
Submitted:
07 June 2023
Posted:
07 June 2023
You are already at the latest version
Abstract
Maternal infections during pregnancy may increase the risk of psychiatric disorders in offspring. We recently demonstrated that activation of peroxisome proliferator-activate receptor‐α (PPARα), with the clinically available agonist fenofibrate, attenuates the neurodevelopmental disturbances induced by maternal immune activation (MIA) in rat offspring. We hypothesized that fenofibrate might reduce MIA-induced cytokine imbalance using a MIA model based on the viral mimetic polyriboinosinic-polyribocytidilic acid [poly (I:C)]. By using the Bio-Plex Multiplex-Immunoassay-System, we measured cytokine/chemokine levels in maternal serum and in the fetal brain of rats treated with fenofibrate, at 6 and 24 hours after poly (I:C). We found that MIA induced time-dependent changes in the levels of several cytokines/chemokines/colony-stimulating factors (CSFs). Specifically, the maternal serum of the poly (I:C)/CTRL group showed increased levels of (i) proinflammatory chemokine macrophage inflammatory protein 1-alpha (MIP-1α), (ii) tumor necrosis factor-alpha (TNF-), the monocyte chemoattractant protein-1 (MCP-1), the macrophage (M-CSF) and granulocyte-macrophage colony-stimulating factor (GM-CSF). Conversely, in the fetal brain of the poly (I:C)/CTRL group, interleukin 12p70 and MIP-1 levels were lower than in veh/CTRL group. Notably, MIP-1, TNF-, GRO/KC, GM-CSF, and M-CSF levels were lower in the poly (I:C)/FEN than in poly (I:C)/CTRL rats, indicating the protective role of the PPARα agonist. PPARα might represent a therapeutic target to attenuate the consequences of MIA.
Keywords:
MIA
; cytokines
; chemokines
; colony-stimulating factors
; Poly (I:C)
; neurodevelopmental disorders
1. Introduction
Maternal infections during pregnancy have been consistently related to neurodevelopmental disorders in offspring [1,2,3]. Currently, the pandemic impact of SARS-CoV-2 on pregnant women raises serious concerns. Among others, maternal immune activation (MIA) might represent a pathway by which infection with SARS-CoV-2 during gestation could influence the developing brain [4,5]. Several studies in animal models have concurred to unveil the relationship between MIA and altered fetal neurodevelopment, which might lead to long-term anatomical and behavioral impairments correlated with inflammation [6,7,8]. Thus, preclinical and clinical studies showed that MIA triggers a cytokine imbalance that increases the risk of neurodevelopmental disorders in the offspring [9,10,11,12]. Both rodent and non-human primate models took advantage of the polyriboinosinic-polyribocytidylic acid [poly (I:C)], a double-stranded synthetic RNA that elicits an innate immune response by simulating a viral infection. Offspring whose mothers were exposed to poly (I:C) during gestation showed schizophrenia- and autism-like traits [13,14,15].
In our previous studies, we reported behavioral impairments correlated with an altered mesolimbic dopamine transmission both in the first and second generation [16,17]. We showed how inflammation caused by MIA alters the endocannabinoid signaling, which negatively influences the dopamine system, eventually leading to schizophrenia-like phenotype in adulthood [18]. In addition, we demonstrated that activation of peroxisome proliferator-activated receptor-α (PPARα) with the clinically available agonist fenofibrate attenuates the neurodevelopmental disturbances induced by MIA in rat offspring [19]. PPARα are nuclear receptor transcription factors abundantly expressed in the central nervous system, where they fine-tune several functions, such as the mesolimbic neuronal activity and other neuronal pathways involved in affective behavior [20,21,22]. Importantly, PPARα modulate genes involved in the anti-inflammatory response [23,24,25]. PPARα-mediated effects involve the nuclear factor kappa-light-chain-enhancer of activated B cells (NFkB) that leads to the inhibition of pro-inflammatory cytokines such as interleukins 1 and 6 (IL-1, IL-6) and tumor necrosis factor-α (TNF-α) [26]. Poly (I:C) acts on the toll-like-receptor 3 (TLR-3) pathway that activates the NFκB through the mitogen-activated protein kinases (MAPK) cascade [27], leading to the synthesis of several proinflammatory cytokines, such as IL-1, IL-6, and TNF-α [14,28]. Most studies reporting the effects of poly (I:C) on cytokine levels in maternal serum, placenta, or fetal brain are performed in mice, reporting conflicting results [29]. Additionally, early studies investigated the levels of proinflammatory cytokines such as IL-1, IL-6, and TNF-α [30,31,32]. Nevertheless, only recently, several cytokines and chemokines have been analyzed in maternal serum and in fetal brains [33]. However, to our knowledge, only two studies have examined the complete profile of cytokine, chemokine and colony-stimulating factors (CSFs) in mice [34] and Long Evans rats [35].
In a rat model of MIA, this study aimed i) to investigate the full cytokines/chemokines/CSFs profile in the maternal serum and in the fetal brain; ii) to test the hypothesis that a treatment with the PPARα agonist fenofibrate, administered during pregnancy, might reduce MIA-induced cytokine imbalance.
2. Results
Figure 1 shows the experimental protocol, illustrating the MIA model, control (CTRL) or fenofibrate treatment (FEN) during gestation and tissue collection at 6 h and 24 h after poly (I:C) or vehicle (Veh) injection.
2.1. Effect of fenofibrate treatment on MIA-induced inflammation in the maternal serum.
To examine whether FEN might rescue the inflammation induced by MIA, we analyzed cytokines, chemokines and CSFs levels in the maternal serum in both CTRL and FEN, 6 h and 24 h after poly (I:C) or veh injection.
Data showed that MIA-induced time-dependent changes in the levels of several maternal serum cytokines, chemokines and CSFs and FEN reverts some of these MIA-induced effects (Table 1).
2.1.1. Chemokines
As shown in figure 2A, at 6 h two way ANOVA showed a significant main effect of MIA for the concentration of the chemokine MIP-1α [F(1,16)=25.27, p=0.0001] revealing a significantly higher concentration in poly (I:C)/CTRL than in veh/CTRL (p<0.01) and in poly (I:C)/ FEN than in veh/FEN (p<0.05).
At 24 h, two-way ANOVA revealed a MIA × treatment interaction [F(1,12)=15.11, p=0.0022], and a significant main effect of MIA [F(1,12)=23.23, p=0.0004] but not of treatment. Tukey’s post hoc test revealed significant differences in the levels of MIP-1α between poly (I:C) and veh administration in the CTRL group (p<0.001), and poly (I:C)/CTRL and veh/FEN group (p<0.01). Remarkably, the levels of this proinflammatory chemokine are reduced in the poly (I:C)/FEN group as compared with the poly (I:C)/CTRL group (p<0.05).
Furthermore, at 6 h two-way ANOVA for GRO/KC showed a significant main effect of MIA [F(1,15)=9.51, p=0.0076] and of treatment [F(1,15)=11.70, p=0.0038] but not MIA × treatment interaction (Figure 2B). Specifically, GRO/KC displayed significantly higher concentrations in poly (I:C)/CTRL than in the veh/CTRL (p<0.01) and poly (I:C)/FEN (p<0.01). No differences among groups were detected at 24 h.
Finally, two way ANOVA for the concentration of the chemokine MCP-1 (Figure 2C) at 6 h and 24 h revealed a main effect of MIA [F(1,16)=20.18, p=0.0004 and F(1,12)=8.67, p=0.0123, respectively]. At 6 h, MCP-1 levels were higher in the poly (I:C)/FEN than in the veh/FEN group (p<0.01), while at 24 h levels were higher in the poly (I:C)/CTRL than in the veh/CTRL group (p< 0.05).
2.1.2. Cytokines
Among the analyzed proinflammatory cytokines, the levels of TNF-α, IL-1α, IL-17, and IL-18 were altered (Figure 3). Specifically, at 6 h after poly (I:C) injection, the two-way ANOVA revealed a MIA × treatment interaction for IL-17 [F(1,15)=8.91, p=0.0093] and IL-18 [F(1,16)=8.20, p=0.0112] and a significant main effect of treatment for IL-18 [F(1,16)=9.62, p=0.0069]. Tukey’s post hoc test revealed a significant increase of both IL-17 (p<0.05) and IL-18 (p<0.01) levels in the maternal serum of the poly (I:C)/FEN as compared to the poly (I:C)/CTRL group (Figure 3A,B).
A significant main effect of treatment was also observed for IL-1α [F(1,16)=5.76, p=0.0289] with poly (I:C)/FEN showing higher levels than poly (I:C)/CTRL group (p<0.05) (Figure 3C).
At 24 h after the injection, no main effect of treatment, MIA nor their interaction, was observed for IL-17, IL-18 and IL-1α. The two-way ANOVA showed a MIA × treatment interaction for TNF-α [F(1,12)=11.83, p=0.0049]. Notably, the levels of TNF-α were reduced in the poly (I:C)/FEN as compared to the poly (I:C)/CTRL group (p<0.05), indicating the protective effect of fenofibrate treatment (Figure 3D).
2.1.3. Colony-stimulating factors
As shown in Figure 4, no difference in the CSFs levels was observed among groups at 6 h after poly (I:C) injection, while at 24 h two-way ANOVA displayed a significant MIA x treatment interaction for GM-CSF [F(1,12)=14.48, p=0.0025], G-CSF [F(1,12)=4.89, p=0.0471], and M-CSF [F(1,12)=9.25, p=0.0102]. The Tukey’s post hoc revealed a decrease of GM-CSF (p<0.05) (Figure 4A) and M-CSF (p<0.05) (Figure 4C) in the poly (I:C)/FEN as compared to the poly (I:C)/CTRL group. Moreover, the levels of GM-CSF were reduced (p< 0.05) in the poly (I:C)/FEN as compared to the veh/FEN group (Figure 4A).
2.2. Effect of fenofibrate treatment on MIA-induced neuroinflammation in the fetal brain.
We analyzed the levels of cytokines, chemokines, and CSFs in the fetal brain from dams injected with veh or poly (I:C) at GD15 and sacrificed 6 h or 24 h later. We also examined whether the FEN treatment might rescue the MIA induced neuroinflammation.
As shown in table 2, at 6 h, all the cytokines, chemokines and CSFs, except IL-6 and VEGF, were found in controls, revealing their physiological presence in the developing brain without an immune response. All cytokines showed levels of expression ranging from 40 to 337 pg/ml, except IL-17, IL-7, and IL-10 whose concentrations were below 35 pg/ml. Chemokines and CSFs were also present in the control brain homogenates at concentrations ranging from 1.34 to 108 pg/ml.
ND: not detectable, cytokines, chemokines or colony-stimulating factors concentrations under the limit of detection. Veh: vehicle; CTRL: control treatment; FEN: Fenofibrate treatment.
