for cutaneous disease (OR=1.425; p=7.6x10-10) [45]. Julia et al. also found a significant association between PsA and rs17716942, a variant in strong linkage disequilibrium with rs1990760 [67].
Our findings highlight the relevance of IFIH1/MDA5 in the risk of developing psoriatic disease with a significant increased risk for EOPs and PsA among carriers of the high expression genotypes. This association was supported by studies that showed increased expression of interferon system proteins, including IFIH1, in skin lesions from psoriasis patients compared with healthy skin [18,45]. These findings point to IFIH1/MDA5 as a pharmacological target whose inhibition could be effective in treating psoriatic disease.
Our study has several limitations. First, it was based on a limited number of patients and from a single Spanish region and would thus require replication in larger cohorts from different populations. The rs35744605 and rs35337543 are predicted to be likely pathogenic by introducing a premature stop codon and an alternative mRNA. The two deleterious alleles were very rare in our population, and no homozygous was found in our cohort. Moreover, based on the observed allele frequencies homozygotes would have a frequency lower than 1 per 1,000 among Europeans. This makes difficult to stablish a significant conclusion about the increased risk for psoriasis among wild-type homozygotes, that would require much larger cohorts.
In conclusion, we found a significant association between a common IFIH1 variant that has been linked to increased gene expression and EOPs. The effect on early-onset disease risk was more pronounced among Cw6-negative patients. The common rs1990760 T allele was also an independent risk factor for PsA in our cohort. This risk allele was in linkage disequilibrium with other variants previously associated with the risk of psoriasis and PsA as well. Our study reinforces the widely reported role of IFIH1 gene variants on psoriatic disease and other immune mediated diseases.
Author Contributions
All the authors contributed to this work by recruiting the study cohorts or performing the genetic and statistical analysis. All the authors approved the submission of this manuscript.
Funding
This work was supported by a grant from the Spanish Instituto de Salud Carlos III-European FEDER funds (no. PI16/01792).
Data Availability Statement
The materials and raw data described in the manuscript will be freely available to any researcher without breaching participant´s confidentiality. To facilitate the revision of the results by other researchers, a file with the patient´s data is available as an excel file upon request to the corresponding author.
Conflicts of Interest
None of the authors have competing interests related to this work.
References
- Harden JL, Krueger JG, Bowcock AM. The immunogenetics of Psoriasis: A comprehensive review. J Autoimmun. 2015; 64:66-73.
- Oka A, Mabuchi T, Ozawa A, Inoko H. Current understanding of human genetics and genetic analysis of psoriasis. J Dermatol. 2012; 39: 231-41.
- Coto E, Santos-Juanes J, Coto-Segura P, Alvarez V. New psoriasis susceptibility genes: momentum for skin-barrier disruption. J Invest Dermatol. 2011; 131:1003-5.
- Nair RP, Stuart PE, Nistor I, et al. Sequence and haplotype analysis supports HLA-C as the psoriasis susceptibility 1 gene. Am J Hum Genet. 2006; 78:827-51.
- Gudjonsson JE, Karason A, Runarsdottir EH, et al. Distinct clinical differences between HLA-Cw*0602 positive and negativepsoriasis patients--an analysis of 1019 HLA-C- and HLA-B-typed patients.J Invest Dermatol. 2006; 126: 740-5.
- Jordan CT, Cao L, Roberson ED, et al. Rare and common variants in CARD14, encoding an epidermal regulator of NF-kappaB, in psoriasis.Am J Hum Genet. 2012; 90: 796-808.
- Nair RP, Duffin KC, Helms C, et al. Genome-wide scan reveals association of psoriasis with IL-23 and NF-kappaB pathways.NatGenet. 2009; 41: 199-204.
- Nikamo P, Lysell J, Ståhle M. Association with Genetic Variants in the IL-23 and NF-κB Pathways Discriminates between Mild and Severe Psoriasis Skin Disease.J Invest Dermatol. 2015; 135: 1969-76.
- González-Lara L, Coto-Segura P, Penedo A, et al. SNP rs11652075 in the CARD14 gene as a risk factor for psoriasis (PSORS2) in a Spanish cohort.DNA Cell Biol. 2013; 32: 601-4.
