
Submitted:
03 June 2023
Posted:
05 June 2023
You are already at the latest version
Abstract
Aerobic exercise training (AET) has been used in the management of heart disease. AET may, totally or partially, restore the activity and/or expression of proteins that regulate calcium (Ca2+) handling, optimize intracellular Ca2+ flow, and attenuate cardiac functional impairment in failing hearts. However, the literature presents conflicting data regarding the effects of AET on Ca2+ transit and cardiac function in rats with heart failure resulting from aortic stenosis. The objective of this study was to evaluate the effects of AET on calcium handling and cardiac function in rats with heart failure due to aortic stenosis. Wistar rats were distributed into two groups: control (Sham; n= 61) and aortic stenosis (AoS; n= 44). After 18 weeks, the groups were redistributed into: non-exposed to exercise training (Sham, n = 28 and AoS, n = 22) and trained (Sham-ET, n = 33 and AoS-ET, n = 22) for 10 weeks. Treadmill exercise training was performed with a velocity equivalent to the lactate threshold. Echocardiogram, isolated papillary muscle, and isolated cardiomyocyte analyzed cardiac function. During isolated papillary muscle assay and isolated cardimyocyte was evaluated Ca2+. The expression of regulatory proteins of diastolic Ca2+ was analyzed via Western Blot. AET attenuated the diastolic dysfunction and benefited the systolic function. AoS-ET animals presented better response to post-rest contraction, and SERCA2a and L-type Ca2+ channels blocked than the AoS. Furthermore, AET was able to improve aspects of the mechanical function and the responsiveness of the myofilaments to the Ca2+ of the AoS-ET animals. AoS animals presented alteration in the protein expression of the SERCA2a and NCX, and AET restored SERCA2a and NCX levels near normal values. Therefore, AET increased SERCA2a activity, improved the cellular Ca2+ influx mechanism, and increased myofilament responsiveness to Ca2+, attenuating cardiac dysfunction at cellular, tissue, and chamber levels in animals with aortic stenosis and heart failure.
Keywords:
exercise training
; aortic stenosis
; heart failure
; isolated papillary muscle
; isolated cardiomyocyte
; calcium handling.
1. Introduction
The practice of exercise training plays an important role 1) in improving the physical fitness of practitioners, which is essential to optimize the performance of daily activities [1,2], 2) in the prevention of diseases, making it unlikely that the active individual in youth develops risk factors for chronic diseases [3], 3) in the mitigation of the effects of possible diseases related to aging [1,4,5,6,7], and 4) and for attenuating the consequences of current diseases [1,5,6,8,9,10].
Lower cardiorespiratory fitness and poorer functional capacity and quality of life are common features of heart failure [11,12,13]. Aerobic exercise training (AET) is an important therapeutic tool in order to improve cardiac function and, consequently, the functionality of individuals with heart failure.
Experimental studies evaluating rats with heart failure show that AET improves physical conditioning, functional capacity and cardiac function by enhancing several physiological mechanisms, with the calcium (Ca2+) handling being one of the main factors indicated in positive cardiac adaptation [14,15,16,17,18,19,20,21,22,23,24,25,26]. Specifically, in cardiac dysfunction resulting from different models of experimental aortic stenosis (AoS), the literature presents a lack of studies and divergence of results regarding the participation of Ca2+ handling and its regulatory proteins in the improvement of cardiac function by AET [14,26]. In heart diseased pigs with AoS and preserved ejection fraction, low-intensity aerobic exercise training improved the functional characteristics of the isolated cardiomyocyte, and this positive adequacy is related to the attenuation of the homeostatic damages of Ca2+ handling [14]; the authors observed an increase in the sarcoplasmic reticulum calcium ATPase (SERCA2a)/phospholamban (PLB) ratio, PLB phosphorylated in serine 16 and Na+/Ca2+ exchanger (NCX), in addition to a decrease in diastolic Ca2+. In contrast, van Deel et al [26], in mice with severe aortic stenosis, found that voluntary exercise, incorporated immediately after surgery, did not restore SERCA2a and RyR levels.
Thus, the aim of this study was to evaluate the effects of AET on calcium handling and cardiac function in rats with heart failure due to aortic stenosis. We hypothesize that aerobic training improves cardiac function through positive mechanistic adaptations intrinsic to the myocardium, emphasizing the protagonism of adaptations of the elements and the functioning of calcium handling. Our findings showed that animals with aortic stenosis developed heart failure, and AET was able to generate positive adaptations in physical fitness, in the heart and in the mechanisms of Ca2+ sarcoplasmic reuptake and cellular influx in the myocardium of these rats.
2. Results
2.1. Aerobic exercise training improves systolic and diastolic cardiac function, and signs of heart failure in animals with heart failure resulting from aortic stenosis
The echocardiographic examination was performed before and after aerobic exercise training, that is, after 18 and 28 weeks of the experimental protocol. The first evaluation showed that 18 weeks of aortic stenosis caused important structural and functional cardiac changes (Table 1), such as: predominantly concentric cardiac hypertrophy (↑PWDT, ↑ISDT and ↑RWT), left atrium dilation (↑LA/Ao), and diastolic (↑E wave, ↑E/A, ↑E/E’, and EDT) and systolic dysfunction (↓PWSV). AoS-ET group also showed a decrease in FS and in EF. In the second evaluation, after adding 10 weeks of experimental protocol, the animals submitted to aortic stenosis remained with structural and functional alterations similar to the first evaluation (Table 1). In addition to the losses verified in the first moment, the AoS group presented in comparison to Sham group: ↑LVDD and LVSD, and ↓A’, E’, and FS). AoS-ET animals presented positive cardiac adaptations after an aerobic exercise training protocol in relation to AoS (Table 1), such as: lower magnitude atrial dilatation (↓LA/Ao), and better systolic (↑PWSV and EF) and diastolic function (↑E’, A’, and EDT, and ↓E wave, E/A and E/E’). Furthermore, at the end of 28 weeks, the clinical and pathological signs of the animals were evaluated. The AoS group had a higher frequency of ascites, pleural effusion, left atrial thrombi, hepatic congestion and tachypnea compared to the AoS-ET group (data not shown).
2.2. Aortic stenosis impairs the rate of tension development and resting tension, whereas exercise is effective in restoring resting tension in exercised cardiac animals
Aortic stenosis impaired the myocardial contractile and relaxation function, observed by reduction of maximum rate of tension development, and increased rest tension and time-to-peak tension (Table 2). AET improved muscle relaxation function whereas rest tension decreased in AoS-ET group compared to AoS.
2.3. Aortic stenosis deteriorates SERCA2a function, whereas AET attenuates the loss of SERCA2a functionality in exercised cardiac animals
Figure 1A–C presents papillary muscle response percentage to post-rest contraction (10, 30, and 60 seconds). AoS group showed functional impairment after the pauses performed in the 3 variables studied in relation to Sham, except for the 10-seconds pause for the variables −dT/dt and +dT/dt. AoS-ET group showed worse performance compared to the Sham-ET group after the 30 and 60 seconds pause for the variables DT and +dT/dt and after the 60 seconds pause for −dT/dt. Both findings from the groups with heart disease suggest that aortic stenosis caused a decrease in the calcium recapture potential by the sarcoplasmic reticulum. On the other hand, AoS-ET animals had better performance compared to AoS animals after a 60 seconds pause for the DT and −dT/dt variables, which suggests a positive adequacy in calcium recapture in the exercised group. Figure 1D–F shows papillary muscle response percentage to elevation of extracellular Ca2+ (1.5, 2.5, and 3.5 mM). There were no changes in the functional response between the experimental groups for all variables analyzed.
Figure 2A–C expresses papillary muscle responses to SERCA2a inhibition and calcium concentration increasing. After SERCA2a inhibition, no differences were observed in the functional response between the four experimental groups for the variables studied. Figure 2D–F presents papillary muscle response to L-type calcium channels inhibition and calcium concentration increasing.
The AoS group showed a significantly greater deterioration in performance than the Sham group for all variables studied at practically all times, except for the concentration of 0.5 mM Ca2+ for TD, +dT/dt and −dT/dt (Figure 2A–C), which indicates that these animals have impaired function of L-type Ca2+ channels after 28 weeks of surgery. AET was able to attenuate these deleterious effects of aortic stenosis, since the papillary muscles of AoS-ET animals expressed similar performance to those of the Sham groups during the maneuver, as well as, with statistical support, lower DT, +dT/ dt and −dT/dt depression when compared to AoS animals at concentrations of 1.5, 2.0 and 3.5 mM of Ca2+ (Figure 2A–C). Additionally, there was no difference between the Sham-ET and AoS-ET groups for all variables at all evaluated moments.
Briefly, the post-rest contraction maneuver showed that after the electrical stimulus was paused, AoS animals did not respond inotropically to the return of stimulation; these response would be expected due to the fact that SERCA2a continues to recapture Ca2+ during the stimulus rest. There are three hypothetical causes for this physiological impairment: 1) functional loss of SERCA2a, 2) impairment of mechanisms for Ca2+ entry into the cell and/or Ca2+ release by RyR, and 3) loss of myofilament sensitivity to Ca2+. The answer could have been clarified by the calcium elevation maneuver, since after it increases in the ion extracellular concentration there was no difference in response between the groups. However, AoS animals showed significant functional loss compared to Sham and AoS-ET groups, for all variables studied, after blocking of L-type calcium channels by diltiazem, which shows that there was also impairment of calcium channels function in these animals. As such, in this context, none of the hypotheses mentioned above as explanations for the worst performance in the post-rest contraction maneuver could be completely discarded.
The experimental maneuvers showed that AoS animals have a deterioration of SERCA2a and L-type calcium channels, and that aerobic exercise is an important therapeutic tool to attenuate these deleterious effects of pressure overload caused by aortic stenosis.
2.4. At a cellular level, AET improved the shortening and relaxation function exercised cardiac animals
Figure 3A–F has shown cardiomyocytes mechanical function. AoS group compared to the Sham group showed impairment in the following variables: fractional shortening, shortening maximum velocity, time to 50% shortening, and time to 50% relaxation. Furthermore, AoS-ET group presented worse shortening maximum velocity than Sham-ET. AET was efficient in improving the shortening and relaxation capacity of cardiomyocytes. AoS-ET group showed a higher velocity of shortening and relaxation, shorter time to 50% relaxation, and better fractional shortening compared to AoS group.
2.5. In the isolated cardiomyocyte experiment, AET improves the responsiveness of myofilaments to calcium
Figure 4A–H summarizes calcium handling functioning after 28 weeks of experimental protocol, which included 10 weeks of exercise training. AoS and AoS-ET animals presented alterations in the time to Ca2+ peak, time to 50% of Ca2+ decay, and time to 50% of Ca2+ decay. Aerobic exercise training seems to have improved the sensitivity of myofilaments to calcium in animals submitted to aortic stenosis, since the AoS-ET group had a lower value for Systolic Ca2+/FS and Systolic Ca2+/SMV ratios in relation to the AoS group.
2.6. Aortic stenosis causes changes in calcium handling proteins, which are restored in animals that participated in the AET
Figure 5A–I shows the expression of proteins that regulate myocardial Ca2+ handling. Aortic stenosis caused an increase in SERCA2a and NCX when comparing the Sham and AoS groups (Figure 5B and G), and in PLBthr17 between Sham-ET and AoS-ET (Figure 5I). On the other hand, AET prevented the increase in NCX and SERCA2a in the AoS-ET group; the animals had a lower amount of these proteins in relation to AoS (Figure 5B and G).
2.7. AET improves the functional capacity and metabolic profile of exercise in cardiac animals exercised
Figure 6A–G presents the assessment of cardiorespiratory fitness. Aortic stenosis caused a decrease in exercise tolerance in the stenosis groups, in relation to the respective Sham, in the four stress tests, verified by the decrease in the variables exhaustion velocity and LT velocity (Figures 6A and C), except in the initial T1 and T3 in the second variable mentioned. In addition, the Sham-ET group showed greater cardiorespiratory fitness than the Sham at T2, T3 and T4 (Figures 6C).
