
Submitted:
08 June 2023
Posted:
09 June 2023
You are already at the latest version
Abstract
Biological pretreatment of lignocellulosic residues has the potential to serve as a sustainable, less energy-intensive alternative to harsh chemical treatments for enhancing cellulose accessibility, despite the requirement of a lengthy incubation period. The study characterized the simultaneous delignification of corn stover by the white-rot fungus Pyrenophora phaeocomes S-1, as well as the co-production of a ligno-hemicellulolytic enzyme cocktail consisting of laccase, xylanase, and mannanase. The maximum yields of all three components of the cocktail were achieved after just 4 days of incubation under solid-state conditions. Following a 40-day fermentation period, we achieved a cellulose recovery of 44.25 ± 1.72%, attributed to the activities of 133.88 U/gds of laccase, 14.93 U/gds of xylanase, and 1.34 U/gds of mannanase co-produced by the fungus in the medium. Subsequently, through the extraction of biologically treated biomass with 0.5N NaOH after 40 days, the cellulose recovery increased to 66.4 ± 1.39%. Enzymatic hydrolysis of the same, employing only 5 FPU/gds of in-house produced cellulases, resulted in the liberation of 397.84 mg/gds of total reducing sugars after 144h. The findings of this study encourage further optimization of biological pretreatment of lignocellulosic residues and enzymatic hydrolysis to enhance the yields of total reducing sugars for their valorization.
Keywords:
Biological pretreatment
; Ligno-hemicellulolytic enzymes
; Corn stover
; Enzymatic hydrolysis
; Pyrenophora phaeocomes S-1
1. Introduction
Lignocellulosic residues represent a promising source of future energy in the form of biofuels. These residues primarily consist of three polymers: cellulose, hemicelluloses, and lignin. To convert raw biomass into biofuels, it is crucial to expose the cellulose content from the complex structure for hydrolysis into monomeric sugars, which can be further fermented into ethanol. The amorphous and highly branched lignin presents a significant obstacle in biomass decomposition, making the removal of lignin a critical and rate-determining step for biofuel production and the generation of value-added products. Various treatment methods are available, including physical techniques (such as mechanical shearing, pyrolysis, and microwave), physio-chemical approaches (such as ammonia fiber explosion, steam explosion, carbon dioxide explosion, and hot water), and chemical treatments (such as acid, alkaline, and organosolvents). However, these methods often involve the use of harsh chemicals or require sophisticated instruments, ultimately adding to the process’s cost [1,2,3].
An attractive alternative to these methods is the use of microbial pretreatment through fungi or their enzymes for lignin breakdown or removal from the holocellulose surface. This approach offers the advantage of being environmentally friendly, avoiding the use of harsh chemicals and high temperatures. Biological pretreatment of lignocellulosic residues has the potential to be a less energy-intensive treatment for improving the accessibility of cellulose. However, it has the drawback of requiring a longer incubation period. The use of fungal pretreatment has gained significant interest among researchers due to its environmental friendliness [4,5,6,7].
The main microorganisms responsible for lignin degradation in nature are white-rot fungi, which belong to the basidiomycetes and possess complex ligninolytic systems. These systems include lignin peroxidase (EC 1.11.1.14), manganese peroxidase (EC 1.11.1.13), and laccase (EC 1.10.3.1), which play a crucial role in wood degradation [7,8,9]. An ideal fungal candidate for biological pretreatment should selectively target lignin degradation while preserving the other carbohydrate components, particularly cellulose content. Some fungi have been reported to selectively degrade lignin and a portion of hemicellulose, thereby enhancing the accessibility of cellulose for subsequent hydrolysis and fermentation [10,11,12,13]. However, this energy-saving pretreatment process suffers from the major drawback of low hydrolysis efficiency [8,10,14,15,16,17]. To overcome the limitations of microbial pretreatment, the combination of microbial and traditional pretreatment approaches has been explored, aiming to enhance efficiency and reduce incubation periods [4,13,18,19].
Considering these challenges, the present study aimed to utilize abundantly available corn stover as a substrate for the production of an industrially important enzyme cocktail, while simultaneously performing pretreatment to preserve most of the cellulose. This was achieved by employing a fungal strain belonging to Pyrenophora phaeocomes S-1, enabling the conversion of the substrate to fermentable sugars through enzymatic hydrolysis.
2. Materials and Methods
2.1. Microorganism
The fungal strain P. phaeocomes S-1, which exhibits the ability to co-produce ligne-hemicellulolytic enzymes, was isolated for this study. The isolation process involved screening decaying wooden samples obtained from the biodiversity of Chandigarh city. The culture of P. phaeocomes S-1 was cultivated on Potato Dextrose Agar (PDA) plates at a temperature of 28°C for duration of 4 days, as described by Rastogi et al. [19].
2.2. Ligno-hemicellulolytic enzyme co-production using corn stover
Five grams of corn stover was moistened using a salt solution consisting of the following components (in g/L): NH4NO3, 4.0; KH2PO4, 0.8; Na2HPO4.7H2O, 0.75; MgSO4.7H2O, 0.5; yeast extract, 2.0; ZnSO4.7H2O, 0.002; FeSO4.7H2O, 0.005; CaCl2.2H2O, 0.06; MnSO4.4H2O, 0.05; and CuSO4.7H2O, 0.5, as described by Rastogi et al. [19]. The flasks containing the mixture were sterilized by autoclaving at 15 psi for 30 min. Subsequently, each flask was inoculated with 5 mycelial discs (7mm in diameter) obtained from the outer edge of actively growing colonies of P. phaeocomes S-1 cultivated on PDA plates. The inoculated flasks were then incubated under static conditions at a temperature of 28°C for a period of 4 days, with manual shaking performed once a day.
2.3. Enzyme extraction and quantification
After specific incubation duration, the contents of the flask were mixed with distilled water at a substrate-to-liquid ratio of 1:40. The mixture was mechanically stirred for 5 min. The fermented residue was then separated from the aqueous phase crude enzyme extract (CEE) through filtration, followed by centrifugation at 10,000 rpm at 4°C for 10 min. This resulted in obtaining a mycelium-free CEE, which was quantified for the activity of lignin-modifying enzyme laccases, as well as hemicellulose-modifying enzymes xylanase and mannanase.
The activity of laccase was determined by measuring the change in absorbance of guaiacol at OD470, resulting from oxidation at a temperature of 50°C for 10 min, as described by Jhadav et al. [20]. The laccase activity was expressed in International Units (IU), representing the amount of enzyme required to oxidize 1 µmoles of guaiacol per minute at a temperature of 50°C and pH 5.0. Xylanase and mannanase activities were determined using oat spent xylan [21] and guar gum [22] as substrates, respectively. The measurements were performed at a temperature of 50°C and pH 5.0, and the release of µmoles of xylose and mannose per minute was quantified using the dinitrosalicylic acid reagent [23]. The productivity of all three enzymes was expressed in terms of International Units (IU) per gram of dry substrate (gds) used in solid-state fermentation.
2.4. Biological pretreatment reaction system
The biological pretreatment of the biomass was conducted by cultivating P. phaeocomes S-1 as solid-state cultures on corn stover. The fungal treatment was carried out in 500 ml Erlenmeyer flasks, each containing 15 g of corn stover and distilled water to maintain an initial moisture level of 83.3% w/v. The sterilized corn stover medium was then inoculated with fifteen mycelial discs of P. phaeocomes S-1 that were previously propagated on PDA plates, as mentioned earlier. These flasks were incubated at a temperature of 28°C in a stationary state, with manual shaking once a day, for duration of 60 days.
