
Submitted:
13 June 2023
Posted:
16 June 2023
You are already at the latest version
Abstract
With the approval of the FDA Modernization Act 2.0, the pharmaceutical industry is poised to expand its research components with a plethora of alternative models, including organ-on-microfluidic chips in pharma and biotechnology, resulting in a personalized approach. Microfluidics opens new possibilities for the study of cell biology, especially for a better understanding of cell-cell interactions and the pathophysiology of neurodegenerative diseases in vitro and use these models to assess the efficacy of novel therapies. These thumb-sized organ-on-a-chip systems have the potential to reduce animal testing and replace simple 2D culture systems. Restoring critical aspects of endothelial-brain immune cell communication in a biomimetic system using microfluidics may accelerate the process of central nervous system (CNS) drug discovery and improve our understanding of the mechanisms of multiple neurodegenerative diseases. These organ-on-chip technologies can be used to optimize drug targets and assess drug efficacy and toxicity in real-time, which can significantly help minimize animal testing requirements, as authorized by the recent FDA Act. Recent advances in modeling cell-to-cell communication in the CNS are described in this review. This Review initially summarizes the fundamental advantages of microfluidic systems in creating a compartmentalized cell culture for the complex three-dimensional architectures of neural tissue cells such as neurons, glial cells, and endothelial cells, and their recapitulation of spatiotemporal biophysicochemical gradients and mechanical microenvironments. Brain endothelial cell-astroglia-on-a-chip models with a focus on neurodegenerative diseases such Alzheimer's disease, Parkinson's disease, and Huntington's disease and amyotrophic lateral sclerosis is introduced. Then, the current limitations of these microfluidic devices and strategies to overcome them are discussed.
Keywords:
astroglia
; endothelial cells
; biomimetic systems
; microfluidics
; neurodegeneration
Introduction
Neurovascular unit and Microfluidics:
Microfluidics is the science and technology of systems that process or manipulate small amounts of fluid to combine biological and sometimes biochemical elements [1]. Before microfluidics was a science, it was closely associated with several other fields that contributed to its rise and development by providing their methods and materials (Figure 1). Understanding the human microenvironment and pathophysiology requires studying how living cells and tissues function, and how they interact with each other [2]. One of the major challenges in developing therapeutics for central nervous system (CNS) disorders is achieving adequate penetration of the blood-brain barrier (BBB) [3]. The BBB, which consists of specialized endothelial cells as well as other components of the neurovascular unit (NVU; pericytes, astrocytes, and neurons) (Figure 2), maintains the proper environment in the CNS by preventing that potentially harmful bloodborne agents, metabolites, drugs, and immune cells enter it. BBB dysfunction has been commonly associated with CNS pathologies [4]. The BBB regulates the movement of essential nutrients, ions and hormones while preventing neurotoxins and pathogens from entering the brain. This helps in maintaining brain homeostasis. The BBB consists of specialized endothelial cells (ECs) connected by connecting complexes such as tight junctions and adherent junctions, and is surrounded by glial cells (such as astrocytes, oligodendrocytes, and microglia), pericytes, neurons, extracellular matrix (ECM), and a basal lamina [5]. However, this complicates the development of drugs that can cross the barrier and reach the brain. Currently, in vivo models are the most used to test therapeutic efficacy because they provide the natural and extremely complex environment of an organ like the brain. However, in addition to ethical concerns, animal testing also has some disadvantages. Most importantly, despite promising results in preclinical studies, many drugs fail in the late stages of development. This could be due to poorly designed preclinical studies or a difference in animal and human response [6]. The BBB plays a vital role in maintaining brain health and protecting it from harmful substances. In vitro BBB models have been developed to improve our understanding of its functions and response to external factors. Traditional static models have limitations in replicating the physiological conditions of the BBB, resulting in poor correlation with real-life data. To overcome this, dynamic in vitro models with medium flow and 3D models with multiple cell types have been developed. These models provide a more realistic representation by incorporating factors like shear stress and cellular interactions. Additionally, microfluidic devices with integrated transendothelial electrical resistance (TEER) sensors allow continuous and non-invasive monitoring of the barrier's integrity. A novel open-microfluidic 3D BBB model can be designed, consisting of a brain unit with astrocytes and pericytes in a 3D hydrogel, and a vascular unit with cerebral endothelial cells. The platform enables parallel measurements under various conditions and induces a gradual and unidirectional flow to enhance the formation of a tight endothelial barrier. The integrated TEER sensor provides real-time monitoring of the barrier's integrity, while transparent electrodes facilitate high-resolution live imaging. This platform successfully replicates the rapid reorganization of the BBB and endothelial cell cytoskeleton observed during ischemic events, offering valuable insights into the underlying mechanisms of BBB disruption. This innovative BBB-on-chip platform serves as an essential tool for studying BBB dynamics and developing therapeutic approaches to stabilize the barrier and reduce neurological damage associated with conditions like stroke [7].
Other testing methods include in vitro studies performed in cell cultures in either a 2D or 3D environment. Cell culture in the commercially available Transwell® insert is an example of a commonly used model for BBB research. This device consists of two chambers separated by a porous membrane that allows cells to be cultivated. A problem with this model is that it is often oversimplified and cannot account for natural physical parameters found in vivo, such as blood flow [8]. As a result, a new frontier of in vitro models known as organs-on-a-chip (OOC) has emerged.
The idea behind OOC is to combine microfluidics and bioengineering to replicate the smallest structure that makes up an organ by culturing cells in a dynamic environment [9]. A BBB-on-chip is still being developed by a number of research organizations [6,7]. The various published models differ in design, materials and cell lines used, but they all attempt to reproduce the basic structure of the BBB, which consists of brain endothelial cells [12]. Polydimethylsiloxane (PDMS), which is elastic and biocompatible, has become the most widely used material for endothelial-astrocytes in the lab-on-a-chip. Endothelial cells are cultured together with other brain-specific cells in some models to make the model more physiologically relevant, since some of these cells are able to enhance the BBB-specific properties of the endothelial cells [9,10]. Particular attention has been paid to a brain-specific cell type called astrocytes, which has the ability to induce BBB-specific properties in the endothelial cells [15]. The cultivation method affects astrocyte function and morphology. When cultured in ordinary plastic dishes, the cells exhibit a state known as reactive astrocytes, which are the main regulators of the brain's inflammatory response [16].
1. Molecular and Cellular BBB Players: Roles of NVU non-endothelial cells in BBB formation and function
The human BBB (hBBB) controls the exchange of chemicals and the transport of immune cells into the CNS, preventing infections. It is the barrier that separates the blood from the brain parenchyma. The extracellular components, namely the extracellular matrix (ECM) and basement membrane as well as cellular components such as endothelial cells, pericytes, astrocytes and microglia form the hBBB. Interactions between these cells, structures and neurons give rise to the complete neurovascular unit (NVU), a functional unit responsible for a dynamic system capable of controlling local blood flow [17].
Astrocytes
Astrocytes are the most common glial cells and are found throughout the brain. They play critical roles in brain homeostasis, cell-to-cell communication, maintenance of the extracellular environment, and endothelial function and they outnumber neurons by almost a factor of five. They tile the entire CNS and perform a variety of critical, intricate tasks in a healthy CNS (Table 1). Reactive astrogliosis, now recognized as a pathologic feature of structural CNS lesions, is the process by which astrocytes respond to various types of CNS trauma [18]. Recently, significant advances have been made in understanding the processes and functions of reactive astrogliosis, as well as the functions of astrocytes in CNS disorders and diseases [19]. A broad molecular arsenal is available to reactive astrocytes, which will now be described. Reactive astrogliosis is now understood as a finely graded continuum of changes taking place in a contextual manner and driven by specific signalling events, rather than a simple all-or-nothing process. These modifications range from transient scarring with reorganization of tissue structure to irreversible changes in gene expression and cell hypertrophy with retention of cell domains and tissue structure. Reactive astrogliosis may contribute to CNS disease or act as an underlying cause of CNS disease by interfering with normal astrocyte function or causing aberrant effects, according to mounting evidence [20]. A polystyrene-based microfluidic device was used to study the effects of a hypoxic and/or nutrient-depleted microenvironment created by ischemic stroke on astrocyte response [21].
Endothelial cells
ECs are mesoderm-derived modified simple squamous epithelial cells that form the walls of blood vessels. A single EC folds over itself to form the arterial lumen in the smallest capillary, but the diameter of major arteries and veins can consist of dozens of ECs [22].
The CNS ECs express two types of transporters. The first type of transporter is an efflux transporter, which is oriented towards the luminal surface and moves many lipophilic molecules away from the cell membrane where they would otherwise diffuse. The second category consists of highly specialized nutrient transporters that allow the passage of certain nutrients through the BBB and into the CNS and the excretion of certain waste products from the CNS into the bloodstream. CNS ECs have more mitochondria than other ECs, which is believed to be necessary for the production of ATP to drive the ion gradients required for transport tasks [23].
Tight junctions
Tight junctions, which are substantial multiprotein complexes maintained by direct contact with astrocytes, connect BBB endothelial cells together [24]. Tight junctions form a continuous, impermeable barrier that regulates the paracellular transport of solutes between endothelial cells. Tight junctions ensure that the BBB has a high transendothelial resistance that can be measured to ensure that the BBB is preventing the free passage of water and solutes. Transmembrane proteins such as claudin, tricellulin, occludin and junctional adhesion molecules (JAMs) are required for the formation of BBB tight junctions and are linked to cytoskeletal filaments via interactions with accessory proteins such as zonula occludens (ZO) proteins [25]. Three tight junction-associated Marvel proteins (TAMPs) have been discovered on the mammalian BBB. These are the proteins tricellulin, MarvelD3 and occludin. Tricellulin plays a key role in controlling paracellular permeability for macromolecules. At the BBB, MarvelD3 appears to serve as a compensating sealant. According to a study, the physiological function of occludin in epithelial barriers involves epithelial differentiation rather than the development of barrier properties. It has also been found that the tight junction composition and operation are significantly regulated by occludin. Occludin has three cytoplasmic domains including an intracellular short turn domain, a long C-terminal domain and a tetra-spanning wonder domain [26]. A study with mutated occludin isoforms in Madin-Darby canine kidney (MDCK) cells revealed that it has two extracellular loops (ECLs) and at least one transmembrane domain that play a crucial role in the selective permeability of tight junctions. The N-terminus of occludin helps maintain the tight junction integrity as evidenced by the discovery that deletion of the N-terminal sequence of occludin increases paracellular permeability and decreases transepithelial/endothelial electrical resistance (TEER). The lateral (cis) homodimerization or oligomerization of the occludin is mediated by the Wonder motif, the cytoplasmic C-terminus and the second extracellular loop. These domains are sensitive to changes in redox conditions because of the disulfide bonds between the cysteine residues on ECL2 and the C-terminal sequence of occludin. The termini of the C-interactions are required for the transport of occludin to the tight junction [27].
Transporters
CNS ECs contain discrete lumenal and abluminal compartments and are highly polarized cells [28]. The limited permeability of the paracellular junctions allows cells to regulate the flow of ions and chemicals between the blood and the brain. The two main types of transporters expressed by CNS ECs are efflux transporters and food transporters. To better understand the external requirements for brain metabolism and function, and to identify targets for drug delivery across the BBB. Multidrug resistance protein (Mdr1) formerly known as permeability glycoprotein, (P-gp), breast cancer resistance protein (BCRP) and multidrug resistance-associated protein (MDRP) are all efflux transporters that use ATP hydrolysis to move their substrates up the concentration gradient [29]. Many of these transporters transport a variety of substrates into the blood compartment and are concentrated on the luminal surface. These transporters can block many tiny lipophilic compounds that would otherwise passively diffuse across the EC membrane due to the wide variety of substrates that can bind to Mdr1 [30]. The ATP-binding cassette (ABC) transporter superfamily of integral membrane proteins is responsible for the ATP-driven translocation of multiple substrates across membranes. The minimally required architecture of ABC transporters consists of two ABC domains (or nucleotide-binding domains) with highly conserved sequence patterns and two transmembrane domains (TMDs) [31].
These core components can be combined with additional domains to confer regulatory activities and a periplasmic binding protein is required for ligand delivery to prokaryotic importers in this family. ABC transporters must bind an ATP molecule to conserved sequence motifs at the junction of the two ABC domains to achieve a catalytically capable nucleotide hydrolysis conformation. As a transporter progresses through the various stages of nucleotide binding and hydrolysis, the interface between the two ABC domains changes from a closed state typical of ATP binding to a more open conformation associated with non-ATP states [32].
There are three distinct sets of folds that have been identified in TMDs that are architecturally distinct. Although the specific folds differ, they all communicate with the helical domains of ABCs via coupling helices, located in the loops between the helices that span the membrane. These interactions link the nucleotide status of the ABC domains and the conformations of the TMDs [33] .
Soluble Lipid Carrier (SLC) Transporters
Many SLC transporters show increased expression in the human BBB. SLC transporters previously unknown to be expressed at the BBB have been discovered [34]. According to a comparative expression analysis, it was shown that drug transporters, which play an important role in renal and hepatic drug clearance, are expressed at the same or higher levels in the BBB [35]. These results suggest a broad spectrum of roles for SLC transporters in the BBB in maintaining homeostasis of vitamins, amino acids, neurotransmitters, and other critical nutrients in the CNS, and the possibility that these transporters could be targeted to deliver drugs to treat to the CNS in neurodegenerative and other CNS diseases. As new SLC genes are discovered and more transporter-directed antibodies become available, future studies with additional probes will be crucial to confirm and extend these results and better elucidate SLC transporter expression [36] (Table 1).

Table 1.
