
Submitted:
08 June 2023
Posted:
09 June 2023
You are already at the latest version
Abstract
Amyotrophic lateral sclerosis (ALS) is a severe and incurable neurodegenerative disease, char-acterized by the progressive death of motor neurons, leading to paralysis and death. It is a rare disease distinguished by high patient-to-patient heterogeneity, making its study arduous and complex. Extracellular vesicles (EVs) emerged as an important player in ALS development. Thus, ALS phenotype-expressing cells can spread their abnormal bioactive cargos through the secretion of EVs, even at far distant tissues. Importantly, due to their nature and composition, EVs’ formation and cargo can be exploited for the better comprehension of this elusive disease and identification of novel biomarkers, as well as for potential therapeutic applications, such as those based on stem cell-derived exosomes. This review highlights the recent advances in the identification of the role of EVs in ALS etiopathology and how EVs can be promising new approaches for therapeutic strategies.
Keywords:
Amyotrophic lateral sclerosis
; neurodegenerative diseases
; extracellular vesicles
; exosomes
; miRNA
; biomarkers
1. The Current State-of-the Art of ALS
Amyotrophic lateral sclerosis (ALS) is a fatal neurodegenerative disease. Described for the first time in the 19th century by Charcot, ALS is characterized by the degeneration of lower (spinal and bulbar) and upper (corticospinal) motor neurons [1]. The selective loss of motor neurons (MNs) in the primary motor cortex, brainstem, and spinal cord progressively leads to severe effects such as loss of motor control, paralysis, and death. Death usually occurs due to respiratory failure. About half of ALS patients show impairments in cognitive functions and behavior, with 5-25% of patients ending up developing frontotemporal dementia (FTD) [2,3,4], which is an uncommon type of dementia characterized by changes in the frontal and temporal lobes.
ALS is considered a rare disease, having an incidence that ranges between 0.6 to 3.8 persons out of 100000, and a prevalence between 4.1 and 8.4 per 100 000 individuals, with the average age of onset between 51 and 66 years old [5]. The life expectancy of ALS patients is short, between 24 to 50 months. However, about 10% of patients manage to live for more than 10 years [5], a fact that reflects the high patient-to-patient phenotypic variability that characterizes ALS. The triggering elements of the disease remain unknown, although in several individuals a genetic cause can be attributed. Some studies point it toward having a possible oligogenic or polygenic nature, as mutations in two or more genes may be required for the disease [6,7]. People with a history of ALS cases in their family, and that carry ALS-related genes, are more likely to develop the disease (familial, fALS), representing 5 to 10% of all the cases. For the remaining 90% to 95%, the illness can occur spontaneously, meaning without a family history (sporadic, sALS) [6], and still be linked to ALS-related genes. Currently, ALS is difficult to diagnose, due to the absence of a test that can, solely, lead to its identification, unless it is a familial form. In the absence of family history, a battery of exams is often performed to discard other possible pathologies. Currently, ALS remains cureless, and the available treatments are sparse and mostly palliative. Merely two approved medications are currently prescribed to patients, Riluzole and Edaravone, with the latter only being approved in some countries. Yet, those present merely small benefits in delaying ALS progression, usually only by some months [8]. Therefore, the discovery of new and more effective drugs is of the utmost importance.
1.1. Risk Factors for ALS Onset and Progression
The likelihood of developing ALS and its progression is influenced by numerous factors, including genetic and non-genetic origins. One important non-genetic factor is age, as individuals who develop ALS in early adulthood tend to experience slower disease progression rates [9,10]. Another factor is gender, with men being about 1.3 times more likely to develop ALS than women, and earlier in life [11]. Gender also plays a role in ALS onset type, being a spinal-onset more common in men, while women are more likely to present a bulbar-onset [10]. In addition to genetic factors, exposure to certain modifiers throughout an individual’s life may also contribute to the risk of developing ALS [12]. Several environmental and lifestyle factors that have been identified as potential risk factors for ALS onset include hazardous smoking habits [13], higher lipid levels [14], prolonged exposure to pollutants [15], heavy metals [16], chemicals [17], electromagnetic fields [17], a history of electric shock [18], and head trauma [19]. Other factors that have been linked to an increased risk of ALS include military service [20], participation in professional sports [21,22], and occupations that involve repetitive physical work [12,23,24]. However, some of these factors are contested due to studies with inconclusive results [25]. These factors can eventually lead to epigenetic and genomic changes that may contribute to ALS onset, for instance, the occurrence of C9ORF72 (chromosome 9 open reading frame 72) somatic mutations [26]. Scientific reports consistently evidence an interaction between genetic and environmental risk factors. Epigenetic alterations, mostly comprising DNA methylations, were identified by screening ALS patients’ biofluids, and postmortem brain and spinal cord tissues. In this regard, [27] reported gene and CpG islands methylations in 38 differentially methylated sites, when studying 10 sALS patients’ brain samples [27]. Similarly, Figueroa-Romero et al. (2012) identified 3574 methylated genes in postmortem sALS patients’ spinal cord [28]. Cai et al. (2022) recently proposed a role for DNA methylation in the pathogenesis of sALS. Their study involved analyzing and comparing the blood of 32 healthy controls with 32 sALS patients, leading to the identification of 12 differentially methylated regions (DMRs) in 12 genes, and 34 differentially methylated positions (DMPs) in 13 genes. The abnormal methylation patterns were primarily associated with genes involved in the regulation of crucial cellular functions that have previously been linked to ALS, including microtubule-based movement, ATP-nucleotide binding, and neuronal apoptosis [29]. Despite research efforts to enlighten the impact of environmental and lifestyle factors on the different cellular and molecular processes involved in ALS onset and progression, the exact mechanisms underlying motor neuron degeneration are still not sufficiently understood [1].
1.2. ALS Genetics and Associated Mechanisms
ALS is a highly heterogeneous disease that can be caused by a wide array of different genes, some having hundreds of possible mutations [30]. Consequently, distinct fundamental cellular processes have been reported to be dysfunctional in different stages of the disease, including DNA repair mechanisms, RNA metabolism, mRNA axonal transport, protein homeostasis, protein trafficking, protein misfolding and aggregation, calcium regulation, mitochondrial function [31], redox signaling, lipid metabolism, glutamate signaling, and autophagy [32]. ALS-related gene mutations may also affect intercellular communication and function, such as neurovascular function [33,34,35], glial-related neuroinflammation, [10,36,37,38], and neuron-glia interaction [39,40]. Among the several genes identified to be ALS-related, some are involved in both fALS and sALS, such as TDP-43 (TAR DNA-binding protein 43), also known as TARDBP (transactive response DNA binding protein), SOD1 (copper-zinc superoxide dismutase 1), C9orf72, and FUS (fused in sarcoma), among others [41]. Nevertheless, for 32% and 89% of the patients with fALS and sALS, respectively, the mutations that are involved are unknown [42,43].
One of the most studied ALS-related genes is SOD1, which encodes for an important antioxidant protein, superoxide dismutase [44], responsible for converting superoxide radicals in hydrogen peroxide and oxygen [45]. Mutant SOD1 (mSOD1) alters different metabolic pathways and also results in the formation of misfolded SOD1 protein aggregates and neurodegeneration [46,47]. Accordingly, mSOD1 aggregates’ accumulation impairs axonal transport and is neurotoxic to spinal cord MNs, from the presymptomatic phase onwards, in the ALS mice SOD1-G93A model [48]. mSOD 1 is also responsible for the alteration of the dynamic interaction between MNs and their surrounding glial cells, evoking a non-cell autonomous toxicity mechanism, driven either by the promotion of the secretion of neurotoxic cytokines, through the loss of glial cell’s supporting properties, or both, leading to the death of MNs [49,50]. In one proposed mechanism, extracellular mSOD1 is endocytosed by microglia and activates caspase-1, leading to upregulation of IL-1β [51,52]. IL-1β is one of the pro-inflammatory cytokines potentially involved in ALS neuroinflammation-related processes [53], like microgliosis or astrogliosis. In ALS patients’ postmortem tissue samples, microglia is in a proinflammatory state [54] and releases several cytokines, such as IL-1α and TNF-α, that induce astrocytes neurotoxicity [54]. Such evidence points to a deleterious crosstalk between microglia and astrocytes, thus tracing for an increased proinflammatory and neurotoxic microenvironment. Therefore, the progressive degeneration of corticospinal and spinal motor neurons may depend on their vulnerability to both mSOD1 aggregates accumulation and the effects of surrounding glial cell’s dysregulation, a fact emphasizing the simultaneous occurrence of a lower and upper MNs degeneration [55].
The most commonly mutated gene in both fALS and sALS patients is the C9ORF72. The C9ORF72 gene contains 11 exons, and (GGGGCC)n is located between exons 1a and 1b. (GGGGCC)n is located in the first intron of V1 and V3 and in the promoter region of variant 2. This gene codes for a protein with the same name whose function is not fully understood but is thought to be involved in different cellular activities, such as protein transport, vesicle formation, autophagy, RNA processing, cell signaling, among others [56,57]. It has been suggested that C9RF72 may partake in the autosomal and lysosomal function of macrophages and microglia through the regulation of inflammatory responses, possibly being related to MNs survival, relevant in ALS [58,59]. Wild-type C9ORF72 forms a complex with SMCR8 (Smith–Magenis syndrome chromosomal region candidate gene 8) and WDR41 (WD40 repeat-containing protein 41), to carry out the mentioned functions, including their effect on macrophages and microglia [57,60]. Due to the nature of this procedure, this function has been proposed to be affected in ALS when in the presence of mutations, but further studies are needed [57,59]. C9ORF72 mutation is non-also the most common mutation in ALS but is also responsible for FTD. This mutation is reflected as an increase in the number of hexanucleotide (G4C2)n repeat expansions (HRE) in the non-coding region of C9ORF72, which results in both loss-of-function linked to C9ORF72 haploinsufficiency and a gain of function resulting in the expression of abnormal bidirectionally transcribed RNAs carrying the repeat [61]. This repeat expansion leads to abnormal RNA molecules biosynthesis, which are then translated into dipeptide repeat proteins (DPRs) that contain multiple copies of the specific amino acid sequence GGGGCC. DPRs such as poly-proline-arginine (poly-PR), poly-glycine-arginine (poly-GR), and poly-glycine-alanine (poly-GA) are cytotoxic [62], and accumulate in neurons [63], and may then spread to glial cells via intercellular communication [64], thus impairing protein folding and transport, inducing oxidative stress and disrupting mitochondrial function [65]. An important player in the pathophysiology of ALS patients carrying C9ORF72 expansion is poly-GA which induces the intracellular aggregation of phosphorylated TDP-43 proteins, through the impairment of TDP43 nuclear translocation and cytoplasm mislocation [66,67]. In fact, a pathological hallmark of these patients is the presence of TDP-43 inclusions in neurons and oligodendroglial cells. The C9ORF72 gene was also recently associated with nucleolar dysfunction [68] and DNA repair inhibition [69]. Another important cellular process that is affected by C9ORF72 gene mutation is vesicular and protein trafficking [70] C9ORF72 HRE was found to reduce the interaction between C9orf72 and the Rab GTPase key regulator Rab7L1, resulting in a decreased extracellular vesicles (EVs) release [70]. The role of C9ORF72 in protein trafficking was further demonstrated in the human spinal cord of an ALS patient (with C9ORF72-intronic repeat expansion mutation), where an increased proportion of motor neurons showed colocalization of C9orf72 with Rab 5, Rab 7 and Rab 11 (when compared to healthy individuals) possibly resulting in the dysregulation of endosomal trafficking [70]. Interestingly, these proteins were recognized to be tangled with vesicle trafficking regulation from the multivesicular bodies (MVB) to the plasma membrane, being involved, among other instances, in autophagy [70].
On its own, TARDBP, which codes for TDP-43, is also another commonly mutated gene in ALS. In normal physiological conditions, TDP-43 is primarily found in the nucleus, where it participates in the regulation of gene expression [71]. However, mutations in this gene in ALS or FTD patients lead to a mislocalization of the corresponding protein, accumulating in the cytoplasm in the form of abnormal TDP-43 aggregates, and generating anomalous ubiquitin-positive inclusions in the nucleus and cytoplasm [72]. Those inclusions can affect physiological functions of p62 (also known as SQSTM), involved in autophagy and proteasome regulation. Sequestration of p62 within the TDP-43 aggregates leads to autophagy and proteasome functions impairment, driving further accumulation of misfolded proteins within the cells [73,74]. Indeed, aggregates co-localizing TDP-43 with p62, and also with SOD1, were found in post-mortem ventral spinal cord tissues of fALS and sALS patients, despite the existence of different aggregation profiles among them [74]. This can occur even in the absence of mutations in the respective genes, which may be attributed to incorrect protein folding, namely of SOD1 [74,75].
Another commonly ALS-linked mutated gene is FUS, which encodes for the RNA- binding protein FUS. In healthy individuals, FUS is found in the nuclei and relates to gene expression regulation, DNA repair, and RNA processing [76]. Yet, in ALS and FTD patients, FUS translocates into the cytoplasm, creating FUS inclusions, that can boost further nefarious effects, such as RNA mislocation associated with sequestering of the motor protein kinesin-1 [77], and axonal transport impairments [78]. FUS mutations in ALS may also impair mitochondrial function through sequestration of respiratory chain complex mRNAs in the cytoplasm [79]. Moreover, FUS loss of function can lead to neuronal dysfunction and death [80]. It is possible that FUS mislocation into the cytoplasm may contribute to their incorporation into EVs, and then, by dissemination to other cells via intercellular transfer, ALS disease’s phenotype is spread onto circulation [81,82].
2. Extracellular Vesicles and Their Role in ALS Onset and Development
2.1. EVs Overview
Extracellular vesicles (EVs) are endogenous bilipid layer, plasma membrane, or endosome-derived nanoparticles released by most eukaryotic cells into the extracellular space [83]. They were first described by [84] and initially thought to be a cellular waste product. Most literature reports that cells can synthesize and secrete three main types of EVs: exosomes or exosome-like vesicles, microvesicles or ectosomes, and apoptotic bodies [85,86]. However, more recently, other types of EVs were reported, such as retrovirus-like vesicles and mitovesicles. The former are 90-100 nm particles that possess a subset of retroviral proteins and carry endogenous retroviral sequences, but not for cellular entry nor retroviral propagation [87]. Mitovesicles are of mitochondrial origin, possessing components of this organelle, such as mitochondrial proteins, lipids, and mitochondrial DNA (mtDNA) [88]. mitovesicles are distinguishable from exosomes and microvesicles by their morphology, size, and content [88]. EVs classification relies on several parameters, such as their size, content, function, biogenesis, and release pathways, among others [85]. EVs’ biological functions depend on their type and their highly specific bioactive cargo, which represents the progenitor cell state [89,90]. There are different ways to identify EVs, such as by physical characterization through microscopy, proteomic analysis, RNA sequencing, functional characterization, and the biochemical analysis of their composition [91]. An important way to identify EVs is through the presence of specific surface protein markers, which may depend on many factors, such as their origin. In the case of exosomes, some proteins tendentially common among them and often used in their identification include annexin, CD9, CD63, CD81, HSP70, and flotillin [92,93].
It has been recognized that EVs play a fundamental role in intercellular communication, functioning as vehicles for transporting and delivering a range of cellular bioactive cargos, including membrane and cytosolic proteins, lipids, DNA, mRNA (messenger RNA), and miRNA (microRNA) [94,95]. Through the action of their specific shuttled content, EVs may directly influence recipient cells’ cellular state. That occurs, for instance, via miRNA-induced gene expression post-transcriptional regulation processes [96], which include numerous cellular epigenetic regulations [97,98]. EVs have a role in the maintenance of cellular homeostasis by being pivotal in cellular uptake mechanisms [99]. An example of this is the ligand/receptor interaction within brain synaptic transmission [100,101]. EVs are also important in the maintenance of stem cell plasticity [102] and in the formation of new tissues, since they are important for angiogenesis [103,104], for the generation of morphologic gradients for tissue genesis along neuronal development [105,106], and neuronal regeneration [107,108].
Regarding their release pathways, EVs are delivered into the extracellular space via SNARE-mediated fusion of multivesicular endosomes with the plasma membrane [109]. The direct budding of vesicles with the plasma membrane results in microvesicles [110,111]. Additionally, those vesicles that may shed from cells undergoing programmed cell death, originate the apoptotic bodies [112]. Following exocytosis, EVs may remain in the extracellular space, surrounding the secreter cell or, instead, travel elsewhere, such as into the brain, by crossing the blood-brain-barrier (BBB) [113], or from the brain into the periphery. As within the brain, exosomes are released by several cell types, such as by neurons [114], microglial cells [115], astrocytes [116], and oligodendrocytes [117].
2.2. The Role of EVs in ALS
EVs have been associated with numerous pathologies, from metastatic cancers [122] to neurodegenerative diseases, such as Alzheimer’s disease, Parkinson’s disease, among many others [123,124,125]. Under such pathological conditions, EVs shuttle enclosed misfolded proteins and other neurotoxic elements that potentially induce dysfunction in the recipient cells [125,126]. EVs are being increasingly recognized as being of great importance also in the pathogenesis of ALS and in the identification of biomarkers, which will be explored in this section (Figure 1).
2.2.1. EVs in ALS Disease Progression and Pathological Mechanisms
EVs have emerged as significant players in the progression of ALS, with increasing evidence pointing to their role in the dissemination of detrimental biocargos. EVs allow for the hypothetical prion-like propagation of ALS-related mutant misfolded proteins and dysregulated miRNAs [81], which are believed to contribute to the severity and progression of the disease [127,128,129]. The most common cargos found in EVs from ALS patients include misfolded proteins such as mSOD1, FUS, TDP43, C9orf72 expansions DPRs, and other neurotoxic elements [64,81]. These harmful cargos have been screened in both astrocytes and neuronal-derived exosomes in different ALS disease models, such as the SOD1-G93A mice model, one of the most commonly used animal models for studying ALS. In this model, the mutated SOD1 gene harbors a substitution of glycine for alanine at the codon 93. Recently, [130] demonstrated that mutant SOD1 (mSOD1) accumulation occurs in cellular vacuoles, which may be constituted by different portions of organelles, and once released, lead to the existence of different types of EVs, particularly mitoEVs. The formation and type of these vacuoles, and resulting EVs, appear to be related to the stage of ALS pathology in this mice model. Interestingly, it seems that before motor symptoms onset, those vacuoles are already present and are mainly of mitochondrial origin, having a high content of mSOD1, ultimately resulting in the release of mSOD1-containing EVs [130]. The authors of this study hypothesized that these EVs, derived from damaged neurons, may be responsible for the initiation of a sequence of signaling cascades that contribute to neuroinflammation, glial-mediated neurotoxicity, and prion-like spreading of the disease. The existence of different vacuoles/EVs phenotypes and associated cell death pathways may have different roles in the onset and severity of symptoms, as well as in the heterogeneity, and progression of the disease.
Exosomal TDP-43 is another significant cargo that plays a crucial role in ALS progression. A longitudinal study conducted on ALS patients demonstrated an increase in the exosomal TDP-43 ratio in peripheral blood during the course of the disease, particularly in the early stages [131]. This rise in TDP-43 ratio is associated with elevated levels of neurofilament light chain (NFL) in the plasma of these patients, which is more prevalent in individuals with a rapid disease progression [131]. Further evidence supports the significance of exosomal TDP-43 in disease propagation. Ding et al. (2015) described the damaging effect of exosomes enclosing TDP-43 C-terminal fragments (CTFs) from the cerebrospinal fluid of ALS patients with FTD (ALS-FTD-CSF) in human glioma cells (U251 cells). After being incubated with the ALS-FTD-CSF-derived exosomes, naive U251 cells developed intracellular TDP-43 aggregates in the form of tunneling nanotubes (TNTs)-like structures [132]. Although in vivo studies are required, this previous work suggests that EVs may act as a vehicle for the spread of TDP-43 aggregates in the context of ALS.
EVs and their toxic payloads not only damage neurons but also spread pathological signaling by transferring them between different cell types, including neurons, astrocytes, and muscle cells. Evidence of these interactions was provided by a study showing that EVs-mediated transfer of DRPs occurred between MNs-like NSC34 cells and rat cortical neurons, and then from these to rat cortical astrocytes [64]. This transfer is relevant to ALS, as EVs carrying C9orf72-encoded DPRs were identified to be involved in the exchange between human C9orf72 induced pluripotent stem cell-derived motor neurons (hiPSC-MNs) and control iPSC-derived spinal MNs [64]. In NSC34 cells transfected with mutant SOD1(G93A) (hSOD1-G93A NSC34 cells), miR-124 was found to be upregulated and transferred to EVs. When these cells were co-cultured with N9-microglial cells, miR-124 contained in mSOD1 exosomes was translocated to N9-microglial cells, resulting in phenotypic alterations such as the reduction in their phagocytic capability and activation of neuroinflammation pathways [133]. Exosomes released by mouse astrocytes overexpressing G93A SOD1 were also previously shown to be responsible for the transfer of mutant SOD1 to mouse spinal neurons and induce MNs death [134]. Moreover, astrocytic-derived exosomes from the plasma of sALS patients were found to transport inflammation-related cargos, including IL-6, a pro-inflammatory interleukin, which was increased in these vesicles and positively associated with the rate of disease progression [135]. The negative impact of EVs and their cargos on the interaction of affected muscle cells with MNs was further demonstrated by evidence showing that multivesicular bodies released from ALS muscle cells were neurotoxic to healthy MNs [136]. In this study, EVs derived from muscle cells obtained from biopsies of sALS patients were exposed to healthy hiPSC-MNs and were shown to be neurotoxic through increased FUS expression, resulting in shorter and less branched neurites, atrophic myotubes, and enhanced cell death [136]. The observed cell death was greatly reduced by immunoblocking the vesicle uptake by MNs with anti-CD63. Finally, a study by Anakor and colleagues supports the cause-and-effect relationship between muscle cell vesicles and MNs. Exposure of MNs to ALS patients’ skeletal muscle cells-derived exosome-like vesicles (MuVs) resulted in reduced neurite length, number of neurite branches, and reduced MNs survival and myotubes by 31% and 18%, respectively. Moreover, adding ALS-derived MuVs to healthy astrocytes led to an increase in the proportion of stellate astrocytes and, thus to mild activation of these cells [137].
2.2.2. miRNAs and Misfolded Proteins EVs Cargo in ALS: Potential ALS Biomarkers
One particular cargo of EVs, microRNAs (miRNAs), has attracted research interest as potential biomarkers for ALS due to their versatile functions in regulating gene expression across a wide range of processes, including neural development, cell proliferation and differentiation, protein ubiquitination, apoptosis, and other transcriptional regulatory processes (summarized in Table 1). Despite their link to ALS progression, the mechanisms underlying alterations in their expression and levels remain inconclusive. In fact, defective RNA metabolism and miRNA dysregulations are closely associated with ALS [138]. miRNA profiles in ALS exhibit significant variations among patients and can be over- or under-expressed, as they are transported by EVs across multiple biofluids and tissues (summarized in Table 1). Most of the research has been focused on screening ALS patients’ plasma-circulating EVs using a variety of research methodologies, ranging from RT-qPCR analysis to microarrays [139].
