
Submitted:
05 June 2023
Posted:
06 June 2023
You are already at the latest version
Abstract
This review examines the evidence supporting the role of dysbiosis in the development of obses-sive-compulsive disorders (OCD). We review the molecular mechanisms and role of the microbiota in the microbiota-gut-brain axis, focusing on the endocrine, nervous, and immune pathways. We then propose a model that positions dysbiosis as the central unifying element in the neurochemi-cal, immunological, genetic, and environmental factors leading to OCD. Based on this, we review the animal and human clinical evidence for the use of microbial reprogramming strategies such as probiotic or fecal microbiota transplants to treat OCD. Finally, we discuss the unique challenges that must be addressed in future clinical interventions.
Keywords:
OCd
; obsessive-compulsive disorder
; microbiota
; gut
; gut-brain axis
; probiotics
; fecal transplants
; microbial reprogramming
1. Introduction
Obsessive-Compulsive Disorder (OCD) is a chronic mental health disorder characterized by the presence of the intrusive and persistent thoughts that cause distress called obsessions; these are followed by compulsions, which are repetitive behaviors or mental acts that individuals feel driven to perform to calm their obsessions [1]. OCD affects approximately 2-3% of the global population, and greatly interferes with quality of life, disturbing the daily functioning of an individual, from eating to bathing to walking or even breathing. For example, an individual with OCD can spend thirty minutes closing a door and verifying it is closed, with the hope that the anxiety might be calmed after a certain number of repetitions. Decision-making of individuals with OCD is greatly affected, as every decision may be felt as a threat leading to maximum indecisiveness [2,3,4]. It is a very debilitating disorder that often hides behind another disease or disorder. Indeed, although OCD is now recognized as an independent disorder category, it often occurs with another disorder such as autism, attention deficit hyperactivity disorder (ADHD), depression, general anxiety disorder, eating disorder, hoarding disorder, Tourette syndrome, panic disorder, or schizophrenia [5]. This category includes other disorders such as hoarding disorder, hair-pulling disorder, and skin-picking disorder [6].
The exact mechanisms underlying OCD are not yet fully understood. Research has highlighted several associations, leading to the conclusion that a combination of genetic, neurobiological, immunological, and environmental factors may contribute to its development. Indeed, studies have identified heritability of OCD through multiple genes such as the serotonin transporter gene (SLC6A4) and the gene encoding the dopamine D2 receptor (DRD2) [7,8]. Also, OCD has been associated with neurobiological changes such as the dysregulation of the cortico-striato-thalamo-cortical (CSTC) circuit. Brain regions involved include the orbitofrontal cortex, anterior cingulate cortex, and basal ganglia, as well as the dysregulation in neurotransmitters like serotonin, dopamine, and glutamate [6,9,10,11]. In addition, environmental factors, such as childhood trauma including physical and/or sexual abuse, have been associated with an increased risk of developing OCD. Finally, stressful life events, such as significant life changes or trauma, have been found to precede the onset or exacerbation of OCD symptoms [6,12].
To date, cognitive-behavioral therapy (CBT) and pharmacotherapy are the primary treatments for OCD [13,14]. CBT typically involves exposure and response prevention, where individuals are gradually encouraged to face their obsessions while refraining from engaging in their compulsive behaviors. This helps to reduce the anxiety associated with the obsessions and weaken the link between the obsession and compulsion. CBT has been shown to be effective in reducing OCD symptoms and improving overall functioning [14]. On the other hand, selective serotonin re-uptake inhibitors (SSRIs), such as fluoxetine, sertraline, and fluvoxamine, are the first-line medications for OCD treatment [13,15]. These medications increase serotonin levels in the brain and help to alleviate symptoms. Additionally, combining SSRIs with antipsychotics or glutamate modulators is sometimes used for individuals who do not respond adequately to SSRIs alone [16]. Despite the availability of treatment options for OCD, there are significant limitations that warrant the exploration of novel therapeutic approaches. While these interventions can be effective for some individuals, many patients experience only partial response to treatment, lingering symptoms, and high rates of relapse [16,17]. CBT is efficient but each treatment plan is specific to an obsession and does not avoid the appearance of another obsession and compulsion later, which would require another course of CBT. Additionally, there are side effects associated with the use of medication, such as gastrointestinal disturbances and sexual dysfunction, which can further impact treatment adherence and quality of life [18].
The limitations of current treatment options emphasize the need for innovative and therapeutic approaches that target the etiology of OCD. To date, several factors have been proposed to contribute to the development of OCD, and it is difficult to point at one single cause. Nevertheless, there is one emerging avenue of investigation that presents itself as promising and key for the understanding and treatment of OCD: the gut microbiota. The gut microbiota comprises trillions of microorganisms residing in the gastrointestinal tract, from bacteria to fungi to viruses, archea or protozoa. These microbes actually outnumber human cells by a factor of 10, and the genes they express form the microbiome [19]. These are usually classified into three categories according to their interaction with their human hosts: beneficial, pathogens and commensal microbes. Because they control each other’s growth, eubiosis (the undefined but balanced composition of the gut microbiota) is essential to prevent the overgrowth of pathogens or lack of growth of certain beneficial microbes from lacking. In contrast, dysbiosis refers to an imbalance in the composition or function of the gut microbiome. It can occur when there are changes in the relative abundance of certain microbial species or alterations in the overall diversity, resulting in alteration of the metabolites produced by the microbiota. Dysbiosis has been associated with various health conditions, including metabolic disorders, mental health disorders, autoimmune diseases and inflammatory bowel diseases [20,21,22,23,24,25,26,27,28,29,30,31,32,33]. Interestingly, dysbiosis has been associated with all disorders where OCD has been found as a comorbidity such as Autism, Tourette Syndrome, Anxiety Disorders, Panic Disorder, Eating Disorders, Depression and hording disorder but also gastrointestinal diseases such as ulcerative colitis or Crohn’s disease[34,35,36].
While we acknowledge that no study has pointed at the prevalence of the co-occurrence of dysbiosis and OCD in anxiety disorders, latest advances in the understanding of the bidirectional communication between the gut and the brain strongly implicate the gut microbiome as a key component for future investigations. In this review, we examine the growing evidence that supports the possible causal role of dysbiosis in these anxiety disorders. We also discuss the emerging clinical studies that aim to modulate the gut microbial composition to increase its diversity and inhibit the growth of pathogens.
We will first review the molecular mechanisms involved in the microbiota-gut-brain axis (MGBA). Then, we will gather the latest evidence that supports our rationale; the latest evidence that shows dysbiosis in OCD and how dysbiosis fits into a model explaining the neurochemical, the genetic, and the immunological and the environmental basis of OCD. Finally, we review recent clinical interventions that support the promising potential of two microbial reprogramming strategies: dietary interventions using prebiotics and probiotics and fecal microbiota transplantation (FMT). We will discuss the challenges of studying such clinical interventions in OCD and identify important considerations for future clinical studies.
2. Mechanisms of the Microbiota-Gut-Brain Axis (MGBA)
The MGBA refers to the bidirectional communication between the gut microbiota, the gastrointestinal tract, and the central nervous system (CNS). From brain to gut, endocrine systems such as the hypothalamic-pituitary-adrenal and hypothalamic-pituitary-gonadal axes regulate the gut microbiota [37,38,39,40]. From gut to brain, the gut microbiome, consisting of the microbes, their genomes and their products, can influence brain function through a variety of mechanisms, summarized in Figure 1. We will describe these below as: 1. the endocrine pathway; 2. the nervous pathway; and 3. the immune pathway.
2.1. The Endocrine Pathway
The endocrine pathway consists of the release into the systemic circulation of microbiota-derived products issued from the metabolic activity of microbes including short chain fatty acids (SCFAs) and neurotransmitters, hormones and inflammatory factors that directly or indirectly regulate the function of central nervous system (CNS).
The production of short chain fatty acids (SCFAs) by gut bacteria, after anaerobic fermentation of indigestible polysaccharides such as dietary fibers and resistant starch, play a crucial role in modulating the metabolic activity of the gut and are pivotal to microbiota-gut-brain crosstalk [41,42,43]. A variety of SCFAs are produced, depending on the nature of the dietary fibers being digested: the most abundant are butyrate, acetate, and propionate [44,45].
Following their production, SCFAs can cross the enterocyte layer and be absorbed by colonocytes. This happens mainly via H+-dependent or Na+-dependent monocarboxylate trans-porters [46]. They can regulate gut barrier integrity and mucosal immunity through various molecular mechanisms involving G protein-coupled receptors such as free fatty acid receptors 2 and 3 or hydrocarboxylic acid receptors [47]. Butyrate has been shown to promote the upregulation of proteins constituting tight junctions such as zonula occludens-1, claudin-5, and occulin, and also promote the inhibition of zonulin to reduce intestinal permeability and maintain the gut barrier integrity [48,49,50,51,52,53,54]. The decrease in abundance of butyrate can lead to a leaky gut syndrome, thereby influencing the immune response, as well as the integrity of both the gut and the blood-brain barrier (BBB) [55,56,57,58,59]. Indeed, expression of claudin and occludin have also been shown to be reduced in the BBB of germ-free mice, leading to increased permeability of the BBB from intrauterine life to adulthood [60]. Brain uptake of SCFAs has previously been shown in rats [61], and studies have shown detectable levels of acetate, propionate and butyrate in the cerebrospinal fluid [62]. In another study, brains of mice supplemented with live Clostridium butyricum had significantly higher concentrations of butyrate than did peripheral blood [63,64]. The recolonization of these adult mice with a complex microbiota, or monocolonization with SCFA-producing bacterial strains, recovered the integrity of the BBB [58,60]. Similarly, treatment of an in vitro model of cerebrovascular endothelial cells with propionate attenuated the permeabilizing effects of exposure to lipopolysaccharide (LPS) [65].
Sometimes, protein fermentation in the distal portion of the intestine can lead to the production of potentially toxic metabolites, such as ammonia, phenols, and sulfides, as well as unique branched-chain fatty acids [66,67,68,69]. By controlling the BBB integrity, SCFAs play a pivotal role in the passage of these and other molecules and nutrients from the circulation to the brain, playing a central role in brain development and the preservation of CNS homeostasis [59,70,71,72].
2.2. The Nervous Pathway
Various gut bacteria have been shown to also produce neurotransmitter precursors and hormones, such as dopamine, acetylcholine, γ-aminobutyric acid, noradrenaline, serotonin, corticotrophin-releasing hormone [73]. In addition to producing peripheral serotonin, gut microbes can affect transmission of central serotonin by modulating the production of tryptophan in the plasma. This has been demonstrated for Bifidobacterium infantis [74]. The enterochromaffin cells can bind several microbial products and secrete serotonin into the lamina propria, increasing colonic and blood concentrations of 5-HT [75,76]. SCFAs regulate the expression levels of tryptophan 5-hydroxylase 1, the enzyme involved in synthesis of serotonin, and tyrosine hydroxylase, which is involved in a rate-limiting step in the biosynthesis of dopamine, noradrenaline and adrenaline. SCFAs thereby exert an effect on brain neurochemistry [75,76,77,78,79,80,81]. The neural pathway involves the bidirectional communication between the gut and the brain via the autonomic nervous system or the vagus nerve[82,83,84,85]. The vagus nerve serves as a major conduit for transmitting signals between the gut and the CNS. When gut bacteria modulate the production of neurotransmitters such as GABA, 5-HT, DA, NE, Glu, and ACh, these can bind to primary afferents of the enteric nervous system and the vagus nerve to influence brain function and behavior. Retrograde transport also plays an important role in gut to brain communication. It is now recognized that the onset of Parkinson’s disease probably starts in the gut, with α-synuclein aggregation upon LPS binding, and retrograde transport of aggregates through the vagus nerve to the brain. There is accumulating evidence that SCFAs may also modulate key neuro-pathological processes underlying Alzheimer’s disease by interfering with the assembly of amyloid-β peptides into the neurotoxic oligomers[86,87,88]. In addition, the metabolites of gut microbes, by controlling the secretion of gut hormones such as glucagon-like peptide 1 and peptide YY, can influence food intake, which will in return influence bacterial fermentation, thereby reinforcing the close relationship between diet and gut microbiome [89].
2.3. The Immune Pathway
2.3.1. From Gut to Host Immune System
Changes in the gut microbiota composition can affect the production and availability of SCFAs, thus impacting the metabolic activity of the gut [90,91,92,93,94]. Dysbiosis, by leading to reduced SCFA production, impaired gut barrier function, and increased intestinal permeability, is a starting point for systemic inflammation and potential neuroinflammation [95,96]. Byproducts of microbiota metabolism can activate immune cells in the gut, leading to the production of pro-inflammatory or anti-inflammatory cytokines [97,98,99,100,101]. These immune signals can then communicate with the brain and affect neural function, as neuroinflammation is an important process shaping brain function.
A good example of this are bacterial lipopolysaccharides (LPS) also known as endotoxins, which are components of the outer membrane of gram-negative bacteria. They are typically produced as part of the bacterial growth and replication process[102]. These endotoxins trigger a pro-inflammatory cascade in the mucosa, mediated by toll-like receptor 4 and cytokines such as tumor necrosis factor α (TNF-α) and interleukin 6 (IL-6). The LPS-induced pro-inflammatory cascades have been shown to be inhibited by butyrate inhibition of histone deacetylase (HDAC) [103]. This intracellular signaling has been found not only in the gut and associated immune tissue, but also in the peripheral nervous system and CNS [53,104,105,106,107]. Perturbations of the gut microbiota by antibiotics in experimental animal models systemically produced altered immune responses with pro-inflammatory profiles [108]. In early life, if the microbiota is depleted using antibiotics, this results in an inflammatory response in the CNS with pro-inflammatory cytokine secretion and altered microglial morphology, which could be reversed by butyrate treatment [109,110,111,112,113,114,115]. Indeed, butyrate has been shown to control the maturation of mucosa-associated lymphoid tissue, and the differentiation of lymphocytes, characterized by the presence of macrophages, B and T cells. Similarly, acetate treatment of microglia primary culture in vitro has been shown to reduce inflammatory signaling by downregulating the expression of IL-1β, IL-6, and TNF-α and the phosphorylation of p38 MAPK, JNK, and NF-κB [116]. The precise signaling involved in the effects of SCFAs on microglia remain unclear, and the histone acetylation or the epigenetically regulated gene expression is considered the main mechanism [117].
2.3.2. From Host Immune System to Gut Microbiota
Several studies investigating the relationship between immunoglobulin A (IgA) and the gut microbiome have concluded that an adequate production of IgA is essential for the colonization of certain “good” bacteria such as Bifidobacterium and Bacteroides [118,119,120,121,122,123,124,125,126] On the other hand, microbial acetate produced by gut microbes is also able to regulate IgA reactivity to commensal bacteria, thus highlighting a bidirectional relationship between gut microbes and the immune system [72,127,128,129,130].
IL-22 and IL-17 have been shown to stimulate gut intraepithelial cells to produce antimicrobial peptides such as α-defensins and β-defensin 1, which can quickly inactivate microorganisms entering the host through a leaky gut [131,132]. Furthermore, mice transgenic for defensins, exhibited an altered microbiota composition [133,134,135,136]. Thus, the antimicrobial peptides affect the microbial composition.
