
Submitted:
07 June 2023
Posted:
08 June 2023
You are already at the latest version
Abstract
Cosmetics have various characters, but there have been no studies which classified the properties of cosmetics based on their effects on skin-domesticated bacteria. In this study, we established an anaerobic culture and short-chain fatty acid measurement system using Staphylococcus epidermidis, a representative bacterium of indigenous skin bacteria. Furthermore, this system was used to extract and classify the characteristics of 9 cosmetics. Cosmetics containing ferments of rice and soy were clustered in the vicinity, although one cosmetic containing soy ferment was clustered distant from other similar cosmetics. Cosmetics from South Asia and those containing natural plant extracts were clustered in the vicinity. These results show that this method can be utilized as a cosmetics pattern map, and will provide a method to prevent growth of opportunistic bacteria.
Keywords:
cosmetics
; skin microbiota
; Staphylococcus epidermidis
; lactic acid
; PCA
1. Introduction
Cosmetics are applied to the skin for the purpose of enhancement of beauty. Before the establishment of modern chemical industry, there were few concepts of make-up and skin care, and most cosmetics used plant-derived ingredients or minerals. In Japan, make-up products such as white powder, safflower, and eyebrow ink, and plant-derived skin care products such as loofah water and flower dew [1,2] began to become popular among the general public during the Edo period (AD 1600-1868). The establishment of the chemical industry in the 18th and early 19th centuries led to the use of glycerin as a moisturizing ingredient and the spread of cosmetics made with chemical products. For this reason, the cosmetics industry established in the 20th century, combined with the petrochemical industry established in the early 20th century, almost exclusively used petroleum-derived chemical products. However, such chemical products have not been applied to human skin in the past, and due to the problem of petroleum-derived trace constituents, they have been causing various health problems since the 1970s. For example, skin diseases such as melanosis [3] and chemical sensitivity have been reported. As a result, from the 1980s, many cosmetics using naturally derived ingredients began to appear on the market. Furthermore, taking advantage of traditional Japanese fermentation technology, many cosmetics containing fermented products were developed in Japan [4,5]. However, there have been no studies to date that have revealed the effects of cosmetics containing such natural or fermentation-derived ingredients on skin bacteria, let alone cosmetics made with chemical products. There are cosmetics from various cultural backgrounds in the world, and they have completely different characteristics. However, with the development of logistics, cosmetics from all over the world are now available, and there have been no studies comparing the effects of these cosmetics to skin microbiota.
Properties of cosmetics vary dependent on the cultures of countries. For example, in Japan, many cosmetics are based on fermented products because of the country's long tradition of fermented foods. On the other hand, in South Asia, cosmetics based on medicinal herbs are considered common due to the Ayurvedic tradition of using medicinal herbs. In Europe, where the cosmetics industry first emerged, many cosmetics are chemical-based, and many are dairy- and plant-based due to the traditions of dairy farming and Unani medicine. However, there are no studies that report such regional characteristics because there is no system to identify the properties of cosmetics.
Skin is said to be "the largest organ of the human body " [6] since it covers our entire body, occupying a total area of about 1.6 square meters (for an adult) and weighing about 15-16% of our body weight. Therefore, it is believed that maintaining healthy skin leads to the maintenance of the overall health of the body. Human skin is inhabited by about 1,000 species of bacteria, which are referred to as dermal commensal bacteria. The skin commensal flora is said to be the second largest flora in the human body after the intestinal flora [7], and it is classified into three major categories according to their functions for the host. They are classified into three categories according to their function on the host: beneficial bacteria, opportunistic bacteria that change their role depending on the balance between beneficial and harmful bacteria on the skin, and harmful bacteria that cause skin problems. Among these, Staphylococcus epidermidis, which is the most ubiquitous bacterium on human skin and is known as the representative of good bacteria, degrades sweat and sebum on the skin and produces glycerin and fatty acids. The glycerin produced by these indigenous skin bacteria strengthens the skin barrier function [8] and the fatty acids keep the pH level of the epidermis acidic and inhibit the growth of bad bacteria [9]. Other short-chain fatty acids derived from indigenous skin bacteria have also been reported to have various effects on the skin. For example, Lactic acid has been reported to have a peeling effect on the skin and a softening effect on the stratum corneum [10]. In addition, it has recently been reported that lactate produced by indigenous skin bacteria [11] and Staphylococcus aureus act on the immune response of the host through the production of IL-10 and control skin pathogens [12]. Succinic acid inhibits the growth of Propionibacterium acnes, a causative agent of acne [13], and propionic acid inhibits the growth of Staphylococcus aureus, a representative of hazardous bacteria [14]. On the other hand, acetic acid and formic acid are known to be highly irritating to the skin [15]. Therefore, insight into the effects of cosmetics on short-chain fatty acids produced by S. epidermidis will have an impact on skin health.
In this study, we attempted to establish a cosmetics pattern map that can extract and compare the characteristics of cosmetics based on the short-chain fatty acids produced by S. epidermidis.
2. Materials and Methods
2.1. Materials
The bacteria used were Staphylococcus epidermidis (NBRC100911) purchased from the National Institute of Biotechnology and Life Science (NBRC).
The nine cosmetics used in this study and the main ingredients used in the cosmetics are listed below.
