
Submitted:
09 June 2023
Posted:
09 June 2023
You are already at the latest version
Abstract
Plants attained cellular homeostasis of phosphate (Pi) through an integrated response pathway regulated by different families of transcription factors including MYB, WRKY, bHLH and ZFP. The systemic response to Pi limitation showed the critical role played by inositol pyrophosphate (PP-InsPs) as signaling molecule and SPX (SYG1/PHO81/XPR1) domain proteins as sensor of cellular Pi status. Binding of SPX to PP-InsPs regulates the transcriptional activity of the MYB-CC proteins, phosphate starvation response factors (PHR/PHL) as the central regulator of Pi-deficiency response in plants. Vacuolar phosphate transporter, VPT may sense the cellular Pi status by its SPX domain, and vacuolar sequestration is activated under Pi replete condition and the stored Pi is an important resource to be mobilized under Pi deficiency. Proteomic approaches led to new discoveries of proteins associated with Pi-deficient response pathways and post-translational events that may influence plants in achieving Pi homeostasis. This review provides current understanding on the molecular mechanisms at the transcriptional and translational levels for achieving Pi homeostasis in plants highlighting the importance of DNA-protein and protein-protein interactions. The potential strategies for employing the CRISPR technology to modify the gene sequences of key regulatory and response proteins for attaining plant Pi homeostasis are discussed.
Keywords:
phosphate homeostasis
; transcription factors
; phosphate starvation response factors
; SPX
; proteomics
; post-translational
; CRISPR
; gene editing
1. Introduction
In over 70% of the world's cultivated land, phosphate (Pi) shortage is a significant limiting factor for crop growth and productivity [1]. Inefficiency in the uptake of phosphorus (P), the second most important macronutrients after nitrogen for plants is a major concern due to the depleting non-renewable Pi rock which is the main resource for P [2]. The situation is worsened by over-fertilising practices in acidic tropical soils as Pi, the P form utilised by plants is loss due to fixation forming insoluble Al and Fe precipitates not suitable for absorption by plants and furthermore run-off of excess P fertiliser damages the natural ecosystem and causes eutrophication [3]. In the rhizosphere, soils microbes convert inorganic Pi into organic form not suitable for acquisition by plants, however symbiotic interaction between plants and beneficial microbes such as arbuscular mycorrhizal fungi and phosphate solubilizing bacteria can help to enhance Pi acquisition efficiency [4]. Thus, development of crops with efficient Pi acquisition and enhanced P use efficiency (PUE) through breeding and biotechnology approaches could be a potential solution for the long term.
Pi is an essential component of biomolecules including nucleic acids, phosphorylated sugars, phospholipids and phosphoproteins and energy-rich compounds such as ATP and NADP. Thus, Pi plays a pivotal role in genetic heredity, membrane structure, cell energy currency, intracellular signaling, and metabolism [5,6]. It is essential for root development, photosynthesis and the entire reproduction process from fertilisation to fruit production [7]. Under limited intracellular P, plants mobilised P stored in vacuoles to the cytosol [8]. While phytic acid (InsP6) represents the storage form of P, inositol pyrophosphates (PP-InsPs) acts as highly charged signalling molecules in plants and other eukaryotic organisms with critical role in regulating cellular Pi homeostasis [9]. Essentially, plant cells are very well-equipped with rapid and dynamic response measures to face any uncertainties in Pi levels that cause imbalance affecting the normal functioning of the plant biological system.
Both Pi starvation and excessive Pi have harmful effects on plants. Cellular Pi homeostasis is achieved in plants through a series of adaptive responses that facilitates Pi acquisition from external environment, reduces Pi consumption and adjusts internal Pi recycling under inadequate Pi supply. Pi starvation leads to reduce plant growth and causes severe effect on young seedlings. Meanwhile, excessive Pi in cytoplasm caused serious cellular damage and growth retardation in plants [10]. Pi starvation responses in plants include reduction in photosynthesis, and accumulation of starch and anthocyanin. Anthocyanin which produces purple color on the stem, petiole, and leaves and also dark/green color in leaves provides protective role against abiotic stresses [11]. Plants can reduce their need for ATP and ADP and substitute non-phosphorus containing lipids such as galactolipids and sulfolipids for membrane phospholipids [12]. In barley, the low-P tolerant line and low-P sensitive lines were distinguishable based on their ability in P acquisition and utilization from the analysis of differential expression of genes involved in the growth of lateral roots, Pi transport within cells as well as from roots to shoots [13].
Plant Pi deficiency response involved signalling molecules and a repertoire of stress responsive genes such as high affinity Pi transporter 1 (PHT1), transporter traffic facilitator 1 (PHF1), purple acid phosphatases (PAP) and SPX (SYG1/PHO81/XPR1) [14,15]. Several transcription factors (TFs) have been identified to be involved in transcriptional regulation of Pi deficiency response including those from myeloblastosis (MYB) [16], WRKY [17], basic helix-loop-helix (bHLH) [18] and zinc finger protein (ZFP) [19] family of TFs. Gene expression or transcriptomics analysis in diverse crop species including perennial crops like oil palm [15,20] and Medicago sativa [21] and annual species such as soybean [22] as well as studies utilising mutant plants in Arabidopsis thaliana [23] and rice [24] showed the central roles played by phosphate starvation response (PHR) transcription factors.
Pi homeostasis relies on SPX domain proteins which also potentially act as sensors of Pi status. Under Pi replete condition, the SPX domain protein is present in a conformation that can bind PP-InsPs. This promotes SPX interaction with PHR and the resulting protein complex inhibits PHR in binding to phosphate starvation response 1 binding site (P1BS) motif found in the promoter regions of phosphate stress response genes (PSR) [25]. On the other hand, binding of PHR to P1BS activate expression of downstream PSR under Pi limited condition. Interaction between PP-InsPs and SPX that regulates the activities of the vacuolar transporter 1 (VPT1) that transports Pi into the vacuole is an important mechanism for Pi homeostasis [26]. Clearly in-depth studies to decipher molecular interactions especially between regulatory proteins and between protein and target DNA is essential for a better comprehension of the molecular mechanism for Pi deficiency response in plants.
The current development in precision genome engineering may potentially enable scientists to produce resource efficient crops [27]. CRISPR technology allows precise modification of gene sequence at specific genomic sites [28,29]. Since the universal mechanism for Pi deficiency response in plants is known, potential target genes for improvement and the specific nucleotides to be altered for potential enhancement of Pi uptake can be systematically evaluated. This review looks at the current understanding on the molecular mechanism on plant Pi deficiency response for identification of potential targets for maintaining plant Pi homeostasis. The different strategies for employing the CRISPR technology that can potentially be used exploiting on the knowledge of molecular regulatory mechanisms for improving Pi homeostasis in plants is elaborated.
2. Molecular Regulation of Phosphate Uptake at the Transcriptional, Translational and Post-translational Levels
2.1. Transcription Factors as Regulators of Systemic Responses to Phosphate (Pi) Starvation
Plants can monitor Pi starvation both at the local and systemic levels. Local signaling involves mechanisms that report the external Pi concentration and sensed at the root tip and responded through root system architecture (RSA) remodeling. Pi starvation activates partly the root apex endoplasmic reticulum (ER) stress-induced autophagy as a consequent of local Pi sensing and the associated modifications in root development. This ER stress-induced autophagy is unlikely to be required for systemic recycling of the macronutrient as previously thought [30,31]. Systemic or long-distance signaling mechanisms sense the internal Pi status of the whole plant. Systemic signaling governs internal Pi status for restoring Pi homeostasis through Pi recycling, uptake and transport [32]. Pi acquired from roots are translocated to shoots, before allocating to different tissues and organs. PHT1 family of transporters are involved in Pi uptake from the soil, translocation across tissues, and remobilization from senescent organs [33,34]. PHOSPHATE1 (PHO1) regulates the root-to-shoot phosphate transport. PHO1 mediates Pi efflux out of cells and Pi loading into the xylem [35]. The adaptive response of plant to Pi deficiency is largely control at the transcriptional level involving major families of TF including MYB, WRKY, bHLH and ZFP that regulate Pi homeostasis and signaling [17,18,36,37]. The promoter sequences of phosphate starvation inducible genes (PSI) such as EgPHT1 and BnPHO1 often harbor different Pi deficiency-responsive elements including the P1BS, W-box and E/G-box, the binding site for PHR (MYB family), WRKY and bHLH, respectively suggesting that these families of TFs are required for regulating and fine tuning their expression through a complex regulatory network [33,38].
The MYB family of TFs is classified into four types based on the number and position of imperfect repeats (R) involved in DNA recognition and has been reported to be involved in abiotic stress responses [39]. PHR and PHL (PHR-like) belonging to 1R-type MYB TF was shown to be the central regulator of systemic PSR in plants [18,40]. AtPHR1 and its rice homologue, OsPHR2 activate expression of a broad range of PSI including PHT1, PHF1, SPX, PAP that contain P1BS (GNATATNC) in their promoter sequences [14,41,42,43]. EgPHR2 expression in oil palm roots showed significant upregulation at low Pi and further increased in the absence of Pi and the expression profile correlated with P1BS containing EgPHT1;4 and EgPHT1;7 expression [20], while Pi-starvation inducible EgPHL7 was discovered through RNA-sequencing of root samples [15]. In Arabidopsis, the role of PHR1 as the major player in regulating Pi deficiency response is supported by other members with MYB-CC structural domain including PHL1, PHL2, PHL3 and PHL4 which demonstrated functional redundancy with PHR1 in Arabidopsis mutant functional knock-out and complementation studies [23,44]. OsPHR4 is a Pi starvation-inducible gene found in the nucleus and its expression is upregulated by OsPHR1/2/3 which can bind to the P1BS elements found in OsPHR4 promoter sequence and like in other plant species, the different OsPHR1 homologues, OsPHR1/2/3/4 regulate the expression of PSI in rice [45]. OsPHR3 is also responsive to different forms of N independent of Pi regimes [46] and is a potential candidate for studying crosstalk between P and N, the two major plant macronutrients. For their transcriptional activation activities, PHR and PHL bind to the DNA in dimeric form through formation of homodimers or heterodimers. Dimerization of Arabidopsis PHR1 is essential for high-affinity binding to P1BS [23]. It was shown that the binding affinity of OsPHR2 to P1BS is affected by variation in nucleotide sequence in the P1BS motif. Transgenic rice harboring tandem copies of P1BS with modified nucleotide motif sequences in the PHF1 promoter for enhancing the binding affinity showed significant improved tolerance to Pi deficiency. This strategy was demonstrated as an innovative way for engineering crop with improved tolerance to Pi deficiency [47].
The R2R3-type MYB members that have been described and reported to be involved in Pi signaling include AtMYB2, AtMYB62, OsMYB2P-1, OsMYB4P, and OsMYB5P [48,49,50]. MicroRNA399 (miR399) is an important regulator of phosphate (Pi) signaling and homeostasis through post-transcriptional mechanism in plants. Pi starvation-induced miR399 precursor, miR399f promotes Pi uptake by cleaving ubiquitin-conjugating E2 enzyme, PHOSPHATE2 (PHO2) mRNA, suppressing its expression [51,52,53,54]. It was demonstrated that AtMYB2 acts as a transcriptional activator for miR399f by binding to the MYB-binding site in the promoter of miR399f for regulating its expression. The temporal and spatial expression profile of AtMYB2 and miR399f is strongly correlated. Overexpression of AtMYB2 suppressed primary root (PR) growth but promoted development of root hairs suggesting its role in regulating PSR by activating miR399f transcript [55]. MYB62 was upregulated upon Pi starvation and repressed the expression of a number of the PSRs including gibberellin biosynthetic genes. The results of over expression of MYB62 affects the RSA, Pi uptake and acid phosphatase activity [48]. In Arabidopsis, AGB1, the β subunit of the heterotrimeric G protein is a versatile component of transmembrane signaling transduction pathways with critical role in gibberellin pathway in plants. Qi et al. 2021 demonstrated that the AGB1–MYB62 pair is involved in regulating the root growth process under Pi starvation. AGB1 inhibited the binding of MYB62 to the GA2ox7 (a gibberellic acid degradation gene) promoter, resulting in down-regulation of MYB62 activity and positively regulating the gibberellin pathway in Arabidopsis [56].
