
Submitted:
05 June 2023
Posted:
05 June 2023
You are already at the latest version
Abstract
We report the characterization of phenotype, genotype, and functional parameters such as CD4+ T cell, viral load, HIV-1 genotype, class I HLA genotype, CCR5 polymorphism, and IFN-γ producing cells from a cohort of 37 people living with HIV-1 (PLWH) (viremic controllers, were compared those on antiretroviral therapy (ART) from two sites in São Paulo, Brazil. Half the PLWH (51.4%) were classified as viremic controllers (Controller group - CG), compared to 48.6% were on ART. As expected, controllers showed baseline higher levels of CD4+ T cells and nadir compared to those under ART, confirming the ability of HIV-1 controllers to preserve these cells even in the absence of ART. IFN-γ-producing cells showed greater magnitude to the HIV peptides Gag in controllers (p<0.03). Our results demonstrated that HLA protective alleles are associated with strong production of IFN-γ to Gag, Nef and RT epitopes in people living with HIV controllers.
Keywords:
Human Immunodeficiency Virus (HIV)
; Long-Term Non-Progressor (LTNP)
; Elite Controllers (EC)
; Viremic Controllers (VC)
; IFN-γ T cell responses
; Human Leukocyte Antigens (HLA)
1. Introduction
Human immunodeficiency virus type 1 (HIV-1) disease is a pandemic which affects approximately 38.4 million people living with HIV (PLWH) worldwide [1]. For the vast majority of those PLWH in the absence of antiretroviral therapy (ART) eventually causes CD4+ T cell depletion and profound immunodeficiency, resulting in high morbidity and mortality [2,3]. Otherwise, one small group (1-2%) may delay the HIV natural history of the infection, actually they are able to maintain stable CD4+ T cell counts for more than 10 years, even in the absence of ART and are defined as controllers [4,5]. Thus, controllers can be categorized according are able to maintain undetectable viral loads (VL<40 copies/mL) as elite controllers (EC), whereas controllers have the ability to suppress viremia to low levels, and both groups present normal peripheral CD4+ T cell counts without ART [6,7]. For this research, both categories were grouped as controllers.
The pathogenesis of the progression of HIV disease can be influenced by several factors such as: 1) replication of HIV in the lymphoid organs after primary infection [8]; 2) persistent viral replication in the lymphoid organs throughout the course of HIV infection [9,10]; 3) chronic stimulation of the immune system, which can cause inappropriate immune activation and progressive exhaustion of the immune response [11,12]; and 4) destruction of lymphoid tissue, which results in damage to the ability to maintain an HIV-specific immune response and to generate immune responses against new pathogens over time [13,14]. Also, the HIV genetic diversity might be responsible for different clinical evolution [15,16].
The factors that determine slow progression in HIV-1 infected individuals are not fully understood, despite extensive studies from many international HIV cohorts [17,18,19]. Several host characteristics and immune and viral mechanisms have been associated with the effective control of the HIV infection [20,21,22]. Deletions or deleterious mutations of the viral genome implicated in the decreasing of the HIV virulence are the putative causes of non-progression PLWH [23,24,25]. From a host point of view, genetic variations in the CCR5 co-receptor [26,27] and certain HLA alleles are associated with slower progression of the course of HIV infection. HLA- A*01, A*02, A*03, A*25, A*32, B*13, B*14, B*27, B*51, B*57, Cw*08, C*14 and DPB1 alleles are found more frequently in controllers [28,29,30]. Nonetheless, host restriction factors such as APOBEC proteins [31], SAMHD [32], Tetherin [33], and TRIM5a [34,35] can contribute to the control of viral replication.
It is possible that high levels of HIV-specific CD8+ T cells, measured by tetramers or cytokine expression, are present in a strong proportion in controllers. Several studies, few of them from Brazilian cohorts [36,37,38], have shown strong amplitude of the responses directed against a large diversity of HIV epitopes [22,39,40,41]. Considering the above stated, we aimed to characterize the clinical, genetic, immunological and epidemiological profile of a small cohort of controllers in São Paulo, southeast Brazil.
2. Methods
2.1. Subjects, sample collection and follow-up
From October 2012 to December 2022, we cross-sectionally studied 37 HIV-infected patients on follow-up at the outpatient clinic ADEE3002 in the Hospital das Clínicas da Faculdade de Medicina da Universidade de São Paulo (ADEE 3002/HC-FM-USP) and Centro de Referência e Treinamento DST/Aids (CRT DST/Aids-SP) in São Paulo, Brazil.
Before enrollment, participant’s ART history and medical history were obtained, as well as the nadir CD4+ T cell count; the highest recorded plasma HIV RNA level; the CD4+ T cell counts and baseline HIV RNA level. Follow-up visits were scheduled yearly. The methods were carried out according to approved guidelines.
A questionnaire on epidemiological and social data was voluntarily answered by all participants, and we revised their medical records, searching for their laboratory immune and virologic monitoring data. At each visit, we interviewed, examined and measured patients' CD4+ T cell count and HIV RNA level. We also typed Class I HLA for all individuals.
2.2. PBMC separation
Peripheral blood mononuclear cells (PBMC) suspensions were separated from 80 mL heparinized venous blood (BD Vacutainer®, BD Biosciences, San Jose, CA, USA) by density gradient sedimentation over Ficoll-Paque Plus, 1.077 g/L (GE Healthcare Bio-Sciences AB, Upsala, Sweden). Isolated PBMC were then washed twice in RPMI 1640 medium [(Advanced RPMI 1640, Gibco®, Grand Island, NY, USA) supplemented with 200 mM glutamine, 100 U/mL antibiotic-antimycotic, 100 mM non-essential amino acids and 100 mM sodium pyruvate (GibcoTM)], and approximately 107 cells were cryopreserved per vial in liquid nitrogen in freezing solution containing 50% heat-inactivated fetal bovine serum (FBS; GibcoTM), 40% RPMI 1640 medium and 10% dimethyl sulfoxide (DMSO; Sigma-Aldrich®, St Louis, MO, USA) until used in the assays. At the time of the assays, PBMC were rapidly thawed in a 37 °C water bath and washed in RPMI 1640 medium supplemented with 10% FBS. Cells were counted, checked for viability and resuspended in RPMI 1640 medium at a concentration of 106 cells/mL.
2.3. CD4+ / CD8+ T cell Immunophenotyping
The two major T-cell subpopulations were classified on the basis of the expression of CD4+ and CD8+ surface molecules from EDTA-peripheral blood samples (BD Vacutainer®, BD Biosciences). The CD4+/CD8+ cell immunophenotyping was performed as recommended by Brazilian National Network for CD4+/CD8+ T cell immunophenotyping. Briefly, total blood was stained with the four-color Multi-test anti-human CD45, anti-human CD3, anti-human CD4, anti-human CD8 (BD Biosciences) in Trucount tubes (BD Biosciences) for 20 minutes, then a lyse-no-wash step was done using FACS lysing solution, and the sample was analyzed in the FACScalibur flow cytometer (BD Biosciences). We used multiset software to count and get the absolute cell number.