In the quest for an ALS molecular biomarker fingerprint, [156] reported a downregulation of miR-27a-3p in ALS patients’ serum-derived exosomes. [143] found 27 differentially expressed miRNAs, 5 of them upregulated and 22 downregulated when comparing, via next-generation sequencing, EVs isolated from ALS patients’ plasma samples with those from healthy controls. Some of these miRNAs were relevant to ALS diagnosis as they were related to Revised ALS Functional Rating Scale (ALSFRS-R) scores. This is the case for miR-193a-5p, which allowed distinguishing patients with a low versus high score. miR-15a-5p was demonstrated to be important in differentiating controls from ALS patients. In a separate study, [151] analyzed miRNA profiles in neuron-derived EVs from ALS patients’ plasma samples via microarrays, and identified 30 differentially expressed miRNAs, 13 up-regulated and 17 down-regulated. In another study, [270] performed miRNA screening in ALS patients’ serum-derived exosomes, using RT-qPCR analysis. Due to the small sample size used in this study, no statistically significant differences were observed in the expression levels of any miRNA. However, a recent study by [153] analyzed the miRNA cargo profiles of EVs isolated from postmortem homogenates of frontal cortex, spinal cord, and serum of sALS patients. The authors found no difference in the number of EVs between patients and controls, but ALS patients presented larger spinal cord vesicles and smaller-sized vesicles in serum. Two miRNAs related to axon guidance and long-term potentiation were significantly dysregulated in all analyzed tissues: miR-342-3p was upregulated, and miR-1254 was downregulated. Furthermore, miRNA levels were reduced in the frontal cortex and spinal cord of sALS patients, while they were increased in serum. Another study by [160] analyzed EVs isolated from MNs cultures, obtained from fibroblasts-reprogrammed iPSCs of ALS patients carrying C9ORFf72, SOD1, and TARDBP mutations. These authors found dysregulation of several miRNAs, specifically upregulations in miR-629-5p and miR-194-5p levels and downregulations in miR-34a-5p, miR-1267, and miR-625-3p. Interestingly, the latter was found to be constantly downregulated in C9orf72 MNs-exosomes and upregulated in EVs from TARDBP-MNs. In the same study, miR-625-3p was predicted to be a mediator in cell-to-cell communication, immune system pathways, and autophagy. Furthermore, in another study from the same authors [271], using iPSC-derived MNs progenitors from fALS and sALS patients, further dysregulation was found, notably of miR-34a, which is involved in cell cycle regulation, autophagy, apoptosis, neurogenesis, and neuronal differentiation [272]. Sproviero et al. (2021) also searched for potential ALS EVs miRNA biomarkers and found dysregulation of hsa-miR-206, hsa-miR-205-5p, miR-1-3p, hsa-miR-205-5p, hsa-miR-200b-3p, hsa-miR-200c-3p, hsa-miR-6888-3p, hsa-miR-31-5p, hsa-miR-141-3p, and hsa-miR-210-3p in the plasma of ALS patients [273]. In a different approach, Yelick et al. (2020), using in situ hybridization analysis, found a downregulation of miR-124-3p in exosomes from SOD1-G93A mice spinal MNs. Moreover, in this study, the authors found a significant correlation between cerebrospinal fluid (CSF) exosomal miR-124-3p expression levels and the disease stage of male ALS patients, as denoted by the ALSFRS-R score [147]. It is worth noting that miR-124-3p is a recognized oncogene [274,275] with an essential role in cell proliferation and apoptosis [274], being linked with poor survival rates in hepatocellular carcinoma patients [276]. Conversely, its upregulation was shown to decrease the metastatic behavior of hepatocarcinoma cells, through the reversion of CRKL expression, which resulted in the suppression of the extracellular signal- regulated kinase (ERK) pathway and the malignant cells proliferation inhibition [277]. Importantly, its upregulation was found to be protective against post-traumatic neurodegeneration through the activation of Rela/ApoE signaling pathway [278] and which downregulation was linked with neurodegeneration and neuroinflammatory states post-traumatic brain injury (TBI) [279].
Other miRNAs differentially expressed in serum-derived extracellular vesicles from 50 ALS patients were reported recently by [148]. Statistically significant robust results yielded a differential expression of seven miRNAs included in extracellular vesicles, two of them upregulated (miR-151a-5p, miR-146a-5p), and three downregulated (miR-4454, miR-10b-5p, miR-29b-3p) [148]. Among the reported functions, those specific miRNAs have been found to be linked with tumorigenesis [280,281,282] and protection against cell apoptosis [283].
Despite the recent advances in understanding the role of miRNAs associated with EVs, in driving the progression of ALS, this field is still in its early stages. Analysis of miRNA expression profiles suggests that the current knowledge is insufficient to predict their involvement in the pathological mechanisms of ALS [284]. In a study that analyzed the results of research from 2013 to 2018, Foggin et al. (2019) reported that most of the dysregulated miRNAs were either up or downregulated. This outcome may be due to intrinsic differences in the methodologies used for miRNA detection or to other factors, such as different miRNA expression across different tissues and sample extraction protocols. Interestingly, eight of the nine most commonly dysregulated miRNAs were predicted to target at least one of the most commonly mutated genes in ALS, but a random sample of unrelated miRNAs that were not found to be dysregulated in ALS patients also retrieved a similar prediction [284]. Nonetheless, the search for miRNAs as potential biomarkers for ALS remains promising, due to their good preservation in different types of biological samples, such as CSF and blood, often with an advantage over several proteins in allowing for a more reliable and faster diagnosis and a closer classification and understanding of each case. In this scope, as suggested by [160], it is important to analyze miRNAs isolated from different human biological samples (e.g., MNs, exosomes, and CSF) of different ALS types. Likewise, miR-206 has been proposed to be a potential biomarker in a study by Toivonen et al. (2014) since it displayed consistent changes towards its upregulation across ALS disease progression in the SOD1 mice [285]. miR-206 is a microRNA that has been identified as a tumor suppressor involved in regulating the transforming growth factor-β (TGF-β) signaling pathway [286]. It is considered a myomiR because it is highly expressed in skeletal muscle [287] and plays a vital role in myogenesis and skeletal tissue regeneration [288,289]. In several studies related to ALS pathology, consistent expression levels of miR-206 have been observed across different biological samples. For example, miR-206 was found to be overexpressed in the serum of sALS patients [290] and in both plasma and skeletal muscles of spinal onset ALS patients [291]. In a study performed with the SOD1-G93A ALS mice model, [288] investigated the role of miR-206 in ALS disease progression. Their findings suggested that miR-206 overexpression was associated with the onset of neurological symptoms, which may be attributed to skeletal muscle denervation. Indeed, when miR-206 was intentionally downregulated, the restoration of neuromuscular synapses was observed, indicating the potential of miR-206 as a therapeutic target for ALS.
In addition to miRNAs, the protein cargo of EVs associated with ALS may also hold potential as novel biomarkers (summarized in Table 2). In a study by [292], 12 proteins were identified as being exclusive to EVs derived from post-mortem motor cortex tissue of ALS patients, including CD177, CHMP4B, CSPG5, DYNC1I2, IGHV3-43, LBP, RPS29, S100A9, SAA1, SCAMP4, SCN2B, and SLC16A1. Additionally, [293] discovered a potential new method for patient stratification based on levels of cyclophilin A, a protein involved in TDP-43 trafficking and function, in combination with the EVs size distribution in plasma-derived EVs from ALS patients. This approach could distinguish between slow and fast progression of the disease. Recently, [294] conducted a pilot study comparing the CSF and CSF-EVs from ALS patients and matched control subjects to search for novel ALS biomarker candidates. They found four differentially expressed proteins in ALS patients’ CSF, including downregulated MB and upregulated JAM-A, TNF-R2, and CHIT1. Although no proteins were differentially expressed in CSF-EVs, there was a trend for the downregulation of perlecan, a proteoglycan of the extracellular matrix involved in cell proliferation, differentiation, adhesion, migration, tissue repair and regeneration [295]. Conversely, Thompson et al. (2020) found no significant differences in terms of CSF-EVs concentration and size distribution between control and ALS groups. However, they did identify altered protein homeostatic mechanisms in ALS patients, including the downregulation of bleomycin hydrolase [296], a cytosolic cysteine protease that has been connected to the release of chemokines in inflammation and wound healing processes [297]. These data, together with those indicating the involvement of EVs in aggregated protein spread, suggest that the analysis of EVs protein content is mandatory for the development of innovative diagnostic/prognostic tools and even the identification of new therapeutic targets for ALS. Overall, while these studies have provided promising results, further research is needed to understand the role of miRNA and proteins transported by EVs in ALS development and progression, and their possible use as biomarkers.
3. Therapeutic Perspectives with EVs in ALS
In recent years, several studies have proposed innovative next-generation EVs-related therapies, which hold great promise for the treatment of human diseases [326]. EVs present several therapeutic advantages due to higher biocompatibility and reduced immunogenicity over alternative carriers, such as some synthetic nanocarriers, that may also be prone to accumulation in the liver and spleen [327,328]. As natural nanoparticles, EVs can be easily isolated from various biofluids and can cross biological barriers to deliver potential therapeutics (Das et al. 2019). Despite some uncertainty regarding their functional mechanisms, it is becoming clear that these bioparticles shuttle diverse cargos capable of recapitulating the benefits of “whole-cell therapy,” either by preventing or mitigating abnormal cellular functions [329]
One of the emerging therapeutic approaches related to EVs is the use of stem cell-derived EVs. These EVs have a positive impact on the pathophysiology of different neurodegenerative diseases [330]. In the case of ALS, these EVs may achieve beneficial effects by modulating the immune system, addressing mitochondrial dysfunction, and boosting MNs’ neuroprotection [331,332,333]. For example, exosomes derived from adipose-derived stem cells (ASCs) obtained from SOD1-G93A mice were shown to have a neuroprotective effect by reducing oxidative stress-related damage in MN-like NSC-34 cells that overexpressed ALS-associated mutations, including SOD1(G93A), SOD1(G37R), and SOD1(A4V) [332]. Furthermore, the same research group observed that NSC-34(G93A) cells internalized ASC-derived exosomes, leading to the downregulation of pro-apoptotic proteins (Bax and cleaved caspase-3), and the upregulation of anti-apoptotic proteins (Bcl-2α), ultimately improving neuronal survival [334]. In a more recent study by these authors, ASC-derived exosomes obtained from SOD1-G93A mice were used to slow the progression of ALS by reducing glial cell activation and improving motor performance. Interestingly, these exosomes showed an affinity towards the lesioned areas of the brain, suggesting targeted delivery, although the exact mechanisms behind this phenomenon still need further elucidation [335]. Similarly, ASCs-derived exosomes were found to increase the expression levels of phospho-CREB/CREB and PGC-1α in neurons derived from neural stem cells of SOD1-G93A mice. This resulted in a reduction of cytosolic SOD1 aggregates and rescued mitochondrial dysfunction [315]. Additionally, the same type of exosomes was shown to rescue the inherent impairment in oxidative phosphorylation (OXPHOS) specifically linked to the mitochondrial complex I in NSC-34(G93A) cells [336]. In their study, human bone marrow endothelial progenitor cells (hBM-EPC)-derived exosomes were shown to restore mouse brain endothelial cells previously damaged through in vitro exposure to SOD1-G93A mutant male mice plasma. The results indicated a significant reduction in microvascular endothelium damage. Interestingly, blocking the β1 integrin of exosomes using an anti-CD29 blocking antibody prevented their internalization by recipient cells, thereby increasing brain endothelial cell death percentage. These findings suggest that hBM-EPC-derived exosomes have the potential to repair endothelial damage in ALS and that their internalization by recipient cells may play a critical role in their therapeutic effects. In another study, Garbuzova-Davis et al. (2020) investigated the potential therapeutic role of exosomes derived from human hBM-EPCs in repairing endothelial damage in ALS. To induce damage, the researchers exposed a mouse brain endothelial cell line to plasma from SOD1-G93A male mice. They found that ALS animals’ plasma-derived exosomes treatment significantly increased endothelium cell death. However, a significant reduction in cell death was obtained by supplementing brain endothelium, previously exposed to ALS plasma derived exosomes, with 1 μg/mL of the hBM-EPC-derived exosomes for 24 hours. Moreover, when those EVs were pre-treated with an anti-CD29 blocking antibody to block β1 integrin, EVs were prevented from being internalized by recipient cells, resulting in a significant increase in brain endothelium cell death. These findings suggest that hBM-EPC-derived exosomes have the potential to reduce the number of damaged endothelial cells in ALS, but their beneficial effects may be dependent on proper internalization by cells [337]. On the other hand, the negative effects of ALS-related EVs were reverted in a study by Varcianna et al. (2019). The authors isolated EVs from human induced astrocytes derived from C9ORF72-ALS sALS patients (C9ORF72-ALS iAstrocytes-derived EVs) and found that originally they compromised both neurite network maintenance and MNs survival of the HB9-GFP+ mouse cultured MNs (Hb9-GFP+ MNs). Such an effect was related to the downregulation of miR-494-3p content in those EVs. Nevertheless, following treatment with the C9ORF72-ALS iAstrocytes-derived EVs, where miR-494-3p levels were intentionally upregulated, HB9-GFP+ mouse cultured motor neurons presented neurite network restoration and decreased MNs death. These beneficial effects of miR-494-3p overexpression may be related to its function as a negative regulator of semaphorin 3A (SEMA3A) and other targets involved in axonal maintenance [203].
Besides the described therapeutic possibilities, EVs have emerged as promising drug carriers, with the potential to deliver synthetic drugs to the brain. This is especially important as many proteic and small-molecule neurological drugs may fail to bypass the blood-brain barrier (BBB), which can hinder their effectiveness [338]. Encapsulating these drugs within EVs could help overcome this limitation by allowing them to cross the BBB and improve drug targeting and efficiency [339]. While EVs are not currently being used to deliver drugs for ALS treatment, they have been employed in the treatment of other brain diseases, such as brain tumors, using doxorubicin-loaded exosomes [340]. Therefore, this approach may also be a promising avenue for future ALS research.
4. Conclusions
ALS is a fatal neurodegenerative disease with a complex and unclear etiopathology that strongly impacts patients’ health and well-being. With no cure available so far, searching for an effective treatment that can improve patients’ life expectancy and quality of life is paramount. ALS presents several important challenges and hurdles to its research, due to the intrinsic complexity and heterogeneity of the disease. Nevertheless, important advances have been made in recent years. Among these are the recent advances in ALS-related EVs research, which are emerging as key players in the surfacing and development of the disease by allowing for the transport of biomolecular cargo from cell to cell, thus spreading the anomalies across the system.
EVs also have promising potential to be employed as a source of potential biomarkers for early detection of ALS and personalized prognostic purposes. Furthermore, they may also be exploitable to tackle existing altered mechanisms and for application in a variety of therapeutic strategies, such as being employed for drug delivery purposes, as they can carry different types of molecules, both natural and artificial. Specifically, within the ALS research area, stem cell-derived EVs’ use is arising for therapeutic purposes, of higher relevance given the disease heterogeneity and allowing for a precision-based approach.
While the use of stem cell-derived EVs for therapeutic purposes is promising, further innovative and consensual approaches are needed to reverse the disease’s biopathologic mechanisms and translate this knowledge into real-life applications that can bring hope to both patients and their families.
Author Contributions
GA, CC, SM, and EF have reviewed the literature and wrote the first draft of the manuscript. JV, SM and EF edited and revised the manuscript. Artwork performed by JV. All authors contributed to manuscript revision, read, and approved the submitted version.
Funding
This work was financed by the European Regional Development Fund (ERDF), through the Centro 2020 Regional Operational Programme, through the COMPETE 2020 - Operational Programme for Competitiveness and Internationalisation and Portuguese national funds via FCT – Fundação para a Ciência e a Tecnologia, under the project PTDC/BTM-ORG/0055/2021, and UIDB/04539/2020, UIDP/04539/2020 and LA/P/0058/2020. CC is supported by FCT (Ref: 2022.13281.BD). SM is supported by FCT (Ref: DL57/2016/CP1448/CT0027). GA is supported by FCT under a fellowship from project PTDC/BTM-ORG/0055/2021. EF was supported by FCT under CEECIND/00322/2017 and is supported by FCT under 2022.00011.CEECIND.
Acknowledgments
The authors want to acknowledge the helpful revision and suggestions given by Dr. Paulo J. Oliveira. The authors also thank the colleagues at MitoXT lab (https://cnc.uc.pt/pt/research-group/mitoxt-toxicologia-e-terapeutica-experimental-mitocondrial) for fruitful discussions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- van Es, M.A.; Hardiman, O.; Chio, A.; Al-Chalabi, A.; Pasterkamp, R.J.; Veldink, J.H.; van den Berg, L.H. Amyotrophic Lateral Sclerosis. Lancet 2017, 390, 2084–2098. [Google Scholar] [CrossRef]
- Cividini, C.; Basaia, S.; Spinelli, E.G.; Canu, E.; Castelnovo, V.; Riva, N.; Cecchetti, G.; Caso, F.; Magnani, G.; Falini, A.; et al. Amyotrophic Lateral Sclerosis-Frontotemporal Dementia: Shared and Divergent Neural Correlates Across the Clinical Spectrum. Neurology 2021, 98, e402–e415. [Google Scholar] [CrossRef] [PubMed]
- Strong, M.J.; Abrahams, S.; Goldstein, L.H.; Woolley, S.; Mclaughlin, P.; Snowden, J.; Mioshi, E.; Roberts-South, A.; Benatar, M.; HortobáGyi, T.; et al. Amyotrophic Lateral Sclerosis - Frontotemporal Spectrum Disorder (ALS-FTSD): Revised Diagnostic Criteria. Amyotroph. Lateral Scler. Frontotemporal Degener. 2017, 18, 153–174. [Google Scholar] [CrossRef] [PubMed]
- Saxon, J.A.; Thompson, J.C.; Jones, M.; Harris, J.M.; Richardson, A.M.; Langheinrich, T.; Neary, D.; Mann, D.M.; Snowden, J.S. Examining the Language and Behavioural Profile in FTD and ALS-FTD. J. Neurol. Neurosurg. Psychiatry 2017, 88, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Longinetti, E.; Fang, F. Epidemiology of Amyotrophic Lateral Sclerosis: An Update of Recent Literature. Curr. Opin. Neurol. 2019, 32, 771–776. [Google Scholar] [CrossRef]
- Renton, A.E.; Chiò, A.; Traynor, B.J. State of Play in Amyotrophic Lateral Sclerosis Genetics. Nat. Neurosci. 2014, 17, 17–23. [Google Scholar] [CrossRef]
- McCann, E.P.; Henden, L.; Fifita, J.A.; Zhang, K.Y.; Grima, N.; Bauer, D.C.; Chan Moi Fat, S.; Twine, N.A.; Pamphlett, R.; Kiernan, M.C.; et al. Evidence for Polygenic and Oligogenic Basis of Australian Sporadic Amyotrophic Lateral Sclerosis. J. Med. Genet. 2020. [CrossRef]
- Jaiswal, M.K. Riluzole and Edaravone: A Tale of Two Amyotrophic Lateral Sclerosis Drugs. Med. Res. Rev. 2019, 39, 733–748. [Google Scholar] [CrossRef]
- Oliveira Santos, M.; Gromicho, M.; Pinto, S.; De Carvalho, M. Clinical Characteristics in Young-Adult ALS - Results from a Portuguese Cohort Study. Amyotroph. Lateral Scler. Frontotemporal Degener. 2020, 21, 620–623. [Google Scholar] [CrossRef]
- Cunha-Oliveira, T.; Montezinho, L.; Mendes, C.; Firuzi, O.; Saso, L.; Oliveira, P.J.; Silva, F.S.G. Oxidative Stress in Amyotrophic Lateral Sclerosis: Pathophysiology and Opportunities for Pharmacological Intervention. Oxid. Med. Cell. Longev. 2020, 2020, 5021694. [Google Scholar] [CrossRef]
- McCombe, P.A.; Henderson, R.D. Effects of Gender in Amyotrophic Lateral Sclerosis. Gend. Med. 2010, 7, 557–570. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, E.; Factor-Litvak, P.; Santella, R.M.; Mitsumoto, H. Clinical Perspective on Oxidative Stress in Sporadic Amyotrophic Lateral Sclerosis. Free Radic. Biol. Med. 2013, 65, 509–527. [Google Scholar] [CrossRef] [PubMed]
- Weisskopf, M.G.; McCullough, M.L.; Calle, E.E.; Thun, M.J.; Cudkowicz, M.; Ascherio, A. Prospective Study of Cigarette Smoking and Amyotrophic Lateral Sclerosis. Am. J. Epidemiol. 2004, 160, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Zeng, P.; Zhou, X. Causal Effects of Blood Lipids on Amyotrophic Lateral Sclerosis: A Mendelian Randomization Study. Hum. Mol. Genet. 2019, 28, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Seelen, M.; Toro Campos, R.A.; Veldink, J.H.; Visser, A.E.; Hoek, G.; Brunekreef, B.; van der Kooi, A.J.; de Visser, M.; Raaphorst, J.; van den Berg, L.H.; et al. Long-Term Air Pollution Exposure and Amyotrophic Lateral Sclerosis in Netherlands: A Population-Based Case-Control Study. Environ. Health Perspect. 2017, 125, 097023. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; Kwee, L.C.; Allen, K.D.; Umbach, D.M.; Ye, W.; Watson, M.; Keller, J.; Oddone, E.Z.; Sandler, D.P.; Schmidt, S.; et al. Association between Blood Lead and the Risk of Amyotrophic Lateral Sclerosis. Am. J. Epidemiol. 2010, 171, 1126–1133. [Google Scholar] [CrossRef] [PubMed]
- Park, R.M.; Schulte, P.A.; Bowman, J.D.; Walker, J.T.; Bondy, S.C.; Yost, M.G.; Touchstone, J.A.; Dosemeci, M. Potential Occupational Risks for Neurodegenerative Diseases. Am. J. Ind. Med. 2005, 48, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Jafari, H.; Couratier, P.; Camu, W. Motor Neuron Disease after Electric Injury. J. Neurol. Neurosurg. Psychiatry 2001, 71, 265–267. [Google Scholar] [CrossRef]
- Gallagher, J.P.; Sanders, M. Trauma and Amyotrophic Lateral Sclerosis: A Report of 78 Patients. Acta Neurol. Scand. 1987, 75, 145–150. [Google Scholar]
- Coffman, C.J.; Horner, R.D.; Grambow, S.C.; Lindquist, J. ; VA Cooperative Studies Program Project #500 Estimating the Occurrence of Amyotrophic Lateral Sclerosis among Gulf War (1990-1991) Veterans Using Capture-Recapture Methods. Neuroepidemiology 2005, 24, 141–150. [Google Scholar]
- Chiò, A.; Benzi, G.; Dossena, M.; Mutani, R.; Mora, G. Severely Increased Risk of Amyotrophic Lateral Sclerosis among Italian Professional Football Players. Brain 2005, 128, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Scarmeas, N.; Shih, T.; Stern, Y.; Ottman, R.; Rowland, L.P. Premorbid Weight, Body Mass, and Varsity Athletics in ALS. Neurology 2002, 59, 773–775. [Google Scholar] [CrossRef] [PubMed]
- Julian, T.H.; Glascow, N.; Barry, A.D.F.; Moll, T.; Harvey, C.; Klimentidis, Y.C.; Newell, M.; Zhang, S.; Snyder, M.P.; Cooper-Knock, J.; et al. Physical Exercise Is a Risk Factor for Amyotrophic Lateral Sclerosis: Convergent Evidence from Mendelian Randomisation, Transcriptomics and Risk Genotypes. EBioMedicine 2021, 68, 103397. [Google Scholar] [CrossRef]
- Oskarsson, B.; Horton, D.K.; Mitsumoto, H. Potential Environmental Factors in Amyotrophic Lateral Sclerosis. Neurol. Clin. 2015, 33, 877–888. [Google Scholar] [CrossRef]
- Zhang, G.; Zhang, L.; Tang, L.; Xia, K.; Huang, T.; Fan, D. Physical Activity and Amyotrophic Lateral Sclerosis: A Mendelian Randomization Study. Neurobiol. Aging 2021, 105, 374–e1. [Google Scholar] [CrossRef]
- Savage, A.L.; Lopez, A.I.; Iacoangeli, A.; Bubb, V.J.; Smith, B.; Troakes, C.; Alahmady, N.; Koks, S.; Schumann, G.G.; Al-Chalabi, A.; et al. Frequency and Methylation Status of Selected Retrotransposition Competent L1 Loci in Amyotrophic Lateral Sclerosis. Mol. Brain 2020, 13, 154. [Google Scholar] [CrossRef] [PubMed]
- Morahan, J.M.; Yu, B.; Trent, R.J.; Pamphlett, R. A Genome-Wide Analysis of Brain DNA Methylation Identifies New Candidate Genes for Sporadic Amyotrophic Lateral Sclerosis. Amyotroph. Lateral Scler. 2009, 10, 418–429. [Google Scholar] [CrossRef]
- Figueroa-Romero, C.; Hur, J.; Bender, D.E.; Delaney, C.E.; Cataldo, M.D.; Smith, A.L.; Yung, R.; Ruden, D.M.; Callaghan, B.C.; Feldman, E.L. Identification of Epigenetically Altered Genes in Sporadic Amyotrophic Lateral Sclerosis. PLoS One 2012, 7, e52672. [Google Scholar] [CrossRef]
- Cai, Z.; Jia, X.; Liu, M.; Yang, X.; Cui, L. Epigenome-Wide DNA Methylation Study of Whole Blood in Patients with Sporadic Amyotrophic Lateral Sclerosis. Chin. Med. J. 2022, 135, 1466–1473. [Google Scholar] [CrossRef]
- Chen, L.-X.; Xu, H.-F.; Wang, P.-S.; Yang, X.-X.; Wu, Z.-Y.; Li, H.-F. Mutation Spectrum and Natural History of ALS Patients in a 15-Year Cohort in Southeastern China. Front. Genet. 2021, 12, 746060. [Google Scholar] [CrossRef]
- Dupuis, L.; Gonzalez de Aguilar, J.-L.; Oudart, H.; de Tapia, M.; Barbeito, L.; Loeffler, J.-P. Mitochondria in Amyotrophic Lateral Sclerosis: A Trigger and a Target. Neurodegener. Dis. 2004, 1, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Otomo, A.; Pan, L.; Hadano, S. Dysregulation of the Autophagy-Endolysosomal System in Amyotrophic Lateral Sclerosis and Related Motor Neuron Diseases. Neurol. Res. Int. 2012, 2012, 498428. [Google Scholar] [CrossRef] [PubMed]
- Devos, D.; Moreau, C.; Lassalle, P.; Perez, T.; De Seze, J.; Brunaud-Danel, V.; Destée, A.; Tonnel, A.B.; Just, N. Low Levels of the Vascular Endothelial Growth Factor in CSF from Early ALS Patients. Neurology 2004, 62, 2127–2129. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Watanabe, T.; Mikami, H.; Nomura, M.; Yamazaki, T.; Irie, T.; Ishikawa, H.; Yasui, K.; Ono, S. Immunohistochemical Studies of Vascular Endothelial Growth Factor in Skin of Patients with Amyotrophic Lateral Sclerosis. J. Neurol. Sci. 2009, 285, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Anand, A.; Thakur, K.; Gupta, P.K. ALS and Oxidative Stress: The Neurovascular Scenario. Oxid. Med. Cell. Longev. 2013, 2013, 635831. [Google Scholar] [CrossRef]
- Mejzini, R.; Flynn, L.L.; Pitout, I.L.; Fletcher, S.; Wilton, S.D.; Akkari, P.A. ALS Genetics, Mechanisms, and Therapeutics: Where Are We Now? Front. Neurosci. 2019, 13, 1310. [Google Scholar] [CrossRef]
- Nagano, S.; Araki, T. Axonal Transport and Local Translation of mRNA in Amyotrophic Lateral Sclerosis. In Amyotrophic Lateral Sclerosis; Araki, T., Ed.; Exon Publications: Brisbane (AU) ISBN 9780645001778.
