
Submitted:
01 June 2023
Posted:
02 June 2023
You are already at the latest version
Abstract
The location of Israel at the junction of three continents leads to a unique faunal combination of Palearctic and Afrotropic zoogeographic origins. Following systematic revisions over the past sixty years and the discovery of new species, the only available key to the pseudoscorpions (Arachnida: Pseudoscorpiones) of Israel (Beier 1963) has become outdated. We provide here an up-to-date checklist of the pseudoscorpion species of Israel including distribution maps, and the first illustrated identification key of the Israeli fauna based on morphological characters. Prior to our study this fauna comprised twelve families, 26 genera and 52 morphospecies, including several “subspecies”. We increase this number and list 61 pseudoscorpion morphospecies that belong to 28 genera and fourteen families. Most species are Palearctic and Mediterranean, and only a few are Afrotropic. Two families new to Israel are reported here for the first time: Syarinidae and Cheiridiidae. Both families are cosmopolitan and have representatives in the Mediterranean region. The putative new species are presented here at a genus level and will be described separately elsewhere.
Keywords:
Cheiridiidae
; False scorpions
; Levant
; Species List
; Syarinidae
; Taxonomy
; Zoogeography
1. Introduction
Pseudoscorpiones is an order of arachnids that comprises 25 families, 472 genera, and more than 4120 species worldwide [1]. They occupy diverse terrestrial habitats such as leaf litter, soil, under stones and logs, in mosses and lichens, under the bark of trees and in caves, as well as inside nests of birds, rodents or insects which they often use as phoretic hosts [2]. Being predators of various soil arthropods, they are important in the soil food web and affect arthropod population dynamics [3]. High diversity of pseudoscorpion species is found in the Mediterranean region, both because the area served as one of the Pleistocene glacial refugia [4,5], and because karstic systems harbour additional cave-adapted species [2,5,6].
Israel’s geographic location in the south-east Mediterranean coast at the junction of three continents (Europe, Asia, and Africa) forms a diverse biogeographic unit with a unique faunal combination of Palearctic and Afrotropic zoogeographic origins. This unit was formed by the crossroads between north-eastern African and north-western Arabian plates and the eastern Mediterranean Levantine basin. Climatically, Israel is situated in the subtropical drylands, between 29.5° and 33.5° N, and is characterized by a single rainy season (November-March) and a hot and dry summer [7]. Despite its small area (ca. 27900km2 including Palestine; Figure 1), Israel experiences sharp climatic gradients, both in the north-south and the west-east directions dividing it to five main climatic regions according to the rainfall gradient: mesic alpine climate at the north-east tip (Mt. Hermon); Mediterranean climate in north and central Israel; semi-arid steppe climate in the south and east of Israel (including the West Bank); arid desert Saharo-Arabian climate in the south of Israel; and the coastal plain, which is susceptible to influence from the Mediterranean [8,9]. The combination of the different zoogeographic regions in addition to Israel’s heterogeneous climate regions, topography, geology, hydrology and soil, lead to a high diversity of biological habitats. These conditions are ideal for rich arachnid diversity [4,10] including relict and endemic species with unique evolutionary history [11,12,13,14].
Pseudoscorpion research in Israel has hardly existed until recently. All the taxonomic work was done outside the country [13,15,16,17] and a methodological survey was never conducted. The first and only extensive taxonomic study of pseudoscorpions was conducted by Austrian zoologist Max Beier who listed a total of 31 species belonging to seven families based on ad hoc collections in Israel [15]. Additional species were identified and described infrequently thereafter [13,16,17], bringing the total count to 52 species (including subspecies) and twelve families [1,18,19].
The primary aim of this paper is to present an updated checklist of the pseudoscorpion fauna of Israel based on examination of all the collection material at the Israel National Arachnid Collection at the Hebrew University of Jerusalem (HUJ), and the identification of newly collected specimens from recent surveys. We also present distribution maps of all species, analyze their records according to season, habitat, and collecting methods, as well as localities in the country in relation to zoogeographic world distribution. Based on this data we present a key for the families and genera occurring in Israel. While the new family records for Israel are reported here, the putative new species will be described separately elsewhere.
2. Materials and Methods
More than 1680 pseudoscorpion specimens were examined, of which ca. 1100 that have been deposited at the Israel Arachnid National Natural History Collection (NNHC) at the Hebrew University of Jerusalem (HUJ) over decades, and ca. 580 fresh specimens collected from agroecosystems, caves and other natural habitats, in recent years.
Specimens were collected by means of pitfall traps, trunk traps (that were attached to tree trunks [20]), visual search (under stones, rocks or tree bark), and sifting (leaf litter and soil). We also include in this checklist species and subspecies that are mentioned in published records or in the World Pseudoscorpiones Catalog [1] but were not deposited in the collection (NNHC, HUJ).
The specimens were examined using a Nikon SMZ25 motorized stereomicroscope or a Bausch & Lomb stereo zoom 7, with a 15 x W.F. stereo lens. Digital images were taken using a Nikon DS Fi2 digital camera mounted on a Nikon SMZ25 stereomicroscope driven by NIS-Elements D. v. 4.20 software (Nikon). Image stacks were combined using Zerene Stacker (Version 1.04). For finer details, certain specimens were examined after clearing with lactic acid or clove oil as temporary glycerine mounts in cavity slides using Olympus BX60 compound microscope. Live specimens were photographed with a Cannon 700D camera equipped with a macro lens. Historical material is preserved in 75% ethanol at room temperature, while freshly collected material is preserved in −20 °C freezer either in 75% for further morphological study or in 99% ethanol for further molecular study. Specimens were identified using taxonomic literature, mainly Beier’s key [15]. Morphological terminology and trichobothrial terminology follows Beier [15]. The taxonomic ranks are according to Benavides et al. [19]. Nomenclature follows WPC [1].
Distribution maps were generated using ArcGIS Pro. Geographic coordinates are given in WGS84. A scheme often used for biogeographical studies in Israel is based on geomorphological data that divides Israel and adjacent areas into 22 biogeographical units (Figure 1a) [21]. In order to visualize the distribution patterns clearly, we grouped together several small units to create eleven zoographical sub-regions that would correspond to the combination of the topographic, climatic and zoogeographic zones discussed in the introduction (Figure 1b, Figure 2).
Figure 1.
Maps of Israel and adjacent countries. (a) Scheme of biogeographical units in Israel and adjacent countries (adopted from Klein 1988): 1. Upper Galilee; 2. Lower Galilee; 3. Carmel Ridge; 4. Northern Coastal Plain; 5. Jezreel Valley; 6. Samaria; 7. Jordan Valley and Sea of Galilee; 8. Central Coastal Plain; 9. Southern Coastal Plain; 10. Judea Foothills; 11. Judean Hills; 12. Judean Desert; 13. Dead Sea Area; 14. ‘Arava Valley; 15. Northern Negev; 16. Southern Negev; 17. Central Negev; 18. Golan Heights; 19. Mount Hermon; 20-22. Sinai (Egypt): (Northern Sinai; Central Sinai Foothills; Sinai Mountains). (b) Map of biogeographic sub-regions in Israel and adjacent countries, that was used by us in this publication (numbers in parentheses relate to biogeographical units in Figure 1a): 1. Hermon (19) 2. Golan (18) 3. Galilee (1 & 2) 4. Hula Valley (included in 1) 5. Coastal Plain (4, 8 & 9) 6. Jezreel Valley (5) 7. Central Mountains (Carmel, Samaria, Judea hills and Judea foothills; 3, 6, 10 & 11) 8. Dead Sea and Jordan Valley (7 & 13) 9. Judean Desert (12) 10. Negev Desert (Northern Negev, Central Negev, Southern Negev; 15, 16 & 17) 11. ‘Arava Valley (14).
Figure 1.
Maps of Israel and adjacent countries. (a) Scheme of biogeographical units in Israel and adjacent countries (adopted from Klein 1988): 1. Upper Galilee; 2. Lower Galilee; 3. Carmel Ridge; 4. Northern Coastal Plain; 5. Jezreel Valley; 6. Samaria; 7. Jordan Valley and Sea of Galilee; 8. Central Coastal Plain; 9. Southern Coastal Plain; 10. Judea Foothills; 11. Judean Hills; 12. Judean Desert; 13. Dead Sea Area; 14. ‘Arava Valley; 15. Northern Negev; 16. Southern Negev; 17. Central Negev; 18. Golan Heights; 19. Mount Hermon; 20-22. Sinai (Egypt): (Northern Sinai; Central Sinai Foothills; Sinai Mountains). (b) Map of biogeographic sub-regions in Israel and adjacent countries, that was used by us in this publication (numbers in parentheses relate to biogeographical units in Figure 1a): 1. Hermon (19) 2. Golan (18) 3. Galilee (1 & 2) 4. Hula Valley (included in 1) 5. Coastal Plain (4, 8 & 9) 6. Jezreel Valley (5) 7. Central Mountains (Carmel, Samaria, Judea hills and Judea foothills; 3, 6, 10 & 11) 8. Dead Sea and Jordan Valley (7 & 13) 9. Judean Desert (12) 10. Negev Desert (Northern Negev, Central Negev, Southern Negev; 15, 16 & 17) 11. ‘Arava Valley (14).
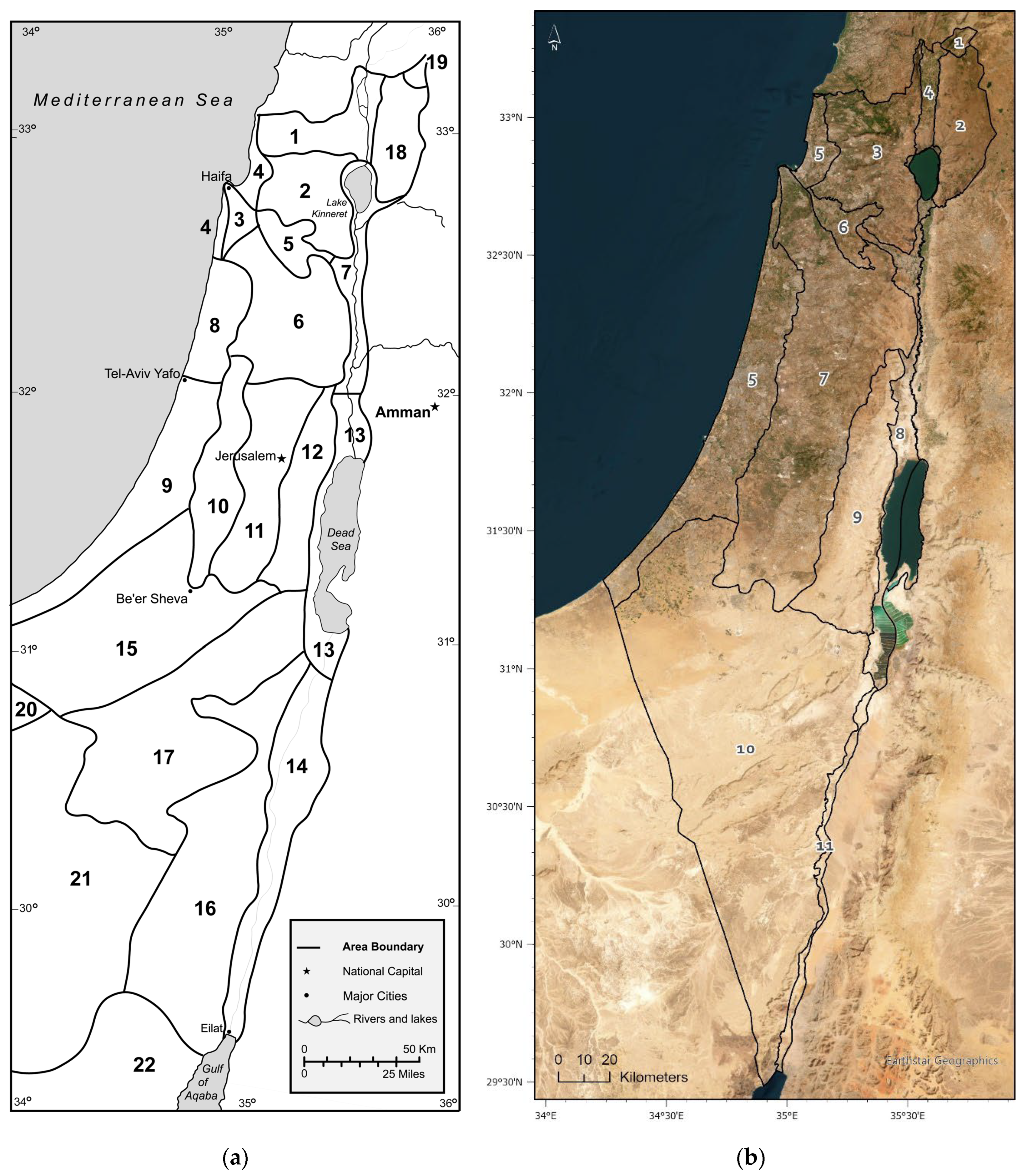
Figure 2.
Israel’s biogeographic sub-regions A. Hermon B. Golan C. Galilee D. Hula Valley E. Coastal Plain F. Jezreel Valley G. Central Mountains H. Jordan Valley I. Dead Sea J. Judean Desert K. Negev Desert L. Arava Valley. Photos by I. Armiach Steinpress.
Figure 2.
Israel’s biogeographic sub-regions A. Hermon B. Golan C. Galilee D. Hula Valley E. Coastal Plain F. Jezreel Valley G. Central Mountains H. Jordan Valley I. Dead Sea J. Judean Desert K. Negev Desert L. Arava Valley. Photos by I. Armiach Steinpress.

Biogeographical assessments of various taxonomic groups often use the approach of global chorotype classification proposed by Vigna Taglianti et al. [22] for the Near East fauna [23,24,25]. We used this concept to classify all pseudoscorpion species found in Israel into fourteen chorological categories detailed in Table 1. We further grouped these chorotypes into six chorological complexes: widely distributed species, Mediterranean, Euro-Asian, Asian, Afrotropic and species endemic to Israel. The world distributions for each species were extracted from Harvey 2013 [18] and WPC 2023 [1] and used for chorological assignment. We interpreted zoogeographical patterns using this approach.
3. Results
3.1. Systematics and Taxonomy
Based on all specimens deposited at the NNHC, HUJ (over 1600, as well as fourteen Type specimens) (Table S1), published records and the World Pseudoscorpiones Catalog [1], 61 pseudoscorpion species of 28 genera in fourteen families are now recorded from Israel. The list of the fourteen families, arranged systematically following Benavides et al. [19], is presented in Table 2. Two families are reported here for the first time from Israel: Syarinidae Chamberlin, 1930 and Cheiridiidae Hansen, 1894. Syarinidae is a widely distributed cosmopolitan family with 125 species known worldwide, most of which occur in America. Yet, several relict species are found sporadically and rarely in Europe and the Mediterranean region, with most of them being troglobionts. Such is the one we report here, which was collected in a cave in Samaria, in the Central Mountains (Figure 1b, sub-region 7). Cheiridiidae is a widely distributed cosmopolitan family with 79 species known worldwide, that has common representatives in the East Mediterranean region including Turkey (three species), Italy and Bulgaria (two species in each) and Greece, Croatia and Montenegro (one species in each), and was expected to occur also in Israel. The species we report here was collected from a cave in the Judean Desert (Figure 1b, sub-region 9).