(1) Cosmetic A containing Soy Milk Fermentation Liquid (Made in Japan) (Soy.fermentA)
Soy milk fermentation liquid
(2) Cosmetics containing Soy Milk Fermentation Liquid (Made in Japan) (Soy.fermentB)
Soy milk fermentation liquid
(3) Antibacterial cosmetics (Made in Japan) (Antimicrobial)
Antimicrobial ingredient
(4) cosmetics containing plant extracts (Made in South Asia)(SouthAsia)
Plant oil
(5) cosmetics claiming natural ingredients (Made in Japan) (Natural)
Soy milk fermentation liquid, vegetable oil
(6) cosmetics containing loofah water (Made in Japan) (Loofar)
Loofah extract
(7) cosmetics containing plant extracts (Made in Japan) (Plant)
Plant extract
(8) Lotion containing rice ferment (Made in Japan) (Rice.fermentA)
Rice fermentation liquid, vegetable oil
(9) cosmetics containing rice ferment (Made in Japan) (Rice.fermentB)
Rice fermentation liquid
2.2. Reagents
The following culture media and reagents were used to culture the bacteria.
Bacterial pre-culture: 702 medium (High polypeptone [Wako Pure Chemical Industries, Ltd.], yeast extract [Wako Pure Chemical Industries, Ltd.], magnesium sulfate heptahydrate MgSO₄-7H₂O [Nakalai Tesque Co.]).After preparation of the medium, pH was adjusted to 7.0.Co-culture of bacteria and cosmetics: GAM medium (Modified GAM Bouillon [Nissui Pharmaceutical Co.]).
2.3. Nitrogen replacement in the culture vessel
A 10 mL headspace vial (vial, 45 mm × 22 mm, Agilent technologies Inc., USA) was used as the culture vessel. Nitrogen was supplied from the connecting tube of a nitrogen gas cylinder to the vials using silicone tubes (9 × 12 mm), 1000-5000 μL Pipet Tip. GAM medium was added to 5 mL vials. 5 µL of resazurin diluent (1 mg/mL), a redox indicator, was added at the time of GAM medium preparation, and the color of the medium changed from reddish purple to yellow derived from the medium as a discrimination of anaerobic condition. The tip of the nitrogen inflow device was inserted into vials. The entire area was covered with aluminum foil, and holes were drilled for oxygen exit. An anaerobic environment was created by flowing nitrogen gas and bubbling for 5 minutes. After gas displacement, a crimp cap (septum type: molded PTFE/butyl, Agilent technologies Inc., USA) was immediately set on the vial and immediately sealed using a hand crimper (G.L. Science Corporation). The culture medium was then sterilized by autoclaving.
2.4. Co-culture of S. epidermidis and cosmetics
S. epidermidis was inoculated into 5 mL of 702 medium and pre-cultured at 37°C for 24 hours. In the main culture, the pre-culture solution was added to the GAM medium prepared in the method to achieve an initial bacterial concentration of 1.0 × 10⁷ cells/5 mL. A micro syringe (Ito Manufacturing Co., Ltd.) was used for the addition. 0.25 mL (5% of the total volume of 5 mL) of the cosmetic product to be studied was added and incubated at 37°C for 24 and 48 h. A total of 6 samples per cosmetic product were prepared: vehicle-added control group (0, 24, 48 h) and cosmetic product-added group (0, 24, 48 h).
2.5. Extraction of exogenous metabolites (removal of foreign substances)
After co-culturing as described above, the culture medium was centrifuged (24,000 rpm, 5 min) and 2.0 mL of the supernatant was collected as exogenous metabolites. Further centrifugation (14000 rpm, 1 min) was performed to ensure removal of bacteria, and 1.5 mL of the supernatant was removed of remnants using a 0.45 µm membrane filter (Shimadzu GLC Corporation). 1.0 mL was added to a vial for HPLC, covered with a lid, and set in an autosampler for HPLC (L-2200, Hitachi, Ltd. Hitachi, Ltd. Randomization was performed using the random function in Microsoft Excel for the analysis, and all analyses of each cosmetic were performed on the same day.
2.6. Short-chain fatty acid (BTB method) analysis by high-performance liquid chromatography (HPLC)
Analysis by HPLC was performed using an RSpak KC-811 column (particle size 6.0 µm, inner diameter 8 mm, length 300 mm, G.L. Science Corporation), column oven: 40°C (FS-420, Pro Devices Corporation), flow rate: 0.5 mL/min (A), 0.6 mL/min (D), buffer A: 3 mM HClO₄, buffer D: 0.1 mM BTB and 15 mM Na₂HPO₄, analysis time: 80 min.
2.7. Peak identification
Peak identification was based on the retention time in the chromatogram. The obtained area value data for short-chain fatty acids were converted to an Excel file and used for subsequent statistical analysis.
2.8. Statistical Analysis
SIMCA-P+ ver. 13.0 (Umetrics, Umeå, Sweden) was used for principal component analysis.
3. Results
1. Establishment of an anaerobic culture and short-chain fatty acid production measurement system for S. epidermidis.
First, to investigate whether metabolites of S. epidermidis, an indigenous skin bacterium, can be quantitatively detected in an anaerobic environment that mimics skin pores, media with or without S. epidermidis cultured anaerobically for 0, 24, and 48 hours were subjected to HPLC for analysis. As a result, six peaks were detected and identified as six short-chain fatty acids (succinic acid, lactic acid, formic acid, acetic acid, propionic acid, and isobutyric acid). This suggested that S. epidermidis metabolites could be quantitatively detected under these culture conditions. The concentrations (µg/mL) of the short-chain fatty acids were calculated from the detected peak area values, calibration curves for each short-chain fatty acid. Using these values, we compared the amount of short-chain fatty acid production by incubation time, and found that lactic acid and acetic acid increased in the 24- and 48-hour incubation samples (Figure 1). From this result, we considered that we could construct a system to simulate the metabolism of indigenous skin bacteria in skin pores.