Among WRKY TFs, few of them function as positive and negative regulators involved in PSRs such as WRKY6, WRKY42, WRKY46 and WRKY75 that play a role in fine tuning of the Pi homeostasis in plants in Pi sufficient and deficient condition [17,48,57]. Devaiah and Raghothama (2007) reported that WRKY75 is upregulated under Pi starvation and positively regulates a number of PSRs, including phosphatase, MT4/TFS1-like genes and PHT1 [36]. A recent report by Li et al. 2021 showed that GmWRKY46 could directly bind to the W-box motif and activating the eukaryotic aspartyl protease family protein (ATAED1) expression [58]. Ectopic expression of GmWRKY46 in transgenic Arabidopsis enhanced tolerance to Pi starvation and improved root development. AtAED1 was one of the DEGs discovered through RNA-seq in the transgenic Arabidopsis overexpressing GmWRKY46. Subsequent functional characterization demonstrated that overexpression of AtAED1 could confer tolerance to Pi starvation in transgenic Arabidopsis. These results are very beneficial for molecular breeding to produce soybean that can grow under Pi deficient condition.
bHLH and zinc finger containing TFs were also reported to be involved in Pi starvation responses and modulating RSA [18]. The TF bhlh32 is a negative regulator of PHR and its Arabidopsis mutant showed significantly increased expression of PSI, more anthocyanin and Pi accumulation and root hair formations under Pi-sufficient condition compared to the wild type Arabidopsis [59]. Over expression of the bHLH TF, B-BOX32 (BBX32), and the mutation of ELONGATED HYPOCOTYL 5 (HY5), which is down-regulated by BBX32, resulted in the tolerance to Pi deficiency in Arabidopsis [60]. HY5 (bZIP TF) binds directly to the promoters of light-inducible genes, to promote their expression for photomorphogenic development. Slight inhibition of the PR lengths of 35S:BBX32 and hy5 plants were observed under Pi deficient condition while the fresh weights were significantly higher in comparison with wild type. Their results suggest that BBX32 and HY5 are involved in the regulation of PSRs and under Pi deficient condition, B light may regulate PR growth inhibition.
ZAT6, a TF of a zinc finger family serves as a negative regulator of Pi starvation response. It is induced upon Pi starvation and ectopic expression of this gene inhibits the transcripts of many PSI and attenuates the PR growth [19]. In Arabidopsis, C2H2 TF ZFP5 is an important regulator of trichome and root hair development. ZFP5 increased root hair elongation under Pi and potassium deficient condition through activating the expression of ETHYLENE INSENSITIVE2 (EIN2), a key regulator of ethylene signaling [61]. The recent examples involving bHLH and zinc finger TF showed the different levels of regulation of gene expression and the regulatory network responding to different abiotic stresses and signaling molecules. Some TFs acting at the upstream position and they are involved in regulating the expression of other TF(s) under Pi starvation to fine tune PSI expression. Table 1 provides a summary of the transcription factors and other stress-responsive signalling pathways under Pi starvation that have been discussed.
2.2. New Insights for Plant Phosphate Starvation Response at the Translational Levels
Proteomics can be used to show a wide range of species-specific metabolites that alter energy metabolism as well as carbon and nitrogen assimilation inside the root cells, which in turn changes the morphology and metabolism of roots that influence Pi uptake. For the development of sustainable agricultural strategies, an understanding of the mechanisms behind plant PUE in crops is essential.
Under P-limiting growth conditions, it is generally accepted that root-secreted acid phosphatases and organic anions which are key players in mobilizing plant P from the rhizosphere. Previous studies suggested that additional mechanisms are involved, particularly in species with fine roots like wheat. Staudinger et al. 2022 profiled root tip mucilage, soluble root secretion, and root tip proteomes in order to characterise the plant-derived extracellular proteome of wheat roots. Selected reaction monitoring was used to focus on extracellular acid phosphatases and enzymes involved in the central carbon metabolism [62]. Mucilage contains more than 140 proteins with predicted extracellular localization. In both root tip and soluble root-secreted proteomes, P starvation-induced proteins that are projected to be localised to the apoplast which are associated with cell wall modification and defence. P limitation in root tips significantly enhanced the quantity of glycolytic enzymes as well as phosphoenolpyruvate carboxylase (PEPC) and plastidial malate dehydrogenase (MDH). There were no soluble acid phosphatases found in samples of extracellular proteins. It was suggested that proteins with the functional capacity to actively influence their immediate environment by altering biotic interactions and plant structural elements exist in the mucilage of root tip. A small role involving acid phosphatases synergizing with rhizobacteria appears to be required in the mobilisation of P from the immediate root surface during Pi starvation condition in rice.
A recent study by Yu Zhang et al 2022 involved the PHO proteins which are known to transport inorganic Pi in plant cells [63]. Two co-orthologs of the PHO1 gene in
Arabidopsis are encoded by the maize (Zea mays ssp. mays) genome and are known as ZmPho1;2a and ZmPho1;2b. Allele Zmpho1;2a0 -m1.1; the transposon footprint, also known as pho1;2a, has been discovered by Yu Zhang et al 2022. In this study, the ZmPho1;2a gene's activator transposable element was removed to create the stable derivative known as the pho1;2a allele. Yu Zhang et al demonstrated that the Pho1;2a allele is associated with mildly reduced seedling growth and a dosage-dependent reduction in Pho1;2a transcript accumulation. A total of 1100 differentially expressed genes between wild-type plants and plants bearing the pho1;2a mutation were found when the shoot and root transcriptomes were characterised under full nutrition, low nitrogen, low phosphorus, and combined low nitrogen and low phosphorus conditions. The fact that 966 of these 1100 genes were elevated in plants expressing PHO1;2a suggests that the wild-type PHO1;2a mostly affects negative gene regulation. Yu Zhang et al suggested that Pho1;2a play a vital role in modifying the transcriptional Pi starvation response, according to the gene set enrichment analysis of the transcripts that are misregulated in pho1;2a mutants.
Quantitative proteomics approach was used to identify proteins involved in the response to Pi starvation in soybean leaves [64]. In order to adapt to Pi shortage, soybean plants use a number of strategies in response to Pi starvation. The underlying biochemical pathways causing the Pi starvation response are, however, still completely unclear. Cheng et al 2021 analysed soybean leaves grown in low and high Pi environments using label-free quantification (LFQ) [65]. From Pi-starved leaves, they discovered 267 induced and 440 decreased differentially expressed proteins. Pi starvation increased proteins were accumulated in the carbon metabolic and chlorophyll biosynthetic processes, and over a quarter of the Pi starvation decreased proteins are involved in translation activities. Only the GmENO2a and GmENO2b genes have the P1BS motif in their promoter regions, out of the 14 enolase genes. Additionally, GmENO2b was particularly increased in the soybean plants overexpressing GmPHR31. Alternative splicing (AS) events in rice as reported by Guo, M., et al 2022 revealed Regulator of Leaf Inclination (RLI1) resulting in two proteins isoforms known as RLI1a and RLI1b; both containing MYB binding domain [66]. Both proteins can adjust the Pi starvation signaling. It was also found that AS event involving RLI1-related genes is abundant in plants. Therefore, the AS event is an important and functionally preserved method to coordinate Pi starving signalling and growth to assist plants in coping with stress caused by Pi starvation. These studies provide new insights into the molecular mechanisms underlying plant responses to nutrient stress.
Muneer et al 2015 evaluated the proteome of tomato leaves treated with and without Pi for 10 days in order to ascertain the adaption and homeostatic response to Pi deficiency [67]. On two-dimensional gels, 46 out of 600 reproducible proteins showed differential expression. Major metabolic processes involving these proteins included photosynthesis, transcriptional and translational controls, glucose and energy metabolism, protein synthesis, immune response, and other secondary metabolisms. The outcomes also demonstrated that under -Pi treatments, the P content was decreased due to the decrease in photosynthetic pigments. Additionally, under -Pi treatments, increased levels of the high-affinity Pi transporters (lePT1 and lePT2) were observed. Additionally, in the epidermis and palisade parenchyma under +Pi treatments as opposed to -Pi treatments, a greater concentration of Pi transporters was seen. According to the findings, tomato plants evolved reactive oxygen species (ROS) scavenging methods to deal with low Pi concentration, including the up-regulation of proteins that are mostly involved in significant metabolic pathways. By expanding the Pi transporter genes and their logical localisation, Pi-starved tomato plants also improved the efficiency of their internal Pi utilisation. Thus, these findings offer crucial information regarding the homeostasis and responses of tomato plants to Pi shortage.
The complex trait of P deficit tolerance in rice is regulated by many genes involving different pathways. In the distinctive Pi starvation tolerance Dongxiang Wild Rice (Oryza Rufipogon, DXWR) germplasm, low P tolerance-related proteins were identified using proteomics analysis, which would serve as the foundation for the investigation of its regulatory mechanism. To find potential novel low Pi response genes in DXWR at seedling stage, proteomics with joint analysis of transcriptome data were carried out. The findings revealed 3589 significant differential accumulation proteins between the DXWR root samples treated with low P and normal P. A total of 60 up-regulated and 15 down-regulated proteins were among those with expression changes of more than 1.5 times, and 24 of them had expression changes of at least 1.5 times in the transcriptome data. Seven genes that correspond to the significantly differentially expressed proteins discovered in this study were unknown and distributed in the low P tolerance QTLs interval. Two of these genes, LOC Os12g09620 and LOC Os03g40670, were found to be present at both the transcriptome and proteome levels. The thorough investigation revealed that in dealing with low P stress condition, DXWR employs various strategies which include enhancing the the expression of PAPs, modifying membrane location of P transporters, rhizosphere area, and AS, and decreasing ROS activity [68]. This research will provide some light on the molecular mechanisms underlying low P resistance in DXWR and enable cloning of the P-deficit tolerance genes from wild rice.
2.3. Roles of Phosphorylated Proteins and Phospholipids in Pi Homeostasis
Plants will take the necessary precautions when Pi levels are low, such as developing an internal defence system to defend themselves from harmful consequences. Using methods like phosphoproteomics, phosphorylation events that affect Pi absorption can be found at the post-translational level. This entails enriching phosphorylated proteins, followed by the identification and measurement of phosphorylation sites using mass spectrometry. Specific phosphosites and phosphoproteins that react to variations in Pi availability can be identified by examining the phosphorylation patterns of cells or tissues under various Pi conditions [69]. Transporters, enzymes, and signalling molecules involved in Pi absorption, utilisation, and homeostasis can be found in these phosphoproteins [70]. Overall, phosphoproteomics offers a potent tool for comprehending the post-translational regulation of Pi absorption.
Soybean (Glycine max) is an important economic legume crop and a valuable source of protein [71,72]. It is well known that soybean adapts to Pi starvation by changing its physiological and morphological characteristics, including enhancing its symbiotic relationship with rhizobia and arbuscular mycorrhizal fungi, increasing malate exudation and root-associated PAP activities, and altering its root system [73,74] The molecular processes driving soybean responses to P deficiency in reversible protein phosphorylation are still unknown. A study by Jiang et al. 2021 employed iTRAQ in conjunction with LC-MS/MS analysis to identify a set of differential phosphoproteins in soybean roots at two Pi levels [75]. Based on the result, 542 phosphopeptides were found, and projected to be involved in a number of biological pathways. Additionally, altered phosphorylation levels of GmNR4, a nitrate reductase that was upregulated in response to Pi deprivation, had a significant impact on its in vitro activity but not on its subcellular location. The findings provided fresh insights into the molecular mechanisms driving plant adaptation to P deficit and strongly supported the complex regulatory networks present in soybean in response to P deficiency through reversible protein phosphorylation. Overall, phosphoproteomics offers a potent tool for comprehending the post-translational regulation of phosphate absorption.