The AoS group showed a higher concentration of basal lactate in relation to the Sham group, while the AoS-ET animals showed a lower concentration of basal lactate in relation to the AoS.
AoS animals showed an increase in the relative amount of lactate at lactate threshold and at exhaustion compared to Sham animals (Figure 6F and G). The AoS-ET group showed changes in relation to Sham-ET only in the relative concentration of lactate at the lactate threshold in T2. At T2, T3 and T4, there was a reduction in the relative amount of lactate at the lactate threshold and at exhaustion in the AoS-ET group in relation to the AoS (Figure 6F and G). These outcomes show that aortic stenosis reduced the aerobic capacity of the animals, since the production of lactate, a marker of the activation of anaerobic metabolism, was increased at the mentioned moments in relation to the control animals. On the other hand, the AET was efficient in increasing the aerobic capacity of the animals with heart disease, which produced less lactate at the lactate threshold and at exhaustion when corrected by the velocities at the respective moments.
3. Discussion
The aim of our study was to evaluate the effects of AET on calcium handling and cardiac function in rats with heart failure due to aortic stenosis. Our findings showed that animals with aortic stenosis developed heart failure, and AET was able to generate positive adaptations in physical fitness, in the heart and in the mechanisms of Ca2+ sarcoplasmic reuptake and cellular influx in the myocardium of these rats. In these animals, AET was able to: 1) improve aerobic potential, functional capacity, and metabolic profile of exercise, 2) enhance left ventricular systolic and diastolic function and signs of heart failure, 3) decrease tension during myocardial rest and attenuate the loss of calcium recapture potential by the sarcoplasmic reticulum, 4) improve the functional capacity of cardiomyocytes, favoring positive adjustments in cardiomyocyte contractility and relaxation, and 5) increase the sensitivity of myofilaments to calcium.
The magnitude of systolic and diastolic cardiac dysfunction and the structural changes verified in the echocardiographic exams after 18 and 28 weeks of the experimental protocol are in agreement with previous studies [11,12,27,28,29,30,31,32,33]. The structural adequacy, evidenced by cardiac hypertrophy, after a period of aortic constriction is not functional, presenting a pathological characteristic. There are several pathophysiological mechanisms that contribute to the dysfunctional hypertrophy response, such as: exacerbation of the extracellular matrix, decreased adrenergic responsiveness, changes in contractile proteins, cytoskeleton, and energy metabolism, loss of myocytes by necrosis, apoptosis or autophagy, reprogramming and impairments in the excitation/contraction/relaxation coupling process [34,35,36,37,38,39,40,41,42,43]. Furthermore, the depression of cardiac function was accompanied by the appearance of clinical and pathological signs of heart failure, including: significant decrease in body weight and exercise intolerance, features in cachexia [30], increased RV water content, altered breathing pattern, ascites, pleural effusion, atrial thrombus and hepatic congestion. Findings from in vitro analyzes support the outcomes of the 28-week echocardiographic examination. The animals submitted to aortic stenosis had myocardial stiffness (↑RT) and slowing and decreased ability to develop tension (↓+dT/dt and DT), shorten (↓Fractional shortening, SMV and ↑TS50%) and relax (↑TR50%). Previous studies are in agreement with the data checked in our research [12,14,44].
Due to the complexity of the pathophysiological process of the disease, the treatment of heart failure has been one of the great challenges of the scientific community in recent decades, and the therapeutic success and/or positive prognosis depend on the type, time and intensity of aggression , and the characteristics of the individual [38,45,46]. In addition to the drugs that have been tested in research and used in the treatment of heart failure, the practice of physical activity has great potential to attenuate and/or reverse the process of transition from pathological hypertrophy to heart failure [35,36,47]. In this sense, aerobic exercise training has been consolidated as a therapeutic tool in the management of cardiovascular diseases, both in the preventive context or in the complementary treatment of heart diseases [1,6,8,9,10,16,35,48,49].
Similar to previous studies, low-volume, moderate-intensity AET was implemented at 18 weeks of AoS, that is, after the establishment of ventricular dysfunction in the animals. The type of protocol is justified by the low exercise tolerance of animals with severe pressure overload [11,12,29,30,31,32,33]. This fact was confirmed in the first cardiorespiratory test, after 18 weeks of surgery, in which the animals with heart disease showed exercise intolerance, visualized by the reduced exhaustion velocity in relation to the respective Sham groups. During and at the end of the experimental protocol, the exercised cardiac animals showed significant improvement in exercise tolerance and lactate levels at different intensities of effort, which shows the efficiency of the exercise procedure for this specific group. The increase in cardiac and musculoskeletal aerobic potential by AET became clear from the observation of lower relative concentrations of lactate at the moments of lactate threshold and exhaustion [30,33,50,51]. In addition, the higher speed of exhaustion of the exercised groups expresses the better functional capacity and quality of life of the animals.
In this study, AET attenuated the systolic dysfunction and, mainly, the diastolic damage resulting from the pressure overload imposed by the aortic stenosis, as in previous studies [11,12,29,30,31,32,33]. Trained rats with heart disease showed higher values of EF, PWST, EDT, E' and A' and lower values of LA/Ao, E wave and E/A and E'/A' ratios in relation to the sedentary group; these outcomes point to an increase in the contractile capacity and less myocardial stiffness of the AoS animals. In the literature, in models of myocardial infarction in rats [17,22,23,25], of genetic sympathetic hyperactivity in mice [15,24] and of heart rate overload by ventricular pacemaker in dogs [19], there was prevention of heart failure by AET; authors related the benefits to the improvement of Ca2+ handling and its regulatory agents. Other studies in mice with aortic stenosis found worsening of ventricular dysfunction associated with changes in Ca2+ transit regulatory proteins [26] and increased collagen deposition [26,52] and oxidative stress [52] after 8 weeks of voluntary exercise, started immediately after surgery; the authors proposed that aortic stenosis blocks the beneficial vasodilatory effect of TFa, via activation of endothelial nitric oxide synthase (eNOS), due to fixed aortic obstruction [52]. However, in our study, even without modifying the aggressor agent (aortic stenosis), important functional, systolic and diastolic benefits were detected, making it possible to infer that the positive adaptations are results of the intrinsic improvement of cells and cardiac tissue constituents. In vitro study is in line with the echocardiogram results. AET decreased myocardial stiffness (↓RT) and increased the ability to shorten (↑Fractional shortening and SMV) and relax (↓TR50%). The improvement of fractional shortening and relaxation time in cardiomyocytes from trained cardiac animals, compared to sedentary, was shown in an important study with infarcted rats [25].
Studies have shown, in different experimental models, that calcium handling adjustments are essential for improving the performance of the heart by the AET in heart failure and dysfunction [14,15,16,17,18,19,20,21,22,23,24,25]. According to Kim et al [45], the restoration of protein expression levels close to the normal heart of the elements related to the excitation-contraction-relaxation coupling is among the positive adaptations to the therapies implemented in the course of HF. Our results confirm the proposition, since AoS-ET animals showed lower protein expression of SERCA2a and NCX in relation to AoS, these values being similar to the control groups; however, the data are in disagreement with the literature, which mostly points to maintenance or increase in the expression of SERCA2a by AET in normal, infarcted and hypertensive animals [16,18,25,53]. In pigs with AoS and preserved ejection fraction submitted to AoS, authors observed an increase in the SERCA2a/PLB ratio, serine 16 phosphorylated PLB and NCX, in addition to a decrease in diastolic Ca2+, associated with functional improvements of cardiomyocytes, in relation to animals sedentary [14]. However, van Deel et al [26] did not identify improvement in cardiac function and positive adjustments in calcium handling by voluntary training in mice with AoS; the explanation may be related to the model and severity of aortic stenosis, and the time of implementation and type of exercise protocol. It is important to highlight that, in our study, the decrease in SERCA2a in trained cardiac animals was accompanied by an improvement in cardiomyocyte function in relation to AoS; in the analysis of isolated cardiomyocytes, these rats expressed better values for RMV and TR50% than the sedentary. In addition, the AoS-ET group showed improvement in SMV and shortening fraction, which may be related to the greater sensitivity of myofilaments to Ca2+, proposed by the improvement in the ratios of systolic Ca2+/Fractional shortening and systolic Ca2+/SMV. In agreement, previous studies have shown increased sensitivity of myofilaments to Ca2+ by exercise training in normal [54,55] and infarcted [25] rats.
The maneuvers performed during isolated papillary muscle provided support regarding the effects of AET on SERCA2a and L-channel activity in cardiac animals. In the post-rest contraction and blocking of the ATP binding site of SERCA2a by cyclopiazonic acid, there was no statistical difference between AoS-ET and AoS groups for all variables and moments studied. As there was a significant decrease in SERCA2a expression in the sarcoplasmic reticulum of AoS-ET mice in relation to AoS, the similarity of response suggests a better function of SERCA2a in the AoS-ET group in relation to AoS; this group should have presented better performance after the electrical stimulus pauses and a lower percentage of inhibition after ACP, due to the higher amount of SERCA2a. Authors have shown, in normal [53,56] and infarcted [25] rats, increased SERCA2a activity by training with interval aerobic exercise. As previously mentioned, said protein is ATPase, depending on cytosolic ATP levels to perform its normal functions [57]; in this context, the number and functioning of mitochondria plays a fundamental regulatory role [58]. The literature shows that AET leads to an improvement in mitochondrial function, which is impaired in heart diseases [59], and to a decrease in oxidative stress [60], which is increased in pathological conditions [38]; therefore, in the present study, these mechanisms may have been related to the attenuation of the functional impairments of SERCA2a.
We believe that AET also generates adjustments in the calcium entry mechanism in the diseased cardiac cell, since in the diltiazem blockade experiment there was a better performance of AoS-ET animals in relation to AoS for the variables DT, +dT/dt and –dT /dt. The literature differs regarding the effects of exercise training on calcium current (ICa), which is governed by L channels, in normal rats [53,56,61,62,63]; while authors have shown increased protein expression [56] and adaptive plasticity of L-channels [63], other studies have not detected training effects on ICa and said protein [61,62].
4. Materials and Methods
4.1. Study design
Twenty one-day-old male Wistar rats were submitted to the simulated (Sham, n = 61) or aortic stenosis induction (AoS, n = 44) surgery. After 18 weeks of experimental protocol, the animals were divided into four groups regarding the practice or not of aerobic exercise training for a period of 10 weeks: simulated surgery (Sham, n = 28); Sham plus exercise training (Sham-ET, n = 33); aortic stenosis surgery (AoS, n = 22); and AoS plus exercise training (AoS-ET, n = 22).
The animals were conducted to two lines of experimental analysis:
- The first experimental line evaluated cardiac function by echocardiogram and isolated papillary muscle, and the expression of Ca2+ handling regulatory protein by Western Blot. SERCA2a and L-type calcium channels activity were analyzed during post-rest contraction and calcium elevation (Sham, n = 22; Sham-ET, n = 20; AoS, n = 16; AoS-ET, n = 16), respectively, and by the cumulative administration of extracellular Ca2+ in the presence of SERCA2a or L-type calcium channels specific blockers in the isolated papillary muscle assay. In addition, in order to assess functional capacity and prescribe aerobic exercise training, the animals were submitted to cardiorespiratory fitness tests at weeks 18, 22, 25 and 28 of the experimental protocol.
- the second experimental line evaluated cardiac function by echocardiogram and isolated cardiomyocyte assay, which measured mechanical function and Ca2+ handling (Sham, n = 6; Sham-ET, n = 6; AoS, n = 6; AoS-ET, n = 6). In these animals, the week 28 cardiorespiratory fitness tests were not performed due to technical issues.