3.1. Change in short-chain fatty acid production by S. epidermidis upon addition of cosmetics
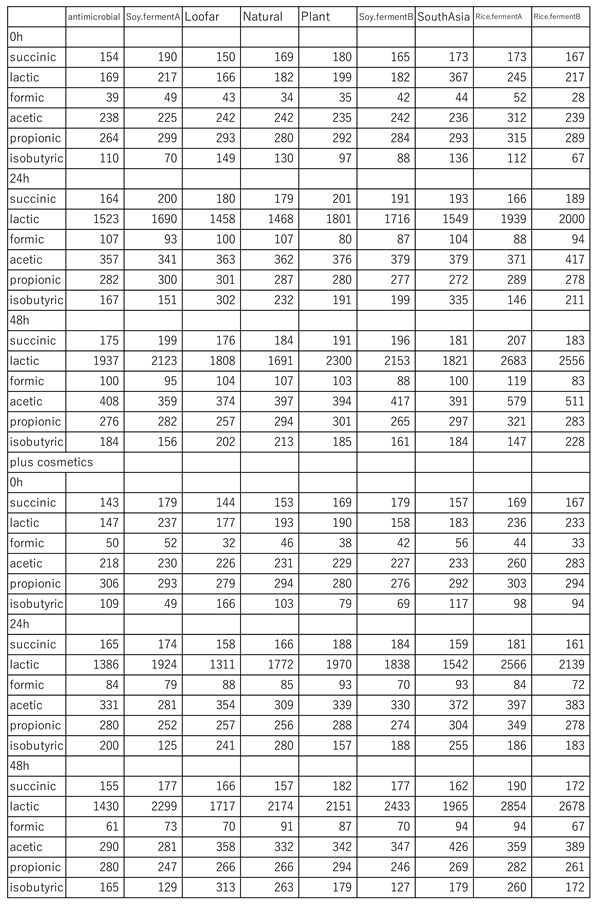
Since we were able to construct a system for analyzing metabolites of S. epidermidis as described above, we decided to examine whether the metabolites changed when actual cosmetics were added to the culture. S. epidermidis alone cultured for 0, 24, and 48 hours, and S. epidermidis added with various cosmetics for 0, 24, 48 hours, were subjected to HPLC and analyzed. The results showed that there were various patterns depending on the cosmetics (Table 1). The results showed that there were various patterns depending on the cosmetics, which may allow us to distinguish and clarify the effects of cosmetics on indigenous skin bacteria.
3.2. Classification of cosmetics based on principal component analysis of the amount of short-chain fatty acids produced by indigenous skin bacteria
In order to gain a comprehensive insight into the pattern of short-chain fatty acid production by each cosmetic product obtained above, a principal component analysis was conducted using the values as explanatory variables. As a result, an axis separating lactic acid and other short-chain fatty acids as the first principal component was obtained, and an axis separating succinic acid, formic acid and isobutyric acid as the second principal component (Figure 2). The contribution of the first and second principal components (PC1 and PC2) was 44.8% and 24.7%, respectively. In the score plot, the samples were divided into two groups. First group was the groups of rice-fermented cosmetics and soy-fermented cosmetics and the second group was South Asian cosmetics, plant extract cosmetics and natural cosmetics. One cosmetic containing soy ferment was clustered in the vicinity of antimicrobial. This finding suggested that it would be possible to classify cosmetics according to their characteristics and to identify the substances that contribute to this classification.
Principal component analysis was performed using concentrations of six short-chain fatty acids obtained from HPLC analysis of cosmetics and cultured S. epidermidis cultures. (A) Score plot (B) Loading plot.
4. Discussion
Until now, there have been no studies classifying the properties of cosmetics based on their effects on indigenous skin bacteria, but in this study, for the first time, we succeeded in establishing a system for classifying the properties of cosmetics based on their effects on S. epidermidis, one of the indigenous skin bacteria.
Among the nine cosmetics investigated in this study, rice- and soy-fermented cosmetics were plotted in the close proximity on the score plot; the fact that these cosmetics were located near succinic acid and formic acid on the loading plot suggests that these cosmetics have similar effects on S. epidermidis. The fact that these cosmetics were located near formic acid, acetic acid in the loading plot suggests that they are highly effective in promoting these acids production towards S. epidermidis. In contrast, South Asian cosmetics, loofar cosmetic, natural cosmetic, and plant-based cosmetic formed clusters at positions near isobutyric acid. These results suggest that these cosmetics may have similar properties and may enhance production of isobutyric acid. Since South Asian cosmetics contain abundant plant-derived ingredients, it was considered that cosmetics with high plant ingredients would be located in the vicinity. There were two bean-fermented cosmetics, but each was located in a different cluster, suggesting that the same bean-fermented cosmetics may have very different properties.
Cosmetics plotted onto the loading plot space generates many insights into the characteristics of the cosmetics. As described above, lactic acid has beneficial effects on the physical properties and immunity of the skin. Succinic acid has been reported to inhibit the growth of Propionibacterium acnes, the causative agent of acne, without causing significant irritation to the skin [16]. On the other hand, acetic acid and formic acid are known to be highly irritating to skin [13]. Based on these reports, we hypothesized that cosmetics showing a positive correlation with lactic acid and succinic acid in the principal component analysis would be highly effective towards S. epidermidis, while cosmetics showing a positive correlation with acetic acid and formic acid would be less effective.
