
Submitted:
31 May 2023
Posted:
01 June 2023
You are already at the latest version
Abstract
Enteric protozoan pathogenic infections significantly contribute to the global burden of gastrointestinal illnesses. Their occurrence is considerable within remote and indigenous communities and regions due to reduced access to clean water and adequate sanitation. The robustness of these pathogens means requirement of harsh treatment methods such as medicinal drugs or antibiotics. However, such treatments impact the gut microbiome, and create dysbiosis, often leading to opportunistic pathogens, anti-microbial resistance, or functional gastrointestinal disorders (FGIDs) such as irritable bowel syndrome (IBS). Recent studies have shown that these impacts do not remain confined to gut, and are reflected across the gut-brain, gut-liver, and gut-lung axes, among others. Therefore, apart from the medicinal treatment, nutritional supplementation is also a key aspect of providing the recovery from this dysbiosis. Future proteins, prebiotics, probiotics, synbiotics, and food formulations offer a good solution to remedy this dysbiosis. Furthermore, the nutritional supplementation also helps to build a resilience against the opportunistic pathogens and potential future infections and disorders that may arise due to the dysbiosis. Systems biology techniques have shown to be highly effective tools to understand the biochemistry of these processes. Systems biology techniques characterises the fundamental host-pathogen interaction biochemical pathways, at various infection and recovery stages. This same mechanism also allows to track the impact of abovementioned treatment methods of gut microbiome remediation. This manuscript is organised in sections delving into system biology approaches and upcoming developments to understand (1) Infection mechanism and current global status; (2) Cross-organ impacts of dysbiosis, particularly within gut-liver and gut-lung axes; (3) Nutritional interventions. It highlights the impact of antimicrobial resistance (AMR) and Multi-drug resistance (MDR) from a perspective of protozoal infections. It also highlights the role of nutritional interventions to add resilience against the chronic problems caused by these phenomena.
Keywords:
Systems Biology
; Multiomics
; Cryptosporidium
; Giardia
; Entamoeba
; Anti-Microbial Resistance
; Multi Drug Resistance
; Probiotic
; Prebiotics
; Synbiotics
; Postbiotics
1. Introduction
One of the biggest concerns in human health is enteric infections. Besides causing considerable malnutrition, they also contribute to the death of a significant number of people worldwide. In spite of the supplementation programs of the World Health Organisation (WHO) in recent decades, 1.2 million annual deaths were still reported in 2006 [1]. Globally, 1.4 million deaths were attributed to intestinal infectious diseases in 2015, including 0.53 million deaths among children under 4 years of age[2] and in developing/underdeveloped countries (Figure 1).
2. Current global status
Protozoan infections contribute towards a large part of global mortality, causing up to 0.84 million deaths annually, as recent as 2015 [3]. Detailed information related to the spread of these organisms is provided in Table 1. While the widespread outbreak of Entamoeba spp. is less highlighted, it causes the second most common parasite-based human death worldwide (after malaria). As a result of Entamoeba spp. infections, about 10% of the human population is infected, resulting in 110,000 annual deaths [4].
The prevalence of Giardia spp. is much higher in developing and underdeveloped countries. Children under 10 years of age account for up to 20% of global outbreaks [5] and 35.2% of global waterborne disease outbreaks between 2004 and 2010.
The number of deaths caused by Cryptosporidium spp. has been estimated at 83,000 (2005), 99,800 (2010), and 64,800 (2015). In 2016, there were approximately 2.7 million reported cases of paediatric cryptosporidiosis in South and Latin America, 3.5 million in Sub-Saharan Africa, and 3.2 million in Asia, with 4.7 million cases in the Indian subcontinent There is a high prevalence of entamoebiasis and cryptosporidiosis in Australia, especially during summer. There were about 18,000 reported cases in 2010, most of them in Aboriginal and Torres Strait Islander people [6]. Out of all the clinical cases from 88 countries between 2000 - 2015, about 10.9% of the population was infected with cryptosporidiosis [6].

Table 1.
Transmission of key protozoal enteric parasites and their current global status (2000 – 2016).
Table 1.
Transmission of key protozoal enteric parasites and their current global status (2000 – 2016).
| Name | Transmission mode/ agent characteristics | Health symptoms | Hosts | Disinfection resistance | Outbreaks/ Cases | References |
|---|---|---|---|---|---|---|
| Cryptosporidium spp. | Water (drinking and recreational), faecal-oral route, | Moderate/ diarrhoea | Humans, cattle, rodents | Very high | 239/65,540(2004 – 2014) | [2,3,6,7] |
| Oocyst, 3.5 – 6.5 µm (Ø) | Ozone (4 ppm/10 min); > 3% hypochlorite | |||||
| Giardia spp. | Water, faecal-oral route | Moderate/ diarrhoea, gas, bloating, anorexia | Human, animals | High | 142/1110(2007 – 2014) | [1,3,7,8] |
| Cyst (ovoid, 8 – 18 × 7 – 10 µm) | Fenbendazole (5 mg/kg); Ozone (0.3 ppm/ 3 min); 1% Na- hypochlorite | |||||
| Entamoeba spp. | Water, food, faecal-oral route | Severe/ Colitis, dysentery, diarrhoea, liver issues | Humans | High | 15/ 9.41 million (2000 - 2015) | [2,4,8,9] |
| Cyst (10 - 16µm Ø) | Chlorine (5 ppm, pH 7, 5 min), 1% sodium hypochlorite | |||||
| Cyst (40 - 60µm Ø), trophozoites (100 – 150 µm) | 1% sodium hypochlorite |
3. Life cycle and infection mechanism
3.1. Life cycle
Cryptosporidium spp., Giardia spp. and Entamoeba spp. spread through aqueous routes, either via drinking water, food, faecal matter, or environmental calamities associated with water. The protozoan parasites spread through cyst bodies. Cysts are often the first step of infection by these protozoan parasites, originating from infected matrices or individuals. The mature cyst in Entamoeba, Cryptosporidium and Giardia consists of 4, 4 and 2 trophozoites (sporozoites in Giardia), respectively. Acidic conditions in the stomach, followed by slightly alkaline conditions in the upper intestine (duodenum) result in the process of excystation, releasing these trophozoites in the duodenum [10,11].
The trophozoites attached to intestinal mucosa are mostly localised and non-invasive. But sometimes, particularly with Entamoeba, the trophozoites do penetrate the mucosal and epidermal layers and are carried into the bloodstream to cause multi-organ infections. [10,12]. Entamoeba trophozoites are also particularly motile because of their pseudopodia. As a result, the trophozoites can move actively towards the larger intestine. The trophozoites then reproduce asexually by binary fission in the large intestine and feed on cellular debris and microbiota. Giardia sporozoites, on the other hand, reproduce by asexual binary fusion and then attach to the apical epithelium layer of the duodenal and jejunal parts of the small intestine [11]. The Cryptosporidium trophozoites can opt for both sexual (gemetogeny) and asexual (merogeny) reproduction. Immature oocysts sporulate inside the host and are released as mature cysts in faeces (Figure 2).
Except for Entamoeba spp., other protozoal parasites are considered stable interaction parasites (or asymptomatic parasites), so they do not cause significant changes in host physiology. This not only results in a delayed diagnosis of infection, but also means that most of the infection is localised within the small intestine. However, the localised nature of pathogens also mean a lower mortality, except for the immune-compromised host [13]. In the case of Entamoeba, since the parasite can penetrate epithelial lining of the intestines and enter the bloodstream, the mortality rate exceeds 50% [12].
3.2. Dysbiosis and target organs
Although most enteric protozoans are limited to intestinal systems, there are numerous variations by which they affect the host. Such as nutrient absorption and host microbiome perturbation which results in impacts to other organs. This may also affect the biochemical and physiological conditions in other organs and systems of the host body (Figure 3).
3.2.1. Dysbiosis
Bacterial species in the large intestine occur as micro-colonies or associative multispecies consortia. Studies of oral biofilms and assemblages in the large intestine have shown that the bacteria reside as biofilm communities in the cavities along mucosal lining on epithelial lining of both clinical [14] and animal [15] models. It is well known that Cryptosporidium spp. is an epithelial lining-based parasite, causing minimal invasion and generally it is unable to penetrate through mucosal layers. Therefore, it is highly likely that its interactions with bacterial biofilms in the gut lining cause behavioural changes in itself and the surrounding bacterial community. Recently, Koh, et al. [16] demonstrated a direct proportionality of Cryptosporidium growth with respect to Pseudomonas aeruginosa biofilm maturation, showing a 2-3 fold increase in population increase of Cryptosporidium under aquatic environment. In a similar study with Coquerel’s sifaka, a Madagascar lemur species, genomic studies indicated a considerable microbial diversity depletion with recovery depending on the host age (older lemurs recovered earlier than younger ones). Number of bacteria associated with human enteric diseases, such as Desulphovibrio spp. and, Enterococcus spp. increased considerably. On the other hand, the population of healthy gut bacteria such as Bifidobacterium spp., Akkermansia spp. and Succinovibrio spp., decreased considerably in infected individuals [17]. It has been reported that a release of several enzymes such as lactate dehydrogenase, proteases, haemolyses and phospholipases is caused during C. parvum infection. A resulting cellular hydrolysis and degradation causes blunting of microvilli and, measurable intestinal epithelium damage, likely affecting the microbiome (rather than C. parvum directly affecting microbiome populations, as seen in giardiasis) [18]. Our multi-omics study of mouse cryptosporidiosis indicated that during the infection, Faecalibaculum and Lachnospiraceae population depleted from duodenum onwards. However, Lactobacillus, Lachnospiraceae, Desulphovibrio, and Coriobacteria populations elevated in the jejunum and ileum [19].
This endosymbiont interaction of parasite and bacterial species has been shown in a number of studies performed with mouse models of Giardia infection [20]. It was shown that the inherent gut microbiota interferes with the Giardia infection. However, in presence of the bacterial endosymbionts such as Escherichia coli and Shigella spp., which are responsible for enteropathogenicity, the intestinal saccharide ligands change, aiding the protozoan parasites to colonise the sites by adhesion [21]. Briefly, the study indicated that the bacterial symbiotes altered the cellular surface saccharides of Crithidia oncopelti, a protozoal parasite, through the fucose binding lecithin, increasing agglutination. Besides protecting sporozoites, this activity also increases Giardia duodenalis parasite expression [22]. However, a similar interaction under in vitro condition has shown to increase pathogen virulence (in addition to sporozoite protection and increasing expression) of Entamoeba spp. infection [23,24].
A more recent study on Giardia’s effect on microbiota was reported by Caenorhabditis elegans model analysis through genomic output [25]. The study involved a multi-pronged approach where interactions between G. lamblia and commensal E. coli were tested. Additionally, the microbiota from healthy and irritable bowel syndrome (IBS) affected human representatives were transplanted in C. elegans intestine and effects of Giardia, E. coli and ‘Giardia + E. coli’ were observed. It was seen that Giardia altered E. coli gene expressions, especially of ribosomal proteins, flagellar, adhesion and transport (taurine) genes. This resulted in an increased virulence in commensal E. coli, converting them from host-microbiome symbiotic species into pathogenic species. The interaction also decreased the expression of cysB genes, responsible for producing H2S [25], which is known to be an anti-inflammatory and cryoprotective metabolite in the intestine [26]. We have also noted that during giardiasis in mice, the populations of Autopobiaceae and Desulphovibrionaceae increased, while that of Akkermansiaceae decreased [27] in the gut.