Validation markers for BBB.
| Category | Property | Relevance | Validation | References |
|---|---|---|---|---|
| Tight junctions | Occludin claudin-5 ZO-1 | Transendothelial transport and uptake investigations, tight junction exploration, and cell polarization research | mRNA and protein expression localization | [37] |
| High junctional tightness | TEER and permeability measurements | [38] | ||
| Efflux transporters | P-pg | Drug delivery to/through the BBB, transendothelial transport and absorption studies and toxicity | mRNA and protein expression Cellular uptake or efflux in absence/presence of inhibitors bi-directional transport studies | [39] |
| BCRP | [40] | |||
| Mrp | [41] | |||
| SLC expression | Glut-1 | Drug distribution to/through the BBB: investigations on transendothelial transport and uptake, studies on brain nutrition | mRNA and protein expression – Cellular uptake in absence/presence of inhibitors – transendothelial transport studies | [42] |
| LAT-1 | [43] | |||
| MCT-1 | [44] | |||
| Receptor systems | Transferrin receptor | Findings on brain nutrition and receptor-mediated transport | mRNA and protein expression – transferrin uptake – transendothelial transport of iron | [45] |
| Responsiveness to regulation from NVU cells | Induction by astrocytes | Review on NVU signalling and cell regulation | Regulation of TEER, P-glycoprotein expression and cell morphology | [46] |
| Induction by pericytes | Regulation of TEER, proteins involved in vesicular transport | [47] |
Extracellular matrix proteins
ECM proteins, which play both structural and modulatory roles in the BBB in addition to cellular participants, are essential components of the barrier. ECM proteins in the basement membrane (BM) of the micro-vessels in the CNS, located between the ECs and the astrocytic end-feet, affect BBB functions [48]. Extracellular matrices, known as BMs, are attached to cell surfaces. They support vascular ECs and epithelia and form a sheet-like structure. Type IV collagen and elastin are structural proteins that make up BMs along with other specialized proteins such as laminins, entactin/nidogen, fibronectin, and vitronectin. The gliovascular basal lamina of the blood-brain barrier consists mainly of laminins. Laminin-mice lack the laminin-211 heterotrimer expressed by astrocytes and pericytes, which tends to result in a defective BBB that allows systemically circulated tracer to enter the brain parenchyma [49]. Significant abnormalities were present in the laminin vascular endothelium, including altered integrity and composition of the basal lamina, misexpression of the embryonic vascular endothelial protein MECA32, significantly reduced pericyte coverage, and aberrant tight junctions. Astrocytic end-feet also lacked correctly polarized aquaporin-4 channels and were hypertrophic [50].
Since preventing dystroglycan expression in neural cells resulted in a similar array of BBB and gliovascular abnormalities, including altered localization of vascular endothelial glucose transporter-1, laminin-211 appears to mediate these effects, at least in part, through interactions with dystroglycan receptors [51]. These results shed light on the cellular and molecular changes resulting from Lama2 mutations or improper post-translational dystroglycan modifications in congenital muscular dystrophies accompanied by seizures and other brain disorders. Laminin-dystroglycan interactions play a unique role in the cooperative integration of astrocytes, endothelial cells, and pericytes in regulation of the BBB, with the typical ultrastructural morphology of tight junction observed between adjacent perineural cells surrounding the minor nerves and between themselves touching perineural cell processes embedded in the tumour stroma neurofibromatosis 1 (NF1) [52]. Immunohistochemistry revealed the presence of claudin-1, claudin- and ZO-1 in the intercellular junctions of a subset of tumour cells [53]. Occludin was mainly found in the perineurium while claudin-5 was concentrated in the blood vessels. A labeling pattern compatible with a perineural cell phenotype can be seen in the claudin-1 positive cells, which were also positive for type IV collagen and epithelial membrane antigen, but not for S-100 protein. The authors showed that cutaneous neurofibromas contain accumulations of perineural cells around the primitive nerves and at the edges of some neurofibromas included clusters of perineural cells around the primitive nerves and at the edges of some neurofibromas using the marker claudin-1 [54].
An organophosphorus substance, chlorpyrifos (CPF), can cross the BBB and disrupt its integrity and function. To identify potential factors contributing to CPF toxicity, an in vitro BBB model using co-cultured neonatal rat astrocytes and bovine microvascular endothelial cells (BMEC) was employed. CPF is metabolized by the BBB, which we theorize inhibits esterase activity and disrupts the BBB. Tight junctions were made by co-culture of BMECs and astrocytes according to electron microscopy, electrical resistance and western blot analysis of two tight junction-associated proteins (ZO-1 and E-cadherin) [55].
2. Microfluidics
2.1. Microfluidic Systems in the Study of Endothelial Astrocyte Interactions
Microfluidics is the science and technology of systems that process or manipulate small amounts of fluid to combine biological and sometimes biochemical elements (Figure 2) [1]. Understanding the human microenvironment and pathophysiology requires studying how living cells and tissue’s function, and how they interact with each other [56].
Fundamental aspects of endothelial glia interactions typically involved 2D culture dishes with and without the support of biomaterials such as hydrogels (to support 3D culture) in which biophysiochemical stimuli were observed. Furthermore, it is often difficult to assess the 3D complexity and function of the human nervous system, as well as to mimic the complex CNS physiological microenvironment, which encompasses cell-to-cell communication and is known to strongly influence CNS physiological function. The traditional 2D culture system, on the other hand, has many limitations, most notably the inability to 3D mimic the cell-interactive microenvironment and other biological features of the nervous system [57]. Table 2 summarizes the advantages and disadvantages of various microfluidic fabrication methods.
In the early 21st century, with the advent of micro-nanofabrication techniques integrated with microfluidics, the limitations of traditional 2D flat cell culture models were overcome, and new techniques were developed to transform the characteristic structure of human or animal tissues or organs beyond the conventional culture model. Standard microfabrication methods such as soft lithography, are used to create microfluidic chip-based systems for neural cell biology and endo-glial models. For example, PDMS has been widely used as a soft lithographic material with advantages such biocompatibility (Table 3). A 3D microfluidic system with a biocompatible biomaterial scaffold (e.g., hydrogels) was developed that can present a 3D microenvironment to cells with well-defined bio-stimuli. Various biomaterials containing ECM components such as collagen, fibrin and Matrigel® can be used in a microfluidic system to mimic brain physiology [65].
Biomimetic platforms have been proposed to study the complex immune system and brain in healthy and neuroinflammatory states. For example, multilayer PDMS channels containing human brain ECs and primary astroglia could be used to mimic neuroimmune communication to model the human endothelial astrocyte [66]. For example, 3D interaction between astrocytes induced pluripotent stem cells (iPSC)-derived neurons and endothelial cells. Recently, a 3D neurovascular hydrogel device was developed to study BBB dysfunction. Another 3D model provides new insights into the study of nanoparticle transport and astrocyte/endothelial interactions [67].
2.2. Astrocyte-Endothelial Interactions and Regulation
Numerous elements specific to the micro-niche of the neurovascular unit maintain and regulate the maintenance of a functional BBB to maturity [68]. Astrocytic BBB-EC interactions are known to control EC morphology, angiogenesis, and barrier phenotype under both normal and pathological conditions. One of these pathways is the glial-derived neurotrophic factor (GDNF), angiopoietin-1 (ANG-1), and Hh signaling cascade, which is known to have a role in embryonic morphogenesis, neuronal guidance, and angiogenesis plays [69]. BBB ECs have the Hh receptor Patched-1, the signal converter Smoothened (Smo) and transcription factors from the Gli family, and astrocytes release Sonic Hh (SH) [70]. Astroglia are multifunctional brain cells that support neuronal synapses and vascular endothelial cells. They act as passive support cells for conducting biological wires and are primarily responsible for the cellular homeostasis of the nervous system. Astrocytes also support and maintain the integrity of the BBB through physical and chemical interactions with endothelial cells and other neural tissue types [71]. Astrocytes also secrete angiogenic substances such as vascular endothelial growth factor (VEGF) that promote vascular growth. Angiopoietins (Ang1) are secreted by perivascular cells including astrocytes and play a crucial role in the intricate process of BBB differentiation by promoting angiogenesis and causing a time-dependent reduction in endothelial permeability. Astrocytes produce angiotensin-converting enzyme-1, an enzyme that converts angiotensin I to angiotensin II and acts on type 1 angiotensin receptors (AT1) expressed by BBB-ECs. Angiotensin II causes vasoconstriction, and activation of AT1 in the CNS reduces BBB permeability and stabilizes the function of connecting proteins by promoting their recruitment to lipid rafts. Occludin is overexpressed at the BBB in angiotensinogen-deficient animals, suggesting that astrocyte-secreted angiotensin II promotes tight junction development [72].
2.3. BBB Function Regulation by Endothelial-Astrocyte-Derived Biochemical Factors
Soluble biochemical factors such as cytokines and chemokines play important roles in the nervous system, as the neuronal wiring and the entire NVU have constant interactions via their paracrine factors that can regulate not only neurodevelopment and inflammation but also neuronal communication. These factors are transported to cells by a variety of physiological conditions, including interstitial fluid, which has a concentration gradient within the connective tissue beneath the basement membrane and is termed the "interstitium" [73]. Several studies suggest that astrocytes play a dual role in controlling BBB function. It was discovered that astroglial connections with blood vessels were lost in an animal model of multiple sclerosis (MS), leading to disruption of the BBB. On the other hand, selective knockout of astrocytic Na+/H+ exchanger isoform 1 reduced astrogliosis after ischemic stroke in mice, resulting in less cerebral vascular damage and improved BBB function [73,74] .
It was also found that ethyl-1-(4-(2,3,3-trichloroacrylamide) phenyl)-5-(trifluoromethyl)-1H-pyrazole-4-carboxylate reduced astrocyte pathology and BBB destruction in intracerebral hemorrhage model rats [76]. As previously mentioned, soluble gradient factors play an important role in the development of both the central and peripheral nervous systems. The microfluidic approach has enabled us to create biochemical factor gradients in spatially defined flow patterns, aiding in the creation of physiologically and/or pathologically relevant fluid states, such as maintaining a constant soluble microenvironment with a high surface area to volume ratio. Furthermore, the ease with which chemical gradients can be generated in a microfluidic approach allows for the dose-response testing of multiple biochemical factors at high content and throughput in the same microfluidic system. For example, various growth factors and chemo-attractants have been generated and maintained using microfluidics under continuous flow and static conditions to study neural stem cell growth and responses [77].
Angiogenesis and vasculogenesis are important biological processes in the nervous system. For example, VEGF or vascular permeability factor (VPF) is one of the most important signaling proteins for blood vessel formation as it has a dramatic impact on endothelial cell-specific signaling events such as proliferation, survival, budding and migration VEGF receptor 2 signaling (VEGFA/VEGFR2) [78]. It is easier to create a linear profile of different growth factors like VEGF and ANG-1 in a microfluidic setup. In vitro experiments have demonstrated the opposite effects of ANG-1 (stabilizing) and VEGF (proliferating) on endothelial cells using these microfluidic devices. This type of microfluidic platform has helped to study interactions and potential synergistic effects of different growth factors and cell types in a variety of physicochemical microenvironments [79].
2.4. Most Common Designs and Features of the Chips for Brain Endothelial-Astrocyte Interactions
Microfluidics is the design or application of devices to impart fluid flow of fluid to channels at least one millimeter wide. Reduced reagent consumption, well-controlled mixing and particle manipulation, assay integration and automation (lab-on-a-chip), and easier imaging and tracking are all advantages of using microfluidic devices. Microchannels, microvalves, and micropumps are examples of miniaturized fluid and flow manipulation components found in microfluidic systems. Microfluidic devices are adaptive technologies, and microfluidic spinning can produce highly controllable, continuously fabricated, cell-embedded fibers. Large sample volumes or stringent polymerization requirements are not necessary for this effective, rapid, and inexpensive approach. Microfluidic BBB technology is an effective method to assess the physiological function of the BBB in vitro and to accelerate drug development to treat brain diseases [80].
The functionality of the final device, biomedical design, endpoint displays, cost and biocompatibility are just a few of the many variables influencing the choice of chip material. Multiple combinations of materials (including plastics, glass, silicone rubber, and resins) are used to create the final OOC/lab-on-a-chip (LOC) device. Glass, silicone rubber such as PDMS and thermoplastics such as polystyrene, poly(methyl methacrylate), polycarbonate or cyclic olefin copolymer are among the most commonly used materials. As a result, it becomes difficult to choose one material as the best, since each material has its own advantages and disadvantages (Table 3). Material selection often represents trade-offs between intended functionality, accessibility to manufacturing resources, and product development phase. Glass is strong and inert, but it is expensive and requires sophisticated processing equipment. Silicon allows complex nanostructures to be fabricated to create on-chip sensors or barrier gratings, but this is costly and time-consuming as clean room equipment is required. Because it's opaque, there are also compatibility issues with traditional inverted microscopes. Although thermoplastics are simple and offer transparency, prototyping complex designs with them is challenging. PDMS is now the material most used for the development of OOC devices because it is so easy to build devices with high resolution micro and nanostructures by duplicate molds in PDMS. The combination of biocompatibility, optical transparency and gas permeability of PDMS makes it the perfect material for biological applications. In addition, the flexibility of PDMS was used to mechanically stimulate cells. Recently, a cutting-edge technology has been proposed that enables fast and high-quality PDMS 3D processing by laser pyrolysis [81].
Unfortunately, this substance is known to adsorb and absorb a variety of (bio)chemicals, which could affect the experimental results, especially in drug testing applications [82].
3. Design Considerations
- The design efficiently mimics the microvascular system of the BBB and to understand the pathophysiological origins of neuro diseases. According to bioengineering theory, an ideal in vitro BBB replica should consist of three main components: A selection of endothelial cells that exhibit the multicellular longitudinal and radial blood vessel architecture. To achieve this, endothelial cells can be ring-shaped or linear in longitudinal section (similar to the radial cross-section of a blood artery). Micropatterning can provide mechanistic information about endothelial cell morphology [83];
- Transwell® chambers that replicate the function of the BBB micro-vessels by maintaining constant and continuous blood flow. The use of a microfluidics-based perfusion system can provide information about disease pathology caused by endothelial functions. In this in vitro NVU, astrocytes maintain apicobasal polarity at the tissue-tissue interface by wrapping their end-feet around the endothelial capillaries on the transluminal surface (i.e., away from the blood vessel lumen) [84].