- Dias Junior AG, Sampaio NG and Rehwinkel J. A Balancing Act: MDA5 in Antiviral Immunity and Autoinflammation. Trends Microbiol 2019;27(1):75–85;. [CrossRef]
- Sampaio NG, Chauveau L, Hertzog J, Bridgeman A, Fowler G, Moonen JP, et al. The RNA sensor MDA5 detects SARS-CoV-2 infection. Sci Rep. 2021; 11: 13638. PMID: 34211037. [CrossRef]
- Thorne LG, Reuschl AK, Zuliani-Alvarez L, Whelan MVX, Turner J, Noursadeghi M, et al. SARS-CoV-2 sensing by RIG-I and MDA5 links epithelial infection to macrophage inflammation. EMBO J. 2021; 40: e107826. PMID: 34101213. [CrossRef]
- Yin X, Riva L, Pu Y, Martin-Sancho L, Kanamune J, Yamamoto Y, et al. MDA5 governs the Innate Immune Response to SARS-CoV-2 in Lung Epithelial Cells.Cell Rep. 2021; 34: 108628. PMID: 33440148 doi: 0.1016/j.celrep.2020.108628.
- Reikine S, Nguyen JB, Modis Y. Pattern Recognition and Signaling Mechanisms of RIG-I and MDA5. Frontiers in immunology. 2014; 5:342. [PubMed: 25101084].
- Vabret N, Blander JM. Sensing microbial RNA in the cytosol. Frontiers in immunology. 2013; 4:468. [PubMed: 24400006].
- Yao Y, Richman L, Morehouse C, de los Reyes M, Higgs BW, Boutrin A, et al. Type I interferon: potential therapeutic target for psoriasis? PloS one. 2008; 3:e2737. [PubMed: 18648529].
- Van der Fits L, van der Wel LI, Laman JD, Prens EP, Verschuren MC. In psoriasis lesional skin the type I interferon signaling pathway is activated, whereas interferon-alpha sensitivity is unaltered. The Journal of investigative dermatology. 2004; 122:51–60. [PubMed: 14962089].
- Schmid P, Itin P, Cox D, McMaster GK, Horisberger MA. The type I interferon system is locally activated in psoriatic lesions. Journal of interferon research. 1994; 14:229–34. [PubMed: 7861026].
- Kim WK, Jain D, Sánchez MD, Koziol-White CJ, Matthews K, Ge MQ, et al. Deficiency of melanoma differentiation-associated protein 5 results in exacerbated chronic postviral lung inflammation. Am J Respir Crit Care Med. 2014; 189: 437-48. PMID: 24417465. [CrossRef]
- Asgari S, Schlapbach LJ, Anchisi S, Hammer C, Bartha I, Junier T, et al. Severe viral respiratory infections in children with IFIH1 loss-of-function mutations. Proc Natl Acad Sci U S A. 2017; 114: 8342-8347. PMID: 28716935. [CrossRef]
- Lamborn IT, Jing H, Zhang Y, Drutman SB, Abbott JK, Munir S, et al. Recurrent rhinovirus infections in a child with inherited MDA5 deficiency. J Exp Med. 2017; 214: 1949-1972. PMID: 28606988. [CrossRef]
- Rebendenne A, Valadão ALC, Tauziet M, Maarifi G, Bonaventure B, McKellar J, et al. SARS-CoV-2 Triggers an MDA-5-Dependent Interferon Response Which Is Unable to Control Replication in Lung Epithelial Cells. J Virol 2021; 95: e02415-20. PMID: 33514628. [CrossRef]
- Amado-Rodríguez L, Salgado Del Riego E, Gomez de Ona J, López Alonso I, Gil-Pena H, López-Martínez C, et al. Effects of IFIH1 Rs1990760 Variants on Systemic Inflammation and Outcome in Critically Ill COVID-19 Patients in an Observational Translational Study. Elife 2022; 11: e73012; PMID: 35060899. [CrossRef]
- Rice GI, Del Toro Duany Y, Jenkinson EM, Forte GM, Anderson BH, Ariaudo G, et al. Gain-of-function mutations in IFIH1 cause a spectrum of human disease phenotypes associated with upregulated type I interferon signaling. Nat Genet. 2014; 46: 503-509. PMID: 24686847. [CrossRef]
- Rice GI, Park S, Gavazzi F, Adang LA, Ayuk LA, Van Eyck L, et al. Genetic and phenotypic spectrum associated with IFIH1 gain-of-function. Hum Mutat. 2020; 41: 837-849. PMID: 31898846. [CrossRef]
- TonuttiA, Motta F, Ceribelli A, Isailovic N, Selmi C, De Santis M. Anti-MDA5 Antibody Linking COVID-19, Type I Interferon, and Autoimmunity: A Case Report and Systematic Literature Review. Front Immunol. 2022; 13: 937667. PMID: 35833112. [CrossRef]
- Kitamura H, Matsuzaki Y, Kimura K, Nakano H, Imaizumi T, Satoh K, et al. Cytokine modulation of retinoic acid-inducible gene-I (RIG-I) expression in human epidermal keratinocytes. Journal of dermatological science. 2007; 45:127–34. [PubMed: 17182220].
