
Submitted:
13 June 2023
Posted:
14 June 2023
You are already at the latest version
Abstract
The plethora of bio-functional compounds present in fermented alcoholic beverages like wine, as well as the valorization of bioactives from their wineries/breweries by-products like grape pomace and grape seed, has gained significant interest in the functional foods sector. Wine has always accompanied humanity, for religion or for health. For example, Christians are using wine in the “Holy Communion”, following what Jesus Christ himself did and said during the Last Supper, while the benefits of the wine consumption in moderation was embedded in all Mediterranean civilizations, as it is mentioned by the wise King-Prophet David in his psalms, but also by Hip-pocrates and other wise men, in a Greek-based philosophy of “All things in Moderation” (Cleobulus of Lindos; 6th century BC). In modern times, and after the outbreak of research on wine benefits since the 90’s, when the term “French Paradox” was introduced in the US public during a CBS show, recent evidence has outlined that these benefits are derived by the synergisms of its bio-functional compounds and their digestion-derived metabolites. Within this article, the proposed health benefits of moderate wine consumption, as a functional component of a balanced diet (i.e. the Mediterra-nean Diet), against inflammation-related chronic disorders, is thoroughly reviewed. The various bio-functional compounds of both wine and wineries’ by-products, such as their bioactive phe-nolics, unsaturated fatty acids, polar lipids, and dietary fibers, and their functional antioxidant, anti-inflammatory and antithrombotic health promoting properties are also thoroughly evaluated. The mechanisms of actions and synergism, by which the health benefits are elicited, are also ex-plored. Functional properties of non-alcoholic wine products are also introduced. Emphasis is also given to applications of wineries’ by-products bioactives, as ingredients of bio-functional foods, supplements and nutraceuticals. Limitations and future perspectives for this popular functional alcoholic beverage (wine) ant its rich in bioactives by-products are also addressed. .
Keywords:
wine
; grape pomace
; grape seed
; by-products
; bioactives
; phenolics
; polar lipids
; anti-inflammatory
; anti-oxidant
; Mediterranean Diet
1. Introduction
Making of wine is one of the most well-established fermentation processes in human history, while the vinification was/is one of the important traditions that passed from one generation to the next one, especially in the Mediterranean and Middle East territories, which usually concluded to established festivities during the Greek-Roman period. Since then, and especially during the industrialization, making of wine has become an industrial process for making several types of this alcoholic beverage in order to be available all around the globe. Most types of wines are fermented alcoholic beverages comprising roughly 12-14% v/v ethanol. Their production is achieved through the fermentation of the juice from crushed ripe grapes in the presence of yeasts. Throughout the vinification process, grapes are processed, and alcoholic fermentation takes place. The methods applied during the process differ among the varieties of grapes used and wines produced [1].
Red wine contains a large variety of phenolic compounds which sets it aside from alternative alcoholic beverages. Maceration is a process unique to red winemaking and it hugely impacts the phenolic composition of wine. The presence of phenolic compounds is important in relation to the sensory and protective properties of wine [1]. Wine polyphenolic composition varies depending on the variety of grape, geographical location, as well as soil and weather conditions. There is a high abundance of flavonoids and phenolic acids present in red wine which can also be described as microbiota metabolites, formed by all wine polyphenols except stilbenes [2].
The abundant group of bioactive compounds known as polyphenols, including resveratrol, phenolic acids, anthocyanins, and several flavonoids have been found to possess potent antioxidant and anti-inflammatory activities, as well as anti-platelet aggregation and anti-atherogenic properties [3,4,5,6]. In addition, such bioactive phenolics have also been found to inhibit the activities and reduce the synthesis and the levels of the potent inflammatory mediator, platelet-activating factor (PAF), by inhibiting its binding on its membrane receptor and thus by inhibiting the PAF-associated inflammatory signaling and by reducing the PAF-related inflammatory manifestations [5,6,7]. Such bioactive compounds that can reduce the levels and/or inhibit the activities of thrombo-inflammatory mediators like PAF and thrombin, may potentially reduce the risk of developing chronic disorders [5,6,7].
Apart from the abundant phenolics, grapes, must, wine and grape pomace have also been found to contain bioactive polar lipids, with strong anti-inflammatory and anti-thrombotic properties that attribute to the bio-functional activities and protective effects of grape products and by-products [1,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23]. According to the analysis of the biological activity of total lipids in wine, the polar lipids, and especially the glycolipids, and some phospholipids rich in unsaturated fatty acids like oleic, linoleic and linolenic acids were the most effective of the wine lipid fractions against specific inflammatory pathways, including those associated with PAF [8,9,10,11]. It seems that such bioactive lipid compounds pre-exist both in grapes and in their yeasts on which fermentation is based and subsequently bioactive lipid compounds also exist in the final wine fermented product [8,9,10,11], as well as in the main by-product of wine-making, grape pomace [16,17,22,23].
Inflammation involves the innate response of the immune system to combat the causative agent of tissue damage and minimize its effects, stimulate the healing of wounds and reinstate tissue homeostasis. When inflammation becomes unresolved and chronic, then it can lead to further loss of tissue dysfunction and homeostasis, increasing the risk and onset of inflammation-related chronic disorders such as atherosclerosis, cardiovascular diseases (CVD), diabetes, and cancer metastasis [6,7,24]. Wine and grape pomace polyphenols and bioactive polar lipids have the potential to reduce inflammatory markers, therefore consumption of wine, wine extracts and extracts of grape pomace can lessen the risk of developing atherosclerosis and other chronic disorders [1,2,3,4,5,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23]. Therefore, the presence of bio-functional compounds in wine, such as polyphenols and polar lipids, with strong anti-inflammatory, antithrombotic and antioxidant properties, further supports the health benefits observed by moderate consumption of wine [1,2,3,4,5,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23].
According to evidence from epidemiological studies and trials, there are many proposed health benefits associated with the moderate consumption of red wine [1,11,12,13,14,18,19,20]. For instance, moderate wine consumption is closely correlated with less endothelial dysfunction and low-grade inflammation, which help prevent the manifestation of inflammation-related chronic disorders [25]. Moreover, apart from the beneficial health effects of such wine’s bio-functional compounds, health benefits have also been attributed to the probiotic potential linked to moderate red wine consumption. For example, it has been shown that polyphenols in red wine can beneficially regulate the growth of select favorable gut microbiota in humans [26].
However, the health benefits associated with red wine are limited to its consumption in moderation. The low to moderate consumption of wine with meals is a characteristic trait of the Mediterranean diet. Adherence to the Mediterranean diet is reported to contribute to health benefits such as attenuating the cardiometabolic risks factors associated with the onset of CVD [7,27,28]. The Mediterranean diet has become more prevalent in recent years as people are better educated in the health benefits arising from the consumption of a glass of wine as part of a balanced diet. Wine differs from other alcoholic beverages and its moderate consumption not only does not increase the risk of chronic degenerative diseases but is also associated with health benefits particularly when included in a Mediterranean diet model due to its rich content in anti-inflammatory, antithrombotic and anti-oxidant compounds [1,11,14,18].
Nevertheless, the association between alcohol consumption and corresponding health effects are represented by a J-shaped curve. the alcohol content of wine should not be disregarded, as excessive consumption of wine can lead to detrimental health effects. Among these undesirable effects is the increased risk of developing CVD [29,30]. Excessive alcohol consumption is also linked with liver cirrhosis and for several types of cancers related to the digestive and respiratory tract [29,30]. In addition, sound clinical judgment should be used in determining whether alcohol consumption is an appropriate recommendation for each patient, taking into account factors such as contraindications with other medications, while there is a small portion of the population that may be prone to addiction, or fail to moderate consumption, and therefore may present a hazard to society via acts of violence, accidents, and spousal, child and elder abuse, as well as incurring detrimental effects on their own health. Therefore, special effort must be made to promote behavioral education to prevent abuse, especially among young people.
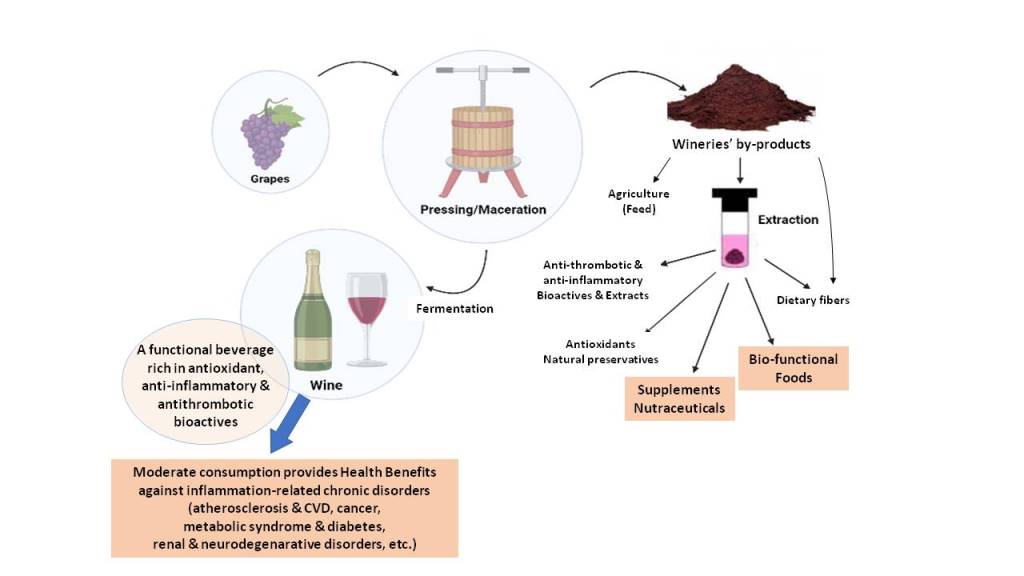
On the other hand, throughout wine making, several by-products are also derived, with the main ones being grape pomace and grape seed oils [22,23]. Many areas of the food industry, including wineries, bring about copious quantities of such waste as by-products. Lately it has been proposed that such by-products, including grape pomace, are attractive sustainable sources of bioactive compounds, which if appropriately recovered can be valorized as ingredients for functional foods, food supplements or even nutraceuticals. Many of these compounds are found to be beneficial to human health and therefore their up-cycling is becoming a new trend in food science. These waste products are available in large quantities in such sustainable sources, at very low cost, making the recovery of these products use both economically viable and environmentally friendly.
More specifically, significant amounts of grape pomace are generated from grapes during wine making, which accounts for 20% of the original grape used [22,23]. This waste generated by the wine industry creates contamination and pollution issues, costly disposal issues, and financial loss. Traditionally, this waste is undervalued and used as animal feed or fertilizer, due to not extensive research on the appropriate methodology for the recovery and valorization of its compounds. Grape pomace is distinguished from other food by-products due to its high content in high-value compounds such as dietary fibre, lipid bioactives, phenolics and other natural antioxidants [22,23]. These compounds are relatively inexpensive to extract and can be employed as ingredients in foods or even in nutritional supplements, providing a significant economic advantage. Furthermore, many of these bioactive compounds have been linked to several health benefits, including anti-inflammatory reactions, cardioprotective and antidiabetic effects [10,16,17,21,22,23].
Attempts to recover bioactive compounds have recently increased, with the goal of utilizing them as added ingredients to help improve functionality. The fortification of existing food products through novel applications aims to enhance functional and biochemical properties, while lowering their environmental impact and food industry costs. Similarly, there is a lot of interest in using grape by-products as food additives, nutraceuticals, and components in functional foods as a result of the growing attention to agricultural sustainability and consumer preference for natural over synthetic substances. Numerous studies have been conducted to understand the bioactivity of grapes, with the main effort focused on its antioxidant activity, which is associated with its high content of polyphenols acting as free radical scavengers [22,23].
Thus, in this review paper, an overview of the current literature on the bioactive compounds, functional properties and associated health benefits and potential applications of the fermented alcoholic beverage, wine, and its grape pomace by-product is thoroughly conducted. Emphasis is given to the composition, nutritional content-value, and health benefits of these wine products and by-products, and especially in their important bio-functional constituents, such as bioactive lipids, phenolics, dietary fibres, probiotics and other beneficial compounds with significant antioxidant, anti-inflammatory, and antithrombotic properties with associated health benefits. Specific frameworks are also considered regarding the industrial valorization of such produce, as well as the production of innovative functional foods and nutraceuticals against the risk of chronic diseases. This review provides a comprehensive picture of the strengths and limitations, which can aid future studies in advancing the applications of wine, grape pomace and of their bioactives.
2. Bio-functional compounds and health benefits of the fermented alcoholic beverage, wine and of its by-products
2.1. Composition, nutritional value, bio-functional components, and functional properties
2.1.1. Types of wine; emphasis to red wine production, composition and nutritional value
Wine is a traditional alcoholic beverage acquired by the processing of grapes and the alcoholic fermentation of the grape must. Following harvest, grapes enter the winery to finalize the vinification process and undergo destemming, crushing, pressing, maceration, and alcoholic fermentation [1]. Wine comprises a vast range of chemical constituents such as aldehydes, ketones, esters, minerals, lipids, phenolics, organic acids, soluble proteins, sugars, and vitamins. The complexity of the compounds present in wine is mainly due to the composition of the grape and the fermentation process involved [1,11,18,23]. Thus, the quality of wine is linked to the composition and variety of the grape [23,30]. Red wines are formed by the alcoholic fermentation of musts combined with solid parts of the berry, the skins, and seeds. In contrast, white wine is exclusively obtained from the fermentation of grape juice [30].
The maceration process has a significant impact on the phenolic composition of wine. It requires the inclusion of skins and seeds with the must to allow for the extraction of polyphenols, enhancing the wine in terms of color and flavor [1]. Maceration facilitates the production of metabolites such as acetaldehyde from yeasts which react with phenolic compounds. The absorption of anthocyanins by yeast cells alters the color and phenolic composition of the wine [1]. By extending the maturation time, elevated alcohol levels can result in the over-extraction of components from the solid element of grapes, particularly the seeds. Increases in phenolic and flavonoid content, in addition to the trans- and cis-resveratrol concentration occur due to the lengthened maturation process [31]. The separation step from the skins and seeds following maceration in combination with the pressure of mash can significantly elevate the phenolic content in wines and their antioxidant properties [31]. Maturation of red wine takes place in barrels and its main purpose is to soften the astringent and bitter taste due to the high phenolic content. Red wines require a more lengthened maturation process as a result of their high phenolic content to enhance the quality, character and flavor of the wine [31]. Regardless of the alterations to red wine during maturation, the level of antioxidant capacity remains consistent throughout the process [31]. Red wine is distinct as it contains approximately 10-fold more phenolic compounds than white wine. The high phenolic content is a result of the fermentation of grape juice combined with skins, grape pieces, and seeds.
The total phenolic content of red wine is strongly correlated with its antioxidant activity. The antioxidant capacity of red wine is not defined by one single compound, instead, there are several components that exert a synergistic effect on the antioxidant activity [32]. Apart from the antioxidant properties, some wine compounds have also shown several other bio-functional properties, including anti-inflammatory and antithrombotic potential, such as the phenolic content of wine and its bioactive polar lipids [1,2,3,4,5,6,7,8,9,10,11,12,13,14,18,19,20,25,30,31,32]. Importance has recently been given to the anti-inflammatory and protective effects of not only wine bioactive phenolics, but also to the various bioactive components present in wine and their positive contribution to health, such as bioactive polar lipids [1,5,6,7,8,9,10,11,12,13,14,16,17,18,19,20,21,22]. The functionality of red wine is strongly associated with its high polyphenolic content that exerts anti-inflammatory and antioxidant effects, with lower but significant levels of bioactive polar lipids contributing to antithrombotic and anti-inflammatory activities [1,2,3,4,5,6,7,8,9,10,11,12,13,14,18,19,20,25,30,31,32].
In addition to red wine, other wine types also exhibit protective effects against atherosclerosis and platelet aggregation as well as potent antioxidant effects. Apart from the dominant focus on red wine, in terms of the anti-inflammatory and protective effects of its high polyphenolic content, there are alternative wines that can also contribute to beneficial health effects. For example, white wines are developed in the absence of grape skins, which directly impacts the concentration of phenolic compounds within the wine, providing a lower distribution of phenolic compounds in white wine compared to red wine, (2567mg/GAE/l vs. 239mg/GAE/L). Nevertheless, the antioxidant capacity of white wine is quite significant [9,10]. The extension of the maceration process of white wines with grape pomace has resulted in the development of an orange-amber color, a distinct tannic flavor as well as the enhancement of the phenolic content and antioxidant capacity [1]. Moreover, consumption of white wine (Robola) along with a meal equally decreased postprandially platelet aggregation induced by the inflammatory mediator, platelet activating factor (PAF), and maintained low triacylglycerol levels during postprandial elevation, similarly to the same observed beneficial effect during the intake of red Cabernet Sauvignon variety along with the same meal. The presence of PAF inhibitors in both white and red wine seems to equally influences such beneficial effects [9,10,11,12]. Overall, the incorporation of moderate wine consumption in the diet may help prevent the clinical manifestations involved in the development of inflammation-related chronic disorders.
2.1.2. Wineries by-products - composition and nutritional value
Grape pomace is the main by-product of wineries and accounts for 20% of processed grape weight, generated through the pressing of grapes for production of must. For every 6 liters of wine, 1kg of grape pomace is produced, though this may depend on the type of press used and the grape varietal. The composition of grapes and its subsequent pomace vary according to intrinsic factors such as variety and maturity, and extrinsic factors, such as soil conditions and agricultural practices [22,23]. This affects not only the sensory properties in the wine it makes, but also the practicality of successive applications of fresh pomace. Furthermore, red wine pomace and white wine pomace are inherently different due to the winemaking process. In red winemaking, the grape skin and pulps are fermented along with the juice before being filtered out, while white winemaking has the juice pressed out and the pomace discarded early in the process. This leads to red wine pomace containing a higher concentration of polyphenols, with white wine pomace retaining their fermentable sugars. The weight of the grapes is made up of 2-5 % seeds and 38-52% of the solid waste matter created during the winemaking process. The seeds include around 40% fibre, 10% protein, 10%-20% lipids, and another 30-40% carbohydrates, complex phenolics, vitamins, and minerals, while overall grape pomace contain around 17-89% total dietary fibre, 16-64% insoluble fibre, 4-14% protein, 1-14% lipids, 12-40% carbohydrates, 0.3-9% total phenolic content and another 2-9% ash (Table 1) [16,22,23,33,34,35,36,37,38,39,40,41,42].
Ultimately, wineries’ by-products pose a difficult waste disposal challenge for the wine industry. Though much of grape pomaces are being composted and reintroduced into the vineyards to complete the carbon cycle, the utilization of grape pomace has been largely inefficient. The pomaces are often converted to cattle feed or used as materials for biofuel digesters, and in some environmentally detrimental cases, for landfill disposal or incineration. Following the growing sustainability concerns, as well as consumer demand for natural products, there is immense potential in bioconversion of these by-products and of their bioactives into functional foods, supplements, or additives. With the integration of in vitro techniques and high-end bioprocess engineering technologies, the continuous production of nutritionally improved wine pomace-derived metabolites could be achieved. In addition, another valuable by-product is also the grape seed oil, which is rich in both unsaturated lipids and lipophilic/amphiphilic phenolics and thus can also promote lower production costs being more competitive than others and represent a new food source for human consumption in the food industry [22,23].
2.2. Bio-functional components and associated health benefits
2.2.1. Wine and its by-products phenolic bioactives with antioxidant, anti-inflammatory and anti-thrombotic beneficial properties
Phenolic compounds are the most abundant bioactive components in wine [1,2,3,4,32,43,44], and are primarily present in the seeds and skins of grapes, except for hydroxycinnamic acids which are found in lesser amounts in the flesh [1,22,23,36,37,42]. The high number of factors influencing phytochemical composition and content leads to a very wide range of phenolic content. In general, red grape varietals have more phenolic compounds than white grape varieties, and the red vinification process, which includes longer maceration durations, supports nearly total extraction. Vineyard interventions similarly may have a significant impact on grape phenolic compositions. Different phenolic compositions can also be observed among different wine varieties, The estimated overall polyphenolic composition in red wines ranges from 1531 and 3192 mg of gallic acid equivalents (GAE) per liter, with white wine having a phenolic composition between 210 and 402 GAE/L [43], while the remaining wineries’ by-products like grape pomace contain also valuable amounts of phenolic compounds, the main classes of which are shown in Table 1 (expressed in GAE/L) [16,22,23,33,34,35,36,37,38,39,40,41,42].
Such phenolic compounds of both wine and grape pomace can be divided into two main groups, flavonoids, and non-flavonoids [36,42,43,44,45]. The structures of the most characteristic classes and compounds of these two groups of phenolics are shown in Figure 1. Flavonoids attribute to 85% of the phenolic constituents in red wine and are primarily present in the skins of grapes, as well as in wineries’ by-products (wine) and by-products (grape pomace) too, while they comprise mainly by flavan-3-ols, anthocyanins, and flavonols (Figure 1). Flavan-3-ols include monomers like catechin, epicatechin, and polymers (polyphenols) like proanthocyanidins [1,2,3,4,36,37,42,43,44]. Flavanols, catechins and epicatechins, are the most complicated subclass of flavonoids that contribute to the different sensory qualities and structure of wines, by interacting with proline-rich proteins in the mouth to produce an astringent sensation and bitter taste [30]. In the non-flavonoids group of phenolic compounds, several phenolic acids (cinnamic/hydroxycinnamic acids and benzoic/hydroxybenzoic acids), phenolic aldehydes, volatile simple phenolics, stilbenes, tannins and coumarins are classified (Figure 1) [36,37,42,43,44,45,46].
Apart from their role on the organoleptic characteristics of wine, a plethora of studies have indicated that phenolic compounds are associated with the health benefits of moderate consumption of wine, as well as with the potential use of wineries’ by-products as a sustainable source of functional compounds, since such phenolic compounds have exhibited potent antioxidant, anti-inflammatory and antithrombotic properties, with the most bioactive ones being the flavonols, flavanols, and anthocyanins from the flavonoids and resveratrol from the non-flavonoids [1,2,3,4,5,6,22,23,25,30,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53]. The health-promoting effects of wine and wineries by-products’ phenolic compounds was initially attributed on their strong antioxidant capacity [1,4,20,21,32,36,37,38,39,40,41,43,51,52,53,54,55]. It has also been proposed that the greater the polyphenolic content, the greater the antioxidant activity [1]. Phenolic bioactives like stilbenes, flavonoids and flavan-3-ols derivatives of wine, can cross cell membranes and effectively demonstrate their antioxidant potential through the reduction in reactive oxygen species (ROS) within cells [54].
The antioxidant activity of wine flavonoids is linked to their ability to scavenge of free radicals, which reduces the vulnerability of low-density lipoprotein (LDL) to oxidation and prevents the manifestation of endothelial dysfunction and atherosclerosis as a result [51,53,54,55,56]. Intake of moderate wine consumption with a meal can significantly enhance plasma total antioxidant capacity (TAC) and counteract oxidative lipid damage and the activation of NF-κB signaling [55], suggesting that the antioxidant effects of moderate wine consumption, influenced by the presence of wine polyphenols. Additionally, red wine and its polyphenolic constituents, play an essential role in CVD prevention by means of its antioxidant, antithrombotic, anti-inflammatory and antiatherogenic properties [1,2,3,4,30,43,48,49,50,51].
This is not surprising since the important pathophysiological processes of our body, oxidative stress, inflammation and thrombosis are intertwined, and if any of them and/or all of these processes is/are unregulated, then it/they can contribute to the unregulation of the other ones, promoting thus a vicious cycle of a continuous induction of oxidative stress and thrombo-inflammatory manifestations, which can conclude to the onset and development of several chronic disorders, including atherosclerosis, CVD, insulin resistance, hypertension and cancer [6,7,9,24,43,56]. This process can be impeded through the incorporation of wine and wineries’ by-products bioactive phenolic compounds in the diet, which promote not only anti-oxidant but also a plethora of anti-inflammatory health benefits [1,2,3,4,5,6,7,14,18,19,20,25,30,42,43,44,46,48,49,50,53,56]. Even though wine and wineries’ by-products phenolics are not essential nutrients, they can contribute to health through several pleiotropic effects, and subsequently they also do not fit in the classic and rigorous pharmacological definitions, as they can be modified by organisms before they interact with targets, can have different targets depending on their concentration, and do not have a univocal pharmacological mechanism of action [42,43,46]. Therefore, a reductionist approach or studying of one single mechanism of action should not be followed with respect to the benefits of wine and wineries’ by-products phenolics, because this can limit their classification as only being free radical scavengers and antioxidants, while they possess manifold mechanisms of actions, including anti-inflammatory actions [43].
More specifically, the polyphenolic composition of wine and wineries’ by-products is of significance in terms of the associated anti-inflammatory and antioxidant actions that attribute to the greater antioxidant serum activity, increased resistance of LDL peroxidation and thus inhibition of LDL oxidation, stimulation of the level of high-density lipoprotein (HDL), the promotion of vasorelaxation, the inhibition of platelet aggregation and lowering of platelet sensitization and adhesiveness, as well as the inhibition of the activities and synthesis of inflammatory mediators like PAF and cytokines, as well as a plethora of anti-inflammatory effects on several inflammatory genes, mediators, receptors, adhesive molecules, signaling pathways and thrombo-inflammatory manifestations [1,2,3,4,5,6,7,14,16,18,19,20,22,23,25,30,33,38,42,43,44,46,48,49,50,53,56,58,59,60,61]
Thus, the moderate consumption of wine is not only linked with an increase in antioxidant capacity, but it is also interlinked with anti-inflammatory beneficial effects and a reduction in pro-inflammatory markers [1,2,4,5,6,18,19,20,25,30,43,48,49,50,56,60,61], which have been attributed to its phenolic compounds. Moreover, according to recent literature, the mode of action by which polyphenols exert their beneficial properties appears to involve the interplay between molecular signaling pathways and regulators of cellular actions involved in inflammation [2,4,5,6,18,30,42,43,48,49,50,59]. Both flavonoids and non flavonoids have exhibited such benefits, while differences on the effects of compounds from each group have also been reported [2,3,30,43]
The different interactions that wine flavonoids may have with key biological targets are linked to the unique structure of each and their structure activity relationships are crucial for the health protective properties of wine and grape pomace based products against several diseases such as CVD, cancer, obesity, neurodegenerative diseases, diabetes, allergies and osteoporosis [4,22,23]. Moreover, several studies have clearly revealed the anti-inflammatory protective effects of flavonoids of wine on health, by lessening the risk of the development and onset of inflammation-related chronic disorders [2,4,18,30,43,44,48,49,50,59]. In addition, pleiotropic anti-inflammatory benefits on vascular health have also been observed in trials based on monomeric or polymeric flavonoids [62]. Furthermore, consumption of polymeric flavonoids like procyanidins (polyphenols that are polymers of catechin and/or epicatechin units) within a diet is correlated to the reduced risk of developing type 2 diabetes (T2D) and CVD [63].
Apart from flavan-3-ols and flavonols, anthocyanins are also one of the most important water-soluble flavonoid pigments in nature, with grapes being amongst the fruits with the highest concentration in anthocyanins [22,23,42]. Subsequently, anthocyanins are found in red wine and especially in red wine’s grape pomace wastes, and in lesser amounts in white wines and their grape pomaces [22,23,42]. Pharmacokinetics studies has shown indication that anthocyanins are rapidly absorbed into the bloodstream shortly after consumption, as well as their positive implication and essential role in the prevention of a variety of diseases, including cancer, neurodegenerative disorders and CVD through anti-inflammatory activities and improvement of the immune system [4,30,42,48,49].
Among the non-flavonoids’ group of phenolic compounds in grapes, wine and grape pomace, a plethora of studies have highlighted the anti-oxidant and anti-inflammatory health benefits and preventative-therapeutic properties of resveratrol [1,2,3,5,6,7,18,42,43,44,50,64,65,66,67]. The benefits of resveratrol have been documented since 90’s as the main bioactive component of wine (concentrations of 0.1–14.3 mg/L), when Drs. Michel de Lorgeril and Serge Renaud talked about the “French Paradox”, the observation of an unusually low rate of heart disease among Southern French people who drink a lot of red wine, despite a high saturated fat diet, and the theoretically inhibitory effects of wine consumption against lipid peroxidation, in the “60 Minutes” CBS show [43,64,66]. At the time the “free radical/antioxidant hypothesis” was in full swing and it was commonplace to believe that eating and drinking (poly)phenols would scavenge free radicals and prevent their noxious effects, for example by inhibiting LDL oxidation [43,64].
This granted red wine (poly)phenols, namely resveratrol, immediate popularity not only as a possible explanation for the “French paradox,”, but also trigger the vast amount of well-funded research [43,64,66]. From then, several studies have highlighted resveratrol as a pan-assay interference compound, hence linking to a wide variety of signaling pathways. Subsequently, resveratrol’s usage as a nutraceutical and a therapeutic agent for a variety of disorders has been extensively investigated in in vitro, ex vivo and in vivo studies in animal models and humans, as well as in preclinical and clinical trials as a natural molecule. Accumulating evidence suggests that resveratrol is a highly pleiotropic molecule that modulates numerous targets and molecular functions [65,66,67,68,69,70,71,72,73,74]. Epidemiological studies indicate that the Mediterranean diet, rich in resveratrol, is associated with a reduced risk of atherosclerosis. Resveratrol is believed to decrease circulating low-density lipoprotein cholesterol levels, reduce cardiovascular disease risk; it reduces lipid peroxidation, platelet aggregation and oxidative stress [65,66,67,68,69,70,71,72,73,74].
For example, resveratrol’s ability to protect against inflammation and oxidative stress occurs via nuclear erythroid 2-related factor 2 (Nrf2) signaling pathway [75]. Moreover, the cardio-protective anti-inflammatory effects of resveratrol have been demonstrated at nutritionally relevant concentrations, by decreasing the over expression of intercellular and vascular cell adhesion molecules, as it inhibits the induced by inflammatory cytokines activation of coronary arterial endothelial cells, such as the inhibition of tumor necrosis factor (TNF)-α-induced nuclear factor Kappa B (NF-Κb) activation of these cells and the subsequent expression of inflammatory genes [68,76,77]. Thus, resveratrol supplementation may partially protect against CVD especially during the early atherosclerotic phase by its anti-inflammatory effects, as well as by reducing circulating levels of important chemotactic chemokines, such as the monocyte chemoattractant protein-1 (MCP-1/CCL2) and macrophage inflammatory protein-1 alpha (MIP-1α/CCL3), which regulate migration and infiltration of monocytes/macrophages and are induced and involved in various diseases [68,76].
Resveratrol has also improved the TNF-α-induced endothelial dysfunction during the interaction of Caco-2 cells and endothelium [78]. Apart from the inhibitory effect of resveratrol on the actions of inflammatory cytokines, resveratrol is also associated with the reduction of the levels of pro-inflammatory cytokines, such as interleukin (IL)-1 and TNF-α, which are related to the pathologies of cancer and CVD [79]. The anti-cancer potential of resveratrol was also demonstrated by the induced autophagic cell death and reduction in cell viability found in oral cancer cells but absent from normal cells [80]. Several other very recent studies within 2023 have also indicated the anti-tumor effects of resveratrol and other wine phenolics, through their (pro-)apoptotic and immune-regulatory effects [6,50,80,81,82,83,84,85,86,87], while it has also previously been reported that resveratrol suppresses tumor metastasis by its effects on platelets and by inhibiting platelet-mediated angiogenic responses, and thus resveratrol is a potential therapeutic drug for the prevention of tumor metastasis by interrupting the platelet-tumor cell amplification loop [87].
