
Submitted:
31 May 2023
Posted:
02 June 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
As the name implies, severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is a single-stranded RNA virus and a member of the corona virus family, primarily affecting the upper respiratory system and the lungs. Like many other respiratory viruses, SARS-CoV-2 can spread to other organ systems. Apart from causing diarrhea, another most common but debilitating complication caused by the SARS-CoV-2 is neurological symptoms and cognitive difficulties, which occur in up to two thirds of hospitalized covid patients and ranging from shortness of concentration, overall declined cognitive speed to executive or memory function impairment. Neuro-cognitive dysfunction and “brain fog” are frequently present in COVID-19 cases, which can last several months after the infection, leading to disruption of daily life. Cumulative evidence suggests that SARS-CoV-2 affects vasculature in the extra pulmonary systems directly or indirectly, leading to impairment of endothelial function and even multi-organ damage. The post COVID-19 long-lasting neurocognitive impairments have not been studied fully; and the underlying mechanism remains elusive. In this review, we summarize the current understanding of the effects of COVID-19 on vascular dysfunction and how vascular dysfunction leads to cognitive impairment in patients.
Keywords:
Long-COVID-19
; SARS-CoV-2
; Endothelial cells
; Cognitive dysfunction
; Blood Brain Barrier
; Neuro-inflammation
Introduction
In the late 2019 a new variant of Coronaviruses, SARS-CoV2 or COVID-19, emerged as a cause of novel severe acute respiratory syndrome, which had quickly spread to entire world and became the newest global health concern [1,2]. SARS-CoV-2 is primarily characterized as a respiratory pathogen transmitting via respiratory droplets, although abnormalities of other organs including the brain, heart, kidneys, liver, skeletal muscle, and skin have also been noticed [3]. A meta-analysis study of 24,410 patients with COVID-19 reported fever, cough, fatigue, and hyposmia as the most common symptoms [4]. Compared to impaired olfaction and gustation as common as 85% of COVID-19 patients, 36.4% of them exhibited neurological abnormalities, such as dizziness, headache and weakened consciousness during hospitalization [3]. A study using murine models indicated that coronavirus could enter CNS after infection [5], corroborating with the neurological symptoms observed clinically. In addition, long-term cognitive dysfunction has been detected in patients recovered from COVID-19 infection and patients with persistent infection [6]. Cognition is defined as the conceptual processing of information that is mandatory for acquiring knowledge and expression of a response. This process includes obtaining information (perception); selecting necessary data (attention), representing (understanding) representing (understanding) and retaining information (memory); leading to behavior control (reasoning and judgment) [7]. Fatigue and lasting impairment of memory, decision-making, and concentration are the most common neurological injuries reported in non-hospitalized COVID-19 patients. Additionally, numerous patients described subtle cognitive and behavioral deficits that are difficult to be characterized. These symptoms are commonly described as “brain fog”; or “mental clouding” [8]. A recent guideline on long-term effects of COVID-19 published by the National Institute of Health and Care Excellence (NICE) suggests use the definition “ongoing symptomatic COVID-19” for indications lasting between 4 and 12 weeks after the acute onset and “post-COVID-19-syndrome” for symptoms lasting more than 12 weeks [9]. Evidence of cohort studies implies the relationship between continuing systemic inflammation during SARS-CoV-2 infectivity and following cognitive failure and hippocampal atrophy. Hence, in COVID19-caused cognitive decline, attention should be paid to the role of inflammation. Through pro-inflammatory cytokines releasing, SARS-CoV-2 can trigger signaling pathways, finally resulting in disrupted TJ proteins and blood-brain-barrier (BBB) leakage. This process further allows the infiltration of immune cells and other particles into the brain and worsening brain injury [10,11]. Herein, we discuss the long-lasting neuro-cognitive impairment caused SARS-CoV-2 and the probable role of vascular dysfunction on proceeding the cognitive deficiency.
Long-term neurological and cognitive dysfunction of COVID-19
During Covid-19 pandemic, cognitive abnormalities were commonly detected. Interests of clinical and public health are therefore no longer restricted to mortality rate and clinical consequences of hospitalized patients but extended to long-term adverse outcomes of COVID-19 disease even after recovery and discharge from the hospital. A growing body of evidence implies on the persistent neurological and cognitive symptoms after COVID-19 illness. Given the persistent neurological features after SARS-CoV-2 infection, several studies implied on the onset of neurodegenerative disease in patients without any previous history. Accordingly, 15% of SARS-CoV-2 cases showed one or more neurological signs for first time including 13% polyneuro/myopathy, 1% Guillain-Barré syndrome, 2% mild encephalopathy, 1% parkinsonism, and 1% ischemic stroke during 3 months after infection [12]. As well, a cohort study of 143 patients revealed headache, hyposmia, and myalgia at least in 5% of population 2 months after SARS-CoV-2 infection [13].
In the context of cognitive deficits, it has been reported that 20–70% of COVID-19 cases display a quantitative or qualitative disturbance of consciousness during the acute phase [14,15]. In one study, the psychopathological and cognitive status of 226 COVID-19 pneumonia survivors were prospectively assessed one- and three-months following hospital discharge. Regardless of clinical and physical severity, 78% of the participants presented poor function in at least one cognitive domain, while 50 to 57% of the sample displayed impaired executive functions and psychomotor coordination [16]. In another case report, cognitive screens from recovered non-hospitalized COVID-19 case showed persistent neuro-cognitive symptoms, particularly difficulties in working memory and executive function [17]. Also, a follow up study of 13001 participants with relatively mild COVID-19 disease informed the significantly higher prevalence of memory problems eight months after the positive SARS-CoV-2 test compared to the control group in favor of the worsening of health even after recovery from infection [18]. In a separate cohort study, only long-term anosmia, ageusia, memory loss, and headache remained in COVID-19 cases after 60 days, yet memory deficit continued in these patients in third months of disease [19]. Similarly, according to follow up of 18 mild to moderate COVID-19 patients 85 days after recovery, prevalence rates of over than 75% reported difficulties in episodic memory, attention, and concentration [20]. As well, comparison of cognitive function in SARS-CoV-2 patients to matched controls suggested significant differences in the sustained attention [21], executive function and visuospatial processing [22], attention, memory, and language [23].
Interestingly a case report describes progress of disease from mild common symptoms like myalgia, fatigue, smelling loss, and memory deficit at the beginning to severe outcomes including right-sided weakness, sensory loss, and worsening cognitive impairment. Neuroimaging has confirmed an ischemic infarct in the middle cerebral artery of this case [1,24]. As well, a recent cross-sectional report studied the connection between COVID-19 disease and cognitive signs in 57 hospitalized participants and found a high prevalence of cognitive impairment (>80% of the sample), primarily in the fields of attention and executive function was observed [25].
Among cognitive performances, fatigue, memory, and concentration difficulties were reported as most common long-lasting symptoms by 36% of symptomatic COVID-19 patients after 4 weeks and mostly observed in women [9,26,27,28].
According to a study, cases with a history of mild symptomatic SARS-CoV-2 infections have more than 18 times higher risk for cognitive deficits than individuals without clinical manifestations of the infection [29]. As well, a recent longitudinal study showed the important role for COVID-19 severity in modulating the long-term cognitive outcomes of disease [30]. Therefore, cognitive changes can occur even after milder COVID-19 illness. However, hospitalized and severely affected patients are in higher risk for persistent cognitive and neurological dysfunction. Besides, amongst diverse cognitive domains, impairment of episodic and working memory, attention and concentration deficits, consciousness, and executive function difficulty are the major cognitive complications in patients suffering COVID-19 infection.