Similar observations have also been seen in Entamoeba histolytica and its relationship with host microbiome. Among the asymptomatic and amoebic liver abscess (ALA) patients, considerable alterations in 11 major microbiome populations were observed. Especially, the populations of Bacteroides spp., Bifidobacterium spp., Lactobacillus spp. and Clostridium spp. were significantly decreased. Similarly, the asymptomatic patients showed considerable decrease in commensal E. coli and increase in Pseudomonas aeruginosa populations [28]. Interestingly, one of the characteristics which sets Entamoeba spp. apart from other protozoans (to a higher degree) is its nature of intestinal mucosal colonisation. The protozoan, probably due to its ancient relationship with human hosts, has been reported to live in a commensal relationship with asymptomatic host, with non-pathogenic trophozoites releasing cysts on a continuous basis [29,30]. However, hamster model studies suggested that the invasiveness is triggered when a considerable amount of trophozoites feed on commensal bacteria. This induces a release of amoebaporic enzymes such as cysteine proteases. The resulting enzymatic activity induces the protozoan to become highly invasive in intestinal environment, causing significant epithelial cell damage (observed as inflammations and lesions). The inflammation triggers the production of several cytokines including interferon gamma (IFNγ), tumour necrosis factor (TNF) and interleukins (IL) 4, 5, 8 and 17. The following phagocytic activity causes dysbiosis (severe alteration of microbiota, disrupting host-microbiome relationship). This has not only been observed to cause FGID symptoms, but also increasing epithelial and endothelial permeability. This event aids Entamoeba spp. to infect liver tissues through portal circulation, causing cell damage, lesions and, hepatic abscess [29,31,32].
4. Cross-organ impacts
4.1. Gut-Liver axis
The portal circulation results in passage of Entamoeba spp. through to blood stream, primarily reaching liver. This event induces a host immune-response, creating severe conditions such as high influx of cytotoxins and, phagocytic nitrogen intermediates and reactive oxygen species (ROS) [33]. A noteworthy work on this interaction and establishment of E. histolytica has been reported by Rigothier, et al. [34]). The study involved a hamster model of E. histolytica infection tracking through 35S labelled protein monitoring. The study, continuing on previous models [35,36], indicated that trophozoites arriving through portal circulation enter liver through sinusoids, causing host neutrophil reaction, resulting in localised inflammation. The host liver hepatocytes degenerate and lyse due to both neutrophil activity and trophozoite cytolytic activities. During this early phase (< 12 hours), a severe environment created by neutrophil activities cause massive trophozoite mortality. However, after 12 hours, the trophozoites enter a commitment phase where considerable multiplication is observed. This causes a creation of numerous infection loci on liver parenchyma, ultimately causing numerous necrotic and inflammatory regions [34].
Unlike Entamoeba spp., Cryptosporidium spp. and Giardia spp. are unable to permeate into blood stream through intestinal epithelium. However, an infection with these parasites numerous organs beyond the gut. One of the early mouse models for Cryptosporidium spp. showed that the infection caused a swollen liver, due to inflammation of hepatic biliary system, possibly causing jaundice-like effects. Similarly, the distension was seen across bile and cystic ducts in addition to gall bladders [37]. Our mouse study indicated spiked oxalate levels in the hepatocytes during cryptosporidiosis, likely causing hyperoxaluria or a hyperoxaluria-like condition [19]. Similar cases have been observed in case of Giardia infections. The alterations of microbiota caused during giardiasis have shown to cause nutrient malabsorption [38]. The observations from Giardia infected children showed vitamin A malabsorption, not only from intestinal parasitism, but also indirectly, via liver-based retinol mobilisation. Our study [27] showed that the depletion of Akkermansiaceae spp. in gut caused an oxidative stress across the gut-liver axis, leading to elevated glutathione metabolism, especially in the small intestine, serum and liver.
4.2. Gut-Lung axis
Although very uncommon, the infection of respiratory system by protozoan parasites has been reported. Cryptosporidium and Entamoeba have been known to cause opportunistic invasion of lungs and bronchi [39]. Case-study subjects infected by E. histolytica displayed productive coughing, breathing issues, chest pain and erythema as major symptoms. This was caused by dry oropharynx mucosal membranes, displaced right lung (especially, right and middle lobes) and right-sided pneumonia combined with multicystic empyema [40]. The examination of respiratory fluids showed Entamoeba infection via cysts. Another case displayed multi-organ infection, with cysts observed in intestine and cerebrospinal fluid. However, the symptoms such as dehydration and bronchiolitis indicated a likelihood of Entamoeba infection in respiratory system as well [41]. Similarly, multi-organ Cryptosporidium infection case studies have been presented. In one study, C. parvum was identified by the 18S rDNA analysis of sputum (tracheal/ bronchial mucus expelled during coughing) and stool samples of two patients, causing deaths [42]. The case-study performed by López-Vélez, et al. [43] and Clavel, et al. [44] involved diagnostics from sputum and broncho-alveolar lavage (BAL) samples and, indicated lung cryptosporidiosis in about 16% and 100%, respectively, of infected patients.
The enteric infection’s triggering of host immune system of adolescents, has shown to cause hyper-activation of IFN and IL proteins, resulting in release of cytokines such as IL-1β and IL-10. The cytokine overexpression has previously shown to cause obstructive issues in respiratory system such as inflammation and asthma [45]. Resulting α-1-antitrypsin release in stool samples [46] has already been used as enteropathy marker. Burgess, et al. [45] reported coupling this effect with wheezing monitoring as an earlier enteric infection biomarker system in 0 – 2-year infants. The study indicated wheezing episodes in 43% of 700 infants observed. Higher wheezing indicated increased IL-10 and IL-β levels, especially during early infancy (by age of 24 weeks), whereas a successive increase in stool α-1-antitrypsin release (by age of 40 weeks) resulted in decreased wheezing episodes. In our studies of both cryptosporidiosis [19] and giardiasis [27], elevated accumulation of short-chain fatty acids (SCFAs) have been seen in the gut during infection. The gut-lung axis studies have shown that SCFAs transfer from gut to lungs via serum, and modulate the immune system [47] through dendritic cells (DCs) impairment. This causes an attenuated allergic response [48,49], and promotes regulatory T cells (Treg cells) differentiation, reducing asthmatic response [50], and reducing neutrophil recruitment during influenza [51].
Walker, et al. [52] indicated a direct relationship between occurrences of diarrhoea and subsequent pneumonia (or pneumonia-like instances), with the relative risk of up to 1.08/day in Ghanaian children. Similarly, the relative risk factors such as zinc deficiency increased the relative risk of mortality by 1.2 in both cases. A correlation between these two, with a zinc deficiency factor can be therefore, in combination with other factors, used as a potential biomarker for early diagnosis of protozoan infection. The review work of Halliez and Buret [53] discusses numerous long-term consequences of giardiasis in extra-intestinal aspects. Some of the primary issues observed been caused by the malnutrition, immune response and solute/mineral losses during diarrhoea [54].
5. Nutritional interventions
5.1. Prebiotics, probiotics and synbiotics supplementation
The phenomenon of FGIDs is not an uncommon among the post-infection patients. The long-term impacts of cryptosporidiosis and giardiasis including IBS, cognitive deficiencies, chronic fatigues, and joint pains have been extensively highlighted [53,55]. The Rome Foundation, through its Rome IV working report, has indicated the significance of post-infection IBS (PI-IBS) [56]. The report indicates a 4 - 36% occurrence of PI-IBS among enteritis patients and highlights a complete absence of pharmacologic strategy to treat PI-IBS.
In this context, probiotics have been investigated to address this problem. It has been suggested that in the cases of post infection diarrhea caused by parasites such as Campylobacter, Salmonella, Cryptosporidium, and Giardia, the supplemented probiotics compete with the gut parasites for nutrition and resources, thereby increasing the anti-parasitic immunity within the gut [57]. This in turn aided towards shortening the diarrheal period and reducing its severity. The predominant species used on commercial levels include Lactobacillus, Streptococcus, Enterococcus, Lactococcus, Pediococcus, Bacillus, Escherichia, and sometimes Saccharomyces [58]. The American Gastroenterology Association (AGA), in its 2020 guideline, although indicated that there are knowledge gaps regarding the impact of probiotics on the post-infection conditions, conditionally recommended the use of probiotics during the antibiotic treatment. Although a reduction in diarrheal period by 21.91 – 28.9 hours was seen in paediatric acute infectious gastroenteritis, the experimental outcomes varied, resulting in AGA recommending against the probiotic use during infectious gastroenteritis [59]. It is likely that this may be due to the symptom control and not targeting pathogenic mechanisms, approach of the current treatments [60]. One of the recent studies exploring the IBS caused from cow milk allergy may shed some light into the change of approach. The study, based on brain-gut immunoendocrine microbiota axis, indicated that the use of extensively hydrolysed casein formula along the Lactobacillus rhamnosus GG probiotic helped to decreased FGIDs in children [61]. Similarly, the intake of low fermentable oligosaccharides, disaccharides, monosaccharides, and polyols (FODMAPs) has shown to help alleviate the IBS [62]. This is not surprising since these molecules are not digestible by the human gut, and act as good fibre sources.
Synbiotics and combination therapies have also been studied, and have indicated to have positive impacts on gut health. Complementary synbiotics are the mixture of prebiotic and probiotic components. In the inflammatory bowel disease (IBD) mouse model, recent study by Shinde, et al. [63] showed that synbiotic additive, created through mixing of prebiotic such as green banana resistant starch, and probiotic such as Bacillus coagulans, led to an increase on colonic SCFAs. This phenomenon was not observed with the addition of probiotic only supplementation, but was observed when prebiotic was added, indicating the important role of prebiotics in production of SCFAs. In addition to the gut modulation, synbiotics feeding has also showed to increase the levels of anti-inflammatory and chemopreventive metabolites such as 2-pentanone [64], shown to inhibit prostaglandin and COX-2 protein expressions in colon cancer cells [65]. Furthermore, the synbiotic therapy has shown to alleviate the chemotherapy effects, as recently reviewed by Singh, et al. [66].
In contrast, synergistic synbiotics consist of adding a stimulated microbe, or enhancing the activity of delivered microbe through adding a specific substrate . A good example can be cited through the work of Boger, et al. [67], who utilised short-chain inulin (sc-inulin) as a prebiotic and Lactobacillus paracasei subsp. paracasei W20 as a Step 1 probiotic. The species was able to ferment the sc-inulin which, through a cross-feeding mechanism, enabled an increased fermentation by step 2 probiotic Lactobacillus salivarius W57. Such synergistic synbiotics, although much difficult to obtain, have the ability to deliver significant impacts in selective, targeted manner [68] such as to counter AMR and MDR. In the case of protozoal infection, early study on rats by Ribeiro, et al. [69] showed that supplementation of synbiotic mixture of Bifidobacterium animalis and Raftilose® P95 fructooligosaccharides by its own, and as an addition to dexamethasone treatment.
Metronidazole (MTZ) is the most used drug to address giardiasis. However, emerging studies are showing an increasingly developing Giardia resistance to MTZ [70,71]. In this context, nutritional sources have shown promise in mitigating the parasite removal. For example, the blueberry polyphenolic extract, under in vitro conditions, has shown to inactivate > 90% Giardia trophozoites at 167µg/mL, with respect to a 100% achieved by 67 µg/mL MTZ [72]. The dichloromethane polyphenolic extracts of ginger and cinnamon, particularly at 20 mg dosage rate in albino rats, have shown to reduce Giardia cyst count by 90.1% and 100% respectively, while reducing cyst count by 75.4% and 34.1%, respectively [73]. In mouse model, BIOintestil® (contains gingergrass (or palmarosa) essential oil, ginger powder and gingerol), when combined with MicrobiomeX® (contains citrus extract flavonoids) at 100 mg/day dosage, eliminated 100% of Giardia cysts in Swiss mouse model within five days, and was twice as effective as Albendazole and Metronidazole [74].