At different time points (10, 20, 30, 40, and 60 days) of incubation, the flasks were withdrawn in duplicate, and the contents were mixed with distilled water. The crude enzyme extract (CEE) was then extracted from the mixture as described in section 2.3. The solid residue was washed 2-3 times with distilled water, followed by drying at 80°C. The dried residue was subsequently analyzed for cellulose content using the method described by Updegraff [24]. The supernatant obtained from the mixture was subjected to analysis for ligno-hemicellulolytic enzyme activities, acid-precipitable polymeric lignin (APPL) content [18], and total reducing sugars [23].
2.5. Biomass regeneration
Based on the observation that the 40-day-old fungal-treated biomass yielded the highest total reducing sugars, a portion of this biomass, along with untreated corn stover, was subjected to extraction using 0.1 N and 0.5 N NaOH. The extraction process involved soaking the biomass for 30 minutes at a solid loading of 6%. The aim of this extraction was to dissolve any loosely bound lignin residues. Subsequently, the biomass was washed repeatedly with distilled water and dried at 80°C. The cellulose content of untreated and pretreated corn stover samples was determined following NREL laboratory analytical procedure [25].
2.6. Enzymatic hydrolysis using in-house produced cellulases
The enzymatic hydrolysis was performed in 250 ml Erlenmeyer flasks. The corn stover obtained after each treatment was dried and milled to a size of 20 mesh. Two grams each of untreated (U), biologically pretreated (B), 0.1 N and 0.5 N NaOH extracted (0.1 N & 0.5 N), as well as biologically treated + NaOH extracted (B+ 0.1 N & B+ 0.5 N) samples, were separately dispensed into the flasks. To each flask, 20 ml of 0.1 M sodium acetate buffer (pH 4.5) was added, and the flasks were subjected to steaming at 15 psi for 3 h to inactivate the fungal enzymes. For enzymatic hydrolysis, an in-house produced cellulase preparation from Aspergillus niger NS-2 [26] was added at a dose of 5 filter paper units (FPU) per gram of dried substrate and made the final volume to 30 ml with buffer. To prevent microbial contamination, 0.1% formaldehyde was also added to each flask. The hydrolysis process was carried out at a temperature of 50°C in a water bath shaker (Grant, UK) at a speed of 150 rpm for duration of 144 hours. Samples were withdrawn at regular intervals of 24 h, centrifuged at 10,000 rpm for 10 min, and analyzed for reducing sugars using the dinitrosalicylic acid (DNSA) method [23].
2.7. Statistical analysis
All the values reported in the study are presented as mean ± standard deviation, based on three independent experiments with three replicates each. Statistical analysis was performed using one-way analysis of variance (ANOVA) with the Holm–Sidak method. The software used for statistical analysis was SigmaPlot from Systat Software (San Jose, CA).
3. Results and Discussion
3.1. Microorganism
The efficiency of microorganisms in delignification depends on their ability to colonize and produce the required enzymatic machinery. In this study, the fungal strain P. phaeocomes S-1 was used for solid-state fermentation on corn stover, resulting in the co-production of lingo-hemicellulolytic enzyme cocktail. Previous research has demonstrated the ability of this fungal strain to grow and co-produce ligne-hemicellulolytic enzymes on various lignocellulosic residues, achieving high productivity. For example, on rice straw, the highest productivities reported were 10,859.51 ± 46.74 IU/gds for laccase, 22.01 ± 1.00 IU/gds for xylanase, and 10.45 ± 0.128 IU/gds for mannanase after 4 days of growth [19].
Solid-state fermentation represents a promising approach for the production of enzymes, as it offers the potential for recycling agro-industrial waste and simultaneously facilitates the delignification process, resulting in the generation of valuable products [27]. In this study, the fungal strain P. phaeocomes S-1 exhibited the ability to initiate the production of all three enzyme components of the enzyme cocktail within 24 h of incubation. The highest levels of laccase, xylanase, and mannanase productivity were achieved after 96 hours of incubation, followed by a gradual decline thereafter (Figure 1). This rapid attainment of substantial enzyme productivity within a relatively short incubation period of four days underscores the suitability of the fungal strain and the combination with corn stover for efficient delignification. Previous investigations into the co-production of diverse enzymes using corn stover have encountered limited success. For instance, Saha et al. [8] investigated the productivity rates of different lignocellulolytic enzymes when utilizing P. revisor NRRL-13108 and corn stover as substrates, and found that laccase and xylanase reached peak productivities after 8 and 27 days, respectively, with no observable mannanase activity. Méndez-Hernández et al. [28] explored the utilization of Fomes sp. EUM1 for enzyme production during the biological pretreatment of corn stover and reported that laccase reached a maximum productivity of 4.2 U/g after 7 days, whereas xylanase peaked at 77.4 U/g after 5 days of treatment. Additional studies have screened various fungal strains for corn stover pretreatment, revealing that the highest productivities of laccase and xylanase were achieved following longer fermentation durations [10]. Table 1 presents a comprehensive analysis of enzyme activities within the cocktail investigated in the current study, utilizing P. phaeocomes S-1, in comparison to similar studies conducted on various lignocellulosic residues documented in existing literature. The results of this comparison reveal favorable enzyme combinations and yields achieved in the present study, highlighting its significance. The inclusion of the selected strain adds an intriguing aspect to the study on the basis of its ability to achieve high enzyme production rates using cost-effective agro-industrial waste, which can subsequently be utilized for biofuel production following its biological pretreatment.
Figure 1.
Time course for the co-production of laccase, xylanase and mannanase of the ligno-hemicellulytic cocktail by P. phaeocomes S-1 during solid state fermentation of corn stover.
Figure 1.
Time course for the co-production of laccase, xylanase and mannanase of the ligno-hemicellulytic cocktail by P. phaeocomes S-1 during solid state fermentation of corn stover.