- Pehar, M.; Vargas, M.R.; Robinson, K.M.; Cassina, P.; Díaz-Amarilla, P.J.; Hagen, T.M.; Radi, R.; Barbeito, L.; Beckman, J.S. Mitochondrial Superoxide Production and Nuclear Factor Erythroid 2-Related Factor 2 Activation in p75 Neurotrophin Receptor-Induced Motor Neuron Apoptosis. J. Neurosci. 2007, 27, 7777–7785. [Google Scholar] [CrossRef]
- Juliani, J.; Vassileff, N.; Spiers, J.G. Inflammatory-Mediated Neuron-Glia Communication Modulates ALS Pathophysiology. J. Neurosci. 2021, 41, 1142–1144. [Google Scholar] [CrossRef]
- Radford, R.A.W.; Vidal-Itriago, A.; Scherer, N.M.; Lee, A.; Graeber, M.; Chung, R.S.; Morsch, M. Evidence for a Growing Involvement of Glia in Amyotrophic Lateral Sclerosis. Spectrums of Amyotrophic Lateral Sclerosis 2021, 123–142. [Google Scholar]
- Laferriere, F.; Polymenidou, M. Advances and Challenges in Understanding the Multifaceted Pathogenesis of Amyotrophic Lateral Sclerosis. Swiss Med. Wkly 2015, 145, w14054. [Google Scholar] [CrossRef] [PubMed]
- Scarrott, J.M.; Herranz-Martín, S.; Alrafiah, A.R.; Shaw, P.J.; Azzouz, M. Current Developments in Gene Therapy for Amyotrophic Lateral Sclerosis. Expert Opin. Biol. Ther. 2015, 15, 935–947. [Google Scholar] [CrossRef] [PubMed]
- Yun, Y.; Ha, Y. CRISPR/Cas9-Mediated Gene Correction to Understand ALS. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Cudkowicz, M.E.; Brown, R.H. , Jr An Update on Superoxide Dismutase 1 in Familial Amyotrophic Lateral Sclerosis. J. Neurol. Sci. 1996, 139 Suppl, 10–15. [Google Scholar] [CrossRef]
- Patterson, D.; Warner, H.R.; Fox, L.M.; Rahmani, Z. Superoxide Dismutase, Oxygen Radical Metabolism, and Amyotrophic Lateral Sclerosis. Mol. Genet. Med. 1994, 4, 79–118. [Google Scholar]
- Andrus, P.K.; Fleck, T.J.; Gurney, M.E.; Hall, E.D. Protein Oxidative Damage in a Transgenic Mouse Model of Familial Amyotrophic Lateral Sclerosis. J. Neurochem. 1998, 71, 2041–2048. [Google Scholar] [CrossRef]
- Culik, R.M.; Sekhar, A.; Nagesh, J.; Deol, H.; Rumfeldt, J.A.O.; Meiering, E.M.; Kay, L.E. Effects of Maturation on the Conformational Free-Energy Landscape of SOD1. Proc. Natl. Acad. Sci. U. S. A. 2018, 115, E2546–E2555. [Google Scholar] [CrossRef]
- Soon, C.P.W.; Crouch, P.J.; Turner, B.J.; McLean, C.A.; Laughton, K.M.; Atkin, J.D.; Masters, C.L.; White, A.R.; Li, Q.-X. Serum Matrix Metalloproteinase-9 Activity Is Dysregulated with Disease Progression in the Mutant SOD1 Transgenic Mice. Neuromuscul. Disord. 2010, 20, 260–266. [Google Scholar] [CrossRef]
- Haidet-Phillips, A.M.; Hester, M.E.; Miranda, C.J.; Meyer, K.; Braun, L.; Frakes, A.; Song, S.; Likhite, S.; Murtha, M.J.; Foust, K.D.; et al. Astrocytes from Familial and Sporadic ALS Patients Are Toxic to Motor Neurons. Nat. Biotechnol. 2011, 29, 824–828. [Google Scholar] [CrossRef]
- Lee, J.; Hyeon, S.J.; Im, H.; Ryu, H.; Kim, Y.; Ryu, H. Astrocytes and Microglia as Non-Cell Autonomous Players in the Pathogenesis of ALS. Exp. Neurobiol. 2016, 25, 233–240. [Google Scholar] [CrossRef]
- Zhao, W.; Beers, D.R.; Appel, S.H. Immune-Mediated Mechanisms in the Pathoprogression of Amyotrophic Lateral Sclerosis. J. Neuroimmune Pharmacol. 2013, 8, 888–899. [Google Scholar] [CrossRef] [PubMed]
- Meissner, F.; Molawi, K.; Zychlinsky, A. Mutant Superoxide Dismutase 1-Induced IL-1beta Accelerates ALS Pathogenesis. Proc. Natl. Acad. Sci. U. S. A. 2010, 107, 13046–13050. [Google Scholar] [CrossRef] [PubMed]
- Luís, J.P.; Simões, C.J.V.; Brito, R.M.M. The Therapeutic Prospects of Targeting IL-1R1 for the Modulation of Neuroinflammation in Central Nervous System Disorders. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.-S.; Peterson, T.C.; et al. Neurotoxic Reactive Astrocytes Are Induced by Activated Microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Genc, B.; Gozutok, O.; Kocak, N.; Ozdinler, P.H. The Timing and Extent of Motor Neuron Vulnerability in ALS Correlates with Accumulation of Misfolded SOD1 Protein in the Cortex and in the Spinal Cord. Cells 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Smeyers, J.; Banchi, E.-G.; Latouche, M. C9ORF72: What It Is, What It Does, and Why It Matters. Front. Cell. Neurosci. 2021, 15, 661447. [Google Scholar] [CrossRef] [PubMed]
- Pang, W.; Hu, F. Cellular and Physiological Functions of C9ORF72 and Implications for ALS/FTD. J. Neurochem. 2021, 157, 334–350. [Google Scholar] [CrossRef] [PubMed]
- O’Rourke, J.G.; Bogdanik, L.; Yáñez, A.; Lall, D.; Wolf, A.J.; Muhammad, A.K.M.G.; Ho, R.; Carmona, S.; Vit, J.P.; Zarrow, J.; et al. C9orf72 Is Required for Proper Macrophage and Microglial Function in Mice. Science 2016, 351, 1324–1329. [Google Scholar] [CrossRef]
- Sullivan, P.M.; Zhou, X.; Robins, A.M.; Paushter, D.H.; Kim, D.; Smolka, M.B.; Hu, F. The ALS/FTLD Associated Protein C9orf72 Associates with SMCR8 and WDR41 to Regulate the Autophagy-Lysosome Pathway. Acta Neuropathol Commun 2016, 4, 51. [Google Scholar] [CrossRef]
- Tang, D.; Sheng, J.; Xu, L.; Zhan, X.; Liu, J.; Jiang, H.; Shu, X.; Liu, X.; Zhang, T.; Jiang, L.; et al. Cryo-EM Structure of C9ORF72-SMCR8-WDR41 Reveals the Role as a GAP for Rab8a and Rab11a. Proc. Natl. Acad. Sci. U. S. A. 2020, 117, 9876–9883. [Google Scholar] [CrossRef]
- DeJesus-Hernandez, M.; Mackenzie, I.R.; Boeve, B.F.; Boxer, A.L.; Baker, M.; Rutherford, N.J.; Nicholson, A.M.; Finch, N.A.; Flynn, H.; Adamson, J.; et al. Expanded GGGGCC Hexanucleotide Repeat in Noncoding Region of C9ORF72 Causes Chromosome 9p-Linked FTD and ALS. Neuron 2011, 72, 245–256. [Google Scholar] [CrossRef]
- Mori, K.; Weng, S.-M.; Arzberger, T.; May, S.; Rentzsch, K.; Kremmer, E.; Schmid, B.; Kretzschmar, H.A.; Cruts, M.; Van Broeckhoven, C.; et al. The C9orf72 GGGGCC Repeat Is Translated into Aggregating Dipeptide-Repeat Proteins in FTLD/ALS. Science 2013, 339, 1335–1338. [Google Scholar] [CrossRef]
- Kato, Y.; Yokogawa, M.; Nakagawa, I.; Onodera, K.; Okano, H.; Inoue, H.; Hattori, M.; Okada, Y.; Tsuiji, H. C9ORF72 Dipeptide Repeat Proteins Disrupt Formation of GEM Bodies and Induce Aberrant Accumulation of Survival of Motor Neuron Protein. bioRxiv 2021.
- Westergard, T.; Jensen, B.K.; Wen, X.; Cai, J.; Kropf, E.; Iacovitti, L.; Pasinelli, P.; Trotti, D. Cell-to-Cell Transmission of Dipeptide Repeat Proteins Linked to C9orf72-ALS/FTD. Cell Rep. 2016, 17, 645–652. [Google Scholar] [CrossRef]
- Lopez-Gonzalez, R.; Lu, Y.; Gendron, T.F.; Karydas, A.; Tran, H.; Yang, D.; Petrucelli, L.; Miller, B.L.; Almeida, S.; Gao, F.B. Poly(GR) in C9ORF72-Related ALS/FTD Compromises Mitochondrial Function and Increases Oxidative Stress and DNA Damage in iPSC-Derived Motor Neurons. Neuron 2016, 92. [Google Scholar] [CrossRef] [PubMed]
- Khosravi, B.; Hartmann, H.; May, S.; Möhl, C.; Ederle, H.; Michaelsen, M.; Schludi, M.H.; Dormann, D.; Edbauer, D. Cytoplasmic Poly-GA Aggregates Impair Nuclear Import of TDP-43 in C9orf72 ALS/FTLD. Human Molecular Genetics 2016, ddw432. [Google Scholar] [CrossRef]
- Prudencio, M.; Humphrey, J.; Pickles, S.; Brown, A.-L.; Hill, S.E.; Kachergus, J.M.; Shi, J.; Heckman, M.G.; Spiegel, M.R.; Cook, C.; et al. Truncated Stathmin-2 Is a Marker of TDP-43 Pathology in Frontotemporal Dementia. J. Clin. Invest. 2020, 130, 6080–6092. [Google Scholar] [CrossRef] [PubMed]
- White, M.A.; Lin, Z.; Kim, E.; Henstridge, C.M.; Pena Altamira, E.; Hunt, C.K.; Burchill, E.; Callaghan, I.; Loreto, A.; Brown-Wright, H.; et al. Sarm1 Deletion Suppresses TDP-43-Linked Motor Neuron Degeneration and Cortical Spine Loss. Acta Neuropathol Commun 2019, 7, 166. [Google Scholar] [CrossRef]
- Andrade, N.S.; Ramic, M.; Esanov, R.; Liu, W.; Rybin, M.J.; Gaidosh, G.; Abdallah, A.; Del’Olio, S.; Huff, T.C.; Chee, N.T.; et al. Dipeptide Repeat Proteins Inhibit Homology-Directed DNA Double Strand Break Repair in C9ORF72 ALS/FTD. Mol. Neurodegener. 2020, 15, 13. [Google Scholar] [CrossRef]
- Ferrara, D.; Pasetto, L.; Bonetto, V.; Basso, M. Role of Extracellular Vesicles in Amyotrophic Lateral Sclerosis. Front. Neurosci. 2018, 12, 574. [Google Scholar] [CrossRef] [PubMed]
- Jo, M.; Lee, S.; Jeon, Y.-M.; Kim, S.; Kwon, Y.; Kim, H.-J. The Role of TDP-43 Propagation in Neurodegenerative Diseases: Integrating Insights from Clinical and Experimental Studies. Exp. Mol. Med. 2020, 52, 1652–1662. [Google Scholar] [CrossRef]
- Hasegawa, M.; Arai, T.; Nonaka, T.; Kametani, F.; Yoshida, M.; Hashizume, Y.; Beach, T.G.; Buratti, E.; Baralle, F.; Morita, M.; et al. Phosphorylated TDP-43 in Frontotemporal Lobar Degeneration and Amyotrophic Lateral Sclerosis. Ann. Neurol. 2008, 64, 60–70. [Google Scholar] [CrossRef]
- Suk, T.R.; Rousseaux, M.W.C. The Role of TDP-43 Mislocalization in Amyotrophic Lateral Sclerosis. Mol. Neurodegener. 2020, 15, 45. [Google Scholar] [CrossRef]
- Trist, B.G.; Fifita, J.A.; Hogan, A.; Grima, N.; Smith, B.; Troakes, C.; Vance, C.; Shaw, C.; Al-Sarraj, S.; Blair, I.P.; et al. Co-Deposition of SOD1, TDP-43 and p62 Proteinopathies in ALS: Evidence for Multifaceted Pathways Underlying Neurodegeneration. Acta Neuropathol Commun 2022, 10, 122. [Google Scholar] [CrossRef]
- Pokrishevsky, E.; Grad, L.I.; Yousefi, M.; Wang, J.; Mackenzie, I.R.; Cashman, N.R. Aberrant Localization of FUS and TDP43 Is Associated with Misfolding of SOD1 in Amyotrophic Lateral Sclerosis. PLoS One 2012, 7, e35050. [Google Scholar] [CrossRef]
- Vance, C.; Scotter, E.L.; Nishimura, A.L.; Troakes, C.; Mitchell, J.C.; Kathe, C.; Urwin, H.; Manser, C.; Miller, C.C.; Hortobágyi, T.; et al. ALS Mutant FUS Disrupts Nuclear Localization and Sequesters Wild-Type FUS within Cytoplasmic Stress Granules. Hum. Mol. Genet. 2013, 22, 2676–2688. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, K.; Clatterbuck-Soper, S.F.; Jackrel, M.E.; Shorter, J.; Mili, S. FUS Inclusions Disrupt RNA Localization by Sequestering Kinesin-1 and Inhibiting Microtubule Detyrosination. J. Cell Biol. 2017, 216, 1015–1034. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, K.R.; Godena, V.K.; Hewitt, V.L.; Whitworth, A.J. Axonal Transport Defects Are a Common Phenotype in Drosophila Models of ALS. Hum. Mol. Genet. 2016, 25. [Google Scholar] [CrossRef]
- Tsai, Y.-L.; Coady, T.H.; Lu, L.; Zheng, D.; Alland, I.; Tian, B.; Shneider, N.A.; Manley, J.L. ALS/FTD-Associated Protein FUS Induces Mitochondrial Dysfunction by Preferentially Sequestering Respiratory Chain Complex mRNAs. Genes Dev. 2020, 34, 785–805. [Google Scholar] [CrossRef] [PubMed]
- Ishigaki, S.; Sobue, G. Importance of Functional Loss of FUS in FTLD/ALS. Front Mol Biosci 2018, 5, 44. [Google Scholar] [CrossRef]
- Sproviero, D.; La Salvia, S.; Giannini, M.; Crippa, V.; Gagliardi, S.; Bernuzzi, S.; Diamanti, L.; Ceroni, M.; Pansarasa, O.; Poletti, A.; et al. Pathological Proteins Are Transported by Extracellular Vesicles of Sporadic Amyotrophic Lateral Sclerosis Patients. Front. Neurosci. 2018, 12, 487. [Google Scholar] [CrossRef] [PubMed]
- Gagliardi, D.; Bresolin, N.; Comi, G.P.; Corti, S. Extracellular Vesicles and Amyotrophic Lateral Sclerosis: From Misfolded Protein Vehicles to Promising Clinical Biomarkers. Cell. Mol. Life Sci. 2021, 78, 561–572. [Google Scholar] [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular Vesicles: Exosomes, Microvesicles, and Friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef]
- Wolf, P. The Nature and Significance of Platelet Products in Human Plasma. Br. J. Haematol. 1967, 13, 269–288. [Google Scholar] [CrossRef]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, Secretion, and Intercellular Interactions of Exosomes and Other Extracellular Vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Akers, J.C.; Gonda, D.; Kim, R.; Carter, B.S.; Chen, C.C. Biogenesis of Extracellular Vesicles (EV): Exosomes, Microvesicles, Retrovirus-like Vesicles, and Apoptotic Bodies. J. Neurooncol. 2013, 113, 1–11. [Google Scholar] [CrossRef] [PubMed]
- D’Acunzo, P.; Pérez-González, R.; Kim, Y.; Hargash, T.; Miller, C.; Alldred, M.J.; Erdjument-Bromage, H.; Penikalapati, S.C.; Pawlik, M.; Saito, M.; et al. Mitovesicles Are a Novel Population of Extracellular Vesicles of Mitochondrial Origin Altered in Down Syndrome. Sci Adv 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological Properties of Extracellular Vesicles and Their Physiological Functions. J Extracell Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Sheng, S.; Wang, Y.; Ding, L.; Xu, X.; Xia, X.; Zheng, J.C. Astrocyte-Derived Extracellular Vesicles: A Double-Edged Sword in Central Nervous System Disorders. Neurosci. Biobehav. Rev. 2021, 125, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Bağcı, C.; Sever-Bahcekapili, M.; Belder, N.; Bennett, A.P.S.; Erdener, Ş.E.; Dalkara, T. Overview of Extracellular Vesicle Characterization Techniques and Introduction to Combined Reflectance and Fluorescence Confocal Microscopy to Distinguish Extracellular Vesicle Subpopulations. Neurophotonics 2022, 9. [Google Scholar] [CrossRef]
- Deng, F.; Miller, J. A Review on Protein Markers of Exosome from Different Bio-Resources and the Antibodies Used for Characterization. J. Histotechnol. 2019, 42, 226–239. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The Biology Function and Biomedical Applications of Exosomes. Science 2020, 367. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, J.; Wysoczynski, M.; Hayek, F.; Janowska-Wieczorek, A.; Ratajczak, M.Z. Membrane-Derived Microvesicles: Important and Underappreciated Mediators of Cell-to-Cell Communication. Leukemia 2006, 20, 1487–1495. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Li, J.; Huang, B.; Liu, J.; Chen, X.; Chen, X.-M.; Xu, Y.-M.; Huang, L.-F.; Wang, X.-Z. Exosomes: Novel Biomarkers for Clinical Diagnosis. ScientificWorldJournal 2015, 2015, 657086. [Google Scholar] [CrossRef] [PubMed]
- Rota, R.; Ciarapica, R.; Giordano, A.; Miele, L.; Locatelli, F. MicroRNAs in Rhabdomyosarcoma: Pathogenetic Implications and Translational Potentiality. Mol. Cancer 2011, 10, 120. [Google Scholar] [CrossRef] [PubMed]
- Camussi, G.; Deregibus, M.-C.; Bruno, S.; Grange, C.; Fonsato, V.; Tetta, C. Exosome/microvesicle-Mediated Epigenetic Reprogramming of Cells. Am. J. Cancer Res. 2011, 1, 98–110. [Google Scholar] [PubMed]
- Sharma, A. Bioinformatic Analysis Revealing Association of Exosomal mRNAs and Proteins in Epigenetic Inheritance. J. Theor. Biol. 2014, 357, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; van Niel, G.; Stahl, P.D. Extracellular Vesicles and Homeostasis-An Emerging Field in Bioscience Research. FASEB Bioadv 2021, 3, 456–458. [Google Scholar] [CrossRef] [PubMed]
- Smalheiser, N.R. Exosomal Transfer of Proteins and RNAs at Synapses in the Nervous System. Biol. Direct 2007, 2, 35. [Google Scholar] [CrossRef]
- Zappulli, V.; Friis, K.P.; Fitzpatrick, Z.; Maguire, C.A.; Breakefield, X.O. Extracellular Vesicles and Intercellular Communication within the Nervous System. J. Clin. Invest. 2016, 126, 1198–1207. [Google Scholar] [CrossRef]
- Lee, Y.; El Andaloussi, S.; Wood, M.J.A. Exosomes and Microvesicles: Extracellular Vesicles for Genetic Information Transfer and Gene Therapy. Hum. Mol. Genet. 2012, 21, R125–R134. [Google Scholar] [CrossRef]
- Cantaluppi, V.; Figliolini, F.; Deregibus, M.C.; Camussi, G. Membrane-Derived Extracellular Vesicles from Endothelial Progenitor Cells Activate Angiogenesis. Tumor Dormancy, Quiescence, and Senescence, Volume 2 2014, 17–25.