Similarly, the mouse model study has indicated that zinc supplementation and Lactobacillus acidophilus + dill seed oil supplementation reduced the Cryptosporidium oocysts by 98.3% and 95.8%, respectively, with respect to prescribed drug, nitazoxanide (91.6%) within eight days of treatment. Furthermore, these treatments were also found to significantly reduce the TNF-α levels in serum [75].
Although supplementation of micronutrients such as zinc have been suggested to control parasitic activity in gut [71], it has been shown that in the infant gut, supplementation of micronutrients such as iron and vitamins (A, C, D, folate), but without zinc, decrease the gut microbiome diversity and aid the growth of protozoal parasites such as Entamoeba [76]. Therefore, it is important to ascertain the impact of certain nutritional interventions.
5.2. Postbiotics and microbiome modulation to improve MDR resilience
Postbiotics, although have been traditionally remained neglected in modern medicine for FGID treatments, have started to gain relevance over last few years. The International Scientific Association for Probiotics and Prebiotics (ISAPP) defines postbiotics as “preparation of inanimate microorganisms and/or their components that confers a health benefit on the host” [77]. They can range from sugar alcohols to amino acids, fatty acids, vitamins, and microbial peptides. A widely known example would be Vitamin K generated by Escherichia coli in gut. Some of the recent additions to the postbiotics include SCFAs, D-amino acids, and small proteins/peptides.
SCFAs and D-amino acids, generated by gut microbes, have been shown to contribute towards the host immune response to infections [63,78,79,80]. In our mouse model study of cryptosporidiosis, the SCFA accumulation elevated in response to the infection. Particularly, significant acetate elevations were seen in duodenum and jejunum, while butyrate levels increased in caecum and colon [19]. However, it appeared that these levels were much higher during giardiasis, particularly the propanoate and butyrate increase in colon [81]. It has been shown that the supplementation of SCFAs such as acetate to the influenza infected mice, through the gut-lung axis, aided to improve the alveolar macrophage activity [82]. On the other hand, propionate production by gut Bacteroides disrupted the intracellular homeostasis, inhibiting pathogenic Salmonella enterica growth [83]. In the aging mice, Lactobacillus acidophilus DDS-1, when added as a probiotic, led to an increase of caecal butyrate levels, leading to downregulation of inflammatory cytokines [84]. Similarly, the D-amino acid levels increased throughout the mouse gut during cryptosporidiosis [19] and giardiasis [81], but their levels were observed to be much higher in small intestine with respect to large intestine. Early assessment has shown that although D-amino acids were unable to prevent the Staphylococcus aureus colonisation, they inhibited the biofilm assembly development under in vitro conditions [85].
In addition to SCFAs and D-amino acids, small proteins such as bacteriocins/colicins, produced by gut Enterobacteriaceae, have shown to competitively inhibit the growth of pathogenic Salmonella enterica [86]. One of the very recent reviews by Upatissa and Mitchell [87] has indicated the utilisation of these small proteins to control specific drug resistant pathogens. For example, microcin J25 has shown to inhibit more than 28 multi-antibiotic resistant Salmonella enterica serovars [88]. The protein also has shown effectiveness against some strains of multi-drug resistant E. coli [89]. The work of Upatissa and Mitchell [87] provides a good insight into these proteins and their action mechanism. In the case of cryptosporidiosis, cathelicidin related antimicrobial peptide (CRAMP) has been indicated to significantly reduce the parasite burden. However, the indigenous CRAMP appeared to be downregulated during cryptosporidiosis. In such a case, oral feeding of 5 µg CRAMP has shown to aid the reduction of Cryptosporidium sporozoites, but not the oocyst [90]. Antimicrobial peptides, particularly from venomous insects such as bees, have shown inhibitory effects on protozoal parasites[91,92] and promise to be applied in protozoal infection treatment.
One of the components that may arguably be categorised as both prebiotics and postbiotics are enteric viruses, especially macrophages. In addition to chronic disorders such as colitis [93], they have been proposed to be effective treatment for AMR and MDR pathogens [94,95,96]. For example, the phages such as Bϕ-B1251 and PD-6A3 have shown to provide lytic activity against MDR resistant Acinetobacter baumannii [97], E. coli and methicillin-resistant Staphylococcus aureus [98], respectively. However, due to their extreme specificity, more studies need to be undertaken to ascertain the impacts of bacteriophage treatment.
In addition, recent reviews of [99] has also covered various new and emerging techniques of improving gut microbiome resilience, and microbiome resurrection post infection and in various other gut and extra-gut inflammations.
6. Application of multiomics in high-throughput analysis of gut microbiome health and inter-organ axes
6.1. Multiomics approaches
Numerous analytical techniques have been used to elaborate the workings of this multi-level complex relationship. Although, the technical specifications of individual omics platforms are beyond the scope of this work, mentioning these platforms is important in the context of analytical assessment of abovementioned systems biology approaches. However, Pinu, et al. [100] provide an excellent coverage of multi-omics integration, including experimental design, data integration and analysis, systems modelling, and challenges.
In metagenomics, the emerging sequencing methods, combined with the robust databases [101], provide a good understanding of microbiome identification and characterisation. The compiled work of Nagarajan [102] has provided further information. Metaproteomics has also been significantly developed over last few years, with its own databases and highly sensitive, high-throughput mass spectrometry-based analytical tools. Detailed information of these instruments, their relevant sample preparation, work-flows, data quantification and synchronisation have been reviewed by Antoine and Bruno [103] and Heyer, et al. [104]. Metabolomics, including lipidomics, glycomics and ionomics, have the potential to provide biochemical information to understand and characterise the various mechanisms. Metabolomics have been applied to investigate bacterial processes related to preventative health [105,106], environmental pollution [107,108] and, food [109] among others. Metabolomics also assist in extricating the correlation between cell phenotypes and their metabolic patterns and stoichiometry [110]. Metabolic flux studies have previously been applied in toxicology and medicine [111,112,113] and, microbial respiratory systems [114,115]. For further information, the reader is suggested to refer to the works of Beale, et al. [116].
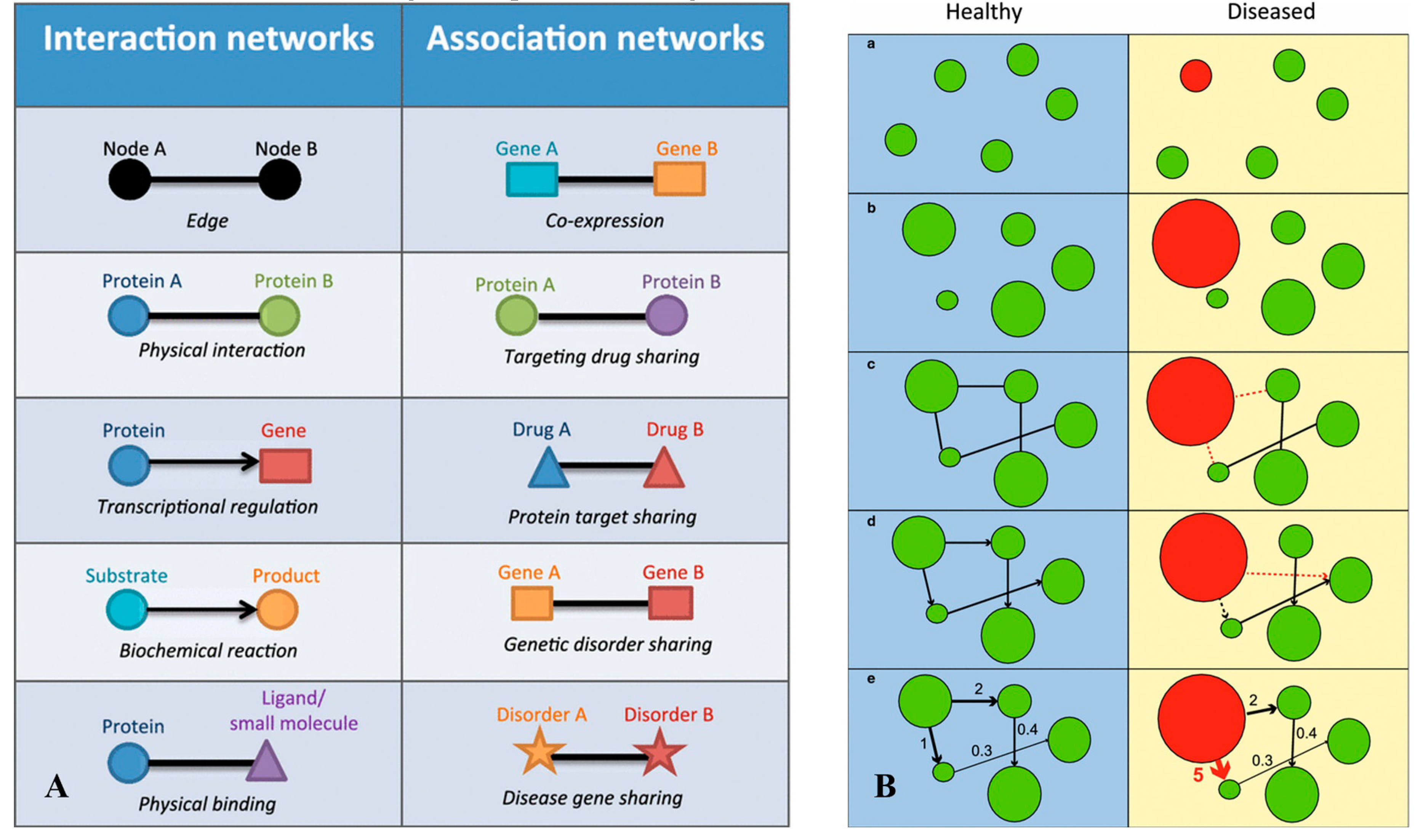
In case of enteric infection, a highly complex host-parasite-host microbiome relationship is developed. Resultantly, a significant change in genomic, transcriptomic, proteomic, and metabolic expression is observed, contributing to expressional pool. This variation causes considerable perturbations at various regulation levels such as gene expression (major) and proteomic translation (considerable, but less than gene expression) (Figure 4).
Multiomics utilises the capability of two or more omic platforms, either in standalone or an integrated manner. Since the individual omic platforms do not provide a big picture of system biology, an integrated approach is increasingly utilised. In the context of gut infections, our recent works have integrated metabolomics with gut metaproteomics and 16S rRNA genomics to understand the interactomics of cryptosporidiosis [19] and giardiasis [27]. Particularly in the giardiasis interactomics study [27], we integrated the 16S rRNA population genetics-GC-MS metabolomics data with LC-MS metaproteomics-GC-MS metabolomics via PICRUSt [118], BURRITO [119], and MetaboAnalyst [120] toolboxes. The integration not only indicated the key microbiome species impacted but also helped to filter the most significant pathways redox pathways, impacting gut-liver axis. Similar strategies have been further employed to understand the extra-gut mechanism of FGIDs such as irritable bowel syndrome (IBS), functional dyspepsia [121], and infections such as SARS-CoV2 [122,123]. In the area of nutritional impacts of prebiotics and probiotics, the study reported by Shinde, et al. [63] included the combined approach of immunohistochemistry, enzyme kinetics, and metabolomics including SCFA analytics. The study showed that the probiotic and prebiotic combinations provide synergistic immune-regulating efficacy and protect epithelial integrity and mediate the reduction in colonic inflammation. The 16S rRNA population genetics data was added to this approach to determine the modulation of key gut microbial species during these treatment regimens to improve the gut health in mouse model [78,84]. The on-field study done by Attia, et al. [124] utilised a less sensitive, but more impactful multiomics approach for determining the impacts of severe acute malnutrition (SAM), contributing to mortality on paediatric patients (age: 6 – 60 months). The study undertook targeted genomic identification, protein, and immune assays, combined with GC-MS-based SCFA analysis. The results indicated a considerable presence of Shigella, Giardia, and Campylobacter, with upregulated calprotectin and depleted butyrate propionate in fatal cases. A more recent omics study [125] indicated that in the case of SAM, the pre-treated paediatric patients had higher number of Proteobacteria, particularly Enterobacteriaceae. Furthermore, they had lower gut microbiome diversity and depleted SCFAs. When fed with high cowpea flour combined with WHO standard feed F75 and F100, impacts of antibiotics decreased, and improved gut integrity. The study indicated that post-antibiotic cessation, children fed with this diet showed increased Firmicutes, correlating to increased SCFA by 28th day after intervention.