- Prens EP, Kant M, van Dijk G, van der Wel LI, Mourits S, van der Fits L. IFN-alpha enhances poly-IC responses in human keratinocytes by inducing expression of cytosolic innate RNA receptors: relevance for psoriasis. The Journal of investigative dermatology. 2008; 128:932–8. [PubMed: 17928888].
- Racz E, Prens EP, Kant M, Florencia E, Jaspers NG, Laman JD, et al. Narrowband ultraviolet B inhibits innate cytosolic double-stranded RNA receptors in psoriatic skin and keratinocytes. The British journal of dermatology. 2011; 164:838–47. [PubMed: 21143460].
- Li Y, Liao W, Cargill M, Chang M, Matsunami N, Feng BJ, et al. Carriers of rare missense variants in IFIH1 are protected from psoriasis. The Journal of investigative dermatology. 2010; 130:2768–72. [PubMed: 20668468].
- Bijlmakers MJ, Kanneganti SK, Barker JN, Trembath RC, Capon F. Functional analysis of the RNF114 psoriasis susceptibility gene implicates innate immune responses to double-stranded RNA in disease pathogenesis. Hum Mol Genet. 2011; 20:3129–37. [PubMed: 21571784].
- Liu S, Wang H, Jin Y, et al. IFIH1 Polymorphisms Are Significantly Associated with Type 1 Diabetes and IFIH1 Gene Expression in Peripheral Blood Mononuclear Cells. Hum Mol Genet 2009; 18:358–365;. [CrossRef]
- Cen H, Wang W, Leng R-X, et al. Association of IFIH1 Rs1990760 Polymorphism with Susceptibility to Autoimmune Diseases: A Meta-Analysis. Autoimmunity 2013; 46:455–462;. [CrossRef]
- Gorman JA, Hundhausen C, Errett JS, et al. The A946T Variant of the RNA Sensor IFIH1 Mediates an Interferon Program That Limits Viral Infection but Increases the Risk for Autoimmunity. Nat Immunol 2017;18:744–752;. [CrossRef]
- Borysewicz-SańczykH, Sawicka B, Wawrusiewicz-Kurylonek N, Głowińska-Olszewska B, Kadłubiska A, Gościk J, et al. Genetic Association Study of IL2RA, IFIH1, and CTLA-4 Polymorphisms with Autoimmune Thyroid Diseases and Type 1 Diabetes. Front Pediatr. 2020 Aug 21; 8:481. [CrossRef]
- Shapiro MR, Thirawatananond P, Peters L, et al. De-Coding Genetic Risk Variants in Type 1 Diabetes. Immunol Cell Biol 2021; 99:496–508;. [CrossRef]
- FunabikiM, Kato H, Miyachi Y, Toki H, Motegi H, Inoue M, et al. Autoimmune disorders associated with gain of function of the intracellular sensor MDA5. Immunity. 2014; 40: 199-212. PMID: 24530055. [CrossRef]
- BammingD, Horvath CM. Regulation of signal transduction by enzymatically inactive antiviral RNA helicase proteins MDA5, RIG-I, and LGP2. J Biol Chem. 2009; 284: 9700–12. PMID: 19211564. [CrossRef]
- Chistiakov DA, Voronova NV, Savost'Anov KV, TurakulovRI. Loss-of-Function Mutations E6 27X and I923V of IFIH1 Are Associated with Lower Poly(I:C)–Induced Interferon-β Production in Peripheral Blood Mononuclear Cells of Type 1 Diabetes Patients. Human Immunology 2010; 71: 1128–34; PMID: 20736039. [CrossRef]
- DownesK, Pekalski M, Angus KL, Hardy M, Nutland S, Smyth DJ, et al. Reduced expression of IFIH1 is protective for type 1 diabetes. PLoS One. 2010; 5: e12646. PMID: 20844740. [CrossRef]
- Forbester JL and Humphreys IR. Genetic Influences on Viral-Induced Cytokine Responses in the Lung. Mucosal Immunol 2021; 14: 14–25;. [CrossRef]
- Nejentsev S, Walker N, Riches D, Egholm M, Todd JA. Rare Variants of IFIH1, a Gene Implicated in Antiviral Responses, Protect against Type 1 Diabetes. Science 2009; 324: 387–9; PMID: 19264985. [CrossRef]
- Mine K, Yoshikai Y, Takahashi H, Mori H, Anzai K, NagafuchiS. Genetic Susceptibility of the Host in Virus-Induced Diabetes. Microorganisms 2020; 8: E1133; PMID: 32727064. [CrossRef]
- Hébert HL, Bowes J, Smith RL, Flynn E, Parslew R, Alsharqi A, McHugh NJ, Barker JN, Griffiths CE, Barton A, Warren RB. Identification of loci associated with late-onset psoriasis using dense genotyping of immune-related regions. Br J Dermatol. 2015 Apr;172(4):933-9. Epub 2015 Feb 5. PMID: 25124732. [CrossRef]
- Stuart PE, Nair RP, Tsoi LC, et al. Genome-wide Association Analysis of Psoriatic Arthritis and Cutaneous Psoriasis Reveals Differences in Their Genetic Architecture. Am J Hum Genet. 2015; 97:816-36.