3. Dysbiosis and the Neurobiology Basis of OCD
3.1. Dysbiosis in OCD
The potential role of the gut-brain axis in the pathophysiology of OCD has been highlighted by several studies and suggests that alterations in gut microbiota composition may impact brain function and behaviour, including obsessive-compulsive symptoms. Studies have found significant differences in the relative abundance of certain bacterial taxa between individuals with OCD as compared to healthy individuals, suggesting a potential association between gut microbiota dysbiosis and OCD[137,138,139,140,141,142,143]. It is important to note that these studies have limitations, including small sample sizes and the presence of confounding factors. Overall, while there is emerging evidence suggesting a correlation between gut microbiota dysbiosis and OCD, more research is needed to clearly establish a causal relationship and determine the clinical implications. Although clinical interventions targeting dysbiosis and focusing specifically on OCD are scarce, a few studies are worth mentioning. A recent scientific report by Domenech et al. (2022) reported dysbiosis in the gut and oropharyngeal microbiomes of OCD patients [140]. They noted an increase of bacteria from the Rikenellaceae family, associated with gut inflammation, and a decrease of bacteria from the Coprococcus genus. A lower bacterial diversity in the gut of OCD patients has been observed, consistent with the lower gut α-diversity in PANS/PANDAS patients reported by Quagliariello, et al. [144], and in OCD by Turna, et al. [143]. The latter observed a decrease in species richness/eveness and a lower relative abundance of three butyrate producing genera (Oscillospira, Odoribacter and Anaerostipes) in OCD patients [143]. Furtrhermore, lower α-diversity has also been reported in subjects with ADHD [145,146,147], and in studies of ASD individuals [28,29,148,149,150,151].
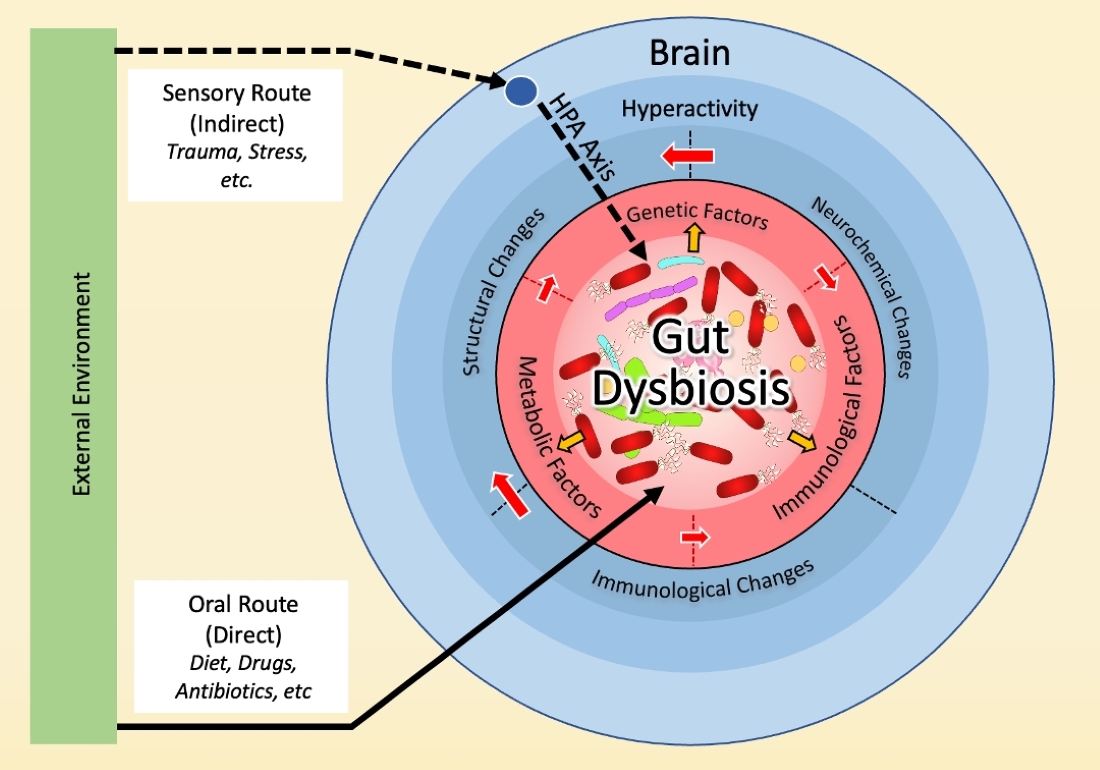
We propose a model where gut dysbiosis is at the center of all factors previously associated with OCD and may be implicated in the neurochemical, immune, genetic, and environmental bases of OCD (Figure 2). Below, we will detail evidence in support of this model.
3.2. Dysbiosis and Hyperactivity in the Cortico-Striato-Thalamo-Cortical Circuit (CSTC)
The CSTC projects from the orbito-frontal-cortical region (OFC) to the striatum, and then onward to thalamic sites, before looping back to the cortex. It is responsible for reward- and motivational-related processes, executive function, motor and response inhibition, and habit-based behaviour [152]. Two pathways within this circuit, direct and indirect, should respectively have opposing net effects on the thalamus, and this balance is critical for initiation and suppression of behaviour [152]. Any imbalance is thought to contribute to OCD pathology. Indeed, an overactivity in the direct pathway results in a hyperactivity in the feedback loop creating an overall hyperactivity within the circuit. Several studies have noted an increased activation of the OFC and striatum and caudate regions [6,152,153,154]. This hyperactivity is believed to involve the neurotransmitters serotonin, glutamate and dopamine [153,155,156].
3.2.1. Serotonin
Adams et al. (2005) pointed at an increased binding of 5-HT2AR, the receptor to serotonin, in the caudate nuclei of unmedicated OCD patients, possibly due to the compensatory effects of low levels of serotonin within the CSTC circuit [157]. Simpson et al. (2021) showed that an earlier onset of OCD was associated with increased 5-HT2AR availability in the circuit [158]. This is supported by the successful treatment using serotonin receptor inhibitors (SRI). Nevertheless, mechanisms whereby SRIs ameliorate symptoms are still not well understood, and only 40–60% of patients improve following SRI intervention [159,160].
The gut microbiota plays a crucial role in the production of serotonin. Specific gut bacteria, such as Lactobacillus and Bifidobacterium species, have been found to produce serotonin in the gut [161]. Changes in the composition of the gut microbiota can disrupt this serotonin production process, leading to imbalances in serotonin levels. Certainly, several gut bacteria (such as Clostridium, Burkholderia, Streptomyces, Pseudomonas, and Bacillus) play a role in the metabolism of tryptophan [155,162,165]. Although in normal conditions peripheral serotonin cannot freely cross the BBB, its precursor, tryptophan, can cross the BBB through specialized transport mechanisms, to then be converted into serotonin by local neurons. Furthermore, dysbiosis can both influence the availability of tryptophan and disrupt the BBB allowing other molecules inside the brain. Certain gut bacteria can metabolize tryptophan, affecting its availability for serotonin production [155,162,163,164,165]. Changes in gut microbial composition can alter tryptophan metabolism, potentially impacting serotonin levels in the brain. As a result, dysbiosis-induced alterations in the gut microbiota could result in reduced serotonin synthesis in the brain, potentially contributing to mood disorders and behavioural changes.
Gut dysbiosis can also influence the serotoninergic system in the brain through other mechanisms. For instance, the gut microbiota can affect serotonin signaling by influencing the expression and activity of serotonin receptors in the brain. Dysbiosis-related changes in the gut microbial composition have been associated with alterations in serotonin receptor expression and function. A study by Hsiao et al. (2013) [166] explored the impact of gut dysbiosis on serotonin signaling. They investigated mice with gut microbiota imbalances and observed abnormal serotonin receptor expression patterns in specific brain regions. These changes were associated with behavioral abnormalities, including altered social interactions and increased anxiety-like behavior. This suggests that dysbiosis-induced disruptions in the gut microbiota can influence serotonin receptor function, potentially contributing to mood disorders and behavioral dysregulation.
3.2.2. Glutamate
The glutamate system is the major excitatory neurotransmitter system in the brain. Studies have shown that untreated OCD patients have elevated glutamate concentrations in the caudate region as compared to healthy individuals; these normalized after 12 weeks of SRI treatment, suggesting that availability of serotonin at the frontal region of the circuit might modulate the concentration of glutamate in the caudate part [167]. Because there are important glutamatergic projections between the frontal cortical part to the striatum, it was proposed that the SRI treatment allowed the increase of serotonin levels that in turn inhibited the glutamate levels in the caudate. In contrast, with low levels of available serotonin, the inhibitory effects within the circuit are reduced, which would allow for an elevated glutamate activity in the circuit [167,168]. Increased glutamate concentrations have also been observed in the cerebrospinal fluid (CSF) of untreated OCD patients [169]. Unsurprisingly, the modulation of glutamate via n-acetyl cysteine showed improvements in double-blind placebo-controlled studies for the obsessive compulsive-related disorders trichotillomania and skin-picking disorder [170,171].
The gut microbiota has been shown to influence the glutamate system through two main mechanisms. Firstly, certain gut bacteria, such as Lactobacillus and Bifidobacterium species, are capable of producing and metabolizing glutamate, thereby influencing its levels in the body. Dysbiosis-induced changes in the composition of the gut microbiota can lead to alterations in glutamate production and metabolism, potentially impacting glutamate signaling in the brain. A study by Strandwitz et al. (2019) demonstrated the significance of gut bacteria in glutamate metabolism [172,173]. They identified specific bacterial enzymes involved in glutamate production and observed that germ-free mice lacking these bacteria had lower levels of brain glutamate compared to control mice, indicating the influence of specific gut bacteria on brain glutamate levels.
Secondly, dysbiosis can affect the expression and function of glutamate receptors and transporters in the brain. Changes in the gut microbiota can lead to modifications in the expression of glutamate receptors, such as N-methyl-D-aspartate (NMDA) receptors, and glutamate transporters, such as EAAT3. In their investigation of the impact of gut dysbiosis on glutamate-related pathways, Sharon et al. (2019) found that mice with disrupted gut microbiota exhibited altered expression of NMDA receptors and EAAT3 transporters in the brain, along with neurobehavioral abnormalities [115]. These findings suggest that dysbiosis-induced changes in the gut microbiota can influence the function of glutamate receptors and transporters, potentially influencing glutamate neurotransmission in the brain.
3.2.3. Dopamine
Imaging studies reveal increased dopamine levels in the basal ganglia of OCD patients as well as enhanced binding to the dopamine transporter [174] Another study found increased density of the dopamine transporter in the left caudate and left putamen of untreated OCD patients [175]. The antipsychotic drugs that are sometimes offered to OCD patients who resist SRI treatments block subcortical dopamine receptor activity and are proposed to target the habit system and compulsive behaviors.
The gut microbiota can also influence the dopamine system. Some gut bacteria, including certain strains of Enterococcus and Lactobacillus, are capable of producing and metabolizing dopamine. Dysbiosis-induced changes in the gut microbial composition can impact dopamine production and metabolism. The influence of dysbiosis on the expression and function of dopamine receptors and transporters in the brain has been an area of growing research interest. Several studies have demonstrated that changes in the gut microbiota composition can indeed lead to alterations in dopamine receptor expression and dopamine transporter activity. One study conducted by Bercik et al. (2011) explored the effects of the gut microbiota on central levels of brain-derived neurotrophic factor (BDNF) and behavior in mice [176]. They found that germ-free mice, lacking gut microbiota, displayed altered dopamine receptor expression in specific brain regions compared to control mice with normal gut microbiota. In another study, Dinan et al. (2013) investigated the role of psychobiotics, a novel class of bacteria with potential mental health benefits, on neurotransmitter systems including dopamine [177]. They found that certain psychobiotics, such as Lactobacillus and Bifidobacterium strains, had the ability to modulate dopamine receptor expression and dopamine transporter activity, highlighting the impact of specific gut bacteria on the dopaminergic system [178,179,180].
3.3. Dysbiosis and the Immune Basis of OCD
Inflammation and immune dysregulation have been implicated in the pathogenesis of OCD. Preliminary studies have noted abnormal concentrations of IgA in children with OCD. IgA mediates microbial composition and homeostasis at the mucosal level, via prevention and promotion of bacterial growth, which influence bacterial gene expression [120].
There is also evidence for the role of immunological processes in the pathophysiology of OCD [9,142,181,182,183]. Commonly known as the “autoimmune OCD subtype”, the Pediatric Autoimmune Neuropsychiatric Disorders Associated with Streptococcal Infections or PANDAS is characterized by OCD that appears after a streptococcal infection, such as scarlet fever or strep throat [181]. The dramatic surge of symptoms which happens overnight includes motor obsessions and compulsions. In addition, children experience mood issues and anxiety attacks. In this context, the streptococcus A infection is clearly a manifestation of dysbiosis, which activates these immunological processes. Through molecular mimicry, the streptococcal bacteria are believed to induce an autoimmune response that attacks the brain via neuroinflammation. Indeed, several studies have found cross-reactive antibodies targeting the brain in children with PANDAS, as well as microglial activation [9,181,184,185,186].
3.4. Dysbiosis and the Genetic Basis of OCD
The neurobiological basis of overactivity involving serotonin, glutamate and dopamine is thought to be mainly rooted in genetic factors on one hand, with a heritability of 27–65%, and environmental factors on the other [187]. Twin studies have shown that OCD can run in families [188,189]. Epigenetic alterations have been suggested to be particularly relevant in OCD. Investigations into the peripheral DNA methylation signatures of OCD are scarce but DNA methylation patterns have been described [190].
Among the genes under investigation, notable examples include serotonergic genes (HTR2A, 5HTTLPR, SLC6A4), glutamatergic genes (SLC1A1, DLGAP3, SAPAP3), and dopaminergic genes (SLC6A3, DRD4)[10,11,189,191,192,193]. Recently, there has been a growing focus on the role of epigenetic mechanisms such as gene methylation, histone deacetylation (HDAC), and histone acetylation (HAT) in psychiatric disorders. Efforts have been made to develop treatment strategies targeting these mechanisms. Studies comparing DNA methylation patterns in OCD [194] patients and control groups have revealed distinct methylation profiles in the promoter regions of genes such as MAOA, GABA, MOG, BDNF, LEPR, OXTR, SLC6A4, and SLC6A3. In a recent study, a statistically significant correlation was observed between certain obsessions/compulsions and polymorphisms in HDAC2, HDAC3, and HDAC4 in the OCD group (p < 0.05) [195]. Indeed, gut microbes have been found to inhibit the histone desacetylase, an enzyme that facilitates transcription of genes by removing the acetyl group from histone proteins on DNA [196].
Other genes have been found to be associated to OCD in some studies, such as the oxytocin receptor (OXTR) gene [190,197,198,199,200] , the monoamine oxidase A (MAOA) gene [201], the brain-derived neurotrophic factor (BDNF) gene [202], the gamma-aminobutyric acid B receptor 1 (GABBR1), the estrogen receptor 1 (ESR1), myelin oligodendrocyte glycoprotein (MOG) genes and again the brain-derived neurotrophic factor (BDNF) gene [203].
The growing body of evidence implicating gut microbes in epigenetics places dysbiosis at the center of our model. However, not all clinical observations can be explained by the genetic hypothesis alone. For example, several studies failed to find significant associations between the dopamine transporter gene (SLC6A3) and OCD [204,205,206,207], the Dopamine receptor 2 (DRD2), or the dopamine receptor 3 (DRD3) [206,208,209]. This suggests that other mechanisms are involved in the observed increase of dopamine in the CSTC circuit.
