
Submitted:
12 June 2023
Posted:
15 June 2023
You are already at the latest version
Abstract
Ribosomal proteins (RPs) mediate protein synthesis and stability of the ribosomal complex. RPs and rRNA form ribosomal subunits and ri-bosomes, the cellular machinery for protein synthesis, a fundamental biological process related to cell growth and proliferation. The differ-ences between paralogs within the same RP family have led them to ac-quire extraribosomal functions involved in plants' growth, develop-ment, and biotic and abiotic stresses. From transcriptomic analysis, it has been revealed that ribosomal proteins have a crucial role during MTs development. The RPLs; RPL11, RPL29 and RPL40 interact with PEBP family members (Phosphatidylethanolamine-binding proteins), poten-tial activators in the MTs development. In potatoes, genome editing is a promising technology to introduce crop breeding traits. Recently, the edition of the genome through the CRISPR/Cas9 system has made it the most efficient and powerful genetic modification method. Based on this, the gene modulation by overexpression and silencing in RPL11, RPL29 and RPL40 will guide us to understand the effect on the microtuberiza-tion process and produce improved potato plants with the capability of growing under adverse environments (biotic and abiotic stresses). By understanding the molecular biology mechanisms that RPs govern, we can improve crops under today's changing climatic conditions.
Keywords:
Ribosome biogenesis
; Potato
; Carbon metabolism
; Genome edition
; CRISPR/Cas
; Microtuberization
1. Introduction
Potato is the crop that produces the most significant amount of food per unit area, helping to fight global hunger and poverty. By 2050, the world population has been estimated to skyrocket to 10 billion. The agricultural production of food shall increase more than 60 percent to meet this rapidly growing demand. Climate change is a principal contributor affecting food production, while agricultural land expansion is not a sustainable solution to feed the world by 2050 [1]. Hence genetic modification of the potato, specifically of specific ribosomal proteins, represents one of the most promising alternatives for the present and the future.
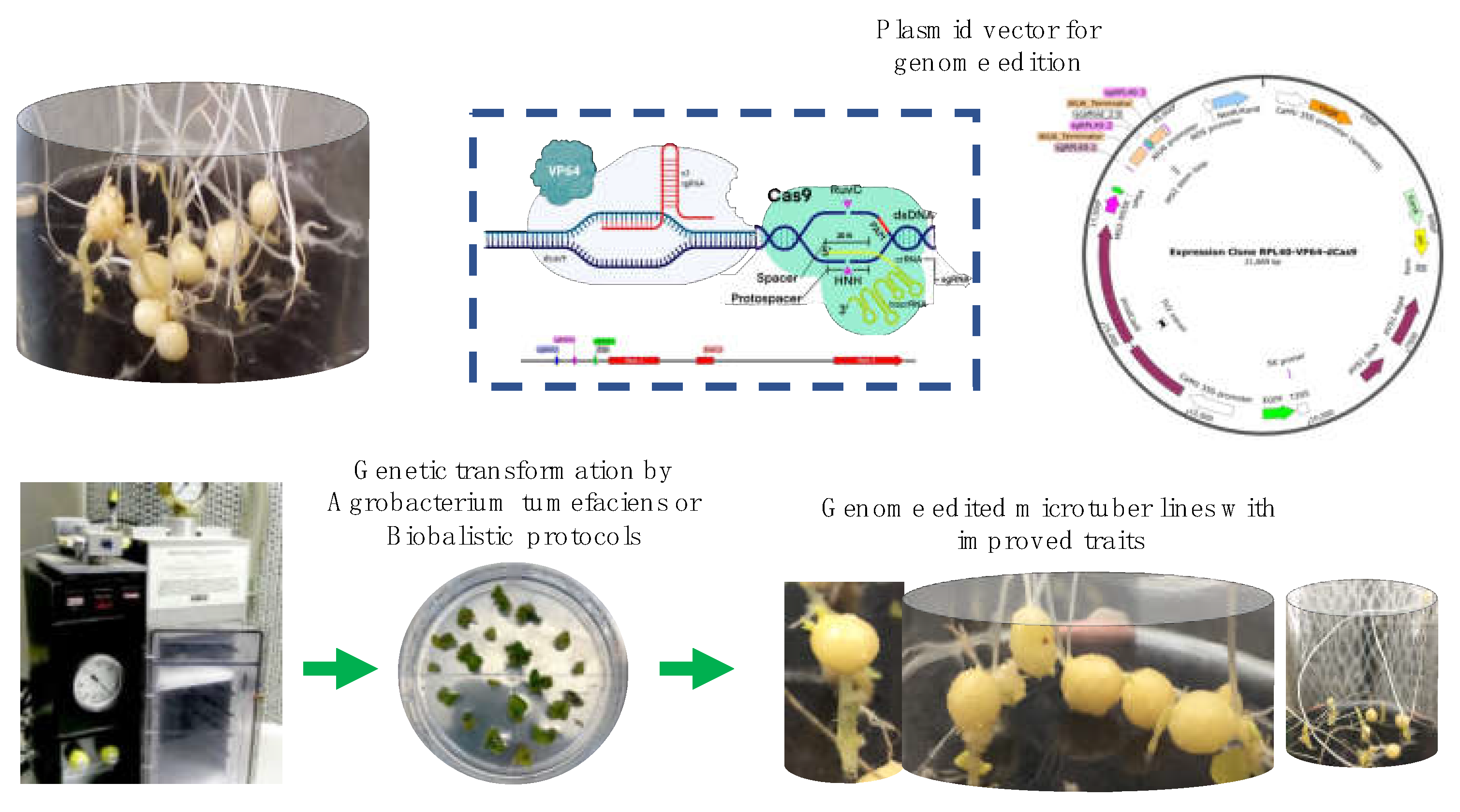
The molecular mechanisms involved in potato tuber development under greenhouse conditions have been extensively studied [for review, see 2]. An alternative technology is the in vitro induction of potato microtubers (MTs). The axillary buds underwent MT induction in a medium containing high sucrose content, plant growth regulators, and several light quality/darkness. The benefits of MT production includes a higher multiplication rate, better storage and transport due to smaller size and weight, and easier, cheaper and faster cultivation than other methods [3,4,5].
A MTs induction protocol for potato S. tuberosum var Alpha by culturing stolon explants in MS medium supplemented with 8% sucrose, high content of gelrite 6.0 g/L, cytokinin (CK) 2iP 10 mg/L under darkness was developed by our research group. This protocol was based upon the underlying mechanisms of the interaction of CK signaling with homeobox transcription factors, RPs, cell cycle (CC), carbon metabolism, auxin-responsive factors and stem cell maintenance proteins also involved in the whole process [6].
Transcriptome analysis of the MTs revealed that ~1700 up-regulated and ~1600 down-regulated genes were regulating the morphogenetic process. The PPI network analyses were performed at highest confidence in the STRING database platform (v11.5, www.string-db.org), revealing ~300 genes were highly associated in tens of clusters. Two essential life proteins groups were discussed: Ribosomal proteins, of which RPL11 interacts with several PEBP family members, and proteins of the cell cycle [7].
The yeast two-hybrid approach for protein interactors screening found that the PEBP - StSP6A, a positive regulator of tuber development, can interact with RPs, other protein synthesis regulators, RNA and DNA interacting proteins, histones, initiation factors, flowering regulation, cellular signaling and carbon metabolism [8,9,10,11,12].
In the tuberization process of S. tuberosum from previous transcriptomic analyses have established the relevance of RPs during the tuberization process [13,14,15,16,17]. In the work of Sharma and Hannapel 2016 [13], a PPI network analysis revealed that RPs interact with SP6A, RP40SA, RP60S, RPS8, RPS4A, RPL10, BEL5, and RPL14. In addition, we applied the highest confidence value (0.900) in the same analysis, which showed 69 transcripts coding for RPs proteins that were up-regulated, 53 of which are directly involved in ribosome biogenesis [13].
In this manuscript, we will discuss the feasibility of activating RPs using CRISPR/Cas technology to induce MTs with several trait advantages such as size, number, stress tolerance, and protein content. We will also discuss the agronomic importance of several genes interacting with RPs derived from the transcriptome analysis performed by Valencia-Lozano et al. 2022 [7].
Transcriptome Analysis of MTs Induction
Transcriptome analysis of MT induction was performed according to Valencia-Lozano et al. 2022 [7]. The study revealed 1699 up-regulated genes, from which 299 were tightly associated with two essential biological processes and highly conserved through organisms: protein synthesis and cell cycle regulation comprising 29 and 117 proteins, respectively.
RPs interact with environment sensor proteins: 6 PEBP members, 21 related to osmotic stress, 9 with oxidative stress, 23 with CK response, 6 with one-carbon metabolism, 38 with carbon metabolism, 16 with TCA cycle, 6 with acyl carrier proteins, 14 fatty acid metabolism, 13 to thylakoid, 9 to redoxins, 8 with sulfur metabolism, 5 disulfide isomerase activity and 12 with immunophilins.