Resveratrol and other phenolics from wine and grape pomace exhibit a plethora of beneficial effects by affecting platelet functions [12,14,16,18,22,23,30,42,43,50,65,66,67,68,69,70,71,72,73,74,87,88,89,90,91]. These effects can take place through several mechanisms, while similar mechanisms exist also in several other cells and tissues, in which resveratrol acts beneficially. For example, resveratrol, and wine and grape pomace phenolics in general have shown strong cardioprotective properties and anti-inflammatory protection against chronic disorders by strongly inhibiting platelet aggregation and the activation and sensitization of platelets induced by the activities and release of thrombotic and inflammatory mediators, such as PAF, thrombin, collagen, fibrinogen, arachidonic acid (AA), Thromboxane A2 (TxA2), TxB2, ADP and epinephrine, and thus by exhibiting potent anti-inflammatory and antithrombotic protection against inflammation and thrombosis related chronic disorders [12,14,16,18,22,23,30,42,43,50,65,66,67,68,69,70,71,72,73,74,87,88,89,90,91].
For example, resveratrol-induced inhibition of platelet metabolism and TXA2 release may lead to a reduction of platelet function and thrombus formation in patients with type 2 diabetes, and thus resveratrol may be beneficial to prevent vascular complications as a future complementary treatment in aspirin-resistant diabetic patients [88]. More specifically, resveratrol reduced collagen-induced thrombi by over 50%, in both the blood of healthy and diabetic patients, TXA2 release by 38% in healthy platelets and by 79% in diabetic platelets. Resveratrol also reduced the activities of enzymes responsible for glycolysis and oxidative metabolism in the platelets of both groups [88]. Such effects of resveratrol on platelets appear to be mediated through cyclooxygenase-1 (COX-1) repression, which results in decreased TxA2 production and thus inhibition of platelet aggregation, rather than through cyclooxygenase-2 (COX-2) that synthesizes prostacyclins as antiplatelet factors in vascular endothelium [69]. Resveratrol can reduce platelet aggregation by forming stable complexes in platelet COX-1 channels, as well as by inhibiting the arachidonate-dependent synthesis of inflammatory agents, such as TXB2, hydroxyheptadecatrienoate, and 12-hydroxyeicosatetraenoate [69,89].
Moreover, resveratrol inhibits thrombin-induced platelet aggregation through decreasing Ca2+ release from its stores and inhibiting store-operated Ca2+ influx into platelets [91], while it has also been proposed that resveratrol may inhibit platelet aggregation induced by epinephrine and other mediators by increasing NO production [70]. In addition the antiplatelet effect of resveratrol, and subsequently its antithrombotic and anti-inflammatory benefits, both in vitro and in vivo, has also been attributed to its potential to modulate gene/protein expression of tissue factor (TF) and its functions, since TF that can be produced by several cells and especially under inflammatory cytokines induction, is a well-known thromboplastin, which activates thrombosis through binding to and further stimulation of coagulation factor VII as a principle initiator of extrinsic coagulation cascade [69].
Furthermore, recent studies have outlined that resveratrol can target and activate AMP-activated protein kinase (AMPK), having an important role in reducing fat accumulation, cholesterol synthesis, and inflammatory cytokines, while it can also activate-stimulate at an amplitude of ~10-fold the mammalian versions of the sirtuin family of proteins, SIRT1, and subsequently all the pathways regulated by SIRT1, which deacetylates histones and non-histone proteins, such as transcription factors, and thus affects important processes like metabolism, stress resistance, cell survival, cellular senescence, inflammation-immune function, endothelial functions, and circadian rhythms [66,67,68]. Stimulation of SIRT1 and AMPK boosts the eNOS activity in human coronary arterial endothelial cells and increases NO production and mitochondrial biogenesis, which triggers vasodilation and decreases atherosclerosis [68], while the activation of SIRT1 by resveratrol seems to protect against disorders such as improper metabolic regulation, inflammation, and cell cycle abnormalities [66].
Interestingly, resveratrol-induced activation of SIRT1 downregulates the expression of the receptor of PAF (PAFR) on platelets via proteosomal and lysosomal pathways, and that this downregulation inhibits platelet aggregation in vitro and pulmonary thrombus formation in vivo [88]. It has also been proposed that resveratrol might improve cardiovascular health by affecting the gene expression for producing PAFs [69].
In addition, resveratrol has also been found to reduce the levels of PAF by inhibiting its synthesis and thus reducing the inflammatory status [5,6,7,92]. For example, resveratrol has inhibited PAF-synthesis in human mesangial cells [5], as well as in U-937 macrophages under inflammatory conditions [92], suggesting a potential antitumor effect of resveratrol [6], as well as protective effects against several PAF-associated inflammatory and thrombotic chronic diseases [7].
Resveratrol content in wines varies from 0.43 to 62.65 µM, and depends on several factors such as grape variety, climate and winemaking process. Resveratrol is highly absorbed in the intestine, presents low bioavailability and is rapidly excreted, while consumption of pure resveratrol at doses higher than those present in wines results in a low content of this compound in plasma. These findings may challenge the common idea that resveratrol is the main phenolic compound associated with cardioprotective effect. Consequently, other stilbenes and other phenolic compounds present in wines such as flavonoids, have also been proposed to act beneficially [70]. Astringin is another stilbene for which early reports have shown promising results, citing anti-oxidant activity and a free radical scavenging ability more potent than resveratrol, while other natural and synthetic analogues/derivatives of resveratrol have also been extensively studied for enhanced beneficial effects against inflammation, thrombosis and associated disorders [66,67,68,69,71,72,93,94,95].
The majority of the reviewed phytochemicals in wine and grape pomace require relatively high doses to be active on several cells and tissues in vitro and these doses may further increase when plasma is present, for instance in case of resveratrol. As Professor Visioli et al (2020) have highlighted, animal studies often employ very high doses of grape/wine (poly)phenols, such as resveratrol, with results that cannot be readily transferred to humans, who would need to ingest several grams of extracts to replicate the same effects, while discrepancies between animal and human effects and potential toxicity of high doses of resveratrol have also been recently reviewed [43]. Therefore, the current high dosing is one of the drawbacks for the testing of such compounds for possible therapeutical intervention [4,43,74].
Several phenolic compounds are very weak (if at all effective) in vivo direct antioxidants, while for kinetic reasons they do not scavenge free radicals and their bioavailability is generally so low that they contribute very little to the integrated cellular antioxidant machinery, which is mostly composed of enzymes [43]. Thus, other limitations are the often unclear bioavailability and metabolic absorption of the phytochemicals and of their metabolites, the pharmacokinetic profile in blood and non-platelet effects, which for the cardiovascular system may be positive or negative [4,43,74]. There is still a gap between the knowledge of wine flavonoids bioavailability and their health-promoting effects The beneficial effects of dietary phenolic compounds are affected by their low intestinal absorption as well as their differential bioavailability and interactions with plasma and gut microbiota that generates broad shifts in the plasma metabolome and gut microbiota composition [4,26,43,74,96,97,98,99,100,101]. It is estimated that the small intestine only absorbs 5-10% of consumed dietary polyphenols following enzymatic glycosylation. The remaining dietary polyphenols enter the colon intact and undergo degradation by the gut microbiota yielding simple phenolic acids that are absorbed into the bloodstream [96]. Thus, it is apparent that the gut microbiota facilitates the bioaccumulation of polyphenols and their associated metabolites
Hence, the molecular forms of phenolic compounds that contribute to health benefits are not limited to those ingested but also include their associated metabolites created in vivo by the intestinal microbiota. Lately, plenty of researchers correctly use (poly)phenols’ metabolites in their in vitro studies, focusing on low-molecular weight (LMW) polyphenol-related components consisting of free anthocyanins, free proanthocyanins, pyranoanthocyanins as well as smaller amounts of phenolic acids and resveratrol. The hurdle here is the difficulty of synthesizing such metabolites, which are often produced by the organism in different forms, while the relatively recent discovery of microbiota-synthesized metabolites amplifies the list of potential biologically-active molecules produced by the body after the ingestion of (poly)phenol-rich foods [43]. Therefore, more in vivo results as well as studies focused on phenolic metabolites are still required. Moreover, it is also necessary to better understand how biological interactions (with microbiota and cells, enzymes or general biological systems) could interfere with phenolics’ bioavailability. Nevertheless, the moderate consumption of red wine is positively correlated with the beneficial modulation of gut microbiota [100]. The ability of red wine consumption to balance the growth of select gut microbiota in humans indicates the potential probiotic benefits linked to the incorporation of red wine polyphenols into the diet. For example, resveratrol plays a significant role in the regulation of the gut microbiome, protection of the intestinal barrier and in the inhibition of intestinal inflammation [96,97,98,99,100,101].
In addition, the acquired knowledge on effects of certain phytochemicals, as platelet-inhibiting and anti-inflammatory compounds, can be employed for the further selection and chemical modification of these in the design of effective antiplatelet and anti-inflammatory drugs. In other words, those phytochemical compounds with a proven effect on platelets, thrombus formation and inflammation can trigger new ways for drug discovery. This could develop into new antiplatelet and anti-inflammatory drugs, and also to potentiate the action of current antiplatelet drugs, as several of the phytochemicals seem to have priming effects on platelets. Interestingly, combinations of wine and grape pomace phytochemicals with other bioactives present can have synergistic effects on platelets and inflammatory signaling, which will further enhance the priming.
2.2.2. Bioactive lipid compounds of wine and wineries’ by-products
Apart from the plentiful phenolics, several other bioactive compounds have also been identified in wine and wineries’ by-products, such as their bio-functional lipid bioactives. The lipids present in grapes/yeast undergo chemical modification throughout the fermentation process with the most bioactive lipids being derived at the final wine product. Even though wine contains lower amounts of lipids compared to grapes and the remaining grape pomace, still for the last 20 years Prof. C.A. Demopoulos and colleagues (Prof. Antonopoulou SA, Dr. Fragopoulou E, Dr. Nomikos T, Dr. Tsoupras A, et al) have highlighted that the wine amphiphilic polar lipids, such as several phospholipids and especially glycolipids (Figure 2), originating from the grapes, yeasts and wine must, as well as due to the fermentation process, are highly bioactive and have exhibited strong anti-inflammatory, antithrombotic and anti-atherogenic cardioprotection [7,8,9,10,11,12,13,14,18,19,20]. Grape pomace has also been found to contain similar bioactive polar lipids along with its rich content on phenolics. Such bioactive polar lipids usually contain bio-functional unsaturated fatty acids (UFA) within their structures in a combination that usually favors an anti-inflammatory potential [7,8,9,10,11,16,17,22].
Such UFA are usually the monounsaturated fatty acid (MUFA) oleic acid (OA; 18:1 omega-9) and the long-chain polyunsaturated fatty acids (PUFA), linoleic acid (LA: 18:2-omega-6), gamma linoleic acid (GLA; 18:3 omega-6) and alpha linolenic acid (ALA; 18:3 omega-3) [16] (Figure 2A). UFA have shown on their own strong anti-inflammatory and antithrombotic properties against several inflammatory and thrombotic mediators, including PAF, promoting an anti-inflammatory potential and health benefits against several inflammation-related disorders [7,102,103,104,105,106,107]. UFA control cell fluidity, the attachment of certain enzymes to cell membranes, and the transmission of signals and other metabolic activities. They are involved in the manufacture of eicosanoids, leukotrienes, prostaglandins, and resolvins, which have anti-inflammatory, anti-arrhythmic, and anti-aggregatory properties [102]. Several of them promote cardiovascular health, and others increase visual function and cognition in newborns and adults [102].
Similarly, grape seed oils are high value by-products for the extraction of bioactives from grape pomace [108]. The seeds consist of up to 20% oil, which is rich in UFA [109]. Inversely, the grape seed oil has a low polyphenol content, which can be traced to the hydrophilic nature of these compounds [110]. The most abundant fatty acid in grape seed oil is LA, but the nutritional role of this omega-6 UFA has grown progressively complex throughout years of study, with important functions in regulating inflammatory cellular processes. In contrast, the omega-3 fatty acids, such as ALA, perform opposing effects with well known cardioprotective effects and anti-inflammatory characteristics, reducing thromboxane synthesis and PAF, and demonstrating immense potential in inhibition of pro-inflammatory cytokine synthesis [7,102,106,107]. Thus, despite the suggestions of the possible health benefits of LA, the anti-inflammatory effect of grape seed oil with regards to its high UFA and LA content remains to be seen [111].
Nevertheless, more research has recently been contacted on the polar lipid bioactives of wine and grape pomace [7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22]. Polar lipids are amphiphilic biomolecules and important structural elements for all cells in nature, with a plethora of diverse bioactivities. Polar lipids generally contain two fatty acids usually esterified or rarely etherified to a glycerol- or a sphingosine-based backbone, and a phosphorus functional group for phospholipids or a sugar for glycolipids that is linked to a head group (Figure 2B). Bioactive polar lipids with UFA in their structures (usually at the sn2 position of their glycerol/sphingosine backbone), such as polar lipids with OA and/or ALA (Figure 2B), possess higher bioavailability of their UFA throughout the body, due to their amphiphilic properties, while most importantly polar lipids themselves possess strong anti-inflammatory and antithrombotic properties against several mediators and inflammatory pathways, by a variety of mechanisms of actions (Figure 4), with promising health benefits against atherosclerosis and CVD, cancer and metastatic procedures, renal and neurodegenerative disorders, persistent infections and associated inflammatory manifestations, allergy and asthma, sepsis, etc. [6,7,11,14,18,22,102].
For example, such bioactive polar lipids have modulated or even reduced the formation of atherosclerotic plaques, by inhibiting the activities and reducing the levels of the inflammatory and thrombotic mediator, PAF, and its atherogenic effects [7]. Since the thrombotic and inflammatory pathways of PAF are implicated in several chronic disorders, including cancer, atherosclerosis and CVD [6,7,24], the inhibition of PAF activities and the reduction of its levels by wine and grape pomace polar lipid bioactives further suggest their anti-atherogenic, cardioprotective and anti-tumor potential [6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22].
Apart from modulating PAF-metabolism towards reduced PAF-levels, such bioactive polar lipids from wine, wine-must, yeasts and grape pomace have also inhibited the inflammatory and thrombotic pathways of both PAF and thrombin, while they have reduced the platelet sensitivity, activation and aggregation induced by well-established platelet agonists, collagen, and ADP [7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,112]. It seems that the anti-inflammatory and anti-thrombotic anti-PAF properties of these polar lipid bioactives occur either through affecting beneficially PAF-metabolism towards reduction of its levels of homeostatic ones and/or through inhibiting the binding of PAF on its receptor and thus inhibiting PAF-related inflammatory and thrombotic pathways and activities (Figure 3), and subsequently reducing the risk for PAF-associated inflammatory chronic disorders such as atherosclerosis, CVD, and cancer [6,7,11,14,18,22,102].
Moreover, apart from the strong anti-inflammatory and anti-thrombotic properties of the whole structures of such bioactive polar lipids, it has also been proposed that once the rich in UFA polar lipids have surpassed the intestine barrier and are bound to and transferred from plasma lipoproteins to the cell-membranes of all tissues, there a cytoplasmic phospholipase A2 (PLA2) releases their UFA content from their structure of these membrane bound polar lipids, while the released UFA interacts with several inflammatory pathways, genes and signaling, such as the eicosanoids pathways (COX-enzymes), for reducing and resolving inflammation and the inflammatory cell-response (Figure 3) [6,7,102]. Based on these findings, several trials have been conducted based on the effects of the consumption of wine or of grape pomace extracts rich phenolics and bioactive polar lipids on thrombo-inflammatory mediators like PAF, on inflammatory cytokines’ release and on metabolic and oxidative stress responses [12,13,15,19,20,21].
Overall, the benefits of wine consumption and of grape pomace extracts are not only attributed to their phenolic content but also to other microconstituents like their bioactive lipid compounds, and especially in their glycolipids, phospholipids and their UFAs, as well as by the synergism of all these bioactives. Although bioavailable amounts of the antioxidant wine and grape pomace polyphenols have little to do with a proposed in vivo protection against oxidative stress, they still play a crucial role as primary antioxidants against oxidation of lipids, including PL and UFA compounds, in several natural sources, foods, beverages, cosmetics, and lipid extracts [113]. In extracts of several sources, a very substantial improvement in oxidative stability, bioavailability, and preservation of the bioactivities of phenolics, UFA and polar phenolic compounds can be achieved by a co-presence and synergism of all these polar compounds. Subsequently, the presence of phenolic compounds in PL extracts from any type of wine and/or grape pomace, seem to facilitate the preservation of the bioactivities of the protected PL compounds. Thus, the synergism of such wine and grape pomace bioactive compounds, which can ameliorate the oxidative stress response and inhibit the activities and/or reduce the levels of thrombo-inflammatory mediators like PAF and thrombin, seem to explain the potential of an extract rich in such polar organic bioactives, rather than of just one molecule like resveratrol, for the prevention/reduction of risk of several inflammation and thrombosis associated chronic disorders [6,7,10,11,14,18,22].
2.2.3. Bio-functional dietary fibres from wineries’ by-products
Dietary fibres are usually defined as “carbohydrate polymers that present ten or more monomeric units, which are not hydrolysed by the endogenous enzymes in the small intestine of humans”, while the main proposed benefits from their inclusion in the diet are associated with prevention against CVD, cancer and diabetes, due to glucose absorption attenuation, improvement of food transit through digestive system, diabetes, blood cholesterol decrease, constipation and obesity prevention [108]. Dietary fibre content has also exhibited antioxidant capacity, as a product that is composed of more than 50% of dietary fibre on a dried matter basis presents a natural antioxidant capacity equivalent to at least 50mg of vitamin E (when measured by the DPPH method) [108]. The daily intake recommendation guidelines of dietary fibre consumption (25–30 g/day), is not easily achieved and a need for other alternative sources than cereals, vegetables and fruits has promoted the development of commercial fibre-rich products and supplements. This brings by-products obtained from fruits processing as a potential sustainable source of dietary fibres, with a higher soluble dietary fibres content, better insoluble/soluble fibre ratio, low caloric content, and better functional properties than those obtained from cereal processing [108].
The winemaking industry by-products have those characteristics, since grape skin is a complex lignocellulosic material containing large amounts of hemicellulosic sugars that, after hydrolysis, produce solutions containing a wide variety of xylose and glucose monomers [108], while the major component of dried wine pomace is dietary fibre, taking up concentrations as high as 85% depending on grape variety, with a major fraction of these dietary fibres being insoluble fibres such as lignin, cellulose and hemi-celluloses [33,108]. Apart from the general cardioprotective, antitumor and antidiabetic health benefits attributed to the intake of the appropriate composition of dietary fibres, grape pomace, dietary fibres have also been proposed to exhibit beneficial effects on the gastrointestinal tracts due to the fibre’s high porosity and low density, while they are also a major energy source for gut microbiota, having a significant impact on the microfloral diversity and indirectly the innate immune response of the gut mucosa [33]. The grape pomace dietary fibres also moulds into a complex formation with the ability to establish a linkage with polyphenols, increasing the capacity for phenolic compounds [33]. Overall, the aforementioned health benefits suggest that dietary fibres of grape pomace have potential to be used in different food products such as bakery products, beverages and meat products, in cosmeceutical and neutraceutical-pharmaceutical areas of applications cosmetics and industries [33,108].
3. Health benefits of moderate red wine consumption and detrimental effects of alcohol abuse; a coin with two sides
The life expectancy in developing countries is continuously increasing however, there is a resulting rise in the burden of chronic diseases related to age and lifestyle, such as diabetes mellitus, cancer, atherosclerosis and CVD, neurodegenerative disorders and amyloid diseases, notably Parkinson’s and Alzheimer’s diseases, among others. Numerous studies support that adherence to Mediterranean diet or to other healthy dietary patterns provide reduction of the risk and protection against such disorders [6,7]. In recent years, extensive epidemiological studies, for instance, the Health Professionals Study, the Nurses’ Health Study (NHS) and the European Prospective Investigation into Cancer and Nutrition (EPIC) study, as well as their follow-ups, have highlighted the health benefits associated with adherence to healthy dietary patterns such as the Mediterranean diet.
The light to moderate consumption of red wine has been characterized as one of the main characteristics of the Mediterranean diet and is proven to provide protective effects against CVD. Since the Lyon Diet Heart Study, the health benefits of wine consumption, mostly red wine, were attributed at the antioxidant protection of lipoproteins and cell membranes due to wine’s rich content in phenolics, also was presented by the main PIs of the study, Drs. Michel de Lorgeril and Serge Renaud, in the “60 Minutes” CBS show since 90’s, while they also talked about the “French Paradox”, the observation of an unusually low rate of heart disease among Southern French people who drink a lot of red wine, despite a high saturated fat diet, and the theoretically inhibitory effects of wine consumption against lipid peroxidation [43,64,66]. Subsequently, the focus on the benefits of wine consumption was mainly given to its antioxidant potency and cardiovascular protection [1,2,3,4,5,6,7,18,19,20,25,30,42,43,44,48,49,50,56,60,61,64,65,66,67]. Nevertheless, as it is outlined in the Lyon Diet Heart Study, the components of food and alcoholic beverages included in the Mediterranean diet exhibit beneficial properties not only against lipid peroxidation, but they also include an inhibitory effect against platelets’ activation and aggregation [64], which was not emphasized during that show and thus was initially neglected.
Since inflammation and thrombo-inflammation have been characterized as the main cause of the onset and development of not only CVD, but also of several other inflammation-related chronic disorders [6,7,24], it is now well established that apart from the antioxidant protection, the effects of moderate wine consumption that promote health benefits include mainly 1. beneficial moderation of lipid metabolism, 2. protection against oxidative stress, inflammation, platelet activation-aggregation and thrombo-inflammation, 3. improvement of endothelial function, and 4. the modulatory effect on the gut microbiota. Moreover, in recent years, the beneficial effects of the inclusion of moderate wine consumption in the diet have been extensively researched, not only in epidemiological studies, but also in several interventions in animal models and humans, including randomized clinical trials, where it has been evidenced that moderate wine consumption with meals based on a Mediterranean diet setting, or on other healthy diets, have exhibited health benefits against not only atherosclerosis and CVD (Supplementary Table 1) [12,13,19,20,25,53,55,60,62,64,99,114,115,116,117,118,119,120,121,122,123,124,125], but also against several other pathologies and all cause mortality (Supplementary Table 2) [3,26,126,127,128,129,130,131,132,133,134,135,136,137,138,139,140,141,142,143,144,145,146,147,148,149,150,151,152,153,154,155].
More specifically, in Supplementary Table 1, several characteristic studies, interventions and clinical trials are outlined with respect to the benefits of moderate consumption of wine and its bio-functional compounds against inflammation, thrombosis, vascular inflammatory activation and adhesion of leukocytes, endothelial dysfunction, atherosclerosis and CVD [12,13,19,20,25,53,55,60,62,64,99,114,115,116,117,118,119,120,121,122,123,124,125], whereas in Supplementary Table 2, other characteristic studies, interventions and clinical trials with respect to the benefits of moderate consumption of wine and its bio-functional compounds against other inflammatory and thrombotic manifestations and inflammation-related chronic disorders, including cancer, metabolic syndrome and diabetes mellitus, .gastrointestinal disorders, pulmonary diseases, stroke, neurodegenerative diseases and depression, as well as all cause mortality, are also presented.
As it is shown in both these supplementary tables, there is vast evidence demonstrating the health benefits associated with moderate wine consumption, specifically the anti-inflammatory and protective effects. The most recent epidemiological studies are partly in agreement with the firstly reported outcomes of the benefits of wine consumption presented at the Lyon Heart study, while numerous clinical studies have now outlined the beneficial effects of wine consumption on inflammatory factors and endothelial function which are paramount in the manifestation of chronic disorders.
The observations from a range of clinical studies have shown that the chemical composition of wine, specifically its bio-functional components, are the main attributes of health benefits associated with moderate wine consumption. More specifically, such benefits are provided by the protective properties of the wine itself and its bioactive compounds, notably its polyphenolic constituents and its bio-functional polar lipids, as well as due to the positive interactions of wine and its compounds with gut microbiota. The potent anti-inflammatory and antioxidant effects of wine and its bioactive components are major contributors to beneficial health outcomes. Moderate red wine consumption has exhibited anti-inflammatory properties by reducing markers of inflammation thus, preventing the likelihood of chronic inflammation.
Therefore, the most prominent mechanisms mediating the effects of moderate wine consumption are the outcomes of the synergistic effects of these wine bioactives and their metabolites produced by digestion, are the induced by wine enhancement of endothelial function and counterbalance of inflammation, by beneficially affecting inflammatory-related biomarkers, mediators and associated thrombo-inflammatory signaling-pathways, including the reduction of the activities and levels of PAF and of other inflammatory chemokines and cytokines, as well as the down-regulation of monocyte adhesion to endothelial cells and the reduced expression of adhesion molecules. Once these mechanistic pathways are improved by moderate wine consumption then all the other mechanisms are also beneficially affected with an overall health benefit against the risk for atherosclerosis, CVD and other inflammation-related chronic disorders (Supplementary Tables 1-2).
There are also studies like the Hoorn Study, in which the anti-inflammatory and endothelial protective health benefits of red wine consumption were emphasized, suggesting even that from the components of the Mediterranean diet, mostly red wine consumption, but not fruit, vegetables, fish or dairy products, mainly protects against factors involved in the pathogenesis of CVD and other diseases, as it was associated with less endothelial dysfunction and less low-grade inflammation [23]. In addition to the anti-inflammatory and antioxidant effects associated with wine and its bio-functional components, the moderate intake of wine can also provide protective effects against the pathogenesis of a range of chronic disorders.
However, in the case of the associations of wine consumption and cancer an intense scientific debate exists. For example, huge epidemiological studies like the Health Professionals Study, the Nurses’ Health Study (NHS) and the European Prospective Investigation into Cancer and Nutrition (EPIC) study and their follow-ups, have outlined that alcohol intake is associated with the presence and development of several types of cancers in either women or men, such as cutaneous basal cell carcinoma, colorectal, skin, pancreatic and mainly breast cancer [156,157,158,159,160,161], whether an increase in the incidence of cancer is observed also in association with moderate consumption levels is yet not definitively ascertained.
Moreover, based on this assumption that alcohol is a leading risk factor for cancer, it has been observed in other studies that awareness about the other side of the coin presenting an alcohol-cancer link is low. Awareness may be influenced by perceptions of potential health benefits of alcohol consumption or certain alcoholic beverage types. A study based on data from the 2020 Health Information National Trends Survey 5 Cycle 4, a nationally representative survey of U.S. adults, estimated awareness of the alcohol-cancer link by beverage type and examined the relationship between this awareness and concomitant beliefs about alcohol and heart disease risk [162]. It was observed that low levels of accurate awareness exist on the cancer harms associated with alcohol use, including wine, beer, and liquor consumption. Awareness of the alcohol-cancer link was higher among those recognizing that alcohol use increased heart disease risk. Based on these observations, it has been proposed that the "health halo" surrounding consumption of wine and other forms of alcohol as reducing heart disease risk has lead the public to over-generalize alcohol health benefits to other diseases, including cancer, and this increases the need to address high levels of perceived risk uncertainty to help the public distinguish between the impact of alcohol on heart disease versus cancer, and to overcome other barriers to including alcohol use reduction as a cancer prevention strategy [163]. Given recent increases in U.S. population drinking rates, as well as morbidity and mortality associated with alcohol use, it was also proposed that there is a need to educate U.S. adults about the alcohol-cancer link, including raising awareness that drinking all alcoholic beverage types increases cancer risk [162,163].
Although it is clearly established that the abuse of alcohol is seriously harmful to health, much epidemiological and clinical evidence seem to underline the protective role of moderate quantities of alcohol and in particular of wine on health [50]. Wine differs from other alcoholic beverages and its moderate consumption not only does not increase the risk of chronic degenerative diseases but is also associated with health benefits particularly when included in a Mediterranean diet model. Moreover, it is worth mentioning that follow-up of such epidemiological studies [126,164,165,166] and several other studies and especially interventions and clinical trials have outlined that adherence on a moderate wine consumption pattern provides antitumor protection against several types of cancer (Supplementary Table 2).
For example, only heavy (but not light or moderate) consumption of alcohol at baseline is associated with intestinal-type noncardia gastric cancer risk in men from the EPIC cohort, while such a positive association was not observed specifically for wine consumption [164]. In addition, in a follow-up of the Health Professionals Study on the effects of alcohol consumption on prostate cancer, it was observed that cancer-free men who consumed alcohol had a slightly lower risk of lethal prostate cancer compared with abstainers, and especially among men with prostate cancer, red wine was associated with a lower risk of progression to lethal disease, while it was proposed that such observed associations seem to provide assurance that moderate alcohol consumption is safe for patients with prostate cancer [126]. Moreover, in another follow up of the Nurses' Health Study, Nurses' Health Study II, and Health Professionals Follow-Up Study, it was also proposed that in these three prospective cohorts, alcohol consumption was associated with reduced risk of pituitary adenoma, compared to almost no consumption of alcohol [165]. In another very recent follow up study of the Nurses' Health Study and the Health Professionals Follow-Up Study it was also proposed that encouraging an increased intake of specific flavonoid-rich foods and beverages, including tea and red wine, even in middle age, may lower early mortality risk [166]
However, all these effects seem primarily to be associated with the amount of alcohol consumed, while the role of the different alcoholic beverages, and of their minor components, in this regard is in fact not clearly defined. It seems that the effect of alcohol consumption on cancer and health in general is far more complex, while the type of alcoholic beverage, the quantity consumed and the frequency of consumption (Supplementary Tables 1-2), as well as the type of cancer and the overall personalized health status and genetic profile of each individual, which seem to also play a significant role in these contradictory associations. It should be mentioned however that wine and wine-derived compounds are promising chemoprotective and chemotherapeutic agents for cancer, as they have been shown to participate in several mechanisms against cancers, including deoxyribonucleic acid damage, oxidative stress, , cell proliferation, cell cycle arrest, cell apoptosis, autophagy, inflammation-related cell invasion and metastasis, immunity and metabolism, regulation of multiple signaling molecules, and gene expression [6,167].
Due to the presence of such beneficial for health bioactives mostly in wine, it has been proposed that the light to moderate consumption of alcohol in the form of wine (≤1 drink/day for women and 1 to 2 drinks/day for men), is associated with a lower risk of inflammation-related disorders, such as atherosclerosis and CVD, stroke, cancer, type 2 diabetes mellitus and all-cause mortality (Supplementary Tables 1-2) [1,6,7,11,14,19,25,43,49,50,53,55,56,60,61,64,70,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139,140,141,142,143,144,145,146,147,148,149,150,151,152,153,154,164,165,166,168,169,170,171]. Conversely, heavy drinking, (>4 drinks/day), is closely correlated with detrimental effects on health and associated with an increased risk of developing cancer, CVD and stroke (both ischemic and hemorrhagic), among others [156,157,158,159,160,161,162,163,164,165,166,168,169,170]. Among males aged 15 to 59 years, alcohol abuse is perhaps the leading cause of premature death. Excessive alcohol intake trails behind only smoking and obesity among the 3 leading causes of premature deaths in the westernized societies. As such, the risk-to-benefit ratio of drinking is less favorable in younger individuals.