3.5. Dysbiosis and the Environmental Basis of OCD
In this proposed model, childhood trauma and stressful life events can modulate the gut microbiota composition and thus trigger dysbiosis. This has been proposed to happen via the hypothalamo-pituitary axis [37,38,39,210,211]. Lifestyle changes, particularly diet, as well as the recurrent use of antibiotics at early stages of life can be determinants in the early development of the gut microbiota and the development of neurodevelopmental disorders [211,212,213,214]. Indeed, gut microbiota and brain maturation, including myelination, occur synchronically between birth and three years of age, and the gut microbiota has been found to play a critical role in myelination [215,216]. The gut microbiota includes bacteria that can synthesize various neurotransmitters besides serotonin. For instance, Lactobacillus and Bifidobacterium produce GABA, Escherichia, Bacillus, and Saccharomyces spp generate norepinephrine, Bacillus synthesizes dopamine, and Lactobacillus produces acetylcholine [217,218,219,220]. Thus, disruption of the microbiota in early years can impact the normal functioning of the gastrointestinal tract and affect the overall health of the individual and is likely to elevate the occurrence of diverse mental disorders [27,148,221,222].
4. Microbial Reprogramming Strategies
4.1. Prebiotics, Probiotics and Postbiotics
If dysbiosis is a central element in the development of OCD, then it would be expected that manipulation of the gut microbiota might influence its occurrence, and offer potential options for its treatment. This does in fact appear to be the case, and in this section, we gather clinical evidence of the use of probiotics and fecal microbiota transplants in the treatment of OCD.
A probiotic is a live organism that, when ingested in adequate amounts, exerts a health benefit for the host [223]. Probiotics use dietary fibers or resistant starch as nutrient sources (or prebiotics) to produce beneficial metabolites (postbiotics). The term synbiotic is used to refer to the mixture of both prebiotics and probiotics [224]. Dietary fibers and resistant starch therefore play an essential role in fermentation and postbiotic production [225,226,227]. Westernized diets are characterized by a relatively low intake in dietary fiber, which could explain the presence of dysbiosis in most modern diseases and disorders. Dietary fibers also include plant-based carbohydrates, such as polyphenols, and non-carbohydrate compounds such as lignin. Probiotics such as Lactobacillus and Bifidobacterium or Akkermansia can use these compounds to produce SCFA, which in turn promote various beneficial effects for the host [228,229,230,231,232,233,234].
Over the last decade, a number of studies have shown promising results for the use of probiotics in the treatment of OCD. However, while a growing number of studies have investigated the potential value of probiotics in treating Autism or ADHD, investigations of probiotic interventions for OCD are still at their very early stages, with most studies being performed in animal models.
Kantak et al. (2014) found that two-week pretreatment with Lactobacillus rhamnosus GG had the ability to reduce obsessive-compulsive disorder (OCD) in mice. Results were comparable to treatment with fluoxetine [236].
In 2018, Tabouy et al. [244], using Shank3 KO mice (a model used to study neurodevelopmental disorders such as autism), found Lactobacillus reuteri to be in a decreased relative abundance in the Shank3 KO. The treatment of Shank3 KO mice with Lactobacillus reuteri induced a significant decrease in repetitive behaviors in both male and female Shank3 KO mice [244].
In a study conducted by Szklany et al. (2020), male mice receiving – from the day of birth onwards - a prebiotic mixture composed of short-chain galactooligosaccharides (scGOS) and long-chain fructo-oligosaccharide (lcFOS), exhibited changes in the serotonergic system [243]. These neurological modulations were associated with behavioral changes such as a reduction in anxiety and repetitive behavior during development and increased social interest in adulthood compared to mice fed a control diet. Brains of the treated group exhibited altered mRNA expression of astrocytic glial fibrillary acidic protein and microglial integrin alpha M. There was also enhanced mRNA expression of BDNF in the prefrontal cortex. Additionally, analysis of the cecal content of treated animals revealed relatively increased levels of SCFA such as butyric acid, and decreased levels of valeric, isobutyric, and isovaleric acid [243].
Another animal study conducted by Sanikhani et al. (2020) demonstrated the effectiveness of Lactobacillus casei Shirota in treating OCD in a rat model. After daily administration of L. casei Shirota (10^9 CFU/mL for four weeks), the probiotic showed beneficial effects, possibly effected through the modulation of genes related to serotonin. Following concurrent treatment with L. casei Shirota and fluoxetine, the expression level of Bdnf significantly increased, while the expression of Htr2a (serotonin receptor 2A) decreased in the orbitofrontal cortex tissues of all rats involved in the study [237].
In 2021, Sunand et al. [242], found that selected probiotic strains and complex treatment with probiotic significantly ameliorated microbial diversity, repetitive behaviour, and the concentrations of NF-a, BDNF, and 5-HT.
Alghamdi et al. in 2022 [235], using an animal model of Autism induced by propionic acid, found that subjects with cognitive dysfunction had altered levels of neurotransmitters in their brains. However, in the group of animals treated with probiotic, neurotransmitter levels were 1.2-fold higher compared to the control group. In the same study, the alpha-melanocyte-stimulating hormone (α-MSH) was monitored. α-MSH acts on melanocortin type 4 receptors (MC4R), a receptor that interacts with neurochemical systems that regulate socioemotional behaviors, including oxytocin and dopamine. Oxytocin can influence social cognition by modulating various neurochemical systems, including serotonin, glutamate, dopamine, and GABA neurotransmitters in specific brain regions such as the hypothalamus, amygdala, and hippocampus. The study observed significantly lower levels of α-MSH in animals treated with propionic acid compared to controls. However, this effect was reversed by the administration of bee pollen and a mixed probiotic bacteria preparation called ProtexinR, which contains beneficial bacteria such as Bifidobacterium infantis, Bifidobacterium breve, Lactobacillus acidophilus, Lactobacillus bulgaricus, Lactobacillus casei, Lactobacillus rhamnosus, and Streptococcus thermophiles. The concentration of the mixed probiotic bacteria in ProtexinR was 1 billion CFU per gram [235].
Pochakom (2022) investigated supplementation with Lacticaseibacillus rhamnosus HA-11 (Lr) and Ligilactobacillus salivarius HA-118 (Ls) in the BTGR mouse model for autism (109 CFU/mL in drinking water for 4 weeks) [239]. Supplementation with Lr, but not Ls, increased the microbial richness and diversity, and increased the concentrations of beneficial neuroactive compounds, such as 5-aminovaleric acid and choline. Both Lr and Ls treatment reduced behavioural deficits in social novelty preference, but no changes in hyperactivity or repetitive behaviour were observed [239]. This suggests that not all probiotic microbes result in the same outcomes and a more complex mix of microbes might actually be required to target various behaviours.
Sen et al. 2022 [240] found that daily oral administration of Blautia stercoris MRx0006 attenuated social and repetitive behavior in a mouse model of autism. The study showed that MRx0006 increases the expression of oxytocin and its receptor in hypothalamic cells in vitro, and hypothalamic oxytocin mRNA in mice, while altering the metabolome profile. It was proposed that biotherapy using Blautia stercoris would be a viable treatment option for autism.
Studies in human subjects are rarer but encouraging. A double blind randomized controlled trial focused on the effect of a synbiotic called Synbiotic 2000, composed of three anti-inflammatory lactic acid bacteria and four anti-inflammatory fibers on patients with ADHD [241]. One of the measured outcomes was repetitive behavior. Synbiotic 2000 reduced both the total score of autism symptoms, and the restricted, repetitive and stereotyped behaviors, as compared to placebo [241]. Similarly, in a case report of a child with Autism, Sacharomyces boulardii was shown to reduce OCD behavior [238].
4.2. Fecal Microbiota Transplants
Fecal microbiota transplantation (FMT), or the transfer of a fecal matter from a healthy donor to a patient, has emerged as another promising therapeutic approach for restoring a healthy gut microbiome and achieving beneficial effects in various diseases [245]. Currently, there are no FMT studies that have been performed specifically to treat OCD. However, several studies have noted the significant changes in microbial ecology, metabolism and behavior observed in patients after FMT, most of them providing strong support for FMT as a therapeutic method to treat OCD [246,247,248,249,250,251].
Kang et al. (2017) published an important follow up after the publication of the first clinical trial results using FMT on autistic children [252]. Spectacular improvements were observed in GI symptoms, autism-related symptoms, and gut microbiota diversity with a higher abundance of Bifidobacteria and Prevotella, and these were sustained after two years [246]. The autism-related symptoms even exhibited further improvement, suggesting that the fecal transplants might have initiated further changes during the two-year period. These findings underscore the long-term safety and effectiveness of FMT as a potential therapy for gut dysbiosis-associated disorders. Although the focus of the study wasn’t OCD behavior, it is particularly relevant considering the close relationship between gut dysbiosis and brain dysfunction. For example, Kilinsarcslan et al. (2020) found that the severity of several factors, including obsession, decreased after FMT in patients with inflammatory bowel disease [253]. This suggests that the restoration of a healthy gut microbial community through FMT can have positive effects on psychological symptoms associated with certain diseases. Alghamdi et al. (2022) conducted a study in a rodent model of Autism and included the use of FMT from healthy donor rats, which resulted in a significant increase in α-MSH levels by 2.7-fold (compared with 1.2 fold for probiotics) and an increase in brain levels of neurotransmitters (1.6 fold) and subtance P (2.2 fold) to above that of the controls [235]. These results suggest that FMT might be superior to probiotics in initiating metabolic changes, however further clinical studies are needed to compare both the efficacy and safety of FMT and probiotics.
Although not focusing on OCD but on Autism, a very interesting and recent study by Wang et al. (2023) highlights important changes after fecal transplants [254]. Fecal microbiota samples from ASD children and healthy donors were transplanted into a mouse model of ASD. The researchers conducted 16S rRNA gene sequencing of fecal samples and untargeted metabolomic analysis of samples to identify differences in gut microbial communities and metabolic pathways related to ASD behaviors. mRNA sequencing analysis was also performed on colon and brain tissues after sacrificing the animals, to identify enriched signaling pathways and potential molecular mechanisms. The study revealed metabolite changes related to serotonergic and glutamatergic synapse pathways. They also demonstrated that these were associated with behavioral changes in ASD: there was an increase in ASD-like behaviours in mice that received FMT from ASD donors but a decrease in such behaviours in mice that received FMT from health donors. Indeed, the colonization of certain bacterial genera, such as Bacteroides, Odoribacter, Turicibacter, and Alistipes, was correlated with the improvement in behavior after FMT, but did not specifically point at OCD behavior. However, the changes in serotoninergic and glutamatergic pathways might also predict positive outcomes for future OCD studies.
The close link between gut dysbiosis and brain function underscores the importance of targeting the gut microbiota for therapeutic interventions. FMT offers a unique opportunity to restore a healthy gut microbial community and potentially alleviate symptoms associated with various disorders. These findings highlight the potential benefits of FMT in improving mental health conditions and support further exploration of this therapeutic approach.
5. Discussion
The correlation between gut dysbiosis and the vast majority of modern diseases is now firmly established [139,147,255,256,257,258,259,260,261,262]. The gut microbiome might actually play a significant role in the development and manifestation of OCD, providing a comprehensive explanation for the multiple factors previously associated with the disorder. The gut microbiome can influence genetic, neurobiological, and environmental factors indirectly, thereby impacting the pathophysiology of OCD.
We have seen that SCFAs can improve gut-brain health via a number of pathways, including: maintening the gut barrier integrity; producing mucus and protecting against inflammation; and communicating with the brain via the vagus nerve and neurohormones[34,263,264,265]. However, the gut bacterial composition is determined by multiple factors, including genetics, immune status, drugs (e.g., metformin), antibiotic courses, diet, pollutants, etc.[211]. Thus, studies investigating the gut microbiota composition must consider multiple variables. Furthermore, the various methods for collection, storage and handling of microbiological materials add even more variability to the studies [266,267]. Microbial reprogramming strategies, using either probiotics or FMT, may also encounter significant challenges arising from inter-individual variations and even temporal variations within a single individual.
The host’s genetic background can modulate bacterial colonization, particularly the genetic variants such as single nucleotide polymorphisms, which could explain the interpersonal variability in circulating levels of SCFA observed after fiber intake. Variations in genes coding the receptors of SCFAs such as GPR41, GPR43 or GPR109A have been proposed to have a significant impact on metabolism in general [101,268,269]. Additionally, genes responsible for transportation, such as the SLC16A family of monocarbohydrate transporters, effector genes like MUC2 involved in colon mucus production, and regulatory genes like NRF2, which regulate the expression of proteins related to antioxidant defense mechanisms, may have significant implications for health outcomes. These effects could arise from compromised absorption of short-chain fatty acids (SCFA) or their intracellular functions [270].
The impact of probiotics on human health has been studied through clinical trials and resulted in numerous suggested health indications and claims [271,272,273,274,275,276]. Nevertheless, there are also studies with contradictory findings, resulting in conflicting, ambiguous conclusions regarding the efficacy of probiotics [277,278]. One of the main reasons for these conflicting results – and also the main challenge for future studies – is that, in contrast to animal models, humans exhibit significant heterogeneity in terms of diet, age range, genetic background, and gut microbiome composition [279,280]. As a result, they may respond differently to the same probiotic intervention. In fact, several studies on probiotics have emphasized the importance of precision in considering host-related factors, microbiome characteristics, and dietary influences, as these factors play a crucial role in determining the varied outcomes observed [281,282]. More specifically, the extent of gut colonization by probiotics can vary significantly among individuals. This variability in colonization levels can contribute to the diverse effects of probiotics on both the hosts themselves and their gut microbiomes[283]. This is understandable since pre-existing microbes can inluence each other’s growth as well as that of the newly ingested microbes. If dysbiosis is already present, such as in atopic dermatitis or milk hypersensitivity, dysbiosis can alter the effects of the probiotic intervention on the host [284,285]. These permissive microbiomes are also more susceptible to compositional and functional changes in response to probiotics, resulting in distinct enrichment of pathways in the gut [286]. Microbiomes that facilitate the colonization of probiotic bacteria are associated with improved clinical responses in various models of colitis and depression [287,288,289].
Therefore, differences in the initial conditions of the host and their gut microbiome, as well as variations in environmental exposures, can lead to contrasting outcomes among individuals who receive the same probiotic supplement [290,291]. In addition, for in vitro studies, the characteristics of probiotic bacteria, such as adhesion, hydrophobicity, and autoaggregation, may vary depending on the source from which they were isolated [292,293].