Plants under Pi starvation have an adaptation mechanism called membrane lipid remodelling: mainly comprises of two primary mechanisms; hydrolysis of phospholipids and the synthesis of glycolipids (Figure 1). In several species, including Arabidopsis [76], rice [77], tobacco [78], camelina [79], and soybean [80], sets of key genes or proteins that contribute to the decrease in phospholipids and increase in non-phosphorus lipids in membrane lipid remodelling have been reported under low-P stress conditions. A perspective in metabolic adaptation during Pi starvation involving lipid remodelling by glycerophosphodiester phosphodiesterases (GDPD6) detected in Arabidopsis roots was performed [81]. PHR1 which is known to be essential for maintaining Pi homeostasis, controls the expression of GDPD6. Research by Ngo and Nakamura 2022 showed that mutant plants lacking GDPD6 had altered root morphology in comparison to wild-type of Arabidopsis thaliana [81]. This allowed the researchers to evaluate the role of GDPD6 in Arabidopsis root growth. Particularly, the mutants lacking GDPD6 had shorter primary roots and less lateral root growth. This shows that GDPD6 controls root growth when there is Pi starvation. Further research found that GDPD6 controls the amounts of specific lipid molecules in the root tissues, particularly glycerophosphodiester complexes. During Pi starvation, it is believed that this lipid modification is crucial for maintaining normal membrane integrity and cellular functions. The study also shows that genes involved in Pi transport and metabolism have their expression levels changed when GDPD6 is disrupted. This suggests that the involvement of GDPD6 in coordinating Pi homeostasis and signalling pathways in Arabidopsis roots may be more extensive. The findings of this work offer important new understandings of the molecular processes underpinning Arabidopsis roots' response to Pi starvation.
In response to Pi starvation, plants slow down shoot growth while accelerating root growth, to improve Pi uptake from the soil. Membrane lipid remodelling is a metabolic adaptation that allows plants to get limited Pi while maintaining the membrane structure by swapping out membrane phospholipids for non-phosphorous galactolipids. Although there are documented metabolic routes that convert phospholipids to galactolipids, it is still unclear how phospholipid production is involved in this process. However, study involving lysophosphatidic acid acyltransferase (LPAT2) overexpression showed an increase in the number of phospholipids in the root membranes, particularly phosphatidylcholine (PC) and phosphatidylethanolamine (PE). This rise in phospholipids resulted in enhanced cellular and membrane integrity, facilitating better adaptability to Pi shortage during Pi starvation. The results demonstrate the beneficial effects of LPAT2 overexpression in boosting de novo phospholipid production on root development in Pi-starved Arabidopsis plants. Under Pi-limiting conditions, it may be possible to improve plant development by manipulating the production of phospholipids. This investigation clarifies the function of phospholipids in plant responses to Pi stress and suggests possible directions for enhancing crop nutrient uptake [82]. In addition, degradation of phospholipid during
During phosphate starvation, the limited availability of phosphate triggers a series of events that lead to the remodeling of lipid membranes. Sensor proteins located in the membrane detect the scarcity of phosphate and initiate signaling pathways. This remodelling is influenced by the expression of LPAT2. An enzyme called LPAT2 is involved in the manufacture of phospholipids, which are a crucial part of biological membranes. Signalling pathways are activated in response to phosphate deficiency, altering the patterns of gene expression. The expression of LPAT2 has increased, which is one of the main results. By increasing LPAT2's enzymatic activity as a result of this overexpression, phospholipid production is made easier. As a result, the lipid membrane's structure and composition change, possibly improving the fluidity and functionality of the membrane in phosphate-deficient settings.
Membrane lipid remodelling during Pi starvation was performed by Ngo et al 2022 [83]. They reported that Pi starvation transcriptionally activates the enzymes phospho-base N-methyltransferases 1 and 2 (PMT1 and PMT2), which change phosphoethanolamine into phosphocholine (PCho). Upon phosphate deprivation, the shoots of Arabidopsis thaliana seedlings with the PMTpmt1 PMTpmt2 double mutant exhibited poor growth; however, membrane lipid profiles were unaffected. It was discovered that Pi deprived PMTpmt1 and PMTpmt2 had reduced PCho content and exhibited abnormal leaf growth and the abnormal growth was corrected by exogenous PCho supplementation. It was suggested that Pi starvation causes PMT1 and PMT2 to synthesise PCho primarily for maintaining leaf development rather than for PC production in membrane lipid remodelling. The results imply that PMT1 and PMT2 are important enzymes involved in phospholipid production and are crucial for preserving leaf development in Arabidopsis under Pi-starved conditions.
3. SPX PP-InsPs in Regulating Cellular Pi Homeostasis in Plants
Inositol is a small, water soluble and versatile intracellular signalling molecules based on a six-carbon ring structure, which is cis-1,2,3,5-trans-4,6-cyclohexanehexol. PP-InsPs, the high-energy derivatives of inositol have fully phosphorylated inositol ring containing one or two pyrophosphate moieties at the C3 and C5 positions [84]. PP-InsPs binds to the small hydrophilic SPX domain that are involved in regulating Pi homeostasis. Recent reports suggest that the InsP8-SPX signalling module may be an evolutionarily conserved mechanism in eukaryotic organisms for Pi sensing [25].
3.1. Phosphorylated Inositols as Signalling Molecules and Roles of Inositol Polyphosphate Kinases for Cellular Homeostasis of Pi in Plants
PP-InsPs are synthesized from phosphorylation of InsP6 [85]. Identification of Arabidopsis inositol 1,3,4-trisphosphate 5/6-kinases, ITPK1 and ITPK2 as putative novel plant InsP6 kinases from in vitro assays was reported [86,87]. ITPK1 stimulates 5-InsP7, InsP8 synthesis and regulates phosphate starvation responses in planta in Pi-dependent manner [88]. Two highly homologous diphosphoinositol pentakisphosphate kinases (VIH1 and VIH2) catalyze the reversible reactions of InsP8 synthesis and hydrolysis. This enzyme referred to as VIH or VIP possesses both kinase and phosphatase domains at the N and C terminal, respectively. Changes in cellular ATP and Pi concentrations are relayed by VIH1 and VIH2 to changes in PP-InsP levels, enabling maintenance of sufficient Pi levels in plants. Phosphorylation of InsP7 to produce InsP8 occurs under high cellular Pi when there is sufficient supply of Pi. Conversely, when the cellular Pi level drops during Pi starvation VIH switch on phosphatase activity and InsP8 is converted to InsP7 and InsP6 [89].
Phenotypic analysis of vih1/vih2 double mutant with disrupted kinase activities in both alleles exhibited reduction of InsP8 and over-accumulation of Pi due to impaired synthesis from InsP7. The growth of the mutant plants was affected and shorter roots and smaller leaves were displayed. Increased expression of PSRs were observed even under Pi replete condition [25,89]. The complexity of the signalling cascades involving InsP8 was observed by Land et al., 2021 which demonstrated that under Pi-limiting condition, both local and systemic responses to Pi limitation are suppressed in the vip1/vip2 double mutant with modest reduction in InsP8 [90]. The mutant plants exhibited shorter root hairs and lateral roots and less accumulation of anthocyanin, sulfolipids and compared to wild type galactolipids plants. However, no effects were observed on the expression of PSRs. The results provided an insight on the complex regulatory mechanisms linking inositol phosphates and pyrophosphates in homeostasis of phosphate in plants. Mutants that block the synthesis of PP-InsP precursors including mutants affecting inositol pentakisphosphate 2-kinase 1 (IPK1) that converts InsP5 into InsP6 and ITPK1 exhibited similar Pi-related phenotype with decreased levels of InsP6 and InsP7. The mutants also showed an increase in the D/L-Ins(3,4,5,6)P4 level. Analysis of InsPs profile in these mutants revealed a correlation between elevated D/L-Ins(3,4,5,6)P4 level and activation of Pi uptake and PSR expression. The study suggests that both IPK1 and ITPK1 play important roles in maintenance of Pi homeostasis under Pi replete condition [91].
Laha et al. 2015 and Glubani et al. 2022 [92,93] identified InsP8 as the common moiety for transducing both jasmonic acid (JA) signaling and PSR and could serve as a versatile signaling molecules for diverse stress exposures. Arabidopsis thaliana ipk1, itpk1 or vih2 mutants showed suppression in basal salicylic acid (SA)-dependent immunity during phosphate starvation. Their results suggest likely diversion to PSR-supportive from defense-promoting roles in the mutants. Upregulated PSR expression could in turn support demand for Pi to support heighten basal defense requirement. The findings demonstrate the essential role played by InsP8 in mediating crosstalk in defense-Pi homeostasis involving different cellular pathways during adaptation to various stress conditions.
3.2. Molecular Mechanism Involving SPX in Regulating Transcriptional Activities of PHR
AtPHR1 and OsPHR2 positively regulate phosphate homeostasis and hairy roots development. In Arabidopsis and rice, AtPHR1/OsPHR2 transcriptional regulatory activity is suppressed through protein-protein interaction by their downstream target, the SPX proteins. Control of PHR transcriptional regulatory activity by SPX is achieved through controlling the translocation of PHR into the nucleus from the cytoplasm based on studies on interaction of rice PHR2 and SPX4 [94]/ SPX6 [95] or through inhibiting PHR from binding to P1BS promoter motif found on PSR through blocking PHR binding site through protein-protein interaction as demonstrated by SPX1 and SPX2. Studies in Arabidopsis and rice showed functional redundancy of nuclear localized SPX1 and SPX2 [96,97] which act as Pi-dependent inhibitors of PHR serving as Pi sensors. SPX4 which is mainly localised in the cytoplasm potentially plays the predominant role in inhibiting translocation of PHR2 while SPX6 is evenly distributed in both the nucleus and cytoplasm. Acceleration of SPX4 degradation under Pi deficiency through the 26S proteasome pathway, releases and enables PHR2 to be translocated into the nucleus and activating the expression of PSI [94]. Qi et al. 2017 demonstrated that under Pi sufficient condition, AtSPX1 binds to monomeric AtPHR1 regulating PSI expression by tuning the AtPHR1-DNA-binding equilibrium [98]. Regulation of AtPHR1-DNA-binding equilibrium which is Pi-dependent, tightly controls PSI gene expression through a negative feedback loop on the expression of AtSPX. Over expression of BnaA2.SPX1 transgenic Arabidopsis suppressed the expression of PSR genes including PHTs, PAP10, and miRNAs supporting its role as a negative regulator in the Pi signaling network [99], consistent with the findings on nuclear-localised Arabidopsis SPX1 which harbours P1BS in its promoter sequence [96].
Direct binding of PP-InsP to SPX domains exerts control on the different cellular components to maintain Pi homeostasis. Under high intracellular Pi, the formation of PHR-SPX complex is enhanced which inhibits PHR from inducing expression of PSR [25]. The basic surface of the SPX domain binds to the negatively charged Pi at low affinity but demonstrates high affinity binding to PP-InsPs [100]. Dong et al 2019 provided evidence suggesting InsP8 as the intracellular Pi signalling molecule serving as the ligand of SPX1 for controlling Pi homeostasis in plants [25]. SPX1 and PHR1 interaction is dependent on the binding of lnsP8 to the SPX domain which occurs under high intracellular Pi that promotes the synthesis of InsP8 by VIH1 and VIH2. This blocks PHR1’s transcriptional activation of PSR. Under limited intracellular Pi, expression of PSR such as genes encoding PHT1 is activated because the PHR is separated from SPX that inhibits its transcriptional regulatory activity.
Reid et al. 2021 which studies the crystal structure of the CC domain of PHR1 showed that InsP8-SPX complex targets the unique plant PHRs CC domain [101]. The unusual four-stranded anti-parallel arrangement of the CC domain of PHR1 was altered to a monomeric form through introduction of interface mutations. Interaction with the SPX receptor both in vitro and in planta was disrupted when basic residues located at the surface of the CC were mutated, leading to constitutive Pi starvation responses. When InsP8 levels are reduced, under Pi starvation, dissociation of the SPX–PHR complexes enables the TFs to acquire the oligomeric state essential for high affinity binding to the promoter. Modulating PP-InsP levels or the affinity/specificity of the SPX domain for PHR could be a potential strategy to engineer crops to maintain high yield under reduced Pi fertilizer input. Steric hindrance prevents the SPX bound PHR TF from retaining the dimeric form essential for interacting with the promoter elements of PSI [102]. It was shown that the monomeric SPX1-PHR2 complex triggered by InsP6 resulted in PHR2-DNA binding inhibition. SPX1 disrupts PHR2 dimers forming a 1:1 SPX1-PHR2 complex upon binding of InsP6. The complex structure reveals that the α1 helix of SPX1 stabilized by InsP6 can impose a steric hindrance when interacting with the PHR2 dimer. It leads to allosteric effects that stabilizes the SPX1-PHR2 through decoupling of the PHR2 dimer. This enables SPX1 to interact with the PHR2 MYB domain blocking it from binding to the PSR promoter motif. This suggests that SPX1 suppresses PHR2 via attenuation of PHR2 dimerization and the PHR2 TF DNA binding activities [103].