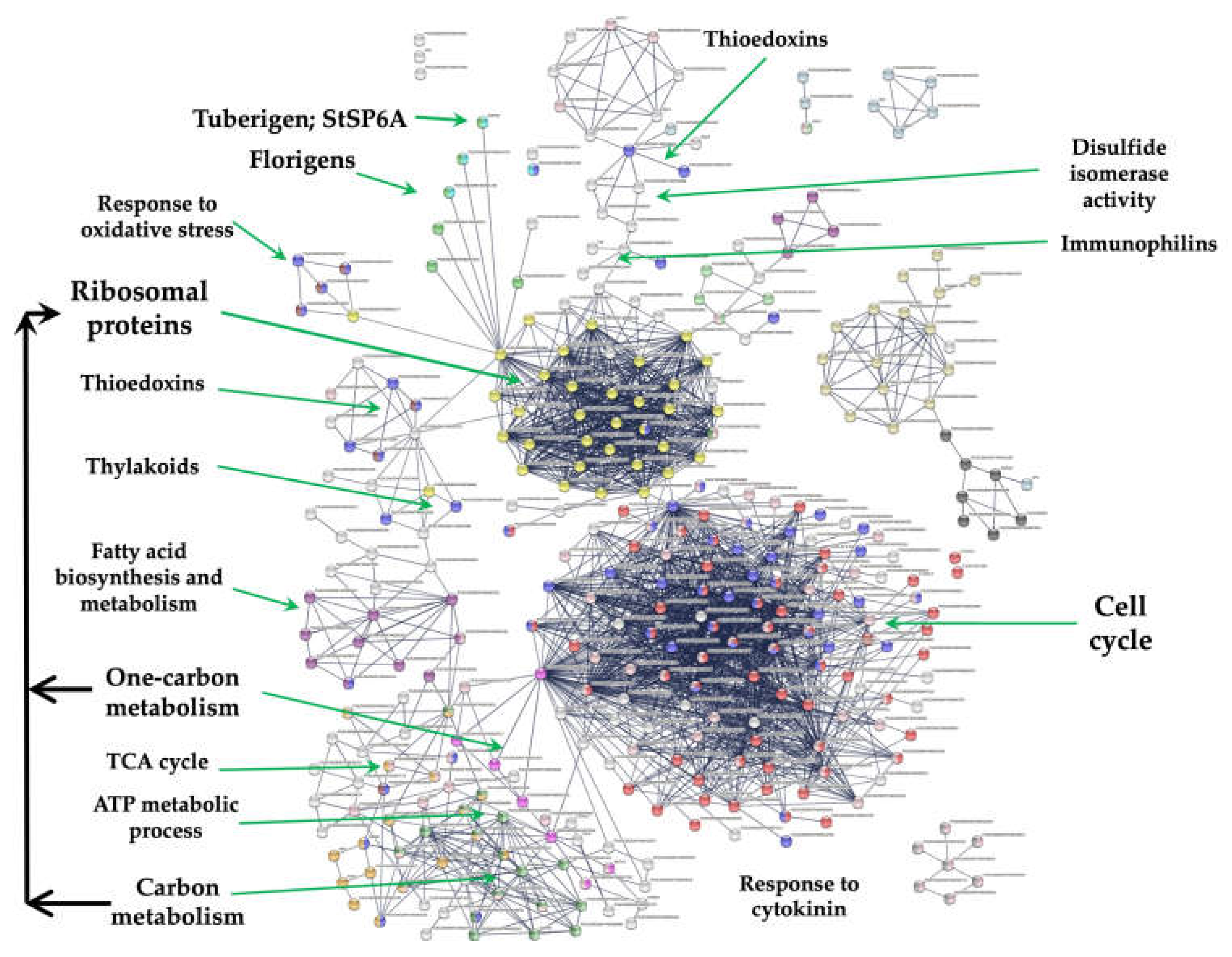
Figure 1.
PPI network of RPL11 interacting with the tuberigen StSP6A and PEBP members during microtuberization of potato. Also, the interaction of RPL11 with thioredoxins, thylakoid biogenesis, fatty acid biosynthesis, and carbon metabolism. The PPI network has the highest confidence (0.900).
Figure 1.
PPI network of RPL11 interacting with the tuberigen StSP6A and PEBP members during microtuberization of potato. Also, the interaction of RPL11 with thioredoxins, thylakoid biogenesis, fatty acid biosynthesis, and carbon metabolism. The PPI network has the highest confidence (0.900).
What is the Importance of Ribosome Biogenesis?
The biogenesis of the ribosome is directly associated with plant growth and development, and reproduction. The loss-of-function in genes encoding ribosomal proteins (RPs) or ribosome biogenesis factors (RBFs) gave rise to decreased growth, delayed flowering, and in more severe cases, are lethal. Loss-of-function of 19 ribosomal proteins and 26 ribosome biogenesis factors are seedling/embryo-lethal (Table 1).
Lethal Mutants
The small subunits RPs, RPS1 and RPS20, are seedling-lethal in rice [18,19], RPS9 and RPS17 are embryo lethal in maize [20,21], RPS16 and RPS27 are embryo lethal in Arabidopsis [22,23], and RPS18A is lethal in tobacco [24] (Table 1).
Large subunit RPs, RPL13, RPL12, RPL13, and RPL21C are seedling/embryo lethal in rice [25,26,27,28] (Table 1). In Arabidopsis, RPL5C, RPL9C,D, RPL10, RPL21C, and RPL28-1 are embryo lethal [29,30,31,32,33,34] (Table 1). At least 29 ribosome biogenesis factors have been shown to be lethal, including proteins involved in chloroplast development, ribosome biogenesis, nucleolar organization, and chlorophyll biosynthesis (Table 1).
In our transcriptome analysis 5 RPLs; RPL1, 12, 13, 27 and RPL35 and RPSs; RPS1, 9, 16 and RPS17 are embryo lethal.
Overexpression of Ribosomal Proteins and Ribosome Biogenesis Factors
Horvath et al. 2006 [35] demonstrated that EBP1 transcriptional profile correlates positively with organ development and ribosome biogenesis related genes upregulation in potatoes. The EBP1 regulates the final stages of rRNA processing. Silenced potato lines showed reduced size, tuber yield and abnormal morphology.
Potato plants transformed with the RP StoL13a from Solanum torvum, SW, a highly resistant plant to Verticillium dhaliae infection, showed a resistance increase to V. dahliae infection compared with wild-type plants. The transformed plants exhibited reactive oxygen species decreased levels achieving an attenuated oxidative damage. In addition, several defence and antioxidant enzyme coding genes were upregulated in those transformed plants. These results support the plant defence role that StoL13a plays against V. dahliae infection [36].
Overexpression of RPL6 in Oryza sativa indica resulted in salt tolerance [37]. The overexpression of RPL23A in rice plants showed resistance to water deficiency, growth suitability and yield parameters compared to the negative control. It was also associated with upregulating many RPs, small and large subunits [38].
The 25S ribosomal RNA requires of the nucleolar methyltransferase activity of OLI2/NOP2A to process its transcript maturation and, in turn to join to the 60S large ribosomal subunit. Plants mutated in OLI2 yielded lighter seeds, displayed delayed germination, developed lesser lateral roots resembling the auxin-related mutants, and with increased sensitivity to sugar concentrations. The OLI2 overexpressing plants produced heavier seeds and showed a reduced sensitivity to high sugar concentration [39].
For the abiotic stress of cold the STCH4/REIL2 triggers the upregulation of this ribosomal biogenesis factor. STCH4 overexpression in Arabidopsis gave rise to chilling and freezing tolerance. Also, the stch4 mutants showed a decrease in the C-repeat-binding factor (CBF) protein levels and, in turn, the expression of the genes induced by CBF was also decreased significantly [40].
Are RPs good candidate genes for improving of multiple abiotic stress tolerance in potato?
Crop plants, such as potato, are negatively affected by drought stress, for any developmental stage of the plant, from seed germination to tuber emergence and bulking to finally impact in tuber yield. Tuber cracking and malformation, hollow heart, vascular discolouration, and reduction in the dry matter accumulation in tubers are physiological disorders as a direct consequence of extensive periods of water deficiency.
Kappachery et al. 2013 [41] used the functional screening approach in yeast for the potential drought tolerance genes identification in potato plants. From potato plants exposed in a hyperosmotic culture media a cDNA expression library was constructed, and the transformed yeast with enhanced ability to survive under hyperosmotic stress expressing the cDNAs from the expression library were selected.
Sixty-nine genes were selected for their ability to grow under drought, salt, and heat stress. Of this, 8 were RPs; RPS7, RPL12, RPL10-like, RPL27, RPL18a, RPL1, RPS11, and RPL36 [41]. Accordingly, with that, the potential of RPs is very promising to obtain potato microtubers lines with desirable traits.
How to produce potato microtubers tolerant to biotic and abiotic stress?
The methodology CRISPR/Cas is a genetic modification tool that can be used to induce the expression of any gene, including the RPs. We found in the transcriptome analysis (RPL11, RPL29, RPL40, RPL16), interacting with several gene clusters, like the tuberigen and flowering, cell cycle, proteasome, immunophilins and oxidative stress.
The system CRISPRa 2.0 [42] and CRISPR-P 2.0 [43] can be used to obtain the plasmid constructs. Three guides RNAs (gRNAs) for the -228 bp and -19 bp from the transcription initiation site for each gene (for the avoidance TATA-box, initiator elements, and DPE), including those with the NGG sequence as PAM, aligning with the promoter region of the S. tuberosum genes SP6A (PGSC0003DMT400060057), RPL11 (PGSC0003DMT400031869), RPL29 (PGSC0003DMT400069470), RPL40 (PGSC0003DMT400047686), RPL16 (PGSC0003DMT400060127), and cloned under methodology described by [44,45] (Lowder et al., 2015; Garcia-Murillo et al., 2023).
The vectors dCas9 fused to the VPG4, and the gRNA cassette, into a binary vector to generate: dCas9-VPG4-SP6A, dCas9-VP64-RPL11, dCas9-VP64-RPL2, dCas9-VP64-RPL40, dCas9-VP64-RPL16; named here SP6Av, RPL11v, RPL29v, RPL40v, RPL16v). Controls for transformation will be the empty entry vector containing only the dCas9 (dC9-v) and plant wilt-type without genetic modifications.