Environmental factors, as described in our model, influence the gut microbiome and thus the response to probiotics. Dietary polyunsaturated fatty acids, for instance, have been found to modulate the adhesion of probiotics in laboratory settings[290]. Similarly, diet can impact clinical outcomes, as preterm infants fed with human milk demonstrate a reduced risk of late-onset sepsis and a shorter time to achieve full enteral feeding compared to formula-fed infants [294]
Consequently, we consider FMT interventions to be an option with less variability in terms of results. This is because, with FMT, an entire community of microbes, including fungi, is transplanted. However, the downside of this is that we do not currently have enough data to predict, manage and control the eventual risks patients are exposed to when transplanted with the gut microbiome of another healthy individual. Indeed, FMT has shown promise with positive outcomes for various diseases [246,249,295,296,297]. However, the escalating problem of antibiotic resistance poses a threat to the use of FMT. Samples screening must follow rigorous guidelines as antibiotic resistance becomes a criterion for donor stool selection. For FMT to become a successful approach in disease treatment and management, advances are necessary in defining the composition of fecal samples and method of administration. There must also be a shift towards personalized fecal sample selection. The future of a safe FMT probably resides in our ability to further elucidate what a “healthy microbiome” is. New analytical techniques, such as machine learning, might become necessary tools to integrate into omics studies, in order to find the best FMT for OCD [298].
6. Conclusions
We presented a model where dysbiosis plays a pivotal role in the pathogenesis of OCD. To validate this model, more clinical studies are needed, and to shed more light on the potential role of gut microbes in the pathogenesis and treatment of OCD. Exploring the gut microbiome as a target for intervention in OCD holds promise for several reasons. Indeed, this is an opportunity to address the limitations of current treatments and potentially improve treatment outcomes for individuals who experience resistance to current approaches. In addition, the gut microbiome represents a modifiable factor that can be influenced in various ways including dietary interventions, probiotics/prebiotics, or even fecal microbiota transplantation.
Although the use of probiotics and FMT in medicine has been used empirically for centuries, it was only once research acknowledged the importance of the role of the gut microbiota in health and diseases that clinical studies started developing its use. Most studies have used Bifidobacterium such as B. longum, B. breve or B. infantis, and Lactobacillus (L. helveticus and L. rhamnosus), with doses between 10⁸ and 10¹⁰ colony-forming units for about 4 weeks. These probiotics have shown efficacy in improving psychiatric disorder-related behaviors including anxiety, depression, ASD, ADHD, and OCD.
However, research in this area is still in its early stages, and more studies are needed to optimize the methods, and assess the efficacy and safety of microbial reprogramming in OCD. Further investigation and clinical trials will pave the way for more personalized and effective interventions for individuals with OCD, perhaps through the development of disorder-specific probiotic mixtures, making microbiome therapeutics a significant part of the precision medicine field.
Author Contributions
Conceptualization, GB. and CM and RM.; methodology, GB and RM.; writing—original draft preparation, GB and RM and CM; writing—review and editing, GB and CM and RM; visualization, CM. All authors have read and agreed to the published version of the manuscript.”.
Conflicts of Interest
The authors declare no conflict of interest.
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders. Published online May 22, 2013. [CrossRef]
- Oren E, Dar R, Liberman N. Obsessive-Compulsive tendencies are related to a maximization strategy in making decisions. Front Psychol. 2018, 9, 778. [CrossRef]
- Nestadt G, Kamath V, Maher BS, et al. Doubt and the Decision-making Process in Obsessive-Compulsive Disorder. Med Hypotheses. 2016, 96, 1. [CrossRef]
- Pushkarskaya H, Tolin D, Ruderman L, et al. Decision-Making Under Uncertainty in Obsessive-Compulsive Disorder. J Psychiatr Res. 2015, 69:166. [CrossRef]
- Murphy DL, Timpano KR, Wheaton MG, Greenberg BD, Miguel EC. Obsessive-compulsive disorder and its related disorders: a reappraisal of obsessive-compulsive spectrum concepts. Dialogues Clin Neurosci. 2010, 12, 131. [CrossRef]
- Jalal B, Chamberlain SR, Sahakian BJ. Obsessive-compulsive disorder: Etiology, neuropathology, and cognitive dysfunction. Brain Behav. Published online 2023, e3000. [CrossRef]
- Johnson PM, Kenny PJ. Addiction-like reward dysfunction and compulsive eating in obese rats: Role for dopamine D2 receptors. Nat Neurosci. 2010, 13, 635. [CrossRef]
- Mattheisen M, Samuels JF, Wang Y, et al. Genome-wide association study in obsessive-compulsive disorder: results from the OCGAS. Mol Psychiatry. 2015, 20, 337–344. [CrossRef]
- Frick L, Pittenger C. Microglial Dysregulation in OCD, Tourette Syndrome, and PANDAS. J Immunol Res. 2016, 2016. [CrossRef]
- Mahjani B, Klei L, Mattheisen M, et al. The genetic architecture of obsessive-compulsive disorder: Alleles across the frequency spectrum contribute liability to OCD. medRxiv. Published online 2021. [CrossRef]
- Purty A, Nestadt G, Samuels J, Viswanath B. Genetics of obsessive-compulsive disorder. Indian J Psychiatry. 2019, 61, S37–S43. [CrossRef]
- Fawcett EJ, Power H, Fawcett JM. Women are at greater risk of OCD than men: A meta-analytic review of OCD prevalence worldwide. Journal of Clinical Psychiatry. 2020, 81. [CrossRef]
- Grassi G, Cecchelli C, Vignozzi L, Pacini S. Investigational and Experimental Drugs to Treat Obsessive-Compulsive Disorder. J Exp Pharmacol. 2020, 12, 695. [CrossRef]
- Allen A. Cognitive behavioral therapy of obsessive-compulsive disorder. Dialogues Clin Neurosci. 2010, 12, 199. [CrossRef]
- Kellner M. Drug treatment of obsessive-compulsive disorder. Dialogues Clin Neurosci. 2010, 12, 187. [CrossRef]
- Fineberg NA, Gale TM, Sivakumaran T. A Review of Antipsychotics in the Treatment of Obsessive Compulsive Disorder. 2007, 5, 354–360. [CrossRef]
- Eisen JL, Goodman WK, Keller MB, et al. Patterns of Remission and Relapse in Obsessive-Compulsive Disorder: A 2-Year Prospective Study. J Clin Psychiatry. 1999, 60, 14024. [CrossRef]
- Ferguson JM. SSRI Antidepressant Medications: Adverse Effects and Tolerability. Prim Care Companion J Clin Psychiatry. 2001, 3, 22. [CrossRef]
- Thursby E, Juge N. Introduction to the human gut microbiota. Biochemical Journal. 2017, 474, 1823. [CrossRef]
- Benjamin JL, Hedin CRH, Koutsoumpas A, et al. Smokers with active Crohnʼs disease have a clinically relevant dysbiosis of the gastrointestinal microbiota*. Inflamm Bowel Dis. 2012, 18, 1092–1100. [CrossRef]
- Kang S, Denman SE, Morrison M, et al. Dysbiosis of fecal microbiota in Crohnʼs disease patients as revealed by a custom phylogenetic microarray. Inflamm Bowel Dis. 2010, 16, 2034–2042. [CrossRef]
- Hou M, Xu G, Ran M, Luo W, Wang H. APOE-ε4 Carrier Status and Gut Microbiota Dysbiosis in Patients With Alzheimer Disease. Front Neurosci. 2021, 15. [CrossRef]
- Pistollato F, Sumalla Cano S, Elio I, Masias Vergara M, Giampieri F, Battino M. Role of gut microbiota and nutrients in amyloid formation and pathogenesis of Alzheimer disease. Nutr Rev. 2016, 74, 624–634. [CrossRef]
- Shabbir U, Arshad MS, Sameen A, Oh DH. Crosstalk between gut and brain in alzheimer’s disease: The role of gut microbiota modulation strategies. Nutrients. 2021, 13, 1–23. [CrossRef]
- Scheperjans F, Aho V, Pereira PAB, et al. Gut microbiota are related to Parkinson’s disease and clinical phenotype. Movement Disorders. 2015, 30, 350–358. [CrossRef]
- Madore C, Leyrolle Q, Lacabanne C, et al. Neuroinflammation in Autism: Plausible Role of Maternal Inflammation, Dietary Omega 3, and Microbiota. Neural Plast. 2016, 2016:3597209. [CrossRef]
- De Angelis M, Francavilla R, Piccolo M, De Giacomo A, Gobbetti M. Autism spectrum disorders and intestinal microbiota. Gut Microbes. 2015, 6, 207–213. [CrossRef]
- Li Q, Zhou JM. The microbiota-gut-brain axis and its potential therapeutic role in autism spectrum disorder. Neuroscience. 2016, 324(March):131-139. [CrossRef]
- Ding HT, Taur Y, Walkup JT. Gut Microbiota and Autism: Key Concepts and Findings. J Autism Dev Disord. 2017, 47, 480–489. [CrossRef]
- Zhu F, Guo R, Wang W, et al. Transplantation of microbiota from drug-free patients with schizophrenia causes schizophrenia-like abnormal behaviors and dysregulated kynurenine metabolism in mice. Mol Psychiatry. 2020, 25, 2905–2918. [CrossRef]
- Frazier TH, Dibaise JK, McClain CJ. Gut microbiota, intestinal permeability, obesity-induced inflammation, and liver injury. JPEN J Parenter Enteral Nutr. 2011, 35(5 Suppl):14S-20S. [CrossRef]
- Musso G, Gambino R, Cassader M. Obesity, diabetes, and gut microbiota: the hygiene hypothesis expanded? Diabetes Care. 2010, 33, 2277–2284. [CrossRef]
- Phimister EG, Jess T. Microbiota, Antibiotics, and Obesity. New England Journal of Medicine. 2014, 371, 2526–2528. [CrossRef]
- Clapp M, Aurora N, Herrera L, Bhatia M, Wilen E, Wakefield S. Gut microbiota’s effect on mental health: The gut-brain axis. Clin Pract. 2017, 7, 987. [CrossRef]
- Iannone LF, Preda A, Blottière HM, et al. Microbiota-gut brain axis involvement in neuropsychiatric disorders. Expert Rev Neurother. 2019, 19, 1037–1050. [CrossRef]
- Sommer F, Bäckhed F. The gut microbiota-masters of host development and physiology. Nat Rev Microbiol. 2013, 11, 227–238. [CrossRef]
- Sudo N. Microbiome, HPA axis and production of endocrine hormones in the gut. Adv Exp Med Biol. 2014, 817:177-194. [CrossRef]
- de Weerth C. Do bacteria shape our development? Crosstalk between intestinal microbiota and HPA axis. Neurosci Biobehav Rev. 2017, 83:458-471. [CrossRef]
- Sudo N. Role of microbiome in regulating the HPA axis and its relevance to allergy. Chem Immunol Allergy. 2012, 98:163-175. [CrossRef]
- Sudo N. Role of Microbiome in Regulating the HPA Axis and Its Relevance to Allergy. Published online 2012:163-175. [CrossRef]
- Schönfeld P, Wojtczak L. Short- and medium-chain fatty acids in energy metabolism: The cellular perspective. J Lipid Res. 2016, 57, 943–954. [CrossRef]
- Macfarlane S, Macfarlane GT. Regulation of short-chain fatty acid production. Proceedings of the Nutrition Society. 2003, 62, 67–72. [CrossRef]
- Dalile B, Van Oudenhove L, Vervliet B, Verbeke K. The role of short-chain fatty acids in microbiota–gut–brain communication. Nat Rev Gastroenterol Hepatol. 2019, 16, 461–478. [CrossRef]
- van der Hee B, Wells JM. Microbial Regulation of Host Physiology by Short-chain Fatty Acids. Trends Microbiol. 2021, 29, 700–712. [CrossRef]
- Dalile B, Van Oudenhove L, Vervliet B, Verbeke K. The role of short-chain fatty acids in microbiota–gut–brain communication. Nat Rev Gastroenterol Hepatol. 2019, 16, 461–478. [CrossRef]
- Vijay N, Morris M. Role of Monocarboxylate Transporters in Drug Delivery to the Brain. Curr Pharm Des. 2014, 20, 1487–1498. [CrossRef]
- Bolognini D, Tobin AB, Milligan G, Moss CE. The pharmacology and function of receptors for short-chain fatty acids. Mol Pharmacol. 2016, 89, 388–398. [CrossRef]
- Asbjornsdottir B, Snorradottir H, Andresdottir E, et al. Zonulin-Dependent Intestinal Permeability in Children Diagnosed with Mental Disorders: A Systematic Review and Meta-Analysis. Nutrients. 2020, 12, 1–27. [CrossRef]
- Caviglia GP, Rosso C, Ribaldone DG, et al. Physiopathology of intestinal barrier and the role of zonulin. Minerva Biotecnol. 2019, 31, 83–92. [CrossRef]
- Louis P, Flint HJ. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol Lett. 2009, 294, 1–8. [CrossRef]
- Rivière A, Selak M, Lantin D, Leroy F, De Vuyst L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front Microbiol. 2016, 7:979. [CrossRef]
- Szentirmai É, Millican NS, Massie AR, Kapás L. Butyrate, a metabolite of intestinal bacteria, enhances sleep. Sci Rep. 2019, 9(1). [CrossRef]
- Stilling RM, van de Wouw M, Clarke G, Stanton C, Dinan TG, Cryan JF. The neuropharmacology of butyrate: The bread and butter of the microbiota-gut-brain axis? Neurochem Int. 2016, 99:110-132. [CrossRef]
- Hamer HM, Jonkers D, Venema K, Vanhoutvin S, Troost FJ, Brummer RJ. Review article: The role of butyrate on colonic function. Aliment Pharmacol Ther. 2008, 27, 104–119. [CrossRef]
- Kelly JR, Kennedy PJ, Cryan JF, Dinan TG, Clarke G, Hyland NP. Breaking down the barriers: the gut microbiome, intestinal permeability and stress-related psychiatric disorders. Front Cell Neurosci. 2015, 9:392. [CrossRef]
- Doney E, Dion-Albert L, Coulombe-Rozon F, et al. Chronic stress exposure alters the gut barrier: sex-specific effects on microbiota and jejunum tight junctions. Biological Psychiatry Global Open Science. Published online May 2023. [CrossRef]
- Viggiano D, Ianiro G, Vanella G, et al. Gut barrier in health and disease: focus on childhood. Eur Rev Med Pharmacol Sci. 2015, 19, 1077–1085. Accessed March 13, 2017. http://www.ncbi.nlm.nih.gov/pubmed/25855935.