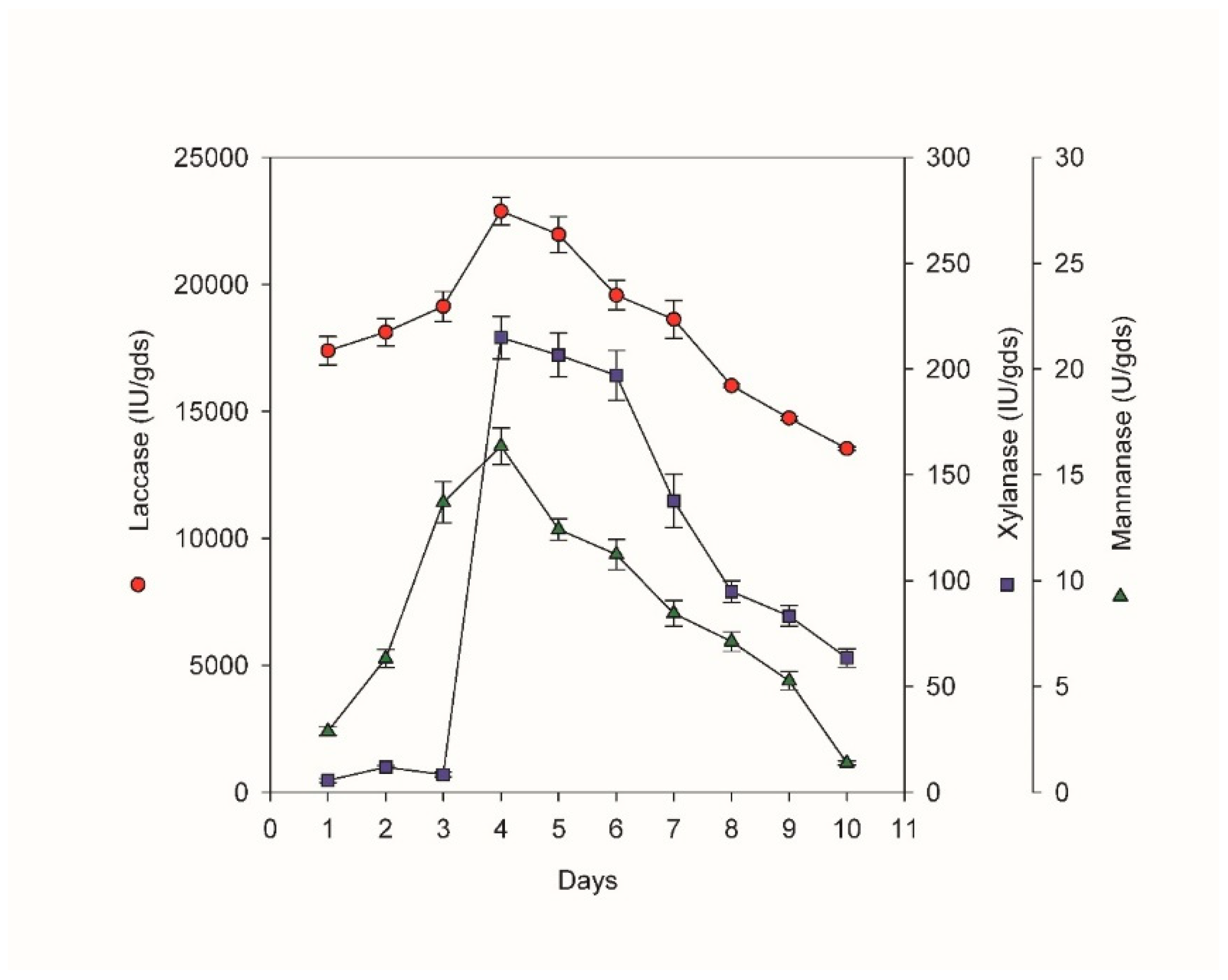
Some research groups have explored co-culturing techniques using commercially known laccase (C. comatus) and xylanase (T. reesei) producers, achieving peak productivities of 2180 U/ml and 160 U/ml, respectively, after 5 days of incubation [29]. The early appearance of high levels of laccases and hemicellulases during fermentation facilitates the delignification process and reduces the duration of biological treatment [30]. The availability of high titers of lingo-hemicellulases at the early stages of fermentation appears to be advantageous for the biological pretreatment of corn stover.

Table 1.
Comparison of ligno-hemicellulytic enzyme yields by solid state cultures of various fungi on different substrates.
Table 1.
Comparison of ligno-hemicellulytic enzyme yields by solid state cultures of various fungi on different substrates.
| Substrate | Laccase | Xylanase | Mannanase | Time (days) | Fermentation type | Fungal strain | Reference |
| (U/gds) | |||||||
| Corn stover | 22884.29 | 214.95 | 16.36 | 4 | SSF | P. phaeocomes S-1 | Present study |
| Rice straw | 10859.51 | 22.01 | 10.45 | 8 | SSF | P. phaeocomes S-1 | [19] |
| Corn stover | 6.61 | - | - | 14 | SSF | Myrothecium verrucaria | [31] |
| Corn stover | 1.5 | 4.8 | - | 7 | SSF | Ceriporiopsis subvermispora | [17] |
| Corn stover | 0.8 | 11.22 | - | 7, | SSF | Trametes hirsuta | [32] |
| Soyabean meal | 47.7 | - | 3 | SSF | Aspergillus niger | [33] | |
| Jerusalem artichoke | 2.0 | 106.5 ± 3.3 | - | 5, 20 | SSF | Ceriporiopsis subvermispora | [34] |
| Rice straw | 316.28 | - | - | 6 | SSF | Schizophyllum commune | [35] |
| Wheat straw | 1360 | - | 33.9 | 21 | SSF | Pleurotus ostreatus | [36] |
| Wheat straw | 72.9 | 98.9 | 35.5 | 21 | SSF | Trametes versicolor | [36] |
| Wheat straw | - | 1924.4 | 1.6 | 42 | SSF | Piptoporus betulinus | [36] |
| Rice bark | 2172.28 | - | - | 25 | SSF | Pleorotus ostreatus AMRL 173–6 | [37] |
| Leaf of corn cob | 29.31 | - | - | 12 | SSF | Pleurotus eryngii Han 1787 | [38] |
| Pinus tabuliformis | 2.46 | - | - | 14 | SSF | Lentinus edodes Han 1788 | [38] |
| Wheat straw | 25.51 | - | - | 5 | SSF | A. niger | [7] |
| Kraft lignin | 5.68 U/mL | - | - | 10 | SmF | Pleurotus ostreatus | [13] |
3.2. Enzyme quantification and correlation with the biomass cellulose recovery during fungal pretreatment
The pretreatment of lignocellulosic biomass is considered a major rate-limiting and cost-determining step for the production of value-added products [39]. Preserving the enriched holocellulose while exposing amorphous cellulose to hydrolytic enzymes is crucial for improving the recovery of reducing sugars. Lignin is the main obstacle in hydrolysis, and pretreatment aims to degrade the lignin-hemicellulose complex and disrupt the crystallinity of cellulose to make it more accessible to enzymes. Microbial treatment offers targeted action on the lignin-hemicellulose complex and provides the advantage of treated biomass with improved hydrolysis activities and a variety of ligno, hemicellulo, or cellulolytic enzyme systems. The productivity profiles of ligno-hemicellulolytic enzymes are critical in determining the efficiency of the biological pretreatment reaction [10,28,30].
In this study, the fungal strain P. phaeocomes S-1 quickly colonized the corn stover within the first 24 h and remained active even after longer incubation periods of 60 days, as evidenced by the profiles of ligno-hemicellulolytic enzyme levels (Table 2). No other ligninolytic enzyme activity was detected in the crude enzyme extracts, indicating that laccase played a major role in the delignification of corn stover. Laccase activity reached its peak of 336.36 U/gds after 4 days, with the lowest activity levels of 53.71 U/gds observed after 60 days. The decline in laccase productivity after 4 days may be attributed to the gradual secretion of proteases [28]. Interestingly, a second peak in laccase levels was observed, with a rapid increase reaching 166.94 U/gds after 40 days of incubation. This reappearance of the peak could be attributed to fungal autolysis, resulting in the release of intracellular enzymes into the medium [19]. Similar trends in laccase production have been observed in fungal pretreatment of corn stover with other fungal strains, albeit with lower yields compared to the present study [10,13,40].
Table 2.
Effect of biological pretreatment of corn stover by the growth of Pyrenophora phaeocomes S-1 on the disintegration of constituents due to the production of laccase, xylanase and mannanase.
Table 2.
