Submitted:
07 June 2023
Posted:
07 June 2023
You are already at the latest version
Abstract
Ovarian tissue cryopreservation (OTC) or testicular tissue cryopreservation (TTC) are effective and often the only options for fertility preservation in female or male patients due to oncological, medical, or social aspects. While TTC and resumption of spermatogenesis, either in vivo or in vitro, has still be considered an experimental approach in humans, OTC and autotransplantation has been applied increasingly to preserve fertility with more than 200 live births worldwide. However, the cryopreservation of reproductive cells followed by the resumption of gametogenesis, either in vivo or in vitro, may interfere with sensitive and highly regulated cellular processes. In particular, the epigenetic profile, which includes not just reversible modifications of the DNA itself but also post-translational histone modifications, small non-coding RNAs, gene expression and availability, and storage of related proteins or transcripts, have to be considered in this context. Due to complex reprogramming and maintenance mechanisms of the epigenome in germ cells, growing embryos, and offspring, OTC and TTC are carried out at very critical moments early in the life cycle. Given this background, the safety of OTC and TTC taking into account the epigenetic profile has to be clarified. Cryopreservation of mature germ cells (including Metaphase II oocytes and mature spermatozoa collected via ejaculation or more invasively after testicular biopsy) or embryos has been used successfully for many years in Medically Assisted Reproduction (MAR). However, tissue freezing followed by in vitro or in vivo gametogenesis has become more attractive in the past, while few human studies have analysed the epigenetic effects, with most data deriving from animal studies. In this review, we highlight the potential influence of the cryopreservation of immature germ cells and subsequent in vivo or in vitro growth and differentiation on the epigenetic profile in humans and animals.
Keywords:
epigenetic
; genomic imprinting
; ovarian tissue cryopreservation
; testicular tissue cryopreservation
; Medically Assisted Reproduction
1. The Epigenome
Exactly 70 years ago, DNA was described as the universal carrier of genetic information through the pioneering work of Francis Crick, James Watson, and Rosalind Franklin. However, gene expression relies on more than just the nucleobase sequence; epigenetic factors control cellular functions at a superimposed level without affecting the genetic code itself. The epigenome includes reversible DNA methylation, post-translational histone modifications (PTMs), and the abundance and availability of relevant transcripts and proteins to establish or maintain DNA methylation (Figure 1). It is not surprising that epigenetic control is crucial for cell and tissue differentiation, reaction to exogenous and endogenous influences, sex chromosome dosage compensation, or fertilisation and embryogenesis [1]. Epigenetic modifications are not rigid during the entire life cycle, rather they are subjected to dynamic reprogramming. In brief, early in life, the entire somatic DNA methylation pattern is erased in primordial germ cells, with subsequent de novo establishment of the germ cell-specific DNA methylation profile. In males, global DNA methylation (gDNA) is completed in pachytene-stage spermatocytes; in females, gDNA is established in mature post-menarche oocytes [2,3]. To establish totipotency during early embryogenesis, gDNA demethylation occurs actively in paternally inherited DNA and passively in maternally inherited DNA in the zygote and growing embryo at each cell division (Figure 2). Later, during embryogenesis, cell and tissue-specific de novo gDNA reprogramming is established.
Imprinted genes represent unique DNA sequences within the genome. Genomic imprinting occurs oppositely in differentially methylated regions (DMR) in male and female germ cells, leading to mono-allelic gene expression after syngamy. This evolutionarily conserved mechanism is essential for metabolic function and embryonic, placental, and postnatal development by expressing one parental allele and inactivating the other parental allele. De novo establishment of parental-specific DNA methylation takes place during gametogenesis in sperm and oocytes at specific CpG (Cytosine guanine dinucleotide) sites; however, conversely to gDNA methylation, the methylation pattern of imprinted genes is maintained after fertilisation and embryogenesis. The progressive establishment of characteristic DNA methylation during gametogenesis depends on the de novo methyltransferases DNMT3A and/or DNMT3B/DNMT3L [4]. Several of the 200 known imprinted genes are organised into imprinted control regions (ICR). After oocyte-to-embryo transition, it is essential to distinguish between gDNA methylation, which becomes erased, and imprinted genes, where the methylation pattern is stable (Figure 2). To this end, a complex machinery is required to protect imprinted genes from demethylation in the early stages of life, which includes Maintenance DNA methyltransferase 1 (DNMT1), zinc finger protein 57 (ZFP57), Mater protein homolog (MATER), or STELLA. Many of these factors belong to the group of maternal effect genes (MEG) and are often stored in distinct compartments like the subcortical maternal complex (SCMC). The accumulation of these factors during gametogenesis and availability after fertilisation is essential for epigenetic control [2]. It is also noteworthy that post-translational histone modifications, mainly by acetylation, methylation, ubiquitylation of specific histone residues, or specific small non-coding RNAs are also involved in this highly coordinated orchestra of epigenetic regulation [5,6]. Thus, epigenetics is much more complex than simple DNA methylation itself, involving a dense network at different cellular levels. Disturbances due to cryopreservation or in vitro treatment may, therefore, interfere with the correct establishment and maintenance of the epigenome. To estimate any potential adverse effects of OTC or TTC, we must consider not just the methylation landscape but also factors located up- and downstream of DNA methylation.
2. Imprinting disorders
A dozen clinical syndromes are known to be caused by imprinting disorders (e.g., Beckwith-Wiedemann syndrome (BWS), Silver-Russell syndrome (SRS), Prader-Willi syndrome (PWS), transient neonatal diabetes mellitus, recurrent miscarriages, and hydatidiform moles). For instance, total or partial loss of maternal methylation at the SNURF-SNRPN imprinting centre can lead to Angelman syndrome, maternal ICR1 hypermethylation to Beckwith-Wiedemann syndrome, and paternal loss of ICR1 methylation to Silver-Russell syndrome [7,8,9]. Aberrant methylation patterns can derive from genomic errors (e.g., DNA mutation/deletion/duplication) or epimutations themselves (e.g., gain/loss of DNA methylation patterns). As mentioned above, DNA methylation is embedded in a complex cellular network to establish stage-specific DNA methylation or to protect imprinted genes from demethylation. Therefore, genomic-based errors often lead to multi-locus imprinting disturbances (MLID), whereby one factor controls and regulates a broad spectrum of differently methylated regions. A genomic mutation in the ZFP57 gene results in abnormal DNA methylation, as seen in patients with neonatal diabetes mellitus; mutations in the NLRP gene family (which includes MATER and NLRP7) result not only in miscarriages and hydatidiform moles but also imprinting disorders such as Beckwith-Wiedemann syndrome or Silver-Russell syndrome—including loss of DNA methylation in several DMRs [10,11,12,13].
Epigenetic DNA methylation is not only influenced indirectly by genomic errors, like altered MEGs or components of the SCMC, but also directly by environmental stressors. Environmental exposure such as in vitro culture, ovarian stimulation, or cryopreservation in early development might induce epimutations, as reported in different species [13,14,15,16,17]. Epimutations were not only reported in vitrified oocytes or embryos but also in later foetuses and placentas derived from cryopreserved embryos [17,18,19]. In humans, several studies also reported an increased risk of rare genomic imprinting disorders in children conceived via MAR techniques, including BWS, AS, PWS, and SRS [16,20,21]. Nevertheless, it is still unclear whether MAR techniques per se or the impaired fertility background of the parents contributed to these rare genomic disorders in MAR children; however, epigenome alterations due to MAR treatment could be a possible mechanism.
3. Ovarian tissue cryopreservation (OTC)
Cryopreservation of mature metaphase II (MII) oocytes, zygotes, or embryos is the common method to safeguard fertility in female patients after controlled ovarian stimulation and oocyte pick-up in MAR. Fertility preservation is primarily indicated in patients undergoing gonadotoxic radiotherapy or chemotherapy treatment, while other medical or social reasons (e.g., individual life planning) may also be of relevance. However, for women undergoing oncological treatment that cannot be delayed or prepubertal girls with no possibility of obtaining mature germ cells, OTC is the only option to preserve fertility. OTC can also be offered to female patients with benign ovarian diseases requiring ovariectomy and conditions with an increased risk of premature ovarian failure.
Ovarian tissue cryopreservation and transplantation were first performed in animal model systems several decades ago [22], while the first human live birth was reported in 2004 by Donnez and colleagues after controlled tissue freezing [23]. To date, more than 200 babies have been born worldwide after OTC. Before gonadotoxic cancer treatment or premature loss of the ovarian reserve, the ovarian cortex with dormant immature follicles and germinal vesicle (GV) oocytes of one or both ovaries are separated into small pieces of approximately 5x5 mm with a thickness of 1-2 mm prior to cryopreservation [24]. OTC can be performed either by slow-freezing protocols or an ultra-fast vitrification technique. Slow-freezing protocols are commonly used for OTC and nearly all live births reported used this technique. Nevertheless, in recent years, vitrification has also become a very promising approach for OTC, with just a few live births reported after vitrification and warming of ovarian tissue [25,26]. Both techniques require cryoprotective agents (CPA) to avoid the uncontrolled and detrimental formation of intracellular ice crystals during the cryopreservation process. Dimethyl sulfoxide (DMSO), glycerol, propanediol, or ethylene glycol are used as permeating agents and nonpermeating agents like proteins, sugars, and other macromolecules are used to achieve controlled cell dehydration and cellular CPA uptake. In both techniques, an optimised balance between CPA concentration and exposure time is necessary to minimise potential cytotoxic CPA stress. Slow-freezing protocols typically use low CPA concentrations combined with a long exposure time during controlled freezing, while CPA exposure during vitrification is much shorter. Colligative combinations of two or more CPAs are used for vitrification, however, significantly higher CPA concentrations are necessary to prevent harmful ice crystal formation during the ultra-fast transition into a glass-like amorphous solid [27]. CPAs are known to have toxic effects on cellular structures and functions, and exposure to DMSO can lead to epimutations in different cell types, including DNA hypo- or hypermethylation, alterations in post-translational histone modifications, or misregulated expression of epigenetically relevant DNA methyltransferases [28,29,30].