6.2. Application of Artificial intelligence and machine learning (AIML) and future aspects
AIML has been making strong inroads in the system biology. In addition to the multivariate statistics, it provides a strong potential for understanding the key biomarkers or pathways involved in the gut processes. A recent study by [126] applied machine learning (ML) to elaborate the metabolome and lipidome associations of glioblastoma patients through graphical network analysis. The ML analysis indicated homogenous networks with lipid cluster linkages between the key metabolic markers. On the other hand, among the patients with unfavourable outcomes, fewer cluster networks were seen, with altered key lipids. Another study assessed the host-pathogen interaction, and its effects on gut microbiota through machine learning [127] . The study involved a minimum curvilinear Markov clustering (MC-MCL) method to analyse mechanisms of bacterial network re-organizations caused by proton pump inhibitor (PPI) intervention, and Helicobacter pylori infection in stomach. MC-MCL indicated that nine bacterial species from Fusobacteria, Proteobacteria, Bacteroides and Firmicutes positively correlated with the PPI treatment. Similarly, six species showed a depletion during the H. pylori infection. Another approach for diagnosing several neglected tropical diseases was reported by [128]. The study used an ensemble algorithm to cross validate antibody response from variable populations to predict pathogenic surveillance. These approaches, as reported previously by CoviRx database for COVID-19 [129] in addition to the multiomics, has a potential to provide a much rapid and reliable outcomes of nutritional solutions to the protozoal MDR infection agents. The recent reviews [130,131,132] highlight the impact of integrating multiomics with AIML to further develop the prebiotic, probiotics, and symbiotic landscape, which the readers can refer to for more information.
Multiomics, combined with AIML have started to show promising results to understand the mechanism of these interventions and their impacts. While multiomics has shown immense potential to get more understanding of infection interactomics and impacts of nutritional interventions, AIML still remains in its infancy. A combination of portable multi-omics equipment, cloud computing, and AIML would expedite our capabilities to address the research and development gaps in this area over the upcoming decade.
Author Contributions
All authors contributed equally to the manuscript.
Funding
Not applicable.
Data Availability Statement
Not applicable
Acknowledgments
The authors would like to thank Dr Crispin Howitt and Dr Xue-Rong Zhou from CSIRO Future Protein Mission and CSIRO Agriculture and Food, for their review and support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Petri, W.A.; Miller, M.; Binder, H.J.; Levine, M.M.; Dillingham, R.; Guerrant, R.L. Enteric infections, diarrhea, and their impact on function and development. J. Clin. Investig. 2008, 118, 1277–1290. [Google Scholar] [CrossRef] [PubMed]
- WHO. Global Health Observatory Data Repository: Diarrhoeal Diseases. Available online: http://apps.who.int/gho/data/view.main.CM1002015WORLD-CH3?lang=enhttp://www.who.int/mediacentre/factsheets/fs310/en/ (accessed on 17 January).
- Efstratiou, A.; Ongerth, J.E.; Karanis, P. Waterborne transmission of protozoan parasites: Review of worldwide outbreaks - An update 2011–2016. Water Res. 2017, 114, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Berger, S. Amoebiasis: Global Status; GIDEON Informatics Inc: Los Angeles, CA, UNITED STATES, 2017. [Google Scholar]
- Berger, S. Giardiasis: Global Status; GIDEON Informatics Inc: Los Angeles, CA, UNITED STATES, 2017. [Google Scholar]
- Berger, S. Cryptosporidiosis: Global Status; GIDEON Informatics Inc: Los Angeles, CA, UNITED STATES, 2017. [Google Scholar]
- Betancourt, W.Q.; Rose, J.B. Drinking water treatment processes for removal of Cryptosporidium and Giardia. Vet. Parasitol. 2004, 126, 219–234. [Google Scholar] [CrossRef]
- Jarroll, E.L.; Hoff, J.C. Effect of disinfectants on Giardia cysts. Crit. Rev. Environ. Sci. Technol. 1988, 18, 1–28. [Google Scholar]
- Rubin, A.J.; Engel, J.P.; Sproul, O.J. Disinfection of amoebic cysts in water with free chlorine. J. (Water Pollut. Control Fed. ) 1983, 55, 1174–1182. [Google Scholar]
- Chalmers, R.M. Chapter Sixteen - Cryptosporidium. In Microbiology of Waterborne Diseases (Second Edition); Academic Press: London, 2014; pp. 287–326. [Google Scholar]
- Lane, S.; Lloyd, D. Current trends in research into the waterborne parasite Giardia. Crit. Rev. Microbiol. 2002, 28, 123–147. [Google Scholar] [CrossRef]
- Chalmers, R.M. Chapter Eighteen - Entamoeba histolytica. In Microbiology of Waterborne Diseases (Second Edition); Academic Press: London, UK, 2014; pp. 355–373. [Google Scholar]
- Duncan, H.E.; Edberg, S.C. Host-Microbe Interaction in the Gastrointestinal Tract. Crit. Rev. Microbiol. 1995, 21, 85–100. [Google Scholar] [CrossRef]
- Macfarlane, S.; Dillon, J.F. Microbial biofilms in the human gastrointestinal tract. J. Appl. Microbiol. 2007, 102, 1187–1196. [Google Scholar] [CrossRef]
- Swidsinski, A.; Ung, V.; Sydora, B.C.; Loening-Baucke, V.; Doerffel, Y.; Verstraelen, H.; Fedorak, R.N. Bacterial overgrowth and inflammation of small intestine after carboxymethylcellulose ingestion in genetically susceptible mice. Inflamm. Bowel Dis. 2009, 15, 359–364. [Google Scholar] [CrossRef]
- Koh, W.; Clode, P.L.; Monis, P.; Thompson, R.A. Multiplication of the waterborne pathogen Cryptosporidium parvum in an aquatic biofilm system. Parasites Vectors 2013, 6, 270. [Google Scholar] [CrossRef]
- McKenney, E.A.; Greene, L.K.; Drea, C.M.; Yoder, A.D. Down for the count: Cryptosporidium infection depletes the gut microbiome in Coquerel’s sifakas. Microb. Ecol. Health Dis. 2017, 28, 1335165. [Google Scholar] [CrossRef] [PubMed]
- Certad, G.; Viscogliosi, E.; Chabé, M.; Cacciò, S.M. Pathogenic Mechanisms of Cryptosporidium and Giardia. Trends Parasitol. 2017, 33, 561–576. [Google Scholar] [CrossRef] [PubMed]
- Karpe, A.V.; Hutton, M.L.; Mileto, S.J.; James, M.L.; Evans, C.; Shah, R.M.; Ghodke, A.B.; Hillyer, K.E.; Metcalfe, S.S.; Liu, J.-W.; et al. Cryptosporidiosis Modulates the Gut Microbiome and Metabolism in a Murine Infection Model. Metabolites 2021, 11, 380. [Google Scholar] [CrossRef] [PubMed]
- Berrilli, F.; Di Cave, D.; Cavallero, S.; D'Amelio, S. Interactions between parasites and microbial communities in the human gut. Front. Cell. Infect. Microbiol. 2012, 2. [Google Scholar] [CrossRef]
- Dwyer, D.M.; Chang, K.P. Surface membrane carbohydrate alterations of a flagellated protozoan mediated by bacterial endosymbiotes. Proc. Natl. Acad. Sci. United States Am. 1976, 73, 852–856. [Google Scholar] [CrossRef]
- Torres, M.F.; Uetanabaro, A.P.T.; Costa, A.F.; Alves, C.A.; Farias, L.M.; Bambirra, E.A.; Penna, F.J.; Vieira, E.C.; Nicoli, J.R. Influence of bacteria from the duodenal microbiota of patients with symptomatic giardiasis on the pathogenicity of Giardia duodenalis in gnotoxenic mice. J. Med. Microbiol. 2000, 49, 209–215. [Google Scholar] [CrossRef]
- Mirelman, D.; Feingold, C.; Wexler, A.; Bracha, R. Interactions between Entamoeba histolytica, bacteria and intestinal cells. In Cytopathology of Parasitic Disease. Pitman Books: London, UK, 1983; Volume 99, pp. 2–30. [Google Scholar]
- Galván-Moroyoqui, J.M.; Del Carmen Dominguez-Robles, M.; Franco, E.; Meza, I. The interplay between Entamoeba and enteropathogenic bacteria modulates epithelial cell damage. PLoS Neglected Trop. Dis. 2008, 2, e266. [Google Scholar] [CrossRef]
- Gerbaba, T.K.; Gupta, P.; Rioux, K.; Hansen, D.; Buret, A.G. Giardia duodenalis-induced alterations of commensal bacteria kill Caenorhabditis elegans: a new model to study microbial-microbial interactions in the gut. Am. J. Physiol. -Gastrointest. Liver Physiol. 2015, 308, G550–G561. [Google Scholar] [CrossRef]
- Wallace, J.L.; Blackler, R.W.; Chan, M.V.; Da Silva, G.J.; Elsheikh, W.; Flannigan, K.L.; Gamaniek, I.; Manko, A.; Wang, L.; Motta, J.-P. Anti-inflammatory and cytoprotective actions of hydrogen sulfide: Translation to therapeutics. Antioxid. Redox Signal. 2015, 22, 398–410. [Google Scholar] [CrossRef]
- Karpe, A.V.; Hutton, M.L.; Mileto, S.J.; James, M.L.; Evans, C.; Ghodke, A.B.; Shah, R.M.; Metcalfe, S.S.; Liu, J.-W.; Walsh, T.; et al. Gut microbial perturbation and host response induce redox pathway upregulation along the Gut-Liver axis during giardiasis in C57BL/6J mouse model. Int. J. Mol. Sci. 2023, 24, 1636. [Google Scholar] [CrossRef]
- Kedia, S.; Rampal, R.; Paul, J.; Ahuja, V. Gut microbiome diversity in acute infective and chronic inflammatory gastrointestinal diseases in North India. J. Gastroenterol. 2016, 51, 660–671. [Google Scholar] [CrossRef]