- Braniste V, Al-Asmakh M, Kowal C, et al. The gut microbiota influences blood-brain barrier permeability in mice. Sci Transl Med. 2014, 6(263). [CrossRef]
- Bien-Ly N, Watts RJ. The blood-brain barrier’s gut check. Sci Transl Med. 2014, 6(263). [CrossRef]
- Smith O. The gut microbiota and the blood-brain barrier. Sci Signal. 2014, 7(353). [CrossRef]
- Oldendorf WH. Carrier-mediated blood-brain barrier transport of short-chain monocarboxylic organic acids. 1973, 224, 1450–1453. [CrossRef]
- Bachmann C, Colombo JP, Berüter J. Short chain fatty acids in plasma and brain: Quantitative determination by gas chromatography. Clinica Chimica Acta. 1979, 92, 153–159. [CrossRef]
- Liu J, Sun J, Wang F, et al. Neuroprotective Effects of Clostridium butyricum against Vascular Dementia in Mice via Metabolic Butyrate. Biomed Res Int. 2015, 2015. [CrossRef]
- Sun J, Ling Z, Wang F, et al. Clostridium butyricum pretreatment attenuates cerebral ischemia/reperfusion injury in mice via anti-oxidation and anti-apoptosis. Neurosci Lett. 2016, 613:30-35. [CrossRef]
- Hoyles L, Snelling T, Umlai UK, et al. Microbiome–host systems interactions: Protective effects of propionate upon the blood–brain barrier. Microbiome. 2018, 6, 1–13. [CrossRef]
- Jin X, Chen D, Wu F, et al. Hydrogen Sulfide Protects Against Ammonia-Induced Neurotoxicity Through Activation of Nrf2/ARE Signaling in Astrocytic Model of Hepatic Encephalopathy. Front Cell Neurosci. 2020, 14:336. [CrossRef]
- Blachier F, Andriamihaja M, Larraufie P, Ahn E, Lan A, Kim E. Production of hydrogen sulfide by the intestinal microbiota and epithelial cells and consequences for the colonic and rectal mucosa. Am J Physiol Gastrointest Liver Physiol. 2021, 320, G125–G135. [CrossRef]
- Tomasova L, Konopelski P, Ufnal M. Gut Bacteria and Hydrogen Sulfide: The New Old Players in Circulatory System Homeostasis. Molecules. 2016, 21(11). [CrossRef]
- Kossewska J, Bierlit K, Trajkovski V. Personality, Anxiety, and Stress in Patients with Small Intestine Bacterial Overgrowth Syndrome. The Polish Preliminary Study. Int J Environ Res Public Health. 2023, 20(1). [CrossRef]
- Frost G, Sleeth ML, Sahuri-Arisoylu M, et al. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat Commun. 2014, 5. [CrossRef]
- Weis AM, Round JL. Microbiota-antibody interactions that regulate gut homeostasis. Cell Host Microbe. 2021, 29, 334–346. [CrossRef]
- Abokor AA, McDaniel GH, Golonka RM, et al. Immunoglobulin A, an Active Liaison for Host-Microbiota Homeostasis. Microorganisms. 2021, 9(10). [CrossRef]
- Lyte M. Microbial Endocrinology in the Microbiome-Gut-Brain Axis: How Bacterial Production and Utilization of Neurochemicals Influence Behavior. PLoS Pathog. 2013, 9(11). [CrossRef]
- Desbonnet L, Garrett L, Clarke G, Kiely B, Cryan JF, Dinan TG. Effects of the probiotic Bifidobacterium infantis in the maternal separation model of depression. Neuroscience. 2010, 170, 1179–1188. [CrossRef]
- Reigstad CS, Salmonson CE, Rainey JF, et al. Gut microbes promote colonic serotonin production through an effect of short-chain fatty acids on enterochromaffin cells. The FASEB Journal. 2015, 29, 1395–1403. [CrossRef]
- Yano JM, Yu K, Donaldson GP, et al. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell. 2015, 161, 264–276. [CrossRef]
- Dalile B, Van Oudenhove L, Vervliet B, Verbeke K. The role of short-chain fatty acids in microbiota–gut–brain communication. Nature Reviews Gastroenterology & Hepatology 2019 16:8. 2019, 16, 461–478. [CrossRef]
- Yano JM, Yu K, Donaldson GP, et al. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell. 2015, 161, 264–276. [CrossRef]
- Reigstad CS, Salmonson CE, Rainey JF, et al. Gut microbes promote colonic serotonin production through an effect of short-chain fatty acids on enterochromaffin cells. The FASEB Journal. 2015, 29, 1395–1403. [CrossRef]
- Nankova BB, Agarwal R, MacFabe DF, La Gamma EF. Enteric Bacterial Metabolites Propionic and Butyric Acid Modulate Gene Expression, Including CREB-Dependent Catecholaminergic Neurotransmission, in PC12 Cells - Possible Relevance to Autism Spectrum Disorders. PLoS One. 2014, 9, e103740. [CrossRef]
- Clarke G, Stilling RM, Kennedy PJ, Stanton C, Cryan JF, Dinan TG. Minireview: Gut Microbiota: The Neglected Endocrine Organ. Molecular Endocrinology. 2014, 28, 1221–1238. [CrossRef]
- Bonaz B, Bazin T, Pellissier S. The Vagus Nerve at the Interface of the Microbiota-Gut-Brain Axis. Front Neurosci. 2018, 12:49. [CrossRef]
- Breit S, Kupferberg A, Rogler G, Hasler G. Vagus Nerve as Modulator of the Brain-Gut Axis in Psychiatric and Inflammatory Disorders. Front Psychiatry. 2018, 9:44. [CrossRef]
- Bravo JA, Forsythe P, Chew M V, et al. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc Natl Acad Sci U S A. 2011, 108, 16050–16055. [CrossRef]
- Wang X, Wang BR, Zhang XJ, Xu Z, Ding YQ, Ju G. Evidences for vagus nerve in maintenance of immune balance and transmission of immune information from gut to brain in STM-infected rats. World J Gastroenterol. 2002, 8, 540–545. [CrossRef]
- Zhang L, Wang Y, Xiayu X, et al. Altered Gut Microbiota in a Mouse Model of Alzheimer’s Disease. Journal of Alzheimer’s Disease. 2017, 60, 1241–1257. [CrossRef]
- Ho L, Ono K, Tsuji M, Mazzola P, Singh R, Pasinetti GM. Protective roles of intestinal microbiota derived short chain fatty acids in Alzheimer’s disease-type beta-amyloid neuropathological mechanisms. Expert Rev Neurother. 2018, 18, 83–90. [CrossRef]
- Yoo DY, Kim W, Nam SM, et al. Synergistic effects of sodium butyrate, a histone deacetylase inhibitor, on increase of neurogenesis induced by pyridoxine and increase of neural proliferation in the mouse dentate gyrus. Neurochem Res. 2011, 36, 1850–1857. [CrossRef]
- Silva YP, Bernardi A, Frozza RL. The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front Endocrinol (Lausanne). 2020, 11:25. [CrossRef]
- Nicholson JK, Holmes E, Kinross J, et al. Host-gut microbiota metabolic interactions. Science (1979). 2012, 336, 1262–1267. [CrossRef]
- Tilg H, Kaser A. Gut microbiome, obesity, and metabolic dysfunction. Journal of Clinical Investigation. 2011, 121, 2126–2132. [CrossRef]
- Koren O, Goodrich JK, Cullender TC, et al. Host remodeling of the gut microbiome and metabolic changes during pregnancy. Cell. 2012, 150, 470–480. [CrossRef]
- Kasubuchi M, Hasegawa S, Hiramatsu T, Ichimura A, Kimura I. Dietary gut microbial metabolites, short-chain fatty acids, and host metabolic regulation. Nutrients. 2015, 7, 2839–2849. [CrossRef]
- Moreira APB, Texeira TFS, Ferreira AB, Peluzio M do CG, Alfenas R de CG. Influence of a high-fat diet on gut microbiota, intestinal permeability and metabolic endotoxaemia. Br J Nutr. 2012, 108, 801–809. [CrossRef]
- Rosean CB, Bostic RR, Ferey JCM, et al. Preexisting commensal dysbiosis is a host-intrinsic regulator of tissue inflammation and tumor cell dissemination in hormone receptor–positive breast cancer. Cancer Res. 2019, 79, 3662–3675. [CrossRef]
- Pai YC, Li YH, Turner JR, Yu LCH. Transepithelial Barrier Dysfunction Drives Microbiota Dysbiosis to Initiate Epithelial Clock-driven Inflammation. J Crohns Colitis. Published online April 2, 2023:1-18. [CrossRef]
- Wang T, He C. Pro-inflammatory cytokines: The link between obesity and osteoarthritis. Cytokine Growth Factor Rev. 2018, 44:38-50. [CrossRef]
- Diamanti AP, Manuela Rosado M, Laganà B, D’Amelio R. Microbiota and chronic inflammatory arthritis: An interwoven link. J Transl Med. 2016, 14(1). [CrossRef]
- Schirmer M, Smeekens SP, Vlamakis H, et al. Linking the Human Gut Microbiome to Inflammatory Cytokine Production Capacity. Cell. 2016, 167, 1125. [CrossRef]
- Liu B, Ding Z, Xiong J, Heng X, Wang H, Chu W. Gut Microbiota and Inflammatory Cytokine Changes in Patients with Ankylosing Spondylitis. Biomed Res Int. 2022, 2022. [CrossRef]
- Maslowski KM, Vieira AT, Ng A, et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature. 2009, 461, 1282–1286. [CrossRef]
- An L, Wirth U, Koch D, et al. The Role of Gut-Derived Lipopolysaccharides and the Intestinal Barrier in Fatty Liver Diseases. Journal of Gastrointestinal Surgery. 2022, 26, 671. [CrossRef]
- De Vadder F, Kovatcheva-Datchary P, Goncalves D, et al. Microbiota-generated metabolites promote metabolic benefits via gut-brain neural circuits. Cell. 2014, 156(1-2):84-96. [CrossRef]
- Fung TC, Olson CA, Hsiao EY. Interactions between the microbiota, immune and nervous systems in health and disease. Nature Neuroscience 2016 20:2. 2017, 20, 145–155. [CrossRef]
- Arpaia N, Campbell C, Fan X, et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013 504:7480. 2013, 504, 451–455. [CrossRef]
- Smith PM, Howitt MR, Panikov N, et al. The microbial metabolites, short-chain fatty acids, regulate colonic T reg cell homeostasis. Science (1979). 2013, 341, 569–573. [CrossRef]
- Haghikia A, Jörg S, Duscha A, et al. Dietary Fatty Acids Directly Impact Central Nervous System Autoimmunity via the Small Intestine. Immunity. 2015, 43, 817–829. [CrossRef]
- Erny D, De Angelis ALH, Jaitin D, et al. Host microbiota constantly control maturation and function of microglia in the CNS. Nature Neuroscience 2015 18:7. 2015, 18, 965–977. [CrossRef]
- Stanisavljević S, Čepić A, Bojić S, et al. Oral neonatal antibiotic treatment perturbs gut microbiota and aggravates central nervous system autoimmunity in Dark Agouti rats. Scientific Reports 2019 9:1. 2019, 9, 1–13. [CrossRef]
- Minter MR, Zhang C, Leone V, et al. Antibiotic-induced perturbations in gut microbial diversity influences neuro-inflammation and amyloidosis in a murine model of Alzheimer’s disease. Scientific Reports 2016 6:1. 2016, 6, 1–12. [CrossRef]
- Minter MR, Hinterleitner R, Meisel M, et al. Antibiotic-induced perturbations in microbial diversity during post-natal development alters amyloid pathology in an aged APPSWE/PS1ΔE9 murine model of Alzheimer’s disease. Scientific Reports 2017 7:1. 2017, 7, 1–18. [CrossRef]
- Jang HM, Lee HJ, Jang SE, Han MJ, Kim DH. Evidence for interplay among antibacterial-induced gut microbiota disturbance, neuro-inflammation, and anxiety in mice. Mucosal Immunol. 2018, 11, 1386–1397. [CrossRef]
- Patnala R, Arumugam T V., Gupta N, Dheen ST. HDAC Inhibitor Sodium Butyrate-Mediated Epigenetic Regulation Enhances Neuroprotective Function of Microglia During Ischemic Stroke. Mol Neurobiol. 2017, 54, 6391–6411. [CrossRef]
- Wang P, Zhang Y, Gong Y, et al. Sodium butyrate triggers a functional elongation of microglial process via Akt-small RhoGTPase activation and HDACs inhibition. Neurobiol Dis. 2018, 111:12-25. [CrossRef]
- Sharon G, Cruz NJ, Kang DW, et al. Human Gut Microbiota from Autism Spectrum Disorder Promote Behavioral Symptoms in Mice. Cell. 2019, 177, 1600–1618.e17. [CrossRef]
- Soliman ML, Puig KL, Combs CK, Rosenberger TA. Acetate reduces microglia inflammatory signaling in vitro. J Neurochem. 2012, 123, 555–567. [CrossRef]
- Reddy DS, Wu X, Golub VM, Dashwood WM, Dashwood RH. Measuring Histone Deacetylase Inhibition in the Brain. Curr Protoc Pharmacol. 2018, 81, e41. [CrossRef]
- Phalipon A, Cardona A, Kraehenbuhl JP, Edelman L, Sansonetti PJ, Corthésy B. Secretory component: A new role in secretory IgA-mediated immune exclusion in vivo. Immunity. 2002, 17, 107–115. [CrossRef]
- Mora JR, Iwata M, Eksteen B, et al. Generation of gut-homing IgA-secreting B cells by intestinal dendritic cells. Science (1979). 2006, 314, 1157–1160. [CrossRef]
- Huus KE, Petersen C, Finlay BB. Diversity and dynamism of IgA−microbiota interactions. Nat Rev Immunol. 2021, 21, 514–525. [CrossRef]
- Senior BW, Dunlop JI, Batten MR, Kilian M, Woof JM. Cleavage of a recombinant human immunoglobulin A2 (IgA2)-IgA1 hybrid antibody by certain bacterial IgA1 proteases. Infect Immun. 2000, 68, 463–469. [CrossRef]
- Mantis NJ, Rol N, Corthésy B. Secretory IgA’s complex roles in immunity and mucosal homeostasis in the gut. Mucosal Immunol. 2011, 4, 603–611. [CrossRef]
- Woof JM, Russell MW. Structure and function relationships in IgA. Mucosal Immunol. 2011, 4, 590–597. [CrossRef]
- Bunker JJ, Erickson SA, Flynn TM, et al. Natural polyreactive IgA antibodies coat the intestinal microbiota. Science (1979). 2017, 358(6361). [CrossRef]
- Fransen F, Zagato E, Mazzini E, et al. BALB/c and C57BL/6 Mice Differ in Polyreactive IgA Abundance, which Impacts the Generation of Antigen-Specific IgA and Microbiota Diversity. Immunity. 2015, 43, 527–540. [CrossRef]
- Sterlin D, Fadlallah J, Adams O, et al. Human IgA binds a diverse array of commensal bacteria. Journal of Experimental Medicine. 2020, 217(3). [CrossRef]
- Yang Y, Palm NW. Immunoglobulin A and the microbiome. Curr Opin Microbiol. 2020, 56:89-96. [CrossRef]
- Janzon A, Goodrich JK, Koren O, Waters JL, Ley RE. Interactions between the Gut Microbiome and Mucosal Immunoglobulins A, M, and G in the Developing Infant Gut. mSystems. 2019, 4(6). [CrossRef]
- Donaldson GP, Ladinsky MS, Yu KB, et al. Gut microbiota utilize immunoglobulin a for mucosal colonization. Science (1979). 2018, 360, 795–800. [CrossRef]
- Sutherland DB, Suzuki K, Fagarasan S. Fostering of advanced mutualism with gut microbiota by Immunoglobulin A. Immunol Rev. 2016, 270, 20–31. [CrossRef]
- Dixon BREA, Radin JN, Piazuelo MB, Contreras DC, Algood HMS. IL-17a and IL-22 Induce Expression of Antimicrobials in Gastrointestinal Epithelial Cells and May Contribute to Epithelial Cell Defense against Helicobacter pylori. PLoS One. 2016, 11, 148514. [CrossRef]
- Keir ME, Yi T, Lu TT, Ghilardi N. The role of IL-22 in intestinal health and disease. J Exp Med. 2020, 217(3). [CrossRef]
- Selsted ME, Miller SI, Henschen AH, Ouellette AJ. Enteric defensins: antibiotic peptide components of intestine host defense. J Cell Biol. 1992, 118, 929–936. [CrossRef]
- Salzman NH, Ghosh D, Huttner KM, Paterson Y, Bevins CL. Protection against enteric salmonellosis in transgenic mice expressing a human intestinal defensin. Nature 2003 422:6931. 2003, 422, 522–526. [CrossRef]
- Boman HG. Peptide antibiotics and their role in innate immunity. Annu Rev Immunol. 1995, 13:61-92. [CrossRef]
- Rajabi M, De Leeuw E, Pazgier M, Li J, Lubkowski J, Lu W. The Conserved Salt Bridge in Human α-Defensin 5 Is Required for Its Precursor Processing and Proteolytic Stability*âTM¦. Published online 2008. [CrossRef]
- Turna J, Grosman Kaplan K, Anglin R, et al. The gut microbiome and inflammation in obsessive-compulsive disorder patients compared to age- and sex-matched controls: a pilot study. Acta Psychiatr Scand. 2020, 142, 337–347. [CrossRef]
- Rees JC. Obsessive-compulsive disorder and gut microbiota dysregulation. Med Hypotheses. 2014, 82, 163–166. [CrossRef]
- Turna J, Grosman Kaplan K, Anglin R, Van Ameringen M. “wHAT’S BUGGING the GUT in OCD?” A REVIEW of the GUT MICROBIOME in OBSESSIVE-COMPULSIVE DISORDER. Depress Anxiety. 2016, 33, 171–178. [CrossRef]
- Domènech L, Willis J, Alemany-Navarro M, et al. Changes in the stool and oropharyngeal microbiome in obsessive-compulsive disorder. Sci Rep. 2022, 12(1). [CrossRef]
- Higher prevalence of irritable bowel syndrome and greater gastrointestinal symptoms in obsessive-compulsive disorder - PubMed. Accessed May 27, 2023. https://pubmed.ncbi.nlm.nih.gov/31437616/. [CrossRef]
- Troyer EA, Kohn JN, Ecklu-Mensah G, Aleti G, Rosenberg DR, Hong S. Searching for host immune-microbiome mechanisms in obsessive-compulsive disorder: A narrative literature review and future directions. Neurosci Biobehav Rev. 2021, 125:517-534. [CrossRef]
- Turna J, Grosman Kaplan K, Anglin R, et al. The gut microbiome and inflammation in obsessive-compulsive disorder patients compared to age- and sex-matched controls: a pilot study. Acta Psychiatr Scand. 2020, 142, 337–347. [CrossRef]
- Quagliariello A, Del Chierico F, Russo A, et al. Gut microbiota profiling and gut-brain crosstalk in children affected by pediatric acute-onset neuropsychiatric syndrome and pediatric autoimmune neuropsychiatric disorders associated with streptococcal infections. Front Microbiol. 2018, 9(APR):675. [CrossRef]
- Jiang H yin, Zhou Y yue, Zhou G ling, et al. Gut microbiota profiles in treatment-naïve children with attention deficit hyperactivity disorder. Behavioural Brain Research. 2018, 347:408-413. [CrossRef]
- Wang LJ, Yang CY, Chou WJ, et al. Gut microbiota and dietary patterns in children with attention-deficit/hyperactivity disorder. Eur Child Adolesc Psychiatry. Published online May 22, 2019. [CrossRef]
- Aarts E, Ederveen THA, Naaijen J, et al. Gut microbiome in ADHD and its relation to neural reward anticipation. PLoS One. 2017, 12, e0183509. [CrossRef]
- Axelsson PB, Clausen TD, Petersen AH, et al. Relation between Infant Microbiota and Autism?: Results from a National Cohort Sibling Design Study. Epidemiology. 2019, 30, 52–60. [CrossRef]
- Wang L, Christophersen CT, Sorich MJ, Gerber JP, Angley MT, Conlon MA. Low relative abundances of the mucolytic bacterium Akkermansia muciniphila and Bifidobacterium spp. in feces of children with autism. Appl Environ Microbiol. 2011, 77, 6718–6721. [CrossRef]
- Lu Bai HMXH. The Development of Native Chinese Affective Picture System--A pretest in 46 College Students. Chinese Mental Health Journal. 2005, 19, 719–722. Accessed August 3, 2021. https://psycnet.apa.org/record/2005-15454-001.