Interaction of RPs cluster with immunophilines
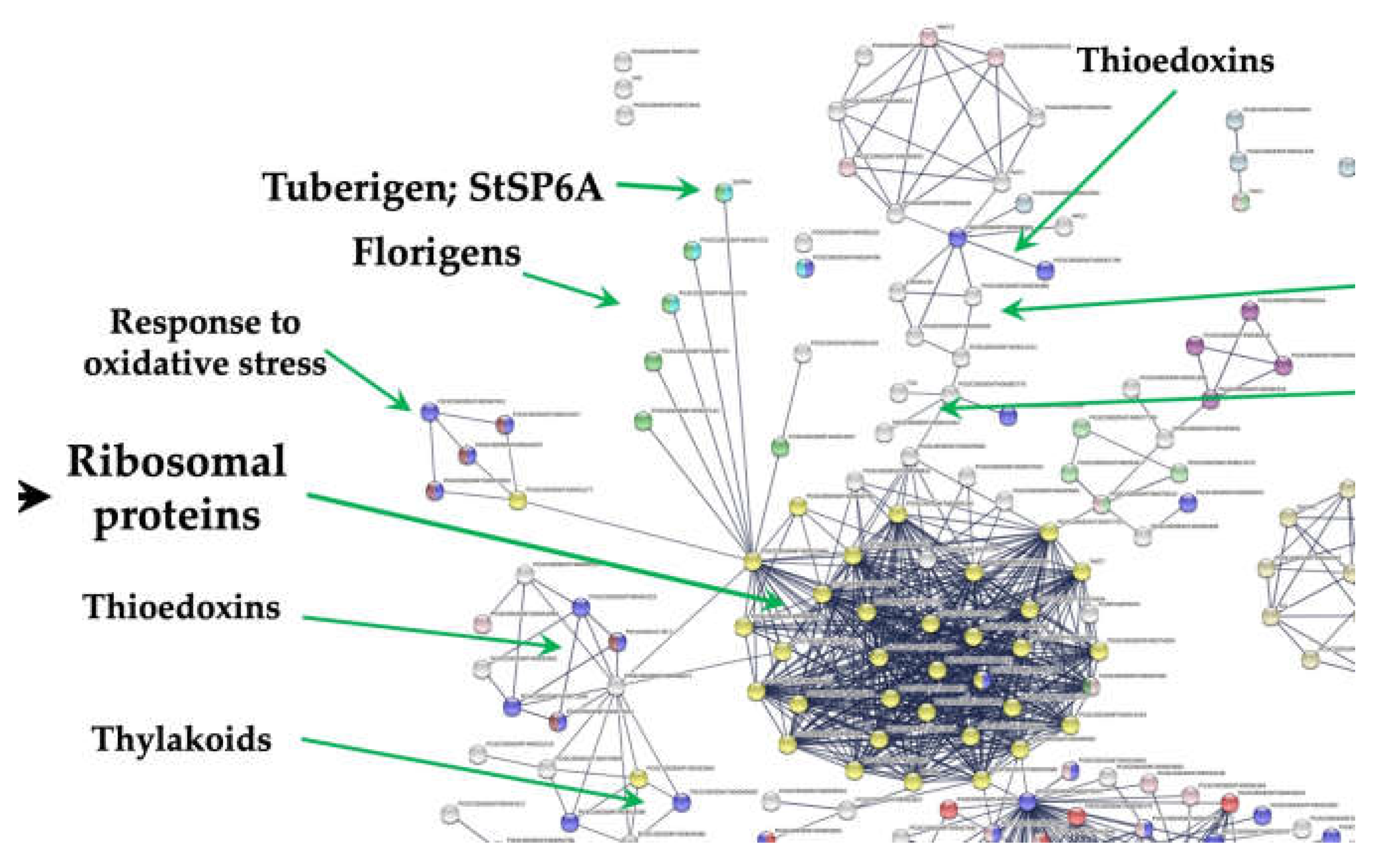
RPL29 (PGSC0003DMT400069847) interacts with FKBP12. This group of proteins includes PPIs or immunophilins and protein disulfide isomerases. The cluster consists of 4 PPI genes interacting with SOS3 (PGSC0003DMT400023568), a calcium sensor calcineurin B, crucial for the salt stress-induced Ca2+ signalling transduction and activation of salt tolerance mechanisms in Arabidopsis. The SOS3 loss-of-function gave rise to salt hypersensitivity and significantly decreases the Ca2+ binding capability [47].
The FKBP12 overexpression in Arabidopsis, has promoted growth under non stress conditions, and has increased the responses to abiotic stress [48]. Also, the CYP21 overexpression in potatoes, which is involved in oxidative stress response, produced longer plants, heavier tubers, and microtuberization yielded more significant amounts in a shorter time and weight per MT [49]. Furthermore, three genes with Disulfide Isomerase-like proteins which are present in this cluster, STPDI1, 2 and 3. They interact with the calnexin (PGSC0003DMT400036920). StPDI1 accumulation triggers the quaternary structure establishment to counteracts salt exposure, enhance the catalyzes of disulfide bonds and contributes to reactive oxygen species response. Thus, they may work in a similar way as HSP for chaperone activity during salt stress [50]. The StPDI1 expression levels reduction in potato plants negatively impacts the abiotic stress tolerance. The malate amount, succinate and 2-oxoglutarate and reducing equivalent NADH content was decreased significantly in StPDI1-inhibited potato plants. In contrast, serine and threonine content were significantly increased compared to wild-type plants [51].
Redoxins
RPs interact with the thioredoxin cluster through the interaction of the PRXQ peroxiredoxin (PGSC0003DMT400035271) with RPL11 (PGSC0003DMT400031869) and RPL18 (PGSC0003DMT400011931). This cluster consists of 7 peroxiredoxin genes. PRXQ carried out an oxidative stress protecting function through peroxides detoxification. And also, this protein is involved in the photosystem II protection against hydrogen peroxide.
To decrease the Reactive Oxygen Species (ROS) negative effects, aerobic organisms have developed counteracting mechanisms, involving catalases, superoxide dismutases, ascorbate peroxidases, glutathione peroxidases and GSTs with antioxidant enzymatic activities as well as the antioxidants synthesis molecules such as ascorbic acid and glutathione (GSH).
The transformed potato plants with the 2-cysteine peroxiredoxin with stress-inducible SWPA2 promoter or under the constitutive CaMV 35S promoter regulation showed a significant increase of tolerance to abiotic stresses, also including high temperatures and MV-induced oxidative stress. The 2-cysteine peroxiredoxin plants under the SWPA2 stress-inducible promoter exhibited the best tolerance [52].
Response to oxidative stress
The cellular production of Reactive O2 species (ROS) are not restricted to an specific physiological status. Plants are the exception and have established physiological mechanisms to counteract ROS accumulation through a tight synthesis regulation and a plethora of enzymes for their remotion. The superoxide dismutases (SODs) in the cell are undoubtedly the first line of defence to prevent ROS accumulation.
The superoxidase dismutase 1 gene is essential in potato response to low temperatures, since its downregulation, through interference RNA, gave rise to a decrease in low-temperature tolerance and its overexpression impacts positively the tolerance to this stress in potatoes [53]. Furthermore, the SOD from Potentilla atrosanguinea (PaSOD) when is overexpressed in potato (S. tuberosum ssp. tuberosum L. cv. Kufri Sutlej) conferred a significant enhancement of the net photosynthetic rates (PN), as well as the stomatal conductance (gs) increase respect to wild-type plants (Pal et al. 2013). Interestingly, potato plants transformed to overexpress SOD, ascorbate peroxidase, and choline oxidase underwent an increase in the lignification process, starch synthesis, and several abiotic stress tolerance enhancement [54,55].
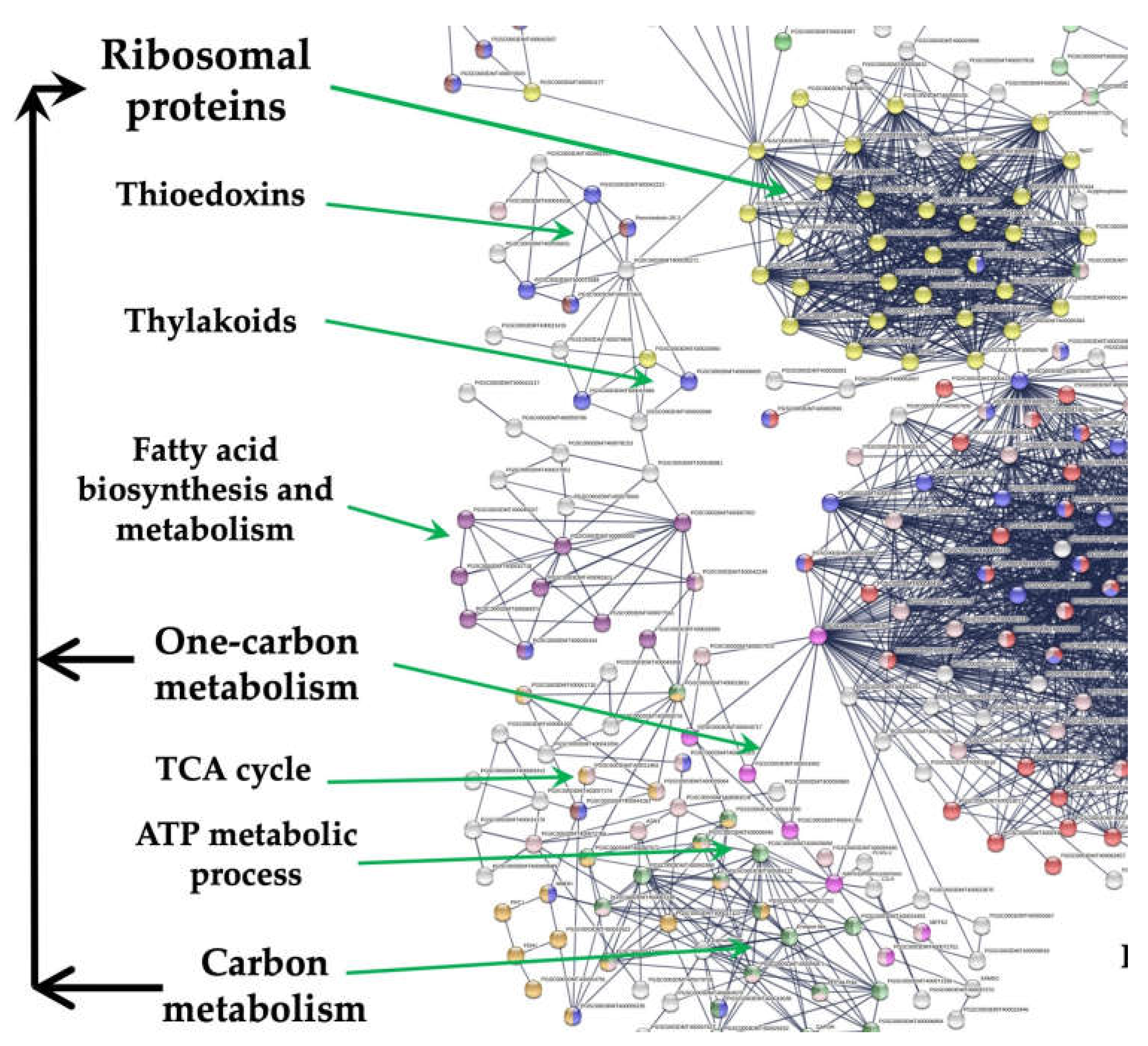
RPs interacting with Carbon metabolism, one-carbon metabolism, TCA-cycle.
In the PPI network, 38 genes were involved in carbon metabolism, 7 in one-carbon metabolism, and 6 in the citrate cycle. This cluster interacts with the CC cluster through the interaction of the DHFR and RPs.
Carbon metabolism
Carbon metabolism (CM) transforms carbon into energy in different amounts through glycolysis, gluconeogenesis, the pentose phosphate pathway, carbon fixation pathways, and the TCA pathway.