3.1. Various Approaches for the Integration to Multi-Organ Devices
There are several methods for integrating multiple organs into the device, as schematized in Figure 3. A static system is a collection of tissues that function as a unit (Figure 3A), while a semi-static sone uses Transwell®-based tissue implants connected by a fluid network (Figure 3B). In addition, a flexible system uses micro-connectors to connect platforms designed specifically for individual organs or tissues (Figure 3C).
3.2. Fabrication of Microfluidics Devices: Types of Materials
Besides desirable mechanical and chemical properties, polymers are also inexpensive, easy to manufacture, biocompatible and more flexible [85]. Polymeric materials such as PDMS, polycarbonates, aliphatic polyesters, polystyrene, poly(ethylene terephthalate glycol), poly(methyl methacrylate), polyolefins and others have found use in manufacturing, evaluated by microchips instead of glass (Table 3). Silicon has good chemical and thermal stability and can be used to create complex 2D and 3D microstructures using photolithography and etching techniques, but is difficult to achieve due to its brittleness, high price, opacity, poor electrical insulation, and complicated limited surface properties (Figure 4). The use of PDMS and poly(methyl methacrylate) for biochips is common in both academic and industrial contexts due to their high efficiency and economical manufacturing. Due to problems with management and resolution of surface properties, they are ineffective for other applications such as using hydrophobic chemicals or those requiring stable surface properties [86]. Poly(methyl methacrylate) channel and cover plates (or cover foils) must be glued together to produce microfluidic chips. Poly(methyl methacrylate) channel plates are fabricated by injection molding, laser ablation, in situ polymerization, solvent etching, hot stamping, room temperature embossing and other techniques. Soft lithography is used to build the microfluidic device from PDMS in a single layer. To create channels, the PDMS layer was plasma bonded to glass coverslips. Polypropylene (PP) is the perfect material for biomedical applications due to its inert surface, but because it is soft, prototyping with PP has historically proven difficult as thin sealing foils tend to sink into channels when heated. Because of its low cost and high etch rate, soda-lime glass is an excellent choice for fabricating disposable glass microfluidic devices [87].
Table 3.
List of common materials used in the fabrication of multicellular system-on-a-chip [88].
Table 3.
List of common materials used in the fabrication of multicellular system-on-a-chip [88].
| Material | Advantage | Disadvantage | References |
| Polydimethylsiloxane (PDMS) | Good Gas permeability Good Biocompatibility Good Optical property (Transparency) Good Mechanical property (Elasticity) |
Poor Chemical resistance Expensive |
[89] |
| Poly(methyl methacrylate) (PMMA) | Good Biocompatibility Good Transparency Good Chemical resistance Inexpensive |
Rigid Elasticity | [90] |
| Glass | High Transmittance High processing accuracy |
Extremely fragile Expensive |
[91] |
| Polylactic acid | Biocompatible Transparent Low-cost |
Lower thermal stability | [92] |
| Epoxy resins (Thermosetting polymers) |
Good Biocompatibility Good mechanical Good Chemical resistance Thermal stability |
Expensive Time-consuming |
[93] |
| Polystyrene (Thermoplastic polymer) |
Good Biocompatible Good Transparency |
Poor chemical resistance Poor Elasticity |
[94] |
| Others Polyimide Polycarbonate Cyclic olefin copolymer |
Good Biocompatibility Good Biocompatibility Good Biocompatibility |
Poor Transparency and Elasticity Poor Transparency Poor Transparency |
[95] |
4. In-vitro models of BBB with endothelial astrocytes
Drug development is complicated by the BBB, a unique CNS-specific selective barrier that prevents most substances from reaching the CNS [96]. Innovative BBB in vitro models can shed light on how drugs are delivered to the BBB and how CNS diseases progress [97]. Both the typical dynamic in vitro BBB and the fluidic shear stress are absent from static Transwell® models, as is the thin bilayer interface. Using a dynamic environment and a relatively thin culture membrane (10 m), a microfluidic BBB very similar to the in vivo one to overcome these two disadvantages has been designed [98] .
BBBs were co-cultured with End3 ECs and either C8-D1A astrocytes or not [99] . TEER, permeability experiments, and optical imaging were used to analyse their key features. After ZO-1 imaging, End3 cells had tight junctions that were clearly expressed, good cell survival was demonstrated by live/dead assay, and the astrocytic morphology of C8-D1A cells was confirmed by environmental scanning electron microscope (ESEM) and glial fibrillary acidic protein (GFAP) immunostaining. In BBB co-cultures, TEER values frequently exceeded 250 Ω cm2 on day 3 of endothelial culture, but those in Transwell® co-cultures typically reached 25 Ω cm2 [100]. Time observation of an immediate transient decrease in TEER in response to histamine administration was followed by a recovery, suggesting that the constructed BBB model was stable. These results indicate that the developed BBB system is a trusted model for multiple studies on BBB function and drug delivery [101].
Hydrogels for Modeling
A hydrogel is a water-based gel-like material composed of cross-linked polymers that form a three-dimensional network. It has the capacity to absorb and retain large quantities of water or biological fluids. The cross-linking of polymers within the hydrogel structure allows it to maintain its shape and integrity while holding a substantial amount of water. This unique property enables hydrogels to have a high-water content, often surpassing 90%. Advanced cell cultures offer the opportunity to mimic a more physiologically relevant environment but pose challenges in ECM scaffold design to support a complex brain environment. Both natural and synthetic hydrogels have been tested, each with their own unique advantages and limitations [102]. Genetically engineered hydrogels are reticulated structures of cross-linked polymer chains with very high-water content and exhibit flexibility in mechano-transduction properties in contrast to nanofibrous or microporous scaffolds and they that have undergone genetic modifications or enhancements using techniques from the field of genetic engineering. The advancement of personalized and regenerative medicine is being enhanced by the progress in creating intelligent hydrogels that can respond to different types of stimuli, including biochemical, electrical, thermal, magnetic, light, and pH changes. These hydrogels, when combined with customized ECM formulations, have the potential to facilitate the development of sophisticated brain-on-a-chip (BOC) models (Table 4). Natural polymer hydrogels, which consist of ECM components such as collagen, fibrin, and hyaluronic acid, offer biocompatibility due to the presence of naturally occurring factors necessary for cell viability, proliferation, and tissue development. By utilizing these hydrogels, researchers can create experimental models that mimic the complex environment of the brain, opening up new possibilities for studying neurological processes and developing innovative therapeutic approaches [103].
Beneficial properties of injectable hydrogels include the ability to inject them into the lesion site, adjust lesion geometry, fill the void space, release therapeutics for extended periods of time, and deliver stem cell-based therapies. Thus, injectable hydrogels conform to the cavity, in contrast to biomaterial matrices that are already manufactured and implanted in the tissue. In addition, injectable hydrogels in the liquid state have the advantage that they can be combined with growth factors or therapeutics prior to injection into the tissue and thus serve as drug carriers to promote regenerative processes in the tissue [104].
Endothelial-Astrocytic Interactions in a microfluidic device:
Table 4.
Key Features of Hydrogel Models Containing Endothelial-Astrocytes [105].
Table 4.
Key Features of Hydrogel Models Containing Endothelial-Astrocytes [105].
| Device design | Cells/Co-culture cells | Physiological function | TEER | Fluorescent tracker (probes for permeability assay) | References |
| PDMS | End3 (mouse)/ C8D1A (mouse) |
Co-culture with astrocytes increases BBB integrity | 250–300 Ω cm2 |
1.4 kDa | [106] |
| PDMS sandwich |
hiPS derived BMEC (human)/ primary astrocyte (rat) | Co-culture with astrocyte will enhance BBB integrity | 4000 to 5000 Ω·cm2 | 70 kDa | [107] |
| Transwell® | Endothelial cells directly interfaced with astrocytes via a system of capillaries. | Plays a role in controlling capillary features and BBB permeability | - | 0.16 | [108] |
| 3D ECM gel-based | RBE4, immortalized rat brain micro vessel endothelial cell line | Transmigration of neutrophils | NA | 40 kDa | [109] |
| 3D scaffolds Hydrogel GelMA and PEGDA | hPSC’s | Appropriate mechanical properties and bioactive sites, which are beneficial for cells viability. | - | - | [110] |
| Microfluidics, hydrogel | Human umbilical vein endothelial cells (HUVECs) | Generating a three-dimensional BBB microfluidic platform which presents both structural and functional properties of the BBB in vivo. | - | 10 kDa | [111] |
| PDMS | cEND (immortalized mouse cerebral endothelial cells) | Strong occludin and claudin-5 expression at the tight junctions | 300 -800 | NA | [112] |
5. CNS dysfunction in disorders
BBB disruption is associated with a variety of different neurological disorders, including MS, stroke, Alzheimer's disease (AD), epilepsy, and traumatic brain trauma (Table 5). The tight junctions, transporters, transcytosis, and expression of LAM, among other BBB properties, can all change because of this dysfunction. This breakdown can result in edema, disturbed ionic balance, abnormal signaling, and immune infiltration, all of which can lead to dysregulated neuronal activity and eventual degeneration (Figure 5), despite the fact that CNS dysfunctions typically develops as a consequence of initial injury, BBB failure has occasionally been implicated as an etiology of some diseases, including MS, epilepsy, and AD [113].
ICAM1: Intercellular Adhesion Molecule 1; HLA genes: Human Leukocyte Antigen genes, CD-1A: Cell Surface Protein, FAS: Cell Surface Protein (CD95 or APO-1), FcGR: Fc Gamma Receptors, NOD: Nucleotide-binding Oli-gomerization Domain, TNF-α: Tumour Necrosis Factor alpha & TLR4: Toll like Receptor 4.
Alzheimer's disease
To describe the syndrome, Alois Alzheimer 1906 used pathological indicators (senile plaques and neurofibrillary tangles) together with the criteria of increasing memory loss, disorientation and dementia [114].
The main neuropathological features of AD appear to be senile plaques and neurofibrillary tangles. Before spreading to other cortical regions during the course of the disease, senile plaques appear to originate in brain regions associated with cognition [115].
Amyloid precursor protein (APP) is a transmembrane protein found in neurons and other cells. It goes through proteolytic processing by secretase enzymes, resulting in the creation of various fragments. A peptide is generated from APP by two sequential cleavage processes: one end of the A-peptide is produced by proteolytic activity by secretase, while the other end is produced by secretase using the same mechanism [116].
There seem to be two different types of A: A40, which is shorter, and A42, which is longer. A42 appears to be deposited first and may play a role in initiating the processes that eventually lead to amyloid deposition [117].
The development of neurofibrillary tangles in neurons is a second feature of AD. Specifically, neurofibrillary tangles are formed when tau protein, a protein involved in microtubule formation, undergoes chemical modification (abnormally folded and phosphorylated [118]. Although extremely rare, they account for less than 1% of the cases of three of these specific genetic mutations from chromosome 21, presenilin-1 [PS 1] on chromosome 14 and presenilin-2 [PS 2] on chromosome 1 have been detected in patients with early onset familial autosomal dominant AD [98,99,100]. All of these genes appear to promote cleavage of APP by beta- or gamma-secretase, which appears to increase cellular synthesis of A-42 [120]. The fourth AD gene is apolipoprotein E (APOE), which has three allelic variations (APOE-2 , -3 and -4) each changing in the amino acid exchanged and located on the long arm of chromosome 19 [121].
Only clinically significant results from cholinesterase inhibitors have been achieved in patients with AD. These drugs increase the amount of acetylcholine available for synaptic transmission by inhibiting the enzymes that break it down (i.e., acetylcholinesterase) [122]. Anti-inflammatories, estrogen replacement therapy, and antioxidants are also used to relieve symptoms. The BBB maintains the tightly regulated internal environment of the brain necessary for optimal synaptic and neuronal function and protects neurons from substances present in the systemic circulation. Breakdown of the BBB promotes the entry of neurotoxic blood-derived substances, cells and infections into the brain and is associated with inflammatory and immunological responses that can initiate numerous neurodegenerative pathways. BBB changes in AD include cortical and hippocampal modifications that include capillary basement membrane changes, tight junction protein decreases, and microvascular reductions [123].
The fluorescence in situ hybridization (FISH)-based microfluidic chip platform can detect Alzheimer's disease (AD) at an early stage and is an effective diagnostic tool with low cost, high speed, and high sensitivity [124]. Researchers developed a 3D human AD triculture system that could show the neuroinflammatory role of microglia in AD pathogenesis. In this model, infiltrating microglia migrated faster and induced greater neuronal toxicity/death via a pathway involving interferon and TLR 4. Cho et al. cultured human microglial cells in a microfluidic platform and studied the role of A in microglial accumulation and its response to chemotaxis [125].
A microfluidic concave microwell array system was used to assess the neurotoxic effects of Aβ on the formed gel-free 3D spheroids of rat cortical neuronal progenitor cells with or without fluid flow [126]. A tripartite microfluidic device was used to study how cell proliferation can induce tau aggregation in downstream neurons [127].
Parkinson’s Disease
James Parkinson first described this disease as shaking palsy in 1817. It is a persistent, progressive neurodegenerative condition with both motor and non-motor symptoms [128]. The motor symptoms of Parkinson's disease are thought to be caused by the death of dopaminergic neurons in the striatum, although the presence of non-motor symptoms is demonstrable for neuronal loss at non-dopaminergic sites [129]. The reduced motor function and clinical signs of the disease are caused by the loss of dopaminergic activity in Parkinson's disease (PD). The motor components of the basal ganglia are affected by the disruption of the extrapyramidal system PD [130] .
Histopathological indicators of PD include Lewy bodies (LBs) and loss of pigmented dopaminergic neurons [131].