- Kang DW, Park JG, Ilhan ZE, et al. Reduced Incidence of Prevotella and Other Fermenters in Intestinal Microflora of Autistic Children. PLoS One. 2013, 8(7). [CrossRef]
- Rǎdulescu A, Herron J, Kennedy C, Scimemi A. Global and local excitation and inhibition shape the dynamics of the cortico-striatal-thalamo-cortical pathway. Sci Rep. 2017, 7(1). [CrossRef]
- Brem S, Grünblatt E, Drechsler R, Riederer P, Walitza S. The neurobiological link between OCD and ADHD. Atten Defic Hyperact Disord. 2014, 6, 175–202. [CrossRef]
- Gao J, Zhou Y, Yang X, et al. Abnormalities within and beyond the cortico-striato-thalamo-cortical circuitry in medication-free patients with OCD revealed by the fractional amplitude of low-frequency fluctuations and resting-state functional connectivity. Neurosci Lett. 2019, 712. [CrossRef]
- O’Mahony SM, Clarke G, Borre YE, Dinan TG, Cryan JF. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behavioural Brain Research. 2015, 277:32-48. [CrossRef]
- Yano JM, Yu K, Donaldson GP, et al. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell. 2015, 161, 264–276. [CrossRef]
- Patients with obsessive-compulsive disorder have increased 5-HT2A receptor binding in the caudate nuclei - PubMed. Accessed May 31, 2023. https://pubmed.ncbi.nlm.nih.gov/15801987/. [CrossRef]
- Simpson CA, Mu A, Haslam N, Schwartz OS, Simmons JG. Feeling down? A systematic review of the gut microbiota in anxiety/depression and irritable bowel syndrome. J Affect Disord. 2020, 266:429-446. [CrossRef]
- Menzies RG, O’Brian S, Onslow M, Packman A, St. Clare T, Block S. An experimental clinical trial of a cognitive-behavior therapy package for chronic stuttering. J Speech Lang Hear Res. 2008, 51, 1451–1464. [CrossRef]
- Dougherty DM, Mathias CW, Marsh DM, Papageorgiou TD, Swann AC, Moeller FG. Laboratory Measured Behavioral Impulsivity Relates to Suicide Attempt History. Suicide Life Threat Behav. 2004, 34, 374–385. [CrossRef]
- Sudo N, Chida Y, Aiba Y, et al. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. Journal of Physiology. 2004, 558, 263–275. [CrossRef]
- Dell’Osso L, Carmassi C, Mucci F, Marazziti D. Depression, Serotonin and Tryptophan. Curr Pharm Des. 2016, 22, 949–954. [CrossRef]
- Cangiano C, Cardelli-Cangiano P, Cascino A, et al. On the stimulation by insulin of tryptophan transport across the blood-brain barrier. Biochem Int. 1983, 7, 617–627. http://europepmc.org/abstract/MED/6091659.
- Waclawiková B, El Aidy S. Role of Microbiota and Tryptophan Metabolites in the Remote Effect of Intestinal Inflammation on Brain and Depression. Pharmaceuticals (Basel). 2018, 11, 63. [CrossRef]
- Morris G, Berk M, Carvalho A, et al. The Role of the Microbial Metabolites Including Tryptophan Catabolites and Short Chain Fatty Acids in the Pathophysiology of Immune-Inflammatory and Neuroimmune Disease. Mol Neurobiol. Published online June 27, 2016. [CrossRef]
- Hsiao EY, McBride SW, Hsien S, et al. Microbiota Modulate Behavioral and Physiological Abnormalities Associated with Neurodevelopmental Disorders. Cell. 2013, 155, 1451–1463. [CrossRef]
- Rosenberg DR, Macmaster FP, Keshavan MS, Fitzgerald KD, Stewart CM, Moore GJ. Decrease in caudate glutamatergic concentrations in pediatric obsessive-compulsive disorder patients taking paroxetine. J Am Acad Child Adolesc Psychiatry. 2000, 39, 1096–1103. [CrossRef]
- Goddard AW, Shekhar A, Whiteman AF, McDougle CJ. Serotoninergic mechanisms in the treatment of obsessive-compulsive disorder. Drug Discov Today. 2008, 13(7-8):325-332. [CrossRef]
- Chakrabarty K, Bhattacharyya S, Christopher R, Khanna S. Glutamatergic dysfunction in OCD. Neuropsychopharmacology. 2005, 30, 1735–1740. [CrossRef]
- Grant JE, Odlaug BL. Update on pathological skin picking. Curr Psychiatry Rep. 2009, 11, 283–288. [CrossRef]
- Grant JE, Chamberlain SR, Redden SA, Leppink EW, Odlaug BL, Kim SW. N-Acetylcysteine in the Treatment of Excoriation Disorder: A Randomized Clinical Trial. JAMA Psychiatry. 2016, 73, 490–496. [CrossRef]
- Strandwitz P. Neurotransmitter modulation by the gut microbiota. Brain Res. 2018, 1693:128-133.
- Strandwitz P, Kim KH, Terekhova D, et al. GABA-modulating bacteria of the human gut microbiota. Nat Microbiol. 2019, 4, 396–403. [CrossRef]
- Denys D, Fluitman S, Kavelaars A, Heijnen C, Westenberg H. Decreased TNF-α and NK activity in obsessive-compulsive disorder. Psychoneuroendocrinology. 2004, 29, 945–952. [CrossRef]
- Van Der Wee NJ, Stevens H, Hardeman JA, et al. Enhanced dopamine transporter density in psychotropic-naive patients with obsessive-compulsive disorder shown by [123I]{beta}-CIT SPECT. Am J Psychiatry. 2004, 161, 2201–2206. [CrossRef]
- Bercik P, Denou E, Collins J, et al. The intestinal microbiota affect central levels of brain-derived neurotropic factor and behavior in mice. Gastroenterology. 2011, 141(2). [CrossRef]
- Dinan TG, Cryan JF. Melancholic microbes: a link between gut microbiota and depression? Neurogastroenterology & Motility. 2013, 25, 713–719. [CrossRef]
- Dinan TG, Cryan JF. The Microbiome-Gut-Brain Axis in Health and Disease. Gastroenterol Clin North Am. 2017, 46, 77–89. [CrossRef]
- Dinan TG, Cryan JF. The impact of gut microbiota on brain and behaviour: implications for psychiatry. Curr Opin Clin Nutr Metab Care. 2015, 18, 552–558. [CrossRef]
- Cryan JF, Dinan TG. Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nat Rev Neurosci. 2012, 13, 701–712. [CrossRef]
- Marazziti D, Palermo S, Arone A, et al. Obsessive-Compulsive Disorder, PANDAS, and Tourette Syndrome: Immuno-inflammatory Disorders. Adv Exp Med Biol. 2023, 1411:275-300. [CrossRef]
- Williams K, Shorser-Gentile L, Sarvode Mothi S, et al. Immunoglobulin A Dysgammaglobulinemia Is Associated with Pediatric-Onset Obsessive-Compulsive Disorder. J Child Adolesc Psychopharmacol. 2019, 29, 268. [CrossRef]
- Westwell-Roper C, Williams KA, Samuels J, et al. Immune-Related Comorbidities in Childhood-Onset Obsessive Compulsive Disorder: Lifetime Prevalence in the Obsessive Compulsive Disorder Collaborative Genetics Association Study. J Child Adolesc Psychopharmacol. 2019, 29, 615. [CrossRef]
- Palm NW, de Zoete MR, Flavell RA. Immune-microbiota interactions in health and disease. Clinical Immunology. 2015, 159, 122–127. [CrossRef]
- McCusker RH, Kelley KW. Immune-neural connections: How the immune system’s response to infectious agents influences behavior. Journal of Experimental Biology. 2013, 216, 84–98. [CrossRef]
- Lazar V, Ditu LM, Pircalabioru GG, et al. Aspects of gut microbiota and immune system interactions in infectious diseases, immunopathology, and cancer. Front Immunol. 2018, 9(AUG). [CrossRef]
- Robbins TW, Vaghi MM, Banca P. Obsessive-Compulsive Disorder: Puzzles and Prospects. Neuron. 2019, 102, 27–47. [CrossRef]
- Iervolino AC, Rijsdijk F V., Cherkas L, Fullana MA, Mataix-Cols D. A multivariate twin study of obsessive-compulsive symptom dimensions. Arch Gen Psychiatry. 2011, 68, 637. [CrossRef]
- Mahjani B, Bey K, Boberg J, Burton C. Genetics of obsessive-compulsive disorder. Psychol Med. 2021, 51, 2247–2259. [CrossRef]
- Schiele MA, Thiel C, Kollert L, et al. Oxytocin Receptor Gene DNA Methylation: A Biomarker of Treatment Response in Obsessive-Compulsive Disorder? Psychother Psychosom. 2020, 90, 57–63. [CrossRef]
- Zilhão NR, Smit DJ, Boomsma DI, Cath DC. Cross-disorder genetic analysis of tic disorders, obsessive-compulsive, and hoarding symptoms. Front Psychiatry. 2016, 7(JUN). [CrossRef]
- Rotge JY, Aouizerate B, Tignol J, Bioulac B, Burbaud P, Guehl D. The glutamate-based genetic immune hypothesis in obsessive-compulsive disorder. An integrative approach from genes to symptoms. Neuroscience. 2010, 165, 408–417. [CrossRef]
- Gassó P, Ortiz AE, Mas S, et al. Association between genetic variants related to glutamatergic, dopaminergic and neurodevelopment pathways and white matter microstructure in child and adolescent patients with obsessive-compulsive disorder. J Affect Disord. 2015, 186:284-292. [CrossRef]
- Mohammadi AH, Karimian M, Mirzaei H, Milajerdi A. Epigenetic modifications and obsessive–compulsive disorder: what do we know? Brain Struct Funct. Published online May 19, 2023:1-11. [CrossRef]
- Dondu A, Caliskan M, Orenay-Boyacioglu S. Link between obsessive-compulsive disorder and polymorphisms in HDAC genes. Brazilian Journal of Psychiatry. 2022, 44, 156. [CrossRef]
- Kratsman N, Getselter D, Elliott E. Sodium butyrate attenuates social behavior deficits and modifies the transcription of inhibitory/excitatory genes in the frontal cortex of an autism model. Neuropharmacology. 2016, 102:136-145. [CrossRef]
- Cappi C, Diniz JB, Requena GL, et al. Epigenetic evidence for involvement of the oxytocin receptor gene in obsessive-compulsive disorder. BMC Neurosci. 2016, 17, 1–8. [CrossRef]
- D’Addario C, Pucci M, Bellia F, et al. Regulation of oxytocin receptor gene expression in obsessive–compulsive disorder: a possible role for the microbiota-host epigenetic axis. Clin Epigenetics. 2022, 14(1). [CrossRef]
- D’Addario C, Pucci M, Bellia F, et al. Regulation of oxytocin receptor gene expression in obsessive–compulsive disorder: a possible role for the microbiota-host epigenetic axis. Clin Epigenetics. 2022, 14(1). [CrossRef]
- Bey K, Campos-Martin R, Klawohn J, et al. Hypermethylation of the oxytocin receptor gene (OXTR) in obsessive-compulsive disorder: further evidence for a biomarker of disease and treatment response. Epigenetics. 2022, 17, 642–652. [CrossRef]
- Schiele MA, Thiel C, Deckert J, Zaudig M, Berberich G, Domschke K. Monoamine Oxidase A Hypomethylation in Obsessive-Compulsive Disorder: Reversibility By Successful Psychotherapy? Int J Neuropsychopharmacol. 2020, 23, 319–323. [CrossRef]
- D’Addario C, Bellia F, Benatti B, et al. Exploring the role of BDNF DNA methylation and hydroxymethylation in patients with obsessive compulsive disorder. J Psychiatr Res. 2019, 114:17-23. [CrossRef]
- Nissen JB, Hansen CS, Starnawska A, et al. DNA Methylation at the Neonatal State and at the Time of Diagnosis: Preliminary Support for an Association with the Estrogen Receptor 1, Gamma-Aminobutyric Acid B Receptor 1, and Myelin Oligodendrocyte Glycoprotein in Female Adolescent Patients with OCD. Front Psychiatry. 2016, 7(MAR). [CrossRef]
- Frisch A, Michaelovsky E, Rockah R, et al. Association between obsessive-compulsive disorder and polymorphisms of genes encoding components of the serotonergic and dopaminergic pathways. Eur Neuropsychopharmacol. 2000, 10, 205–209. [CrossRef]
- Hemmings SMJ, Kinnear CJ, Niehaus DJH, et al. Investigating the role of dopaminergic and serotonergic candidate genes in obsessive-compulsive disorder. European Neuropsychopharmacology. 2003, 13, 93–98. [CrossRef]
- Billett EA, Richter MA, Sam F, et al. Investigation of dopamine system genes in obsessive-compulsive disorder. Psychiatr Genet. 1998, 8, 163–169. [CrossRef]
- Miguita K, Cordeiro Q, Siqueira-Roberto J, et al. Association analysis between a VNTR intron 8 polymorphism of the dopamine transporter gene (SLC6A3) and obsessive- compulsive disorder in a Brazilian sample. Arq Neuropsiquiatr. 2007, 65(4A):936-941. [CrossRef]
- Cruz C, Orozco B. DRD2, DRD3 and 5HT2A receptor genes polymorphisms in Obsessive-Compulsive Disorder. Accessed May 28, 2023. https://www.researchgate.net/publication/14064809.