3.1.1. Figures and Tables
Based on the results presented in the following tables and figures (Table 3 and Figure 3, Figure 4 and Figure 5), we provide hereafter information on each recorded family, followed by a taxonomic key to all families.

Table 3.
List of the pseudoscorpion species in Israel arranged by alphabetical order of family, genus and species, their record from Israel (in [ ]), chorotype classification, chorological complexes, and global zoographic category (based on Harvey [18] and WPC [1]). The chorotype classifications and respective codes, as well as their attribution to chorological complexes are detailed in Table 1. New records for Israel are marked with an asterisk *, e indicates species endemic to Israel, T indicates Type specimen deposited at the NNHC, HUJ and ¢ indicates that no specimens are deposited at the HUJ.
Table 3.
List of the pseudoscorpion species in Israel arranged by alphabetical order of family, genus and species, their record from Israel (in [ ]), chorotype classification, chorological complexes, and global zoographic category (based on Harvey [18] and WPC [1]). The chorotype classifications and respective codes, as well as their attribution to chorological complexes are detailed in Table 1. New records for Israel are marked with an asterisk *, e indicates species endemic to Israel, T indicates Type specimen deposited at the NNHC, HUJ and ¢ indicates that no specimens are deposited at the HUJ.
| Family | Species | Global Zoogeographic category | Chorotype | Chorological Complex |
|---|---|---|---|---|
| Atemnidae | ||||
| Atemnus sp. * | Palearctic and Afrotropic | OLD | Widely distributed | |
| Atemnus syriacus (Beier, 1955) ¢ [16] | Palearctic | EMED | Mediterranean | |
| Cheiridiidae * | ||||
| Apocheiridium sp. * | Palearctic | COS | Widely distributed | |
| Cheliferidae | ||||
| Chelifer cancroides (Linnaeus, 1758) [15] | Palearctic and Afrotropic | COS | Widely distributed | |
| Chelifer cancroides cancroides (Linnaeus, 1758) ¢ [1] | Palearctic and Afrotropic | COS | Widely distributed | |
| Dactylochelifer kussariensis (Daday, 1889) [15] | Palearctic | CWA | Asian | |
| Dactylochelifer kussariensis kussariensis (Daday, 1889) ¢ [1] | Palearctic | CWA | Asian | |
| Dactylochelifer pallidus Beier, 1963 e, T [15] | Endemic (Palearctic origin) | END | Endemic | |
| Hysterochelifer cyprius (Beier, 1929) [15] | Palearctic | EMED | Mediterranean | |
| Hysterochelifer distinguendus (Beier, 1929) e [39] | Endemic (Palearctic origin) | END | Endemic | |
| Hysterochelifer gracilimanus Beier, 1949 ¢ [17] | Palearctic | EMED | Mediterranean | |
| Hysterochelifer tuberculatus(Lucas, 1849) [15] | Palearctic | MED | Mediterranean | |
| Hysterochelifer tuberculatus tuberculatus (Lucas, 1849) ¢ [1] | Palearctic | MED | Mediterranean | |
| Rhacochelifer corcyrensis (Beier, 1930) ¢ [17] | Palearctic | MED | Mediterranean | |
| Rhacochelifer corcyrensis bicolor Beier, 1963 T [15] | Palearctic | LEV | Mediterranean | |
| Rhacochelifer corcyrensis corcyrensis (Beier, 1930) ¢ [17] | Palearctic | EMED | Mediterranean | |
| Rhacochelifer lobipes (Beier, 1929) [15] | Palearctic | EMED | Mediterranean | |
| Rhacochelifer longeunguiculatus Beier, 1963 T [15] | Afrotropic | APA | Afrotropic | |
| Rhacochelifer maculatus (L. Koch, 1873) [17] | Palearctic | MED | Mediterranean | |
| Rhacochelifer peculiaris (L. Koch, 1873) [1] | Palearctic | SEM | Mediterranean | |
| Rhacochelifer peculiaris latissimus Beier, 1963 T [15] | Palearctic | EMED | Mediterranean | |
| Chernetidae | ||||
| Allochernes masi (Navás, 1923) [15] | Palearctic | MED | Mediterranean | |
| Chernes rhodinus Beier, 1966 [16] | Palearctic | EMED | Mediterranean | |
| Chernes sp. * | Palearctic | HOL | Widely distributed | |
| Lamprochernes nodosus (Schrank, 1803) [15] | Palearctic and Afrotropic | OLD | Widely distributed | |
| Lamprochernes nodosus nodosus (Schrank, 1803) ¢ [1] | Palearctic and Afrotropic | OLD | Widely distributed | |
| Lamprochernes savignyi (Simon, 1881) [15] | Afrotropic | COS | Widely distributed | |
| Lasiochernes turcicus Beier, 1949 [15] | Palearctic | EMED | Mediterranean | |
| Nudochernes spalacis Beier, 1955 e, T [41] | Endemic (Afrotropic origin) | END | Endemic | |
| Pselaphochernes scorpioides (Hermann, 1804) ¢ [16] | Palearctic | HOL | Widely distributed | |
| Chthoniidae | ||||
| Ayyalonia dimentmani Ćurčić, 2008 e [13] | Endemic (Afrotropic origin) | END | Endemic | |
| Chthonius jonicus Beier, 1931 ¢ [16] | Palearctic | MED | Mediterranean | |
| Chthonius shulovi Beier, 1963 e ¢ [15] | Endemic (Palearctic origin) | END | Endemic | |
| Chthonius sp. * | Palearctic | COS | Widely distributed | |
| Ephippiochthonius sacer (Beier, 1963) e, T [15] | Endemic (Palearctic origin) | END | Endemic | |
| Ephippiochthonius tetrachelatus (Preyssler, 1790) [15] | Palearctic | COS | Widely distributed | |
| Ephippiochthonius spp. * | Palearctic | COS | Widely distributed | |
| Garypidae | ||||
| Garypus beauvoisii (Audouin, 1826) [15] | Palearctic | MED | Mediterranean | |
| Garypus levantinus Navás, 1925 [15] | Palearctic | MED | Mediterranean | |
| Garypinidae | ||||
| Garypinus asper Beier, 1955 [16] | Palearctic | EMED | Mediterranean | |
| Garypinus dimidiatus (L. Koch, 1873) [15] | Palearctic | EMED | Mediterranean | |
| Geogarypidae | ||||
| Geogarypus pulcher Beier, 1963 e, T [15] | Endemic (Palearctic origin) | END | Endemic | |
| Geogarypus shulovi Beier, 1963 T [15] | Palearctic | CWA | Asian | |
| Hesperolpiidae | ||||
| Calocheirus atopos Chamberlin, 1930 T [15] | Afrotropic | APA | Afrotropic | |
| Cardiolpium stupidum (Beier, 1963) T [15] | Palearctic | EMCA | Euro-Asia | |
| Menthidae | ||||
| Paramenthus shulovi Beier, 1963 e, T [15] | Endemic (Afrotropic origin) | END | Endemic | |
| Neobisiidae | ||||
| Neobisium (Neobisium) validum (L. Koch, 1873) [15] | Palearctic | EMCA | Euro-Asia | |
| Neobisium sp. * | Palearctic | EMCA | Euro-Asia | |
| Olpiidae | ||||
| Calocheiridius libanoticus Beier, 1955 [15] | Palearctic | EMWA | Mediterranean | |
| Halominniza aegyptiaca (Ellingsen, 1910) ¢ [1] | Palearctic | EMED | Mediterranean | |
| Halominniza aegyptiaca litoralis (Beier, 1963) [15] | Palearctic | LEV | Mediterranean | |
| Minniza babylonica Beier, 1931 [15] | Palearctic and Afrotropic | CWAP | Asian | |
| Minniza babylonica babylonica Beier, 1931 ¢ [1] | Palearctic and Afrotropic | CWAP | Asian | |
| Minniza lindbergi Beier, 1957 [15] | Palearctic | EMED | Mediterranean | |
| Olpium kochi Simon, 1881 [26] | Palearctic | EMED | Mediterranean | |
| Olpium pallipes (Lucas, 1849) [39,40] | Palearctic | MCA | Mediterranean | |
| Olpium pallipes balcanicum Beier, 1931 [15] | Palearctic | EMED | Mediterranean | |
| Olpium pallipes pallipes (Lucas, 1849) ¢ [1] | Palearctic | MCA | Mediterranean | |
| Syarinidae * | ||||
| Hadoblothrus sp. * | Palearctic | EMED | Mediterranean | |
| Withiidae | ||||
| Nannowithius wahrmani (Beier, 1963) e, T [15] | Endemic (Afrotropic origin) | END | Endemic | |
Figure 3.
The geographic distribution of the pseudoscorpion families (percentage of specimens) in each of the eleven biogeographic sub-regions of Israel (Figure 1b) (data of 1548 specimens in the collection, see Supplementary Materials Table S2).
Figure 3.
The geographic distribution of the pseudoscorpion families (percentage of specimens) in each of the eleven biogeographic sub-regions of Israel (Figure 1b) (data of 1548 specimens in the collection, see Supplementary Materials Table S2).
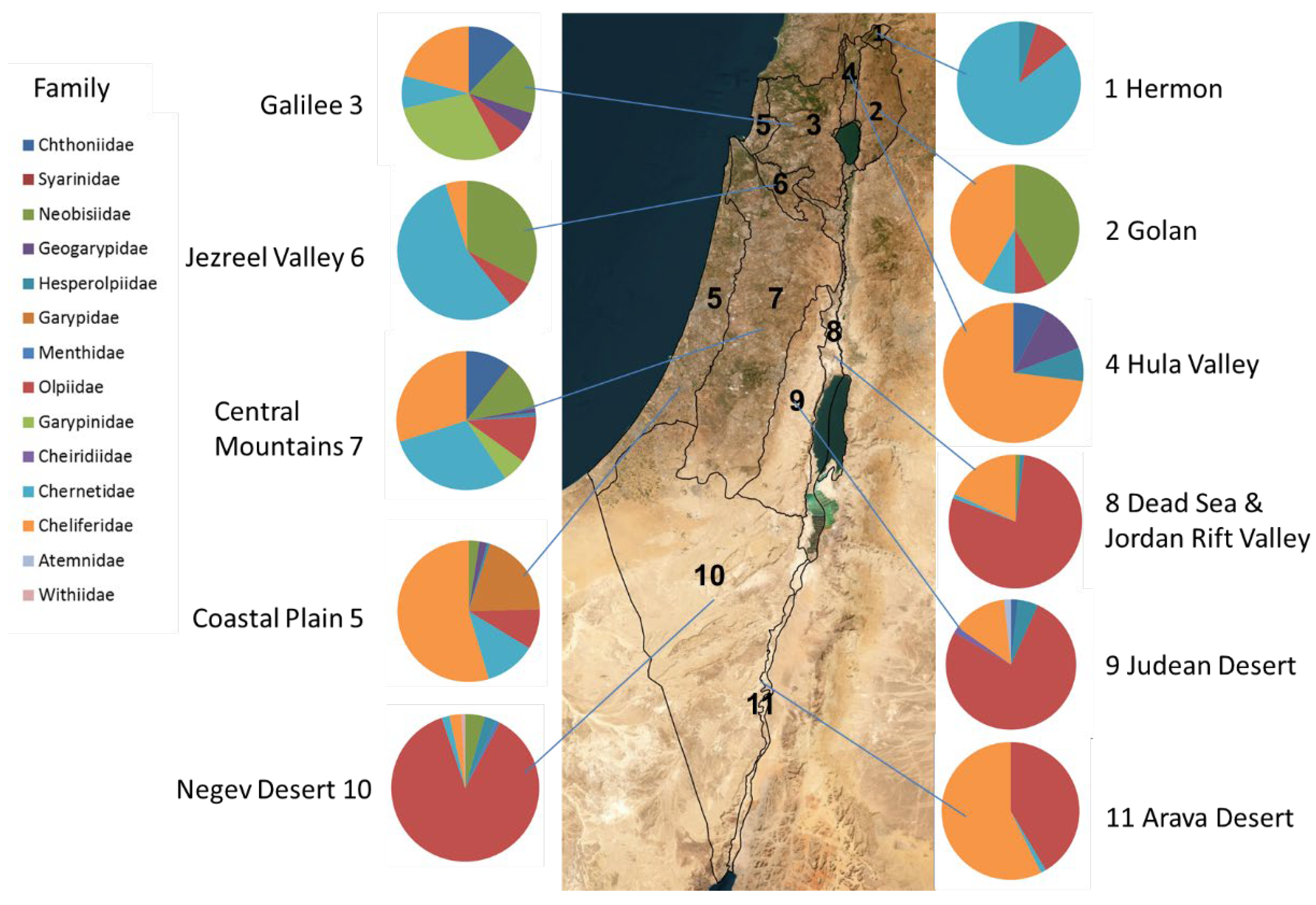
Figure 4.
Geographic distribution maps of the pseudoscorpion species in Israel. Localities of 1397 identified specimens deposited in the collection (circle) (Supplementary Materials Table S3), and localities specified in: Beier [15], Halperin & Mahnert [17], Mahnert [16] and Šťáhlavský et al. [26], (square). Endemic species are indicated by a star symbol. (a) Cheliferidae; (b) Cheliferidae; (c) Olpiidae; (d) Cheliferoidea (Atemnidae, Chernetidae and Withiidae); (e) Neobisioidea (Neobisiidae and Syarinidae); (f) Chthoniidae; (g) Cheiridiidae and Garypinidae; (h) Garypoidea (Garypidae, Geogarypidae, Hesperolpiidae and Menthidae).
Figure 4.
Geographic distribution maps of the pseudoscorpion species in Israel. Localities of 1397 identified specimens deposited in the collection (circle) (Supplementary Materials Table S3), and localities specified in: Beier [15], Halperin & Mahnert [17], Mahnert [16] and Šťáhlavský et al. [26], (square). Endemic species are indicated by a star symbol. (a) Cheliferidae; (b) Cheliferidae; (c) Olpiidae; (d) Cheliferoidea (Atemnidae, Chernetidae and Withiidae); (e) Neobisioidea (Neobisiidae and Syarinidae); (f) Chthoniidae; (g) Cheiridiidae and Garypinidae; (h) Garypoidea (Garypidae, Geogarypidae, Hesperolpiidae and Menthidae).
