Submitted:
09 June 2023
Posted:
09 June 2023
You are already at the latest version
Abstract
The size of seeds is particularly important for agricultural development, as it is a key trait that determines yield. It is controlled by the coordinated development of the integument, endosperm, and embryo. Large seeds are one of the important ways to improve the ultimate “sink strength” of crops, providing more nutrients for early plant growth and showing certain tolerance to abiotic stresses. There are several pathways for regulating plant seed size, including the HAIKU (IKU) pathway, ubiquitin-proteasome pathway, G (Guanosine triphosphate) protein regulatory pathway, mitogen-activated protein kinase (MAPK) pathway, transcriptional regulators pathway, phytohormone regulatory pathways including auxin, brassinosteroid (BR), gibberellin (GA), jasmonic acid (JA), cytokinin (CK), Abscisic acid (ABA), and MicroRNAs (miRNAs) regulatory pathway. This article summarized the seed size regulatory network and prospected ways to improve yield. We expect to provide valuable reference to researcher in the related field.
Keywords:
seed size
; IKU
; ubiquitin
; G protein
; MAPK
; transcription factor
; phytohormone
; miRNA
1. Introduction
Crops such as rice, corn, and wheat urgently need to increase their yield [1]. Seed size and number determine the final yield, and seed shape also plays an important role in seed development and weight [2]. Seed development is coordinated by the embryo, endosperm, and integument [3] and begins with double fertilization, one sperm cell fuses with the egg cell to produce a diploid embryo, and the other sperm cell fuses with the central cell to form a triploid endosperm [4]. The embryo and endosperm develop within the ovule, and the ovule integument eventually becomes the seed coat of the seed [5]. The endosperm plays a major role in regulating seed size [6]. Starch synthesis during endosperm development in rice affects the size of the grain [7]. Seed coat development largely determines the final size and shape of the seed [8].
Plants regulate seed size through multiple signaling pathways [9]. Transcription factors regulate cell growth in the ovule, resulting in seed growth. Plant hormones not only regulate plant growth and development, but also regulate seed size [10]. In rice, grain size is the main determinant of yield, and the spikelet hull is a key determinant of grain size [11]. Several genes that regulate seed size have been identified in different crops through various pathways, including the IKU pathway, ubiquitin-proteasome pathway, G protein regulatory pathway, MAPK pathway, transcriptional regulators pathway, phytohormone regulatory pathways and miRNA regulatory pathway. This article reviews the research results on the mechanisms that regulate seed size in plants such as Arabidopsis thaliana L., Triticum aestivum L., and Oryza sativa L..
2. HAIKU (IKU) pathway
The IKU pathway genes include IKU1, IKU2, MINISEED3 (MINI3), and SHORT HYPOCOTYL UNDER BLUE1 (SHB1). Wang et al. cloned the IKU1 gene (At2g35230) in Arabidopsis, and the iku1 mutant resulted in reduced endosperm growth and small seeds [12]. IKU1 encoded a protein containing a VQ motif, which was a plant-specific motif. IKU1 was expressed in early endosperm and the central cell, and the VQ motif was an essential regulatory element for seed size regulation [12]. IKU2 encoded a leucine-rich repeat (LRR) kinase (At3g19700), and overexpression of IKU2 in Arabidopsis led to an increase in seed size, weight, and oil content [13]. MINI3 is a WRKY10 transcription factor [14]. Mutants of iku2 and mini3 had a small-seeded phenotype, which was associated with reduced growth of endosperm and early cellularization [14], similar to the iku1 phenotype [12]. SHB1 is a gene that affects endosperm development, and normal expression of IKU2 and MINI3 requires SHB1 [15]. The shb1-D (a gain-of-function overexpression allele) increased seed size, while shb1 (a loss-of-function allele) decreased seed size. IKU1 interacted with MINI3, and SHB1 bound to the promoters of IKU2 and MINI3 to promote endosperm growth during early seed development [15].
3. Ubiquitin-proteasome pathway
Grain width 2 (GW2), a RING-type E3 ubiquitin ligase, regulated seed development by affecting cell growth [16,17]. GhGW2-2D in cotton (Gossypium spp.) and AtDA2 in Arabidopsis share similar functional domains and highly conserved sequences [16]. Overexpression of GhGW2-2D in Arabidopsis significantly reduced seed and seedling size [16]. The ubiquitin receptor DA1 is a negative regulator of seed size [18]. In Arabidopsis, downregulation of AtDA1 increased seeds and organs size by increasing cell division in the integuments [18]. Overexpression of AtDA1R358K in Brassica napus L. to downregulate BnDA1, the transgenic plants significantly increased biomass and size of the seeds, cotyledons, leaves, flowers, and siliques [18]. Liu et al. identified TaDA1 as a negative regulator of grain size in wheat (Triticum aestivum) [19]. Overexpression of TaDA1 reduced grain size and weight, while downregulation of TaDA1 using RNA interference (RNAi) had the opposite effect [19]. NGATHA-like protein (NGAL2), a transcriptional repressor encoded by the inhibitor of da1-1 (SOD7), regulated seed size by limiting cell proliferation in the integuments and developing seeds [20]. Overexpression of SOD7 significantly reduced seed size [20]. SOD7 and DEVELOPMENT-RELATED PcG TARGET IN THE APEX 4 (DPA4/NGAL3) interacted with the seed size regulator KLUH (KLU) to regulate seed growth, independently of DA1 [20]. GhDA1-1A in cotton and AtDA1 in Arabidopsis share highly conserved sequences and functional domains [21]. Overexpression of GhDA1-1AR301K in Arabidopsis significantly increased seed size and weight [21]. DA2 in Arabidopsis encoded a RING-type protein with E3 ubiquitin ligase activity, which regulated seed size by limiting cell proliferation in the maternal integuments [22]. The da2-1 mutant produced larger seeds, while overexpression of DA2 reduced seed size [22]. DA1 and its homologous genes DAR1 and DAR2 were redundantly required for endoreduplication during organs development [23]. The triple mutant da1-ko1 dar1-1 dar2-1 showed very small plants and leaves, but flowers and seeds were still larger compared with wild type (WT) [23].
UBIQUITIN-SPECIFIC PROTEASE15 (UBP15), encoded by SUPPRESSOR2 OF DA1 (SOD2), regulated seed size by promoting cell proliferation in the integuments of ovules and developing seeds [24]. The sod2/ubp15 mutants formed small seeds, while overexpression of UBP15 increased seed size [24]. UBP15 and DA1 had antagonistic effects in a common pathway that affected seed size, but this effect was independent of DA2 [24]. The dominant large-grain mutant, large grain1-D (lg1-D), had a 30.8% increase in seed width and a 34.5% increase in 1,000-grain weight compared to the WT [25]. Map-based cloning showed that LG1 encoded a constitutively expressed OsUBP15 with deubiquitinated activity in vitro [25]. Loss-of-function and down-regulation of OsUBP15 resulted in narrower and smaller grains than the control [25]. OsDA1 directly interacted with OsUBP15 [25]. OsUBP15 is a positive regulator of rice grain width and size. These results indicate that the ubiquitin-proteasome pathway plays an important regulatory role in seed size.
4. G (Guanosine triphosphate) protein regulatory pathway
There are three types of Gγ proteins in rice, DEP1, GGC2, and GS3, which antagonistically regulate grain size. DEP1 and GGC2, individually or in combination, increased grain length when complexed with Gβ [26]. GS3 had no direct effect on grain size by itself, but reduced grain length by competitively interacting with Gβ [26]. The two Gγ proteins, DEP1 and GS3, antagonistically regulated grain yield. Gene editing of DEP1 significantly increased the number of grains per panicle, while gene editing of GS3 reduced the number of grains per panicle but significantly increased grain length [27]. Yang et al. demonstrated that Chang Li Geng 1 (CLG1), encoding an E3 ligase, regulated grain size by targeting the negative regulator of grain length factor GS3 for degradation [28]. Overexpression of CLG1 led to an increase in grain length, and CLG1 interacted with GS3 and ubiquitinated it, which then underwent degradation via the endosome degradation pathway, resulting in an increase in grain size [28]. Gγ protein AGG3, which was localized to the plasma membrane, promoted seed and organ growth by increasing the proliferation growth period in Arabidopsis [29,30]. The homologous genes of AGG3 in rice (GS3 and DEP1/qPE9-1) had been identified as important quantitative trait loci for seed size and yield [29]. GS3 and DEP1 affected seed and organ growth by inhibiting cell proliferation [29]. Overexpression of AGG3 significantly increased the length of siliques, the number and size of seeds per silique in Arabidopsis [30]. In Camelina sativa, constitutive or specific expression of AGG3 in seed tissue increased seed size, seed mass, and seed number by 15%-40%, effectively increasing oil yield per plant [31].
The qLGY3 locus, which encoded a MADS-domain transcription factor OsMADS1, was associated with the quantitative and qualitative traits of rice grain yield [32]. OsMADS1 was a downstream effector of the G-protein βγ dimers and its alternatively spliced isoform OsMADS1lgy3 was related to the formation of long and slender grains, which could increase rice yield and quality [32]. The Gγ subunits GS3 and DEP1 directly interacted with the conserved keratin-like domain of MADS transcription factors and acted as cofactors to enhance the transcriptional activity of OsMADS1, promoting the coordinated activation of target genes and thereby regulating grain size and shape [32]. WIDE GRAIN 7 (WG7) activated OsMADS1 expression by directly binding to its promoter, increasing the enrichment of histone H3K4me3 in the promoter and ultimately increasing grain size [33].
The rice genome contains a single Gα (RGA1) and Gβ (RGB1) subunit, as well as five Gγ subunits (RGG1, RGG2, GS3, DEP1/qPE9-1, and GGC2) [34]. DEP1/qPE9-1 was an atypical putative Gγ protein that controlled grain size, dense and erect panicles [34]. Zhang et al. showed that rice plants carrying DEP1/qPE9-1 had more endosperm cells per grain than those carrying dep1/qpe9-1 allele [34]. The DEP1/qPE9-1 line also had higher levels of ABA, auxin, and cytokinin in grains. Exogenous application of auxin or cytokinin could enhance starch accumulation and the expression of grain-filling-related enzymes in dep1/qpe9-1 grains, but had no effect on ABA [34]. DEP1/qPE9-1 mainly positively regulated starch accumulation during grain-filling stage through auxin and cytokinin pathways, enhancing the expression of starch biosynthesis genes in the mid to late stage of grain-filling and prolonging the duration of grain filling [34].