The role of cytokines and other mediators in the development of PD is the subject of ongoing research. Possible role for inflammatory responses associated with loss of dopaminergic neurons in PD etiology [127,128]. In vitro experiments have shown that damage to dopaminergic neurons causes microglial and astrocyte activation [134]. A variety of molecular signaling pathways may all play a role in the neuro-pathophysiology of PD, a complex neurodegenerative disorder [135] in the subthalamic nucleus includes microvascular degeneration, tight junction reduction and disruption, and changes in the capillary basement membrane [136].
Microfluidic systems enable research into PD with neurobiological relevance by incorporating 3D cell cultures that better represent the complexity of the brain. Also, facilitate high-throughput drug screening and evaluation, particularly by creating microenvironments that mimic the BBB to assess the efficacy of potential therapeutics [137]. Additionally, these systems allow for precise fluid flow control and integration of on-chip sensors and detectors, enabling real-time monitoring and analysis of cellular responses and dopaminergic neuronal activity [138]. Furthermore, the combination of microfluidics with biosensors and lab-on-a-chip devices may lead to the development of personalized diagnostic and monitoring tools for PD that analyze biofluids for disease-specific biomarkers and help in early tracking of disease progression [139].
Researchers developed microfluidic platforms to study PD and associated mitochondrial impairment and synuclein pathology [140]. In one of the platforms, the nigrostriatal circuitry was mimicked, with two compartments housing midbrain dopaminergic neurons and medium-sized striatal spiny neurons. These compartments were connected to a central synaptic chamber via microgrooves, allowing active connectivity between the two neuronal populations. The study conducted on this platform found that the PD-OPA1 mutation resulted in abnormal morphology, reduced mobility, loss of mitochondria and deterioration in functional synapses. In another report, the microfluidic device focused on drug screening for PD and examined the effects of the CLR01 molecular tweezers on dopaminergic neurons. It consisted of two chambers, an insult chamber, and a home chamber, housing induced pluripotent stem cell (iPSC)-derived dopaminergic neurons. The chambers were connected by microgrooves, allowing for axonal separation. Treatment with α-synuclein oligomers in the insult chamber resulted in damage observed in the home chamber, indicating active transport of the oligomers between the two compartments. The presence of CLR01 significantly reduced this damage, demonstrating its neuroprotective properties. Furthermore, the device showed that CLR01 inhibited axonal transport of α-synuclein through interaction with dynein and kinesin, resulting in a reduction in oligomeric pathology of α-synuclein. Both microfluidic platforms provided valuable insights into the mechanisms underlying mitochondrial impairment and α-synuclein pathology in PD. By offering a controlled and interconnected neuronal environment, these platforms facilitated the study of disease-related processes and the screening of potential therapeutic interventions. Ultimately, these advancements contribute to a better understanding of PD pathogenesis and the development of novel treatment strategies [132,133].
Multiple Sclerosis
MS, the most common neurological impairment, is an autoimmune-mediated disease affecting the CNS [142]. MS often leads to severe neurological problems, physical or cognitive impairments in young people [143]. Focal infiltrations of T lymphocytes and macrophages, multifocal zones of inflammation and oligodendrocyte death are the main causes of myelin sheath degradation. Degradation of the myelin sheath occurs in both white and gray matter, leading to the development of CNS plaques composed of inflammatory cells and their byproducts, demyelinated and severed axons, and astrogliosis [144]. CNS white and gray matter tissues become inflamed because of localized infiltration of immune cells and related cytokines, which is the major cause of damage in MS. Interleukin (IL)-12, IL-23, and IL-4 are some of the cytokines produced when pathogen-associated molecules bind to toll-like receptors on antigen-presenting cells (APCs). Microfluidic systems offer valuable tools for studying MS by modeling the BBB, recreating immune cell migration and activation, enabling high-throughput drug screening, detecting disease-specific biomarkers, and supporting personalized medicine approaches. These advancements have the potential to deepen our understanding of MS pathogenesis and contribute to the development of more effective treatments for this complex autoimmune disease [145].
Huntington’s Disease
Huntington's disease (HD), a degenerative brain disorder, is caused by a single defective gene on chromosome 4, one of the 23 human chromosomes that contains a person's complete genetic code [148]. The disease will eventually strike anyone who inherits this dominant defect from a parent who carries the disease. The disease is in honor of Dr. George Huntington, who first recognized the condition in the late 1800s. A defective gene produces the protein known as huntingtin. The protein is known as huntingtin, although its typical function is unknown because studies have linked Huntington's disease to a defective form of the protein. Defective huntingtin protein causes irregular involuntary movements, dramatic loss of thinking and reasoning skills, irritability, depression and other mood problems [149]. HD is mainly identified by uncontrolled movements of the arms, legs, head, face, and upper body. It also affects cognitive functions such as memory, concentration, judgment, and the ability to plan and organize. In the putamen, tight junction protein expression is reduced and disrupted as a result of endothelial damage [150]. Microfluidics systems can be utilized to create microenvironments that mimic the complex cellular interactions and biochemical conditions found in the brain affected by HD. By incorporating patient-derived cells or genetically modified cell lines, researchers can investigate disease mechanisms, study neuronal dysfunction and degeneration, and test potential therapeutic strategies. Microfluidic platforms also allow for high-throughput drug screening and evaluation, enabling the rapid screening of potential therapeutics and optimization of treatment strategies. Additionally, microfluidics can be integrated with biosensors and detection systems to analyze HD-specific biomarkers, aiding in early diagnosis, disease monitoring, and assessing treatment response. Overall, microfluidics provides a versatile and powerful platform for studying Huntington's disease, contributing to a deeper understanding of the disease and the development of effective therapeutic interventions [151]. Vatine et al. used diseased iPSC lines from patients to create an HD-on-a-Chip and study BBB dysfunction [152].
Autism Spectrum Disorder
Challenges in social contact, verbal and nonverbal communication, the occurrence of repetitive behavior, and narrowed interests are features of autism spectrum disorder (ASD) or autism spectrum condition (ASC), a spectrum of neurodevelopmental problems [153]. Two more common symptoms include difficulty making friends and strange responses to sensory input. The three main signs of autism are: (a) a qualitative impairment in social interaction (an inability to relate to others, often without eye contact); (b) stereotypical, ritualistic, repetitive, and restrictive patterns of interests, behaviors and activities; and (c) severe deficiencies in language development and other communication skills.
Human embryonic stem cell (hESC) research has emerged as a promising area of developmental research, offering the opportunity to study early human development in vitro and to gain insight into neurodevelopment. However, progress in understanding neurodevelopmental diseases has been limited by the lack of disease-specific hESCs with genetic alterations associated with disorders such as ASD. To overcome this challenge, induced pluripotent stem cells (iPSCs) were developed using human cells. In this breakthrough technique, the genetic network landscape is reprogrammed into a pluripotent state using a set of transcription factors, offering a simpler and more ethical approach. iPSCs provide a valuable model for the study of complex hereditary and sporadic diseases by allowing the recapitulation of an individual's development in the laboratory. This in vitro model of human neurodevelopment has the potential to reveal specific cellular and biochemical features of human cognition and behavior that are difficult to replicate in other models. In addition, iPSC technology opens up opportunities to manipulate phenotypic changes in drug candidates and paves the way for future drug screening platforms [154]. Microfluidic systems can be utilized for the analysis of biomarkers in blood or cerebrospinal fluid samples, aiding in early diagnosis and monitoring of ASD. The integration of microfluidics with other technologies, such as biosensors and lab-on-a-chip devices, holds promise for personalized diagnostics and treatment strategies in ASD [144,145].
Neurofibromatosis
A neurocutaneous disease called neurofibromatosis is characterized by tumors on the skin and nervous system. The most common forms of neurofibromatosis are types 1 and 2, which are separate diseases. An autosomal dominant disease is von Recklinghausen disease, often known as neurofibromatosis type 1 [146,147]. A condition known as neurofibromatosis type 2 (NF2) is characterized by meningiomas and bilateral vestibular Schwannomas (VS). It is inherited in an autosomal dominant manner [159].
A loss-of-function mutation on the neurofibromin 1 (NF1) gene, either acquired or de novo, leads to neurofibromatosis type 1. This gene codes for neurofibromin and is located on band 17q11.2. A tumor suppressor called neurofibromin acts in the mTOR and RAS/MAPK pathways [160]. Schwann cells, perineural cells, and fibroblasts are among the mixed cell types found in neurofibromas, which are benign tumors. Mast cells, axonal processes and a collagenous extracellular matrix are also present in the tumors [161]. Microfluidics allows for detailed cellular and molecular analysis, including the study of cell migration, proliferation, and differentiation involved in neurofibromatosis pathology. Although further research is needed, microfluidic systems hold promise for advancing our understanding of neurofibromatosis and developing improved diagnostic and therapeutic approaches [162].
Guillain-Barre Syndrome
A rare but severe immune-mediated neuropathy caused by an infection is known as Guillain-Barre syndrome (GBS). Numbness, tingling, and weakness that might progress to paralysis are among the symptoms caused by the autoimmune destruction of peripheral nervous system nerves [163].
The scientific community has developed a variety of study models throughout the years, ranging from genetically engineered animals to two- and three-dimensional cell culture, to examine the functionality of human brain physiology and pathophysiology. These models do, however, have some shortcomings that make it difficult to examine neurodegeneration's pathophysiological characteristics precisely, impeding both therapeutic research and drug development. To correctly replicate neural circuits and characterize the human central nervous system in vitro, compartmentalized microfluidic devices offer simple settings. Our ability to enhance neurodegenerative models and understand the cellular and molecular factors behind the advancement of these issues is made possible by BoC [164] .
Budgets for healthcare and society are significantly impacted by neurodegenerative diseases. Many recent clinical studies for disease-modifying medicines have had a significant failure rate. A major factor in this failure is the challenge of translating findings from studies using cell models based on animals to human patients. 2D Models of newborn mouse neurons and glia have been used for most studies on non-animal neurodegenerative disorders. Since patient-derived 3D brain cell models can now be produced using human stem cell technologies in conjunction with significantly improved microfluidic technology, these systems' usefulness has practically come to an end. These systems have illuminated neural cell function and malfunction [165].
The interactions between different patient-derived brain cell types are made possible by them, and they also offer a micro physiological system that is more like the in vivo brain environment. Yet before these model systems replace 2D rodent models as the foundation of research on neurodegenerative disorders, there are still a lot of challenges to be solved (Table 3). The contrary is true, with these challenges, research will soon move from outmoded models to new patient-derived 3D brain cell systems, which will undoubtedly improve the outcomes of disease therapeutics [166].
Microfluidics: A powerful tool for studying the intricate aspects of rare neurological conditions at a microscale level
Rare diseases encompass a wide range of disorders that can affect almost any tissue, organ system, or biological process in the body. Human-based multi-organ models are flexible in terms of the organ modules, cell types and microfluidics to be included and enable the development of platforms for the recapitulation of highly specific disease states. In rare diseases with a genetic component, iPSC technology can be used to study the effect of relevant mutations or patient-specific cells on relevant tissues [167]. The development of drugs for orphan diseases is hampered by the current problems facing the entire pharmaceutical industry, namely the high costs of bringing new therapeutics to market combined with the low success rate for regulatory approval. Over the past few decades, advances in micro-nano fabrication and neuronal cell biology techniques have enabled the development of micro physiological systems such as OOC and HOC models. These platforms, combined with induced pluripotent stem cell technology, enable the development of patient-specific phenotypic models of rare diseases such rare hemolyticanemia [168]. These technical advances enabled the 2012 NIH and DARPA Micro-physiological Systems for Drug Efficacy and Safety Screening program initiatives and have encouraged the maturation and commercialization of this microfluidic technology [169].
The characteristics of astroglia in more common neuropathologies such as AD and PD have been extensively studied. However, there are still numerous rare neurological disorders in which the involvement of astrocytes and endothelia is unknown and needs further investigation, which is explored in the following review [170]. In addition, the remaining neuropathologies in which the involvement of astrocytes along with endothelial cells have yet to be explored with this technology using patient-derived cells.
Outlook
Microfluidics in neuronal cell biology is still in its infancy, as stringent culture conditions pose a challenge for maintaining and preventing cell death, especially when primary neuroglial cultures are used [171]. In contrast, it provides a controllable microenvironment to study the interactions of glial cells with chemical or physical stimuli, as well as the growth and direct connections of neurons, brain endothelium, dendrites, and synapses [172]. To study the neuromuscular synapse, neurons, glial cells, and muscle cells were combined in another microfluidic model. Another type of in vitro model that effectively replicates the BBB involves the insertion of capillary endothelial cells, basement membrane, neuroglial membrane, and glial podocytes [173]. This review article gives an overview of the applications of microfluidic in vitro systems in the interaction between endothelial cells and immune cells. Despite decades of significant effort, the fundamental processes underlying cell-cell communication and endothelial cell migration are still being identified and often unknown [174]. Both benefit from in vitro reconstitution of in vivo similar physiology and disease [175]. Before describing some of the models offered in the literature, we first outlined the advantages of microfluidic devices in this review. The use of microfluidic devices to explore the effects of biomechanical variables and techniques to study endothelial immune system function were then covered in detail. In order to replicate the entire nervous system, understanding 3D models and knowing how to bio-fabricate vascular and neural networks to create a neurovascular BBB unit and muscle-motor neuron (neuromuscular) connections is crucial. We need multidisciplinary expertise to understand the disease mechanisms [176]. These components can now be integrated using microfluidics into tiny lab-on-a-chips that could serve a range of purposes, from CNS drug discovery [177].
Acknowledgments
M.K. thank the support of the department of pharmaceuticals, Ministry of chemicals & fertilizers, Government of India is gratefully acknowledged. A.S. thanks the support of the Tamara and Harry Handelsman Academic Chair.