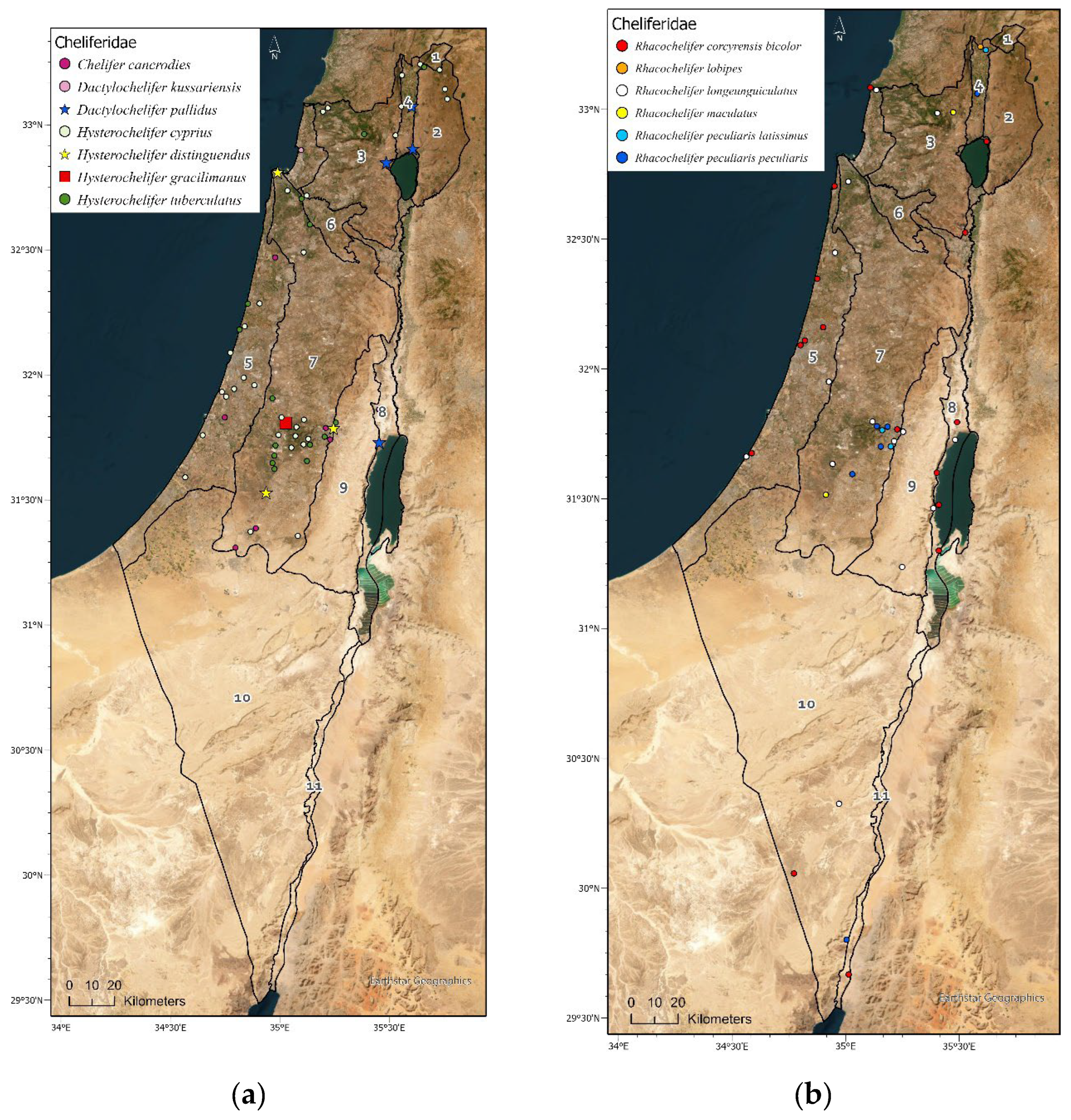
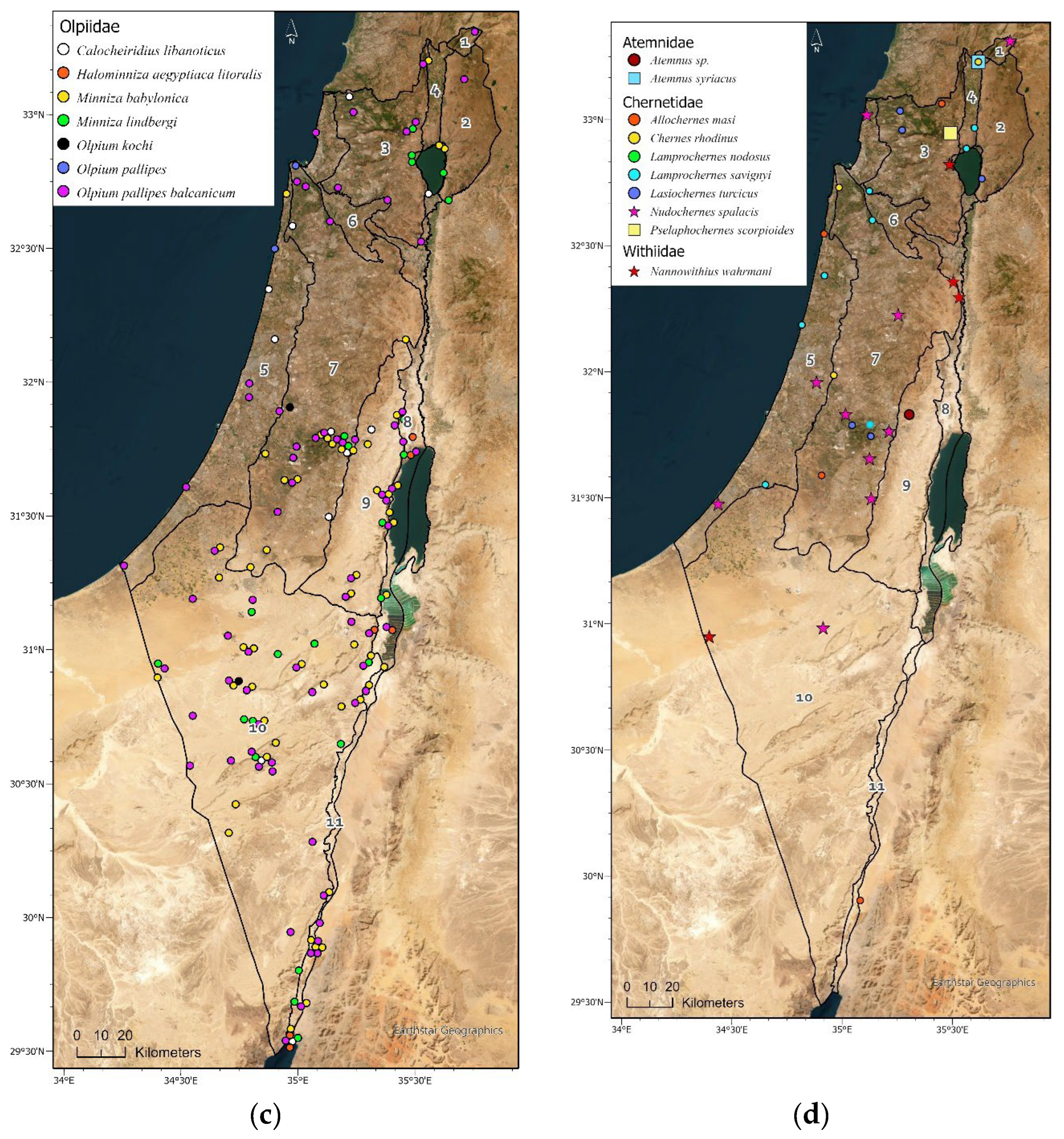
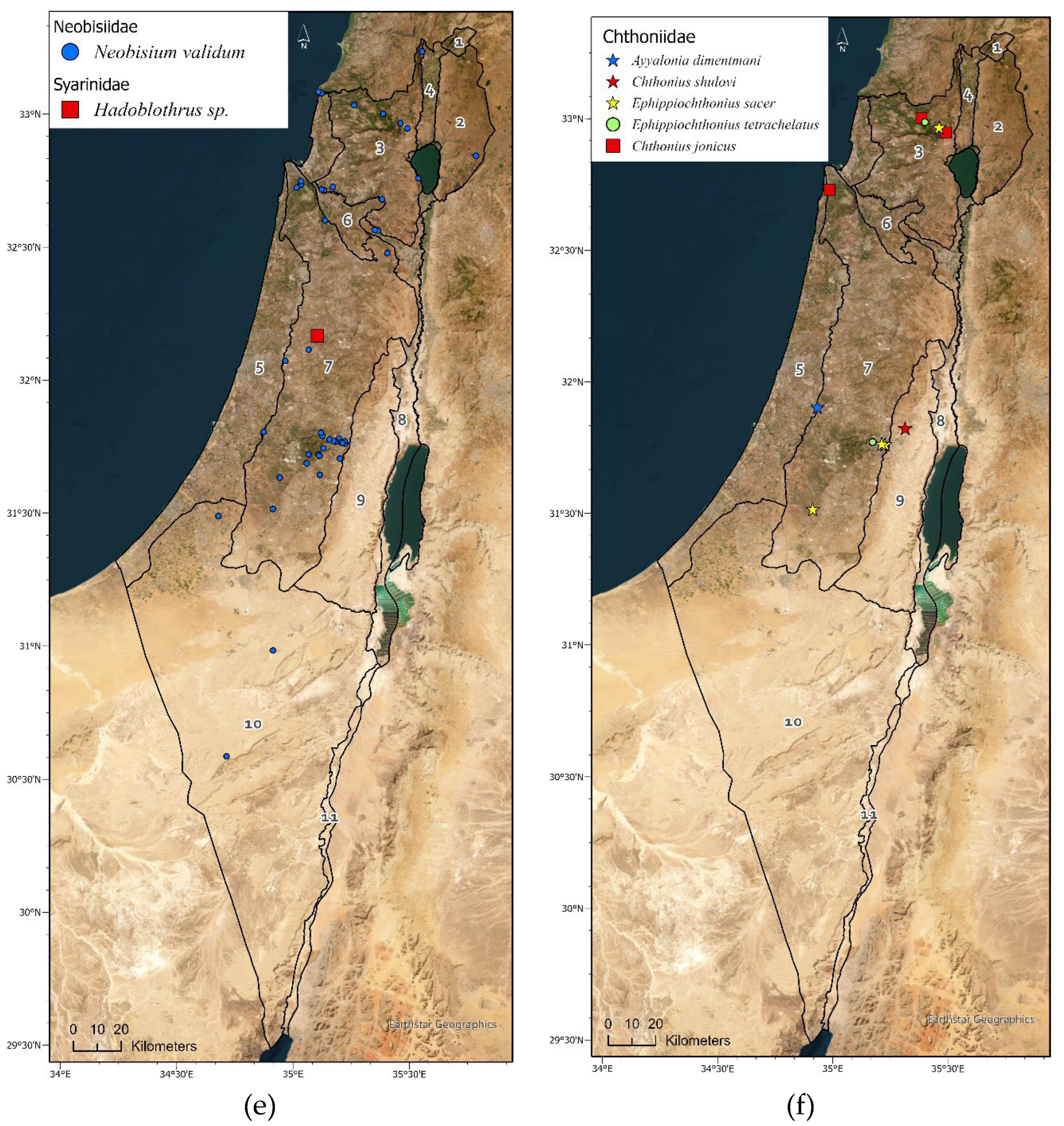
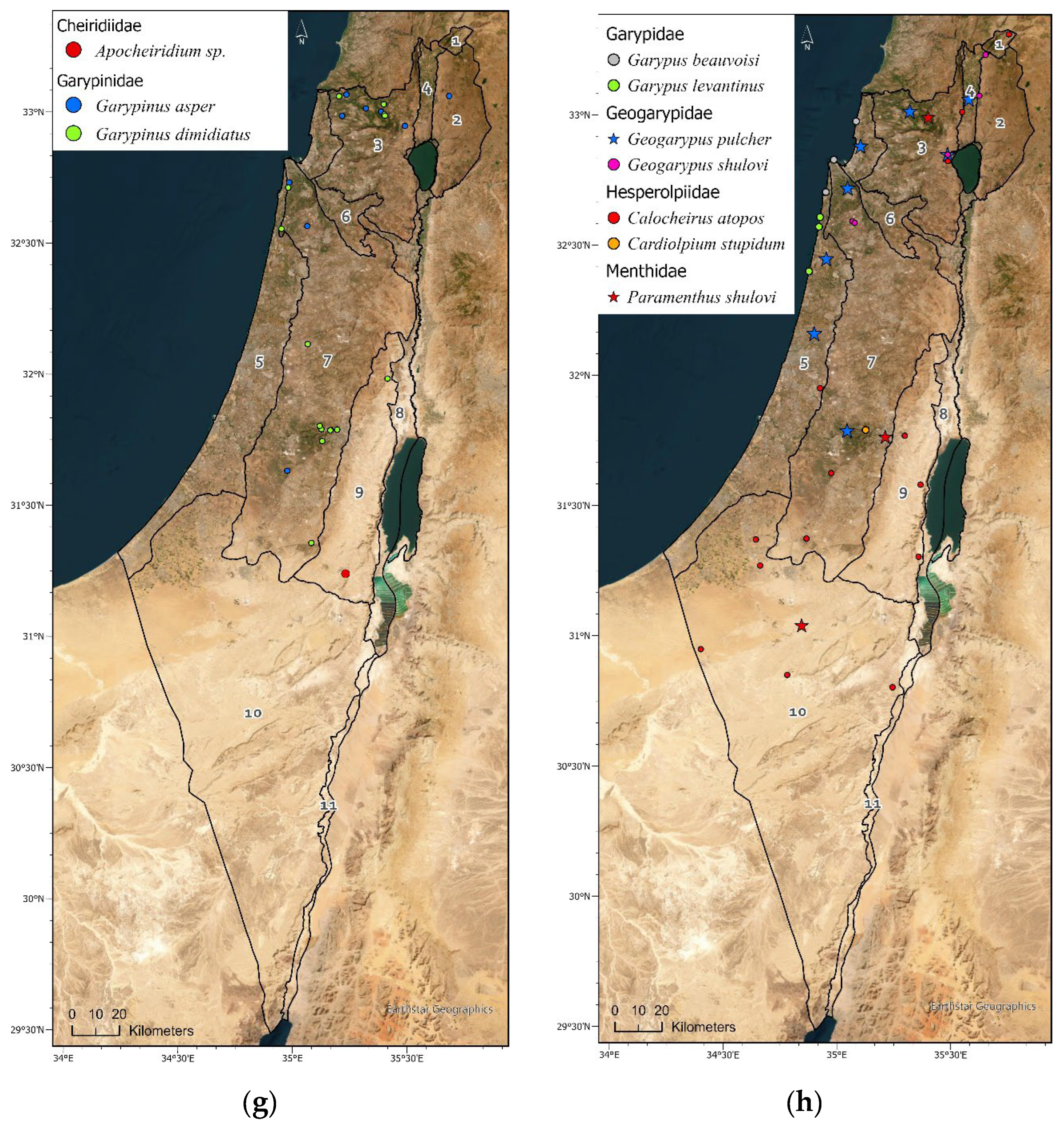
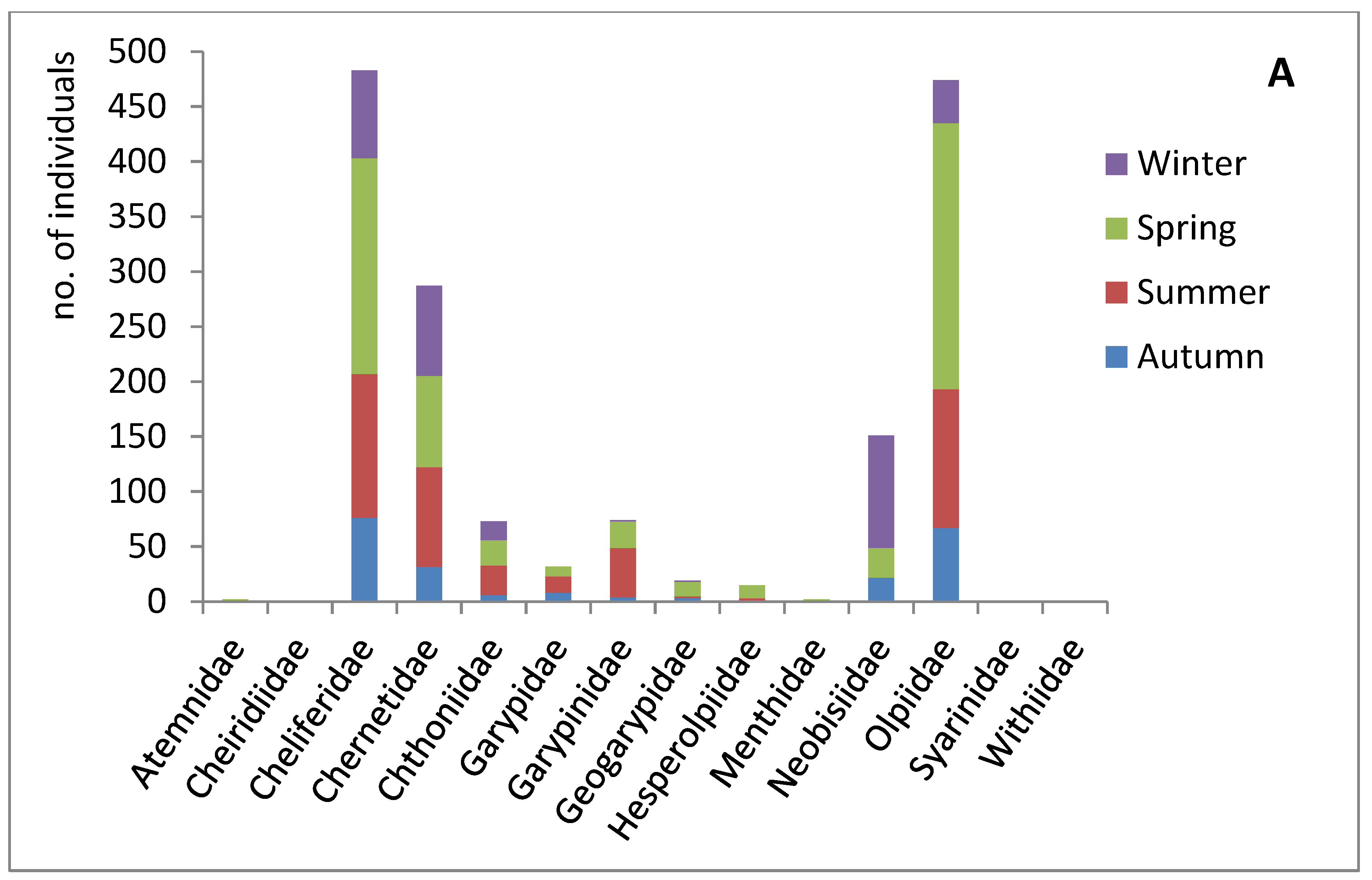
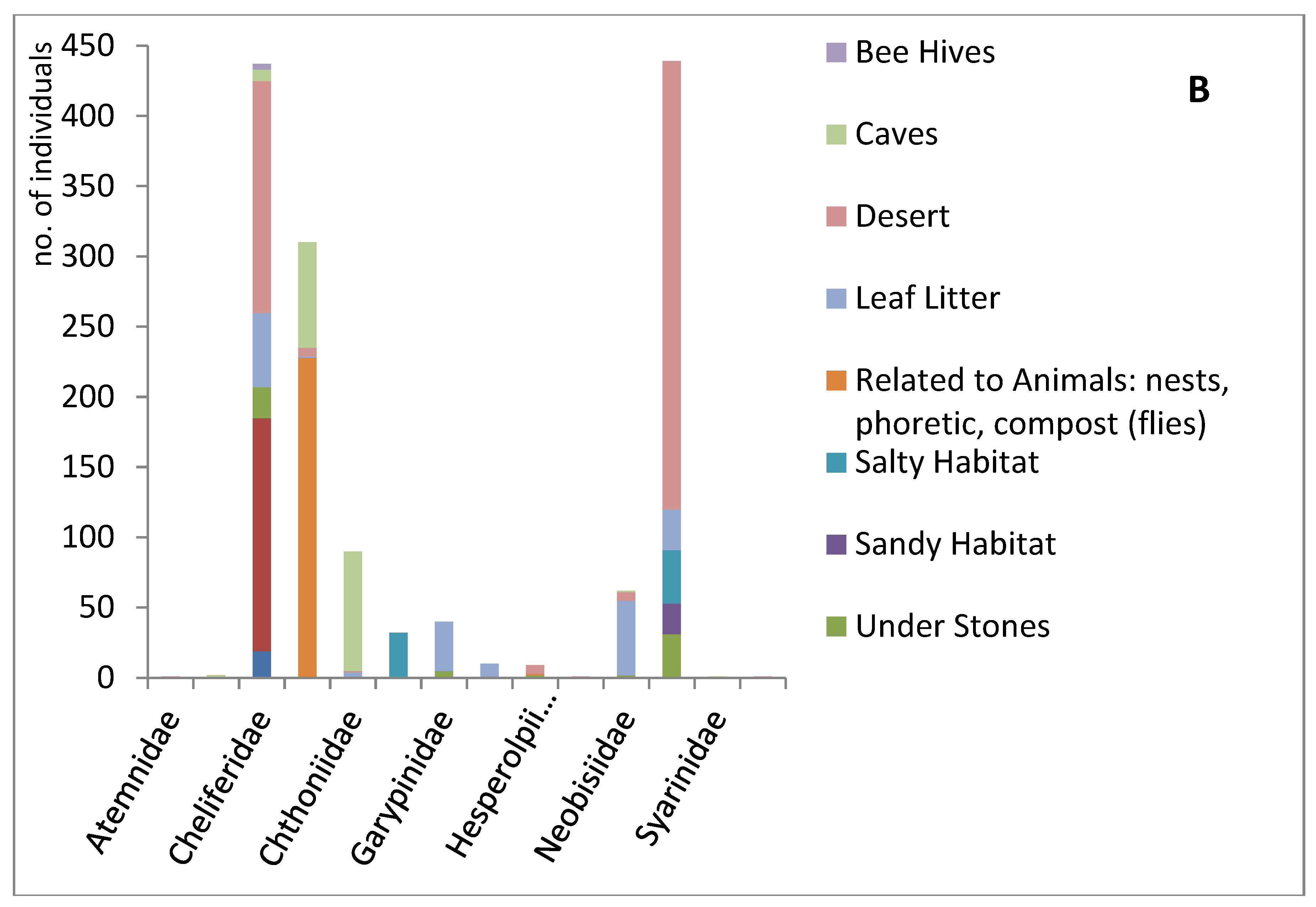
Figure 5.
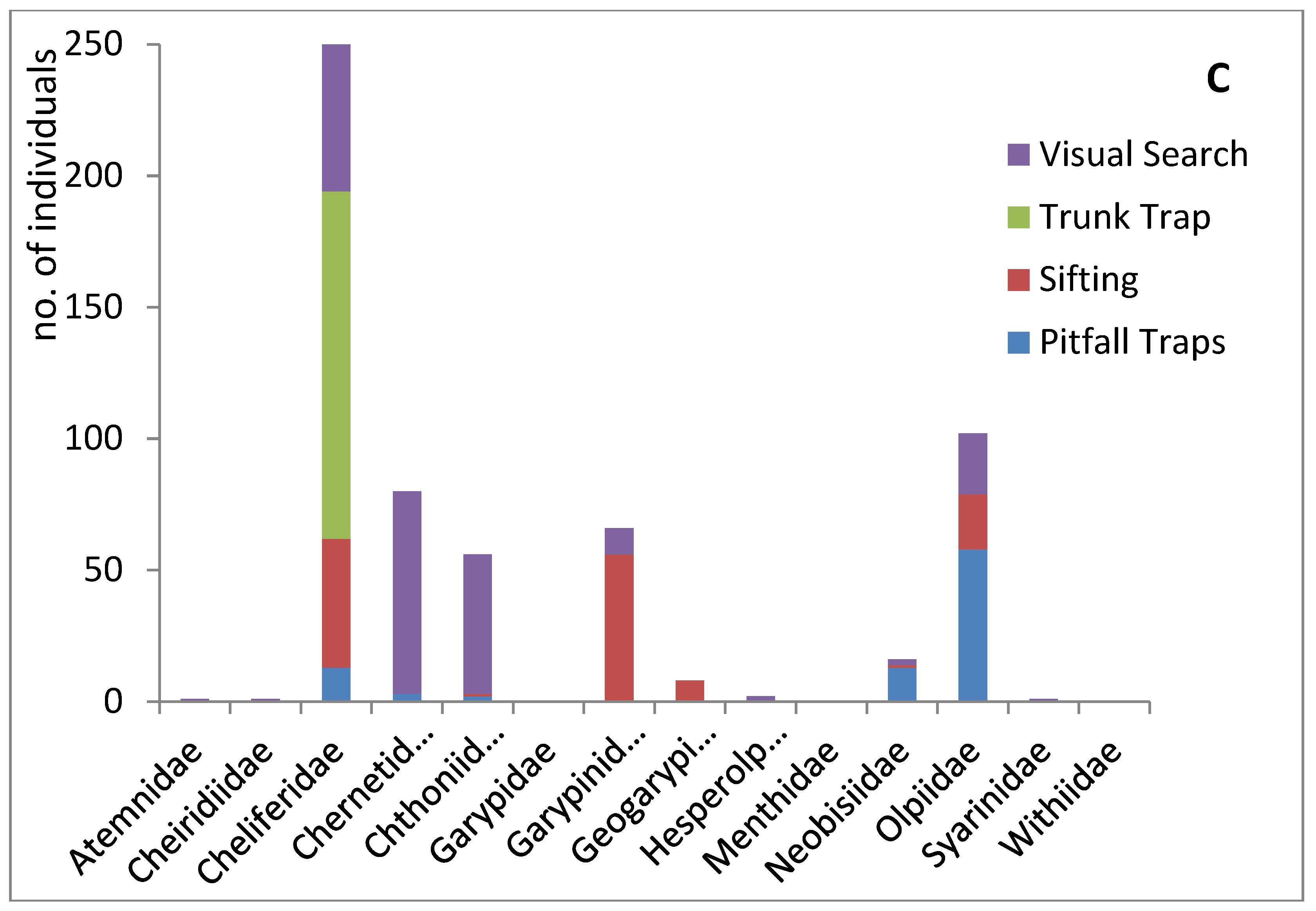
Seasonality, habitat, and collection methodology of pseudoscorpions deposited at the HUJ collection. (A) Seasonal distribution of the pseudoscorpion families in Israel (winter: December-February; spring: March-May; summer: June-August; autumn: September-November) (based on data of 1614 specimens in the collection, Supplementary Materials Table S4). (B) Habitats occupied by pseudoscorpion families in Israel (according to data of specimens in the collection). Data of the same specimen can contribute to more than one category, e.g. desert and under a stone; or cave and related to animals (Supplementary Materials Table S5). (C) Collection method (according to data of 583 specimens in the collection, Supplementary Materials Table S6).
Figure 5.
Seasonality, habitat, and collection methodology of pseudoscorpions deposited at the HUJ collection. (A) Seasonal distribution of the pseudoscorpion families in Israel (winter: December-February; spring: March-May; summer: June-August; autumn: September-November) (based on data of 1614 specimens in the collection, Supplementary Materials Table S4). (B) Habitats occupied by pseudoscorpion families in Israel (according to data of specimens in the collection). Data of the same specimen can contribute to more than one category, e.g. desert and under a stone; or cave and related to animals (Supplementary Materials Table S5). (C) Collection method (according to data of 583 specimens in the collection, Supplementary Materials Table S6).
3.1.2. The pseudoscorpion families
Cheliferidae is the largest and most common family deposited in the NNHC HUJ collection. Four genera and eighteen species and subspecies are documented from Israel (Table 3, Figure 4a,b). Only one species is Afrotropic, and the rest are Palearctic, with eleven Mediterranean, two Asian, two cosmopolitan and two endemic species (Table 3). Cheliferids were collected all over the country, in all regions except the Hermon (Figure 3, Figure 4a,b) and all year round. Most cheliferids were collected in spring and summer, and fewer individuals were collected in autumn and winter (Figure 5A). Cheliferids were collected using all methods. They were often found under bark and in leaf litter (Figure 