Following cryopreservation, CPAs are removed after thawing/warming for slow freezing as well as for vitrification. It should be noted that, due to the higher CPA concentrations needed for vitrification, residual CPAs are higher in ovarian tissue after vitrification/warming compared with slow freezing and thawing [31]. Thus, ovarian tissue vitrification might pose a higher risk of cytotoxic CPA exposure compared with slow-freezing protocols.
Several in vivo or in vitro options are available to resume folliculogenesis and oogenesis after OTC, with ovarian tissue autotransplantation still being the method of choice (Figure 3). After orthotopic or heterotopic transplantation, endogenous functionality is commonly restored after up to six months and resumption of folliculogenesis and oogenesis can occur in vivo. Mature MII oocytes are available for fertilisation either spontaneously (orthotopic autotransplantation) or via MAR techniques (heterotopic autotransplantation). Besides in vivo strategies, in some cases, in vitro treatment is necessary to avoid the risk of remission of malignant cells or to enhance the activation of resting follicles after ovarian tissue autotransplantation. In vitro culture of immature follicles (IVC), in vitro maturation of immature oocytes (IVM), artificial ovaries including isolated follicles enclosed in extracellular matrix tissue and in vivo growth (AO), or in vitro activation of immature follicles (IVA) prior to autotransplantation afford new strategies to preserve female fertility [32]. Most human in vitro techniques are still experimental, however, live births have been reported after IVA in patients with idiopathic primary ovarian insufficiency [25]. In vitro follicle culture systems are options when tissue autotransplantation is contraindicated. Healthy live births were reported in animal models after IVC of immature follicles several years ago [33]; however, human IVC is still challenging due to different culture conditions including growth factors and hormones, or simply due to unequal follicle sizes between species [34]. Nevertheless, human IVC is a promising option and progress was reported by McLaughlin and colleagues [35]. Here, fertilisable human MII oocytes were obtained after complete in vitro growth and maturation. The translation of these techniques from animal models to humans is ambitious and further research is needed, however, these studies are also indispensable to understanding the direct and long-term effects of the cryopreservation of immature germ cells combined with interrupted gametogenesis.
Overall, OTC and subsequent folliculogenesis and oogenesis can preserve fertility in women facing gonadotoxic treatment, premature loss of the ovarian reserve due to benign conditions, or in prepubertal girls, regardless of the cryopreservation protocols applied. Autotransplantation of ovarian tissue is commonly performed; however, in vitro techniques are promising, particularly when tissue autotransplantation is contraindicated. In each case, folliculogenesis and oogenesis are interrupted and long-term effects on the epigenome cannot be excluded, which could be due to extracorporeal treatment, exposure to potentially harmful substances, or the cryopreservation treatment itself.
4. OTC and the Epigenome
The cryopreservation of mature oocytes or pre-implantation embryos, mostly via vitrification, is a very common and routinely applied MAR technique in humans. Survival and clinical outcomes are good, however, the adverse effects on epigenetic patterns are still of concern as several epigenome alterations have been reported in recent years, especially in animal models but also in humans [14,15,17,36,37,38]. To estimate the potential effect of the cryopreservation of immature (and not fully grown) oocytes, different phases must be considered: i) immediate effects of cryopreservation directly after treatment, ii) mid-term effects during in vivo/in vitro growth, and iii) long-term effects in embryos/offspring. Moreover, since most cellular factors required for oocyte-to-embryo transition rely on maternal accumulation and storage in distinct compartments, like the SCMC during oocyte growth, not just i) DNA methylation patterns alone but also ii) the abundance and availability of epigenome-related proteins and transcripts, or iii) post-translational histone modifications are of relevance.
Gene expression and protein abundance
In general, cryopreservation of ovarian tissue can result in changes in gene expression and protein abundance at different stages [39,40]. More precisely, cryopreservation also hampers several specific epigenetically relevant factors. In mice, vitrification and warming of juvenile ovaries resulted in significantly reduced mRNA expression and protein abundance of DNMT1 [41]. Vitrification of fully grown immature ovine GV oocytes induced alterations in a subset of genes implicated in epigenetic control during oocyte maturation and early embryo development. These included DNA methyltransferase DNMT3B and histone deacetylase HDAC1 [42]. Slow freezing or vitrification of murine ovarian tissue followed by orthotopic transplantation also led to gene expression differences in the imprinted genes Igf2r, H19, and PLAGL1 in various tissue types in the offspring compared with natural controls [43]. In contrast, after vitrification of immature pre-antral follicles and subsequent in vitro follicle culture, the abundance of nearly 2000 proteins is similar between vitrified and non-vitrified controls in fully grown GV or MII oocytes [44]. This dataset also included several MEGs including MATER, Histone acetyltransferase Hat1, and DNMT1. As a consequence, the cellular machinery establishing epigenetic patterns after vitrification of immature GV oocytes followed by in vitro growth might not be altered (or is even restored), although it is not representative of the entire proteome in mature oocytes [45]. Compared to the entire proteome, effects on lower-abundance proteins essential for epigenetic control cannot be excluded.
DNA methylation patterns
The immediate effects of cryopreservation treatment without subsequent grafting or IVC were reported after vitrification and warming of juvenile murine ovaries. Here, the Growth factor receptor-binding protein 10 (Grb10) promoter was hypermethylated compared with controls [41]. Similarly, 16 different single CpGs in the Snrpn DMR were analysed in GV oocytes from juvenile murine ovaries directly after cryopreservation. The Snrpn methylation status in vitrified/warmed GV oocytes did not vary from fresh controls [46]. Although allotransplantation was carried out afterwards, no epigenetic data regarding pre-implantation embryos or offspring were available. Certainly, the cryopreservation of fully grown GV oocytes followed by IVM is quite different to classic OTC approaches, especially as regards the unequal epigenetic status of early-stage GV and fully grown GV oocytes; however, freezing of GV oocytes might, in some cases, also be an option for fertility patients. Moreover, due to comparable chromatin organisation in early and fully grown GV oocytes (chromatin vs. condensed chromosomes), epigenetic effects after GV cryopreservation followed by IVM might be relevant at this point. Therefore, in brief, normal gDNA methylation patterns were observed in mouse MII oocytes after GV cryopreservation/IVM [47] and in the human imprinted genes H19 and KCNQ1OT1 [48]. In bovines, gDNA methylation was not affected after GV vitrification and IVM in MII oocytes, yet it was significantly reduced in pre-implantation embryos compared with fresh controls [49].
Some studies have been carried out in animal model systems concerning the mid-term effects during in vivo/in vitro growth or long-term effects in embryos/offspring. In a setup using vitrified and IVC murine pre-antral follicles, H19, Igf2r, and Snrpn imprinting patterns were analysed in fully grown GV oocytes. H19 and Igf2r DNA methylation was comparable between in vivo/in vitro controls and the vitrified group, while some single CpG errors were reported in the maternally imprinted Snrpn gene [50]. Although indirect, different methylation patterns for Inhba and Inhbb were reported in mice after vitrification and IVC in granulosa cells from large antral follicles [51]. In the pioneering study of Sauvat et al., [52], two epigenetic marks in murine offspring were analysed for the first time after cryopreservation and grafting of immature tissue. No differences in the imprinted genes H19 and Lit1 were found in muscle, kidney, and tongue in offspring from grafted mice compared to controls. Similarly, a normal Igf2r methylation status was reported in ovine offspring after grafting of cryopreserved immature ovaries [53]. Controversially, in a recent study by Yan and colleagues [43], the methylation rates of four imprinted genes were analysed after slow freezing or vitrification of murine ovarian tissue followed by orthotopic transplantation. While the methylation pattern was stable in Snrpn, alterations were reported in Igf2r, H19, and PLAGL1 in brain and liver tissue in the offspring compared with natural controls. These alterations were combined with different gene expression levels for Igf2r, H19, and PLAGL1, yet with no significant morphological/functional differences (e.g., birth defects, body weight gain, exercise capacity, or anti-fatigue ability) in offspring derived from either the cryopreserved or non-cryopreserved group.
Post-translational histone modifications
Post-translational histone modifications also occur in a stage-specific manner and are essential for gene activation/silencing not just due to their ability to act as mediators between the enzymatic machinery and DNA itself. For instance, H3K4me3 is commonly associated with gene expression and cell differentiation, while H3K9me3 refers to heterochromatic DNA. Thus, PTMs are also crucial for epigenetic control and direct or indirect alterations of the histone landscape after cryopreservation could be of interest [54,55,56]. Data regarding OTC and complete in vitro/in vivo resumption of gametogenesis is scarce. In one study, Tian et al., reported that H3K9me3, H3K4me3, and H3K27ac levels in murine pre-implantation embryos after cryopreservation and in vitro folliculogenesis were comparable to fresh controls [57]. When analysing PTMs after cryopreservation of immature GV, post-translational histone modifications (H3K9me3), either at the MII stage or in blastocysts, were not different after vitrification of immature bovine GV oocytes followed by in vitro maturation compared with fresh controls [49]. In contrast, in another study by Lee and Comizzoli [58], histone H3 trimethylation at lysine 4 (H3K4me3) was dramatically reduced after vitrification of immature GV oocytes in domestic cats compared with fresh controls, while H3K9me3 levels were unaffected.

Table 1.