RGB1 was a positive regulator of cell proliferation, as evidenced by the shorter stature of d1-5, a mutant lacking the RGB1 gene, in addition to the loss of RGA1 [35]. The number of sterile seeds also increased in the RGB1 knockout lines [35]. Overexpression of RGG2 in Nipponbare (NIP) led to reduced plant height and smaller grain size, as RGG2 was a negative regulator of plant growth and organ size in rice [36]. RGG2 was also involved in GA signaling as measured by the length of the second leaf sheath and GA-induced α-amylase activity, suggesting that RGG2 might regulate grain and organ size through the GA pathway [36].
5. Mitogen-activated protein kinase (MAPK) pathway
MAPK cascades are conserved and involved in plant signal transduction in eukaryotes [37]. The MAPK cascades consist of three functional protein kinases: MAPKs, MAPK kinases (MAPKKs), and MAPK kinase kinases (MAPKKKs) [38]. MPK3, MPK6, and MPK10 form a small group of MPKs family in Arabidopsis [39]. The phosphorylation of MPK3/6 inactivated DA1 and made it unstable to increase the abundance of UBP15, promoting outer integument cell proliferation and increasing seed size [40]. WRKY10/MINI3 is a member of the IKU pathway related to endosperm development, and MPK10 is specifically expressed in the early developmental endosperm, but the expression gradient is opposite. MPK10 and WRKY10 inhibited each other's expression. The inhibitory effect of MPK10 on the expression of WRKY10 and its downstream target genes was achieved by interacting with WRKY10 to inhibit its transcriptional activity [39]. The mpk10 mutant produced large seeds, while WRKY10/MINI3 positively regulated seed growth [39].
Rice OsMKK3 encoded a MAP kinase kinase that controlled grain size and chalkiness by affecting cell proliferation in the hull [41]. OsMKKK70 positively regulated rice grain size, since overexpression of OsMKKK70 resulted in longer seeds compared with WT [42]. The osmkkk62/70 and osmkkk55/62/70 mutants showed significantly smaller seeds than WT, indicating functional redundancy between OsMKKK70 and its homologs OsMKKK62 and OsMKKK55 [42]. OsMKKK70 was an active kinase that interacted with OsMKK4 and promoted OsMAPK6 phosphorylation [42]. Overexpression of constitutively active OsMKK4, OsMAPK6, and OsWRKY53 partially rescued the smaller seed size of the osmkkk62/70 double mutant [42]. OsMKKK70 might regulate rice grain size by activating OsMAPK6, and that OsMKKK70, OsMKK4, OsMAPK6, and OsWRKY53 acted together to control the signal pathway of grain shape [42].
Dwarf and small grain 1 (DSG1) encodes a mitogen-activated protein kinase, OsMAPK6, which plays a critical role in rice grain size through cell proliferation, BR signaling, and homeostasis [43]. OsMAPK6 interacted strongly with OsMKK4, indicating that OsMKK4 was likely to be the upstream MAPK kinase of OsMAPK6 [43]. In rice, loss of function of OsMKKK10 led to small and light grains, short panicles, and semi-dwarf plants, while overexpression of the constitutively active OsMKKK10 (CA-OsMKKK10) resulted in large and heavy grains, long panicles, and tall plants [44]. OsMKKK10 interacted with OsMKK4 and phosphorylated OsMKK4 [44]. OsMKK4A227T encoded by the large11-1D allele had stronger kinase activity than OsMKK4 [44]. Plants overexpressing the constitutively active form of OsMKK4 (OsMKK4-DD) also produced larger grains [44]. The OsMKKK10-OsMKK4-OsMAPK6 signaling pathway positively regulated rice grain size and weight.
GSN1 is a negative regulator of the OsMKKK10-OsMKK4-OsMPK6 cascade that determines panicle architecture [45]. GSN1 encodes the mitogen-activated protein kinase phosphatase OsMKP1, a dual-specificity phosphatase with unknown function [45]. Decreased GSN1 expression led to larger but fewer grains, while overexpression resulted in more but smaller grains [45]. GSN1 directly interacted with OsMPK6 and inactivated it by dephosphorylation [45]. The lectin receptor-like kinase LecRK-VIII.2, which acted upstream of MPK6, was a specific receptor-like kinase that regulated the final yield of Arabidopsis seeds by controlling silique number, seed size, and seed number [46]. Plants with the lecrk-VIII.2 mutation had smaller seeds but more siliques and seeds number, leading to increased yield [46]. However, overexpression of LecRK-VIII.2 resulted in larger seeds but fewer siliques and seeds, resulting in a yield similar to that of WT plants [46].
6. Transcriptional regulators pathway
NAC proteins are one of the largest families of plant-specific transcription factors (TFs) and play important roles in plant growth and development. TaNAC020s were mainly expressed in developing grains [47]. Transgenic rice overexpressing TaNAC020-B exhibited higher starch density and lower amylose contents than those of WT [47]. In wheat (Triticum aestivum L.), TaNAC020s positively regulated starch synthesis and accumulation and were critical regulators of grain size and number [47]. Overexpression of TaNAC100 in wheat suppressed plant height, prolonged the heading date, and promoted seed size and 1,000-grain weight [48]. Transgenic rice overexpressing an OsNAC2 mutant (OErN) had better plant architecture and longer panicles, and produced more grains [49]. In rice, loss of function of OsNAC129 significantly increased grain length, weight, apparent amylose content (AAC), and plant height, while overexpression of OsNAC129 had the opposite effect [50]. The expression of OsNAC129 was induced by ABA and overexpression of OsNAC129 in plants reduced sensitivity to exogenous BR, indicating that OsNAC129 negatively regulated seed development and plant growth and participated in the BR signaling pathway.
WRINKLED1 (WRI1) is an APETALA2 (AP2) transcription factor. Two wheat TaWRI1Ls genes, TaWRI1L1 and TaWRI1L2, were cloned based on the Arabidopsis AtWRI1 sequence [51]. Overexpression of TaWRI1L2 could compensate for the loss of AtWRI1 function in Arabidopsis mutants, restoring seed shape and fatty acid accumulation [51]. Knockout of TaWRI1L2 reduced grain size, 1,000-grain weight, and grain fatty acid synthesis in wheat [51]. In Arabidopsis, overexpression of RAV1, a plant-specific B3 domain and AP2 domain-containing transcription factor, resulted in reduced seed size, weight, and number in a silique [52]. RAV1 repressed the expression of MINI3 and IKU2 by directly bond to their promoters [52].
The Alfin-like (AL) family was a group of small plant-specific transcriptional factors and the seeds of the osal7.1 and osal11 mutants were larger in rice [53]. ATBS-INTERACTING FACTOR 2 (AIF2) is a non-DNA-binding basic-helix-loop-helix (bHLH) transcription factor [54]. AIF2ox plants had fewer siliques and fewer seeds per silique, leading to a significant decrease in total grain weight and yield [54]. Conversely, aif2-1/aif4-1 plants exhibited opposite siliques number and seed phenotypes [54]. The positive regulation factors of seed size, such as SHB1, IKU1, MINI3 were suppressed in AIF2ox [54]. BRASSINAZOLE RESISTANT 1 (BZR1) is one of the important transcription factors that regulates organ size in the BR signaling pathway in Arabidopsis [55]. AIF2 negatively regulated BR-induced BZR1-mediated pollen development and seed formation [54]. Sucrose and BR inhibited AIF2 ectopic accumulation, thereby increasing silique length and seed number [54]. Sucrose and BR induced AIF2 to inhibit positive regulation of pollen production and seed formation in Arabidopsis [54]. Overexpression of ZmBZR1 in Arabidopsis showed phenotypes of enlarged cotyledons, rosette leaves, floral organs, and seed size [55]. The cells in transgenic ZmBZR1 lines were significantly larger in rosette leaves and other organs than those of the WT [55]. ZmBES1/BZR1-5 positively regulated grain size as overexpression of ZmBES1/BZR1-5 significantly increased seed size and weight in Arabidopsis and rice [56]. ZmBES1/BZR1-5 bound to the promoters of AP2/EREBP to inhibit the transcription [56]. ZmBES1/BZR1-5 interacted with casein kinase II subunit β4 (ZmCKIIβ4) and ferredoxin 2 (ZmFdx2) in vitro and in vivo [56].
GATA represented a highly conserved family of transcription factors, and OsGATA8 increased seed size and tolerance to abiotic stresses in Arabidopsis and rice, and also helped to maintain yield under stress, with overexpressing plants yielding about 46% higher than the WT [57]. WRKY6, a WRKY6 family transcription factor, had a high expression level in developing seeds and played an important role in regulating the accumulation of fatty acids (FAs) in Arabidopsis [58]. Mutation of WRKY6 led to significantly increased seed size, accompanied by increased FA content and changes in FA composition [58]. In Arabidopsis, MYB56, an R2R3 MYB transcription factor, positively controlled seed size, since loss-of-function or knock-down of MYB56 yielded smaller seeds, while overexpression of MYB56 produced larger seeds than the WT [59]. Microscopic observation showed that myb56 formed smaller endothelial cells with reduced cell numbers in the outer integument, whereas MYB56 overexpressing lines had expanded endothelial cells and increased cell number in the outer integument [59]. MYB37, a R2R3 MYB subgroup 14 transcription factor, played a positive role in the regulation of seed production [60]. Overexpression of MYB37 delayed the flowering time, as the appearance of flower buds in the OE1 and OE6 transgenic plants was delayed 8 and 12 days, respectively, under long day conditions [60]. However, mature OE1 and OE6 plants exhibited greater growth than the WT plants in the later developmental stages, with significantly higher stem height and weight or bigger biomass. Furthermore, transgenic OE1 and OE6 plants produced more siliques, leading to higher seed production than the WT plants [60].
Auxin Response Factor 12 of maize (ZmARF12), a member transcription factor, which mediated the expression of auxin-responsive genes, regulated seed size development, as ZmARF12 mutant exhibited larger seed size and higher grain weight [61]. Growth-regulating factor (GRF) interacting factor 1(GIF1) is a plant-specific transcriptional cofactor positively regulates the grain size in rice [62]. Overexpression and functional knock-out using CRISPR/Cas9 strategies showed that OsGIF1 not only positively regulated the size of rice leaves, stems, and grains, but also affected rice reproduction [62]. Grain length and width 2 (GLW2) encoded growth-regulating factor 4 (OsGRF4), which was regulated by miR396c in vivo [63]. Mutation in OsGRF4 disrupted the targeted regulation of OsGRF4 by OsmiR396, resulting in larger seeds and increased grain yield [63]. OsGIF1 directly interacted with OsGRF4 and increasing its expression to improve seed size [63].