- Partida-Rodríguez, O.; Serrano-Vázquez, A.; Nieves-Ramírez, M.E.; Moran, P.; Rojas, L.; Portillo, T.; González, E.; Hernández, E.; Finlay, B.B.; Ximenez, C. Human intestinal microbiota: Interaction between parasites and the host immune response. Arch. Med. Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Zermeño, V.; Ximénez, C.; Morán, P.; Valadez, A.; Valenzuela, O.; Rascón, E.; Diaz, D.; Cerritos, R. Worldwide genealogy of Entamoeba histolytica: An overview to understand haplotype distribution and infection outcome. Infect. Genet. Evol. 2013, 17, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Silva, M.A.; Santos, H.L.C.; Peralta, R.S.; Peralta, J.M.; de Macedo, H.W. Experimental amoebic liver abscess in hamsters caused by trophozoites of a Brazilian strain of Entamoeba dispar. Exp. Parasitol. 2013, 134, 39–47. [Google Scholar] [CrossRef]
- Dolabella, S.S.; Serrano-Luna, J.; Navarro-García, F.; Cerritos, R.; Ximénez, C.; Galván-Moroyoqui, J.M.; Silva, E.F.; Tsutsumi, V.; Shibayama, M. A mo ebic liver abscess production by Entamoeba dispar. parasite 2012, 13, 15. [Google Scholar]
- Weber, C.; Koutero, M.; Dillies, M.-A.; Varet, H.; Lopez-Camarillo, C.; Coppée, J.Y.; Hon, C.-C.; Guillén, N. Extensive transcriptome analysis correlates the plasticity of Entamoeba histolytica pathogenesis to rapid phenotype changes depending on the environment. Sci. Rep. 2016, 6, 35852. [Google Scholar] [CrossRef]
- Rigothier, M.-C.; Khun, H.; Tavares, P.; Cardona, A.; Huerre, M.; Guillén, N. Fate of Entamoeba histolytica during establishment of amoebic liver abscess analyzed by quantitative radioimaging and histology. Infect. Immun. 2002, 70, 3208–3215. [Google Scholar] [CrossRef]
- Stanley, S.L. Pathophysiology of amoebiasis. Trends Parasitol. 2001, 17, 280–285. [Google Scholar] [CrossRef]
- Tsutsumi, V.; Mena-Lopez, R.; Anaya-Velazquez, F.; Martinez-Palomo, A. Cellular bases of experimental amebic liver abscess formation. Am. J. Pathol. 1984, 117, 81. [Google Scholar]
- Ungar, B.L.; Burris, J.A.; Quinn, C.A.; Finkelman, F.D. New mouse models for chronic Cryptosporidium infection in immunodeficient hosts. Infect. Immun. 1990, 58, 961–969. [Google Scholar] [CrossRef]
- Astiazaran-Garcia, H.; Lopez-Teros, V.; Valencia, M.E.; Vazquez-Ortiz, F.; Sotelo-Cruz, N.; Quihui-Cota, L. Giardia lamblia infection and its implications for vitamin a liver stores in school children. Ann. Nutr. Metab. 2010, 57, 228–233. [Google Scholar] [CrossRef]
- Swann, J.; Jamshidi, N.; Lewis, N.E.; Winzeler, E.A. Systems analysis of host–parasite interactions. Wiley Interdiscip. Rev. : Syst. Biol. Med. 2015, 7, 381–400. [Google Scholar] [CrossRef]
- Zakaria, A.; Al-Share, B.; Al Asad, K. Primary Pulmonary Amebiasis Complicated with Multicystic Empyema. Case Rep. Pulmonol. 2016, 2016, 4. [Google Scholar] [CrossRef]
- Goh, L.M.L.; Marrone, J.R. Entamoeba histolytica meningoencephalitis diagnosed by trophozoites in cerebrospinal fluid. New Microbes New Infect. 2013, 1, 16–17. [Google Scholar] [CrossRef]
- Reina, F.T.R.; Ribeiro, C.A.; Araújo, R.S.D.; Matté, M.H.; Castanho, R.E.P.; Tanaka, I.I.; Viggiani, A.M.F.S.; Martins, L.P.A. INTESTINAL AND PULMONARY INFECTION BY Cryptosporidium parvum IN TWO PATIENTS WITH HIV/AIDS. Rev. Do Inst. De Med. Trop. De São Paulo 2016, 58. [Google Scholar] [CrossRef]
- López-Vélez, R.; Tarazona, R.; Camacho, A.G.; Gomez-Mampaso, E.; Guerrero, A.; Moreira, V.; Villanueva, R. Intestinal and extraintestinal cryptosporidiosis in AIDS patients. Eur. J. Clin. Microbiol. Infect. Dis. 1995, 14, 677–681. [Google Scholar] [CrossRef]
- Clavel, A.; Arnal, A.C.; Sánchez, E.C.; Castillo, F.J.; Varea, M.; Gómez-Lus, R.; Cuesta, J.; Letona, S.; Amiguet, J.A. Respiratory cryptosporidiosis: Case series and review of the literature. Infection 1996, 24, 341–346. [Google Scholar] [CrossRef]
- Burgess, S.L.; Lu, M.; Ma, J.Z.; Naylor, C.; Donowitz, J.R.; Kirkpatrick, B.D.; Haque, R.; Petri, W.A., Jr. Inflammatory markers predict episodes of wheezing during the first year of life in Bangladesh. Respir. Med. 2015, 110, 53–57. [Google Scholar] [CrossRef]
- Braamskamp, M.J.A.M.; Dolman, K.M.; Tabbers, M.M. Clinical practice. Eur. J. Pediatr. 2010, 169, 1179–1185. [Google Scholar] [CrossRef]
- Wypych, T.P.; Wickramasinghe, L.C.; Marsland, B.J. The influence of the microbiome on respiratory health. Nat. Immunol. 2019, 20, 1279–1290. [Google Scholar] [CrossRef]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Cait, A.; Hughes, M.R.; Antignano, F.; Cait, J.; Dimitriu, P.A.; Maas, K.R.; Reynolds, L.A.; Hacker, L.; Mohr, J.; Finlay, B.B.; et al. Microbiome-driven allergic lung inflammation is ameliorated by short-chain fatty acids. Mucosal Immunol. 2018, 11, 785–795. [Google Scholar] [CrossRef] [PubMed]
- Thorburn, A.N.; McKenzie, C.I.; Shen, S.; Stanley, D.; Macia, L.; Mason, L.J.; Roberts, L.K.; Wong, C.H.Y.; Shim, R.; Robert, R.; et al. Evidence that asthma is a developmental origin disease influenced by maternal diet and bacterial metabolites. Nat. Commun. 2015, 6, 7320. [Google Scholar] [CrossRef] [PubMed]
- Trompette, A.; Gollwitzer, E.S.; Pattaroni, C.; Lopez-Mejia, I.C.; Riva, E.; Pernot, J.; Ubags, N.; Fajas, L.; Nicod, L.P.; Marsland, B.J. Dietary Fiber Confers Protection against Flu by Shaping Ly6c<sup>−</sup> Patrolling Monocyte Hematopoiesis and CD8<sup>+</sup> T Cell Metabolism. Immunity 2018, 48, 992–1005.e1008. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.L.F.; Rudan, I.; Liu, L.; Nair, H.; Theodoratou, E.; Bhutta, Z.A.; O'Brien, K.L.; Campbell, H.; Black, R.E. Global burden of childhood pneumonia and diarrhoea. Lancet 2013, 381, 1405–1416. [Google Scholar] [CrossRef]
- Halliez, M.C.M.; Buret, A.G. Extra-intestinal and long term consequences of Giardia duodenalis infections. World J. Gastroenterol. : WJG 2013, 19, 8974–8985. [Google Scholar] [CrossRef]
- Buret, A.G.; Reti, K. Acute enteric infections alter commensal microbiota: new mechanisms in post-infectious intestinal inflammatory disorders. In Proceedings of the Old Herborn University Seminar Monograph: Persisting Consequences of Intestinal Infection; 2014; pp. 87–100. [Google Scholar]
- Hanevik, K. Long-Term Consequences of Cryptosporidium and Giardia Gastroenteritis. Curr. Trop. Med. Rep. 2016, 3, 89–93. [Google Scholar] [CrossRef]
- Barbara, G.; Grover, M.; Bercik, P.; Corsetti, M.; Ghoshal, U.C.; Ohman, L.; Rajilić-Stojanović, M. Rome Foundation Working Team Report on Post-Infection Irritable Bowel Syndrome. Gastroenterology 2019, 156, 46–58.e47. [Google Scholar] [CrossRef]
- Allen, S.J.; Martinez, E.G.; Gregorio, G.V.; Dans, L.F. Probiotics for treating acute infectious diarrhoea. Cochrane Database Syst. Rev. 2010. [Google Scholar] [CrossRef]
- Depoorter, L.; Vandenplas, Y. Chapter 21 - Probiotics in pediatrics. In Probiotics; Brandelli, A., Ed.; Academic Press: London, UK, 2022; pp. 425–450. [Google Scholar]
- Su, G.L.; Ko, C.W.; Bercik, P.; Falck-Ytter, Y.; Sultan, S.; Weizman, A.V.; Morgan, R.L. AGA Clinical Practice Guidelines on the Role of Probiotics in the Management of Gastrointestinal Disorders. Gastroenterology 2020, 159, 697–705. [Google Scholar] [CrossRef]
- Di Nardo, G.; Cremon, C.; Staiano, A.; Stanghellini, V.; Borrelli, O.; Strisciuglio, C.; Romano, C.; Mallardo, S.; Scarpato, E.; Marasco, G.; et al. Role of inflammation in pediatric irritable bowel syndrome. Neurogastroenterol. Motil. 2023, 35, e14365. [Google Scholar] [CrossRef]
- Nocerino, R.; Di Costanzo, M.; Bedogni, G.; Cosenza, L.; Maddalena, Y.; Di Scala, C.; Della Gatta, G.; Carucci, L.; Voto, L.; Coppola, S.; et al. Dietary Treatment with Extensively Hydrolyzed Casein Formula Containing the Probiotic Lactobacillus rhamnosus GG Prevents the Occurrence of Functional Gastrointestinal Disorders in Children with Cow's Milk Allergy. J Pediatr 2019, 213, 137–142.e132. [Google Scholar] [CrossRef]
- Ford, A.C.; Lacy, B.E.; Talley, N.J. Irritable Bowel Syndrome. N Engl J Med 2017, 376, 2566–2578. [Google Scholar] [CrossRef]
- Shinde, T.; Perera, A.P.; Vemuri, R.; Gondalia, S.V.; Beale, D.J.; Karpe, A.V.; Shastri, S.; Basheer, W.; Southam, B.; Eri, R. Synbiotic supplementation with prebiotic green banana resistant starch and probiotic Bacillus coagulans spores ameliorates gut inflammation in mouse model of inflammatory bowel diseases. Eur. J. Nutr. 2020, 59, 3669–3689. [Google Scholar] [CrossRef]
- Vitali, B.; Ndagijimana, M.; Cruciani, F.; Carnevali, P.; Candela, M.; Guerzoni, M.E.; Brigidi, P. Impact of a synbiotic food on the gut microbial ecology and metabolic profiles. BMC Microbiol. 2010, 10, 4. [Google Scholar] [CrossRef]