Assessment of epigenome-related effects after OTC.
| Reference | Type/Species | Analysis | Main Outcome |
|---|---|---|---|
| Shirazi et al. (2016) [42] | Ovine GV oocytes, IVM | Epigenetically-relevant mRNA abundance in GV/embryos | Alteration of DNMT3B and HDAC1 |
| Demant et al. (2012) [44] | Murine pre-antral follicles, IVC | Proteome analysis in GV/MII | No differences between vitrified and non-vitrified GV/MII |
| He at al. (2018) [41] | Murine OTC | mRNA expression and protein abundance | Decreased mRNA/ protein levels for Dnmt1 |
| Yan et al. (2020) [43] | Murine OTC and orthotopic transplantation | Epigenetically relevant mRNA abundance in offspring | mRNA differences in H19, Igf2r and PLAGL1 but normal Snrpn expression |
| Yodrug et al. (2021) [49] | Bovine GV oocytes, IVM | Global DNA methylation in MII and embryos | Normal gDNA pattern in MII but altered in blastocysts |
| Al-Khtib et al. (2011) [48] | Human GV oocytes, IVM | Imprinted genes in MII | Normal pattern for H19 and KCNQ1OT1 |
| Yan et al. (2014) [47] | Murine GV oocytes, IVM | Global DNA methylation in MII | Normal gDNA pattern |
| Trapphoff et al. (2010) [50] | Murine pre-antral follicles, IVC | Imprinted genes in GV | Normal establishment of H19 and Igf2r imprinting but some single CpG errors in Snrpn |
| Yan et al. (2020) [43] | Murine OTC and orthotopic transplantation | Imprinted genes in offspring | Significant variations in H19, Igf2r, and PLAGL1 but normal Snrpn methylation |
| He et al. (2018) [41] | Murine OTC | Methylation pattern | Hypermethylation of the Grb10 promoter region |
| Wang et al. (2013) [46] | Murine OTC | Methylation pattern in GV after vitrification/warming | Normal Snrpn methylation |
| Sauvat et al. (2008) [52] | Murine OTC and grafting | Imprinted genes in offspring | Normal H19 and Lit1 methylation |
| Sauvat et al. (2013) [53] | OTC and grafting in ewes | Imprinted gene in offspring | Normal Igf2r methylation |
| Damavandi et al. (2021) [51] | Murine pre-antral follicles, IVC | CpG methylation in granulosa cells | Altered Inhba/Inhbb methylation |
| Yodrug et al. (2021) [49] | Bovine GV oocytes, IVM | Histone modifications | Normal H3K9me pattern in MII/blastocysts |
| Tian et al. (2022) [57] | Murine pre-antral follicles, IVC | Histone modifications in embryos | Normal histone pattern (H3K9me3, H3K4me3, H3K27ac) |
| Lee and Comizzoli (2019) [58] | Domestic cat GV | Histone modifications after vitrification | Normal H3K9me3 but altered H3K4me3 |
5. Conclusion - OTC and the Epigenome
Overall, data regarding epigenome patterns at different levels (imprinted genes, gDNA methylation, PTMs, or abundance of relevant transcripts and proteins) after OTC are controversial. While the epigenome, in most studies, presented (at best) no or only minor alterations, some studies revealed imprinting defects in pre-implantation embryos or offspring that could lead to detrimental imprinting disorders. This has to be taken into account, especially since several studies in humans have reported an increased risk of rare genomic imprinting disorders in children conceived by MAR techniques [16,20,21]. The cryopreservation of mature MII oocytes or pre-implantation embryos can lead to epimutation, as seen in several studies; this effect can certainly also be transferred to immature oocytes, as highlighted herein. OTC affords the possibility of restoring initial (or transient) epimutations after subsequent growth and differentiation, however, long-term effects cannot be excluded. Additionally, it has to be considered that epigenetic control is normally not an all-or-nothing mechanism. Single CpG errors or (slightly) reduced DNA methylation patterns may not ultimately lead to imprinting disorders as there is a great distance between (epigenetic) genotype and (functional) phenotype. As for OTC, differences between studies might be due to different cell stages (prepubertal vs. adult) or cell composition (isolated follicles vs. tissue), different species, or methods to resume gametogenesis (in vitro vs. in vivo), different cryopreservation protocols (vitrification vs. slow freezing), or the sensitivity of the analysis tools (bisulphite treatment and pyrosequencing vs. Southern blotting). Studies addressing the question of OTC and imprinting disorders after cryopreservation are rare, particularly in humans, and further research are needed to exclude any potential long-lasting adverse effects derived from cryopreservation techniques.
6. Testicular tissue cryopreservation (TTC)
In males, the option of choice for fertility preservation is the cryopreservation of mature spermatozoa. Sperm can be collected directly via ejaculation or more invasively after testicular biopsy. Later, mature sperms can be used for different MAR techniques including in vitro fertilisation (IVF), intracytoplasmic sperm injection (ICSI), or, after testicular biopsy, via testicular sperm extraction followed by intracytoplasmic sperm injection (TESE-ICSI). Freezing/thawing of mature sperms has been used successfully in MAR (including IVF, ICSI, TESE-ICSI) for many decades [59]. However, mature spermatozoa can only be obtained after the onset of final spermatogenesis. In prepubertal cancer patients without active spermatogenesis, testicular tissue cryopreservation is the only option currently available prior to gonadotoxic radiotherapy or chemotherapy treatment. TTC offers the possibility of preserving spermatogonial stem cells (SSC) and resumption of gametogenesis, either in vivo or in vitro, for later treatment [60]. Males without active spermatogenesis have been offered testicular tissue banking for more than 20 years to potentially restore fertility after successful treatment (or provide the option to do so in the future). Comparable to OTC, the resumption of spermatogenesis can either occur in vivo after autotransplantation of thawed testicular tissue or isolated SSCs, or in vitro under appropriate culture conditions (Figure 4) [61,62,63]. Transplantation or culture of isolated SSC could avoid the remission of malignant cells and would be the better option for patients with metastatic malignancies or haematological cancer. Moreover, spermatogonial stem cell autotransplantation (SSCT) allows (so far only in animal model systems) the restoration of spermatogenesis in vivo (including natural conception without requiring MAR techniques); however, additional in vitro propagation is necessary to increase cell numbers. In contrast to tissue grafting, SSC isolation requires enzymatic digestion by collagenase and trypsin treatment or mechanical disaggregation; enzymatic treatment in particular might interfere with susceptibility to the cryopreservation process and cell viability [60,64].
Cryopreservation can be carried out via different protocols, including slow freezing or vitrification (or variants thereof), however, these protocols are still under development to optimise outcomes. Accordingly, in a mouse model, cryopreservation of testicular tissue might be more effective than testicular cell suspension cryopreservation [65]. Overall, the resumption of spermatogenesis in humans (and undoubtedly also in non-human model systems) is still challenging and viable offspring have so far only been reported in animal model systems [66,67,68].
7. TTC and the Epigenome
Compared with OTC, TTC and resumption of gametogenesis, either in vivo or in vitro, is still in the developmental phase and current research is mainly focused on the generation of healthy offspring (or the way towards it), mainly via fresh tissue or cells. Great progress has been made over the last two decades, yet studies including fresh/frozen and in vivo controls are still scarce and the assessment of the effects of cryopreservation might, at best, only constitute the next step. The need for proper controls is underlined by the fact that extracorporeal in vitro SSC propagation itself might induce epimutations [69]. Epigenetic instability was shown in several imprinted genes after cryopreservation of human testicular tissue and long-term IVC. Demethylation of paternally imprinted genes (H19, H19-DMR, and MEG3) along with increased methylation of maternally imprinted genes (PEG3 and KCNQ1OT) were found during in vitro SSC culture. Controversially, a stable epigenetic profile was reported in mouse and marmoset SSC cultures without cryopreservation [70,71]. Whether epigenetic instability relies on cryopreservation or species differences remains unclear. Data regarding epigenetic effects after TTC must be considered with caution unless suitable controls are performed to distinguish between de novo epimutations due to cryopreservation or limited artificial spermatogenesis.
In recent years, proof-of-principle regarding tissue/SSC transplantation was demonstrated for different techniques. Live offspring were reported after SSC autotransplantation and grafting or IVC of prepubertal testis in different species [66,67,72,73]. A normal DNA methylation pattern in the offspring was reported in mice after IVC of testicular tissue [73,74] or SSCT with fresh samples [67,75]. The more recent study Serrano et al., found no major DNA methylation differences between SSCT-derived offspring and their corresponding controls.
Since healthy mouse offspring with normal methylation patterns in several imprinted genes were produced after testicular tissue cryopreservation, thawing, culture, and fertilisation in vitro, one could speculate that cryopreservation of testicular tissue does not induce de novo epimutations per se [73,74]. This might be true for a single biological endpoint, however, epigenomic control is much more complex, being embedded in a dense epigenetic network and (transient) alterations in intermediate stages cannot be excluded. So far, to the best of our knowledge, only two studies have assessed the direct influence of cryopreservation on the epigenetic profile (or factors related to it). Oblette et al., [76] cultured fresh and slow-frozen testicular tissue in vitro and compared them to in vivo controls. Cryopreservation limited the spermatogenesis and fertilisation capacity, yet embryonic development was initiated after intracytoplasmic sperm injection. Post-translational histone modifications (H3K4me3, H3K27me3, H3K9ac) in embryos were comparable between spermatozoa generated in vitro and in vivo, while DNA differences in gDNA methylation were found after in vitro spermatogenesis (with or without cryopreservation) during early embryogenesis. In another study from the same group, different cryopreservation protocols (controlled slow freezing vs. solid surface vitrification) were also compared after IVC of fresh or frozen mouse prepubertal testes [77]. Relative mRNA levels of epigenetically relevant enzymes, including the DNA methyltransferases DNMT1 and DMT3A and several post-translational histone modifications (H3K4me3, H3K9ac, and H4K8ac), were altered compared with unfrozen or in vivo controls, while gDNA methylation was comparable in spermatozoa after cryopreservation. Unfortunately, the methylation pattern analysis of two imprinted genes (H19 and Igf2r) was unsuccessful due to limited cell numbers. To date, there is no direct data regarding methylation patterns of imprinted genes following cryopreservation. However, cryopreservation might, therefore, pose a risk of inducing de novo epimutations; at which level, especially in later stages, remains unclear.