Watkinson et al. 2006 [56] assessed three accessions-genotypes of S. tuberosum ssp. Andigena under drought stress conditions and through a transcriptomic analysis determined the relevance of carbon metabolism, citrate cycle and oxidative stress gene activation. In agreement with their results, we found similar genes compared with “intermediate” genotypes, with 20 genes upregulated under stress conditions, similar to our analysis. The term “intermediate” means plants that recovered the photosynthetic index after one cycle of stress and performed even better in the second, yielding similar results to the control plants unaffected by the stress. This may explain why molecular mechanisms involved in field conditions are very similar to those under in vitro conditions.
This includes PGK, Phosphoglycerate kinase, cytosolic; TPI, Triosephosphate isomerase, cytosolic; IAR4, Pyruvate dehydrogenase e1, GAPC2, Glyceraldehyde 3-phosphate dehydrogenase; LOS2, Enolase and PKP3, Pyruvate kinase. However, they showed that genes like Hexokinase-1 and Fructose-bisphosphate aldolase were down-regulated. In contrast, IAR4 did not show significant changes in expression; otherwise, it did not happen when we induced stress. Regarding the TCA cycle, we found that the up-regulated genes, like Watkinson et al. 2006 [56], are MMDH1, Malate dehydrogenase and MDH, NAD-malate dehydrogenase. Overexpression of these genes have shown their agricultural relevance in different plant breeding programs. The phosphoglycerate kinase gene promotes biomass and yields in tobacco under salt stress conditions [57], while in rice, it improves thermotolerance [58]. Similarly, the overexpression of the pyruvate dehydrogenase in rice and barley plants impacted positively in drought stress tolerance, and grain size and weight [59,60,61]. Moreover, the overexpression of pyruvate kinase negatively affects root growth in maize [62], contrasting when it is silenced, in rice leads to sucrose translocation defects and grain filling inhibition [63] and decreases grain starch content [64]. Additionally, the triosephosphate isomerase gene overexpression in rice, pigeon peas, and maize plants enhance photosynthesis under elevated CO2 levels [58], increases salt stress resilience, as well as better drought stress tolerance [65,66,67].
Enolase contributes to salt stress tolerance, as determined in Mesembryanthemum crystallinum L. Likewise, the presence of the fructose-bisphosphate aldose gene is associated with salt stress response in Brassica napus [68] and tobacco [69], as well as biomass accumulation in tobacco [70]. The acetyl-CoA carboxylase overexpression leads to increased lipid content in microalgae, including Dunaliella sp. [71], Chlamydomonas reinhardtii [72], and Scenedesmus sp. [73], and increased seed yield in tobacco [74].
The sucrose phosphate synthase overexpression enhances growth, thermotolerance [75,76], sink strength in tomatoes [77], potato yield characteristics [78], and cold tolerance in Arabidopsis [79]. It also positively impacts biomass production in sugarcane [80] and in tomatoes under saturating light and CO2 conditions [81] and also enhances foliar sucrose/starch levels in Arabidopsis [82]. Similarly, the ATP-citrate synthase gene is directly involved in the salt stress response in Halogeton glomeratus [83] and sugar beet [84].
The glyceraldehyde 3-phosphate dehydrogenase overexpression enhances drought tolerance in potato [85] and salt tolerance in soybean [86], rice [87], and potato [88]. The 6-phosphogluconate dehydrogenase overexpression gene contributes to resistance against Nilaparvata lugens in rice [89], as well as salt tolerance in barley [90], and is also involved in starch accumulation in maize [91]. Additionally, malate dehydrogenase gene overexpression boosts organic acid synthesis and aluminium tolerance in alfalfa [92], salt tolerance in rice [93], apple, and tomato, along with cold tolerance [94,95]. However, it is worth noting that the gene is embryo-lethal in Arabidopsis [96].
Lastly, the overexpression of the phosphoenolpyruvate carboxylase gene increases photosynthetic efficiency in rice [97], fatty acid production in N. tabacum [98], protein content in Vicia narbonensis [99], and affects dark and light respiration in potato [100].
In cerium stress-treated microalgae, Nannocloropsis oculata gave rise to lipid accumulation with the carbon metabolism prominently activated and ribosome biogenesis genes [101].
In summary, overexpressing specific genes for CM in different plant species improves stress tolerance, yield, biomass production, and metabolic processes.
One-carbon-metabolism
One-carbon metabolism is a metabolic process in which multiple enzymatic reactions provide methyl groups (one carbon) for nucleotide metabolism, purine and pyrimidine synthesis, and amino acid metabolism. These effects involve many cellular activities, such as cell growth, differentiation, and development. One-carbon metabolism carries out the mobilization of a carbon unit to the tetrahydrofolate (THF) from serine or glycine to yield methylene-THF for DNA synthesis.
The S-adenosylmethionine synthetase (SHM4 and SHM1) overexpression enhances cold and salt tolerance in tobacco [102, 103], lipid production in Chlamydomonas [104], salt, H2O2 and drought tolerance in Arabidopsis [105, 106 Kim et al. 2015), and alkali tolerance in tomato [107]. The adenosylhomocysteinase gene (HOG1), when overexpressed, affects early flowering and reduced biomass in Arabidopsis [108], and it increases lycopene and reduces the ripening time in tomatoes [109]. Furthermore, the overexpression of the serine hydroxymethyltransferase gene (MAT3) confers salt tolerance [110], increases root growth and sugar levels and decreases H2O2 levels in Arabidopsis [111], cold tolerance [112] and antioxidant ability in rice [113].
5. Conclusions
1.- Gene modulation of ribosomal proteins by genome edition mediated by CRISPR/Cas technology can potentially enhance potato challenge to stress and increase nutritional value.
2.- The sets of genes involved in carbon metabolism: PGK, TPI, IAR4, GAPC2, LOS2, and PKP3 that match with plant field experiments and our conditions have the potential to improve biomass and yield.
3.- The cluster of immunophilins and disulfide isomerase interacting with RPs will allow the activation of alternative mechanisms for survival enhancement under adverse conditions.
4.- The gene modulation of one-carbon metabolism pathways will favour survival under adverse environments.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
E.V.-L. and L.H.-I.: Conceptualization, methodology, validation, formal analysis, investigation, data curation, writing original draft preparation, review and editing, funding acquisition; J.A.F.-L. and O.S.R.-M.: Methodology, software, validation, investigation; A.B.: Software, data curation, formal analysis, supervision, writing, review and editing; J.L.C.-P.: Conceptualization, methodology, validation, formal analysis, investigation, data curation, writing original draft preparation, review and editing, funding acquisition, writing, review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded, in part, by IPN (project SIP 20222061, proyectos de desarrollo tecnológico o inovación para alumnos del IPN # 120).
Acknowledgments
We are very grateful to MSc Diana Marcela Rivera-Toro and BSc Alex Ricardo Bermudez-Valle for the academic debate during the writing of this manuscript.
Conflicts of Interest
The authors declare No conflict of interest.
References
- Food, E. I. T. (2019). About EIT food.
- Dutt, S.; Manjul, A.S.; Raigond, P.; Singh, B.; Siddappa, S.; Bhardwaj, V.; Kawar, P.G.; Patil, V.U.; Kardile, H.B. Key players associated with tuberization in potato: potential candidates for genetic engineering. Crit. Rev. Biotechnol. 2017, 37, 942–957. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, D. J., Coleman, W. K., & Coleman, S. E. (2003). Potato microtuber production and performance: A review. American journal of potato research, 80, 103-115.
- Hannapel, D. J. (2007). Signalling the induction of tuber formation. In Potato biology and biotechnology (pp. 237-256). Elsevier Science BV. [CrossRef]
- Vinterhalter, D., Dragicevic, I., & Vinterhalter, B. (2008). Potato in vitro culture techniques and biotech-nology. Fruit, vegetable and cereal science and biotechnology, 2(1), 16-45.
- Herrera-Isidron, L., Valencia-Lozano, E., Rosiles-Loeza, P. Y., Robles-Hernández, M. G., Napsuciale-Heredia, A., & Cabrera-Ponce, J. L. (2021). Gene expression analysis of microtubers of potato Solanum tu-berosum L. induced in cytokinin containing medium and osmotic stress. Plants, 10(5), 876.
- Valencia-Lozano, E.; Herrera-Isidrón, L.; Flores-López, J.A.; Recoder-Meléndez, O.S.; Barraza, A.; Cabre-ra-Ponce, J.L. Solanum tuberosum Microtuber Development under Darkness Unveiled through RNAseq Transcriptomic Analysis. Int. J. Mol. Sci. 2022, 23, 13835. [Google Scholar] [CrossRef] [PubMed]
- Purwestri, Y. A., Susanto, F. A., & Tsuji, H. (2017). Hd3a florigen recruits different proteins to reveal its function in plant growth and development. In Plant Engineering. IntechOpen. [CrossRef]
- Wang, E., Liu, T., Sun, X., Jing, S., Zhou, T., Liu, T., & Song, B. (2022). Profiling of the Candidate Interact-ing Proteins of SELF-PRUNING 6A (SP6A) in Solanum tuberosum. International Journal of Molecular Scien-ces, 23(16), 9126.
- Abelenda, J. A. , Navarro, C., & Prat, S. (2011). From the model to the crop: genes controlling tuber for-mation in potato. Current Opinion in Biotechnology, 22(2), 287-292.
- Abelenda, J. A., Bergonzi, S., Oortwijn, M., Sonnewald, S., Du, M., Visser, R. G., ... & Bachem, C. W. (2019). Source-sink regulation is mediated by interaction of an FT homolog with a SWEET protein in po-tato. Current Biology, 29(7), 1178-1186.