- Pettersson, J.; Karlsson, P.C.; ouml; ransson, U.; Rafter, J.J.; Bohlin, L. The Flavouring Phytochemical 2-Pentanone Reduces Prostaglandin Production and COX-2 Expression in Colon Cancer Cells. Biol. Pharm. Bull. 2008, 31, 534–537. [Google Scholar] [CrossRef]
- Singh, N.K.; Beckett, J.M.; Kalpurath, K.; Ishaq, M.; Ahmad, T.; Eri, R.D. Synbiotics as Supplemental Therapy for the Alleviation of Chemotherapy-Associated Symptoms in Patients with Solid Tumours. Nutrients 2023, 15. [Google Scholar] [CrossRef]
- Boger, M.C.L.; Bueren, A.L.v.; Dijkhuizen, L. Cross-Feeding among Probiotic Bacterial Strains on Prebiotic Inulin Involves the Extracellular <i>exo</i>-Inulinase of Lactobacillus paracasei Strain W20. Appl. Environ. Microbiol. 2018, 84, e01539–01518. [Google Scholar] [CrossRef]
- Gomez Quintero, D.F.; Kok, C.R.; Hutkins, R. The Future of Synbiotics: Rational Formulation and Design. Front. Microbiol. 2022, 13. [Google Scholar] [CrossRef]
- Ribeiro, C.M.; Costa, V.M.; Gomes, M.I.F.V.; Golim, M.A.; Modolo, J.R.; Langoni, H. Effects of synbiotic-based Bifidobacterium animalis in female rats experimentally infected with Toxoplasma gondii. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 111–114. [Google Scholar] [CrossRef]
- Emery, S.J.; Baker, L.; Ansell, B.R.E.; Mirzaei, M.; Haynes, P.A.; McConville, M.J.; Svärd, S.G.; Jex, A.R. Differential protein expression and post-translational modifications in metronidazole-resistant Giardia duodenalis. GigaScience 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, S. Herbal, Nutritional, and Traditional Remedies for Giardiasis. In Neglected Tropical Diseases and Phytochemicals in Drug Discovery; 2021; pp. 135–169. [Google Scholar]
- Anthony, J.P.; Fyfe, L.; Stewart, D.; McDougall, G.J.; Smith, H.V. The effect of blueberry extracts on Giardia duodenalis viability and spontaneous excystation of Cryptosporidium parvum oocysts, in vitro. Methods 2007, 42, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.; Attia, R.; Said, S.; Ibraheim, Z. Ginger and cinnamon: can this household remedy treat giardiasis? Parasitological and histopathological studies. Iran J Parasitol 2014, 9, 530–540. [Google Scholar]
- de Almeida, C.R.; Bezagio, R.C.; Colli, C.M.; Romera, L.I.L.; Ferrari, A.; Gomes, M.L. Elimination of Giardia duodenalis BIV in vivo using natural extracts in microbiome and dietary supplements. Parasitol. Int. 2022, 86, 102484. [Google Scholar] [CrossRef]
- Gaber, M.; Galal, L.A.A.; Farrag, H.M.M.; Badary, D.M.; Alkhalil, S.S.; Elossily, N. The Effects of Commercially Available Syzygium aromaticum, Anethum graveolens, Lactobacillus acidophilus LB, and Zinc as Alternatives Therapy in Experimental Mice Challenged with Cryptosporidium parvum. Infect. Drug Resist. 2022, 15, 171–182. [Google Scholar] [CrossRef]
- Popovic, A.; Bourdon, C.; Wang, P.W.; Guttman, D.S.; Soofi, S.; Bhutta, Z.A.; Bandsma, R.H.J.; Parkinson, J.; Pell, L.G. Micronutrient supplements can promote disruptive protozoan and fungal communities in the developing infant gut. Nat. Commun. 2021, 12, 6729. [Google Scholar] [CrossRef]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Shinde, T.; Vemuri, R.; Shastri, S.; Perera, A.P.; Gondalia, S.V.; Beale, D.J.; Karpe, A.V.; Eri, R.; Stanley, R. Modulating the microbiome and immune responses using whole plant fibre in synbiotic combination with fibre-digesting probiotic attenuates chronic colonic inflammation in spontaneous colitic mice model of IBD. Nutrients 2020, 12, 2380. [Google Scholar] [CrossRef]
- Kobayashi, J. D-amino acids and lactic acid bacteria. Microorganisms 2019, 7, 690. [Google Scholar] [CrossRef]
- Sasabe, J.; Miyoshi, Y.; Rakoff-Nahoum, S.; Zhang, T.; Mita, M.; Davis, B.M.; Hamase, K.; Waldor, M.K. Interplay between microbial d-amino acids and host d-amino acid oxidase modifies murine mucosal defence and gut microbiota. Nat. Microbiol. 2016, 1, 1–7. [Google Scholar] [CrossRef]
- Karpe, A.V.; Hutton, M.L.; Mileto, S.J.; James, M.L.; Evans, C.; Ghodke, A.B.; Shah, R.M.; Metcalfe, S.S.; Liu, J.-W.; Walsh, T.; et al. Gut Microbial Perturbation and Host Response Induce Redox Pathway Upregulation along the Gut–Liver Axis during Giardiasis in C57BL/6J Mouse Model. Int. J. Mol. Sci. 2023, 24, 1636. [Google Scholar] [PubMed]
- Sencio, V.; Barthelemy, A.; Tavares, L.P.; Machado, M.G.; Soulard, D.; Cuinat, C.; Queiroz-Junior, C.M.; Noordine, M.-L.; Salomé-Desnoulez, S.; Deryuter, L.; et al. Gut Dysbiosis during Influenza Contributes to Pulmonary Pneumococcal Superinfection through Altered Short-Chain Fatty Acid Production. Cell Rep. 2020, 30, 2934–2947.e2936. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, A.; Lam, L.; Rajendram, M.; Tamburini, F.; Honeycutt, J.; Pham, T.; Van Treuren, W.; Pruss, K.; Stabler, S.R.; Lugo, K.; et al. A Gut Commensal-Produced Metabolite Mediates Colonization Resistance to Salmonella Infection. Cell Host Microbe 2018, 24, 296–307.e297. [Google Scholar] [CrossRef]
- Vemuri, R.; Gundamaraju, R.; Shinde, T.; Perera, A.P.; Basheer, W.; Southam, B.; Gondalia, S.V.; Karpe, A.V.; Beale, D.J.; Tristram, S.; et al. Lactobacillus acidophilus DDS-1 Modulates Intestinal-Specific Microbiota, Short-Chain Fatty Acid and Immunological Profiles in Aging Mice. Nutrients 2019, 11, 1297. [Google Scholar] [CrossRef] [PubMed]
- Hochbaum, A.I.; Kolodkin-Gal, I.; Foulston, L.; Kolter, R.; Aizenberg, J.; Losick, R. Inhibitory effects of D-amino acids on Staphylococcus aureus biofilm development. J Bacteriol 2011, 193, 5616–5622. [Google Scholar] [CrossRef]
- Sassone-Corsi, M.; Nuccio, S.-P.; Liu, H.; Hernandez, D.; Vu, C.T.; Takahashi, A.A.; Edwards, R.A.; Raffatellu, M. Microcins mediate competition among Enterobacteriaceae in the inflamed gut. Nature 2016, 540, 280–283. [Google Scholar] [CrossRef]
- Upatissa, S.; Mitchell, R.J. The “Cins” of Our Fathers: Rejuvenated Interest in Colicins to Combat Drug Resistance. J. Microbiol. 2023, 61, 145–158. [Google Scholar] [CrossRef]
- Ben Said, L.; Emond-Rheault, J.-G.; Soltani, S.; Telhig, S.; Zirah, S.; Rebuffat, S.; Diarra, M.S.; Goodridge, L.; Levesque, R.C.; Fliss, I. Phenomic and genomic approaches to studying the inhibition of multiresistant Salmonella enterica by microcin J25. Environ. Microbiol. 2020, 22, 2907–2920. [Google Scholar] [CrossRef]
- Martin-Gómez, H.; Jorba, M.; Albericio, F.; Viñas, M.; Tulla-Puche, J. Chemical Modification of Microcin J25 Reveals New Insights on the Stereospecific Requirements for Antimicrobial Activity. Int. J. Mol. Sci. 2019, 20, 5152. [Google Scholar] [CrossRef] [PubMed]
- Guesdon, W.; Pezier, T.; Menard, S.; Nicolosi, A.; Le Vern, Y.; Silvestre, A.; Diana, J.; Laurent, F.; Lacroix-Lamandé, S. Cryptosporidium parvum Subverts Antimicrobial Activity of CRAMP by Reducing Its Expression in Neonatal Mice. Microorganisms 2020, 8, 1635. [Google Scholar] [CrossRef] [PubMed]
- Sabiá Júnior, E.F.; Menezes, L.F.S.; de Araújo, I.F.S.; Schwartz, E.F. Natural Occurrence in Venomous Arthropods of Antimicrobial Peptides Active against Protozoan Parasites. Toxins 2019, 11, 563. [Google Scholar] [CrossRef] [PubMed]
- Mahdavi Abhari, F.; Pirestani, M.; Dalimi, A. Anti-amoebic activity of a cecropin-melittin hybrid peptide (CM11) against trophozoites of Entamoeba histolytica. Wien. Klin. Wochenschr. 2019, 131, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-Y.; Kim, M.-S.; Kim, E.; Cheon, J.H.; Lee, Y.-S.; Kim, Y.; Lee, S.-H.; Seo, S.-U.; Shin, S.-H.; Choi, Sun S.; et al. Enteric Viruses Ameliorate Gut Inflammation via Toll-like Receptor 3 and Toll-like Receptor 7-Mediated Interferon-β Production. Immunity 2016, 44, 889–900. [Google Scholar] [CrossRef]
- Tsigalou, C.; Konstantinidis, T.; Stavropoulou, E.; Bezirtzoglou, E.E.; Tsakris, A. Potential elimination of human gut resistome by exploiting the benefits of functional foods. Front. Microbiol. 2020, 11, 50. [Google Scholar] [CrossRef]
- Nath, A.; Bhattacharjee, R.; Nandi, A.; Sinha, A.; Kar, S.; Manoharan, N.; Mitra, S.; Mojumdar, A.; Panda, P.K.; Patro, S.; et al. Phage delivered CRISPR-Cas system to combat multidrug-resistant pathogens in gut microbiome. Biomed. Pharmacother. 2022, 151, 113122. [Google Scholar] [CrossRef]
- Zhang, Y.; Lin, Y.; Galgano, S.; Houdijk, J.; Xie, W.; Jin, Y.; Lin, J.; Song, W.; Fu, Y.; Li, X.; et al. Recent Progress in Phage Therapy to Modulate Multidrug-Resistant Acinetobacter baumannii, including in Human and Poultry. Antibiotics 2022, 11, 1406. [Google Scholar] [CrossRef]
- Jeon, J.; Kim, J.-w.; Yong, D.; Lee, K.; Chong, Y. Complete Genome Sequence of the Podoviral Bacteriophage YMC/09/02/B1251 ABA BP, Which Causes the Lysis of an OXA-23-Producing Carbapenem-Resistant Acinetobacter baumannii Isolate from a Septic Patient. J. Virol. 2012, 86, 12437–12438. [Google Scholar] [CrossRef]