Molecules contribute to COVID-19 penetration into the CNS
Since none of the COVID-19 cases with severe neurological complains reveals evidence of SARS-CoV-2 virus in the cerebrospinal fluid, both direct and indirect mechanisms may participate in CNS dysfunction [17]. The covid virus infects via binding of spike glycoprotein (S protein) of the virus to the angiotensin converting enzyme 2 (ACE2) receptor on the host cell surface, followed by the incorporation of viral genome into the host cell genome. ACE2, a cardio-cerebro- vascular protective factor that converts angiotensin (Ang) I to Ang II to stimulate Ang II receptor type1 (AT1R), is expressed by neurons, astrocytes, oligodendrocytes, and endothelial cells of CNS (2), and highly concentrated in the olfactory bulb, substantia nigra, middle temporal gyrus, and posterior cingulate gyrus [31]. Activated AT1R can promote the inflammation and neurodegeneration and increase blood pressure [32]. Therefore, the widespread presentation of the ACE2 as SARS-CoV-2 receptor on the neural and endothelial cells makes the CNS susceptible to SARS-CoV-2 invasion [33,34]. Since direct access of COVID-19 disease is possible through BBB free sites in the brain, its interaction with ACE-2 of endothelial cells in disrupted BBB provides another penetration route for virus [35], While, the transmembrane protease serine 2 (TMPRSS2) facilitates virus entrance through aiding S protein priming. TMPRSS2 inhibitors are beneficial for mental recovery [31]. TMPRSS2 is detectable all over the olfactory epithelium and choroid plexus of rodents and human [36,37]. In addition to ACE2, basigin or CD147 and neuropilin-1 (NRP1) also operate as docking receptors for SARS-CoV-2 virus. Furthermore, several proteases like cathepsin B and L (CatB/L), and furin contribute in the viral entrance to cell and its replication [38]. NRP1, a transmembrane receptor without cytosolic protein kinase domain, is highly expressed in the olfactory epithelium and mediates the SARSCoV2 invasion into the CNS [39]. As mentioned above, alternative receptor for penetration of SARS-CoV-2 into the brain is CD147 which is expressed in neural cells [40]. Additionally, CatB/L, an endosomal cysteine protease, mediates the priming of the SARSCoV2 spike protein [41]. It suggests that TMPRSS2 and lysosomal cathepsins are able to trigger SARS-CoV-2 invasion; likewise, furin facilitates this process [42].
Endothelial cell infection and endotheliitis in COVID-19
The vascular endothelia cells are the innermost lining of blood vessels and fundamental for maintaining vessel structural and functional integrity by regulating vascular tone and permeabilization, responses to oxidative stress or inflammation to achieve tissue homeostasis [43]
A recent study displayed the relationship between the levels of endothelia biomarker and the levels of pro-inflammatory cytokines and chemokines [44]. Along the same vein, recent research indicated that pro-inflammatory mediators released by spike-activated macrophages augment the endothelia cells activation, probably contributing to the impairment of vascular integrity and development of a pro-coagulative endothelium [45].
Accumulating evidence indicates the presence of extensive microthrombi and endothelia cells damage all over the pulmonary vasculature, suggesting the involvement of vasculopathy in the pathogenesis of COVID 19. Although one study failed to find viral RNA in endothelial cells from COVID-19 patients’ autopsy tissue [46], another study detected SARS-CoV-2 RNA in endothelial cells by in situ hybridization (ISH) in 2 out of 32 cases [47] [44]. It has shown that endothelial cells and pericytes throughout the body express ACE-2 as well as BSG, CD147, and NRP1 [38,48,49,50], supporting the trophism for SARS-CoV-2. Autopsy tissues from COVID-19 patients shown the high expression of ACE2, TMPRSS2 and endothelial cells inflammation agents in capillaries but less presence in arteries and veins. The existent data suggests the development of COVID-19-causes endotheliitis in small vessels like capillaries; nevertheless, the involvement of main coronary vessels principally arises from indirect mechanisms of SARS-CoV-2 infection [51].
A recent study utilizing the COVID-19 rhesus macaques model showed that SARS-CoV-2 infection can mediate indirect endothelial cell dysfunction by immune and inflammatory pathways interaction [52]. In another COVID-19 study, combined activation of NF-κB signaling cascade and changes in mitochondrial quality control led to endothelia cells activation and enhanced neuroinflammation [53]. As a result, SARS-CoV-2-infected human brain microvascular endothelial cells showed augmented caspase 3 cleavage and apoptotic cell death of endothelial cells.
RNA sequencing data of covid 19 subjects showed the higher expression of endothelia cells activation markers, RELB and TNF-α [54]. Accordingly, it is assumed that hyper-inflammation is responsible for endothelia cell injury and death following SARS-CoV-2 infection. As well, significantly high levels of Cxcl9 and Cxcl10 in the endothelia cell cluster in SARS-CoV-2 infected K18 mice (SARS-CoV-2-infected human cytokeratin 18 (K18)-hACE2) was reported compared to infected WT mice, suggesting the interferon pathway upregulation. The Kras pathway was also upregulated. Additionally, upregulation of Vcam1 and Icam1 genes in the lysosome/apoptosis and complement pathways in viral RNA positive cells was evident [55]. A more recent study reported signs of damage and activation of endothelia cells in the form of enhanced gene expression cascades regarding EMT/EndoMT in the hearts of COVID-19 infected patients followed by an upregulation of tissue hypoxia-related pathways and the formation of ultra-structurally detectable thrombi (uTh). Endothelia cells induce the recruitment of monocytes/macrophages to the site of injury by upregulation of adhesion-molecules and activation of SDF-1/CXCR4 signaling [56]. Overall, these results shed lights on the importance of SARS-CoV-2-mediated immune activation on the endothelial dysfunction and injury.
Degradation of endothelial glycocalyx makes them vulnerable to SARS-CoV-2 entry.
The glycocalyx is a layer covering the luminal surface of vascular endothelia cells, which contributes to the maintenance of vascular homeostasis via regulating vascular tone, permeability, thrombosis and leukocyte adhesion to endothelium [57].The glycocalyx is comprised of numerous proteoglycans, glycosaminoglycans, glycoproteins, and associated plasma proteins. The intact highly sulfated glycocalyx structure of the endothelium can prevent SARS-CoV-2 but its disruption increases susceptibility to SARS-CoV-2 infection, resulting in hyperinflammation and oxidative stress [58]. In this regard, a recent study has reported the interaction of intact glycocalyx with Spike protein and its ability in prohibiting the binding of S protein and ACE2. Conversely, damaged glycocalyx encouraged S protein and ACE2 binding and enabled viral entry [59]. In vivo evidence suggests that COVID-19 patients with severe symptoms showed endothelial glycocalyx disruption and syndecan-1 (SDC-1) secretion (Yamaoka-Tojo 2020). Even among patients recovered from COVID-19, the levels of SDC-1 were significantly elevated compared to healthy controls, exhibiting the existence of persistent endothelial damage after COVID-19 progression [60].
Endothelial cells inflammation and blood brain barrier disruption in COVID-19
Cranial nerves and the BBB are main avenues between the brain and SARS-CoV-2. However, viruses such as SARS-CoV-2 cannot easily enter the brain parenchyma through the endothelial cells that line the lumen side of the capillaries of the systemic circulatory system due to the unique physiology of the BBB. The BBB is a functional unit mostly consists of brain endothelia cells linked by tight junctions, vascular basement membrane, pericytes, astrocytes end processes, neural cells, and microglia [61]. This dynamic structure operates as a bridge between the plasma and brain parenchyma, which jointly considered as the neurovascular units (NVUs). In BBB, endothelial cells are fastened by tight junction (TJ) proteins including ZO scaffolding proteins, claudin-5, and occludin as well as junctional adhesion molecules to prevent the extracellular and transcellular diffusion of molecules in the CNS [62]. Hence, in addition to neuroinvasion, disturbance of BBB integrity including endothelial TJs disruption may expose the brain in danger to SARS-CoV-2 penetration from infected blood, influencing neuronal function in the CNS [63]. Hyper-inflammatory condition after COVID-19 infection may contribute to BBB damage [64]. In support of this notion, human brain microvascular endothelia cells were reported to over express pro-inflammatory cytokines, chemokines, and adhesion molecules and with lower expression of tight junction protein coincided with increased BBB permeability following SARS-CoV-2 infection [65].