7. Phytohormone regulatory pathways
7.1. Auxin regulatory pathway
Auxin is an essential phytohormone in plant development [64], mainly involved in regulating different signaling pathways in embryos, endosperm, and seed coat, leading to various changes in seed size or weight [9].
Rice YUCCA (YUC) flavin-containing monooxygenase encoding gene OsYUC11 was a key factor for auxin biosynthesis in endosperm [64]. Both osyuc11 and osnf-yb1 showed reduced seed size and increased chalkiness, accompanied by reduced indole-3-acetic acid level [64]. OsYUC11-mediated auxin biosynthesis was critical for endosperm development in rice [64]. Mutations of the wheat Aux/IAA gene TaIAA21 significantly increased grain length, width, and weight [65]. Mutations of the tetraploid wheat ARF25 gene significantly reduced grain size and weight [65]. Transcriptional activation experiments showed that ARF25 promoted the transcription of ERF3, while TtERF3 mutations led to reduced grain size and weight [65].
The modified expression of TaCYP78A5 could accumulate auxin to increase wheat grain weight and yield per plant [66]. Constitutive overexpression of TaCYP78A5 led to significantly increased seed size and weight, but not yield per plant due to the enhancement of apical dominance [66]. Auxin Response Factor 19 (JcARF19) was demonstrated to control seed size and seed yield in the woody plant Jatropha curcas [67]. Overexpression of JcARF19 significantly increased seed size and seed yield in Arabidopsis and Jatropha curcas, indicating the importance of auxin pathway in controlling seed yield [67].
7.2. Brassinosteroids (BRs) regulatory pathway
BRs are a group of plant steroid hormones that play an important role in regulating organ size [68]. Sun et al. identified an ortholog of rice DWARF11 (D11) in maize (Zea mays) called ZmD11 [68]. They found that constitutive expression of ZmD11 significantly increased seed length, seed area, seed weight, and both seed starch and protein contents in rice and maize [68]. Overexpression of the BR-synthesis gene D11 or the BR signaling factor OsBZR1 led to the accumulation of more sugar in developing anthers and seeds, resulting in higher grain yield [69]. Conversely, knockdown of D11 or OsBZR1 led to defective pollen maturation, reduced seed size and weight, with less starch accumulation [69]. The CLUSTERED PRIMARY BRANCH 1 (CPB1) gene is a new allele of D11 in rice. Mutations in the CPB1/D11 gene specifically affected rice panicle architecture and seed size development, and the CPB1/D11 transgenic plants driven by panicle-specific promoters could enlarge seed size and enhanced grain yield [70]. TaD11 affected wheat grain size and root length [71]. Ectopic expression of TaD11-2A rescued the abnormal panicle structure and plant height of the cpb1 mutant, and also increased endogenous BR levels, resulting in improved grain yield and quality in rice [71]. The tad11-2a-1 mutant showed dwarfism, smaller grains, and reduced endogenous BR levels [71]. The semi-dominant mutant Small and round seed 5 (Srs5) encodes an alpha-tubulin protein [72]. Mutant in the BR receptor BRI1 (d61-2) resulted in short seed phenotype due to impaired cell length [72]. Seeds of the Srs5 d61-2 double mutant were smaller than those of either single mutant [72]. Overexpression of SRS5 could promote grain length and rescue the shortened grain length of BR-related mutants [73].
BR plays a crucial role in determining the seed size, quality, and shape by transcriptionally modulating specific seed developmental pathways. The seeds of BR-deficient mutant de-etiolated 2 (det2) exhibited decreased seed cavity, reduced endosperm volume and integument cell length, resulting in smaller and less elongated seeds compared with WT [74]. The det2 mutant also showed delayed embryo development, with reduced size and number of embryo cell [74]. BR activated the expression of positive regulators of seed size, such as SHB1, MINI3 and IKU2, while suppressed the expression of negative regulators of seed size, such as AP2 and ARF2 [74]. In rice, glycogen synthase kinase-2 (GSK2) phosphorylated WRKY53 and reduced its stability, indicating that WRKY53 is a substrate of GSK2 in BR signaling [75]. WRKY53 interacted with BZR1 and cooperated to regulate BR-related developmental processes [75]. WRKY53 and the MAPKKK10-MAPKK4-MAPK6 cascade pathway jointly participated in controlling leaf angle and seed size, suggesting that WRKY53 was a direct substrate of this pathway [75]. GSK2 phosphorylated MAPKK4 to inhibit MAPK6 activity, suggesting that GSK2-mediated BR signaling might also regulate the MAPK pathway [75].
7.3. Gibberellin (GA) regulatory pathway
GA is an important phytohormones that plays a crucial role in controlling seed germination, stem elongation, leaf development, and floral induction [76]. Transgenic plant overexpressing Arabidopsis GA2-oxidase gene (AtGA2ox8) in Brassica napus L. exhibited a significant increase in seed yield by 9.6-12.4% [77]. Gibberellic acid-stimulated Arabidopsis 4 (GASA4) was one of the 14 members of the small polypeptide family in Arabidopsis, which regulated flowering and seed development and affected seed size, weight, and yield [78]. The gasa4-1 null mutant had significantly reduced seed weight, while, the 35S::GASA4-overexpressing lines had significantly increased seed weight and yield [78]. Even in the gasa4-1 plants with smaller seeds, higher total seed yield was also observed compared to WT due to increased branching, resulting in more seed number [78]. TaGW2-6A encoded a RING E3 ubiquitin-ligases with actively involved in GA perception and signal transduction and negatively regulated grain size [79]. TaGW2-6A allelic variation increased seed size through the GA signaling regulatory pathway [79]. TaGW2-6A allelic variation regulated GA synthesis through GA3-oxidase, resulting in high expression of GASA4 to control endosperm cell elongation and division during grain filling [79].
7.4. Jasmonate (JA) regulatory pathway
JA is a key phytohormone essential for multiple developmental processes in plants [80]. Seed size was regulated positively by the JA pathway factors COI1, MYC2 (and its homologs), MED25, and JAR1, but promoted by the JA signaling repressor JAZ [80,81]. It was found that the JA receptor mutants, both coi1-2 and coi1-8 had obviously enlarged seeds compared WT in Arabidopsis and OsCOI1-RNAi lines coi1-13 and coi1-18 also exhibited much larger grains than WT in rice [80]. MYCs are key transcription factors responsible for diverse JA responses [80]. The myc2 single mutant, myc2/3 double mutant, myc2/3/4 triple mutant and myc2/3/4/5 quadruple mutant all exhibited large seed size phenotype, with increased 100-grain weight, seed length and width compared with WT, indicating MYC2, MYC3, MYC4 and MYC5 acted redundantly to repress seed size [80]. MYC-MED25/PFT1 complex was required for activation of JA-responsive gene expression. The med25 mutants, both pft1-2 and pft1-3 displayed a significant increase in seed size and 100-grain weight [80].
JA stimulated the degradation of JASMONATE ZIM-DOMAIN (JAZ) protein, thereby relieving the inhibition of transcription factors coincident with reduced growth and fecundity. The mutant of JA signaling repressor JAZ6 (jaz6-3) exhibited decreased seed length, width and 100-grain weight compared to WT [80]. The JA signaling repressor OsJAZ11 was involved in regulating seed width and weight, since transgenic plants overexpressing OsJAZ11 showed up to a 14% increase in seed width and ~30% increase in seed weight compared to WT [81]. It was demonstrated that OsJAZ11 interacted with OsMADS29 and OsMADS68. OsGW7, a key negative regulator of grain size, was downregulated in OsJAZ11-overexpressing plants, suggesting that OsJAZ11 participated in regulating seed size by coordinating the expression of JA-related, OsGW7, and MADS genes [81]. In Arabidopsis, JAZ-deficient mutants (jazD) with mutations in three MYC genes (mycT) exhibited enhanced defense and reduced seed yield [82]. The de-repression of MYCs in jazD reduced fruit size and seed yield [82]. JA plays an essential role in plant seed development, but the regulation of seed size by JA is still not very clear.
7.5. Cytokinin (CK) regulatory pathway
CK is a phytohormone that plays important roles in multiple biological processes [83] and is crucial for determining grain yield [84]. Exogenous application of CK, ectopic expression of IPT, or down-regulation of cytokinin oxidase/dehydrogenase (CKX) led to increased seed yield [85]. ONAC096 regulated grain yield by affecting leaf senescence and panicle number in rice [86]. Inhibition OsCKX2 expression in onac096 mutants led to a 15% increase in panicle number [86]. OsCKX2 regulated rice grain yield by controlling cytokinin levels and modulating floral primordial activity under normal and abiotic stress conditions [84]. Yeh et al. used artificial short hairpin RNAs (shRNA) to specifically silence OsCKX2 and found that both CX3 and CX5 transgenic plants produced more tillers and grains per plant and had a heavier 1,000-grain weight than WT [87]. In cotton, suppression of CKX led to increased endogenous cytokinin levels and the transgenic plants with moderate inhibition resulted in delayed leaf senescence, enhanced photosynthesis, increased fruiting branches and bolls, and larger seed size [88].
Treatment of thidiazuron (TDZ), a novel and efficient cytokinin, could increase seed diameter and silique length, as well as the yield since the embryos and cotyledon epidermal cells were obviously larger than the control under TDZ treatment in Brassica napus [89]. TDZ upregulated genes related to maternal tissue development, including two G-protein signaling genes (AGG3 and RGA1) and two transcription factors (ANT and GS2) and might regulate key genes involved in auxin metabolism, and endosperm growth, resulting in enlargement of cotyledon epidermal cells and seed size in Brassica napus [89].
Arabidopsis histidine kinases (AHKs) are receptors in the CK signaling pathway. The ahk2 ahk3 mutant showed an increase in seed size but a decrease in seed yield, indicating a negative regulation of CK on seed size and a positive regulation on seed yield [90]. Bartrina et al. isolated the constitutively active gain-of-function variants of AHK2 and AHK3 genes, named repressor of cytokinin deficiency2 (rock2) and rock3 [91]. The transgenic AHK2: rock2 lines showed almost twice as many siliques as the WT. The number and yield of siliques were increased in AHK3: rock3 due to an extended flowering period [91]. Riefler et al. found that the seed length and width of ahk2 ahk3 ahk4 triple mutant were 30% larger than those of WT, while the seed volume of ahk2 ahk3 ahk4 increased to ~250% of the WT seed volume [92].