2.2.1. Cytokines
At 6 h, two-way ANOVA for the cytokine IL-12p70 revealed a MIA x treatment interaction [F(1,13)=9.16, p=0.0097], and a significant main effect of MIA [F(1,13)=16.00, p=0.0015]. Tukey’s post hoc displayed a significant decrease of IL-12p70 fetal brain levels in the poly (I:C)/CTRL as compared to the veh/CTRL (p<0.01) and poly (I:C)/FEN as compared to the veh/CTRL group (p<0.05) (Figure 5A). Moreover, two-way ANOVA revealed a significant main effect of MIA for IL-4 [F(1,13)=5.08, p=0.0421] and IFN-γ [F(1,13)=5.28, p=0.0387] with fetus brain from poly (I:C)/CTRL dam showing lower levels than veh/CTRL dam (p<0.05) (Figure 5B,C). A significant main effect of MIA was also observed for IL-1β [F(1,13)=4.90, p=0.0453] showing an upward trend of its value in fetus brain from poly (I:C)/CTRL and poly (I:C)/FEN dam (Figure 5D).
2.2.2. Chemokines and colony-stimulating factors
At 24 h after poly (I:C) injection two-way ANOVA showed a main effect of MIA for MCP-1 [F(1,12)=8.08, p=0.0148] (Figure 5E) and MIP-1α [F(1,12)=36.71, p<0.0001], showing for the latter a decrease in the poly (I:C)/CTRL and poly (I:C)/FEN group as compared to veh/CTRL (p<0.05) and veh/FEN group (p<0.001), respectively (Figure 5F).
No differences among groups were detected for CSFs both at 6 and 24 h after poly (I:C) injection.
3. Discussion
The main findings of this study are: i) MIA alters cytokines, chemokines and CSFs levels in the maternal serum and fetal brain and ii) MIA-induced effects are partially reverted by fenofibrate.
Several studies have used MIA models to induce neurodevelopmental disorders in the offspring, but only a few reported a full immune response profile to poly (I:C) injection (see [29] for a review). Here, using the multiplex assay that maximizes the simultaneous detection of cytokines, chemokines and CSFs in a single sample, we identified and monitored several molecules both in the maternal serum and fetal brain, thus providing a complete maternal and fetal immune profile.
In agreement with other groups [30,34,35], our results show that MIA produces a time-dependent increase of some cytokines in maternal serum. Specifically, the levels of chemokine GRO/KC, MCP-1, and MIP-1α showed rapid increase at 6 h, while TNF-α and CSFs levels were significantly upregulated at 24 h, suggesting that their induction required a longer time to develop and are more persistent.
Chemokines are a family of small cytokines, initially studied because of their ability to manage the migration of leukocytes in the respective anatomical locations during inflammatory and homeostatic processes (see [36,37] for a review). Besides their contribution in inflammation, chemokines also play a role in brain development [34,38] and in neurodevelopment disorders, such as autism and schizophrenia [39,40,41,42]. Our findings are in line with those obtained by Arrode-Brusés (2012) [34] in mice administered with poly (I:C) at GD16, which reported increased levels of the chemokines GRO/KC, MCP-1, and MIP-1α, and of CSFs in maternal serum at 6 or 24 h.
Additionally, in agreement with our findings, several studies described increased levels of : i) TNF-α in maternal plasma/serum after poly (I:C) or LPS challenge both in mice and rats [31,32,43]; ii) CXCL1 (GRO/KC), but not IL-6, with a 3 h delay after subcutaneous injection of poly (I:C) (1.0 mg/kg) into the tail [44].
Contrary to our results, other groups showed increased levels of IL-1β, IL-2, IL-5, IL-6, IL-10 and IL-12 in maternal plasma at 3-6 h or 24 h after poly (I:C) administration in pregnant rodents [30,33,34,43,45,46]. However, no change or even a decrease of cytokines/chemokine has been described in maternal serum after poly (I:C) injection during pregnancy [29].
These discrepancies observed among studies could depend on the wide range of protocols that vary in the type, mode of delivery, timing, and dose of the immunogen used [47]. Several studies demonstrated that the serum cytokine responses in pregnant mice after systemic administration of poly (I:C) can be either increased (IL-6, IL-12p40) [34] or unaltered (i.e.: IL-6, IL-10, IL-17A) [48].
Considering its critical role as a molecular mediator of MIA, we expected the rise of IL-6 in the maternal serum after poly (I:C) injection. Murray et al. (2019) [49] demonstrated in rats that poly (I:C) induced a variable increase of IL-6 in the plasma at 3 h post-injection, and this response is strain-dependent. Indeed, high levels of IL-6 and fewer “non-responders” were consistently observed in Wistar rats, compared to Sprague Dawley or Lister Hooded rats. Thus, other rat strain and/or sample collection timing than ours could be optimal to study IL-6.
Lipopolysaccharide (LPS) and poly (I:C) maternal administration may increase, reduce or have no effect on fetal brain and placental cytokines, chemokines and CSFs levels [31,32,50,51,52]. Notably, the reduction or elevation of fetal brain cytokines might originate maternally or be attributable to endogenous production.
Out of the 14 pro- and anti-inflammatory cytokines examined in this study, only IL-12p70, IL-4, and IFN-γ were down-regulated in the fetal brain 6 h after poly (I:C), while among the chemokines analyzed, only MIP1-α and MCP-1 were decreased in the fetal brain of poly (I:C)/CTRL and poly (I:C)/FEN rats. Accordingly, maternal exposure to LPS increased placental and amniotic fluid TNF-α, but decreased it in fetal brain, indicating that maternal infection might directly impact cytokine levels in the developing fetal brain [32]. It could be argued that the observed downregulation of chemokines MIP1-α and MCP-1, together with the downregulation of the cytokine IL-12p70, IL-4, IFN-γ or TNF-α (all pro-inflammatory), may act as a compensatory mechanism occurring in the brain in response to maternal inflammation.
FEN is a clinically PPARα agonist, approved by both Food and Drug Administration (FDA) and European Medicines Agency (EMA) to treat patients with hypertriglyceridemia, primary hypercholesterolemia, or mixed dyslipidemia [53,54]. Additionally, several studies have revealed that FEN exhibits a robust anti-inflammatory effect on diseases of various systems, including the nervous system [55,56]. It exerts an anti-inflammatory effect through direct or indirect mechanisms, either secondary to its lipid-lowering effect (i.e., reduction of inflammation caused by hyperlipidemia in the circulatory system) or by activating PPARα and inhibiting downstream inflammatory signaling pathways. Specifically, FEN suppresses different inflammatory circuits, including NF-κB [57], mitogen-activated protein kinase (AMPK) [58], toll-like receptor 4 (TLR4) [59], or IL-1 or IL-6 [60]. Recently, our group demonstrated that prenatal FEN administration attenuated behavioral impairments and dopaminergic dysfunction in MIA offspring [19].
Our present findings showed that FEN treatment prevented the increase of MIA-induced cytokine TNF-α and chemokines MIP-1α and GRO/KC. Besides the neuroinflammatory processes, the above-mentioned chemokines are involved in cerebral ischemia/reperfusion injury [61,62], and in ASD [63], in which they have been associated with behavioral deficits [39,64,65,66].
Furthermore, MIA-induced altered CSF-s serum levels returned to basal values in the FEN group. Importantly, besides their role in the control, proliferation, and differentiation of hemopoietic cell population, evidence suggests that CSFs (GM-CSF, G-CSF and M-CSF) are also involved in inflammatory processes, including those taking place in the CNS [67,68,69].
By contrast, unexpectedly, we observed that FEN induced a significant increase of IL-17 and IL-18 in poly (I:C) treated dam compared to poly (I:C)/CTRL. Of note, although FEN displays anti-inflammatory effects in many diseases, it has also been reported to exert no effects [70,71] or even exacerbate the inflammatory response in an experimental mouse model of acute colitis [72] and in spontaneously hypertensive rats expressing the human C-reactive protein transgene [73]. In humans, FEN does not suppress the clinical or inflammatory response to low-dose endotoxin when administered at an effective dosage for heart disease or lipid disorders [71], suggesting that its systemic properties are limited.
Lastly, our results indicate that FEN does not reverse the observed decrease of proinflammatory cytokine and chemokine induced by MIA in the fetal brain.
To our knowledge, this is the first investigation of the effect of FEN treatment on cytokines, chemokines, and CSFs levels in the maternal serum and the fetal brain after MIA induced by poly (I:C) or LPS.
A limitation of this study could be the lack of information about the effects of FEN on cytokines, chemokines, and CSFs levels in the placenta during different time-windows during gestation (e.g., GD9 vs GD15). Further studies are needed to elucidate the cytokines transmission at the interface between the placenta and fetus. In conclusion, our findings suggest that PPARα might represent an attractive therapeutic target to attenuate the consequences of MIA.
4. Materials and Methods
4.1. Animals.
All animal care and experimental procedures were carried out in accordance with European Council directives (63/2010) and in compliance with the animal policies approved by the Italian Ministry of Health and the Ethics Committee for Animal Experiments (CESA, University of Cagliari). We made all efforts to minimise pain and suffering, and to reduce the number of animals used. Adult female Sprague-Dawley (SD) rats (250-300 g) (Envigo, Italy) were housed at temperature of 22 ± 2 °C and 60% humidity, under a 12 h light/dark cycle (light on at 7:00 a.m.) with food and water available ad libitum.
4.2. Drugs and Treatments.
Rats were mated at the age of 3 months. The first day after the copulation was defined as Gestational Day 0 (GD 0). Pregnant dams were randomly assigned to two experimental groups: the first group received a treatment consisting in a diet enriched with the PPARα agonist fenofibrate (0.2% w/w) ad libitum from GD 8 to the day of the sacrifice, while the second group received a control diet for the whole pregnancy.
Poly (I:C) was purchased from InvivoGen and dissolved in endotoxin-free saline solution. Pregnant dams on GD 15 were randomly assigned to receive either a single injection of poly (I:C) (4.0 mg/kg, i.v) or an equivalent volume of endotoxin-free saline solution (veh) in the lateral vein of the tail (for more details see SF).
4.3. Maternal blood and fetal brain collection.
6 or 24 h after poly (I:C) or saline injection, rats were deeply anesthetized with isoflurane (5%) (Merial, Toulouse, France) and sacrificed by decapitation. Maternal trunk blood was collected into 5 ml tube and allowed to clot at room temperature for 45 min. Fetuses were surgically delivered and killed by decapitation, and heads were placed in ice-cold PBS for immediate harvesting of the brains. Using a stereomicroscope, the brain was collected and immediately frozen in liquid nitrogen and stored at -80°C. An average of 15 pups per dam was obtained and a pool of 4 brain fetuses was used for cytokine/chemokine/CSFs measurements.
4.4. Blood and brain tissue processing.
Maternal blood was centrifuged at 1,000g for 15 min at 4°C. The supernatant was collected and recentrifuged at 10,000g for 10 min at 4°C and extracted serum was stored at -80°C for later cytokines/chemokines/CSFs measurements.