- Tsoi LC, Spain SL, Knight J, Ellinghaus E, Stuart PE, Capon F, et al. Identification of 15 new psoriasis susceptibility loci highlights the role of innate immunity. Nat Genet. 2012 Dec;44(12):1341-8. Epub 2012 Nov 11. PMID: 23143594. [CrossRef]
- Strange A, Capon F, Spencer CC, Knight J, Weale ME, Allen MH, et al A genome-wide association study identifies new psoriasis susceptibility loci and an interaction between HLA-C and ERAP1. Nat Genet. 2010 Nov;42(11):985-90. Epub 2010 Oct 17.PMID: 20953190. [CrossRef]
- Patrick MT, Stuart PE, Raja K, Gudjonsson JE, Tejasvi T, Yang J, Chandran V, Das S, Callis-Duffin K, Ellinghaus E, Enerbäck C, Esko T, Franke A, Kang HM, Krueger GG, Lim HW, Rahman P, Rosen CF, Weidinger S, Weichenthal M, Wen X, Voorhees JJ, Abecasis GR, Gladman DD, Nair RP, Elder JT, Tsoi LC. Genetic signature to provide robust risk assessment of psoriatic arthritis development in psoriasis patients. Nat Commun. 2018 Oct 9;9(1):4178. PMID: 30301895. [CrossRef]
- Tazi Ahnini R, Camp NJ, Cork MJ, et al. Novel genetic association between the corneodesmosin (MHC S) gene and susceptibility to psoriasis. Hum Mol Genet. 1999; 8: 1135-40.
- Gladman DD, C. Antoni, P. Mease, D. O. Clegg, and P. Nash, “Discussion: clinical features, epidemiology, classification criteria, and quality of life in psoriasis and psoriatic arthritis,” Ann Rheum Dis, vol. 64, no. 2, pp. ii24-ii25, 2005.
- Pang L, Gong X, Liu N, Xie G, Gao W, Kong G, Li X, Zhang J, Jin Y, Duan Z. A polymorphism in melanoma differentiation-associated gene 5 may be a risk factor for enterovirus 71 infection. Clin Microbiol Infect. 2014 Oct;20(10): O711-7. Epub 2014 Apr 10.PMID: 24621100. [CrossRef]
- Yao Y, Shen Y, Shao H, Liu Y, Ji Y, Du G, Ye X, Huang P, Chen H. Polymorphisms of RIG-I-like receptor influence HBV clearance in Chinese Han population.J Med Virol. 2021 Aug;93(8):4957-4965. Epub 2021 Apr 3. PMID: 33783003. [CrossRef]
- Domsgen E, Lind K, Kong L, Hühn MH, Rasool O, van Kuppeveld F, Korsgren O, Lahesmaa R, Flodström-Tullberg M.An IFIH1 gene polymorphism associated with risk for autoimmunity regulates canonical antiviral defence pathways in Coxsackievirus infected human pancreatic islets. Sci Rep. 2016 Dec 21;6:39378. PMID: 28000722. [CrossRef]
- Dou Y, Yim HC, Kirkwood CD, Williams BR, Sadler AJ.The innate immune receptor MDA5 limits rotavirus infection but promotes cell death and pancreatic inflammation. EMBO J. 2017 Sep 15;36(18):2742-2757. Epub 2017 Aug 29. PMID: 28851763. [CrossRef]
- Dieter C, de Almeida Brondani L, Lemos NE, Schaeffer AF, Zanotto C, Ramos DT, Girardi E, Pellenz FM, Camargo JL, Moresco KS, da Silva LL, Aubin MR, de Oliveira MS, Rech TH, Canani LH, Gerchman F, Leitão CB, Crispim D. Polymorphisms in ACE1, TMPRSS2, IFIH1, IFNAR2, and TYK2 Genes Are Associated with Worse Clinical Outcomes in COVID-19. Genes (Basel). 2022 Dec 22;14(1):29. PMID: 36672770. [CrossRef]
- Nln I, Fernandez-Ruiz R, Muskardin TLW, Paredes JL, Blazer AD, Tuminello S, Attur M, Iturrate E, Petrilli CM, Abramson SB, Chakravarti A, Niewold TB. Interferon pathway lupus risk alleles modulate risk of death from acute COVID-19. Transl Res. 2022 Jun;244:47-55. Epub 2022 Jan 31. PMID: 35114420. [CrossRef]