- Teo, C.-J.; Takahashi, K.; Shimizu, K.; Shimamoto, K.; Taoka, K.-I. Potato Tuber Induction is Regulated by Interactions Between Components of a Tuberigen Complex. Plant Cell Physiol. 2016, 58, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Lin, T.; Hannapel, D.J. Targets of the StBEL5 Transcription Factor Include the FT Ortholog StSP6A. Plant Physiol. 2015, 170, 310–324. [Google Scholar] [CrossRef]
- Salvato, F.; Havelund, J.F.; Chen, M.; Rao, R.S.P.; Rogowska-Wrzesinska, A.; Jensen, O.N.; Gang, D.R.; Thelen, J.J.; Møller, I.M. The Potato Tuber Mitochondrial Proteome. Plant Physiol. 2013, 164, 637–653. [Google Scholar] [CrossRef] [PubMed]
- van Dijk, J.P.; Cankar, K.; Scheffer, S.J.; Beenen, H.G.; Shepherd, L.V.T.; Stewart, D.; Davies, H.V.; Wilko-ckson, S.J.; Leifert, C.; Gruden, K.; et al. Transcriptome Analysis of Potato TubersEffects of Different Agricultural Practices. J. Agric. Food Chem. 2009, 57, 1612–1623. [Google Scholar] [CrossRef]
- Shan, J.; Song, W.; Zhou, J.; Wang, X.; Xie, C.; Gao, X.; Xie, T.; Liu, J. Transcriptome analysis reveals novel genes potentially involved in photoperiodic tuberization in potato. Genomics 2013, 102, 388–396. [Google Scholar] [CrossRef]
- Vulavala, V. K., Fogelman, E., Faigenboim, A., Shoseyov, O., & Ginzberg, I. (2019). The transcriptome of potato tuber phellogen reveals cellular functions of cork cambium and genes involved in periderm for-mation and maturation. Scientific reports, 9(1), 10216.
- Zhou, K., Zhang, C., Xia, J., Yun, P., Wang, Y., Ma, T., & Li, Z. (2021). Albino seedling lethality 4; Chloro-plast 30S ribosomal protein S1 is required for chloroplast ribosome biogenesis and early chloroplast de-velopment in rice. Rice, 14(1), 1-12.
- Gong, X.; Jiang, Q.; Xu, J.; Zhang, J.; Teng, S.; Lin, D.; Dong, Y. Disruption of the Rice Plastid Ribosomal Protein S20 Leads to Chloroplast Developmental Defects and Seedling Lethality. G3 Ge-nes|Genomes|Genetics 2013, 3, 1769–1777. [Google Scholar] [CrossRef]
- Ma, Z.; Dooner, H.K. A mutation in the nuclear-encoded plastid ribosomal protein S9 leads to early em-bryo lethality in maize. Plant J. 2004, 37, 92–103. [Google Scholar] [CrossRef]
- Schultes, N.P.; Sawers, R.J.H.; Brutnell, T.P.; Krueger, R.W. Maize high chlorophyll fluorescent 60 muta-tion is caused by an Ac disruption of the gene encoding the chloroplast ribosomal small subunit protein 17. Plant J. 2000, 21, 317–327. [Google Scholar] [CrossRef]
- Tsugeki, R.; Kochieva, E.Z.; Fedoroff, N.V. A transposon insertion in the Arabidopsis SSR16 gene causes an embryo-defective lethal mutation. Plant J. 1996, 10, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Revenkova, E.; Masson, J.; Koncz, C.; Afsar, K.; Jakovleva, L.; Paszkowski, J. Involvement of Arabidopsis thaliana ribosomal protein S27 in mRNA degradation triggered by genotoxic stress. EMBO J. 1999, 18, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Rogalski, M. , Ruf, S. ( 34(16), 4537–4545. [PubMed]
- Lee, J.; Jang, S.; Ryu, S.; Lee, S.; Park, J.; Lee, S.; An, G.; Park, S.K. Mutation of Plastid Ribosomal Protein L13 Results in an Albino Seedling-Lethal Phenotype in Rice. Plant Breed. Biotechnol. 2019, 7, 395–404. [Google Scholar] [CrossRef]
- Lee, J., Jang, S., Ryu, S., Lee, S., Park, J., Lee, S., ... & Park, S. K. (2019). Impaired plastid ribosomal pro-tein L3 causes albino seedling lethal phenotype in rice. Journal of Plant Biology, 62, 419-428.
- Zhao, D. S., Zhang, C. Q., Li, Q. F., Yang, Q. Q., Gu, M. H., & Liu, Q. Q. (2016). A residue substitution in the plastid ribosomal protein L12/AL1 produces defective plastid ribosome and causes early seedling le-thality in rice. Plant molecular biology, 91, 161-177.
- Lin, D., Jiang, Q., Zheng, K., Chen, S., Zhou, H., Gong, X., ... & Dong, Y. (2015). Mutation of the rice ASL 2 gene encoding plastid ribosomal protein L21 causes chloroplast developmental defects and seedling death. Plant biology, 17(3), 599-607.
- Dupouy, G.; McDermott, E.; Cashell, R.; Scian, A.; McHale, M.; Ryder, P.; de Groot, J.; Lucca, N.; Brych-kova, G.; McKeown, P.C.; et al. Plastid ribosome protein L5 is essential for post-globular embryo develo-pment in Arabidopsis thaliana. Plant Reprod. 2022, 35, 189–204. [Google Scholar] [CrossRef] [PubMed]
- Devis, D., Firth, S. M., Liang, Z., & Byrne, M. E. (2015). Dosage sensitivity of RPL9 and concerted evolu-tion of ribosomal protein genes in plants. Frontiers in Plant Science, 6, 1102.
- Ferreyra, M. L. F., Pezza, A., Biarc, J., Burlingame, A. L., & Casati, P. (2010). Plant L10 ribosomal proteins have different roles during development and translation under ultraviolet-B stress. Plant physiology, 153(4), 1878-1894.
- Pesaresi, P.; Varotto, C.; Meurer, J.; Jahns, P.; Salamini, F.; Leister, D. Knock-out of the plastid ribosomal protein L11 in Arabidopsis: effects on mRNA translation and photosynthesis. Plant J. 2001, 27, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Yin, J., Ibrahim, S., Petersen, F., & Yu, X. (2021). Autoimmunomic signatures of aging and age-related neurodegenerative diseases are associated with brain function and ribosomal proteins. Frontiers in Aging Neuroscience, 13, 679688.
- Romani, I.; Tadini, L.; Rossi, F.; Masiero, S.; Pribil, M.; Jahns, P.; Kater, M.; Leister, D.; Pesaresi, P. Versati-le roles of Arabidopsis plastid ribosomal proteins in plant growth and development. Plant J. 2012, 72, 922–934. [Google Scholar] [CrossRef]
- Horváth, B.M.; Magyar, Z.; Zhang, Y.; Hamburger, A.W.; Bakó, L.; Visser, R.G.F.; Bachem, C.W.B.; Bögre, L. EBP1 regulates organ size through cell growth and proliferation in plants. EMBO J. 2006, 25, 4909–4920. [Google Scholar] [CrossRef]
- Yang, L.; Xie, C.; Li, W.; Zhang, R.; Jue, D.; Yang, Q. Expression of a wild eggplant ribosomal protein L13a in potato enhances resistance to Verticillium dahliae. Plant Cell, Tissue Organ Cult. (PCTOC) 2013, 115, 329–340. [Google Scholar] [CrossRef]
- Moin, M.; Saha, A.; Bakshi, A.; Madhav, M.; Kirti, P. Constitutive expression of Ribosomal Protein L6 modulates salt tolerance in rice transgenic plants. Gene 2021, 789, 145670. [Google Scholar] [CrossRef] [PubMed]
- Moin, M.; Bakshi, A.; Madhav, M.S.; Kirti, P.B. Expression Profiling of Ribosomal Protein Gene Family in Dehydration Stress Responses and Characterization of Transgenic Rice Plants Overexpressing RPL23A for Water-Use Efficiency and Tolerance to Drought and Salt Stresses. Front. Chem. 2017, 5, 97. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, S.; Yanagisawa, S. Ribosome biogenesis factor OLI2 and its interactor BRX1-2 are associated with morphogenesis and lifespan extension in Arabidopsis thaliana. Plant Biotechnol. 2021, 38, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Yu, H., Kong, X., Huang, H., Wu, W., Park, J., Yun, D. J., ... & Zhu, J. K. (2020). STCH4/REIL2 confers cold stress tolerance in Arabidopsis by promoting rRNA processing and CBF protein translation. Cell Reports, 30(1), 229-242.
- Kappachery, S.; Yu, J.W.; Baniekal-Hiremath, G.; Park, S.W. Rapid identification of potential drought tolerance genes from Solanum tuberosum by using a yeast functional screening method. Comptes Rendus Biol. 2013, 336, 530–545. [Google Scholar] [CrossRef]
- Lei, Y.; Lu, L.; Liu, H.-Y.; Li, S.; Xing, F.; Chen, L.-L. CRISPR-P: A Web Tool for Synthetic Single-Guide RNA Design of CRISPR-System in Plants. Mol. Plant 2014, 7, 1494–1496. [Google Scholar] [CrossRef]
- Liu, H.; Ding, Y.; Zhou, Y.; Jin, W.; Xie, K.; Chen, L.-L. CRISPR-P 2.0: An Improved CRISPR-Cas9 Tool for Genome Editing in Plants. Mol. Plant 2017, 10, 530–532. [Google Scholar] [CrossRef] [PubMed]
- Lowder, L. G., Zhang, D., Baltes, N. J., Paul III, J. W., Tang, X., Zheng, X., ... & Qi, Y. (2015). A CRISPR/Cas9 toolbox for multiplexed plant genome editing and transcriptional regulation. Plant physio-logy, 169(2), 971-985.
- García-Murillo, L.; Valencia-Lozano, E.; Priego-Ranero, N.A.; Cabrera-Ponce, J.L.; Duarte-Aké, F.P.; Vizuet-De-Rueda, J.C.; Rivera-Toro, D.M.; Herrera-Ubaldo, H.; de Folter, S.; Alvarez-Venegas, R. CRIS-PRa-mediated transcriptional activation of the SlPR-1 gene in edited tomato plants. Plant Sci. 2023, 329, 111617. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Ponce, J. L., López, L., Assad-Garcia, N., Medina-Arevalo, C., Bailey, A. M., & Herrera-Estrella, L. (1997). An efficient particle bombardment system for the genetic transformation of asparagus (Aspara-gus officinalis L.). Plant cell reports, 16, 255-260.
- Mahajan, S.; Pandey, G.K.; Tuteja, N. Calcium- and salt-stress signaling in plants: Shedding light on SOS pathway. Arch. Biochem. Biophys. 2008, 471, 146–158. [Google Scholar] [CrossRef]
- Alavilli, H., Lee, H., Park, M., Yun, D. J., y Lee, B. H. (2018). Enhanced multiple stress tolerance in Ara-bidopsis by overexpression of the polar moss peptidyl prolyl isomerase FKBP12 gene. Plant cell reports, 37(3), 453–465.