PURINE PERMEASE1 (OsPUP1), involved in importing CK into endoplasmic reticulum (ER) to unload CK from the vascular tissues by cell-to-cell transport, and overexpression of which resulted in reduced leaf length, plant height, grain weight, panicle length, and grain number in rice [83]. A dominant mutant, big grain3 (bg3-D) showed significant increase of grain size caused by activation of the OsPUP4 which involved in the long-distance transport of CK [93]. The T-DNA mutant of OsPUP7, which involved in transporting the CK derivative caffeine, exhibited various phenotypic changes, such as increased plant height, larger seeds, and delayed flowering [94]. These results indicate that PUP transport system involved in CK transport is critical for seed development.
7.6. Abscisic acid (ABA) regulatory pathway
ABA regulates various aspects of plant growth and development as well as responses to abiotic stress [95]. LOS5/ABA3 is involved in ABA biosynthesis by encoding molybdenum co-factor sulfurase, which is required by aldehyde oxidase (AO) in the last step of ABA biosynthesis in plants [96]. Transgenic plants overexpressing LOS5/ABA3 showed at least a 21% increase in seed yield compared to the WT under drought stress conditions [96]. OsAO3 was essential for regulation of grain yield in rice, since osao3 mutant increased grain yield while overexpression of OsAO3 reduced grain yield by affecting panicle number per plant, spikelet number per panicle, and spikelet fertility [95]. Cytosolic ABA receptors PYRABACTIN RESISTANCE 1 LIKE/REGULATORY COMPONENTS OF ABA RECEPTORS (PYL/RCARs) could regulate ABA-dependent gene expression in rice [97]. Constitutive expression of OsPYL/RCAR5 slightly reduced plant height and severely decreased seed yield under paddy field conditions although improved abiotic stress tolerance [97].
ABSCISIC ACID DEFICIENT2, a unique short-chain dehydrogenase/reductase involved in ABA biosynthesis, regulated seed development, since the aba2-1 mutant exhibited increased seed size, mass, and embryo cell number, but delayed endosperm cellularization [98]. RNA level of SHB1, MINI3, and IKU2 involved in seed size control was significantly increased in aba2-1 and ABSCISIC ACID-INSENSITIVE (ABI5) involved in ABA signaling was decreased in aba2-1 [98]. Functional loss of ABI5 led to increased seed size. ABA2 regulated the endogenous levels of ABA and controlled the transcription of ABI5. ABI5 inhibited the expression of SHB1, which further regulated MINI3 and IKU2 [98].
8. MicroRNAs (miRNAs) regulatory pathway
MiRNAs are a class of small noncoding RNAs that play key regulatory roles in plant seed development, and the Zma-miRNA169 family is highly expressed during maize seed development [99]. Overexpression of maize zma-miR169o increased seed size and weight, while inhibiting its expression had the opposite effect [99]. Zma-miR169 negatively regulated its target gene ZmNF-YA13, which played a crucial role in determining seed size and regulated the expression of the auxin biosynthesis gene ZmYUC1, thereby regulating maize seed size and ultimately yield [99]. OsmiR530 negatively regulated grain yield, as suppressing OsmiR530 increased grain yield, while overexpression of OsmiR530 significantly reduced grain size and panicle branching, leading to yield decrease [100]. Phytochrome-interacting factor-like 3 (OsPL3) encoded a protein containing a PLUS3 domain and was directly targeted by OsmiR530 and knocking out OsPL3 reduced grain yield [100]. Both overexpressing (miR529a-OE) and suppressing (miR529a-MIMIC) transgenic plants of miR529a resulted in narrower grain size and lower 1,000-grain weight, while miR529a-OE resulted in longer and slenderer grains, but miR529a-MIMIC led to shorter grains [101]. MiR529a regulated grain sizes by altering the expression of SQUAMOSA PROMOTER BINDING-LIKE (SPL) transcription factors which could control rice tiller, panicle architecture and grain size [101].
Zhao et al. successfully suppressed the expression of mature miR159 by using Short Tandem Target Mimic (STTM), leading to increased expressions of its two target genes, OsGAMYB and OsGAMYBL1 (GAMYB-LIKE 1) [102]. STTM159 transgenic plants were dwarfed with reduced organ size, stem diameter, length of flag leaf, main panicle, spikelet hulls and grain size [102]. Transgenic plants overexpressing miR398 (OX-miR398a) could increase rice panicle length, grain number and grain size, however, silencing of miR398 (STTM398) had the opposite effects [103]. Os07g46990, one of the miR398 targets, was modified, and the mOs07g46990 transgenic plants showed decreased panicle length and grain number per panicle like the phenotypes of STTM lines [103]. Wang et al. knocked down miR160 and miR165/166 (STTM160 and STTM165/166) and found that STTM160 showed significantly smaller grain size, lower grain weight, shorter siliques length, shorter plant height, and more serrated leaves, while STTM165/166 showed reduced seed numbers, disabled siliques, and curled upward leaves [104].
9. Conclusion and prospect
Seed size is particularly important for agricultural production and is regulated by the coordinated control of the ovule, endosperm, and embryo, while the development of these tissues is regulated by various pathways (Figure 1). The ubiquitin-proteasome pathway and the MAPK signaling pathway primarily influence integument development. The IKU pathway mainly regulates endosperm development. Furthermore, several pathways involved in seed size regulation may cross-talk through certain genes or phytohormones, such as the auxin signaling pathway and BR signaling pathway (ARF2 and BZR1), the IKU pathway and cytokinin signaling pathway (MINI3 and CKX2), the BR signaling pathway and MAPK pathway (OsMKK4, OsMAPK6, and BR-related genes), the G-protein pathway and BR signaling pathway (Gα influences BR signal cascade), and so on. However, despite this, the molecular mechanisms governing seed size regulation are still not fully understood, and the connections between various pathways are scarce, thus failing to elucidate how the development of the ovule, endosperm, and embryo is coordinated with each other.
Seed size is regulated by a complex regulatory network, and although many genes have been identified (Table 1), the regulatory mechanisms are still not well understood, and there is still much to be explored regarding upstream and downstream genes and proteins. For instance, the regulatory processes of some genes are still unclear, and their target substrates remain unknown. Are they involved in signal transduction processes? What are the specific ligands and regulatory factors? Do these regulatory genes exist in other crops? How are they expressed during seed development? Modern biotechnologies such as gene editing techniques and systems biology approaches like transcriptomics, proteomics, metabolomics, and epigenomics will facilitate researchers in unraveling the molecular mechanisms that regulate seed size more rapidly.
Seeds are the most crucial source of food for human survival. The study of genes regulating seed size development has significant theoretical value and provides essential genetic information for improving food production through genetic engineering techniques. It holds great potential for various areas, such as functional genetic improvement, and offers important prospects for applications in increasing crop yield and other related fields.
Author Contributions
J.Z. and X.W. conceptualized and wrote the manuscript. X.Z. and Q.P. prepared the figure and table. X.L. and Y.W. collected the literature. All authors have read and agreed to the published version of the manuscript.
Funding
The work was supported by the National Natural Science Foundation of China (32000244), the Research Start Up Fund of Henan Agricultural University (Grant No. 30500484), Program for Innovative Research Team (in Science and Technology) in University of Henan Province (Grant No. 221RTSTHN0230).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sakamoto, T.; Matsuoka, M. Identifying and exploiting grain yield genes in rice. Current opinion in plant biology 2008, 11, 209–214. [Google Scholar] [CrossRef]
- Jiang, W.B.; Lin, W.H. Brassinosteroid functions in Arabidopsis seed development. Plant signaling & behavior 2013, 8. [Google Scholar] [CrossRef]
- Yu, A.; Wang, Z.; Zhang, Y.; Li, F.; Liu, A. Global Gene Expression of Seed Coat Tissues Reveals a Potential Mechanism of Regulating Seed Size Formation in Castor Bean. International journal of molecular sciences 2019, 20, 1282. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yue, L.; Wu, X.; Liu, H.; Wang, W. Function of Small Peptides During Male-Female Crosstalk in Plants. Frontiers in plant science 2021, 12, 671196. [Google Scholar] [CrossRef]
- Chaudhury, A.M.; Koltunow, A.; Payne, T.; Luo, M.; Tucker, M.R.; Dennis, E.S.; Peacock, W.J. Control of early seed development. Annual review of cell and developmental biology 2001, 17, 677–699. [Google Scholar] [CrossRef]