It is noteworthy that common TTC cryopreservation protocols include DMSO as the main CPA [64]. As seen in different cells types, DMSO treatment can induce de novo epimutations, including misregulated expression of epigenetically relevant DNA methyltransferases, DNA hypo- or hypermethylation, or alterations in affected post-translational histone modifications [28,29,30,78]. Accordingly, cryopreservation of mature spermatozoa can lead to epimutations in Igf2 in boar [79]; yet, to date, no effects have been reported in humans in Snrpn, Snurf, Ebe3a, or H19 [80,81]. Nevertheless, TTC might also require enzymatic digestion via trypsin/collagenase or mechanical disaggregation for SSC isolation, and this enzymatic treatment may also increase/potentiate susceptibility to the cryopreservation process of immature germ cells, including epigenetic alterations as seen after cryopreservation of mature spermatozoa in boar.
Table 2.
Assessment of epigenome-related effects after TTC.
| Reference | Type/Species | Analysis | Main Outcome |
|---|---|---|---|
| Oblette et al. (2019) [77] | Murine in vitro culture of TT | Testicular tissue after IVC | Normal expression of epigenetic modification enzymes and gDNA methylation, but differences in histone modification |
| Oblette et al. (2021) [76] | Murine in vitro culture of TT | Pre-implantation embryo | Normal post-translational histone modifications and altered gDNA methylation |
8. Conclusion - TTC and the Epigenome
Testicular tissue transplantation affords the possibility of restoring fertility and producing healthy offspring in model systems. No major epigenetic alterations in offspring were reported, although minor changes were present. Artificial spermatogenesis per se could pose a risk of epimutations, even without cryopreservation, thus making the estimation of environmental effects caused by cryopreservation on the epigenome quite difficult. Few studies in the literature have addressed whether the effects of enzymatic treatment, CPA exposure, processing for cryopreservation treatment, or cryopreservation itself interfere with the epigenome. Since TTC is still an experimental approach, it is evident that further research is needed to assess the potential, long-lasting, adverse effects of TTC cryopreservation per se before the transition to human clinical trials can occur.
Author Contributions
Conceptualization, T.T. and S.D.; writing—original draft preparation, T.T, S.D.; writing—review and editing, T.T, S.D.; funding acquisition, S.D. All authors have read and agreed to the published version of the manuscript.
Funding
This project was funded by Theramex Germany GmbH.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
The authors would like to thank Nele Ullenbroeck for creating the illustrations.
Conflicts of Interest
The authors declare that this study received funding from Theramex Germany GmbH. The funder was not involved in the study design, collection, analysis, interpretation of data, the writing of this article or the decision to submit it for publication.
References
- Tucci, V.; Isles, A.R.; Kelsey, G.; Ferguson-Smith, A.C. Genomic Imprinting and Physiological Processes in Mammals. Cell 2019, 176, 952–965. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Gao, Z.; Qin, D.; Li, L. A Maternal Functional Module in the Mammalian Oocyte-To-Embryo Transition. Trends Mol. Med. 2017, 23, 1014–1023. [Google Scholar] [CrossRef] [PubMed]
- Swales, A.K.E.; Spears, N. Genomic Imprinting and Reproduction. Reproduction 2005, 130, 389–399. [Google Scholar] [CrossRef]
- Anvar, Z.; Chakchouk, I.; Demond, H.; Sharif, M.; Kelsey, G.; Van den Veyver, I.B. DNA Methylation Dynamics in the Female Germline and Maternal-Effect Mutations That Disrupt Genomic Imprinting. Genes (Basel). 2021, 12, 1214. [Google Scholar] [CrossRef] [PubMed]
- Demond, H.; Kelsey, G. The Enigma of DNA Methylation in the Mammalian Oocyte. F1000Research 2020, 9, F1000. [Google Scholar] [CrossRef] [PubMed]
- Weaver, J.R.; Bartolomei, M.S. Chromatin Regulators of Genomic Imprinting. Biochim. Biophys. Acta 2014, 1839, 169–177. [Google Scholar] [CrossRef]
- Nicholls, R.D.; Saitoh, S.; Horsthemke, B. Imprinting in Prader-Willi and Angelman Syndromes. Trends Genet. 1998, 14, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Poole, R.L.; Leith, D.J.; Docherty, L.E.; Shmela, M.E.; Gicquel, C.; Splitt, M.; Temple, I.K.; Mackay, D.J.G. Beckwith-Wiedemann Syndrome Caused by Maternally Inherited Mutation of an OCT-Binding Motif in the IGF2/H19-Imprinting Control Region, ICR1. Eur. J. Hum. Genet. 2012, 20, 240–243. [Google Scholar] [CrossRef]
- Ishida, M. New Developments in Silver-Russell Syndrome and Implications for Clinical Practice. Epigenomics 2016, 8, 563–580. [Google Scholar] [CrossRef]
- Begemann, M.; Rezwan, F.I.; Beygo, J.; Docherty, L.E.; Kolarova, J.; Schroeder, C.; Buiting, K.; Chokkalingam, K.; Degenhardt, F.; Wakeling, E.L.; et al. Maternal Variants in NLRP and Other Maternal Effect Proteins Are Associated with Multilocus Imprinting Disturbance in Offspring. J. Med. Genet. 2018, 55, 497–504. [Google Scholar] [CrossRef]
- Boonen, S.E.; Mackay, D.J.G.; Hahnemann, J.M.D.; Docherty, L.; Grønskov, K.; Lehmann, A.; Larsen, L.G.; Haemers, A.P.; Kockaerts, Y.; Dooms, L.; et al. Transient Neonatal Diabetes, ZFP57, and Hypomethylation of Multiple Imprinted Loci: A Detailed Follow-Up. Diabetes Care 2013, 36, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Kou, Y.C.; Shao, L.; Peng, H.H.; Rosetta, R.; del Gaudio, D.; Wagner, A.F.; Al-Hussaini, T.K.; Van den Veyver, I.B. A Recurrent Intragenic Genomic Duplication, Other Novel Mutations in NLRP7 and Imprinting Defects in Recurrent Biparental Hydatidiform Moles. Mol. Hum. Reprod. 2008, 14, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Mackay, D.J.G.; Callaway, J.L.A.; Marks, S.M.; White, H.E.; Acerini, C.L.; Boonen, S.E.; Dayanikli, P.; Firth, H.V.; Goodship, J.A.; Haemers, A.P.; et al. Hypomethylation of Multiple Imprinted Loci in Individuals with Transient Neonatal Diabetes Is Associated with Mutations in ZFP57. Nat. Genet. 2008, 40, 949–951. [Google Scholar] [CrossRef] [PubMed]
- Barberet, J.; Binquet, C.; Guilleman, M.; Doukani, A.; Choux, C.; Bruno, C.; Bourredjem, A.; Chapusot, C.; Bourc’his, D.; Duffourd, Y.; Fauque, P. Do Assisted Reproductive Technologies and in Vitro Embryo Culture Influence the Epigenetic Control of Imprinted Genes and Transposable Elements in Children? Hum. Reprod. 2021, 36, 479–492. [Google Scholar] [CrossRef] [PubMed]
- Barberet, J.; Romain, G.; Binquet, C.; Guilleman, M.; Bruno, C.; Ginod, P.; Chapusot, C.; Choux, C.; Fauque, P. Do Frozen Embryo Transfers Modify the Epigenetic Control of Imprinted Genes and Transposable Elements in Newborns Compared with Fresh Embryo Transfers and Natural Conceptions? Fertil. Steril. 2021, 116, 1468–1480. [Google Scholar] [CrossRef]
- Hattori, H.; Hiura, H.; Kitamura, A.; Miyauchi, N.; Kobayashi, N.; Takahashi, S.; Okae, H.; Kyono, K.; Kagami, M.; Ogata, T.; Arima, T. Association of Four Imprinting Disorders and ART. Clin. Epigenetics 2019, 11, 21. [Google Scholar] [CrossRef]
- Yao, J.-F.; Huang, Y.-F.; Huang, R.-F.; Lin, S.-X.; Guo, C.-Q.; Hua, C.-Z.; Wu, P.-Y.; Hu, J.-F.; Li, Y.-Z. Effects of Vitrification on the Imprinted Gene Snrpn in Neonatal Placental Tissue. Reprod. Dev. Med. 2020, 4, 25–31. [Google Scholar] [CrossRef]
- Ma, Y.; Ma, Y.; Wen, L.; Lei, H.; Chen, S.; Wang, X. Changes in DNA Methylation and Imprinting Disorders in E9.5 Mouse Fetuses and Placentas Derived from Vitrified Eight-Cell Embryos. Mol. Reprod. Dev. 2019, 86, 404–415. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, L.; He, F. Embryo Vitrification Affects the Methylation of the H19/Igf2 Differentially Methylated Domain and the Expression of H19 and Igf2. Fertil. Steril. 2010, 93, 2729–2733. [Google Scholar] [CrossRef]
- Henningsen, A.A.; Gissler, M.; Rasmussen, S.; Opdahl, S.; Wennerholm, U.B.; Spangsmose, A.L.; Tiitinen, A.; Bergh, C.; Romundstad, L.B.; Laivuori, H.; et al. Imprinting Disorders in Children Born after ART: A Nordic Study from the CoNARTaS Group. Hum. Reprod. 2020, 35, 1178–1184. [Google Scholar] [CrossRef]