2.4. Quantification of HIV-1 Viral Load
HIV-1 RNA present in plasma was quantified from EDTA-peripheral blood samples and fractionated by amplifying gag region of the viral RNA with homogeneous real-time fluorescence detection, four hours after collection (Abbott RealTime HIV-1®, Abbott Molecular, Illinois, USA), according to the manufacturer's instructions. The detection limit of this test was 40 RNA copies/mL.
2.5. CCR5-Δ32 and CCR5-P-59029A/G Polymorphisms
Genomic DNA was extracted from the buffy coat of the individual’s samples, collected in EDTA-sterile tubes (BD Vacutainer®, BD Biosciences) using a commercial kit QIAamp® genomic DNA Kit (Qiagen, Hilden, Germany) following the manufacturer’s instructions.
CCR5-Δ32 and CCR5-P-59029A/G alleles were determined by the QIAGEN® Multiplex PCR kit. Forward and reverse primers used were: F-5’-CTTCATCATCCTCCTGACAATCG-3’ and R-5’-GACCAGCCCCAAGTTGACTATC-3’ for CCR5-Δ32, F-5’- TGGGGTGGGATAGGGGATAC-3’ and R-5’- TGTATTGAAGGCGAAAAGAATCAG-3’ (Life technologies, Carlsbad, CA, USA) for CCR5-P-59029A/G as described by Kristiansen et al. [42].
The amplification of the wild type allele of CCR5 resulted in 262 bp fragments while the mutant allele (CCR5-Δ32) resulted in amplicons of 230 bp. The Amplification product of CCR5-P-59029A/G (453 pb) was digested with the endonuclease Bsp1286I 5U (New England Biolabs®, UK). The wild type genotype resulted in a 408 bp fragment and the allele CCR5-P-59029A/G was cleaved in two fragments of 45 bp and 363 bp. The amplified and digested products were visualized under UV light in agarose gel 2% stained with Sybr® Safe (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA). All PLWH disclosed were subtype B, as predominant in our cohort in Sao Paulo, Brazil.
2.6. Typing of HLA class I
The HLA class I typing (HLA-A, HLA-B and HLA-C) was performed at the HLA Laboratory of the Fundação Hemocentro de Ribeirão Preto (Fundherp), São Paulo, Brazil, by Sequence-Specific Oligonucleotides PCR amplifications (PCR-SSO), using the commercial LABType® SSO Class I kits: RSSO1A (LABType SSO Class I A Locus), RSSO1B (LABType SSO Class I B Locus) e RSSO1C (LABType SSO Class I C Locus; One Lambda, Inc., USA) according to the manufacturer’s protocol (LABType One-Lambda) (Supplementary Table 1).
2.7. T Cell Responses
2.7.1. HIV antigens
Synthetic 15-mer peptides overlapping by 11 amino acids (aa) and corresponding to the entire HIV-1 Gag, Nef, and reverse transcriptase (RT) sequence, consensus B derived from the HXB2 AA coordinates, from the NIH AIDS Reagent Program (Germantown, USA) and pooled into 23 peptide pools: 5 p24 pools, 3 p17 pools, and 3 PP (i.e., p2/p7/p1/p6) pools for Gag, 4 Nef pools and 8 pools RT. We used 1.0 mg of each of the 281 lyophilized peptides (Supplementary Table 1).
2.7.2. IFN-gamma Enzyme-Linked Immunospot (Elispot) Assays
Interferon (IFN)-γ Elispot assay was performed as described previously [43,44,45]. Briefly, 96-well Multiscreen® ELISPOT plate (EMD Millipore Corporation, Burlington, MA, USA) were coated with 50 μL/well of anti-human IFN-γ antibody (anti-human IFN-γ mAb 1-D1K; Mabtech AB, Nacka Strand, Sweden) overnight. After blocking with 10% foetal bovine serum (FBS), triplicate wells were filled with 1 x 105 PBMC and 4 μg/mL HIV-1 pool peptides. Plates were incubated at 37 °C in 5% CO2 for 20 h. Phytohemagglutinin (PHA; Sigma-Aldrich®; 0.5 μg/mL) and medium alone served as positive and negative controls, respectively. Wells were then washed and spots were detected after the addition of biotinylated anti-human IFN-γ mAb (anti-human IFN-γ mAb 7-B6-1; Mabtech AB; 1.0 μg/mL) (2 h, 37 °C in 5% CO2), followed by streptavidin alkaline phosphatase (Mabtech AB; 1 h, 37 °C in 5% CO2) and substrate (5-bromo-4-chloro-3-indolyl-phosphate//nitroblue tetrazolium (Sigma-Aldrich) incubated at room temperature (25 °C) until the appearance of blue spots. Antigen-specific spot forming cells (SFC)/106 PBMC frequencies were measured on a computer-based system (AID EliSpot Reader System; Autoimmun Diagnostika GmbH, Stranberg, Germany). The SFC number was calculated after subtracting the negative control values (wells with cells in the absence of peptide) from the same subject. The positivity cut-off was ≥ 30 IFN-γ SFC/106 PBMCs after subtracting background.
2.8. Statistical Analysis
We performed the statistical analyses with the aid of GraphPad Prism (version 8.0.1; GraphPad Software, La Jolla, CA, USA). We used one-way analysis of variance (ANOVA) and the nonparametric Kruskal-Wallis and Friedman tests to compare the studied groups. The correlation between variables was quantified using the non-parametric Spearman correlation. P value <0.05 was considered statistically significant.
2.9. Ethics Approval
This study was approved by the Ethical Committee Boards of Hospital das Clínicas da Faculdade de Medicina da Universidade de São Paulo (HC/FMUSP - CAPPesq protocol CAAE 05749912.6.0000.0068) and Centro de Referência e Treinamento DST/AIDS (CRT/DST/Aids-SP - protocol 023/2010).
2.10. Data Availability
Clinical and laboratory data of all HIV-infected patients cohort of controllers (LTNP and EC), from the outpatient clinic ADEE3002, Hospital das Clínicas da Faculdade de Medicina da Universidade de São Paulo (ADEE3002/HC-FM-USP) and Centro de Referência e Treinamento DST/Aids (CRT DST/Aids-SP), which were used and/or analyzed during the current study, such as: CD4+ and CD8+T cell counts; plasma HIV RNA levels; Class I HLA typing, CCR5-Δ32 and CCR5-P-59029A/G polymorphisms and T cell responses to HIV antigens by IFN-γ Elispot, can be requested to the corresponding author.
3. Results
Participants meeting the inclusion criteria were 19 individuals who were defined as controllers, asymptomatic for at least ten years with stable CD4+ T cell counts ≥ 500 cells/µL and no history of ART. These controllers were compared to 18 patients on ART, with stable CD4+ T cell counts ≥ 350 cells/µL independently of VL levels.