- Catalano M, Sciuto G, Di Bella D, Novelli E, Nobile M, Bellodi L. Lack of association between obsessive-compulsive disorder and the dopamine D3 receptor gene: some preliminary considerations. Am J Med Genet. 1994, 54, 253–255. [CrossRef]
- Rosin S, Xia K, Azcarate-Peril MA, et al. A Preliminary Study of Gut Microbiome Variation and HPA Axis Reactivity in Healthy Infants. Psychoneuroendocrinology. 2021, 124:105046. [CrossRef]
- Rothschild D, Weissbrod O, Barkan E, et al. Environment dominates over host genetics in shaping human gut microbiota. Nature. 2018, 555, 210–215. [CrossRef]
- Diamanti T, Prete R, Battista N, Corsetti A, De Jaco A. Exposure to Antibiotics and Neurodevelopmental Disorders: Could Probiotics Modulate the Gut-Brain Axis? Antibiotics (Basel). 2022, 11(12). [CrossRef]
- Njotto LL, Simin J, Fornes R, et al. Maternal and Early-Life Exposure to Antibiotics and the Risk of Autism and Attention-Deficit Hyperactivity Disorder in Childhood: a Swedish Population-Based Cohort Study. Drug Saf. Published online May 1, 2023. [CrossRef]
- Slob EMA, Brew BK, Vijverberg SJH, et al. Early-life antibiotic use and risk of attention-deficit hyperactivity disorder and autism spectrum disorder: results of a discordant twin study. Int J Epidemiol. 2021, 50, 475–484. [CrossRef]
- Ahmed S, Travis SD, Díaz-Bahamonde F V., et al. Early Influences of Microbiota on White Matter Development in Germ-Free Piglets. Front Cell Neurosci. 2021, 15:520. [CrossRef]
- Hoban AE, Stilling RM, Ryan FJ, et al. Regulation of prefrontal cortex myelination by the microbiota. Transl Psychiatry. 2016, 6, e774. [CrossRef]
- Tette FM, Kwofie SK, Wilson MD. Therapeutic Anti-Depressant Potential of Microbial GABA Produced by Lactobacillus rhamnosus Strains for GABAergic Signaling Restoration and Inhibition of Addiction-Induced HPA Axis Hyperactivity. Curr Issues Mol Biol. 2022, 44, 1434. [CrossRef]
- Bravo JA, Forsythe P, Chew M V, et al. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc Natl Acad Sci U S A. 2011, 108, 16050–16055. [CrossRef]
- Chávez-Castillo M. Depression as a neuroendocrine disorder: Emerging neuropsychopharmacological approaches beyond monoamines. Adv Pharmacol Pharm Sci. 2019, 2019:7943481.
- Associations of neurotransmitters and the gut microbiome with emotional distress in mixed type of irritable bowel syndrome | Scientific Reports. Accessed May 31, 2023. https://www.nature.com/articles/s41598-022-05756-0. [CrossRef]
- Laswi I, Shafiq A, Al-Ali D, et al. A comparative pilot study of bacterial and fungal dysbiosis in neurodevelopmental disorders and gastrointestinal disorders: Commonalities, specificities and correlations with lifestyle. Microorganisms. 2021, 9, 741. [CrossRef]
- Bendriss G, Al-Ali D, Shafiq A, et al. Targeting the gut microbiome: A brief report on the awareness, practice, and readiness to engage in clinical interventions in Qatar. Qatar Med J. 2021, 2020(3). [CrossRef]
- Sanders ME. Probiotics: Definition, Sources, Selection, and Uses. Clinical Infectious Diseases. 2008, 46(s2):S58-S61. [CrossRef]
- Skott E, Yang LL, Stiernborg M, et al. Effects of a synbiotic on symptoms, and daily functioning in attention deficit hyperactivity disorder – A double-blind randomized controlled trial. Brain Behav Immun. 2020, 89:9-19. [CrossRef]
- Zhao L, Zhang F, Ding X, et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science (1979). 2018, 359, 1151–1156. [CrossRef]
- Wu Z, Xu Q, Wang Q, et al. The impact of dietary fibers on Clostridioides difficile infection in a mouse model. Front Cell Infect Microbiol. 2022, 12. [CrossRef]
- Chen K, Chen H, Faas MM, et al. Specific inulin-type fructan fibers protect against autoimmune diabetes by modulating gut immunity, barrier function, and microbiota homeostasis. Mol Nutr Food Res. 2017, 61, 1601006. [CrossRef]
- Everard A, Belzer C, Geurts L, et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proceedings of the National Academy of Sciences. 2013, 110, 9066–9071. [CrossRef]
- Demirci M, Tokman HB, Uysal HK, et al. Reduced Akkermansia muciniphila and Faecalibacterium prausnitzii levels in the gut microbiota of children with allergic asthma. Allergol Immunopathol (Madr). 2019, 47, 365–371. [CrossRef]
- Wang L, Christophersen CT, Sorich MJ, Gerber JP, Angley MT, Conlon MA. Low relative abundances of the mucolytic bacterium Akkermansia muciniphila and Bifidobacterium spp. in feces of children with autism. Appl Environ Microbiol. 2011, 77, 6718–6721. [CrossRef]
- Klaenhammer TR, Altermann E, Pfeiler E, et al. Functional Genomics of Probiotic Lactobacilli. J Clin Gastroenterol. 2008, 42(September):S160-S162. [CrossRef]
- Maassen CBM, Claassen E. Strain-dependent effects of probiotic lactobacilli on EAE autoimmunity. Vaccine. 2008, 26, 2056–2057. [CrossRef]
- Martín R, Delgado S, Maldonado A, et al. Isolation of lactobacilli from sow milk and evaluation of their probiotic potential. Journal of Dairy Research. 2009, 76, 418–425. [CrossRef]
- Moorthy G, Murali MR, Devaraj SN. Lactobacilli facilitate maintenance of intestinal membrane integrity during Shigella dysenteriae 1 infection in rats. Nutrition. 2009, 25, 350–358. [CrossRef]
- Alghamdi MA, Al-Ayadhi L, Hassan WM, Bhat RS, Alonazi MA, El-Ansary A. Bee Pollen and Probiotics May Alter Brain Neuropeptide Levels in a Rodent Model of Autism Spectrum Disorders. Metabolites. 2022, 12(6). [CrossRef]
- Kantak PA, Bobrow DN, Nyby JG. Obsessive-compulsive-like behaviors in house mice are attenuated by a probiotic (Lactobacillus rhamnosus GG). Behavioural Pharmacology. 2014, 25, 71–79. [CrossRef]
- Sanikhani NS, Modarressi MH, Jafari P, et al. The Effect of Lactobacillus casei Consumption in Improvement of Obsessive–Compulsive Disorder: an Animal Study. Probiotics Antimicrob Proteins. 2020, 12, 1409–1419. [CrossRef]
- Kobliner V, Mumper E, Baker SM. Reduction in obsessive compulsive disorder and self-injurious behavior with saccharomyces boulardii in a child with autism: A case report. Integrative Medicine (Boulder). 2018, 17, 38–41.
- Pochakom A, Mu C, Rho JM, Tompkins TA, Mayengbam S, Shearer J. Selective Probiotic Treatment Positively Modulates the Microbiota-Gut-Brain Axis in the BTBR Mouse Model of Autism. Brain Sci. 2022, 12(6). [CrossRef]
- Sen P, Sherwin E, Sandhu K, et al. The live biotherapeutic Blautia stercoris MRx0006 attenuates social deficits, repetitive behaviour, and anxiety-like behaviour in a mouse model relevant to autism. Brain Behav Immun. 2022, 106:115-126. [CrossRef]
- Skott E, Yang LL, Stiernborg M, et al. Effects of a synbiotic on symptoms, and daily functioning in attention deficit hyperactivity disorder - A double-blind randomized controlled trial. Brain Behav Immun. 2020, 89:9-19. [CrossRef]
- Enrichment of Gut Ecosystem by Daily Supplementation of Selective Probiotic Strains and Probiotic Complex in Dysbiosis Condition of Autism. International Journal of Pharmaceutical Research. 2021, 13(01). [CrossRef]
- Szklany K, Wopereis H, de Waard C, et al. Supplementation of dietary non-digestible oligosaccharides from birth onwards improve social and reduce anxiety-like behaviour in male BALB/c mice. Nutr Neurosci. 2020, 23, 896–910. [CrossRef]
- Tabouy L, Getselter D, Ziv O, et al. Dysbiosis of microbiome and probiotic treatment in a genetic model of autism spectrum disorders. Brain Behav Immun. 2018, 73:310-319. [CrossRef]
- Al-Ali D, Ahmed A, Shafiq A, et al. Fecal microbiota transplants: A review of emerging clinical data on applications, efficacy, and risks (2015-2020). Qatar Med J. 2021, 2021(1). [CrossRef]
- Kang DW, Adams JB, Coleman DM, et al. Long-term benefit of Microbiota Transfer Therapy on autism symptoms and gut microbiota. Sci Rep. 2019, 9, 5821. [CrossRef]
- Ananthaswamy A. Faecal transplant eases symptoms of Parkinson’s disease. New Sci (1956). 2011, 209, 8–9. [CrossRef]
- Ianiro G, Segal JP, Mullish BH, et al. Fecal microbiota transplantation in gastrointestinal and extraintestinal disorders. Future Microbiol. 2020, 15, 1173–1186. [CrossRef]
- Li N, Chen H, Cheng Y, et al. Fecal Microbiota Transplantation Relieves Gastrointestinal and Autism Symptoms by Improving the Gut Microbiota in an Open-Label Study. Front Cell Infect Microbiol. 2021, 11. [CrossRef]
- Wang J, Liu X, Li Q. Interventional strategies for ischemic stroke based on the modulation of the gut microbiota. Front Neurosci. 2023, 17:1158057. [CrossRef]
- Khanna S, Vazquez-Baeza Y, González A, et al. Changes in microbial ecology after fecal microbiota transplantation for recurrent C. difficile infection affected by underlying inflammatory bowel disease. Microbiome. 2017, 5(1). [CrossRef]
- Kang DW, Adams JB, Gregory AC, et al. Microbiota Transfer Therapy alters gut ecosystem and improves gastrointestinal and autism symptoms: An open-label study. Microbiome. 2017, 5, 1–16. [CrossRef]
- Kilinçarslan S. The effect of fecal microbiota transplantation on psychiatric symptoms among patients with inflammatory bowel disease: an experimental study. Actas Esp Psiquiatr. 2020, 48, 1–7.