5B) usually in habitats with trees. The subspecies Rhacochelifer corcyrensis bicolor Beier, 1963 was found almost exclusively in the desert, mainly on Acacia trees (Vachellia Wight & Arn) using trunk traps (Figure 4a,b, Figure 5A,C). The cosmopolitan species Chelifer cancroides (Linnaeus 1758) is phoretic and often found in beehives. The genus Dactylochelifer Beier, 1932 (the species D. kussariensis (Daday1889) and the endemic species D. pallidus Beier, 1963) was typically found near wetlands (swamps and riverbanks) (Figure 4a, Figure 5A). The other endemic cheliferid Hysterochelifer distinguendus (Beier, 1929) was found only in the Judea foothills (Figure 4a). Several specimens of Rhacochelifer longeunguiculatus Beier 1963 as well as unidentified species were collected from cave entrances but no troglobiont species were recorded from Israel thus far (Figure 6A).
Olpiidae is the second largest and most common family deposited in the NNHC HUJ collection, both in number of specimens and number of species and subspecies. Four genera and ten species are documented from Israel (Table 3, Figure 4c). Although globally Olpiidae species are usually found in xeric habitats (desert and semi-desert), the species in Israel are mostly Palearctic, with eight species from the Mediterranean and two species from Central and Western Asia and the Arabian Peninsula, thus from both Palearctic and Afrotropic regions (Table 3). Olpiids were collected all over the country, in all regions except the Hula Valley, but were significantly more common in the desert, where more than 75% of the specimens were found, while less than 10% were found in all the northern regions of Israel and the coastal plain (Supplementary Materials Tables S2-S3). They were collected all year, but the highest number was collected in spring while very little were collected in winter (Figure 5A). Specimens were collected by visual search under stones, sifting and in pitfall traps (Figure 5C). None of the olpiid species are endemic to Israel, yet Halominniza aegyptiaca litoralis (Beier, 1963) is endemic to the southern Levant (Israel, Jordan and Sinai), and was found in Israel only in the desert near the Dead Sea and the Red Sea, in association with sea (salty) water (Figure 4c). The Asian species Minniza babylonica Beier, 1931, which is found in both Palearctic and Afrotropic regions, is typically found in Israel in the desert (Figure 4c). This family has not been recorded from caves or moist epigean habitats (Figure 6B).