3.1. Long-term non-progressors maintain CD4+ T cell counts similar to uninfected donors and TCD4/CD8+ ratio is higher in elite controllers compared to treated patients
We studied the evolution of annual CD4+, CD8+ T cells counts and VL, for 31 of the 37 patients, over a four-year period. CD4+ T cell counts and VL from patients in each group are shown in Figure 1 and Table 1. Median CD4+ T cell counts for controllers was 850 cells/µL (range: 621 – 11693 cells/µL), and 637 cells/µL (401 – 1332 cells/µL) for ART patients, respectively [Figure 1 (A)]. The controller individuals had a low VL (median: 704 copies/mL), and in six of these individuals detectable VL were not found whereas ART patients showed variable detection of VL [Figure 1 (B)]. As expected, controllers had higher T CD4+ cell counts and T CD4+/T CD8+ ratio than patients on ART during the whole four-year period (p<0.05 for both comparisons) (Data not shown for CD4 counts).
3.2. Patients with protective HLA alleles presented higher T-cell responses to HIV-1 Gag, Nef and RT pool of peptides
Controllers were grouped according to class 1 HLA typing (A, B and C locus) and separated into two groups: individuals who carry protective alleles against progression to AIDS and individuals who carry alleles susceptible to progression to the disease. We could find statistically significant responses for the Gag pools (P17 Gag 45-99) versus the C1 locus; P24 (Gag 221-275) versus C1 locus; P2/P1/P7/P6 (Gag 353-407) versus B1 locus, Nef (93-155) versus B1 locus; Nef (145-206) versus A1 locus and B1 locus; RT (213-283) versus B1 locus; RT (333-395) versus B2 locus; and RT (541-599) versus locus B2, respectively. See Table 2.
4. Discussion
In our study, we evaluated clinical, genetic, immunological and epidemiological data profiles controllers, compared with cART-treated individuals and healthy HIV-negative volunteers. By evaluating T cell counts, we identified higher numbers of CD4+ T cell baseline and nadir, and CD4+/CD8+ T cell ratio in controllers compared to cART-treated individuals. These findings may indicate the ability of HIV-1 controllers to preserve CD4+ T cell counts with low or undetectable viremia in the absence of ART. It is important highlighted that most controllers present more than twenty years of known HIV seropositivity.
In the present study, IFN-γ-Elispot assay was used to evaluate HIV-1-specific T cell responses. In this context, PBMCs obtained from our cohort of HIV-1+ subjects mainly recognized a pool of Gag and Nef peptides. Several studies indicate that these pools are commonly recognized due to highly conserved epitope regions [46,47,48]. Furthermore, anti-Gag T cell responses have been inversely correlated with plasma viral load [46,47,48,49,50]. We did not find any significant association between VL or CD4+ T count and breadth and magnitude of the total HIV-1 responses.
IFN-γ production and non-progression to Aids is controversial, for example, a previous study associated non-progression to Aids to Gag-specific CD4+ T cell proliferation, but not T-cells IFN-γ production [51]. Emu et al. [52] reported no difference between controllers and non-controllers individuals in terms of IFN-γ–producing CD4+ T cell response to Gag peptide pools. Likewise, other groups suggested that the impact of T cell responses on the control of viral replication cannot be explained only by quantification of the magnitude and breadth of the T cell response [47,48,53,54,55]. Additionally, secreting IFN-γ is not the only feature of T cells, as they are also involved in proliferative ability; production of other cytokines (e.g., IL-2 and TNF-α), and chemokines (e.g., MIP-1β); and antiviral cytotoxic activity [51,56,57]. In this context, the simultaneous production of two or more immune mediators has been associated with a better HIV control replication [10].
The suppression of HIV without cART depends on viral fitness differences, due to gene mutations or deletions and host genetic factors [20,58]. In this context, some HLA class I alleles (e.g. HLA-B*57, HLA-B*27, HLA-B*14 and HLA-B*51) have been associated with protection against disease [17,18,20,29,30,63,64,65]. In our work, when patients were grouped according to the expression of protective HLA alleles, specifically, HLA-B*57 and/or HLA-B*27, the magnitude of IFN-γ T-cell responses against the pool of peptides Gag, Nef and RT was higher than in individuals presenting HLA alleles not related to HIV replication. However, only two in six controllers individuals from our cohort present HLA-B*57+. In fact, it has been shown that only a few HLA-B*57+ or HLA-B*27+ patients become controllers, although their plasma viral load is lower than that of individuals who do not have protective HLA alleles [61,62,63,64].
HLA genes are the strongest genetic factors that can affect immune response and HIV disease progression [28,29,59,60,64]. The selective pressure on HIV-1 and the disease outcome is strongly associated with HLA-B protective alleles [65]. PLWH that carry HLA-B protective alleles presented higher IFN-γ production against viral antigens [29,59,60,61,65], as shown in our results considering controllers and normal progressors under ART. Other groups have reported different cell-intrinsic mechanisms with the potential to block HIV-1 replication cycles. The cyclin-dependent kinase (CDK) inhibitor p21 is upregulated in PBMC from controllers when compared to both cART–suppressed and uninfected individuals. It is possible to infer this protein reduces the efficacy of HIV-1 reverse transcription [66,67]. Moreover, single nucleotide polymorphisms (SNPs) in the RICH2 and CXCR6 genes have been associated with natural control in different populations [68,69,70]. On the other hand, less-expressed HLA-C variants are associated with poor HIV-1 control and unstable HLA-C variants are associated with higher HIV-1 infectivity. The results showed a significant correlation between rapid progression to AIDS and the presence of two or one unstable HLA-C variants. Thus, unstable HLA-C variants both at genotype and at allele levels and rapid progression to Aids [71].
Genes differentially expressed by controllers, but not by cART-treated individuals, may be involved in the host's intracellular defenses. Luque et al. [72] observed upregulation of genes associated with cell survival in controllers, while genes related to induction of apoptosis were found to be upregulated in patients undergoing cART treatment. In fact, anti-apoptotic profiles of controllers had already been suggested by Bottarel et al. [73].
In conclusion, controller individuals present normal physiologic CD4+ T-cell counts and low or undetectable viral load, even when almost all of them have already more than twenty years of known seropositivity. Furthermore, only seven of 19 controllers display at least one of a classical protective HLA class I allele (B*27/B*57) and none of them presented the CCR5-Δ32 deletion in homozygous alleles. Our results confirm the importance of controllers cohort’s studies. Based on them, different parameters involved in non-progression to Aids were found, and the study of controllers individuals still constitutes an important field of discoveries, being considered a model to vaccine and/or to functional cure [20,21,74].
Acknowledgments/financial disclosure
The authors would like to thank Neifi Hassan Saloum Deghaide, HLA Laboratory, Fundação Hemocentro de Ribeirão Preto (Fundherp), for providing the HLA-I typing, José Eduardo Martins Noemia Mie Orii, and Liã Bárbara Arruda, Laboratory of Dermatology and Immunodeficiencies (LIM 56), Hospital das Clinicas HCFMUSP, for providing viral load assays, flow cytometry immunophenotyping, and support in molecular biology assays, respectively, and Lucas Alberto Medeiros, Clinic ADEE3002, (ADEE3002/HC-FM-USP) for recruitment of HIV seropositive patients and NIH AIDS Reagent Program for providing the HIV-1 peptides. This work was supported by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), fellowship 150854/2012-9 for B. C. M. Da Silva; Departamento de Vigilância, Prevenção e Controle das IST, do HIV/Aids e das Hepatites Virais of Ministry of Health of Brazil, grant 749396/2010, and by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), grant 2013/06584-4, and fellowships 2014/09623-3 for B. C. M. Da Silva; 2015/21701-2 for G. F. Hildebrando and 2018/12460-0 for L. T. Da Silva.