Effect of biological pretreatment of corn stover by the growth of Pyrenophora phaeocomes S-1 on the disintegration of constituents due to the production of laccase, xylanase and mannanase.
| Fermentation Duration (days) |
Weight loss (%) |
Cellulose (%) |
APPL (mg/g) |
Total reducing sugars |
Ligno-hemicellulolytic enzymes (IU/gds) |
||
| (mg/g) | Laccase | Xylanase | Mannanase | ||||
| 0 | 7.28 | 17.09 | 80 | 19.26 | 0 | 0 | 0 |
| 4 | 15.80 | 19.21 | 192.8 | 15.78 | 336.3636 | 54.27 | 3.62 |
| 8 | 19.80 | 21.09 | 237.6 | 24.37 | 264.4628 | 46.94 | 3.29 |
| 10 | 21.80 | 22.17 | 255.2 | 28.78 | 227.2727 | 39.61 | 2.94 |
| 15 | 23.20 | 23.36 | 298 | 38.60 | 179.3388 | 37.20 | 2.59 |
| 20 | 25.80 | 25.28 | 333.6 | 42.01 | 183.4711 | 13.71 | 3.01 |
| 30 | 26.13 | 30.12 | 365.6 | 44.21 | 154.5455 | 10.76 | 2.31 |
| 40 | 28.26 | 44.25 | 384.8 | 51.83 | 133.8843 | 14.93 | 1.34 |
| 60 | 30.53 | 43.96 | 224.8 | 49.18 | 53.71901 | 3.98 | 0.44 |
During pretreatment, xylanase and mannanase were also produced, which were responsible for the degradation of the hemicellulose content of corn stover. Maximum production of all three components of the enzyme cocktail was observed after 4 days, with a gradual decline in activity during longer incubations. Xylanase activity has been observed by different researchers during fungal pretreatment of corn stover using [10] and Fomes sp. EUM1 [28] (Table 2). However, no mannanase activity has been reported during fungal pretreatment of lignocellulosic substrates.
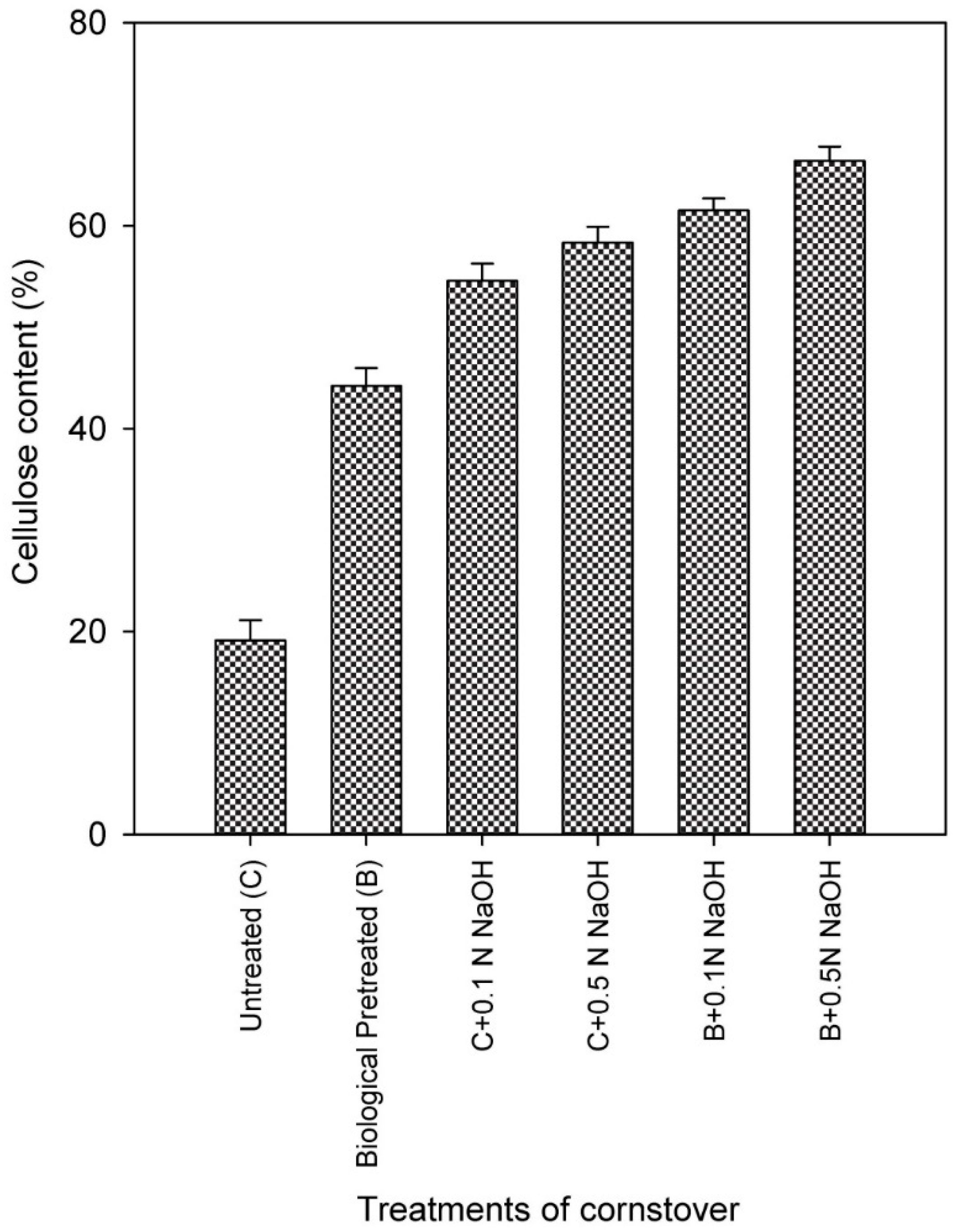
The mutual interaction between fungal growth and the generation of ligno-hemicellulolytic enzymes synergistically operates to structurally break down the biomass [41]. The progressive augmentation in weight reduction observed during extended incubation periods in this investigation corresponds to these findings. The efficiency patterns of the enzymes were utilized to evaluate the rates of substrate delignification and cellulose retrieval. Subjecting the biomass to treatment for 40 days resulted in a 28.26% decline in substrate weight and an overall retrieval of 44.25% cellulose (Table 2). As the maximum quantity of lignin is situated on the surface of the substrate, the fungal strain initiated colonization, leading to the formation of pores on the substrate. With prolonged fermentation duration, the liberated enzymes acted upon the lignin and hemicellulose, resulting in their targeted disruption, as evidenced by the amplified presence of acid-precipitated lignin (APPL) and reducing sugars in the crude enzyme extracts (Table 2). Following 40 days of fungal treatment, a maximum of 384.8 mg/g of APPL and 51.83 mg/g of total sugars were recovered in the treatment supernatant CEE. The presence of APPL in the filtrate indicates the degradation of lignin during the treatment. A direct positive correlation was observed between the released APPL content in the filtrate and the laccase productivity profile during fungal treatment of corn stover, confirming the degradation of ligno-hemicellulolytic components facilitated by fungal enzyme machinery. This explains the role of extracellular enzymes produced by the organism, which is involved in both lignin degradation and polymerization, rendering it partially soluble in water and leading to mineralization into CO2 [42]. These yields are significantly higher compared to previous reports on the treatment of rice straw with Trametes hirsuta and Myrothecium roridum for 7 days [18]. The recovery of APPLs can contribute to economic benefits, as it can be employed as an agrochemical for promoting plant growth or utilized as a raw material for plastics and resins [43]. The biologically treated biomass for 40 days was subsequently subjected to mild alkali extraction (0.1 N NaOH) at room temperature, followed by repetitive rinsing with water. This combined biological and chemical pretreatment led to further modifications in the structure, resulting in an increased cellulose content of 61.52% and 66.40% after extraction with 0.1N and 0.5N NaOH, respectively (Figure 2). Additional losses accounted for by alkali extraction and subsequent water rinsing are likely due to the removal of biologically degraded soluble lignins, hemicelluloses, and fungal proteins, along with modifications in cellulose crystallinity and hemicellulose linkages. This step is crucial as these soluble lignins and other compounds may act as inhibitors for enzymatic hydrolysis [44]. The considerable degradation of lignin and hemicelluloses observed in this study surpasses numerous prior investigations of biological pretreatment documented in the literature, thus providing a novel contribution. In a study by Ghorbani et al. [45], a maximum of 74% lignin degradation was observed after 30 days of cultivating T. viride on rice straw, even after optimizing various factors. In a recent study focusing on the biological treatment of corn stover with 26 different white rot fungi for 30 days, Saha et al. [46] reported lignin losses ranging from 1.5% to 51.4%, hemicellulose losses ranging from 14.9% to 52.1%, and higher cellulose losses of 12.8% to 50.1%, while emphasizing the significance of selective lignin degradation.