Increased pro-inflammatory cytokines levels causes TJ function alteration and disruption of BBB. For example, IL-1 enhancement damages BBB integrity [66], whereas IL-1β upregulates matrix metalloproteinase (MMP)-9 and promotes TJ proteins disruption via activating extracellular signal regulated kinases [67]. Additionally, increased TNF-α, IL-6, and IL-12 cytokines levels result in the deprivation of TJ proteins and consequent BBB permeability impairment [68]. cytokines probably disturb the integrity of different sorts of junctional proteins such as VE-cadherin, ZO-1, β-catenin and gap junction, leading to penetration of inflammatory and immune cells [69]. Inflammatory BBB injury enhances the passing of immune cells through the BBB and finally increases viral particles and proinflammatory cytokines in the CNS, causing cytokines activation and vascular endothelial growth factor (VEGF) production in astrocytes [70]. By triggering the phosphoinositide 3 (PI3)-kinase and AKT signaling cascades and MMP-9 upregulation, VEGF is capable of disrupting TJ proteins in brain capillary endothelia cells and BBB deficiency [71] (Figure 1).
Proinflammatory cytokines enhance the passing of immune cells through the BBB that leads to VEGF production in astrocytes. By triggering the phosphoinositide 3 (PI3)-kinase and AKT signaling cascades and MMP-9 upregulation, VEGF can disrupt tight junction (TJ) proteins in brain capillary endothelial cells and leading to BBB breakdown.
Disseminating intravascular coagulation and blood brain barrier disruption in COVID-19
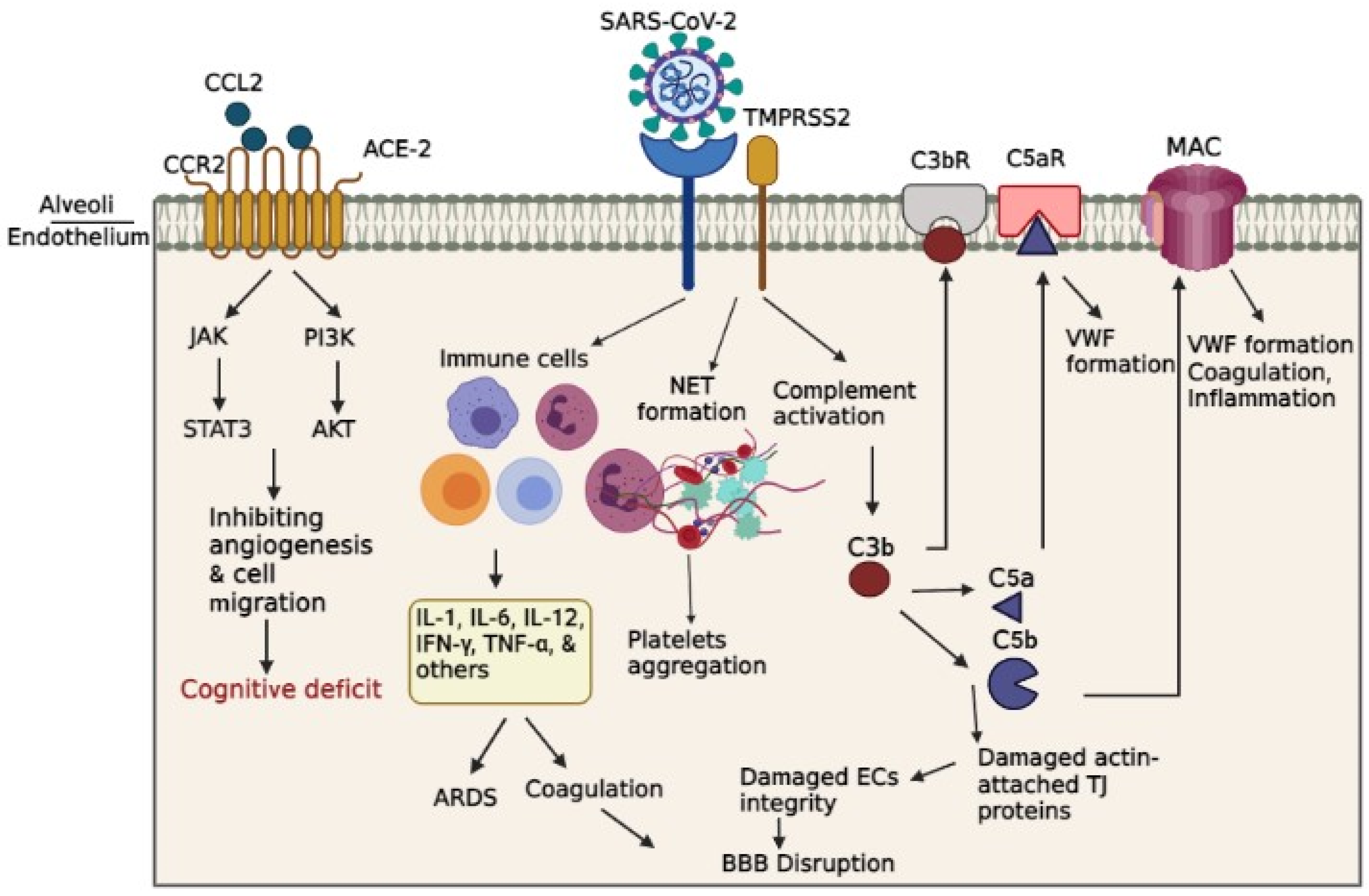
Both overactive coagulation as well as hyper-inflammation have been proposed as the main mechanisms underlying endothelia cells damage and thrombus development in COVID-19 [72]. Coagulation is frequently impaired in COVID-19 patients, resulting in a common hypercoagulable state in patients, which may be related to the incidence of stroke. Scientific records imply on increased intravascular coagulation, blood clot formation and bleeding in severe COVID-19 patients. Since thromboembolic single or multiple infarcts is reported in nearly 20% of dementia cases [73], thromboembolic occlusion of cerebral blood vessels is a potential causative of neurological manifestations, including cognitive deficit or dementia, especially in younger healthy adult COVID-19 subjects [74]. As shown in Figure. 2, several pathways are involved in intravascular coagulation in COVID-19 patients. (1) The release of pro-inflammatory cytokines can exert the platelets release, activation, and accumulation. Amongst cytokines, some agents like IL-6 and Cathepsin G, a serine protease generated by neutrophils, has more thrombogenic capacity and can stimulate platelets aggregation. In addition, TNF-ɑ induces the surge of plasminogen-activator inhibitor-1 (PAI-1), leading to subsequent decreased activity of plasmin and reduced fibrinolysis [75]. (2) Activation of complement system is a principal inducer of coagulation. SARS-CoV2 interacts with ACE-2 and activates the complement system including the lectin and classical pathways, resulting in the production of C3a and C3b. C3a mediates inflammation and activates the platelets, while C3b contributes to the production of the C5a and C5b. Binding of C5a to C5 receptor mediates the platelet activation, aggregations, discharge of the procoagulant microparticles (PMP), and the development of the blood clots. In addition, it may also contribute to the recruitment of neutrophils. The C5b generates the membrane attack complexes (MACs), serving as a transmembrane channel to initiate lysis of the embedded cells. The MAC activates the microvascular complement deposition, coagulation, and inflammation. Besides, cell lysis and death of target cells are involved in coagulopathy by enhancement of prothrombin activity as well as von Willebrand factor (VWF) formation [76]. Another mechanism by which complement activation is involved in coagulation is the binding of C3b to CR1 receptor on the platelet’s membrane. This process triggers the release of short-chain polyphosphate (polyP) from platelets, inducing the expression of tissue factor (TF). (3) Whereas formation of neutrophil extracellular traps (NET), composed of chromatin and microbicidal proteins as well as neutrophils, is a key mechanism of conglutination since neutrophils are crucial player in the production of thromboses. NETs participate in the pathobiology of thrombosis, by which histones, as a main element of NETs, attract and bind to platelets, leading to their aggregation [77]. As indicated in a recent study, neutrophils contribute in the immune response to SARS-CoV-2 invasion. Since these cells are much bigger than erythrocytes and the average capillary diameter, neutrophils can plug capillaries and cause significant blood flow disruption [78]. The adhesion of hyper-activated neutrophils in brain capillaries diminishes the cerebral blood flow in animal models of AD and consequently causes memory dysfunction [79]. Neutrophil-induced disruption of capillary blood flow within the lungs, brain, heart, and other organs is implicated in the poor prognosis of COVID-19 illness [80].