Because of the opposite direction of the association between alcohol consumption and cardiovascular and cancer events, the association with all-cause mortality is complex, and J-shaped [168,169,170], with a consumption window theoretically associated with a reduction in all-cause mortality, up to 25 g alcohol per day. The J-shaped curve indicates that light to moderate alcohol consumption leads to lesser risk against chronic disorders than abstainers and heavy consumers of alcohol who are at the most risk in terms of health. A daily habit of light to moderate drinking is ideal for those who choose to consume alcohol regularly. Red wine in particular before or during the evening meal is linked with the best long-term health outcomes. Most of the studies on alcohol and health are observational, and correlation does not prove causation [156,157,158,159,160,161,162,163,164,165,166], while the recent increase of interventions and trials conducted on wine consumption have provided more robust evidence for the health benefits of wine consumption in moderation (Supplementary Tables 1-2). Subsequently, health care professionals need to advise with precaution nondrinkers to begin drinking because of the paucity of randomized outcome data coupled with the potential for alcohol abuse even among seemingly low risk individuals, and every effort must be made to promote behavioral education to prevent abuse, especially among young people. Moreover, additional research is required to evaluate and clarify the doubts that still exist.
For example, in all type of studies, including epidemiological studies, interventions and trials, the abuse of alcoholic beverages has been associated with an increased risk of chronic-degenerative diseases, including diabetes mellitus, so that there is a general diffidence towards the low/moderate consumption of wine by individuals with type-2 diabetes or at risk of developing it. Whether wine/grape derivatives must be excluded or if their low/moderate consumption could be part of the daily diet of individuals with type-2 diabetes, is still studied. Although further intervention studies on the consumption of alcoholic beverages and the development or control of type-2 diabetes are needed, the burden of evidence suggests that low/moderate wine consumption seems to provide beneficial effects (Supplementary Table 2). It has also been proposed that for older adult drinkers, registered dietitians and physicians may find benefit in inquiring about the specific alcohol types consumed, as opposed to inquiring merely about holistic alcohol intake, as part of patient consultations or routine visits, while they should also encourage older adult alcohol drinkers to consume alcohol in moderation and to consume a higher proportion of total alcohol intake from red wine in order to help reduce the risk for weight gain, metabolic syndrome, adiposity-associated health risks and diabetes.
Overall, wine is actually an alchemy of unique properties, with a rich and original composition in terms of polyphenols, other phenolic and nonphenolic bioactive components, such as polar lipid bioactived and UFA that are usually less considered, which may contribute to the alleged health effects and a protective association between low-to-moderate wine consumption and inflammation-related chronic disorders like CVD, type 2 diabetes and neurodegenerative disorder, while it does not appreciably influence the overall risk of cancer. There is therefore strong scientific evidence from Mediterranean and non-Mediterranean countries that moderate wine consumption provides protection against all cause of mortality and thus increases longevity.
Bioactive components are not the only reason for the beneficial effects associated with wine consumption; social factors also matter. The Mediterranean diet is a dietary model that is also considered healthy because it suggests consuming wine during meals. When consumed during meals, wine tends to be sipped more slowly than other alcoholic beverages and this may provide metabolic benefits. In addition, the concomitant presence of food in the stomach slows the absorption of ethanol, aiding metabolism and hepatic clearance, and lowering the peak blood alcohol concentration. The concomitant presence of food may also reduce the amount of alcohol available to the oral microbiota, which has the ability to metabolize ethanol to acetaldehyde, a compound associated with the tumor effects of ethanol in the upper gastrointestinal tract. In addition, the presence of alcohol may improve the bioavailability of wine bioactives in the food bolus, making them more assimilable and may reduce glucose bioaccessibility, which is consistent with the hypoglycemic effects observed in intervention and observational studies of moderate wine consumption [50].
Wine has always accompanied humanity, for religion or for health. For example, wine is mentioned in the Bible, where it is written that Jesus Christ himself chose wine for his Last Supper, and subsequently Christians also use wine for the re-enactment of the “Holy Communion”, while the wise King-Prophet David had also emphasized in several of his psalms the benefits of food with wine and oil, with characteristic examples being the 4th (8th verse; “ἔδωκας εὐφροσύνην εἰς τὴν καρδίαν μου· ἀπὸ καρποῦ σίτου, οἴνου καὶ ἐλαίου αὐτῶν ἐπληθύνθησαν”) and 103rd psalms (15th verse; καὶ οἶνος εὐφραίνει καρδίαν ἀνθρώπου τοῦ ἱλαρῦναι πρόσωπον ἐν ἐλαίῳ, καὶ ἄρτος καρδίαν ἀνθρώπου στηρίζει).
The reason why the original Greek versions of these psalms are extended here, is because unfortunately, the most common English translation of the original Greek phrases used in these psalms, “εὐφροσύνην εἰς τὴν καρδίαν μου” and “εὐφραίνει καρδίαν ανθρώπου” is usually the use of the simple phrase “gladdens the heart”. However, we should not neglect that the Greek language and thus several Greek words and phrases like the aforementioned ones are conceptual and thus provide a deeper meaning derived from the synthesis of their root-words (εὐ-φροσύνην and εὐ-φραίνει; from the origin-word “εὐ-“, which means “in a good way (καλώς)” and the verb “φρονώ”, which means “wisely taking care”), and subsequently the phrases mentioned in these psalms seem to have a more appropriate meaning like for example “consumption of food with wine and oil wisely takes care of and gladdens men’s heart”. All the above, further indicate the deeper philosophy of the Mediterranean diet basis, which seems to be the famous Greek phrase “Μέτρον Άριστον” (All things in Moderation), firstly mentioned by the Greek poet Cleobulus of Lindos in 6th century BC, but was engulfed since then by all Mediterranean civilizations as a way of life. I should not also be neglected that Hippocrates, the “father of modern medicine”, had also emphasized this in his phrase "Wine is a thing wonderfully appropriate to man if, in health as in disease, it is administered with appropriate and just measure according to the individual constitution" [171].
In modern times wine was responsible for the lot of ink flowed to explain the French or the Crete paradox, while a lot of research tried to study wine and its bioactives, in order to solve the mystery of its benefits. Beyond its cardiovascular effects, there are also effects on longevity, metabolism, cancer prevention, and neuroprotection, and the list goes on. Wine is a preserve of antioxidants and anti-inflammatory compounds for the winter and wine acts mostly as an alcoholic solution. Even though several epidemiological studies have suggested an association of alcohol, including alcoholic wine, with cancer and other diseases [156,157,158,159,160,161,162,163,164,165,166], still such studies only bring up associations but not evidence. There are also studies proposing health benefits from alcohol free wines [155], while the plethora of recent interventions and trials have provided more robust evidence for the health benefits of wine consumption in moderation (Supplementary Tables 1-2), making researchers to suggest that a wine without alcohol is like a “pure heresy” [171].
Wine is the elixir that by design, over millennials, has acted as a pharmacopeia that enabled man to heal and prosper on the planet [171]. From Hippocrates to Michel de Lorgeril and Serge Renaud, nutrition was the key to health and longevity, whether the Cretan or Okinawa diet, it is the small dose of alcohol (wine or sake) that allows the appropriate beneficial interaction(s) with gut microbiota and the bioavailability of their bioactive compounds and bio-functional metabolites. Moderate drinking gives a protection for diseases and a longevity potential. In conclusion, let us drink fewer, but drink better, to live older [171].
4. Recovery and valorization of bioactive compounds from wineries’ by-products as ingredients for developing health promoting functional foods, supplements and nutraceuticals
Almost a hundred million tons of grapes are globally produced annually, with the majority of them (approx. 55-65%) being used for wine production. The winemaking process generates large amounts of grape pomace, the main winery by-product that supposes an economic and environmental problem. It is estimated that every 100 kg of grapes used for winemaking yields a quarter of this amount (20–25 kg) of grape pomace, with an annually estimate of grape pomace production ranging approximately from 10 to 15 million tons worldwide [46,70]. Grape pomace is constituted by mainly of grape skin, seeds, stems, and remaining pulp. Moreover, in grape seeds, an easily obtained byproduct of grape juice and wine processing, the majority of polyphenolic content of grape is stored, which if valorized efficiently exhibits rich bioactivities, with promising application prospects in the food industry, while it can also produce grape seed oil, another valuable by-product from wineries [46,108]. The harvesting and winemaking processes are short, and a large quantity of grape pomace and/or grape seed is generated in a very short time [46,108]. Even though grape pomace and grape seed can be used for animal feeding and compost, only a small amount is reused and its disposal raises environmental concerns while representing a high cost to the industry [33,46,70,108].
After the fermentation-maceration process of wine making not all the valuable compounds of grapes are transferred to wine, and thus grape pomace and grape seed are usually rich in bioactive phenolic compounds, UFA, polar lipids and functional dietary fibers [22,33,39,42,44,46,70,108,172,173,174]. These bio-functional compounds present in grape pomace and some in grape seed and its oil exhibit a variety of bioactivities, such as antioxidant, anti-inflammatory, and anti-cancer effects, while they are also beneficial in alleviating metabolic syndrome and regulating intestinal flora [22,33,39,42,44,46,70,108,172,173,174]. In this sense, research has now been focused on the potential health properties of such wineries’ by-products in the prevention of disorders associated with oxidative stress, inflammation and thrombosis, such as endothelial dysfunction, atherosclerosis and CVD, hypertension, hyperglycemia, metabolic syndrome, diabetes, obesity, cancer and neurodegenerative disorders, among others [10,16,17,21,22,33,39,42,44,46,70,108,172,173,174,175]. Those effects are due to the bioactive compounds of wineries’ by-products and the mechanisms concern especially modulation of antioxidant/prooxidant activity, improvement of nitric oxide bioavailability, inhibition of platelet aggregation, reduction of pro-inflammatory cytokines and modulation of antioxidant/inflammatory signal pathways, as well as beneficial interactions with gut microbiota.
Grape pomace extracts can be applied in food, pharmaceutical, cosmetics and others products in the form of liquid extracts, concentrate or powder (Figure 4) [46,108]. In addition, grape seed extract, has been generally recognized as safe (GRAS) by the Food and Drug Administration (FDA) and is commercially sold as a dietary additive listed on the “Everything Added to Food in the United States (EAFUS)” to improve the overall quality of products and extend their shelf life [46,173].
Figure 4.
Applications of wineries’ by-products and their bioactives in the food industry, as ingredient(s) for the fortification/production of existing/novel functional food products, supplements and nutraceuticals with increased antioxidant protection - shelf life, and antithrombotic and anti-inflammatory health promoting properties against chronic disorders.
Figure 4.
Applications of wineries’ by-products and their bioactives in the food industry, as ingredient(s) for the fortification/production of existing/novel functional food products, supplements and nutraceuticals with increased antioxidant protection - shelf life, and antithrombotic and anti-inflammatory health promoting properties against chronic disorders.
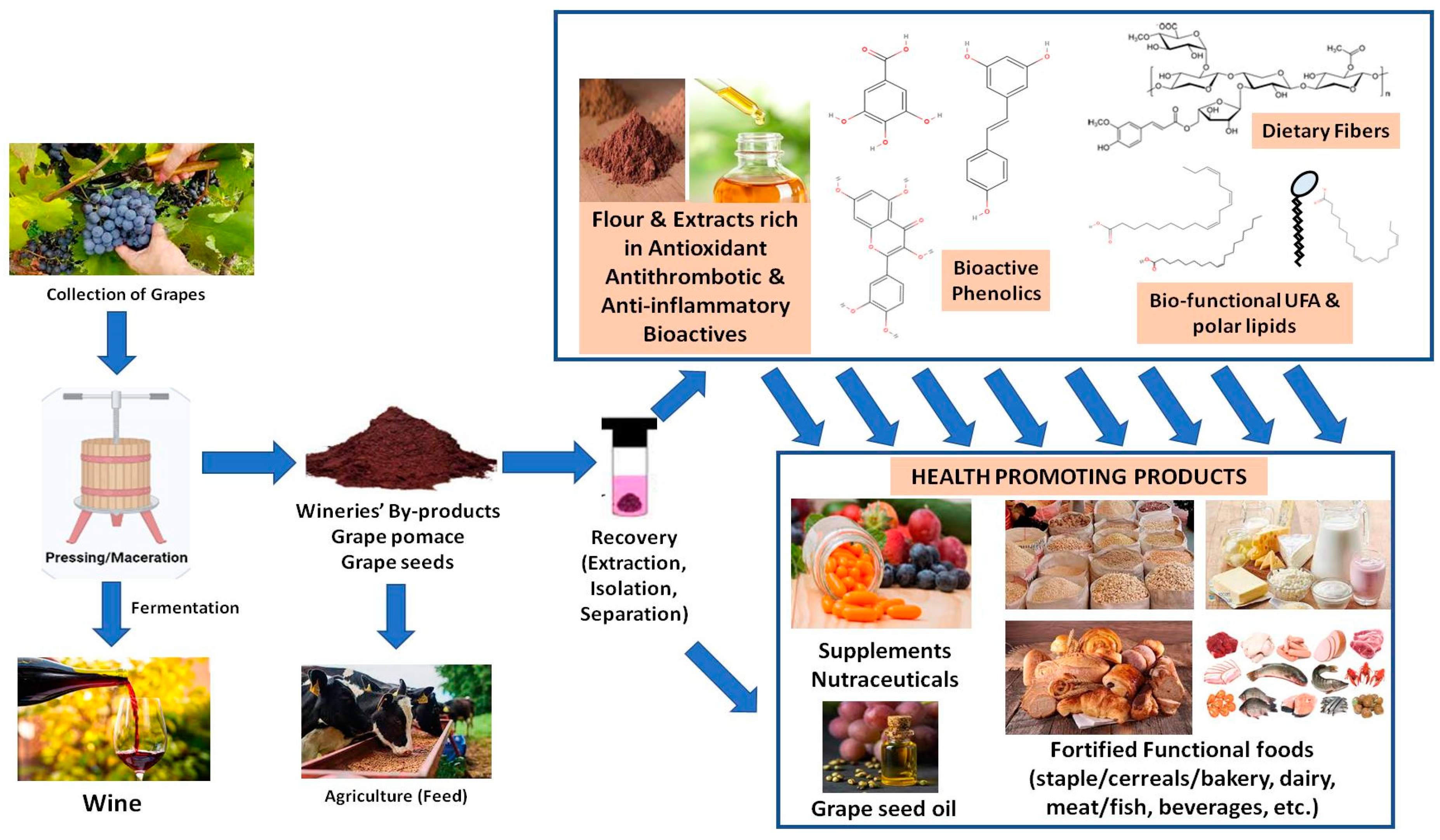
Nowadays, several commercialized cosmetic products, such as day or night cream, and face serum, mattifying fluid, anti-wrinkle and protect fluid, declare to use polyphenols from grape pomace and/or grape seed. In the food supplementation field there are few brands claiming to use grape polyphenols, mainly resveratrol. Such products confirm the commercial potential of bioactive compounds extracted from grape, or grape by-products. Nevertheless, despite the potential health benefits of resveratrol and other wineries’ by-products derived phenolics, their employment as nutraceutical ingredients within the food industry has specific limitations, due to their poor water-solubility, chemical instability and limited bioavailability. For these reasons, several delivery systems in the form of emulsions have been employed as promising encapsulation approaches, since encapsulation of such amphiphilic/lipophilic bioactive ingredients inside the hydrophobic core of the lipid droplets can protect them from degradation during storage and liberate them only after ingestion.
Moreover, discrepancies have been observed with extracts from wineries’ by-products against several disorders, mainly because the bioactives of the extracts have not been well quantified and characterized for each subclass of functional molecules that target specific molecular, cellular and biological processes. Moreover, some studies used enzymatic extraction while other used different solvents such methanol, ethanol or acetone, and the different method applied provided different content and bio-functional outcomes. For this reason, several investigations have focused on the appropriate extraction methods and recovery of wineries’ by-products bioactive compounds [108,176,177,178,179,180], as well as on their potential use as antioxidant and functional agents in the food, cosmetic, and pharmaceutical industries [22,33,70,108]. Recovery and clarification of wineries’ by-products bioactive compounds profile, their bioavailability and the action mechanisms for maintaining the redox cell balance and anti-inflammatory index involved in health benefits, is of great importance for their potential use as good and inexpensive functional ingredients of supplements and nutraceuticals for the prevention of inflammation-related chronic disorders [22].
4.1. Characteristic extraction methods for the recovery of wineries’ by-products bioactive compounds
The vast volumes of wine pomace produced during the winemaking process, as well as their potential market, has prompted food scientists to search for novel ways to use this by-product. However, in order to acquire suitable applications for the food sector, a number of aspects must be addressed, and especially the extraction method and its efficiency [108,176,177,178,179,180]. Pomace collection should also be preceded by preliminary care selection to determine the best pomace for the manufacturing of food-grade components based on functional constituent composition. In typical extraction processes, the medicinally active sections of plant or animal tissue are separated from the inactive or inert components using selected solvents [176]. Soxhlet extraction, maceration, and reflux extraction are some of the traditional procedures that have been employed for a long time [108,176]. As these procedures need a substantial volume of solvent, they are not suitable for commercial use. Furthermore, the high heat required to attain these methods leads to the loss of polyphenol and UFA products, as well as consume high amounts of energy. Traditional mechanical techniques damage cell walls and also release polyphenols, but they also speed up the release of enzymes like polyphenoloxidase and peroxidase, which might speed up polyphenol oxidation and destruction. Similar methodology is also used for the production of grape seed oil, but the use of cold temperatures and alternative pressing approaches may optimize the yield and quality [109]. Nevertheless, due to the drawbacks, non-conventional green and sustainable extraction methods had to be developed/used for extracting grape pomace bioactives in such a way so that they remain unaffected by the process, with highest possible yield and low energy cost [34,35,176].
For example, solid-liquid extraction through mechanical agitation and Soxhlet extraction with ethanolic or methanolic solutions as solvent are the most often utilized extraction methods for value-added by-products from grape pomace [108,178,179]. The extraction method is an extensively used unit operation to recover many important food components, including phenolics, lipids and dietary fibres [108]. Alterations on the extraction process of grape pomace under various different experimental conditions, such as solvent type (ethanol, methanol, water, etc.), temperature and time produce different yield of extracted compounds and antiradical activity of extracts, depending on the desirable extracted content of grape pomace. For example, a significant amount of anthocyanins or phenolics in general can be extracted by specific methods [108,180], while dietary fibres or grape seed oil will need similar to much different methodology for higher yield [108]. Especially for extraction of all phenolics, there are few available standardized procedures on solid-liquid extraction, but more understanding and systematic research for a high yield of unaffected extracted compounds is required [108,181].
Due to an increasing need for "natural" methods that do not introduce any leftover organic compounds, such as the use of nanofiltration membaranes or the supercritical fluid extraction, have emerged as appealing separation approaches for the extraction of valuable compounds from food wastes [34,176], including grape pomace, to be used in the food and pharmaceutical sectors [108]. The basic premise of supercritical fluid extraction involves a fluid at its supercritical state where discrete liquid and gas phases do not exist, which occurs at temperatures and pressures over its critical point [176]. Supercritical carbon dioxide's unusual solvent qualities as well as its cheap costs have made it a popular choice for extracting antioxidants, pigments and fatty acids from plant and animal sources, such as the grape seed oil that is rich in such lipid bioactives [182]. When modified with ethanol, it drastically increases polyphenols extraction while reducing extraction times, especially when compared to other methods such as the Soxhlet extraction or the solid-liquid extraction [183,184]. In another study, modification of ethanol (60%) extracted more polyphenols and anthocyanins from pomace, increasing the extract's anti-oxidant, anti-proliferation, and anti-inflammation activities. Furthermore, the functional activities of wine were increased when the crushed seeds in the fruit extract were included [185]. A similar assessment with supercritical fluid extraction compared the yield of resveratrol with carbon dioxide and the ethanol combination, with the latter extracting significantly more [186].
The extraction of bioactives from food by-products like grape pomace through microwave assisted extraction have also been thoroughly investigated [187,188], revealing its many advantages in natural compound valorizations. With the use of microwave techniques, polyphenol content extracted from grape pomace had a higher polyphenol content compared to traditional methods [189].
Ultrasound has been utilized more for the process of valuable compounds extraction from food by-products, including grape pomace, with high product yield whilst maintaining low solvent consumption and a shorter processing time [108]. Thus, when done with ultrasound assisted techniques, the extraction of oil from grape seeds was very efficient compared to Soxhlet methods, while using less time [190,191].
The recovery of valuable chemicals from food wastes and by-products like grape pomace using pulsed electric field treatment is a promising technology. Pulsed electric field processing is a non-thermal food processing technology that uses brief, high voltage pulses that results in cell disintegration and microbial inactivation [192]. The process of pulsed electric field is instantaneous, targeted and minimizes heat, the procedure is energy efficient whilst resulting in high-quality products [193]. Pulsed electric field allows for the recovery of polyphenols without addition of liquids, even in cases of fermented grape pomace, which has low relative humidity [194]. The shelf-life of the product is also significantly extended due to the inactivation of microorganisms from the electromechanical instability [195]. When pulsed electric field is used, total anthocyanin contents of up to 45% were extracted from grape by-products [196,197].
Overall, even though the traditional methods are still the most used for the extractions of compounds of interest from grape pomace, alternative technologies are emerging to recover bioactive compounds from this winemaking by-product, in order to promote faster and “greener” technologies [108,176,177,178,179,180].
4.2. Applications of wineries’ by-products and their bioactives in the food industry, as ingredient(s) for the fortification/production of existing/novel functional food products
The utilization of wineries’ by-products and their bioactive compounds as a source of functional ingredients is a promising field [22,33,42,46,70,108,110,111,172,182]. In this context, the recovery and valorization of bioactive compounds, dietary fibres and functional oil from wineries’ by-products as ingredients to fortify foods can still be an interesting alternative of environmental and economic approaches in the agro-food and pharmaceutical sector. Recently, the exploitation and application of wineries’ by-products as important candidates and sustainable sources of bioactive ingredients in the production of fortified foods has gained increasing scientific and industrial interest (Supplementary Table 3). The addition of wineries’ by-products bioactive compounds to a wide range of food products, such as plant origin food, meat, fish, and dairy products, could improve the nutritional composition of the final products and increase their nutritional values, mainly due to higher polyphenols and dietary fibers’ contents (Figure 4) [22,33,42,46,70,108,172,182]. Nevertheless, the change of color and undesirable changes in texture and sensory properties of the final products, caused by increased levels of polyphenolic compounds and/or dietary fibers, are the mainly observed adverse effects of the fortification of foods with wineries’ by-products [46]. Apart from these adverse effects, the most valuable influence of the wineries’ by-products bioactives addition is certainly a higher nutritional quality and oxidative stability of fortified products, reflected as higher polyphenol and total dietary fiber content [33,108]. Thus, wineries’ by-products bioactive compounds may undoubtedly be used as functional components in foods and beverages, providing health advantages in addition to nutritional value and other physiochemical applications, with a wide range of applications in both the food and pharmaceutical sectors [22,33,42,46,70,108,172,182]. Additional in vitro, in vivo and clinical studies are needed to examine the health efficacy of such products.
4.2.1. Applications of wineries’ by-products and their bioactive ingredients for the fortification/production of functional flour/cereals-based foods
The fortification of flour/cereals-based staple foods such as bread and pasta with grape pomace extracts/flour has been investigated in an effort to improve nutritional value of such foods. Grape pomace flour has been studied in different staple foods fortification, due to its high antioxidant capacity and delaying lipid oxidation properties [108]. Thus, research has also evaluated the use of grape pomace and seed flours in different flour/cereals-based pastry products and snacks too, such as popsicles, cereal bars, biscuits and cookies, extruded snacks, and muffin, obtaining products rich in fibre with antioxidant capacity potential and good acceptances by consumers [46,108]. With incorporation of grape pomace flour, it enhanced the antioxidant properties of different baked products, as well as increased total dietary fibre content [198]. The antioxidant capacity was also improved when products were enriched with phenols such as anthocyanins [199]. Nevertheless, biscuits were shown to be less brittle, reporting reduced hardness after incorporation of grape pomace phenols and dietary fibre [200], while in cookies consumer acceptance was highest at 5%, with further additions of grape pomace flour causing textural property changes [201].
Similar approaches have been done on bread through replacement or enrichment of wheat flour with grape pomace flour and extract. In these studies, the extract greatly increased the antioxidant activity of the final product despite some losses of phenolic compounds due to thermal treatment [202,203,204]. However, in some cases grape pomace powder negatively influenced the shelf-life of bread-based products like breadsticks, and thus showed an increase in oxidation rates [205], which was attributed to the grape pomace powder used, which measures at 9g/100g dry matter of fat with 50% being UFA, which are prone to oxidative stress [206]. Although the increase in phenolic compounds and nutrients is valuable, all the studies done on bread reported negative impacts on textural properties of the product, with reports of noticeable astringency and acidity that could affect consumer acceptance. Hardness and porosity of the bread has also been observed, likely due to the reduction of yeast activity, which lowered the gassing power of yeast, as phenolics can limit endogenous amylase activity in dough, resulting in insufficient maltose release for yeast activity during proving [207,208].
Fortification of pasta with winemaking by-products has shown promising results too. Substitution of flour or semolina with grape pomace powder at different rates has shown significant increase in dietary fibres. However, the studies showed different proportion of anthocyanins retained after the cooking process [209], with one showing no detection of anthocyanin after the cooking process, while other phenols and condensed tannin content in the pasta were not affected, which could be due to the highly bound phenol-fibre matrix [210]. The loss of phenolic content and antioxidant activity could also be attributed to the loss of the gluten protein network from grape pomace fibre interference [211]. Due to the same effect, cooking time was also shown to have reduced [212].
An easily overlooked aspect of wine pomace is its mineral composition, as it has been undervalued and there are no applications yet that focus on utilizing the mineral content, which is likely due to the intense focus on polyphenols and dietary fibre. However, incorporation of grape pomace flour has led to observations of improved mineral content that brings product stability. The calcium concentration obtained from grape seed flour aided in the stabilization of enzymes such as proteases and α-amylases, improving the quality of bread and cereal products [213,214].
Overall, there is a tendency that new applications of grape pomace in this area of functional flour/cereals-based products may arise in the coming years, especially on plan-based dietary patterns.
4.2.2. Applications of wineries’ by-products and their bioactive ingredients for the fortification/production of functional dairy-based foods
Improved physicochemical parameters, sensory properties and biological activities have also been proposed for the fortification of dairy products with grape pomace ingredients [215]. For example fortification of phenolics in dairy products, asides from the enhanced nutritional benefits, has also demonstrated increased stability in physicochemical properties, as well as anti-microbial and anticancer effects [215,216,217].
More specifically, the incorporation of phenolic compounds in yogurt showed beneficial modifications in its sensory properties and antioxidant capacity [216]. The incorporation of grape pomace powder into yogurt improved total phenolic content, while the total phenolic content was increased in yogurt too, as the grape pomace powder ratio increased [218]. Grape fibre can be used as an alternative source of antioxidants and dietary fibre when added to yogurt and salad dressings, not only for increasing the fibre and phenolic content, but also for delaying lipid oxidation during storage, extending the shelf life of these products ([219]. The addition of grape pomace powder to yogurt (1, 3 - 5%) significantly improved the yogurt color, even at the end of the storage period [218], as well as a greater perception of sweetness and grape aroma in probiotic goat milk yogurts caused, which assisted in facilitating the consumption of these products by consumers unfamiliar with goat milk products by decreasing the perception of acidity and goat milk aroma [220]. Nevertheless, when grape pomace powder was added into semi-hard and hard cheeses, asides from increased total phenolic content and radical scavenging activity, it produced a significant pH decrease due to the presence of organic acids from the grape pomace powder [221], which was also observed in grape pomace powder-fortified yogurt that showed a relatively higher acidity too [218]
Fortification of buffalo stirred-type yogurt with 0.5% of grape seed extract significantly increased the inhibition of free radicals, in comparison to control, indicating that the high phytochemical content found in grape seed extract was most likely responsible for increasing the antiradical ability of the grape seed extract-containing yogurt [222]. Fortified cheeses with grape pomace displayed increased antioxidant activity too without affecting starter development and acid generation that is crucial in the production [223]. Stirred-type yogurt supplemented with grape seed extract showed also antibacterial properties against several pathogens, such as E. coli and S. aureus [222]. This effect was significantly increased by increasing the concentration of grape seed extract in yogurt, suggesting that besides the endogenous presence of milk bioactive peptides, grape seed extract provided its antibacterial components in the yogurt product [222].
In fortification of both full-fat and non-fat yoghurts with grape pomace, the commercial shelf life was extended without affecting product stability or lactobacilli count [224]. The total phenol content, anti-oxidant activity and lactic acid bacteria trend were also retained throughout the shelf life [225,226]. It was also proposed that the production of functional yogurts supplemented with grape-seed antioxidants is feasible, given that the supplementation is carried out in the fermented product and not in milk prior to fermentation [224]. Moreover, yogurt containing 0.5% grape seed extract significantly increased cytotoxicity activity, achieving 62.47 and 70.36% cell death against MCF-7 and HCT-116 cancer cell lines, respectively, with the phenolic substances found in fortified yogurt derived from grape seed extract playing an important role in the observed increased anticancer activity, in comparison to the anticancer activity of 0% grape seed extract yogurt [222].
4.2.3. Applications of wineries’ by-products and their bioactive ingredients for the fortification/production of functional meat-/fish-based foods
Meat, fish and their products are the most prominent dietary categories in which wineries’ by-products have been used to prevent mostly lipid oxidation and thus prolong storage or shelf life [33,46,108]. Several studies have used grape pomace extracts as food protectors due to their antibacterial capacity against different bacterial spectrum, while seedless grape pomace products showed also bactericidal effects against total aerobic mesophilic bacteria, lactic-acid and Enterobacteriacea [108]. Thus, research on the influence of grape pomace fortification in meat and fish products were primarily conducted to determine the effects on shelf life and storage stability as well as anti-oxidative capacities [46]. These fortifications were done largely through addition of grape pomace powder or extract, or marination in grape pomace solutions.
Grape pomace dietary fibres and the rich in UFAs grape seed oil has also been utilized in meat products [108]. For example, the addition of grape seed extract to Western-style smoked sausage improved its color and extended its shelf life due to the strong antioxidant properties of grape seed extract [46,227]. In particular, grape seed extract could significantly substitute and thus decrease residual nitrite, which may inhibit the formation of harmful and carcinogenic substances, such as N-nitrosamine [46,227]. Such modifications in the formulation of meat products have been made in order to attend an increased interest from consumer for healthier and functional foods, such as the reduction of some compounds such as fat, saturated fatty acids, and the addition of ingredients such as fibre, UFA and antioxidants into products, which can be achieved by the addition of grape pomace, grape seed oil or their functional ingredients [108].
For example, there is an increasing consumer demand to limit the utilization of nitrite and other preservatives and synthetic antioxidants in processed meat products due to associated carcinogenic activities, therefore incentivizing researchers to use natural antioxidants as a substitute in processed meats to achieve similar shelf stability and antimicrobial qualities. In several studies incorporating grape pomace into preserved meat products, the antioxidant capacity scaled accordingly to different concentrations, with most inhibiting lipid oxidation [228,229,230]. A low microbial count was also achieved [231], likely due to the bacteriostatic or bactericidal effects exhibited at low or high phenolic concentrations respectively. This result showed similar effects to the usage of nitrite in preserved meats, suggesting that grape pomace could be used as an alternative additive for preservation.
Phenolic extracts from grape pomace have also effectively been used in meat products as preservatives - natural antioxidant replacers to synthetic antioxidants added usually for the preservation of meat, such as the use of red grape pomace in pork burgers [232]. In the addition of grape seed extract into raw pork at 1% w/w, oxidative stability was achieved without negatively affecting sensory qualities of the meat [233]. Grape seed extract also effectively reduced lipid oxidation and improved the shelf life of storage lamb meat, equally to similar effects observed by the addition of vitamin E [234]. Grape pomace extract has also been effectively used to improve shelf life of both raw and cooked chicken meat [235,236], in which such a fortification resulted in strong antioxidant activity during storage time, comparable to the common antioxidant food additive, butyl hydroxy toluene (BHT). However, negative changes in sensory properties were observed following such a fortification in chicken meat.