- Li, J.; Nie, X.; Tan, J.L.; Berger, F. Integration of epigenetic and genetic controls of seed size by cytokinin in Arabidopsis. Proceedings of the National Academy of Sciences of the United States of America 2013, 110, 15479–15484. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Jiang, S.; Jiang, L.; Li, W.; Tang, Y.; He, W.; Wang, M.; Xing, J.; Cui, Y.; Lin, Q.; et al. Transcription factor OsSGL is a regulator of starch synthesis and grain quality in rice. Journal of experimental botany 2022, 73, 3417–3430. [Google Scholar] [CrossRef] [PubMed]
- Paolo, D.; Orozco-Arroyo, G.; Rotasperti, L.; Masiero, S.; Colombo, L.; de Folter, S.; Ambrose, B.A.; Caporali, E.; Ezquer, I.; Mizzotti, C. Genetic Interaction of SEEDSTICK, GORDITA and AUXIN RESPONSE FACTOR 2 during Seed Development. Genes 2021, 12, 1189. [Google Scholar] [CrossRef]
- Cao, J.; Li, G.; Qu, D.; Li, X.; Wang, Y. Into the Seed: Auxin Controls Seed Development and Grain Yield. International journal of molecular sciences 2020, 21, 1662. [Google Scholar] [CrossRef]
- Alam, I.; Batool, K.; Huang, Y.; Liu, J.; Ge, L. Developing Genetic Engineering Techniques for Control of Seed Size and Yield. International journal of molecular sciences 2022, 23, 3256. [Google Scholar] [CrossRef]
- Yang, X.; Zhao, X.; Dai, Z.; Ma, F.; Miao, X.; Shi, Z. OsmiR396/growth regulating factor modulate rice grain size through direct regulation of embryo-specific miR408. Plant physiology 2021, 186, 519–533. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Garcia, D.; Zhang, H.; Feng, K.; Chaudhury, A.; Berger, F.; Peacock, W.J.; Dennis, E.S.; Luo, M. The VQ motif protein IKU1 regulates endosperm growth and seed size in Arabidopsis. The Plant journal : for cell and molecular biology 2010, 63, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Fatihi, A.; Zbierzak, A.M.; Dormann, P. Alterations in seed development gene expression affect size and oil content of Arabidopsis seeds. Plant physiology 2013, 163, 973–985. [Google Scholar] [CrossRef]
- Luo, M.; Dennis, E.S.; Berger, F.; Peacock, W.J.; Chaudhury, A. MINISEED3 (MINI3), a WRKY family gene, and HAIKU2 (IKU2), a leucine-rich repeat (LRR) KINASE gene, are regulators of seed size in Arabidopsis. Proceedings of the National Academy of Sciences of the United States of America 2005, 102, 17531–17536. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, X.; Kang, X.; Zhao, X.; Zhang, X.; Ni, M. SHORT HYPOCOTYL UNDER BLUE1 associates with MINISEED3 and HAIKU2 promoters in vivo to regulate Arabidopsis seed development. The Plant cell 2009, 21, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Yang, S.; Wu, L.; Xin, Y.; Song, J.; Wang, L.; Pei, W.; Wu, M.; Yu, J.; Ma, X.; et al. Genome-Wide Analysis of the GW2-Like Genes in Gossypium and Functional Characterization of the Seed Size Effect of GhGW2-2D. Frontiers in plant science 2022, 13, 860922. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Park, S.W.; Kim, Y.J.; Koo, Y.J.; Song, J.T.; Seo, H.S. Grain width 2 (GW2) and its interacting proteins regulate seed development in rice (Oryza sativa L.). Botanical studies 2018, 59, 23. [Google Scholar] [CrossRef]
- Wang, J.L.; Tang, M.Q.; Chen, S.; Zheng, X.F.; Mo, H.X.; Li, S.J.; Wang, Z.; Zhu, K.M.; Ding, L.N.; Liu, S.Y.; et al. Down-regulation of BnDA1, whose gene locus is associated with the seeds weight, improves the seeds weight and organ size in Brassica napus. Plant biotechnology journal 2017, 15, 1024–1033. [Google Scholar] [CrossRef]
- Liu, H.; Li, H.; Hao, C.; Wang, K.; Wang, Y.; Qin, L.; An, D.; Li, T.; Zhang, X. TaDA1, a conserved negative regulator of kernel size, has an additive effect with TaGW2 in common wheat (Triticum aestivum L.). Plant biotechnology journal 2020, 18, 1330–1342. [Google Scholar] [CrossRef]
- Zhang, Y.; Du, L.; Xu, R.; Cui, R.; Hao, J.; Sun, C.; Li, Y. Transcription factors SOD7/NGAL2 and DPA4/NGAL3 act redundantly to regulate seed size by directly repressing KLU expression in Arabidopsis thaliana. The Plant cell 2015, 27, 620–632. [Google Scholar] [CrossRef]
- Yang, S.; Huang, L.; Song, J.; Liu, L.; Bian, Y.; Jia, B.; Wu, L.; Xin, Y.; Wu, M.; Zhang, J.; et al. Genome-Wide Analysis of DA1-Like Genes in Gossypium and Functional Characterization of GhDA1-1A Controlling Seed Size. Frontiers in plant science 2021, 12, 647091. [Google Scholar] [CrossRef] [PubMed]
- Xia, T.; Li, N.; Dumenil, J.; Li, J.; Kamenski, A.; Bevan, M.W.; Gao, F.; Li, Y. The ubiquitin receptor DA1 interacts with the E3 ubiquitin ligase DA2 to regulate seed and organ size in Arabidopsis. The Plant cell 2013, 25, 3347–3359. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Chen, L.; Lu, Y.; Wu, Y.; Dumenil, J.; Zhu, Z.; Bevan, M.W.; Li, Y. The ubiquitin receptors DA1, DAR1, and DAR2 redundantly regulate endoreduplication by modulating the stability of TCP14/15 in Arabidopsis. The Plant cell 2015, 27, 649–662. [Google Scholar] [CrossRef]
- Du, L.; Li, N.; Chen, L.; Xu, Y.; Li, Y.; Zhang, Y.; Li, C.; Li, Y. The ubiquitin receptor DA1 regulates seed and organ size by modulating the stability of the ubiquitin-specific protease UBP15/SOD2 in Arabidopsis. The Plant cell 2014, 26, 665–677. [Google Scholar] [CrossRef]
- Shi, C.; Ren, Y.; Liu, L.; Wang, F.; Zhang, H.; Tian, P.; Pan, T.; Wang, Y.; Jing, R.; Liu, T.; et al. Ubiquitin Specific Protease 15 Has an Important Role in Regulating Grain Width and Size in Rice. Plant physiology 2019, 180, 381–391. [Google Scholar] [CrossRef]
- Sun, S.; Wang, L.; Mao, H.; Shao, L.; Li, X.; Xiao, J.; Ouyang, Y.; Zhang, Q. A G-protein pathway determines grain size in rice. Nature communications 2018, 9, 851. [Google Scholar] [CrossRef]
- Wu, L.; Wang, X.; Yu, Z.; Cui, X.; Xu, Q. Simultaneous Improvement of Grain Yield and Quality through Manipulating Two Type C G Protein Gamma Subunits in Rice. International journal of molecular sciences 2022, 23, 1463. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Wu, K.; Wang, B.; Liu, H.; Guo, S.; Guo, X.; Luo, W.; Sun, S.; Ouyang, Y.; Fu, X.; et al. The RING E3 ligase CLG1 targets GS3 for degradation via the endosome pathway to determine grain size in rice. Molecular plant 2021, 14, 1699–1713. [Google Scholar] [CrossRef]
- Li, S.; Liu, W.; Zhang, X.; Liu, Y.; Li, N.; Li, Y. Roles of the Arabidopsis G protein gamma subunit AGG3 and its rice homologs GS3 and DEP1 in seed and organ size control. Plant signaling & behavior 2012, 7, 1357–1359. [Google Scholar] [CrossRef]
- Li, S.; Liu, Y.; Zheng, L.; Chen, L.; Li, N.; Corke, F.; Lu, Y.; Fu, X.; Zhu, Z.; Bevan, M.W.; et al. The plant-specific G protein gamma subunit AGG3 influences organ size and shape in Arabidopsis thaliana. The New phytologist 2012, 194, 690–703. [Google Scholar] [CrossRef]
- Roy Choudhury, S.; Riesselman, A.J.; Pandey, S. Constitutive or seed-specific overexpression of Arabidopsis G-protein gamma subunit 3 (AGG3) results in increased seed and oil production and improved stress tolerance in Camelina sativa. Plant biotechnology journal 2014, 12, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Han, R.; Wu, K.; Zhang, J.; Ye, Y.; Wang, S.; Chen, J.; Pan, Y.; Li, Q.; Xu, X.; et al. G-protein betagamma subunits determine grain size through interaction with MADS-domain transcription factors in rice. Nature communications 2018, 9, 852. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Bai, X.; Cheng, N.; Xiao, J.; Li, X.; Xing, Y. Wide Grain 7 increases grain width by enhancing H3K4me3 enrichment in the OsMADS1 promoter in rice (Oryza sativa L.). The Plant journal : for cell and molecular biology 2020, 102, 517–528. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, M.; Zhou, Y.; Wang, Y.; Shen, J.; Chen, H.; Zhang, L.; Lu, B.; Liang, G.; Liang, J. The Rice G Protein gamma Subunit DEP1/qPE9-1 Positively Regulates Grain-Filling Process by Increasing Auxin and Cytokinin Content in Rice Grains. Rice 2019, 12, 91. [Google Scholar] [CrossRef]
- Utsunomiya, Y.; Samejima, C.; Takayanagi, Y.; Izawa, Y.; Yoshida, T.; Sawada, Y.; Fujisawa, Y.; Kato, H.; Iwasaki, Y. Suppression of the rice heterotrimeric G protein beta-subunit gene, RGB1, causes dwarfism and browning of internodes and lamina joint regions. The Plant journal : for cell and molecular biology 2011, 67, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Yang, Z.; Zhang, D.; Wang, Y.; Xu, M.; Zhou, L.; Wang, J.; Wu, S.; Yao, Y.; Du, X.; et al. Mutation of RGG2, which encodes a type B heterotrimeric G protein gamma subunit, increases grain size and yield production in rice. Plant biotechnology journal 2019, 17, 650–664. [Google Scholar] [CrossRef]
- Chen, T.; Zhou, B.; Duan, L.; Zhu, H.; Zhang, Z. MtMAPKK4 is an essential gene for growth and reproduction of Medicago truncatula. Physiologia plantarum 2017, 159, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Osuna, A.; Calatrava, V.; Galvan, A.; Fernandez, E.; Llamas, A. Identification of the MAPK Cascade and its Relationship with Nitrogen Metabolism in the Green Alga Chlamydomonas reinhardtii. International journal of molecular sciences 2020, 21, 3417. [Google Scholar] [CrossRef]