A study utilizing rat model of intraventricular hemorrhage indicated BBB disruption followed by thrombin-caused activation of Src kinase phosphorylation. Src triggering increases BBB permeability by MMPs phosphorylation and TJ proteins disruption and VEGF upregulation [81,82]. As well, fibrinogen can harm endothelia cells integrity by damaging actin filaments-attached TJ proteins [83]. On the other hand, enhanced generation of actin probably results in cellular stiffness, actin filaments retraction, and spreading of endothelia cells junctions, thus interrupting endothelia cell integrity [84]. In this regard, Yepes et al. discovered vascular leakage in a dose-dependent manner following the intraventricular infusion of endogenous tissue plasminogen activator (tPA) [85] (Figure 2).
Pneumonia and BBB disruption in COVID-19
It has been suggested that cerebral hypoxia triggers hypometabolic, cognitive and degenerative changes in the brain and is contributory to the pathology of Alzheimer’s disease (AD) [86].
A large body of evidence suggests the bidirectional relationship between cognitive outcomes and ARDS, so that, pneumonia may influence cognitive performance depending on the patient populations and clinical contexts [87]. In contrast, pre-existing cognitive dysfunction is linked to the enhanced rate of mortality by pneumonia [88]. Hypoxemia and hypoxia are considered as the causes of neuronal atrophy, consequent enlargement of ventricles, and related cognitive dysfunction, mostly memory loss [89]. In this line, CT scans of COVID-19 patients with ARDS, exhibited widespread cerebral atrophy and widening of bilateral temporal horn compared to control group (Hopkins & Jackson, 2006). Consistently pneumonia has been reported to promote the risk of developing dementia among elderly patients [90]. Even in non-elderly cases with single episode of pneumonia without main medical comorbidities are vulnerable to enhanced danger of cognitive decline [90]. According to the report by Wilcox et al. 70 to 100% of ARDS- hospitalized patients indicated cognitive impairment, ranging from difficulties in attention, concentration, and memory, to executive function. These changes persisted in 46–78% one year, 25–47% two years, and almost 20% of cases five years after infection [91]. The weakening of memory may result from the susceptibility of hippocampal neurons to oxygen shortage [92]. Therefore, ARDS and hypoxia are vital contributors to the cognitive deficits specially memory loss in COVID-19 subjects.
Blood vessels supply oxygen and nutrients to neurons. Continuous hypoxic condition of brain tissue will ultimately cause irreversible neural damage [93]. Postmortem analyses of COVID-19 patients with hypoxic brain damage indicated neuronal injure in the neocortex, hippocampus, and cerebellum areas. Oligodendrocyte demise and widespread gliosis were also reported [94]. A body of evidence demonstrates an important role of hypoxia in BBB disruption [95]. Based on evidence, through changing the actin distribution and attenuating TJ proteins, the hypoxia rises the paracellular permeability of brain capillary endothelial cells (Figure 2).
Mitochondria are organelles operating as an energy delivery system. Mitochondrial energy metabolism is directly associated with the hypoxic condition. As well, hypoxia in the brain may strength the proliferative ability of the virus [96]. After hypoxia, a high virus presence in COVID-19 subjects with CNS involvement leads to the compromise of neurons with high-level energy metabolism. Hence, it has indicated that targeting selective neuronal mitochondrial in SARS-CoV-2 infection induces ‘brain fog’ and causes cognitive and behavioral deficits [8].
In another way, SARS-CoV-2 virus can result in mitochondrial energy metabolism impairment through targeting oxygen availability and consumption. Notably, the integration of the viral genome into the host cell mitochondrial matrix and creating a viral-mitochondrial interaction leads to these effects. The interaction between virus and mitochondria increases energy and reduces host immune reaction, resulting in enhanced replication and survival of virus [97,98]. Thus, this pathological influence of SARS-CoV-2 infection may elucidate the long-term psychiatric, cognitive, and neurodegenerative outcomes. Also, reduction of available mitochondrial energy leads to undesired host immune response [99]. Therefore, Impaired mitochondrial energy metabolism may be considered as a chief factor in cognitive manifestations in COVID-19 patients.
Vascular dysfunction, brain inflammation, and cognitive impairment
Normally, viruses such as SARS-CoV-2 cannot easily enter the brain parenchyma through the endothelial cells that line inside of the capillaries in the systemic circulatory system due to the unique physiology of the BBB. VE-cadherin, ZO-1, β-catenin and gap junction caused by hyper-inflammatory condition of SARS-CoV-2 virus results in dysfunction and enhanced leakage of BBB, which exacerbates penetration of virus through the disrupted BBB [63]. Once SARS-CoV-2 reaches cerebral tissue, it causes neuro-inflammation through activation of microglia and macrophages, leading to release of local pro-inflammatory cytokines in addition to the circulatory cytokines. Due to huge amount of pro-inflammatory cytokines in the circulation, integrity of BBB is disrupted, and brain becomes more vulnerable to the ischemic, hypoxic, thrombolytic threats along with invasion of various pathogens [100,101] (Figure 2).
Based on research, neurological dysfunction levels and BBB injury seems to be associated with the grade of cognitive loss and of COVID-19 infection severity. A recent study found the BBB permeability in 58% of COVID-19 subjects in 31 cases with neurological indications [102], suggesting that SARSCoV-2 can cause BBB dysfunction. The data of a recent research suggested that loss of BBB integrity might contribute to progressive impairment of cognitive in the diabetic rats. The increase of TNF-α and IL-6 expression might trigger the disruption of BBB in the brain, which eventually caused cognitive impairment in the 8-week STZ rats [103]. In support of BBB disruption influence on cognitive impairment, a recent findings suggested that BBB breakdown takes part in APOE4-connected cognitive weakening independently of Alzheimer’s disease pathology, and might be a therapeutic target in APOE4 carriers [104].
Likewise, regarding the regulatory role of ACE2 in controlling of blood pressure, occupying of ACE2 by viral spike protein may result in imbalance of angiotensin system and affect normal blood pressure. Primary pneumonia and pulmonary infection also cause oxygen deprivation in the brain parenchyma and produce hypoxic condition and subsequent metabolic disruption [105].
The cytokine storm phenomenon is an increase of pro-inflammatory cytokines levels in the serum, such as IL-2, IL-6 and IL-1β, IL-17, IL-8, G-CSF, GM-CSF, IP10, MCP1, MIP1α (also known as CCL3), and TNF [33], and causes acute respiratory distress syndrome (ARDS) in COVID-19 severe cases [106]. It has indicated that increased cytokines concentration, especially IL-6, TNFα, and IL-1β has a strong effect on working memory and attention. Damage of these cognitive abilities is a typical feature of delirium and approves key role of these cytokines in the etiology of COVID-19-associated cognitive impairments [107].
Prolonged neuro-inflammation, and continued hypoxia have been postulated as the potential primary causes of both acute and chronic cognitive features of COVID-19 [101]
Chemokines are small molecules primarily known for regulating chemoattraction of leukocytes and modulating immune reaction [108]. Additionally, chemokines are involved in different phases of CNS development via supporting cells migration, proliferation, and survival [109,110]. Four families of Chemokines include C, CC, CXC, and CX3C, characterized based on the conserved cysteine residues position [111]. CCL11 acts through activating PI3K/AKT, MAPK/p38, and JAK/STAT3 signaling pathways to prevents apoptosis, and promote angiogenesis and cell migration [112]. CCL11 also can trigger oxidative stress via microglial NOX1 stimulation and potentiate glutamate-mediated neurotoxicity (Parajuli, Horiuchi et al. 2015). During adulthood, the CCL2/CCR2 axis is able to modulate neurotransmission and neuromodulation [112]. It has indicated that treatment with CCL11 is capable of inhibiting neurogenesis in the adult brain, resulting in cognitive weakening [113].
Enhanced CCL11 concentration was found in plasma samples of long COVID cases suffering from cognitive symptoms, as well as white matter microglial reactivity was detected in patients with SARS-CoV-2 infection [114]. In addition, elevated levels of CCL11 are noted in Alzheimer’s disease and schizophrenia. Accordingly, targeted therapy to normalize CCL11 levels might develop mental and physical health among patients with schizophrenia and Alzheimer’s [115].