Grape antioxidant dietary fibre added on chicken breast hamburger and on fish muscle showed an improvement on the oxidative stability and on the radical scavenging activity of both products [237,238]. When grape seed flour was used at different concentrations into sausages, similar decreased levels of oxidation can be observed, followed by enhanced protein and increased total dietary fibre [239]. However, water holding capacity was also increased likely due to the increase in fibre-matrix complexes, which could increase the water activity of the product, and thereby increase susceptibility to microbial growth. Of these studies, most saw alterations in color with other sensory properties such as texture and taste unaffected.
Pre-emulsified grape seed oil has also been used to replace pork back fat in frankfurter sausages resulting in beneficially affected fatty acid composition and overall reduction of the fat content, which was then closer to the target value of 20% (fat content), while these reduced fat frankfurters had consumers acceptability similar to regular fat frankfurters, as there was also an increase in gumminess and chewiness, improving textural properties [240]. Similar results were obtained when different concentrations of grape seed oil replaced animal fat in frankfurters sausages and even though the decrease in the lipid oxidation was not significant, however it was well accepted by panelists [239].
Whole grape pomace was used in minced fish frozen successfully to lipid oxidation prevention [108], as it was also shown by the fortification of fish products with grape pomace at concentrations of 1% [239] and 2% [242]. Other studies used also grape pomace extracts as food protectors due to their antioxidant ability to prevent lipid oxidation in fish-based products [243]. Similar observations were also observed regarding oxidation inhibition and storage stability, followed by increase in dietary fibres after the addition of grape pomace flour into salmon patties [242]. In the addition of grape seed extract into minced fish muscle, the extract demonstrated inhibition of lipid oxidation throughout cold storage too, but as with the meat products, a deeper color of the finished product was noticed in the enriched products, leading to lower consumer acceptance. The application of fibre from grape pomace has also been performed in fish products, such as codfish and seafood, where the addition of these compounds has been shown to be a promising tool in minimizing flavor changes, color, texture and lipid oxidation during freezing storage [238,244,245].
In the application of wine pomace in preserved meat products, the mineral content allowed the product to have a lower level of sodium, higher levels of calcium and potassium, whilst maintaining product shelf stability [228], which may help to lower blood pressure, minimize the risk of stroke, and prevent CVD, particularly in hypertensive people. Finally, the use of grape pomace flour for coating has also improved the nutritional value of meat or fish. For example, daily consumption of beef burgers prepared with wine grape pomace flour for one month in 27 male volunteers with metabolic syndrome, improved their blood biochemical parameters, such as fasting glucose and insulin resistance, including plasma antioxidant levels and oxidative stress biomarkers, in comparison to the other months of the intervention, when they consumed daily either no burgers (in the second month), and one control-burger (in the third month) [246].
Therefore, it seems that such grape pomace and grape seed oil functional ingredients have the potential as a dietary supplement and fortifying agents of meat and fish foods to manage chronic disease risk in humans, without affecting the overall nutritional and sensory properties of the fortified foods.
4.2.4. Applications of wineries’ by-products and their bioactive ingredients for the fortification/production of other plant-based functional foods and beverages
Wineries’ by-products and their bioactive ingredients has also been effectively used for the fortification/production of other plant-based functional foods and beverages. For example, supplementation with grape skin powder of tomato puree with varied particle sizes at a concentration of 3.2%, even though no significant impact on polyphenol concentrations was observed, still the antioxidant potential of the fortified tomato puree was greater due to the introduction of grape pomace anthocyanins, while the reduced particle size of grape skin powder received higher results in sensory studies too [247]. Moreover, when lycopene was used together with anthocyanins in vitro, the combination seems to work synergistically, demonstrating not only antioxidant improvement, but also strong anti-inflammatory potential through a significant increase in inhibition of cytokine IL-8 [248]. In a similar evaluation, walnut paste was encapsulated and emulsified with grape skin extract to evaluate the oxidative stability and phenolic retention. However, results showed that though the antioxidant capacity was incorporated into the walnut paste, further thermal stress did not bring significant benefits, and most antioxidant properties were traced back to tocopherols that were originally present in the walnuts [249]. Thereby, the combined synergistic abilities for different antioxidant bioactives requires further investigation.
Asides from antioxidant capacity, the possibility of inhibiting acrylamide formation through grape pomace addition was also studied. Acrylamide is a molecule produced during the formation of browning in Maillard reactions that is found to exhibit carcinogenicity and neurotoxicity numerous human and animal studies [250]. Through incorporation of grape pomace extract into potato chips, a significant reduction of 90% less production of acrylamide was observed in the final product [251]. This was reflected in another study, observing the decrease in acrylamides through not only the free radical scavenging effects, but also reaction with carbonyl species [252].
Apart from the traditional application of grape pomace in the food industry as a substrate for the production of some spirits by distillation [46], grape pomace and its extracts can also be used as functional supplement in food production, to enrich beverages, or even as ingredient of osmotic solution to obtain dehydrated fruit incremented in phenolic content [253]. Due to their ability to absorb tannin, a study suggested that grape fibre addition in red wine production would be able to remove up to 38% of tannins generated during the process [254]. Grape ingredients have also been successfully used to address the unpleasant taste and flavor faced by produced fermented products from plant-based dairy alternative drinks. Thus, supplementing such plant based yogurt-like fermented products with some natural functional ingredients, including grape bioactives, can overcome the alleged unpleasant or poor taste in this kind of plant-based products [215].
4.3. Health benefits and applications of wineries’ by-products and their bioactives, as ingredients of bio-functional food products, supplements and nutraceuticals
The application of wineries’ byproducts, such as grape pomace and grape seed, has shown promising outcomes, with progress reported in their valorization and/or the incorporation of their bioactive ingredients in producing bio-functional foods, supplements and neutraceuticals with several promising observed benefits and health promoting effects (Figure 4) [22,33,42,46,70,108,110,111,172,182,255]. Each of the bioactive ingredients of these by-products, such as their phenolics, UFA, polar lipids and dietary fibers, or even several forms of the by-products themselves or extracts of these by-products containing several of these bioactives, which usually act synergistically, have been assessed for health benefits by either improving the fortified bio-functional foods or in the form of supplements-nutraceuticals. For example, several products derived from these by-products have been developed, such as supplements containing grape pomace extracts or grape seed extract capsule, while grape seed oil on its own has been characterized and sold as a supplement product ((Figure 4) [11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,182,255].
The most characteristic bioactive ingredient(s) of the wineries’ by-products used in such applications are several of their extracts rich in phenolics or the recovered and isolated from them specific phenolic ingredients with specific metabolic and bio-functional properties and health promoting effects [16,22,33,38,39,40,41,42,46,47,70,108,110,172,174,176,182]. Apart from vitamins and minerals, phenolic compounds may be one of the most popular dietary supplement categories. Wineries’ by-products like grape pomace and grape seeds seem to be sustainable sources of phenolic bioactives like resveratrol for developing bio-functional food products and promoting health supplements and neutraceuticals, while such by-products contain also other bioactive ingredients like UFAs, polar lipid bioactives and dietary fibres, which have also shown several health benefits (Figure 4) [10,16,17,21,22,33,38,39,40,41,42,46,47,70,108,110,111,172,173,174].
However, it is still necessary to implement more clinical trials to be able to draw safe conclusions on the possible use of winery by-products and of their bioactives as single isolated compounds with a known concentration or as a validated standardized mixture of these compounds, with increased bioavailability, to be used for the production of protective and/or therapeutic bio-functional products, supplements and nutraceuticals, with antioxidant, antithrombotic and anti-inflammatory health promoting effects.
4.3.1. Anti-oxidant, anti-inflammatory and antithrombotic health promoting effects of grape pomace and of its bioactives, extracts and relevant bio-functional products
Grape pomace is an important source of phenolic compounds, and many effects on health such as free radicals scavenging activity, anti-platelet and anti-inflammatory properties, anticancer, and cardioprotective activity have been reported [22,33,42,46,70,108,172,174,176]. Flavonoids, stilbenes, lignans, and phenolic acids, classes of polyphenols found in grape pomace, have been investigated as an important alternative source for active substances that could be used in the management of oxidative stress, platelet activation, inflammation and of many other pathological manifestations [22,38,39,40,41,42,46,47,68,70,108,172,174,176,256,257].
A large variety of in vitro and in vivo studies has exhibited the beneficial antioxidant and anti-inflammatory actions of grape pomace and its bioactive ingredients. Several in vitro works have presented the antioxidant effects of grape pomace and its bioactive ingredients, such as the observed decrease in ROS, MDA, and TBARS levels, as well as an increase in GSH levels, while the observed in vivo antioxidant benefits usually include the increase in CAT, SOD and GPx4 levels, a stimulation of endothelial eNOS gene expression, as well as a beneficial modification of uric acid (UA), protein carbonyls (PC) and TBARS levels [21,38,39,40,41,42,108,172,257,258,259].
The in vitro anti-inflammatory actions of grape pomace and its bioactive ingredients, and especially those of the grape pomace phenolics, usually include the inhibition of NFκβ and PGE2 associated inflammatory pathways, as well as the decrease of some inflammatory markers such as IL-8 [42,108]. In addition, the inhibition of platelet activation and aggregation induced by several inflammatory and thrombotic mediators like PAF, thrombin, collagen and ADP, by grape pomace and/or grape seed extracts rich in bioactive phenolics, UFAs and polar lipids further support their protective effects against several thrombotic and -inflammatory manifestations and associated disorders [6,7,10,16,17,22,42,70,74,90,91,92,108,260,261,262,263,264,265]. The in vivo anti-inflammatory properties of grape pomace and its bioactive ingredients include also the inhibition of the release of several inflammatory markers such as IL-1𝛼, IL-1β, IL-6, IFN-𝛾, TNF-α, and CRP [42], as well as the protection against thrombo-inflammatory activation of platelets [15].
For example, grape pomace extract rich in phenolic compounds like quercetin, catechin, epicatechin and gallic acid, have shown better inhibition of platelet aggregation induced by ADP, compared with wine’s effect on platelets, nevertheless such phenomena could be explained due to the higher concentration of phenolic compounds present in the pomace extract [261]. In addition, extracts of both red grape pomace and white grape pomace have also been found to inhibit platelet aggregation activity using ADP as agonist [265]. Strong anti-platelet effects of red grape pomace extracts, not only against ADP, but also against the platelet aggregatory thrombo-inflammatory signaling of PAF and thrombin has also been found [16,17]. Such extracts were not also rich in bioactive phenolics, but they were also found to contain considerable amounts of bio-functional polar lipids and UFA, which explain their strong anti-inflammatory and antithrombotic potency [16]. In a randomized trial in healthy individuals that was based on the consumption of a supplement containing several plant extracts, including grape pomace extract, showed a strong protective effect against PAF-associated thrombo-inflammatory signaling, by reducing both PAF-activities (reduced PAF-induced platelet aggregation) and PAF-levels (increased PAF-catabolism) [15].
Several human studies have shown that consumption of grape pomace products significantly reduce risks of atherosclerosis, hypertension and diabetes related conditions [70,108]. For example, grape pomace extract presented protective vascular and antioxidant properties [258], as such extracts induced a relaxation or inhibition in the contraction of aortic rings in a dose-dependent manner, mainly due to antioxidant activity of the phenolic compounds present in the extract and the induced activation of eNOS through a NO-dependent mechanism. Supplementation with red grape pomace flour in atherogenic diet-fed SR-B1 KO/ApoER61h/h mice, a model of lethal ischemic heart disease, increased plasma antioxidant activity, changed TNF-α and IL-10 levels, decreased atheromatous aortic and brachiocephalic plaque sizes, attenuated myocardial infarction and dysfunction and thus reduced premature death [266].
Grape pomace extracts and products have also exhibited antioxidant and anti-postprandial hyperglycemic protective activities in vitro and in vivo, suggesting its use as a functional ingredient against these manifestations [21,267]. For example, intervention study showed that as a dietary supplement, consumption of wine grape pomace flour improved the blood pressure of patients, as well as enhanced insulin sensitivity and antioxidant protection [268]. More specifically, the consumption of grape pomace flour rich in fiber and polyphenol antioxidants, , in males with at least one component of metabolic syndrome, as a food supplement in a regular diet or as an ingredient of functional foods fortified with this winery by-product, improved fasting glucose levels, glycaemia and postprandial insulin, systolic and diastolic blood pressure and plasma antioxidant levels (i.e. it increased the levels of plasma γ-tocopherol and δ-tocopherol, which provide antioxidant protection and decreased oxidative protein damage), indicating attenuation of oxidative stress and protection against metabolic syndrome in humans [246,268].
In another double-blind placebo controlled study, blood pressure was significantly lowered when polyphenol rich grape extracts were administered [269]. A double-blind randomized placebo-controlled trial showed also that foods fortified with wine pomace, or with its bioactive flavanols, can attenuate hyperglycaemia-induced endothelial dysfunction and oxidative damage in endothelial cells and thus can significantly improve vascular function [270,271]. The antioxidant activity demonstrated in the various applications of wine grape pomace and fortification into different functional foods seems to be beneficial, as it was also shown in another trial in healthy normal and overweight/obese women based on the consumption of a grape pomace extract, which resulted in postprandial metabolic benefits and protection against oxidative stress responses [21]. Moreover, treatment of intestinal cells with grape pomace extract provided anti-oxidant protection, as it neutralized the production of reactive oxygen induced by tert-butyl hydroperoxide [272].
The presence of high concentrations of grape pomace phenolics (polyphenols, tannins, flavonoids and anthocyanins) can also provide plasma lipid lowering protective effects, since such a polyphenolic grape pomace extract reduced cholesterol levels by lowering the enzymatic activities of enzymes involved in intracellular cholesterol production [38]. Moreover, in a triple-blind, randomized controlled trial, grape extract consumption reduced atherogenic markers and exhibited additional cardioprotective effects, including decreased LDL and apolipoprotein-B [273].
Even though glucose, total cholesterol, HDL-C and LDL-C levels were not affected in vivo in male Wistar rats treated with a rich in phenolics grape pomace extract (malvidin, quercetin, gallic acid, and a procyanidin dimer type B were the main compounds identified), still other biomarkers of CVD, such as VLDL-C and triacylglycerols levels, were decreased with such a grape pomace extract treatment [274]. On the other hand, in another randomized crossover clinical trial based on consumption of a grape seed extract by hyperlipidemia patients (21–64 years old) a decrease in total cholesterol, LDL-C and the Ox-LDL-C levels was observed, while triacyclglycerols, HDL-C and VLDL-C levels were not modified [275]. The observed differences between these two studies may be explained either by the different winery by-product source used for obtaining the rich in phenolics extracts, or by the organisms of the trials and/or by the amount supplemented, because higher dosages were given to the rats compared with those given to the hyperlipidemia patients.
Most of the studies have presented the effects of specific concentrations of single grape pomace polyphenols, such as resveratrol, quercetin, and gallic acid, against oxidative and inflammatory damage, but recent experimental data have shown that grape pomace extracts are more effective than a single bioactive ingredient, possibly because of their synergistic action that interferes with more than one pathophysiological mechanism [15,16,17,21,22,42,46,70,108,172]. All these studies have presented that grape pomace as a whole extract, but also different individual bioactive ingredients contained in grape pomace can modulate the endogenous pathway responsible in reducing oxidative stress and chronic inflammation [22], indicating that they may be successfully utilized as valuable therapeutic candidates capable to reduce the thrombo-inflammatory and oxidative stress associated pathological processes. Although further research is needed, the bioactive molecules from wineries by-products show promise for use as antioxidant supplements [21,38,39,40,41,42,108,172,257,258,259]. The ethanolic extracts of grape pomace have been proposed as candidates for food supplements and nutraceuticals with antioxidant protection and anti-platelet benefits as well [15,21].
4.3.2. Anti-oxidant, anti-inflammatory and antithrombotic health promoting effects of grape seeds and of their bioactives, extracts and relevant bio-functional products
Considerable interest has also been given on the use of grape seed and its extracts as raw material to develop products with nutritional value and health benefits [46,108,173,262,263,264,275,276,277,278,279,280,281,282,283], with considerable popularization and application prospects [46,108,173]. For example, grape seed extract capsules have been developed and sold by some companies as dietary nutritional supplement products to protect humans from oxidative damage and to maintain health. Moreover, due to its solubility in water and ethanol, grape seed extract has considerable potential as a beverage added to produce products with health benefits that could satisfy the taste standards of the public [46,276,277].
Grape seed products and extracts have shown potent antioxidant and anti-inflammatory vascular protection, lipid lowering effects and subsequent cardiovascular benefits [46,108,173,262,263,264,275,276,277,278,279,280,281,282,283]. For example, administration of grape seed extract to hamsters for 12 weeks resulted in a substantial reduction of plasma cholesterol [256]. Moreover, grape seeds hold a large amount of proanthocyanidins, also known as condensed tannins [277], while the pharmacokinetics and biological metabolism of these grape seed bioactives have shown that they possess significant antioxidant and anti-inflammatory capacities, demonstrated by the suppression of IL-3 activity, inhibition of inflammatory mediators and lipoxygenase, as well as reduced LDL oxidation [283,284,285]. In in vivo studies, grape seed powder and proanthocyanidin consumption is shown to exert anti-oxidative, anti-inflammatory and cardioprotective effects [282,285].
Grape seed extracts have also shown potent antiplatelet cardioprotective effects [70,108,262,263,264]. For example, grape seed extracts (containing a mixture of polyphenols including gallic acid, falvan-3-ols, and proanthocyanins) inhibited more potently than resveratrol the platelet activation induced by proteolytic (thrombin-induced) and non-proteolytic (thrombin receptor activation peptide – induced) signaling, while the formation of platelet microparticle ((PMP) generation was also reduced [262]. It seems that the presence of a variety of phenolic bioactives in grape seed extracts provides the ability of this wineries’ by-product to interact with several pathways in platelet activation, leading to decreased degranulation and PMP shedding, thus contributing to its higher inhibition than pure resveratrol. The specific method by which such phenolic compounds limit platelet activity has yet to be determined, however it appears that these phytochemicals alter the fluidity of membranes, the affinity of the ligand-receptor, and the signaling pathways inside cells. These effects might be mediated by antioxidant processes, as well as the control of pathways linked to nitric oxide generation and release, which alleviate the oxidative stress-induced platelet activation and PMP shedding by scavenging free radicals. Similarly, flavan-3-ol-rich extracts from grape seed were shown to have a platelet-inhibiting effect under flow in response to collagen/ADP [263]. Interestingly, grape seed and grape skin extracts elicit a greater antiplatelet effect against on collagen-induced platelet aggregation when used in combination than when used individually in both dogs and humans [264].
As aforementioned, wineries’ by-products, their products and extracts, as well as several of their fortified functional foods, have also exhibited potent anti-diabetic benefits [108]. For example, the incorporation of extracts from grape pomace, grape seed or grape skin, in yogurt resulted in a significant increase in total phenolic content and antioxidant and antidiabetic properties, compared to the controls, with the functional yogurt fortified with grape skin extracts showing the highest phenolic content and antioxidant capacity, as well as the highest inhibition of the activity of α-glucosidase [286], an enzyme that is associated with the digestion of carbohydrates as well as with the elevation of postprandial glucose levels in diabetic patients . Furthermore, a study also shows that resveratrol-amplified grape skin extracts to inhibit fat accumulation and fatty acid synthesis, as well as modulate insulin sensitivity, all of which lead to risk reduction in obesity rates [287]. With respect to grape seed, an extract of this winery by-product has in vitro inhibited the intestinal α-glucosidases and α-pancreatic amylase that may delay carbohydrate digestion and absorption, resulting in the suppression of postprandial glycemia, while this extract has reduced in vivo postprandial plasma glucose in healthy participants receiving a high carbohydrate meal, in an acute, randomized, controlled crossover study-design [278]. Moreover, oligomers of grape-seed procyanidin extract were able to enhance insulin-mediated glucose uptake by their interaction on the GLUT4-mediated glucose-uptake process [279], which further support the anti-diabetic protective properties of grape seed bioactive phenolics.
Several studies have also shown the potential use of grape seed extracts rich in bioactive phenolics, as a therapeutic against several neurodegenerative disorders. For example, a commercially available grape seed polyphenolic extract significantly attenuated Alzheimer’s diseases type cognitive deterioration and reduced cerebral amyloid deposition [281].
Grape seed is also a valuable source of bio-functional oils [46,108,110,111,182,255]. Grape seed oil also contains phenolics, but it is also rich in UFAs (especially linoleic acid) and lipid-soluble vitamins (such as vitamins A, D and E), conferring grape seeds oil with various health-promoting activities [18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,182,255]. Recently, some lipid-embedded substances in grape seed oil, such as tocopherols, phytosterols and phenols, have attracted increasing attention from researchers because of their diverse biological activities, such as antioxidant and anti-inflammatory activities [108,110,111,182]. Recent reported data has proved beneficial effects of grape seed oil, such as hepatoprotective, neuroprotective and reducing liver cholesterol [108,288,289,290]. The potent antioxidant, anti-inflammatory, antiapoptotic and lipid lowering activities of grape seed oil exhibited a protective effect on acute liver injury, while similar protective mechanisms of grape seed oil against oxidative damage and inflammatory cascades in the brain, such as its ability to scavenge free radicals, improve the activity of antioxidant enzymes, down-regulate the gene expression levels of xanthine oxidase and inducible nitric oxide synthase and suppress the inflammatory responses, seem to also be responsible for its neuroprotective effects too [288,289,290].
In this respect, the recovery of oil from grape seeds is of considerable significance with respect to the exploitation and utilization of grape seed oil as a functional food and supplement on its own [108,110,111]. Some grape seed oil products have been sold by companies and are consumed as part of the daily diet.
4.3.3. Anticancer protective effects of wineries’ by-products and of their bioactives, extracts and relevant bio-functional products
Extracts and products derived from winery by-products, containing several of their phenolic bioactive compounds, and especially anthocyanins and/or resveratrol, have also exhibited anti-tumor protective properties against several types of cancers [22,33,46,58,65,66,67,72,81,82,83,84,85,86,87,108,172,280,291,292,293,294,295,296,297,298]. For example, grape seed extracts have exhibited anticancer actions against different cancer types (skin, prostate, breast) [280]. In addition, a daily dose of 80 g resveratrol-containing freeze-dried grape powder for 14 days in 8 colon cancer patients resulted in a significant inhibition of the Wnt signaling pathway, which is one of the key cascades regulating development and stemness, and has also been tightly associated with cancer [291]. The chemopreventive mechanisms of resveratrol containing extracts of winery by-products has also been attributed to resveratrol’s capacity to control inflammation, remove reactive oxygen species and deactivate pro-carcinogens [6,81,82,83,84,85,86,87,291].
Anthocyanins similarly exhibit anti-carcinogenic activity against cancer cells in vitro and in vivo [293]. In vitro, the chemopreventive properties of anthocyanins demonstrated include radical scavenging actions, decreased cell proliferation, decreased inflammation as well as induction of apoptosis, while, dietary anthocyanins have also inhibited in vivo gastrointestinal cancers. More specifically, anthocyanin-rich extracts from grapes, and several pure anthocyanins and anthocyanidins, seem to exhibit their pro-apoptotic effects in multiple cancer cell types in vitro, through both intrinsic (mitochondrial) and extrinsic (FAS type I transmembrane protein of the TNFα superfamily) pathways. In the intrinsic pathway, anthocyanin treatment of cancer cells results in an increase in mitochondrial membrane potential, cytochrome c release and modulation of caspase-dependent anti- and pro-apoptotic proteins. In the extrinsic pathway, anthocyanins modulate the expression of FAS and FAS-ligand pathway in cancer cells resulting in apoptosis. In addition, treatment of cancer cells, but not normal cells, with anthocyanins leads to an accumulation of ROS and subsequent apoptosis, suggesting that the ROS-mediated mitochondrial caspase-independent pathway is important for anthocyanin-induced apoptosis too.
For example, administration of an anthocyanin-rich extract from bilberry, chokeberry and grape (containing 3.85 g anthocyanins per kg diet) in rats with colon cancer induced by azoxumethane, significantly reduced the formation of the azoxymethane (AOM) induced colonic aberrant crypt foci (the earliest identifiable intermediate precancerous lesions during colon carcinogenesis). This reduction was associated with decreased cell proliferation and COX-2 gene expression, however, the levels of urinary 8-OHdG were similar among rats fed the different diets [294]. Administration of grape pomace in female mice, with AOM/dextran sulfate sodium (DSS) induced colorectar cancer, showed protective effects against colitis-associated carcinogenesis, since such an administration ameliorated the disease activity index (DAI) score, reduced tumor number, tumor size and pathological scores, as well as it suppressed cell proliferation and inflammation (suppression of colonic expression of inflammatory cytokines, IL-1β and TNF-α, inhibition of NF-κB inflammatory signaling, increase of anti-inflammatory cytokine TGF-β mRNA expression) and it alleviated DNA methylation of the promoter region of Cdx2 gene and hypermethylation of CpG island methylator phenotype (CIMP), which commonly occurs during CRC carcinogenesis [295].
Recently, two phenolic-rich extracts from biotransformed grape pomace, which was derived by a 15-day solid-state fermentation with the white-rot fungi Phanerochaete chrysosporium and Trametes gibbosa, respectively, have exhibited potent antioxidant and antiproliferative potentials against colorectal cancer in vitro, in both Caco-2 and SW620 colorectal cancer cell lines [296]. In addition, an anthocyanin-rich grape extract and a component grape anthocyanin have shown breast cancer chemopreventive potential due in part to their capacity to block carcinogen-DNA adduct formation, modulate activities of carcinogen-metabolizing enzymes, and suppress ROS in these noncancerous human breast cells [297]. Fractions of a grape pomace derived from the muscadine 'Noble' variety, which contained a mixture of anthocyanidins and ellagic acids, exhibited the strongest anti-oxidative activity, as determined at both low and high concentrations in the 2,2-diphenyl-1-picrylhydrazyl (DPPH) and ferric-reducing antioxidant power (FRAP) assays, while they were also able to potently induce cell cycle arrest and apoptosis in MDA-MB-231 breast cancer cells, via caspase activation and by downregulating cyclin A and upregulating p21 [298].
4.3.4. Antimicrobial protective effects of wineries’ by-products and of their bioactives, extracts and relevant bio-functional products
Natural products as sources of potential antimicrobials, and especially from food wastes like the rich in phenolics wineries’ by-products (pomace, skins, stalks, leaves, and especially seeds), seem to provide an alternative sustainable way to counteract bacterial infections. Some phenolics like resveratrol are bacteriostatic/bactericidal against several pathogenic bacteria and may have a synergistic action towards antibiotics, mitigating or reverting bacterial resistance to these drugs. Complex phenolic mixtures, such as those present in these winemaking residues, are even more effective as chemotherapeutic antibacterial agents and could be used in combined therapy, thereby contributing to management of the antibiotic resistance crisis [299], while they can also be successfully used to replace the requirement for artificial preservatives.
For example, grape seed extracts were efficient in their antibacterial activities after inoculation into cheese and demonstrated bactericidal effects against Escherichia coli [300]. The antibacterial impact was shown at greater concentrations than in vitro experiments, suggesting that the antimicrobial effect was reduced when the items were introduced to meals. The reduced impact is most likely due to polyphenols' limited solubility in specific meals and their interaction with other dietary components [301]. In a study, resveratrol showed significant antibacterial activity against gram-positive bacteria through disruption of phospholipid bilayer and causing significant cell membrane damage when tested against Listeria monocytogenes [302]. This observation was reflected similarly in other studies against gram-positive bacteria [299,303].
4.3.5. Bio-delivery systems to improve the bioavailability and bio-functionality of wineries’ by-products and of their bioactives, extracts and relevant bio-functional products
Delivery of phenolic bioactives of winery by-products and of their bioactives into the body system is limited due to their poor water solubility, bioavailability and chemical stability [303,304]. With the use of different encapsulation as an approach, the delivery for such bioactives may be achieved. Emulsion-based delivery methods, in particular, are a potential encapsulation method. Lipophilic bioactive substances can be contained inside the hydrophobic core of lipid droplets, preserving them from destruction while allowing them to be released after consumption [305]. The value of an encapsulation technique for stabilizing bioactive chemicals derived from winemaking by-product was also shown in a study, where crude extract polyphenols were observed to degrade faster than those encapsulated [304].
For example, formulation of cosmetic emulsions with grape seed oil and diluted wine in the aqueous phase presented advantages such as direct inclusion of natural antioxidants, aroma and color compounds that enhanced their organoleptic characteristics [306]. In another cosmetic application of grape pomace, inactivation of proteolytic enzymes related to skin aging, such as collagenase and elastase, was observed, due to the higher availability of hydrophilic polyphenols, such as low molecular weight phenolic acids, especially gallic acid [307]. An antioxidant therapeutic nanoplatform consisting of nano-sized functionalized liposomes loaded with selected polyphenol-rich extracts from grape seeds and skins, with high blood-brain barrier crossing capabilities, successfully reduced oxidative stress (decreased ROS levels), prevented the aggregation of α-synuclein fibrils, and restored cell viability, in a relevant in vitro model of Parkinson's disease [308].
In another nanoemulsion, grape seed oil and grape skin extract were combined to encapsulate resveratrol, and thus created a stable delivery system for resveratrol, with minimal damage against UV-light isomerization and degradation, which reduced oxidation damage [309]. In another study, the synergistic effect between a sunscreen system containing UV filters and grape pomace extract on improving the antioxidant activity and UVB protection of this winery by-product, has also been observed [310]. More specifically, both samples (control formulation containing UV filters and sample formulation containing UV filters + grape pomace extract at 10.0% w/w) were considered safe, while sample formulation containing UV filters + grape pomace extract was more efficient in protecting skin against UVB radiation, taking approximately more time for UVB to induce erythema compared to the extract-free control
Dietary fibres of wineries’ by-products seem to be another popular grape-derived product with significant antioxidant effects, which can also be used as a natural system that increases the bio-delivery of wineries’ by-products bioactives [108,175,311,312]. Wineries’ by-products dietary fibre antioxidant activity may differ significantly depending on the extraction method used [108,175,313,314,315,316].
For example, the overall antioxidant activity of generated fibres was increased through a method of hot aqueous extraction of white grape pomace followed by membrane concentration [313]. Fibre components in grape pomace have shown to form chemical bonds with phenolic substances through a complex matrix, thus increasing the phenols’ ability to scavenge free radicals as they form an antioxidant dietary fibre compound, and thus enhanced the nutraceutical properties of the fortified bio-functional food product [200,311,312]. Furthermore, enhanced bio-functionality, nutritional value and storability of yogurt and salad dressing were observed when these food products were fortified with grape pomace fibres [219], while when grape pomace was used as a dietary fibre supplement for the use of prebiotic and bio-preservative, an increase in probiotic population was observed [317].
In human studies, though the dietary fibre delayed the absorption of phenolics, it showed that the dietary fibre intake increased plasma antioxidant capacity, demonstrating that the combination makes it partially bioavailable to humans [312]. This makes dietary fibres from wineries’ by-products an appealing source for the development of nutraceuticals with the goal of long-term enhanced bio-delivery of their bioactives.