Chernetidae is the third largest and most common family deposited in the NNHC HUJ collection in both number of specimens and number of species, with six genera and eight species, in addition to undescribed species from caves. Five of these species are widely distributed and three are Mediterranean whereas one is endemic from an Afrotropic origin (Table 3, Figure 4d). Most of the specimens (80%), were collected in the mountains of central Israel and Jezreel valley, while hardly any were collected in the desert (Figure 4d, Supplementary Materials Tables S2-S3). Chernetids were found evenly throughout the year (Figure 5A) and usually by visual search (Figure 5C). Species of Chernetidae are often phoretic on rodents or insects (Figure 5B) and tend to aggregate in their nests as evident by 80% of the family’s specimens in Israel: Lamprochernes savignyi (Simon, 1881) is phoretic on flies, Lasiochernes turcicus Beier, 1949 (Figure 6C) is found in nests of Apodemus Kaup, 1829, and Nudochernes spalacis Beier, 1955 in nests of Nannospalax species. Nearly half of the specimens collected from caves in Israel (44%) belong to the Chernetidae family, being a quarter of the family’s specimens collected in Israel, from the species Allochernes masi, Chernes rhodinus, Lasiochernes turcicus and other unidentified species (Figure 5B). They were found mainly in caves with bats, both frugivorous and insectivorous, and L. turcicus also in an Apodemus nest.
Neobisiidae is the fourth most common family deposited in the NNHC HUJ collection, in terms of number of specimens; however, it is represented in Israel with only one genus and one species throughout the country, the Palearctic Neobisium validum (L. Koch, 1873) (Figure 6D), which is found in the East-Mediterranean as well as Central and Western Asia (Table 3, Figure 4e), with the type locality in Syria. Only 4% of the specimens collected from the desert, while 93% were collected in the mountains of north and central Israel and Jezreel valley (Figure 4e, Supplementary Materials Tables S2-S3). The remaining 3% were collected in the coastal region. Preliminary DNA sequencing (not presented here) indicates a species complex, which needs to be examined further. As a hygrophilous family, Neobisiidae specimens were collected four times more in winter than in spring and autumn, while none were collected during the summer (Figure 5A). They were usually found by sifting soil and leaf litter, visual search under stones or using pitfall traps (Figure 5B,C). One specimen of a presently unidentified troglobitic species was found in a cave.
Chthoniidae is represented in Israel by five species from three genera in addition to several undescribed species from caves. Three species are endemic, one Mediterranean and one cosmopolitan (Table 3, Figure 4f). Chthoniids were found exclusively in the mountains of north and central Israel (Figure 3, Figure 4f, Supplementary Materials Tables S2-S3) or in caves (Figure 5B). Half of the pseudoscorpion specimens we found in Israel’s caves belong to the Chthoniidae, representing 75% of the family’s specimens collected in Israel. Being a family with high short-range endemism [6], we expect several new species to science among the unidentified cave specimens (Figure 6E). As a hygrophilous family, only a few Chthoniidae specimens were collected in the driest season in Israel, autumn (Figure 5A). Chthoniids were collected mostly by visual search under stones, and on bats guano in caves (Figure 5B,C), but they also occur in leaf litter, and some were collected by sifting and pitfall traps. Ayyalonia dimentmani Ćurčić, 2008 is an example of an endemic relict species found only in a single unique chemoautotrophic cave. Another endemic species, Chthonius shulovi Beier, 1963 was found only once in one locality in Wadi Qelt [15]. Both species were never found again. However, the endemic species Ephippiochthonius sacer (Beier, 1963) seems to be more widespread, and was found several times in the Mediterranean region of north and central Israel.
Garypinidae is represented in Israel by two species of Garypinus Dadday, 1889 that have an eastern Mediterranean to Palearctic distribution (Table 3, Figure 4g). Both were found exclusively in the mountains of north and central Israel (Figure 3, Figure 4g, Supplementary Materials Tables S2-S3), mostly in spring and summer (Figure 5A), often by sifting leaf litter or visual search under stones (Figure 5B,C).
Garypidae is represented in Israel by two species of Garypus L. Koch, 1873 that are halophilous and widespread in seashore habitats of the Mediterranean (Table 3, Figure 4h). Both were found in all seasons but winter (Figure 5A), exclusively in the salty habitat of the Mediterranean coast (Figure 3, Figure 4h, Figure 5B). Some specimens were found in algae debris at the coastline.
Geogarypidae is represented in Israel by two species from one genus: Geogarypus shulovi Beier, 1963 is Palearctic from Central and Western Asia, and Geogarypus pulcher Beier, 1963 is endemic to Israel (Table 3, Figure 4h). Most of the specimens were found in the mountains of north and central Israel, but some also in the coastal plain and Hula valley (Figure 3, Figure 4h, Figure 5B, Supplementary Materials Tables S2-S3). They were usually found by sifting leaf litter, mostly in spring (Figure 5B,C).
Hesperolpiidae is represented in Israel by two species from two genera, Calocheirus atopos Chamberlin, 1930 is Afrotropic from the Arabian Peninsula and Sudan, and Cardiolpium stupidum (Beier, 1963) is Palearctic from East-Mediterranean and Central-West Asia (Table 3, Figure 4h). They are found mostly in spring (Figure 5A), in small numbers but in a wide distribution along Israel (Figure 3, Figure 4h, Figure 5B).
Menthidae is an infrequently collected family and represented in our collection by only three specimens; all belong to the endemic species Paramenthus shulovi Beier, 1963; an Afrotropic genus. Each of the specimens was collected from a different sub-region of the country; the Negev desert, the Central Mountains, and the Galilee (Table 3, Figure 4 h).
Atemnidae is represented in our collection by only two specimens, each from a different species and a different region: Atemnus syriacus (Beier, 1955) in the north of Israel, and an undescribed species from the Judean desert (Table 3, Figure 4d) .
Withiidae is represented in our collection by the endemic species Nannowithius wahrmani (Beier, 1963), which belongs to the Afrotropic genus Nannowithius Beier. It is a small myrmecophilous species that was found in nests of the ant Messor semirufus (André, 1883) in the Negev desert, under a stone near the sea of Galilee, and phoretic on the myrmecophilous scorpion Birulatus israelensis which lives only in Messor ebeninus Santschi, 1927 nests (Figure 4d, Figure 6f) [27].
Cheiridiidae is represented in our collection by a single specimen that was collected in a cave in the Judean desert (Figure 4g).
Syarinidae is represented in our collection by a single specimen that was collected in a cave in the Central Mountains (Figure 4e).
Figure 6.
Habitus of live pseudoscorpion representatives from Israel. A. Hysterochelifer cyprius (Cheliferidae). B. Olpium pallipes balcanicum (Olpidae). C. Lasiochernes turcicus (Chernetidae). D. Neobisium validum (Neobisiidae). E. Ephippiochthonius sp. (Chthoniidae). F. Nannowithius wahrmani (Withiidae) phoretic on Birulatus israelensis. Photos A, B, D, E, F by S. Aharon, C by J. A. Ballesteros.
Figure 6.
Habitus of live pseudoscorpion representatives from Israel. A. Hysterochelifer cyprius (Cheliferidae). B. Olpium pallipes balcanicum (Olpidae). C. Lasiochernes turcicus (Chernetidae). D. Neobisium validum (Neobisiidae). E. Ephippiochthonius sp. (Chthoniidae). F. Nannowithius wahrmani (Withiidae) phoretic on Birulatus israelensis. Photos A, B, D, E, F by S. Aharon, C by J. A. Ballesteros.

3.1.3. A dichotomous key to the pseudoscorpion families and genera of Israel (applying to adults) (in parentheses no. of genera and no. of species in the family).
1. a. Legs with different number of tarsal segments: legs I and II with one tarsal segment and legs III and IV with two tarsal segments; pedipalpal chelal fingers slender and without venom apparatus; two trichobothria (ib and isb) medially on dorsal face of pedipalpal chela hand; coxal spines present (Figure 7A) . . . . . . . . . . . . . . . . . . . . . . . . . suborder Heterosphyronida . . . . . . . . . . . . . . . . . . . . . superfamily Chthonioidea . . . . . . . . . family Chthoniidae (3, 7) 2
1. b. All legs with same number of tarsal segments; at least one pedipalpal chelal finger with venom apparatus (Figure 7B); trichobothria ib and isb usually located on the lateral face of the fixed chelal finger (occasionally ib is on the dorsal surface); coxal spines absent (Figure 7C) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .suborder Iocheirata 4
2. a. Coxae III without coxal spines; only coxae II each with 11 fine elongated coxal spines, middle spines longest; apex of pedipal pal coxa with three setae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Ayyalonia
2. b. Coxae II and III with coxal spines; apex of pedipal pal coxa with two setae . . . . . . . . . . Chthonius-related genera 3
3. a. Teeth of pedipalpal chelal fingers pointed, aligned, erect and spaced; pedipal pal hand with dorsal step-like outline (dorsodistal saddle-shaped constriction) (Figure 7D) between the groups of trichobothria ib-isb ang eb-esb-ist . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Ephippiochthonius
3. b. Teeth of pedipalpal chelal fingers blunt, dense and distinctly reclined backwards; pedipal pal hand with evenly rounded weakly curved dorsal outline (Figure 7E) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Chthonius
4. a. Rectangular carapace (anterior and posterior margins similar in length); robust chelicerae, their bases almost as wide as posterior side of carapace; all legs with two tarsal segments; two pairs of eyes (epigean species) or none (caves); setae present on the apical margin of pedipalp coxa (Figure 7F,G) . . . . . . . . . . . . . . . .infraorder Hemictenata . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . superfamily Neobisioidea 5
4. b. Carapace not rectangular (anterior margin shorter than posterior margin); width of chelicerae bases smaller than posterior margin of carapace (less than half its length) (Figure 7H); the legs have either all one tarsal segment or all two tarsal segments; either no eyes or one or two pairs of eyes or eyespots; setae on the apical margin of pedipalp coxa may be present or absent . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .infraorder Panctenata 6
5. a. Apex of pedipalpal coxa rounded and bears three or more setae (Figure 7F) . . . . . . . . . . . family Neobisiidae (1, 3)
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Neobisium
5. b. Apex of pedipalpal coxa not rounded and bears two long subequal setae (Figure 7G) . . . . family Syarinidae (1, 1)
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .Hadoblothrus
6. a. All legs with two tarsal segments (Figure 7I); two pairs of eyes with lenses; usually elongated carapace (longer than wide); pedipalpal femur with or without trichobothria . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .7
6. b. All legs with one tarsal segment (Figure 7J); one pair of eyes or eye-spots, or eyes absent; carapace sub-triangular; pedipalpal femur without trichobothria . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16
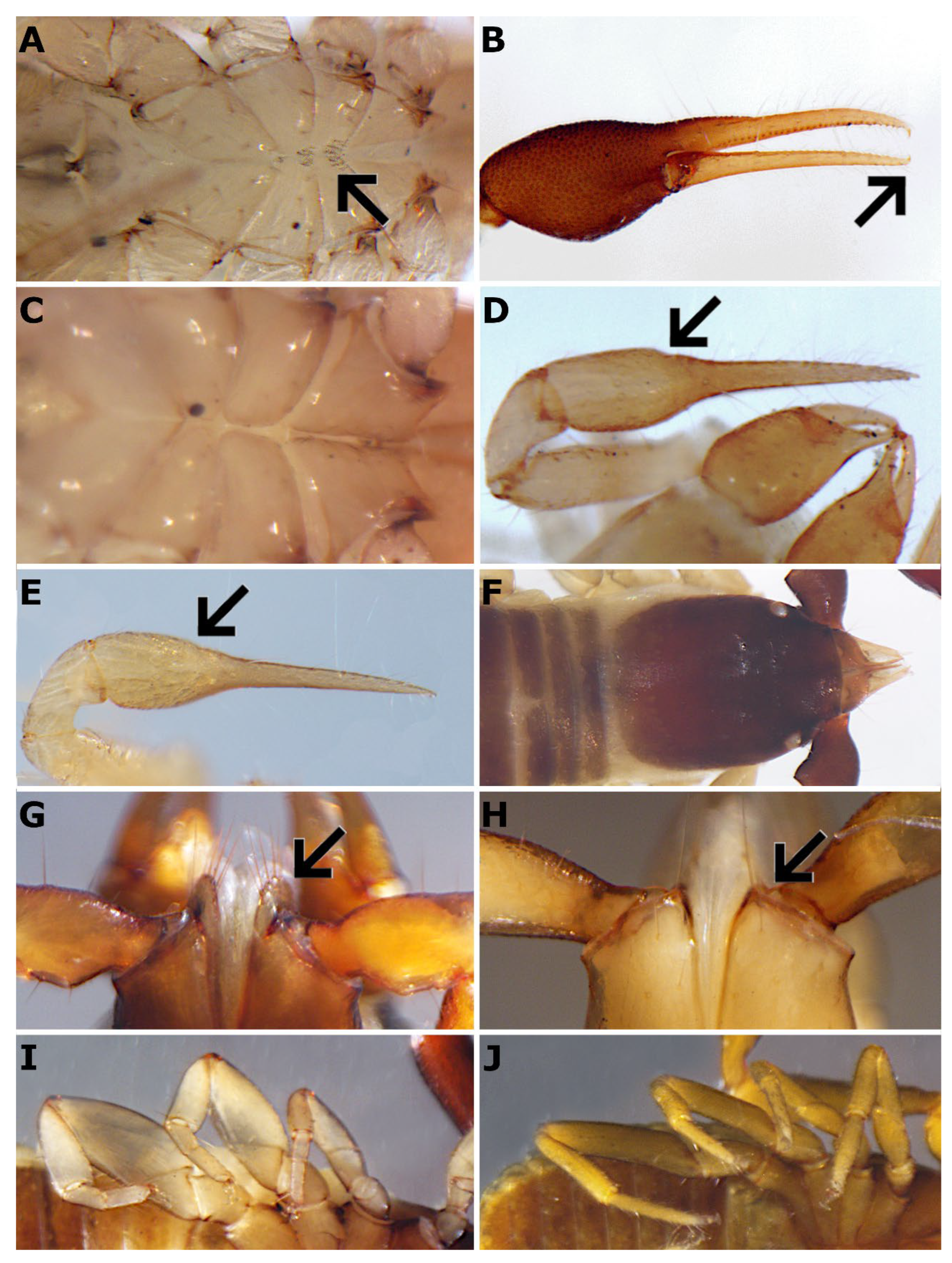
Figure 7.
A. Ephippiochthonius tetrachelatus with coxal spines present. B. Geogarypus pulcher with venom apparatus on both pedipalpal chelal fingers. C. Neobisium validum with no coxal spines. D. Ephippiochthonius sp. with dorsal step-like outline of pedipalpal hand. E. Cthonius sp. with evenly rounded weakly curved dorsal outline of pedipalpal hand. F. Neobisium validum with five setae on rounded pedipalp coxa apex. G. Hadoblothrus sp. pedipalp coxal apex not rounded and bears two long subequal setae. H. Garypinus sp. with the width of the chelicerae bases smaller than the width of the posterior margin of carapace. I. Garypinus sp. with two tarsal segments in all legs. J. Hysterochelifer tuberculatus with one tarsal segment in all legs.
Figure 7.
A. Ephippiochthonius tetrachelatus with coxal spines present. B. Geogarypus pulcher with venom apparatus on both pedipalpal chelal fingers. C. Neobisium validum with no coxal spines. D. Ephippiochthonius sp. with dorsal step-like outline of pedipalpal hand. E. Cthonius sp. with evenly rounded weakly curved dorsal outline of pedipalpal hand. F. Neobisium validum with five setae on rounded pedipalp coxa apex. G. Hadoblothrus sp. pedipalp coxal apex not rounded and bears two long subequal setae. H. Garypinus sp. with the width of the chelicerae bases smaller than the width of the posterior margin of carapace. I. Garypinus sp. with two tarsal segments in all legs. J. Hysterochelifer tuberculatus with one tarsal segment in all legs.