References
- Kovarik, M. L.; Gach, P. C.; Ornoff, D. M.; Wang, Y.; Balowski, J.; Farrag, L.; Allbritton, N. L. Micro Total Analysis Systems for Cell Biology and Biochemical Assays. Anal. Chem. 2012, 84, 516–540. [Google Scholar] [CrossRef]
- Yesil-Celiktas, O.; Hassan, S.; Miri, A. K.; Maharjan, S.; Al-kharboosh, R.; Quiñones-Hinojosa, A.; Zhang, Y. S. Mimicking Human Pathophysiology in Organ-on-Chip Devices. Adv. Biosyst. 2018, 2, 1–25. [Google Scholar] [CrossRef]
- Yildirimer, L.; Zhang, Q.; Kuang, S.; Cheung, C. W. J.; Chu, K. A.; He, Y.; Yang, M.; Zhao, X. Engineering Three-Dimensional Microenvironments towards in Vitro Disease Models of the Central Nervous System. Biofabrication 2019, 11. [Google Scholar] [CrossRef]
- Villabona-Rueda, A.; Erice, C.; Pardo, C. A.; Stins, M. F. The Evolving Concept of the Blood Brain Barrier (BBB): From a Single Static Barrier to a Heterogeneous and Dynamic Relay Center. Front. Cell. Neurosci. 2019, 13. [Google Scholar] [CrossRef]
- Obermeier, B.; Daneman, R.; Ransohoff, R. M. Development, Maintenance and Disruption of the Blood-Brain Barrier. Nat. Med. 2013, 19, 1584–1596. [Google Scholar] [CrossRef]
- Lippmann, E. S.; Azarin, S. M.; Kay, J. E.; Nessler, R. A.; Wilson, H. K.; Al-Ahmad, A.; Palecek, S. P.; Shusta, E. V. Derivation of Blood-Brain Barrier Endothelial Cells from Human Pluripotent Stem Cells. Nat. Biotechnol. 2012, 30, 783–791. [Google Scholar] [CrossRef]
- Wei, W.; Cardes, F.; Hierlemann, A.; Modena, M. M. 3D In Vitro Blood-Brain-Barrier Model for Investigating Barrier Insults. Adv. Sci. 2023, 2205752, 1–15. [Google Scholar] [CrossRef]
- Pijuan, J.; Barceló, C.; Moreno, D. F.; Maiques, O.; Sisó, P.; Marti, R. M.; Macià, A.; Panosa, A. In Vitro Cell Migration, Invasion, and Adhesion Assays: From Cell Imaging to Data Analysis. Front. Cell Dev. Biol. 2019, 7, 1–16. [Google Scholar] [CrossRef]
- Leung, C.M.; de Haan, P.; Ronaldson-Bouchard, K.; Kim, G.A.; Ko, J.; Rho, H.S.; Chen, Z.; Habibovic, P.; Jeon, N.L.; Takayama, S.; et al. A Guide to the Organ-on-a-Chip. Nat. Rev. Methods Prim. 2022, 2. [Google Scholar] [CrossRef]
- Stewart, P. A.; Wiley, M. J. Developing Nervous Tissue Induces Formation of Blood-Brain Barrier Characteristics in Invading Endothelial Cells: A Study Using Quail--Chick Transplantation Chimeras. Dev. Biol. 1981, 84, 183–192. [Google Scholar] [CrossRef]
- Janzer, R. C.; Raff, M. C. Astrocytes Induce Blood-Brain Barrier Properties in Endothelial Cells. Nature 1987, 325, 253–257. [Google Scholar] [CrossRef]
- Etc, M.C.S.; Das, C.; Lucia, M.S.; H.K. and T.J. 乳鼠心肌提取 HHS Public Access. Physiol. Behav. 2019, 176, 139–148. [Google Scholar]
- Tontsch, U.; Bauer, H. C. Glial Cells and Neurons Induce Blood-Brain Barrier Related Enzymes in Cultured Cerebral Endothelial Cells. Brain Res. 1991, 539, 247–253. [Google Scholar] [CrossRef]
- Wang, C.-W.; Fischer, W. B. Rotational Dynamics of The Transmembrane Domains Play an Important Role in Peptide Dynamics of Viral Fusion and Ion Channel Forming Proteins-A Molecular Dynamics Simulation Study. Viruses 2022, 14. [Google Scholar] [CrossRef]
- Kubotera, H.; Ikeshima-Kataoka, H.; Hatashita, Y.; Allegra Mascaro, A. L.; Pavone, F. S.; Inoue, T. Astrocytic Endfeet Re-Cover Blood Vessels after Removal by Laser Ablation. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Lin, L.; Yee, S. W.; Kim, R. B.; Giacomini, K. M. SLC Transporters as Therapeutic Targets: Emerging Opportunities. Nat. Rev. Drug Discov. 2015, 14, 543–560. [Google Scholar] [CrossRef]
- Guarino, V.; Zizzari, A.; Bianco, M.; Gigli, G.; Moroni, L.; Arima, V. Advancements in Modelling Human Blood Brain-Barrier on a Chip. Biofabrication 2023, 15. [Google Scholar] [CrossRef]
- Potjewyd, G.; Kellett, K. A. B.; Hooper, N. M. 3D Hydrogel Models of the Neurovascular Unit to Investigate Blood-Brain Barrier Dysfunction. Neuronal Signal. 2021, 5, NS20210027. [Google Scholar] [CrossRef]
- Ge, S.; Pachter, J. S. Isolation and Culture of Microvascular Endothelial Cells from Murine Spinal Cord. J. Neuroimmunol. 2006, 177, (1–2). [Google Scholar] [CrossRef]
- Sofroniew, M. V.; Vinters, H. V. Astrocytes: Biology and Pathology. Acta Neuropathol. 2010, 119, 7. [Google Scholar] [CrossRef]
- Chan, C. Y.; Goral, V. N.; DeRosa, M. E.; Huang, T. J.; Yuen, P. K. A Polystyrene-Based Microfluidic Device with Threedimensional Interconnected Microporous Walls for Perfusion Cell Culture. Biomicrofluidics 2014, 8, 1–7. [Google Scholar] [CrossRef]
- Jiang, R.; Du, X.; Brink, L.; Lönnerdal, B. The Role of Orally Ingested Milk Fat Globule Membrane on Intestinal Barrier Functions Evaluated with a Suckling Rat Pup Supplementation Model and a Human Enterocyte Model. J. Nutr. Biochem. 2022, 108, 109084. [Google Scholar] [CrossRef]
- Middelkamp, H.H.T.; Verboven, A.H.A.; De Sá Vivas, A.G.; Schoenmaker, C.; Klein Gunnewiek, T.M.; Passier, R.; Albers, C.A.; ’t Hoen, P.A.C.; Nadif Kasri, N.; van der Meer, A.D. Cell Type-Specific Changes in Transcriptomic Profiles of Endothelial Cells, IPSC-Derived Neurons and Astrocytes Cultured on Microfluidic Chips. Sci. Rep. 2021, 11, 2281. [Google Scholar] [CrossRef]
- A, L.; C, A.; F, G.; K, G.; P, C. Tight Junctions at the Blood-Brain Barrier: Physiological Architecture and Disease-Associated Dysregulation. Fluids Barriers CNS 2012, 9, 23. [Google Scholar]
- Haseloff, R. F.; Dithmer, S.; Winkler, L.; Wolburg, H.; Blasig, I. E. Transmembrane Proteins of the Tight Junctions at the Blood-Brain Barrier: Structural and Functional Aspects. Semin. Cell Dev. Biol. 2015, 38, 16–25. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, L.; Bao, L.; Liu, J.; Zhu, H.; Lv, Q.; Liu, R.; Chen, W.; Tong, W.; Wei, Q.; et al. SARS-CoV-2 Crosses the Blood-Brain Barrier Accompanied with Basement Membrane Disruption without Tight Junctions Alteration. Signal Transduct. Target. Ther. 2021, 6, 337. [Google Scholar] [CrossRef]
- Lochhead, J. J.; Yang, J.; Ronaldson, P. T.; Davis, T. P. Structure, Function, and Regulation of the Blood-Brain Barrier Tight Junction in Central Nervous System Disorders. Front. Physiol. 2020, 11, 914. [Google Scholar] [CrossRef]
- Visigalli, R.; Rotoli, B. M.; Ferrari, F.; Di Lascia, M.; Riccardi, B.; Puccini, P.; Dall’Asta, V.; Barilli, A. Expression and Function of ABC Transporters in Human Alveolar Epithelial Cells. Biomolecules 2022, 12. [Google Scholar] [CrossRef]
- Rees, D. C.; Johnson, E.; Lewinson, O. ABC Transporters: The Power to Change. Nat. Rev. Mol. Cell Biol. 2009, 10, 218–227. [Google Scholar] [CrossRef]
- De Stefani, D.; Patron, M.; R. R. 乳鼠心肌提取 HHS Public Access. Physiol. Behav. 2015, 176, 139–148. [Google Scholar]
- Xiao, Y.; Zeng, B.; Berner, N.; Frishman, D.; Langosch, D.; Teese, M. G. Experimental Determination and Data-Driven Prediction of Homotypic Transmembrane Domain Interfaces. Comput. Struct. Biotechnol. J. 2020, 18, 3230–3242. [Google Scholar] [CrossRef]
- Wilkens, S. Structure and Mechanism of ABC Transporters. F1000Prime Rep. 2015, 7, 1–9. [Google Scholar] [CrossRef]
- Sharpe, H. J.; Stevens, T. J.; Munro, S. A Comprehensive Comparison of Transmembrane Domains Reveals Organelle-Specific Properties. Cell 2010, 142, 158–169. [Google Scholar] [CrossRef]
- Geier, E. G.; Chen, E. C.; Webb, A.; Papp, A. C.; Yee, S. W.; Sadee, W.; Giacomini, K. M. Profiling Solute Carrier Transporters in the Human Blood-Brain Barrier. Clin. Pharmacol. Ther. 2013, 94, 636–639. [Google Scholar] [CrossRef]
- Mizuno, N.; Niwa, T.; Yotsumoto, Y.; Sugiyama, Y. Impact of Drug Transporter Studies on Drug Discovery and Development. Pharmacol. Rev. 2003, 55, 425–461. [Google Scholar] [CrossRef]
- Hur, J.; Smith-Warner, S.A.; Rimm, E.B.; Willett, W.C.; Wu, K.; Cao, Y. E. G. 乳鼠心肌提取 HHS Public Access. J. Int. Soc. Burn Inj. 2017, 43, 909–932. [Google Scholar]
- Nitta, T.; Hata, M.; Gotoh, S.; Seo, Y.; Sasaki, H.; Hashimoto, N.; Furuse, M.; Tsukita, S. Size-Selective Loosening of the Blood-Brain Barrier in Claudin-5-Deficient Mice. J. Cell Biol. 2003, 161, 653–660. [Google Scholar] [CrossRef]
- Smith, Q. R.; Rapoport, S. I. Cerebrovascular Permeability Coefficients to Sodium, Potassium, and Chloride. J. Neurochem. 1986, 46, 1732–1742. [Google Scholar] [CrossRef]
- Cordon-Cardo, C.; O’Brien, J. P.; Casals, D.; Rittman-Grauer, L.; Biedler, J. L.; Melamed, M. R.; Bertino, J. R. Multidrug-Resistance Gene (P-Glycoprotein) Is Expressed by Endothelial Cells at Blood-Brain Barrier Sites. Proc. Natl. Acad. Sci. U. S. A. 1989, 86, 695–698. [Google Scholar] [CrossRef]
- Zhang, W.; Mojsilovic-Petrovic, J.; Andrade, M. F.; Zhang, H.; Ball, M.; Stanimirovic, D. B. The Expression and Functional Characterization of ABCG2 in Brain Endothelial Cells and Vessels. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2003, 17, 2085–2087. [Google Scholar] [CrossRef]
- Miller, D. S.; Nobmann, S. N.; Gutmann, H.; Toeroek, M.; Drewe, J.; Fricker, G. Xenobiotic Transport across Isolated Brain Microvessels Studied by Confocal Microscopy. Mol. Pharmacol. 2000, 58, 1357–1367. [Google Scholar] [CrossRef]
- Nualart, F.; Godoy, A.; Reinicke, K. Expression of the Hexose Transporters GLUT1 and GLUT2 during the Early Development of the Human Brain. Brain Res. 1999, 824, 97–104. [Google Scholar] [CrossRef]
- Zheng, P.-P.; Romme, E.; van der Spek, P. J.; Dirven, C. M. F.; Willemsen, R.; Kros, J. M. Glut1/SLC2A1 Is Crucial for the Development of the Blood-Brain Barrier in Vivo. Ann. Neurol. 2010, 68, 835–844. [Google Scholar] [CrossRef]
- Gerhart, D. Z.; Enerson, B. E.; Zhdankina, O. Y.; Leino, R. L.; Drewes, L. R. Expression of Monocarboxylate Transporter MCT1 by Brain Endothelium and Glia in Adult and Suckling Rats. Am. J. Physiol. 1997, 273 Pt 1, E207–13. [Google Scholar] [CrossRef]
- Yu, Y.J.; Atwal, J.K.; Zhang, Y.; Tong, R.K.; Wildsmith, K.R.; Tan, C.; Bien-Ly, N.; Hersom, M.; Maloney, J.A.; Meilandt, W.J.; et al. Therapeutic Bispecific Antibodies Cross the Blood-Brain Barrier in Nonhuman Primates. Sci. Transl. Med. 2014, 6, 261ra154. [Google Scholar] [CrossRef]