- Muñiz-Banciella MG, Albaiceta GM, Amado-Rodríguez L, Del Riego ES, Alonso IL, López-Martínez C, Martín-Vicente P, García-Clemente M, Hermida-Valverde T, Enríquez-Rodriguez AI, Hernández-González C, Cuesta-Llavona E, Alvarez V, Gómez J, Coto E. Age-dependent effect of the IFIH1/MDA5 gene variants on the risk of critical COVID-19. Immunogenetics. 2023 Apr;75(2):91-98. Epub 2022 Nov 25. PMID: 36434151. [CrossRef]
- Zurawek M, Fichna M, Fichna P, Skowronska B, Dzikiewicz-Krawczyk A, et al. Cumulative effect of IFIH1 variants and increased gene expression associated with type 1 diabetes. Diabetes Res Clin Pract. 2015; 107: 259-66. PMID: 25515714. [CrossRef]
- Baldwin HM, Pallas K, King V, Jamieson T, McKimmie CS, Nibbs RJ, et al. Microarray analyses demonstrate the involvement of type I interferons in psoriasiform pathology development in D6-deficient mice. J Biol Chem. (2013) 288:36473–83. [CrossRef]
- Nestle FO, Conrad C, Tun-Kyi A, Homey B, Gombert M, Boyman O, et al. Plasmacytoid predendritic cells initiate psoriasis through interferon-alpha production. J Exp Med. (2005) 202:135–43. [CrossRef]
- Gregorio J, Meller S, Conrad C, Di Nardo A, Homey B, Lauerma A, et al.. Plasmacytoid dendritic cells sense skin injury and promote wound healing through type I interferons. J Exp Med. (2010) 207:2921–30. [CrossRef]
- Lu Y, Kane S, Chen H, Leon A, Levin E, Nguyen T, Debbaneh M, Millsop JW, Gupta R, Huynh M, Butler D, Cordoro K, Liao W.The role of 39 psoriasis risk variants on age of psoriasis onset. ISRN Dermatol. 2013 Sep 23; 2013:203941. eCollection 2013.PMID: 24175098. [CrossRef]
- Reich K, Mössner R, König IR, Westphal G, Ziegler A, Neumann C. Promoter polymorphisms of the genes encoding tumor necrosis factor-alpha and interleukin-1beta are associated with different subtypes of psoriasis characterized by early and late disease onset. J Invest Dermatol. 2002 Jan;118(1):155-63. PMID: 11851889. [CrossRef]
- Zhang C, Qin Q, Li Y, Zheng X, Chen W, Zhen Q, Li B, Wang W, Sun L.Multifactor dimensionality reduction reveals the effect of interaction between ERAP1 and IFIH1 polymorphisms in psoriasis susceptibility genes. Front Genet. 2022 Nov 8;13:1009589. eCollection 2022.PMID: 36425068. [CrossRef]
- Budu-Aggrey A, Bowes J, Stuart PE, Zawistowski M, Tsoi LC, Nair R, Jadon DR, McHugh N, Korendowych E, Elder JT, Barton A, Raychaudhuri S.A rare coding allele in IFIH1 is protective for psoriatic arthritis. Ann Rheum Dis. 2017 Jul;76(7):1321-1324. Epub 2017 May 13.PMID: 28501801. [CrossRef]
- Bowes J, Budu-Aggrey A, Huffmeier U, Uebe S, Steel K, Hebert HL, Wallace C, Massey J, Bruce IN, Bluett J, Feletar M, Morgan AW, Marzo-Ortega H, Donohoe G, Morris DW, Helliwell P, Ryan AW, Kane D, Warren RB, Korendowych E, Alenius GM, Giardina E, Packham J, McManus R, FitzGerald O, McHugh N, Brown MA, Ho P, Behrens F, Burkhardt H, Reis A, Barton A. Dense genotyping of immune-related susceptibility loci reveals new insights into the genetics of psoriatic arthritis. Nat Commun. 2015 Feb 5;6:6046. PMID: 25651891. [CrossRef]
- Julià A, Tortosa R, Hernanz JM, Cañete JD, Fonseca E, Ferrándiz C,.Risk variants for psoriasis vulgaris in a large case-control collection and association with clinical subphenotypes. Hum Mol Genet. 2012 Oct 15;21(20):4549-57. Epub 2012 Jul 19.PMID: 22814393. [CrossRef]
- Shigemoto T, Kageyama M, Hirai R, Zheng J, Yoneyama M, Fujita T. Identification of loss of function mutations in human genes encoding RIG-I and MDA5: implications for resistance to type I diabetes. J Biol Chem. 2009 May 15;284(20):13348-13354. Epub 2009 Mar 26.PMID: 19324880. [CrossRef]
Figure 1.