As noted, echocardiogram evaluation contemplated all animals of study (Sham, n = 28; Sham-ET, n = 33; AoS, n = 22; AoS-ET, n = 22).
4.2. Animals
The Wistar rats obtained from the Animal Center of Botucatu Medical School (Botucatu, São Paulo, Brazil) were allocated in collective cages at a 23 °C room temperature, with a 12 hours light/dark cycle, a relative humidity of 60%, and water ad libitum. The research was approved by the "Committee for Experimental Research Ethics of the Faculty of Medicine in Botucatu – UNESP”, in accordance with the "Guide for the Care and Use of Laboratory Animals” (protocol 1138/2015).
4.3. Aortic stenosis surgery
AoS was induced surgically as described previously [64,65]. The rats were anesthetized with a mixture of ketamine (50 mg/kg; Dopalen, Sespo Indústria e Comércio Ltda, Divisão Vetbrands, Jacareí, São Paulo, Brazil) and xylazine (10 mg/kg; Anasedan, Sespo Indústria e Comércio Ltda- Divisão Vetbrands, Jacareí, São Paulo, Brazil), and the heart was exposed via a median thoracotomy. Then, a silver clip (0.62 mm of internal diameter) was placed on the ascending aorta at approximately 3 mm from its root, constituting the AoS group. Clinical and pathological signs of heart failure were analyzed by the same investigator (tachypnea, ascites, pleural effusion, left atrium thrombi, and hepatic congestion), who was blind to the different groups.
4.4. Cardiorespiratory fitness test (CFT)
The prescription of aerobic exercise training (AET) and the assessment of capacidade funcional were performed using the cardiorespiratory fitness test (CFT). After 1 week of adaptation to the treadmill, the Sham, Sham-ET, AoS and AoS-ET groups underwent CFT. Before the beginning of the AET, after 18 weeks of surgery, the CFT was performed (T1), with the aim of prescribing the training initial workload. After the 4th and 7th week of AET (T2 and T3), CFTs were performed in order to readjust the training workloads. At the end of the 10th week of AET, CFT was performed again to assess the final physical fitness level of the animals (T4).
The exercise test was started at a speed of 6 m/min, being progressively increased (3 m/min) after 3 minutes, until exhaustion, as described previously [66]. The end of the test was defined by the exhaustion of the animal; the criterion adopted was the non-maintenance of the race, at the imposed speed, for 5 seconds. During the tests, at baseline and after each speed increment, 25 μl of blood was collected from the animal's tail for analysis of lactate concentrations. For the collection, glass slides, 25 × 75 × 1 mm, (Sigma Chemical Company®- USA, modelo Techware S8902), micropipettes, 20 to 200 μl, (Nichiryo Co.®- Japão, Modelo Nichipet NPX 200) and disposable tips were used. Blood samples were stored in 0.5 ml Eppendorf tubes containing 50 μl of 1% sodium fluoride and kept in a freezer until the analysis period. Lactate concentration was determined by the electro-enzymatic method using a lactimeter (Yellow Springs Instruments®, 2300 Stat Plus Glucose & L-Lactate Analyzer- USA).
The AET prescription was based on the analysis of lactate curves, with the daily training speed being equivalent to the lactate threshold (LT). The determination of this point was performed through the graphic plotting of lactate concentrations versus the running speed of the stages. The moment when there was a change in the linearity of the curve as a function of the increase in speed, established by visual inspection, was considered the LT [33]. The variables analyzed to determine the functional capacity of the animals were: velocities of exhaustion and at lactate threshold (LT velocity), lactate concentrations at baseline ([LAC]basal), and at lactate threshold ([LAC]LT), the ratios of lactate concentrations at lactate threshold and at exhaustion by respective velocities at each moment ([LAC](LT)/Vel(LT), and [LAC](Ex)/Vel(Ex)).
4.5. Aerobic exercise training (AET)
AET was started 18 weeks after surgery and involved a rat-specific treadmill running program (Insight Instrumentos – Ribeirão Preto, São Paulo, Brasil) five days a week for 10 consecutive weeks [29,32,33]. The velocity equivalent to the LT was determined as the training intensity. The daily volume progressively increased during AET, presenting the following characteristics: 10, 12, 14 and 16 minutes in duration in weeks 1, 2, 3 and 4, respectively, and 16, 18 and 20 minutes in duration in weeks 5, 6 and 7, respectively. AET volume was maintained at 20 minutes until the end of 10 weeks.
4.6. Cardiac function
4.6.1. Echocardiogram
The echocardiographic study provided data on cardiac structure and function before and after aerobic exercise training, that is, after 18 and 28 weeks of experimental surgery. Commercially available echocardiography (General Electric Medical Systems, Vivid S6, Tirat Carmel, Israel) equipped with a 5–11.5 MHz multifrequency probe was used as previously described [29,31,64]. Rats were anesthetized via intraperitoneal injection with a mixture of ketamine (50 mg/kg; Dopalen®, Sespo Indústria e Comércio Ltda- Divisão Vetbrands, Jacareí, São Paulo, Brasil) and xylazine (0.5 mg/kg; Anasedan®, Sespo Indústria e Comércio Ltda- Divisão Vetbrands, Jacareí, São Paulo, Brasil). The following variables evaluated cardiac structure: LA normalized to the aortic diameter (LA/Ao), left ventricle diastolic diameter (LVDD), left ventricular systolic diameter (LVSD), posterior wall diastolic thickness (PWDT), interventricular septum diastolic thickness (ISDT), and relative wall thickness (RWT). The following parameters assessed ventricular function: heart rate (HR), midwall fraction shortening (FS); ejection fraction (EF); posterior wall systolic velocity (PWSV), early diastolic mitral inflow velocity (E wave), ratio between E wave and atrial contraction flow peak (A wave), E-wave deceleration time (EDT), velocity of the mitral annulus during early ventricular filling (E’), mitral velocity annulus during atrial contraction (A’), and ratio between filling flow peak and mitral annulus velocity during early ventricular filling (E/E’).
4.6.2. Isolated papillary muscle assay
The cardiac contractile performance was evaluated by studying isolated papillary muscles from LV as previously described [27,64]. The following mechanical parameters were measured during isometric contraction: maximum developed tension (DT; g/mm2) and maximum rate of tension development (+dT/dt; g/mm2/s) and decline (−dT/dt; g/mm2/s). Regulator mechanisms of Ca2+ influx and L-type calcium channels activity were analyzed by Ca2+ concentration extracellular elevation maneuver (response percentage), and elevation of extracellular Ca2+ concentrations (0.5, 1.5, 2.5, and 3.5 mM) in the presence and absence of diltiazem (10-5 M; Diltiazem Hydrochoride, Sigma® Aldrich, St Louis, MO, USA), a specific blocker of L-type calcium channels. Post-rest contraction maneuver (response percentage), and elevation of extracellular Ca2+ concentrations (0.5, 1.5, 2.5, and 3.5 mM) in the presence and absence of cyclopiazonic acid (CPA, 30mM; Penicillium cyclopium, Sigma® Aldrich, St Louis, MO, USA), a highly specific blocker of SERCA2a was performed to assess the potential of SERCA2a function. All variables were normalized per cross-sectional area of the papillary muscle (CSA). Seven papillary muscles with CSA >1.5 mm2 were excluded from analysis as they can present central core hypoxia and impaired functional performance [64,65].
4.6.3. Isolated cardiomyocyte assay
- Cardiomyocyte contractility
Cardiomyocyte preparation for isolated cardiomyocyte assessments was performed as described previously [27]. Briefly, isolated cells were placed in an experimental chamber with a glass coverslip base mounted on the stage of an inverted microscope (IonOptix, Milton, MA, USA) edge detection system with a 40× objective lens (Nikon Eclipse – TS100, USA). Cells were immersed in Tyrode’s solution, and field stimulated at 1 Hz (20 V, 5 ms duration square pulses). Cell shortening in response to electrical stimulation was measured with a video-edge detection system at a 240-Hz frame rate (Ionwizard, Ion Optix, Milton, MA, USA) and the contractile parameters were evaluated. Sarcomere length, fractional shortening (expressed as a percentage of resting cell length), maximum shortening velocity, maximum relaxation velocity, and time to 50% shortening (time to 50% peak) and 50% relaxation (time for 50% relaxation) were measured in 6 cells per animal in each experimental group.
- Intracellular Ca2+ measurements
Myocytes were loaded with 1.0 μM Fura2-acetoxymethyl (AM) ester (Molecular Probes, Eugene, OR, USA) for 10 minutes at room temperature, washed with Tyrode solution and allowed to rest for an additional 10 min to allow the desesterification of dye. Subsequently, the cardiomyocytes were stimulated at 1 Hz (Myopacer 100, Ionoptix Inc.) and fluorescence images were obtained using excitation of 340 to 380 nm wavelengths using a Hyper Switch system (Ionwizard, IonOptix, Milton, MA). Background-corrected Fura 2AM ratios, reflecting intracellular Ca2+ concentration detected at approximately 510 nm. Diastolic and systolic Ca2+, time to Ca2+ peak, and time to 50% Ca2+ peak and decay were also analyzed. In addition, the ratios of systolic Ca2+ by shortening fraction and by shortening maximum velocity were performed to evaluate the myofilaments Ca2+ responsiveness.
4.7. Expression of calcium handling protein
The protein expression of the elements responsible for the regulation of calcium handling was analyzed by Western Blot. Fragments of the LV were frozen in liquid nitrogen and stored at −80°C. Frozen samples were subsequently homogenized in RIPA buffer containing protease (SigmaAldrich, St. Louis, MO, USA) and phosphatase (Roche Diagnostics, Indianapolis, IN, USA) inhibitors using a bead beater homogenizer (Bullet Blender®, Next Advance, Inc., NY, USA). The homogenized product was centrifuged (5804R Eppendorf, Hamburg, Germany) at 12,000 rpm for 20 minutes at 4 °C, and the supernatant was transferred to Eppendorf tubes and stored at -80°C. Protein concentration was determined using the Pierce BCA Protein Assay Kit. SDS-PAGE was used to resolve a total of 25 µg protein lysate from each sample. Electrophoresis was performed with biphasic gel stacking (240 mm Tris-HCl pH 6.8, 30% polyacrylamide, APS and TEMED) and resolving (240 mm Tris-HCl pH 8.8, 30% polyacrylamide, APS and TEMED) at a concentration of 6 to 10%, depending on the molecular weight of the analysed protein. The Kaleidoscope Prestained Standard (Bio-Rad, Hercules, CA, USA) was used to identify band sizes. Electrophoresis was performed at 120 V (Power Pac HC 3.0 A, Bio-Rad, Hercules, CA, USA) for 3 h with running buffer (0.25 M Tris, 192 mM glycine, and 1% SDS). Proteins were transferred to a nitrocellulose membrane (Armsham Biosciences, Piscataway, NJ, USA) using a Mini Trans-Blot (Bio-Rad, Hercules, CA, USA) system with transfer buffer (25 mM Tris, 192 mM glycine, 20% methanol, and 0,1% SDS). Membranes were blocked with 5% non-fat dry milk in TBS-T buffer (20 mM Tris-HCl pH 7.4, 137 mM NaCl, and 0.1% Tween 20) for 120 minutes at room temperature under constant agitation. The membrane was washed three times with TBS-T and incubated for 12 h at 4–8 °C under constant agitation with the following primary antibodies: Serca2 ATPase (1:2500; ABR, Affinity BioReagents, Golden, CO, USA), Phospholamban (1:5000; ABR), Phospho-Phospholamban (Ser16) (1:5000; Badrilla, Leeds, West Yorkshire, UK), Phospho–Phospholamban (Thr17) (1:5000; Badrilla), Exchanger Na+/Ca2+ (1:2000; Upstate, Lake Placid, NY, USA), Calcium Channel, VoltageGated Alpha 1C (1:100; Chemicon International, Temecula, CA, USA), Ryanodine Receptor (1:5000; ABR, Affinity Bioreagents, Golden, CO, USA), and GAPDH (1:1000; Santa Cruz Biotechonology Inc., CA, USA). After incubation with the primary antibody, membranes were washed three times in TBS-T and incubated with peroxidase-conjugated secondary antibodies (anti-rabbit or anti-mouse IgG; 1:5000–1:10,000; Abcam) for 2 h under constant agitation. Membranes were then washed three times with TBS-T to remove excess secondary antibody. Blots were incubated with ECL (Enhanced Chemi-Luminescence, Amersham Biosciences, Piscataway, NJ) for chemiluminescence detection by ImageQuant™ LAS 4000 (GE Healthcare). Quantification analysis of blots was performed using Scion Image software (Scion Corporation, Frederick, MD, USA). The immunoblots were quantified by densitometry using ImageJ Analysis software (NIH), and target band results were normalized to the expression of cardiac GAPDH [12]. The Rianodine Receptor is expressed without normalization because it is not possible to evaluate GAPDH as a normalizer in the same gel as the Rianodine Receptor due to the difference between the molecular weight of the two proteins.