SPX4 functions as a negative regulator not only of PHR1-dependent but also of PHR1-independent responses. SPX4 may also interact with other TFs besides PHR1, such as suppressor of overexpression of CONSTANS1 and Arabidopsis NAC DOMAIN CONTAINING PROTEIN55 involved in regulating shoot development. However, in the roots, transcriptional regulation of P status is independent of SPX4 [104]. SPX4 interacts with the MYB TF, PAP1 that regulates the anthocyanin biosynthetic pathway, in an InsPs-dependent manner. SPX4 acts as a negative regulator of anthocyanin biosynthesis under Pi sufficient condition through formation of SPX4-PAP1 complex that prevents PAP1 from binding to the promoter of dihydroflavonol 4- reductase, thus suppressing anthocyanin biosynthesis. During Pi starvation, PAP1 was released from SPX enables activation of anthocyanin biosynthesis. In addition, the phr1spx4 mutant studies indicated PAP1 expression is induced in a PHR1-dependent manner under Pi starvation, promoting anthocyanin production [103].
3.3. Regulatory Mechanism Involving SPX in Controlling Vacuolar Phosphate Transporter (VPT) Activities for Plant Pi Homeostasis
Vacuoles play important role in cellular Pi homeostasis and play dual role as source and sink of Pi. The vacuoles serve to buffer Pi concentrations in the cytoplasm against fluctuations in external Pi. Low Pi induced the expression of SPX-MFS domain-containing genes in different species such as Elaeis guineensis [15] and Brassica napus [99] even though different genes may respond differently in different tissues and at different duration following Pi scarcity treatment. Under severe Pi starvation, VPT (PHT5), the root vacuolar influx transporter that harbours SPX-MFS domain showed significant upregulated expression and there was a significant increase in the total P content in the roots [15] of oil palm seedlings indicating that the seedlings have started mobilising Pi from other tissues. This suggests that the oil palm VPT is involved in Pi homeostasis after prolonged Pi scarcity stress, to respond to the changes in cellular Pi. Both Arabidopsis and rice have three VPT genes (VPT1, 2 and 3). It was demonstrated that Pi accumulation in vacuoles is impaired under high Pi status in plants lacking VPTs thus, compromising plant ability to cope under low Pi. In Arabidopsis, vpt1 vpt3 double mutant shows more severe defects than the vpt1 single mutant in Pi homeostasis, [8] indicating VPT3 plays important supportive role to VPT1 in protecting the plant cells from Pi toxicity. Distinct and conserved roles in cellular Pi homoeostasis of two Brassica napus BnPHT5 in comparison to the homologous gene in Arabidopsis were revealed through functional characterization of their mutants [105]. The vacuolar Pi sequestration capacity was severely impaired in rice VPT double and triple mutants resulting in increased Pi allocation to aerial organs. Alteration of vacuolar Pi sequestration could be a novel and effective strategy to improve tolerance to low phosphorus in rice [24].
It was found that VPTs’ role in vacuolar Pi sequestration is linked with fine tuning of the systemic Pi homeostasis during the reproductive stage. Under sufficient Pi, impaired reproductive development with shortened siliques and abnormal seed set was observed in vpt1 vpt3 double mutant, with over-accumulation of Pi detected in the floral organs [8]. This suggests more Pi is allocated into the reproductive organs when the transport activities of VPT1 and VPT3 are impaired enabling long-distance control of Pi transport through regulating vacuole transporter. Genetic manipulation of the transport activities of VPT-type proteins in crops could be used as a strategy for low-Pi containing grains due to their involvement in systemic Pi distribution. In order to reduce seeds total P content, Sun et al. 2023 genetically regulated VPT1 during the flowering stage and found that P content in seeds can be reduced without detrimental effects on production and seed vigor [106]. Downregulation of Pi accumulation into the grain would potentially decrease phytate in seeds and enhance grain filling for rice production.
The transport capabilities of many transporters are activated by covalent modifications and/or binding of ligands that lead to conformational change that produce the active form [107]. Detailed structure-function analyses suggests that under low cellular Pi, Arabidopsis VPT1 remains in the auto-inhibitory state. Binding of InsPs to the SPX domain at the N-terminal of VPT1 under high cellular Pi switch on VPT1 activity for transporting Pi into the vacuole. Mutation of key residues involved in ligand binding in the SPX domain severely impaired the transport activity of VPT1 [108]. The coupling of Pi status to vacuolar sequestration could potentially be an important mechanism for maintaining Pi homeostasis in plant cells. Other important players in the maintenance of Pi homeostasis are vacuolar Pi efflux transporters (VPE). The VPT and VPE transporters enable vacuolar Pi storage to be managed based on intracellular Pi status [109] (Figure 2).
4.0. CRISPR-mediated genome editing for improving phosphorus use efficiency
The advent of genome editing technologies in the past decade has enabled desirable changes be made to economically important traits in agriculture through precise genetic modifications of the plant genome. Meganucleases, zinc finger nucleases (ZFNs), and transcriptional activator-like effector nucleases (TALENs) are examples of first-generation site-specific nucleases (SSNs), while clustered regularly interspaced short palindromic repeat/CRISPR-associated system (CRISPR) has garnered more attention as the second generation due to its ease of use, efficiency, and cost effectiveness [28].
All of the aforementioned genome editing systems share the same underlying principle of producing DNA double-strand breaks (DSBs) at a specific genomic site, which are then repaired by plant endogenous repair mechanisms using either non-homologous end joining (NHEJ) or homology-directed repair (HDR) [110]. While the majority of DSBs are repaired back to their original state, sometimes the repair is imperfect, resulting in DNA modifications such as substitution and insertion/deletion (indels) [111]. The changes in
Under high intracellular Pi and ATP, the signalling molecule InsP8 is produced in plant cells by VIH1 and VIH2. InsP8 binds to SPX that acts as intracellular Pi sensor. The InsP8 bound SPX (SPX4/ SPX6) binds to the CC domain of PHR/PHL. The formation of PHR-SPX complex in the cytoplasm inhibits PHR translocation into the nucleus. While binding of SPX (SPX1/ SPX2) to PHR in the nucleus disrupts formation of PHR/PHL homodimers or heterodimers dimers essential for high affinity binding to the P1BS motif found in PSI promoters, suppressing the expression of the PSI. InsP8 binds to SPX in the SPX-MFS domain found in VPT. This activates the Pi influx transport activity of VPT1 and VPT3, enables excess Pi to be stored in the vacuoles.
Under Pi deficiency, SPX-PHR complex dissociates enables PHR to be translocated into the nucleus. In the nucleus PHR/PHL homodimers or heterodimers are formed. The MYB domains of the dimeric protein are involved in high affinity binding with the P1BS motif of PSI in the nucleus, activating the expression of PSI such as high affinity PHT1, PHF1, SPX and PAP to respond to the Pi deficient condition. SPX in the SPX-MFS domain found in VPT is free from InsP8, this inactivates the Pi influx transport activity of VPT.
DNA sequences may alter gene function, in which some of them code for proteins and carry instructions for particular traits, resulting in the modification of physical characteristics.
Since its introduction, CRISPR/Cas9-mediated targeted gene knock out has been routinely used to functionally characterize candidate genes that may have potential roles in molecular mechanisms underpinning various key traits for future crop improvement. Although mutant lines generated by T-DNA insertion and RNAi-mediated gene silencing have been conventionally used for deciphering the functions of genes in previous studies, their wide application is often hampered by technical limitations such as unwanted chromosomal rearrangement [112], possible adverse effects on the expression of the neighboring genes [113], variable levels of knock down efficacy, and unpredictable off-target effects [28].
The mutation of MYB1 in rice using CRISPR/Cas9 resulted in enhanced Pi uptake and accumulation, as well as altered expression of a subset of genes associated with Pi transporters and Pi starvation signaling [114]. To provide insights into the gene-regulatory networks governing phosphate homeostasis in crop plants, CRISPR/Cas9 technology can be employed to study the various phosphate transporters and transcription factors through targeted mutagenesis of amino acid residues. Through deletion mutants generated by CRISPR, the Pi transport role of AtPHO1 homolog in tomato, SlPHO1;1 was confirmed as demonstrated by typical Pi starvation responses including decreased shoot fresh weight, increased root biomass and root-to-shoot ratio as previously shown for Arabidopsis [115]. Apart from SlPHO1;1, there are another five AtPHO1 homologs identified in the tomato genome, which may play a similar role in Pi transport. Rapid mutant generation using CRISPR should provide some insight into their transport function.
When plant sensed low Pi in the soil, morphological and biochemical adaptive responses are initiated, such as alteration of RSA [116,117] and enhancement of fine root structures [112], in which the hormone ethylene plays a pivotal role during this process. Using CRISPR, the roles of two members of the OsACS family; OsACS1 and OsACS2 in ethylene biosynthesis were studied during low Pi conditions in rice. The results showed that editing of both genes resulted in several Pi deficiency-induced adaptive responses demonstrated as altered RSA [118]. It has been hypothesized that involvement of ethylene in stimulating lateral root elongation during low Pi levels in rice is possibly through crosstalk with other plant hormones such as cytokinin. Identification followed by characterization of candidate genes involved in this crosstalk using CRISPR may shed light on Pi-induced lateral root development in rice. In addition, a pair of rice vacuolar Pi efflux transporters have been identified in a recent study, namely OsVPE1 and OsVPE2. The CRISPR/Cas9 double mutant of these genes appeared to accumulate higher vacuolar Pi content under low Pi stress than the wild type [119].
Apart from transporters, TFs represent promising targets for improving phosphate uptake in plants owing to their role in modulating plants’ responses through controlling growth and development. Functional studies and editing of SPX-domain containing proteins using CRISPR may open up a new avenue for breeding efforts to generate low-Pi tolerant crop varieties. Another negative regulator of Pi signaling is PHO2, in which its transcript level is reduced by the expression of miR399 regulated by PHR [51,120]. Single, double, or triple mutants of OsPHR1, OsPHR2, and OsPHR3 using CRISPR/Cas9 displayed growth retardation under low Pi conditions [24]. Pi uptake and starvation signaling is regulated by N availability through the NIGT1-SPX-PHR cascade [121]. By inhibiting the expression of SPX, NIGT1 and NIGT1.2 can indirectly activate PHR expression [121,122,123]. This was confirmed by the nigt1.1 nigt1.2 double mutant generated by CRISPR/Cas9 that demonstrated reduced P uptake and improved N under Pi-deficient conditions [123]. The list of genes/proteins involved in phosphate homeostasis of plants that have been functional characterized using CRISPR/Cas9-mediated targeted mutagenesis is given in Table 2.
All of the abovementioned examples illustrate that highly efficient targeted mutagenesis in various target genes can be achieved using CRISPR/Cas9. However, there has been some concern about the high-frequency off-target mutagenesis induced by this method, which gets further accentuated in the T2 progeny [124]. Although the majority of the earlier studies did not observe any off-target effects, the risk can be minimized by genome searching of highly specific target sequences [125,126]. Moreover, the unwanted off-target mutations can be reduced by employing different engineered Cas9 variants with altered PAM specificities [127]. With less stringent dependence on the presence of the original PAM sequence (5’-NGG-3’), the efficacy of the Cas9 system to target a specific sequence can be enhanced. Additionally, the Cas9 paired nickase can also improve the specificity and eliminate off-target mutations [128].
TFss are often members of the multigene families, and their functional redundancy is common. Therefore, an explicit understanding of the highly complex and intricate co-regulatory networks of various TF families is essential for enhancing Pi uptake efficiency in crop plants. This complexity can be addressed by utilizing CRISPR/Cas9-mediated multiplex genome editing. Numerous different sequence-specific gRNAs coupled with monomeric Cas9 proteins allow for simultaneous editing of multiple TFs. In addition, the availability of efficient cloning techniques such as Golden Gate Cloning [129] and Gibson Assembly [97], facilitates the assembly of different components into a single T-DNA. Apart from TFs, other gene regulons such as protein regulators and microRNAs controlling Pi homeostasis in plants can also be simultaneously targeted to remodel Pi-starvation responses in crops.