- Lazaraviciute, G.; Kauser, M.; Bhattacharya, S.; Haggarty, P.; Bhattacharya, S. A Systematic Review and Meta-Analysis of DNA Methylation Levels and Imprinting Disorders in Children Conceived by IVF/ICSI Compared with Children Conceived Spontaneously. Hum. Reprod. Update 2014, 20, 840–852. [Google Scholar] [CrossRef]
- Deanesly, R. Immature Rat Ovaries Grafted after Freezing and Thawing. J. Endocrinol. 1954, 11, 197–200. [Google Scholar] [CrossRef] [PubMed]
- Donnez, J.; Dolmans, M.M.; Demylle, D.; Jadoul, P.; Pirard, C.; Squifflet, J.; Martinez-Madrid, B.; van Langendonckt, A. Livebirth after Orthotopic Transplantation of Cryopreserved Ovarian Tissue. Lancet 2004, 364, 1405–1410. [Google Scholar] [CrossRef] [PubMed]
- Yding Andersen, C.; Mamsen, L.S.; Kristensen, S.G. Fertility preservation: Freezing of Ovarian Tissue and Clinical Opportunities. Reproduction 2019, 158, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, K.; Cheng, Y.; Suzuki, N.; Deguchi, M.; Sato, Y.; Takae, S.; Ho, C.; Kawamura, N.; Tamura, M.; Hashimoto, S.; et al. Hippo Signaling Disruption and Akt Stimulation of Ovarian Follicles for Infertility Treatment. Proc. Natl. Acad. Sci. U. S. A. 2013, 110, 17474–17479. [Google Scholar] [CrossRef]
- Suzuki, N.; Yoshioka, N.; Takae, S.; Sugishita, Y.; Tamura, M.; Hashimoto, S.; Morimoto, Y.; Kawamura, K. Successful Fertility Preservation Following Ovarian Tissue Vitrification in Patients with Primary Ovarian Insufficiency. Hum. Reprod. 2015, 30, 608–615. [Google Scholar] [CrossRef]
- Rivas Leonel, E.C.; Lucci, C.M.; Amorim, C.A. Cryopreservation of Human Ovarian Tissue: A Review. Transfus. Med. hemotherapy 2019, 46, 173–181. [Google Scholar] [CrossRef]
- Cheng, H.; Han, Y.; Zhang, J.; Zhang, S.; Zhai, Y.; An, X.; Li, Q.; Duan, J.; Zhang, X.; Li, Z.; Tang, B.; Shen, H. Effects of Dimethyl Sulfoxide (DMSO) on DNA Methylation and Histone Modification in Parthenogenetically Activated Porcine Embryos. Reprod. Fertil. Dev. 2022, 34, 598–607. [Google Scholar] [CrossRef]
- Iwatani, M.; Ikegami, K.; Kremenska, Y.; Hattori, N.; Tanaka, S.; Yagi, S.; Shiota, K. Dimethyl Sulfoxide Has an Impact on Epigenetic Profile in Mouse Embryoid Body. Stem Cells 2006, 24, 2549–2556. [Google Scholar] [CrossRef]
- Verheijen, M.; Lienhard, M.; Schrooders, Y.; Clayton, O.; Nudischer, R.; Boerno, S.; Timmermann, B.; Selevsek, N.; Schlapbach, R.; Gmuender, H.; et al. DMSO Induces Drastic Changes in Human Cellular Processes and Epigenetic Landscape in Vitro. Sci. Rep. 2019, 9, 4641. [Google Scholar] [CrossRef]
- Nakamura, Y.; Obata, R.; Okuyama, N.; Aono, N.; Hashimoto, T.; Kyono, K. Residual Ethylene Glycol and Dimethyl Sulphoxide Concentration in Human Ovarian Tissue during Warming/Thawing Steps Following Cryopreservation. Reprod. Biomed. Online 2017, 35, 311–313. [Google Scholar] [CrossRef]
- Dolmans, M.-M.; Donnez, J.; Cacciottola, L. Fertility Preservation: The Challenge of Freezing and Transplanting Ovarian Tissue. Trends Mol. Med. 2021, 27, 777–791. [Google Scholar] [CrossRef]
- O’Brien, M.J.; Pendola, J.K.; Eppig, J.J. A Revised Protocol for in Vitro Development of Mouse Oocytes from Primordial Follicles Dramatically Improves Their Developmental Competence. Biol. Reprod. 2003, 68, 1682–1686. [Google Scholar] [CrossRef]
- Telfer, E.E.; Andersen, C.Y. In Vitro Growth and Maturation of Primordial Follicles and Immature Oocytes. Fertil. Steril. 2021, 115, 1116–1125. [Google Scholar] [CrossRef]
- McLaughlin, M.; Albertini, D.F.; Wallace, W.H.B.; Anderson, R.A.; Telfer, E.E. Metaphase II Oocytes from Human Unilaminar Follicles Grown in a Multi-Step Culture System. Mol. Hum. Reprod. 2018, 24, 135–142. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, L.; Wang, Z.; Chang, H.; Xie, X.; Fu, L.; Zhang, Y.; Quan, F. Resveratrol Improved the Developmental Potential of Oocytes after Vitrification by Modifying the Epigenetics. Mol. Reprod. Dev. 2019, 86, 862–870. [Google Scholar] [CrossRef]
- Cheng, K.-R.; Fu, X.-W.; Zhang, R.-N.; Jia, G.-X.; Hou, Y.-P.; Zhu, S.-E. Effect of Oocyte Vitrification on Deoxyribonucleic Acid Methylation of H19, Peg3, and Snrpn Differentially Methylated Regions in Mouse Blastocysts. Fertil. Steril. 2014, 102, 1183–1190. [Google Scholar] [CrossRef]
- Ma, Y.; Long, C.; Liu, G.; Bai, H.; Ma, L.; Bai, T.; Zuo, Y.; Li, S. WGBS Combined with RNA-Seq Analysis Revealed That Dnmt1 Affects the Methylation Modification and Gene Expression Changes during Mouse Oocyte Vitrification. Theriogenology 2022, 177, 11–21. [Google Scholar] [CrossRef]
- Gu, R.; Ge, N.; Huang, B.; Fu, J.; Zhang, Y.; Wang, N.; Xu, Y.; Li, L.; Peng, X.; Zou, Y.; Sun, Y.; Sun, X. Impacts of Vitrification on the Transcriptome of Human Ovarian Tissue in Patients with Gynecological Cancer. Front. Genet. 2023, 14, 1114650. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, W.; Todorov, P.; Pei, C.; Isachenko, E.; Rahimi, G.; Mallmann, P.; Nawroth, F.; Isachenko, V. RNA Transcripts in Human Ovarian Cells: Two-Time Cryopreservation Does Not Affect Developmental Potential. Int. J. Mol. Sci. 2023, 24, 6880. [Google Scholar] [CrossRef]
- He, Z.-Y.; Wang, H.-Y.; Zhou, X.; Liang, X.-Y.; Yan, B.; Wang, R.; Ma, L.-H.; Wang, Y.-L. Evaluation of Vitrification Protocol of Mouse Ovarian Tissue by Effect of DNA Methyltransferase-1 and Paternal Imprinted Growth Factor Receptor-Binding Protein 10 on Signaling Pathways. Cryobiology 2018, 80, 89–95. [Google Scholar] [CrossRef]
- Shirazi, A.; Naderi, M.M.; Hassanpour, H.; Heidari, M.; Borjian, S.; Sarvari, A.; Akhondi, M.M. The Effect of Ovine Oocyte Vitrification on Expression of Subset of Genes Involved in Epigenetic Modifications during Oocyte Maturation and Early Embryo Development. Theriogenology 2016, 86, 2136–2146. [Google Scholar] [CrossRef]
- Yan, Z.; Li, Q.; Zhang, L.; Kang, B.; Fan, W.; Deng, T.; Zhu, J.; Wang, Y. The Growth and Development Conditions in Mouse Offspring Derived from Ovarian Tissue Cryopreservation and Orthotopic Transplantation. J. Assist. Reprod. Genet. 2020, 37, 923–932. [Google Scholar] [CrossRef]
- Demant, M.; Trapphoff, T.; Fröhlich, T.; Arnold, G.J.; Eichenlaub-Ritter, U. Vitrification at the Pre-Antral Stage Transiently Alters Inner Mitochondrial Membrane Potential but Proteome of in Vitro Grown and Matured Mouse Oocytes Appears Unaffected. Hum. Reprod. 2012, 27, 1096–1111. [Google Scholar] [CrossRef]
- Pfeiffer, M.J.; Siatkowski, M.; Paudel, Y.; Balbach, S.T.; Baeumer, N.; Crosetto, N.; Drexler, H.C.A.; Fuellen, G.; Boiani, M. Proteomic Analysis of Mouse Oocytes Reveals 28 Candidate Factors of the “Reprogrammome”. J. Proteome Res. 2011, 10, 2140–2153. [Google Scholar] [CrossRef]
- Wang, H.-Y.; Li, Y.-H.; Sun, L.; Gao, X.; You, L.; Wang, Y.; Ma, J.-L.; Chen, Z.-J. Allotransplantation of Cryopreserved Prepubertal Mouse Ovaries Restored Puberty and Fertility without Affecting Methylation Profile of Snrpn-DMR. Fertil. Steril. 2013, 99, 241–247. [Google Scholar] [CrossRef]
- Yan, J.; Zhang, L.; Wang, T.; Li, R.; Liu, P.; Yan, L.; Qiao, J. Effect of Vitrification at the Germinal Vesicle Stage on the Global Methylation Status in Mouse Oocytes Subsequently Matured in Vitro. Chin. Med. J. (Engl). 2014, 127, 4019–4024. [Google Scholar] [CrossRef]
- Al-Khtib, M.; Perret, A.; Khoueiry, R.; Ibala-Romdhane, S.; Blachère, T.; Greze, C.; Lornage, J.; Lefèvre, A. Vitrification at the Germinal Vesicle Stage Does Not Affect the Methylation Profile of H19 and KCNQ1OT1 Imprinting Centers in Human Oocytes Subsequently Matured in Vitro. Fertil. Steril. 2011, 95, 1955–1960. [Google Scholar] [CrossRef]
- Yodrug, T.; Parnpai, R.; Hirao, Y.; Somfai, T. Effect of Vitrification at Different Meiotic Stages on Epigenetic Characteristics of Bovine Oocytes and Subsequently Developing Embryos. Anim. Sci. J. 2021, 92, e13596. [Google Scholar] [CrossRef]
- Trapphoff, T.; El Hajj, N.; Zechner, U.; Haaf, T.; Eichenlaub-Ritter, U. DNA Integrity, Growth Pattern, Spindle Formation, Chromosomal Constitution and Imprinting Patterns of Mouse Oocytes from Vitrified Pre-Antral Follicles. Hum. Reprod. 2010, 25, 3025–3042. [Google Scholar] [CrossRef]
- Damavandi, M.; Farrokh, P.; Zavareh, S. Effect of Mouse Ovarian Vitrification on Promoter Methylation of Inhba and Inhbb in Granulosa Cells of Follicles. Cryo Letters 2021, 42, 67–72. [Google Scholar]
- Sauvat, F.; Capito, C.; Sarnacki, S.; Poirot, C.; Bachelot, A.; Meduri, G.; Dandolo, L.; Binart, N. Immature Cryopreserved Ovary Restores Puberty and Fertility in Mice without Alteration of Epigenetic Marks. PLoS One 2008, 3, e1972. [Google Scholar] [CrossRef]