Overall, this study demonstrates the efficacy of the fungal strain P. phaeocomes S-1 in colonizing corn stover and producing ligno-hemicellulolytic enzymes, particularly laccase, for the purpose of delignification. The production profiles of these enzymes, combined with the degradation of lignin and hemicellulose, contribute to the structural breakdown of biomass and the retrieval of cellulose.
Figure 2.
Changes in percentage composition of cellulose in corn stover after sole biological treatment (40 d), sole extraction with 0.1 N NaOH (30 min; room temperature), sole extraction with 0.5N NaOH (30 min; room temperature), biological pretreatment followed by extraction with 0.1 N NaOH and biological pretreatment followed by extraction with 0.5 N NaOH.
Figure 2.
Changes in percentage composition of cellulose in corn stover after sole biological treatment (40 d), sole extraction with 0.1 N NaOH (30 min; room temperature), sole extraction with 0.5N NaOH (30 min; room temperature), biological pretreatment followed by extraction with 0.1 N NaOH and biological pretreatment followed by extraction with 0.5 N NaOH.
3.3. Enzymatic hydrolysis of pretreated biomass
The sugar yields obtained from enzymatic saccharification of fungal-pretreated corn stover of varying durations, using 5 FPU/g at pH 5.0 and 50°C, are presented in Table 3. The hydrolysis data reveals a direct correlation between lignin loss and hydrolysis efficiency. Each treatment group exhibited different levels of hydrolysis efficiency, indicating variations in the effectiveness of the treatments. Fungal pretreatment resulted in a reduction of lignin, which in turn increased the pore size on the substrate’s surface, enhancing the accessibility of cellulases and hemicellulases [30].
Untreated biomass showed resistance to cellulases, resulting in minimal yields of reducing sugars and glucose after 120 hours (5 days) of incubation (Table 3), with values of 46.67 mg /g revealing a hydrolysis efficiency of 21.98%. However, pretreating the biomass with P. phaeocomes S-1 for 40 days led to structural modifications that improved enzymatic hydrolysis. This resulted in 5.38-fold enhancements in the recovery of total reducing sugars, compared to the untreated biomass with hydrolysis efficiency of 49.11%. It should be noted that the sugar recovery achieved through biological pretreatment cannot be directly compared to conventional chemical treatments. One possible explanation for this difference is the limited disruption of the structural ligno-hemicellulose complex caused by the synergistic action of the fungus with its enzyme production profile [31].
Table 3.
Pattern of reducing sugars and glucose formation after enzymatic hydrolysis of corn stover after various pretreatments.
Table 3.
Pattern of reducing sugars and glucose formation after enzymatic hydrolysis of corn stover after various pretreatments.
| Treatment of corn stover | Time (days) | |||||
| 0 | 1 | 4 | 5 | 6 | ||
| S. No. | Total reducing sugars (mg/ml) | |||||
| 1 | Untreated (C) | 0.05 | 1.71 | 2.30 | 3.05 | 3.11 |
| 2 | Biologically pretreated (B) | 1.10 | 3.12 | 6.48 | 12.49 | 16.08 |
| 3 | C +0.1 N NaOH treated | 0.78 | 2.59 | 5.26 | 10.02 | 14.83 |
| 4 | C +0.5 N NaOH treated | 0.93 | 3.21 | 6.10 | 11.58 | 16.32 |
| 5 | B + 0.1N NaOH treated | 1.11 | 4.10 | 12.46 | 19.65 | 23.86 |
| 6 | B + 0.5N NaOH treated | 1.16 | 4.69 | 12.94 | 20.31 | 26.52 |
| Total reducing sugars (mg/g) | ||||||
| 1 | Untreated (C) | 0.78 | 25.79 | 34.54 | 45.75 | 46.67 |
| 2 | Biologically pretreated (B) | 16.61 | 46.85 | 97.32 | 187.38 | 251.22 |
| 3 | C +0.1 N NaOH treated | 11.83 | 38.98 | 79.04 | 150.34 | 222.47 |
| 4 | C +0.5 N NaOH treated | 14.07 | 48.15 | 91.51 | 173.77 | 244.83 |
| 5 | B + 0.1N NaOH treated | 16.67 | 61.53 | 186.91 | 294.83 | 358.02 |
| 6 | B + 0.5N NaOH treated | 17.52 | 70.46 | 194.18 | 304.65 | 397.84 |
During the process of biological pretreatment, the lignin present in the biomass undergoes degradation, resulting in the formation of lignin fragments. These fragments can condense and reabsorb onto the surface of cellulose, leading to the non-productive binding of cellulolytic enzymes [47]. It was observed that the sole biological treatment with P. phaeocomes S-1 did not induce any modifications in cellulose crystallinity, re-adsorption of depolymerized lignin, or the need for laccase inactivation. Therefore, certain modifications were required in the biomass prior to hydrolysis. Moreover, the presence of laccases in the hydrolysis medium can hinder the action of cellulases during saccharification [31].
To address these challenges, the treated biomass was subjected to mild alkali extraction. This step aimed to disrupt the hydrogen bonds between cellulose molecules and remove polyphenols, thereby enhancing the accessibility of the biomass to enzymatic treatment [19,31]. Extraction of the untreated biomass using 0.1 N and 0.5 N NaOH resulted in a 4.76-fold and 5.24-fold improvement, respectively, in the recovery of reducing sugars after enzymatic hydrolysis, indicating hydrolysis efficiencies of 36.73% and 37.81%, respectively. Encouraged by these enhancements, the fungal-treated biomass was also subjected to 0.1 N and 0.5 N NaOH extractions. This mild alkali treatment further enhanced hydrolysis efficiency and the action of cellulases, resulting in increased yields of reducing sugars and glucose. Extraction of the biologically treated biomass with 0.1 N and 0.5 N NaOH significantly increased the recovery of total reducing sugars, reaching values of 358.02 mg/gds and 397.84 mg/gds, respectively, corresponding to hydrolysis efficiencies of 52.42% and 53.97%, respectively. These values were 7.67-fold and 8.52-fold higher than those obtained with the untreated biomass. The 0.1 N and 0.5 N NaOH extraction of the fungal-treated biomass led to a 1.42-fold and 1.58-fold increase, respectively, in total reducing sugar recovery compared to the fungal-pretreated biomass, indicating the necessity of laccase inactivation and removal of delignified lignin residues from corn stover [31]. Since the biological pretreatment with P. phaeocomes S-1 did not alter cellulose crystallinity, it was followed by extraction with dilute NaOH to disrupt inter- and intra-hydrogen bonds between cellulose molecules and remove polyphenols, thereby rendering cellulose more accessible for enzymatic treatment [18].