7. a. Arolia bifurcate (Figure 8A); at least a few tergites and sternites are divided. . . . . . . . . .superfamily Garypinoidea
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . family Garypinidae (1, 2)
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Garypinus
7. b. Arolia simple (Figure 8B) (not bifurcate); tergites and sternites can (in certain families) be all undivided . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . superfamily Garypoidea . . . . . . . . . . . . . . . . . .8
8. a. Venom apparatus present only in the fixed chelal finger; fixed chelal finger with 11 trichobothria, of which two are on the dorsal surface of the hand; specialised joint present between coxae II and III . . . . . . family Menthidae (1, 1)
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Paramenthus shulovi
8. b. Venom apparatus present in both chelal fingers; fixed chelal finger with 8 or (rarely) 7 trichobothria, including 3 on internal face; dorsal surface of the hand without trichobothria; articulation furrow between coxae II and III absent . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9
9. a. All tergites and sternites undivided (Figure 8C,D); abdomen oblong, not much broader than the carapace; the carapace sub-rectangular and slightly tapering orally; eyes positioned near anterior margin of carapace, not sitting on mounds; arolia longer than claws; vestitural bristles of pedipalps rather long, erect, and thinly pointed; pleural membrane of abdomen with smooth and even longitudinal striation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
9. b. At least a few tergites and sternites divided (Figure 8E); abdomen broadly oval, distinctly broader than the carapace; carapace triangular and strongly tapering orally; eyes protruding, sitting on mounds, removed from anterior margin of the carapace; arolia can be longer or shorter than the claws; vestitural bristles of pedipalps very short, fine, bent, and abruptly pointed; pleural membrane of abdomen granulate or with short wavy undulating striation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .15
10. a. Palpal fingers distinctly bent; palpal femur with or without two fairly long dorsal tactile bristles . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . family Olpiidae (4, 10) . . . . . . . .11
10. b. Palpal fingers straight; palpal femur with a single fairly long dorsosubbasal tactile bristle . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .family Hesperolpiidae (2, 2) . . . . . . . 14
11. a. Carapace without submedian transverse furrow, longer than broad maximum 1.5 times . . . . . . . . . . . . . . . . . . . .12
11. b. Carapace with a submedian transverse furrow; either just slightly or 1.7 to 1.8 times longer than broad. . . . . . . 13
12. a. Carapace and pedipalps dark brown; palpal femur 0.6 to 0.7 mm long; fingers about as long as the hand with pedicel . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .Olpium
12. b. Carapace and pedipalps light yellowish-brown; palpal femur 0.9 to 1 mm long; fingers much longer than hand with pedicel . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Halominniza
13. a. Body length about 2 to 3 mm; carapace 1.7 to 1.8 times longer than broad; femur, tibia and chela of pedipalps dark brown, with pedicel and tips reddish, rarely entirely reddish; desert species with worm-shaped abdomen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Minniza
13. b. Body length 1.5 to 1.7 mm; carapace but slightly longer than broad; femur and tibia of pedipalps light brownish-yellow, chela blackish-olivebrown . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .Calocheiridius
14. a. Body length 2.3 to 2.8 mm; eyes about half their diameter distant from one another; carapace with a weak transverse furrow; palpal hand very broad, abruptly constricted medially and laterally towards the base of the fingers; fingers at least as long as the femur; trichobothria est situated near the middle of fixed finger; three trichobothria ist, it and et form a group near tip of finger . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Calocheirus
14. b. Body length 1.2 to 1.9 mm; eyes touching, the anterior eyes slightly excavated behind, the posterior eyes conical and flattened; carapace without a transverse furrow; palpal hand not very broad, medially obliquely narrowed towards the base of fingers; fingers shorter than femur; trichobothria est situated rather basally, proximal of the middle, near trichobothria isb, close to the group formed of eb-esb-ib-isb . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Cardiolpium
15. a. Carapace triangular and medially concave (Figure 8E); maxilla of palpal coxae with developed shoulder; coxal area not expanding posteriorly: coxa IV approximately same width as coxa I (broad and short); body 2 to 2.5 mm long; arolia longer than the claws; tergites usually with dark spots; occur in leaf litter and under stones or bark in the Mediterranean shrubland . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . family Geogarypidae (1, 2) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .Geogarypus
15. b. Carapace triangular but not medially concave (Figure 8F); maxillar shoulder of palpal coxae not developed; coxal area expanding posteriorly: i.e. coxa IV distinctly longer and narrower than coxa I; body 5 to 6 mm long; arolia shorter than claws; tergites usually with dark bands; occur only in littoral or supralittoral zones . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . family Garypidae (1, 2)
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Garypus
16. a. Small body size (< 2 mm); carapace triangular and coarsely granulate with one medial deep transverse furrow (Figure 8G); one pair of small eyes, if not absent, distinctly removed from anterior margin; femur and patella of all legs fused, with suture between them hardly visible; coxa IV much wider than coxa I; pedipalps with reduced number of trichobothria: at most seven on fixed chelal finger and one or two on moveable finger. . .superfamily Cheiridioidea . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .family Cheiridiidae (1, 1)
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Apocheiridium
16. b. Body size larger than 1.5 mm; eyes (if not absent) positioned near the anterior margin of the carapace (Figure 8H); femur and patella of legs not fused; coxa IV not wider than coxa I; pedipalps usually with eight trichobothria on fixed chelal finger and four on moveable finger . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . superfamily Cheliferoidea. . . . 17
17. a. Venom apparatus present in both chelal fingers; one pair of eyes or eye spots; males with or without coxal sacs or ram’s horn organs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .18
17. b. Venom apparatus absent from one chelal finger; no eyes or one pair of eyes or eye spots; males without coxal sacs or ram’s horn organs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .22
18. a. Articulation of femur and patella of leg I and II narrow and transverse, therefore these joints scarcely movable; male (and occasionally females) abdominal posterior sternites with discrete patches of glandular sensory setae; hind coxae of male without coxal sacks and ram’s horn organs absent; very small myrmecophilous specimens . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .family Withiidae (1, 1)
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Nannowithius wahrmani
18. b. Articulation of femur and patella of leg I and II well developed (Figure 8I), great and oblique and therefore well movable; male abdominal sternites without patches of sensory setae; hind coxae of male excavate caudally, male genitalia with coxal sacks and ram’s horn shaped organs (Figure 8I); not myrmecophilous . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . .family Cheliferidae (4, 18) . . . . . . . . . . . . . . . .19
19. a. Carapace with dense granulation and greater bristle-bearing granules; abdominal tergites of male with lateral carinae and elongate hind-angles. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20
19. b. Carapace with dense granulation without greater granules; abdominal tergites of male without lateral carinae, their hind-angles not elongate. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
20. a. Pedipalpal femur with coarse bristle-bearing medial granules; subbasal bristle of chelicera present . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .Hysterochelifer
20. b. Pedipalpal femur without coarse medial granules; no subbasal bristle of chelicera . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .Chelifer
21. a. At least submedian transverse furrow of the carapace deeply incised; pedipalps slender. . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Dactylochelifer
21. b. Both transverse furrows of carapace shallow; pedipalps clumsy and strong . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .Rhacochelifer
22. a. No eyes or eye spots (Figure 8J); venom apparatus developed only in movable chelal finger; carapace with grooves; robust (Figure 8J); abdomen broadly oval, distinctly broader than the carapace . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . family Chernetidae (6, 9) . . . . . . . . . . .23
22. b. One pair of eyes or eye spots; venom apparatus developed only in fixed chelal finger; longitudinal grooves may be present; slender and elongate abdomen, not broader than the carapace . . . . . . . . . . . . . . . . . . family Atemnidae (1, 2)
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .Atemnus
23. a. Body and pedipalp setae long, thin and pointed; carapace almost smooth; subbasal transverse furrow on carapace indistinct; tibia and hand of pedipalps with fairly long lateral pseudotactile setae; often phoretic on flies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .Lamprochernes
23. b. Body and pedipalp setae short, stout, dentate or clavate; carapace granulate; tibia and hand of pedipalps without lateral pseudotactile setae; not phoretic on flies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24
24. a. Tarsus of leg IV with long erect tactile seta near the middle, distinctly longer than width of tarsus . . . . . . . . . . . 25
24. b. Tarsus of leg IV without erect tactile seta near the middle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27
25. a. Carapace rather coarsely granulated, with two distinct transverse furrows, of which the posterior is flatter than the anterior; body and pedipalp setae always slightly but clearly clavate, relatively long; male pedipalp without dense long setation; palpal femur at most 0.5 mm long; small 1.5-2 mm; in ground litter and detritus, in moist places . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .Pselaphochernes
25. b. Carapace with dense and fine granulation with posterior transverse furrow situated scarcely nearer to hind border than to the anterior furrow; body setae dentate, but not clavate; palpal femur longer than 0.7 mm; body length 2-4 mm; in nests of small mammals and in caves with bats . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .26
26. a. Tactile bristle near the middle of the hind tarsus long and simple; body setae rather long serrated to slightly truncate not clavate, those of the pedipalps only serrated; setae on femur and tibia of the pedipalps of the male very long and dense like a mane; fairly large and robust: body length 4 mm, palpal femur at least 1 mm long; in nests of Apodemus. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Lasiochernes
26. b. Tactile bristle near the middle of the hind tarsus dentate and scarcely twice as long as the other bristles and about one third as long as width of tarsus; body setae very short; medial bristles of the palpal femur of the male but scarcely more dense than in the female; body length 2-3 mm, femur length 0.69-0.74 mm; in nests of the rodent Nannospalax . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Nudochernes
27. a. Carapace quite coarsely granulated; body and pedipalp setae serrated and strongly culled usually quite short; slightly clavate; pedialps are moderately slender, granulated . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Allochernes
27. b. Carapace moderately coarsely granulated, sometimes partially reticulated; body and pedipalp setae dentate and more or less truncate; pedialps robust, granulated; pedipalp coxae usually not granulated . . . . . . . . . . . . . . . . . . Chernes
Figure 8.
A. Bifurcate arolia. B. Simple arolia. C. Minniza babylonica with all tergites undivided. D. Minniza babylonica with all sternites undivided. E. Geogarypus sp. with five tergites divided. F. Geogarypus sp. with carapace triangular and medially concave. G. Garypus beauvoisi with carapace triangular but not medially concave. H. Cheiridiidae sp. with one medial deep transverse furrow in the coarsely granulate and triangular carapace. I. Hysterochelifer tuberculatus with eyes positioned near the anterior margin of the carapace. J. Hysterochelifer tuberculatus with articulation of femur and patella of leg I and II well developed and male genitalia with ram’s horn shaped organs. K. Chernetidae sp. robust with two grooves in the carapace and without eyes or eye spots.
Figure 8.
A. Bifurcate arolia. B. Simple arolia. C. Minniza babylonica with all tergites undivided. D. Minniza babylonica with all sternites undivided. E. Geogarypus sp. with five tergites divided. F. Geogarypus sp. with carapace triangular and medially concave. G. Garypus beauvoisi with carapace triangular but not medially concave. H. Cheiridiidae sp. with one medial deep transverse furrow in the coarsely granulate and triangular carapace. I. Hysterochelifer tuberculatus with eyes positioned near the anterior margin of the carapace. J. Hysterochelifer tuberculatus with articulation of femur and patella of leg I and II well developed and male genitalia with ram’s horn shaped organs. K. Chernetidae sp. robust with two grooves in the carapace and without eyes or eye spots.

3.2. Ecology and Distribution
3.2.1. Biogeography and biodiversity
Despite Israel being largely arid or semi-arid, the zoogeographic origin of the pseudoscorpion fauna in Israel is mostly Mediterranean and Palearctic, based on our analyses. The most dominant chorological complex is Mediterranean, where half of the pseudoscorpion species are found (29 species). One fifth of the pseudoscorpion species are classified as widely distributed species (twelve species), while only 15% have Asiatic, African or European origin (nine species in total) (Figure 9). For each species, the chorotype classification and chorological complex are listed in Table 3, based on Harvey [18] and WPC [1], as well as their global zoographic category (Palearctic or Afrotropic). Forty species (including subspecies and genera of undescribed species) from twelve families are Palearctic, three species are Afrotropic and six species (including three subspecies) are found both in the Palearctic and in the Afrotropic zoogeographic zones (Table 3).
Only nine (15%) described species from six families are endemic to Israel; six are of Palearctic origin and three are of Afrotropic origin (Table 3). Most of the endemic species are rare and restricted to a certain biogeographic sub-region or habitat (Figure 3). Most of the undescribed species, which were all collected in caves, are probably endemic, but a further taxonomic study is needed to verify this.
3.2.2. Seasonality, habitat, and collection methodology
Although pseudoscorpions can be found all year round, we found variability in family seasonality in the pseudoscorpions deposited in the collection (Figure 5A). For example, Neobisiidea were mostly collected in the winter, some individuals in spring and autumn, but none in the summer. At the same time, some families such as Garypidae and Garypinidae were not collected at all in winter. Other families were collected in higher numbers during springtime (Olpiidae, Cheliferidae, Geogarypidea, and Hesperolpiidae). Families that reside in habitats with stable conditions, such as Chernetidae species associated with host nests and Chthoniidae species from subterranean habitats, were collected almost evenly in winter, spring and summer, and less in autumn.
We also found that the habitat varied between some families (Figure 5B). Cheliferidae were often collected under bark of trees, while phoretic species of the family Chernetidae were often collected in nests of rodents. Many epigean species, of different families, were collected from soil and leaf litter, while in subterranean habitats the families Chernetidae and Chthoniidae dominate.
Information regarding the collection method was available for only a third of the specimens. Four main methods were reported: pitfall traps, trunk traps, visual search and sifting (Figure 5C). Only Cheliferidae were collected by trunk traps while visual search was an effective method for most families. Pitfall traps were very useful to collect Olpiidae, and also Cheliferidae and Neobisiidae. Sifting was very useful for soil and leaf litter inhabitants of several families, while visual search was almost the only method used for collecting Chernetidae and Chthoniidae.
4. Discussion
This study is based on historical collection material that has been collected sporadically over decades, and on freshly collected specimens from recent years. However, no methodological survey of the pseudoscorpion fauna of Israel has ever been conducted. The uneven species richness in different regions of Israel might therefore reflect the degree of exploration by researchers and sampling biases, in addition to geographic differences. For instance, the collection harbours fewer records from the Hermon and Golan sub-regions in north-east Israel, compared to other regions of the country (Supplementary Materials Table S2), which makes the evaluation of species richness in these regions impossible. Prior to this study, 52 species of twelve pseudoscorpion families had been documented from Israel. This is less than expected compared to some other Mediterranean countries such as Turkey (114 species), Greece (141), Bosnia & Herzegovina (91), Croatia (147), Italy (272), and Spain (286) although Israel is also a smaller country and without any islands [1]. Apart from environmental and geographical differences between the countries, as well as their relative size, this gap is probably due to extensive research and exploration in these countries over the years.