- Deregibus, M.C.; Cantaluppi, V.; Calogero, R.; Lo Iacono, M.; Tetta, C.; Biancone, L.; Bruno, S.; Bussolati, B.; Camussi, G. Endothelial Progenitor Cell Derived Microvesicles Activate an Angiogenic Program in Endothelial Cells by a Horizontal Transfer of mRNA. Blood 2007, 110, 2440–2448. [Google Scholar] [CrossRef]
- Cruz, L.; Romero, J.A.A.; Iglesia, R.P.; Lopes, M.H. Extracellular Vesicles: Decoding a New Language for Cellular Communication in Early Embryonic Development. Front Cell Dev Biol 2018, 6, 94. [Google Scholar] [CrossRef] [PubMed]
- Draebing, T.; Heigwer, J.; Juergensen, L.; Katus, H.A.; Hassel, D. Extracellular Vesicle-Delivered Bone Morphogenetic Proteins: A Novel Paracrine Mechanism during Embryonic Development. bioRxiv 2018, 321356. [Google Scholar]
- Xiao, T.; Zhang, W.; Jiao, B.; Pan, C.-Z.; Liu, X.; Shen, L. The Role of Exosomes in the Pathogenesis of Alzheimer’ Disease. Transl. Neurodegener. 2017, 6, 3. [Google Scholar] [CrossRef] [PubMed]
- Upadhya, R.; Zingg, W.; Shetty, S.; Shetty, A.K. Astrocyte-Derived Extracellular Vesicles: Neuroreparative Properties and Role in the Pathogenesis of Neurodegenerative Disorders. J. Control. Release 2020, 323, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Prada, I.; Furlan, R.; Matteoli, M.; Verderio, C. Classical and Unconventional Pathways of Vesicular Release in Microglia. Glia 2013, 61, 1003–1017. [Google Scholar] [CrossRef] [PubMed]
- Cocucci, E.; Racchetti, G.; Rupnik, M.; Meldolesi, J. The Regulated Exocytosis of Enlargeosomes Is Mediated by a SNARE Machinery That Includes VAMP4. J. Cell Sci. 2008, 121, 2983–2991. [Google Scholar] [CrossRef] [PubMed]
- Turola, E.; Furlan, R.; Bianco, F.; Matteoli, M.; Verderio, C. Microglial Microvesicle Secretion and Intercellular Signaling. Front. Physiol. 2012, 3, 149. [Google Scholar] [CrossRef]
- Kakarla, R.; Hur, J.; Kim, Y.J.; Kim, J.; Chwae, Y.-J. Apoptotic Cell-Derived Exosomes: Messages from Dying Cells. Exp. Mol. Med. 2020, 52, 1–6. [Google Scholar] [CrossRef]
- Busatto, S.; Morad, G.; Guo, P.; Moses, M.A. The Role of Extracellular Vesicles in the Physiological and Pathological Regulation of the Blood-Brain Barrier. FASEB Bioadv 2021, 3, 665–675. [Google Scholar] [CrossRef]
- Fauré, J.; Lachenal, G.; Court, M.; Hirrlinger, J.; Chatellard-Causse, C.; Blot, B.; Grange, J.; Schoehn, G.; Goldberg, Y.; Boyer, V.; et al. Exosomes Are Released by Cultured Cortical Neurones. Mol. Cell. Neurosci. 2006, 31, 642–648. [Google Scholar] [CrossRef]
- Potolicchio, I.; Carven, G.J.; Xu, X.; Stipp, C.; Riese, R.J.; Stern, L.J.; Santambrogio, L. Proteomic Analysis of Microglia-Derived Exosomes: Metabolic Role of the Aminopeptidase CD13 in Neuropeptide Catabolism. J. Immunol. 2005, 175, 2237–2243. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.R.; Robinson, M.B.; Gifondorwa, D.J.; Tytell, M.; Milligan, C.E. Regulation of Heat Shock Protein 70 Release in Astrocytes: Role of Signaling Kinases. Dev. Neurobiol. 2007, 67, 1815–1829. [Google Scholar] [CrossRef] [PubMed]
- Krämer-Albers, E.-M.; Bretz, N.; Tenzer, S.; Winterstein, C.; Möbius, W.; Berger, H.; Nave, K.-A.; Schild, H.; Trotter, J. Oligodendrocytes Secrete Exosomes Containing Major Myelin and Stress-Protective Proteins: Trophic Support for Axons? Proteomics Clin. Appl. 2007, 1, 1446–1461. [Google Scholar] [CrossRef]
- Baranyai, T.; Herczeg, K.; Onódi, Z.; Voszka, I.; Módos, K.; Marton, N.; Nagy, G.; Mäger, I.; Wood, M.J.; El Andaloussi, S.; et al. Isolation of Exosomes from Blood Plasma: Qualitative and Quantitative Comparison of Ultracentrifugation and Size Exclusion Chromatography Methods. PLoS One 2015, 10, e0145686. [Google Scholar] [CrossRef]
- Royo, F.; Zuñiga-Garcia, P.; Sanchez-Mosquera, P.; Egia, A.; Perez, A.; Loizaga, A.; Arceo, R.; Lacasa, I.; Rabade, A.; Arrieta, E.; et al. Different EV Enrichment Methods Suitable for Clinical Settings Yield Different Subpopulations of Urinary Extracellular Vesicles from Human Samples. J Extracell Vesicles 2016, 5, 29497. [Google Scholar] [CrossRef]
- Zonneveld, M.I.; Brisson, A.R.; van Herwijnen, M.J.C.; Tan, S.; van de Lest, C.H.A.; Redegeld, F.A.; Garssen, J.; Wauben, M.H.M.; Nolte-’t Hoen, E.N.M. Recovery of Extracellular Vesicles from Human Breast Milk Is Influenced by Sample Collection and Vesicle Isolation Procedures. J Extracell Vesicles 2014, 3. [Google Scholar] [CrossRef] [PubMed]
- Street, J.M.; Barran, P.E.; Mackay, C.L.; Weidt, S.; Balmforth, C.; Walsh, T.S.; Chalmers, R.T.A.; Webb, D.J.; Dear, J.W. Identification and Proteomic Profiling of Exosomes in Human Cerebrospinal Fluid. J. Transl. Med. 2012, 10, 5. [Google Scholar] [CrossRef]
- Morad, G.; Moses, M.A. Brainwashed by Extracellular Vesicles: The Role of Extracellular Vesicles in Primary and Metastatic Brain Tumour Microenvironment. Journal of Extracellular Vesicles 2019, 8, 1627164. [Google Scholar] [CrossRef]
- Brites, D.; Vaz, A.R. Microglia Centered Pathogenesis in ALS: Insights in Cell Interconnectivity. Front. Cell. Neurosci. 2014, 8, 117. [Google Scholar] [CrossRef]
- Brites, D.; Fernandes, A. Neuroinflammation and Depression: Microglia Activation, Extracellular Microvesicles and microRNA Dysregulation. Front. Cell. Neurosci. 2015, 9, 476. [Google Scholar] [CrossRef]
- Gupta, A.; Pulliam, L. Exosomes as Mediators of Neuroinflammation. J. Neuroinflammation 2014, 11, 68. [Google Scholar] [CrossRef]
- Schneider, A.; Simons, M. Exosomes: Vesicular Carriers for Intercellular Communication in Neurodegenerative Disorders. Cell Tissue Res. 2013, 352, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, T.; Masuda-Suzukake, M.; Arai, T.; Hasegawa, Y.; Akatsu, H.; Obi, T.; Yoshida, M.; Murayama, S.; Mann, D.M.A.; Akiyama, H.; et al. Prion-like Properties of Pathological TDP-43 Aggregates from Diseased Brains. Cell Rep. 2013, 4, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Grad, L.I.; Yerbury, J.J.; Turner, B.J.; Guest, W.C.; Pokrishevsky, E.; O’Neill, M.A.; Yanai, A.; Silverman, J.M.; Zeineddine, R.; Corcoran, L.; et al. Intercellular Propagated Misfolding of Wild-Type Cu/Zn Superoxide Dismutase Occurs via Exosome-Dependent and -Independent Mechanisms. Proc. Natl. Acad. Sci. U. S. A. 2014, 111, 3620–3625. [Google Scholar] [CrossRef] [PubMed]
- Cicardi, M.E.; Marrone, L.; Azzouz, M.; Trotti, D. Proteostatic Imbalance and Protein Spreading in Amyotrophic Lateral Sclerosis. EMBO J. 2021, 40, e106389. [Google Scholar] [CrossRef] [PubMed]
- Salvany, S.; Casanovas, A.; Piedrafita, L.; Gras, S.; Calderó, J.; Esquerda, J.E. Accumulation of Misfolded SOD1 Outlines Distinct Patterns of Motor Neuron Pathology and Death during Disease Progression in a SOD1 Mouse Model of Amyotrophic Lateral Sclerosis. Brain Pathol. 2022, 32, e13078. [Google Scholar] [CrossRef]
- Chen, P.-C.; Wu, D.; Hu, C.-J.; Chen, H.-Y.; Hsieh, Y.-C.; Huang, C.-C. Exosomal TAR DNA-Binding Protein-43 and Neurofilaments in Plasma of Amyotrophic Lateral Sclerosis Patients: A Longitudinal Follow-up Study. J. Neurol. Sci. 2020, 418, 117070. [Google Scholar] [CrossRef]
- Ding, X.; Ma, M.; Teng, J.; Teng, R.K.F.; Zhou, S.; Yin, J.; Fonkem, E.; Huang, J.H.; Wu, E.; Wang, X. Exposure to ALS-FTD-CSF Generates TDP-43 Aggregates in Glioblastoma Cells through Exosomes and TNTs-like Structure. Oncotarget 2015, 6, 24178–24191. [Google Scholar] [CrossRef] [PubMed]
- Pinto, S.; Cunha, C.; Barbosa, M.; Vaz, A.R.; Brites, D. Exosomes from NSC-34 Cells Transfected with hSOD1-G93A Are Enriched in miR-124 and Drive Alterations in Microglia Phenotype. Front. Neurosci. 2017, 11, 273. [Google Scholar] [CrossRef]
- Basso, M.; Pozzi, S.; Tortarolo, M.; Fiordaliso, F.; Bisighini, C.; Pasetto, L.; Spaltro, G.; Lidonnici, D.; Gensano, F.; Battaglia, E.; et al. Mutant Copper-Zinc Superoxide Dismutase (SOD1) Induces Protein Secretion Pathway Alterations and Exosome Release in Astrocytes: Implications for Disease Spreading and Motor Neuron Pathology in Amyotrophic Lateral Sclerosis. J. Biol. Chem. 2013, 288, 15699–15711. [Google Scholar] [CrossRef]
- Chen, Y.; Xia, K.; Chen, L.; Fan, D. Increased Interleukin-6 Levels in the Astrocyte-Derived Exosomes of Sporadic Amyotrophic Lateral Sclerosis Patients. Front. Neurosci. 2019, 13, 574. [Google Scholar] [CrossRef] [PubMed]
- Le Gall, L.; Duddy, W.J.; Martinat, C.; Mariot, V.; Connolly, O.; Milla, V.; Anakor, E.; Ouandaogo, Z.G.; Millecamps, S.; Lainé, J.; et al. Muscle Cells of Sporadic Amyotrophic Lateral Sclerosis Patients Secrete Neurotoxic Vesicles. J. Cachexia Sarcopenia Muscle 2022, 13, 1385–1402. [Google Scholar] [CrossRef] [PubMed]
- Anakor, E.; Milla, V.; Connolly, O.; Martinat, C.; Pradat, P.F.; Dumonceaux, J.; Duddy, W.; Duguez, S. The Neurotoxicity of Vesicles Secreted by ALS Patient Myotubes Is Specific to Exosome-like and Not Larger Subtypes. Cells 2022, 11, 845. [Google Scholar] [CrossRef] [PubMed]
- Laneve, P.; Tollis, P.; Caffarelli, E. RNA Deregulation in Amyotrophic Lateral Sclerosis: The Noncoding Perspective. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Barbo, M.; Ravnik-Glavač, M. Extracellular Vesicles as Potential Biomarkers in Amyotrophic Lateral Sclerosis. Genes 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; He, H.; Shen, X.; Zhao, J.; Cao, X.; Han, S.; Cui, C.; Chen, Y.; Wei, Y.; Xia, L.; et al. miR-9-5p Inhibits Skeletal Muscle Satellite Cell Proliferation and Differentiation by Targeting IGF2BP3 through the IGF2-PI3K/Akt Signaling Pathway. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Shu, Z.; Gao, F.; Xia, Q.; Zhang, M. MiR-9-5p Promotes Cell Proliferation and Migration of Hepatocellular Carcinoma by Targeting CPEB3. Biomark. Med. 2021, 15, 97–108. [Google Scholar]
- Wang, M.; Gao, Q.; Chen, Y.; Li, Z.; Yue, L.; Cao, Y. PAK4, a Target of miR-9-5p, Promotes Cell Proliferation and Inhibits Apoptosis in Colorectal Cancer. Cell. Mol. Biol. Lett. 2019, 24, 58. [Google Scholar] [CrossRef]
- Saucier, D.; Wajnberg, G.; Roy, J.; Beauregard, A.-P.; Chacko, S.; Crapoulet, N.; Fournier, S.; Ghosh, A.; Lewis, S.M.; Marrero, A.; et al. Identification of a Circulating miRNA Signature in Extracellular Vesicles Collected from Amyotrophic Lateral Sclerosis Patients. Brain Res. 2019, 1708, 100–108. [Google Scholar] [CrossRef]
- Singh, R.; Ha, S.E.; Wei, L.; Jin, B.; Zogg, H.; Poudrier, S.M.; Jorgensen, B.G.; Park, C.; Ronkon, C.F.; Bartlett, A.; et al. miR-10b-5p Rescues Diabetes and Gastrointestinal Dysmotility. Gastroenterology 2021, 160, 1662–1678. [Google Scholar] [CrossRef]
- Ge, G.; Yang, D.; Tan, Y.; Chen, Y.; Jiang, D.; Jiang, A.; Li, Q.; Liu, Y.; Zhong, Z.; Li, X.; et al. miR-10b-5p Regulates C2C12 Myoblasts Proliferation and Differentiation. Biosci. Biotechnol. Biochem. 2019, 83, 291–299. [Google Scholar] [CrossRef]
- Faur, C.I.; Roman, R.C.; Jurj, A.; Raduly, L.; Almășan, O.; Rotaru, H.; Chirilă, M.; Moldovan, M.A.; Hedeșiu, M.; Dinu, C. Salivary Exosomal MicroRNA-486-5p and MicroRNA-10b-5p in Oral and Oropharyngeal Squamous Cell Carcinoma. Medicina 2022, 58. [Google Scholar] [CrossRef]
- Yelick, J.; Men, Y.; Jin, S.; Seo, S.; Espejo-Porras, F.; Yang, Y. Elevated Exosomal Secretion of miR-124-3p from Spinal Neurons Positively Associates with Disease Severity in ALS. Exp. Neurol. 2020, 333, 113414. [Google Scholar] [CrossRef] [PubMed]
- Banack, S.A.; Dunlop, R.A.; Stommel, E.W.; Mehta, P.; Cox, P.A. miRNA Extracted from Extracellular Vesicles Is a Robust Biomarker of Amyotrophic Lateral Sclerosis. J. Neurol. Sci. 2022, 442, 120396. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yang, Q.; Li, J.; Chen, W.; Jin, X.; Wang, Y. MicroRNA-15a-5p Inhibits Endometrial Carcinoma Proliferation, Invasion and Migration via Downregulation of VEGFA and Inhibition of the Wnt/β-Catenin Signaling Pathway. Oncol. Lett. 2021, 21, 310. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Zhang, H.; Wang, X.; Zhou, L.; Li, H.; Deng, T.; Qu, Y.; Duan, J.; Bai, M.; Ge, S.; et al. The miR-24-Bim Pathway Promotes Tumor Growth and Angiogenesis in Pancreatic Carcinoma. Oncotarget 2015, 6, 43831–43842. [Google Scholar] [CrossRef] [PubMed]
- Katsu, M.; Hama, Y.; Utsumi, J.; Takashina, K.; Yasumatsu, H.; Mori, F.; Wakabayashi, K.; Shoji, M.; Sasaki, H. MicroRNA Expression Profiles of Neuron-Derived Extracellular Vesicles in Plasma from Patients with Amyotrophic Lateral Sclerosis. Neurosci. Lett. 2019, 708, 134176. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Hu, C.; Li, J.; Liu, L.; Jing, W.; Tang, W.; Tian, W.; Long, J. Effect of miR-26a-5p on the Wnt/Ca(2+) Pathway and Osteogenic Differentiation of Mouse Adipose-Derived Mesenchymal Stem Cells. Calcif. Tissue Int. 2016, 99, 174–186. [Google Scholar] [CrossRef]
- Lo, T.-W.; Figueroa-Romero, C.; Hur, J.; Pacut, C.; Stoll, E.; Spring, C.; Lewis, R.; Nair, A.; Goutman, S.A.; Sakowski, S.A.; et al. Extracellular Vesicles in Serum and Central Nervous System Tissues Contain microRNA Signatures in Sporadic Amyotrophic Lateral Sclerosis. Front. Mol. Neurosci. 2021, 14, 739016. [Google Scholar] [CrossRef]
- Zhou, L.; Liang, X.; Zhang, L.; Yang, L.; Nagao, N.; Wu, H.; Liu, C.; Lin, S.; Cai, G.; Liu, J. MiR-27a-3p Functions as an Oncogene in Gastric Cancer by Targeting BTG2. Oncotarget 2016, 7, 51943–51954. [Google Scholar] [CrossRef]
- Xi, T.; Jin, F.; Zhu, Y.; Wang, J.; Tang, L.; Wang, Y.; Liebeskind, D.S.; Scalzo, F.; He, Z. miR-27a-3p Protects against Blood-Brain Barrier Disruption and Brain Injury after Intracerebral Hemorrhage by Targeting Endothelial Aquaporin-11. J. Biol. Chem. 2018, 293, 20041–20050. [Google Scholar] [CrossRef]
- Xu, Q.; Zhao, Y.; Zhou, X.; Luan, J.; Cui, Y.; Han, J. Comparison of the Extraction and Determination of Serum Exosome and miRNA in Serum and the Detection of miR-27a-3p in Serum Exosome of ALS Patients. Intractable Rare Dis Res 2018, 7, 13–18. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, M.; Ye, L.; Chen, C.; She, J.; Song, Y. MiR-29b-3p Promotes Particulate Matter-Induced Inflammatory Responses by Regulating the C1QTNF6/AMPK Pathway. Aging 2020, 12, 1141–1158. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Xiong, F.; Zhang, S.; Guo, W.; He, Y. Crucial Roles of miR-625 in Human Cancer. Front. Med. 2022, 9, 845094. [Google Scholar] [CrossRef] [PubMed]
- Slabáková, E.; Culig, Z.; Remšík, J.; Souček, K. Alternative Mechanisms of miR-34a Regulation in Cancer. Cell Death Dis. 2017, 8, e3100. [Google Scholar] [CrossRef] [PubMed]
- Rizzuti, M.; Melzi, V.; Gagliardi, D.; Resnati, D.; Meneri, M.; Dioni, L.; Masrori, P.; Hersmus, N.; Poesen, K.; Locatelli, M.; et al. Insights into the Identification of a Molecular Signature for Amyotrophic Lateral Sclerosis Exploiting Integrated microRNA Profiling of iPSC-Derived Motor Neurons and Exosomes. Cell. Mol. Life Sci. 2022, 79, 189. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Zhu, T.; Liu, H.; Zhu, Y.; Peng, Y.; Tang, T.; Zhou, S.; Hu, C.; Chen, H.; Guo, A.; et al. The miR-100-5p Targets SMARCA5 to Regulate the Apoptosis and Intracellular Survival of BCG in Infected THP-1 Cells. Cells 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Zhang, Y.; Huang, S.; Yan, M.; Li, J.; Jin, T.; Bao, S. MiR-100-5p, miR-199a-3p and miR-199b-5p Induce Autophagic Death of Endometrial Carcinoma Cell through Targeting mTOR. Int. J. Clin. Exp. Pathol. 2017, 10, 9262–9272. [Google Scholar] [PubMed]
- Zeng, J.; Wang, L.; Zhao, J.; Zheng, Z.; Peng, J.; Zhang, W.; Wen, T.; Nie, J.; Ding, L.; Yi, D. MiR-100-5p Regulates Cardiac Hypertrophy through Activation of Autophagy by Targeting mTOR. Hum. Cell 2021, 34, 1388–1397. [Google Scholar] [CrossRef]
- Dubes, S.; Soula, A.; Benquet, S.; Tessier, B.; Poujol, C.; Favereaux, A.; Thoumine, O.; Letellier, M. miR-124-Dependent Tagging of Synapses by Synaptopodin Enables Input-Specific Homeostatic Plasticity. EMBO J. 2022, 41, e109012. [Google Scholar] [CrossRef]
- Zhang, Z.-B.; Xiong, L.-L.; Xue, L.-L.; Deng, Y.-P.; Du, R.-L.; Hu, Q.; Xu, Y.; Yang, S.-J.; Wang, T.-H. MiR-127-3p Targeting CISD1 Regulates Autophagy in Hypoxic-Ischemic Cortex. Cell Death Dis. 2021, 12, 279. [Google Scholar] [CrossRef]
- Shen, L.; Li, Q.; Wang, J.; Zhao, Y.; Niu, L.; Bai, L.; Shuai, S.; Li, X.; Zhang, S.; Zhu, L. miR-144-3p Promotes Adipogenesis Through Releasing C/EBPα From Klf3 and CtBP2. Front. Genet. 2018, 9, 677. [Google Scholar]
- Pan, J.; Du, M.; Cao, Z.; Zhang, C.; Hao, Y.; Zhu, J.; He, H. miR-146a-5p Attenuates IL-1β-Induced IL-6 and IL-1β Expression in a Cementoblast-Derived Cell Line. Oral Dis. 2020. [CrossRef] [PubMed]
- Banack, S.A.; Dunlop, R.A.; Cox, P.A. An miRNA Fingerprint Using Neural-Enriched Extracellular Vesicles from Blood Plasma: Towards a Biomarker for Amyotrophic Lateral Sclerosis/motor Neuron Disease. Open Biol. 2020, 10, 200116. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Zhu, H.; Lei, Q.; Chen, L.; Yang, D.; Sui, W. MicroRNA-149-3p Inhibits Cell Proliferation by Targeting AKT2 in Oral Squamous Cell Carcinoma. Mol. Med. Rep. 2021, 23. [Google Scholar] [CrossRef]