- Hayashi, Y.; Nomura, M.; Yamagishi, S.; Harada, S.; Yamashita, J.; Yamamoto, H. Induction of Various Blood-Brain Barrier Properties in Non-Neural Endothelial Cells by Close Apposition to Co-Cultured Astrocytes. Glia 1997, 19, 13–26. [Google Scholar]
- Daneman, R.; Zhou, L.; Kebede, A. A.; Barres, B. A. Pericytes Are Required for Blood-Brain Barrier Integrity during Embryogenesis. Nature 2010, 468, 562–566. [Google Scholar] [CrossRef]
- Menezes, M. J.; McClenahan, F. K.; Leiton, C. V; Aranmolate, A.; Shan, X.; Colognato, H. The Extracellular Matrix Protein Laminin A2 Regulates the Maturation and Function of the Blood-Brain Barrier. J. Neurosci. Off. J. Soc. Neurosci. 2014, 34, 15260–15280. [Google Scholar] [CrossRef]
- Barraza-Flores, P.; Bates, C. R.; Oliveira-Santos, A.; Burkin, D. J. Laminin and Integrin in LAMA2-Related Congenital Muscular Dystrophy: From Disease to Therapeutics. Front. Mol. Neurosci. 2020, 13, 1–9. [Google Scholar] [CrossRef]
- Xu, L.; Nirwane, A.; Xu, T.; Kang, M.; Devasani, K.; Yao, Y. Fibroblasts Repair Blood-Brain Barrier Damage and Hemorrhagic Brain Injury via TIMP2. Cell Rep. 2022, 41, 111709. [Google Scholar] [CrossRef] [PubMed]
- Cameron, T.; Bennet, T.; Rowe, E. M.; Anwer, M.; Wellington, C. L.; Cheung, K. C. Review of Design Considerations for Brain-on-a-chip Models. Micromachines 2021, 12. [Google Scholar] [CrossRef]
- Pummi, K. P.; Aho, H. J.; Laato, M. K.; Peltonen, J. T. K.; Peltonen, S. A. Tight Junction Proteins and Perineurial Cells in Neurofibromas. J. Histochem. Cytochem. 2006, 54, 53–61. [Google Scholar] [CrossRef]
- Wilson, E. R.; Della-Flora Nunes, G.; Weaver, M. R.; Frick, L. R.; Feltri, M. L. Schwann Cell Interactions during the Development of the Peripheral Nervous System. Dev. Neurobiol. 2021, 81, 464–489. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, Z.; Jia, J. Mechanisms of Smoothened Regulation in Hedgehog Signaling. Cells 2021, 10. [Google Scholar] [CrossRef]
- Akhshi, T.; Shannon, R.; Trimble, W. S. The Complex Web of Canonical and Non-Canonical Hedgehog Signaling. Bioessays 2022, 44, e2100183. [Google Scholar] [CrossRef]
- Sackmann, E. K.; Fulton, A. L.; Beebe, D. J. The Present and Future Role of Microfluidics in Biomedical Research. Nature 2014, 507, 181–189. [Google Scholar] [CrossRef]
- Wang, J. D.; Khafagy, E.-S.; Khanafer, K.; Takayama, S.; ElSayed, M. E. H. Organization of Endothelial Cells, Pericytes, and Astrocytes into a 3D Microfluidic in Vitro Model of the Blood-Brain Barrier. Mol. Pharm. 2016, 13, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Kim, P.; Kwon, K. W.; Park, M. C.; Lee, S. H.; Kim, S. M.; Suh, K. Y. Soft Lithography for Microfluidics: A Review. Biochip J. 2008, 2, 1–11. [Google Scholar]
- Gong, M. M.; Sinton, D. Turning the Page: Advancing Paper-Based Microfluidics for Broad Diagnostic Application. Chem. Rev. 2017, 117, 8447–8480. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, N. P.; Cabot, J. M.; Smejkal, P.; Guijt, R. M.; Paull, B.; Breadmore, M. C. Comparing Microfluidic Performance of Three-Dimensional (3D) Printing Platforms. Anal. Chem. 2017, 89, 3858–3866. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Cook, B. S.; Fang, Y.; Tentzeris, M. M. Fully Inkjet-Printed Microfluidics: A Solution to Low-Cost Rapid Three-Dimensional Microfluidics Fabrication with Numerous Electrical and Sensing Applications. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, Q.; Gu, M. Three-Dimensional Direct Laser Writing of Biomimetic Neuron Structures. Opt. Express 2018, 26, 32111. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Jiang, H. R. A Review on Continuous-Flow Microfluidic PCR in Droplets: Advances, Challenges and Future. Anal. Chim. Acta 2016, 914, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Guo, M. T.; Rotem, A.; Heyman, J. A.; Weitz, D. A. Droplet Microfluidics for High-Throughput Biological Assays. Lab Chip 2012, 12, 2146–2155. [Google Scholar] [CrossRef]
- Adriani, G.; Ma, D.; Pavesi, A.; Kamm, R. D.; Goh, E. L. K. A 3D Neurovascular Microfluidic Model Consisting of Neurons, Astrocytes and Cerebral Endothelial Cells as a Blood-Brain Barrier. Lab Chip 2017, 17, 448–459. [Google Scholar] [CrossRef]
- Lauranzano, E.; Rasile, M.; Matteoli, M. Integrating Primary Astrocytes in a Microfluidic Model of the Blood-Brain Barrier. Methods Mol. Biol. 2022, 2492, 225–240. [Google Scholar] [CrossRef]
- Sreekanthreddy, P.; Gromnicova, R.; Davies, H.; Phillips, J.; Romero, I. A.; Male, D. A Three-Dimensional Model of the Human Blood-Brain Barrier to Analyse the Transport of Nanoparticles and Astrocyte/Endothelial Interactions. F1000Research 2015, 4, 1279. [Google Scholar] [CrossRef]
- Ding, J.; Li, H.-Y.; Zhang, L.; Zhou, Y.; Wu, J. Hedgehog Signaling, a Critical Pathway Governing the Development and Progression of Hepatocellular Carcinoma. Cells 2021, 10. [Google Scholar] [CrossRef]
- Kim, H.; Leng, K.; Park, J.; Sorets, A.G.; Kim, S.; Shostak, A.; Embalabala, R.J.; Mlouk, K.; Katdare, K.A.; Rose, I.V.L.; et al. Reactive Astrocytes Transduce Inflammation in a Blood-Brain Barrier Model through a TNF-STAT3 Signaling Axis and Secretion of Alpha 1-Antichymotrypsin. Nat. Commun. 2022, 13. [Google Scholar] [CrossRef]
- Yang, H.; Feng, G.-D.; Olivera, C.; Jiao, X.-Y.; Vitale, A.; Gong, J.; You, S.-W. Sonic Hedgehog Released from Scratch-Injured Astrocytes Is a Key Signal Necessary but Not Sufficient for the Astrocyte de-Differentiation. Stem Cell Res. 2012, 9, 156–166. [Google Scholar] [CrossRef]
- Osaki, T.; Shin, Y.; Sivathanu, V.; Campisi, M.; Kamm, R. D. In Vitro Microfluidic Models for Neurodegenerative Disorders. Adv. Healthc. Mater. 2018, 7, 1–29. [Google Scholar] [CrossRef]
- Wosik, K.; Cayrol, R.; Dodelet-Devillers, A.; Berthelet, F.; Bernard, M.; Moumdjian, R.; Bouthillier, A.; Reudelhuber, T. L.; Prat, A. Angiotensin II Controls Occludin Function and Is Required for Blood-Brain Barrier Maintenance: Relevance to Multiple Sclerosis. J. Neurosci. 2007, 27, 9032–9042. [Google Scholar] [CrossRef]
- Kuo, Y. M.; Lee, Y. H. Epoxyeicosatrienoic Acids and Soluble Epoxide Hydrolase in Physiology and Diseases of the Central Nervous System. Chin. J. Physiol. 2022, 65, 1–11. [Google Scholar] [CrossRef]
- Eilam, R.; Segal, M.; Malach, R.; Sela, M.; Arnon, R.; Aharoni, R. Astrocyte Disruption of Neurovascular Communication Is Linked to Cortical Damage in an Animal Model of Multiple Sclerosis. Glia 2018, 66, 1098–1117. [Google Scholar] [CrossRef]
- Wurdeman, S. R.; Stevens, P. M.; Campbell, J. H.; Davie-Smith, F.; Coulter, E.; Kennon, B.; Wyke, S.; Paul, L.; Ingegneria, F.; Magistrale, L.; et al. 乳鼠心肌提取 HHS Public Access. Physiol. Behav. 2017, 176, 498–503. [Google Scholar]
- Michinaga, S.; Koyama, Y. Dual Roles of Astrocyte-Derived Factors in Regulation of Blood-Brain Barrier Function after Brain Damage. Int. J. Mol. Sci. 2019, 20, 1–22. [Google Scholar] [CrossRef]
- Galan, E. A.; Zhao, H.; Wang, X.; Dai, Q.; Huck, W. T. S.; Ma, S. Intelligent Microfluidics: The Convergence of Machine Learning and Microfluidics in Materials Science and Biomedicine. Matter 2020, 3, 1893–1922. [Google Scholar] [CrossRef]
- Nakatsu, M. N.; Sainson, R. C. A.; Pérez-Del-Pulgar, S.; Aoto, J. N.; Aitkenhead, M.; Taylor, K. L.; Carpenter, P. M.; Hughes, C. C. W. VEGF121 and VEGF165 Regulate Blood Vessel Diameter Through Vascular Endothelial Growth Factor Receptor 2 in an in Vitro Angiogenesis Model. Lab. Investig. 2003, 83, 1873–1885. [Google Scholar] [CrossRef]
- Shin, Y.; Jeon, J. S.; Han, S.; Jung, G. S.; Shin, S.; Lee, S. H.; Sudo, R.; Kamm, R. D.; Chung, S. In Vitro 3D Collective Sprouting Angiogenesis under Orchestrated ANG-1 and VEGF Gradients. Lab Chip 2011, 11, 2175–2181. [Google Scholar] [CrossRef] [PubMed]
- Sivanand. 乳鼠心肌提取 HHS Public Access. Physiol. Behav. 2019, 176, 139–148. [Google Scholar]
- Mou, L.; Jiang, X. Materials for Microfluidic Immunoassays: A Review. Adv. Healthc. Mater. 2017, 6, 1–20. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, L.; Chen, G. Fabrication, Modification, and Application of Poly(Methyl Methacrylate) Microfluidic Chips. Electrophoresis 2008, 29, 1801–1814. [Google Scholar] [CrossRef] [PubMed]
- Osipova, E. D.; Komleva, Y. K.; Morgun, A. V.; Lopatina, O. L.; Panina, Y. A.; Olovyannikova, R. Y.; Vais, E. F.; Salmin, V. V.; Salmina, A. B. Designing in Vitro Blood-Brain Barrier Models Reproducing Alterations in Brain Aging. Front. Aging Neurosci. 2018, 10, 1–14. [Google Scholar] [CrossRef]
- Saunders, N. R.; Dziegielewska, K. M.; Unsicker, K.; Ek, C. J. Delayed Astrocytic Contact with Cerebral Blood Vessels in FGF-2 Deficient Mice Does Not Compromise Permeability Properties at the Developing Blood-Brain Barrier. Dev. Neurobiol. 2016, 76, 1201–1212. [Google Scholar] [CrossRef] [PubMed]
- Sollier, E.; Murray, C.; Maoddi, P.; Di Carlo, D. Rapid Prototyping Polymers for Microfluidic Devices and High Pressure Injections. Lab Chip 2011, 11, 3752–3765. [Google Scholar] [CrossRef] [PubMed]
- Borók, A.; Laboda, K.; Bonyár, A. PDMS Bonding Technologies for Microfluidic Applications: A Review. Biosensors 2021, 11. [Google Scholar] [CrossRef]
- Wang, J.; Yang, L.; Wang, H.; Wang, L. Application of Microfluidic Chips in the Detection of Airborne Microorganisms. Micromachines 2022, 13, 1–18. [Google Scholar] [CrossRef]
- Azizipour, N.; Avazpour, R.; Rosenzweig, D. H.; Sawan, M.; Ajji, A. Evolution of Biochip Technology: A Review from Lab-on-a-Chip to Organ-on-a-Chip. Micromachines 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Gao, Y.; Broussard, J.; Haque, A.; Revzin, A.; Lin, T. Functional Imaging of Neuron–Astrocyte Interactions in a Compartmentalized Microfluidic Device. Microsystems Nanoeng. 2016, 2, 1–9. [Google Scholar] [CrossRef]
- Persson, H.; Park, S.; Mohan, M.; Cheung, K. K.; Simmons, C. A.; Young, E. W. K. Rapid Assembly of PMMA Microfluidic Devices with PETE Membranes for Studying the Endothelium. Sensors Actuators B Chem. 2022, 356, 131342. [Google Scholar] [CrossRef]