- Park, H.J.; Lee, A.; Lee, S.S.; An, D.-J.; Moon, K.-B.; Ahn, J.C.; Kim, H.-S.; Cho, H.S. Overexpression of Golgi Protein CYP21-4s Improves Crop Productivity in Potato and Rice by Increasing the Abundance of Mannosidic Glycoproteins. Front. Plant Sci. 2017, 8, 1250. [Google Scholar] [CrossRef]
- Andème Ondzighi, C., Christopher, D. A., Cho, E. J., Chang, S. C., & Staehelin, L. A. (2008). Arabidopsis protein disulfide isomerase-5 inhibits cysteine proteases during trafficking to vacuoles before pro-grammed cell death of the endothelium in developing seeds. The Plant Cell, 20(8), 2205-222.
- Eggert, E., Obata, T., Gerstenberger, A., Gier, K., Brandt, T., Fernie, A. R., ... & Kühn, C. (2016). A sucrose transporter-interacting protein disulphide isomerase affects redox homeostasis and links sucrose parti-tioning with abiotic stress tolerance. Plant, cell & environment, 39(6), 1366-1380.
- Kim, M. D., Kim, Y. H., Kwon, S. Y., Jang, B. Y., Lee, S. Y., Yun, D. J., ... & Lee, H. S. (2011). Overexpres-sion of 2-cysteine peroxiredoxin enhances tolerance to methyl viologen-mediated oxidative stress and high temperature in potato plants. Plant Physiology and Biochemistry, 49(8), 891-897.
- Che, Y.; Zhang, N.; Zhu, X.; Li, S.; Wang, S.; Si, H. Enhanced tolerance of the transgenic potato plants overexpressing Cu/Zn superoxide dismutase to low temperature. Sci. Hortic. 2020, 261, 108949. [Google Scholar] [CrossRef]
- Shafi, A.; Pal, A.K.; Sharma, V.; Kalia, S.; Kumar, S.; Ahuja, P.S.; Singh, A.K. Transgenic Potato Plants Overexpressing SOD and APX Exhibit Enhanced Lignification and Starch Biosynthesis with Improved Salt Stress Tolerance. Plant Mol. Biol. Rep. 2017, 35, 504–518. [Google Scholar] [CrossRef]
- Ahmad, R., Kim, Y. H., Kim, M. D., Kwon, S. Y., Cho, K., Lee, H. S., & Kwak, S. S. (2010). Simultaneous expression of choline oxidase, superoxide dismutase and ascorbate peroxidase in potato plant chloro-plasts provides synergistically enhanced protection against various abiotic stresses. Physiologia Plantarum, 138(4), 520-533.
- Watkinson, J. I., Hendricks, L., Sioson, A. A., Vasquez-Robinet, C., Stromberg, V., Heath, L. S., ... & Grene, R. (2006). Accessions of Solanum tuberosum ssp. andigena show differences in photosynthetic recovery after drought stress as reflected in gene expression profiles. Plant Science, 171(6), 745-758.
- Joshi, R., Karan, R., Singla-Pareek, S. L., & Pareek, A. (2016). Ectopic expression of Pokkali phospho-glycerate kinase-2 (OsPGK2-P) improves yield in tobacco plants under salinity stress. Plant Cell Reports, 35, 27-41.
- Suzuki, Y., Konno, Y., Takegahara-Tamakawa, Y., Miyake, C., & Makino, A. (2022). Effects of suppression of chloroplast phosphoglycerate kinase on photosynthesis in rice. Photosynthesis Research, 153(1-2), 83-91.
- Lei, J., Teng, X., Wang, Y., Jiang, X., Zhao, H., Zheng, X., Ren, Y., Dong, H., Wang, Y., Duan, E., Zhang, Y., Zhang, W., Yang, H., Chen, X., Chen, R., Zhang, Y., Yu, M., Xu, S., Bao, X., Zhang, P., Liu, S., Tian, Y., Jiang, L., Wang, Y. & Wan, J. (2022). Plastidic pyruvate dehydrogenase complex E1 component subunit Alpha1 is involved in galactolipid biosynthesis required for amyloplast development in rice. Plant Bio-technology Journal, 20(3), 437-453.
- Guo, P., Baum, M., Grando, S., Ceccarelli, S., Bai, G., Li, R., ... & Valkoun, J. (2009). Differentially ex-pressed genes between drought-tolerant and drought-sensitive barley genotypes in response to drought stress during the reproductive stage. Journal Of Experimental Botany, 60(12), 3531-3544.
- Huang, S.; Xin, S.; Xie, G.; Han, J.; Liu, Z.; Wang, B.; Zhang, S.; Wu, Q.; Cheng, X. Mutagenesis reveals that the rice OsMPT3 gene is an important osmotic regulatory factor. Crop. J. 2020, 8, 465–479. [Google Scholar] [CrossRef]
- He, K.; Zhao, Z.; Ren, W.; Chen, Z.; Chen, L.; Chen, F.; Mi, G.; Pan, Q.; Yuan, L. Mining genes regulating root system architecture in maize based on data integration analysis. Theor. Appl. Genet. 2023, 136, 127. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Tu, B.; Yang, W.; Yuan, H.; Li, J.; Guo, L.; Zheng, L.; Chen, W.; Zhu, X.; Wang, Y.; et al. Mitochon-dria-Associated Pyruvate Kinase Complexes Regulate Grain Filling in Rice. Plant Physiol. 2020, 183, 1073–1087. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y., Li, S., Jiao, G., Sheng, Z., Wu, Y., Shao, G., Xie, L., Peng, C., Xu, J., Tang, S., Wei, X. & Hu, P. (2018). Os PK 2 encodes a plastidic pyruvate kinase involved in rice endosperm starch synthesis, com-pound granule formation and grain filling. Plant Biotechnology Journal, 16(11), 1878-1891.
- Awana, M.; Jain, N.; Samota, M.K.; Rani, K.; Kumar, A.; Ray, M.; Gaikwad, K.; Praveen, S.; Singh, N.K.; Singh, A. Protein and gene integration analysis through proteome and transcriptome brings new insight into salt stress tolerance in pigeonpea (Cajanus cajan L. ). Int. J. Biol. Macromol. 2020, 164, 3589–3602. [Google Scholar] [CrossRef] [PubMed]
- Salekdeh, G. H., Siopongco, J., Wade, L. J., Ghareyazie, B., & Bennett, J. (2002). Proteomic analysis of rice leaves during drought stress and recovery. PROTEOMICS: International Edition, 2(9), 1131-1145.
- Riccardi, F., Gazeau, P., de Vienne, D., & Zivy, M. (1998). Protein changes in response to progressive water deficit in maize: quantitative variation and polypeptide identification. Plant physiology, 117(4), 1253-1263.
- Xing, Q.; Bi, G.; Cao, M.; Belcour, A.; Aite, M.; Mo, Z.; Mao, Y. Comparative Transcriptome Analysis Pro-vides Insights into Response of Ulva compressa to Fluctuating Salinity Conditions. J. Phycol. 2021, 57, 1295–1308. [Google Scholar] [CrossRef] [PubMed]
- Perveen, N.; Dinesh, M.R.; Sankaran, M.; Ravishankar, K.V.; Krishnajee, H.G.; Hanur, V.S.; Alamri, S.; Kesawat, M.S.; Irfan, M. Comparative transcriptome analysis provides novel insights into molecular re-sponse of salt-tolerant and sensitive polyembryonic mango genotypes to salinity stress at seedling stage. Front. Plant Sci. 2023, 14. [Google Scholar] [CrossRef]
- Uematsu, K.; Suzuki, N.; Iwamae, T.; Inui, M.; Yukawa, H. Increased fructose 1,6-bisphosphate aldolase in plastids enhances growth and photosynthesis of tobacco plants. J. Exp. Bot. 2012, 63, 3001–3009. [Google Scholar] [CrossRef]
- Talebi, A. F., Tohidfar, M., Bagheri, A., Lyon, S. R., Salehi-Ashtiani, K., & Tabatabaei, M. (2014). Manipu-lation of carbon flux into fatty acid biosynthesis pathway in Dunaliella salina using AccD and ME genes to enhance lipid content and to improve produced biodiesel quality. Biofuel Research Journal, 1(3), 91-97.
- Chen, D. , Yuan, X., Liang, L., Liu, K., Ye, H., Liu, Z.,... & Xue, T. (2019). Overexpression of acetyl-CoA carboxylase increases fatty acid production in the green alga Chlamydomonas reinhardtii. Biotechnology letters, 41, 1133-1145.
- Ma, C.; Ren, H.; Xing, D.; Xie, G.; Ren, N.; Liu, B. Mechanistic understanding towards the effective lipid production of a microalgal mutant strain Scenedesmus sp. Z-4 by the whole genome bioinformation. J. Hazard. Mater. 2019, 375, 115–120. [Google Scholar] [CrossRef]
- Madoka, Y., Tomizawa, K. I., Mizoi, J., Nishida, I., Nagano, Y., & Sasaki, Y. (2002). Chloroplast transfor-mation with modified accD operon increases acetyl-CoA carboxylase and causes extension of leaf longev-ity and increase in seed yield in tobacco. Plant and cell physiology, 43(12), 1518-1525.
- Zhang, Y.; Zeng, D.; Liu, Y.; Zhu, W. SlSPS, a Sucrose Phosphate Synthase Gene, Mediates Plant Growth and Thermotolerance in Tomato. Horticulturae 2022, 8, 491. [Google Scholar] [CrossRef]
- Liu, H., Xiu, Z., Yang, H., Ma, Z., Yang, D., Wang, H., & Tan, B. C. (2022). Maize Shrek1 encodes a WD40 protein that regulates pre-rRNA processing in ribosome biogenesis. The Plant Cell, 34(10), 4028-4044.
- Nguyen-Quoc, B.; N'Tchobo, H.; Foyer, C.H.; Yelle, S. Overexpression of sucrose phosphate synthase increases sucrose unloading in transformed tomato fruit. J. Exp. Bot. 1999, 50, 785–791. [Google Scholar] [CrossRef]
- Ishimaru, K., Hirotsu, N., Kashiwagi, T., Madoka, Y., Nagasuga, K., Ono, K., & Ohsugi, R. (2008). Over-expression of a maize SPS gene improves yield characters of potato under field conditions. Plant Produc-tion Science, 11(1), 104-107.