Safety of cosmetics is evaluated by several indices, such as skin stimulation, allergic potential, light toxicity, toxicity, and eye stimulation [17]. However, there has been no study to elucidate the effects of cosmetics on skin microbiota. Indeed, some patients resort skin inflammation upon local administration of cosmetics, but the cause was not known. These symptoms might be explained by the disorder of skin microbiota in the future, considering these researches.
Several ingredients might influence the growth and short chain fatty acid production by S. epidermidis. For example, oligosaccharides and amino acids might enhance the growth and short chain fatty acid production. Antibiotics contained in cosmetics might hinder the growth of S. epidermidis.
It is now not known if ingredients in cosmetics such as emollients, lubricants, emulsifiers, fatty acids, alcohols, gelling agents, thickeners and antibiotics might affect growth or metabolism of skin microbiota. This study will shed light on the effect of these constituents towards skin microbiota, and will enable design of a more functional cosmetics that function on skin microbiota.
Recently, many cosmetics that appeal improvement of skin microbiota are commercially available. However, there is no indices that objectively evaluate the competence of the cosmetics on skin microbiota. This confused situation gives rise to survival of low quality cosmetics, and is unbeneficial for consumers. Therefore, this research will be an effective method to evaluate those cosmetics in terms of their function on skin microbiota, which is beneficial for consumers.
The problem of multidrug resistance bacteria is one of the strongest threat to the human beings now. MRSA is generated from Staphylococcus aureus, one of skin microbiota. It is known that S. aureus is sensitive to organic acids including lactic acid, acetic acid and propionic acid [18]. Indeed, S. epidermidis is known to prevent colonization of S. aureus through delta-lysin, esp and antimicrobial peptide [19,20,21,22], and skin microbiota abundant in S. epidermidis or Corynebacterium acnes is proposed to be healthy [23]. Imbalance in skin microbiota generates nonpathogenic but unfavorable status of the skin, such as dry skin and sensitivity [24]. Moreover, S. epidermidis act on TLRs and enhances production of antimicrobial peptides, which also leads to prevention of opportunistic pathogenic bacteria [25]. S. epidermidis also prevents adhesion and biofilm formation by S. aureus [26]. It is proposed that the lipoteichoic acid of S. epidermidis can prevent skin inflammation mediated by TLR2 and TLR3 [27]. Therefore, regulation of lactic acid by S. epidermidis will be a potent approach to the regulation of S. aureus, which leads to prevention of the rise of MRSA.
So far, in vitro on mono- or co-cultures of skin bacteria to measure the effects of cosmetics by growth measurement using qPCR has been described [28]. Studies using 3D skin models have been applied to elucidate the interaction between skin microbiota and cosmetics [29]. These are considered ex vivo models, and the knowledge obtained in this study can be in the future utilized in these ex vivo models. However, this is the first study to extract information on cosmetics by using data obtained from in vitro culture of skin microbiota.
Cosmetic ingredients used that are either functional ingredients, such as preservatives, oils and emulsifiers, or active ingredients, impact the skin microbiota and require caution. Indeed, conventional skincare or hygiene products such as soap, gel and cream contain preservatives and natural and synthetic chemicals that impact microbiota even if these effects are not fully investigated in detail until now [30,31].
This research has several limitations. For example, it did not adopt 3D-structured skin model [29]. However, when focusing onto the metabolism of skin microbiota, this research will provide some information on the properties of cosmetics. Secondly, this research used only S. epidermidis. Skin microbiota contain other bacteria, such as Propionibacterium acnes, Corynebacterium acnes and S. aureus. Therefore, in the future, these bacteria should also be taken into account. The composition of media is artificial. Media with which bacteria grow in the actual skin pores are triglycerides and amino acids secreted from the sebaceous glands. However, since the growth using these components requires enormous time, media containing glucose was used to facilitate the analysis. Therefore, knowledge obtained through this study should take these into account.
Skin pH varies dependent on the parts of the skin. For example, pHs of the skin of Axilliary vault, and antecubital crease tend to be high. In contrast, pH of the skin of glabella and alar crease tend to be low [32,33]. Therefore, administration of cosmetics and decrease of pH through S. epidermidis should be considered in terms of the control of pH.
Recently, fungi which proliferate in skin pores, such as Malassezia furfur [34], cause pancreatic oncogenesis and is a serious threat to the human health. Since fungi generally cannot grow in an acidic environment, increase of short-chain fatty acids by S. epidermidis might be favorable to prevent proliferation of these fungi, which is a significant target of future research.
Some cosmetics contain detergents. Since detergents interfere with skin bacteria, these might have impacted the results obtained in this research. This should be considered in the future research.
It was not clear from this study which components of these cosmetics stimulated the production of these acids. However, plant extracts and fermented products contain plant and microbial components and microbial metabolites, these components may have stimulated the production of the short-chain fatty acids by S. epidermidis.