4.8. Statistical analysis
Kolmogorov-Smirnov test was used to evaluate the data normality. Data from cardiorespiratory fitness tests, cardiac function, in vivo and in vitro, and calcium handling protein expression analysis are reported as means ± SD or median (25 percentile; 75 percentile). Comparisons between four groups were evaluated using two-way analysis of variance (ANOVA) for independent samples or Kruskal-Wallis, complemented with Bonferroni or Dunn's multiple comparison tests, respectively. Calcium elevation, post-rest contraction, and SERCA2a and L-channels blockage maneuvers are reported as means ± standard deviation, and studied using analysis of variance for repeated measures, complemented with Bonferroni’s multiple comparison test. Data were evaluated at a significance level of 0.05. The statistical analyses were performed using SigmaStat 3.5, and graphics were generated using GraphPad Prism 5.
5. Conclusions
Aerobic exercise training improves myocardial Ca2+ handling in animals with supravalvular aortic stenosis and ventricular dysfunction, since in vitro cardiac performance analysis techniques showed that there was an improvement in the capacity of Ca2+ recapture by the sarcoplasmic reticulum, consequent to the positive adequacy of the activity of the SERCA2a and L-type calcium channels, and increased responsiveness of myofilaments to Ca2+, which supports the positive responses observed in echocardiographic examination and cardiorespiratory fitness tests. Our findings reinforce the importance of controlled exercise training not only in health promotion and disease prevention, but also as a possible tool in the treatment of heart failure. The loss of quality of life and functional capacity of individuals with heart failure is evident, and it is essential to develop strategies to improve this scenario and ensure greater longevity for this population. In addition, the full knowledge of the molecular mechanisms related to the benefits of the failing heart by the different types and intensities of exercise is fundamental for structuring the proposal for intervention with exercise, which must be specific for each type and severity of cardiac pathology. Cardiac improvement on a macro point of view - heart as a pump - is a product of the positive adjustments observed on a micro level - molecular and cellular view. A better heart along with the improvement of other organs by exercise training generates a greater quality of life for individuals with heart failure
Author Contributions
Conceptualization, V.L.d.S., D.H.S.d.C. and A.C.C.; Methodology, V.L.d.S., D.H.S.d.C., S.L.B.d.S., G.A.F.M., A.B.M., D.F.V., P.M.C., P.G.S., and S.G.Z.B.; Software, V.L.d.S., D.H.S.d.C., A.B.M., P.M.C., A.P.L.L., and A.S.L.; Validation, V.L.d.S., D.H.S.d.C., S.L.B.d.S., and G.A.F.M.; Formal Analysis, V.L.d.S., C.P. and A.C.C.; Investigation, V.L.d.S., D.H.S.d.C., S.L.B.d.S., and G.A.F.M.; Resources, V.L.d.S., D.H.S.d.C., S.L.B.d.S., G.A.F.M., and A.C.C.; Data Curation, V.L.d.S., and A.C.C.; Writing – Original Draft Preparation, V.L.d.S.; Writing – Review & Editing, V.L.d.S., G.A.F.M., S.L.B.d.S.,D.H.S.d.C., A.B.M., D.F.V.,P.M.C., P.G.S., C.P., A.P.L.L., S.G.Z.B., A.S.L., A.C.C.; Visualization, V.L.d.S., G.A.F.M., S.L.B.d.S.,D.H.S.d.C., A.B.M., D.F.V.,P.M.C., P.G.S., C.P., A.P.L.L., S.G.Z.B., A.S.L., A.C.C.; Supervision, A.C.C.; Project Administration, V.L.d.S., S.L.B.d.S., G.A.F.M., and A.C.C.; Funding Acquisition, V.L.d.S., D.H.S.d.C., S.L.B.d.S., G.A.F.M, and A.C.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by the São Paulo Research Foundation – FAPESP (2015/20013-5).
Institutional Review Board Statement
The animal study protocol was approved by the Ethics Committee on Animals Experiments of the Botucatu Medical School, São Paulo State University, UNESP (protocol 1138/2015).
Informed Consent Statement
Not applicable.
Data Availability Statement
All data were included in this manuscript. Additional datasets generated during and/or analyzed during the current study are available from the corresponding author upon request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Blair, S.N. Exercício, Envelhecimento Bem-Sucedido e Prevenção das Doenças. In: Fisiologia do Exercício: Nutrição, Energia e Desempenho Humano, 7th edition. Guanabara Koogan: Barueri, São Paulo, 2011, pp. 854–957.
- Egan, B.; Zierath, J.R. Exercise metabolism and the molecular regulation of skeletal muscle adaptation. Cell. Metab. 2013, 17, 162–84. [Google Scholar] [CrossRef]
- Anderson, E.; Durstine, J.L. Physical activity, exercise, and chronic disease: A brief review. SMHS. 2019, 3–10. [Google Scholar] [CrossRef]
- World Health Organization. Available online: www.who.int/news-room/fact-sheets/detail/physical-activity (accessed on 30 May 2023).
- Warburton, D.E.; Nicol, C.W.; Bredin, S.S. Health benefits of physical activity: the evidence. CMAJ. 2006, 174, 801–9. 77. [Google Scholar] [CrossRef] [PubMed]
- Booth, F.W.; Roberts, C.K.; Laye, M.J. Lack of exercise is a major cause of chronic diseases. Compr. Physiol. 2012, 2, 1143–211. [Google Scholar] [PubMed]
- Leskinen, T.; Stenholm, S.; Aalto, V.; Head, J.; Kivimäki, M.; Vahtera, J. Physical activity level as a predictor of healthy and chronic disease-free life expectancy between ages 50 and 75. Age Ageing. 2018, 47, 423–9. [Google Scholar] [CrossRef]
- Zanesco, A.; Antunes, E. Effects of exercise training on the cardiovascular system: pharmacological approaches. Pharmacol. Ther. 2007, 114, 307–17. [Google Scholar] [CrossRef]
- Myers, J. Exercise and Cardiovascular Health. Circulation. 2003, 107, 81. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.K.; Saltin, B.K. Exercise as medicine - evidence for prescribing exercise as 95 therapy in 26 different chronic diseases. Scand. J. Med. Sci. Sports. 2015, 25, 1–72. [Google Scholar] [CrossRef]
- Mota, G.A.F.; de Souza, S.L.B.; da Silva, V.L.S.; Gato, M.; de Campos, D.H.S.; Sant’Ana, P.G.; et al. Cardioprotection Generated by Aerobic Exercise Training is Not Related to the Proliferation of Cardiomyocytes and Angiotensin-(1-7) Levels in the Hearts of Rats with Supravalvar Aortic Stenosis. Cell. Physiol. Biochem. 2020, 54, 719–35. [Google Scholar] [CrossRef]
- de Souza, S.L.B.; Mota, G.A.F.; da Silva, V.L.S.; Sant’Ana, P.G.; Vileigas, D.F.; de Campos, D.H.S.; et al. Adjustments in β-Adrenergic Signaling Contribute to the Amelioration of Cardiac Dysfunction by Exercise Training in Supravalvular Aortic Stenosis. Cell. Physiol. Biochem. 2020, 54, 665–81. [Google Scholar] [CrossRef]
- Kaminsky, L.A.; Arena, R.; Ellingsen, O.; Harber, M.P.; Myers, J.; Ozemek, C.; et al. Cardiorespiratory fitness and cardiovascular disease – The past, present and future. Prog. Cardiovasc. Dis. 2019, 62, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Hiemstra, J.A.; Veteto, A.B.; Lambert, M.D.; Olver, T.D.; Ferguson, B.S.; McDonald, K.S.; et al. Chronic low-intensity exercise attenuates cardiomyocyte contractile dysfunction and impaired adrenergic responsiveness in aortic-banded mini-swine. J. Appl. Physiol. (1985). 2018, 124, 1034–44. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, A.; Rolim, N.P.L.; Oliveira, R.S.F.; Rosa, K.T.; Mattos, K.C.; Casarini, D.E.; et al. Exercise training delays cardiac dysfunction and prevents calcium handling abnormalities in sympathetic hyperactivity-induced heart failure mice. J. Appl. Physiol. 2008, 104, 103–9. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, B.C.; Ooi, J.Y.Y.; Weeks, K.L.; Patterson, N.L.; McMullen, L.R. Understanding key 96 mechanisms of exercise-induced cardiac protection to mitigate disease: current knowlegde and emerging concepts. Physiol. Rev. 2018, 98, 419–75. [Google Scholar] [CrossRef]
- Melo, S.F.S.; Barauna, V.G.; Neves, V.J.; Fernandes, T.; Lara, L.S.; Mazzotti, D.R.; et al. Exercise training restores the cardiac microRNA-1 and -214 levels regulating Ca2+ handling after myocardial infarction. BMC Cardiovasc. Disord. 2015, 15, 166. [Google Scholar] [CrossRef]
- Garciarena, C.D.; Pinilla, A.O.; Nolly, M.B.; Laguens, R.P.; Escudero, E.M.; Cingolani, H.E.; et al. Endurance training in the spontaneously hypertensive rat: conversion of pathological into physiological cardiac hypertrophy. Hypertension. 2009, 53, 708–14. [Google Scholar] [CrossRef]
- Lu, L.; Mei, D.F.; Gu, A.G.; Wang, S.; Lentzner, B.; Gutstein, D.E.; et al. Exercise training normalizes altered calcium-handling proteins during development of heart failure. J. Appl. Physiol. 2002, 92, 1524–30. [Google Scholar] [CrossRef]
- Høydal, M.A.; Stølen, T.O.; Kettlewell, S.; Maier, L.S.; Brown, J.H.; Sowa, T.; et al. Exercise training reverses myocardial dysfunction induced by CaMKII overexpression by restoring Ca2+ homeostasis. J. Appl. Physiol. 2016, 121, 212–20. [Google Scholar] [CrossRef]
- Carneiro-Júnior, M.A.; Quintão-Júnior, J.F.; Drummond, L.R.; Lavorato, V.N.; Drummond, F.R.; Amadeu, M.A.; et al. Effect of exercise training on Ca+2 release units of left ventricular myocytes of spontaneously hypertensive rats. Braz. J. Med. Biol. Res. 2014. [Google Scholar] [CrossRef]
- Kemi, O.J.; MacQuaide, N.; Hoydal, M.A.; Ellingsen, O.; Smith, G.L.; Wisløff, U. Exercise training corrects control of spontaneous calcium waves in hearts from myocardial infarction heart failure rats. J. Cell. Physiol. 2012, 227, 20–6. [Google Scholar] [CrossRef]
- Bito, V.; de Waard, M.C.; Biesmans, L.; Lenaerts, I.; Ozdemir, S.; van Deel, E.; et al. Early exercise training after myocardial infarction prevents contractile but not electrical remodeling or hypertrophy. Cardiovasc. Res. 2010, 86, 72–81. [Google Scholar] [CrossRef]
- Rolim, N.P.; Medeiros, A.; Rosa, K.T.; Mattos, K.C.; Irigoyen, M.C.; Krieger, E.M.; et al. Exercise training improves the net balance of cardiac Ca+2 handling protein expression in heart failure. Physiol. Genomics. 2007, 29, 246–52. [Google Scholar] [CrossRef] [PubMed]