- Sauvat, F.; Bouilly, J.; Capito, C.; Lefèvre, A.; Blachère, T.; Borenstein, N.; Sarnacki, S.; Dandolo, L.; Binart, N. Ovarian Function Is Restored after Grafting of Cryopreserved Immature Ovary in Ewes. FASEB J. 2013, 27, 1511–1518. [Google Scholar] [CrossRef]
- Bonnet-Garnier, A.; Feuerstein, P.; Chebrout, M.; Fleurot, R.; Jan, H.-U.; Debey, P.; Beaujean, N. Genome Organization and Epigenetic Marks in Mouse Germinal Vesicle Oocytes. Int. J. Dev. Biol. 2012, 56, (10–12). [Google Scholar] [CrossRef]
- Kageyama, S.; Liu, H.; Kaneko, N.; Ooga, M.; Nagata, M.; Aoki, F. Alterations in Epigenetic Modifications during Oocyte Growth in Mice. Reproduction 2007, 133, 85–94. [Google Scholar] [CrossRef]
- Stewart, K.R.; Veselovska, L.; Kim, J.; Huang, J.; Saadeh, H.; Tomizawa, S.; Smallwood, S.A.; Chen, T.; Kelsey, G. Dynamic Changes in Histone Modifications Precede de Novo DNA Methylation in Oocytes. Genes Dev. 2015, 29, 2449–2462. [Google Scholar] [CrossRef]
- Tian, C.; Shen, L.; Gong, C.; Cao, Y.; Shi, Q.; Zhao, G. Microencapsulation and Nanowarming Enables Vitrification Cryopreservation of Mouse Preantral Follicles. Nat. Commun. 2022, 13, 7515. [Google Scholar] [CrossRef]
- Lee, P.-C.; Comizzoli, P. Desiccation and Supra-Zero Temperature Storage of Cat Germinal Vesicles Lead to Less Structural Damage and Similar Epigenetic Alterations Compared to Cryopreservation. Mol. Reprod. Dev. 2019, 86, 1822–1831. [Google Scholar] [CrossRef]
- Tamburrino, L.; Traini, G.; Marcellini, A.; Vignozzi, L.; Baldi, E.; Marchiani, S. Cryopreservation of Human Spermatozoa: Functional, Molecular and Clinical Aspects. Int. J. Mol. Sci. 2023, 24, 4656. [Google Scholar] [CrossRef]
- Picton, H.M.; Wyns, C.; Anderson, R.A.; Goossens, E.; Jahnukainen, K.; Kliesch, S.; Mitchell, R.T.; Pennings, G.; Rives, N.; Tournaye, H.; et al. A European Perspective on Testicular Tissue Cryopreservation for Fertility Preservation in Prepubertal and Adolescent Boys. Hum. Reprod. 2015, 30, 2463–2475. [Google Scholar] [CrossRef]
- Delgouffe, E.; Braye, A.; Goossens, E. Testicular Tissue Banking for Fertility Preservation in Young Boys: Which Patients Should Be Included? Front. Endocrinol. (Lausanne). 2022, 13, 854186. [Google Scholar] [CrossRef]
- Sanou, I.; van Maaren, J.; Eliveld, J.; Lei, Q.; Meißner, A.; de Melker, A.A.; Hamer, G.; van Pelt, A.M.M.; Mulder, C.L. Spermatogonial Stem Cell-Based Therapies: Taking Preclinical Research to the Next Level. Front. Endocrinol. (Lausanne). 2022, 13, 850219. [Google Scholar] [CrossRef]
- Wyns, C.; Kanbar, M.; Giudice, M.G.; Poels, J. Fertility Preservation for Prepubertal Boys: Lessons Learned from the Past and Update on Remaining Challenges towards Clinical Translation. Hum. Reprod. Update 2021, 27, 433–459. [Google Scholar] [CrossRef]
- Onofre, J.; Baert, Y.; Faes, K.; Goossens, E. Cryopreservation of Testicular Tissue or Testicular Cell Suspensions: A Pivotal Step in Fertility Preservation. Hum. Reprod. Update 2016, 22, 744–761. [Google Scholar] [CrossRef]
- Onofre, J.; Kadam, P.; Baert, Y.; Goossens, E. Testicular Tissue Cryopreservation Is the Preferred Method to Preserve Spermatogonial Stem Cells Prior to Transplantation. Reprod. Biomed. Online 2020, 40, 261–269. [Google Scholar] [CrossRef]
- Fayomi, A.P.; Peters, K.; Sukhwani, M.; Valli-Pulaski, H.; Shetty, G.; Meistrich, M.L.; Houser, L.; Robertson, N.; Roberts, V.; Ramsey, C.; et al. Autologous Grafting of Cryopreserved Prepubertal Rhesus Testis Produces Sperm and Offspring. Science 2019, 363, 1314–1319. [Google Scholar] [CrossRef]
- Goossens, E.; De Rycke, M.; Haentjens, P.; Tournaye, H. DNA Methylation Patterns of Spermatozoa and Two Generations of Offspring Obtained after Murine Spermatogonial Stem Cell Transplantation. Hum. Reprod. 2009, 24, 2255–2263. [Google Scholar] [CrossRef]
- Serrano, J.B.; van Eekelen, R.; de Winter-Korver, C.M.; van Daalen, S.K.M.; Tabeling, N.C.; Catsburg, L.A.E.; Gijbels, M.J.J.; Mulder, C.L.; van Pelt, A.M.M. Impact of Restoring Male Fertility with Transplantation of in Vitro Propagated Spermatogonial Stem Cells on the Health of Their Offspring throughout Life. Clin. Transl. Med. 2021, 11, e531. [Google Scholar] [CrossRef]
- Nickkholgh, B.; Mizrak, S.C.; van Daalen, S.K.M.; Korver, C.M.; Sadri-Ardekani, H.; Repping, S.; van Pelt, A.M.M. Genetic and Epigenetic Stability of Human Spermatogonial Stem Cells during Long-Term Culture. Fertil. Steril. 2014, 102, 1700–1707. [Google Scholar] [CrossRef] [PubMed]
- Kanatsu-Shinohara, M.; Ogonuki, N.; Iwano, T.; Lee, J.; Kazuki, Y.; Inoue, K.; Miki, H.; Takehashi, M.; Toyokuni, S.; Shinkai, Y.; et al. Genetic and Epigenetic Properties of Mouse Male Germline Stem Cells during Long-Term Culture. Development 2005, 132, 4155–4163. [Google Scholar] [CrossRef]
- Langenstroth-Röwer, D.; Gromoll, J.; Wistuba, J.; Tröndle, I.; Laurentino, S.; Schlatt, S.; Neuhaus, N. De Novo Methylation in Male Germ Cells of the Common Marmoset Monkey Occurs during Postnatal Development and Is Maintained in Vitro. Epigenetics 2017, 12, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Katagiri, K.; Gohbara, A.; Inoue, K.; Ogonuki, N.; Ogura, A.; Kubota, Y.; Ogawa, T. In Vitro Production of Functional Sperm in Cultured Neonatal Mouse Testes. Nature 2011, 471, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Yokonishi, T.; Sato, T.; Komeya, M.; Katagiri, K.; Kubota, Y.; Nakabayashi, K.; Hata, K.; Inoue, K.; Ogonuki, N.; Ogura, A.; Ogawa, T. Offspring Production with Sperm Grown in Vitro from Cryopreserved Testis Tissues. Nat. Commun. 2014, 5, 4320. [Google Scholar] [CrossRef]
- Wu, X.; Goodyear, S.M.; Abramowitz, L.K.; Bartolomei, M.S.; Tobias, J.W.; Avarbock, M.R.; Brinster, R.L. Fertile Offspring Derived from Mouse Spermatogonial Stem Cells Cryopreserved for More than 14 Years. Hum. Reprod. 2012, 27, 1249–1259. [Google Scholar] [CrossRef] [PubMed]
- Serrano, J.B.; Tabeling, N.C.; de Winter-Korver, C.M.; van Daalen, S.K.M.; van Pelt, A.M.M.; Mulder, C.L. Sperm DNA Methylation Is Predominantly Stable in Mice Offspring Born after Transplantation of Long-Term Cultured Spermatogonial Stem Cells. Clin. Epigenetics 2023, 15, 58. [Google Scholar] [CrossRef] [PubMed]
- Oblette, A.; Rives-Feraille, A.; Dumont, L.; Delessard, M.; Saulnier, J.; Rives, N.; Rondanino, C. Dynamics of Epigenetic Modifications in ICSI Embryos from in Vitro-Produced Spermatozoa. Andrology 2021, 9, 640–656. [Google Scholar] [CrossRef] [PubMed]
- Oblette, A.; Rondeaux, J.; Dumont, L.; Delessard, M.; Saulnier, J.; Rives, A.; Rives, N.; Rondanino, C. DNA Methylation and Histone Post-Translational Modifications in the Mouse Germline Following in-Vitro Maturation of Fresh or Cryopreserved Prepubertal Testicular Tissue. Reprod. Biomed. Online 2019, 39, 383–401. [Google Scholar] [CrossRef]
- Kawai, K.; Li, Y.-S.; Song, M.-F.; Kasai, H. DNA Methylation by Dimethyl Sulfoxide and Methionine Sulfoxide Triggered by Hydroxyl Radical and Implications for Epigenetic Modifications. Bioorg. Med. Chem. Lett. 2010, 20, 260–265. [Google Scholar] [CrossRef]
- Zeng, C.; Peng, W.; Ding, L.; He, L.; Zhang, Y.; Fang, D.; Tang, K. A Preliminary Study on Epigenetic Changes during Boar Spermatozoa Cryopreservation. Cryobiology 2014, 69, 119–127. [Google Scholar] [CrossRef]
- Khosravizadeh, Z.; Hassanzadeh, G.; Tavakkoly Bazzaz, J.; Alizadeh, F.; Totonchi, M.; Salehi, E.; Khodamoradi, K.; Khanehzad, M.; Hosseini, S.R.; Abolhassani, F. The Effect of Cryopreservation on DNA Methylation Patterns of the Chromosome 15q11-Q13 Region in Human Spermatozoa. Cell Tissue Bank. 2020, 21, 433–445. [Google Scholar] [CrossRef]
- Kläver, R.; Bleiziffer, A.; Redmann, K.; Mallidis, C.; Kliesch, S.; Gromoll, J. Routine Cryopreservation of Spermatozoa Is Safe-Evidence from the DNA Methylation Pattern of Nine Spermatozoa Genes. J. Assist. Reprod. Genet. 2012, 29, 943–950. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Scheme of epigenetic control. The epigenome includes reversible DNA methylation, post-translational histone modifications (PTMs), and the abundance and availability of relevant transcripts and proteins to establish or maintain DNA methylation. Methylation (Me), acetylation (Ac) and ubiquitylation (Ub).