Tissue derived from a pool of 4 brain fetuses was processed using Bio-Plex Cell Lysis Kit (Bio-Rad Laboratories, Inc., USA), according to the manufacturer’s instructions. Specifically, brain frozen tissues were disrupted in Bioplex cell lysis buffer (BioRad, Hercules, CA, USA) containing factors 1 and 2 (protease and phosphatase inhibitors, respectively; BioRad) and the protease inhibitor phenyl-methylsulfonyl fluoride (500 mM; Sigma–Aldrich). Samples were homogenized on ice with a potter and homogenate frozen for at least 30 min at -80 °C. After thawing and sonication for 15 sec, samples were centrifuged at 4°C and 4,500g (Thermo Scientific Micro 17R Refrigerated Centrifuge) for 8-10 min. The supernatant was collected, and the protein content of each sample was determined using the BioRad Protein Assay (BioRad), with bovine serum albumin as a standard, according to the manufacturer’s protocol. The final supernatant was stored at -80°C until the day of the experiment and then used for cytokines/chemokines/CSFs measurements.
4.5. Cytokines, chemikines and CSFs measurements.
As previously described [18,74] sera and brain tissue homogenates were assayed for cytokines, chemokines and CSFs using a Luminex xMAP-based multiplex bead-based immunoassay, the Bio-Plex ProTM Rat Cytokine Group I Panel 23-Plex (Bio-Rad Laboratories, Inc., USA), which detects cytokines: [Interleukins (IL)-1α, IL-1β, IL-2, IL-4, IL-5, IL- 6, IL-7, IL-10, IL-12(p70), IL-13, IL-17A, IL-18, interferon (IFN)γ, tumor necrosis factor (TNF)α; chemokines: monocyte chemotactic protein-1 (MCP-1), macrophage inflammatory protein (MIP)1α, MIP-3α, regulated on activation normal T cells expressed and secreted (RANTES), keratinocyte derived chemokine (GRO/KC or CXCL1), and growth factors: granulocytes macrophage colony- stimulating factor (GM)-CSF, granulocyte (G)-CSF, macrophage (M)-CSF and vascular endothelial growth factor (VEGF)].
Assays were performed in 96-well plates; briefly, each assay plate layout consisted of eight lyophilized cytokines standards in duplicate, two blank wells and up to 19 brain fetus homogenate samples run at 300 μg proteins/well and 20 maternal serum samples (diluted 1:4 prior to assay). Each tissue and serum sample were run in duplicate and samples from veh/CTRL-, poly (I:C)/CTRL-, veh/FEN- and poly (I:C)/FEN-treated animals were analyzed in the same plate. All the wash steps were performed on a Bio-Plex Pro wash station at room temperature. A Bio-Plex MagPix Multiplex System by Luminex was used to read the plate and data were analyzed using BioPlex manager 4.1 software with 5-parametric logistic regression (5PL) curve fitting to determine the standard curve (pg/ml) from 8 standards in duplicate and extrapolate the sample concentrations from the standard curve. Only standards and samples with coefficients of variance under 5% were included in the analysis, and when readings fell under the detection limit (below the background value) they were excluded from analyses.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, SF: Maternal Immune Activation Model Reporting Guidelines Checklist.
Author Contributions
Conceptualization, M.S., R.M., M.P and M.P.C; methodology, R.M., M.S., V.O. and V.S.; validation, M.S. and R.M.; formal analysis, M.S, R.M and M.P.C.; investigation, M.S., R.M, V.O. and V.S.; writing-original draft preparation, M.S. and R.M.; writing – review and editing, M.P., M.P.C. and C.S.; supervision, M.P. and M.P.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by “FSC 2014–2020—Patto per lo Sviluppo della Regione Sardegna, Legge Regionale n. 7 del 7 agosto 2007 (Bando 2017)”, by “Progetti di Rilevante Interesse Nazionale” (PRIN) 2017 (2017YH3SXK), by a grant, with the code POC01_00069 from the Italian Ministry of University and Research (MUR) within the program Proof of Concept and by “FONDAZIONE di SARDEGNA (FdS) bando 2020 (F75F21001290007).
Institutional Review Board Statement
All animal care and experimental procedures were carried out in accordance with European Council directives (63/2010) and in compliance with the animal policies approved by the Italian Ministry of Health and the Ethics Committee for Animal Experiments (CESA, University of Cagliari).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article and supplementary material.
Acknowledgments
We gratefully acknowledge animal housing and care at CeSASt (Centro Servizi di Ateneo per gli Stabulari) of the University of Cagliari, Italy, and the CeSAR (Centro Servizi d'Ateneo per la Ricerca) of the University of Cagliari, Italy for the experiments performed with Bio-Plex 200 (BIO-Rad Laboratories S.r.l.).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Estes, M.L.; McAllister, A.K. Maternal Immune Activation: Implications for Neuropsychiatric Disorders. Science 2016, 353, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Han, V.X.; Patel, S.; Jones, H.F.; Nielsen, T.C.; Mohammad, S.S.; Hofer, M.J.; Gold, W.; Brilot, F.; Lain, S.J.; Nassar, N.; et al. Maternal Acute and Chronic Inflammation in Pregnancy Is Associated with Common Neurodevelopmental Disorders: A Systematic Review. Transl Psychiatry 2021, 11, 71. [Google Scholar] [CrossRef] [PubMed]
- Han, V.X.; Patel, S.; Jones, H.F.; Dale, R.C. Maternal Immune Activation and Neuroinflammation in Human Neurodevelopmental Disorders. Nat Rev Neurol 2021, 17, 564–579. [Google Scholar] [CrossRef]
- Shook, L.L.; Sullivan, E.L.; Lo, J.O.; Perlis, R.H.; Edlow, A.G. COVID-19 in Pregnancy: Implications for Fetal Brain Development. Trends Mol Med 2022, 28, 319–330. [Google Scholar] [CrossRef]
- Edlow, A.G.; Castro, V.M.; Shook, L.L.; Kaimal, A.J.; Perlis, R.H. Neurodevelopmental Outcomes at 1 Year in Infants of Mothers Who Tested Positive for SARS-CoV-2 During Pregnancy. JAMA Netw Open 2022, 5, e2215787. [Google Scholar] [CrossRef]
- Patterson, P.H. Immune Involvement in Schizophrenia and Autism: Etiology, Pathology and Animal Models. Behav Brain Res 2009, 204, 313–321. [Google Scholar] [CrossRef]
- Meyer, U. Developmental Neuroinflammation and Schizophrenia. Prog Neuropsychopharmacol Biol Psychiatry 2013, 42, 20–34. [Google Scholar] [CrossRef]
- Kreitz, S.; Zambon, A.; Ronovsky, M.; Budinsky, L.; Helbich, T.H.; Sideromenos, S.; Ivan, C.; Konerth, L.; Wank, I.; Berger, A.; et al. Maternal Immune Activation during Pregnancy Impacts on Brain Structure and Function in the Adult Offspring. Brain Behav Immun 2020, 83, 56–67. [Google Scholar] [CrossRef]
- Miller, A.H.; Haroon, E.; Raison, C.L.; Felger, J.C. Cytokine Targets in the Brain: Impact on Neurotransmitters and Neurocircuits. Depress Anxiety 2013, 30, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Mueller, F.S.; Polesel, M.; Richetto, J.; Meyer, U.; Weber-Stadlbauer, U. Mouse Models of Maternal Immune Activation: Mind Your Caging System! Brain Behav Immun 2018, 73, 643–660. [Google Scholar] [CrossRef]
- Allswede, D.M.; Yolken, R.H.; Buka, S.L.; Cannon, T.D. Cytokine Concentrations throughout Pregnancy and Risk for Psychosis in Adult Offspring: A Longitudinal Case-Control Study. Lancet Psychiatry 2020, 7, 254–261. [Google Scholar] [CrossRef]
- Carbone, E.; Buzzelli, V.; Manduca, A.; Leone, S.; Rava, A.; Trezza, V. Maternal Immune Activation Induced by Prenatal Lipopolysaccharide Exposure Leads to Long-Lasting Autistic-like Social, Cognitive and Immune Alterations in Male Wistar Rats. Int J Mol Sci 2023, 24, 3920. [Google Scholar] [CrossRef] [PubMed]
- Zuckerman, L.; Rehavi, M.; Nachman, R.; Weiner, I. Immune Activation during Pregnancy in Rats Leads to a Postpubertal Emergence of Disrupted Latent Inhibition, Dopaminergic Hyperfunction, and Altered Limbic Morphology in the Offspring: A Novel Neurodevelopmental Model of Schizophrenia. Neuropsychopharmacology 2003, 28, 1778–1789. [Google Scholar] [CrossRef]
- Meyer, U. Prenatal Poly(i:C) Exposure and Other Developmental Immune Activation Models in Rodent Systems. Biol Psychiatry 2014, 75, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Rose, D.R.; Careaga, M.; Van de Water, J.; McAllister, K.; Bauman, M.D.; Ashwood, P. Long-Term Altered Immune Responses Following Fetal Priming in a Non-Human Primate Model of Maternal Immune Activation. Brain Behav Immun 2017, 63, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Luchicchi, A.; Lecca, S.; Melis, M.; De Felice, M.; Cadeddu, F.; Frau, R.; Muntoni, A.L.; Fadda, P.; Devoto, P.; Pistis, M. Maternal Immune Activation Disrupts Dopamine System in the Offspring. Int J Neuropsychopharmacol 2016, 19, pyw007. [Google Scholar] [CrossRef] [PubMed]
- Santoni, M.; Frau, R.; Pistis, M. Transgenerational Sex-Dependent Disruption of Dopamine Function Induced by Maternal Immune Activation. Front Pharmacol 2022, 13, 821498. [Google Scholar] [CrossRef] [PubMed]
- Santoni, M.; Sagheddu, C.; Serra, V.; Mostallino, R.; Castelli, M.P.; Pisano, F.; Scherma, M.; Fadda, P.; Muntoni, A.L.; Zamberletti, E.; et al. Maternal Immune Activation Impairs Endocannabinoid Signaling in the Mesolimbic System of Adolescent Male Offspring. Brain Behav Immun 2023, 109, 271–284. [Google Scholar] [CrossRef] [PubMed]
- De Felice, M.; Melis, M.; Aroni, S.; Muntoni, A.L.; Fanni, S.; Frau, R.; Devoto, P.; Pistis, M. The PPARα Agonist Fenofibrate Attenuates Disruption of Dopamine Function in a Maternal Immune Activation Rat Model of Schizophrenia. CNS Neurosci Ther 2019, 25, 549–561. [Google Scholar] [CrossRef]