These results were obtained after a 40-day treatment period, using a low dose of in-house produced crude cellulase (5 FPU/g) for enzymatic hydrolysis. This approach demonstrates a cost-effective enzymatic hydrolysis of biologically pretreated biomass utilizing low-cost in-house produced cellulase preparations. In contrast, most studies in the literature have relied on expensive commercial enzymes [30,31,47]. For instance, Ding et al. [10] investigated the analysis of C. gallica, T. versicolor, and P. sajor-caju for corn stover pretreatment and reported the highest concentration of reducing sugars (13.65 g/L) using 20 FPU/g of commercial cellulases after 60 hours of hydrolysis with P. sajorcaju over 25 days. Similarly, Saha et al. [8] compared 26 white-rot fungi for the biological pretreatment of corn stover and achieved a maximum of 394 ± 13 mg/g of reducing sugars after 30 days of C. stercoreus fermentation, utilizing a commercial enzyme cocktail consisting of 2 FPU cellulase, 5 U β-glucosidase, and 530 U xylanase per biomass. Recent research has suggested that combining chemical and biological pretreatments is more effective than using either method alone [48]. The yields obtained in this study were significantly higher than those obtained by enzymatic hydrolysis of rice straw co-treated with Sphingobacterium sp. LD-1 and 4%/6% NaOH/Urea [49]. Saritha et al. [42] achieved a 52.59% saccharification efficiency of rice straw subjected to fungal pretreatment followed by extraction with mild alkali after the addition of commercial cellulase preparation. The saccharification efficiency is directly proportional to the amount of enzyme used for hydrolysis. In our study, an appreciable efficiency of 53.97% was obtained with a significantly low enzyme dose of in-house produced cellulose preparation compared to 56% achieved by Ghorbani et al. [45] using 15 FPU/g of commercial cellulase and 75.6% achieved by Arora et al. [44] employing 29 FPU of commercial cellulose preparation per gram of biologically + chemically pretreated rice straw. A comparison with some earlier studies on the hydrolysis of biologically pretreated corn stover is presented in Table 4.
The recovery of 397.84 mg/gds of reducing sugars achieved through the combined biological pretreatment and mild alkali extraction, along with the addition of 5 FPU/g of crude cellulases, indicates the potential for further improvement in saccharification efficiency. One way to enhance this efficiency is by optimizing the dosage of enzymes used. By determining the optimal dose levels of in-house or commercial enzyme preparations, the sugar yield can be significantly increased.
Additionally, optimizing the solid-state cultivation conditions can contribute to improved saccharification efficiency. Factors such as the particle size of the feedstock, inoculum size, temperature, and moisture levels can be fine-tuned to create the ideal conditions for maximizing sugar extraction. By carefully adjusting these parameters, the accessibility of cellulose to enzymatic treatment can be enhanced, leading to higher yields of reducing sugars.
Moreover, considering cost reduction is important in the overall process. By utilizing in-house enzyme preparations or exploring cost-effective commercial enzyme options, the economic feasibility of the saccharification process can be improved. This may involve evaluating different enzyme sources, production methods, and dosage levels to find the most efficient and cost-effective approach. Overall, through careful optimization of enzyme dosage, solid-state cultivation conditions, and cost considerations, it is possible to significantly enhance the saccharification efficiency and increase the sugar yield from the biomass.
Table 4.
Comparison of fungi aided biological pretreatments of corn stover as reported by various workers.
Table 4.
Comparison of fungi aided biological pretreatments of corn stover as reported by various workers.
| Substrate |
Fungi and duration |
FPU |
Hydrolysis efficiency (%)/ reducing sugars (mg/g) |
Reference |
| Corn stover alkali extracted |
P. phaeocomes S-1 40 days |
5 FPU/g | 53.97% 397.84 23.86g/L |
Present study |
| Corn stover | Bacillus sp. | 20 FPU/ g | 56% 55.50±0.74 |
[30] |
|
Corn stover |
P. sajor-caju 25 days |
20 FPU Sigma Cellic® CTec2 |
71.24% 13.65 g/L |
[10] |
| Corn stover | Fomes sp. EUM1 | 0.5 FPU/ml | 34.1% 147.4 |
[28] |
| Corn stover | Ceriporiopsis subvermispora | 5 FPU/g 20 xylanase |
21.02% glucose yield | [50] |
| Corn stover |
Cyathus stercoreus NRRL-6573 30 days |
3 commercial enzyme (cellulase, β-glucosidase, hemicellulase) | 394 ± 13 g/L | [46] |
| Corn stover | Pycnoporus sanguineus FP-10356-Sp | 3 commercial enzyme (cellulase, β-glucosidase, hemicellulase) | 393 ± 17 g/L | [46] |
| Corn stover | Phlebia brevispora NRRL-13108). | 3 commercial enzyme (cellulase, β-glucosidase, hemicellulase) | 383 ± 13 g/L | [46] |
This study presents innovative strategies to enhance the enzymatic saccharification of biomass that has undergone fungal pretreatment. The researchers observed that reducing the lignin content of the biomass improved the efficiency of hydrolysis. Furthermore, the fungal pretreatment increased the size of the pores in the biomass, thereby enhancing the accessibility of enzymes. However, additional modifications were necessary to address the issues related to cellulose crystallinity and laccase inhibition. To overcome these challenges, a mild alkali extraction process was employed. This process aimed to disrupt the hydrogen bonds and remove polyphenols, thereby increasing the susceptibility of the biomass to enzymatic treatment. Both the untreated biomass and the fungal-pretreated biomass exhibited significantly improved recovery of reducing sugars following extraction with NaOH. Remarkably, the study demonstrated that a low-cost crude cellulase preparation, produced in-house, was as effective as commercial enzymes. This finding offers a cost-effective solution for enzymatic hydrolysis. These findings provide valuable insights for optimizing the process of enzymatic saccharification and reducing production costs in the biofuel and biorefinery industries. The combination of fungal pretreatment, mild alkali extraction, and the utilization of low-cost cellulase preparations holds great promise for achieving efficient saccharification of biomass.
4. Conclusions
Significant progress has been made in utilizing corn stover as a substrate for the cost-effective production of ligno-hemicellulolytic enzymes. This study stands out by demonstrating the simultaneous production of laccase, xylanase, and mannanase on corn stover through a four-day solid-state fermentation process. Notably, P. phaeocomes S-1 was able to selectively degrade the lignin-hemicellulose complex in corn stover that had undergone a 40-day pretreatment, resulting in enhanced cellulose recovery. Subsequent hydrolysis using an in-house produced cellulase preparation, after combining P. phaeocomes S-1 treatment with 0.5 N NaOH, yielded a remarkable 8.52-fold increase in reducing sugars. This combined treatment approach holds great promise for the advancement of biological pretreatment methods. It offers an exciting avenue for the development of efficient and advanced strategies in the field of biomass processing. The current study presents a potential avenue to enhance the efficiency of hydrolysis by optimizing the dosage of enzymes, which can result in a substantial reduction in hydrolysis costs through the utilization of in-house enzyme preparations.