A recent study reported white matter microglial reactivity in mouse model of mild SARS-CoV-2 infection. Furthermore, the same pattern of prominent white matter-selective microglial responses was found in SARS-CoV-2 infected human brain tissue. In mouse model, pro-inflammatory CSF CCL11 enhanced at least 7-weeks after infection. As well, humans undergoing long-COVID with cognitive deficits showed higher CCL11 levels related to the cases with long-COVID without cognitive indications. Likewise, following mild SARS-CoV-2 infection in mice, hippocampal neurogenesis impairment, decreased oligodendrocytes, and myelin loss in subcortical white matter were detected after 1 week of infection, which persevered till at least 7 weeks [116]. The results of this study revealed a crucial role for long-term enhanced CCL11 levels in astrocyte-mediated microglial activation during SARS-CoV-2- induced CNS inflammation, leading to the extended mental and cognitive dysfunction on COVID-19 patients [117]. It has been also suggested that cerebral hypoxia triggers hypometabolic, cognitive and degenerative changes in the brain and is contributory to the pathology of Alzheimer’s disease (AD) and related cognitive impairment [118].
Conclusion and future directions
In summary, SARS-CoV-2 virus can invade the brain and exert the neurological manifestation through the binding to ACE2 on nerve cells and endothelial cells. A sound body of evidence shows the contribution of SARS-CoV-2 to vasculature integrity by direct or indirect viral infection, leading to endothelium damage and augmented vascular penetrability in peripheral vessels [120], leading to the disruption of the BBB [119].
Given the evidence, SARS-CoV2 pathogen can induce cognitive impairment via vascular dysfunction, disruption of BBB, interruption of oxygen supply, dissemination of intravascular coagulation, and neuro-inflammation. Taken together, the long-term cognitive consequences of SARS-CoV-2, to some extent, may be due to possible disruption of micro-structural and functional brain integrity during COVID-19 contamination and in the recovery stages of illness. In addition to present evidence, future studies are needed to discover exact long term cognitive deficits in patients with COVID-19 and their probable mediator mechanisms.
Funding
This study was supported by the National Institutes of Health R01 HL122774 (HS) NS027713 (HS), NS112819 (HS), NIH NINDS R01NS102886 (JL), Department of Veterans Affairs Administration BLR&D 2I01BX003335 (JL), IK6BX004600 (JL) and by the Michael Ryan Zodda Foundation (HS).
Conflict of Interest
'Declarations of interest: none”.
Ethical Approval
This article does not contain any studies with human participants or animals performed by any of the authors.
References
- Garg, A., et al., A case of COVID-19 with memory impairment and delayed presentation as stroke. Cureus, 2020. 12(8). [CrossRef]
- Wu, Z. and J.M. McGoogan, Characteristics of and important lessons from the coronavirus disease 2019 (COVID-19) outbreak in China: summary of a report of 72 314 cases from the Chinese Center for Disease Control and Prevention. jama, 2020. 323(13): p. 1239-1242. [CrossRef]
- Pilotto, A., et al., Long-term neurological manifestations of COVID-19: prevalence and predictive factors. Neurological Sciences, 2021. 42: p. 4903-4907. [CrossRef]
- Grant, M.C., et al., The prevalence of symptoms in 24,410 adults infected by the novel coronavirus (SARS-CoV-2; COVID-19): A systematic review and meta-analysis of 148 studies from 9 countries. PloS one, 2020. 15(6): p. e0234765. [CrossRef]
- Dubé, M., et al., Axonal transport enables neuron-to-neuron propagation of human coronavirus OC43. Journal of virology, 2018. 92(17): p. e00404-18. [CrossRef]
- Goërtz, Y.M., et al., Persistent symptoms 3 months after a SARS-CoV-2 infection: the post-COVID-19 syndrome? ERJ open research, 2020. 6(4). [CrossRef]
- Raja, C.P., et al., Science behind Usefulness of Bacopa monnieri for Memory and Cognition, in Phytopharmaceuticals for Brain Health. 2017, CRC Press. p. 225-250.
- Stefano, G.B., et al., Selective neuronal mitochondrial targeting in SARS-CoV-2 infection affects cognitive processes to induce ‘brain fog’and results in behavioral changes that favor viral survival. Medical science monitor: international medical journal of experimental and clinical research, 2021. 27: p. e930886-1. [CrossRef]
- Bliddal, S., et al., Acute and persistent symptoms in non-hospitalized PCR-confirmed COVID-19 patients. Scientific reports, 2021. 11(1): p. 13153. [CrossRef]
- Iwashyna, T.J., et al., Long-term cognitive impairment and functional disability among survivors of severe sepsis. Jama, 2010. 304(16): p. 1787-1794. [CrossRef]
- Semmler, A., et al., Persistent cognitive impairment, hippocampal atrophy and EEG changes in sepsis survivors. Journal of Neurology, Neurosurgery & Psychiatry, 2013. 84(1): p. 62-69. [CrossRef]
- Rass, V., et al., Neurological outcome and quality of life 3 months after COVID-19: A prospective observational cohort study. European journal of neurology, 2021. 28(10): p. 3348-3359. [CrossRef]
- Carfì, A., R. Bernabei, and F. Landi, Persistent symptoms in patients after acute COVID-19. Jama, 2020. 324(6): p. 603-605. [CrossRef]
- Helms, J., et al., Neurologic features in severe SARS-CoV-2 infection. New England Journal of Medicine, 2020. 382(23): p. 2268-2270. [CrossRef]
- Mao, L., et al., Neurologic manifestations of hospitalized patients with coronavirus disease 2019 in Wuhan, China. JAMA neurology, 2020. 77(6): p. 683-690. [CrossRef]
- Mazza, M.G., et al., Persistent psychopathology and neurocognitive impairment in COVID-19 survivors: effect of inflammatory biomarkers at three-month follow-up. Brain, behavior, and immunity, 2021. 94: p. 138-147. [CrossRef]
- Hellmuth, J., et al., Persistent COVID-19-associated neurocognitive symptoms in non-hospitalized patients. Journal of neurovirology, 2021. 27: p. 191-195. [CrossRef]
- Garrigues, E., et al., Post-discharge persistent symptoms and health-related quality of life after hospitalization for COVID-19. Journal of Infection, 2020. 81(6): p. e4-e6. [CrossRef]
- Han, Q., et al., Long-term sequelae of COVID-19: a systematic review and meta-analysis of one-year follow-up studies on post-COVID symptoms. Pathogens, 2022. 11(2): p. 269. [CrossRef]
- Ritchie, K. and D. Chan, The emergence of cognitive COVID. World Psychiatry, 2021. 20(1): p. 52. [CrossRef]
- Zhou, H., et al., The landscape of cognitive function in recovered COVID-19 patients. Journal of psychiatric research, 2020. 129: p. 98-102. [CrossRef]