4.4. Limitations in the applications of wineries by-products and of their bioactive ingredients
The first impediment to widespread industrial utilization of grape pomace is its limited chemical stability largely due to its high water content, which is likely to also cause microbial and enzymatic degradation. As discussed in the extraction processes, the low thermal stability of the bioactive compounds in the pomace is susceptible to degradation under high heat. Due to these reasons, dehydration of wet grape pomace becomes an essential procedure prior to any subsequent application. Many conventional process techniques have been suggested, such as infrared drying, freeze drying and gamma irradiation [318,319,320]. Furthermore, non-conventional extraction methods such as ultrasound, supercritical fluid extraction and pulsed electric fields have also been brought up, only to be limited by high initial costs and integrations. Therefore, traditional drying procedures are still the most popular ways of treating wine pomace due to costs. A study has revealed that drying grape pomace, instead of seeds and skins individually, in a vacuum drier at 70℃ gave the quickest drying time and energy savings. The process also showed high polyphenol stability despite their volatility to heat when compared to conventional drying methods [321]. Further processing with surfactants could also increase polyphenol stability. In a demonstration to decrease anthocyanin color change and degradation rate, both polysorbate 20 and sorbic acid were used as surfactants to process the phenolic compound with great success [322].
Overall, numerous studies have demonstrated the successful incorporation of grape pomace and its bioactive compounds into many plant-derived and animal-derived foodstuffs for the purpose of fortification and supplements. The result of this addition includes increased overall polyphenolic concentration and as a result oxidative stability, leading to longer and more stable shelf life in most products. However, phenols exhibit sensory properties that include bitterness, acidity and astringency and are likely not well accepted by consumers, especially when presented in meat derivatives. Discoloration of meat products have generally led to lowered consumer acceptance. Of course, the amount of discoloration generated by grape pomace extract may vary depending on the grape varietal, and antioxidants with less anthocyanin contents may be more suitable in these conditions. However, textural differences have also been described in the addition of grape pomace especially in baked products, showing decreased chewiness or hardness, demonstrating a commercial challenge if incorporation of grape pomace were to be included on a large-scale production. Textural issues were also shown in pasta products, as the addition of grape pomace increased water-binding capacity, water retention rates were higher in products containing grape pomace. In cheese products, the fortified samples displayed a marbling effect, as well as negative sensory properties such as increased granularity and saltiness [323].
In addition to sensory issues, polyphenol interaction with other compounds plays a critical part in the formulation of successful functional food products. The most extensively studied are wine and grape derived tannins, also found in high amounts in wineries’ by-products, which demonstrate the ability to bind to residual proteins and polysaccharides. The astringency of such products is decreased during an increase in polysaccharides, rather than a reduction in tannin content [324]. However, the addition of the sweetener aspartame had no effect on the perceived astringency [325]. Similarly, a study showed that the higher the addition of grape pomace, the lower the protein digestibility in cookies, signaling a related issue [326]. These examples exhibit the requirement for extending our knowledge of polyphenol structure and binding, as it would have important implications in the application for novel food products.
Another issue that bears mentioning is whether the effects of added grape pomace or isolated phenols and its subsequent health benefits are obtainable by the body. The bioavailability may depend on bioaccessibility of compounds in the food matrix [327]. As aforementioned, a solution to this issue that is currently under research is the use of several delivery systems like emulsions may enhance the bio-delivery and bioavailability of winery by-products bioactives [303,304,305,306,307,308,309,310]. Another natural bio-delivery system of such bioactives has also been found to be the winery by-products dietary fibers [108,200,219,311,312,313,314,315,316,317] However, it was also observed that an increase in dietary fibre presence lowered polyphenol bioaccessibility in the small intestine [311]. This is extremely interesting as the same fibre complex that forms with polyphenols was also able to increasing the holding capacity for phenolic compounds [175], while simultaneously decreasing digestibility of the compounds. This could potentially increase the quantity of polyphenols that could reach the lower portions of the intestines. However, more research is needed to understand the influence of dietary fibres and further beneficial activities in the colon.
Another primary concern for using a waste product such as grape pomace is the possibility of harmful residues such as pesticides or heavy metals. Fortunately, vinification process such as pressing and separation, and post processes such the drying process drastically reduce pesticide and fungicide residue ([328,329]. However, usage of grape skin powders gave rise to high amounts of sodium, as well as lead and cadmium though the latter two were well below the legal limits [330]. The source of grape pomace would have to be constantly monitored for any contaminants, and the extracts have to be carefully selected as to avoid any harmful residues.
5. Conclusion
Within this article, a holistic review was conducted on wine’s bioactives, their mode of action and the associated health benefits of this bio-functional beverage when consumed in moderation, as well as on the valorization of the wineries’ by-products bioactive compounds in several health promoting applications, as functional ingredients for fortification of foods and the development of supplements and nutraceuticals.
With respect to the health benefits of moderate wine consumption, as part of a healthy, balanced diet, apart from observed associations mentioned in a plethora of epidemiological studies, a broad range of in vitro and in vivo studies, including randomized control clinical trials, have also outlined the various mechanisms and synergistic actions by which wine’s bioactives (phenolics, UFA and polar lipids, as well as their metabolites after digestion) mediate their antioxidant, anti-thrombotic and anti-inflammatory effects against several bio-molecular, biochemical and biological inflammatory manifestations and cell/tissue/body responses, which further support the bio-functional health promoting effects of wine consumption in moderation against inflammation-related chronic disorders. On the other hand, excessive consumption of this alcoholic beverage is not recommended, as it may lead to several negative implications on health, observed in other alcoholic drinks too. Focus has also been given to the putative benefits of de-alcoholized wines, but more targeted research is needed to fully evidence them.
The wine industry is responsible for a considerable part of environmental issues, as they dispose large amounts of residues that are rich in polyphenols and fibers, such as grape pomace and seeds. On the other hand, the rich content of such wine industry by-products in several of these compounds, has made them promising sustainable sources of natural anti-inflammatory bioactives and antioxidants, as well as a great sources of dietary fibre and plant-based bio-functional oils, which if appropriately recovered can be valorized as functional ingredients in food products and supplements/nutraceuticals, or even for improving cosmetics.
The potential of the key bioactive components of wineries’ by-products, dietary fibre, polyphenols, UFA and polar lipids, as well as several of their extracts, to be employed for the fortification of existing/novel nutraceutical food products and supplements, with higher overall polyphenolic content, improved oxidative stability and shelf life, and in some cases with enhanced health promoting effects, has also been outlined. These compounds can replace synthetic additives, adding multifunctional concepts by combining health benefits to technological use. The positive biochemical qualities and appropriate recovery and utilization of these bioactive compounds and extracts from wineries’ by-products, have been connected to a number of health benefits, including inflammatory pathway suppression, oxidative stress reduction and lipid lowering effects, protection against tumors, neurodegenerative disorders and several pathogens, and an overall prevention against several chronic diseases.
The large number of applications mentioned in this review demonstrates the tremendous untapped potential for the food sector to re-evaluate and promote further research on the valorization of wineries’ by-products and/or of their bioactives. Not only does this improve environmental issues and encourage sustainable use of waste products, but it could also potentially lower production costs for antioxidants and preservatives and provide new methods to diversify food productions and the development of supplements. More bioactive compounds in wine residues should be isolated and identified and their underlying mechanisms of action should be further evaluated, while their applications should further be explored in more food products and more clinical trials should also be carried out to further confirm the health benefits of thus produced functional foods.
Notably, the undesirable effects of the extraction and isolation processes on the recovered bioactives, as well as the drawbacks of negative sensory properties should also be considered. Traditional methods are still the most used for the extractions of compounds of interest, while alternative “greener” technologies are emerging to recover bioactive compounds from wineries’ by-products, while the food industry should also incorporate consumer response and their acceptance towards such products closely. Thus, further research is required in the optimization and modification of recovery processes and food formulas, so that they may be used to their maximum potential, while every attempt to study and improve the usage of bioactive compounds should be made and assessed with care and with consumers’ acceptance, in order to appropriately address the demand for new and improved sustainable natural products.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, A.T.; methodology, A.T.,; software, E.O.M., V.L.J.N., M.K. and A.T.; validation, A.T. and M.K.; investigation, A.T.; writing—original draft preparation, E.O.M., V.L.J.N., M.K. and A.T.; writing—review and editing, A.T.; visualization, A.T.; supervision, A.T.; project administration, A.T.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to thank Department of Chemistry of the International Hellenic University (Kavale, Greece), and especially the Head of the Department, Prof G. Kyzas, as well as the Department of Biological Sciences of the School of Science and Engineering of the University of Limerick (Limerick, Ireland), and especially the Dean og the School of Sciences & Engineering, Prof Sean Arkins, for their continuous support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Basalekou, M.; Kallithraka, S.; Kyraleou, M. Wine bioactive compounds. In Functional Foods and Their Implications for Health Promotion, 1st ed.; Zabetakis, I., Tsoupras, A., Lordan, R., Ramji, D., Eds.; Academic Press, 2023; Volume 13, pp. 341–63. [Google Scholar]
- Calabriso, N.; Scoditti, E.; Massaro, M.; Pellegrino, M.; Storelli, C.; Ingrosso, I.; et al. Multiple anti-inflammatory and anti-atherosclerotic properties of red wine polyphenolic extracts: differential role of hydroxycinnamic acids, flavonols and stilbenes on endothelial inflammatory gene expression. Eur J Nutr 2016, 55, 477–89. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Fidalgo, S.; Cárdeno, A.; Villegas, I.; Talero, E.; de la Lastra, C.A. Dietary supplementation of resveratrol attenuates chronic colonic inflammation in mice. Eur J Pharmac. 2010, 633, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, I.; Pérez-Gregorio, R.; Soares, S.; Mateus, N.; De Freitas, V. Wine Flavonoids in Health and Disease Prevention. Molecules. 2017, 22, 292. [Google Scholar] [CrossRef] [PubMed]
- Tsoupras, A.B.; Fragopoulou, E.; Nomikos, T.; Iatrou, C.; Antonopoulou, S.; Demopoulos, C.A. Characterization of the de novo biosynthetic enzyme of platelet activating factor, DDT-insensitive cholinephosphotransferase, of human mesangial cells. Mediators Inflamm. 2007, 2007, 27683. [Google Scholar] [CrossRef]
- Tsoupras, A.B.; Iatrou, C.; Frangia, C.; Demopoulos, C.A. The implication of platelet activating factor in cancer growth and metastasis: potent beneficial role of PAF-inhibitors and antioxidants. Infect Disord Drug Targets. 2009, 9, 390–9. [Google Scholar] [CrossRef]
- Tsoupras, A.; Lordan, R.; Zabetakis, I. Inflammation, not Cholesterol, Is a Cause of Chronic Disease. Nutrients. 2018, 10, 604. [Google Scholar] [CrossRef]
- Fragopoulou, E.; Nomikos, T.; Tsantila, N.; Mitropoulou, A.; Zabetakis, I.; Demopoulos, C.A. Biological Activity of Total Lipids from Red and White Wine/Must. J Agr Food Chem. 2001, 49, 5186–93. [Google Scholar] [CrossRef] [PubMed]
- Fragopoulou, E.; Antonopoulou, S.; Demopoulos, C.A. Biologically Active Lipids with Antiatherogenic Properties from White Wine and Must. J Agr Food Chem. 2002, 50, 2684–94. [Google Scholar] [CrossRef]
- Fragopoulou, E.; Antonopoulou, S.; Tsoupras, A.; Tsantila, N.; Grypioti, A.; Gribilas, G.; Gritzapi, H.; Konsta, E.; Skandalou, E.; Papadopoulou, A. Antiatherogenic properties of red/white wine, musts, grape-skins, and yeast. In Proceedings of the 45th International Conference on the Bioscience of Lipids, University of, Ioannina, Ioannina, Greece, 25–29 May 2004; p. 66. [Google Scholar]
- Fragopoulou, E.; Demopoulos, C.A.; Antonopoulou, S. Lipid Minor Constituents in Wines. A Biochemical Approach in the French Paradox. Int. J. Wine Res. 2009, 1, 131–143. [Google Scholar]
- Xanthopoulou, M.N.; Kalathara, K.; Melachroinou, S.; Arampatzi-Menenakou, K.; Antonopoulou, S.; Yannakoulia, M.; Fragopoulou, E. Wine Consumption Reduced Postprandial Platelet Sensitivity against Platelet Activating Factor in Healthy Men. Eur. J. Nutr. 2017, 56, 1485–1492. [Google Scholar] [CrossRef]
- Argyrou, C.; Vlachogianni, I.; Stamatakis, G.; Demopoulos, C.A.; Antonopoulou, S.; Fragopoulou, E. Postprandial Effects of Wine Consumption on Platelet Activating Factor Metabolic Enzymes. Prostaglandins Other Lipid Mediat. 2017, 130, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Fragopoulou, E.; Choleva, M.; Antonopoulou, S.; Demopoulos, C.A. Wine and its metabolic effects. A comprehensive review of clinical trials. Metabolism. 2018, 83, 102–19. [Google Scholar] [CrossRef] [PubMed]
- Gavriil, L.; Detopoulou, M.; Petsini, F.; Antonopoulou, S.; Fragopoulou, E. Consumption of plant extract supplement reduces platelet activating factor-induced platelet aggregation and increases platelet activating factor catabolism: a randomised, double-blind and placebo-controlled trial. Br J Nutr 2019, 121, 982–991. [Google Scholar] [CrossRef]
- Choleva, M.; Boulougouri, V.; Panara, A.; Panagopoulou, E.; Chiou, A.; Thomaidis, N.S.; Antonopoulou, S.; Fragopoulou, E. Evaluation of Anti-Platelet Activity of Grape Pomace Extracts. Food Funct. 2019, 10, 8069–8080. [Google Scholar] [CrossRef]
- Choleva, M.; Tsota, M.; Boulougouri, V.; Panara, A.; Thomaidis, N.; Antonopoulou, S.; & Fragopoulou, E. Anti-platelet and anti-inflammatory properties of an ethanol-water red grape pomace extract. Proceedings of the Nutrition Society, 2020, 79(OCE2), E370.
- Fragopoulou, E.; Antonopoulou, S. The French paradox three decades later: Role of inflammation and thrombosis. Clin Chim Acta. 2020, 510, 160–9. [Google Scholar] [CrossRef]
- Fragopoulou, E.; Argyrou, C.; Detopoulou, M.; Tsitsou, S.; Seremeti, S.; Yannakoulia, M.; Antonopoulou, S.; Kolovou, G.; Kalogeropoulos, P. The Effect of Moderate Wine Consumption on Cytokine Secretion by Peripheral Blood Mononuclear Cells: A Randomized Clinical Study in Coronary Heart Disease Patients. Cytokine 2021, 146, 155629. [Google Scholar] [CrossRef]
- Choleva, M.; Argyrou, C.; Detopoulou, M.; Donta, M.-E.; Gerogianni, A.; Moustou, E.; Papaemmanouil, A.; Skitsa, C.; Kolovou, G.; Kalogeropoulos, P.; et al. Effect of Moderate Wine Consumption on Oxidative Stress Markers in Coronary Heart Disease Patients. Nutrients 2022, 14, 1377. [Google Scholar] [CrossRef]
- Choleva, M.; Matalliotaki, E.; Antoniou, S.; Asimomyti, E.; Drouka, A.; Stefani, M.; Yannakoulia, M.; Fragopoulou, E. Postprandial Metabolic and Oxidative Stress Responses to Grape Pomace Extract in Healthy Normal and Overweight/Obese Women: A Randomized, Double-Blind, Placebo-Controlled Crossover Study. Nutrients 2023, 15, 156. [Google Scholar] [CrossRef]
- Karantonis, H.C.; Tsoupras, A.; Moran, D.; Zabetakis, I.; Nasopoulou, C. Olive, apple, and grape pomaces with antioxidant and anti-inflammatory bioactivities for functional foods. In Functional Foods and Their Implications for Health Promotion, 1st ed.; Zabetakis, I., Tsoupras, A., Lordan, R., Ramji, D., Eds.; Academic Press; 2023; Volume 5, pp. 131–159.
- Sabra, A.; Netticadan, T.; Wijekoon, C. Grape bioactive molecules, and the potential health benefits in reducing the risk of heart diseases. Food Chem X. 2021, 12, 100149. [Google Scholar] [CrossRef]
- Tsoupras, A; Lordan, R.; Zabetakis, I. Inflammation and Cardiovascular Diseases. In The Impact of Nutrition and Statins on Cardiovascular Diseases, 1st ed. Zabetakis, I., Lordan, R., Tsoupras, A., Eds. Academic Press; 2019. Volume 3, pp. 53–117.
- van Bussel, B.C.T.; Henry, R.M.A.; Schalkwijk, C.G.; Dekker, J.M.; Nijpels, G.; Feskens, E.J.M.; et al. Alcohol and red wine consumption, but not fruit, vegetables, fish or dairy products, are associated with less endothelial dysfunction and less low-grade inflammation: the Hoorn Study. Eur J Nutr. 2018, 57, 1409–19. [Google Scholar] [CrossRef] [PubMed]
- Queipo-Ortuño, M.I.; Boto-Ordóñez, M.; Murri, M.; Gomez-Zumaquero, J.M.; Clemente-Postigo, M.; Estruch, R.; et al. Influence of red wine polyphenols and ethanol on the gut microbiota ecology and biochemical biomarkers. Am J Clin Nutr. 2012, 95, 1323–34. [Google Scholar] [CrossRef]
- Panagiotakos, D.B.; Pitsavos, C.; Arvaniti, F.; Stefanadis, C. Adherence to the Mediterranean Food Pattern Predicts the Prevalence of Hypertension, Hypercholesterolemia, Diabetes and Obesity, among Healthy Adults; the Accuracy of the MedDietScore. Prev. Med. 2007, 44, 335–340. [Google Scholar] [CrossRef]
- Esposito, K.; Maiorino, M.I.; Bellastella, G.; Panagiotakos, D.B.; Giugliano, D. Mediterranean diet for type 2 diabetes: cardiometabolic benefits. Endocrine. 2017, 56, 27–32. [Google Scholar] [CrossRef]
- Piano, M.R. Alcohol's Effects on the Cardiovascular System. Alcohol Res. 2017, 38, 219–41. [Google Scholar]
- Markoski, M.M.; Garavaglia, J.; Oliveira, A.; Olivaes, J.; Marcadenti, A. Molecular Properties of Red Wine Compounds and Cardiometabolic Benefits. Nutr Metabol Ins. 2016, 9, 51–7. [Google Scholar] [CrossRef]
- Poklar Ulrih, N.; Opara, R.; Skrt, M.; Košmerl, T.; Wondra, M.; Abram, V. Part I. Polyphenols composition and antioxidant potential during 'Blaufränkisch' grape maceration and red wine maturation, and the effects of trans-resveratrol addition. Food Chem Toxicol. 2020, 137, 111122. [Google Scholar] [CrossRef]
- Xiang, L.; Xiao, L.; Wang, Y.; Li, H.; Huang, Z.; He, X. Health benefits of wine: Don’t expect resveratrol too much. Food Chem. 2014, 156, 258–63. [Google Scholar] [CrossRef] [PubMed]
- Antonić, B.; Jančíková, S.; Dordević, D.; Tremlová, B. Grape Pomace Valorization: A Systematic Review and Meta-Analysis. Foods 2020, 9, 1627. [Google Scholar] [CrossRef] [PubMed]
- Arboleda Mejia, J.A.; Ricci, A.; Figueiredo, A.S.; Versari, A.; Cassano, A.; Parpinello, G.P.; De Pinho, M.N. Recovery of Phenolic Compounds from Red Grape Pomace Extract through Nanofiltration Membranes. Foods 2020, 9, 1649. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, M.S.; Torres, J.S.; Nuñez, M.J. ;. Extraction of polyphenols from white distilled grape pomace: Optimization and modelling. Biores Tech 2008, 99, 1311–1318. [Google Scholar] [CrossRef]
- Lingua, M.S.; Fabani, M.P.; Wunderlin, D.A.; Baroni, M.V. From grape to wine: Changes in phenolic composition and its influence on antioxidant activity. Food Chem 2016, 208, 228–38. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, Y.; Göksel, Z.; Erdoğan, S.S.; Öztürk, A.; Atak, A.; Özer, C. Antioxidant Activity and Phenolic Content of Seed, Skin and Pulp Parts of 22 Grape (Vitis vinifera L.) Cultivars (4 Common and 18 Registered or Candidate for Registration). J Food Proc Preserv. 2015, 39, 1682–91. [Google Scholar] [CrossRef]
- Ferri, M.; Bin, S.; Vallini, V.; Fava, F.; Michelini, E.; Roda, A.; Minnucci, G.; Bucchi, G.; Tassoni, A. Recovery of polyphenols from red grape pomace and assessment of their antioxidant and anti-cholesterol activities. New Biotechnol. 2016, 33, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Iora, S.R.F.; Maciel, G.M.; Zielinski, A.A.F.; da Silva, M.V.; Pontes, P.V. de A.; Haminiuk, C.W.I.; Granato, D. Evaluation of the bioactive compounds and the antioxidant capacity of grape pomace. Int J Food Sci Tech 2015, 50, 62–69. [Google Scholar] [CrossRef]
- Negro, C.; Tommasi, L.; Miceli, A. Phenolic compounds and antioxidant activity from red grape marc extracts. Biores Tech 2003, 87, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Szabó, É.; Marosvölgyi, T.; Szilágyi, G.; Kőrösi, L.; Schmidt, J.; Csepregi, K.; Márk, L.; Bóna, Á. Correlations between Total Antioxidant Capacity, Polyphenol and Fatty Acid Content of Native Grape Seed and Pomace of Four Different Grape Varieties in Hungary. Antioxidants 2021, 10, 1101. [Google Scholar] [CrossRef]
- Chedea, V.S.; Macovei, Ș.O.; Bocșan, I.C.; Măgureanu, D.C.; Levai, A.M.; Buzoianu, A.D.; Pop, R.M. Grape Pomace Polyphenols as a Source of Compounds for Management of Oxidative Stress and Inflammation—A Possible Alternative for Non-Steroidal Anti-Inflammatory Drugs? Molecules 2022, 27, 6826. [Google Scholar] [CrossRef]
- Visioli, F.; Panaite, S.A.; Tomé-Carneiro, J. Wine’s Phenolic Compounds and Health: A Pythagorean View. Molecules 2020, 25, 4105. [Google Scholar] [CrossRef]
- Kennedy, J.A.; Saucier, C.; Glories, Y. Grape and Wine Phenolics: History and Perspective. Am J Enol Vitic. 2006, 57, 239–248. [Google Scholar] [CrossRef]
- Lago-Vanzela, E.; Alves Baffi, M.; Castilhos, M.; Ribeiro-Pinto, M.; Del Bianchi, V.; Ramos, A.; Stringheta, P.; Hermosín-Gutiérrez, I.; Da Silva, R. Phenolic compounds in grapes and wines: Chemical and biochemical characteristics and technological quality. In Grapes: production, phenolic composition and potential biomedical effects, 1st ed.; Câmara J.S., Eds.; Nova Science Publishers Inc.; 2014; Volume 3, pp. 1–18.
- Zhou, D.-D.; Li, J.; Xiong, R.-G.; Saimaiti, A.; Huang, S.-Y.; Wu, S.-X.; Yang, Z.-J.; Shang, A.; Zhao, C.-N.; Gan, R.-Y.; Li, H.-B. Bioactive Compounds, Health Benefits and Food Applications of Grape. Foods 2022, 11, 2755. [Google Scholar] [CrossRef]
- Panzella, L.; Moccia, F.; Nasti, R.; Marzorati, S.; Verotta, L.; Napolitano, A. Bioactive Phenolic Compounds From Agri-Food Wastes: An Update on Green and Sustainable Extraction Methodologies. Front. Nutr. 2020, 7, 60. [Google Scholar] [CrossRef]
- Cordova, A.C.; Sumpio, B.E. Polyphenols are medicine: Is it time to prescribe red wine for our patients? Int J Angiol 2009, 18, 111–117. [Google Scholar] [CrossRef]
- Snopek, L.; Mlcek, J.; Sochorova, L.; Baron, M.; Hlavacova, I.; Jurikova, T.; Kizek, R.; Sedlackova, E.; Sochor, J. Contribution of Red Wine Consumption to Human Health Protection. Molecules 2018, 23. [Google Scholar] [CrossRef]
- Hrelia, S.; Di Renzo, L.; Bavaresco, L.; Bernardi, E.; Malaguti, M.; Giacosa, A. Moderate Wine Consumption and Health: A Narrative Review. Nutrients 2023, 15, 175. [Google Scholar] [CrossRef]
- Aviram, M.; Fuhrman, B. Wine Flavonoids Protect against LDL Oxidation and Atherosclerosis. Ann N Y Acad Sci 2002, 957, 146–161. [Google Scholar] [CrossRef]
- Nigdikar, S.V.; Williams, N.R.; Griffin, B.A.; Howard, A.N. Consumption of red wine polyphenols reduces the susceptibility of low-density lipoproteins to oxidation in vivo. Am J Clin Nutr 1998, 68, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Apostolidou, C.; Adamopoulos, K.; Lymperaki, E.; Iliadis, S.; Papapreponis, P.; Kourtidou-Papadeli, C. Cardiovascular risk and benefits from antioxidant dietary intervention with red wine in asymptomatic hypercholesterolemics. Clin Nutr ESPEN. 2015, 10, e224–e33. [Google Scholar] [CrossRef] [PubMed]
- Stranieri, C.; Guzzo, F.; Gambini, S.; Cominacini, L.; Fratta Pasini, A.M. Intracellular Polyphenol Wine Metabolites Oppose Oxidative Stress and Upregulate Nrf2/ARE Pathway. Antioxidants 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Schrieks, I.C.; van den Berg, R.; Sierksma, A.; Beulens, J.W.; Vaes, W.H.; Hendriks, H.F. Effect of red wine consumption on biomarkers of oxidative stress. Alcohol Alcohol. 2013, 48, 153–9. [Google Scholar] [CrossRef] [PubMed]
- Chiva-Blanch, G.; Arranz, S.; Lamuela-Raventos, R.M.; Estruch, R. Effects of wine, alcohol and polyphenols on cardiovascular disease risk factors: Evidences from human studies. Alcohol Alcohol. 2013, 48, 270–7. [Google Scholar] [CrossRef] [PubMed]
- Mangge, H.; Becker, K.; Fuchs, D.; Gostner, J.M. Antioxidants, inflammation and cardiovascular disease. World J Cardiol 2014, 6, 462–477. [Google Scholar] [CrossRef] [PubMed]
- Leri, M.; Scuto, M.; Ontario, M.L.; Calabrese, V.; Calabrese, E.J.; Bucciantini, M.; et al. Healthy Effects of Plant Polyphenols: Molecular Mechanisms. Int J Mol Sci. 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Habauzit, V.; Morand, C. Evidence for a protective effect of polyphenols-containing foods on cardiovascular health: an update for clinicians. Ther Adv Chronic Dis. 2012, 3, 87–106. [Google Scholar] [CrossRef] [PubMed]
- Di Renzo, L,; Marsella, L. T.; Carraro, A.; Valente, R.; Gualtieri, P.; Gratteri, S.; et al. Changes in LDL Oxidative Status and Oxidative and Inflammatory Gene Expression after Red Wine Intake in Healthy People: A Randomized Trial. Mediators Inflamm. 2015, 2015. [Google Scholar]
- Torres, A.; Cachofeiro, V.; Millán, J.; Lahera, V.; Nieto, M.L.; Martín, R.; Bello, E.; Alvarez-Sala, L.A. Red wine intake but not other alcoholic beverages increases total antioxidant capacity and improves pro-inflammatory profile after an oral fat diet in healthy volunteers. Rev Clin Esp (Barc). 2015, 215, 486–94. [Google Scholar] [CrossRef]
- Weseler, A.R.; Ruijters, E.J.B.; Drittij-Reijnders, M-J. ; Reesink, K.D.; Haenen, G.R.M.M.; Bast, A. Pleiotropic Benefit of Monomeric and Oligomeric Flavanols on Vascular Health - A Randomized Controlled Clinical Pilot Study. Plos One. 2011, 6, e28460. [Google Scholar] [CrossRef]
- Yang, H.; Xiao, L.; Yuan, Y.; Luo, X.; Jiang, M.; Ni, J.; et al. Procyanidin B2 inhibits NLRP3 inflammasome activation in human vascular endothelial cells. Biochem Pharmacol. 2014, 92, 599–606. [Google Scholar] [CrossRef]
- de Lorgeril, M.; Salen, P.; Martin, J.L.; Monjaud, I.; Delaye, J.; Mamelle, N. Mediterranean diet, traditional risk factors, and the rate of cardiovascular complications after myocardial infarction: final report of the Lyon Diet Heart Study. Circulation. 1999, 99, 779–85. [Google Scholar] [CrossRef]
- Meng, X.; Zhou, J.; Zhao, C.N.; Gan, R.Y.; Li, H.B. Health benefits and molecular mechanisms of resveratrol: A narrative review. Foods 2020, 9, 340. [Google Scholar] [CrossRef]
- Khattar, S.; Khan, S.A.; Zaidi, S.A.A.; Darvishikolour, M.; Farooq, U.; Naseef, P.P.; Kurunian, M.S.; Khan, M.Z.; Shamim, A.; Khan, M.M.U.; Iqbal, Z.; Mirza, M.A. Resveratrol from Dietary Supplement to a Drug Candidate: An Assessment of Potential. Pharmaceuticals 2022, 15, 957. [Google Scholar] [CrossRef]
- Zhang, L.X.; Li, C.X.; Kakar, M.U.; Khan, M.S.; Wu, P.F.; Amir, R.M.; Dai, D.F.; Naveed, M.; Li, Q.Y.; Saeed, M.; Shen, J.Q.; Rajput, S.A.; Li, J.H. Resveratrol (RV): A pharmacological review and call for further research. Biomed Pharmacother. 2021, 143, 112164. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Quispe, C.; Zam, W.; Kumar, M.; Cardoso, S.M.; Pereira, O.R.; Ademiluyi, A.O.; Adeleke, O.; Moreira, A.C.; Živković, J. Phenolic bioactives as antiplatelet aggregation factors: the pivotal ingredients in maintaining cardiovascular health. Oxid. Med. Cell. Longev., 2021, 2021, 2195902. [Google Scholar] [CrossRef]
- Parsamanesh, N.; Asghari, A.; Sardari, S.; Tasbandi, A.; Jamialahmadi, T.; Xu, S.; Sahebkar, A. Resveratrol and endothelial function: a literature review. Pharmacol. Res., 2021, 170, 105725. [Google Scholar] [CrossRef]
- Muñoz-Bernal, Ó.A.; Coria-Oliveros, A.J.; de la Rosa, L.A.; Rodrigo-García, J.; Del Rocío Martínez-Ruiz, N.; Sayago-Ayerdi, S.G.; Alvarez-Parrilla, E. Cardioprotective effect of red wine and grape pomace. Food Res Int. 2021, 140, 110069. [Google Scholar] [CrossRef] [PubMed]
- Wiciński, M.; Socha, M.; Walczak, M.; Wódkiewicz, E.; Malinowski, B.; Rewerski, S.; Górski, K.; Pawlak-Osińska, K. Beneficial Effects of Resveratrol Administration-Focus on Potential Biochemical Mechanisms in Cardiovascular Conditions. Nutrients. 2018, 10, 1813. [Google Scholar] [CrossRef]
- Kuršvietienė, L.; Stanevičienė, I.; Mongirdienė, A.; Bernatonienė, J. Multiplicity of effects and health benefits of resveratrol. Medicina (Kaunas). 2016, 52, 148–55. [Google Scholar] [CrossRef]
- Riccioni, G.; Gammone, M.A.; Tettamanti, G.; Bergante, S.; Pluchinotta, F.R.; D'Orazio, N. Resveratrol and anti-atherogenic effects. Int J Food Sci Nutr. 2015, 66, 603–10. [Google Scholar] [CrossRef] [PubMed]
- Tamer, F.; Tullemans, B.M.E.; Kuijpers, M.J.E.; Claushuis, T.A.M.; Heemskerk, J.W.M. Nutrition Phytochemicals Affecting Platelet Signaling and Responsiveness: Implications for Thrombosis and Hemostasis. Thromb Haemost. 2022, 122, 879–894. [Google Scholar] [CrossRef] [PubMed]
- Shahcheraghi, S.H.; Salemi, F.; Small, S.; Syed, S.; Salari, F.; Alam, W.; et al. Resveratrol regulates inflammation and improves oxidative stress via Nrf2 signaling pathway: Therapeutic and biotechnological prospects. Phytother Res. 2023. [Google Scholar] [CrossRef]
- Rius, C.; Abu-Taha, M.; Hermenegildo, C.; et al. , “Trans- but Not Cis-Resveratrol impairs angiotensin-II-mediated vascular inflammation through inhibition of NF-κB activation and peroxisome proliferator-activated Receptor-γ upregulation,” J Immunol 2010, 185, 3718–3727.