After Cas9 has generated the sgRNA-directed DSB, a fragment of DNA that carries a desired sequence can also be inserted or replaced into a precise location via HDR. This method has been used to introduce one or multiple genes into different crop plants [130] but with low efficiency, making it challenging to implement [26]. This may explain the lack of research on PUE using this approach. When more efficient methods become available, targeted gene knock in could be used to insert/swap promoters or add/delete motifs in the plant genome using donor templates to enhance phosphate use efficiency.
Following the CRISPR/Cas systems, another highly efficient genome editing method without creating DSBs was developed, termed base editing (BE) [131,132,133]. The functional amino acid residues can be altered at the target site and result in the point mutation of key residues. Although CRISPR/Cas BE has been successfully applied in various crop plants, targeted base editing for functional characterization of genes associated with the nutrient use-efficiency of plants is limited [134]. So far, only one study has reported the precise base editing of a nitrate transporter gene in rice [135]. Therefore, the key residues of the phosphate transporter could be engineered specifically to enhance the Pi transport efficiency in plants. AtPHT1 was mutated at a key residue involved in the binding of Pi, Tyr312 which resulted in increased Pi transport in yeast [136]. CRISPR/Cas BE could be employed to replicate the work in the native plant system.
Another cutting-edge genome editing method is prime editing (PE), which facilitates targeted insertions, deletions, and base-to-base conversions within a targeted sequence without inducing DSBs [137]. Similar to BEs, the PE tool allows for programmable nucleotide substitutions without the need for a donor template [137]. The advantages of prime editing tools offer enormous potential for crop improvement [138]. However, the application of this technology is yet to be reported for studying and enhancing Pi uptake in crop plants. CRISPR/Cas PE can be utilized for the engineering of promoters and adding more motifs to enhance the binding of transcription factors involved in Pi uptake and transport. For instance, enhancing P1BS, MYCS, and W-Box motifs in the promoter of PHT1 family transporters could help upregulate the PHT1 expression for increased Pi uptake from soil [44,139].
Since its first appearance, the application of the CRISPR/Cas system has expanded beyond targeted gene knock out and gene knock in. The development of catalytically inactive or dead Cas9 (dCas9) enables researchers to knock down or alter the expression of their desired genes in a highly specific manner [29]. Due to the introduced mutations in the nuclease domain, dCas9 is no longer able to cleave DNA, but when directed by the gRNA, it can still bind to the target site with the same specificity. The targeted genes are fused with dCas9 to repress (CRISPRi) or activate (CRISPRa) transcription by RNA polymerase, thereby upregulating or downregulating the gene expression. This modified system is useful for precise gene silencing of a negative regulator of the stress response pathway [140], while enhancing the activity of a positive regulator [141]. In the case of Pi homeostasis in crop plants, the activation or repression of the genes that will in turn activate the expression of the transporters encoded by PHT1 may improve the Pi uptake [34]. The amplification of PHT1 activity can be achieved by enhancing the binding of PHR1 to P1BS or MYCS to P1BS [139].
Another interesting target are the negative regulators PHO2 and SPX1/2 which are repressed by miR399 and NIGT1 and NIGT1.2, respectively [14,142,143]. The binding of dCas9 with specific gRNA targeted to the promoter sequence of these genes could lead to gene repression and increase Pi foraging from soil. Other potential targets of Pi signaling negative regulators include IPS1/2, AtWRKY6, AtWRKY43, and miR827. In contrast, activation of the different positive regulators such as AtPHR1, AtPHR2, PHO1, among others will increase PUE.
5. Summary and Future Prospects
Significant advancements have been made in deciphering the molecular network underlying the maintenance of cellular Pi homeostasis in plants. The regulation of inadequate Pi supply involved a range of transporters, transcription factors, signalling molecules, domain proteins, and enzymes that operate through an integrated response pathway at the transcriptional, translational, or post-translational levels. The key transporters such as PHT1 and PHO1 play significant roles in transporting Pi from the root to other plant parts. Plants exhibit growth and developmental adaptive responses upon sensing a deficiency of Pi in the soil. These responses are primarily controlled at the transcriptional level by various transcription factor families such as MYB, WRKY, bHLH and ZAT. The maintenance of Pi homeostasis is regulated by the direct binding of SPX domain proteins to the PP-InsPs signalling molecule, which in turn regulates different cellular components. Acting as sensors of cellular Pi status, the SPX domain also binds to VPT, a Pi transporter located in the vacuole, to transport Pi into the vacuole under high intracellular Pi.
Proteomic approaches have led to the discovery of regulatory proteins that participate in DNA-protein and protein-protein interactions in Pi homeostasis, operating both at translational and post-translational levels. In response to Pi deficiency, phosphatases secreted by roots have been demonstrated to play a crucial role in the mobilisation of Pi from the root. Further utilisation of quantitative proteomics should reveal more candidate proteins associated with low Pi tolerance, which can provide insights on the regulatory mechanisms of Pi homeostasis. The role of phosphorylation and/or succinylation of regulatory proteins in controlling the response of major metabolic pathways to Pi starvation and recovery in the roots has been revealed through protein-protein interaction network analysis.
In addition to proteomics, the progress in elucidating key regulatory molecules governing the complex regulations of Pi homeostasis in plants has been facilitated by CRISPR/Cas-based gene editing strategies. To date, several genes and proteins associated with Pi homeostasis have been targeted for mutagenesis using this approach in model plants, such as Arabidopsis and rice. It is also anticipated that the other CRISPR/Cas system applications, including gene knock in, base editing, prime editing, CRISPRi, and CRISPRa technology will be extensively utilized to precisely engineer key genes encoding Pi transporters, their transcriptional regulators, and motifs in promoters of target PSIs to fine tune plant Pi metabolism and root systems for Pi acquisition and PUE improvement.
CRISPR/Cas technology could provide numerous opportunities for improving phosphate transport in crop plants by engineering key functional residues of transporters, transcriptional regulation of transporters and regulatory genes, as well as engineering motifs in promoters and transcription factors. We envision that as more reference genomes become available in the coming years, along with improved tissue culture systems, CRISPR-based functional studies will be expanded to economically important crops to elucidate the conserved mechanisms regulating Pi uptake and transport. CRISPR-based engineering of plant phosphate transport not only helps to study the process in native plants with conserved regulatory system but also aids in the development of non-transgenic crops with enhanced PUE by enabling the generation of marker-free plants via non-integral delivery of CRISPR/Cas components. However, the availability of a fully annotated genome, as well as efficient plant regeneration and transformation systems, will be critical in allowing the delivery of CRISPR constructs, which will ultimately aid researchers in fine-tuning plant Pi metabolism to increase Pi uptake and usage efficiency. The generation of plants with a high PUE through the CRISPR/Cas system would reduce the need for chemical fertilizers and promote sustainable agriculture under climate change.
Author Contributions
N. A and S.N.A.A conceived of the review and led the manuscript preparation. M.A.M.H and N.K contributed to writing and editing. All authors have agreed to the published version of the manuscript.
Funding
Funding for the review was provided by the Ministry of Higher Education Malaysia under the Long-Term Research Grant Scheme (LRGS) LRGS/1/2020/UPM/01/2.
Conflict of Interest
The authors declare no conflict of interest.
References
- Lynch, J.P., Root phenes for enhanced soil exploration and phosphorus acquisition: tools for future crops. Plant physiology, 2011. 156(3): p. 1041-1049. [CrossRef]
- Shrivastav, P., et al., Role of nutrients in plant growth and development. Contaminants in agriculture: Sources, impacts and management, 2020: p. 43-59.
- Yadav, D.S., et al., Soil acidification and its impact on plants. Plant responses to soil pollution, 2020: p. 1-26.
- Etesami, H., B.R. Jeong, and B.R. Glick, Contribution of arbuscular mycorrhizal fungi, phosphate–solubilizing bacteria, and silicon to P uptake by plant. Frontiers in Plant Science, 2021. 12: p. 699618. [CrossRef]
- Muhammad, I.I., et al., The dynamic responses of oil palm leaf and root metabolome to phosphorus deficiency. Metabolites, 2021. 11(4): p. 217. [CrossRef]
- Trejo-Téllez, L.I. and F.C. Gómez-Merino, Phosphite as an inductor of adaptive responses to stress and stimulator of better plant performance. Biotic and Abiotic Stress Tolerance in Plants, 2018: p. 203-238.
- Malhotra, H., et al., Phosphorus nutrition: plant growth in response to deficiency and excess. Plant nutrients and abiotic stress tolerance, 2018: p. 171-190. [CrossRef]
- Luan, M., et al., Vacuolar phosphate transporters contribute to systemic phosphate homeostasis vital for reproductive development in Arabidopsis. Plant physiology, 2019. 179(2): p. 640-655. [CrossRef]
- Lorenzo-Orts, L., D. Couto, and M. Hothorn, Identity and functions of inorganic and inositol polyphosphates in plants. New Phytologist, 2020. 225(2): p. 637-652. [CrossRef]
- Pan, Y., et al., The genetic basis of phosphorus utilization efficiency in plants provide new insight into woody perennial plants improvement. International Journal of Molecular Sciences, 2022. 23(4): p. 2353. [CrossRef]
- Li, H., et al., Molecular mechanism of phosphorous signaling inducing anthocyanin accumulation in Arabidopsis. Plant Physiology and Biochemistry, 2023. 196: p. 121-129. [CrossRef]
- Verma, L., et al., Phosphate deficiency response and membrane lipid remodeling in plants. Plant Physiology Reports, 2021: p. 1-12. [CrossRef]
- Long, L., et al., Root plasticity and Pi recycling within plants contribute to low-P tolerance in Tibetan wild barley. BMC plant biology, 2019. 19(1): p. 1-13. [CrossRef]
- Gu, M., et al., Complex regulation of plant phosphate transporters and the gap between molecular mechanisms and practical application: what is missing? Molecular Plant, 2016. 9(3): p. 396-416. [CrossRef]
- Kong, S.-L., et al., Comparative transcriptome analysis reveals novel insights into transcriptional responses to phosphorus starvation in oil palm (Elaeis guineensis) root. BMC genomic data, 2021. 22: p. 1-15. [CrossRef]
- Chien, P.-S., et al., Sensing and signaling of phosphate starvation: from local to long distance. Plant and Cell Physiology, 2018. 59(9): p. 1714-1722. [CrossRef]
- Su, T., et al., WRKY42 modulates phosphate homeostasis through regulating phosphate translocation and acquisition in Arabidopsis. Plant Physiology, 2015. 167(4): p. 1579-1591. [CrossRef]
- Chiou, T.-J. and S.-I. Lin, Signaling network in sensing phosphate availability in plants. Annual review of plant biology, 2011. 62: p. 185-206. [CrossRef]
- Devaiah, B.N., V.K. Nagarajan, and K.G. Raghothama, Phosphate homeostasis and root development in Arabidopsis are synchronized by the zinc finger transcription factor ZAT6. Plant Physiology, 2007. 145(1): p. 147-159. [CrossRef]
- Hamzah, M.L., S.N.A. Abdullah, and A.M. Azzeme, Genome-wide molecular characterization of Phosphate Transporter 1 and Phosphate Starvation Response gene families in Elaeis guineensis Jacq. and their transcriptional response under different levels of phosphate starvation. Acta Physiologiae Plantarum, 2021. 43: p. 1-15. [CrossRef]
- Li, Z., et al., Integrated mRNA and microRNA expression analysis of root response to phosphate deficiency in Medicago sativa. Frontiers in Plant Science, 2022: p. 3444. [CrossRef]
- Xue, Y.-B., et al., GmPHR25, a GmPHR member up-regulated by phosphate starvation, controls phosphate homeostasis in soybean. Journal of Experimental Botany, 2017. 68(17): p. 4951-4967. [CrossRef]
- Sun, L., et al., Arabidopsis PHL2 and PHR1 act redundantly as the key components of the central regulatory system controlling transcriptional responses to phosphate starvation. Plant Physiology, 2016. 170(1): p. 499-514. [CrossRef]
- Guo, M., et al., Integrative comparison of the role of the PHOSPHATE RESPONSE1 subfamily in phosphate signaling and homeostasis in rice. Plant Physiology, 2015. 168(4): p. 1762-1776. [CrossRef]
- Dong, J., et al., Inositol pyrophosphate InsP8 acts as an intracellular phosphate signal in Arabidopsis. Molecular plant, 2019. 12(11): p. 1463-1473. [CrossRef]
- Liu, N., et al., Evolution of the SPX gene family in plants and its role in the response mechanism to phosphorus stress. Open biology, 2018. 8(1): p. 170231. [CrossRef]
- Sedeek, K.E., A. Mahas, and M. Mahfouz, Plant genome engineering for targeted improvement of crop traits. Frontiers in plant science, 2019. 10: p. 114. [CrossRef]
- Doudna, J.A. and E. Charpentier, The new frontier of genome engineering with CRISPR-Cas9. Science, 2014. 346(6213): p. 1258096. [CrossRef]
- Moradpour, M. and S.N.A. Abdulah, CRISPR/dC as9 platforms in plants: strategies and applications beyond genome editing. Plant Biotechnology Journal, 2020. 18(1): p. 32-44.