- Wu, M.; Hu, K.; Xie, Y.; Liu, Y.; Mu, D.; Guo, H.; Zhang, Z.; Zhang, Y.; Chang, D.; Shi, Y. A novel phage PD-6A3, and its endolysin Ply6A3, with extended lytic activity against Acinetobacter baumannii. Front. Microbiol. 2019, 9, 3302. [Google Scholar] [CrossRef]
- Strati, F.; Lattanzi, G.; Amoroso, C.; Facciotti, F. Microbiota-targeted therapies in inflammation resolution. Semin. Immunol. 2022, 59, 101599. [Google Scholar] [CrossRef]
- Pinu, F.R.; Beale, D.J.; Paten, A.M.; Kouremenos, K.; Swarup, S.; Schirra, H.J.; Wishart, D. Systems Biology and Multi-Omics Integration: Viewpoints from the Metabolomics Research Community. Metabolites 2019, 9, 76. [Google Scholar] [CrossRef]
- Balvočiūtė, M.; Huson, D.H. SILVA, RDP, Greengenes, NCBI and OTT — how do these taxonomies compare? BMC Genom. 2017, 18, 114. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, M. Metagenomics: Perspectives, methods, and applications. In Metagenomics; Academic Press: London, UK, 2018. [Google Scholar]
- Antoine, L.; Bruno, D. Advances in high-resolution accurate mass spectrometry application to targeted proteomics. PROTEOMICS 2015, 15, 880–890. [Google Scholar] [CrossRef]
- Heyer, R.; Schallert, K.; Zoun, R.; Becher, B.; Saake, G.; Benndorf, D. Challenges and perspectives of metaproteomic data analysis. J. Biotechnol. 2017, 261, 24–36. [Google Scholar] [CrossRef]
- Bi, H.; Krausz, K.; Manna, S.; Li, F.; Johnson, C.; Gonzalez, F. Optimization of harvesting, extraction, and analytical protocols for UPLC-ESI-MS-based metabolomic analysis of adherent mammalian cancer cells. Anal. Bioanal. Chem. 2013, 405, 5279–5289. [Google Scholar] [CrossRef]
- Marcinowska, R.; Trygg, J.; Wolf-Watz, H.; Mortiz, T.; Surowiec, I. Optimization of a sample preparation method for the metabolomic analysis of clinically relevant bacteria. J. Microbiol. Meth. 2011, 87, 24–31. [Google Scholar] [CrossRef]
- Beale, D.; Barratt, R.; Marlow, D.; Dunn, M.; Palombo, E.; Morrison, P.; Key, C. Application of metabolomics to understanding biofilms in water distribution systems: a pilot study. Biofouling 2013, 29, 283–294. [Google Scholar] [CrossRef]
- Beale, D.; Karpe, A.; Ahmed, W.; Cook, S.; Morrison, P.; Staley, C.; Sadowsky, M.; Palombo, E. A community multi-omics approach towards the assessment of surface water quality in an urban river system. Int. J. Environ. Res. Public Health 2017, 14, 303. [Google Scholar] [CrossRef] [PubMed]
- Beale, D.J.; Morrison, P.D.; Palombo, E.A. Detection of Listeria in milk using non-targeted metabolic profiling of Listeria monocytogenes: A proof-of-concept application. Food Control 2014, 42, 343–346. [Google Scholar] [CrossRef]
- Meijer, S.; Otero, J.; Olivares, R.; Andersen, M.R.; Olsson, L.; Nielsen, J. Overexpression of isocitrate lyase—glyoxylate bypass influence on metabolism in Aspergillus niger. Metab. Eng. 2009, 11, 107–116. [Google Scholar] [CrossRef]
- Maier, K.; Hofmann, U.; Bauer, A.; Niebel, A.; Vacun, G.; Reuss, M.; Mauch, K. Quantification of statin effects on hepatic cholesterol synthesis by transient 13C-flux analysis. Metab. Eng. 2009, 11, 292–309. [Google Scholar] [CrossRef]
- Niklas, J.; Schneider, K.; Heinzle, E. Metabolic flux analysis in eukaryotes. Curr. Opin. Biotech. 2010, 21, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Beale, D.J.; Morrison, P.D.; Karpe, A.V.; Dunn, M.S. Chemometric Analysis of Lavender Essential Oils Using Targeted and Untargeted GC-MS Acquired Data for the Rapid Identification and Characterization of Oil Quality. Molecules 2017, 22, 1339. [Google Scholar] [CrossRef] [PubMed]
- Driouch, H.; Melzer, G.; Wittmann, C. Integration of in vivo and in silico metabolic fluxes for improvement of recombinant protein production. Metab. Eng. 2012, 14, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, H.; Carlsen, M.; Nielsen, J. Identification of enzymes and quantification of metabolic fluxes in the wild type and in a recombinant Aspergillus oryzae strain. Appl. Environ. Microb. 1999, 65, 11–19. [Google Scholar] [CrossRef]
- Beale, D.J.; Kouremenos, K.A.; Palombo, E.A. Microbial metabolomics; Springer Int. Publ: Basel, Switzerland, 2016. [Google Scholar]
- Jinawath, N.; Bunbanjerdsuk, S.; Chayanupatkul, M.; Ngamphaiboon, N.; Asavapanumas, N.; Svasti, J.; Charoensawan, V. Bridging the gap between clinicians and systems biologists: from network biology to translational biomedical research. J. Transl. Med. 2016, 14. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- McNally, C.P.; Eng, A.; Noecker, C.; Gagne-Maynard, W.C.; Borenstein, E. BURRITO: An interactive multi-omic tool for visualizing taxa–function relationships in microbiome data. Front. Microbiol. 2018, 9, 365. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef]
- Karpe, A.V.; Liu, J.-W.; Shah, A.; Koloski, N.; Holtmann, G.; Beale, D.J. Utilising lipid and, arginine and proline metabolism in blood plasma to differentiate the biochemical expression in functional dyspepsia (FD) and irritable bowel syndrome (IBS). Metabolomics 2022, 18, 38. [Google Scholar] [CrossRef]
- Karpe, A.V.; Nguyen, T.V.; Shah, R.M.; Au, G.G.; McAuley, A.J.; Marsh, G.A.; Riddell, S.; Vasan, S.S.; Beale, D.J. A Time-Series Metabolomic Analysis of SARS-CoV-2 Infection in a Ferret Model. Metabolites 2022, 12, 1151. [Google Scholar] [CrossRef]
- Beale, D.J.; Shah, R.; Karpe, A.V.; Hillyer, K.E.; McAuley, A.J.; Au, G.G.; Marsh, G.A.; Vasan, S.S. Metabolic Profiling from an Asymptomatic Ferret Model of SARS-CoV-2 Infection. Metabolites 2021, 11, 327. [Google Scholar] [CrossRef] [PubMed]
- Attia, S.; Versloot, C.J.; Voskuijl, W.; van Vliet, S.J.; Di Giovanni, V.; Zhang, L.; Richardson, S.; Bourdon, C.; Netea, M.G.; Berkley, J.A.; et al. Mortality in children with complicated severe acute malnutrition is related to intestinal and systemic inflammation: an observational cohort study. Am. J. Clin. Nutr. 2016, 104, 1441–1449. [Google Scholar] [CrossRef] [PubMed]
- Calder, N.; Walsh, K.; Olupot-Olupot, P.; Ssenyondo, T.; Muhindo, R.; Mpoya, A.; Brignardello, J.; Wang, X.; McKay, E.; Morrison, D.; et al. Modifying gut integrity and microbiome in children with severe acute malnutrition using legume-based feeds (MIMBLE): A pilot trial. Cell Rep. Med. 2021, 2, 100280. [Google Scholar] [CrossRef]
- Muller Bark, J.; Karpe, A.V.; Doecke, J.D.; Leo, P.; Jeffree, R.L.; Chua, B.; Day, B.W.; Beale, D.J.; Punyadeera, C. A pilot study: Metabolic profiling of plasma and saliva samples from newly diagnosed glioblastoma patients. Cancer Medicine n/a. [CrossRef]
- Durán, C.; Ciucci, S.; Palladini, A.; Ijaz, U.Z.; Zippo, A.G.; Sterbini, F.P.; Masucci, L.; Cammarota, G.; Ianiro, G.; Spuul, P.; et al. Nonlinear machine learning pattern recognition and bacteria-metabolite multilayer network analysis of perturbed gastric microbiome. Nat. Commun. 2021, 12, 1926. [Google Scholar] [CrossRef] [PubMed]
- Arnold, B.F.; van der Laan, M.J.; Hubbard, A.E.; Steel, C.; Kubofcik, J.; Hamlin, K.L.; Moss, D.M.; Nutman, T.B.; Priest, J.W.; Lammie, P.J. Measuring changes in transmission of neglected tropical diseases, malaria, and enteric pathogens from quantitative antibody levels. PLOS Neglected Trop. Dis. 2017, 11, e0005616. [Google Scholar] [CrossRef] [PubMed]
- Jain, H.A.; Agarwal, V.; Bansal, C.; Kumar, A.; Faheem; Mohammed, M.-U.-R.; Murugesan, S.; Simpson, M.M.; Karpe, A.V.; Chandra, R.; et al. CoviRx: A User-Friendly Interface for Systematic Down-Selection of Repurposed Drug Candidates for COVID-19. Data 2022, 7, 164. [Google Scholar] [CrossRef]
- Cunningham, M.; Azcarate-Peril, M.A.; Barnard, A.; Benoit, V.; Grimaldi, R.; Guyonnet, D.; Holscher, H.D.; Hunter, K.; Manurung, S.; Obis, D.; et al. Shaping the Future of Probiotics and Prebiotics. Trends Microbiol. 2021, 29, 667–685. [Google Scholar] [CrossRef]
- Gibbons, S.M.; Gurry, T.; Lampe, J.W.; Chakrabarti, A.; Dam, V.; Everard, A.; Goas, A.; Gross, G.; Kleerebezem, M.; Lane, J.; et al. Perspective: Leveraging the Gut Microbiota to Predict Personalized Responses to Dietary, Prebiotic, and Probiotic Interventions. Adv. Nutr. 2022, 13, 1450–1461. [Google Scholar] [CrossRef]
- Kumar, R.; Sood, U.; Kaur, J.; Anand, S.; Gupta, V.; Patil, K.S.; Lal, R. The rising dominance of microbiology: what to expect in the next 15 years? Microb. Biotechnol. 2022, 15, 110–128. [Google Scholar] [CrossRef]
Figure 1.
A global distribution of death caused due to diarrhoeal infections in 2012. (Courtesy: Chris55 [CC BY-SA 4.0 (https://creativecommons.org/licenses/by-sa/4.0)], via Wikimedia Commons; and http://www.who.int/healthinfo/global_burden_disease/estimates/en/index1.htm).
Figure 1.
A global distribution of death caused due to diarrhoeal infections in 2012. (Courtesy: Chris55 [CC BY-SA 4.0 (https://creativecommons.org/licenses/by-sa/4.0)], via Wikimedia Commons; and http://www.who.int/healthinfo/global_burden_disease/estimates/en/index1.htm).