Author Contributions
S.R.: investigation, writing—original draft preparation, writing—review and editing; R.S.: investigation, writing—review, and editing; A.S.: investigation, writing—original draft preparation, writing—review and editing; S.K.S.: Conceptualization, writing—review and editing, supervision. All authors have read and agreed to the published version of the manuscript.
Funding
The authors gratefully acknowledge the financial assistance provided by University Grants Commission (UGC) and Department of Science and Technology (DST), Ministry of Science & Technology, Government of India under SAP and PURSE-II respectively. Ms. Shubhangi Rastogi is also thankful to University Grants Commission (UGC), India for providing her a research Fellowship under UGC-NET scheme.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sarkar, N.; Ghosh, S.K.; Bannerjee, S.; Aikat, K. Bioethanol production from agricultural wastes : An overview. Renew. Energy. 2012, 37, 19–27. [CrossRef]
- Soni, S. K.; Parkash, O.; Manhas, R.; Tewari, R.; Soni, R. Value added products from lignocellulosic agricultural residues: an overview. Int. j. food ferment. technol. 2019, 9(2), 101-115.
- Yadav, M.; Balan, V.; Varjani, S. Multidisciplinary Pretreatment Approaches to Improve the Bio-methane Production from Lignocellulosic Biomass. Bioenerg. Res. 2023, 16, 228–247. [CrossRef]
- Tian, L.; Branford-White, C.; Wang, W.; Nie, H.; Zhu, L. Laccase-mediated system pretreatment to enhance the effect of hydrogen peroxide bleaching of cotton fabric. Int. J. Biol. Macromol. 2012, 50, 782–787. [CrossRef]
- Kaur, M.; Gill, M. K.; Sharma, S.; Kocher, G. S.; Sodhi, H. S. Biological Pretreatment Strategies for Second-Generation Lignocellulosic Biomass to Enhance Ethanol Production. In Agroindustrial Waste for Green Fuel Application; Singapore: Springer Nature Singapore. 2023, pp. 169-203.
- Sobti, R. C.; Sharma, A.; Soni, S. K. Applications of Biotechnological Techniques in Mitigating Environmental Concerns.; CRC Press: Boca Raton, FL, USA, 2022; pp. 249–312.
- Hasan, S.; Anwar, Z. Khalid, W. Afzal, F. Zafar, M. Ali, U. Aljobair, M. O. Laccase production from local biomass using solid state fermentation. Fermentation, 2023, 9(2), 179.
- Saha, B.C.; Kennedy, G.J.; Qureshi, N.; Cotta, M.A. Biological pretreatment of corn stover with Phlebia brevispora NRRL-13108 for enhanced enzymatic hydrolysis and efficient ethanol production. Biotechnol. Prog. 2017, 33, 365–374. [CrossRef]
- Waghmare, P.R.; Khandare, R. V.; Jeon, B.H.; Govindwar, S.P. Enzymatic hydrolysis of biologically pretreated sorghum husk for bioethanol production. Biofuel Res. J. 2018, 5, 846–853. [CrossRef]
- Ding, C.; Wang, X.; Li, M. Evaluation of six white-rot fungal pretreatments on corn stover for the production of cellulolytic and ligninolytic enzymes, reducing sugars, and ethanol. Appl. Microbiol. Biotechnol. 2019, 103, 5641–5652. [CrossRef]
- Qin, X.; Su, X.; Luo, H.; Ma, R.; Yao, B.; Ma, F. Deciphering lignocellulose deconstruction by the white rot fungus Irpex lacteus based on genomic and transcriptomic analyses. Biotechnol. Biofuels, 2018, 11, 1–14. [CrossRef]
- Sun, S.; Sun, S.; Cao, X.; Sun, R. The role of pretreatment in improving the enzymatic hydrolysis of lignocellulosic materials. Bioresour. Technol. 2016, 199, 49-58.
- Franco, P. C.; Shiraishi, I. S.; Dekker, R. F.; Barbosa-Dekker, A. M.; Borsato, D.; Angilelli, K. B.; Daniel, J. F. Optimization of laccase production by Pleurotus ostreatus Florida and evaluation of metabolites generated during Kraft lignin biotransformation. Waste Biomass Valori. 2023, 1-9. [CrossRef]
- Bak, J.S.; Kim, M.D.; Choi, I.G.; Kim, K.H. Biological pretreatment of rice straw by fermenting with Dichomitus squalens. N. Biotechnol. 2010, 27, 424–434. [CrossRef]
- Taniguchi, M.; Suzuki, H.; Watanabe, D.; Sakai, K.; Hoshino, K.; Tanaka, T. Evaluation of pretreatment with Pleurotus ostreatus for enzymatic hydrolysis of rice straw. J. Biosci. Bioeng. 2005, 100, 637–643. [CrossRef]
- Tiwari, R.; Rana, S.; Singh, S.; Arora, A.; Kaushik, R.; Agrawal, V.V.; Saxena, A.K.; Nain, L. Biological delignification of paddy straw and Parthenium sp. using a novel micromycete Myrothecium roridum LG7 for enhanced saccharification. Bioresour. Technol. 2013, 135, 7–11. [CrossRef]
- Wan, C.; Li, Y. Microbial delignification of corn stover by Ceriporiopsis subvermispora for improving cellulose digestibility. Enzyme Microb. Technol. 2010, 47, 31–36. [CrossRef]
- Mohanram, S.; Rajan, K.; Carrier, D. J.; Nain, L.; Arora, A. Insights into biological delignification of rice straw by Trametes hirsuta and Myrothecium roridum and comparison of saccharification yields with dilute acid pretreatment. Biomass Bioenerg. 2015, 76, 54-60. [CrossRef]
- Rastogi, S.; Soni, R.; Kaur, J.; Soni, S.K. Unravelling the capability of Pyrenophora phaeocomes S-1 for the production of ligno-hemicellulolytic enzyme cocktail and simultaneous bio-delignification of rice straw for enhanced enzymatic saccharification. Bioresour. Technol. 2016, 222, 458–469. [CrossRef]
- Jhadav, A.; Vamsi, K.K.; Khairnar, Y.; Boraste, A.; Gupta, N.; Trivedi, S.; Patil, P.; Gupta, G.; Gupta, M. Optimization of production and partial purification of laccase by Phanerochaete chrysosporium using submerged fermenation. Int. J. Microbiol. Res. 2009, 1, 9–12. [CrossRef]
- Bailey, M.J.; Biely, P.; Poutanen, K. Interlaboratory testing of methods for assay of xylanase activity. J. Biotechnol. 1992, 23, 257–270. [CrossRef]
- Markovič, O.; Slezárik, A.; Labudová, I. Purification and characterization of pectinesterase and polygalacturonase from Trichoderma reesei. FEMS microbiol. Lett. 1985, 27(3), 267-271.
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [CrossRef]
- Updegraff, D.M. Semimicro determination of cellulose inbiological materials. Anal. Biochem. 1969, 32, 420–424. [CrossRef]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Crocker, D. Laboratory Analytical Procedure: Determination of Structural carbohydrates and Lignin in Biomass. Golden, colorado: National Renewable Energy Laboratory, 2011, pp. 1-15.
- Bansal, N.; Janveja, C.; Tewari, R.; Soni, R.; Soni, S.K. Highly thermostable and pH-stable cellulases from Aspergillus niger NS-2: properties and application for cellulose hydrolysis. Appl. Biochem. Biotechnol. 2014, 172, 141–56.