- Luo, H.; Ye, G.; Liu, Y.; Huang, D.; Luo, Q.; Chen, W.; Qi, Z. miR-150-3p Enhances Neuroprotective Effects of Neural Stem Cell Exosomes after Hypoxic-Ischemic Brain Injury by Targeting CASP2. Neurosci. Lett. 2022, 779, 136635. [Google Scholar]
- Wang, H.; Shu, J.; Zhang, C.; Wang, Y.; Shi, R.; Yang, F.; Tang, X. Extracellular Vesicle-Mediated miR-150-3p Delivery in Joint Homeostasis: A Potential Treatment for Osteoarthritis? Cells 2022, 11. [Google Scholar] [CrossRef]
- Zhang, Y.; Hao, T.; Zhang, H.; Wei, P.; Li, X. [Over-expression of miR-151a-3p inhibits proliferation and migration of PC-3 prostate cancer cells]. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi 2018, 34, 247–252. [Google Scholar] [PubMed]
- Zhou, R.; Wang, R.; Qin, Y.; Ji, J.; Xu, M.; Wu, W.; Chen, M.; Wu, D.; Song, L.; Shen, H.; et al. Mitochondria-Related miR-151a-5p Reduces Cellular ATP Production by Targeting CYTB in Asthenozoospermia. Sci. Rep. 2015, 5, 17743. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Qiu, X.; Wang, D.; Li, Y.; Zhang, B.; Yuan, T.; Wei, J.; Zhao, B.; Zhao, X.; Lou, J.; et al. MiR-181a-5p Inhibits Cell Proliferation and Migration by Targeting Kras in Non-Small Cell Lung Cancer A549 Cells. Acta Biochim. Biophys. Sin. 2015, 47, 630–638. [Google Scholar]
- Cao, Y.; Zhao, D.; Li, P.; Wang, L.; Qiao, B.; Qin, X.; Li, L.; Wang, Y. MicroRNA-181a-5p Impedes IL-17-Induced Nonsmall Cell Lung Cancer Proliferation and Migration through Targeting VCAM-1. Cell. Physiol. Biochem. 2017, 42, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, M.; Liu, G.; Xiong, C.; Rao, J. microRNA-181a-5p Impedes the Proliferation, Migration, and Invasion of Retinoblastoma Cells by Targeting the NRAS Proto-Oncogene. Clinics 2022, 77, 100026. [Google Scholar] [CrossRef] [PubMed]
- Assmann, J.L.J.C.; Leon, L.G.; Stavast, C.J.; van den Bogaerdt, S.E.; Schilperoord-Vermeulen, J.; Sandberg, Y.; Bellido, M.; Erkeland, S.J.; Feith, D.J.; Loughran, T.P., Jr; et al. miR-181a Is a Novel Player in the STAT3-Mediated Survival Network of TCRαβ+ CD8+ T Large Granular Lymphocyte Leukemia. Leukemia 2022, 36, 983–993. [Google Scholar] [CrossRef]
- Li, X.; Han, J.; Zhu, H.; Peng, L.; Chen, Z. miR-181b-5p Mediates TGF-β1-Induced Epithelial-to-Mesenchymal Transition in Non-Small Cell Lung Cancer Stem-like Cells Derived from Lung Adenocarcinoma A549 Cells. Int. J. Oncol. 2017, 51, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Guo, Z.; Gao, W. miR-181b-5p Promotes Proliferation and Inhibits Apoptosis of Hypertrophic Scar Fibroblasts through Regulating the MEK/ERK/p21 Pathway. Exp. Ther. Med. 2019, 17, 1537–1544. [Google Scholar] [CrossRef] [PubMed]
- Yeon, M.; Kim, Y.; Pathak, D.; Kwon, E.; Kim, D.Y.; Jeong, M.S.; Jung, H.S.; Jeoung, D. The CAGE-MiR-181b-5p-S1PR1 Axis Regulates Anticancer Drug Resistance and Autophagy in Gastric Cancer Cells. Front Cell Dev Biol 2021, 9, 666387. [Google Scholar] [CrossRef]
- Graham, A.; Holbert, J.; Nothnick, W.B. miR-181b-5p Modulates Cell Migratory Proteins, Tissue Inhibitor of Metalloproteinase 3, and Annexin A2 During In Vitro Decidualization in a Human Endometrial Stromal Cell Line. Reprod. Sci. 2017, 24, 1264–1274. [Google Scholar] [CrossRef]
- Zhang, X.; Yu, J.; Zhao, C.; Ren, H.; Yuan, Z.; Zhang, B.; Zhuang, J.; Wang, J.; Feng, B. MiR-181b-5p Modulates Chemosensitivity of Glioma Cells to Temozolomide by Targeting Bcl-2. Biomed. Pharmacother. 2019, 109, 2192–2202. [Google Scholar] [CrossRef]
- Li, C.; Chen, Y.; Chen, X.; Wei, Q.; Ou, R.; Gu, X.; Cao, B.; Shang, H. MicroRNA-183-5p Is Stress-Inducible and Protects Neurons against Cell Death in Amyotrophic Lateral Sclerosis. J. Cell. Mol. Med. 2020, 24, 8614–8622. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.; Sheng, L.; Qu, W.; Xue, X.; Chen, H.; Zheng, G.; Chen, W. MiR-194-5p Enhances the Sensitivity of Nonsmall-Cell Lung Cancer to Doxorubicin through Targeted Inhibition of Hypoxia-Inducible Factor-1. World J. Surg. Oncol. 2021, 19, 174. [Google Scholar] [CrossRef]
- Shen, X.; Gong, A. The Expression of microRNA-197-3p Regulates the Proliferation of Ovarian Granulosa Cells through CUL3 in Polycystic Ovarian Syndrome. Acta Biochim. Pol. 2022, 69, 599–604. [Google Scholar]
- Jiang, J.; Tang, Q.; Gong, J.; Jiang, W.; Chen, Y.; Zhou, Q.; Aldeen, A.; Wang, S.; Li, C.; Lv, W.; et al. Radiosensitizer EXO-miR-197-3p Inhibits Nasopharyngeal Carcinoma Progression and Radioresistance by Regulating the AKT/mTOR Axis and HSPA5-Mediated Autophagy. Int. J. Biol. Sci. 2022, 18, 1878–1895. [Google Scholar] [CrossRef]
- Chang, R.-M.; Fu, Y.; Zeng, J.; Zhu, X.-Y.; Gao, Y. Cancer-Derived Exosomal miR-197-3p Confers Angiogenesis via Targeting TIMP2/3 in Lung Adenocarcinoma Metastasis. Cell Death Dis. 2022, 13, 1032. [Google Scholar] [CrossRef]
- Liu, W.; Zheng, J.; Dong, J.; Bai, R.; Song, D.; Ma, X.; Zhao, L.; Yao, Y.; Zhang, H.; Liu, T. Association of miR-197-5p, a Circulating Biomarker for Heart Failure, with Myocardial Fibrosis and Adverse Cardiovascular Events among Patients with Stage C or D Heart Failure. Cardiology 2018, 141, 212–225. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Dasgupta, D.; Ghosh, A.; Roychoudhury, S.; Kumar, D.; Gorain, M.; Butti, R.; Datta, S.; Agarwal, S.; Gupta, S.; et al. MiRNA199a-3p Suppresses Tumor Growth, Migration, Invasion and Angiogenesis in Hepatocellular Carcinoma by Targeting VEGFA, VEGFR1, VEGFR2, HGF and MMP2. Cell Death Dis. 2017, 8, e2706. [Google Scholar] [CrossRef]
- Ren, K.; Li, T.; Zhang, W.; Ren, J.; Li, Z.; Wu, G. miR-199a-3p Inhibits Cell Proliferation and Induces Apoptosis by Targeting YAP1, Suppressing Jagged1-Notch Signaling in Human Hepatocellular Carcinoma. J. Biomed. Sci. 2016, 23, 79. [Google Scholar] [CrossRef]
- Liu, J.; Quan, Z.; Gao, Y.; Wu, X.; Zheng, Y. MicroRNA-199b-3p Suppresses Malignant Proliferation by Targeting Phospholipase Cε and Correlated with Poor Prognosis in Prostate Cancer. Biochem. Biophys. Res. Commun. 2021, 576, 73–79. [Google Scholar] [CrossRef]
- Huang, G.-H.; Shan, H.; Li, D.; Zhou, B.; Pang, P.-F. MiR-199a-5p Suppresses Tumorigenesis by Targeting Clathrin Heavy Chain in Hepatocellular Carcinoma. Cell Biochem. Funct. 2017, 35, 98–104. [Google Scholar]
- Zhang, R.; Li, Y.-Z.; Zhao, K.-X. MiR-298 Suppresses the Malignant Progression of Osteosarcoma by Targeting JMJD6. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 2250–2258. [Google Scholar]
- Ye, L.; Wang, F.; Wu, H.; Yang, H.; Yang, Y.; Ma, Y.; Xue, A.; Zhu, J.; Chen, M.; Wang, J.; et al. Functions and Targets of miR-335 in Cancer. Onco. Targets. Ther. 2021, 14, 3335–3349. [Google Scholar] [CrossRef]
- Jiang, H.; Moro, A.; Wang, J.; Meng, D.; Zhan, X.; Wei, Q. MicroRNA-338-3p as a Novel Therapeutic Target for Intervertebral Disc Degeneration. Exp. Mol. Med. 2021, 53, 1356–1365. [Google Scholar] [CrossRef]
- Zhang, C.; Li, H.; Wang, J.; Zhang, J.; Hou, X. MicroRNA-338-3p Suppresses Cell Proliferation, Migration and Invasion in Human Malignant Melanoma by Targeting MACC1. Exp. Ther. Med. 2019, 18, 997–1004. [Google Scholar]
- Fu, Y.; Hu, X.; Zheng, C.; Sun, G.; Xu, J.; Luo, S.; Cao, P. Intrahippocampal miR-342-3p Inhibition Reduces β-Amyloid Plaques and Ameliorates Learning and Memory in Alzheimer’s Disease. Metab. Brain Dis. 2019, 34, 1355–1363. [Google Scholar] [CrossRef]
- Montag, J.; Hitt, R.; Opitz, L.; Schulz-Schaeffer, W.J.; Hunsmann, G.; Motzkus, D. Upregulation of miRNA Hsa-miR-342-3p in Experimental and Idiopathic Prion Disease. Mol. Neurodegener. 2009, 4, 36. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.Y.; Calin, G.A.; Yuen, K.S.; Jin, D.Y.; Chim, C.S. Epigenetic Silencing of miR-342-3p in B Cell Lymphoma and Its Impact on Autophagy. Clin. Epigenetics 2020, 12, 150. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Luo, R.; Yang, W.; Zhou, Z.; Li, C. miR-363-3p Is Activated by MYB and Regulates Osteoporosis Pathogenesis via PTEN/PI3K/AKT Signaling Pathway. In Vitro Cell. Dev. Biol. Anim. 2019, 55, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Qiu, X.; Jin, P.; Li, Z.; Zhang, Y.; Lv, L.; Song, F. MiR-371a-5p Positively Associates with Hepatocellular Carcinoma Malignancy but Sensitizes Cancer Cells to Oxaliplatin by Suppressing BECN1-Dependent Autophagy. Life 2022, 12. [Google Scholar] [CrossRef]
- Muys, B.R.; Sousa, J.F.; Plaça, J.R.; de Araújo, L.F.; Sarshad, A.A.; Anastasakis, D.G.; Wang, X.; Li, X.L.; de Molfetta, G.A.; Ramão, A.; et al. miR-450a Acts as a Tumor Suppressor in Ovarian Cancer by Regulating Energy Metabolism. Cancer Res. 2019, 79, 3294–3305. [Google Scholar] [CrossRef]
- Varcianna, A.; Myszczynska, M.A.; Castelli, L.M.; O’Neill, B.; Kim, Y.; Talbot, J.; Nyberg, S.; Nyamali, I.; Heath, P.R.; Stopford, M.J.; et al. Micro-RNAs Secreted through Astrocyte-Derived Extracellular Vesicles Cause Neuronal Network Degeneration in C9orf72 ALS. EBioMedicine 2019, 40, 626–635. [Google Scholar] [CrossRef]
- Peng, X.; Wu, M.; Liu, W.; Guo, C.; Zhan, L.; Zhan, X. miR-502-5p Inhibits the Proliferation, Migration and Invasion of Gastric Cancer Cells by Targeting SP1. Oncol. Lett. 2020, 20, 2757–2762. [Google Scholar]
- Cao, B.; Tan, S.; Tang, H.; Chen, Y.; Shu, P. miR-512-5p Suppresses Proliferation, Migration and Invasion, and Induces Apoptosis in Non-small Cell Lung Cancer Cells by Targeting ETS1. Mol. Med. Rep. 2019, 19, 3604–3614. [Google Scholar] [CrossRef]
- Chen, J.-Q.; Huang, Z.-P.; Li, H.-F.; Ou, Y.-L.; Huo, F.; Hu, L.-K. MicroRNA-520f-3p Inhibits Proliferation of Gastric Cancer Cells via Targeting SOX9 and Thereby Inactivating Wnt Signaling. Sci. Rep. 2020, 10, 6197. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, Q.; Dai, Y.; Jiang, T.; Zhou, Y. miR-532-3p Inhibits Proliferation and Promotes Apoptosis of Lymphoma Cells by Targeting β-Catenin. J. Cancer 2020, 11, 4762–4770. [Google Scholar] [CrossRef] [PubMed]
- Ji, T.; Gao, L.; Yu, Z. Tumor-Suppressive microRNA-551b-3p Targets H6PD to Inhibit Gallbladder Cancer Progression. Cancer Gene Ther. 2021, 28, 693–705. [Google Scholar] [CrossRef] [PubMed]
- Xuan, Z.; Chen, C.; Tang, W.; Ye, S.; Zheng, J.; Zhao, Y.; Shi, Z.; Zhang, L.; Sun, H.; Shao, C. TKI-Resistant Renal Cancer Secretes Low-Level Exosomal miR-549a to Induce Vascular Permeability and Angiogenesis to Promote Tumor Metastasis. Front Cell Dev Biol 2021, 9, 689947. [Google Scholar]
- Chen, M.; Wang, D.; Liu, J.; Zhou, Z.; Ding, Z.; Liu, L.; Su, D.; Li, H. Functions as a Tumor Suppressor in Hepatocellular Carcinoma by Targeting Ribosomal Protein SA. Biomed Res. Int. 2020, 2020, 3280530. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Ma, R.; Wang, Q.; Zhang, P.; Li, D.; Wang, Q.; Wang, J.; Li, H.; Liu, H.; Wang, Z. MiR-625-3p Promotes Cell Migration and Invasion via Inhibition of SCAI in Colorectal Carcinoma Cells. Oncotarget 2015, 6, 27805–27815. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhao, S.; Wang, J.; Zhu, Z.; Luo, L.; Xiang, Q.; Zhou, M.; Ma, Y.; Wang, Z.; Zhao, Z. MiR-629-5p Promotes Prostate Cancer Development and Metastasis by Targeting AKAP13. Front. Oncol. 2021, 11, 754353. [Google Scholar]
- Cong, J.; Liu, R.; Wang, X.; Jiang, H.; Zhang, Y. MiR-634 Decreases Cell Proliferation and Induces Apoptosis by Targeting mTOR Signaling Pathway in Cervical Cancer Cells. Artif. Cells Nanomed. Biotechnol. 2015. [CrossRef]
- Chen, D.; Wu, X.; Zhao, J.; Zhao, X. MicroRNA-634 Functions as a Tumor Suppressor in Pancreatic Cancer via Directly Targeting Heat Shock-Related 70-kDa Protein 2. Exp. Ther. Med. 2019, 17, 3949–3956. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Wu, M.; Wang, H.; Wu, L.; Xu, B.; Zhou, W.; Fan, X.; Shao, J.; Yang, T. MicroRNA-664a-5p Promotes Osteogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stem Cells by Directly Downregulating HMGA2. Biochem. Biophys. Res. Commun. 2020, 521, 9–14. [Google Scholar] [CrossRef]
- Watanabe, K.; Yamaji, R.; Ohtsuki, T. MicroRNA-664a-5p Promotes Neuronal Differentiation of SH-SY5Y Cells. Genes Cells 2018, 23, 225–233. [Google Scholar] [CrossRef]
- Hayakawa, K.; Kawasaki, M.; Hirai, T.; Yoshida, Y.; Tsushima, H.; Fujishiro, M.; Ikeda, K.; Morimoto, S.; Takamori, K.; Sekigawa, I. MicroRNA-766-3p Contributes to Anti-Inflammatory Responses through the Indirect Inhibition of NF-κB Signaling. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef]
- Gao, J.; Fei, L.; Wu, X.; Li, H. MiR-766-3p Suppresses Malignant Behaviors and Stimulates Apoptosis of Colon Cancer Cells via Targeting TGFBI. Can J Gastroenterol Hepatol 2022, 2022, 7234704. [Google Scholar] [CrossRef]
- Yang, B.; Diao, H.; Wang, P.; Guan, F.; Liu, H. microRNA-877-5p Exerts Tumor-Suppressive Functions in Prostate Cancer through Repressing Transcription of Forkhead Box M1. Bioengineered 2021, 12, 9094–9102. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Chen, G.; Gao, H.; Li, Y.; Zhuang, L.; Meng, Z.; Liu, L. miR-939-5p Contributes to the Migration and Invasion of Pancreatic Cancer by Targeting ARHGAP4. Onco. Targets. Ther. 2020, 13, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Bertolazzi, G.; Cipollina, C.; Benos, P.V.; Tumminello, M.; Coronnello, C. miR-1207-5p Can Contribute to Dysregulation of Inflammatory Response in COVID-19 Targeting SARS-CoV-2 RNA. Front. Cell. Infect. Microbiol. 2020, 10, 586592. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Xie, D.; Gao, S.; Bai, C.-J.; Zhu, M.-X.; Guan, H.; Zhou, P.-K. A Biomarker Panel of Radiation-Upregulated miRNA as Signature for Ionizing Radiation Exposure. Life 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Mo, L.-J.; Song, M.; Huang, Q.-H.; Guan, H.; Liu, X.-D.; Xie, D.-F.; Huang, B.; Huang, R.-X.; Zhou, P.-K. Exosome-Packaged miR-1246 Contributes to Bystander DNA Damage by Targeting LIG4. Br. J. Cancer 2018, 119, 492–502. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Shi, L.; Yang, C.; Ge, Y.; Lin, L.; Fan, H.; He, Y.; Zhang, D.; Miao, Y.; Yang, L. miR-1254 Inhibits Cell Proliferation, Migration, and Invasion by down-Regulating Smurf1 in Gastric Cancer. Cell Death Dis. 2019, 10, 32. [Google Scholar] [CrossRef] [PubMed]
- Xin, Y.; Wang, X.; Meng, K.; Ni, C.; Lv, Z.; Guan, D. Identification of Exosomal miR-455-5p and miR-1255a as Therapeutic Targets for Breast Cancer. Biosci. Rep. 2020, 40. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Zhen, S.; Jin, L.; Chen, J.; Han, Y.; Lei, W.; Zhang, F. miRNA-1260b Promotes Breast Cancer Cell Migration and Invasion by Downregulating CCDC134. Curr. Gene Ther. 2023, 23, 60–71. [Google Scholar]
- Lei, T.; Zhang, L.; Song, Y.; Wang, B.; Shen, Y.; Zhang, N.; Yang, M. Transcriptionally Modulated by an Enhancer Genetic Variant Improves Efficiency of Epidermal Growth Factor Receptor-Tyrosine Kinase Inhibitors in Advanced Lung Adenocarcinoma. DNA Cell Biol. 2020, 39, 1111–1118. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, Y.; Ren, J.; Deng, S.; Yi, G.; Guo, M.; Shu, S.; Zhao, L.; Peng, Y.; Qi, S. miR-1268a Regulates ABCC1 Expression to Mediate Temozolomide Resistance in Glioblastoma. J. Neurooncol. 2018, 138, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.-J.; Chen, X.; Wang, Y.-W.; Liu, H.-T.; Ma, R.-R.; Gao, P. MiR-1268b Confers Chemosensitivity in Breast Cancer by Targeting ERBB2-Mediated PI3K-AKT Pathway. Oncotarget 2017, 8, 89631–89642. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Zhang, Z.; Zheng, P.; Zhao, W.; Han, N. MicroRNA-1285-5p Influences the Proliferation and Metastasis of Non-Small-Cell Lung Carcinoma Cells via Downregulating CDH1 and Smad4. Tumour Biol. 2017, 39, 1010428317705513. [Google Scholar] [CrossRef]
- Li, M.; He, X.-Y.; Zhang, Z.-M.; Li, S.; Ren, L.-H.; Cao, R.-S.; Feng, Y.-D.; Ji, Y.-L.; Zhao, Y.; Shi, R.-H. MicroRNA-1290 Promotes Esophageal Squamous Cell Carcinoma Cell Proliferation and Metastasis. World J. Gastroenterol. 2015, 21, 3245–3255. [Google Scholar]
- miRDB - MicroRNA Target Prediction Database Available online:. Available online: https://mirdb.org/ (accessed on 6 June 2023).
- Byun, Y.J.; Piao, X.M.; Jeong, P.; Kang, H.W.; Seo, S.P.; Moon, S.K.; Lee, J.Y.; Choi, Y.H.; Lee, H.Y.; Kim, W.T.; et al. Urinary microRNA-1913 to microRNA-3659 Expression Ratio as a Non-Invasive Diagnostic Biomarker for Prostate Cancer. Investig Clin Urol 2021, 62, 340–348. [Google Scholar] [CrossRef]
- Xu, J.; Wan, X.; Chen, X.; Fang, Y.; Cheng, X.; Xie, X.; Lu, W. miR-2861 Acts as a Tumor Suppressor via Targeting EGFR/AKT2/CCND1 Pathway in Cervical Cancer Induced by Human Papillomavirus Virus 16 E6. Sci. Rep. 2016, 6, 28968. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Deng, L.; Ji, Y.; Cheng, G.; Su, D.; Qiu, B. Novel Long Noncoding RNA (lncRNA) Panel as Biomarkers for Prognosis in Lung Squamous Cell Carcinoma via Competitive Endogenous RNA (ceRNA) Network Analysis. Transl. Cancer Res. 2021, 10, 393–405. [Google Scholar] [CrossRef]
- miRDB - MicroRNA Target Prediction Database Available online:. Available online: https://mirdb.org/ (accessed on 6 June 2023).