- Wisløff, U.; Loennechen, J.P.; Currie, S.; Smith, G.L.; Ellingsen, O. Aerobic exercise reduces cardiomyocyte hypertrophy and increases contractility, Ca+2 sensitivity and SERCA2a in rat after myocardial infarction. Cardiovasc. Res. 2002, 54, 162–74. [Google Scholar] [CrossRef] [PubMed]
- van Deel, E.D.; de Boer, M.; Kuster, D.W.; Boontje, N.M.; Holemans, P.; Sipido, K.R.; et al. Exercise training does not improve cardiac function in compensated or decompensated left ventricular hypertrophy induced by aortic stenosis. J. Mol. Cell. Cardiol. 2011, 50, 1017–25. [Google Scholar] [CrossRef] [PubMed]
- da Silva, V.L.; de Souza, S.L.B.; Mota, G.A.F.; de Campos, D.H.S.; Melo, A.B.; Vileigas, D.F.; et al. The Dysfunctional Scenario of the Major Components Responsible for Myocardial Calcium Balance in Heart Failure Induced by Aortic Stenosis. Arq. Bras. Cardiol. 2022, 118, 464–75. [Google Scholar]
- de Tomasi, L.C.; Campos, D.H.S.; Sant’Ana, P.G.; Okoshi, K.; Padovani, C.R.; Murata, G.M.; et al. Pathological hypertrophy and cardiac dysfunction are linked to aberrant endogenous unsaturated fatty acid metabolism. PLoS One. 2018, 13. [Google Scholar]
- Pacagnelli, F.L.; Okoshi, K.; Campos, D.H.S.; Souza, R.W.A.; Padovani, C.R.; Carvalho, R.F.; et al. Physical training attenuates cardiac remodeling in rats with supra-aortic stenosis. Exp. Clin. Cardiol. 2014, 20, 1–17. [Google Scholar]
- Gomes, M.J.; Martinez, P.F.; Campos, D.H.; Pagan, L.U.; Bonomo, C.; Lima, A.R.; et al. Beneficial Effects of Physical Exercise on Functional Capacity and Skeletal Muscle Oxidative Stress in Rats with Aortic Stenosis-Induced Heart Failure. Oxid Med Cell Longev. 2016. [Google Scholar] [CrossRef]
- Souza, R.W.A.; Fernandez, G.J.; Cunha, J.P.Q.; Piedade, W.P.; Soares, L.C.; Souza, P.A.T.; et al. Regulation of cardiac microRNAs induced by aerobic exercise training during heart failure. Am. J. Physiol. Heart. Circ. Physiol. 2015, 309, H1629–H1641. [Google Scholar] [CrossRef]
- de Souza, P.A.T.; de Souza, R.W.A.; Soares, L.C.; Piedade, W.P.; Campos, D.H.S.; Carvalho, R.F.; et al. Aerobic training attenuates nicotinic acethylcholine receptor changes in the diaphragm muscle during heart failure. Histol. Histopathol. 2015, 30, 801–11. [Google Scholar]
- Souza, R.W.A.; Piedade, W.P.; Soares, L.C.; Souza, P.A.T.; Aguiar, A.F.; Vechetti-Júnior, I.J.; et al. Aerobic exercise training prevents heart failure-induced skeletal muscle atrophy by anti-catabolic, but not anabolic actions. PloS one. 2014, 9. [Google Scholar] [CrossRef]
- Mann, D.L. Pathophysiology of Heart Failure. In: Braunwald’s Heart Disease - A Textbook of Cardiovascular Medicine, 11th edition. Elsevier. 2012. pp. 487–504.
- Maillet, M.; van Berlo, J.H.; Molkentin, J.D. Molecular basis of physiological heart growth: fundamental concepts and new players. Nat. Rev. Mol. Cell. Biol. 2013, 14, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Sadoshima, J. Mechanisms of physiological and pathological cardiac hypertrophy. Nat. Rev. Cardiol. 2018, 15, 387–407. [Google Scholar] [CrossRef]
- Schirone, L.; Forte, M.; Palmerio, S.; Yee, D.; Nocella, C.; Angelini, F.; et al. A Review of the Molecular Mechanisms Underlying the Development and Progression of Cardiac Remodeling. Oxid. Med. Cell. Longev. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Diwan, A.; Hill, J.A.; Force, T.L. Molecular Basis for Heart Failure. In: Heart Failure: A Companion to Braunwald’s Heart Disease, 3th edition. Elsevier. 2016. pp. 1-27.
- Katz, A.M. Heart Failure. In: Physiology of the Heart, 5th edition. Wolters Kluwer. 2011. p. 510-47.
- Plitt, G.D.; Spring, J.T.; Moulton, M.J.; Agrawal, D.K. Mechanisms, diagnosis, and treatment of heart failure with preserved ejection fraction and diastolic dysfunction. Expert. Rev. Cardiovasc. Ther. 2018, 16, 579–89. [Google Scholar] [CrossRef]
- van Heerebeek, L.; Paulus, W.J. Alterations in Ventricular Function: Diastolic Heart Failure. In: Heart Failure: A Companion to Braunwald’s Heart Disease, 3th edition. Elsevier. 2016. p. 156- 76.
- Gu, J.; Zhao, F.; Wang, Y.; Gao, J.; Wang, X.; Xue, J.; et al. The Molecular Mechanism of Diastolic Heart Failure. Integr. Med. Int. 2015, 143–8. [Google Scholar] [CrossRef]
- Souders, C.A.; Borg, T.K.; Banerjee, I.; Baudino, T.A. Pressure overload induces early morphological changes in the heart. Am. J. Pathol. 2012, 181, 1226–35. [Google Scholar] [CrossRef]
- Silveira, C.F.S.M.P.; Campos, D.H.S.; Freire, P.P.; Deus, A.F.; Okoshi, K.; Padovani, C.R.; et al. Importance of SERCA2a on early isolated diastolic dysfunction induced by supravalvular aortic stenosis in rats. Braz. J. Med. Biol. Res. 2017, 50. [Google Scholar] [CrossRef]
- Kim, G.H.; Uriel, N.; Burkhoff, D. Reverse remodeling and myocardial recovery in heart failure. Nat. Rev. Cardiol. 2018, 15, 83–96. [Google Scholar] [CrossRef]
- Wikman-Coffelt, J.; Parmley, W.W.; Mason, D.T. The Cardiac Hypertrophy Process. Analyses of Factors Determining Pathological Vs. Physiological Development. Circ. Res. 1979, 45, 697–707. [Google Scholar] [CrossRef]
- Scheuer, J.; Malhotra, A.; Hirsch, C.; Capasso, J. Physiologic Cardiac Hypertrophy Corrects Contratile Protein Abnormalities Associated with Pathologic Hypertrophy in Rats. J. Clin. Invest. 1982, 70, 1300–5. [Google Scholar] [CrossRef]
- Agarwal, S.K. Cardiovascular benefits of exercise. Int. J. Gen. Med. 2012, 5, 541–5. [Google Scholar] [CrossRef] [PubMed]
- Gielen, S.; Schuler, G.; Adams, V. Cardiovascular effects of exercise training: molecular mechanisms. Circulation. 2010, 122, 1221–38. [Google Scholar] [CrossRef]
- Brito Vieira, W.H.; Halsberghe, M.J.; Schwantes, M.L.; Perez, S.E.; Baldissera, V.; Prestes, J.; et al. Increased lactate threshold after five weeks of treadmill aerobic training in rats. Braz. J. Biol. 2014, 74, 444–9. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A. Anaerobic threshold: its concept and role in endurance sport. Malays J. Med. Sci. 2004, 11, 24–36. [Google Scholar]
- van Deel, E.D.; Octavia, Y.; Waard, M.C.; Boer, M.; Duncker, D.J. Exercise training has contrasting effects in myocardial infarction and pressure overload due to divergent endothelial nitric oxide synthase regulation. Int. J. Mol. Sci. 2018, 19, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kemi, O.J.; Ceci, M.; Condorelli, G.; Smith, G.L.; Wisløff, U. Myocardial sarcoplasmic reticulum Ca2+ ATPase function is increased by aerobic interval training. Eur. J. Cardiovasc. Prev. Rehabil. 2008, 15, 145–8. [Google Scholar] [CrossRef]
- Wisløff, U.; Loennechen, J.P.; Falck, G.; Beisvag, V.; Currie, S.; Smith, G.; et al. Increased contractility and calcium sensitivity in cardiac myocytes isolated from endurance trained rats. Cardiovasc Res. 2001, 50, 495–508. [Google Scholar] [CrossRef]
- Kemi, O.J.; Haram, P.M.; Loennechen, J.P.; Osnes, J.B.; Skomedal, T.; Wisløff, U.; et al. Moderate vs. high exercise intensity: Differential effects on aerobic fitness, cardiomyocyte contractility, and endothelial function. Cardiovasc Res 2005, 67, 161–72. [Google Scholar] [CrossRef]
- Morán, M.; Saborido, A.; Megías, A. Ca+2 regulatory systems in rat myocardium are altered by 24 weeks treadmill training. Pflugers Arch. 2003, 446, 161–8. [Google Scholar] [CrossRef]
- Katz, A.M. Excitation-Contraction Coupling: Extracellular and Intracellular Calcium Cycles. In: Physiology of the Heart, 5th edition. Wolters Kluwer. 2011. p. 143-76.
- Wang, K.; Xu, Y.; Sun, Q.; Long, J.; Liu, J.; Ding, J. Mitochondria regulate cardiac contraction through ATP-dependent and independent mechanisms. Free Radic. Res. 2018, 3, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Campos, J.C.; Queliconi, B.B.; Bozi, L.H.M.; Bechara, L.R.G.; Dourado, P.M. M, Andres AM. Exercise reestablishes autophagic flux and mitochondrial quality control in heart failure. Autophagy. 2017, 13, 1304–17. [Google Scholar] [CrossRef] [PubMed]
- Silva Jr, S.D.; Jara, Z.P.; Peres, R.; Lima, L.S.; Scavone, C.; Montezano, A.C. Temporal changes in cardiac oxidative stress, inflammation and remodeling induced by exercise in hypertension: Role for local angiotensin II reduction. PLoS One. 2017, 12. [Google Scholar] [CrossRef]
- Mokelke, E.A.; Palmer, B.M.; Cheung, J.Y.; Moore, R.L. Endurance training does not affect intrinsic calcium current characteristics in rat myocardium. Am. J. Physiol. – Heart Circ. Physiol. 1997, 42, H1193–7. [Google Scholar] [CrossRef] [PubMed]
- Delgado, J.; Saborido, A.; Morán, M.; Megías, A. Chronic and acute exercise do not alter Ca+2 regulatory systems and ectonucleotidase activities in rat heart. J. Appl. Physiol. 1999, 87, 152–6. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zheng, J.; Shu, M.; Zhu, S.; Jie, D. Swimming training can a V ect intrinsic calcium current characteristics in rat myocardium. Eur. J. Appl. Physiol. 2008, 104, 549–55. [Google Scholar] [CrossRef] [PubMed]
- Bregagnollo, E.A.; Zornoff, L.A.M.; Okoshi, K.; Sugizaki, M.; Mestrinel, M.A.; Padovani, C.R.; et al. Myocardial contractile dysfunction contributes to the development of heart failure in rats with aortic stenosis. Int. J. Cardiol. 2007, 117, 109–14. [Google Scholar] [CrossRef]
- Bregagnollo, E.A.; Mestrinel, M.A.; Okoshi, K.; Carvalho, F.C.; Bregagnollo, I.F.; Padovani, C.R.; et al. Relative role of left ventricular geometric remodeling and of morphological and functional myocardial remodeling in the transition from compensated hypertrophy to heart failure in rats with supravalvar aortic stenosis. Arq. Bras. Cardiol. 2007, 88, 225–33. [Google Scholar] [CrossRef]
- Rodrigues, B.; Figueroa, D.M.; Mostarda, C.T.; Heeren, M.V.; Irigoyen, M.C.; De Angelis, K. Maximal exercise test is a useful method for physical capacity and oxygen consumption determination in streptozotocin-diabetic rats. Cardiovasc Diabetol. 2007, 6. [Google Scholar] [CrossRef]
Figure 1.