- Xi, X.; Hu, Z.; Nie, X.; Meng, M.; Xu, H.; Li, J. Cross Inhibition of MPK10 and WRKY10 Participating in the Growth of Endosperm in Arabidopsis thaliana. Frontiers in plant science 2021, 12, 640346. [Google Scholar] [CrossRef]
- Wu, X.; Cai, X.; Zhang, B.; Wu, S.; Wang, R.; Li, N.; Li, Y.; Sun, Y.; Tang, W. ERECTA regulates seed size independently of its intracellular domain via MAPK-DA1-UBP15 signaling. The Plant cell 2022, 34, 3773–3789. [Google Scholar] [CrossRef]
- Pan, Y.; Chen, L.; Zhao, Y.; Guo, H.; Li, J.; Rashid, M.A.R.; Lu, C.; Zhou, W.; Yang, X.; Liang, Y.; et al. Natural Variation in OsMKK3 Contributes to Grain Size and Chalkiness in Rice. Frontiers in plant science 2021, 12, 784037. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Mei, E.; Tian, X.; He, M.; Tang, J.; Xu, M.; Liu, J.; Song, L.; Li, X.; Wang, Z.; et al. OsMKKK70 regulates grain size and leaf angle in rice through the OsMKK4-OsMAPK6-OsWRKY53 signaling pathway. Journal of integrative plant biology 2021, 63, 2043–2057. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Hua, L.; Dong, S.; Chen, H.; Zhu, X.; Jiang, J.; Zhang, F.; Li, Y.; Fang, X.; Chen, F. OsMAPK6, a mitogen-activated protein kinase, influences rice grain size and biomass production. The Plant journal : for cell and molecular biology 2015, 84, 672–681. [Google Scholar] [CrossRef]
- Xu, R.; Duan, P.; Yu, H.; Zhou, Z.; Zhang, B.; Wang, R.; Li, J.; Zhang, G.; Zhuang, S.; Lyu, J.; et al. Control of Grain Size and Weight by the OsMKKK10-OsMKK4-OsMAPK6 Signaling Pathway in Rice. Molecular plant 2018, 11, 860–873. [Google Scholar] [CrossRef]
- Guo, T.; Chen, K.; Dong, N.Q.; Shi, C.L.; Ye, W.W.; Gao, J.P.; Shan, J.X.; Lin, H.X. GRAIN SIZE AND NUMBER1 Negatively Regulates the OsMKKK10-OsMKK4-OsMPK6 Cascade to Coordinate the Trade-off between Grain Number per Panicle and Grain Size in Rice. The Plant cell 2018, 30, 871–888. [Google Scholar] [CrossRef]
- Xiao, W.; Hu, S.; Zou, X.; Cai, R.; Liao, R.; Lin, X.; Yao, R.; Guo, X. Lectin receptor-like kinase LecRK-VIII.2 is a missing link in MAPK signaling-mediated yield control. Plant physiology 2021, 187, 303–320. [Google Scholar] [CrossRef] [PubMed]
- Majeed, U.; Hou, J.; Hao, C.; Zhang, X. TaNAC020 homoeologous genes are associated with higher thousand kernel weight and kernel length in Chinese wheat. Frontiers in genetics 2022, 13, 956921. [Google Scholar] [CrossRef]
- Li, J.; Xie, L.; Tian, X.; Liu, S.; Xu, D.; Jin, H.; Song, J.; Dong, Y.; Zhao, D.; Li, G.; et al. TaNAC100 acts as an integrator of seed protein and starch synthesis exerting pleiotropic effects on agronomic traits in wheat. The Plant journal : for cell and molecular biology 2021, 108, 829–840. [Google Scholar] [CrossRef]
- Jiang, D.; Chen, W.; Dong, J.; Li, J.; Yang, F.; Wu, Z.; Zhou, H.; Wang, W.; Zhuang, C. Overexpression of miR164b-resistant OsNAC2 improves plant architecture and grain yield in rice. Journal of experimental botany 2018, 69, 1533–1543. [Google Scholar] [CrossRef]
- Jin, S.K.; Zhang, M.Q.; Leng, Y.J.; Xu, L.N.; Jia, S.W.; Wang, S.L.; Song, T.; Wang, R.A.; Yang, Q.Q.; Tao, T.; et al. OsNAC129 Regulates Seed Development and Plant Growth and Participates in the Brassinosteroid Signaling Pathway. Frontiers in plant science 2022, 13, 905148. [Google Scholar] [CrossRef]
- Yang, F.; Liu, G.; Wu, Z.; Zhang, D.; Zhang, Y.; You, M.; Li, B.; Zhang, X.; Liang, R. Cloning and Functional Analysis of TaWRI1Ls, the Key Genes for Grain Fatty Acid Synthesis in Bread Wheat. International journal of molecular sciences 2022, 23, 5293. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.Y.; Nam, K.H. RAV1 Negatively Regulates Seed Development by Directly Repressing MINI3 and IKU2 in Arabidopsis. Molecules and cells 2018, 41, 1072–1080. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Ma, X.; Xia, H.; Wang, L.; Chen, S.; Xu, K.; Yang, F.; Zou, Y.; Wang, Y.; Zhu, J.; et al. Natural variation of Alfin-like family affects seed size and drought tolerance in rice. The Plant journal : for cell and molecular biology 2022, 112, 1176–1193. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kim, S.H.; Shin, D.M.; Kim, S.H. ATBS1-INTERACTING FACTOR 2 Negatively Modulates Pollen Production and Seed Formation in Arabidopsis. Frontiers in plant science 2021, 12, 704958. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Guo, W.; Du, D.; Pu, L.; Zhang, C. Overexpression of a maize BR transcription factor ZmBZR1 in Arabidopsis enlarges organ and seed size of the transgenic plants. Plant science : an international journal of experimental plant biology 2020, 292, 110378. [Google Scholar] [CrossRef]
- Sun, F.; Ding, L.; Feng, W.; Cao, Y.; Lu, F.; Yang, Q.; Li, W.; Lu, Y.; Shabek, N.; Fu, F.; et al. Maize transcription factor ZmBES1/BZR1-5 positively regulates kernel size. Journal of experimental botany 2021, 72, 1714–1726. [Google Scholar] [CrossRef]
- Nutan, K.K.; Singla-Pareek, S.L.; Pareek, A. The Saltol QTL-localized transcription factor OsGATA8 plays an important role in stress tolerance and seed development in Arabidopsis and rice. Journal of experimental botany 2020, 71, 684–698. [Google Scholar] [CrossRef]
- Song, G.; Li, X.; Munir, R.; Khan, A.R.; Azhar, W.; Yasin, M.U.; Jiang, Q.; Bancroft, I.; Gan, Y. The WRKY6 transcription factor affects seed oil accumulation and alters fatty acid compositions in Arabidopsis thaliana. Physiologia plantarum 2020, 169, 612–624. [Google Scholar] [CrossRef]
- Zhang, Y.; Liang, W.; Shi, J.; Xu, J.; Zhang, D. MYB56 encoding a R2R3 MYB transcription factor regulates seed size in Arabidopsis thaliana. Journal of integrative plant biology 2013, 55, 1166–1178. [Google Scholar] [CrossRef]
- Yu, Y.T.; Wu, Z.; Lu, K.; Bi, C.; Liang, S.; Wang, X.F.; Zhang, D.P. Overexpression of the MYB37 transcription factor enhances abscisic acid sensitivity, and improves both drought tolerance and seed productivity in Arabidopsis thaliana. Plant molecular biology 2016, 90, 267–279. [Google Scholar] [CrossRef]
- Wang, Y.; Nie, L.; Ma, J.; Zhou, B.; Han, X.; Cheng, J.; Lu, X.; Fan, Z.; Li, Y.; Cao, Y. Transcriptomic Variations and Network Hubs Controlling Seed Size and Weight During Maize Seed Development. Frontiers in plant science 2022, 13, 828923. [Google Scholar] [CrossRef]
- He, Z.; Zeng, J.; Ren, Y.; Chen, D.; Li, W.; Gao, F.; Cao, Y.; Luo, T.; Yuan, G.; Wu, X.; et al. OsGIF1 Positively Regulates the Sizes of Stems, Leaves, and Grains in Rice. Frontiers in plant science 2017, 8, 1730. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Gao, F.; Xie, K.; Zeng, X.; Cao, Y.; Zeng, J.; He, Z.; Ren, Y.; Li, W.; Deng, Q.; et al. The OsmiR396c-OsGRF4-OsGIF1 regulatory module determines grain size and yield in rice. Plant biotechnology journal 2016, 14, 2134–2146. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; E, Z.; Zhang, D.; Yun, Q.; Zhou, Y.; Niu, B.; Chen, C. OsYUC11-mediated auxin biosynthesis is essential for endosperm development of rice. Plant physiology 2021, 185, 934–950. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Li, Y.; Wang, Z.; Tao, S.; Sun, G.; Kong, X.; Wang, K.; Ye, X.; Liu, S.; Geng, S.; et al. TaIAA21 represses TaARF25-mediated expression of TaERFs required for grain size and weight development in wheat. The Plant journal : for cell and molecular biology 2021, 108, 1754–1767. [Google Scholar] [CrossRef]
- Guo, L.; Ma, M.; Wu, L.; Zhou, M.; Li, M.; Wu, B.; Li, L.; Liu, X.; Jing, R.; Chen, W.; et al. Modified expression of TaCYP78A5 enhances grain weight with yield potential by accumulating auxin in wheat (Triticum aestivum L.). Plant biotechnology journal 2022, 20, 168–182. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, C.; Wang, N.; Jiang, X.; Mao, H.; Zhu, C.; Wen, F.; Wang, X.; Lu, Z.; Yue, G.; et al. Manipulation of Auxin Response Factor 19 affects seed size in the woody perennial Jatropha curcas. Scientific reports 2017, 7, 40844. [Google Scholar] [CrossRef]
- Sun, H.; Xu, H.; Li, B.; Shang, Y.; Wei, M.; Zhang, S.; Zhao, C.; Qin, R.; Cui, F.; Wu, Y. The brassinosteroid biosynthesis gene, ZmD11, increases seed size and quality in rice and maize. Plant physiology and biochemistry : PPB 2021, 160, 281–293. [Google Scholar] [CrossRef]
- Zhu, X.; Liang, W.; Cui, X.; Chen, M.; Yin, C.; Luo, Z.; Zhu, J.; Lucas, W.J.; Wang, Z.; Zhang, D. Brassinosteroids promote development of rice pollen grains and seeds by triggering expression of Carbon Starved Anther, a MYB domain protein. The Plant journal : for cell and molecular biology 2015, 82, 570–581. [Google Scholar] [CrossRef]
- Wu, Y.; Fu, Y.; Zhao, S.; Gu, P.; Zhu, Z.; Sun, C.; Tan, L. CLUSTERED PRIMARY BRANCH 1, a new allele of DWARF11, controls panicle architecture and seed size in rice. Plant biotechnology journal 2016, 14, 377–386. [Google Scholar] [CrossRef]
- Xu, H.; Sun, H.; Dong, J.; Ma, C.; Li, J.; Li, Z.; Wang, Y.; Ji, J.; Hu, X.; Wu, M.; et al. The brassinosteroid biosynthesis gene TaD11-2A controls grain size and its elite haplotype improves wheat grain yields. TAG. Theoretical and applied genetics. Theoretische und angewandte Genetik 2022, 135, 2907–2923. [Google Scholar] [CrossRef] [PubMed]