- Gao, W.; Lei, Z.; Tabil, L.G.; Zhao, R. Biological Pretreatment by Solid-State Fermentation of Oat Straw to Enhance Physical Quality of Pellets. Hindawi J. Chem. 2020, 3060475, 1-13. [CrossRef]
- Méndez-Hernández, J.E.; Loera, O.; Méndez-Hernández, E.M.; Herrera, E.; Arce-Cervantes, O.; Soto-Cruz, N.Ó. Fungal Pretreatment of Corn Stover by Fomes sp. EUM1: Simultaneous Production of Readily Hydrolysable Biomass and Useful Biocatalysts. Waste Biomass Valori. 2019, 10, 2637–2650. [CrossRef]
- Ma, K.; Ruan, Z. Production of a lignocellulolytic enzyme system for simultaneous bio-delignification and saccharification of corn stover employing co-culture of fungi. Bioresour. Technol. 2015, 175, 586–593. [CrossRef]
- Wu, Y.; Guo, H.; Rahman, S.; Chen, X.; Zhang, J.; Liu, Y. Biological pretreatment of corn stover for enhancing enzymatic hydrolysis using Bacillus sp. P3. Bioresour. Bioprocess. 2021, 8, 1-10. [CrossRef]
- Su, Y.; Yu, X.; Sun, Y.; Wang, G.; Chen, H.; Chen, G. Evaluation of Screened Lignin-degrading Fungi for the Biological Pretreatment of Corn Stover. Sci. Rep. 2018, 8, 1–11. [CrossRef]
- Sun, F. H.; Li, J.; Yuan, Y. X.; Yan, Z. Y. Liu, X. F. Effect of biological pretreatment with Trametes hirsuta yj9 on enzymatic hydrolysis of corn stover. Int. Biodeterior. Biodegradation, 2011, 65(7), 931-938. [CrossRef]
- Vitcosque, G. L.; Fonseca, R. F.; Rodríguez-Zúñiga, U. F.; Bertucci Neto, V.; Couri, S.; Farinas, C. S. Production of biomass-degrading multienzyme complexes under solid-state fermentation of soybean meal using a bioreactor. Enzyme Res. 2012. 248983, 1-9. [CrossRef]
- Zhu, N.; Liu, J.; Yang, J.; Lin, Y.; Yang, Y.; Ji, L.; Yuan, H. Comparative analysis of the secretomes of Schizophyllum commune and other wood-decay basidiomycetes during solid-state fermentation reveals its unique lignocellulose-degrading enzyme system. Biotechnol. biofuels, 2016, 9, 1-22. [CrossRef]
- Asgher, M.; Wahab, A.; Bilal, M.; Iqbal, H. M. N. Lignocellulose degradation and production of lignin modifying enzymes by Schizophyllum commune IBL-06 in solid-state fermentation. Biocatal. Agric. Biotechnol. 2016, 6, 195-201. [CrossRef]
- Valášková, V.; Baldrian, P. Estimation of bound and free fractions of lignocellulose-degrading enzymes of wood-rotting fungi Pleurotus ostreatus, Trametes versicolor and Piptoporus betulinus. Res. microbial. 2006, 157(2), 119-124. [CrossRef]
- Melanouri, E. M.; Dedousi, M,M; Diamantopoulou, P. Cultivating Pleurotus ostreatus and Pleurotus eryngii mushroom strains on agro-industrial residues in solid-state fermentation. Part I: Screening for growth, endoglucanase, laccase and biomass production in the colonization phase. Carbon Resour. Convers. 2022, 5, 61-70. [CrossRef]
- An, Q.; Li, C.S.; Yuan, Y.N.; Dou, X.Y.; Wang, Y.H.; Guo, S.; Chen, Z.; Ping, A.Q.; Zhang, T.X.; Yang, Q.Y.; Han, M.L. Utilization of Agroindustrial Wastes for the Production of Laccase by Pleurotus eryngii Han 1787 and Lentinus edodes Han1788. BioResources, 2023, 18(1), 570-583. [CrossRef]
- Pinheiro, V.E.; Preto, R.; Preto, R. Screening and cocktail optimization using experimental mixture design : enzymatic saccharification as a biological pretreatment strategy. Biofuels, Bioprod. Biorefining, 2021, 15(5), 1–14.
- Sankaralingam, R.; Sengottuvelan, B.; Venkat, P.; Selvaraj, M. Arunachalam, V. Natarajan, J. Experimental investigation on varying flame characteristics of benzoic resin solid fuel pellets. Renew. Energy, 2020, 147, 1500–1510. [CrossRef]
- Moiceanu, G.; Bianca, S.; Ferdes, M. Microorganisms and Enzymes Used in the Biological Pretreatment of the Substrate to Enhance Biogas Production : A Review. Sustainability, 2020, 12(17), 7205. [CrossRef]
- Saritha, M.; Arora, A.; Nain, L. Pretreatment of paddy straw with Trametes hirsuta for improved enzymatic saccharification. Bioresour. Technol. 2012, 104, 459-465. [CrossRef]
- Savy, D.; Piccolo, A. Physical–chemical characteristics of lignins separated from biomasses for second-generation ethanol. Biomass bioenergy. 2014, 62, 58-67. [CrossRef]
- Arora, A.; Priya, S.; Sharma, P.; Sharma, S.; Nain, L. Evaluating biological pretreatment as a feasible methodology for ethanol production from paddy straw. Biocatal. Agric. Biotechnol. 2016, 8, 66-72. [CrossRef]
- Ghorbani, F.; Karimi, M.; Biria, D.; Kariminia, H. R.; Jeihanipour, A. Enhancement of fungal delignification of rice straw by Trichoderma viride sp. to improve its saccharification. Biochem. Eng. J. 2015, 101, 77-84. [CrossRef]
- Saha, B. C.; Qureshi, N.; Kennedy, G. J.; Cotta, M. A. Biological pretreatment of corn stover with white-rot fungus for improved enzymatic hydrolysis. Int Biodeterior Biodegradation, 2016, 109, 29-35. [CrossRef]
- Baramee, S.; Siriatcharanon, A.; Ketbot, P.; Teeravivattanakit, T.; Waeonukul, R.; Pason, P.; Tachaapaikoon, C.; Ratanakhanokchai, K.; Phitsuwan, P. Biological pretreatment of rice straw with cellulase-free xylanolytic enzyme-producing Bacillus firmus K-1: Structural modification and biomass digestibility. Renew. Energy. 2020, 160, 555-563. [CrossRef]
- Martínez-Patiño, J.C.; Lu-Chau, T.A.; Gullón, B.; Ruiz, E.; Romero, I.; Castro, E.; Lema, J.M. Application of a combined fungal and diluted acid pretreatment on olive tree biomass. Ind. Crops Prod. 2018, 121, 10–17. [CrossRef]
- Dai, Y.; Si, M.; Chen, Y.; Zhang, N.; Zhou, M.; Liao, Q.; Liu, Y. Combination of biological pretreatment with NaOH/Urea pretreatment at cold temperature to enhance enzymatic hydrolysis of rice straw. Bioresour. Technol. 2015, 198, 725-731. [CrossRef]
- Huang, W.; Wachemo, A. C.; Yuan, H.; Li, X. Modification of corn stover for improving biodegradability and anaerobic digestion performance by Ceriporiopsis subvermispora. Bioresour. Technol. 2019, 283, 76-85. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