- Naumann, C., et al., The local phosphate deficiency response activates endoplasmic reticulum stress-dependent autophagy. Plant physiology, 2019. 179(2): p. 460-476. [CrossRef]
- Pandey, B.K., P. Mehra, and J. Giri, Phosphorus starvation response in plants and opportunities for crop improvement. Climate change and plant abiotic stress tolerance, 2013: p. 991-1012.
- Liu, T.-Y., et al., Identification of plant vacuolar transporters mediating phosphate storage. Nature communications, 2016. 7(1): p. 11095. [CrossRef]
- Ahmadi, F., et al., Functional characterization of the gene promoter for an Elaeis guineensis phosphate starvation-inducible, high affinity phosphate transporter in both homologous and heterologous model systems. Plant Physiology and Biochemistry, 2018. 127: p. 320-335. [CrossRef]
- Victor Roch, G., et al., The role of PHT1 family transporters in the acquisition and redistribution of phosphorus in plants. Critical Reviews in Plant Sciences, 2019. 38(3): p. 171-198. [CrossRef]
- Chiou, T.-J., The diverse roles of rice PHO1 in phosphate transport: from root to node to grain. Plant and Cell Physiology, 2020. 61(8): p. 1384-1386. [CrossRef]
- Devaiah, B.N. and K.G. Raghothama, Transcriptional regulation of Pi starvation responses by WRKY75. Plant signaling & behavior, 2007. 2(5): p. 424-425. [CrossRef]
- Kumar, A., V. Gahlaut, and M. Nagaraju, Transcription factors and their roles in phosphorus stress tolerance in crop plants, in Transcription factors for abiotic stress tolerance in plants. 2020, Elsevier. p. 201-224.
- Zhang, J.-F., et al., Comprehensive Evolution and Expression anaLysis of PHOSPHATE 1 Gene Family in Allotetraploid Brassica napus and Its Diploid Ancestors. Biochemical Genetics, 2023: p. 1-18. [CrossRef]
- Ambawat, S., et al., MYB transcription factor genes as regulators for plant responses: an overview. Physiology and Molecular Biology of Plants, 2013. 19: p. 307-321. [CrossRef]
- Sega, P. and A. Pacak, Plant PHR transcription factors: put on a map. Genes, 2019. 10(12): p. 1018. [CrossRef]
- Bustos, R., et al., A central regulatory system largely controls transcriptional activation and repression responses to phosphate starvation in Arabidopsis. PLoS genetics, 2010. 6(9): p. e1001102. [CrossRef]
- Gonzalez, E., et al., PHOSPHATE TRANSPORTER TRAFFIC FACILITATOR1 is a plant-specific SEC12-related protein that enables the endoplasmic reticulum exit of a high-affinity phosphate transporter in Arabidopsis. The Plant Cell, 2005. 17(12): p. 3500-3512. [CrossRef]
- Mehra, P., B.K. Pandey, and J. Giri, Improvement in phosphate acquisition and utilization by a secretory purple acid phosphatase (OsPAP21b) in rice. Plant Biotechnology Journal, 2017. 15(8): p. 1054-1067. [CrossRef]
- Wang, Z., et al., Functional characterization of Arabidopsis PHL4 in plant response to phosphate starvation. Frontiers in Plant Science, 2018. 9: p. 1432. [CrossRef]
- Ruan, W., et al., Phosphate starvation induced OsPHR4 mediates Pi-signaling and homeostasis in rice. Plant molecular biology, 2017. 93: p. 327-340. [CrossRef]
- Sun, Y., et al., OsPHR3 affects the traits governing nitrogen homeostasis in rice. BMC plant biology, 2018. 18(1): p. 1-15. [CrossRef]
- Ruan, W., et al., Genetic manipulation of a high-affinity PHR1 target cis-element to improve phosphorous uptake in Oryza sativa L. Plant molecular biology, 2015. 87: p. 429-440. [CrossRef]
- Devaiah, B.N., et al., Phosphate starvation responses and gibberellic acid biosynthesis are regulated by the MYB62 transcription factor in Arabidopsis. Molecular plant, 2009. 2(1): p. 43-58. [CrossRef]
- Yang, W.T., et al., Overexpression of OsMYB4P, an R2R3-type MYB transcriptional activator, increases phosphate acquisition in rice. Plant Physiology and Biochemistry, 2014. 80: p. 259-267. [CrossRef]
- Yang, W.T., et al., Rice OsMYB5P improves plant phosphate acquisition by regulation of phosphate transporter. PLoS One, 2018. 13(3): p. e0194628. [CrossRef]
- Bari, R., et al., PHO2, microRNA399, and PHR1 define a phosphate-signaling pathway in plants. Plant physiology, 2006. 141(3): p. 988-999. [CrossRef]
- Chiou, T.-J., et al., Regulation of phosphate homeostasis by microRNA in Arabidopsis. The Plant Cell, 2006. 18(2): p. 412-421. [CrossRef]
- Hu, B., et al., LEAF TIP NECROSIS1 plays a pivotal role in the regulation of multiple phosphate starvation responses in rice. Plant Physiology, 2011. 156(3): p. 1101-1115. [CrossRef]
- Xu, F., et al., Genome-wide identification of soybean microRNAs and their targets reveals their organ-specificity and responses to phosphate starvation. BMC genomics, 2013. 14: p. 1-30. [CrossRef]
- Baek, D., et al., Regulation of miR399f transcription by AtMYB2 affects phosphate starvation responses in Arabidopsis. Plant physiology, 2013. 161(1): p. 362-373. [CrossRef]
- Qi, X., et al., Arabidopsis G-Protein β Subunit AGB1 Negatively Regulates DNA Binding of MYB62, a Suppressor in the Gibberellin Pathway. International Journal of Molecular Sciences, 2021. 22(15): p. 8270. [CrossRef]
- Chen, Y.-F., et al., The WRKY6 transcription factor modulates PHOSPHATE1 expression in response to low Pi stress in Arabidopsis. The Plant Cell, 2009. 21(11): p. 3554-3566. [CrossRef]
- Li, C., et al., Transcription factor GmWRKY46 enhanced phosphate starvation tolerance and root development in transgenic plants. Frontiers in Plant Science, 2021: p. 1984. [CrossRef]
- Chen, Z.-H., et al., BHLH32 modulates several biochemical and morphological processes that respond to Pi starvation in Arabidopsis. Biochemical Journal, 2007. 405(1): p. 191-198. [CrossRef]
- Yeh, C.-M., et al., Blue light regulates phosphate deficiency-dependent primary root growth inhibition in Arabidopsis. Frontiers in Plant Science, 2020. 10: p. 1803. [CrossRef]
- Huang, L., et al., Zinc finger protein 5 (ZFP5) associates with ethylene signaling to regulate the phosphate and potassium deficiency-induced root hair development in Arabidopsis. Plant molecular biology, 2020. 102: p. 143-158. [CrossRef]
- Staudinger, C., et al., The wheat secreted root proteome: Implications for phosphorus mobilisation and biotic interactions. Journal of Proteomics, 2022. 252: p. 104450. [CrossRef]
- Zhang, Y., et al., Abscisic acid facilitates phosphate acquisition through the transcription factor ABA INSENSITIVE5 in Arabidopsis. The Plant Journal, 2022. 111(1): p. 269-281. [CrossRef]
- Zhou, Y., et al., Proteomic Investigation of Molecular Mechanisms in Response to PEG-Induced Drought Stress in Soybean Roots. Plants, 2022. 11(9): p. 1173. [CrossRef]
- Cheng, L., et al., Quantitative proteomics reveals that GmENO2 proteins are involved in response to phosphate starvation in the leaves of Glycine max L. International Journal of Molecular Sciences, 2021. 22(2): p. 920. [CrossRef]
- Guo, M., et al., Alternative splicing of REGULATOR OF LEAF INCLINATION 1 modulates phosphate starvation signaling and growth in plants. The Plant Cell, 2022. 34(9): p. 3319-3338. [CrossRef]
- Muneer, S. and B.R. Jeong, Proteomic analysis provides new insights in phosphorus homeostasis subjected to Pi (inorganic phosphate) starvation in tomato plants (Solanum lycopersicum L.). PLoS One, 2015. 10(7): p. e0134103. [CrossRef]
- Deng, Q., et al., Identification of phosphorus stress related proteins in the seedlings of Dongxiang wild rice (Oryza rufipogon Griff.) using label-free quantitative proteomic analysis. Genes, 2022. 13(1): p. 108. [CrossRef]
- Roustan, V., et al., Quantitative in vivo phosphoproteomics reveals reversible signaling processes during nitrogen starvation and recovery in the biofuel model organism Chlamydomonas reinhardtii. Biotechnology for biofuels, 2017. 10(1): p. 1-24. [CrossRef]
- Hetz, R., et al., Excessive Inorganic Phosphate Burden Perturbed Intracellular Signaling: Quantitative Proteomics and Phosphoproteomics Analyses. Frontiers in Nutrition, 2022: p. 1101. [CrossRef]
- Graham, P.H. and C.P. Vance, Legumes: importance and constraints to greater use. Plant physiology, 2003. 131(3): p. 872-877.
- Madhu, M. and J.L. Hatfield, Interaction of carbon dioxide enrichment and soil moisture on photosynthesis, transpiration, and water use efficiency of soybean. Agricultural Sciences, 2014. 2014. [CrossRef]
- Guo, W., et al., Identification of temporally and spatially phosphate-starvation responsive genes in Glycine max. Plant Science, 2008. 175(4): p. 574-584. [CrossRef]
- Yang, Z., et al., GmPTF1 modifies root architecture responses to phosphate starvation primarily through regulating GmEXPB2 expression in soybean. The Plant Journal, 2021. 107(2): p. 525-543. [CrossRef]
- Jiang, W., et al., Soybean responds to phosphate starvation through reversible protein phosphorylation. Plant Physiology and Biochemistry, 2021. 167: p. 222-234. [CrossRef]
- Fan, R., et al., Insights into the mechanism of phospholipid hydrolysis by plant non-specific phospholipase C. Nature Communications, 2023. 14(1): p. 1-11. [CrossRef]
- Yang, M., et al., Chloroplastic Sec14-like proteins modulate growth and phosphate deficiency responses in Arabidopsis and rice. Plant Physiology, 2023: p. kiad212. [CrossRef]
- Zhao, Q., et al., Flavonoids Mediate the Modulation of Phosphate Uptake and Phosphate-Starvation Signaling in Tobacco. Journal of Plant Growth Regulation, 2023: p. 1-11. [CrossRef]
- Li, J., et al., Phosphate deficiency modifies lipid composition and seed oil production in camelina. Plant Science, 2023. 330: p. 111636. [CrossRef]
- Okazaki, Y., K. Takano, and K. Saito, Lipidomic analysis of soybean leaves revealed tissue-dependent difference in lipid remodeling under phosphorus-limited growth conditions. Plant Biotechnology, 2017. 34(1): p. 57-63. [CrossRef]
- Ngo, A.H. and Y. Nakamura, Phosphate starvation-inducible GLYCEROPHOSPHODIESTER PHOSPHODIESTERASE6 is involved in Arabidopsis root growth. Journal of experimental botany, 2022. 73(9): p. 2995-3003. [CrossRef]
- Angkawijaya, A.E., V.C. Nguyen, and Y. Nakamura, Enhanced root growth in phosphate-starved Arabidopsis by stimulating de novo phospholipid biosynthesis through the overexpression of LYSOPHOSPHATIDIC ACID ACYLTRANSFERASE 2 (LPAT2). Plant, Cell & Environment, 2017. 40(9): p. 1807-1818.