- Csiszar, A.; Smith, K.; Labinskyy, N.; Orosz, Z.; Rivera, A.; Ungvari, Z. Resveratrol attenuates TNF-alpha-induced activation of coronary arterial endothelial cells: role of NF-kappaB inhibition. Am J Physiol Heart Circ Physiol. 2006, 291, H1694–9. [Google Scholar] [CrossRef]
- Toaldo, I.M.; Van Camp, J.; Gonzales, G.B.; Kamiloglu, S.; Bordignon-Luiz, M.T.; Smagghe, G.; et al. Resveratrol improves TNF-α-induced endothelial dysfunction in a coculture model of a Caco-2 with an endothelial cell line. J Nutr Biochem. 2016, 36, 21–30. [Google Scholar] [CrossRef]
- Chalons, P.; Amor, S.; Courtaut, F.; Cantos-Villar, E.; Richard, T.; Auger, C.; et al. Study of potential anti-inflammatory effects of red wine extract and resveratrol through a modulation of interleukin-1-beta in macrophages. Nutrients. 2018, 10. [Google Scholar] [CrossRef]
- Fukuda, M.; Ogasawara, Y.; Hayashi, H.; Inoue, K.; Sakashita, H. Resveratrol Inhibits Proliferation and Induces Autophagy by Blocking SREBP1 Expression in Oral Cancer Cells. Molecules. 2022, 27. [Google Scholar] [CrossRef]
- Zucchi, A.; Claps, F.; Pastore, A.L.; Perotti, A.; Biagini, A.; Sallicandro, L.; Gentile, R.; Caglioti, C.; Palazzetti, F.; Fioretti, B. Focus on the Use of Resveratrol in Bladder Cancer. Int J Mol Sci. 2023, 24, 4562. [Google Scholar] [CrossRef]
- Buljeta, I.; Pichler, A.; Šimunović, J.; Kopjar, M. Beneficial Effects of Red Wine Polyphenols on Human Health: Comprehensive Review. Curr Issues Mol Biol. 2023, 45, 782–798. [Google Scholar] [CrossRef]
- Chimento, A.; D'Amico, M.; De Luca, A.; Conforti, F.L.; Pezzi, V.; De Amicis, F. Resveratrol, Epigallocatechin Gallate and Curcumin for Cancer Therapy: Challenges from Their Pro-Apoptotic Properties. Life 2023, 13, 261. [Google Scholar] [CrossRef]
- Lalani, A.R.; Fakhari, F.; Radgoudarzi, S.; Rastegar-Pouyani, N.; Moloudi, K.; Khodamoradi, E.; Taeb, S.; Najafi, M. Immunoregulation by resveratrol; implications for normal tissue protection and tumour suppression. Clin Exp Pharmacol Physiol. 2023, 50, 353–368. [Google Scholar] [CrossRef] [PubMed]
- Angellotti, G.; Di Prima, G.; Belfiore, E.; Campisi, G.; De Caro, V. Chemopreventive and Anticancer Role of Resveratrol against Oral Squamous Cell Carcinoma. Pharmaceutics. 2023, 15, 275. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.S.; Gadi, V.; Kaur, G.; Chintamaneni, M.; Tuli, H.; Ramniwas, S.; Sethi, G. Resveratrol and Its Role in the Management of B-Cell Malignancies-A Recent Update. Biomedicines. 2023, 11, 221. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Fan, F.; Hou, X.; Gao, C.; Meng, L.; Meng, S.; Huang, S.; Wu, H. Resveratrol suppresses pulmonary tumor metastasis by inhibiting platelet-mediated angiogenic responses. J Surg Res. 2017, 217, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Bae, J.U.; Kim, I.S.; Chang, C.L.; Oh, S.O.; Kim, C.D. SIRT1 prevents pulmonary thrombus formation induced by arachidonic acid via downregulation of PAF receptor expression in platelets. Platelets. 2016, 27, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Michno, A.; Grużewska, K.; Ronowska, A.; Gul-Hinc, S.; Zyśk, M.; Jankowska-Kulawy, A. Resveratrol Inhibits Metabolism and Affects Blood Platelet Function in Type 2 Diabetes. Nutrients. 2022, 14, 1633. [Google Scholar] [CrossRef] [PubMed]
- Crescente, M.; Jessen, G.; Momi, S.; et al. , “Interactions of gallic acid, resveratrol, quercetin and aspirin at the platelet cyclooxygenase-1 level. Functional and modelling studies. Thromb Haem, 2009, 102, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Marumo, M.; Ekawa, K.; Wakabayashi, I. Resveratrol inhibits Ca2+ signals and aggregation of platelets. Environ Health Prev Med. 2020, 25, 70. [Google Scholar] [CrossRef] [PubMed]
- Vlachogianni, I.C.; Fragopoulou, E.; Stamatakis, G.M.; Kostakis, I.K.; Antonopoulou, S. Platelet Activating Factor (PAF) biosynthesis is inhibited by phenolic compounds in U-937 cells under inflammatory conditions. Prostaglandins Other Lipid Mediat. 2015, 121, 176–83. [Google Scholar] [CrossRef] [PubMed]
- Eräsalo, H.; Hämäläinen, M.; Leppänen, T.; Mäki-Opas, I.; Laavola, M.; Haavikko, R.; Yli-Kauhaluoma, J.; Moilanen, E. Natural Stilbenoids Have Anti-Inflammatory Properties in Vivo and Down-Regulate the Production of Inflammatory Mediators NO, IL6, and MCP1 Possibly in a PI3K/Akt-Dependent Manner. J Nat Prod. 2018, 81, 1131–1142. [Google Scholar] [CrossRef]
- Dutra, L.A.; Guanaes, J.F.O.; Johmann, N.; Lopes Pires, M.E.; Chin, C.M.; Marcondes, S.; Dos Santos, J.L. Synthesis, antiplatelet and antithrombotic activities of resveratrol derivatives with NO-donor properties. Bioorg. Med. Chem. Lett. 2017, 27, 2450–2453. [Google Scholar] [CrossRef]
- Deng, Y.-H.; Alex, D.; Huang, H.-Q.; et al. Inhibition of TNF-α-mediated endothelial cell-monocyte cell adhesion and adhesion molecules expression by the resveratrol derivative, trans-3,5,4'-trimethoxystilbene. Phytotherapy Research. 2011, 25, 451–457. [Google Scholar] [CrossRef]
- Nash, V.; Ranadheera, C.S.; Georgousopoulou, E.N.; Mellor, D.D.; Panagiotakos, D.B.; McKune, A.J.; et al. The effects of grape and red wine polyphenols on gut microbiota – A systematic review. Food Res Internat. 2018, 113, 277–87. [Google Scholar] [CrossRef]
- Dueñas, M.; Cueva, C.; Muñoz-González, I.; Jiménez-Girón, A.; Sánchez-Patán, F.; Santos-Buelga, C.; et al. Studies on Modulation of Gut Microbiota by Wine Polyphenols: From Isolated Cultures to Omic Approaches. Antioxidants. 2015, 4, 1–21. [Google Scholar] [CrossRef]
- O'Callaghan, A.; van Sinderen, D. Bifidobacteria and Their Role as Members of the Human Gut Microbiota. Front Microbiol. 2016, 7, 925. [Google Scholar] [CrossRef]
- Haas, E.A.; Saad, M.J.A.; Santos, A.; Vitulo, N.; Lemos, W.J.F.; Martins, A.M.A.; et al. A red wine intervention does not modify plasma trimethylamine N-oxide but is associated with broad shifts in the plasma metabolome and gut microbiota composition. Am J Clin Nutr. 2022, 116, 1515–29. [Google Scholar] [CrossRef]
- Suo, H.; Shishir, M.R.I.; Xiao, J.; Wang, M.; Chen, F.; Cheng, K-W. Red Wine High-Molecular-Weight Polyphenolic Complex: An Emerging Modulator of Human Metabolic Disease Risk and Gut Microbiota. J Agr Food Chem. 2021, 69, 10907–19. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, J.; Yin, N.; Wele, P.; Li, F.; Dave, S.; et al. Resveratrol in disease prevention and health promotion: A role of the gut microbiome. Crit Rev Food Sci Nutr. 2023, 1–18. [Google Scholar] [CrossRef]
- Tsoupras, A.; Brummell, C.; Kealy, C.; Vitkaitis, K.; Redfern, S.; Zabetakis, I. Cardio-Protective Properties and Health Benefits of Fish Lipid Bioactives; The Effects of Thermal Processing. Mar Drugs. 2022, 20, 187. [Google Scholar] [CrossRef]
- Nunez, D.; Randon, J.; Gandhi, C.; Siafaka-Kapadai, A.; Olson, M.S.; Hanahan, D.J. The inibition of platelet-activating factor-induced platelet activation by oleic acid is associated with a decrease in polyphosphoinositide metabolism. J. Biol. Chem 1990, 265, 18330–18338. [Google Scholar] [CrossRef]
- Perdomo, L.; Beneit, N.; Otero, Y.F.; Escribano, O.; Diaz-Castroverde, S.; Gómez-Hernández, A.; Benito, M. Protective role of oleic acid against cardiovascular insulin resistance and in the early and late cellular atherosclerotic process. Cardiovasc. Diabetol. 2015, 14, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Delgado, G.E.; Krämer, B.K.; Lorkowski, S.; März, W.; Von Schacky, C.; Kleber, M.E. Individual omega-9 monounsaturated fatty acids and mortality—The Ludwigshafen Risk and Cardiovascular Health Study. J. Clin. Lipidol. 2017, 11, 126–135. [Google Scholar] [CrossRef]
- Holy, E.W.; Forestier, M.; Richter, E.K.; Akhmedov, A.; Leiber, F.; Camici, G.C.; Mocharla, P.; Lüscher, T.F.; Beer, J.H.; Tanner, F.C. Dietary α-linolenic acid inhibits arterial thrombus formation, tissue factor expression, and platelet activation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 1772–1780. [Google Scholar] [CrossRef] [PubMed]
- Bazán-Salinas, I.L.; Matías-Pérez, D.; Pérez-Campos, E.; Pérez-Campos Mayoral, L.; García-Montalvo, I.A. Reduction of platelet aggregation from ingestion of oleic and linoleic acids found in Vitis vinifera and Arachis hypogaea Oils. Am. J. Ther. 2016, 23, e1315–e1319. [Google Scholar] [CrossRef]
- Beres, C.; Costa, G.N.S.; Cabezudo, I.; da Silva-James, N.K.; Teles, A.S.C.; Cruz, A.P.G.; Mellinger-Silva, C.; Tonon, R.V.; Cabral, L.M.C.; Freitas, S.P. Towards integral utilization of grape pomace from winemaking process: A review. Waste Manag. 2017, 68, 581–594. [Google Scholar] [CrossRef]
- Rombaut, N.; Savoire, R.; Thomasset, B.; Castello, J.; Van Hecke, E.; Lanoisellé, J.-L. Optimization of oil yield and oil total phenolic content during grape seed cold screw pressing. Industr Crops Prod 2015, 63, 26–33. [Google Scholar] [CrossRef]
- Garavaglia, J.; Markoski, M.M.; Oliveira, A.; Marcadenti, A. ;. Grape Seed Oil Compounds: Biological and Chemical Actions for Health. Nutr Metab Insights 2016, 9, NMI–S32910. [Google Scholar] [CrossRef]
- Matthäus, B. . Virgin grape seed oil: Is it really a nutritional highlight? Europ J Lipid Sci Techn 2008, 110, 645–650. [Google Scholar] [CrossRef]
- Moran, D.; Fleming, M.; Daly, E.; Gaughan, N.; Zabetakis, I.; Traas, C.; Tsoupras, A. Anti-Platelet Properties of Apple Must/Skin Yeasts and of Their Fermented Apple Cider Products. Beverages, 2021, 7, 54. [Google Scholar] [CrossRef]
- Tsoupras, A.; Lordan, R.; Harrington, J.; Pienaar, R.; Devaney, K.; Heaney, S.; Koidis, A.; Zabetakis, I. The Effects of Oxidation on the Antithrombotic Properties of Tea Lipids against PAF, Thrombin, Collagen, and ADP. Foods 2020, 9, 385. [Google Scholar] [CrossRef]
- Janssen, I.; Landay, A.L; Ruppert, K.; Powell, L.H. Moderate wine consumption is associated with lower hemostatic and inflammatory risk factors over 8 years: The study of women's health across the nation (SWAN). Nutr Aging. 2014, 2, 91–9. [Google Scholar] [CrossRef] [PubMed]
- Panagiotakos, D.B.; Kouli, G-M. ; Magriplis, E.; Kyrou, I.; Georgousopoulou, E.N.; Chrysohoou, C.; et al. Beer, wine consumption, and 10-year CVD incidence: the ATTICA study. Eur J Clin Nutr. 2019, 73, 1015–23. [Google Scholar] [CrossRef] [PubMed]
- Taborsky, M.; Ostadal, P.; Adam, T.; Moravec, O.; Gloger, V.; Schee, A.; et al. Red or white wine consumption effect on atherosclerosis in healthy individuals (In Vino Veritas study). Bratisl Lek Listy. 2017, 118, 292–8. [Google Scholar] [CrossRef] [PubMed]
- Salazar, H.M.; de Deus Mendonça, R.; Laclaustra, M.; Moreno-Franco, B.; Åkesson, A.; Guallar-Castillón, P.; et al. The intake of flavonoids, stilbenes, and tyrosols, mainly consumed through red wine and virgin olive oil, is associated with lower carotid and femoral subclinical atherosclerosis and coronary calcium. Eur J Nutr. 2022, 61, 2697–709. [Google Scholar] [CrossRef] [PubMed]
- Chiva-Blanch, G.; Urpi-Sarda, M.; Llorach, R.; Rotches-Ribalta, M.; Guillén, M.; Casas, R.; et al. Differential effects of polyphenols and alcohol of red wine on the expression of adhesion molecules and inflammatory cytokines related to atherosclerosis: a randomized clinical trial. Am J Clin Nutr. 2011, 95, 326–34. [Google Scholar] [CrossRef]
- Canali, R.; Comitato, R.; Ambra, R.; Virgili, F. Red wine metabolites modulate NF-κB, activator protein-1 and cAMP response element-binding proteins in human endothelial cells. Br J Nutr. 2010, 103, 807–14. [Google Scholar] [CrossRef]
- Nallasamy, P.; Kang, Z.Y.; Sun, X.; Anandh Babu, P.V.; Liu, D.; Jia, Z. Natural compound resveratrol attenuates TNF-alpha-induced vascular dysfunction in mice and human endothelial cells: The involvement of the NF-κB signaling pathway. Int J Mol Sci. 2021, 22, 22. [Google Scholar] [CrossRef] [PubMed]
- Kechagias, S.; Zanjani, S.; Gjellan, S.; Leinhard, O.D.; Kihlberg, J.; Smedby, Ö.; et al. Effects of moderate red wine consumption on liver fat and blood lipids: a prospective randomized study. Ann Med. 2011, 43, 545–54. [Google Scholar] [CrossRef]
- Roth, I.; Casas, R.; Medina-Remón, A.; Lamuela-Raventós, R.M.; Estruch, R. Consumption of aged white wine modulates cardiovascular risk factors via circulating endothelial progenitor cells and inflammatory biomarkers. Clin Nutr. 2019, 38, 1036–44. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.H.; Chen, Y.H.; Tsai, H.Y.; Chen, J.S.; Wu, T.C.; Lin, F.Y.; et al. Intake of Red Wine Increases the Number and Functional Capacity of Circulating Endothelial Progenitor Cells by Enhancing Nitric Oxide Bioavailability. Art Thromb Vasc Biol. 2010, 30, 869–77. [Google Scholar] [CrossRef]
- Tomé-Carneiro, J.; Gonzálvez, M.; Larrosa, M.; Yáñez-Gascón, M.J.; García-Almagro, F.J.; Ruiz-Ros, J.A.; et al. Grape resveratrol increases serum adiponectin and downregulates inflammatory genes in peripheral blood mononuclear cells: A triple-blind, placebo-controlled, one-year clinical trial in patients with stable coronary artery disease. Cardiovasc Drugs Ther. 2013, 27, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Cosmi, F.; Di Giulio, P.; Masson, S.; Finzi, A.; Marfisi, R.M.; Cosmi, D.; et al. Regular Wine Consumption in Chronic Heart Failure: Impact on Outcomes, Quality of Life, and Circulating Biomarkers. Circ Heart Fail. 2015, 8, 428–37. [Google Scholar] [CrossRef]
- Downer, M.K.; Kenfield, S.A.; Stampfer, M.J.; Wilson, K.M.; Dickerman, B.A.; Giovannucci, E.L.; Rimm, E.B.; Wang, M.; Mucci, L.A.; Willett, W.C.; Chan, J.M.; Van Blarigan, E.L. Alcohol Intake and Risk of Lethal Prostate Cancer in the Health Professionals Follow-Up Study. J Clin Oncol. 2019, 37, 1499–1511. [Google Scholar] [CrossRef]
- Crockett, S.D.; Long, M.D.; Dellon, E.S.; Martin, C.F.; Galanko, J.A.; Sandler, R.S. Inverse relationship between moderate alcohol intake and rectal cancer: analysis of the North Carolina Colon Cancer Study. Dis Colon Rectum. 2011, 54, 887–94. [Google Scholar] [CrossRef]
- Liu, Z.; Song, C.; Suo, C.; Fan, H.; Zhang, T.; Jin, L.; Chen, X. Alcohol consumption and hepatocellular carcinoma: novel insights from a prospective cohort study and nonlinear Mendelian randomization analysis. BMC Med. 2022, 20, 413. [Google Scholar] [CrossRef]
- Wang, X.; Jia, M.; Mao, Y.; Jia, Z.; Liu, H.; Yang, G.; Wang, S.; Sun, B.; Zhang, H. Very-light alcohol consumption suppresses breast tumor progression in a mouse model. Food Funct. 2022, 13, 3391–3404. [Google Scholar] [CrossRef]
- Schaefer, S.M.; Kaiser, A.; Behrendt, I.; Eichner, G.; Fasshauer, M. Association of alcohol types, coffee and tea intake with mortality: prospective cohort study of UK Biobank participants. Br J Nutr. 2022, 129, 1–11. [Google Scholar] [CrossRef]
- Turati, F.; Carioli, G.; Bravi, F.; Ferraroni, M.; Serraino, D.; Montella, M.; et al. Mediterranean Diet and Breast Cancer Risk. Nutrients 2018, 10. [Google Scholar] [CrossRef]
- Shufelt, C.; Merz, C.N.; Yang, Y.; Kirschner, J.; Polk, D.; Stanczyk, F.; Paul-Labrador, M.; Braunstein, D. Red versus white wine as a nutritional aromatase inhibitor in premenopausal women: a pilot study. J Womens Health. 2012, 21, 281–4. [Google Scholar] [CrossRef]
- Armstrong, M.J.; Mellinger, J.L.; Trivedi, P.J. Alcohol Consumption in Patients with Non-alcoholic Fatty Liver Disease: Convenient vs. Inconvenient Truths. Am J Gastroenterol. 2018, 113, 1437–1439. [Google Scholar] [CrossRef]
- Zhu, W.; Qin, W.; Zhang, K.; Rottinghaus, G.E.; Chen, Y.C.; Kliethermes, B.; et al. Trans-resveratrol alters mammary promoter hypermethylation in women at increased risk for breast cancer. Nutr Cancer. 2012, 64, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Tresserra-Rimbau, A.; Medina-Remón, A.; Lamuela-Raventós, R.M.; Bulló, M.; Salas-Salvadó, J.; Corella, D.; et al. Moderate red wine consumption is associated with a lower prevalence of the metabolic syndrome in the PREDIMED population. Br J Nutr. 2015, 113, S121–30. [Google Scholar] [CrossRef]
- Larsen, B.A.; Klinedinst, B.S.; Le, S.T.; Pappas, C.; Wolf, T.; Meier, N.F.; Lim, Y.L.; Willette, A.A. Beer, wine, and spirits differentially influence body composition in older white adults-a United Kingdom Biobank study. Obes Sci Pract. 2022, 8, 641–656. [Google Scholar] [CrossRef]
- Gepner, Y.; Golan, R.; Harman-Boehm, I.; Henkin, Y.; Schwarzfuchs, D.; Shelef, I.; Durst, R.; Kovsan, J.; Bolotin, A.; Leitersdorf, E.; Shpitzen, S.; Balag, S.; Shemesh, E.; Witkow, S.; Tangi-Rosental, O.; Chassidim, Y.; Liberty, I.F.; Sarusi, B.; Ben-Avraham, S.; Helander, A.; Ceglarek, U.; Stumvoll, M.; Blüher, M.; Thiery, J.; Rudich, A.; Stampfer, M.J.; Shai, I. Effects of Initiating Moderate Alcohol Intake on Cardiometabolic Risk in Adults With Type 2 Diabetes: A 2-Year Randomized, Controlled Trial. Ann Intern Med. 2015, 163, 569–79. [Google Scholar] [CrossRef] [PubMed]
- Golan, R.; Shai, I.; Gepner, Y.; Harman-Boehm, I.; Schwarzfuchs, D.; Spence, J.D.; Parraga, G.; Buchanan, D.; Witkow, S.; Friger, M.; Liberty, I.F.; Sarusi, B.; Ben-Avraham, S.; Sefarty, D.; Bril, N.; Rein, M.; Cohen, N.; Ceglarek, U.; Thiery, J.; Stumvoll, M.; Blüher, M.; Stampfer, M.J.; Rudich, A.; Henkin, Y. Effect of wine on carotid atherosclerosis in type 2 diabetes: a 2-year randomized controlled trial. Eur J Clin Nutr. 2018, 72, 871–878. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Wang, X.; Li, X.; Heianza, Y.; Qi, L. Moderate alcohol drinking with meals is related to lower incidence of type 2 diabetes. Am J Clin Nutr. 2022, 116, 1507–14. [Google Scholar] [CrossRef] [PubMed]
- Chiva-Blanch, G.; Urpi-Sarda, M.; Ros, E.; Valderas-Martinez, P.; Casas, R.; Arranz, S.; et al. Effects of red wine polyphenols and alcohol on glucose metabolism and the lipid profile: a randomized clinical trial. Clin Nutr. 2013, 32, 200–6. [Google Scholar] [CrossRef] [PubMed]
- Ismail, M.; Stagling, S.; Lundberg, A.; Nystrom, F.H. A cross-over study of postprandial effects from moist snuff and red wine on metabolic rate, appetite-related hormones and glucose. Drug Alcohol Depend. 2022, 236, 109479. [Google Scholar] [CrossRef] [PubMed]
- Sattarinezhad, A.; Roozbeh, J.; Shirazi Yeganeh, B.; Omrani, G.R.; Shams, M. Resveratrol reduces albuminuria in diabetic nephropathy: A randomized double-blind placebo-controlled clinical trial. Diabetes Metab. 2019, 45, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Liu, Y.; Ren, G.; Yin, L.; Liang, X.; Geng, T.; Dang, H.; An, R. Resveratrol limits diabetes-associated cognitive decline in rats by preventing oxidative stress and inflammation and modulating hippocampal structural synaptic plasticity. Brain Res. 2016, 1650, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Fischer, K.; Melo van Lent, D.; Wolfsgruber, S.; Weinhold, L.; Kleineidam, L.; Bickel, H.; Scherer, M.; Eisele, M.; van den Bussche, H.; Wiese, B.; König, H.H.; Weyerer, S.; Pentzek, M.; Röhr, S.; Maier, W.; Jessen, F.; Schmid, M.; Riedel-Heller, S.G.; Wagner, M. Prospective Associations between Single Foods, Alzheimer's Dementia and Memory Decline in the Elderly. Nutrients. 2018, 10, 852. [Google Scholar] [CrossRef] [PubMed]
- Mendes, D.; Oliveira, M.M.; Moreira, P.I.; Coutinho, J.; Nunes, F.M.; Pereira, D.M.; Valentão, P.; Andrade, P.B.; Videira, R.A. Beneficial effects of white wine polyphenols-enriched diet on Alzheimer's disease-like pathology. J Nutr Biochem. 2018, 55, 165–177. [Google Scholar] [CrossRef]
- Xu, W.; Wang, H.; Wan, Y.; Tan, C.; Li, J.; Tan, L.; Yu, J.T. Alcohol consumption and dementia risk: a dose-response meta-analysis of prospective studies. Eur J Epidemiol. 2017, 32, 31–42. [Google Scholar] [CrossRef]
- Ho, L.; Ferruzzi, M.G.; Janle, E.M.; Wang, J.; Gong, B.; Chen, T.Y.; Lobo, J.; Cooper, B.; Wu, Q.L.; Talcott, S.T.; Percival, S.S.; Simon, J.E.; Pasinetti, G.M. Identification of brain-targeted bioactive dietary quercetin-3-O-glucuronide as a novel intervention for Alzheimer's disease. FASEB J. 2013, 27, 769–81. [Google Scholar] [CrossRef]
- Valls-Pedret, C.; Lamuela-Raventós, R.M.; Medina-Remón, A.; Quintana, M.; Corella, D.; Pintó, X.; Martínez-González, M.Á.; Estruch, R.; Ros, E. Polyphenol-rich foods in the Mediterranean diet are associated with better cognitive function in elderly subjects at high cardiovascular risk. J Alzheimers Dis. 2012, 29, 773–82. [Google Scholar] [CrossRef]
- Smyth, A.; O'Donnell, M.; Rangarajan, S.; Hankey, G.J.; Oveisgharan, S.; Canavan, M.; McDermott, C.; Xavier, D.; Zhang, H.; Damasceno, A.; Avezum, A.; Pogosova, N.; Oguz, A.; Ryglewicz, D.; Iversen, H.K.; Lanas, F.; Rosengren, A.; Yusuf, S.; Langhorne, P. INTERSTROKE Investigators. Alcohol Intake as a Risk Factor for Acute Stroke: The INTERSTROKE Study. Neurology. 2023, 100, e142–e153. [Google Scholar] [CrossRef] [PubMed]
- Kaluza, J.; Harris, H.R.; Linden, A.; Wolk, A. Alcohol Consumption and Risk of Chronic Obstructive Pulmonary Disease: A Prospective Cohort Study of Men. Am J Epidemiol. 2019, 188, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Godos, J.; Castellano, S.; Ray, S.; Grosso, G.; Galvano, F. ; Dietary Polyphenol Intake and Depression: Results from the Mediterranean Healthy Eating, Lifestyle and Aging (MEAL) Study. Molecules. 2018, 23, 999. [Google Scholar] [CrossRef] [PubMed]
- Barbería-Latasa, M.; Bes-Rastrollo, M.; Pérez-Araluce, R.; Martínez-González, M.Á.; Gea, A. Mediterranean Alcohol-Drinking Patterns and All-Cause Mortality in Women More Than 55 Years Old and Men More Than 50 Years Old in the "Seguimiento Universidad de Navarra" (SUN) Cohort. Nutrients. 2022, 14, 5310. [Google Scholar] [CrossRef] [PubMed]
- Carballo-Casla, A.; Ortolá, R.; García-Esquinas, E.; Oliveira, A.; Sotos-Prieto, M.; Lopes, C.; Lopez-Garcia, E.; Rodríguez-Artalejo, F. The Southern European Atlantic Diet and all-cause mortality in older adults. BMC Med. 2021, 19, 36. [Google Scholar] [CrossRef]
- Grønbaek, M.; Becker, U.; Johansen, D.; Gottschau, A.; Schnohr, P.; Hein, H.O.; et al. Type of alcohol consumed and mortality from all causes, coronary heart disease, and cancer. Ann Intern Med. 2000, 133, 411–9. [Google Scholar] [CrossRef] [PubMed]
- Noguer, M.A.; Cerezo, A.B.; Donoso Navarro, E.; Garcia-Parrilla, M.C. Intake of alcohol-free red wine modulates antioxidant enzyme activities in a human intervention study. Pharmacol Res. 2012, 65, 609–14. [Google Scholar] [CrossRef]
- Giovannucci, E.; Stampfer, M.J.; Colditz, G.A.; Manson, J.E.; Rosner, B.A.; Longnecker, M.P.; Speizer, F.E.; Willett, W.C. ; Recall and selection bias in reporting past alcohol consumption among breast cancer cases. Cancer Causes Control. 1993, 4, 441–8. [Google Scholar] [CrossRef]