- Melis, M.; Carta, S.; Fattore, L.; Tolu, S.; Yasar, S.; Goldberg, S.R.; Fratta, W.; Maskos, U.; Pistis, M. Peroxisome Proliferator-Activated Receptors-Alpha Modulate Dopamine Cell Activity through Nicotinic Receptors. Biol Psychiatry 2010, 68, 256–264. [Google Scholar] [CrossRef]
- Skerrett, R.; Malm, T.; Landreth, G. Nuclear Receptors in Neurodegenerative Diseases. Neurobiol Dis 2014, 72 Pt A, 104–116. [Google Scholar] [CrossRef]
- Scheggi, S.; Melis, M.; De Felice, M.; Aroni, S.; Muntoni, A.L.; Pelliccia, T.; Gambarana, C.; De Montis, M.G.; Pistis, M. PPARα Modulation of Mesolimbic Dopamine Transmission Rescues Depression-Related Behaviors. Neuropharmacology 2016, 110, 251–259. [Google Scholar] [CrossRef]
- Duval, C.; Chinetti, G.; Trottein, F.; Fruchart, J.-C.; Staels, B. The Role of PPARs in Atherosclerosis. Trends Mol Med 2002, 8, 422–430. [Google Scholar] [CrossRef]
- Sagheddu, C.; Torres, L.H.; Marcourakis, T.; Pistis, M. Endocannabinoid-Like Lipid Neuromodulators in the Regulation of Dopamine Signaling: Relevance for Drug Addiction. Front Synaptic Neurosci 2020, 12, 588660. [Google Scholar] [CrossRef]
- Iannotti, F.A.; Vitale, R.M. The Endocannabinoid System and PPARs: Focus on Their Signalling Crosstalk, Action and Transcriptional Regulation. Cells 2021, 10, 586. [Google Scholar] [CrossRef]
- Decara, J.; Rivera, P.; López-Gambero, A.J.; Serrano, A.; Pavón, F.J.; Baixeras, E.; Rodríguez de Fonseca, F.; Suárez, J. Peroxisome Proliferator-Activated Receptors: Experimental Targeting for the Treatment of Inflammatory Bowel Diseases. Front Pharmacol 2020, 11, 730. [Google Scholar] [CrossRef]
- Jiang, Z.; Zamanian-Daryoush, M.; Nie, H.; Silva, A.M.; Williams, B.R.G.; Li, X. Poly(I-C)-Induced Toll-like Receptor 3 (TLR3)-Mediated Activation of NFkappa B and MAP Kinase Is through an Interleukin-1 Receptor-Associated Kinase (IRAK)-Independent Pathway Employing the Signaling Components TLR3-TRAF6-TAK1-TAB2-PKR. J Biol Chem 2003, 278, 16713–16719. [Google Scholar] [CrossRef] [PubMed]
- Reisinger, S.; Khan, D.; Kong, E.; Berger, A.; Pollak, A.; Pollak, D.D. The Poly(I:C)-Induced Maternal Immune Activation Model in Preclinical Neuropsychiatric Drug Discovery. Pharmacol Ther 2015, 149, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Hameete, B.C.; Fernández-Calleja, J.M.S.; de Groot, M.W.G.D.M.; Oppewal, T.R.; Tiemessen, M.M.; Hogenkamp, A.; de Vries, R.B.M.; Groenink, L. The Poly(I:C)-Induced Maternal Immune Activation Model; a Systematic Review and Meta-Analysis of Cytokine Levels in the Offspring. Brain Behav Immun Health 2021, 11, 100192. [Google Scholar] [CrossRef] [PubMed]
- Meyer, U.; Nyffeler, M.; Engler, A.; Urwyler, A.; Schedlowski, M.; Knuesel, I.; Yee, B.K.; Feldon, J. The Time of Prenatal Immune Challenge Determines the Specificity of Inflammation-Mediated Brain and Behavioral Pathology. J Neurosci 2006, 26, 4752–4762. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, J.H.; Jarskog, L.F.; Vadlamudi, S. Maternal Poly I:C Exposure during Pregnancy Regulates TNFα, BDNF, and NGF Expression in Neonatal Brain and the Maternal–Fetal Unit of the Rat. J Neuroimmunol 2005, 159, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Urakubo, A.; Jarskog, L.F.; Lieberman, J.A.; Gilmore, J.H. Prenatal Exposure to Maternal Infection Alters Cytokine Expression in the Placenta, Amniotic Fluid, and Fetal Brain. Schizophr Res 2001, 47, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Openshaw, R.L.; Kwon, J.; McColl, A.; Penninger, J.M.; Cavanagh, J.; Pratt, J.A.; Morris, B.J. JNK Signalling Mediates Aspects of Maternal Immune Activation: Importance of Maternal Genotype in Relation to Schizophrenia Risk. J Neuroinflammation 2019, 16, 18. [Google Scholar] [CrossRef] [PubMed]
- Arrode-Brusés, G.; Brusés, J.L. Maternal Immune Activation by Poly(I:C) Induces Expression of Cytokines IL-1β and IL-13, Chemokine MCP-1 and Colony Stimulating Factor VEGF in Fetal Mouse Brain. J Neuroinflammation 2012, 9, 605. [Google Scholar] [CrossRef] [PubMed]
- Ballendine, S.A.; Greba, Q.; Dawicki, W.; Zhang, X.; Gordon, J.R.; Howland, J.G. Behavioral Alterations in Rat Offspring Following Maternal Immune Activation and ELR-CXC Chemokine Receptor Antagonism during Pregnancy: Implications for Neurodevelopmental Psychiatric Disorders. Prog Neuropsychopharmacol Biol Psychiatry 2015, 57, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Cartier, L.; Hartley, O.; Dubois-Dauphin, M.; Krause, K.-H. Chemokine Receptors in the Central Nervous System: Role in Brain Inflammation and Neurodegenerative Diseases. Brain Res Brain Res Rev 2005, 48, 16–42. [Google Scholar] [CrossRef] [PubMed]
- Charo, I.F.; Ransohoff, R.M. The Many Roles of Chemokines and Chemokine Receptors in Inflammation. N Engl J Med 2006, 354, 610–621. [Google Scholar] [CrossRef]
- Deverman, B.E.; Patterson, P.H. Cytokines and CNS Development. Neuron 2009, 64, 61–78. [Google Scholar] [CrossRef]
- Ashwood, P.; Krakowiak, P.; Hertz-Picciotto, I.; Hansen, R.; Pessah, I.N.; Van De Water, J. Associations of Impaired Behaviors with Elevated Plasma Chemokines in Autism Spectrum Disorders. J Neuroimmunol 2011, 232, 196–199. [Google Scholar] [CrossRef]
- Reale, M.; Patruno, A.; De Lutiis, M.A.; Pesce, M.; Felaco, M.; Di Giannantonio, M.; Di Nicola, M.; Grilli, A. Dysregulation of Chemo-Cytokine Production in Schizophrenic Patients versus Healthy Controls. BMC Neurosci 2011, 12, 13. [Google Scholar] [CrossRef] [PubMed]
- Stuart, M.J.; Baune, B.T. Chemokines and Chemokine Receptors in Mood Disorders, Schizophrenia, and Cognitive Impairment: A Systematic Review of Biomarker Studies. Neurosci Biobehav Rev 2014, 42, 93–115. [Google Scholar] [CrossRef]
- Cieślik, M.; Gąssowska-Dobrowolska, M.; Jęśko, H.; Czapski, G.A.; Wilkaniec, A.; Zawadzka, A.; Dominiak, A.; Polowy, R.; Filipkowski, R.K.; Boguszewski, P.M.; et al. Maternal Immune Activation Induces Neuroinflammation and Cortical Synaptic Deficits in the Adolescent Rat Offspring. Int J Mol Sci 2020, 21, 4097. [Google Scholar] [CrossRef]
- Ding, S.; Hu, Y.; Luo, B.; Cai, Y.; Hao, K.; Yang, Y.; Zhang, Y.; Wang, X.; Ding, M.; Zhang, H.; et al. Age-Related Changes in Neuroinflammation and Prepulse Inhibition in Offspring of Rats Treated with Poly I:C in Early Gestation. Behav Brain Funct 2019, 15, 3. [Google Scholar] [CrossRef] [PubMed]
- Kamerman, P.; Skosana, M.; Loram, L.; Mitchell, B.; Weber, J. Fever and Inflammatory Cytokine Response in Rats Injected Subcutaneously with Viral Double-Stranded RNA Analog, Polyinosinic:Polycytidylic Acid (Poly-I:C). J Therm Biol 2011, 36, 397–402. [Google Scholar] [CrossRef]
- Koga, K.; Cardenas, I.; Aldo, P.; Abrahams, V.M.; Peng, B.; Fill, S.; Romero, R.; Mor, G. Activation of TLR3 in the Trophoblast Is Associated with Preterm Delivery. Am J Reprod Immunol 2009, 61, 196–212. [Google Scholar] [CrossRef] [PubMed]
- McColl, E.R.; Piquette-Miller, M. Poly(I:C) Alters Placental and Fetal Brain Amino Acid Transport in a Rat Model of Maternal Immune Activation. Am J Reprod Immunol 2019, 81, e13115. [Google Scholar] [CrossRef] [PubMed]
- Kentner, A.C.; Bilbo, S.D.; Brown, A.S.; Hsiao, E.Y.; McAllister, A.K.; Meyer, U.; Pearce, B.D.; Pletnikov, M.V.; Yolken, R.H.; Bauman, M.D. Maternal Immune Activation: Reporting Guidelines to Improve the Rigor, Reproducibility, and Transparency of the Model. Neuropsychopharmacology 2019, 44, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Valtanen, P.; Diermen, B.A.; Lakhan, N.; Lousberg, E.L.; Robertson, S.A.; Hayball, J.D.; Diener, K.R. Maternal Host Responses to Poly(I:C) during Pregnancy Leads to Both Dysfunctional Immune Profiles and Altered Behaviour in the Offspring. Am J Reprod Immunol 2020, 84. [Google Scholar] [CrossRef] [PubMed]
- Murray, E.; Sharma, R.; Smith, K.B.; Mar, K.D.; Barve, R.; Lukasik, M.; Pirwani, A.F.; Malette-Guyon, E.; Lamba, S.; Thomas, B.J.; et al. Probiotic Consumption during Puberty Mitigates LPS-Induced Immune Responses and Protects against Stress-Induced Depression- and Anxiety-like Behaviors in Adulthood in a Sex-Specific Manner. Brain Behav Immun 2019, 81, 198–212. [Google Scholar] [CrossRef] [PubMed]
- Ashdown, H.; Dumont, Y.; Ng, M.; Poole, S.; Boksa, P.; Luheshi, G.N. The Role of Cytokines in Mediating Effects of Prenatal Infection on the Fetus: Implications for Schizophrenia. Mol Psychiatry 2006, 11, 47–55. [Google Scholar] [CrossRef]
- Carpentier, P.A.; Dingman, A.L.; Palmer, T.D. Placental TNF-α Signaling in Illness-Induced Complications of Pregnancy. Am J Pathol 2011, 178, 2802–2810. [Google Scholar] [CrossRef] [PubMed]
- O’Loughlin, E.; Pakan, J.M.P.; Yilmazer-Hanke, D.; McDermott, K.W. Acute in Utero Exposure to Lipopolysaccharide Induces Inflammation in the Pre- and Postnatal Brain and Alters the Glial Cytoarchitecture in the Developing Amygdala. J Neuroinflammation 2017, 14, 212. [Google Scholar] [CrossRef] [PubMed]
- U.S. Department of Health and Human Services Food and Drug Administration Center for Drug Evaluation and Research. Drug Approvals and Databases Tricor (Fenofibrate) Tablets https://www.accessdata.fda.gov/drugsatfda_docs/nda/2004/021656s000_TricorTOC.cfm. (accessed on 30 May 2023).