- Segami, S.; Kono, I.; Ando, T.; Yano, M.; Kitano, H.; Miura, K.; Iwasaki, Y. Small and round seed 5 gene encodes alpha-tubulin regulating seed cell elongation in rice. Rice 2012, 5, 4. [Google Scholar] [CrossRef] [PubMed]
- Segami, S.; Takehara, K.; Yamamoto, T.; Kido, S.; Kondo, S.; Iwasaki, Y.; Miura, K. Overexpression of SRS5 improves grain size of brassinosteroid-related dwarf mutants in rice (Oryza sativa L.). Breeding science 2017, 67, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.B.; Huang, H.Y.; Hu, Y.W.; Zhu, S.W.; Wang, Z.Y.; Lin, W.H. Brassinosteroid regulates seed size and shape in Arabidopsis. Plant physiology 2013, 162, 1965–1977. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; He, M.; Mei, E.; Zhang, B.; Tang, J.; Xu, M.; Liu, J.; Li, X.; Wang, Z.; Tang, W.; et al. WRKY53 integrates classic brassinosteroid signaling and the mitogen-activated protein kinase pathway to regulate rice architecture and seed size. The Plant cell 2021, 33, 2753–2775. [Google Scholar] [CrossRef]
- Yu, S.; Wang, J.W. The Crosstalk between MicroRNAs and Gibberellin Signaling in Plants. Plant & cell physiology 2020, 61, 1880–1890. [Google Scholar] [CrossRef]
- Zhou, B.; Lin, J.Z.; Peng, W.S.; Peng, D.; Zhuo, Y.H.; Zhu, D.F.; Huang, X.Q.; Tang, D.Y.; Guo, M.; He, R.Q.; Zhang, J.H.; Li, X.S.; Zhao, X.Y.; Liu, X.M. Dwarfism in Brassica napus L. induced by the over-expression of a gibberellin 2-oxidase gene from Arabidopsis thaliana. Molecular breeding 2012, 29, 115–127. [Google Scholar] [CrossRef]
- Roxrud, I.; Lid, S.E.; Fletcher, J.C.; Schmidt, E.D.; Opsahl-Sorteberg, H.G. GASA4, one of the 14-member Arabidopsis GASA family of small polypeptides, regulates flowering and seed development. Plant & cell physiology 2007, 48, 471–483. [Google Scholar] [CrossRef]
- Li, Q.; Li, L.; Liu, Y.; Lv, Q.; Zhang, H.; Zhu, J.; Li, X. Influence of TaGW2-6A on seed development in wheat by negatively regulating gibberellin synthesis. Plant science : an international journal of experimental plant biology 2017, 263, 226–235. [Google Scholar] [CrossRef]
- Hu, S.; Yang, H.; Gao, H.; Yan, J.; Xie, D. Control of seed size by jasmonate. Science China. Life sciences 2021, 64, 1215–1226. [Google Scholar] [CrossRef]
- Mehra, P.; Pandey, B.K.; Verma, L.; Prusty, A.; Singh, A.P.; Sharma, S.; Malik, N.; Bennett, M.J.; Parida, S.K.; Giri, J.; et al. OsJAZ11 regulates spikelet and seed development in rice. Plant direct 2022, 6, e401. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Major, I.T.; Kapali, G.; Howe, G.A. MYC transcription factors coordinate tryptophan-dependent defence responses and compromise seed yield in Arabidopsis. The New phytologist 2022, 236, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Zhang, J.; Yu, G.; Lu, X.; Mei, W.; Deng, H.; Zhang, G.; Chen, G.; Chu, C.; Tong, H.; et al. Endoplasmic Reticulum-Localized PURINE PERMEASE1 Regulates Plant Height and Grain Weight by Modulating Cytokinin Distribution in Rice. Frontiers in plant science 2020, 11, 618560. [Google Scholar] [CrossRef]
- Joshi, R.; Sahoo, K.K.; Tripathi, A.K.; Kumar, R.; Gupta, B.K.; Pareek, A.; Singla-Pareek, S.L. Knockdown of an inflorescence meristem-specific cytokinin oxidase - OsCKX2 in rice reduces yield penalty under salinity stress condition. Plant, cell & environment 2018, 41, 936–946. [Google Scholar] [CrossRef]
- Jameson, P.E.; Song, J. Cytokinin: a key driver of seed yield. Journal of experimental botany 2016, 67, 593–606. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.; Shim, Y.; Gi, E.; An, G.; Paek, N.C. Mutation of ONAC096 Enhances Grain Yield by Increasing Panicle Number and Delaying Leaf Senescence during Grain Filling in Rice. International journal of molecular sciences 2019, 20, 5241. [Google Scholar] [CrossRef]
- Yeh, S.Y.; Chen, H.W.; Ng, C.Y.; Lin, C.Y.; Tseng, T.H.; Li, W.H.; Ku, M.S. Down-Regulation of Cytokinin Oxidase 2 Expression Increases Tiller Number and Improves Rice Yield. Rice 2015, 8, 36. [Google Scholar] [CrossRef]
- Zhao, J.; Bai, W.; Zeng, Q.; Song, S.; Zhang, M.; Li, X.; Hou, L.; Xiao, Y.; Luo, M.; Li, D.; et al. Moderately enhancing cytokinin level by down-regulation of GhCKX expression in cotton concurrently increases fiber and seed yield. Molecular breeding : new strategies in plant improvement 2015, 35, 60. [Google Scholar] [CrossRef]
- Zhai, L.; Xie, L.; Xu, J.; Xu, B.; Dong, J.; Zhang, X. Study on exogenous application of thidiazuron on seed size of Brassica napus L. Frontiers in plant science 2022, 13, 998698. [Google Scholar] [CrossRef]
- Li, W.; Nguyen, K.H.; Ha, C.V.; Watanabe, Y.; Tran, L.P. Crosstalk between the cytokinin and MAX2 signaling pathways in growth and callus formation of Arabidopsis thaliana. Biochemical and biophysical research communications 2019, 511, 300–306. [Google Scholar] [CrossRef]
- Bartrina, I.; Jensen, H.; Novak, O.; Strnad, M.; Werner, T.; Schmulling, T. Gain-of-Function Mutants of the Cytokinin Receptors AHK2 and AHK3 Regulate Plant Organ Size, Flowering Time and Plant Longevity. Plant physiology 2017, 173, 1783–1797. [Google Scholar] [CrossRef]
- Riefler, M.; Novak, O.; Strnad, M.; Schmulling, T. Arabidopsis cytokinin receptor mutants reveal functions in shoot growth, leaf senescence, seed size, germination, root development, and cytokinin metabolism. The Plant cell 2006, 18, 40–54. [Google Scholar] [CrossRef]
- Xiao, Y.; Liu, D.; Zhang, G.; Gao, S.; Liu, L.; Xu, F.; Che, R.; Wang, Y.; Tong, H.; Chu, C. Big Grain3, encoding a purine permease, regulates grain size via modulating cytokinin transport in rice. Journal of integrative plant biology 2019, 61, 581–597. [Google Scholar] [CrossRef]
- Qi, Z.; Xiong, L. Characterization of a purine permease family gene OsPUP7 involved in growth and development control in rice. Journal of integrative plant biology 2013, 55, 1119–1135. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Tian, Q.; Deng, P.; Zhang, W.; Jing, W. The rice aldehyde oxidase OsAO3 gene regulates plant growth, grain yield, and drought tolerance by participating in ABA biosynthesis. Biochemical and biophysical research communications 2021, 548, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, J.; Zhang, J.; Hao, L.; Hua, J.; Duan, L.; Zhang, M.; Li, Z. Expression of an Arabidopsis molybdenum cofactor sulphurase gene in soybean enhances drought tolerance and increases yield under field conditions. Plant biotechnology journal 2013, 11, 747–758. [Google Scholar] [CrossRef]
- Kim, H.; Lee, K.; Hwang, H.; Bhatnagar, N.; Kim, D.Y.; Yoon, I.S.; Byun, M.O.; Kim, S.T.; Jung, K.H.; Kim, B.G. Overexpression of PYL5 in rice enhances drought tolerance, inhibits growth, and modulates gene expression. Journal of experimental botany 2014, 65, 453–464. [Google Scholar] [CrossRef]
- Cheng, Z.J.; Zhao, X.Y.; Shao, X.X.; Wang, F.; Zhou, C.; Liu, Y.G.; Zhang, Y.; Zhang, X.S. Abscisic acid regulates early seed development in Arabidopsis by ABI5-mediated transcription of SHORT HYPOCOTYL UNDER BLUE1. The Plant cell 2014, 26, 1053–1068. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zheng, H.; Jin, L.; Xing, L.; Zou, J.; Zhang, L.; Liu, C.; Chu, J.; Xu, M.; Wang, L. miR169o and ZmNF-YA13 act in concert to coordinate the expression of ZmYUC1 that determines seed size and weight in maize kernels. The New phytologist 2022, 235, 2270–2284. [Google Scholar] [CrossRef]
- Sun, W.; Xu, X.H.; Li, Y.; Xie, L.; He, Y.; Li, W.; Lu, X.; Sun, H.; Xie, X. OsmiR530 acts downstream of OsPIL15 to regulate grain yield in rice. The New phytologist 2020, 226, 823–837. [Google Scholar] [CrossRef]
- Yan, Y.; Wei, M.; Li, Y.; Tao, H.; Wu, H.; Chen, Z.; Li, C.; Xu, J.H. MiR529a controls plant height, tiller number, panicle architecture and grain size by regulating SPL target genes in rice (Oryza sativa L.). Plant science : an international journal of experimental plant biology 2021, 302, 110728. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wen, H.; Teotia, S.; Du, Y.; Zhang, J.; Li, J.; Sun, H.; Tang, G.; Peng, T.; Zhao, Q. Suppression of microRNA159 impacts multiple agronomic traits in rice (Oryza sativa L.). BMC plant biology 2017, 17, 215. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, J.; Yan, J.; Gou, F.; Mao, Y.; Tang, G.; Botella, J.R.; Zhu, J.K. Short tandem target mimic rice lines uncover functions of miRNAs in regulating important agronomic traits. Proceedings of the National Academy of Sciences of the United States of America 2017, 114, 5277–5282. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shi, C.; Yang, T.; Zhao, L.; Chen, J.; Zhang, N.; Ren, Y.; Tang, G.; Cui, D.; Chen, F. High-throughput sequencing revealed that microRNAs were involved in the development of superior and inferior grains in bread wheat. Scientific reports 2018, 8, 13854. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Various signaling pathways involved in seeds development.