Figure 1.
Scheme of epigenetic control. The epigenome includes reversible DNA methylation, post-translational histone modifications (PTMs), and the abundance and availability of relevant transcripts and proteins to establish or maintain DNA methylation. Methylation (Me), acetylation (Ac) and ubiquitylation (Ub).
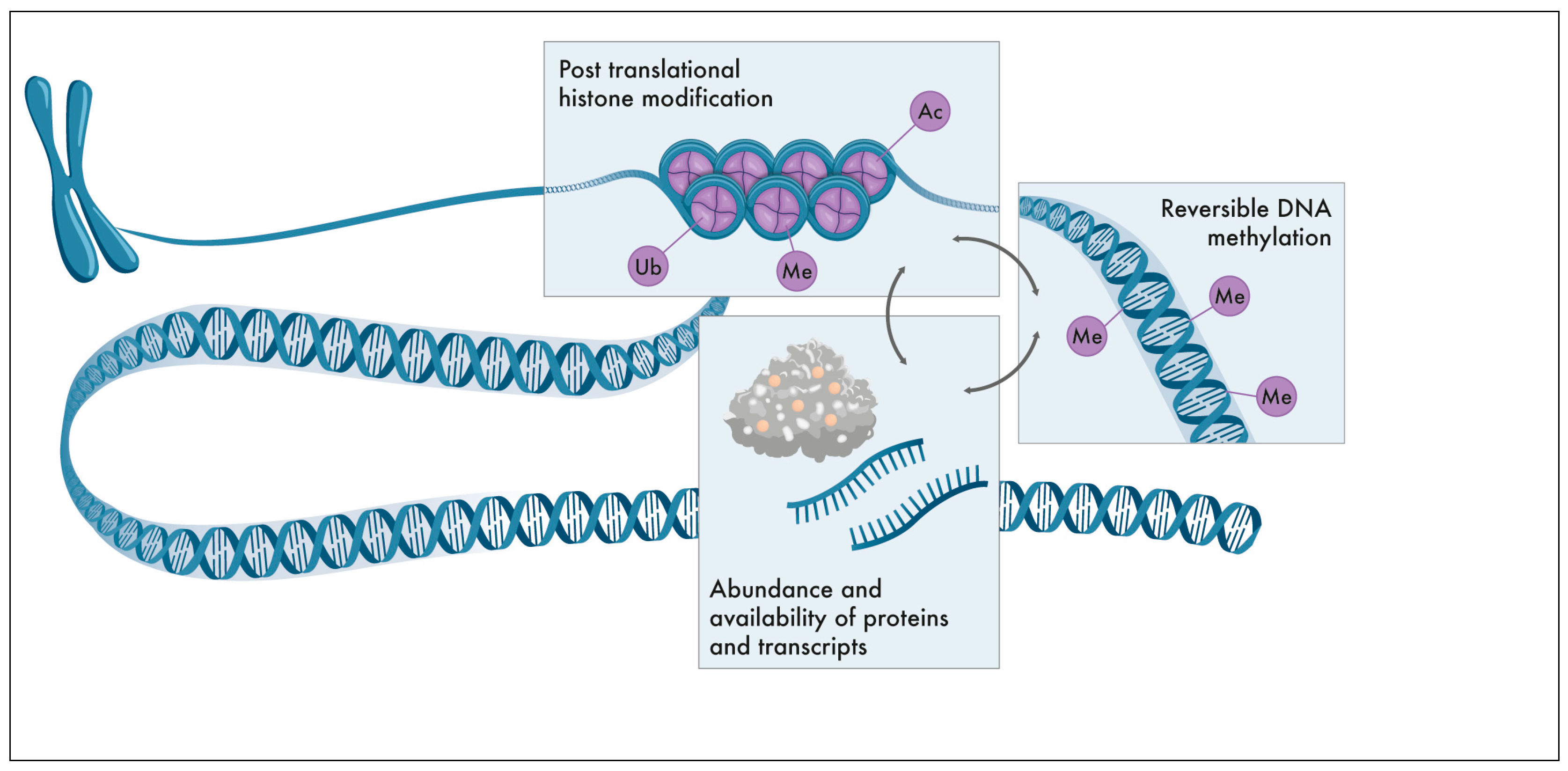
Figure 2.
DNA methylation pattern in gametogenesis and early embryogenesis. DNA methylation pattern is erased in primordial germ cells (PGC) followed by subsequent de novo establishment of the germ cell-specific DNA methylation profile for imprinted genes and gDNA. During early embryogenesis, global DNA demethylation occurs actively in paternally inherited DNA and passively in maternally inherited DNA in the zygote and growing embryo. During embryogenesis, cell and tissue-specific de novo gDNA reprogramming is established. Methylation pattern of imprinted genes is maintained after fertilisation and embryogenesis. Medically Assisted Reproduction (MAR) techniques that may interfere with the epigenetic profile including controlled ovarian stimulation (COS), ovarian tissue cryopreservation (OTC), testicular tissue cryopreservation (TTC), in vitro fertilisation (IVF) and intracytoplasmic sperm injection (ICSI).
Figure 2.
DNA methylation pattern in gametogenesis and early embryogenesis. DNA methylation pattern is erased in primordial germ cells (PGC) followed by subsequent de novo establishment of the germ cell-specific DNA methylation profile for imprinted genes and gDNA. During early embryogenesis, global DNA demethylation occurs actively in paternally inherited DNA and passively in maternally inherited DNA in the zygote and growing embryo. During embryogenesis, cell and tissue-specific de novo gDNA reprogramming is established. Methylation pattern of imprinted genes is maintained after fertilisation and embryogenesis. Medically Assisted Reproduction (MAR) techniques that may interfere with the epigenetic profile including controlled ovarian stimulation (COS), ovarian tissue cryopreservation (OTC), testicular tissue cryopreservation (TTC), in vitro fertilisation (IVF) and intracytoplasmic sperm injection (ICSI).