Frequencies of the rs1990760 C/T genotypes in the early and late-onset psoriasis according to the presence of Cw6. Among the Cw6 positive the TT was more frequent in the EOPs (45% vs 27%; p=0.06). Among the Cw6 negative the TT+TC were significantly more common in the EOPs (93% vs 72%; p<0.001).
Figure 1.
Frequencies of the rs1990760 C/T genotypes in the early and late-onset psoriasis according to the presence of Cw6. Among the Cw6 positive the TT was more frequent in the EOPs (45% vs 27%; p=0.06). Among the Cw6 negative the TT+TC were significantly more common in the EOPs (93% vs 72%; p<0.001).
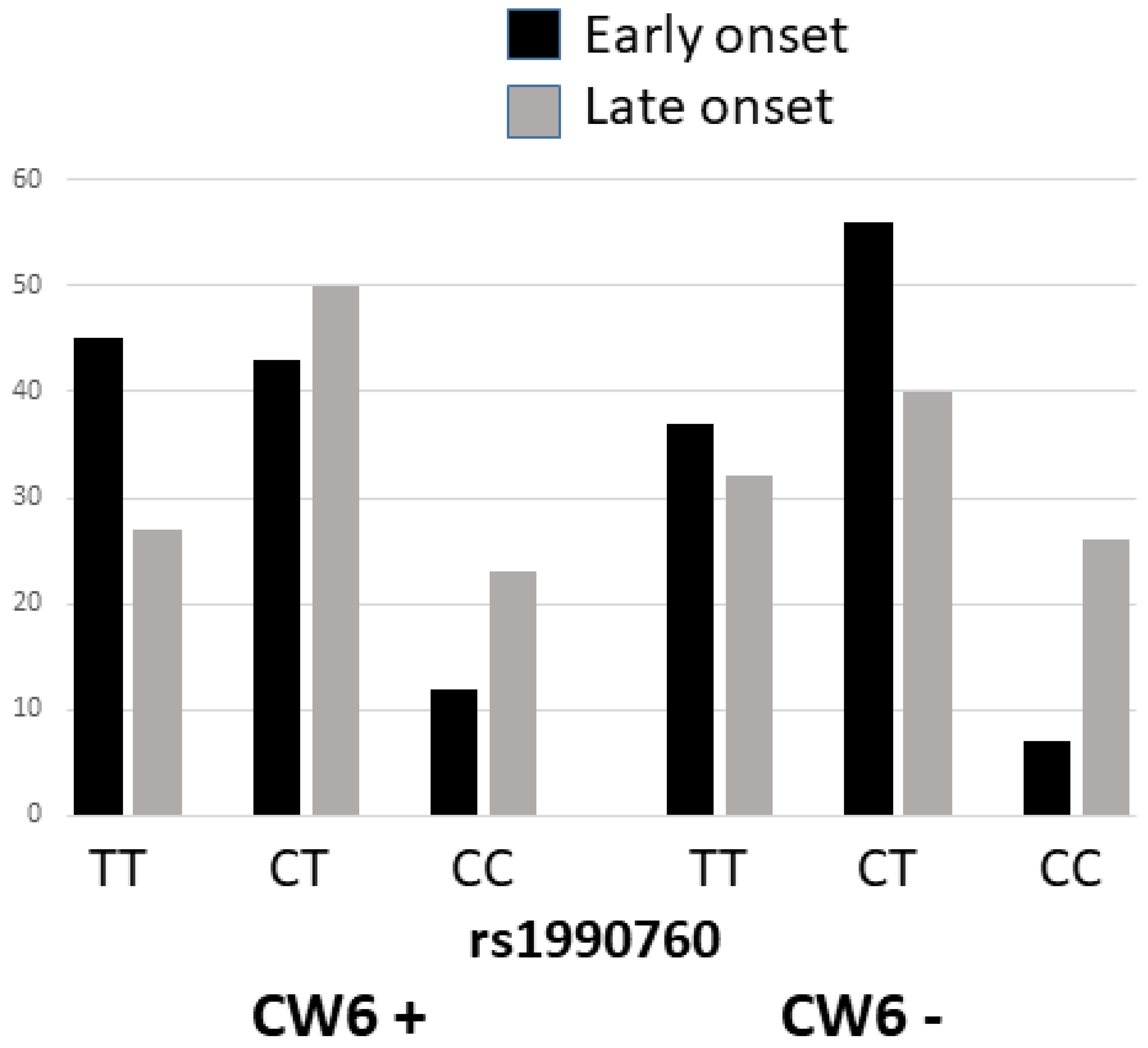
Figure 2.