Conflicts of Interest
All authors declared that there is no conflict of interest.
References
- WHO-UNAIDS – Global Aids update. Geneva 2021. Available online: https://www.unaids.org/en/resources/documents/2021/2021-global-aids-update (accessed on 18 April 2023).
- Mindel, A.; Tenant-Flowers, M. ABC of AIDS: Natural history and management of early HIV infection. BMJ 2001, 322, 1290–1293. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo-Gil, E.; Ikediobi, U.; Sutton, R.E. Mechanisms of Virologic Control and Clinical Characteristics of HIV+ Elite/Viremic Controllers. Yale J Biol Med. 2017, 90, 245–259. [Google Scholar]
- Grabar, S.; Selinger-Leneman, H.; Abgrall, S.; Pialoux, G.; Costagliola, D. Prevalence and comparative characteristics of long-term nonprogressors and HIV controller patients in the French Hospital Database on HIV. AIDS 2009, 23, 1163–1169. [Google Scholar] [CrossRef]
- Mandalia, S.; Westrop, S.J.; Beck, E.J.; Nelson, M.; Gazzard, B.G; Imami, N. Are long-term non-progressors very slow progressors? Insights from the Chelsea and Westminster HIV cohort, 1988-2010. PLoS One 2012, 7, e29844. [Google Scholar] [CrossRef] [PubMed]
- Gurdasani, D.; Iles, L.; Dillon, D.G.; Young, E.H.; Olson, A.D.; Naranbhai, V.; Fidler, S.; Gkrania-Klotsas, E.; Post, F.A.; Kellam, P.; Porter, K.; Sandhu, M.S.; UK HIV Genomics Consortium. A systematic review of definitions of extreme phenotypes of HIV control and progression. AIDS 2014, 28, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Salwe, S.; Padwal, V.; Velhal, S.; Sutar, J.; Bhowmick, S.; Mukherjee, S.; Nagar, V.; Patil, P.; Patel, V. Delineation of Homeostatic Immune Signatures Defining Viremic Non-progression in HIV-1 Infection. Front Immunol. 2020, 11, 182. [Google Scholar] [CrossRef]
- Graziosi, C.; Pantaleo, G.; Butini, L.; Demarest, J.F.; Saag, M.S.; Shaw, G.M.; Fauci, A.S. Kinetics of human immunodeficiency virus type 1 (HIV-1) DNA and RNA synthesis uuring Primary HIV-1 Infection. Proc Natl Acad Sci USA 1993, 90, 6405–6409. [Google Scholar] [CrossRef] [PubMed]
- Mzingwane, M.L.; Tiemessen, C.T. Mechanisms of HIV persistence in HIV reservoirs. Rev Med Virol. 2017, 27, e1924. [Google Scholar] [CrossRef]
- Virgilio, M.C.; Collins, K.L. The Impact of Cellular Proliferation on the HIV-1 Reservoir. Viruses 2020, 12, 127. [Google Scholar] [CrossRef]
- Khaitan, A.; Unutmaz, D. Revisiting immune exhaustion during HIV infection. Curr HIV/AIDS Rep. 2011, 8, 4–11. [Google Scholar] [CrossRef]
- Fenwick, C.; Joo, V.; Jacquier, P.; Noto, A.; Banga, R.; Perreau, M.; Pantaleo, G. T-cell exhaustion in HIV infection. Immunol Rev. 2019, 292, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Zeng, M.; Southern, P.J.; Reilly, C.S.; Beilman, G.J.; Chio=pman, J.G.; Schacker, T.W.; Haase, A.T. Lymphoid tissue damage in HIV-1 infection depletes naïve T cells and limits T cell reconstitution after antiretroviral therapy. PLoS Pathog. 2012, 8, e1002437. [Google Scholar] [CrossRef] [PubMed]
- Estes, J.D. Pathobiology of HIV/SIV-associated changes in secondary lymphoid tissues. Immunol Rev. 2013, 254, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Palmer, D.S.; Turner, I.; Fidler, S.; Frater, J.; Goedhals, D.; Gouder, P.; Huang, K-H.G.; Oxenius, A.; Phillips, R.; Shapiro, R.; van Vuuren, C.; McLean, A.R.; McVean, G. Mapping the drivers of within-host pathogen evolution using massive data sets. Nat Commun. 2019, 10, 3017.
- Zhao, J.; Lv, X.; Zhang, L. Ji, H. HIV-1 molecular epidemiology and drug resistance-associated mutations among treatment-naïve blood donors in China. Sci Rep. 2020, 10, 7571. [Google Scholar] [CrossRef] [PubMed]
- Magierowska, M.; Theodorou, I.; Debre, P.; Sanson, F.; Autran, B.; Riviere, Y.; Charron, D.; Franch ALT, IMMUNOCO Study Groups; Costagliola, D. Combined genotypes of CCR5, CCR2, SDF1, and HLA genes can predict the long-term nonprogressor status in human immunodeficiency virus-1-infected individuals. Blood 1999, 93, 936–941.