- Wu, S.; Li, W.Q.; Qureshi, A.A.; Cho, E. Alcohol consumption and risk of cutaneous basal cell carcinoma in women and men: 3 prospective cohort studies. Am J Clin Nutr. 2015, 102, 1158–66. [Google Scholar] [CrossRef]
- Papadimitriou, N.; Bouras, E.; van den Brandt, P.A.; Muller, D.C.; Papadopoulou, A.; Heath, A.K.; et al. A Prospective Diet-Wide Association Study for Risk of Colorectal Cancer in EPIC. Clin Gastroenterol Hepatol. 2022, 20, 864–873.e13. [Google Scholar] [CrossRef] [PubMed]
- Mahamat-Saleh, Y.; Al-Rahmoun, M.; Severi, G.; Ghiasvand, R.; Veierod, M.B.; Caini, S.; Palli, D.; Botteri, E,; et al. Baseline and lifetime alcohol consumption and risk of skin cancer in the European Prospective Investigation into Cancer and Nutrition cohort (EPIC). Int J Cancer. 2023, 152, 348–362. [Google Scholar] [CrossRef] [PubMed]
- Heath, A.K.; Muller, D.C.; van den Brandt, P.A.; Papadimitriou, N.; Critselis, E.; et al. Nutrient-wide association study of 92 foods and nutrients and breast cancer risk. Breast Cancer Res. 2020, 22, 5. [Google Scholar] [CrossRef] [PubMed]
- Naudin, S.; Li, K.; Jaouen, T.; Assi, N.; Kyrø, C.; Tjønneland, A.; Overvad, K.; Boutron-Ruault, M.C.; et al. Lifetime and baseline alcohol intakes and risk of pancreatic cancer in the European Prospective Investigation into Cancer and Nutrition study. Int J Cancer. 2018, 143, 801–812. [Google Scholar] [CrossRef] [PubMed]
- Seidenberg, A.B.; Wiseman, K.P.; Klein, W.M.P. Do Beliefs about Alcohol and Cancer Risk Vary by Alcoholic Beverage Type and Heart Disease Risk Beliefs? Cancer Epidemiol Biomarkers Prev. 2023, 32, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Hay, J.L.; Kiviniemi, M.T.; Orom, H.; Waters, E.A. Moving beyond the "Health Halo" of Alcohol: What Will it Take to Achieve Population Awareness of the Cancer Risks of Alcohol? Cancer Epidemiol Biomarkers Prev. 2023, 32, 9–11. [Google Scholar] [CrossRef] [PubMed]
- Duell, E.J.; Travier, N.; Lujan-Barroso, L.; Clavel-Chapelon, F.; Boutron-Ruault, M.C.; Morois, S.; et al. Alcohol consumption and gastric cancer risk in the European Prospective Investigation into Cancer and Nutrition (EPIC) cohort. Am J Clin Nutr. 2011, 94, 1266–75. [Google Scholar] [CrossRef] [PubMed]
- Cote, D.J.; Smith, T.R.; Kaiser, U.B.; Laws, E.R. Jr.; Stampfer, M.J. Alcohol intake and risk of pituitary adenoma. Cancer Causes Control. 2022, 33, 353–361. [Google Scholar] [CrossRef]
- Bondonno, N.P.; Liu, Y.L.; Zheng, Y.; Ivey, K.; Willett, W.C.; Stampfer, M.J.; Rimm, E.B.; Cassidy, A. Change in habitual intakes of flavonoid-rich foods and mortality in US males and females. BMC Med. 2023, 21, 181. [Google Scholar] [CrossRef]
- Duan, J.; Guo, H.; Fang, Y.; Zhou, G. The mechanisms of wine phenolic compounds for preclinical anticancer therapeutics. Food Nutr Res. 2021, 65. [Google Scholar] [CrossRef]
- O'Keefe, J.H.; Bybee, K.A.; Lavie, C.J. Alcohol and cardiovascular health: the razor-sharp double-edged sword. J Am Coll Cardiol. 2007, 50, 1009–14. [Google Scholar] [CrossRef] [PubMed]
- O'Keefe, E.L.; DiNicolantonio, J.J.; O'Keefe, J.H.; Lavie, C.J. Alcohol and CV Health: Jekyll and Hyde J-Curves. Prog Cardiov Dis. 2018, 61, 68–75. [Google Scholar] [CrossRef]
- Poli, A. Is drinking wine in moderation good for health or not? Eur Heart J Suppl. 2022, 24, I119–I122. [Google Scholar] [CrossRef] [PubMed]
- Rifler, J.P. Is a Meal without Wine Good for Health? Diseases. 2018, 6, 105. [Google Scholar] [CrossRef]
- Gerardi, G.; Cavia-Saiz, M.; Muñiz, P. From winery by-product to healthy product: bioavailability, redox signaling and oxidative stress modulation by wine pomace product. Crit Rev Food Sci Nutr. 2022, 62, 7427–7448. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wen, J.Y.; Deng, Z.X.; Pan, X.Q.; Xie, X.F.; Peng, C. Effective utilization of food wastes: Bioactivity of grape seed extraction and its application in food industry. J. Funct. Foods 2020, 73, 104113. [Google Scholar] [CrossRef]
- Yang, C.; Han, Y.; Tian, X.; Sajid, M.; Mehmood, S.; Wang, H.; Li, H. Phenolic composition of grape pomace and its metabolism. Crit Rev Food Sci Nutr. 2022, 17, 1–17. [Google Scholar] [CrossRef]
- Spinei, M.; Oroian, M. ;. The Potential of Grape Pomace Varieties as a Dietary Source of Pectic Substances. Foods 2021, 10, 867. [Google Scholar] [CrossRef]
- Panzella, L.; Moccia, F.; Nasti, R.; Marzorati, S.; Verotta, L.; and Napolitano, A. Bioactive Phenolic Compounds From Agri-Food Wastes: An Update on Green and Sustainable Extraction Methodologies. Front. Nutr. 2020, 7, 60. [Google Scholar] [CrossRef]
- Ilyas, T.; Chowdhary, P.; Chaurasia, D.; Gnansounou, E.; Pandey, A.; Chaturvedi, P. Sustainable green processing of grape pomace for the production of value-added products: An overview. Envir Tech Innov 2021, 23, 101592. [Google Scholar] [CrossRef]
- Chowdhary, P.; Gupta, A.; Gnansounou, E.; Pandey, A.; Chaturvedi, P. Current trends and possibilities for exploitation of Grape pomace as a potential source for value addition. Envir Pollution 2021, 278. [Google Scholar] [CrossRef]
- Drevelegka, I.; Goula, A.M. Recovery of grape pomace phenolic compounds through optimized extraction and adsorption processes. Chem Engin Proc - Process Intens 2020, 149, 107845. [Google Scholar] [CrossRef]
- Barba, F.; Zhu, Z.; Koubaa, M.; Sant’Ana, A.; Orlien, V. Green alternative methods for the extraction of antioxidant bioactive compounds from winery wastes and by-products: A review. Trends Food Sci Tech 2016, 49, 96–109. [Google Scholar] [CrossRef]
- Jokić, S.; Velić, D.; Bilić, M.; Bucić-Kojić, A.; Planinić, M.; Tomas, S. Modelling of the Process of Solid-Liquid Extraction of Total Polyphenols from Soybeans. Czech J. Food Sci. 2010, 28, 7. [Google Scholar] [CrossRef]
- Yang, C.L.; Shang, K.; Lin, C.C.; Wang, C.; Shi, X.Q.; Wang, H.; Li, H. Processing technologies, phytochemical constituents, and biological activities of grape seed oil (gso): A review. Trends Food Sci. Technol. 2021, 116, 1074–1083. [Google Scholar] [CrossRef]
- Herrero, M.; Mendiola, J.A.; Cifuentes, A.; Ibáñez, E. Supercritical fluid extraction: Recent advances and applications. J Chromatogr A, Extraction Techniques 2010, 1217, 2495–2511. [Google Scholar] [CrossRef] [PubMed]
- Pinelo, M.; Ruiz-Rodríguez, A.; Sineiro, J.; Señoráns, F.J.; Reglero, G.; Núñez, M.J. Supercritical fluid and solid–liquid extraction of phenolic antioxidants from grape pomace: a comparative study. Eur Food Res Technol 2007, 226, 199–205. [Google Scholar] [CrossRef]
- Jeong, J.; Jung, H.; Lee, S.; Lee, H.; Hwang, K.; Kim, T. Anti-oxidant, anti-proliferative and anti-inflammatory activities of the extracts from black raspberry fruits and wine. Food Chem 2010, 123, 338–344. [Google Scholar] [CrossRef]
- Casas, L.; Mantell, C.; Rodríguez, M.; Ossa, E.J.M. de la; Roldán, A.; Ory, I.D.; Caro, I.; Blandino, A. Extraction of resveratrol from the pomace of Palomino fino grapes by supercritical carbon dioxide. J Food Engin 2010, 96, 304–308. [Google Scholar] [CrossRef]
- Liazid, A.; Guerrero, R.F.; Cantos, E.; Palma, M.; Barroso, C.G. Microwave assisted extraction of anthocyanins from grape skins. Food Chem 2011, 124, 1238–1243. [Google Scholar] [CrossRef]
- Yu, H.B.; Ding, L.F.; Wang, Z.; Shi, L.X. Study on Extraction of Polyphenol from Grape Peel Microwave-Assisted Activity. Adv Mater Res 2014, 864–867, 520–525. [Google Scholar] [CrossRef]
- Al Bittar, S.; Périno-Issartier, S.; Dangles, O.; Chemat, F. An innovative grape juice enriched in polyphenols by microwave-assisted extraction. Food Chem 2013, 141, 3268–3272. [Google Scholar] [CrossRef] [PubMed]
- Da Porto, C.; Porretto, E.; Decorti, D. Comparison of ultrasound-assisted extraction with conventional extraction methods of oil and polyphenols from grape (Vitis vinifera L.) seeds. Ultrasonics Sonochem 2013, 20, 1076–1080. [Google Scholar] [CrossRef] [PubMed]
- Drosou, C.; Kyriakopoulou, K.; Bimpilas, A.; Tsimogiannis, D.; Krokida, M. A comparative study on different extraction techniques to recover red grape pomace polyphenols from vinification byproducts. Industrial Crops and Products, Advances in Industrial Crops and Products Worldwide: AAIC 2014 international conference 2015, 75, 141–149. [Google Scholar] [CrossRef]
- Wang, J.; Wang, K.; Wang, Y.; Lin, S.; Zhao, P.; Jones, G. A novel application of pulsed electric field (PEF) processing for improving glutathione (GSH) antioxidant activity. Food Chem 2014, 161, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Jeyamkondan, S.; Jayas, D.S.; Holley, R.A. Pulsed electric field processing of foods: a review. J Food Prot 1999, 62, 1088–1096. [Google Scholar] [CrossRef] [PubMed]
- Brianceau, S.; Turk, M.; Vitrac, X.; Vorobiev, E. Combined densification and pulsed electric field treatment for selective polyphenols recovery from fermented grape pomace. Innov Food Sci Emerg Tech, App Food Process 2015, 29, 2–8. [Google Scholar] [CrossRef]
- Wan, J.; Coventry, J.; Swiergon, P.; Sanguansri, P.; Versteeg, C. Advances in innovative processing technologies for microbial inactivation and enhancement of food safety – pulsed electric field and low-temperature plasma. Trends Food Sci Tech, Natural Safe Foods 2009, 20, 414–424. [Google Scholar] [CrossRef]
- Barba, F.J.; Brianceau, S.; Turk, M.; Boussetta, N.; Vorobiev, E. Effect of Alternative Physical Treatments (Ultrasounds, Pulsed Electric Fields, and High-Voltage Electrical Discharges) on Selective Recovery of Bio-compounds from Fermented Grape Pomace. Food Bioprocess Technol 2015, 8, 1139–1148. [Google Scholar] [CrossRef]
- Corrales, M.; Toepfl, S.; Butz, P.; Knorr, D.; Tauscher, B. Extraction of anthocyanins from grape by-products assisted by ultrasonics, high hydrostatic pressure or pulsed electric fields: A comparison. Innov Food Sci Emerg Tech 2008, 9, 85–91. [Google Scholar] [CrossRef]
- Dordoni, R.; Duserm Garrido, G.; Marinoni, L.; Torri, L.; Piochi, M.; Spigno, G. Enrichment of Whole Wheat Cocoa Biscuits with Encapsulated Grape Skin Extract. Int J Food Sci 2019, 2019, 9161840. [Google Scholar] [CrossRef]
- Pasqualone, A.; Bianco, A.M.; Paradiso, V.M. Production trials to improve the nutritional quality of biscuits and to enrich them with natural anthocyanins. CyTA – J Food 2013, 11, 301–308. [Google Scholar] [CrossRef]
- Mildner-Szkudlarz, S.; Bajerska, J.; Zawirska-Wojtasiak, R.; Górecka, D. White grape pomace as a source of dietary fibre and polyphenols and its effect on physical and nutraceutical characteristics of wheat biscuits. J. Sci. Food Agric. 2013, 93, 389–395. [Google Scholar] [CrossRef]
- Acun, S.; Gül, H. Effects of grape pomace and grape seed flours on cookie quality. Qual Assur Saf Crops Foods 2014, 6, 81–88. [Google Scholar] [CrossRef]
- Bender, A.B.B.; Speroni, C.S.; Salvador, P.R.; Loureiro, B.B.; Lovatto, N.M.; Goulart, F.R.; Lovatto, M.T.; Miranda, M.Z.; Silva, L.P.; Penna, N.G. Grape Pomace Skins and the Effects of Its Inclusion in the Technological Properties of Muffins. J Culinary Sci Tech 2017, 15, 143–157. [Google Scholar] [CrossRef]
- Šporin, M.; Avbelj, M.; Kovač, B.; Možina, S.S. Quality characteristics of wheat flour dough and bread containing grape pomace flour. Food Sci. Technol. Int. 2018, 24, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Tolve, R.; Simonato, B.; Rainero, G.; Bianchi, F.; Rizzi, C.; Cervini, M.; Giuberti, G. Wheat Bread Fortification by Grape Pomace Powder: Nutritional, Technological, Antioxidant, and Sensory Properties. Foods 2021, 10, 75. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, F.; Lomuscio, E.; Rizzi, C.; Simonato, B. Predicted Shelf-Life, Thermodynamic Study and Antioxidant Capacity of Breadsticks Fortified with Grape Pomace Powders. Foods 2021, 10, 2815. [Google Scholar] [CrossRef] [PubMed]
- Nowak, J.Z. Oxidative stress, polyunsaturated fatty acids-derived oxidation products and bisretinoids as potential inducers of CNS diseases: focus on age-related macular degeneration. Pharmacol Rep 2013, 65, 288–304. [Google Scholar] [CrossRef] [PubMed]
- Hoye, C.; Ross, C. Total Phenolic Content, Consumer Acceptance, and Instrumental Analysis of Bread Made with Grape Seed Flour. J Food Sci 2011, 76, S428–S436. [Google Scholar]
- Lavelli, V.; Kerr, W.L.; García-Lomillo, J.; González-SanJosé, M.L. Applications of Recovered Bioactive Compounds in Food Products, In Handbook of Grape Processing By-Products, Galanakis, C.M., Ed.; Academic Press, 2017, Volume 10, pp. 233–266.
- Tolve, R.; Pasini, G.; Vignale, F.; Favati, F.; Simonato, B. Effect of Grape Pomace Addition on the Technological, Sensory, and Nutritional Properties of Durum Wheat Pasta. Foods 2020, 9, 354. [Google Scholar] [CrossRef] [PubMed]
- Sant’Anna, V.; Christiano, F.D.P.; Marczak, L.D.F.; Tessaro, I.C.; Thys, R.C.S. The effect of the incorporation of grape marc powder in fettuccini pasta properties. LWT - Food Sci Techn 2014, 58, 497–501. [Google Scholar] [CrossRef]
- Lavelli, V.; Torri, L.; Zeppa, G.; Fiori, L.; Spigno, G. Recovery Of Winemaking By-Products For Innovative Food Applications No. 4 (): Ital J Food Sci. 2016, 28. [Google Scholar]
- Balli, D.; Cecchi, L.; Innocenti, M.; Bellumori, M.; Mulinacci, N. Food by-products valorisation: Grape pomace and olive pomace (pâté) as sources of phenolic compounds and fiber for enrichment of tagliatelle pasta. Food Chem 2021, 355, 129642. [Google Scholar] [CrossRef]
- Codină, G.G.; Zaharia, D.; Stroe, S.-G.; Ropciuc, S. Influence of calcium ions addition from gluconate and lactate salts on refined wheat flour dough rheological properties. CyTA - J Food 2018, 16, 884–891. [Google Scholar] [CrossRef]
- Mironeasa, S.; Codina, G.; Mironeasa, C. The effects of wheat flour substitution with grape seed flour on the rheological parameters of the dough assessed by Mixolab. J Texture Studies 2012, 43. [Google Scholar] [CrossRef]
- Rashwan, A.K.; Osman, A.I.; Chen, W. Natural nutraceuticals for enhancing yogurt properties: a review. Environ Chem Lett 2023, 21, 1907–1931. [Google Scholar] [CrossRef]
- Barakat, H.; Y. Hassan, M. Chemical, Nutritional, Rheological, and Organoleptical Characterizations of Stirred Pumpkin-Yoghurt. Food Nutr Sci 2017, 8, 746–759. [Google Scholar]
- O’Connell, J.E.; Fox, P.F. Significance and applications of phenolic compounds in the production and quality of milk and dairy products: a review. Int Dairy J 2001, 11, 103–120. [Google Scholar] [CrossRef]
- Demirkol, M.; Tarakci, Z. Effect of grape (Vitis labrusca L.) pomace dried by different methods on physicochemical, microbiological and bioactive properties of yoghurt. LWT Food Sci Technol 2018, 97, 770–777. [Google Scholar] [CrossRef]
- Tseng, A. ; Zhao. Y.Wine grape pomace as antioxidant dietary fibre for enhancing nutritional value and improving storability of yogurt and salad dressing Food Chem., 2013, 138, 356–365. [Google Scholar]
- Silva, F.A.; et al. Incorporation of phenolic-rich ingredients from integral valorization of Isabel grape improves the nutritional, functional and sensory characteristics of probiotic goat milk yogurt. Food Chem 2022, 369, 130957. [Google Scholar] [CrossRef] [PubMed]
- Marchiani, R.; Bertolino, M.; Ghirardello, D.; McSweeney, P.L.H.; Zeppa, G. Physicochemical and nutritional qualities of grape pomace powder-fortified semi-hard cheeses. J Food Sci Technol 2016, 53, 1585–1596. [Google Scholar] [CrossRef]
- Tami, S.H.; et al. Buffalo stirred yoghurt fortified with grape seed extract: new insights into its functional properties. Food Biosci 2022, 47, 101752. [Google Scholar] [CrossRef]
- Gaglio, R.; Restivo, I.; Barbera, M.; Barbaccia, P.; Ponte, M.; Tesoriere, L.; Bonanno, A.; Attanzio, A.; Di Grigoli, A.; Francesca, N.; Moschetti, G.; Settanni, L. Effect on the Antioxidant, Lipoperoxyl Radical Scavenger Capacity, Nutritional, Sensory and Microbiological Traits of an Ovine Stretched Cheese Produced with Grape Pomace Powder Addition. Antioxidants 2021, 10, 306. [Google Scholar] [CrossRef]
- Chouchouli, V.; Kalogeropoulos, N.; Konteles, S.J.; Karvela, E.; Makris, D.P.; Karathanos, V.T. Fortification of yoghurts with grape (Vitis vinifera) seed extracts. LWT - Food Sci Techn 2013, 53, 522–529. [Google Scholar] [CrossRef]
- Marchiani, R.; Bertolino, M.; Belviso, S.; Giordano, M.; Ghirardello, D.; Torri, L.; Piochi, M.; Zeppa, G. Yogurt Enrichment with Grape Pomace: Effect of Grape Cultivar on Physicochemical, Microbiological and Sensory Properties. J Food Qual 2016, 3977–89. [Google Scholar] [CrossRef]
- Karaaslan, M.; Ozden, M.; Vardin, H.; Turkoglu, H. Phenolic fortification of yogurt using grape and callus extracts. LWT - Food Sci Techn 2011, 44, 1065–1072. [Google Scholar] [CrossRef]
- Zhou, Y.J.; Wang, Q.Y.; Wang, S.J. Effects of rosemary extract, grape seed extract and green tea polyphenol on the formation of n-Nitrosamines and quality of western-Style smoked sausage. J. Food Process. Preserv. 2020, 44, e14459. [Google Scholar] [CrossRef]
- García-Lomillo, J.; González-SanJosé, M.L. Applications of Wine Pomace in the Food Industry: Approaches and Functions. Compreh Rev Food Sci Food Safety 2017, 16, 3–22. [Google Scholar] [CrossRef]
- García-Lomillo, J.; Gonzalez-SanJose, M.L.; Del Pino-García, R.; Ortega-Heras, M.; Muñiz-Rodríguez, P. Antioxidant effect of seasonings derived from wine pomace on lipid oxidation in refrigerated and frozen beef patties. LWT 2017, 77, 85–91. [Google Scholar] [CrossRef]
- Ryu, K.; Shim, K.; Shin, D. Effect of Grape Pomace Powder Addition on TBARS and Color of Cooked Pork Sausages during Storage. Kor J Food Sci An Res 2014, 34. [Google Scholar] [CrossRef] [PubMed]
- Riazi, F.; Zeynali, F.; Hoseini, E.; Behmadi, H. Effect of Dry Red Grape Pomace as a Nitrite Substitute on the Microbiological and Physicochemical Properties and Residual Nitrite of Dry-cured Sausage. Nutr Food Sci Res 2016, 3, 37–44. [Google Scholar] [CrossRef]
- Garrido, J.; Borges, F. Wine and grape polyphenols — A chemical perspective. Food Res Intern 2013, 54, 1844–1858. [Google Scholar] [CrossRef]
- Bobko, M.; Haščí k, P.; Kročko, M.; Trembecká, L.; Mendelova, A.; Tkáčová, J.; Czako, P.; Tóth, T. Effect of grape seed extract on quality of raw-cooked meat products. Potr Slovak J Food Sci 2017, 11, 517–521. [Google Scholar] [CrossRef] [PubMed]
- Guerra-Rivas, C.; Vieira, C.; Rubio, B.; Martínez, B.; Gallardo, B.; Mantecón, A.R.; Lavín, P.; Manso. T. Effects of grape pomace in growing lamb diets compared with vitamin E and grape seed extract on meat shelf life. Meat Sci. 2016, 116, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Selani, M.M.; Contreras-Castillo, C.J.; Shirahigue, L.D.; Gallo, C.R.; Plata-Oviedo, M.; Montes-Villanueva, N.D. Wine industry residues extracts as natural antioxidants in raw and cooked chicken meat during frozen storage. Meat Sci 2011, 88, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Shirahigue, L.D.; Plata-Oviedo, M.; De Alencar, S.M.; D’Arce, M.A.B.R.; De Souza Vieira, T.M.F.; Oldoni, T.L.C.; Contreras-Castillo, C.J. Wine industry residue as antioxidant in cooked chicken meat. Int J Food Sci Techn 2010, 45, 863–870. [Google Scholar] [CrossRef]
- Sáyago-Ayerdi, S.G.; Brenes, A.; Goñi. I. Effect of grape antioxidante dietary fibre on the lípido oxidation of raw and cooked Chicken hamburgers. LWT- Food Sci. Technol. 2009, 42, 971–976. [Google Scholar] [CrossRef]
- Sánchez-Alonso, I.; Jiménez-Escrig, A.; Saura-Calixto, F.; Borderías. A.J. Effect of grape antioxidant dietary fibre on the prevention of lipid oxidation in miced fish: Evaluation by different methodologies. Food Chem., 2007, 101, 372–378. [Google Scholar] [CrossRef]
- Özvural, E.; Vural, H. Grape seed flour is a viable ingredient to improve the nutritional profile and reduce lipid oxidation of frankfurters. Meat Sci 2011, 88, 179–83. [Google Scholar] [CrossRef]
- Choi, Y.; Choi, J.; Han, D.; Kim, H.; Lee, M.; Kim, H.; Lee, J.; Chung, H.; Kim. C. Optimization of replacing pork back fat with grape seed oil and rice bran fibre for reduced-fat meat emulsion systems. Meat Sci. 2010, 84, 212–218. [Google Scholar] [CrossRef]
- Özalp Özen, B.; Eren, M.; Pala, A.; Özmen, İ.; Soyer, A. Effect of plant extracts on lipid oxidation during frozen storage of minced fish muscle. Intern J Food Sci Techn 2011, 46, 724–731. [Google Scholar] [CrossRef]
- Cilli, L.; Contini, L.; Sinnecker, P.; Lopes, P.; Andréo, M.; Neiva, C.; Nascimento, M.; Yoshida, C.; Venturini, A. Effects of grape pomace flour on quality parameters of salmon burger. J Food Process Preserv 2019, 44. [Google Scholar] [CrossRef]
- Pazos, M.; Gallardo, J.M.; Torres, J.L.; Medina. I. Activity of grape polyphenols as inhibitors of the oxidation of fish lipids and frozen fish muscle. Food Chem. 2005, 92, 547–557. [Google Scholar] [CrossRef]
- Sánchez-Alonso, I.; Jiménez-Escrig, A.; Saura-Calixto, F.; Borderías. A.J. Antioxidant protection of white grape pomace on restructured fish products during frozen storage. LWT Food Sci. Technol. 2008, 41, 42–50. [Google Scholar] [CrossRef]
- Sánchez-Alonso, I.; Borderías, J.; Larsson, K.; Undeland. I. Inhibition of hemoglobin-mediated oxidation of regular and lipid-fortified washed cod mince by a white grape dietary fibre. J. Agric. Food Chem. 2007, 55, 5299–5305. [Google Scholar] [CrossRef] [PubMed]
- Urquiaga, I.; Troncoso, D.; Mackenna, M.J.; Urzúa, C.; Pérez, D.; Dicenta, S.; de la Cerda, P.M.; Amigo, L.; Carreño, J.C.; Echeverría, G.; Rigotti, A. The Consumption of Beef Burgers Prepared with Wine Grape Pomace Flour Improves Fasting Glucose, Plasma Antioxidant Levels, and Oxidative Damage Markers in Humans: A Controlled Trial. Nutrients. 2018, 10, 1388. [Google Scholar] [CrossRef] [PubMed]
- Lavelli, V.; Sri Harsha, P.S.C.; Torri, L.; Zeppa, G. Use of winemaking by-products as an ingredient for tomato puree: The effect of particle size on product quality. Food Chem 2014, 152, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Phan, M.A.T.; Bucknall, M.P.; Arcot, J. Interferences of anthocyanins with the uptake of lycopene in Caco-2 cells, and their interactive effects on anti-oxidation and anti-inflammation in vitro and ex vivo. Food Chem 2019, 276, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Dordoni, R.; Cantaboni, S.; Spigno, G. Walnut paste: oxidative stability and effect of grape skin extract addition. Heliyon 2019, 5, e02506. [Google Scholar] [CrossRef]
- Friedman, M. Chemistry, Biochemistry, and Safety of Acrylamide. A Review. J. Agric. Food Chem. 2003, 51, 4504–4526. [Google Scholar] [CrossRef]
- Xu, C.; Yagiz, Y.; Marshall, S.; Li, Z.; Simonne, A.; Lu, J.; Marshall, M.R. Application of muscadine grape (Vitis rotundifolia Michx.) pomace extract to reduce carcinogenic acrylamide. Food Chem 2015, 182, 200–208. [Google Scholar] [CrossRef]