- European Medicines Agency Science Medicines Health European Commission final decision Fibrates https://www.ema.europa.eu/en/medicines/human/referrals/fibrates. (accessed on 30 May 2023).
- Jin, L.; Hua, H.; Ji, Y.; Jia, Z.; Peng, M.; Huang, S. Anti-Inflammatory Role of Fenofibrate in Treating Diseases. Biomol Biomed 2023, 23, 376–391. [Google Scholar] [CrossRef]
- Sagheddu, C.; Melis, M.; Muntoni, A.L.; Pistis, M. Repurposing Peroxisome Proliferator-Activated Receptor Agonists in Neurological and Psychiatric Disorders. Pharmaceuticals (Basel) 2021, 14, 1025. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Ren, X.; Han, T.; Chen, Y.; Qiu, H.; Liu, W.; Hu, Y. Fenofibrate Attenuates Fatty Acid-Induced Islet β-Cell Dysfunction and Apoptosis via Inhibiting the NF-ΚB/MIF Dependent Inflammatory Pathway. Metabolism 2017, 77, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Tomizawa, A.; Hattori, Y.; Inoue, T.; Hattori, S.; Kasai, K. Fenofibrate Suppresses Microvascular Inflammation and Apoptosis through Adenosine Monophosphate-Activated Protein Kinase Activation. Metabolism 2011, 60, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Dai, F.; Jiang, T.; Bao, Y.-Y.; Chen, G.-J.; Chen, L.; Zhang, Q.; Lu, Y.-X. Fenofibrate Improves High-Fat Diet-Induced and Palmitate-Induced Endoplasmic Reticulum Stress and Inflammation in Skeletal Muscle. Life Sci 2016, 157, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, R.M.; Baugé, E.; Staels, B.; Gervois, P. Systemic and Distal Repercussions of Liver-Specific Peroxisome Proliferator-Activated Receptor-Alpha Control of the Acute-Phase Response. Endocrinology 2008, 149, 3215–3223. [Google Scholar] [CrossRef]
- Guo, Y.-Q.; Zheng, L.-N.; Wei, J.-F.; Hou, X.-L.; Yu, S.-Z.; Zhang, W.-W.; Jing, J.-M. Expression of CCL2 and CCR2 in the Hippocampus and the Interventional Roles of Propofol in Rat Cerebral Ischemia/Reperfusion. Exp Ther Med 2014, 8, 657–661. [Google Scholar] [CrossRef]
- Song, H.; Zhang, X.; Chen, R.; Miao, J.; Wang, L.; Cui, L.; Ji, H.; Liu, Y. Cortical Neuron-Derived Exosomal MicroRNA-181c-3p Inhibits Neuroinflammation by Downregulating CXCL1 in Astrocytes of a Rat Model with Ischemic Brain Injury. Neuroimmunomodulation 2019, 26, 217–233. [Google Scholar] [CrossRef]
- Suzuki, K.; Matsuzaki, H.; Iwata, K.; Kameno, Y.; Shimmura, C.; Kawai, S.; Yoshihara, Y.; Wakuda, T.; Takebayashi, K.; Takagai, S.; et al. Plasma Cytokine Profiles in Subjects with High-Functioning Autism Spectrum Disorders. PLoS One 2011, 6, e20470. [Google Scholar] [CrossRef]
- Shen, Y.; Ou, Ji.; Liu, M.; Shi, L.; Li, Y.; Xiao, L.; Dong, H.; Zhang, F.; Xia, K.; Zhao, J. Altered Plasma Levels of Chemokines in Autism and Their Association with Social Behaviors. Psychiatry Res 2016, 244, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Masi, A.; Breen, E.J.; Alvares, G.A.; Glozier, N.; Hickie, I.B.; Hunt, A.; Hui, J.; Beilby, J.; Ravine, D.; Wray, J.; et al. Cytokine Levels and Associations with Symptom Severity in Male and Female Children with Autism Spectrum Disorder. Mol Autism 2017, 8, 63. [Google Scholar] [CrossRef] [PubMed]
- Heuer, L.S.; Croen, L.A.; Jones, K.L.; Yoshida, C.K.; Hansen, R.L.; Yolken, R.; Zerbo, O.; DeLorenze, G.; Kharrazi, M.; Ashwood, P.; et al. An Exploratory Examination of Neonatal Cytokines and Chemokines as Predictors of Autism Risk: The Early Markers for Autism Study. Biol Psychiatry 2019, 86, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Chitu, V.; Stanley, E.R. Colony-Stimulating Factor-1 in Immunity and Inflammation. Curr Opin Immunol 2006, 18, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.A. Colony-Stimulating Factors in Inflammation and Autoimmunity. Nat Rev Immunol 2008, 8, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Becher, B.; Tugues, S.; Greter, M. GM-CSF: From Growth Factor to Central Mediator of Tissue Inflammation. Immunity 2016, 45, 963–973. [Google Scholar] [CrossRef] [PubMed]
- Paiva, A.A.; Raposo, H.F.; Wanschel, A.C.B.A.; Nardelli, T.R.; Oliveira, H.C.F. Apolipoprotein CIII Overexpression-Induced Hypertriglyceridemia Increases Nonalcoholic Fatty Liver Disease in Association with Inflammation and Cell Death. Oxid Med Cell Longev 2017, 2017, 1838679. [Google Scholar] [CrossRef] [PubMed]
- Mulvey, C.K.; Ferguson, J.F.; Tabita-Martinez, J.; Kong, S.; Shah, R.Y.; Patel, P.N.; Master, S.R.; Usman, M.H.U.; Propert, K.J.; Shah, R.; et al. Peroxisome Proliferator–Activated Receptor-α Agonism With Fenofibrate Does Not Suppress Inflammatory Responses to Evoked Endotoxemia. J Am Heart Assoc 2012, 1, e002923. [Google Scholar] [CrossRef]
- Gu, X.; Song, Y.; Chai, Y.; Lu, F.; Gonzalez, F.J.; Fan, G.; Qi, Y. GC-MS Metabolomics on PPARα-Dependent Exacerbation of Colitis. Mol Biosyst 2015, 11, 1329–1337. [Google Scholar] [CrossRef]
- Škop, V.; Trnovská, J.; Oliyarnyk, O.; Marková, I.; Malínská, H.; Kazdová, L.; Zídek, V.; Landa, V.; Mlejnek, P.; Šimáková, M.; et al. Hepatotoxic Effects of Fenofibrate in Spontaneously Hypertensive Rats Expressing Human C-Reactive Protein. Physiol Res 2016, 65, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Margiani, G.; Castelli, M.P.; Pintori, N.; Frau, R.; Ennas, M.G.; Zottola, A.C.P.; Orrù, V.; Serra, V.; Fiorillo, E.; Fadda, P.; et al. Correction to: Adolescent Self-administration of the Synthetic Cannabinoid Receptor Agonist JWH-018 Induces Neurobiological and Behavioral Alterations in Adult Male Mice. Psychopharmacology (Berl) 2023, 240, 1199–1199. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Representation of the experimental protocol. MIA model consisted of a single i.v. injection of poly (I:C) (4 mg/kg) or veh (sterile pyrogen-free saline) at GD15. CTRL or FEN (0.2% w/w) treatment from GD8 to the day of sacrifice at 6 h or 24 h after injection, when maternal serum and fetal brains were collected. Abbreviations: Veh = vehicle; CTRL = Control treatment; FEN = Fenofibrate treatment.
Figure 1.
Representation of the experimental protocol. MIA model consisted of a single i.v. injection of poly (I:C) (4 mg/kg) or veh (sterile pyrogen-free saline) at GD15. CTRL or FEN (0.2% w/w) treatment from GD8 to the day of sacrifice at 6 h or 24 h after injection, when maternal serum and fetal brains were collected. Abbreviations: Veh = vehicle; CTRL = Control treatment; FEN = Fenofibrate treatment.
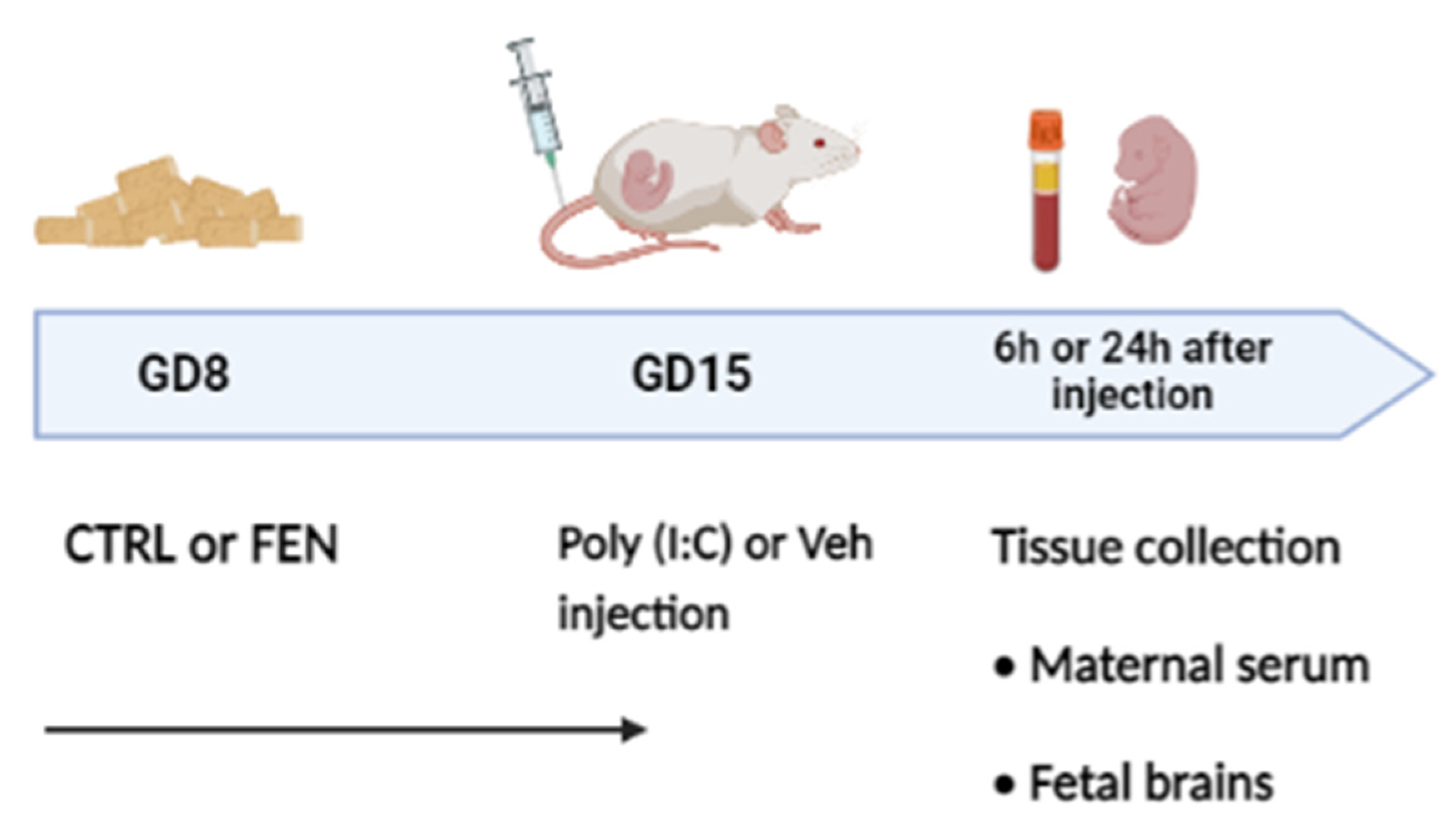
Figure 2.