Atopic dermatitis is one of the major skin diseases in the developed countries. It is caused by skin barrier dysfunction such as 1) deficiency in ceramide, cholesterol and free fatty acids, 2) deficiency in differeitiation of keratinocyte proteins (filaggrin, loricin and involcrin), 3) sensitivity to immune stimulus, 4) decrease in antimicrobial peptides and 5) decrease in the diversity of skin microbiota [35]. Indeed, it is reported that S. epidermidis is less and S. aureus is abundant in atopic dermatitis patients. Both S. epidermidis and S. aureus share the characteristics of biofilm formation, but S. aureus is more frequent in atopic dermatitis patients [36]. Therefore, usage of specific cosmetics might cause dysfunction of skin microbiota and lead to atopic dermatitis.
Cosmetics market is rapidly growing. The global cosmetics market size was estimated to be USD 375.30 billion in 2022. It is expected to grow at a compound annual growth rate (CAGR) of 5.1% from 2021 to 2030 [37]. Therefore, people in the world will more and more use cosmetics. However, since legal regulations of cosmetics vary greatly depending on the countries, some subjective criteria for evaluation of the influence of cosmetics on the skin microbiota is needed. The method proposed in this study will be one such criteria.
It is reported that with increasing age, at the phylum level, Proteobacteria increased and Actinobacteria decreased, and at the genus level, Corynebacterium increased and Cutibacterium decreased in Caucasian women [38]. Therefore, it is hypothesized that alteration of skin microbiota is related to aging, and manipulation of skin microbiota might lead to anti-aging.
So far, the effects of cosmetics on skin microbiota have been analyzed. For example, the effects of cosmetics on the growth, virulence, cytotoxicity on HaCat keratinocytes, biofilm formation of S. epidermidis and Cutibacterium acnes have been reported [39,40,41]. However, there has been no study on the short-chain fatty acids of S. epidermidis and this report is the first study to report it.
In summary, a cosmetics-pattern map using S. epidermidis was established in this study. This novel method can be used to pattern cosmetics and infer their effects on skin microbiota.
Author Contributions
Conceptualization, H.K.; validation, H.K., K.T., and Y.T.; investigation, K.T., S.T., C.M., N.F., N.H., Y.S., Y.H. and H.D.; writing—original draft preparation, H.K., and K.T., writing— review and editing, H.K., K.T., and Y.T.; visualization, H.K., N.F.; supervision, H.K.; project administration, H.K.; funding acquisition, H.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to thank Ms. Sonoko Iwashita for her contributions to culture and HPLC experiments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yamamura, H., History of Japanese cosmetics, Yoshikawa Kobunkan, 2016, 19-30.
- Miki, M. Types and ways of using cosmetics-skin care cosmetics-. J. Jpn. Cosmet. Sci. Ind. 2018, 42, 109–124. [Google Scholar]
- Ryuen, H. Cosmetic industry and consumers. J. Jpn. Res. Assoc. Text. End-Uses 2012, 53, 1032–1037. [Google Scholar]
- Miyagawa, M.; Fujikawa, A.; Nagadome, M.; Kohama, K.; Ogami, T.; Kitamura, S.; Kitagaki, H. Glycosylceramides Purified from the Japanese Traditional Non-Pathogenic Fungus Aspergillus and Koji Increase the Expression of Genes Involved in Tight Junctions and Ceramide Delivery in Normal Human Epidermal Keratinocytes. Fermentation 2019, 5, 43. [Google Scholar] [CrossRef]
- Otsuka, A.; Moriguchi, C.; Shigematsu, Y.; Tanabe, K.; Haraguchi, N.; Iwashita, S.; Tokudome, Y.; Kitagaki, H. Fermented Cosmetics and Metabolites of Skin Microbiota—A New Approach to Skin Health. Fermentation 2022, 8, 703. [Google Scholar] [CrossRef]
- Matt Venus, et al. Basic physiology of the skin. Surgery (Oxford) 2010, 28, 469–472. [Google Scholar] [CrossRef]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Joachim W, et al. Glycerol Regulates Stratum Corneum Hydration in Sebaceous Gland Deficient (Asebia) Mice. Investigate Dermatory. 2003, 120, 728–737. [Google Scholar] [CrossRef] [PubMed]
- Nodake, Y. , et al. Pilot study on novel skin care method by augmentation with Staphylococcus epidermidis, an autologous skin microbe – A blinded randomized clinical trial. Science Direct. 2015, 79, 119–126. [Google Scholar]
- Mizukoshi, K. Effects of lactic acid on the flexibility of the stratum corneum. Ski. Res. Technol. 2020, 26, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Naik, S.; et al. Compartmentalized Control of Skin Immunity by Resident Commensals. Science 2012, 337, 1115–1119. [Google Scholar] [CrossRef] [PubMed]
- Prince, A. Staphylococcus aureus metabolites promote IL-10. Nat. Microbiol. 2020, 5, 1183–1184. [Google Scholar] [CrossRef]
- Y Wang, et al. Staphylococcus epidermidis in the human skin microbiome mediates fermentation to inhibit the growth of Propionibacterium acnes: implications of probiotics in acne vulgaris”. Microb. Cell Physiol. 2014, 98, 411‒424.