- Raman, B., et al., Medium-term effects of SARS-CoV-2 infection on multiple vital organs, exercise capacity, cognition, quality of life and mental health, post-hospital discharge. EClinicalMedicine, 2021. 31: p. 100683. [CrossRef]
- Woo, M.S., et al., Frequent neurocognitive deficits after recovery from mild COVID-19. Brain communications, 2020. 2(2): p. fcaa205. [CrossRef]
- Kumar, V., et al., The incidence of anosmia in patients with laboratory-confirmed COVID 19 infection in India: An observational study. Journal of Anaesthesiology, Clinical Pharmacology, 2021. 37(1): p. 51. [CrossRef]
- Jaywant, A., et al., Frequency and profile of objective cognitive deficits in hospitalized patients recovering from COVID-19. Neuropsychopharmacology, 2021. 46(13): p. 2235-2240. [CrossRef]
- Villani, E.R., et al., Impact of COVID-19-related lockdown on psychosocial, cognitive, and functional well-being in adults with down syndrome. Frontiers in Psychiatry, 2020. 11: p. 578686. [CrossRef]
- Zhou, J., et al., Cognitive disorders associated with hospitalization of COVID-19: Results from an observational cohort study. Brain, Behavior, and Immunity, 2021. 91: p. 383-392. [CrossRef]
- Heesakkers, H., et al., Clinical outcomes among patients with 1-year survival following intensive care unit treatment for COVID-19. Jama, 2022. 327(6): p. 559-565. [CrossRef]
- Del Brutto, O.H., et al., Cognitive decline among individuals with history of mild symptomatic SARS-CoV-2 infection: A longitudinal prospective study nested to a population cohort. European journal of neurology, 2021. 28(10): p. 3245-3253. [CrossRef]
- Cristillo, V., et al., Premorbid vulnerability and disease severity impact on Long-COVID cognitive impairment. Aging clinical and experimental research, 2022: p. 1-4. [CrossRef]
- Hoffmann, M., et al., SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. cell, 2020. 181(2): p. 271-280. e8. [CrossRef]
- Sparks, M.A., et al., Classical renin-angiotensin system in kidney physiology. Comprehensive Physiology, 2014. 4(3): p. 1201. [CrossRef]
- Shabani, Z., Demyelination as a result of an immune response in patients with COVID-19. Acta Neurologica Belgica, 2021. 121(4): p. 859-866. [CrossRef]
- Doobay, M.F., et al., Differential expression of neuronal ACE2 in transgenic mice with overexpression of the brain renin-angiotensin system. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology, 2007. 292(1): p. R373-R381. [CrossRef]
- Baig, A.M., et al., Evidence of the COVID-19 virus targeting the CNS: tissue distribution, host–virus interaction, and proposed neurotropic mechanisms. ACS chemical neuroscience, 2020. 11(7): p. 995-998. [CrossRef]
- Wang, Q., et al., Structural and functional basis of SARS-CoV-2 entry by using human ACE2. Cell, 2020. 181(4): p. 894-904. e9. [CrossRef]
- Seyran, M., et al., The structural basis of accelerated host cell entry by SARS-CoV-2. The FEBS journal, 2021. 288(17): p. 5010-5020. [CrossRef]
- Iadecola, C., J. Anrather, and H. Kamel, Effects of COVID-19 on the nervous system. Cell, 2020. 183(1): p. 16-27. e1. [CrossRef]
- Davies, J., et al., Neuropilin-1 as a new potential SARS-CoV-2 infection mediator implicated in the neurologic features and central nervous system involvement of COVID-19. Molecular medicine reports, 2020. 22(5): p. 4221-4226. [CrossRef]
- Ulrich, H. and M.M. Pillat, CD147 as a target for COVID-19 treatment: suggested effects of azithromycin and stem cell engagement. Stem cell reviews and reports, 2020. 16(3): p. 434-440. [CrossRef]
- Istifli, E.S., et al., Interaction of certain monoterpenoid hydrocarbons with the receptor binding domain of 2019 novel coronavirus (2019-nCoV), transmembrane serine protease 2 (TMPRSS2), cathepsin B, and cathepsin L (CatB/L) and their pharmacokinetic properties. Turkish Journal of Biology, 2020. 44(7): p. 242-264. [CrossRef]
- Shang, J., et al., Cell entry mechanisms of SARS-CoV-2. Proceedings of the National Academy of Sciences, 2020. 117(21): p. 11727-11734. [CrossRef]
- Shabani, Z., J. Schuerger, and H. Su, Cellular loci involved in the development of brain arteriovenous malformations. Frontiers in Human Neuroscience, 2022. [CrossRef]
- Osburn, W.O., et al., Markers of endothelial cell activation are associated with the severity of pulmonary disease in COVID-19. Plos one, 2022. 17(5): p. e0268296. [CrossRef]
- Rotoli, B.M., et al., Endothelial cell activation by sars-cov-2 spike s1 protein: A crosstalk between endothelium and innate immune cells. Biomedicines, 2021. 9(9): p. 1220. [CrossRef]
- Szekely, L., et al., Pulmonary stromal expansion and intra-alveolar coagulation are primary causes of COVID-19 death. Heliyon, 2021. 7(5): p. e07134. [CrossRef]
- Bhatnagar, J., et al., Evidence of severe acute respiratory syndrome coronavirus 2 replication and tropism in the lungs, airways, and vascular endothelium of patients with fatal coronavirus disease 2019: an autopsy case series. The Journal of infectious diseases, 2021. 223(5): p. 752-764. [CrossRef]
- Hamming, I., et al., Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. The Journal of Pathology: A Journal of the Pathological Society of Great Britain and Ireland, 2004. 203(2): p. 631-637. [CrossRef]
- Chen, L. and G. Hao, The role of angiotensin-converting enzyme 2 in coronaviruses/influenza viruses and cardiovascular disease. Cardiovascular research, 2020. 116(12): p. 1932-1936. [CrossRef]
- Wang, K., et al., SARS-CoV-2 invades host cells via a novel route: CD147-spike protein. biorxiv, 2020: p. 2020.03. 14.988345. [CrossRef]
- Maccio, U., et al., SARS-CoV-2 leads to a small vessel endotheliitis in the heart. EBioMedicine, 2021. 63: p. 103182. [CrossRef]
- Choudhary, S., et al., Modeling SARS-CoV-2: comparative pathology in rhesus macaque and Golden Syrian hamster models. Toxicologic Pathology, 2022. 50(3): p. 280-293. [CrossRef]
- Adesse, D., et al., Role of aging in Blood–Brain Barrier dysfunction and susceptibility to SARS-CoV-2 infection: impacts on neurological symptoms of COVID-19. Fluids and Barriers of the CNS, 2022. 19(1): p. 63. [CrossRef]
- Motta, C.S., et al., Human Brain Microvascular Endothelial Cells Exposure to SARS-CoV-2 Leads to Inflammatory Activation through NF-κB Non-Canonical Pathway and Mitochondrial Remodeling. Viruses, 2023. 15(3): p. 745. [CrossRef]
- Qin, Z., et al., Endothelial cell infection and dysfunction, immune activation in severe COVID-19. Theranostics, 2021. 11(16): p. 8076. [CrossRef]
- Werlein, C., et al., Inflammation and vascular remodeling in COVID-19 hearts. Angiogenesis, 2022: p. 1-16. [CrossRef]
- Potje, S.R., et al., Heparin prevents in vitro glycocalyx shedding induced by plasma from COVID-19 patients. Life sciences, 2021. 276: p. 119376. [CrossRef]