- Saina, M.C.; Bi, X.; Lihana, R.; Lwembe, R.; Ishizaki, A.; Panikulam, A.; Palakudy, T.; Musoke, R.; Owens, M.; Songok, E.M.; Ichimura, H. Comparison of HIV-1 nef and gag Variations and Host HLA Characteristics as Determinants of Disease Progression among HIV-1 Vertically Infected Kenyan Children. PLoS One 2015, 10, e0137140. [Google Scholar] [CrossRef]
- Loureiro Dos Reis, M.M.; Queiroz, M.A.F.; Da Silva, B.C.M.; Duarte, A.J.S.; Arganaraz, G.A.; Vallinoto, A.C.R.; Arganaraz, E.R. IL6 and FAS/FASL gene polymorphisms may be associated with disease progression in HIV-1-positive ethnically mixed patients. J Med Virol. 2020, 92, 1148–1157. [Google Scholar] [CrossRef]
- Autran, B.; Descours, B.; Avettand-Fenoel, V.; Rouzioux, C. Elite controllers as a model of functional cure. Curr Opin HIV AIDS 2011, 6, 181–187. [Google Scholar] [CrossRef]
- Cockerham, L.R.; Hatano, H. Elite control of HIV: is this the right model for a functional cure? Trends Microbiol. 2015, 23, 71–75. [Google Scholar] [CrossRef]
- Lopez-Galindez, C.; Pernas, M.; Casado, C.; Olivares, I.; Lorenzo-Redondo, R. Elite controllers and lessons learned for HIV-1 cure. Curr Opin Virol. 2019, 38, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Hassaïne, G.; Agostini, I.; Candotti, D.; Bessou, G.; Caballero, M.; Agut, H.; Autran, B.; Barthalay, Y.; Vigne, R. Characterization of human immunodeficiency virus type 1 vif gene in long-term asymptomatic individuals. Virology 2000, 276, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Alexander, L.; Aquino-DeJesus, M.J.; Chan, M.; Andiman, W.A. Inhibition of human immunodeficiency virus type 1 (HIV-1) replication by a two-amino-acid insertion in HIV-1 Vif from a non progressing mother and child. J Virol. 2002, 76, 10533–10539. [Google Scholar] [CrossRef] [PubMed]
- Kondo, M.; Shima, T.; Nishizawa, M.; Sudo, K.; Iwamuro, S.; Okabe, T.; Takebe, Y.; Imai, M. Identification of attenuated variants of HIV-1 circulating recombinant form 01_AE that are associated with slow disease progression due to gross genetic alterations in the nef/long terminal repeat sequences. J Infect Dis. 2005, 192, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Winkler, C.A.; Hendel, H.; Carrington, M.; Smith, M.W.; Nelson, G.W.; O'brien, S.J.; Phair, J.; Vlahov, D.; Jacobson, L.P.; Rappaport, J.; Vasilescu, A.; Bertin-Maghit, S.; An, P.; Lu, W.; Andrieu, J.M.; Schächter, F.; Therwath, A.; Jean-François Zagury, J.F. Dominant effects of CCR2-CCR5 haplotypes in HIV-1 disease progression. J Acquir Immune Defic Syndr. 2004, 37, 1534–1538. [Google Scholar] [CrossRef] [PubMed]
- McLaren, P.J.; Coulonges, C.; Bartha, I.; Lenz, T.L.; Deutsch, A.J.; Bashirova, A.; Buchbinder, S.; Carrington, M.N.; Cossarizza, A.; Dalmau, J. Polymorphisms of large effect explain the majority of the host genetic contribution to variation of HIV-1 virus load. Proc Natl Acad Sci U S A 2015, 112, 14658–14663. [Google Scholar] [CrossRef] [PubMed]
- Fellay, J.; Ge, D.; Shianna, K.V.; Colombo, S.; Ledergerber, B.; Cirulli, E.T.; Urban, T.J.; Zhang, K.; Gumbs, C.E.; Smith, J.P. Common genetic variation and the control of HIV-1 in humans. PLoS Genet. 2009, 5, e1000791. [Google Scholar] [CrossRef]
- Antoni, G.; Guergnon, J.; Meaudre, C.; Samri, A.; Boufassa, F.; Goujard, C.; Lambotte, O.; Autran, B.; Rouzioux, C.; Costagliola, D.; Meyer, L. MHC-driven HIV-1 control on the long run is not systematically determined ar early times post HIV-1 infection. AIDS 2013, 27, 1707–1716. [Google Scholar] [CrossRef]
- Sundaramurthi, J.C., Ashokkumar, M., Swaminathan, S., Hanna, L.E. HLA based selection of epitopes offers a potential window of opportunity for vaccine design against HIV. Vaccine 2017, 35, 5568–5575. [CrossRef]
- Goila-Gaur, R.; Strebel, K. HIV-1 Vif, APOBEC, and intrinsic immunity. Retrovirology 2008, 5, 51. [Google Scholar] [CrossRef]
- Ruffin, N.; Brezar, V.; Ayinde, D.; Lefebvre, C.; Wiesch, J.S.Z.; van Lunzen, J.; Bockhorn, M.; Schwartz, O.; Hocini, H.; Lelievre, J-D. et al. Low SAMHD1 expression following T-cell activation and proliferation renders CD4+ T cells susceptible to HIV-1. AIDS 2015, 29, 519–530. [CrossRef] [PubMed]
- Liberatore, R.A.; Bieniasz, P.D. Tetherin is a key effector of the antiretroviral activity of type I interferon in vitro and in vivo. Proc Natl Acad Sci U S A. 2011, 108, 18097–18101. [Google Scholar] [CrossRef] [PubMed]
- Stremlau, M.; Owens, C.M.; Perron, M.J.; Kiessling, M.; Autissier, P.; Sodroski, J. The cytoplasmic body component TRIM5alpha restricts HIV-1 infection in Old World monkeys. Nature 2004, 427, 848–853. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, S.M., Malik, H.S.; Emerman, M. Restriction of an extinct retrovirus by the human TRIM5alpha antiviral protein. Science 2007, 316, 1756–1758. [CrossRef] [PubMed]
- Côrtes, F.H.; Bello, G.; Vorsatz, C.; Pilotto, J.H.; Guimarães, M.L.; Grinsztejn, B.; Veloso, V.G.; Pinto, A.R.; Morgado, M.G. Higher cross-subtype IFN-γ ELISpot responses to Gag and Nef peptides in Brazilian HIV-1 subtype B- and F1- than in C-infected subjects. Vaccine 2013, 31, 1106–1112. [Google Scholar] [CrossRef] [PubMed]
- Côrtes, F.H.; Passaes, C.P.B.; Bello, G.; Teixeira, S.L.M.; Vorsatz, C.; Babic, D.; Sharkey, M.; Grinsztejn, B.; Veloso, V.; Stevenson, M. HIV controllers with different viral load cutoff levels have distinct virologic and immunologic profiles. J Acquir Immune Defic Syndr. 2015, 68, 377–385. [Google Scholar] [CrossRef]
- Almeida, R.R.; Rosa, D.S.; Ribeiro, S.P.; Santana, V.C.; Kallás, E.G.; Sidney, J.; Sette, A.; Kalil, J.; Cunha-Neto, E. Broad and cross-clade CD4+ T-cell responses elicited by a DNA vaccine encoding highly conserved and promiscuous HIV-1 M-group consensus peptides. PLoS One 2012, 7, e45267. [Google Scholar] [CrossRef] [PubMed]
- Laher, F.; Ranasinghe, S.; Porichis, F.; Mewalal, N.; Pretorius, K.; Ismail, N.; Buus, S.; Stryhn, A.; Carrington, M.; Walker, B.D. HIV Controllers Exhibit Enhanced Frequencies of Major Histocompatibility Complex Class II Tetramer+Gag-Specific CD4+ T Cells in Chronic Clade C HIV-1 Infection. J Virol. 2017, 91, e02477–16. [Google Scholar] [CrossRef]
- Hersperger, A.R., Migueles, S.A., Betts, M.R.; Connors, M. Qualitative features of the HIV-specific CD8+ T-cell response associated with immunologic control. Curr Opin HIV AIDS 2011, 6, 169–173. [CrossRef]
- Ferre, A.L.; Lemongello, D.; Hunt, P.W.; Morris, M.M.; Garcia, J.C.; Pollard, R.B.; Yee, H.F.Jr.; Martin, J.N.; Deeks, S.G.; Shacklett, B.L. Immunodominant HIV-specific CD8+ T-cell responses are common to blood and gastrointestinal mucosa, and Gag-specific responses dominate in rectal mucosa of HIV controllers. J Virol. 2010, 84, 10354–10365. [Google Scholar] [CrossRef]
- Kristiansen, T.B.; Knudsen, T.B., Ohlendorff, S., Eugen-Olsen, J. A new multiplex PCR strategy for the simultaneous determination of four genetic polymorphisms affecting HIV-1 disease progression. J Immunol Methods 2001, 252, 147–151. [CrossRef] [PubMed]
- Dalod, M.; Dupuis, M.; Deschemin, J.C.; Goujard, C.; Deveau, C.; Meyer, L.; Ngo, N.; Rouzioux, C.; Guillet, J.G.; Delfraissy, J.F.; Sinet, M.; Venet, A. Weak anti-HIV CD8(+) T-cell effector activity in HIV primary infection. J Clin Invest. 1999, 104, 1431–1439. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Lu, W.; Samri, A.; Costagliola, D.; Schnuriger, A.; da Silva, B.C.M.; Blanc, C.; Larsen, M.; Theodorou, I; Rouzioux, C. et al. Distinct differentiation profiles of HIV-Gag and Nef-specific central memory CD8+ T cells associated with HLA-B57/5801 and virus control. AIDS 2010, 24, 2323–2329.