- Y Wang, et al. Propionic acid and its esterified derivative suppress the growth of methicillin-resistant Staphylococcus aureus USA300. Benef. Microbes 2014, 5, 161–168. [Google Scholar] [CrossRef]
- Sigurdsson, J. , Björnsson, A., Gudmundsson, S. T. Formic acid burn--local and systemic effects. Report of a case. Burns Incl Therm Inj. 1983, 9, 358–361. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J. H. , Lee, S., Lee, H. G., Choi, D., Lim, K. M. Evaluation of Skin Irritation of Acids Commonly Used in Cleaners in 3D-Reconstructed Human Epidermis Model, KeraSkinTM. Toxics. 2022, 10, 558. [Google Scholar] [CrossRef]
- Becker, L. C. , Bergfeld, W. F., Belsito, D. V., Hill, R. A., Klaassen, C. D., Liebler, D. C., Marks, J. G., Jr, Shank, R. C., Slaga, T. J., Snyder, P. W., Gill, L. J., & Heldreth, B. Safety Assessment of Glycerin as Used in Cosmetics. Int. J. Toxicol. 2019, 38, 6S–22S. [Google Scholar] [CrossRef]
- Dubal, Z. B., Paturkar, A. M., Waskar, V. S., Zende, R. J., Latha, C., Rawool, D. B., & Kadam, M. M. Effect of food grade organic acids on inoculated S. aureus, L. monocytogenes, E. coli and S. Typhimurium in sheep/goat meat stored at refrigeration temperature. Meat science 2004, 66, 817–821. [CrossRef]
- Lowy F., D. Staphylococcus aureus infections. The New England journal of medicine 1998, 339, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Iwase, T., Uehara, Y., Shinji, H., Tajima, A., Seo, H., Takada, K., Agata, T., & Mizunoe, Y. Staphylococcus epidermidis Esp inhibits Staphylococcus aureus biofilm formation and nasal colonization. Nature 2010, 465, 346–349. [CrossRef]
- Al-Mahrous, M. , Sandiford, S. K., Tagg, J. R., & Upton, M. Purification and characterization of a novel delta-lysin variant that inhibits Staphylococcus aureus and has limited hemolytic activity. Peptides 2010, 31, 1661–1668. [Google Scholar] [CrossRef] [PubMed]
- Cogen, A. L. , Yamasaki, K., Sanchez, K. M., Dorschner, R. A., Lai, Y., MacLeod, D. T., Torpey, J. W., Otto, M., Nizet, V., Kim, J. E., & Gallo, R. L. Selective antimicrobial action is provided by phenol-soluble modulins derived from Staphylococcus epidermidis, a normal resident of the skin. The Journal of investigative dermatology 2010, 130, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Christensen, G.J.M. , Scholz, C.F.P., Enghild, J. et al. Antagonism between Staphylococcus epidermidis and Propionibacterium acnes and its genomic basis. BMC Genomics 2016, 17, 152. [Google Scholar] [CrossRef] [PubMed]
- Grice, E. A. , & Segre, J. A. The skin microbiome. Nature reviews. Microbiology 2011, 9, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Wiesner, J. , & Vilcinskas, A. Antimicrobial peptides: the ancient arm of the human immune system. Virulence 2010, 1, 440–464. [Google Scholar] [CrossRef] [PubMed]
- Christensen, G. J. , & Brüggemann, H. Bacterial skin commensals and their role as host guardians. Beneficial microbes 2014, 5, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y. , Di Nardo, A., Nakatsuji, T., Leichtle, A., Yang, Y., Cogen, A. L., Wu, Z. R., Hooper, L. V., Schmidt, R. R., von Aulock, S., Radek, K. A., Huang, C. M., Ryan, A. F., & Gallo, R. L. Commensal bacteria regulate Toll-like receptor 3-dependent inflammation after skin injury. Nat. Med. 2009, 15, 1377–1382. [Google Scholar] [CrossRef] [PubMed]
- van der Krieken, D. A. , Ederveen, T. H., van Hijum, S. A., Jansen, P. A., Melchers, W. J., Scheepers, P. T., Schalkwijk, J., & Zeeuwen, P. L. An In vitro Model for Bacterial Growth on Human Stratum Corneum. Acta dermato-venereologica 2016, 96, 873–879. [Google Scholar] [CrossRef] [PubMed]
- Niehues, H. , Bouwstra, J. A., El Ghalbzouri, A., Brandner, J. M., Zeeuwen, P. L. J. M., & van den Bogaard, E. H. 3D skin models for 3R research: The potential of 3D reconstructed skin models to study skin barrier function. Experimental dermatology 2018, 27, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Sfriso, R. , Egert, M., Gempeler, M., Voegeli, R., & Campiche, R. Revealing the secret life of skin - with the microbiome you never walk alone. International journal of cosmetic science 2020, 42, 116–126. [Google Scholar] [CrossRef]
- Holland, K. T. , & Bojar, R. A. Cosmetics: what is their influence on the skin microflora? American journal of clinical dermatology 2002, 3, 445–449. [Google Scholar] [CrossRef]
- Kong, H. H., & Segre, J. A. Skin microbiome: looking back to move forward. The Journal of investigative dermatology 2012, 132(3 Pt 2), 933–939. [CrossRef]
- Harris-Tryon, T. A., & Grice, E. A. (2022). Microbiota and maintenance of skin barrier function. Science (New York, N.Y.), 376(6596), 940–945. [CrossRef]
- Aykut, B., Pushalkar, S., Chen, R., Li, Q., Abengozar, R., Kim, J. I., Shadaloey, S. A., Wu, D., Preiss, P., Verma, N., Guo, Y., Saxena, A., Vardhan, M., Diskin, B., Wang, W., Leinwand, J., Kurz, E., Kochen Rossi, J. A., Hundeyin, M., Zambrinis, C., … Miller, G. The fungal mycobiome promotes pancreatic oncogenesis via activation of MBL. Nature, 2019, 574(7777), 264–267. [CrossRef]
- David Boothe, W., Tarbox, J.A., Tarbox, M.B. (2017). Atopic Dermatitis: Pathophysiology. In: Fortson, E., Feldman, S., Strowd, L. (eds) Management of Atopic Dermatitis. Advances in Experimental Medicine and Biology, vol 1027. Springer, Cham. [CrossRef]
- Clausen M, Agner T, Lilje B, Edslev SM, Johannesen TB, Andersen PS. Association of Disease Severity With Skin Microbiome and Filaggrin Gene Mutations in Adult Atopic Dermatitis. JAMA Dermatol. 2018;154(3):293–300. [CrossRef]
- https://www.precedenceresearch.com/cosmetics-market accessed 2023.6.3.