- Wang, K.; Bai, Z.-Y.; Liang, Q.-Y.; Liu, Q.-L.; Zhang, L.; Pan, Y.-Z.; Liu, G.-L.; Jiang, B.-B.; Zhang, F.; Jia, Y. Transcriptome analysis of chrysanthemum (Dendranthema grandiflorum) in response to low tempera-ture stress. BMC Genom. 2018, 19, 319. [Google Scholar] [CrossRef]
- Anur, R. M., Mufithah, N., Sawitri, W. D., Sakakibara, H., & Sugiharto, B. (2020). Overexpression of su-crose phosphate synthase enhanced sucrose content and biomass production in transgenic sugarcane. Plants, 9(2), 200.
- Galtier, N., Foyer, C. H., Murchie, E., Aired, R., Quick, P., Voelker, T. A., ... & Betsche, T. (1995). Effects of light and atmospheric carbon dioxide enrichment on photosynthesis and carbon partitioning in the leaves of tomato (Lycopersicon esculentum L.) plants over-expressing sucrose phosphate synthase. Journal of Exper-imental Botany, 46(special issue), 1335-1344.
- Signora, L.; Galtier, N.; Skøt, L.; Lucas, H.; Foyer, C.H. Over-expression of sucrose phosphate synthase in Arabidopsis thaliana results in increased foliar sucrose/starch ratios and favours decreased foliar carbo-hydrate accumulation in plants after prolonged growth with CO2 enrichment. J. Exp. Bot. 1998, 49, 669–680. [Google Scholar] [CrossRef]
- Wang, J.; Li, B.; Meng, Y.; Ma, X.; Lai, Y.; Si, E.; Yang, K.; Ren, P.; Shang, X.; Wang, H. Transcriptomic profiling of the salt-stress response in the halophyte Halogeton glomeratus. BMC Genom. 2015, 16, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Stevanato, P.; Lv, C.; Li, R.; Geng, G. Comparative Physiological and Proteomic Analysis of Two Sugar Beet Genotypes with Contrasting Salt Tolerance. J. Agric. Food Chem. 2019, 67, 6056–6073. [Google Scholar] [CrossRef] [PubMed]
- Kappachery, S.; Baniekal-Hiremath, G.; Yu, J.W.; Park, S.W. Effect of over-and under-expression of glyceraldehyde 3-phosphate dehydrogenase on tolerance of plants to water-deficit stress. Plant Cell, Tis-sue Organ Cult. (PCTOC) 2014, 121, 97–107. [Google Scholar] [CrossRef]
- Zhao, X., Hong, H., Wang, J., Zhan, Y., Teng, W., Li, H., ... & Han, Y. (2022). Genome-wide identification and analysis of glyceraldehyde-3-phosphate dehydrogenase family reveals the role of GmGAPDH14 to improve salt tolerance in soybean (Glycine max L.).
- Lim, H.; Hwang, H.; Kim, T.; Kim, S.; Chung, H.; Lee, D.; Kim, S.; Park, S.; Cho, W.; Ji, H.; et al. Tran-scriptomic Analysis of Rice Plants Overexpressing PsGAPDH in Response to Salinity Stress. Genes 2021, 12, 641. [Google Scholar] [CrossRef] [PubMed]
- Jeong, M.-J.; Park, S.-C.; Byun, M.-O. Improvement of Salt Tolerance in Transgenic Potato Plants by Glyceraldehyde-3 Phosphate Dehydrogenase Gene Transfer. Mol. Cells 2001, 12, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Kuai, P.; Ye, M.; Zhou, S.; Lu, J.; Lou, Y. Overexpression of a Cytosolic 6-Phosphogluconate Dehydrogenase Gene Enhances the Resistance of Rice to Nilaparvata lugens. Plants 2020, 9, 1529. [Google Scholar] [CrossRef]
- Witzel, K., Weidner, A., SURABHI, G. K., Varshney, R. K., Kunze, G., BUCK-SORLIN, G. H., ... & MOCK, H. P. (2010). Comparative analysis of the grain proteome fraction in barley genotypes with contrasting sa-linity tolerance during germination. Plant, Cell & Environment, 33(2), 211-222.
- Spielbauer, G., Li, L., Römisch-Margl, L., Do, P. T., Fouquet, R., Fernie, A. R., ... & Settles, A. M. (2013). Chloroplast-localized 6-phosphogluconate dehydrogenase is critical for maize endosperm starch accumu-lation. Journal of experimental botany, 64(8), 2231-2242.
- Tesfaye, M., Temple, S. J., Allan, D. L., Vance, C. P., & Samac, D. A. (2001). Overexpression of malate dehydrogenase in transgenic alfalfa enhances organic acid synthesis and confers tolerance to aluminum. Plant physiology, 127(4), 1836-1844.
- Zhang, Y.; Wang, Y.; Sun, X.; Yuan, J.; Zhao, Z.; Gao, J.; Wen, X.; Tang, F.; Kang, M.; Abliz, B.; et al. Ge-nome-Wide Identification of MDH Family Genes and Their Association with Salt Tolerance in Rice. Plants 2022, 11, 1498. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.-X.; Dong, Q.-L.; Zhai, H.; You, C.-X.; Hao, Y.-J. The functions of an apple cytosolic malate dehy-drogenase gene in growth and tolerance to cold and salt stresses. Plant Physiol. Biochem. 2011, 49, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q. J., Sun, H., Dong, Q. L., Sun, T. Y., Jin, Z. X., Hao, Y. J., & Yao, Y. X. (2016). The enhancement of tolerance to salt and cold stresses by modifying the redox state and salicylic acid content via the cytosolic malate dehydrogenase gene in transgenic apple plants. Plant biotechnology journal, 14(10), 1986-1997.
- Beeler, S.; Liu, H.-C.; Stadler, M.; Schreier, T.; Eicke, S.; Lue, W.-L.; Truernit, E.; Zeeman, S.C.; Chen, J.; Kötting, O. Plastidial NAD-Dependent Malate Dehydrogenase Is Critical for Embryo Development and Heterotrophic Metabolism in Arabidopsis. Plant Physiol. 2014, 164, 1175–1190. [Google Scholar] [CrossRef]
- Behera, D., Swain, A., Karmakar, S., Dash, M., Swain, P., Baig, M. J., & Molla, K. A. (2023). Overexpres-sion of Setaria italica phosphoenolpyruvate carboxylase gene in rice positively impacts photosynthesis and agronomic traits. Plant Physiology and Biochemistry, 194, 169-181.
- Fan, Z.; Li, J.; Lu, M.; Li, X.; Yin, H. Overexpression of phosphoenolpyruvate carboxylase from Jatropha curcas increases fatty acid accumulation in Nicotiana tabacum. Acta Physiol. Plant. 2013, 35, 2269–2279. [Google Scholar] [CrossRef]
- Rolletschek, H.; Borisjuk, L.; Radchuk, R.; Miranda, M.; Heim, U.; Wobus, U.; Weber, H. Seed-specific expression of a bacterial phosphoenolpyruvate carboxylase in Vicia narbonensis increases protein content and improves carbon economy. Plant Biotechnol. J. 2004, 2, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Häusler, R. E., Kleines, M., Uhrig, H., Hirsch, H. J., & Smets, H. (1999). Overexpression of phospho enol pyruvate carboxylase from Corynebacterium glutamicum lowers the CO2 compensation point (Γ*) and enhances dark and light respiration in transgenic potato. Journal of Experimental Botany, 50(336), 1231-1242.
- Wu, D., Hou, Y., Cheng, J., Han, T., Hao, N., Zhang, B., ... & Chen, S. (2022). Transcriptome analysis of lipid metabolism in response to cerium stress in the oleaginous microalga Nannochloropsis oculata. Sci-ence of The Total Environment, 838, 156420.
- Seong, E.S.; Jeon, M.R.; Choi, J.H.; Yoo, J.H.; Lee, J.G.; Na, J.K.; Kim, N.Y.; Yu, C.Y. Overexpression of S-Adenosylmethionine Synthetase Enhances Tolerance to Cold Stress in Tobacco. Russ. J. Plant Physiol. 2020, 67, 242–249. [Google Scholar] [CrossRef]
- Zhu, H.; He, M.; Jahan, M.S.; Wu, J.; Gu, Q.; Shu, S.; Sun, J.; Guo, S. CsCDPK6, a CsSAMS1-Interacting Protein, Affects Polyamine/Ethylene Biosynthesis in Cucumber and Enhances Salt Tolerance by Overex-pression in Tobacco. Int. J. Mol. Sci. 2021, 22, 11133. [Google Scholar] [CrossRef] [PubMed]
- Kim, J. H., Ahn, J. W., Park, E. J., & Choi, J. I. (2023). Overexpression of S-Adenosylmethionine Synthetase in Recombinant Chlamydomonas for Enhanced Lipid Production. Journal of Microbiology and Biotechnolo-gy, 33(3), 310.
- Ma, C.; Wang, Y.; Gu, D.; Nan, J.; Chen, S.; Li, H. Overexpression of S-Adenosyl-l-Methionine Synthetase 2 from Sugar Beet M14 Increased Arabidopsis Tolerance to Salt and Oxidative Stress. Int. J. Mol. Sci. 2017, 18, 847. [Google Scholar] [CrossRef]
- Kim, E. Y., Park, K. Y., Seo, Y. S., & Kim, W. T. (2016). Arabidopsis small rubber particle protein homolog SRPs play dual roles as positive factors for tissue growth and development and in drought stress re-sponses. Plant physiology, 170(4), 2494-2510.
- Gong, B., Li, X., VandenLangenberg, K. M., Wen, D., Sun, S., Wei, M., ... & Wang, X. (2014). Overexpres-sion of S-adenosyl-l-methionine synthetase increased tomato tolerance to alkali stress through polyamine metabolism. Plant biotechnology journal, 12(6), 694-708.
- Godge, M. R., Kumar, D., & Kumar, P. P. (2008). Arabidopsis HOG1 gene and its petunia homolog PET-CBP act as key regulators of yield parameters. Plant cell reports, 27, 1497-1507.