Effect of fenofibrate treatment and MIA on the cytokines and chemokines expression in the maternal serum. Levels of cytokines and chemokines were evaluated by Bio-Plex ProTM Rat Cytokine Group I Panel 23-Plex (see materials and methods section) 6 h (left) and 24 h (right) after poly (I:C) injection. Data, expressed as pg/ml concentrations relative to standards containing known amounts of target proteins, are mean ± SEM for each cytokine and chemokine. *p<0.05; **p<0.01; ***p<0.001 Veh/CTRL: n = 4-5; Poly (I:C)/CTRL: n = 4-5; Veh/FEN: n = 4-5; Poly (I:C)/FEN: n = 4-5. (Two-way ANOVA; Tukey’s post hoc test for M x T; Bonferroni’s post hoc test for M and T). M, significant effect of MIA; T, significant effect of the treatment; M x T, significant interaction of MxT.
Figure 2.
Effect of fenofibrate treatment and MIA on the cytokines and chemokines expression in the maternal serum. Levels of cytokines and chemokines were evaluated by Bio-Plex ProTM Rat Cytokine Group I Panel 23-Plex (see materials and methods section) 6 h (left) and 24 h (right) after poly (I:C) injection. Data, expressed as pg/ml concentrations relative to standards containing known amounts of target proteins, are mean ± SEM for each cytokine and chemokine. *p<0.05; **p<0.01; ***p<0.001 Veh/CTRL: n = 4-5; Poly (I:C)/CTRL: n = 4-5; Veh/FEN: n = 4-5; Poly (I:C)/FEN: n = 4-5. (Two-way ANOVA; Tukey’s post hoc test for M x T; Bonferroni’s post hoc test for M and T). M, significant effect of MIA; T, significant effect of the treatment; M x T, significant interaction of MxT.
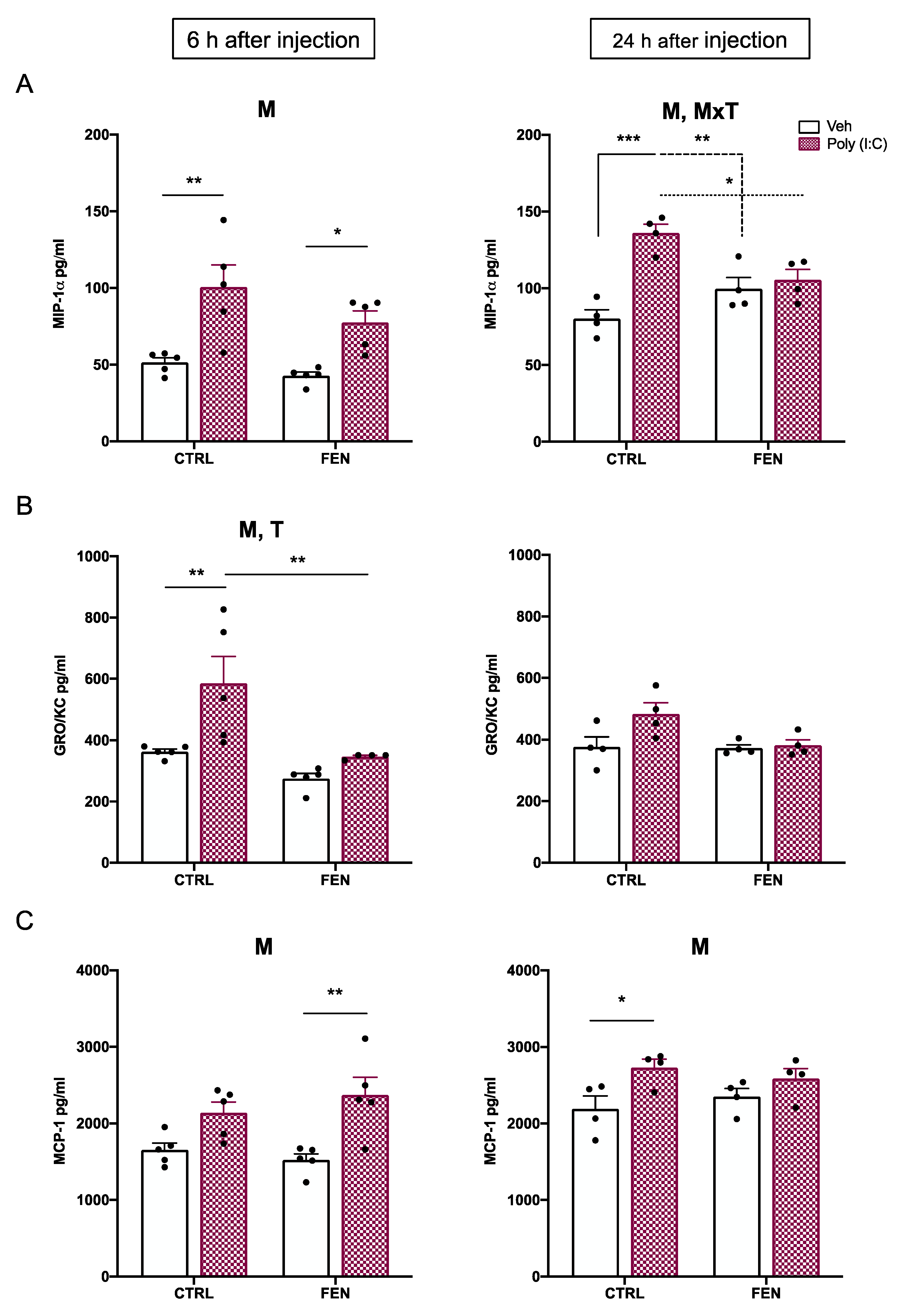
Figure 3.
Effect of fenofibrate treatment and MIA on cytokines expression in the maternal serum. Levels of cytokines were evaluated by Bio-Plex ProTM Rat Cytokine Group I Panel 23-Plex (see materials and methods section) 6 h (left) and 24 h (right) after poly (I:C) injection. Data, expressed as pg/ml concentrations relative to standards containing known amounts of target proteins, are mean ± SEM for each cytokine. *p<0.05; **p<0.01; Veh/CTRL: n = 4-5; Poly (I:C)/CTRL: n = 4-5; Veh/FEN: n = 4-5; Poly (I:C)/FEN: n = 4-5. (Two-way ANOVA; Tukey’s post hoc test for MxT; Bonferroni’s post hoc test for T). M, significant effect of MIA; T, significant effect of the treatment; M x T, significant interaction of MxT.
Figure 3.
Effect of fenofibrate treatment and MIA on cytokines expression in the maternal serum. Levels of cytokines were evaluated by Bio-Plex ProTM Rat Cytokine Group I Panel 23-Plex (see materials and methods section) 6 h (left) and 24 h (right) after poly (I:C) injection. Data, expressed as pg/ml concentrations relative to standards containing known amounts of target proteins, are mean ± SEM for each cytokine. *p<0.05; **p<0.01; Veh/CTRL: n = 4-5; Poly (I:C)/CTRL: n = 4-5; Veh/FEN: n = 4-5; Poly (I:C)/FEN: n = 4-5. (Two-way ANOVA; Tukey’s post hoc test for MxT; Bonferroni’s post hoc test for T). M, significant effect of MIA; T, significant effect of the treatment; M x T, significant interaction of MxT.
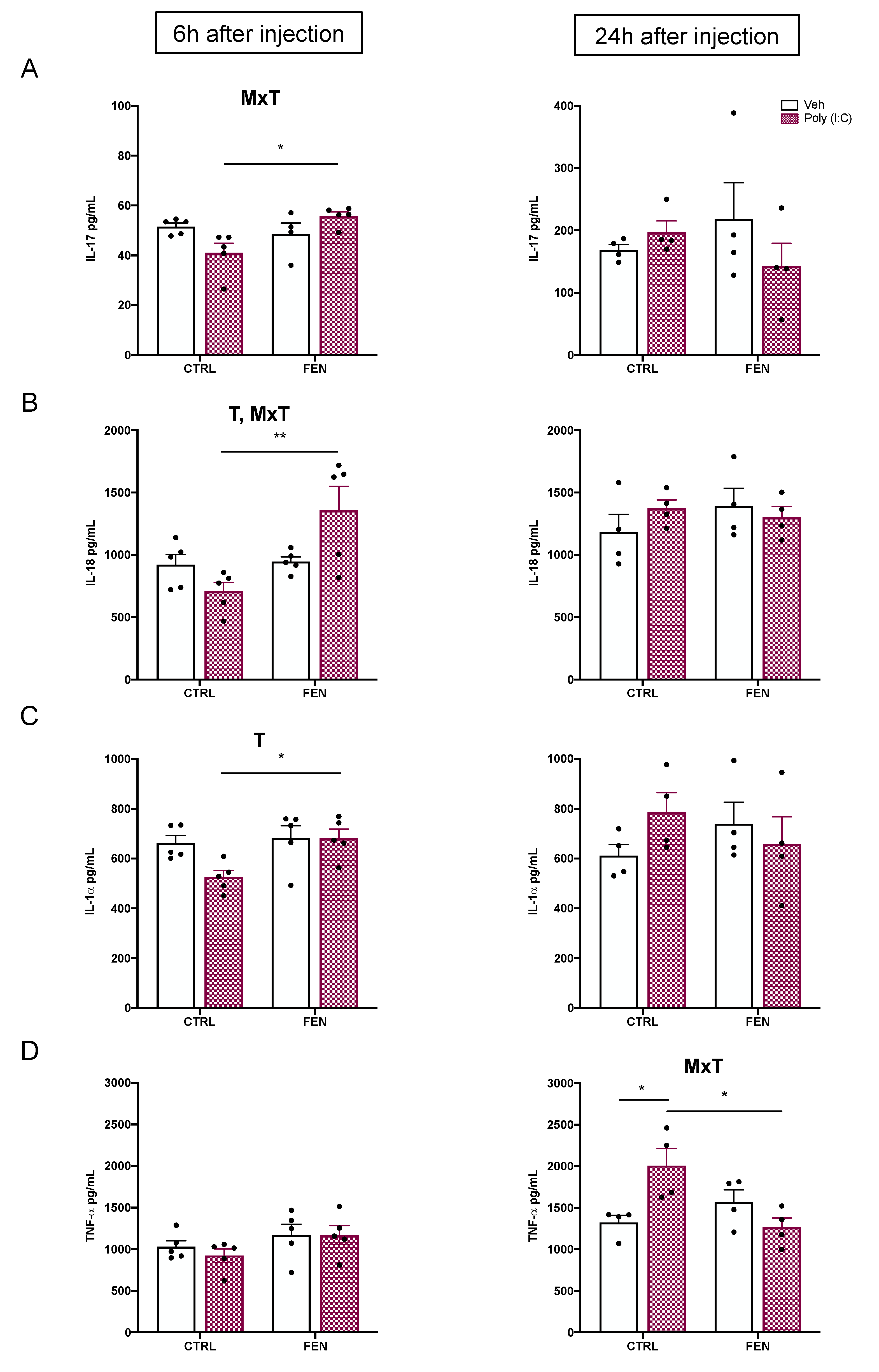
Figure 4.