- Liu, X.; He, Y.; Feng, Z.; Sheng, J.; Dong, A.; Zhang, M.; Cao, L. miR-345-5p Regulates Adipogenesis via Targeting VEGF-B. Aging 2020, 12, 17114–17121. [Google Scholar] [CrossRef]
- Yu, S.; Cao, S.; Hong, S.; Lin, X.; Guan, H.; Chen, S.; Zhang, Q.; Lv, W.; Li, Y.; Xiao, H. miR-3619-3p Promotes Papillary Thyroid Carcinoma Progression via Wnt/β-Catenin Pathway. Ann Transl Med 2019, 7, 643. [Google Scholar] [CrossRef] [PubMed]
- Persson, H.; Kvist, A.; Rego, N.; Staaf, J.; Vallon-Christersson, J.; Luts, L.; Loman, N.; Jonsson, G.; Naya, H.; Hoglund, M.; et al. Identification of New microRNAs in Paired Normal and Tumor Breast Tissue Suggests a Dual Role for the ERBB2/Her2 Gene. Cancer Res. 2011, 71, 78–86. [Google Scholar] [CrossRef]
- Komina, A.; Palkina, N.; Aksenenko, M.; Tsyrenzhapova, S.; Ruksha, T. Antiproliferative and Pro-Apoptotic Effects of MiR-4286 Inhibition in Melanoma Cells. PLoS One 2016, 11, e0168229. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, S.; Wang, H.; Zhang, B.; Zhang, P. miR-4286 Promotes Prostate Cancer Progression via Targeting the Expression of SALL1. J. J. Gene Med. 2019, e3127. [Google Scholar]
- He, Z.; Xue, H.; Liu, P.; Han, D.; Xu, L.; Zeng, X.; Wang, J.; Yang, B.; Luo, B. miR-4286/TGF-β1/Smad3-Negative Feedback Loop Ameliorated Vascular Endothelial Cell Damage by Attenuating Apoptosis and Inflammatory Response. J. Cardiovasc. Pharmacol. 2020, 75, 446–454. [Google Scholar] [CrossRef]
- Ling, C.; Wang, X.; Zhu, J.; Tang, H.; Du, W.; Zeng, Y.; Sun, L.; Huang, J.-A.; Liu, Z. MicroRNA-4286 Promotes Cell Proliferation, Migration, and Invasion via PTEN Regulation of the PI3K/Akt Pathway in Non-Small Cell Lung Cancer. Cancer Med. 2019, 8, 3520–3531. [Google Scholar] [CrossRef]
- Yang, H.; Zhang, W.; Luan, Q.; Liu, Y. miR-4284 Promotes Cell Proliferation, Migration, and Invasion in Non-Small Cell Lung Cancer Cells and Is Associated with Postoperative Prognosis. Cancer Manag. Res. 2021, 13, 5865–5872. [Google Scholar] [CrossRef]
- Zuo, X.-M.; Sun, H.-W.; Fang, H.; Wu, Y.; Shi, Q.; Yu, Y.-F. miR-4443 Targets TRIM14 to Suppress Metastasis and Energy Metabolism of Papillary Thyroid Carcinoma (PTC) in Vitro. Cell Biol. Int. 2021, 45, 1917–1925. [Google Scholar]
- Herrero-Aguayo, V.; Jiménez-Vacas, J.M.; Sáez-Martínez, P.; Gómez-Gómez, E.; López-Cánovas, J.L.; Garrido-Sánchez, L.; Herrera-Martínez, A.D.; García-Bermejo, L.; Macías-González, M.; López-Miranda, J.; et al. Influence of Obesity in the miRNome: miR-4454, a Key Regulator of Insulin Response Via Splicing Modulation in Prostate. J. Clin. Endocrinol. Metab. 2021, 106, e469–e484. [Google Scholar] [CrossRef]
- Dasari, S.; Pandhiri, T.; Grassi, T.; Visscher, D.W.; Multinu, F.; Agarwal, K.; Mariani, A.; Shridhar, V.; Mitra, A.K. Signals from the Metastatic Niche Regulate Early and Advanced Ovarian Cancer Metastasis through miR-4454 Downregulation. Mol. Cancer Res. 2020, 18, 1202–1217. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hu, H.; Luo, Z.; Liu, S.; Wu, W.; Zhu, M.; Wang, J.; Liu, Y.; Lu, Z. miR-4454 up-Regulated by HPV16 E6/E7 Promotes Invasion and Migration by Targeting ABHD2/NUDT21 in Cervical Cancer. Biosci. Rep. 2020, 40. [Google Scholar] [CrossRef]
- Kannathasan, T.; Kuo, W.-W.; Chen, M.-C.; Viswanadha, V.P.; Shen, C.-Y.; Tu, C.-C.; Yeh, Y.-L.; Bharath, M.; Shibu, M.A.; Huang, C.-Y. Chemoresistance-Associated Silencing of miR-4454 Promotes Colorectal Cancer Aggression through the GNL3L and NF-κB Pathway. Cancers 2020, 12. [Google Scholar] [CrossRef]
- Nawaz, Z.; Patil, V.; Thinagararjan, S.; Rao, S.A.; Hegde, A.S.; Arivazhagan, A.; Santosh, V.; Somasundaram, K. Impact of Somatic Copy Number Alterations on the Glioblastoma miRNome: miR-4484 Is a Genomically Deleted Tumour Suppressor. Mol. Oncol. 2017, 11, 927–944. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, Y.; Wang, L.; Kang, Q.; Yu, G.; Wan, X.; Wang, J.; Zhu, K. MiR-4505 Aggravates Lipopolysaccharide-Induced Vascular Endothelial Injury by Targeting Heat Shock Protein A12B. Mol. Med. Rep. 2018, 17, 1389–1395. [Google Scholar]
- Zhao, M.; Tang, Z.; Wang, Y.; Ding, J.; Guo, Y.; Zhang, N.; Gao, T. MIR-4507 Targets to Facilitate the Malignant Progression of Non-Small-Cell Lung Cancer. J. Cancer 2021, 12, 6600–6609. [Google Scholar] [CrossRef]
- Herdoiza Padilla, E.; Crauwels, P.; Bergner, T.; Wiederspohn, N.; Förstner, S.; Rinas, R.; Ruf, A.; Kleemann, M.; Handrick, R.; Tuckermann, J.; et al. Mir-124-5p Regulates Phagocytosis of Human Macrophages by Targeting the Actin Cytoskeleton via the ARP2/3 Complex. Front. Immunol. 2019, 10, 2210. [Google Scholar] [CrossRef]
- Yang, L.; Hou, Y.; Du, Y.-E.; Li, Q.; Zhou, F.; Li, Y.; Zeng, H.; Jin, T.; Wan, X.; Guan, S.; et al. Mirtronic miR-4646-5p Promotes Gastric Cancer Metastasis by Regulating ABHD16A and Metabolite Lysophosphatidylserines. Cell Death Differ. 2021, 28, 2708–2727. [Google Scholar] [CrossRef]
- Jia, J.; Ouyang, Z.; Wang, M.; Ma, W.; Liu, M.; Zhang, M.; Yu, M. MicroRNA-361-5p Slows down Gliomas Development through Regulating UBR5 to Elevate ATMIN Protein Expression. Cell Death Dis. 2021, 12, 746. [Google Scholar] [CrossRef]
- Icli, B.; Li, H.; Pérez-Cremades, D.; Wu, W.; Ozdemir, D.; Haemmig, S.; Guimaraes, R.B.; Manica, A.; Marchini, J.F.; Orgill, D.P.; et al. MiR-4674 Regulates Angiogenesis in Tissue Injury by Targeting p38K Signaling in Endothelial Cells. Am. J. Physiol. Cell Physiol. 2020, 318, C524–C535. [Google Scholar] [CrossRef]
- Wang, W.; Ji, J.; Li, J.; Ren, Q.; Gu, J.; Zhao, Y.; Hong, D.; Guo, Q.; Tan, Y. Several Critical Genes and microRNAs Associated with the Development of Polycystic Ovary Syndrome. Ann. Endocrinol. 2020, 81, 18–27. [Google Scholar] [CrossRef]
- Su, Y.; Xu, C.; Liu, Y.; Hu, Y.; Wu, H. Circular RNA hsa_circ_0001649 Inhibits Hepatocellular Carcinoma Progression Multiple miRNAs Sponge. Aging 2019, 11, 3362–3375. [Google Scholar] [CrossRef]
- Deng, G.; Wang, R.; Sun, Y.; Huang, C.-P.; Yeh, S.; You, B.; Feng, C.; Li, G.; Ma, S.; Chang, C. Targeting Androgen Receptor (AR) with Antiandrogen Enzalutamide Increases Prostate Cancer Cell Invasion yet Decreases Bladder Cancer Cell Invasion via Differentially Altering the AR/circRNA-ARC1/miR-125b-2-3p or miR-4736/PPARγ/MMP-9 Signals. Cell Death Differ. 2021, 28, 2145–2159. [Google Scholar] [CrossRef]
- Song, Y.; Meng, Z.; Zhang, S.; Li, N.; Hu, W.; Li, H. miR-4739/ITGA10/PI3K Signaling Regulates Differentiation and Apoptosis of Osteoblast. Regen Ther 2022, 21, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.-P.; Fu, M.; Masula, M.; Xing, C.-S.; Zhou, Q.; Tan, J.-T.; Wang, J. miR-3064-5p and miR-4745-5p Affect Heparin Sensitivity in Patients Undergoing Cardiac Surgery by Regulating AT-III and Factor X mRNA Levels. Front. Physiol. 2022, 13, 914333. [Google Scholar] [CrossRef] [PubMed]
- miRDB - MicroRNA Target Prediction Database Available online:. Available online: https://mirdb.org/ (accessed on 6 June 2023).
- Mercuri, R.L.; Conceição, H.B.; Guardia, G.D.A.; Goldstein, G.; Vibranovski, M.D.; Hinske, L.C.; Galante, P.A.F. Retro-miRs: Novel and Functional miRNAs Originated from mRNA Retrotransposition. bioRxiv 2023.
- Dai, Y.-C.; Pan, Y.; Quan, M.-M.; Chen, Q.; Pan, Y.; Ruan, Y.-Y.; Sun, J.-G. MicroRNA-1246 Mediates Drug Resistance and Metastasis in Breast Cancer by Targeting NFE2L3. Front. Oncol. 2021, 11, 677168. [Google Scholar]
- Reza, A.M.M.T.; Choi, Y.-J.; Yuan, Y.-G.; Das, J.; Yasuda, H.; Kim, J.-H. MicroRNA-7641 Is a Regulator of Ribosomal Proteins and a Promising Targeting Factor to Improve the Efficacy of Cancer Therapy. Sci. Rep. 2017, 7, 8365. [Google Scholar] [CrossRef] [PubMed]
- MicroRNAs in Drug Addiction: Current Status and Future Perspectives. Pharmacol. Ther. 2022, 236, 108215. [CrossRef]
- Yoshida, M.; Horiguchi, H.; Kikuchi, S.; Iyama, S.; Ikeda, H.; Goto, A.; Kawano, Y.; Murase, K.; Takada, K.; Miyanishi, K.; et al. miR-7977 Inhibits the Hippo-YAP Signaling Pathway in Bone Marrow Mesenchymal Stromal Cells. PLoS One 2019, 14, e0213220. [Google Scholar] [CrossRef]
- Gao, Z.; Wang, Z.; Zhu, H.; Yuan, X.; Sun, M.; Wang, J.; Zuo, M.; Cui, X.; Han, Y.; Zhang, Y.; et al. Hyperinsulinemia Contributes to Impaired-Glucose-Tolerance-Induced Renal Injury Mir-7977/SIRT3 Signaling. Ther. Adv. Chronic Dis. 2020, 11, 2040622320916008. [Google Scholar] [CrossRef]
- Ni, J.; Wang, X.; Chen, S.; Liu, H.; Wang, Y.; Xu, X.; Cheng, J.; Jia, J.; Zhen, X. MicroRNA Let-7c-5p Protects against Cerebral Ischemia Injury via Mechanisms Involving the Inhibition of Microglia Activation. Brain Behav. Immun. 2015, 49, 75–85. [Google Scholar]
- Pregnolato, F.; Cova, L.; Doretti, A.; Bardelli, D.; Silani, V.; Bossolasco, P. Exosome microRNAs in Amyotrophic Lateral Sclerosis: A Pilot Study. Biomolecules 2021, 11. [Google Scholar] [CrossRef]
- Rizzuti, M.; Filosa, G.; Melzi, V.; Calandriello, L.; Dioni, L.; Bollati, V.; Bresolin, N.; Comi, G.P.; Barabino, S.; Nizzardo, M.; et al. MicroRNA Expression Analysis Identifies a Subset of Downregulated miRNAs in ALS Motor Neuron Progenitors. Sci. Rep. 2018, 8, 10105. [Google Scholar] [CrossRef] [PubMed]
- Agostini, M.; Tucci, P.; Steinert, J.R.; Shalom-Feuerstein, R.; Rouleau, M.; Aberdam, D.; Forsythe, I.D.; Young, K.W.; Ventura, A.; Concepcion, C.P.; et al. microRNA-34a Regulates Neurite Outgrowth, Spinal Morphology, and Function. Proc. Natl. Acad. Sci. U. S. A. 2011, 108, 21099–21104. [Google Scholar] [CrossRef] [PubMed]
- Sproviero, D.; Gagliardi, S.; Zucca, S.; Arigoni, M.; Giannini, M.; Garofalo, M.; Olivero, M.; Dell’Orco, M.; Pansarasa, O.; Bernuzzi, S.; et al. Different miRNA Profiles in Plasma Derived Small and Large Extracellular Vesicles from Patients with Neurodegenerative Diseases. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef]
- Deng, D.; Wang, L.; Chen, Y.; Li, B.; Xue, L.; Shao, N.; Wang, Q.; Xia, X.; Yang, Y.; Zhi, F. MicroRNA-124-3p Regulates Cell Proliferation, Invasion, Apoptosis, and Bioenergetics by Targeting PIM1 in Astrocytoma. Cancer Sci. 2016, 107, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Liu, S.; Yan, J.; Sun, M.-Z.; Greenaway, F.T. The Potential Role of miR-124-3p in Tumorigenesis and Other Related Diseases. Mol. Biol. Rep. 2021, 48, 3579–3591. [Google Scholar] [CrossRef] [PubMed]
- Long, H.-D.; Ma, Y.-S.; Yang, H.-Q.; Xue, S.-B.; Liu, J.-B.; Yu, F.; Lv, Z.-W.; Li, J.-Y.; Xie, R.-T.; Chang, Z.-Y.; et al. Reduced Hsa-miR-124-3p Levels Are Associated with the Poor Survival of Patients with Hepatocellular Carcinoma. Mol. Biol. Rep. 2018, 45, 2615–2623. [Google Scholar] [CrossRef]
- Majid, A.; Wang, J.; Nawaz, M.; Abdul, S.; Ayesha, M.; Guo, C.; Liu, Q.; Liu, S.; Sun, M.-Z. miR-124-3p Suppresses the Invasiveness and Metastasis of Hepatocarcinoma Cells via Targeting CRKL. Front. Mol. Biosci. 2020, 7. [Google Scholar] [CrossRef]
- Ge, X.; Guo, M.; Hu, T.; Li, W.; Huang, S.; Yin, Z.; Li, Y.; Chen, F.; Zhu, L.; Kang, C.; et al. Increased Microglial Exosomal miR-124-3p Alleviates Neurodegeneration and Improves Cognitive Outcome after rmTBI. Mol. Ther. 2020, 28, 503–522. [Google Scholar] [CrossRef]
- Vuokila, N.; Lukasiuk, K.; Bot, A.M.; van Vliet, E.A.; Aronica, E.; Pitkänen, A.; Puhakka, N. miR-124-3p Is a Chronic Regulator of Gene Expression after Brain Injury. Cell. Mol. Life Sci. 2018, 75, 4557–4581. [Google Scholar] [CrossRef]
- Iacona, J.R.; Lutz, C.S. miR-146a-5p: Expression, Regulation, and Functions in Cancer. Wiley Interdiscip. Rev. RNA 2019, 10, e1533. [Google Scholar]
- Lin, H.; Zhang, R.; Wu, W.; Lei, L. miR-4454 Promotes Hepatic Carcinoma Progression by Targeting Vps4A and Rab27A. Oxid. Med. Cell. Longev. 2021, 2021, 9230435. [Google Scholar]
- Zhang, Y.; Wang, L.-J.; Yang, H.-Q.; Wang, R.; Wu, H.-J. MicroRNA-10b Expression Predicts Long-Term Survival in Patients with Solid Tumor. J. Cell. Physiol. 2019, 234, 1248–1256. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yi, N.; Chen, R.; Meng, Y.; Wang, Y.; Liu, H.; Cao, W.; Hu, Y.; Gu, Y.; Tong, C.; et al. miR-29b-3p Protects Cardiomyocytes against Endotoxin-Induced Apoptosis and Inflammatory Response through Targeting FOXO3A. Cell. Signal. 2020, 74, 109716. [Google Scholar] [CrossRef] [PubMed]
- Foggin, S.; Mesquita-Ribeiro, R.; Dajas-Bailador, F.; Layfield, R. Biological Significance of microRNA Biomarkers in ALS-Innocent Bystanders or Disease Culprits? Front. Neurol. 2019, 10, 578. [Google Scholar]
- Toivonen, J.M.; Manzano, R.; Oliván, S.; Zaragoza, P.; García-Redondo, A.; Osta, R. MicroRNA-206: A Potential Circulating Biomarker Candidate for Amyotrophic Lateral Sclerosis. PLoS One 2014, 9, e89065. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Lee, Y.S.; Sivaprasad, U.; Malhotra, A.; Dutta, A. Muscle-Specific microRNA miR-206 Promotes Muscle Differentiation. J. Cell Biol. 2006, 174, 677–687. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, J.J. MicroRNA-206: The Skeletal Muscle-Specific myomiR. Biochim. Biophys. Acta 2008, 1779, 682–691. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.H.; Valdez, G.; Moresi, V.; Qi, X.; McAnally, J.; Elliott, J.L.; Bassel-Duby, R.; Sanes, J.R.; Olson, E.N. MicroRNA-206 Delays ALS Progression and Promotes Regeneration of Neuromuscular Synapses in Mice. Science 2009, 326, 1549–1554. [Google Scholar] [CrossRef] [PubMed]
- Przanowska, R.K.; Sobierajska, E.; Su, Z.; Jensen, K.; Przanowski, P.; Nagdas, S.; Kashatus, J.A.; Kashatus, D.F.; Bhatnagar, S.; Lukens, J.R.; et al. miR-206 Family Is Important for Mitochondrial and Muscle Function, but Not Essential for Myogenesis in Vitro. FASEB J. 2020, 34, 7687–7702. [Google Scholar] [CrossRef] [PubMed]
- Waller, R.; Goodall, E.F.; Milo, M.; Cooper-Knock, J.; Da Costa, M.; Hobson, E.; Kazoka, M.; Wollff, H.; Heath, P.R.; Shaw, P.J.; et al. Serum miRNAs miR-206, 143-3p and 374b-5p as Potential Biomarkers for Amyotrophic Lateral Sclerosis (ALS). Neurobiol. Aging 2017, 55, 123–131. [Google Scholar]
- de Andrade, H.M.T.; de Albuquerque, M.; Avansini, S.H.; de S Rocha, C.; Dogini, D.B.; Nucci, A.; Carvalho, B.; Lopes-Cendes, I.; França, M.C. , Jr MicroRNAs-424 and 206 Are Potential Prognostic Markers in Spinal Onset Amyotrophic Lateral Sclerosis. J. Neurol. Sci. 2016, 368, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Vassileff, N.; Vella, L.J.; Rajapaksha, H.; Shambrook, M.; Kenari, A.N.; McLean, C.; Hill, A.F.; Cheng, L. Revealing the Proteome of Motor Cortex Derived Extracellular Vesicles Isolated from Amyotrophic Lateral Sclerosis Human Postmortem Tissues. Cells 2020, 9. [Google Scholar] [CrossRef]