- Wang, J. D.; Khafagy, E. S.; Khanafer, K.; Takayama, S.; Elsayed, M. E. H. Organization of Endothelial Cells, Pericytes, and Astrocytes into a 3D Microfluidic in Vitro Model of the Blood-Brain Barrier. Mol. Pharm. 2016, 13, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Shakeri, A.; Jarad, N. A.; Khan, S.; F Didar, T. Bio-Functionalization of Microfluidic Platforms Made of Thermoplastic Materials: A Review. Anal. Chim. Acta 2022, 1209, 339283. [Google Scholar] [CrossRef]
- Lee, J.; Kim, M. Polymeric Microfluidic Devices Fabricated Using Epoxy Resin for Chemically Demanding and Day-Long Experiments. Biosensors 2022, 12. [Google Scholar] [CrossRef]
- Apelgren, P.; Amoroso, M.; Säljö, K.; Montelius, M.; Lindahl, A.; Stridh Orrhult, L.; Gatenholm, P.; Kölby, L.; Arulkumar, S.; Parthiban, S.; et al. Accepted Muspt. Mater. Today Proc. 2019, 27, 1–31. [Google Scholar]
- McCall, J. G.; Jeong, J.-W. Minimally Invasive Probes for Programmed Microfluidic Delivery of Molecules in Vivo. Curr. Opin. Pharmacol. 2017, 36, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W. M. The Blood-Brain Barrier: Bottleneck in Brain Drug Development. NeuroRx 2005, 2, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Prabhakarpandian, B.; Shen, M. C.; Nichols, J. B.; Mills, I. R.; Sidoryk-Wegrzynowicz, M.; Aschner, M.; Pant, K. SyM-BBB: A Microfluidic Blood Brain Barrier Model. Lab Chip 2013, 13, 1093–1101. [Google Scholar] [CrossRef]
- Sivandzade, F.; Cucullo, L. In-Vitro Blood–Brain Barrier Modeling: A Review of Modern and Fast-Advancing Technologies. J. Cereb. Blood Flow Metab. 2018, 38, 1667–1681. [Google Scholar] [CrossRef] [PubMed]
- Abbott, N. J. Astrocyte-Endothelial Interactions and Blood-Brain Barrier Permeability. J. Anat. 2002, 200, 629–638. [Google Scholar] [CrossRef]
- Dewi, B. E.; Takasaki, T.; Kurane, I. In Vitro Assessment of Human Endothelial Cell Permeability: Effects of Inflammatory Cytokines and Dengue Virus Infection. J. Virol. Methods 2004, 121, 171–180. [Google Scholar] [CrossRef]
- Wu, Y. C.; Sonninen, T. M.; Peltonen, S.; Koistinaho, J.; Lehtonen, Š. Blood–Brain Barrier and Neurodegenerative Diseases—Modeling with Ipsc-derived Brain Cells. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef]
- Castillo Ransanz, L.; Van Altena, P. F. J.; Heine, V. M.; Accardo, A. Engineered Cell Culture Microenvironments for Mechanobiology Studies of Brain Neural Cells. Front. Bioeng. Biotechnol. 2022, 10, 1–26. [Google Scholar] [CrossRef]
- Insperger, T.; Stépán, G. Engineering Applications. Appl. Math. Sci. 2011, 178, 93–149. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, D.; Ren, Y.; Guo, S.; Li, J.; Ma, S.; Yao, M.; Guan, F. Injectable Hyaluronic Acid Hydrogel Loaded with BMSC and NGF for Traumatic Brain Injury Treatment. Mater. Today Bio 2022, 13. [Google Scholar] [CrossRef]
- Spampinato, S. F.; Merlo, S.; Costantino, G.; Sano, Y.; Kanda, T.; Sortino, M. A. Decreased Astrocytic CCL2 Accounts for BAF - 312 Effect on PBMCs Transendothelial Migration Through a Blood Brain Barrier in Vitro Model. J. Neuroimmune Pharmacol. 2022, 427–436. [Google Scholar] [CrossRef]
- Helms, H.C.; Abbott, N.J.; Burek, M.; Cecchelli, R.; Couraud, P.O.; Deli, M.A.; Förster, C.; Galla, H.J.; Romero, I.A.; Shusta, E. V.; et al. In Vitro Models of the Blood-Brain Barrier: An Overview of Commonly Used Brain Endothelial Cell Culture Models and Guidelines for Their Use. J. Cereb. Blood Flow Metab. 2015, 36, 862–890. [Google Scholar] [CrossRef]
- Förster, C.; Silwedel, C.; Golenhofen, N.; Burek, M.; Kietz, S.; Mankertz, J.; Drenckhahn, D. Occludin as Direct Target for Glucocorticoid-Induced Improvement of Blood-Brain Barrier Properties in a Murine in Vitro System. J. Physiol. 2005, 565, 475–486. [Google Scholar] [CrossRef]
- Hickman, J. Transepithelial/Endothelial Electrical Resistance (TEER) Theory and Applications for Microfluidic Body-on-a-Chip Devices. J. Rare Dis. Res. Treat. 2016, 1, 46–52. [Google Scholar] [CrossRef]
- Sackett, S.D.; Tremmel, D.M.; Ma, F.; Feeney, A.K.; Maguire, R.M.; Brown, M.E.; Zhou, Y.; Li, X.; O’Brien, C.; Li, L.; et al. Extracellular Matrix Scaffold and Hydrogel Derived from Decellularized and Delipidized Human Pancreas. Sci. Rep. 2018, 8, 1–16. [Google Scholar] [CrossRef]
- Liu, H.; Wang, Y.; Cui, K.; Guo, Y.; Zhang, X.; Qin, J. Advances in Hydrogels in Organoids and Organs-on-a-Chip. Adv. Mater. 2019, 31, 1–28. [Google Scholar] [CrossRef]
- Wan, Z.; Zhang, S.; Zhong, A.X.; Shelton, S.E.; Campisi, M.; Sundararaman, S.K.; Offeddu, G.S.; Ko, E.; Ibrahim, L.; Coughlin, M.F.; et al. A Robust Vasculogenic Microfluidic Model Using Human Immortalized Endothelial Cells and Thy1 Positive Fibroblasts. Biomaterials 2021, 276, 121032. [Google Scholar] [CrossRef] [PubMed]
- Sivarapatna, A.; Ghaedi, M.; Xiao, Y.; Han, E.; Aryal, B.; Zhou, J.; Fernandez-Hernando, C.; Qyang, Y.; Hirschi, K. K.; Niklason, L. E. Engineered Microvasculature in PDMS Networks Using Endothelial Cells Derived from Human Induced Pluripotent Stem Cells. Cell Transplant. 2017, 26, 1365–1379. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Lutz, M. W.; Xing, Y. A Systems-Based Model of Alzheimer’s Disease. Alzheimers. Dement. 2019, 15, 168–171. [Google Scholar] [CrossRef]
- Fan, L.; Mao, C.; Hu, X.; Zhang, S.; Yang, Z.; Hu, Z.; Sun, H.; Fan, Y.; Dong, Y.; Yang, J.; Shi, C.; Xu, Y. New Insights Into the Pathogenesis of Alzheimer’s Disease. Front. Neurol. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Verma, M.; Wills, Z.; Chu, C. T. Excitatory Dendritic Mitochondrial Calcium Toxicity: Implications for Parkinson’s and Other Neurodegenerative Diseases. Front. Neurosci. 2018, 12, 523. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Koo, E. H. Biology and Pathophysiology of the Amyloid Precursor Protein. Mol. Neurodegener. 2011, 6, 1–16. [Google Scholar] [CrossRef]
- Haapasalo, A.; Hiltunen, M. A report from the 8th Kuopio Alzheimer Symposium. Neurodegener. Dis. Manag. 2018, 18, 289–299. [Google Scholar] [CrossRef]
- Kim, H. Detection of Severity in Alzheimer’s Disease (AD) Using Computational Modeling. Bioinformation 2018, 14, 259–264. [Google Scholar] [CrossRef]
- Salmina, A. B.; Inzhutova, A. I.; Malinovskaya, N. A.; Petrova, M. M. Endothelial Dysfunction and Repair in Alzheimer-Type Neurodegeneration: Neuronal and Glial Control. J. Alzheimer’s Dis. 2010, 22, 17–36. [Google Scholar] [CrossRef]
- González-Reyes, R. E.; Nava-Mesa, M. O.; Vargas-Sánchez, K.; Ariza-Salamanca, D.; Mora-Muñoz, L. Involvement of Astrocytes in Alzheimer’s Disease from a Neuroinflammatory and Oxidative Stress Perspective. Front. Mol. Neurosci. 2017, 10, 1–20. [Google Scholar] [CrossRef]
- Ávila-Villanueva, M.; Marcos Dolado, A.; Gómez-Ramírez, J.; Fernández-Blázquez, M. Brain Structural and Functional Changes in Cognitive Impairment Due to Alzheimer’s Disease. Front. Psychol. 2022, 13, 1–6. [Google Scholar] [CrossRef]
- Khoury, R.; Grysman, N.; Gold, J.; Patel, K.; Grossberg, G. T. The Role of 5 HT6-Receptor Antagonists in Alzheimer’s Disease: An Update. Expert Opin. Investig. Drugs 2018, 27, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Hussain, B.; Fang, C.; Chang, J. Blood–Brain Barrier Breakdown: An Emerging Biomarker of Cognitive Impairment in Normal Aging and Dementia. Front. Neurosci. 2021, 15, 1–22. [Google Scholar] [CrossRef]
- Devadhasan, J. P.; Kim, S.; An, J. Fish-on-a-Chip: A Sensitive Detection Microfluidic System for Alzheimer’s Disease. J. Biomed. Sci. 2011, 18, 1–11. [Google Scholar] [CrossRef]
- Sugama, S.; Takenouchi, T.; Cho, B. P.; Joh, T. H.; Hashimoto, M.; Kitani, H. Possible Roles of Microglial Cells for Neurotoxicity in Clinical Neurodegenerative Diseases and Experimental Animal Models. Inflamm. Allergy - Drug Targets 2009, 8, 277–284. [Google Scholar] [CrossRef]
- Yi, Y. Y.; Park, J. S.; Lim, J.; Lee, C. J.; Lee, S. H. Central Nervous System and Its Disease Models on a Chip. Trends Biotechnol. 2015, 33, 762–776. [Google Scholar] [CrossRef]
- Wu, J.W.; Hussaini, S.A.; Bastille, I.M.; Rodriguez, G.A.; Mrejeru, A.; Rilett, K.; Sanders, D.W.; Cook, C.; Fu, H.; Boonen, R.A.C.M.; et al. Neuronal Activity Enhances Tau Propagation and Tau Pathology in Vivo. Nat. Neurosci. 2016, 19, 1085–1092. [Google Scholar] [CrossRef]
- Alexoudi, A.; Alexoudi, I.; Gatzonis, S. Parkinson’s Disease Pathogenesis, Evolution and Alternative Pathways: A Review. Rev. Neurol. (Paris). 2018, 174, 699–704. [Google Scholar] [CrossRef]
- Cai, P.; Zheng, Y.; Sun, Y.; Zhang, C.; Zhang, Q.; Liu, Q. New Blood-Brain Barrier Models Using Primary Parkinson’s Disease Rat Brain Endothelial Cells and Astrocytes for the Development of Central Nervous System Drug Delivery Systems. ACS Chem. Neurosci. 2021, 12, 3829–3837. [Google Scholar] [CrossRef]
- Wang, C.; Yang, T.; Liang, M.; Xie, J.; Song, N. Astrocyte Dysfunction in Parkinson’s Disease: From the Perspectives of Transmitted α-Synuclein and Genetic Modulation. Transl. Neurodegener. 2021, 10, 1–17. [Google Scholar] [CrossRef]
- Yang, Y.; Song, J.J.; Choi, Y.R.; Kim, S.H.; Seok, M.J.; Wulansari, N.; Darsono, W.H.W.; Kwon, O.C.; Chang, M.Y.; Park, S.M.; et al. Therapeutic Functions of Astrocytes to Treat α-Synuclein Pathology in Parkinson’s Disease. Proc. Natl. Acad. Sci. U. S. A. 2022, 119, 1–12. [Google Scholar] [CrossRef]
- Gratton, C.; Koller, J. M.; Shannon, W.; Greene, D. J.; Maiti, B.; Snyder, A. Z.; Petersen, S. E.; Perlmutter, J. S.; Campbell, M. C. Emergent Functional Network Effects in Parkinson Disease. Cereb. Cortex 2019, 29, 2509–2523. [Google Scholar] [CrossRef]
- Creaby, M. W.; Cole, M. H. Gait Characteristics and Falls in Parkinson’s Disease: A Systematic Review and Meta-Analysis. Parkinsonism Relat. Disord. 2018, 57, 1–8. [Google Scholar] [CrossRef]
- Wanneveich, M.; Moisan, F.; Jacqmin-Gadda, H.; Elbaz, A.; Joly, P. Projections of Prevalence, Lifetime Risk, and Life Expectancy of Parkinson’s Disease (2010-2030) in France. Mov. Disord. 2018, 33, 1449–1455. [Google Scholar] [CrossRef]