Effect of fenofibrate treatment and MIA on the colony-stimulating factors expression in the maternal serum. Levels of colony-stimulating factors were evaluated by Bio-Plex ProTM Rat Cytokine Group I Panel 23-Plex (see materials and methods section) 6 h (left) and 24 h (right) after poly (I:C) injection. Data, expressed as pg/ml concentrations relative to standards containing known amounts of target proteins, are mean ± SEM for each colony stimulating factor. *p<0.05 Veh/CTRL: n = 4-5; Poly (I:C)/CTRL: n = 4-5; Veh/FEN: n = 4-5; Poly (I:C)/FEN: n = 4-5. (Two-way ANOVA; Tukey’s post hoc test for MxT). M, significant effect of MIA; T, significant effect of the treatment; M x T, significant interaction of MxT.
Figure 4.
Effect of fenofibrate treatment and MIA on the colony-stimulating factors expression in the maternal serum. Levels of colony-stimulating factors were evaluated by Bio-Plex ProTM Rat Cytokine Group I Panel 23-Plex (see materials and methods section) 6 h (left) and 24 h (right) after poly (I:C) injection. Data, expressed as pg/ml concentrations relative to standards containing known amounts of target proteins, are mean ± SEM for each colony stimulating factor. *p<0.05 Veh/CTRL: n = 4-5; Poly (I:C)/CTRL: n = 4-5; Veh/FEN: n = 4-5; Poly (I:C)/FEN: n = 4-5. (Two-way ANOVA; Tukey’s post hoc test for MxT). M, significant effect of MIA; T, significant effect of the treatment; M x T, significant interaction of MxT.
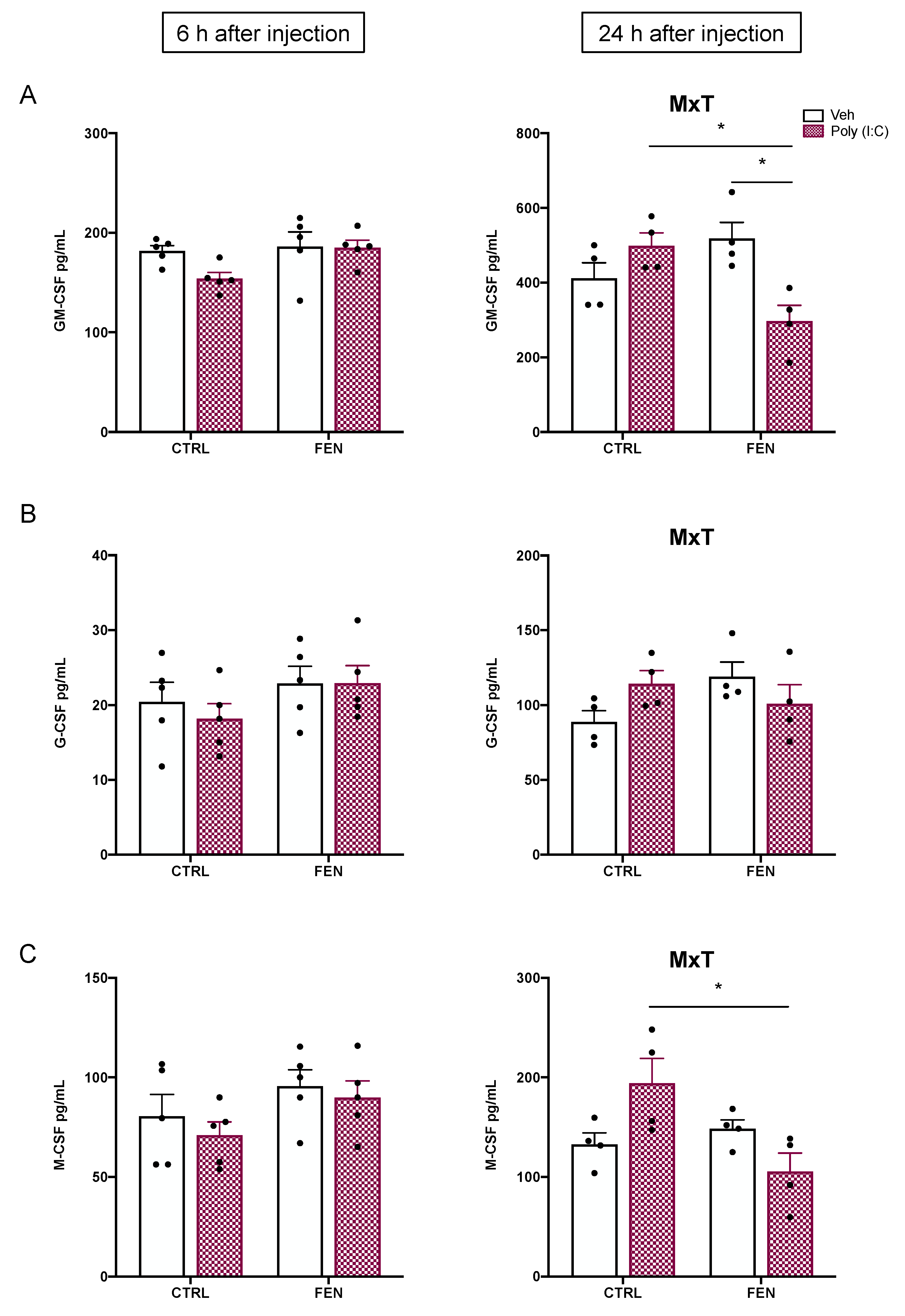
Figure 5.
Effect of fenofibrate treatment and MIA on the cytokines and chemokines expression in the prenatal brain homogenates. Levels of cytokines and chemokines were evaluated by Bio-Plex ProTM Rat Cytokine Group I Panel 23-Plex (see materials and methods section) 6 h (top) and 24 h (bottom) after poly (I:C) injection. Data, expressed as pg/ml concentrations relative to standards containing known amounts of target proteins, are mean ± SEM for each cytokine and chemokine. *p<0.05; **p<0.01; Veh/CTRL: n = 4; Poly (I:C)/CTRL: n = 4; Veh/FEN: n = 4; Poly (I:C)/FEN: n = 4-5. (Two-way ANOVA; Tukey’s post hoc test for M x T; Bonferroni’s post hoc test for M) M, significant effect of MIA; T, significant effect of the treatment; M x T, significant interaction of MxT.
Figure 5.
Effect of fenofibrate treatment and MIA on the cytokines and chemokines expression in the prenatal brain homogenates. Levels of cytokines and chemokines were evaluated by Bio-Plex ProTM Rat Cytokine Group I Panel 23-Plex (see materials and methods section) 6 h (top) and 24 h (bottom) after poly (I:C) injection. Data, expressed as pg/ml concentrations relative to standards containing known amounts of target proteins, are mean ± SEM for each cytokine and chemokine. *p<0.05; **p<0.01; Veh/CTRL: n = 4; Poly (I:C)/CTRL: n = 4; Veh/FEN: n = 4; Poly (I:C)/FEN: n = 4-5. (Two-way ANOVA; Tukey’s post hoc test for M x T; Bonferroni’s post hoc test for M) M, significant effect of MIA; T, significant effect of the treatment; M x T, significant interaction of MxT.
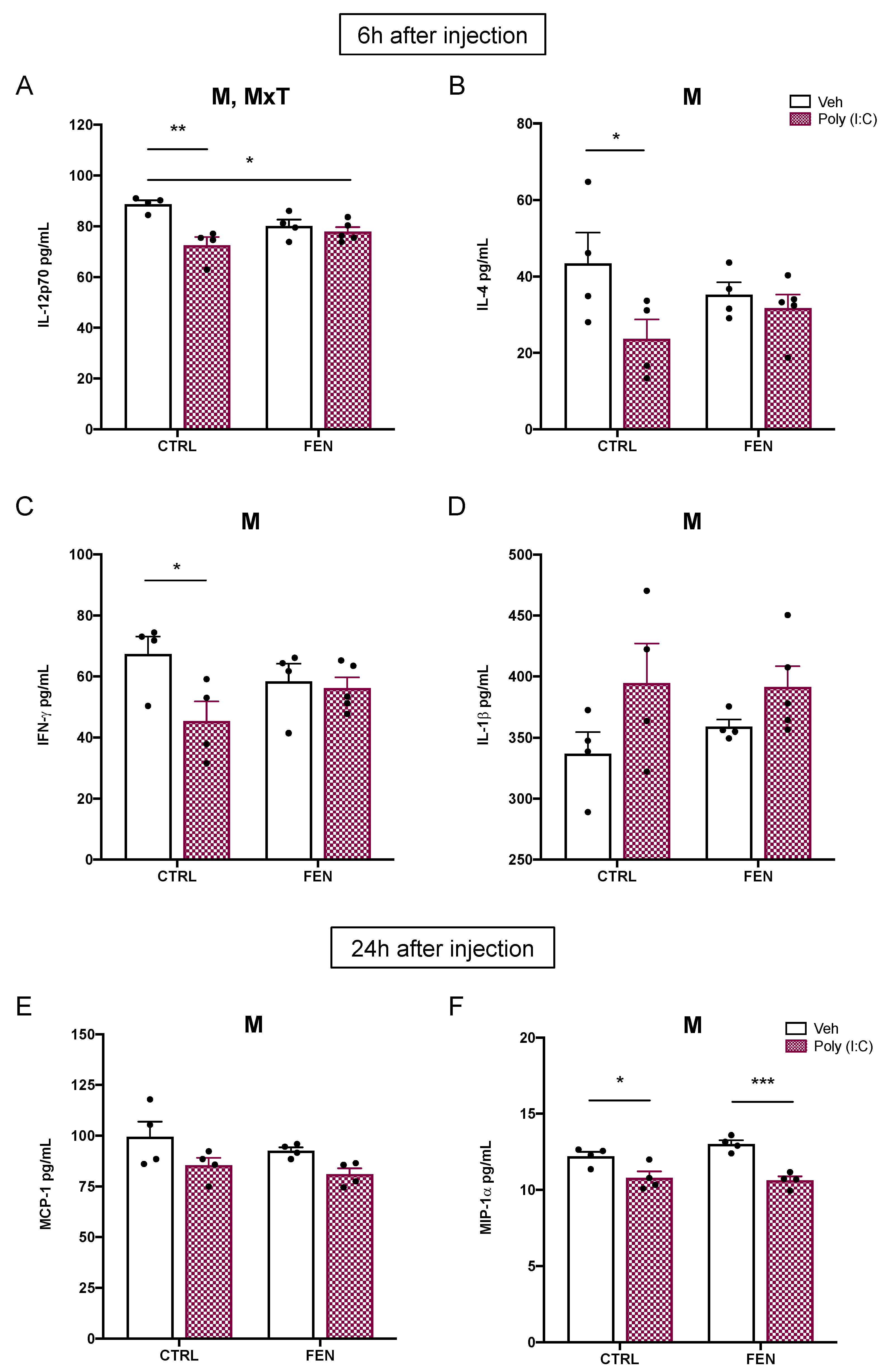

Table 1.