Response percentage to post-rest contraction (A–C) and elevation of extracellular calcium concentration (D–F) from baseline (Ca2+ concentration: 0.5 mM). DT: maximum developed tension; +dT/dt: maximum rate of tension development; -dT/dt: maximum rate of tension decline. Data are expressed as means ± SD of maneuver response percentage. Sham: animals submitted to simulated surgery (n = 22); Sham-ET: animals submitted to simulated surgery and exercise training (n = 20); AoS: animals submitted to aortic stenosis surgery (n= 16); AoS-ET: animals submitted to aortic stenosis surgery and exercise training (n = 16). Analysis of variance for repeated measures and Bonferroni post-hoc test. p < 0.05. a vs. 10 seconds, b vs. 10 seconds and 30 seconds, a vs. 1.5 Ca2+, b vs. 1.5 and 2.5 Ca2+, * AoS vs. Sham, # AoS-ET vs. Sham-ET, § AoS-ET vs. AoS.
Figure 1.
Response percentage to post-rest contraction (A–C) and elevation of extracellular calcium concentration (D–F) from baseline (Ca2+ concentration: 0.5 mM). DT: maximum developed tension; +dT/dt: maximum rate of tension development; -dT/dt: maximum rate of tension decline. Data are expressed as means ± SD of maneuver response percentage. Sham: animals submitted to simulated surgery (n = 22); Sham-ET: animals submitted to simulated surgery and exercise training (n = 20); AoS: animals submitted to aortic stenosis surgery (n= 16); AoS-ET: animals submitted to aortic stenosis surgery and exercise training (n = 16). Analysis of variance for repeated measures and Bonferroni post-hoc test. p < 0.05. a vs. 10 seconds, b vs. 10 seconds and 30 seconds, a vs. 1.5 Ca2+, b vs. 1.5 and 2.5 Ca2+, * AoS vs. Sham, # AoS-ET vs. Sham-ET, § AoS-ET vs. AoS.
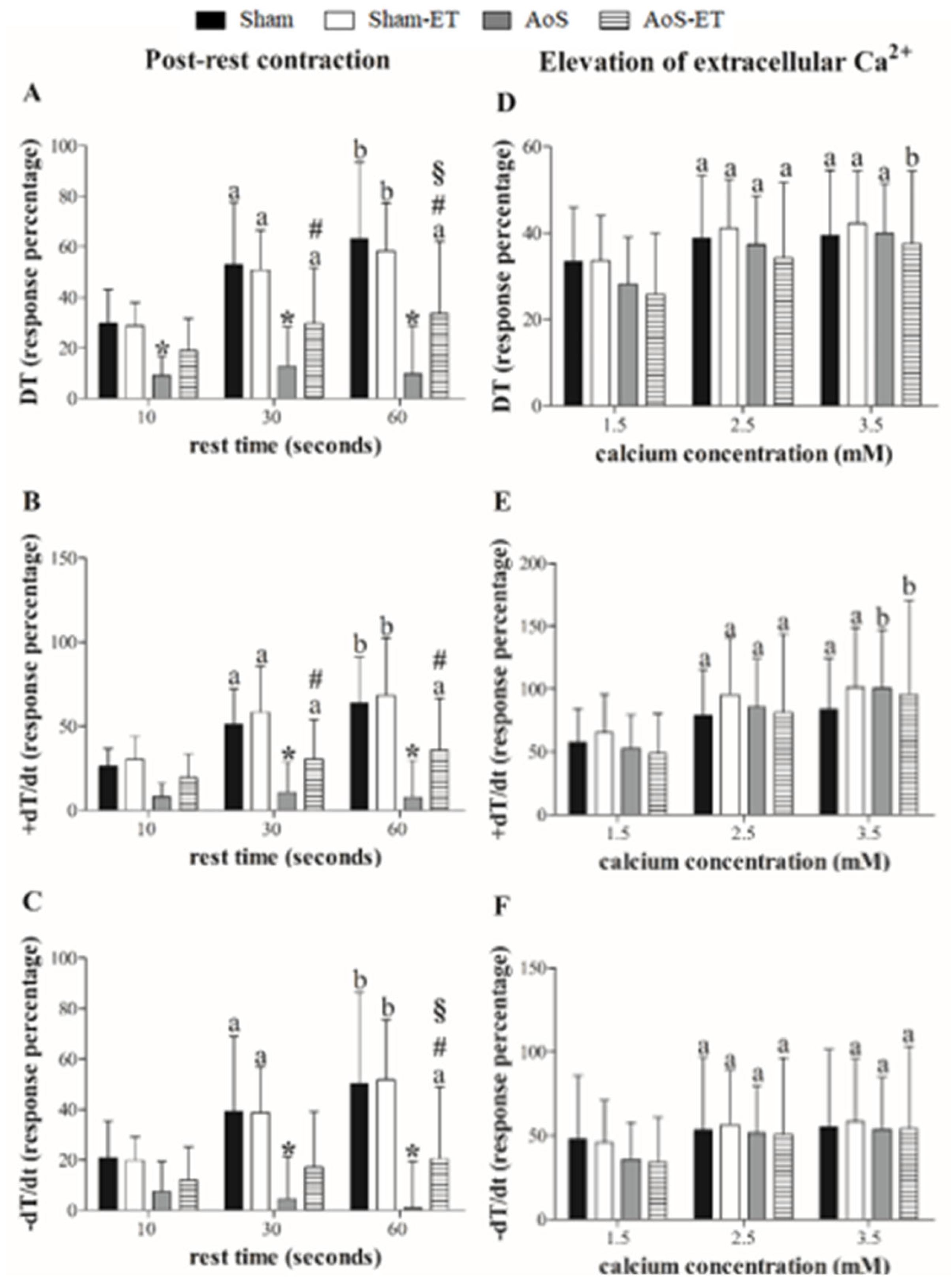
Figure 2.
Inhibition percentage of DT (maximum developed tension), +dT/dt (maximum rate of tension development), and –dT/dt (maximum rate of tension decline) to cyclopiazonic acid (SERCA2a blocker; A–C) and diltiazem (L-type calcium channels blocker; D–F) plus incremental calcium concentration. Data are expressed as means ± SD. Sham: animals submitted to simulated surgery (n = 11); Sham-ET: animals submitted to simulated surgery and exercise training (n = 10); AoS: animals submitted to aortic stenosis surgery (n = 8); AoS-ET: animals submitted to aortic stenosis surgery and exercise training (n = 8). Analysis of variance for repeated measures and Bonferroni post-hoc test. p < 0.05. a vs. 0.5 Ca2+, b vs. 0.5 and 1.5 Ca2+, c vs. 0.5, 1.5, and 2.5 Ca2+, * AoS vs. Sham, § AoS-ET vs. AoS.
Figure 2.
Inhibition percentage of DT (maximum developed tension), +dT/dt (maximum rate of tension development), and –dT/dt (maximum rate of tension decline) to cyclopiazonic acid (SERCA2a blocker; A–C) and diltiazem (L-type calcium channels blocker; D–F) plus incremental calcium concentration. Data are expressed as means ± SD. Sham: animals submitted to simulated surgery (n = 11); Sham-ET: animals submitted to simulated surgery and exercise training (n = 10); AoS: animals submitted to aortic stenosis surgery (n = 8); AoS-ET: animals submitted to aortic stenosis surgery and exercise training (n = 8). Analysis of variance for repeated measures and Bonferroni post-hoc test. p < 0.05. a vs. 0.5 Ca2+, b vs. 0.5 and 1.5 Ca2+, c vs. 0.5, 1.5, and 2.5 Ca2+, * AoS vs. Sham, § AoS-ET vs. AoS.
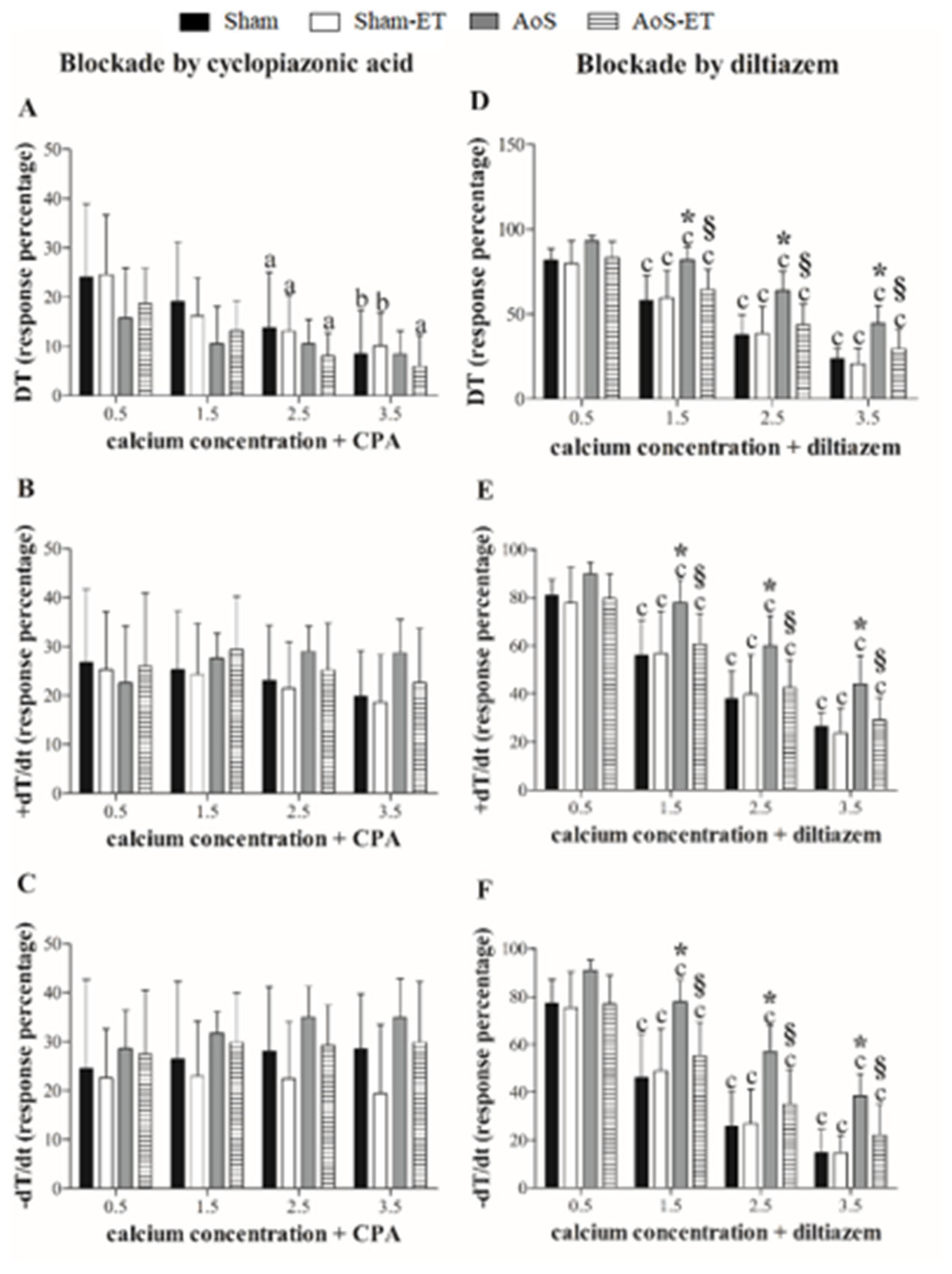
Figure 3.