- Ngo, A.H., et al., The phospho-base N-methyltransferases PMT1 and PMT2 produce phosphocholine for leaf growth in phosphorus-starved Arabidopsis. Journal of experimental botany, 2022. 73(9): p. 2985-2994. [CrossRef]
- Shears, S.B., Intimate connections: Inositol pyrophosphates at the interface of metabolic regulation and cell signaling. Journal of Cellular Physiology, 2018. 233(3): p. 1897-1912. [CrossRef]
- Cridland, C. and G. Gillaspy, Inositol pyrophosphate pathways and mechanisms: what can we learn from plants? Molecules, 2020. 25(12): p. 2789. [CrossRef]
- Adepoju, O., et al., Inositol trisphosphate kinase and diphosphoinositol pentakisphosphate kinase enzymes constitute the inositol pyrophosphate synthesis pathway in plants. Biorxiv, 2019: p. 724914.
- Laha, D., et al., Arabidopsis ITPK1 and ITPK2 have an evolutionarily conserved phytic acid kinase activity. ACS chemical biology, 2019. 14(10): p. 2127-2133. [CrossRef]
- Laha, D., et al., Regulation of plant biotic interactions and abiotic stress responses by inositol polyphosphates. Frontiers in Plant Science, 2022: p. 2835. [CrossRef]
- Zhu, J., et al., Two bifunctional inositol pyrophosphate kinases/phosphatases control plant phosphate homeostasis. Elife, 2019. 8: p. e43582. [CrossRef]
- Land, E.S., et al., A role for inositol pyrophosphates in the metabolic adaptations to low phosphate in arabidopsis. Metabolites, 2021. 11(9): p. 601. [CrossRef]
- Kuo, H.F., et al., Arabidopsis inositol phosphate kinases IPK 1 and ITPK 1 constitute a metabolic pathway in maintaining phosphate homeostasis. The Plant Journal, 2018. 95(4): p. 613-630. [CrossRef]
- Gulabani, H., et al., Arabidopsis inositol polyphosphate kinases IPK1 and ITPK1 modulate crosstalk between SA-dependent immunity and phosphate-starvation responses. Plant Cell Reports, 2021: p. 1-17. [CrossRef]
- Laha, D., et al., VIH2 regulates the synthesis of inositol pyrophosphate InsP8 and jasmonate-dependent defenses in Arabidopsis. The Plant Cell, 2015. 27(4): p. 1082-1097. [CrossRef]
- Lv, Q., et al., SPX4 negatively regulates phosphate signaling and homeostasis through its interaction with PHR2 in rice. The Plant Cell, 2014. 26(4): p. 1586-1597. [CrossRef]
- Zhong, Y., et al., Rice SPX6 negatively regulates the phosphate starvation response through suppression of the transcription factor PHR2. New Phytologist, 2018. 219(1): p. 135-148. [CrossRef]
- Puga, M.I., et al., SPX1 is a phosphate-dependent inhibitor of Phosphate Starvation Response 1 in Arabidopsis. Proceedings of the National Academy of Sciences, 2014. 111(41): p. 14947-14952. [CrossRef]
- Wang, J.-W., et al., CRISPR/Cas9 nuclease cleavage combined with Gibson assembly for seamless cloning. BioTechniques, 2015. 58(4): p. 161-170. [CrossRef]
- Qi, W., et al., AtSPX1 affects the AtPHR1–DNA-binding equilibrium by binding monomeric AtPHR1 in solution. Biochemical Journal, 2017. 474(21): p. 3675-3687. [CrossRef]
- Du, H., et al., Genome-wide identification and characterization of SPX domain-containing members and their responses to phosphate deficiency in Brassica napus. Frontiers in plant science, 2017. 8: p. 35. [CrossRef]
- Jung, J.-Y., et al., Control of plant phosphate homeostasis by inositol pyrophosphates and the SPX domain. Current opinion in biotechnology, 2018. 49: p. 156-162. [CrossRef]
- Ried, M.K., et al., Inositol pyrophosphates promote the interaction of SPX domains with the coiled-coil motif of PHR transcription factors to regulate plant phosphate homeostasis. Nature communications, 2021. 12(1): p. 384. [CrossRef]
- Zhou, J., et al., Mechanism of phosphate sensing and signaling revealed by rice SPX1-PHR2 complex structure. Nature communications, 2021. 12(1): p. 7040. [CrossRef]
- He, Y., et al., SPX4 interacts with both PHR1 and PAP1 to regulate critical steps in phosphorus-status-dependent anthocyanin biosynthesis. New Phytologist, 2021. 230(1): p. 205-217. [CrossRef]
- Osorio, M.B., et al., SPX4 acts on PHR1-dependent and-independent regulation of shoot phosphorus status in Arabidopsis. Plant physiology, 2019. 181(1): p. 332-352. [CrossRef]
- Han, B., et al., Identification of vacuolar phosphate influx transporters in Brassica napus. Plant, Cell & Environment, 2022. 45(11): p. 3338-3353. [CrossRef]
- Sun, G., et al., Genetically controlling VACUOLAR PHOSPHATE TRANSPORTER 1 contributes to low-phosphorus seeds in Arabidopsis. Plant Signaling & Behavior, 2023. 18(1): p. 2186641. [CrossRef]
- Diallinas, G., Understanding transporter specificity and the discrete appearance of channel-like gating domains in transporters. Frontiers in pharmacology, 2014. 5: p. 207. [CrossRef]
- Luan, M., et al., A SPX domain vacuolar transporter links phosphate sensing to homeostasis in Arabidopsis. Molecular Plant, 2022. 15(10): p. 1590-1601. [CrossRef]
- Jia, X., et al., Insights of intracellular/intercellular phosphate transport and signaling in unicellular green algae and multicellular land plants. New Phytologist, 2021. 232(4): p. 1566-1571. [CrossRef]
- Kimura, S. and K. Sakaguchi, DNA repair in plants. Chemical reviews, 2006. 106(2): p. 753-766.
- Jinek, M., et al., A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. science, 2012. 337(6096): p. 816-821. [CrossRef]
- Nacry, P., et al., Major chromosomal rearrangements induced by T-DNA transformation in Arabidopsis. Genetics, 1998. 149(2): p. 641-650. [CrossRef]
- Tamura, K., et al., Decreased expression of a gene caused by a T-DNA insertion in an adjacent gene in arabidopsis. Plos one, 2016. 11(2): p. e0147911. [CrossRef]
- Gu, M., et al., Maintenance of phosphate homeostasis and root development are coordinately regulated by MYB1, an R2R3-type MYB transcription factor in rice. Journal of experimental botany, 2017. 68(13): p. 3603-3615. [CrossRef]
- Zhao, P., Q. You, and M. Lei, A CRISPR/Cas9 deletion into the phosphate transporter SlPHO1; 1 reveals its role in phosphate nutrition of tomato seedlings. Physiologia plantarum, 2019. 167(4): p. 556-563. [CrossRef]
- Pérez-Torres, C.-A., et al., Phosphate availability alters lateral root development in Arabidopsis by modulating auxin sensitivity via a mechanism involving the TIR1 auxin receptor. The Plant Cell, 2008. 20(12): p. 3258-3272.
- Williamson, L.C., et al., Phosphate availability regulates root system architecture in Arabidopsis. Plant physiology, 2001. 126(2): p. 875-882. [CrossRef]
- Lee, H.Y., et al., Editing of the OsACS locus alters phosphate deficiency-induced adaptive responses in rice seedlings. Journal of experimental botany, 2019. 70(6): p. 1927-1940. [CrossRef]
- Xu, L., et al., Identification of vacuolar phosphate efflux transporters in land plants. Nature Plants, 2019. 5(1): p. 84-94. [CrossRef]
- Wu, P. and X. Wang, Role of OsPHR2 on phosphorus homoestasis and root hairs development in rice (Oryza sativa L.). Plant signaling & behavior, 2008. 3(9): p. 674-675. [CrossRef]
- Ueda, Y., T. Kiba, and S. Yanagisawa, Nitrate-inducible NIGT1 proteins modulate phosphate uptake and starvation signalling via transcriptional regulation of SPX genes. The Plant Journal, 2020. 102(3): p. 448-466. [CrossRef]
- Poza-Carrión, C. and J. Paz-Ares, When nitrate and phosphate sensors meet. Nature plants, 2019. 5(4): p. 339-340. [CrossRef]
- Wang, X., et al., The transcription factor NIGT1. 2 modulates both phosphate uptake and nitrate influx during phosphate starvation in Arabidopsis and maize. Plant Cell, 2020. 32(11): p. 3519-3534. [CrossRef]
- Zhang, Q., et al., Potential high-frequency off-target mutagenesis induced by CRISPR/Cas9 in Arabidopsis and its prevention. Plant molecular biology, 2018. 96: p. 445-456. [CrossRef]
- Lei, Y., et al., CRISPR-P: a web tool for synthetic single-guide RNA design of CRISPR-system in plants. Molecular plant, 2014. 7(9): p. 1494-1496. [CrossRef]
- Xie, K., J. Zhang, and Y. Yang, Genome-wide prediction of highly specific guide RNA spacers for CRISPR–Cas9-mediated genome editing in model plants and major crops. Molecular plant, 2014. 7(5): p. 923-926. [CrossRef]
- Meng, X., et al., Robust genome editing of CRISPR-Cas9 at NAG PAMs in rice. Science China. Life Sciences, 2018. 61(1): p. 122-125. [CrossRef]
- Cong, L., et al., Multiplex genome engineering using CRISPR/Cas systems. Science, 2013. 339(6121): p. 819-823. [CrossRef]
- Engler, C. and S. Marillonnet, Golden gate cloning. DNA cloning and assembly methods, 2014: p. 119-131.
- Van Vu, T., et al., Challenges and perspectives in homology-directed gene targeting in monocot plants. Rice, 2019. 12(1): p. 1-29. [CrossRef]
- Kim, Y.B., et al., Increasing the genome-targeting scope and precision of base editing with engineered Cas9-cytidine deaminase fusions. Nature biotechnology, 2017. 35(4): p. 371-376. [CrossRef]
- Komor, A.C., et al., Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature, 2016. 533(7603): p. 420-424. [CrossRef]
- Rees, H.A., et al., Improving the DNA specificity and applicability of base editing through protein engineering and protein delivery. Nature communications, 2017. 8(1): p. 15790. [CrossRef]
- Bharat, S.S., et al., Base editing in plants: Current status and challenges. The Crop Journal, 2020. 8(3): p. 384-395. [CrossRef]
- Lu, Y. and J.-K. Zhu, Precise editing of a target base in the rice genome using a modified CRISPR/Cas9 system. Molecular plant, 2017. 10(3): p. 523-525. [CrossRef]
- Fontenot, E.B., et al., Increased phosphate transport of A rabidopsis thaliana P ht1; 1 by site-directed mutagenesis of tyrosine 312 may be attributed to the disruption of homomeric interactions. Plant, Cell & Environment, 2015. 38(10): p. 2012-2022.
- Anzalone, A.V., et al., Search-and-replace genome editing without double-strand breaks or donor DNA. Nature, 2019. 576(7785): p. 149-157. [CrossRef]
- Zhan, X., et al., Genome editing for plant research and crop improvement. Journal of integrative plant biology, 2021. 63(1): p. 3-33. [CrossRef]
- Chen, A., et al., Identification of two conserved cis-acting elements, MYCS and P1BS, involved in the regulation of mycorrhiza-activated phosphate transporters in eudicot species. New Phytologist, 2011. 189(4): p. 1157-1169.