- du Preez, H.N., et al., Pathogenesis of COVID-19 described through the lens of an undersulfated and degraded epithelial and endothelial glycocalyx. The FASEB Journal, 2022. 36(1): p. e22052. [CrossRef]
- Targosz-Korecka, M., et al., Endothelial glycocalyx shields the interaction of SARS-CoV-2 spike protein with ACE2 receptors. Scientific reports, 2021. 11(1): p. 12157. [CrossRef]
- Vollenberg, R., et al., Indications of persistent glycocalyx damage in convalescent COVID-19 patients: a prospective multicenter study and hypothesis. Viruses, 2021. 13(11): p. 2324. [CrossRef]
- Langen, U.H., S. Ayloo, and C. Gu, Development and cell biology of the blood-brain barrier. Annual review of cell and developmental biology, 2019. 35: p. 591-613. [CrossRef]
- Greene, C. and M. Campbell, Tight junction modulation of the blood brain barrier: CNS delivery of small molecules. Tissue barriers, 2016. 4(1): p. e1138017. [CrossRef]
- Bleau, C., et al., Brain invasion by mouse hepatitis virus depends on impairment of tight junctions and beta interferon production in brain microvascular endothelial cells. Journal of virology, 2015. 89(19): p. 9896-9908. [CrossRef]
- Alquisiras-Burgos, I., et al., Neurological complications associated with the blood-brain barrier damage induced by the inflammatory response during SARS-CoV-2 infection. Molecular neurobiology, 2021. 58(2): p. 520-535. [CrossRef]
- Yang, R.-C., et al., SARS-CoV-2 productively infects human brain microvascular endothelial cells. Journal of Neuroinflammation, 2022. 19(1): p. 149. [CrossRef]
- Almutairi, M.M., et al., Factors controlling permeability of the blood–brain barrier. Cellular and molecular life sciences, 2016. 73: p. 57-77. [CrossRef]
- Ranaivo, H.R., et al., Mild stretch-induced injury increases susceptibility to interleukin-1β-induced release of matrix metalloproteinase-9 from astrocytes. Journal of neurotrauma, 2011. 28(9): p. 1757-1766. [CrossRef]
- Erickson, M.A., et al., Interactions of SARS-CoV-2 with the blood–brain barrier. International Journal of Molecular Sciences, 2021. 22(5): p. 2681. [CrossRef]
- Rauti, R., et al., Effect of SARS-CoV-2 proteins on vascular permeability. Elife, 2021. 10: p. e69314. [CrossRef]
- Erickson, M.A. and W.A. Banks, Neuroimmune axes of the blood–brain barriers and blood–brain interfaces: bases for physiological regulation, disease states, and pharmacological interventions. Pharmacological reviews, 2018. 70(2): p. 278-314. [CrossRef]
- Kilic, E., et al., The phosphatidylinositol-3 kinase/Akt pathway mediates VEGF's neuroprotective activity and induces blood brain barrier permeability after focal cerebral ischemia. FASEB journal, 2006. 20(8): p. 1185. [CrossRef]
- Ahmad, S.J., et al., Neurological sequelae of COVID-19. Journal of Integrative Neuroscience, 2022. 21(3): p. 77. [CrossRef]
- Staekenborg, S.S., et al., Neurological signs in relation to type of cerebrovascular disease in vascular dementia. Stroke, 2008. 39(2): p. 317-322. [CrossRef]
- Miners, S., P.G. Kehoe, and S. Love, Cognitive impact of COVID-19: looking beyond the short term. Alzheimer's research & therapy, 2020. 12(1): p. 1-16. [CrossRef]
- Loo, J., D.A. Spittle, and M. Newnham, COVID-19, immunothrombosis and venous thromboembolism: biological mechanisms. Thorax, 2021. 76(4): p. 412-420. [CrossRef]
- Magro, C., et al., Complement associated microvascular injury and thrombosis in the pathogenesis of severe COVID-19 infection: a report of five cases. Translational Research, 2020. 220: p. 1-13. [CrossRef]
- Zuo, Y., et al., Neutrophil extracellular traps in COVID-19. JCI insight, 2020. 5(11). [CrossRef]
- Harris, A.G. and T.C. Skalak, Leukocyte cytoskeletal structure determines capillary plugging and network resistance. American Journal of Physiology-Heart and Circulatory Physiology, 1993. 265(5): p. H1670-H1675. [CrossRef]
- Cruz Hernández, J.C., et al., Neutrophil adhesion in brain capillaries reduces cortical blood flow and impairs memory function in Alzheimer’s disease mouse models. Nature neuroscience, 2019. 22(3): p. 413-420. [CrossRef]
- Wang, S., et al., Pericytes regulate vascular basement membrane remodeling and govern neutrophil extravasation during inflammation. 2012. [CrossRef]
- Paul, R., et al., Src deficiency or blockade of Src activity in mice provides cerebral protection following stroke. Nature medicine, 2001. 7(2): p. 222-227. [CrossRef]
- Liu, D.Z., et al., Blood–brain barrier breakdown and repair by Src after thrombin-induced injury. Annals of neurology, 2010. 67(4): p. 526-533. [CrossRef]
- Tyagi, N., et al., Fibrinogen induces endothelial cell permeability. Molecular and cellular biochemistry, 2008. 307: p. 13-22. [CrossRef]
- Trepat, X., et al., Thrombin and histamine induce stiffening of alveolar epithelial cells. Journal of Applied Physiology, 2005. 98(4): p. 1567-1574. [CrossRef]
- Yepes, M., et al., Tissue-type plasminogen activator induces opening of the blood-brain barrier via the LDL receptor–related protein. The Journal of clinical investigation, 2003. 112(10): p. 1533-1540. [CrossRef]
- Grammas, P., P.G. Samany, and L. Thirumangalakudi, Thrombin and inflammatory proteins are elevated in Alzheimer's disease microvessels: implications for disease pathogenesis. Journal of Alzheimer's Disease, 2006. 9(1): p. 51-58. [CrossRef]
- Shah, F.A., et al., Bidirectional relationship between cognitive function and pneumonia. American journal of respiratory and critical care medicine, 2013. 188(5): p. 586-592. [CrossRef]
- Salive, M.E., et al., Disability and cognitive impairment are risk factors for pneumonia-related mortality in older adults. Public Health Reports, 1993. 108(3): p. 314.
- Herridge, M.S., et al., Recovery and outcomes after the acute respiratory distress syndrome (ARDS) in patients and their family caregivers. Intensive care medicine, 2016. 42: p. 725-738. [CrossRef]
- Tate, J.A., et al., Infection hospitalization increases risk of dementia in the elderly. Critical care medicine, 2014. 42(5): p. 1037. [CrossRef]
- Wilcox, M.E., et al., Cognitive dysfunction in ICU patients: risk factors, predictors, and rehabilitation interventions. Critical care medicine, 2013. 41(9): p. S81-S98. [CrossRef]
- Cervos-Navarro, J. and N. Diemer, Selective vulnerability in brain hypoxia. Critical reviews in neurobiology, 1991. 6(3): p. 149-182.
- Moskowitz, M.A., E.H. Lo, and C. Iadecola, The science of stroke: mechanisms in search of treatments. Neuron, 2010. 67(2): p. 181-198. [CrossRef]
- Fernando, M.S., et al., White matter lesions in an unselected cohort of the elderly: molecular pathology suggests origin from chronic hypoperfusion injury. Stroke, 2006. 37(6): p. 1391-1398. [CrossRef]
- Yang, Y. and G.A. Rosenberg, Blood–brain barrier breakdown in acute and chronic cerebrovascular disease. Stroke, 2011. 42(11): p. 3323-3328. [CrossRef]