Cardiomyocytes mechanical function. SMV: shortening maximum velocity; RMV: relaxation maximum velocity; TS50%: time to 50% shortening; TR50%: time to 50% relaxation. Data are expressed as means ± SD or median (25 percentile; 75 percentile). Sham: animals submitted to simulated surgery (n = 6; number of cells = 71); Sham-ET: animals submitted to simulated surgery and exercise training (n = 6; number of cells = 76); AoS: animals submitted to aortic stenosis surgery (n = 6; number of cells = 96); AoS-ET: animals submitted to aortic stenosis surgery and exercise training (n = 6; number of cells = 100). Analysis of variance and Bonferroni post-hoc test or Kruskal-Wallis and Dunn’s method post-hoc. p < 0.05. & Sham vs. Sham-ET; * AoS vs. Sham; # AoS-ET vs. Sham-ET; § AoS-ET vs. AoS.
Figure 3.
Cardiomyocytes mechanical function. SMV: shortening maximum velocity; RMV: relaxation maximum velocity; TS50%: time to 50% shortening; TR50%: time to 50% relaxation. Data are expressed as means ± SD or median (25 percentile; 75 percentile). Sham: animals submitted to simulated surgery (n = 6; number of cells = 71); Sham-ET: animals submitted to simulated surgery and exercise training (n = 6; number of cells = 76); AoS: animals submitted to aortic stenosis surgery (n = 6; number of cells = 96); AoS-ET: animals submitted to aortic stenosis surgery and exercise training (n = 6; number of cells = 100). Analysis of variance and Bonferroni post-hoc test or Kruskal-Wallis and Dunn’s method post-hoc. p < 0.05. & Sham vs. Sham-ET; * AoS vs. Sham; # AoS-ET vs. Sham-ET; § AoS-ET vs. AoS.
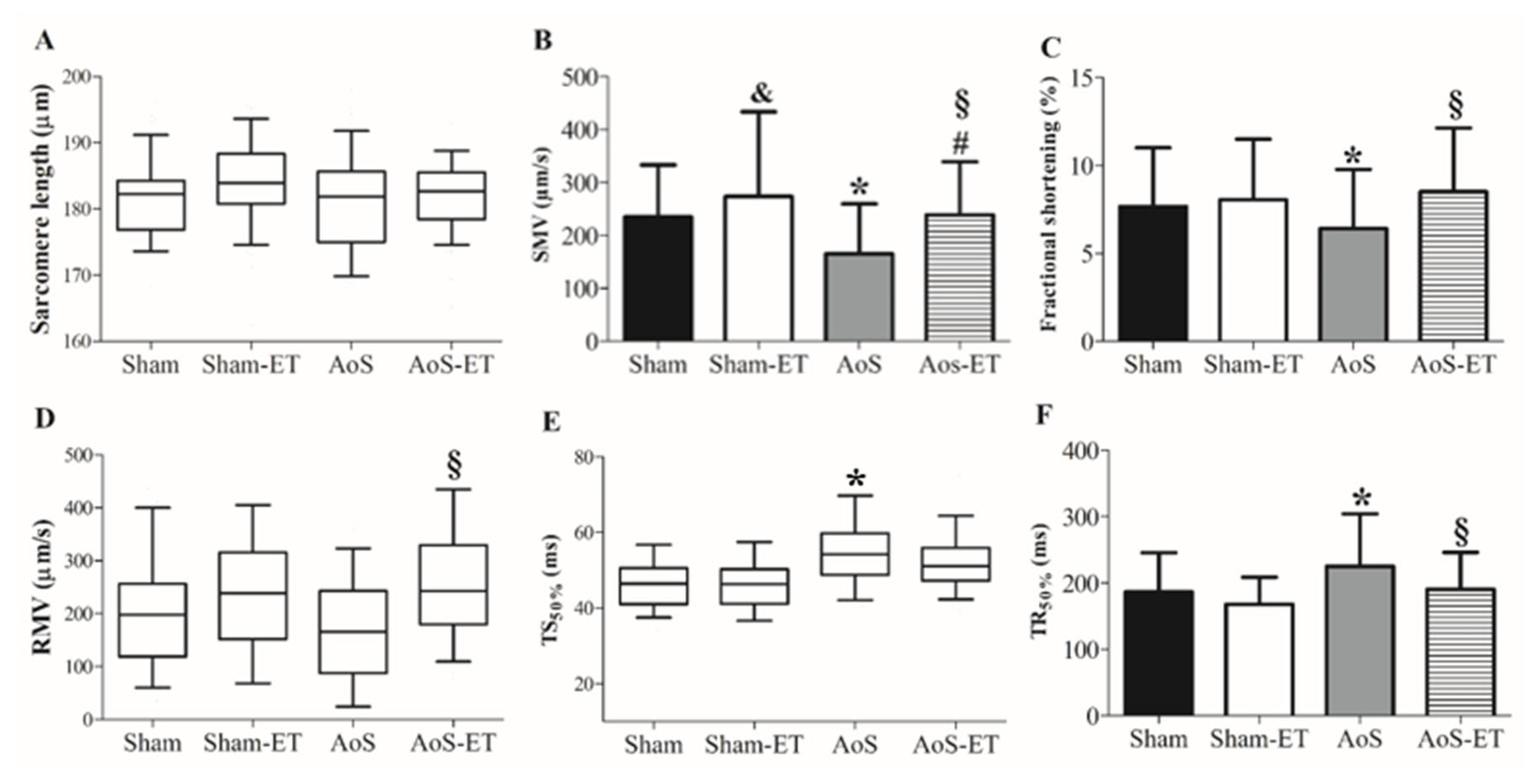
Figure 4.
Cardiomyocyte calcium handling. TCP50%: time to 50% of Ca2+ peak; TCD50%: time to 50% of Ca2+ decay; SMV: shortening maximum velocity; RMV: relaxation maximum velocity. Data are expressed as means ± SD or median (25 percentile; 75 percentile). Sham: animals submitted to simulated surgery (n = 6; number of cells = 29); Sham-ET: animals submitted to simulated surgery and exercise training (n = 6; number of cells = 36); AoS: animals submitted to aortic stenosis surgery (n = 6; number of cells = 41); AoS-ET: animals submitted to aortic stenosis surgery and exercise training (n = 6; number of cells = 46). Analysis of variance and Bonferroni post-hoc test or Kruskal-Wallis and Dunn’s method post-hoc. p < 0.05. * AoS vs. Sham; # AoS-ET vs. Sham-ET; § AoS-ET vs. AoS.
Figure 4.
Cardiomyocyte calcium handling. TCP50%: time to 50% of Ca2+ peak; TCD50%: time to 50% of Ca2+ decay; SMV: shortening maximum velocity; RMV: relaxation maximum velocity. Data are expressed as means ± SD or median (25 percentile; 75 percentile). Sham: animals submitted to simulated surgery (n = 6; number of cells = 29); Sham-ET: animals submitted to simulated surgery and exercise training (n = 6; number of cells = 36); AoS: animals submitted to aortic stenosis surgery (n = 6; number of cells = 41); AoS-ET: animals submitted to aortic stenosis surgery and exercise training (n = 6; number of cells = 46). Analysis of variance and Bonferroni post-hoc test or Kruskal-Wallis and Dunn’s method post-hoc. p < 0.05. * AoS vs. Sham; # AoS-ET vs. Sham-ET; § AoS-ET vs. AoS.

Figure 5.
Expression of calcium handling protein. Data are expressed as means ± SD. RyR: ryanodine receptor; SERCA2a: Sarcoplasmic Reticulum Calcium Pump; NCX: Sodium and calcium exchanger; PLB: Phosfolamban; PLBser16: Phosfolamban phosphorylated at serine 16; PLBthr17: Phosfolamban phosphorylated on threonine 17; CSQ: Calsequestrin; LTCC’s: L-type calcium channels. * p <0.05 AoS vs Sham; § p <0.05 AoS-ET vs AoS; # p <0.05 AoS-ET vs Sham-ET. Sham: animals submitted to simulated surgery (n = 7); AoS: animals submitted to aortic stenosis surgery (n = 7). Sham: animals submitted to simulated surgery (n = 7); Sham-ET: animals submitted to simulated surgery and exercise training (n = 7); AoS: animals submitted to aortic stenosis surgery (n = 7); AoS-ET: animals submitted to aortic stenosis surgery and exercise training (n = 7). Analysis of variance and Bonferroni post-hoc test. p < 0.05.
Figure 5.
Expression of calcium handling protein. Data are expressed as means ± SD. RyR: ryanodine receptor; SERCA2a: Sarcoplasmic Reticulum Calcium Pump; NCX: Sodium and calcium exchanger; PLB: Phosfolamban; PLBser16: Phosfolamban phosphorylated at serine 16; PLBthr17: Phosfolamban phosphorylated on threonine 17; CSQ: Calsequestrin; LTCC’s: L-type calcium channels. * p <0.05 AoS vs Sham; § p <0.05 AoS-ET vs AoS; # p <0.05 AoS-ET vs Sham-ET. Sham: animals submitted to simulated surgery (n = 7); AoS: animals submitted to aortic stenosis surgery (n = 7). Sham: animals submitted to simulated surgery (n = 7); Sham-ET: animals submitted to simulated surgery and exercise training (n = 7); AoS: animals submitted to aortic stenosis surgery (n = 7); AoS-ET: animals submitted to aortic stenosis surgery and exercise training (n = 7). Analysis of variance and Bonferroni post-hoc test. p < 0.05.
Figure 6.
Cardiorespiratory fitness. T1: first cardiorespiratory fitness test; T2: second cardiorespiratory fitness test, after four weeks of AET; T3: third cardiorespiratory fitness test, after seven weeks of AET; T4: fourth cardiorespiratory fitness test, after termination of the AET protocol. LT velocity: velocity at lactate threshold; LT intensity: intensity at lactate threshold; [LAC]basal: lactate concentration at baseline; [LAC]LT: lactate concentration at lactate threshold; [LAC](LT)/Vel(LT): ratio of lactate concentration at lactate threshold to lactate threshold velocity; [LAC](Ex)/Vel(Ex): ratio of lactate concentration at exhaustion to exhaustion velocity. * p < 0.05 AoS vs Sham; & p < 0.05 Sham vs Sham-ET; § p < 0.05 AoS-ET vs AoS; # p < 0.05 AoS-ET vs Sham-ET. a vs. T1; b vs. T1 and T2. Analysis of variance for repeated measures and Bonferroni post-hoc test. p < 0.05.
Figure 6.
Cardiorespiratory fitness. T1: first cardiorespiratory fitness test; T2: second cardiorespiratory fitness test, after four weeks of AET; T3: third cardiorespiratory fitness test, after seven weeks of AET; T4: fourth cardiorespiratory fitness test, after termination of the AET protocol. LT velocity: velocity at lactate threshold; LT intensity: intensity at lactate threshold; [LAC]basal: lactate concentration at baseline; [LAC]LT: lactate concentration at lactate threshold; [LAC](LT)/Vel(LT): ratio of lactate concentration at lactate threshold to lactate threshold velocity; [LAC](Ex)/Vel(Ex): ratio of lactate concentration at exhaustion to exhaustion velocity. * p < 0.05 AoS vs Sham; & p < 0.05 Sham vs Sham-ET; § p < 0.05 AoS-ET vs AoS; # p < 0.05 AoS-ET vs Sham-ET. a vs. T1; b vs. T1 and T2. Analysis of variance for repeated measures and Bonferroni post-hoc test. p < 0.05.