Cytokines, chemokines and colony-stimulating factors concentration in maternal serum. Data are mean ± SEM calculated from one experiment performed in duplicate (n = 4-5 rats per group); values are expressed as pg/ml concentrations relative to standards containing known amounts of target proteins.
Table 1.
Cytokines, chemokines and colony-stimulating factors concentration in maternal serum. Data are mean ± SEM calculated from one experiment performed in duplicate (n = 4-5 rats per group); values are expressed as pg/ml concentrations relative to standards containing known amounts of target proteins.
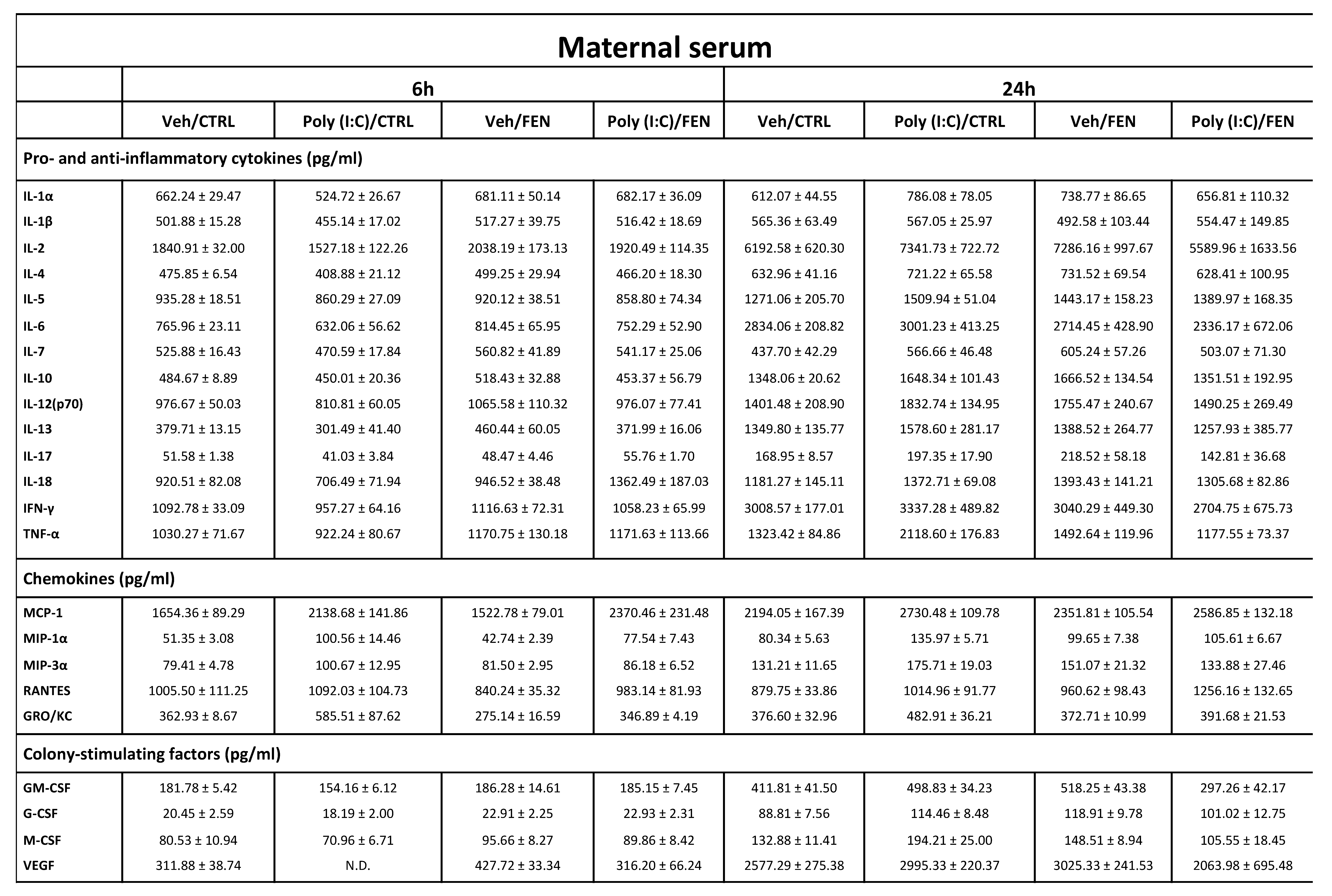 |
ND: not detectable, cytokines, chemokines or colony-stimulating factors concentrations under the limit of detection. Veh: vehicle; CTRL: control treatment; FEN: Fenofibrate treatment.
Table 2.
Cytokines, chemokines and colony-stimulating factors concentration in prenatal brain homogenates. Data, are mean ± SEM calculated from one experiment performed in duplicate (n = 4-5 rats per group); values are expressed as pg/ml concentrations relative to standards containing known amounts of target proteins.
Table 2.
Cytokines, chemokines and colony-stimulating factors concentration in prenatal brain homogenates. Data, are mean ± SEM calculated from one experiment performed in duplicate (n = 4-5 rats per group); values are expressed as pg/ml concentrations relative to standards containing known amounts of target proteins.
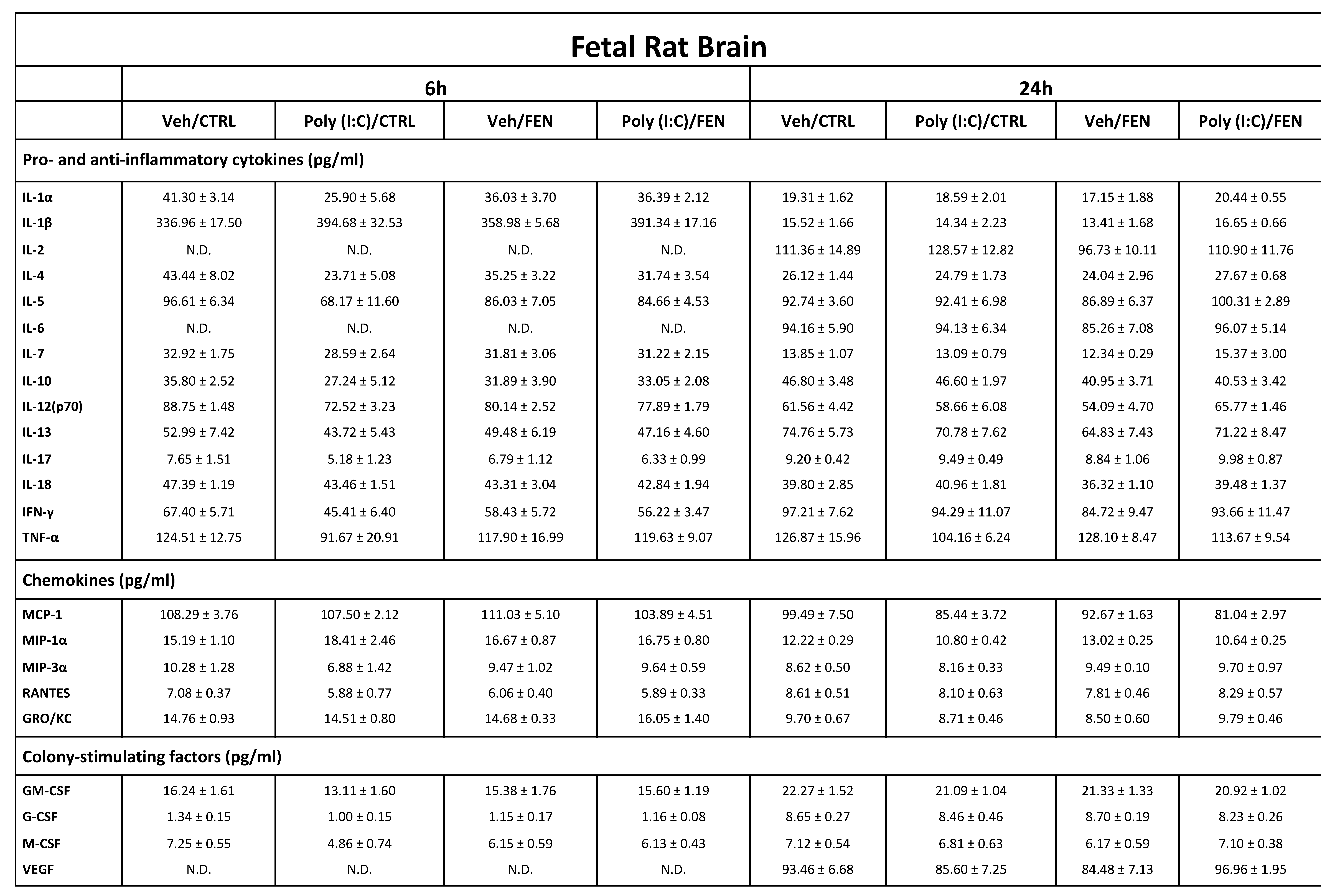 |
ND: not detectable, cytokines, chemokines or colony-stimulating factors concentrations under the limit of detection. Veh: vehicle; CTRL: control treatment; FEN: Fenofibrate treatment.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