- Silva, B.C.; Grassi, M.F.R.; Coutinho, R.; Mascarenhas, R.E.M.; Olavarria, V.N.; Coutinho-Borgo, A.; Kalil, J.; Cunha-Neto, E.; Fonseca, S.G. Mycobacterium tuberculosis epitope-specific interferon-g production in healthy Brazilians reactive and non-reactive to tuberculin skin test. Mem Inst Oswaldo Cruz 2014, 109, 999–1004. [Google Scholar] [CrossRef] [PubMed]
- Edwards, B.H.; Bansal, A.; Sabbaj, S.; Bakari, J.; Mulligan, M.J.; Goepfert, P.A.; Magnitude of functional CD8+ T-cell responses to the gag protein of human immunodeficiency virus type 1 correlates inversely with viral load in plasma. J Virol. 2002, 76, 2298–2305. [CrossRef] [PubMed]
- Addo, M.M.; Yu, X.G.; Rathod, A.; Cohen, D.; Eldridge, R.L.; Strick, D.; Johnston, M.N.; Cordoran, C.; Wurcel, A.G.; Fitzpatrick, C.A. Comprehensive epitope analysis of human immunodeficiency virus type 1 (HIV-1)-specific T-cell responses directed against the entire expressed HIV-1 genome demonstrate broadly directed responses, but no correlation to viral load. J Virol. 2003, 77, 2081–2092. [Google Scholar] [CrossRef]
- Thakar, M.R.; Bhonge, L.S.; Lakhashe, S.K.; Shankarkumar, U.; Sane, S.S.; Kulkarni, S.S.; Mahajan, B.A.; Paranjape, R.S. Cytolytic T lymphocytes (CTLs) from HIV-1 subtype C-infected Indian patients recognize CTL epitopes from a conserved immunodominant region of HIV-1 Gag and Nef. J Infect Dis. 2005, 192, 749–759. [Google Scholar] [CrossRef]
- Kiepiela, P.; Ngumbela, K.; Thobakgale, C.; Ramduth, D.; Honeyborne, E.; Moodley, E.; Reddy, S.; de Pierres, C.; Mncube, Z.; Mkhwanazi, N. CD8+ T-cell responses to different HIV proteins have discordant associations with viral load. Nat Med. 2007, 13, 46–53. [Google Scholar] [CrossRef]
- Zuñiga, R.; Lucchetti, A.; Galvan, P.; Sanchez, S.; Sanchez, C.; Hernandez, A.; Sanchez, H.; Frahm, N.; Linde, C.H.; Hewitt, H.S. Relative dominance of Gag p24-specific cytotoxic T lymphocytes is associated with human immunodeficiency virus control. J Virol. 2006, 80, 3122–3125. [Google Scholar] [CrossRef]
- Dyer, W.B.; Zaunders, J.J.; Yuan, F.F.; Wang, B.; Learmont, J.C.; Geczy, A.F.; Saksena, N.K.; McPhee, D.A.; Gorry, P.R.; Sullivan, J.S. Mechanisms of HIV non-progression; robust and sustained CD4+ T-cell proliferative responses to p24 antigen correlate with control of viraemia and lack of disease progression after long-term transfusion-acquired HIV-1 infection. Retrovirology 2008, 5, 112. [Google Scholar] [CrossRef]
- Emu, B.; Sinclair, E.; Favre, D.; Moretto, W.J.; Hsue, P.; Hoh, R.; Martin, J.N.; Nixon, D.F.; McCune, J.M.; Deeks, S.G. Phenotypic, functional, and kinetic parameters associated with apparent T-cell control of human immunodeficiency virus replication in individuals with and without antiretroviral treatment. J Virol. 2005, 79, 14169–14178. [Google Scholar] [CrossRef] [PubMed]
- Betts, M.R.; Ambrozak, D.R.; Douek, D.C.; Bonhoeffer, S.; Brenchley, J.M.; Casazza, J.P.; Koup, R.A.; Picker, L.J. Analysis of total human immunodeficiency virus (HIV)-specific CD4(+) and CD8(+) T-cell responses: relationship to viral load in untreated HIV infection. J Virol. 2001, 75, 11983–11991. [Google Scholar] [CrossRef] [PubMed]
- Frahm, N.; Korber, B.T.; Adams, C.M.; Szinger, J.J.; Draenert, R.; Addo, M.M.; Feeney, M.E.; Yusim, K.; Sango, K.; Brown, N.V. Consistent cytotoxic-T-lymphocyte targeting of immunodominant regions in human immunodeficiency virus across multiple ethnicities. J Virol. 2004, 78, 2187–2200. [Google Scholar] [CrossRef] [PubMed]
- Adland, E.; Carlson, J.M.; Paioni, P.; Kloverpris, H.; Shapiro, R.; Ogwu, A.; Riddell, L.; Luzzi, G.; Chen, F.; Balachandran, T. Nef-specific CD8+ T cell responses contribute to HIV-1 immune control. PLoS One 2013, 8, e73117. [Google Scholar] [CrossRef] [PubMed]
- Betts, M.R.; Nason, M.C.; West, S.M.; De Rosa, S.C.; Migueles, S.A.; Abraham, J.; Lederman, M.M.; Benito, J.M.; Goepfeert, P.A.; Connors, M. HIV nonprogressors preferentially maintain highly functional HIV-specific CD8(+) T cells. Blood 2006, 107, 4781–4789. [Google Scholar] [CrossRef] [PubMed]
- Deeks, S.G.; Walker, B.D. Human immunodeficiency virus controllers: Mechanisms of durable virus control in the absence of antiretroviral therapy. Immunity 2007, 27, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Lassen, K.G.; Lobritz, M.A.; Bailey, J.R.; Johnston, S.; Nguyen, S.; Lee, B; Chou, T.; Siliciano,R.F.; Markowitz, M.; Arts, E.J. Elite suppressor-derived HIV-1 envelope glycoproteins exhibit reduced entry efficiency and kinetics. PLoS Pathog. 2009, 5, e1000377.