- Pasetto, L.; Callegaro, S.; Corbelli, A.; Fiordaliso, F.; Ferrara, D.; Brunelli, L.; Sestito, G.; Pastorelli, R.; Bianchi, E.; Cretich, M.; et al. Decoding Distinctive Features of Plasma Extracellular Vesicles in Amyotrophic Lateral Sclerosis. Mol. Neurodegener. 2021, 16, 52. [Google Scholar] [CrossRef]
- Sjoqvist, S.; Otake, K. A Pilot Study Using Proximity Extension Assay of Cerebrospinal Fluid and Its Extracellular Vesicles Identifies Novel Amyotrophic Lateral Sclerosis Biomarker Candidates. Biochem. Biophys. Res. Commun. 2022, 613, 166–173. [Google Scholar] [CrossRef]
- Hayes, A.J.; Farrugia, B.L.; Biose, I.J.; Bix, G.J.; Melrose, J. Perlecan, A Multi-Functional, Cell-Instructive, Matrix-Stabilizing Proteoglycan With Roles in Tissue Development Has Relevance to Connective Tissue Repair and Regeneration. Front Cell Dev Biol 2022, 10, 856261. [Google Scholar] [CrossRef]
- Thompson, A.G.; Gray, E.; Mäger, I.; Thézénas, M.-L.; Charles, P.D.; Talbot, K.; Fischer, R.; Kessler, B.M.; Wood, M.; Turner, M.R. CSF Extracellular Vesicle Proteomics Demonstrates Altered Protein Homeostasis in Amyotrophic Lateral Sclerosis. Clin. Proteomics 2020, 17, 31. [Google Scholar] [CrossRef] [PubMed]
- Riise, R.; Odqvist, L.; Mattsson, J.; Monkley, S.; Abdillahi, S.M.; Tyrchan, C.; Muthas, D.; Yrlid, L.F. Bleomycin Hydrolase Regulates the Release of Chemokines Important for Inflammation and Wound Healing by Keratinocytes. Sci. Rep. 2019, 9, 20407. [Google Scholar] [CrossRef] [PubMed]
- Skytthe, M.K.; Graversen, J.H.; Moestrup, S.K. Targeting of CD163 Macrophages in Inflammatory and Malignant Diseases. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef]
- Thome, A.D.; Thonhoff, J.R.; Zhao, W.; Faridar, A.; Wang, J.; Beers, D.R.; Appel, S.H. Extracellular Vesicles Derived From Expanded Regulatory T Cells Modulate and Inflammation. Front. Immunol. 2022, 13, 875825. [Google Scholar] [CrossRef]
- Chang, D.; Sharma, L.; Dela Cruz, C.S. Chitotriosidase: A Marker and Modulator of Lung Disease. Eur. Respir. Rev. 2020, 29. [Google Scholar] [CrossRef]
- Man, J.; Zhang, X. CUEDC2: An Emerging Key Player in Inflammation and Tumorigenesis. Protein Cell 2011, 2, 699–703. [Google Scholar] [CrossRef]
- Otake, K.; Kamiguchi, H.; Hirozane, Y. Identification of Biomarkers for Amyotrophic Lateral Sclerosis by Comprehensive Analysis of Exosomal mRNAs in Human Cerebrospinal Fluid. BMC Med. Genomics 2019, 12, 7. [Google Scholar] [CrossRef]
- Kamelgarn, M.; Chen, J.; Kuang, L.; Arenas, A.; Zhai, J.; Zhu, H.; Gal, J. Proteomic Analysis of FUS Interacting Proteins Provides Insights into FUS Function and Its Role in ALS. Biochim. Biophys. Acta 2016, 1862, 2004–2014. [Google Scholar] [CrossRef]
- Zuehlke, A.D.; Moses, M.A.; Neckers, L. Heat Shock Protein 90: Its Inhibition and Function. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2018, 373. [Google Scholar] [CrossRef] [PubMed]
- Sugano, Y.; Takeuchi, M.; Hirata, A.; Matsushita, H.; Kitamura, T.; Tanaka, M.; Miyajima, A. Junctional Adhesion Molecule-A, JAM-A, Is a Novel Cell-Surface Marker for Long-Term Repopulating Hematopoietic Stem Cells. Blood 2008, 111, 1167–1172. [Google Scholar] [CrossRef] [PubMed]
- Burke, S.J.; Updegraff, B.L.; Bellich, R.M.; Goff, M.R.; Lu, D.; Minkin, S.C., Jr; Karlstad, M.D.; Collier, J.J. Regulation of iNOS Gene Transcription by IL-1β and IFN-γ Requires a Coactivator Exchange Mechanism. Mol. Endocrinol. 2013, 27, 1724–1742. [Google Scholar] [CrossRef]
- Ordway, G.A.; Garry, D.J. Myoglobin: An Essential Hemoprotein in Striated Muscle. J. Exp. Biol. 2004, 207, 3441–3446. [Google Scholar] [CrossRef]
- Garry, D.J.; Mammen, P.P.A. Molecular Insights into the Functional Role of Myoglobin. Adv. Exp. Med. Biol. 2007, 618, 181–193. [Google Scholar]
- Bunton-Stasyshyn, R.K.A.; Saccon, R.A.; Fratta, P.; Fisher, E.M.C. SOD1 Function and Its Implications for Amyotrophic Lateral Sclerosis Pathology: New and Renascent Themes. Neuroscientist 2015, 21, 519–529. [Google Scholar] [CrossRef]
- Heyne, K.; Förster, J.; Schüle, R.; Roemer, K. Transcriptional Repressor NIR Interacts with the p53-Inhibiting Ubiquitin Ligase MDM2. Nucleic Acids Res. 2014, 42, 3565–3579. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, N.; Doi, H.; Kurata, Y.; Kagawa, H.; Atobe, Y.; Funakoshi, K.; Tada, M.; Katsumoto, A.; Tanaka, K.; Kunii, M.; et al. Proteomic Analysis of Exosome-Enriched Fractions Derived from Cerebrospinal Fluid of Amyotrophic Lateral Sclerosis Patients. Neurosci. Res. 2020, 160, 43–49. [Google Scholar] [CrossRef]
- Vomhof-Dekrey, E.E.; Picklo, M.J. , Sr The Nrf2-Antioxidant Response Element Pathway: A Target for Regulating Energy Metabolism. J. Nutr. Biochem. 2012, 23, 1201–1206. [Google Scholar] [CrossRef] [PubMed]
- Provenzano, F.; Nyberg, S.; Giunti, D.; Torazza, C.; Parodi, B.; Bonifacino, T.; Usai, C.; Kerlero de Rosbo, N.; Milanese, M.; Uccelli, A.; et al. Micro-RNAs Shuttled by Extracellular Vesicles Secreted from Mesenchymal Stem Cells Dampen Astrocyte Pathological Activation and Support Neuroprotection in In-Vitro Models of ALS. Cells 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Bourtchuladze, R.; Frenguelli, B.; Blendy, J.; Cioffi, D.; Schutz, G.; Silva, A.J. Deficient Long-Term Memory in Mice with a Targeted Mutation of the cAMP-Responsive Element-Binding Protein. Cell 1994, 79, 59–68. [Google Scholar] [CrossRef]
- Lee, M.; Ban, J.-J.; Kim, K.Y.; Jeon, G.S.; Im, W.; Sung, J.-J.; Kim, M. Adipose-Derived Stem Cell Exosomes Alleviate Pathology of Amyotrophic Lateral Sclerosis in Vitro. Biochem. Biophys. Res. Commun. 2016, 479, 434–439. [Google Scholar] [CrossRef]
- Liang, H.; Ward, W.F. PGC-1alpha: A Key Regulator of Energy Metabolism. Adv. Physiol. Educ. 2006, 30, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.E. An Overview of Phenylalanine and Tyrosine Kinetics in Humans. J. Nutr. 2007, 137, 1549S–1555S; discussion 1573S–1575S. [Google Scholar] [CrossRef] [PubMed]
- Morasso, C.F.; Sproviero, D.; Mimmi, M.C.; Giannini, M.; Gagliardi, S.; Vanna, R.; Diamanti, L.; Bernuzzi, S.; Piccotti, F.; Truffi, M.; et al. Raman Spectroscopy Reveals Biochemical Differences in Plasma Derived Extracellular Vesicles from Sporadic Amyotrophic Lateral Sclerosis Patients. Nanomedicine 2020, 29, 102249. [Google Scholar] [CrossRef] [PubMed]
- Butler, R.E.; Krishnan, N.; Garcia-Jimenez, W.; Francis, R.; Martyn, A.; Mendum, T.; Felemban, S.; Locker, N.; Salguero, F.J.; Robertson, B.; et al. Susceptibility of Mycobacterium Tuberculosis-Infected Host Cells to Phospho-MLKL Driven Necroptosis Is Dependent on Cell Type and Presence of TNFα. Virulence 2017, 8, 1820–1832. [Google Scholar] [CrossRef]
- Ren, L.-Q.; Liu, W.; Li, W.-B.; Liu, W.-J.; Sun, L. Peptidylprolyl Cis/trans Isomerase Activity and Molecular Evolution of Vertebrate Cyclophilin A. Yi Chuan 2016, 38, 736–745. [Google Scholar]
- Boylan, K.B.; Glass, J.D.; Crook, J.E.; Yang, C.; Thomas, C.S.; Desaro, P.; Johnston, A.; Overstreet, K.; Kelly, C.; Polak, M.; et al. Phosphorylated Neurofilament Heavy Subunit (pNF-H) in Peripheral Blood and CSF as a Potential Prognostic Biomarker in Amyotrophic Lateral Sclerosis. J. Neurol. Neurosurg. Psychiatry 2013, 84, 467–472. [Google Scholar] [CrossRef]
- Scotter, E.L.; Chen, H.-J.; Shaw, C.E. TDP-43 Proteinopathy and ALS: Insights into Disease Mechanisms and Therapeutic Targets. Neurotherapeutics 2015, 12, 352–363. [Google Scholar] [CrossRef]
- Feneberg, E.; Steinacker, P.; Lehnert, S.; Schneider, A.; Walther, P.; Thal, D.R.; Linsenmeier, M.; Ludolph, A.C.; Otto, M. Limited Role of Free TDP-43 as a Diagnostic Tool in Neurodegenerative Diseases. Amyotroph. Lateral Scler. Frontotemporal Degener. 2014, 15, 351–356. [Google Scholar] [CrossRef]
- Medler, J.; Wajant, H. Tumor Necrosis Factor Receptor-2 (TNFR2): An Overview of an Emerging Drug Target. Expert Opin. Ther. Targets 2019, 23, 295–307. [Google Scholar] [CrossRef]
- Groen, E.J.N.; Gillingwater, T.H. UBA1: At the Crossroads of Ubiquitin Homeostasis and Neurodegeneration. Trends Mol. Med. 2015, 21, 622–632. [Google Scholar]
- Bobis-Wozowicz, S.; Marbán, E. Editorial: Extracellular Vesicles as Next Generation Therapeutics. Front Cell Dev Biol 2022, 10, 919426. [Google Scholar] [CrossRef]
- Cataldi, M.; Vigliotti, C.; Mosca, T.; Cammarota, M.; Capone, D. Emerging Role of the Spleen in the Pharmacokinetics of Monoclonal Antibodies, Nanoparticles and Exosomes. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wu, X.; Chandra, S.; Lyon, C.; Ning, B.; Jiang, L.; Fan, J.; Hu, T.Y. Extracellular Vesicles: Emerging Tools as Therapeutic Agent Carriers. Acta Pharm Sin B 2022, 12, 3822–3842. [Google Scholar] [CrossRef] [PubMed]
- Bonafede, R.; Mariotti, R. ALS Pathogenesis and Therapeutic Approaches: The Role of Mesenchymal Stem Cells and Extracellular Vesicles. Front. Cell. Neurosci. 2017, 11, 80. [Google Scholar] [CrossRef]
- Belkozhayev, A.M.; Al-Yozbaki, M.; George, A.; Ye Niyazova, R.; Sharipov, K.O.; Byrne, L.J.; Wilson, C.M. Extracellular Vesicles, Stem Cells and the Role of miRNAs in Neurodegeneration. Curr. Neuropharmacol. 2022, 20, 1450–1478. [Google Scholar]
- Sadanandan, N.; Lee, J.-Y.; Garbuzova-Davis, S. Extracellular Vesicle-Based Therapy for Amyotrophic Lateral Sclerosis. Brain Circ 2021, 7, 23–28. [Google Scholar] [PubMed]
- Bonafede, R.; Scambi, I.; Peroni, D.; Potrich, V.; Boschi, F.; Benati, D.; Bonetti, B.; Mariotti, R. Exosome Derived from Murine Adipose-Derived Stromal Cells: Neuroprotective Effect on in Vitro Model of Amyotrophic Lateral Sclerosis. Exp. Cell Res. 2016, 340, 150–158. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Y.; Jin, T.; Botchway, B.O.A.; Fan, R.; Wang, L.; Liu, X. Adipose-Derived Mesenchymal Stem Cells Combined With Extracellular Vesicles May Improve Amyotrophic Lateral Sclerosis. Front. Aging Neurosci. 2022, 14, 830346. [Google Scholar] [CrossRef]
- Bonafede, R.; Brandi, J.; Manfredi, M.; Scambi, I.; Schiaffino, L.; Merigo, F.; Turano, E.; Bonetti, B.; Marengo, E.; Cecconi, D.; et al. The Anti-Apoptotic Effect of ASC-Exosomes in an In Vitro ALS Model and Their Proteomic Analysis. Cells 2019, 8. [Google Scholar] [CrossRef]
- Bonafede, R.; Turano, E.; Scambi, I.; Busato, A.; Bontempi, P.; Virla, F.; Schiaffino, L.; Marzola, P.; Bonetti, B.; Mariotti, R. ASC-Exosomes Ameliorate the Disease Progression in SOD1(G93A) Murine Model Underlining Their Potential Therapeutic Use in Human ALS. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef]
- Calabria, E.; Scambi, I.; Bonafede, R.; Schiaffino, L.; Peroni, D.; Potrich, V.; Capelli, C.; Schena, F.; Mariotti, R. ASCs-Exosomes Recover Coupling Efficiency and Mitochondrial Membrane Potential in an Model of ALS. Front. Neurosci. 2019, 13, 1070. [Google Scholar] [CrossRef] [PubMed]
- Garbuzova-Davis, S.; Willing, A.E.; Ehrhart, J.; Wang, L.; Sanberg, P.R.; Borlongan, C.V. Cell-Free Extracellular Vesicles Derived from Human Bone Marrow Endothelial Progenitor Cells as Potential Therapeutics for Microvascular Endothelium Restoration in ALS. Neuromolecular Med. 2020, 22, 503–516. [Google Scholar] [CrossRef]
- Niu, X.; Chen, J.; Gao, J. Nanocarriers as a Powerful Vehicle to Overcome Blood-Brain Barrier in Treating Neurodegenerative Diseases: Focus on Recent Advances. Asian J. Pharm. Sci. 2019, 14, 480–496. [Google Scholar] [CrossRef]
- Wang, K.; Li, Y.; Ren, C.; Wang, Y.; He, W.; Jiang, Y. Extracellular Vesicles as Innovative Treatment Strategy for Amyotrophic Lateral Sclerosis. Front Cell Dev Biol 2021, 9, 754630. [Google Scholar] [CrossRef] [PubMed]
- Thakur, A.; Sidu, R.K.; Zou, H.; Alam, M.K.; Yang, M.; Lee, Y. Inhibition of Glioma Cells’ Proliferation by Doxorubicin-Loaded Exosomes via Microfluidics. Int. J. Nanomedicine 2020, 15, 8331–8343. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The role of EVs in ALS. EVs contribute to the pathogenesis of ALS (left-reddish side). EVs are produced by different cell types of the central nervous system and the neuromuscular junction. In the context of ALS, EVs may carry disease-related biological molecules (proteins and miRNAs) involved in the transformation and degeneration of brain and neuromuscular elements, thus contributing to the spread of the pathology between different cell types. Furthermore, EVs can reach long distances in the body, contributing to the exchange of harmful molecules between the brain and the neuromuscular junction. Considering this, the molecules transported by EVs circulating in the bloodstream and cerebrospinal fluid have been considered potential biomarkers for the diagnosis and prognosis of ALS. Finally, EVs also have therapeutic potential. Blocking the exchange of EVs carrying harmful molecules and administering EVs with a neuroprotective cargo may slow the progression of ALS or revert its pathological effects (right-greenish side). ALS: Amyotrophic Lateral Sclerosis, EV: extracellular vesicle, miRNA: micro RNA. Figure created with BioRender.com.
Figure 1.
The role of EVs in ALS. EVs contribute to the pathogenesis of ALS (left-reddish side). EVs are produced by different cell types of the central nervous system and the neuromuscular junction. In the context of ALS, EVs may carry disease-related biological molecules (proteins and miRNAs) involved in the transformation and degeneration of brain and neuromuscular elements, thus contributing to the spread of the pathology between different cell types. Furthermore, EVs can reach long distances in the body, contributing to the exchange of harmful molecules between the brain and the neuromuscular junction. Considering this, the molecules transported by EVs circulating in the bloodstream and cerebrospinal fluid have been considered potential biomarkers for the diagnosis and prognosis of ALS. Finally, EVs also have therapeutic potential. Blocking the exchange of EVs carrying harmful molecules and administering EVs with a neuroprotective cargo may slow the progression of ALS or revert its pathological effects (right-greenish side). ALS: Amyotrophic Lateral Sclerosis, EV: extracellular vesicle, miRNA: micro RNA. Figure created with BioRender.com.
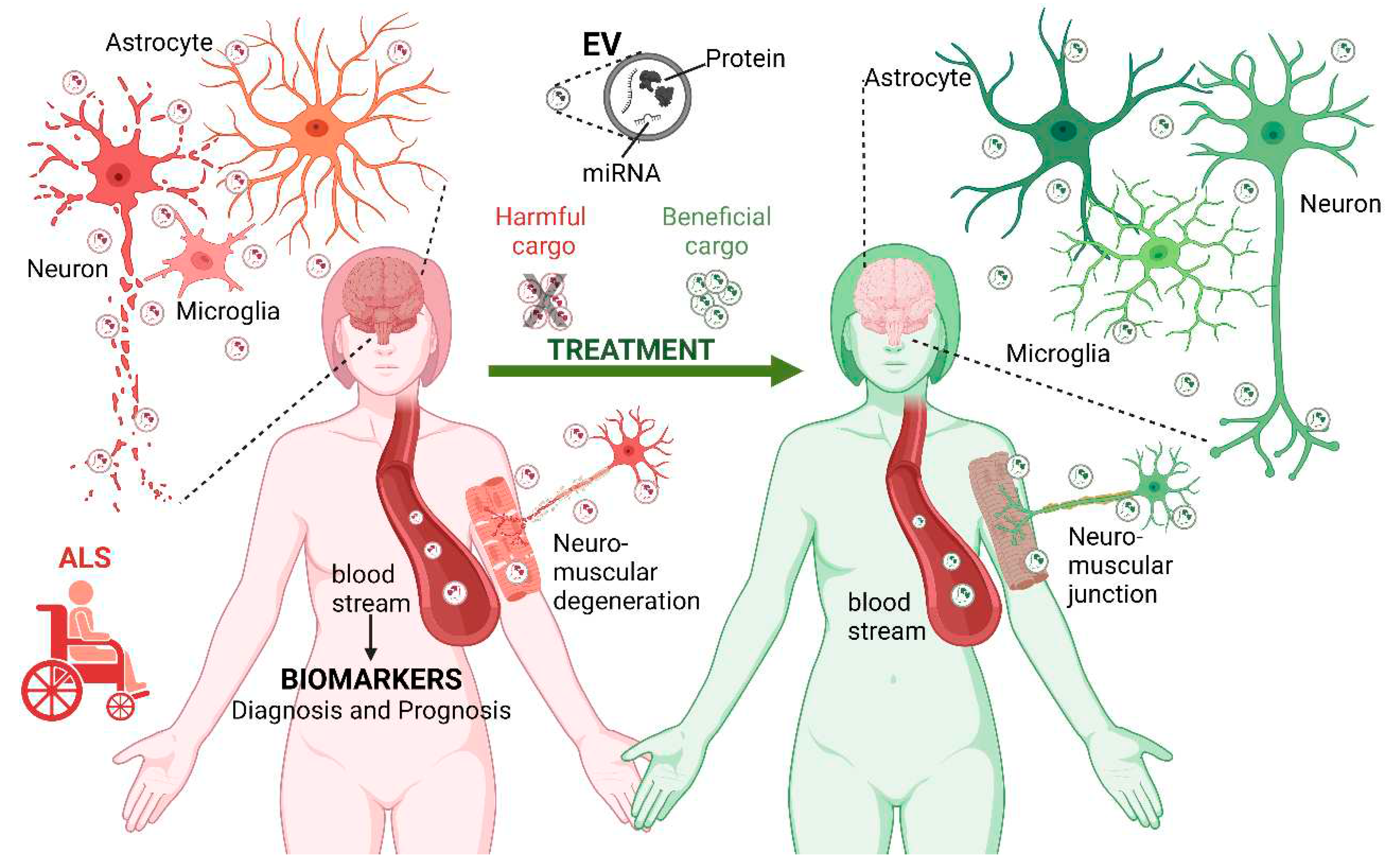

Table 1.
miRNAs cargo in Amyotrophic lateral sclerosis-related extracellular vesicles.