- Wang J, Cao Y, Hou W, et al. Fecal microbiota transplantation improves VPA-induced ASD mice by modulating the serotonergic and glutamatergic synapse signaling pathways. Transl Psychiatry. 2023, 13(1). [CrossRef]
- Wilkins LJ, Monga M, Miller AW. Defining Dysbiosis for a Cluster of Chronic Diseases. Sci Rep. 2019, 9, 12918. [CrossRef]
- Petersen C, Round JL. Defining dysbiosis and its influence on host immunity and disease. Cell Microbiol. 2014, 16, 1024–1033. [CrossRef]
- Mirsepasi-Lauridsen HC, Vrankx K, Engberg J, et al. Disease-Specific Enteric Microbiome Dysbiosis in Inflammatory Bowel Disease. Front Med (Lausanne). 2018, 5:304. [CrossRef]
- Coker OO, Nakatsu G, Dai RZ, et al. Enteric fungal microbiota dysbiosis and ecological alterations in colorectal cancer. Gut. 2019, 68, 654–662. [CrossRef]
- Li L, Wang F, Liu Y, Gu F. Intestinal microbiota dysbiosis in children with recurrent respiratory tract infections. Microb Pathog. 2019, 136:103709. [CrossRef]
- Hou M, Xu G, Ran M, Luo W, Wang H. APOE-ε4 Carrier Status and Gut Microbiota Dysbiosis in Patients With Alzheimer Disease. Front Neurosci. 2021, 15. [CrossRef]
- Cassani E, Barichella M, Cancello R, et al. Increased urinary indoxyl sulfate (indican): New insights into gut dysbiosis in Parkinson’s disease. Parkinsonism Relat Disord. 2015, 21, 389–393. [CrossRef]
- Wood H. New models show gut–brain transmission of Parkinson disease pathology. Nat Rev Neurol. 2019, 15, 491. [CrossRef]
- CR Martin VOAKEM. The brain-gut-microbiome axis. Cell Mol Gastroenterol Hepatol. 2018, 6:133-148. [CrossRef]
- O’ Mahony SM, Stilling RM, Dinan TG, Cryan JF. The microbiome and childhood diseases: Focus on brain-gut axis. Birth Defects Res C Embryo Today. 2015, 105, 296–313. [CrossRef]
- Kim N, Yun M, Oh YJ, Choi HJ. Mind-altering with the gut: Modulation of the gut-brain axis with probiotics. Journal of Microbiology. 2018, 56, 172–182. [CrossRef]
- Shao T, Hsu R, Hacein-Bey C, et al. The Evolving Landscape of Fecal Microbial Transplantation. Clin Rev Allergy Immunol. Published online 2023. [CrossRef]
- Nicco C, Paule A, Konturek P, Edeas M. From Donor to Patient: Collection, Preparation and Cryopreservation of Fecal Samples for Fecal Microbiota Transplantation. Diseases. 2020, 8, 9. [CrossRef]
- Nøhr MK, Egerod KL, Christiansen SH, et al. Expression of the short chain fatty acid receptor GPR41/FFAR3 in autonomic and somatic sensory ganglia. Neuroscience. 2015, 290:126-137. [CrossRef]
- Singh N, Gurav A, Sivaprakasam S, et al. Activation of Gpr109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis. Immunity. 2014, 40, 128–139. [CrossRef]
- Oleskin A V., Shenderov BA. Neuromodulatory effects and targets of the SCFAs and gasotransmitters produced by the human symbiotic microbiota. Microb Ecol Health Dis. 2016, 27(0). [CrossRef]
- Billeci L, Callara AL, Guiducci L, et al. A randomized controlled trial into the effects of probiotics on electroencephalography in preschoolers with autism. Autism. 2023, 27, 117. [CrossRef]
- Gao XW, Mubasher M, Fang CY, Reifer C, Miller LE. Dose-response efficacy of a proprietary probiotic formula of lactobacillus acidophilus CL1285 and lactobacillus casei LBC80R for antibiotic-associated diarrhea and clostridium difficile-associated diarrhea prophylaxis in adult patients. American Journal of Gastroenterology. 2010, 105, 1636–1641. [CrossRef]
- Ruszczyński M, Radzikowski A, Szajewska H. Clinical trial: effectiveness of Lactobacillus rhamnosus (strains E/N, Oxy and Pen) in the prevention of antibiotic-associated diarrhoea in children. Aliment Pharmacol Ther. 2008, 28, 154–161. [CrossRef]
- Canani RB, Cirillo P, Terrin G, et al. Probiotics for treatment of acute diarrhoea in children: randomised clinical trial of five different preparations. BMJ. 2007, 335, 340. [CrossRef]
- Kruis W, Frič P, Pokrotnieks J, et al. Maintaining remission of ulcerative colitis with the probiotic Escherichia coli Nissle 1917 is as effective as with standard mesalazine. Gut. 2004, 53, 1617–1623. [CrossRef]
- Suez J, Zmora N, Segal E, Elinav E. The pros, cons, and many unknowns of probiotics. Nat Med. 2019, 25, 716–729. [CrossRef]
- Dietrich CG, Kottmann T, Alavi M. Commercially available probiotic drinks containing Lactobacillus casei DN-114001 reduce antibiotic-associated diarrhea. World J Gastroenterol. 2014, 20, 15837–15844. [CrossRef]
- Pereg D, Kimhi O, Tirosh A, Orr N, Kayouf R, Lishner M. The effect of fermented yogurt on the prevention of diarrhea in a healthy adult population. Am J Infect Control. 2005, 33, 122–125. [CrossRef]
- Lichtman JS, Sonnenburg JL, Elias JE. Monitoring host responses to the gut microbiota. ISME J. 2015, 9, 1908–1915. [CrossRef]
- Carmody RN, Gerber GK, Luevano JM, et al. Diet dominates host genotype in shaping the murine gut microbiota. Cell Host Microbe. 2015, 17, 72–84. [CrossRef]
- Gill HS, Rutherfurd KJ, Cross ML. Dietary probiotic supplementation enhances natural killer cell activity in the elderly: An investigation of age-related immunological changes. J Clin Immunol. 2001, 21, 264–271. [CrossRef]
- Chuong KH, Mack DR, Stintzi A, O’Doherty KC. Human microbiome and learning healthcare systems: Integrating research and precision medicine for inflammatory bowel disease. OMICS. 2018, 22, 119–126. [CrossRef]
- Han S, Lu Y, Xie J, et al. Probiotic Gastrointestinal Transit and Colonization After Oral Administration: A Long Journey. Front Cell Infect Microbiol. 2021, 11. [CrossRef]
- Pelto L, Isolauri E, Lillus EM, Nuutila J, Salminen S. Probiotic bacteria down-regulate the milk-induced inflammatory response in milk-hypersensitive subjects but have an immunostimulatory effect in healthy subjects. Clinical & Experimental Allergy. 1998, 28, 1474–1479. [CrossRef]
- Roessler A, Friedrich U, Vogelsang H, et al. The immune system in healthy adults and patients with atopic dermatitis seems to be affected differently by a probiotic intervention. Clin Exp Allergy. 2008, 38, 93–102. [CrossRef]
- Zmora N, Zilberman-Schapira G, Suez J, et al. Personalized Gut Mucosal Colonization Resistance to Empiric Probiotics Is Associated with Unique Host and Microbiome Features. Cell. 2018, 174, 1388–1405.e21. [CrossRef]
- Hod K, Dekel R, Aviv Cohen N, et al. The effect of a multispecies probiotic on microbiota composition in a clinical trial of patients with diarrhea-predominant irritable bowel syndrome. Neurogastroenterol Motil. 2018, 30(12). [CrossRef]
- Suwal S, Wu Q, Liu W, et al. The probiotic effectiveness in preventing experimental colitis is correlated with host gut microbiota. Front Microbiol. 2018, 9(NOV):2675. [CrossRef]
- Abildgaard A, Kern T, Pedersen O, Hansen T, Wegener G, Lund S. The antidepressant-like effect of probiotics and their faecal abundance may be modulated by the cohabiting gut microbiota in rats. Eur Neuropsychopharmacol. 2019, 29, 98–110. [CrossRef]
- Ferrario C, Taverniti V, Milani C, et al. Modulation of Fecal Clostridiales Bacteria and Butyrate by Probiotic Intervention with Lactobacillus paracasei DG Varies among Healthy Adults. J Nutr. 2014, 144, 1787–1796. [CrossRef]
- Johnson KVA. Gut microbiome composition and diversity are related to human personality traits. Hum Microb J. 2020, 15:None. [CrossRef]
- He F, Ouwehand AC, Isolauri E, Hosoda M, Benno Y, Salminen S. Differences in composition and mucosal adhesion of bifidobacteria isolated from healthy adults and healthy seniors. Curr Microbiol. 2001, 43, 351–354. [CrossRef]
- Andriantsoanirina V, Teolis AC, Xin LX, Butel MJ, Aires J. Bifidobacterium longum and Bifidobacterium breve isolates from preterm and full term neonates: Comparison of cell surface properties. Anaerobe. 2014, 28:212-215. [CrossRef]
- Aceti A, Maggio L, Beghetti I, et al. Probiotics Prevent Late-Onset Sepsis in Human Milk-Fed, Very Low Birth Weight Preterm Infants: Systematic Review and Meta-Analysis. Nutrients. 2017, 9, 904. [CrossRef]
- He L, Chen R, Zhang B, et al. Fecal microbiota transplantation treatment of autoimmune-mediated type 1 diabetes mellitus. Front Immunol. 2022, 13. [CrossRef]
- Biazzo M, Deidda G. Fecal Microbiota Transplantation as New Therapeutic Avenue for Human Diseases. J Clin Med. 2022, 11(14). [CrossRef]
- Kang DW, Adams JB, Gregory AC, et al. Microbiota Transfer Therapy alters gut ecosystem and improves gastrointestinal and autism symptoms: An open-label study. Microbiome. 2017, 5(1). [CrossRef]
- Goodswen SJ, Barratt JLN, Kennedy PJ, Kaufer A, Calarco L, Ellis JT. Machine learning and applications in microbiology. FEMS Microbiol Rev. 2021, 45, 1–19. [CrossRef]
Figure 1.
Mechanisms of the MGBA.
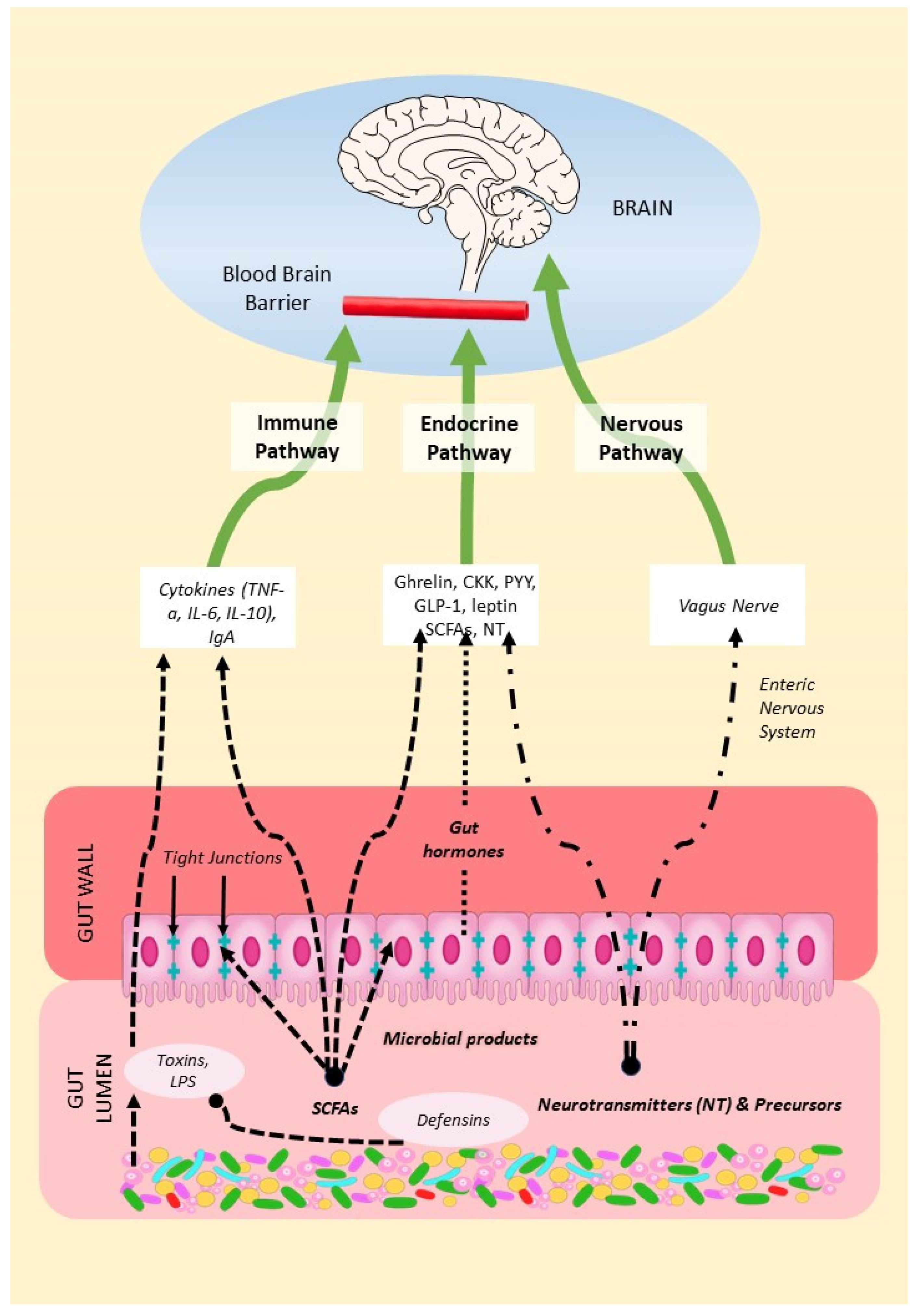
Figure 2.
Model of the microbiological basis of OCD.
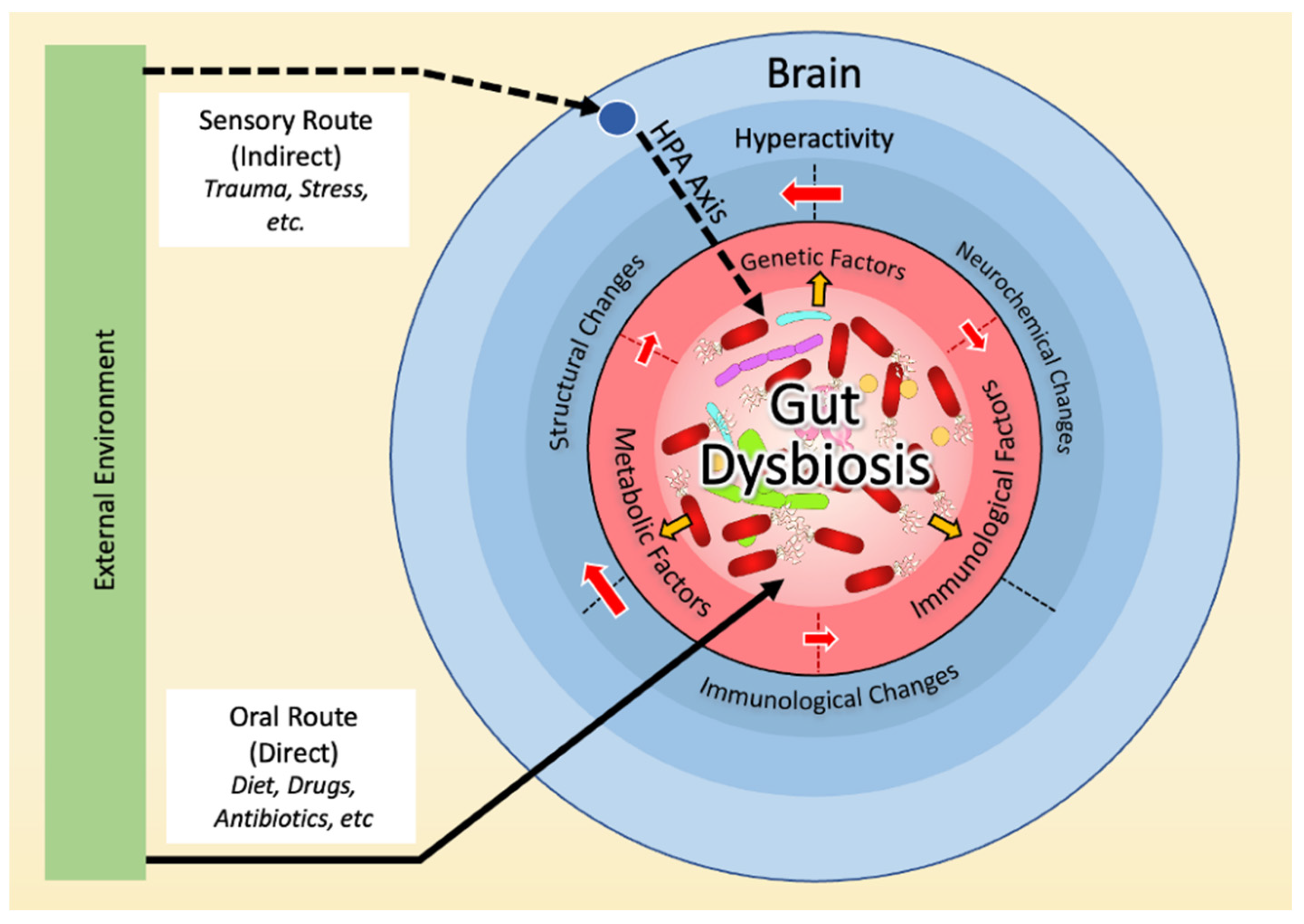
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