Map of the IFIH1 gene indicating the position of the polymorphisms determined in this study and others that were associated with psoriasis and PsA by others. The solid bar indicates the gene, exons plus introns. The location of these variants relative to the MDA5 protein domains is also indicated. The rare rs35337543 C causes skipping of exon 8 (IFIH1-Δ8) removing 39 amino acids of the helicase 1 and the linker part between helicase 1 and helicase 2 domains. The rare 588Ter allele would result in a premature stop codon and a truncated mutant protein lacking the C-terminal RNA-helicase domain.
Figure 2.
Map of the IFIH1 gene indicating the position of the polymorphisms determined in this study and others that were associated with psoriasis and PsA by others. The solid bar indicates the gene, exons plus introns. The location of these variants relative to the MDA5 protein domains is also indicated. The rare rs35337543 C causes skipping of exon 8 (IFIH1-Δ8) removing 39 amino acids of the helicase 1 and the linker part between helicase 1 and helicase 2 domains. The rare 588Ter allele would result in a premature stop codon and a truncated mutant protein lacking the C-terminal RNA-helicase domain.
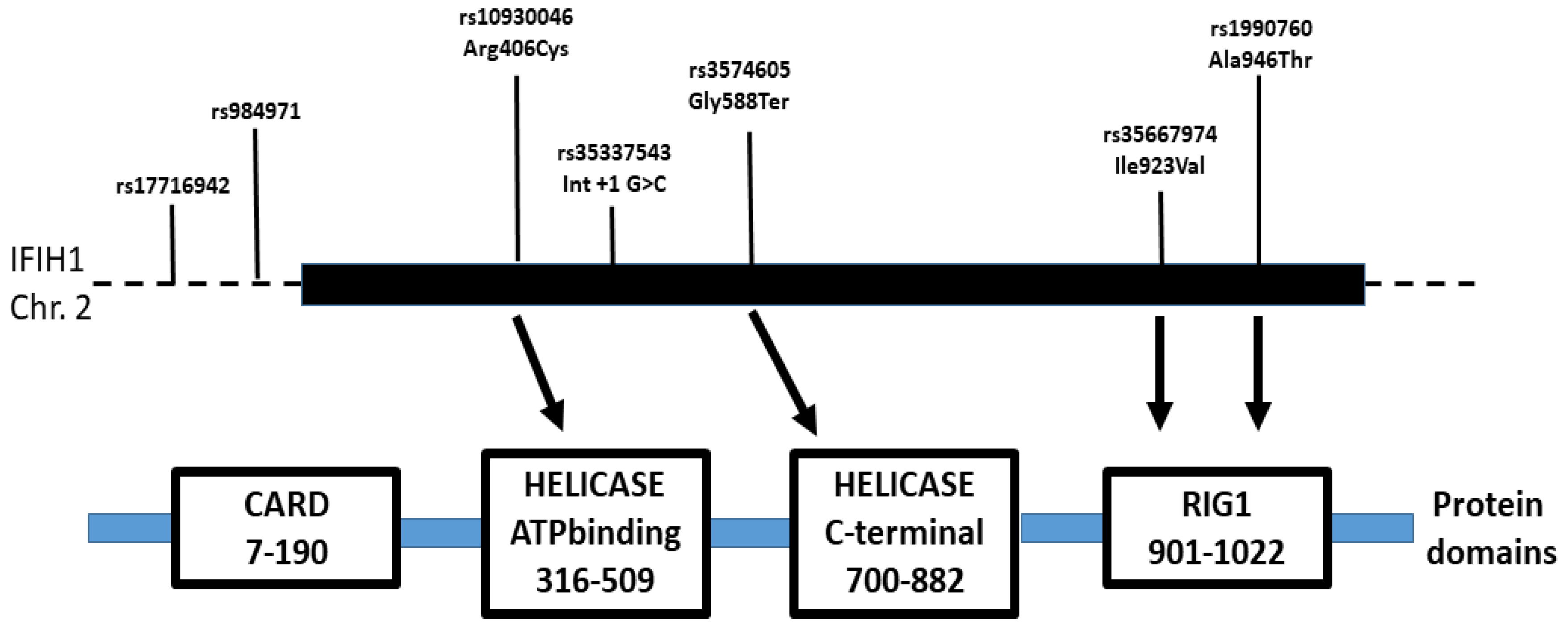

Table 1.