A survey conducted in the last years in Israeli caves, habitats that are often refugial and have relict species [14], yielded pseudoscorpion species new to science, which will be described and discussed elsewhere. Most of the pseudoscorpions found in Israeli caves belong to two cave-dwelling families, the Chthoniidae and Chernetidae. In addition, two families new to Israel were revealed in caves, Cheiridiidae and Syarinidae, as well as a non-troglobitic species new to Israel from the Atemnidae family that was found in a small open cave.
Cheiridiidae is a widely distributed cosmopolitan family, commonly found in neighbouring countries, that was expected to occur also in Israel, but was not recorded until our recent discovery of the yet to be determined species of the putative genus Apocheiridium Chamberlin, 1924.
For Syarinidae, although a widely distributed cosmopolitan family of many species, only several relict species have been found in Europe and the Mediterranean region, and these are rare troglobiont species. One of these is the relict genus Hadoblothrus Beier, 1952, which we report, thereby extending significantly its known range. Hadoblothrus was previously found only in Greece and Italy. Hadoblothrus aegaeus Beron, 1985 was found only in the caves of Iraklia and Santorini but not in any of the well explored numerous caves on the mainland of the Balkan Peninsula, and Hadoblothrus gigas (Caporiacco, 1951) was found only in two caves in southern Italy [28]. This range extension of the putative genus Hadoblothrus suggests a further link of the Israeli fauna with the Palearctic realm.
Atemnidae is a widely distributed cosmopolitan family of 186 species known worldwide, with representatives known from most countries in the East Mediterranean region, such as Syria (two species), Egypt (one species), Cyprus (one species) and Turkey (five species) [1,18]. A single atemnid from a different species, Atemnus syriacus, had already been recorded by Mahnert [16], but being a tritonymph, he left a question mark on its exact identification.
- Distribution and Zoogeography
Four main zoogeographic regions are usually used for terrestrial animals in Israel: Palearctic, Palaeoeremic, Afrotropic, and Oriental. The Palearctic region is in northern Israel, while the Palaeoeremic (Old World desert fauna) reaches Israel from the south into the Negev desert and the Judean desert and stretches up in two ‘arms’ along the coastal plain and along the Jordan Rift Valley. The Palaeoeremic faunal inventory is more related to the Afrotropic than to the Palearctic fauna [29]. Between the Palearctic and Palaeoeremic regions, south of the Jezreel Valley and north of the Negev desert, lies a transition zone that includes both Palearctic and Palaeoeremic elements. The Afrotropic (Ethiopian) zoogeographical region stretches along the Jordan Rift valley and the Dead Sea, while the Oriental zoogeographical elements are scattered without specific geographical affinity [29].
The pseudoscorpion fauna of Israel is mostly Palearctic; fifty precent of the species belong to the Mediterranean chorological complex (of which half belong to the Eastern Mediterranean chorotype) (Figure 9). This is typical in the Mediterranean region, including the Levant [4,29]. Twenty percent of the pseudoscorpion fauna of Israel are widely distributed (e.g., the synanthropic Chelifer cancroides (Linnaeus, 1758), Lamprochernes savignyi (Simon, 1881) and Pselaphochernes scorpioides (Hermann, 1804) [30,31]), and 15% are endemics (see below). The remaining 15% are Asian and Afrotropics (Figure 9). Only two pseudoscorpion species found in Israel are classified as Afrotropic, namely, Calocheirus atopos that occurs in the Arabian Peninsula, Egypt and Sudan, and Rhacochelifer longeunguiculatus Beier, 1963 that occurs in the Arabian Peninsula (Table 3). As Israel is a transitional area between the Palearctic and Afrotropic zoogeographic zones, we would expect more Afrotropic species, however, the Paleoeremic desert belt, that penetrates Israel from the south and east, may serve as a barrier between the Palearctic and Afrotropic faunal regions for species with low dispersal abilities such as pseudoscorpions (except for phoretic species) [29,32]. Thus, Afrotropic pseudoscorpion species found in Israel may be Paleoeremic (Palearctic and Arabia/Sahara) or relicts of fauna that preceded the desertification and found shelter in refugial habitats with stable conditions, such as the endemic species Ayyalonia dimentmani Ćurčić, 2008. The latter shares some morphological features with the chthoniid genera such as Tyrannochthonius, Pseudochthonius, or Paraliochthonius, all are rather tropical in distribution, which indicates that this might be a relict genus of a warmer paleoclimates. This is also the case of the Opiliones genus Haasus Roewer, 1949 and especially H. naasane Aharon et al., 2019. Haasus naasane is found in a humid hot cave in the Judea desert, while the surrounding region is very dry. It was suggested that H. naasane is relict from the period when climate was more humid in this region [11].
The climatic gradient that occurs in Israel allows species of different zoogeographic regions to inhabit the area. Within the transition between climatic regions, species of different zoogeographies can be found alongside one another. Thus, the distribution of pseudoscorpion families (Figure 3) and species (Figure 4) among the eleven biogeographic sub-regions in Israel reflects the climatic gradient. The transition from the more mesic and Mediterranean climate in the north and west of Israel to the drier desert climate in the south and east of the country is reflected by distributional patterns in families of more temperate climates, such as Chthoniidae, Neobisiidae, Garypinidae, Geogarypidae that prevail in the north and west of Israel, and as aridity increases toward the south and east, they become less abundant. In arid regions the Olpiidae species become more dominant, and species of mesic regions disappear (Supplementary Materials Tables S2-S3).
- Endemism
In Israel only nine described species (15%) are endemic, including three species that belong to Afrotropic genera. These three species are not restricted to a certain geographic region, and two of them were found in a host nest, which is considered a habitat with stable conditions. Nannowithius wahrmani was described by Beier [15] as Myrmecowithius, a small myrmecophilous species related to the East African genus Nannowithius (to which it was transferred later by Mahnert [33], which was found in a nest of Messor semirufus in the Negev desert. Messor semirufus was recorded from the Middle East, Mediterranean Basin, Arabian Peninsula, Iran, Afghanistan and as east as Kashmir, but it is believed that it consists of a group of species and needs comprehensive revision [34]. Recently we found N. wahrmani phoretic on the myrmecophilous scorpion Birulatus israelensis which lives only in Messor ebeninus nests in the Jordan Valley [27]. Messor ebeninus is known from Libya, Egypt, Turkey, Arabian Peninsula, Israel, Lebanon, Syria and Iran; and like Messor semirufus group, it is speciose in the Middle East and needs comprehensive revision [34]. Also N. pakistanicus was found in nests of Messor sp., and it is likely that all species of Nannowithius are associated with ants [35]. The second pseudoscorpion species, Nudochernes spalacis, was commonly found in nests of the Israeli blind mole-rat species-complex (Nannospalax spp.). The third pseudoscorpion species, Paramenthus shulovi, belongs to one of the smallest pseudoscorpion families (by means of species richness), that is found in xeric habitats, Menthidae, comprising five genera. Paramenthus shulovi was the first member of the family to be found outside America, but in 2007 Mahnert described from Socotra Island, Yemen, a second Paramenthus species, which was later found also in south Iran [36,37]. Thus, the genus Paramenthus is assumed Paleoeremic and of Afrotropic origin. Five Israeli endemic pseudoscorpions belong to genera of Palearctic origin and were found mostly in relatively mesic habitats such as caves, maquis or near fresh water.
The degree of pseudoscorpion endemism in Israel is very low compared to the endemism pattern in nearby Mediterranean regions such as Turkey and the Balkans, the latter ranked as one of the richest and most versatile fauna of pseudoscorpions in the world [6,23,38]. The Balkan region has high diversity of ecosystems and habitats that offer biological isolates with stable conditions that enhance endemism and support relict and hypogean species.
The Levant pseudoscorpion fauna includes, apart from Israel and Palestine, 43 recorded species; Cyprus with sixteen species, Syria with ten, Lebanon with eleven, Jordan with six, and Egypt with fifteen recorded species. Many of the species are common to the region and are found also in Israel, and some are endemic to a country or region. This low recorded species richness suggests that the pseudoscorpion faunas has hardly been explored in the Levant, and therefore an evaluation of this region’s species richness, zoogeographical patterns and historical biogeography is impossible at this stage.
Following a survey in Israeli caves, new cave-dwelling pseudoscorpions were found, and the endemic chorological category in Israel is expected to increase. In addition, the Hermon and Golan regions that include unique habitats have been less explored and have the potential to harbour species new to the region and new to science.
Endemic rare species, especially those that are limited to a single cave, are often very sensitive to changes in their environment. Ayyalonia dimentmani, for instance, that was found in a previously sealed unique chemoautotrophic cave in 2006 likely became extinct after its discovery, and was never collected again despite targeted surveys. Several of the new species found in caves are represented by a single specimen from a single cave, such as the syarinid, cheiridiid, neobisiid and atemnid. These will require more sampling efforts along with promoting conservation management plans.
- Seasonality
Though there are four distinct seasons in Israel (winter: December-February, spring: March-May, summer: June-August and autumn: September-November), the climate in Israel is characterised by a long dry and hot season starting in spring (April) and ending towards the end of autumn (November), and a short cool season with sporadic rainfall between November to April. Data from this study can only indicate the time of the year a specimen was collected, which, given no methodological survey was carried out, is rather random, and greatly depends on the incidental finding occasion. Yet, exploring the data of ca. 1600 specimens reveals that although pseudoscorpions were found all year round, most of the specimens were found in spring (roughly twice the number in winter, three times the number in autumn, and 1 ½ times the number in summer) (Figure 5A). This pattern is mainly due to the two most common and rather thermophilic families, Cheliferidae and Olpiidae. Even though these random explorations are not evenly distributed along the year, this pattern suits what we would expect regarding vertical movement of soil invertebrates. Disappearance from the surface during the dry season is a common temporal response of soil invertebrates to drought stress in the Mediterranean region [39]. It is therefore not surprising that during the dry summer, there was a sharp decrease in the occurrence of pseudoscorpions, that comprised 0.3% of the total micro-arthropod fauna of the soil, in a survey of soil micro-arthropod fauna in pine forests at the Mediterranean Mount Carmel (Figure 1b, sub-region 7) [40].
- Habitats and Collection methods
All four methods that were reported (pitfall traps, sifting, trunk traps and visual search) were fruitful (Figure 5C). They complement each other and give information regarding the species habitat [41]. Trunk traps is a useful method to capture bark inhabiting pseudoscorpions which include mostly cheliferid species: Dactylochelifer kussariensis, D. pallidus, Hysterochelifer tuberculatus (Lucas, 1849), Hysterochelifer gracilimanus Beier, 1949, Rhacochelifer corcyrensis bicolor Beier 1963, Rhacochelifer corcyrensis (Beier, 1930), Rhacochelifer corcyrensis corcyrensis (Beier, 1930), Rhacochelifer longeunguiculatus, Rhacochelifer peculiaris latissimus Beier 1963 as well as Chernes rhodinus Beier, 1966 and Atemnus syriacus [17]. Pitfall traps are useful for leaf litter and soil inhabitants such as: Olpiidae (rocks and dry litter), Cheliferidae (leaf litter), Neobisiidae and Chthoniidae (under rocks and in moist leaf litter). Sifting has a slightly broader range, captures also Geogarypidae and Garypinidae who reside in moist leaf litter, but is still limited to leaf litter and soil inhabitants. The most fruitful method in terms of diversity and quantity was visual search that can be conducted in all habitats, including specialist habitats such as: under rocks at the seashore for Garypidae, caves for hypogean species, animal nests for Chernetidae or beehives for Chelifer cancroides. This method also yields ecological information, but it is the most time and manpower consuming technique.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, S.W., E.G.-R. and D.H.; resources, S.W., S.A., E.G.-R., I.A.S, P.P.S. and D.H.; investigation, S.W.; formal analysis, S.W.; writing—original draft preparation, S.W.; visualization, S.W. and I.A.S.; Map preparation, S.A.; supervision, E.G.-R. and D.H.; writing—review and editing, E.G.-R., D.H. and P.P.S.; funding acquisition, E.G.-R. and P.P.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by: Fieldwork in Israel was supported by the Israel Taxonomy Initiative (ITI) biodiversity survey grant to E.G.-R. and Y.L. A National Geographic Society Expeditions Council grant no. NGS-271R-18 to P.P.S., J.A.B. and E.G.-R. Our study was supported by the US-Israel Binational Science Foundation grant no. BSF-2019216 to P.P.S. and E.G.-R.
Data Availability Statement
The data presented in this study are available in Supplementary Tables S1-S6.
Acknowledgments
Specimens were collected under permits 2013/40027, 2013/40085, 2014/40313, 2017/41718, 2020/42450, and 2022/43117 from the Israel Nature and Parks Authority to Efrat Gavish-Regev. We thank the Israel Cave Research Center (ICRC), and especially A. Frumkin, S. Ya’aran, B. Langford; The Israel National Arachnid Collection staff, and especially, Z. Ganem, M. Cohen, Y. Zaltz, Y. Zvik, E. Zvik, D.J. Alfred; The Sharma laboratory and especially J.A. Ballesteros, G. Gainett, S. Kulkarni, B. Klementz, H. Steiner, as well as Martina Pavlek and Stefano Mammola for assistance in the field and valuable discussions; We thank Giulio Gardini and Dora Hlebec for the assistance in identifying the syarinid specimen.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- WPC—World Pseudoscorpiones Catalog. Natural History Museum Bern. Available online: https://wac.nmbe.ch/order/pseudoscorpiones/3 (accessed on 12/02/2023).
- Vitali-di Castri, V. Biogeography of Pseudoscorpions in the Mediterranean regions of the world. In Mediterranean Type Ecosystems: Origin and Structure; di Castri, F., Mooney, H.A., Eds.; Springer: Berlin, Heidelberg, Germany, 1973; Ecological Studies; Volume 7, pp. 295–305. [Google Scholar]
- Liebke, D.F.; Harms, D.; Widyastuti, R.; Scheu, S.; Potapov, A.M. Impact of rainforest conversion into monoculture plantation systems on pseudoscorpion density, diversity and trophic niches. Soil Organisms 2021, 93, 83–96. [Google Scholar] [CrossRef]
- Blondel, J.; Aronson, J.; Bodiou, J.Y.; Boeuf, G. The Mediterranean region: biological diversity in space and time. Oxford University Press 2010.
- Harvey, M.S. The smaller arachnid orders: diversity, descriptions and distributions from Linnaeus to the present (1758 to 2007). Zootaxa 2007, 1668, 363–380. [Google Scholar] [CrossRef]
- Hlebec, D.; Podnar, M.; Kučinić, M.; Harms, D. Molecular analyses of pseudoscorpions in a subterranean biodiversity hotspot reveal cryptic diversity and microendemism. Scientific Reports 2023, 13, 430–444. [Google Scholar] [CrossRef]
- Kloog, I.; Sorek-Hamer, M.; Lyapustin, A.; Coull, B.; Wang, Y.; Just, A.C.; Schwartz, J.; Broday, D.M. Estimating daily PM2.5 and PM10 across the complex geo-climate region of Israel using MAIAC satellite-based AOD data. Atmospheric environment 2015, 122, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Goldreich, Y. Climate regions and seasons in Israel. In The Climate of Israel: Observation, Research and Application; Springer: Boston, MA, USA, 2003; pp. 12–22. [Google Scholar]
- Israel Meteorological Service (IMS): map of annual mean precipitation Israel (1991–2020) Climate Atlas of Israel. Available online: https://ims.gov.il/en/ClimateAtlas (accessed on 10/02/2023).