- R. Jugé and others, Shift in skin microbiota of Western European women across aging, Journal of Applied Microbiology, Volume 125, Issue 3, 1 September 2018, Pages 907–916. 1 September; 3. [CrossRef]
- Gannesen, A. V., Borrel, V., Lefeuvre, L., Netrusov, A. I., Plakunov, V. K., & Feuilloley, M. G. J. (2019). Effect of two cosmetic compounds on the growth, biofilm formation activity, and surface properties of acneic strains of Cutibacterium acnes and Staphylococcus aureus. MicrobiologyOpen, 8(3), e00659. 3. [CrossRef]
- Filaire, E.; Vialleix, C.; Cadoret, J.-P.; Guénard, S.; Muller, C.; Dreux-Zigha, A.; Berthon, J.-Y. Characterization of Reactive and Sensitive Skin Microbiota: Effect of Halymenia durvillei (HD) Extract Treatment. Cosmetics 2019, 6, 69. [Google Scholar] [CrossRef]
- Saising, J., & Voravuthikunchai, S. P. (2012). Anti Propionibacterium acnes activity of rhodomyrtone, an effective compound from Rhodomyrtus tomentosa (Aiton) Hassk. leaves. Anaerobe, 18(4), 400–404. [CrossRef]
Figure 1.
Short-chain fatty acid concentration of the culture of S. epidermidis. S. epidermidis was cultured in nitrogen-purged GAM medium anaerobically. Concentrations of cultures incubated with S. epidermidis for 0, 24 and 48 h were analyzed using HPLC.
Figure 1.
Short-chain fatty acid concentration of the culture of S. epidermidis. S. epidermidis was cultured in nitrogen-purged GAM medium anaerobically. Concentrations of cultures incubated with S. epidermidis for 0, 24 and 48 h were analyzed using HPLC.
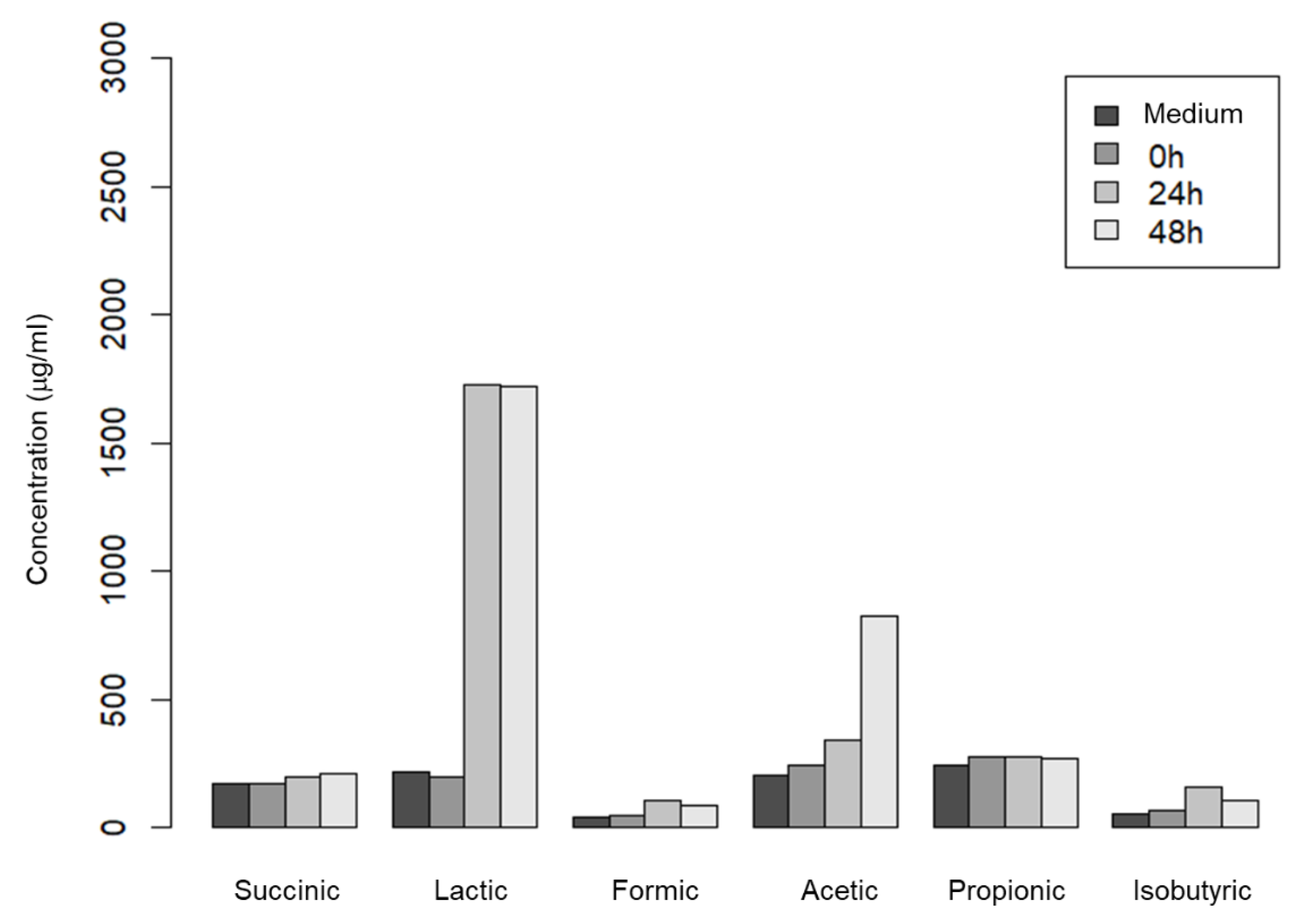
Figure 2.