- Fellay, J.; Shianna, K.V.; Ge, D.; Colombo, S.; Ledergerber, B.; Weale, M.; Zhang, K.; Gumbs, C.; Castagna, A.; Cossarizza, A. A whole-genome association study of major determinants for host control of HIV-1. Science 2007, 317, 944–947. [Google Scholar] [CrossRef]
- Blankson, J.N. Effector mechanisms in HIV-1 infected elite controllers: highly active immune responses? Antiviral Res. 2010, 85, 295–302. [Google Scholar] [CrossRef]
- Lécuroux, C.; Sáez-Cirión, A.; Girault, I.; Versmisse, P.; Boufassa, F.; Avettand-Fenoël, V.; Rouzioux, C; Meyer, L.; Pancino, G.; Lambotte, O. et al. Both HLA-B*57 and plasma HIV RNA levels contribute to the HIV-specific CD8+ T cell response in HIV controllers. J Virol. 2014, 88, 176–187.
- Pereyra, F.; Addo, M.M.; Kaufmann, D.E.; Liu, Y.; Miura, T.; Rathod, A.; Baker, B.; Trocha, A.; Rosenberg, R.; Mackey, E. Genetic and immunologic heterogeneity among persons who control HIV infection in the absence of therapy. J Infect Dis. 2008, 197, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Brener, J.; Gall, A.; Hurst, J.; Batorsky, R.; Lavandier, N.; Chen, F.; Edwards, A.; Bolton, C.; Dsouza, R.; Allen, T. Rapid HIV disease progression following superinfection in an HLA-B*27:05/B*57:01-positive transmission recipient. Retrovirology 2018, 15, 7. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, S.L.M.; De Sá, N.B.R.; Campos, D.P.; Coelho, A.B.; Guimarães, M.L.; Leite, T.C.N.F.; Veloso, V.G.; Morgado, M.G. Association of the HLA-B*52 allele with non-progression to AIDS in Brazilian HIV-1-infected individuals. Genes Immun. 2014, 15, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Kiepiela, P.; Leslie, A.; Honeyborne, I.; Ramduth, D.; Thobakgale, C.; Chetty, S.; Rathnavalu, P.; Moore, C.; Pfafferott, K; Hilton, L. et al. Dominant influence of HLA-B in mediating the potential co-evolution of HIV and HLA. Nature 2004, 432, 769–775.
- de Azevedo, S.S.D.; Ribeiro-Alves, M.; Côrtes, F.H.; Delatorre, E.; Spangenberg, L.; Naya, H.; Seito, L.N.; Hoagland, B.; Grinsztejn, B.; Veloso, V.G. Increased expression of CDKN1A/p21 in HIV-1 controllers is correlated with upregulation of ZC3H12A/MCPIP1. Retrovirology 2020, 17, 18. [Google Scholar] [CrossRef] [PubMed]
- Leng, J.; Ho, H.P.; Buzon, M.J.; Pereyra, F.; Walker, B.D.; Yu, X.G.; Chang, E.J.; Lichterfeld, M. A cell-intrinsic inhibitor of HIV-1 reverse transcription in CD4(+) T cells from elite controllers. Cell Host Microbe. 2014, 15, 717–728. [Google Scholar] [CrossRef]
- Paximadis, M.; Ngqobe, R.N.; Chaisson, R.E.; Martinson, N.A.; Tiemessen, C.T. RICH2 is implicated in viraemic control of HIV-1 in black South African individuals. Infect Genet Evol. 2017, 49, 78–87. [Google Scholar] [CrossRef]
- Le Clerc, S.; Coulonges, C.; Delaneau, O.; van Manen, D.; Herbeck, J.T.; Limou, S.; An, P.; Martinson, J.J.; Spadoni, J.L.; Therwath, A. Screening low-frequency SNPS from genome-wide association study reveals a new risk allele for progression to AIDS. J Acquir Immune Defic Syndr. 2011, 56, 279–284. [Google Scholar] [CrossRef]
- Limou, S.; Coulonges, C.; Herbeck, J.T.; van Manen, D.; An, P.; Le Clerc, S.; Delaneau, O.; Diop, G.; Taing, L.; Montes, M. Multiple-cohort genetic association study reveals CXCR6 as a new chemokine receptor involved in long-term non progression to AIDS. J Infect Dis. 2010, 202, 908–915. [Google Scholar] [CrossRef]
- Stefani, C.; Sangalli, A.; Locatelli, E.; Federico, T.; Malerba, G.; Romanelli, M.G.; Argañaraz, G.A.; Da Silva, B.C.M.; Da Silva, A.J.D.; Casseb, J. Increased Prevalence of Unstable HLA-C Variants in HIV-1 Rapid-Progressor Patients. Int J Mol Sci. 2022, 23, 14852. [Google Scholar] [CrossRef]
- Luque, M.C.; Santos, C.C.; Mairena, E.C.; Wilkinson, P.; Boucher, G.; Segurado, A.C.; Fonseca, L.A.; Sabino, E.; Kalil, J.E.; Cunha-Neto, E. Gene expression profile in long-term non progressor HIV infected patients: in search of potential resistance factors. Mol Immunol. 2014, 62, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Bottarel, F.; Bonissoni, S.; Lucia, M.B.; Bragardo, M.; Bensi, T.; Buonfiglio, D.; Mezzatesta, C.; DiFranco, D.; Balotta, C.; Capobianchi, M.R. Decreased function of Fas in patients displaying delayed progression of HIV-induced immune deficiency. Hematol J. 2001, 2, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Sáez-Cirión, A.; Pancino, G.; Sinet, M.; Venet, M.; Lambotte, O.; ANRS EP36 HIV Controllers Study Group. HIV controllers: how do they tame the virus? Trends Immunol. 2007, 28, 532–540. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Immune response anti-Gag using Elispot IFN-γ. The production of IFN-γ by T cell stimulated with the pools of peptides of HIV-1 is shown. Box and whiskers show min and max value, line at median. The mean value of triplicate wells for each individual measured as spot forming units (SFC/106 PBMC). T-test and the nonparametric Mann-Whitney test were used to calculate p values, *p<0.05.
Figure 1.
Immune response anti-Gag using Elispot IFN-γ. The production of IFN-γ by T cell stimulated with the pools of peptides of HIV-1 is shown. Box and whiskers show min and max value, line at median. The mean value of triplicate wells for each individual measured as spot forming units (SFC/106 PBMC). T-test and the nonparametric Mann-Whitney test were used to calculate p values, *p<0.05.