- Yang, Y.; Zhu, G.; Li, R.; Yan, S.; Fu, D.; Zhu, B.; Tian, H.; Luo, Y.; Zhu, H. The RNA Editing Factor SlORRM4 Is Required for Normal Fruit Ripening in Tomato. Plant Physiol. 2017, 175, 1690–1702. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.; Bhoomika, K.; Dubey, R.S. Differential responses of antioxidative defense system to prolonged salinity stress in salt-tolerant and salt-sensitive Indica rice (Oryza sativa L. ) seedlings. Protoplasma 2011, 250, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Xu, D.; Xiang, D.; Jiang, L.; Hu, H. Serine Hydroxymethyltransferase 1 Is Essential for Primary-Root Growth at Low-Sucrose Conditions. Int. J. Mol. Sci. 2022, 23, 4540. [Google Scholar] [CrossRef] [PubMed]
- Fang, T.; Rao, Y.; Wang, M.; Li, Y.; Liu, Y.; Xiong, P.; Zeng, L. Characterization of the SWEET Gene Family in Longan (Dimocarpus longan) and the Role of DlSWEET1 in Cold Tolerance. Int. J. Mol. Sci. 2022, 23, 8914. [Google Scholar] [CrossRef] [PubMed]
- Wang, S., Li, X., Zhu, J., Liu, H., Liu, T., Yu, G., & Shao, M. (2021). Covalent interaction between high hydrostatic pressure-pretreated rice bran protein hydrolysates and ferulic acid: Focus on antioxidant ac-tivities and emulsifying properties. Journal of Agricultural and Food Chemistry, 69(27), 7777-7785.
- Gorelova, V., De Lepeleire, J., Van Daele, J., Pluim, D., Meï, C., Cuypers, A., ... & Van Der Straeten, D. (2017). Dihydrofolate reductase/thymidylate synthase fine-tunes the folate status and controls redox ho-meostasis in plants. The Plant Cell, 29(11), 2831-2853.
- Maniga, A. (2017). Studies on the meristematic and E2F-dependent gene expression in Arabidopsis thali-ana plants. Cellular Biology, 17(9), 5077-5086.
- Albani, D.; Giorgetti, L.; Pitto, L.; Luo, M.; Cantoni, R.M.; Pujada, M.E.; Rotino, G.L.; Cella, R. Prolifera-tion-dependent pattern of expression of a dihydrofolate reductase-thymidylate synthase gene from Dau-cus carota. Eur. J. Histochem. 2005, 49, 107–116. [Google Scholar] [PubMed]

Table 1.
Loss-of-function of ribosomal proteins and chloroplast/ribosome biogenesis factors in plants.
Table 1.
Loss-of-function of ribosomal proteins and chloroplast/ribosome biogenesis factors in plants.
| RPS5A | Ribosomal protein. Arabidopsis | Growth retardation and floral and vascular defects and the recessive, embryo lethality | Weijers et al. 2001 [2] |
| RPS9 | Ribosomal protein. Maize | Embryo lethal | Ma and Dooner, 2004 [3] |
| RPS13A | Ribosomal protein | Aberrant leaf and trichome morphology, retarded root growth and late flowering. | Ito et al. 2000 [4] |
| RPS16 | Ribosomal protein. Arabidopsis | Embryo lethal | Tsugeki et al. 1996 [5] |
| RPS17 | Ribosomal protein. Maize | Seedling lethal | Schultes et al. 2000 [6] |
| RPS18A | Ribosomal protein. Tobacco | Essential for survival, lethal. | Rogalski et al. 2006 [7] |
| RPS20 | Ribosomal protein. Rice | Seedling lethal | Gong et al. 2013 [8] |
| RPS21 | Ribosomal protein. | Decreased leaf pigmentation, plant growth and photosynthetic activity | Morita et al. 2004 [9] |
| RPS22 | Ribosomal protein. Arabidopsis | No detectable alteration in growth | Tiller et al. 2012 [10] |
| RPS23 | Ribosomal protein. Arabidopsis | Light-green phenotype and retarded growth severely disrupted mesophyll differentiation | Tiller et al. 2012 [10] |
| RPS27 | Ribosomal protein. Arabidopsis | Embryo lethal | Revenkova et al. 1999 [11] |
| RPL3 | Ribosomal protein. Rice | Seedling lethal | Lee et al. 2019 [12] |
| RPL5C | Ribosomal. Arabidopsis | Embryo lethal | Dupouy et al. 2022 [13] |
| RPL9C, RPL9D | Ribosomal protein. Arabidopsis | Embryo lethal | Devis et al. 2015 [14] |
| RPL10 | Ribosomal protein. Arabidopsis, Maize | Embryo lethal | Falcone et al. 2010 [15] |
| RPL11 | Ribsomal protein. Arabidopsis | Significantly decreased leaf pigmentation, plant growth and photosynthetic activity | Pesaresi et al. 2001 [16] |
| RPL12 | Ribosomal protein. Rice | Seeling lethal | Zhao et al. 2016 [17] |
| RPL13 | Ribsomal protein. Rice | Embryo lethal | Lee et al. 2019 [18] |
| RPL15C | Ribosomal protein. | Embryo lethal | Bobik et al. 2019 [19] |
| RPL21C | Ribosomal protein. Arabidopsis, Rice | Embryo lethal | Yin et al. 2021 [20] , Lin et al. 2015 [21] |
| RPL23a | Ribosomal protein, ribosome biogenesis, Arabidopsis | RPL23aA RNAi: growth delay, irregularities in morphology of leaves, roots, phyllotaxy and vasculature, and loss of apical dominance | Degenhardt and Bonham-Smith, 2008 [22] |
| RPL24B | Ribosomal protein. Arabidopsis | Auxin-related developmental defects, in cotyledon number and vascularization | Zhou et al. 2010. [23] |
| RPL28-1 | Ribosomal protein. Arabidopsis | Embryo lethal | Romani et al. 2012 [24] |
| RPL35-1 | Ribosomal protein. Maize | Embryo lethal | Magnard et al. 2004 [25] |
| RPS20, RPL1, RPL4, RPL27 and RPL35 | Ribosomal proteins. Arabidopsis | Embryo lethal | Romani et al. 2012 [24] |
| Chloroplast/Ribosome biogenesis factors | |||
| EDD1 (GlyRS9) | Glycyl tRNA synthetase. Arabidopsis | Embryo lethal | Uwer et al. 1998 [26] |
| CFG1, CFG2 | Chloroplast development. Arabidopsis | Seedling lethal | Zhu et al. 2020 [27] |
| DCL-M | Defective chloroplast and leaf-mutable. Tomato | Embryo lethal | Bellaoui et al. 2003 [28] |
| CPN21 | Chaperonin: Tomato, Tobacco | Seed abortion | Hanania et al. 2006 [29] |
| AtBRX-1-1, AtBRX-1-2 | Maturation of the large pre-60S ribosomal subunit | Pointed leaf and delay early growth. | Weis et al. 2015 [30] |
| AtNuc-L1-AtNuc-L2 | Ribosome biogenesis. Arabidopsis | Seedling lethal | Durut et al. 2015 [31] |
| AtTHAL | Nucleolar organization | thal2 embryo lethal | Chen et al. 2016 [32] |
| AtNMD3 | Nuclear export adaptor of 60S pre-ribosome export and maturation | Lethal | Chen et al. 2012 [33] |
| 30 Ribosome biogenesis factors | Pre-rDNA transcription, pre-rRNA processing, modification, folding, and assembly with RPs | Gene disruptions: infertility, embryo lethality, impaired growth and gametophyte development, aberrant cotyledon, leaf and root development | Weis et al. 2015 [34] |
| RID1 | DEAH-box RNA helicase, Pre-mRNA Splicing | rid1-1: abnormalities in shoot and root apical meristem maintenance, leaf and root morphogenesis | Ohtani et al. 2013 [35] |
| TIC32 | Translocon of the inner envelope of chloroplasts | Embryo lethal | Hörmann et al. 2007 [36] |
| ATS2 | Phosphatidic acid as intermediate for chloroplast membrane lipid biosynthesis. | Embryo lethal | Yu et al. 2004 [37] |
| TIC110 | Translocon of the inner envelope of chloroplasts | Embryo lethal | Kovacheva et al. 2005 [38] |
| CHL27 | Chlorophyl biosynthesis | Retarded growth and chloroplast developmental defects | Bang et al. 2008 [39] |
| DG1 | Early chloroplast development | Delayed greening phenotype | Chi et al. 2008 [40] |
| OEP80 | Chloroplast Outer Envelope Protein | Embryo lethal | Patel et al. 2008 [41] |
| EMB5067/AKRP | Embryo development chloroplast protein | Embryo lethal | Garcion et al. 2006 [42] |
| SPC1 | Carotenoid biosynthesis | Embryo lethal | Dong et al. 2007 [43] |
| PDS3 | phytoene desaturase gene, | Embryo lethal | Qian et al. 2007 [44] |
| EMB1303-1 | Chloroplast biogenesis | Embryo lethal | Huang et al. 2009 [45] |
| EMB1211 | Chloroplast biogenesis | Seedling lethality | Liang et al. 2010 [46] |
| BPG2 | Chloroplast protein accumulation induced by Brassinazole | Decreased number of stacked grana thylakoids | Komatsu et al. 2010 [47] |
| 119 Nuclear genes-assoc. w/chloroplast | Embryo deffective mutants/ associated to chloroplast | Embryo lethal | Bryant et al. 2011 [48] |
| IRM | Involved in RNA processing | Embryo lethal | Palm et al. 2019 [49] |
| ZMRH3 | The RH3 DEAD Box Helicase | Embryo lethal | Asakura et al. 2012 [50] |
| HSP90C | Chloroplast biogenesis | Embryo lethal | Inoue et al. 2013 [51] |
| FTSHI4 | Thylakoid membrane-associated protein | Embryo lethal | Lu et al. 2014 [52] |
| RNAJ | Ribonuclease J (RNase J) required for chloroplast and embryo development | Embryo lethal | Chen et al. 2015 [53] |
| DER | Chloroplast ribosomal RNA processing | Embryo lethal | Jeon et al. 2014 [54] |
| Rrp5, Pwp2, Nob1, Enp1 and Noc4 | Ribosome biogenesis factors | Embryo lethal | Missbach et al. 2013 [55] |
| SHREK1 | Ribosome biogenesis factor | Embryo lethal | Liu et al. 2022 [56] |
| NOP2A,NOP2B | tRNA and rRNA methylation profiles | Embryo lethal | Burgess et al. 2015 [57] |
| RH22 | RNA helicase22 | Embryo lethal | Chi et al. 2012 [58] |
| MDN1 | The AAA-ATPase MIDASIN 1 functions in ribosome biogenesis | Embryo lethal | Li et al. 2019 [59] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