| miRNA | Main Targets | Biological role | Expression |
|
miR-9-1-5p, miR-9-2-5p, miR-9-3-5p* |
PAK4, CoREST, CPEB3, ECAD*, elavl3, FoxG1, Hes1, IGF2BP3, Nr2e1/TLX, REST, Sirt1, Zic5; IGF2-PI3K/Akt signaling | Cell differentiation regulation, neuronal function, synaptic plasticity neurotransmitter release; skeletal muscle cell proliferation and differentiation inhibition regulation [140]; apoptosis inhibition [142] | downregulated: blood plasma-derived EVs [143] |
| miR-10b-5p | NFAT5; KLF11-KIT signaling | Regulation of insulin production, lipid metabolism and gastrointestinal motility [144], myoblasts differentiation [145]. Tumorigenic inhibitor [146] | downregulated: CSF exosomes and blood plasma-derived EVs [147,148] |
| miR-15a-5p | BCL2, Cyclin D1, FEAT, PD-1, ROR1, CXCL10-ERK-LIN28a-let-7 axis, NF-κB signaling, Wtn/β-catenin signaling | Tumor progression inhibition [149] | upregulated: blood plasma-derived EVs [143] |
| miR-24-3p | eNOS, GATA2, PAK4 | Tumor suppression, angiogenesis regulation and cell protection against apoptosis [150] | upregulated: blood plasma-derived EVs [151] |
| miR-26a-5p | ADAM17, Bid, FAF1, SERBP1, Wnt5; TGF-β signaling | Osteogenic differentiation and cell proliferation regulation [152] | upregulated: serum [153] |
| miR-27a-3p | AQP11, BTG2 | Tumor suppression [154], protection against blood-brain barrier and brain injury after brain hemorrhage [155] | downregulated: serum-derived exosomes [156] |
| miR-29b-3p | C1QTNF6/AMPK signaling | Modulation of inflammatory response [157] | downregulated: CSF exosomes and blood plasma-derived EVs [147,148] |
| miR-34a | AXIN2, BCL2, BIRC5, CD44, DGKζ, E2F3, MDMX, MET; MYCN, NOTCH1, NANOG, PD-L1, SIRT1, SNAI1, SOX2; cyclins, cyclin-dependent kinases, TGF-β1/Smad signaling | Cell proliferation, apoptosis, autophagy and cellular senescence regulation [158,159], matrix proteins deposition [159] | downregulated: ALS iMNs- derived exosomes and CSF [160] |
| miR-100-5p | ANKAR, AP1AR, EPDR1, ICK, NR1I3, SMARCA5, ST6GALNAC4, TMPRSS13, TTC39A; mTOR signaling | Cell survival regulation (e.g, apoptosis) [161], and autophagy [162,163] | downregulated: blood plasma- derived EVs [143] |
| miR-124–3p | CDK6, EfnB1, PTBP1, REST, SCP1, Sox9; NeuroD1; | Synaptic connectivity and plasticity regulation [164] | upregulated: CSF exosomes [147] |
| miR-127-3p | BCCIP, BOLA1, FAM27D1, KCNK2, LOC100134822, MTCP1, PSD95, RBPMS, SIRT3, SLC25A2, TPTE2, ZNF3, NeuroD1, NR2A-subunit | Neurogenesis, synapse formation and motor neuron integrity maintenance, mitophagy, ROS and misfolded proteins accumulation [165] | upregulated: serum [153]; downregulated: blood plasma [143,153] |
| miR-144-3p | ABCA1, CCNT2, FoxO1, FST, GABRA1, HGF, IGIP, NFE2L2, ST3GAL6, UBE2D1, UBR3 | Adipogenesis regulation, metastasis and cell proliferation inhibition [166] | upregulated: blood plasma- derived EVs [143] |
| miR-146a-5p | IRAK-1; NF-κB signaling | Immune cell activity, hematopoiesis and malignant transformation regulation [167] | upregulated: blood plasma-derived EVs [148,168] |
| miR-149-3p | AKT2 | Cell proliferation inhibition in cancer [169] | upregulated in blood plasma-derived EVs [151] |
| miR-150-3p | CASP2, SP1 | Neuroprotection of neural stem cells exosomes after ischemic insult and cell proliferation inhibition [170,171] | downregulated: blood plasma-derived EVs [151] |
| miR-151a-3p | SOCS5, SP3, YTHDF3; JAK2/STAT3 signaling | Tumorigenic inhibitor [172] | upregulated: blood plasma-derived EVs [148,168] |
| miR-151a-5p | AGMAT, CYTB, SMARCA5 | Cellular ATP production regulation [173] | upregulated: blood plasma-derived EVs [148,168] |
| miR-181a-1-5p | Kras, NRAS, VCAM-1, ZNF780A, ZNF780B, ZNF204P, ZNF439, ZNF527, ZNF559, ZNF594, ZNF781, ZNF844 | Tumorigenic suppressor, immune response regulation and cell proliferation [174,175,176] | downregulated: blood plasma- derived EVs [143] |
| miR-181a-2-5p | STAT3, TGFβR3 | Tumorigenic suppressor [177] | downregulated: blood plasma- derived EVs [143] |
| miR-181b-1-5p | BAZ2B, NOVA1, TGFβ1, ZNF780A, ZNF780B, ZNF439, ZNF527, ZNF559, ZNF594, ZNF781, ZNF844; MEK/ERK/p21 pathway | Cell proliferation, invasion and metastasis in cancer [178], apoptosis inhibition [179] and autophagy [180] | downregulated: blood plasma- derived EVs [143] |
| miR-181b-2-5p | BCL2, TIMP3; annexin A2 | Cell migratory proteins modulation [181] and chemosensitivity in cancer cells [182] | downregulated: blood plasma- derived EVs [143] |
| miR-183-5p | AKAP12, CCDC121, DHRSX, FKSG83, GNG5, NUDT4, PFN2, PDCD4, PSEN2, RIPK3, SLAIN1, XPNPEP3, | Neuron protection against motor cell death in ALS (under stress conditions) [183] | upregulated: blood plasma- derived EVs [143] |
| miR-194-5p | HIF-1, NR2F2, NR2F6, PAK2; MAPK1/PTEN/AKT signaling | Tumorigenic inhibition [184] | upregulated: ALS iMNs- derived exosomes [143,160] |
| miR-197-5p | HSPA5, KIAA0101, TIMP2/3; AKT/mTOR axis signaling | Tumor suppression, cell proliferation [185], autophagy regulation [186], angiogenesis promotion [187]. Recognized biomarker for myocardial fibrosis and heart failure [188] | downregulated: postmortem frontal cortex and spinal cord [153] |
| miR-199a-1-3p | BCAR3, CDNF, DNMT3a, FABP12, HVCN1, KLHL3, RAP2a; SERPINE2SRR, TMEM161B, TSGA10, WFDC8 | Growth and angiogenesis inhibition in tumors [189] | downregulated: blood plasma- derived EVs [143] |
| miR-199a-2-3p | caveolin-2, Ppargc1a, Sirt1 | Regulation of cell proliferation and survival [190] | downregulated: blood plasma- derived EVs [143] |
| miR-199a-3p | CCND1, CD44, c-MYC, DNMT3a, EGFR, ETNK1, YAP1; mTOR | Cell proliferation regulation and apoptosis induction [190] | upregulated: blood plasma-derived EVs [148,168] |
| miR-199b-3p | CDNF, BCAR3, FABP12, HVCN1, KLHL3, SERPINE2 TSGA10, SRR, TMEM161B, WFDC8; Phospholipase Cε | Tumor suppression [191] | downregulated: blood plasma- derived EVs [143] |
| miR-199a-5p | DDK1, ITGA3, WTN2; CREB/BDNF signaling, NF-κB signaling | Tumorigenic inhibitor [192] | upregulated: blood plasma-derived EVs [148,168] |
| miR-298 | JMJD6 | Tumor suppression, cell proliferation and metastasis inhibition [193] | downregulated: postmortem frontal cortex, spinal cord and serum [153] |
| miR-335 | ROCK1, survivin | Tumor suppression [194] | downregulated: ALS iMNs- derived exosomes [160] |
| miR-338-3p | C5orf47, C6orf141, DGKB, IDNK PREX2, IZUMO3, PIM1, ROBO1, SP3, TAX1BP3,ZNF141, ZNF208 | Tumor suppression; cell proliferation, migration and invasion inhibition [195,196] | downregulated: blood plasma- derived EVs [143] |
| miR-342-3p | ATF3, FOXQ1, RAP2B, MAP1LC3B; HDAC7/PTEN axis signaling, RhoC GTPase | Prion-based neurodegeneration and intracellular motor proteins, axon guidance, cell proliferation and apoptosis regulation [197,198], tumor suppression, autophagy and reduction of cell stemness [199] | upregulated: postmortem frontal cortex, spinal cord and serum- derived EVs [153] |
| miR-363-3p | CD69, DCAF6, FAM24A, FBXW7, FNIP1, MAN2A1, FBXW7, KLF4, PTEN; PI3K/AKT signaling | Osteogenic differentiation [200] | upregulated: blood plasma- derived EVs [143] |
| miR-371a-5p | BCL2; BECN1, SOX2 | Tumor suppression; cell proliferation, migration and autophagy [201] | upregulated in blood plasma-derived EVs [151] |
| miR-450a-2-3p | FOXP3, IGF2, MAPK1, KSR2 | Tumorigenic inhibition [202] | upregulated: postmortem spinal cord and serum [153] |
| miR-494-3p | SEMA3A | Axonal maintenance [negative regulation of semaphorin 3A (SEM3A)] [203] | downregulated: astrocyte-derived EVs and in cortico-spinal tract tissue [203] |
| miR-502-5p | SP1 | Tumor suppression, regulation of cell proliferation and migration [204] | downregulated: postmortem frontal cortex and spinal cord [153] |
| miR-512-5p | ETS1, hTERT, SOD2; Wnt/β-catenin signaling | Tumor suppression, and apoptosis induction [205] | upregulated: postmortem frontal cortex [153] |
| miR-520f-3p | C2orf69, NDST4, SOX9, Wnt signaling | Tumor suppression [206] | upregulated: serum [153] |
| miR-532-3p | C13orf34, C22orf46, DNAL4, ENSA, FOXP3, KLHL12; OPHN1, RPRML, RPS3, ZNF514; β-catenin | Cell proliferation, metastasis inhibition and apoptosis enhancing [207] | upregulated: blood plasma-derived EVs [143] |
| miR-551b-3p | H6PD, Cyclin D1, TRIM31/Akt signaling | Tumor inhibition [208] | upregulated: serum [153] |
| miR-549a | yet to be studied | Angiogenesis and metastasis induction [209] | downregulated: postmortem frontal cortex and spinal cord [153] |
| miR-587 | RPSA | Tumor suppression [210] | downregulated: serum-derived EVs [153] |
| miR-625-3p | GABBR2, SCAI | Tumor suppression [158]. Cancer cells migration and invasion inhibition [211] | downregulated: ALS iMNs-derived exosomes and CSF [160] |
| miR-629-5p | AKAP13, CAV1, SFRP2 | Tumor cell growth regulation [212] | upregulated: ALS iMNs- derived exosomes [143,160] |
| miR-634 | HSPA2; mTOR signaling | Tumor suppression and apoptosis enhancing [213,214] | downregulated: blood plasma-derived EVs [151] |
| miR-664a-5p | AC093802.1, ANKRD36; CCNDBP1, DNASE2, FBXO17, HMGA2, IDH2, PTCD3, SEPT7, ZNF256, ZNF772 | Osteogenic differentiation, controlled apoptosis [215] and neuronal differentiation [216] | downregulated: blood plasma- derived EVs [143] |
| miR-766-3p | NF-κB signaling, TGFBI signaling | Inhibition of inflammatory responses [217] and apoptosis promotion in cancer [218] | downregulated: serum-derived EVs [153] |
| miR-877-5p | FOXM1 | Tumor suppression, cell proliferation, migration and invasion reduction [219] | downregulated: serum-derived EVs [153] |
| miR-939-5p | ARHGAP4, HIF-1 alpha, IGF-1R; PI3K/Akt signaling | Cell migration and invasion in certain types of cancer [220] | upregulated: blood plasma-derived EVs [151] |
| miR-1207-5p | CX3CR1; NF-κB signaling, SARS-CoV-2 RNA | inflammatory response regulation [221] | upregulated: blood plasma-derived EVs [151] |
| miR-1246 | CDR1as, DNAH, FAM53C, FAM169B, GSG1L, KIAA1370, LIG4; NFE2L3, NR2F2, SGOL1, WDR77, ZNF23, ZNF267; NHEJ signaling | Modulation of DNA damage following ionizing radiation exposure [222,223] | upregulated: blood plasma-derived EVs [151] |
| miR-1254 | Smurf1; PIK/Akt signaling, | Cell proliferation, migration and invasion inhibition [224] | downregulated: postmortem frontal cortex, spinal cord and serum [153] |
| miR-1255a | SMAD4; TGF-β signaling | Related with breast cancer malignant phenotype and downstream effector of TGF-β [225] | upregulated in serum [153] |
| miR-1260b | C2orf48, CASP8, CTAGE1, GOLGA8A, MED13L, PABPN1, USP48, ZNF256, ZNF594, ZNF788; MAPK pathway | Tumorigenesis promotion [226] | downregulated: blood plasma-derived EVs [143] |
| miR-1262 | SCL2A1, ULK1 | Tumor suppression [227] | upregulated in serum [153] |
| miR-1268a | ABCC1 | Mediation of temozolomide resistance in glioblastoma [228] | downregulated: blood plasma-derived EVs [151] |
| miR-1268b | AKT, BCL2, ERBB2, PI3KCA, PI3K-AKT signaling | Apoptosis inhibition [229] | Upregulated: serum [153] |
| miR-1285-5p | CDH1, Smad4, TMEM194A | Cell proliferation regulation [230] | upregulated: postmortem frontal cortex [153] |
| miR-1290 | AKAP7, CDR1as, FAM19A5, HIGD2A, OGN MYO10, OSBPL6, RP11-1167A19.2, SGOL1, WDR77 | Cell proliferation, migration and invasion regulation in cancer [231] | downregulated: blood plasma-derived EVs [143] |
| miR-1913 | not yet studied, but 732 predicted targets in [232] | Potential non-invasive biomarker for prostate cancer [233] | downregulated: blood plasma-derived EVs [151] |
| miR-2861 | STAT3, MMP2, EGFR/AKT2/CCND1 signaling | Tumor suppression, cell proliferation regulation and apoptosis [234] | downregulated: blood plasma-derived EVs [151] |
| miR-3176 | AR, PTEN | Promotion of tumorigenesis and tumor progression [235] | downregulated: blood plasma-derived EVs [151] |
| miR-3177-3p | not yet studied, but 65 predicted targets in [236] | to be studied | downregulated: blood plasma-derived EVs [151] |
| miR-3605-5p | SCABR2 | Adipocyte lipolysis regulation [237] | downregulated: blood plasma-derived EVs [151] |
| miR-3619-3p | Wnt/β-catenin signaling | Cell migration and invasion promotion [238] | upregulated: blood plasma-derived EVs [151] |
| miR-3911 | not yet studied | Possible sALS biomarker [139] | downregulated: blood plasma-derived EVs [151] |
| miR-3940-3p | BIRC5, IL-2Ry, KCNA5, Integrin α6 | Regulation of maternal insulin resistance, T cell activity promotion and metastasis inhibition in cancers [239] | downregulated: blood plasma-derived EVs [151] |
| miR-4286 | APLN, C15orf34, CBX2, FAM222B, HKDC1, INPP4A, ZFP36L1, PARVG, PRX PTEN, RNF43, SALL1; TMSB4X, JAK2/STAT3 signaling, PI3K/Akt signaling, TGF-ß/TGF-ß1/Smad signaling | Cell proliferation, apoptosis and inflammatory response modulation [240,241,242,243] | downregulated: blood plasma-derived EVs [151] |
| miR-4298 | SOD2, TGIF2 | Cell proliferation, migration and invasion of cancer cells [244] | upregulated: blood plasma-derived EVs [151] |
| miR-4443 | INPP4A, METLL3, TRIM14; JAK2/STAT3 signaling, NF-κB signaling, Ras signaling, TGF-β1 signaling | Metastasis and energy metabolism suppression [245] | downregulated: postmortem frontal cortex and spinal cord [153] |
| miR-4454 | ABHD2/ NUDT21, Vps4a, Rab27A; GNL3L/ NF-κB signaling; TGF-β/MAPK pathway | Insulin signaling [246], metastasis progression in cancer [247,248] and apoptosis regulation [249] | downregulated:CSF exosomes and blood plasma-derived EVs [143,147,148]; upregulated: serum [153] |
| miR-4505 | HSPA12B; galectin-9 | Tumorigenesis [[251[M1] ] | upregulated: blood plasma-derived EVs [151] |
| miR-4507 | TP53; PI3K-AKT signaling | Cell proliferation and migration in lung cancer [252] | downregulated: blood plasma-derived EVs [151] |
| miR-4508 | ABL1, ASB6, CAPN15, HOOK3, IRAK3, KIAA0754, LEPROTL1, PEX26, RGS6, U2AF2,VAV3,YES1 | Immune response regulation, phagocytosis and cellular protein modification [253] | downregulated: blood plasma-derived EVs [151] |
| miR-4646-5p | ABHD16A; PHD3; PHD3/HIF1A signaling | Ubiquitination and cell proliferation and invasion regulation [254,255] | downregulated: blood plasma-derived EVs [151] |
| miR-4674 | p38k | Angiogenesis regulation [256] | downregulated: blood plasma-derived EVs [151] |
| miR-4687-5p | ATP10D, THRSP | metastasis and cancer progression [257] | downregulated: blood plasma-derived EVs [151] |
| miR-4688 | not yet studied | miRNA sponge and cancer progression [258] | upregulated blood plasma-derived EVs [151] |
| miR-4700-5p | not yet studied | not yet studied | upregulated blood plasma-derived EVs [151] |
| miR-4736 | AR | Inflammatory response regulation [259] | upregulated blood plasma-derived EVs [151] |
| miR-4739 | BMP-7; ITGA10/PI3K signaling | Apoptosis and differentiation regulation [260] | upregulated blood plasma-derived EVs [151] |
| miR-4745-5p | SIRT6/PCSk9 signaling | sensibility to anesthetics regulation [261] | upregulated blood plasma-derived EVs [151] |
| miR-4788 | not yet studied, but 29 predicted targets in [262] | Polycystic ovary development, nervous system development, neurotransmitter levels regulation and transport and synapsis [263] | downregulated: blood plasma-derived EVs [151] |
| miR-7641-1 | BCL2, CAS9, C9orf153, PARP, RAB7L1, RPS16, TMEM156, TMPRSS11BNL, TNFSF10 | Oncogenic miRNA, cell proliferation, migration and invasion regulation [264] and apoptotic signaling in cancer [265] | downregulated: blood plasma- derived EVs [143] |
| miR-7975 | C9orf41, GDNF | Glutamatergic neurotransmission regulation [266] | upregulated: serum of sALS patients [153] |
| miR-7977 | CD84, MRPS12, MRPL27, MUC19, TRAPPC2, SIRT3, Hyppo-YAP signaling | Hematopoiesis regulation [267], oxidative stress and insulin resistance [268] | downregulated: blood plasma-derived EVs [143] |
| let-7c-5p | ARID3B, C14orf28, DNA2, FIGN, HMGA2, LIN28B, TRIM71 SMARCAD1; CTHRC1/AKT/ERK signaling | Microglia activation inhibition. Cell migration and proliferation Inhibition and apoptosis enhancement [269] | downregulated: blood plasma-derived EVs [143] |
Table 2.
Protein content in ALS-derived extracellular vesicles.
| Protein content | Biological function | Vesicle/Sample type | Main results |
| BLMH | Enzyme with proteolytic activity. Involved in release of inflammatory chemokines and in wound healing [297] | EVs from CSF of ALS patients (C9orf72 mutation) | Downregulation in ALS patients CSF-derived EVs |
| CD163, FOXP3, IL2RA, MRC1 | Anti-inflammatory transcripts [298] | Treg-derived EVs from spinal cord from SOD1G93A mice model and iPSC-derived from myeloid cells | - Upregulation in spleen-derived myeloid cells after Treg- derived EVs treatment [299]; - Intranasal administration of enriched Treg EVs slowed disease progression, increased survival, and modulated inflammation within the SOD1G93A mice spinal cord [299] |
| CD177, CHMP4B, CSPG5, DYNC1I2, IGHV3-43, LBP, RPS29, S100A9, SAA1, SCAMP4, SCN2B, SLC16A1, STAU1, FXYD6, DYNC1I1, DHX30 | Involvement in stress granule dynamics [292] | MCEVs from ALS patients’ postmortem motor cortex tissue | - 12 RNA-binding proteins only found in MCEVs from ALS patients (mainly downregulated) [292]; - 4 proteins significantly upregulated in MCEVs from ALS patients (DYNC1I1, DHX30, FXYD6, STAU1) [292] |
| CHIT1 | Cleavage of chitin (protein found in cell walls of various pathogens). Expressed during the later stages of macrophage differentiation. Important in inflammation and tissue remodeling. In the ALS context, plays a role in the feed-forward loop that maintains inflammation [300] | EVs from ALS patients’ CSF | Upregulation in ALS patients CSF-derived EVs [294] |
| CUEDC2 | Regulates ubiquitin-proteasome pathways and inflammatory response [301] | Exos from sALS patients’ CSF | Only expressed in ALS group [302] |
| FUS and pFUS | DNA repair, RNA splicing, dendritic RNA transport, miRNA function and biogenesis [303] | MVs and Exos from sALS patients’ plasma | Protein levels are higher in ALS patients’ plasma-derived MVs than Exos [81] |
| HSP90 | Chaperone protein involved in protein folding [304] | EVs from ALS patients and symptomatic SOD1G93A and TDP-43Q331K ALS mice models plasma | Downregulation in EVs from sALS patients [293] |
| JAM-A | Regulation of several processes including paracellular permeability, platelet activation, angiogenesis and the modulation of junctional tightness in the blood brain barrier (BBB) [305] | EVs from ALS patients CSF | Downregulation in ALS patients CSF-derived EVs [294] |
| IL-6, iNOS, IL-1b, IFN-y | Pro-inflammatory cytokines (IL-1b, IFN-y, IL-6) and enzyme (iNOS) produced in response to cytokines [306] | Treg-derived EVs from spinal cord from SOD1G93A mice model and iPSC-derived from myeloid cells | Downregulation in spleen-derived myeloid cells after Treg-derived EVs treatment [299] |
| MB | Oxygen-binding molecule expressed in skeletal and cardiac muscle tissue [307,308] | EVs from ALS patients CSF | Downregulation in ALS patients CSF-derived EVs [294] |
| mfSOD1 | Antioxidant enzyme, protects cells from ROS [309] | Vacuoles and EVs from spinal cord samples of SOD1G93A mice model | Accumulation of mfSOD1-vacuoles in degenerating MNs, released into the extracellular space in the form of extracellular vesicles [130] |
| NIR | Translocates from the nucleolus to the nucleoplasm in response to the nucleolar stress [310] | Exos from sALS patients CSF and anterior horn postmortem tissue sections | Upregulation in sALS patients CSF-derived exos [311] |
| Nrf2 | Antioxidant factor [312] | EVs from spinal cord tissue of SOD1G93A mice model | Upregulation after exposure to MSCs-derived EVs, with consequent reduction of ROS [313] |
| pCREB | Involved in the synthesis of proteins required for LTP [314] | Exos from SOD1G93A mice model SVZ-derived NSCs, differentiated into G93A neuronal cells | Downregulation in G93A cells, normalized with ADSC-exos treatment [315] |
| PGC-1α | Involved in the regulation cell metabolism [316] | Exos from SOD1G93A mice model SVZ-derived NSCs, differentiated into G93A neuronal cells | Downregulation levels in G93A cells, normalized with ADSC-exos treatment [315] |
| Phenylalanine | Precursor for tyrosine [317], the monoamine neurotransmitters dopamine, norepinephrine, and epinephrine | lEVs and sEVs from sALS patients’ plasma | Downregulation in EVs from sALS patients [318] |
| pMLKL | Effector of necroptotic pathways [319] | Vacuoles and EVs from spinal cord samples of SOD1G93A mice model | Upregulation in vacuoles of degenerating MNs (necroptotic pathway activation/phenotype 3) [130] |
| PPIA | Ubiquitous protein involved in protein folding, transport and signaling (e.g apoptosis, inflammation, etc) [320] | EVs from ALS patients and symptomatic SOD1G93A and TDP-43Q331K ALS mice models plasma | Protein levels and EV size distribution distinguish fast and slow ALS disease progression [293] |
| pNFH | Chaperone involved in TDP-43 trafficking and function [321] | EVs from ALS patients and symptomatic SOD1G93A and TDP-43Q331K ALS mice models plasma | Upregulation in EVs from sALS patients [293] |
| SOD1 | Binds copper and zinc ions, responsible for freeing superoxide radicals from cells [309] | Exos from SOD1G93A mice model SVZ-derived NSCs, differentiated into G93A neuronal cells | ADSC-exos alleviated aggregation of cytosolic SOD1 in G93A ALS mice isolated neuronal cells [315] |
| MVs and Exos from sALS patients’ plasma | Protein levels are higher in ALS patients’ plasma-derived Exos than MVs [81] | ||
| TDP-43 and pTDP-43 | RNA regulation (transcriptional regulation, alternative splicing and mRNA stabilization) [322] | Exos from ALS patients CSF | TDP-43 accumulation [323] |
| MVs and Exos from sALS patients’ plasma | Protein levels are higher in ALS patients’ plasma-derived MVs than Exos [81] | ||
| TNF-R2 | Proinflammatory proteins activation [324] | EVs from ALS patients’ CSF | Upregulation in serum of ALS patients. TNF-R2 knocking down in ALS mouse model results in motor neuron protection [294] |
| UBA1 | Involved in ubiquitination of proteins for degradation by the UPS [325] | EVs from ALS patients’ CSF (C9orf72 mutation) | Upregulation in ALS patients’ CSF-derived EVs [294] |
Abbreviations: ALS (amyotrophic lateral sclerosis); ADSC (exo-adipose-derived stem cell exosomes); BLMH (pentameric proteasome-like protein Bleomycin hydrolase); CD206 (MRC1; mannose receptor C-Type 1); CHIT1 (chitotriosidase/chitinase 1); CHMP4B (charged multivesicular body protein 4B); CSPG5 (chondroitin sulfate proteoglycan 5); CSF (cerebrospinal fluid); CSF-EVs (CSF-derived extracellular vesicles); CUEDC2 (CUE domain-containing protein 2); DHX30 (DExH-box helicase 30); DYNC1I1 (dynein cytoplasmic 1 intermediate chain 1); DYNC1I2 (dynein cytoplasmic 1 intermediate chain 2); Exos (exosomes); EVs (extracellular vesicles); FOXP3 (forkhead box P3); FUS (fused in sarcoma); p-FUS (phosphorylated fused in sarcoma); FXYD6 (FXYD domain-containing ion transport regulator 6); lEVs (large extracellular vesicles); IFN-y (interferon-γ); IL-1β (interleukin-1β); IL-2RA (interleukin-2 receptor subunit α); IL-6 (interleukin-6); IGHV3-43 (immunoglobulin heavy variable 3-43); IGHV3-43 (immunoglobulin heavy variable 3-43); iNOS (inducible nitric oxide synthase); JAM-A (junctional adhesion molecule-A); LBP (lipopolysaccharide-binding protein); MB (myoglobin); MCEVs (motor cortex extracellular vesicles); mfSOD1 (misfolded protein SOD1); MNs (motor neurons); MRC1 (mannose receptor C-type 1/CD206); MSCs (mesenchymal stem cells); MVs (microvesicles); NIR (INHAT repressor); Nrf2 (nuclear factor E2-related factor 2); NSCs (neuronal stem cells); pCREB (phosphorylated cAMP response element-binding protein); PGC-1α (peroxisome proliferator-activated receptor-γ coactivator); pMLKL (phosphorylated mixed lineage kinase domain-like protein); pNFH (phosphorylated neurofilament protein heavy unit); PPIA (cyclophilin A); ROS (reactive oxygen species); RPS29 (ribosomal protein S29); sALS (sporadic amyotrophic lateral sclerosis); sEVs (small extracellular vesicles); SCAMP4 (secretory carrier membrane protein 4); SCN2B (sodium channel subunit β-2); SLC16A1 (solute carrier family 16 member 1); SOD1 (superoxide dismutase 1); STAU1 (staufen double-stranded RNA binding protein 1); SVZ (subventricular zone); S100A9 (S100 calcium-binding protein A9); TNF-R2 (tumor necrosis factor receptor 2); TDP-43 (TAR DNA-binding protein 43); Treg (regulatory T cells); UBA1 (Ubiquitin-activating enzyme E); UPS (Ubiquitin-Proteasome System).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