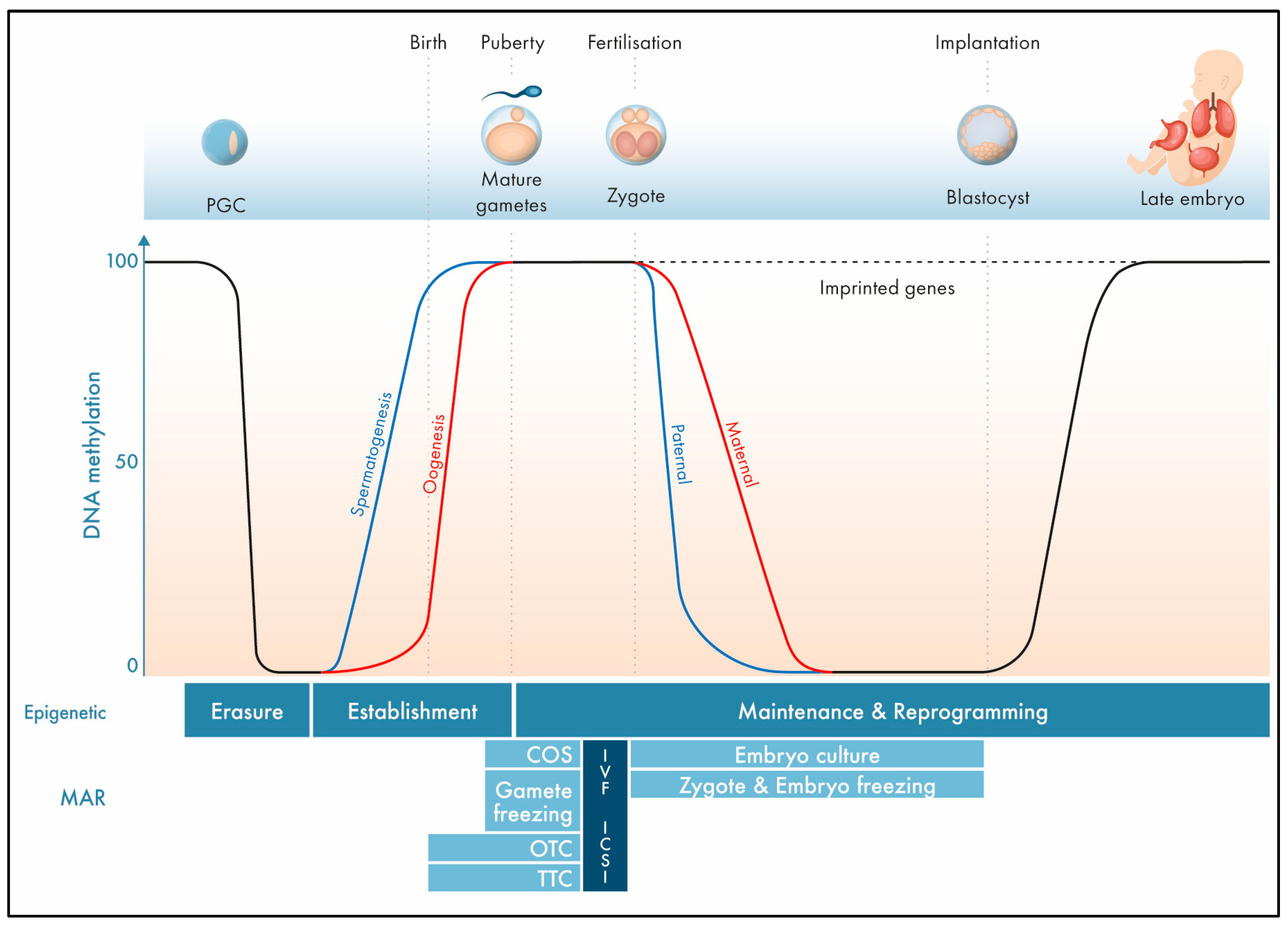
Figure 3.
Strategies for fertility preservation in female patients. Conventional oocyte/zygote/embryo cryopreservation (left pathway) and ovarian tissue cryopreservation and in vivo/in vitro oogenesis (right pathway). Left pathway: Cryopreservation of mature MII oocytes or embryos after controlled ovarian stimulation, oocyte pick-up, and Medically Assisted Reproduction (MAR) techniques. Right pathway: Ovarian tissue cryopreservation followed by in vivo or in vitro folliculogenesis and oogenesis. After orthotopic or heterotopic transplantation, fertilisation can occur spontaneously (orthotopic autotransplantation) or via MAR techniques (heterotopic autotransplantation). Different in vitro techniques to avoid the risk of remission of malignant cells including (experimental) in vitro culture of immature follicles (IVC), in vitro maturation of immature oocytes (IVM), and artificial ovaries (3D AO) followed by MAR or autotransplantation.
Figure 3.
Strategies for fertility preservation in female patients. Conventional oocyte/zygote/embryo cryopreservation (left pathway) and ovarian tissue cryopreservation and in vivo/in vitro oogenesis (right pathway). Left pathway: Cryopreservation of mature MII oocytes or embryos after controlled ovarian stimulation, oocyte pick-up, and Medically Assisted Reproduction (MAR) techniques. Right pathway: Ovarian tissue cryopreservation followed by in vivo or in vitro folliculogenesis and oogenesis. After orthotopic or heterotopic transplantation, fertilisation can occur spontaneously (orthotopic autotransplantation) or via MAR techniques (heterotopic autotransplantation). Different in vitro techniques to avoid the risk of remission of malignant cells including (experimental) in vitro culture of immature follicles (IVC), in vitro maturation of immature oocytes (IVM), and artificial ovaries (3D AO) followed by MAR or autotransplantation.
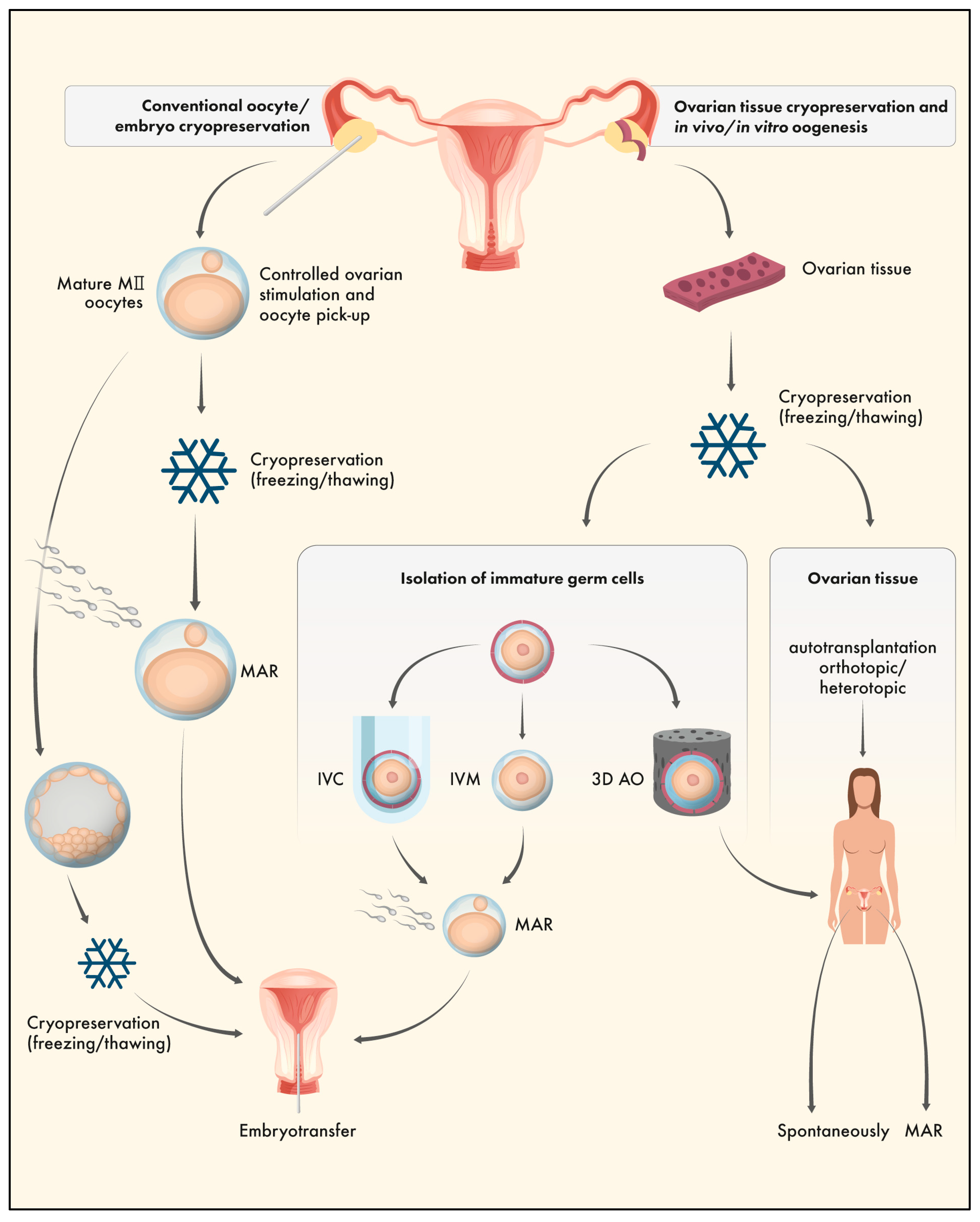
Figure 4.
Strategies for fertility preservation in male patients. Cryopreservation of mature spermatozoa (left pathway) and (experimental) testicular tissue cryopreservation and in vivo/in vitro spermatogenesis (right pathway). Left pathway: Cryopreservation of mature spermatozoa or testicular tissue followed by Medically Assisted Reproduction (MAR) techniques (including IVF, ICSI, TESE-ICSI). Right pathway: Testicular tissue cryopreservation followed by in vivo or in vitro spermatogenesis. Resumption of gametogenesis can occur after tissue grafting or, to avoid the risk of remission of malignant cells, after isolation of spermatogonial stem cells (SSC), in vitro proliferation/spermatogenesis, SSC transplantation, and/or MAR techniques.
Figure 4.
Strategies for fertility preservation in male patients. Cryopreservation of mature spermatozoa (left pathway) and (experimental) testicular tissue cryopreservation and in vivo/in vitro spermatogenesis (right pathway). Left pathway: Cryopreservation of mature spermatozoa or testicular tissue followed by Medically Assisted Reproduction (MAR) techniques (including IVF, ICSI, TESE-ICSI). Right pathway: Testicular tissue cryopreservation followed by in vivo or in vitro spermatogenesis. Resumption of gametogenesis can occur after tissue grafting or, to avoid the risk of remission of malignant cells, after isolation of spermatogonial stem cells (SSC), in vitro proliferation/spermatogenesis, SSC transplantation, and/or MAR techniques.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