- Feng, Z., et al., Efficient genome editing in plants using a CRISPR/Cas system. Cell research, 2013. 23(10): p. 1229-1232. [CrossRef]
- Zhou, J., et al., CRISPR-Cas9 based genome editing reveals new insights into microRNA function and regulation in rice. Frontiers in Plant Science, 2017. 8: p. 1598. [CrossRef]
- Liu, P., et al., A putative high affinity phosphate transporter, CmPT1, enhances tolerance to Pi deficiency of chrysanthemum. BMC plant biology, 2014. 14(1): p. 1-9. [CrossRef]
- Zhou, Z., et al., SPX proteins regulate Pi homeostasis and signaling in different subcellular level. Plant signaling & behavior, 2015. 10(9): p. e1061163. [CrossRef]
Figure 1.
Schematic representation of lipid membrane remodelling in plant cells during phosphate starvation condition.
Figure 1.
Schematic representation of lipid membrane remodelling in plant cells during phosphate starvation condition.
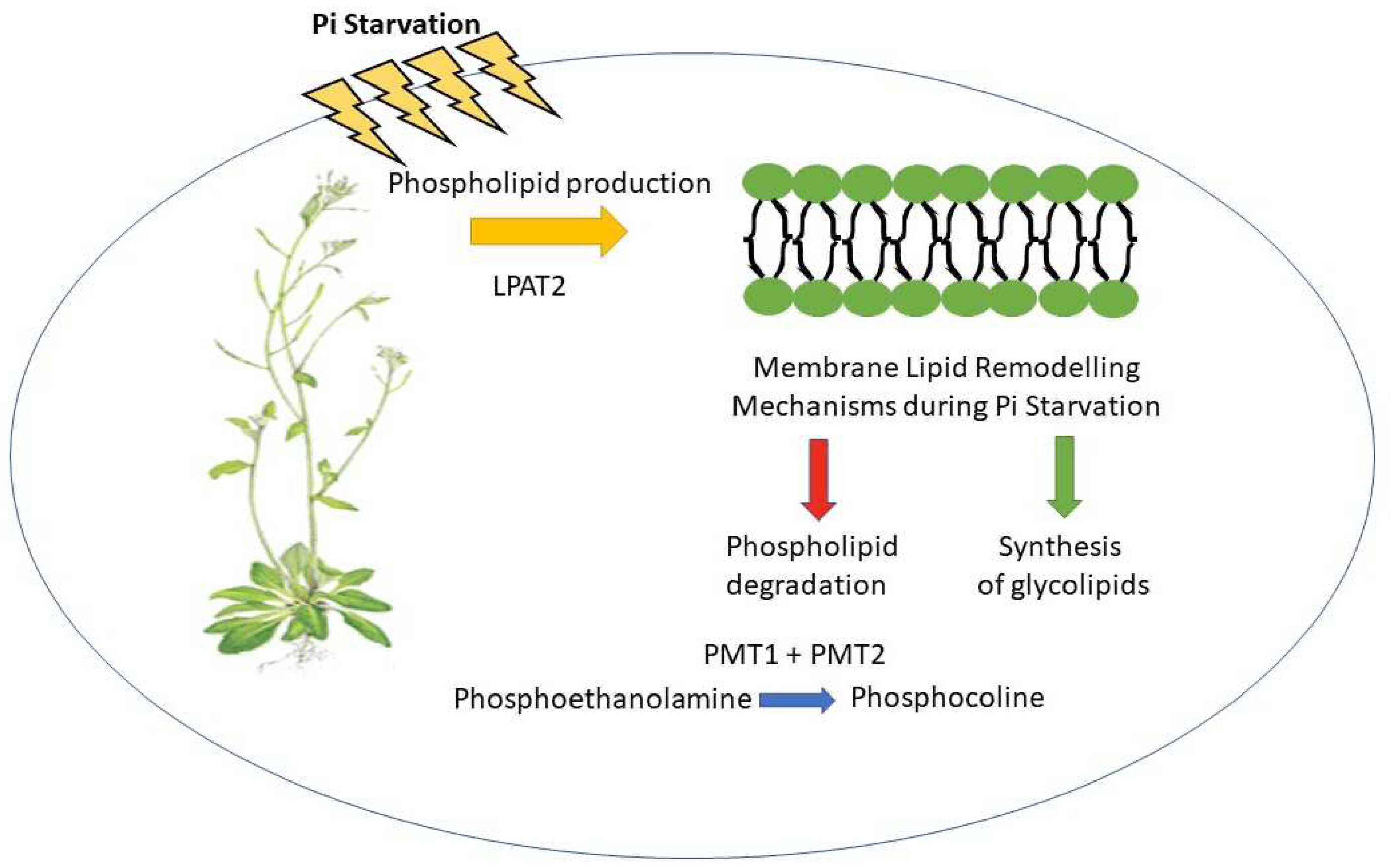
Figure 2.
Mechanism for maintenance of Pi homeostasis in plant cells through Pi-dependent regulation by PHRs/PHLs and VPTs:.
Figure 2.
Mechanism for maintenance of Pi homeostasis in plant cells through Pi-dependent regulation by PHRs/PHLs and VPTs:.
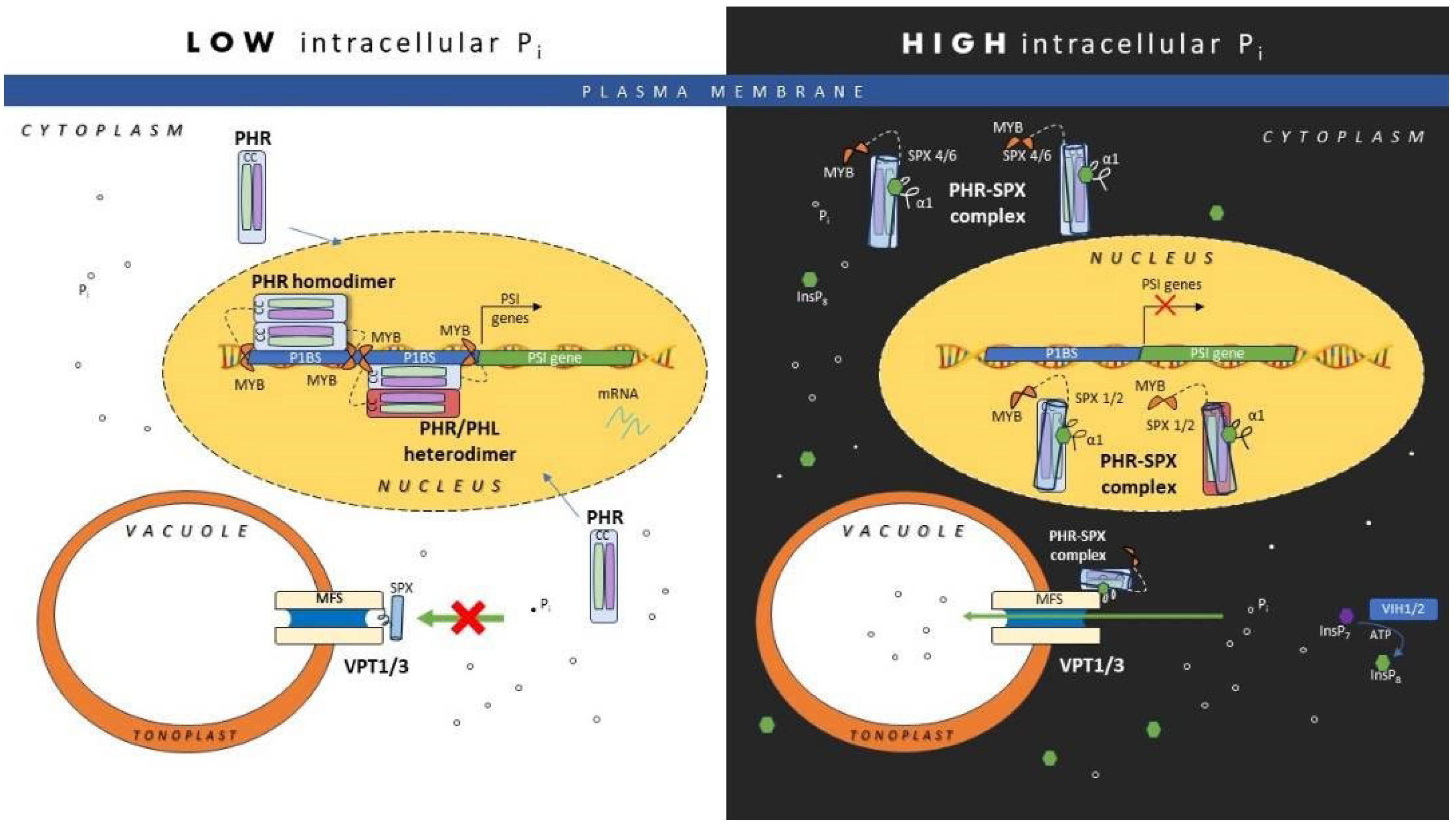

Table 1.
Transcription factors and other stress-responsive signaling pathways under Pi starvation.
| Type of Factor | Transcription | Species | Responses | References |
|---|---|---|---|---|
| MYB Family | MYB-CC (R1-type) - PHL1, PHL2, PHL 3 and PHL4 | Arabidopsis | Pi starvation | Bustos at al., (2010); Sun et al., (2016); Wang et al., (2018) |
| MYB-CC (R2R3-type) -MYB2 | Pi starvation, salt/ABA/drought response | Baek et al., (2013) | ||
| MYB-CC (R2R3-type) - MYB62, | Pi starvation, GA defiency, root development | Devaiah et al., (2009); Qi et al., (2021) | ||
| MYB-CC (R2R3-type) -MYB2P-1, MYB4P | Rice | Pi starvation | Yang et al., (2014); Yang et al., (2018) | |
| W-box and E/G-box, the binding site for PHR | Elaeis guineensis | Pi Starvation, root development | Ahmadi et al., (2018) | |
| WRKY Family | WRKY6 | Arabidopsis | Pi starvation, anthocyanin accumulation and root hair formation | Chen at al., (2009) |
| WRKY42 | Pi starvation | Su et al., (2005) | ||
| WRKY46 | Pi Starvation, root development | Li et al., (2021) | ||
| WRKY75 | Pi starvation | Devaiah et al., (2009) | ||
| bHLH Family | bHLH32 | Arabidopsis | Pi starvation, anthocyanin accumulation, root hair formation | Chen et al., (2007); Yeh et al., (2020) |
| ZFP Family | Zinc Finger (C2H2-type) -ZAT6 | Arabidopsis | Pi starvation | Devaiah et al., (2009) |
| Zinc Finger (C2H2-type) -ZFP5 | Pi starvation, regulation of trichome and root hair development. | Huang et al., (2019) |
Table 2.
Functional characterization of gene/protein involved in phosphate homeostasis of plants using CRISPR/Cas9-mediated targeted mutagenesis.
Table 2.
Functional characterization of gene/protein involved in phosphate homeostasis of plants using CRISPR/Cas9-mediated targeted mutagenesis.
| Target | Gene/ protein |
Function | Effects of targeted editing | References |
|---|---|---|---|---|
| SlPHO1;1 | Gene | Regulates root-to-shoot phosphate transport | Decreased shoot fresh weight, increased root biomass, higher root-to-shoot ratio | Zhao et al., (2019) |
| OsACS1 and OsACS2 | Gene | Regulates ethylene biosynthesis during low Pi | Altered root system architecture | Lee et al., (2019) |
| OsVPE1 and OsVPE2 | Transporter | Vacuolar Pi efflux transporters | Higher accumulation of vacuolar Pi content under low Pi stress than the wild type | Xu et al., (2019) |
| NIGT1 and NIGT1.2 | Trancriptional repressor | Negative regulator of SPX to activate PHR expression | Reduced P uptake and improved N under Pi-deficient conditions | Wang et al., (2020b) |
| OsPHR1, OsPHR2, OsPHR3 | Transcription factor | Regulates expression of miR399, which reduces the expression of a negative regulator, PHO2 | Growth retardation under low Pi conditions | Guo et al., (2015) |
| MYB1 | Transcription factor | Maintain phosphate homeostasis and root development | Enhanced Pi uptake and accumulation, altered expression of a subset of genes associated with Pi transporters and Pi starvation signaling | Gu et al., (2017) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