- Teng, J.; Hu, X.; Tao, N.; Wang, M. Impact and inhibitory mechanism of phenolic compounds on the formation of toxic Maillard reaction products in food. Front. Agr. Sci. Eng. 2018, 5, 321. [Google Scholar] [CrossRef]
- Rózek, A.; Achaerandio, I.; Güell, C.; López, F.; Ferrando. M. Use of commercial grape phenolic extracts to supplement solid foodstuff. LWT Food Sci. Technol. 2010, 43, 623–631. [Google Scholar] [CrossRef]
- Guerrero, R.L.F.; Smith, P.; Bindon. K.A. Application of insoluble fibres in the fining of wine phenolics. J. Agric. Food Chem. 2013, 61, 4424–4432. [Google Scholar] [CrossRef]
- Shinagawa, F.B.; de Santana, F.C.; Torres, L.R.O.; Mancini, J. Grape seed oil: A potential functional food? Food Sci. Technol. 2015, 35, 399–406. [Google Scholar] [CrossRef]
- Auger, C.; Gérain, P.; Laurent-Bichon, F.; Portet, K.; Bornet, A.; Caporiccio, B.; Rouanet. J.M. Phenolics from commercialized grape extracts prevent early atherosclerotic lesions in hamsters by mechanisms other than antioxidant effect. J. Agric. Food Chem. 2004, 52, 5297–5302. [Google Scholar] [CrossRef]
- Tournour, H.H.; Segundo, M.A.; Magalhães, L.M.; Barreiros, L.; Queiroz, J.; Cunha. L.M. Valorization of grape pomace: Extraction of bioactive phenolics with antioxidant properties. Ind Crops Prod, 2015, 74, 397–406. [Google Scholar] [CrossRef]
- Rodriguez-Rodriguez, R.; Justo, M.L.; Claro, C.M.; Vila, E.; Parrado, J.; Herrera, M.D.; de Sotomayor, M.A. . Endothelium-dependent vasodilator and antioxidant properties of a novel enzymatic extract of grape pomace from wine industrial waste. Food Chem. 2012, 135, 1044–1051. [Google Scholar] [CrossRef]
- Scola, G.; Conte, D.; Spada, P.W.D.-S.; Dani, C.; Vanderlinde, R.; Funchal, C.; Salvador, M. Flavan-3-ol Compounds from Wine Wastes with in Vitro and in Vivo Antioxidant Activity. Nutrients 2012, 2, 1048–1059. [Google Scholar] [CrossRef]
- Ed Nignpense, B.; Chinkwo, K.A.; Blanchard, C.L.; Santhakumar, A.B. Polyphenols: Modulators of Platelet Function and Platelet Microparticle Generation? Int J Mol Sci 2019, 21, 146. [Google Scholar] [CrossRef]
- de Lange, D.W.; van Golde, P.H.; Scholman, W.L.G.; Kraaijenhagen, R.J.; Akkerman, J.W.N.; Van De Wiel. A. Red wine and red wine polyphenolic compounds but not alcohol inhibit ADP-induced platelet aggregation. Eur J Inter Med 2003, 14, 361–366. [Google Scholar] [CrossRef]
- Olas, B.; Wachowicz, B.; Stochmal, A.; Oleszek, W. The polyphenol-rich extract from grape seeds inhibits platelet signaling pathways triggered by both proteolytic and non-proteolytic agonists. Platelets 2012, 23, 282–289. [Google Scholar] [CrossRef]
- Bijak, M.; Sut, A.; Kosiorek, A.; Saluk-Bijak, J.; Golanski, J. Dual anticoagulant/antiplatelet activity of polyphenolic grape seeds extract. Nutrients 2019, 11, 93. [Google Scholar] [CrossRef]
- Shanmuganayagam, D.; Beahm, M.R.; Osman, H.E.; Krueger, C.G.; Reed, J.D.; Folts, J.D. Grape seed and grape skin extracts elicit a greater antiplatelet effect when used in combination than when used individually in dogs and humans. J Nutr 2002, 132, 3592–3598. [Google Scholar] [CrossRef]
- Moschona, A.; Liakopoulou-Kyriakides. M. Encapsulation of biological active phenolic compounds extracted from wine wastes in alginate-chitosan microbeads. J Microencaps 2018, 35, 229–240. [Google Scholar] [CrossRef]
- Rivera, K.; Salas-Pérez, F.; Echeverría, G.; Urquiaga, I.; Dicenta, S.; et al. Red Wine Grape Pomace Attenuates Atherosclerosis and Myocardial Damage and Increases Survival in Association with Improved Plasma Antioxidant Activity in a Murine Model of Lethal Ischemic Heart Disease. Nutrients. 2019, 11, 2135. [Google Scholar] [CrossRef]
- Hogan, S.; Zhang, L.; Li, J.; Sun, S.; Canning, C.; Zhou. K. Antioxidant rich grape pomace extract suppresses postprandial hyperglycemia in diabetic mice by specifically inhibiting alpha-glucosidase. Nutr. Metab. 2010, 7, 1. [Google Scholar] [CrossRef]
- Urquiaga, I.; D'Acuña, S.; Pérez, D.; Dicenta, S.; Echeverría, G.; Rigotti, A.; Leighton, F. Wine grape pomace flour improves blood pressure, fasting glucose and protein damage in humans: a randomized controlled trial. Biol Res. 2015, 48, 49. [Google Scholar] [CrossRef]
- Draijer, R.; de Graaf, Y.; Slettenaar, M.; de Groot, E.; Wright, C. Consumption of a Polyphenol-Rich Grape-Wine Extract Lowers Ambulatory Blood Pressure in Mildly Hypertensive Subjects. Nutrients 2015, 7, 3138–3153. [Google Scholar] [CrossRef]
- Del Pino-García, R.; Gerardi, G.; Rivero-Pérez, M.D.; González-SanJosé, M.L.; García-Lomillo, J.; Muñiz, P. Wine pomace seasoning attenuates hyperglycaemia-induced endothelial dysfunction and oxidative damage in endothelial cells. J Funct Foods 2016, 22, 431–445. [Google Scholar] [CrossRef]
- Flammer, A.J.; Sudano, I.; Wolfrum, M.; Thomas, R.; Enseleit, F.; Périat, D.; Kaiser, P.; et al. Cardiovascular effects of flavanol-rich chocolate in patients with heart failure. Eur Heart J 2012, 33, 2172–2180. [Google Scholar] [CrossRef]
- Wang, S.; Mateos, R.; Goya, L.; Amigo-Benavent, M.; Sarriá, B.; Bravo. L. A phenolic extract from grape by-products and its main hydroxybenzoic acids protect Caco-2 cells against pro-oxidant induced toxicity. Food Chem. Toxicol. 2016, 88, 65–74. [Google Scholar] [CrossRef]
- Tomé-Carneiro, J.; Gonzálvez, M.; Larrosa, M.; García-Almagro, F.J.; Avilés-Plaza, F.; Parra, S.; Yáñez-Gascón, M.J.; Ruiz-Ros, J.A.; García-Conesa, M.T.; Tomás-Barberán, F.A.; Espín, J.C. Consumption of a grape extract supplement containing resveratrol decreases oxidized LDL and ApoB in patients undergoing primary prevention of cardiovascular disease: A triple-blind, 6-month follow-up, placebo-controlled, randomized trial. Mol Nutr Food Res 2012, 56, 810–821. [Google Scholar] [CrossRef]
- de Oliveira, W.P.; Biasoto, A.C.T.; Marques, V.F.; dos Santos, I.M.; Magalhães, K.; Correa, L.C.; Shahidi. F. Phenolics from Winemaking By-Products Better Decrease VLDL-Cholesterol and Triacylglycerol Levels than Those of Red Wine in Wistar Rats. J Food Sci 2017, 82, 2432–2437. [Google Scholar] [CrossRef]
- Razavi, S.-M.; Gholamin, S.; Eskandari, A.; Mohsenian, N.; Ghorbanihaghjo, A.; Delazar, A.; Argani. H. Red grape seed extract improves lipid profiles and decreases oxidized low-density lipoprotein in patients with mild hyperlipidemia. J Med Food 2013, 16, 255–258. [Google Scholar] [CrossRef]
- Park, E.; Edirisinghe, I.; Choy, Y.Y.; Waterhouse, A.; Burton-Freeman, B. Effects of grape seed extract beverage on blood pressure and metabolic indices in individuals with pre-Hypertension: A randomised, double-Blinded, two-Arm, parallel, placebo-Controlled trial. Br. J. Nutr. 2016, 115, 226–238. [Google Scholar] [CrossRef]
- Pasini, F.; Chinnici, F.; Caboni, M.F.; Verardo, V. Recovery of oligomeric proanthocyanidins and other phenolic compounds with established bioactivity from grape seed by-Products. Molecules 2019, 24, 677. [Google Scholar] [CrossRef]
- Sapwarobol, S.; Adisakwattana, S.; Changpeng, S.; Ratanawachirin, W.; Tanruttanawong, K.; Boonyarit, W. Postprandial blood glucose response to grape seed extract in healthy participants: A pilot study. Pharmacogn Mag. 2012, 8, 192–6. [Google Scholar] [CrossRef]
- Montagut, G.; Onnockx, S.; Vaqué, M.; Bladé, C.; Blay, M.; Fernández-Larrea, J.; Pujadas, G.; Salvadó, M.J.; Arola, L.; Pirson, I. Oligomers of grape-seed procyanidin extract activate the insulin receptor and key targets of the insulin signaling pathway differently from insulin. J. Nutr. Biochem. 2010, 21, 476–481. [Google Scholar] [CrossRef]
- Kaur, M.; Agarwal, C.; Agarwa. R. Anticancer and cancer chemopreventive potential of grape seed extract and other grape-based products. J. Nutr. 2009, 139, 1806S–1812S. [Google Scholar] [CrossRef]
- Ono, K.; Condron, M.M.; Ho, L.; Wang, J.; Zhao, W.; Pasinetti, G.M.; Teplow, D.B. Effects of grape seed-derived polyphenols on amyloid beta-protein self-assembly and cytotoxicity. J Biol Chem. 2008, 283, 32176–87. [Google Scholar] [CrossRef]
- Charradi, K.; Mahmoudi, M.; Bedhiafi, T.; Jebari, K.; El May, M.; Limam, F.; Aouani, E. Safety evaluation, anti-oxidative and anti-inflammatory effects of subchronically dietary supplemented high dosing grape seed powder (GSP) to healthy rat. Biomed Pharmac 2018, 107, 534–546. [Google Scholar] [CrossRef]
- Cádiz-Gurrea, M.D.L.L.; Borrás-Linares, I.; Lozano-Sánchez, J.; Joven, J.; Fernández-Arroyo, S.; Segura-Carretero, A. Cocoa and Grape Seed Byproducts as a Source of Antioxidant and Anti-Inflammatory Proanthocyanidins. Int J Mol Sci 2017, 18, 376. [Google Scholar] [CrossRef]
- Rauf, A.; Imran, M.; Abu-Izneid, T.; Iahtisham,-Ul-Haq. ; Patel, S.; Pan, X.; Naz, S.; Sanches Silva, A.; Saeed, F.; Rasul Suleria, H.A. Proanthocyanidins: A comprehensive review. Biomed Pharm 2019, 116, 108999. [Google Scholar]
- Kruger, M.J.; Davies, N.; Myburgh, K.H.; Lecour, S. Proanthocyanidins, anthocyanins and cardiovascular diseases. Food Res Int 2014, 59, 41–52. [Google Scholar] [CrossRef]
- Iriondo-DeHond, M.; Blázquez-Duff, J.M.; Del Castillo, M.D.; Miguel, E. Nutritional Quality, Sensory Analysis and Shelf Life Stability of Yogurts Containing Inulin-Type Fructans and Winery Byproducts for Sustainable Health. Foods. 2020, 9, 1199. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-H.; Huang, B.; Choi, S.-K.; Seo, J.-S. Anti-obesity effect of resveratrol-amplified grape skin extracts on 3T3-L1 adipocytes differentiation. Nutr Res Pract 2012, 6, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Ismail, A.F.M.; Moawed, F.S.M.; Mohamed. M.A. Protective mechanism of grape seed oil on carbon tetrachloride-induced brain damage in γ-irradiated rats. J. Photoch. Photobio. 2015, 153, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Ismail, A.F.M.; Salem, A.A.M.; Eassawy. M.M.T. Hepatoprotective effect of grape seed oil against carbon tetrachloride induced oxidative stress in liver of γ-irradiated rat. J. Photoch. Photobio. 2016, 160, 1–10. [Google Scholar] [CrossRef]
- Asadi, F.; Shahriari, A.; Chahardah-Cheric. M. Effect of long-term optimal ingestion of canola oil, grape seed oil, corn oil and yogurt butter on serum, muscle and liver cholesterol status in rats. Food Chem. Toxicol. 2010, 48, 2454–2457. [Google Scholar] [CrossRef]
- Nguyen, A.V.; Martinez, M.; Stamos, M.J.; Moyer, M.P.; Planutis, K.; Hope, C.; Holcombe. R.F. Results of a phase I pilot clinical trial examining the effect of plant-derived resveratrol and grape powder on Wnt pathway target gene expression in colonic mucosa and colon cancer. Cancer Manag. Res. 2009, 1, 25–37. [Google Scholar]
- Elshaer, M.; Chen, Y.; Wang, X.J.; Tang, X. Resveratrol: An overview of its anti-cancer mechanisms. Life Sci 2018, 207, 340–349. [Google Scholar] [CrossRef]
- Wang, L.-S.; Stoner, G.D. Anthocyanins and their role in cancer prevention. Cancer Lett 2008, 269, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Lala, G.; Malik, M.; Zhao, C.; He, J.; Kwon, Y.; Giusti, M.M.; Magnuson. B.A. Anthocyanin-rich extracts inhibit multiple biomarkers of colon cancer in rats. Nutr Cancer 2006, 54, 84–93. [Google Scholar] [CrossRef]
- Tian, Q.; Xu, Z.; Sun, X.; Deavila, J.; Du, M.; Zhu, M. Grape pomace inhibits colon carcinogenesis by suppressing cell proliferation and inducing epigenetic modifications. J Nutr Biochem. 2020, 84, 108443. [Google Scholar] [CrossRef]
- Mišković Špoljarić, K.; Šelo, G.; Pešut, E.; Martinović, J.; Planinić, M.; Tišma, M.; Bucić-Kojić, A. Antioxidant and antiproliferative potentials of phenolic-rich extracts from biotransformed grape pomace in colorectal Cancer. BMC Complement Med Ther. 2023, 23, 29. [Google Scholar] [CrossRef] [PubMed]
- Singletary, K.W.; Jung, K.J.; Giusti, M. Anthocyanin-rich grape extract blocks breast cell DNA damage. J Med Food. 2007, 10, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Song, S.; Wei, Z.; Huang, Y.; Zhang, Y.; Lu, J. The comparative study among different fractions of muscadine grape 'Noble' pomace extracts regarding anti-oxidative activities, cell cycle arrest and apoptosis in breast cancer. Food Nutr Res. 2017, 61, 1412795. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.; Silva, V.; Igrejas, G.; Gaivao, I.; Aires, A.; Klibi, N.; Dapkevicius, M.; Valentao, P.; Falco, V.; Poeta, P. Valorization of Winemaking By-Products as a Novel Source of Antibacterial Properties: New Strategies to Fight Antibiotic Resistance. Molecules. 2021, 26, 2331. [Google Scholar] [CrossRef]
- Shan, B.; Cai, Y.-Z.; Brooks, J.D.; Corke, H. Potential application of spice and herb extracts as natural preservatives in cheese. J Med Food 2011, 14, 284–290. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects – A review. J Funct Foods, Natural Antioxidants 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Mattio, L.M.; Dallavalle, S.; Musso, L.; Filardi, R.; Franzetti, L.; Pellegrino, L.; D’Incecco, P.; Mora, D.; Pinto, A.; Arioli, S. Antimicrobial activity of resveratrol-derived monomers and dimers against foodborne pathogens. Sci Rep 2019, 9, 19525. [Google Scholar] [CrossRef]
- de Vries, K.; Strydom, M.; Steenkamp, V. Bioavailability of resveratrol: Possibilities for enhancement. J Herbal Med 2018, 11, 71–77. [Google Scholar] [CrossRef]
- Tsali, A.; Goula, A.M. Valorization of grape pomace: Encapsulation and storage stability of its phenolic extract. Powder Tech 2018, 340, 194–207. [Google Scholar] [CrossRef]
- Domínguez, R.; Pateiro, M.; Munekata, P.E.S.; McClements, D.J.; Lorenzo, J.M. Encapsulation of Bioactive Phytochemicals in Plant-Based Matrices and Application as Additives in Meat and Meat Products. Molecules 2021, 26, 3984. [Google Scholar] [CrossRef]
- Glampedaki, P.; Dutschk. V. Stability studies of cosmetic emulsions prepared from natural products such as wine, grape seed oil and mastic resin. Colloid. Surf. A, 2014, 460, 306–311. [Google Scholar] [CrossRef]
- Wittenauer, J.; Mäckle, S.; Sußmann, D.; Schweiggert-Weisz, U.; Carle. R. Inhibitory effects of polyphenols from grape pomace extract on collagenase and elastase activity. Fitoterapia 2015, 101, 179–187. [Google Scholar] [CrossRef]
- Marino, A.; Battaglini, M.; Desii, A.; Lavarello, C.; Genchi, G.; Petretto, A.; Ciofani, G. Liposomes loaded with polyphenol-rich grape pomace extracts protect from neurodegeneration in a rotenone-based in vitro model of Parkinson's disease. Biomater Sci. 2021, 9, 8171–8188. [Google Scholar] [CrossRef]
- Davidov-Pardo, G. ; McClements. D.J. Nutraceutical delivery systems: Resveratrol encapsulation in grape seed oil nanoemulsions formed by spontaneous emulsification. Food Chem 2015, 167, 205–212. [Google Scholar]
- Hübner, A.A.; Sarruf, F.D.; Oliveira, C.A.; Neto, A.V.; Fischer, D.C.H.; Kato, E.T.M.; Lourenço, F.R.; Baby, A.R.; Bacchi, E.M. Safety and Photoprotective Efficacy of a Sunscreen System Based on Grape Pomace (Vitis vinifera L.) Phenolics from Winemaking. Pharmaceutics. 2020, 12, 1148. [Google Scholar] [CrossRef]
- Jakobek, L.; Matić, P. Non-covalent dietary fiber - Polyphenol interactions and their influence on polyphenol bioaccessibility. Trends Food Sci Tech 2019, 83, 235–247. [Google Scholar] [CrossRef]
- Pérez-Jiménez, J.; Serrano, J.; Tabernero, M.; Arranz, S.; Díaz-Rubio, M.E.; García-Diz, L.; Goñi, I.; Saura-Calixto, F. Bioavailability of Phenolic Antioxidants Associated with Dietary Fiber: Plasma Antioxidant Capacity After Acute and Long-Term Intake in Humans. Plant Foods Hum Nutr 2009, 64, 102–107. [Google Scholar] [CrossRef]
- Ferreira, C.S.; Pinho, M.N.; Cabral, L. Solid-liquid extraction and concentration with processes of membrane technology of soluble fibers from wine grape pomace. Técnico Lisboa, 2013; 1–9. [Google Scholar]
- Zhang, L.; Zhu, M.; Shi, T.; Guo, C.; Huang, Y.; Chen, Y.; Xie, M. Recovery of dietary fiber and polyphenol from grape juice pomace and evaluation of their functional properties and polyphenol compositions. Food Funct. 2017, 8, 341–351. [Google Scholar] [CrossRef]
- Zhu, F.; Du, B.; Zheng, L.; Li, J. Advance on the bioactivity and potential applications of dietary fibre from grape pomace. Food Chemistry, ISPMF 2015: International Symposium on Phytochemicals in Medicine and Food (Shanghai, China, June 26th –29th, 2015, 186, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Valiente, C.; Arrigoni, E.; Esteban, R.; Amadò, R. Grape Pomace as a Potential Food Fiber. J Food Sci 1995, 60, 818–820. [Google Scholar] [CrossRef]
- Dias, J.F.; Simbras, B.D.; Beres, C.; dos Santos, K.O.; Cabral, L.M.C.; Miguel, M.A.L. Acid Lactic Bacteria as a Bio-Preservant for Grape Pomace Beverage. Front Sust Food Syst 2018, 2. [Google Scholar] [CrossRef]
- Ageyeva, N.; Tikhonova, A.; Burtsev, B.; Biryukova, S.; Globa, E. Grape pomace treatment methods and their effects on storage. Foods Raw Mat 2021, 9, 215–223. [Google Scholar] [CrossRef]
- Ayed, N.; Yu, H.-L.; Lacroix, M. Using gamma irradiation for the recovery of anthocyanins from grape pomace. Rad Phys Chem 2000, 57, 277–279. [Google Scholar] [CrossRef]
- Tseng, A.; Zhao, Y. Effect of different drying methods and storage time on the retention of bioactive compounds and antibacterial activity of wine grape pomace (Pinot Noir and Merlot). J Food Sci 2012, 77, H192–H201. [Google Scholar] [CrossRef]
- Sokač, T.; Gunjević, V.; Pušek, A.; Tušek, A.J.; Dujmić, F.; Brnčić, M.; Ganić, K.K.; Jakovljević, T.; Uher, D.; Mitrić, G.; Redovniković, I.R. Comparison of Drying Methods and Their Effect on the Stability of Graševina Grape Pomace Biologically Active Compounds. Foods 2022, 11, 112. [Google Scholar] [CrossRef] [PubMed]
- MohdMaidin, N.; Oruna-Concha, M.J.; Jauregi, P. Surfactant TWEEN20 provides stabilisation effect on anthocyanins extracted from red grape pomace. Food Chem. 2019, 271, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Torri, L.; Piochi, M.; Marchiani, R.; Zeppa, G.; Dinnella, C.; Monteleone, E. A sensory- and consumer-based approach to optimize cheese enrichment with grape skin powders. J Dairy Sci 2016, 99, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Soares, Susana, Brandão, E. ; Guerreiro, C.; Soares, Sónia, Mateus, N.; de Freitas, V. Tannins in Food: Insights into the Molecular Perception of Astringency and Bitter Taste. Molecules 2020, 25, 2590. [Google Scholar] [CrossRef] [PubMed]
- McRae, J.M.; Kennedy, J.A. Wine and Grape Tannin Interactions with Salivary Proteins and Their Impact on Astringency: A Review of Current Research. Molecules 2011, 16, 2348–2364. [Google Scholar] [CrossRef] [PubMed]
- Canett-Romero, R.; Osuna, A.; Sánchez, M.; Castro, R.; León-Martínez, L.; León-Gálvez, R. Characterization of cookies made with deseeded grape pomace. Arch Latinoam Nutr 2004, 54, 93–9. [Google Scholar] [PubMed]
- Caponio, G.R.; Noviello, M.; Calabrese, F.M.; Gambacorta, G.; Giannelli, G.; De Angelis, M. Effects of Grape Pomace Polyphenols and In Vitro Gastrointestinal Digestion on Antimicrobial Activity: Recovery of Bioactive Compounds. Antioxidants 2022, 11, 567. [Google Scholar] [CrossRef] [PubMed]
- Čuš, F.; Česnik, H.B.; Bolta, Š.V.; Gregorčič, A. Pesticide residues in grapes and during vinification process. Food Control 2010, 21, 1512–1518. [Google Scholar] [CrossRef]
- Hou, X.; Xu, Z.; Zhao, Y.; Liu, D. Rapid analysis and residue evaluation of six fungicides in grape wine-making and drying. J Food Comp Anal 2020, 89, 103465. [Google Scholar] [CrossRef]
- Moncalvo, A.; Marinoni, L.; Dordoni, R.; Duserm Garrido, G.; Lavelli, V.; Spigno, G. Waste grape skins: evaluation of safety aspects for the production of functional powders and extracts for the food sector. Food AddCont: Part A 2016, 33, 1116–1126. [Google Scholar] [CrossRef]
Figure 1.
The structures of the most characteristic non-flavonoid (A) and flavonoid (B) phenolic compounds present in wine and/or grape pomace. Structures were obtained from https://molview.org/ (accessed on 24th of April 2023).
Figure 1.
The structures of the most characteristic non-flavonoid (A) and flavonoid (B) phenolic compounds present in wine and/or grape pomace. Structures were obtained from https://molview.org/ (accessed on 24th of April 2023).
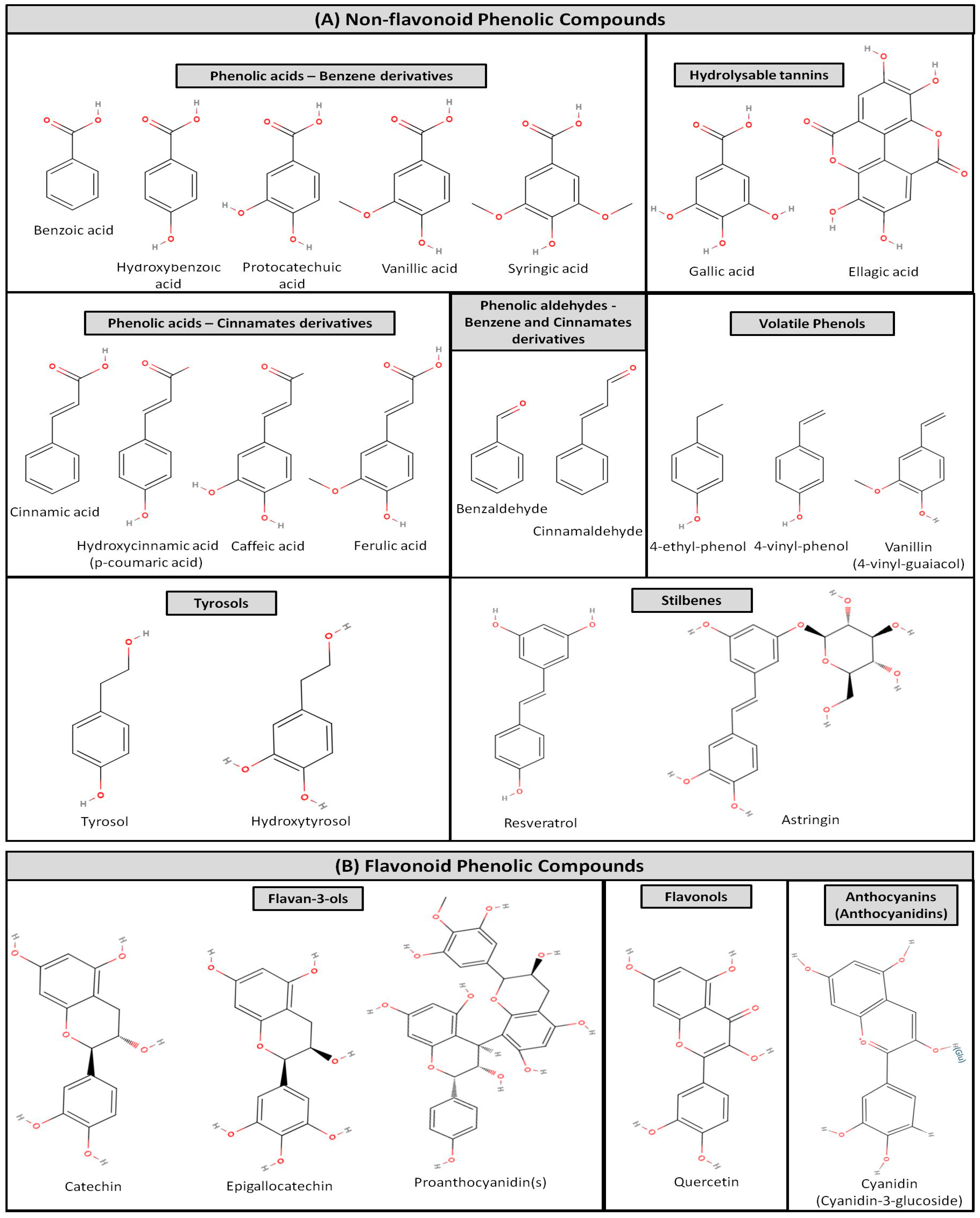
Figure 2.
Structures of bioactive UFA (A) and bio-functional PL with UFA at their sn2 position of their glycerol backbone (B), with anti-inflammatory and antithrombotic properties and health benefits found in wine and grape pomace; ALA = alpha linolenic acid (C18:3 omega-3); OA = oleic acid (C18:1 omega-9).
Figure 2.
Structures of bioactive UFA (A) and bio-functional PL with UFA at their sn2 position of their glycerol backbone (B), with anti-inflammatory and antithrombotic properties and health benefits found in wine and grape pomace; ALA = alpha linolenic acid (C18:3 omega-3); OA = oleic acid (C18:1 omega-9).
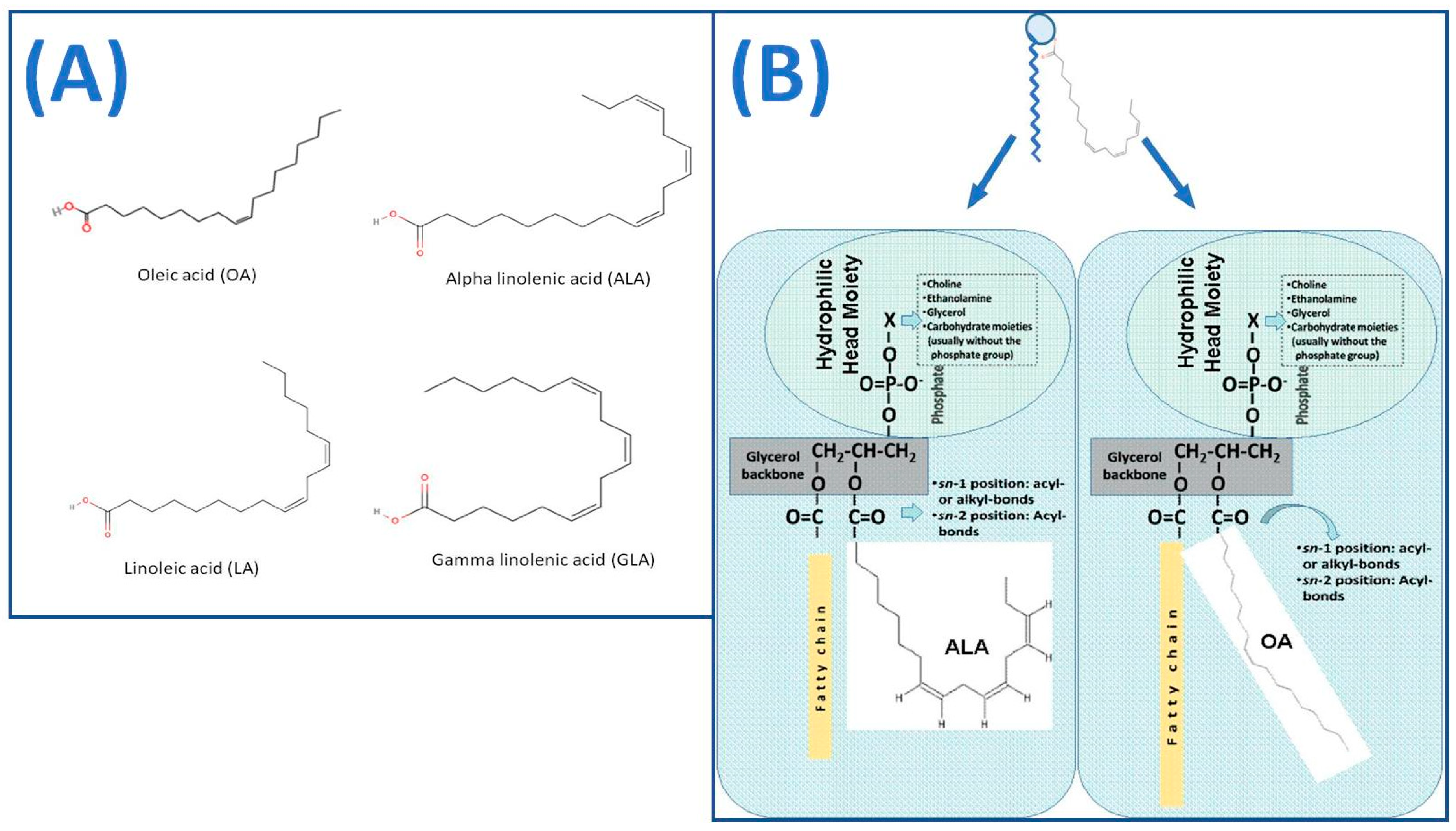
Figure 3.
Modes of beneficial actions of the bioactive polar lipids (in blue color) against the thrombo-inflammatory related pathways and cell-responses (red color). PL = polar lipids; PAF = platelet-activating factor; ADP = adenosine 5′ diphosphate; GPCMR = G-protein coupled membrane receptors; cPLA2 = cytoplasmic Phospholipase A2; LPCAT & PAF-CPT = Basic regulatory biosynthetic enzymes of PAF; ARA = arachidonic acid; COX = cycloxygenases.
Figure 3.
Modes of beneficial actions of the bioactive polar lipids (in blue color) against the thrombo-inflammatory related pathways and cell-responses (red color). PL = polar lipids; PAF = platelet-activating factor; ADP = adenosine 5′ diphosphate; GPCMR = G-protein coupled membrane receptors; cPLA2 = cytoplasmic Phospholipase A2; LPCAT & PAF-CPT = Basic regulatory biosynthetic enzymes of PAF; ARA = arachidonic acid; COX = cycloxygenases.
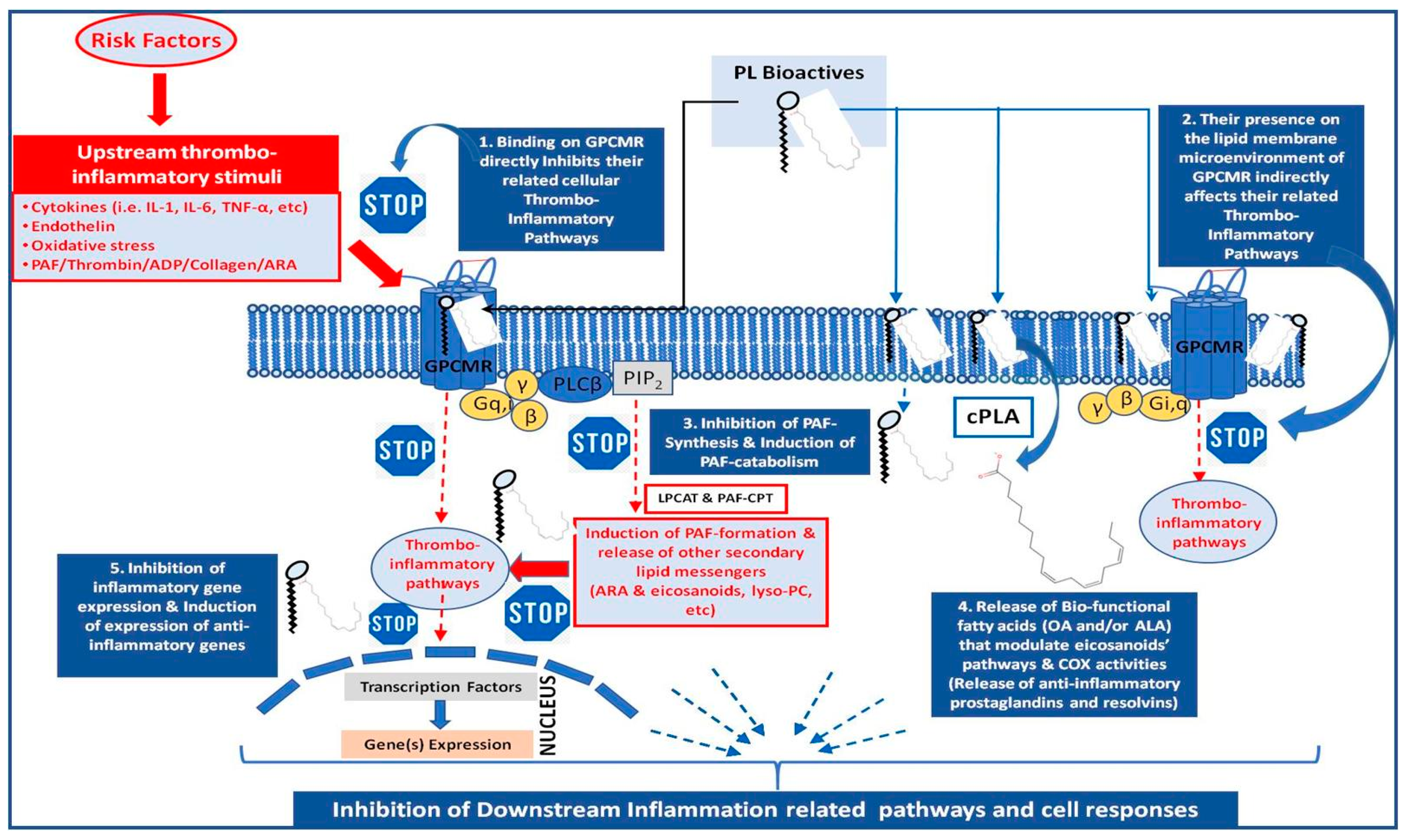

Table 1.
Proximate composition of grape pomace based on dry weight, and phenolic compounds expressed as mg of Gallic Acid equivalent (GAE) per liter of extract (mg GAE/L)*.
Table 1.
Proximate composition of grape pomace based on dry weight, and phenolic compounds expressed as mg of Gallic Acid equivalent (GAE) per liter of extract (mg GAE/L)*.
| Compounds | Quantity g/100g | Phenolic compounds | mg GAE/L |
|---|---|---|---|
| Ash | 1.73 – 9.10 | Flavonoids | 1.31 – 8.36 |
| Protein | 3.57-14.17 | Anthocyanins | 0.33 – 6.8 |
| lipids | 1.14-13.90 | Tannins | 0.74 – 6.41 |
| Total dietary fibre | 17.28-88.70 | Proanthocyanins | 0.43 – 12.00 |
| Insoluble fibre | 16.44-63.70 | Catechin | 0.69 – 8.06 |
| Carbohydrates | 12.20-40.53 | Epicatechin | 0.61-0.99 |
| Total polyphenolic content | 0.28-8.70 | Resveratrol | 0.58 – 37.93 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