- Zonstein, S.; Marusik, Y.M. Checklist of the spiders (Araneae) of Israel. Zootaxa 2013, 3671, 1–127. [Google Scholar] [CrossRef] [PubMed]
- Aharon, S.; Ballesteros, J.A.; Crawford, A.R.; Friske, K.; Gainett, G.; Langford, B.; Santibáñez-López, C.E.; Ya’aran, S.; Gavish-Regev, E.; Sharma, P.P. The anatomy of an unstable node: a Levantine relict precipitates phylogenomic dissolution of higher-level relationships of the armoured harvestmen (Arachnida: Opiliones: Laniatores). Invertebrate Systematics 2019, 33, 697–717. [Google Scholar] [CrossRef]
- Aharon, S.; Ballesteros, J.A.; Gainett, G.; Hawlena, D.; Sharma, P.P.; Gavish-Regev, E. In the land of the blind: Exceptional subterranean speciation of cryptic troglobitic spiders of the genus Tegenaria (Araneae: Agelenidae) in Israel. Molecular Phylogenetics and Evolution 2023, 183, 107705. [Google Scholar] [CrossRef] [PubMed]
- Ćurčić, B.P.M. Ayyalonia dimentmani, n. g., n. sp. (Ayyaloniini n. trib., Chthoniidae, Pseudoscorpiones) from a cave in Israel. Archives of Biological Sciences, Belgrade 2008, 60, 331–339. [Google Scholar] [CrossRef]
- Gavish-Regev, E.; Aharon, S.; Armiach Steinpress, I.; Seifan, M.; Lubin, Y. A Primer on Spider Assemblages in Levantine Caves: The Neglected Subterranean Habitats of the Levant—A Biodiversity Mine. Diversity 2021, 13, 179. [Google Scholar] [CrossRef]
- Beier, M. Die Pseudoscorpioniden-fauna Israels und einiger angrenzender Gebiete. Israel Journal of Ecology and Evolution 1963, 12, 183–212. [Google Scholar]
- Mahnert, V. Einige Pseudoskorpione aus Israel. Revue suisse de Zoologie 1974, 81, 377–386. [Google Scholar] [CrossRef]
- Halperin, J.; Mahnert, V. On some bark-inhabiting pseudoscorpiones (Arachnida) from Israel. Israel journal of Entomology 1987, 21, 127–128. [Google Scholar]
- Harvey, M.S. Pseudoscorpions of the World, version 3.0. Western Australian Museum, Perth. 2013. Available online: https://museum.wa.gov.au/catalogues-beta/pseudoscorpions (accessed on 12/02/2023).
- Benavides, L.R.; Cosgrove, J.G.; Harvey, M.S.; Giribet, G. Phylogenomic interrogation resolves the backbone of the Pseudoscorpiones tree of life. Molecular phylogenetics and Evolution 2019, 139, 106509. [Google Scholar] [CrossRef]
- Salman, I.N.; Ferrante, M.; Möller, D.M.; Gavish-Regev, E.; Lubin, Y. Trunk refugia: A simple, inexpensive method for sampling tree trunk arthropods. Journal of Insect Science 2020, 20, 5. [Google Scholar] [CrossRef]
- Klein, M. The geomorphology of Israel. In The zoogeography of Israel—The distribution and abundance at a zoogeographical crossroad; Yom-Tov, Y., Tchernov, E., Eds.; Dr W. Junk Publishers: Dordrecht, Boston, Lancaster, 1988; pp. 59–78. [Google Scholar]
- Vigna Taglianti, A.; Audisio, P.A.; Biondi, M.; Bologna, M.A.; Carpaneto, G.M.; De Biase, A.; Fattorini, S.; Piattella, E.; Sindaco, R.; Venchi, A.; Zapparoli, M. A proposal for a chorotype classification of the Near East fauna, in the framework of the Western Palearctic region. Biogeographia—The Journal of Integrative Biogeography 1999, 20, 31–59. [Google Scholar] [CrossRef]
- Ćurčić, N.B.; Dimitrijević, R.N.; Ćurčić, S.B. Checklist of the pseudoscorpions (Arachnida: Pseudoscorpiones) of Serbia. Arthropoda Selecta. Русский артрoпoдoлoгический журнал 2020, 1, 13–27. [Google Scholar] [CrossRef]
- Elverici, M. Impact of olive grove management on spider (Araneae) community in Northwestern Turkey. PhD Thesis, Middle East Technical University, Ankara, Turkey, 2018. [Google Scholar]
- Komnenov, M. Spider fauna of the Osogovo Mt. Range, Northeastern Macedonia. Fauna Balkana 2014, 2, 1–267. [Google Scholar]
- Šťáhlavský, F.; Král, J.; Harvey, M.S.; Haddad, C.R. A karyotype study on the pseudoscorpion families Geogarypidae, Garypinidae and Olpiidae (Arachnida: Pseudoscorpiones). European Journal of Entomology 2006, 103, 277–289. [Google Scholar] [CrossRef]
- Zvik, Y. First record of myrmecophyly in the scorpion Birulatus israelensis (Scorpiones: Buthidae). Arachnologische Mitteilungen 2017, 54, 21–23. [Google Scholar] [CrossRef]
- Beron, P. Regional Arachnogeography. In Zoogeography of Arachnida; Springer: Cham, 2018; pp. 319–624. [Google Scholar]
- Por, F.D. An outline of the zoogeography of the Levant. Zoologica Scripta 1975, 4, 5–20. [Google Scholar] [CrossRef]
- Christophoryová, J.; Gruľa, D.; Krajčovičová, K. New records of pseudoscorpions (Arachnida: Pseudoscorpiones) associated with animals and human habitats in Slovakia and the Czech Republic. Arachnologische Mitteilungen 2017, 53, 67–76. [Google Scholar] [CrossRef]
- Christophoryová, J.; Gruľa, D.; Jablonski, D. First record of the genus Lamprochernes (Pseudoscorpiones: Chernetidae) in Albania. Biharean Biologist 2017, 11, 62–64. [Google Scholar]
- Beron, P. Arachnogeographical comparison between West Palearctic and Afrotropical Areas. Ecologica Montenegrina 2016, 7, 464–506. [Google Scholar] [CrossRef]
- Mahnert, V. Die Pseudoskorpione (Arachnida) Kenyas. Familien Withiidae und Cheliferidae. Tropical Zoology 1988, 1, 39–89. [Google Scholar] [CrossRef]
- Borowiec, L.; Salata, S. Review of ants (Hymenoptera: Formicidae) from Jordan. Annals of the Upper Silesian Museum in Bytom, Entomology 2020, 29, 1–26. [Google Scholar] [CrossRef]
- Harvey, M.S. A review of the taxonomy and biology of pseudoscorpions of Nannowithius and Termitowithius (Pseudoscorpiones, Withiidae), inquilines of social insects. The Journal of Arachnology 2015, 43, 342–352. [Google Scholar] [CrossRef]
- Mahnert, V. Pseudoscorpions (Arachnida: Pseudoscorpiones) of the Socotra Archipelago, Yemen. Fauna of Arabia 2007, 23, 271–307. [Google Scholar]
- Nassirkhani, M.; Vafai Shoushtari, R. The first record of the family Menthidae Chamberlin (Arachnida: Pseudoscorpiones) from Iran. International Journal of Research Studies in Zoology 2015, 1, 27–31. [Google Scholar]
- Kunt, K.B.; Bayram, A.; Yağmur, E.A.; Danışman, T. Checklist of the pseudoscorpions of Turkey (Pseudoscorpionida; Arachnida). Turkish Journal of Arachnology 2008, 1, 70–84. [Google Scholar]
- Legakis, A.; Adamopoulou, C. Temporal responses of soil invertebrate communities to drought stress in two semiarid ecosystems of the Mediterranean. Israel Journal of Zoology 2005, 51, 331–348. [Google Scholar] [CrossRef]
- Broza, M.; Poliakov, D.; Conde, B. The first record of the order Palpigradida (Arachnida) in Israel and the occurrence of arachnids in soils of Mediterranean pine forests. Israel Journal of Zoology 1993, 39, 147–151. [Google Scholar]
- Bedoya-Roqueme, E.; Tizo-Pedroso, E. Techniques for collection and sampling of pseudoscorpions (Arthropoda: Arachnida). In Measuring Arthropod Biodiversity; Springer: Cham, 2021; pp. 341–363. [Google Scholar]
- Beier, M. Die Pseudoskorpione des Wiener Naturhistorischen Museums. II. Panctenodactyli. Annalen des Naturhistorischen Museums in Wien 1929, 341–367. [Google Scholar]
- Beier, M. Über Pseudoscorpione aus Syrien und Palästina. Annalen des Naturhistorischen Museums in Wien 1955, 60, 212–219. [Google Scholar]
Figure 9.
The zoogeographic composition of the pseudoscorpion fauna in Israel. See Table 1 for the Chorotype classification and codes.
Figure 9.
The zoogeographic composition of the pseudoscorpion fauna in Israel. See Table 1 for the Chorotype classification and codes.
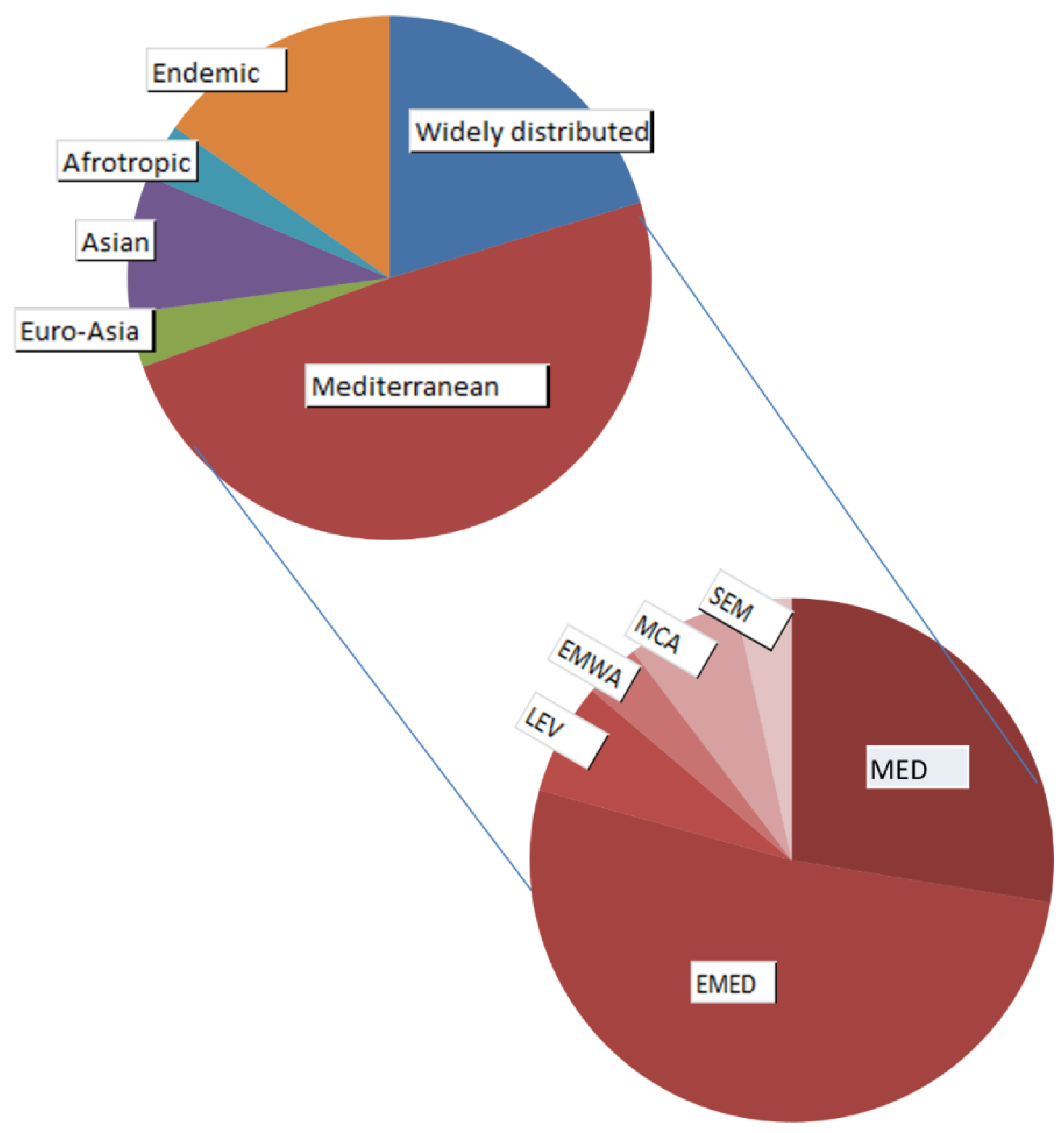
Table 1.
Chorotype classification for the pseudoscorpion fauna in Israel.
| Chorological Complex | Chorotype Classification | Code |
|---|---|---|
| Widely distributed | Cosmopolitan | COS |
| Holarctic | HOL | |
| Old world | OLD | |
| Mediterranean | Circum-Mediterranean | MED |
| East Mediterranean | EMED | |
| Levant | LEV | |
| East-Mediterranean & Western Asia | EMWA | |
| Mediterranean & Central Asia | MCA | |
| South-European & Mediterranean | SEM | |
| Euro-Asia | East-Mediterranean & Central & Western Asia | EMCA |
| Asian | Central & Western Asia | CWA |
| Central & Western Asia & Arabian Peninsula | CWAP | |
| Afrotropic | Arabian Peninsula & Africa | APA |
| Endemic | Endemic to Israel | END |
Table 2.
List of the fourteen pseudoscorpion families of Israel arranged systematically following Benavides et al. [19]. Information on the number of genera and species in Israel, for each family is included.
Table 2.
List of the fourteen pseudoscorpion families of Israel arranged systematically following Benavides et al. [19]. Information on the number of genera and species in Israel, for each family is included.
| Suborder | Infraorder | Superfamily | Family | No. of genera | No. of species | |
|---|---|---|---|---|---|---|
| Valid | Putative | |||||
| HETEROSPHYRONIDA | Chthonioidea | Chthoniidae | 3 | 5 | 2 | |
| IOCHEIRATA | Hemictenata | Neobisioidea | Syarinidae | 1 | 1 | |
| Neobisiidae | 1 | 1 | 2 | |||
| Panctenata | Garypoidea | Geogarypidae | 1 | 2 | ||
| Hesperolpiidae | 2 | 2 | ||||
| Garypidae | 1 | 2 | ||||
| Menthidae | 1 | 1 | ||||
| Olpiidae | 4 | 10 | ||||
| Garypinoidea | Garypinidae | 1 | 2 | |||
| Cheiridioidea | Cheiridiidae | 1 | 1 | |||
| Cheliferoidea | Chernetidae | 6 | 8 | 1 | ||
| Cheliferidae | 4 | 18 | ||||
| Atemnidae | 1 | 1 | 1 | |||
| Withiidae | 1 | 1 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