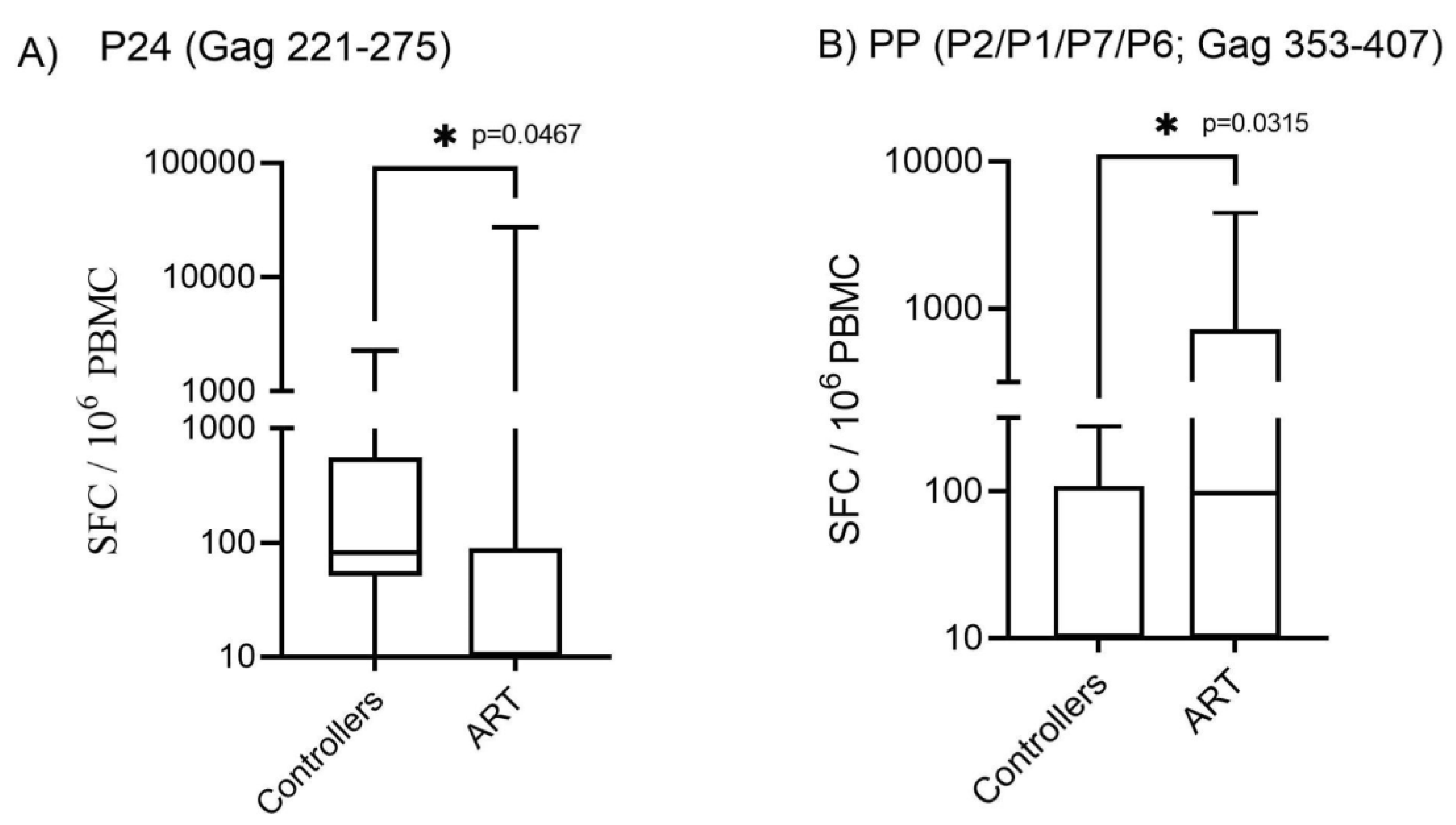

Table 1.
Demographic and baseline data of people living with HIV-1 enrolled in this study (controllers vs ART groups).
Table 1.
Demographic and baseline data of people living with HIV-1 enrolled in this study (controllers vs ART groups).
| Variables | Controllers (C) (n = 19) |
Antiretroviral Treated Individuals (ART) (n = 18) |
P value |
|---|---|---|---|
| Age (years) [median (±SD)] | 51 (±12.08) | 56 (±10.08) | 0.6374 |
| Diagnosis (years) | 17 (±6.405) | 19.5 (±9.41) | 0.8072 |
| Nadir CD4 T cell count (x 106/L) [median (±SD)] | 596 (±186) | 315 (±201) | <0.001 |
| CD4 T cell count (x 106/L) [median (±SD)] | 850 (±331) | 637 (±317) | 0.0073 |
| CD8 T cell count (x 106/L) [median (±SD)] | 978 (±612) | 1104 (±445) | 0.5655 |
| Log10 plasma VL (copies/mL) [median (±SD)] | 704 (±3702) | 25 (±23704) | 0.2239 |
aMedian (standard-deviation) values for age, years of HIV-1 diagnosis, T-cells count and RNA viral load are shown. C: Controllers; ART: antiretroviral treated individuals; VL: viral load. One-way ANOVA and the nonparametric Kruskal-Wallis test were used to calculate P values.
Table 2.
Anti-HIV immune response using Elispot IFN-γ versus HLA immunophenotyping in infection control subjects and elite controllers.
Table 2.
Anti-HIV immune response using Elispot IFN-γ versus HLA immunophenotyping in infection control subjects and elite controllers.
| Peptides | HLA | |||||
|---|---|---|---|---|---|---|
| Locus A1 | Locus A2 | Locus B1 | Locus B2 | Locus C1 | Locus C2 | |
|
P17-1 (Gag 1-55) |
.515 | .077 | .623 | .308 | .620 | .236 |
|
P17-2 (Gag 45-99) |
.523 | .374 | .834 | .104 | .032 | .568 |
|
P17-3 (Gag 89-143) |
.880 | .738 | .304 | .847 | .242 | .774 |
|
P24-1 (Gag 133-187) |
.601 | .602 | .510 | .398 | .726 | .855 |
|
P24-2 (Gag 177-231) |
.231 | .896 | .305 | .152 | .841 | .247 |
|
P24-3 (Gag 221-275) |
.508 | .512 | .175 | .462 | .021 | .459 |
|
P24-4 (Gag 265-319) |
.494 | .413 | .273 | .579 | .181 | .533 |
|
P24-5 (Gag 309-363) |
.532 | .434 | .159 | .906 | .745 | .375 |
|
P2/P1/P7/P6 1 (Gag 353-407) |
.067 | .454 | .009 | .221 | .578 | .127 |
|
P2/P1/P7/P6 2 (Gag 397-451) |
.319 | .572 | .444 | .202 | .801 | .536 |
|
P2/P1/P7/P6 3 (Gag 441-500) |
.291 | .586 | .429 | .549 | .979 | .257 |
|
Nef1 (Nef 1-55) |
.844 | .312 | .145 | .791 | .537 | .071 |
|
Nef2 (Nef 45-103) |
.654 | .507 | .088 | .415 | .863 | .110 |
|
Nef3 (Nef 93-155) |
.093 | .637 | .033 | .094 | .080 | .062 |
|
Nef4 (Nef 145-206) |
.026 | .686 | .002 | .070 | .932 | .053 |
|
RT1 (RT 153-223) |
.070 | .055 | .095 | .590 | .380 | .431 |
|
RT2 (RT 213-283) |
.442 | .880 | .012 | .063 | .770 | .712 |
|
RT3 (RT 273-343) |
.449 | .771 | .065 | .786 | .761 | .372 |
|
RT4 (RT 333-395) |
.134 | .249 | .395 | .027 | .666 | .323 |
|
RT5 (RT 385-455) |
.224 | .944 | .299 | .485 | .851 | .510 |
|
RT6 (RT 445-503) |
.630 | .537 | .347 | .810 | .626 | .280 |
|
RT7 (RT 493-551) |
.714 | .521 | .127 | .390 | .567 | .991 |
|
RT8 (RT 541-599) |
.207 | .805 | .510 | .023 | .975 | .919 |
*Kruskal-wallis test; p=0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