- Esch, T., et al., Emerging roles of blood-borne intact and respiring mitochondria as bidirectional mediators of pro-and anti-inflammatory processes. Medical Science Monitor: International Medical Journal of Experimental and Clinical Research, 2020. 26: p. e924337-1. [CrossRef]
- Shenoy, S., Coronavirus (Covid-19) sepsis: revisiting mitochondrial dysfunction in pathogenesis, aging, inflammation, and mortality. Inflammation research, 2020. 69: p. 1077-1085. [CrossRef]
- Singh, K.K., et al., Decoding SARS-CoV-2 hijacking of host mitochondria in COVID-19 pathogenesis. American Journal of Physiology-Cell Physiology, 2020. [CrossRef]
- Ptacek, R., et al., Psychiatric manifestations of COVID-19 and their social significance. Medical Science Monitor: International Medical Journal of Experimental and Clinical Research, 2020. 26: p. e930340-1. [CrossRef]
- Bostancıklıoğlu, M., SARS-CoV2 entry and spread in the lymphatic drainage system of the brain. Brain, behavior, and immunity, 2020. 87: p. 122. [CrossRef]
- Steardo, L., et al., Neuroinfection may contribute to pathophysiology and clinical manifestations of COVID-19. Acta Physiologica (Oxford, England), 2020. 229(3). [CrossRef]
- Bellon, M., et al., Cerebrospinal fluid features in severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) reverse transcription polymerase chain reaction (RT-PCR) positive patients. Clinical Infectious Diseases, 2021. 73(9): p. e3102-e3105. [CrossRef]
- Geng, J., et al., Blood-brain barrier disruption induced cognitive impairment is associated with increase of inflammatory cytokine. Frontiers in aging neuroscience, 2018. 10: p. 129. [CrossRef]
- Montagne, A., et al., APOE4 leads to blood–brain barrier dysfunction predicting cognitive decline. Nature, 2020. 581(7806): p. 71-76. [CrossRef]
- Wu, Y., et al., Nervous system involvement after infection with COVID-19 and other coronaviruses. Brain, behavior, and immunity, 2020. 87: p. 18-22. [CrossRef]
- Anwar, M.M., Immunotherapies and COVID-19 related neurological manifestations: A comprehensive review article. Journal of Immunoassay and Immunochemistry, 2020. 41(6): p. 960-975. [CrossRef]
- Alnefeesi, Y., et al., Impact of SARS-CoV-2 infection on cognitive function: a systematic review. Frontiers in Psychiatry, 2021: p. 1629. [CrossRef]
- Gao, W., et al., Systematic analysis of chemokines reveals CCL18 is a prognostic biomarker in glioblastoma. Journal of Inflammation Research, 2022: p. 2731-2743. [CrossRef]
- Kolodziej, A., et al., Tonic activation of CXC chemokine receptor 4 in immature granule cells supports neurogenesis in the adult dentate gyrus. Journal of Neuroscience, 2008. 28(17): p. 4488-4500. [CrossRef]
- Senf, K., et al., Chemokine signaling is required for homeostatic and injury-induced neurogenesis in the olfactory epithelium. Stem Cells, 2021. 39(5): p. 617-635. [CrossRef]
- Karimabad, M.N., et al., The Chemokines CXC, CC and C in the Pathogenesis of COVID-19 Disease and as Surrogates of Vaccine-Induced Innate and Adaptive Protective Responses. Vaccines, 2022. 10(8): p. 1299. [CrossRef]
- Yasui, H., et al., CCL2 secreted from cancer-associated mesothelial cells promotes peritoneal metastasis of ovarian cancer cells through the P38-MAPK pathway. Clinical & Experimental Metastasis, 2020. 37: p. 145-158. [CrossRef]
- Villeda, S.A., et al., The ageing systemic milieu negatively regulates neurogenesis and cognitive function. Nature, 2011. 477(7362): p. 90-94. [CrossRef]
- Minton, K., Mechanistic insights into Long COVID in hamsters. Nature Reviews Immunology, 2022. 22(8): p. 463-463. [CrossRef]
- Hong, S., et al., Abnormalities in chemokine levels in schizophrenia and their clinical correlates. Schizophrenia research, 2017. 181: p. 63-69. [CrossRef]
- Fernández-Castañeda, A., et al., Mild respiratory SARS-CoV-2 infection can cause multi-lineage cellular dysregulation and myelin loss in the brain. BioRxiv, 2022. [CrossRef]
- Xu, J., et al., Astrocyte-derived CCL2 participates in surgery-induced cognitive dysfunction and neuroinflammation via evoking microglia activation. Behavioural Brain Research, 2017. 332: p. 145-153. [CrossRef]
- Thirumangalakudi, L., et al., Angiogenic proteins are expressed by brain blood vessels in Alzheimer's disease. Journal of Alzheimer's Disease, 2006. 10(1): p. 111-118. [CrossRef]
- Teuwen, L.-A., et al., COVID-19: the vasculature unleashed. Nature Reviews Immunology, 2020. 20(7): p. 389-391. [CrossRef]
- Varga, Z., et al., Endothelial cell infection and endotheliitis in COVID-19. The Lancet, 2020. 395(10234): p. 1417-1418. [CrossRef]
Figure 1.
Schematic representation of SARS-CoV-2 infection causing vascular dysfunction and BBB disruption.
Figure 1.
Schematic representation of SARS-CoV-2 infection causing vascular dysfunction and BBB disruption.
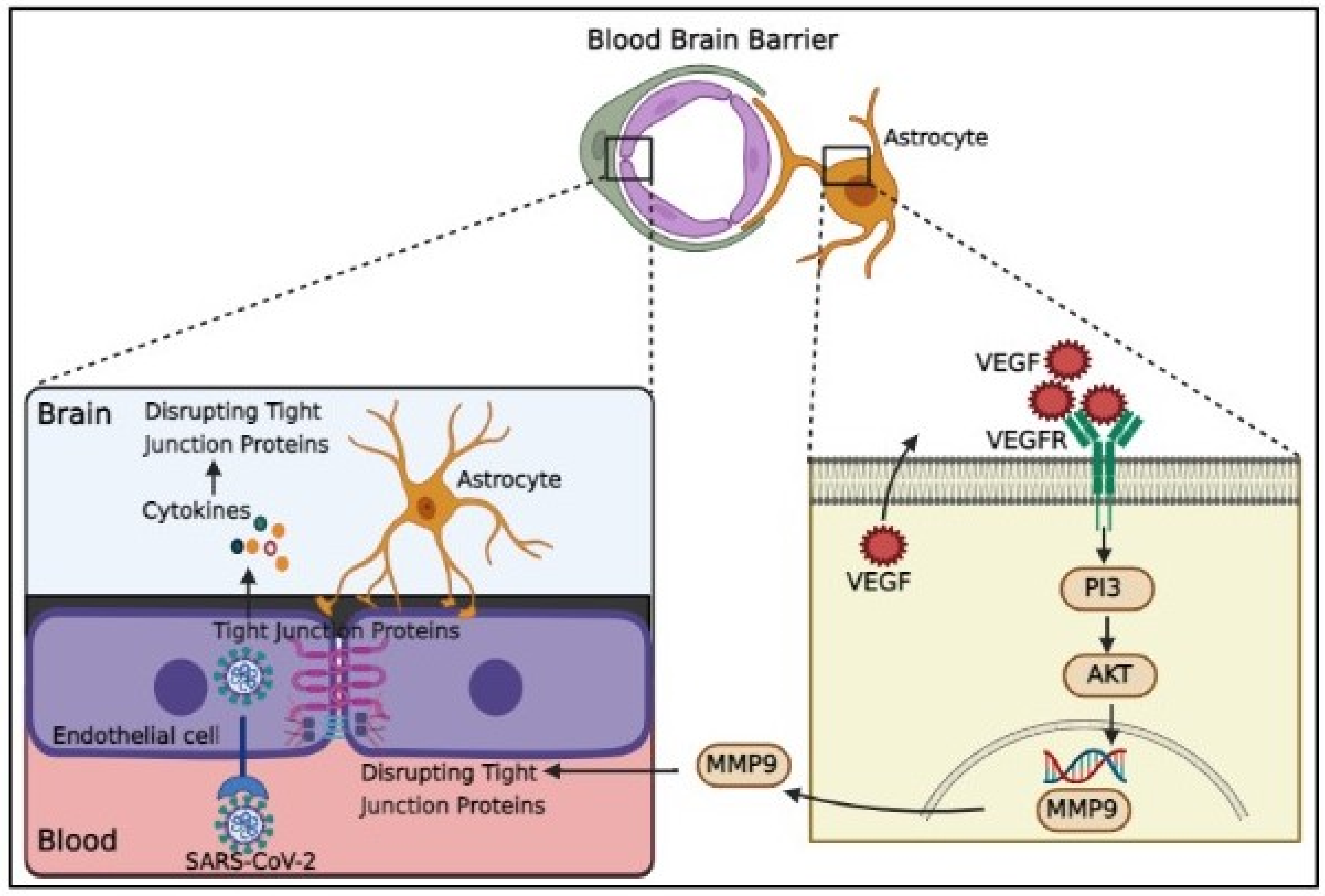
Figure 2.
Schematic representation of indirect BBB disruption resulting from vascular dysfunction in COVID-19. The virus acts via binding of spike glycoprotein of the virus to the angiotensin converting enzyme 2 (ACE2) on cell surface. Disharmonic and disturbed immune reaction in COVID-19 patients recruit a wide local and systemic inflammation by generating cytokine storm, leading to ARDS and vascular coagulation. Alternatively, it led to the formation of neutrophil extracellular traps (NETs), composed of chromatin and microbicidal proteins, which participate in the pathobiology of thrombosis and platelets aggregation. Activation of complement system results in the generation of C5a and C5b, which are involved in the inflammation and activation of the platelets. This C5b forms the membrane attack complex (MAC) to activate the microvascular coagulation, and inflammation.
Figure 2.
Schematic representation of indirect BBB disruption resulting from vascular dysfunction in COVID-19. The virus acts via binding of spike glycoprotein of the virus to the angiotensin converting enzyme 2 (ACE2) on cell surface. Disharmonic and disturbed immune reaction in COVID-19 patients recruit a wide local and systemic inflammation by generating cytokine storm, leading to ARDS and vascular coagulation. Alternatively, it led to the formation of neutrophil extracellular traps (NETs), composed of chromatin and microbicidal proteins, which participate in the pathobiology of thrombosis and platelets aggregation. Activation of complement system results in the generation of C5a and C5b, which are involved in the inflammation and activation of the platelets. This C5b forms the membrane attack complex (MAC) to activate the microvascular coagulation, and inflammation.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