Main disease characteristics of the early (≤40 years) vs late onset disease. The odds ratio (OR) and confidence intervals (95%CI) corresponded to the multivariate analysis (R-linear generalized model).
Table 1.
Main disease characteristics of the early (≤40 years) vs late onset disease. The odds ratio (OR) and confidence intervals (95%CI) corresponded to the multivariate analysis (R-linear generalized model).
| Early Onset N=440 |
Late Onset N=132 |
p-Value Univariate |
OR (95%CI) Multivariate |
||
|---|---|---|---|---|---|
| Male | 232 (53%) | 77 (58%) | 0.26 | ||
| Female | 208 (47%) | 55 (42%) | 1.24 (0.81-1.90) | ||
| Onset Age range | 18-40 | 41-78 | |||
| Median PASI (range) | 11 (1-75) | 6.1 (4.1-7.2) | |||
| Severe (PASI>10) | 248 (56%) | 46 (35%) | <0.001 | 2.35 (1.52-3.66) | |
| Arthritis yes | 139 (32%) | 32 (24%) | 0.11 | 1.22 (0.76-2.01) | |
| Cw6 positive | 211 (48%) | 30 (23%) | <0.001 | 3.25 (2.05-5.29) | |
|
rs1990760 C>T * (Ala946Thr) |
|||||
| TT | 179 (41%) | 40 (30%) |
<0.001 |
4.07 (2.37-7.04) |
|
| CT | 220 (50%) | 56 (42%) | |||
| CC | 41 (9%) | 36 (27%) | |||
| ALLELE T (946Thr) Eur: T=0.60-0.64 |
0.66 | 0.52 | |||
|
rs35337543 int8 +1G>C |
|||||
| GG | 430 (98%) | 128 (97%) | 0.62 | 1.12 (0.29-3.65) | |
| GC | 10 (2%) | 4 (3%) | |||
| ALLELE C Eur: C=0.02 |
0.01 | 0.02 | |||
|
rs35744605 G>T (Glu627Stop) |
|||||
| GG | 432 (98%) | 129 (98%) | 0.74 | 1.11 (0.23.4.10) | |
| GT | 8 (2%) | 3 (2%) | |||
| ALLELE T Eur: T=0.01 |
0.01 | 0.01 | |||
*TT+CT vs CC.
Table 2.
Main characteristics of the patients with/without arthritis. The odds ratio (OR) and confidence intervals (95%CI) corresponded to the multivariate analysis (R-linear generalized model).
Table 2.
Main characteristics of the patients with/without arthritis. The odds ratio (OR) and confidence intervals (95%CI) corresponded to the multivariate analysis (R-linear generalized model).
| Arthritis Yes 171 |
Arthritis No 401 |
p-Value | OR (95%CI) Multivariate |
|
|---|---|---|---|---|
| Male | 81 | 228 |
0.04 |
|
| Female | 90 (52%) | 173 (43%) | 1.63 (1.12-2.38) | |
| Early onset psoriasis | 139 (81%) | 301 (75%) | 0.11 | 1.29 (0.81-2.09) |
| Severe (PASI≥10) | 110 (64%) | 184 (46%) | <0.001 | 2.14 (1.46-3.16) |
| Cw6 positive | 65 (38%) | 176 (44%) | 0.19 | 0.65 (0.44-0.95) |
|
rs1990760C>T * (Ala946Thr) |
||||
| TT | 80 (47%) | 139 (35%) | 0.006 | 1.62 (1.11-2.37) |
| CT | 73 (43%) | 203 (51%) | ||
| CC | 18 (11%) | 59 (15%) | ||
| Allele C | 0.32 | 0.40 | ||
|
rs35337543 int8 +1G>C |
||||
| GG | 169 (99%) | 389 (97%) | 0.20 | 3.04 (0.77-20.36) |
| GC | 2 (1%) | 12 (3%) | ||
|
rs35744605 G>T (Glu627Stop) |
||||
| GG | 168 (98%) | 393 (98%) | 0.85 | 1.23 (0.32-6.04) |
| GT | 3 (2%) | 8 (2%) |
*TT vs CC+CT.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