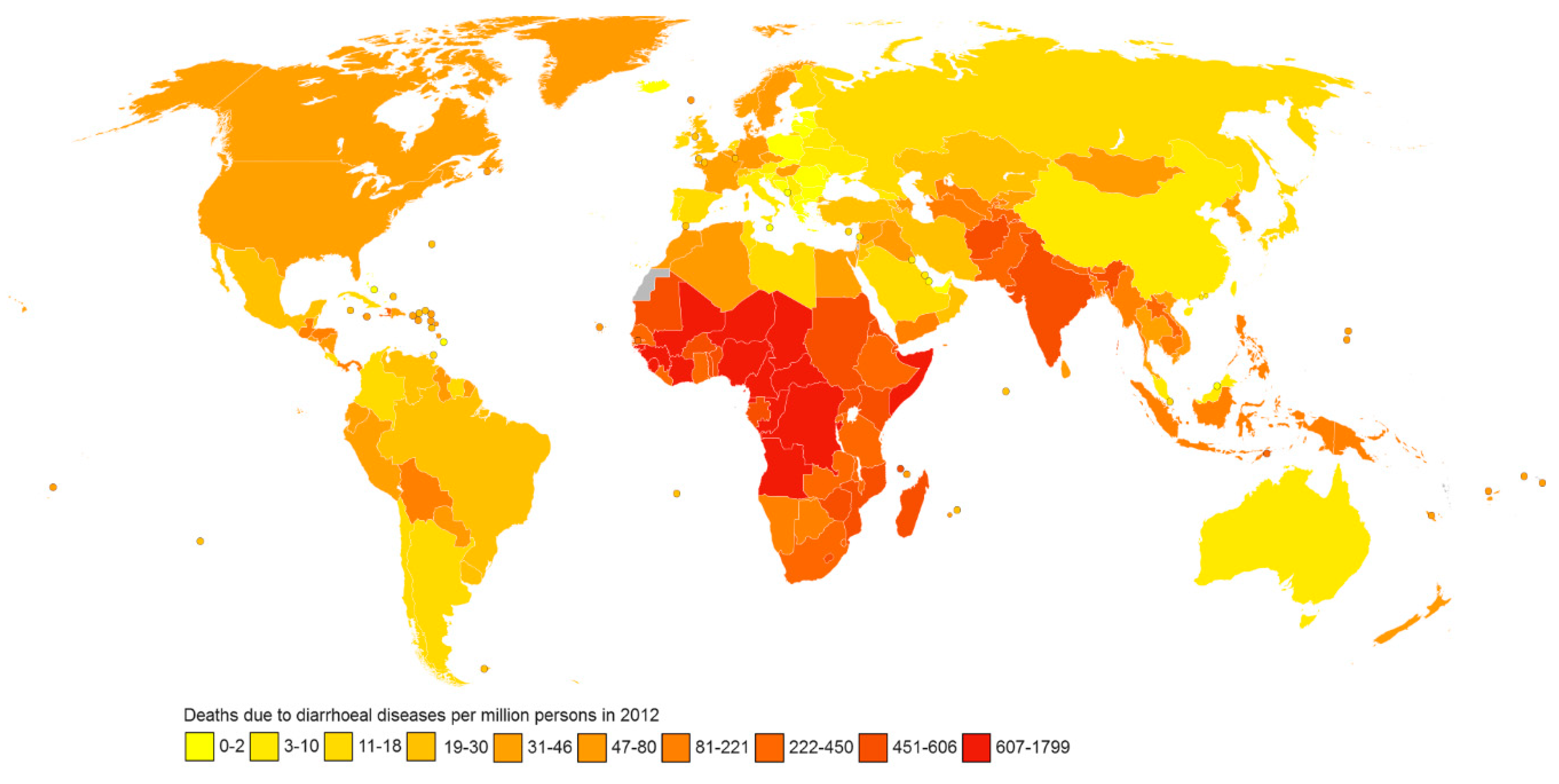
Figure 2.
A general life cycle and, growth stages of protozoan enteric parasites. Note: This is a generalised outlay, with each protozoan discussed here showing minor variations in their respective cycles.
Figure 2.
A general life cycle and, growth stages of protozoan enteric parasites. Note: This is a generalised outlay, with each protozoan discussed here showing minor variations in their respective cycles.
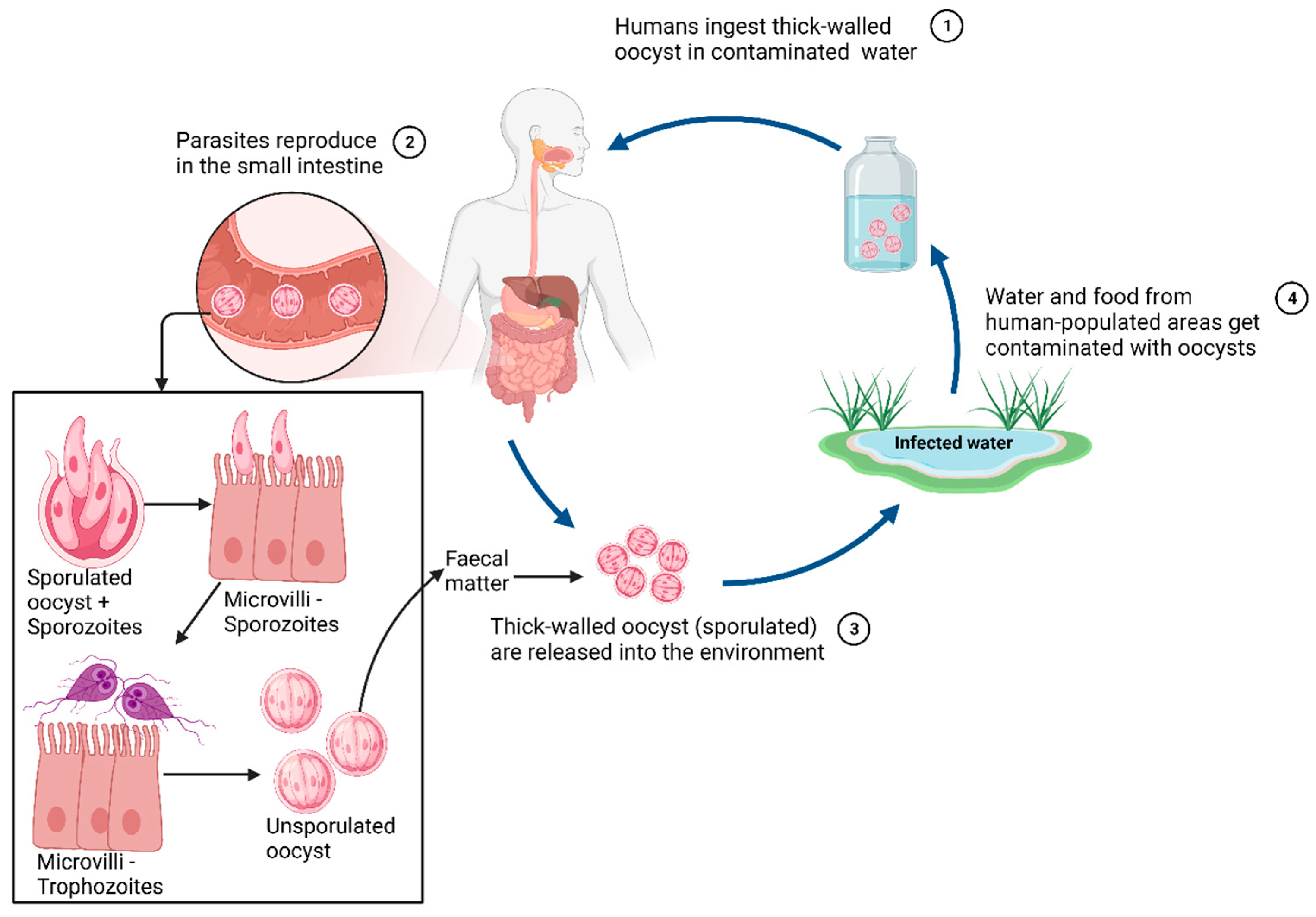
Figure 2.
A general representation of host-organs directly and indirectly affected by the enteric protozoan parasite infection.
Figure 2.
A general representation of host-organs directly and indirectly affected by the enteric protozoan parasite infection.
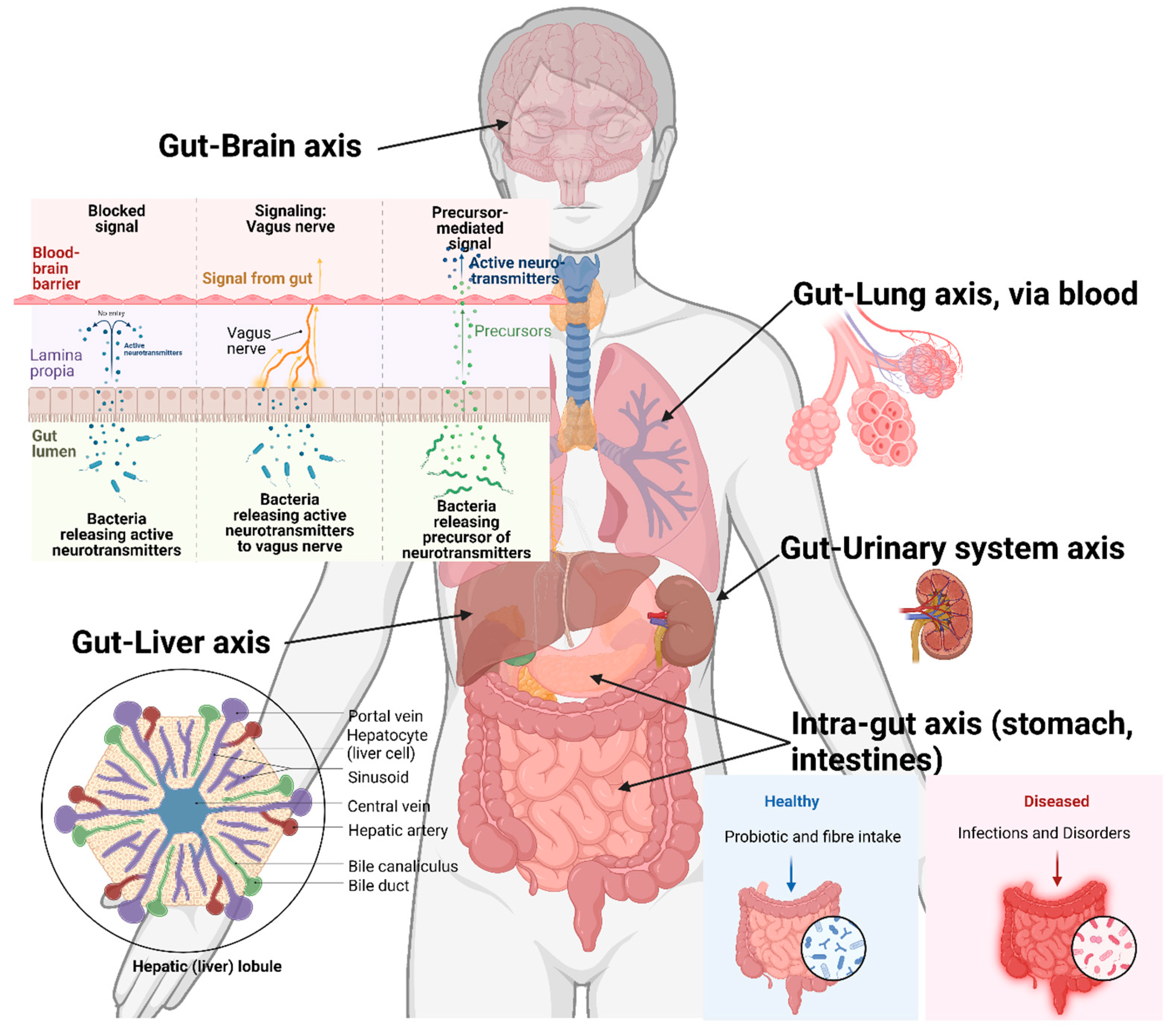
Figure 4.
Interaction and association networks. (A) representing regulatory functions and functional expression, respectively at various levels and (B) representing the changes caused during disease, with regulatory components of (a) present/ absent key component (green: presence, red: absence); (b) misregulated gene expression causing over/ under-expression (node size: expression level); (c) absence/ erroneous interactions (dotted lines represent erroneous interactions); (d) misregulated directions (misdirected arrows); (e) interaction impact (arrow thicknesses + numbers). Figure courtesy of Jinawath, et al. [117]. Distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/).
Figure 4.
Interaction and association networks. (A) representing regulatory functions and functional expression, respectively at various levels and (B) representing the changes caused during disease, with regulatory components of (a) present/ absent key component (green: presence, red: absence); (b) misregulated gene expression causing over/ under-expression (node size: expression level); (c) absence/ erroneous interactions (dotted lines represent erroneous interactions); (d) misregulated directions (misdirected arrows); (e) interaction impact (arrow thicknesses + numbers). Figure courtesy of Jinawath, et al. [117]. Distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