Results of principal component analysis of short-chain fatty acids in cosmetics and cultured S. epidermidis.
Figure 2.
Results of principal component analysis of short-chain fatty acids in cosmetics and cultured S. epidermidis.
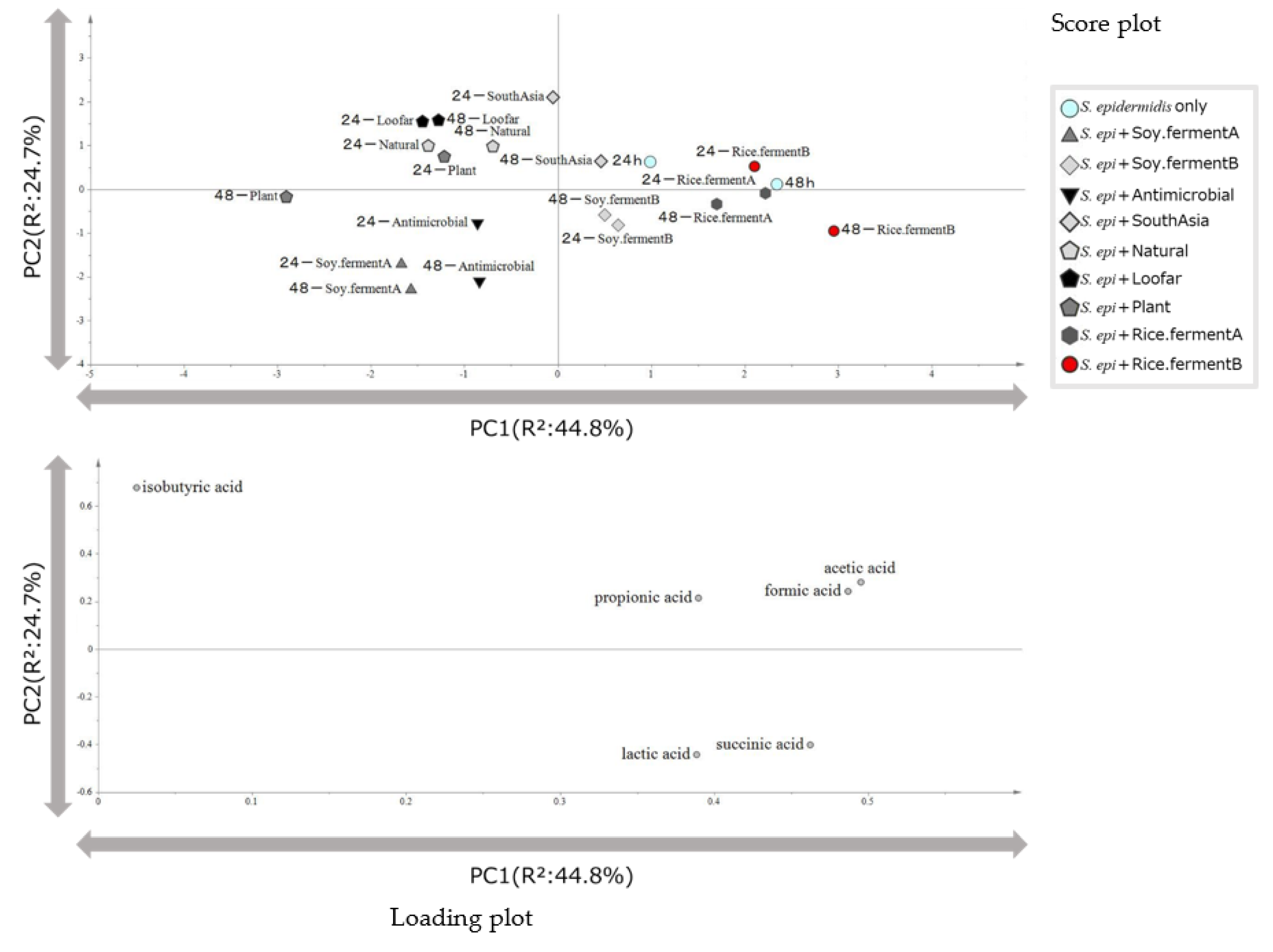

Table 1.
Short-chain fatty acid concentrations (¦Ìg/ml) of S. epidermidis cultured with various cosmetics.
Table 1.
Short-chain fatty acid concentrations (¦Ìg/ml) of S. epidermidis cultured with various cosmetics.
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