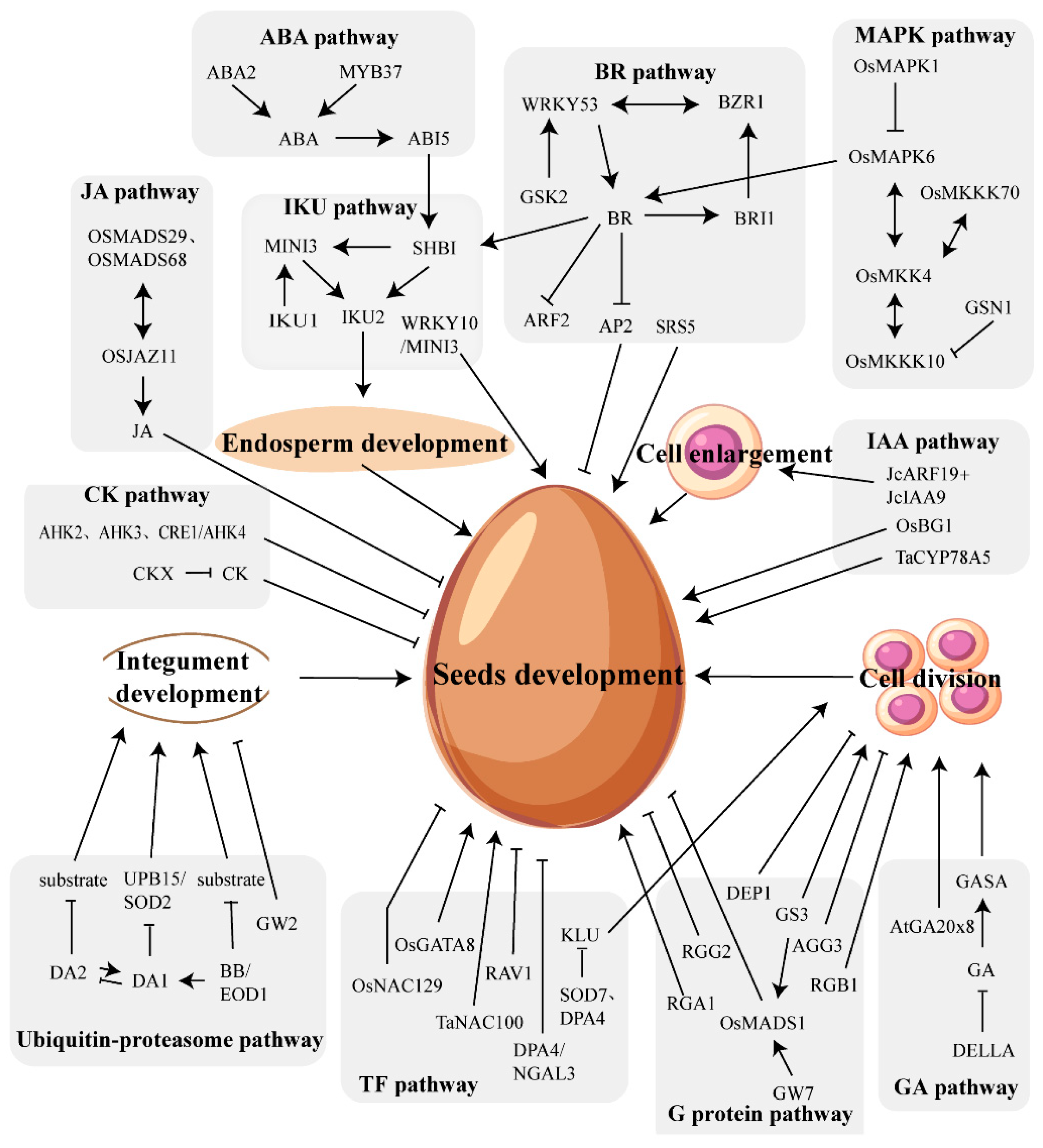

Table 1.
Regulatory factors involved in seeds development.
| Pathway | Protein | Species | Phenotype of loss-of-function | Phenotype of gain-of-function | Reference |
|---|---|---|---|---|---|
| IKU pathway | IKU1 | Arabidopsis thaliana | Small seeds | —— | [12] |
| IKU2 | Arabidopsis thaliana | —— | Big seeds | [13] | |
| MINI3 | Arabidopsis thaliana | Small seeds | —— | [14] | |
| SHB1 | Arabidopsis thaliana | Small seeds | Big seeds | [15] | |
| Ubiquitin-proteasome pathway | GhGW2-2D | Gossypium spp. | —— | Small seeds | [16] |
| AtDA1 | Arabidopsis thaliana | Big seeds | —— | [18] | |
| BnDA1 | Brassica napus | Big seeds | —— | [18] | |
| TaDA1 | Triticum aestivum | Big seeds | Small seeds | [19] | |
| SOD7 | Arabidopsis thaliana | —— | Small seeds | [20] | |
| GhDA1-1A | Gossypium spp. | —— | Big seeds | [21] | |
| DA2 | Arabidopsis thaliana | Big seeds | Small seeds | [22] | |
| UBP15 | Arabidopsis thaliana | Small seeds | Big seeds | [24] | |
| OsUBP15 | Oryza sativa | Small seeds | —— | [25] | |
| GW2 | Oryza sativa | —— | Small seeds | [25] | |
| G protein regulatory pathway | DEP1 | Oryza sativa | —— | Big seeds | [26] |
| GGC2 | Oryza sativa | —— | Big seeds | [26] | |
| GS3 | Oryza sativa | Big seeds | —— | [26] | |
| CGL1 | Oryza sativa | —— | Big seeds | [28] | |
| AGG3 | Arabidopsis thaliana | Big seeds | —— | [30] | |
| RGG2 | Oryza sativa | —— | Small seeds | [36] | |
| MAPK pathway | MPK10 | Arabidopsis thaliana | Big seeds | —— | [39] |
| OsMKK3 | Oryza sativa | —— | Longer seeds | [41] | |
| OsMKKK70 | Oryza sativa | —— | Longer seeds | [42] | |
| OsMKKK10 | Oryza sativa | Small seeds | Big seeds | [44] | |
| OsMKK4 | Oryza sativa | —— | Big seeds | [44] | |
| OsMKP1 | Oryza sativa | Big seeds | Small seeds | [45] | |
| Transcriptional regulators pathway | TaNAC100 | Triticum aestivum | —— | Big seeds | [48] |
| OsNAC129 | Oryza sativa | Big seeds | Small seeds | [50] | |
| TaWRI1L2 | Triticum aestivum | Small seeds | —— | [51] | |
| RAV1 | Arabidopsis thaliana | —— | Small seeds | [52] | |
| OsAL7.1 | Oryza sativa | Big seeds | —— | [53] | |
| OsAL11 | Oryza sativa | Big seeds | —— | [53] | |
| AIF2 | Arabidopsis thaliana | —— | Fewer seeds | [54] | |
| ZmBZR1 | Arabidopsis thaliana | —— | Big seeds | [55] | |
| ZmBES1/BZR1-5 | Arabidopsis thaliana | —— | Big seeds | [56] | |
| OsGATA8 | Oryza sativa | —— | Big seeds | [57] | |
| WRKY6 | Arabidopsis thaliana | Big seeds | —— | [58] | |
| MYB56 | Arabidopsis thaliana | Small seeds | Big seeds | [59] | |
| MYB37 | Arabidopsis thaliana | —— | Higher seed yield | [60] | |
| ZmARF12 | Zea mays | Big seeds | —— | [61] | |
| GIF1 | Oryza sativa | Small seeds | Big seeds | [62] | |
| OsGRF4 | Oryza sativa | Big seeds | —— | [63] | |
| Auxin regulatory pathway | OSYUC11 | Oryza sativa | Small seeds | —— | [64] |
| TaIAA21 | Triticum aestivum | Big seeds | —— | [65] | |
| TtARF25 | Triticum turgidum | Small seeds | —— | [65] | |
| TtERF3 | Triticum turgidum | Small seeds | —— | [65] | |
| TaCYP78A5 | Triticum aestivum | —— | Big seeds | [66] | |
| JcARF19 | Jatropha curcas | —— | Big seeds | [67] | |
| BRs regulatory pathway | ZmD11 | Zea mays | —— | Big seeds | [68] |
| D11 | Oryza sativa | Small seeds | Higher grain yield | [69] | |
| OsBZR1 | Oryza sativa | Small seeds | Higher grain yield | [69] | |
| CPB1 | Oryza sativa | —— | Big seeds | [70] | |
| TaD11-2A | Triticum aestivum | Small seeds | Big seeds | [71] | |
| D61-2 | Oryza sativa | Short seeds | —— | [72] | |
| SRS5 | Oryza sativa | —— | Big seeds | [73] | |
| DET2 | Arabidopsis thaliana | Small seeds | —— | [74] | |
| WRKY53 | Oryza sativa | —— | Big seeds | [75] | |
| GA regulatory pathway | GA2ox8 | Arabidopsis thaliana | —— | Higher seed yield | [77] |
| GASA4 | Arabidopsis thaliana | Small seeds | Big seeds | [78] | |
| JA regulatory pathway | COI1 | Arabidopsis thaliana | Big seeds | —— | [80] |
| MYC2 | Arabidopsis thaliana | Big seeds | —— | [80] | |
| MED25 | Arabidopsis thaliana | Big seeds | —— | [80] | |
| JAZ6 | Arabidopsis thaliana | Small seeds | —— | [80] | |
| OsJAZ11 | Oryza sativa | —— | Big seeds | [81] | |
| CK regulatory pathway | OsCKX2 | Oryza sativa | More grains | —— | [87] |
| AHK2/3/4 | Arabidopsis thaliana | Big seeds | Higher seed yield | [90,91,92] | |
| OsPUP1 | Oryza sativa | —— | Reduced grain weight and number | [83] | |
| OsPUP4 | Oryza sativa | —— | Big seeds | [93] | |
| OsPUP7 | Oryza sativa | Big seeds | —— | [94] | |
| ABA regulatory pathway | LOS5/ABA3 | Arabidopsis thaliana | —— | Higher seed yield | [96] |
| OsAO3 | Oryza sativa | Higher yield | Reduced grain yield | [95] | |
| OsPYL/RCAR5 | Oryza sativa | —— | Reduced grain yield | [97] | |
| ABA2 | Arabidopsis thaliana | Big seeds | —— | [98] | |
| ABI5 | Arabidopsis thaliana | Big seeds | —— | [98] | |
| miRNA regulatory pathway | zma-miR169o | Zea mays | Small seeds | Big seeds | [99] |
| OsmiR530 | Oryza sativa | Higher yield | Small seeds | [100] | |
| miR529a | Oryza sativa | Narrower grain | Narrower and longer grain | [101] | |
| miR159 | Oryza sativa | Small seeds | —— | [102] | |
| miR398 | Oryza sativa | Small seeds | Big seeds | [103] | |
| miR160 | Triticum aestivum | Small seeds | —— | [104] | |
| miR165/166 | Triticum aestivum | Reduced seed number | —— | [104] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