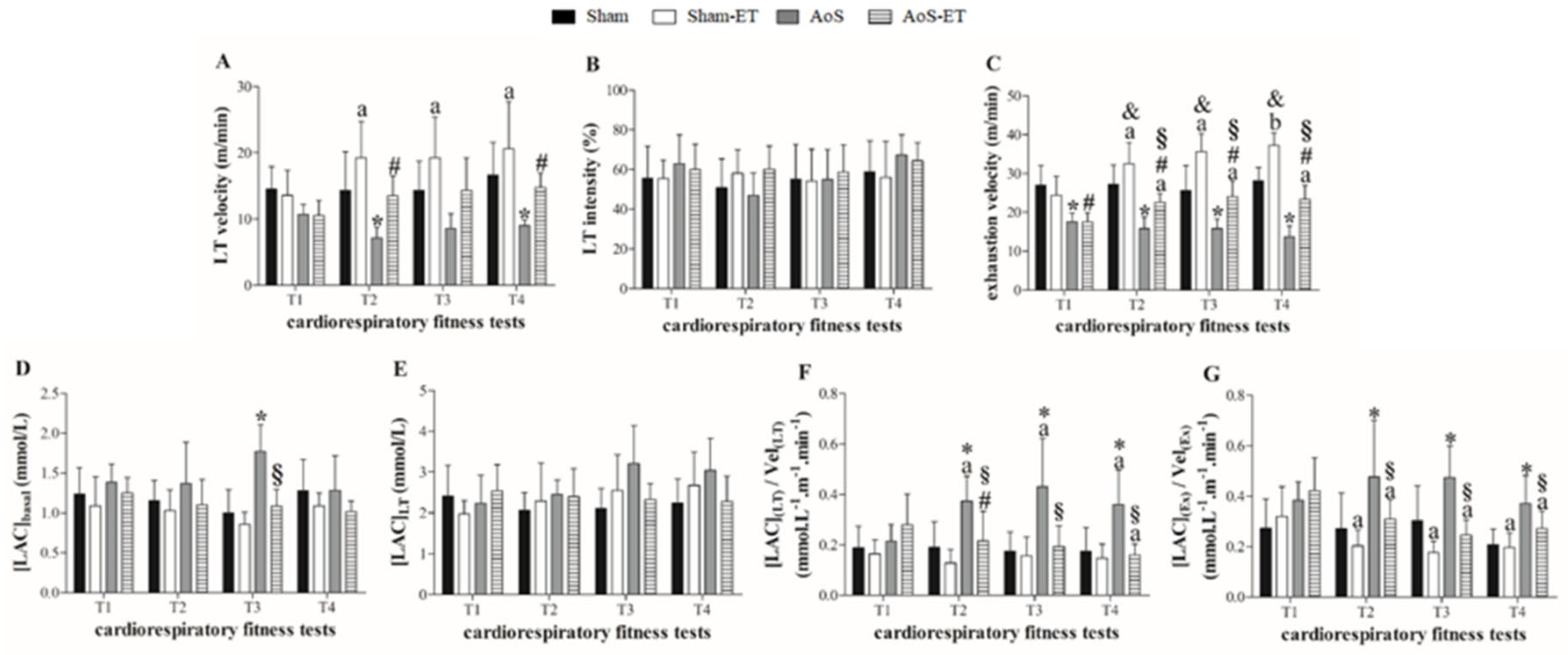

Table 1.
Echocardiogram data before and after exercise training.
| Before exercise training | After exercise training | |||||||
|---|---|---|---|---|---|---|---|---|
| Sham | Sham-ET | AoS | AoS-ET | Sham | Sham-ET | AoS | AoS-ET | |
| BW (g) | 469 ± 38 | 472 ± 55 | 450 ± 54 | 452 ± 59 | 519 ± 45 | 494 ± 60 | 494 ± 70 | 487 ± 59 |
| HR (bpm)& | 285 (280; 301) | 290 (269; 301) | 290 (262; 312) | 290 (280; 301) | 298 ± 42 | 292 ± 37 | 293 ± 43 | 305 ± 36 |
| LVDD (mm)& | 7.41 (6.90; 7.66) | 7.30 (6.90; 7.59) | 7.88 (7.21; 8.46) | 7.41 (6.32; 7.95) | 7.41 (7.15; 7.66) | 7.41 (7.09; 7.92) | 8.43 (7.27; 9.13) ** | 7.92 (7.34; 8.49) |
| LVSD (mm)& | 3.07 (2.81; 3.32) | 2.81 (2.55; 3.07) | 3.58 (3.24; 4.02) | 3.32 (2.74; 3.58) | 3.07 (2.81; 3.32) | 3.07 (2.81; 3.38) | 3.65 (3.32; 5.40) ** | 3.58 (2.81; 4.52) |
| PWDT (mm)& | 1.53 (1.53; 1.53) | 1.53 (1.53; 1.57) | 3.07 (2.81; 3.07) * | 2.92 (2.81; 3.07) # | 1.53 (1.53; 1.65) | 1.53 (1.53; 1.61) | 2.81 (2.55; 3.03) ** | 2.30 (2.55; 2.81) ## |
| ISDT (mm)& | 1.56 (1.53; 1.65) | 1.55 (1.53; 1.65) | 3.07 (2.98; 3.24) * | 3.07 (3.07; 3.32) # | 1.62 (1.53; 1.70) | 1.60 (1.53; 1.65) | 2.95 (2.81; 3.07) ** | 2.81 (2.55; 3.07) ## |
| RWT | 0.43 ± 0.03 | 0.43 ± 0.04 | 0.76 ± 0.13 * | 0.81 ± 0.11 # | 0.43 ± 0.03 | 0.42 ± 0.03 | 0.68 ± 0.14 ** | 0.68 ± 0.11 ## |
| LA/Ao& | 1.21 (1.19; 1.30) | 1.23 (1.19; 1.29) | 1.93 (1.67; 2.00) * | 1.87 (1.52; 2.11) # | 1.23 ± 0.09 | 1.24 ± 0.09 | 1.93 ± 0.19 ** | 1.79 ± 0.19 ##§ |
| E wave (cm/s) | 81 ± 7 | 83 ± 8 | 124 ± 24 * | 127 ± 22 # | 85 ± 7 | 82 ± 9 | 130 ± 20 ** | 120 ± 25 ##§ |
| E/A | 1.64 ± 0.22 | 1.64 ± 0.25 | 3.55 ± 1.67 * | 3.82 ± 2.00 # | 1.50 ± 0.20 | 1.54 ± 0.18 | 5.22 ± 1.42 ** | 3.82 ± 1.85 ##§ |
| E’ (cm/s)& | 5.60 (5.30; 5.80) | 5.60 (3.20; 6.10) | 5.40 (3.70; 6.25) | 5.40 (3.70; 6.12) | 6.13 ± 0.77 | 6.20 ± 0.73 | 5.19 ± 0.99 ** | 5.40 ± 1.18 ## |
| A’ (cm/s)& | 3.75 (3.70; 3.90) | 3.70 (3.35; 4.20) | 3.30 (3.03; 5.35) | 3.20 (3.20; 3.75) | 4.24 ± 0.72 | 4.10 ± 0.60 | 3.08 ± 1.20 ** | 3.95 ± 1.50 § |
| E/E’ | 14.3 ± 1.58 | 14.9 ± 1.98 | 25.9 ± 6.78 * | 26.3 ± 7.82 # | 14.1 ± 2.24 | 13.4 ± 1.6 | 25.6 ± 4.85 ** | 22.5 ± 3.66 ##§ |
| EDT (ms) | 48 ± 7 | 48 ± 5 | 34 ± 11 * | 36 ± 13 # | 49 ± 6 | 50 ± 4 | 28 ± 8 ** | 34 ± 11 ##§ |
| PWSV (cm/s) | 67 ± 9 | 71 ± 7 | 53 ± 12 * | 55 ± 8 # | 68 ± 9 | 70 ± 9 | 37 ± 10 ** | 49 ± 9 ##§ |
| FS (%) | 26 ± 3 | 28 ± 4 | 25 ± 4 | 25 ± 4 # | 26 ± 3 | 25 ± 4 | 23 ± 5 ** | 24 ± 4 |
| EF (%)& | 93 (91; 94) | 94 (92; 95) | 90 (87; 92) | 89 (79; 92) # | 93 ± 2 | 92 ± 3 | 86 ± 9 ** | 90 ± 4 § |
Data are expressed as means ± SD or median (25 percentile; 75 percentile)&. Sham: animals submitted to simulated surgery (n = 28); Sham-ET: animals submitted to simulated surgery and exercise training (n= 33); AoS: animals submitted to aortic stenosis surgery (n = 22); AoS-ET: animals submitted to aortic stenosis surgery and exercise training (n = 22). BW: body weight; HR: heart rate; LVDD: left ventricle diastolic diameter; LVSD: left ventricle systolic diameter; PWDT: posterior wall diastolic thickness; ISDT: interventricular septum diastolic thickness; RWT: left ventricle relative wall thickness; LA: left atrium; AO: aorta diameter; E/A: ratio between filling flow peak (E wave) and atrial contraction flow peak (A wave); EF: ejection fraction; FS: midwall fraction shortening; PWSV: posterior wall systolic velocity; E’: velocity of the mitral annulus during early ventricular filling; A’: mitral velocity annulus during atrial contraction; EDT: E-wave deceleration time; E/E’: ratio between filling flow peak and mitral annulus velocity during early ventricular filling. Analysis of variance and Bonferroni post-hoc test or Kruskal-Wallis and Dunn’s method post-hoc. Before exercise training: * p < 0.05 vs. Sham, # p < 0.05 vs. Sham-ET. After exercise training: ** p < 0.05 vs. Sham, ## p < 0.05 vs. Sham-ET, § p < 0.05 vs. AoS.
Table 2.
Baseline data.
| Sham | Sham-ET | AoS | AoS-ET | |
|---|---|---|---|---|
| CSA (mm2) | 1.14 ± 0.13 | 1.15 ± 0.14 | 1.16 ± 0.16 | 1.20 ± 0.19 |
| DT (g/mm2) | 6.23 ± 1.43 | 6.12 ± 1.61 | 5.20 ± 1.11 | 5.23 ± 1.96 |
| RT (g/mm2) | 0.57 ± 0.19 | 0.61 ± 0.18 | 0.82 ± 0.22 * | 0.66 ± 0.17 § |
| +dT/dt (g/mm2/s) | 66.6 ± 17.2 | 65.9 ± 18.1 | 46.9 ± 10.3 * | 47.9 ± 20.0 # |
| −dT/dt (g/mm2/s) | 21.8 ± 5.50 | 22.1 ± 5.39 | 24.2 ± 5.57 | 23.0 ± 7.61 |
| TPT (ms) # | 180 (180; 200) | 180 (160; 180) | 210 (182; 220) | 200 (180; 210) # |
Data are expressed as means ± SD or median (25 percentile; 75 percentile)&. Sham: animals submitted to simulated surgery (n = 22); Sham-ET: animals submitted to simulated surgery and exercise training (n = 20); AoS: animals submitted to aortic stenosis surgery (n = 16); AoS-ET: animals submitted to aortic stenosis surgery and exercise training (n = 16). CSA: papillary cross-sectional area; DT: maximum developed tension; RT: resting tension; +dT/dt: maximum rate of tension development; -dT/dt: maximum rate of tension decline; TPT: time-to-peak tension. Analysis of variance and Bonferroni post-hoc test or Kruskal-Wallis and Dunn’s method post-hoc. * p < 0.05 AoS vs. Sham; # p < 0.05 AoS-ET vs. Sham-ET; § p < 0.05 AoS-ET vs. AoS.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