- Eggers, C.; Dano, R.; Schill, J.; Fink, G. R.; Timmermann, L.; Voltz, R.; Golla, H.; Lorenzl, S. Access to End-of Life Parkinson’s Disease Patients Through Patient-Centered Integrated Healthcare. Front. Neurol. 2018, 9, 1–6. [Google Scholar] [CrossRef]
- Hughes, R. C. Parkinson’s Disease and Its Management. Bmj 1994, 308, 281. [Google Scholar] [CrossRef]
- Holloway, P. M.; Willaime-Morawek, S.; Siow, R.; Barber, M.; Owens, R. M.; Sharma, A. D.; Rowan, W.; Hill, E.; Zagnoni, M. Advances in Microfluidic in Vitro Systems for Neurological Disease Modeling. J. Neurosci. Res. 2021, 99, 1276–1307. [Google Scholar] [CrossRef]
- Rahmani Dabbagh, S.; Rezapour Sarabi, M.; Birtek, M. T.; Mustafaoglu, N.; Zhang, Y. S.; Tasoglu, S. 3D Bioprinted Organ-on-chips. Aggregate 2023, 4, 1–26. [Google Scholar] [CrossRef]
- Fanizza, F.; Campanile, M.; Forloni, G.; Giordano, C.; Albani, D. Induced Pluripotent Stem Cell-Based Organ-on-a-Chip as Personalized Drug Screening Tools: A Focus on Neurodegenerative Disorders. J. Tissue Eng. 2022, 13. [Google Scholar] [CrossRef]
- Pediaditakis, I.; Kodella, K.R.; Manatakis, D. V.; Le, C.Y.; Hinojosa, C.D.; Tien-Street, W.; Manolakos, E.S.; Vekrellis, K.; Hamilton, G.A.; Ewart, L.; et al. Modeling Alpha-Synuclein Pathology in a Human Brain-Chip to Assess Blood-Brain Barrier Disruption. Nat. Commun. 2021, 12, 1–17. [Google Scholar] [CrossRef]
- Fernandes, J. T. S.; Chutna, O.; Chu, V.; Conde, J. P.; Outeiro, T. F. A Novel Microfluidic Cell Co-Culture Platform for the Study of the Molecular Mechanisms of Parkinson’s Disease and Other Synucleinopathies. Front. Neurosci. 2016, 10, 1–11. [Google Scholar] [CrossRef]
- Noyes, K.; Weinstock-Guttman, B. Impact of Diagnosis and Early Treatment on the Course of Multiple Sclerosis. Am. J. Manag. Care 2013, 19, s321–31. [Google Scholar]
- Sintzel, M. B.; Rametta, M.; Reder, A. T. Vitamin D and Multiple Sclerosis: A Comprehensive Review. Neurol. Ther. 2018, 7, 59–85. [Google Scholar] [CrossRef]
- Solaro, C.; Trabucco, E.; Messmer Uccelli, M. Pain and Multiple Sclerosis: Pathophysiology and Treatment Topical Collection on Demyelinating Disorders. Curr. Neurol. Neurosci. Rep. 2013, 13. [Google Scholar] [CrossRef]
- Ghasemi, N.; Razavi, S.; Nikzad, E. Multiple Sclerosis: Pathogenesis, Symptoms, Diagnoses and Cell-Based Therapy Citation: Ghasemi N, Razavi Sh, Nikzad E. Multiple Sclerosis: Pathogenesis, Symptoms, Diagnoses and Cell-Based Therapy. Cell J. 2017, 19, 1–10. [Google Scholar] [CrossRef]
- Hosmane, S.; Tegenge, M. A.; Rajbhandari, L.; Uapinyoying, P.; Kumar, N. G.; Thakor, N.; Venkatesan, A. Toll/Interleukin-1 Receptor Domain-Containing Adapter Inducing Interferon-β Mediates Microglial Phagocytosis of Degenerating Axons. J. Neurosci. 2012, 32, 7745–7757. [Google Scholar] [CrossRef]
- Kerman, B.E.; Kim, H.J.; Padmanabhan, K.; Mei, A.; Georges, S.; Joens, M.S.; Fitzpatrick, J.A.J.; Jappelli, R.; Chandross, K.J.; August, P.; et al. In Vitro Myelin Formation Using Embryonic Stem Cells. Dev. 2015, 142, 2213–2225. [Google Scholar] [CrossRef]
- Finkbeiner, S. Huntington’s Disease. Cold Spring Harb. Perspect. Biol. 2011, 3. [Google Scholar] [CrossRef]
- Nopoulos, P. C. Huntington Disease: A Single-Gene Degenerative Disorder of the Striatum. Dialogues Clin. Neurosci. 2016, 18, 91–98. [Google Scholar] [CrossRef]
- Bachoud-Lévi, A.C.; Ferreira, J.; Massart, R.; Youssov, K.; Rosser, A.; Busse, M.; Craufurd, D.; Reilmann, R.; De Michele, G.; Rae, D.; et al. International Guidelines for the Treatment of Huntington’s Disease. Front. Neurol. 2019, 10, 1–18. [Google Scholar] [CrossRef]
- Nix, C.; Ghassemi, M.; Crommen, J.; Fillet, M. Overview on Microfluidics Devices for Monitoring Brain Disorder Biomarkers. TrAC Trends Anal. Chem. 2022, 155, 116693. [Google Scholar] [CrossRef]
- Teixeira, M. I.; Amaral, M. H.; Costa, P. C.; Lopes, C. M.; Lamprou, D. A. Recent Developments in Microfluidic Technologies for Central Nervous System Targeted Studies. Pharmaceutics 2020, 12, 1–37. [Google Scholar] [CrossRef]
- Yochum, A. Autism Spectrum/Pervasive Developmental Disorder. Prim. Care 2016, 43, 285–300. [Google Scholar] [CrossRef]
- Rashighi, M.; Harris, J. E. 乳鼠心肌提取 HHS Public Access. Physiol. Behav. 2017, 176, 139–148. [Google Scholar]
- Feliciano, P. Promoting Resistance Repeat Expansion in ALS and FTD Epiallele Accumulation. Nat. Genet. 2011, 43, 1053. [Google Scholar] [CrossRef]
- Chahboun, S.; Stenseng, F.; Page, A. G. The Changing Faces of Autism: The Fluctuating International Diagnostic Criteria and the Resulting Inclusion and Exclusion—A Norwegian Perspective. Front. Psychiatry 2022, 13. [Google Scholar] [CrossRef]
- Gutmann, D. H.; Ferner, R. E.; Listernick, R. H.; Korf, B. R.; Wolters, P. L.; Johnson, K. J. Neurofibromatosis Type 1. Nat. Rev. Dis. Prim. 2017, 3, 1–18. [Google Scholar] [CrossRef]
- Boyd, K. P.; Korf, B. R.; Theos, A. Neurofibromatosis Type 1. J. Am. Acad. Dermatol. 2009, 61, 1–6. [Google Scholar] [CrossRef]
- Wang, W.; Wei, C. J.; Cui, X. W.; Li, Y. H.; Gu, Y. H.; Gu, B.; Li, Q. F.; Wang, Z. C. Impacts of NF1 Gene Mutations and Genetic Modifiers in Neurofibromatosis Type 1. Front. Neurol. 2021, 12, 1–12. [Google Scholar] [CrossRef]
- Hirbe, A. C.; Gutmann, D. H. Neurofibromatosis Type 1: A Multidisciplinary Approach to Care. Lancet Neurol. 2014, 13, 834–843. [Google Scholar] [CrossRef]
- Ferner, R. E. Neurofibromatosis 1. Eur. J. Hum. Genet. 2007, 15, 131–138. [Google Scholar] [CrossRef] [PubMed]
- emov, A.; Sung, H.; Hyland, P.L.; Sloan, J.L.; Ruppert, S.L.; Baldwin, A.M.; Boland, J.F.; Bass, S.E.; Lee, H.J.; Jones, K.M.; et al. Genetic Modifiers of Neurofibromatosis Type 1-Associated ´ -au-Lait Macule Count Identified Using Multi-platform Cafe Analysis. 2014, 10. [Google Scholar] [CrossRef]
- Van Den Berg, B.; Walgaard, C.; Drenthen, J.; Fokke, C.; Jacobs, B. C.; Van Doorn, P. A. Guillain-Barré Syndrome: Pathogenesis, Diagnosis, Treatment and Prognosis. Nat. Rev. Neurol. 2014, 10, 469–482. [Google Scholar] [CrossRef]
- Miny, L.; Maisonneuve, B. G. C.; Quadrio, I.; Honegger, T. Modeling Neurodegenerative Diseases Using In Vitro Compartmentalized Microfluidic Devices. Front. Bioeng. Biotechnol. 2022, 10, 1–17. [Google Scholar] [CrossRef]
- Korhonen, P.; Malm, T.; White, A. R. 3D Human Brain Cell Models: New Frontiers in Disease Understanding and Drug Discovery for Neurodegenerative Diseases. Neurochem. Int. 2018, 120, 191–199. [Google Scholar] [CrossRef]
- Lee, C. T.; Bendriem, R. M.; Wu, W. W.; Shen, R. F. 3D Brain Organoids Derived from Pluripotent Stem Cells: Promising Experimental Models for Brain Development and Neurodegenerative Disorders Julie Y.H. Chan. J. Biomed. Sci. 2017, 24, 1–12. [Google Scholar] [CrossRef]
- de Mello, C. P. P.; Rumsey, J.; Slaughter, V.; Hickman, J. J. A Human-on-a-Chip Approach to Tackling Rare Diseases. Drug Discov. Today 2019, 24, 2139–2151. [Google Scholar] [CrossRef]
- Mencattini, A.; Giuseppe, D.; D’Orazio, M.; Rizzuto, V.; Pereira, M.; Comes, M. C.; Lopez-Martinez, M.; Samitier, J.; Martinelli, E. A Microfluidic Device for Shape Measurement in Red Blood Cells (RBCs). In Proceedings of the 2020 IEEE International Symposium on Medical Measurements and Applications (MeMeA); 2020. [Google Scholar] [CrossRef]
- Herrmann, K.; Pistollato, F.; Stephens, M. L. Beyond the 3Rs: Expanding the Use of Human-Relevant Replacement Methods in Biomedical Research. ALTEX - Altern. to Anim. Exp. 2019, 36, 343–352. [Google Scholar] [CrossRef]
- Ravi, K.; Paidas, M. J.; Saad, A.; Jayakumar, A. R. Astrocytes in Rare Neurological Conditions: Morphological and Functional Considerations. J. Comp. Neurol. 2021, 529, 2676–2705. [Google Scholar] [CrossRef]
- Gupta, N.; Liu, J. R.; Patel, B.; Solomon, D. E.; Vaidya, B.; Gupta, V. Microfluidics-based 3D Cell Culture Models: Utility in Novel Drug Discovery and Delivery Research. Bioeng. Transl. Med. 2016, 1, 63–81. [Google Scholar] [CrossRef]
- Chiareli, R.A.; Carvalho, G.A.; Marques, B.L.; Mota, L.S.; Oliveira-Lima, O.C.; Gomes, R.M.; Birbrair, A.; Gomez, R.S.; Simão, F.; Klempin, F.; et al. The Role of Astrocytes in the Neurorepair Process. Front. Cell Dev. Biol. 2021, 9, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Strazielle, N.; Ghersi-Egea, J. F. Physiology of Blood-Brain Interfaces in Relation to Brain Disposition of Small Compounds and Macromolecules. Mol. Pharm. 2013, 10, 1473–1491. [Google Scholar] [CrossRef] [PubMed]
- Combedazou, A.; Gayral, S.; Colombié, N.; Fougerat, A.; Laffargue, M.; Ramel, D. Small GTPases Orchestrate Cell-Cell Communication during Collective Cell Movement. Small GTPases 2020, 11, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Mi, G.; Wang, M.; Webster, T. J. Since January 2020 Elsevier Has Created a COVID-19 Resource Centre with Free Information in English and Mandarin on the Novel Coronavirus COVID- 19 . The COVID-19 Resource Centre Is Hosted on Elsevier Connect , the Company ’ s Public News and Information. 2020. [Google Scholar]
- Regmi, S.; Poudel, C.; Adhikari, R.; Luo, K. Q. Applications of Microfluidics and Organ-on-a-Chip in Cancer Research. Biosensors 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Peng, Y.; Li, H.; Chen, W. Organ-on-a-Chip: A New Paradigm for Drug Development. Trends Pharmacol. Sci. 2021, 42, 119–133. [Google Scholar] [CrossRef]
Figure 1.
Key milestones of Microfluidic devices. Created in Biorender.com 2022 (https://app.biorender.com/illustrations/6380499d2e82d2aaf66a47ab).
Figure 1.
Key milestones of Microfluidic devices. Created in Biorender.com 2022 (https://app.biorender.com/illustrations/6380499d2e82d2aaf66a47ab).
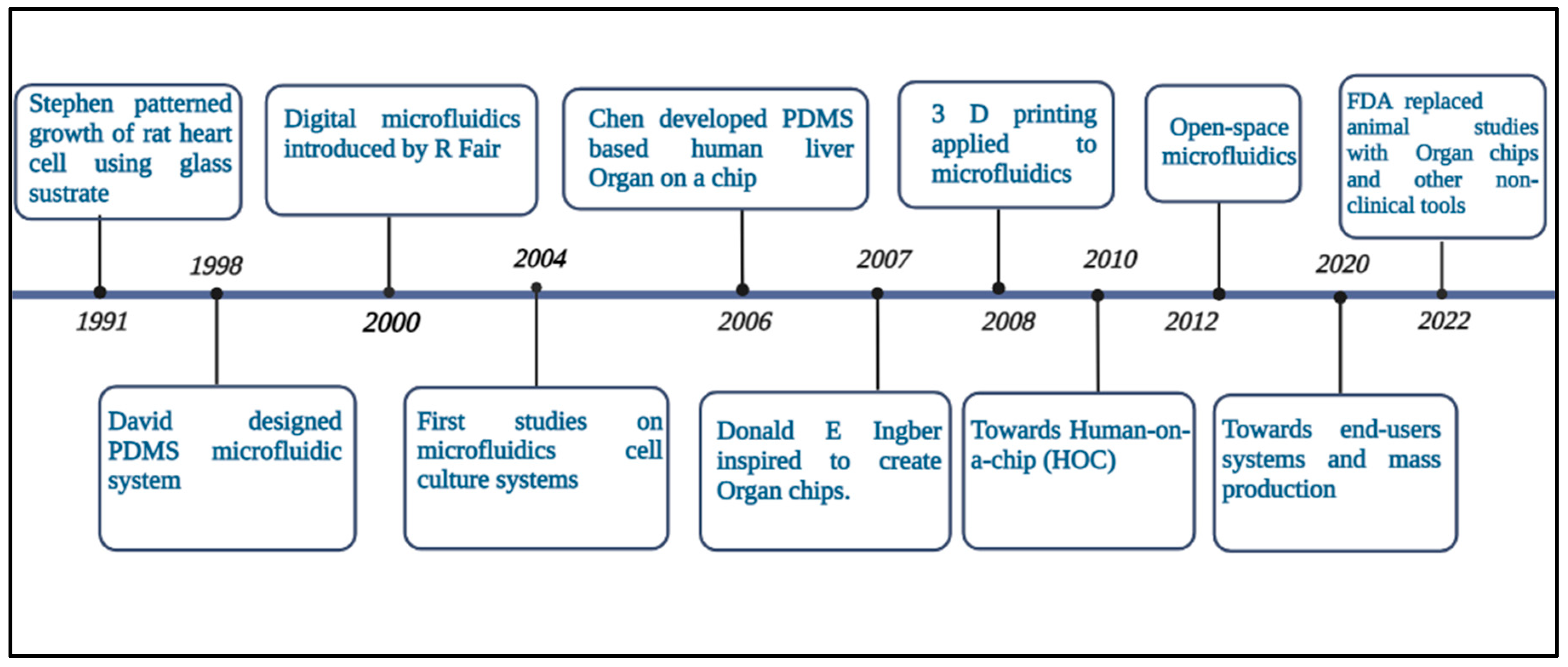
Figure 2.
Schematic representation of Endothelial-Astrocytic Interactions. Created in Biorender.com 2022 (https://app.biorender.com/biorender-templates).
Figure 2.
Schematic representation of Endothelial-Astrocytic Interactions. Created in Biorender.com 2022 (https://app.biorender.com/biorender-templates).
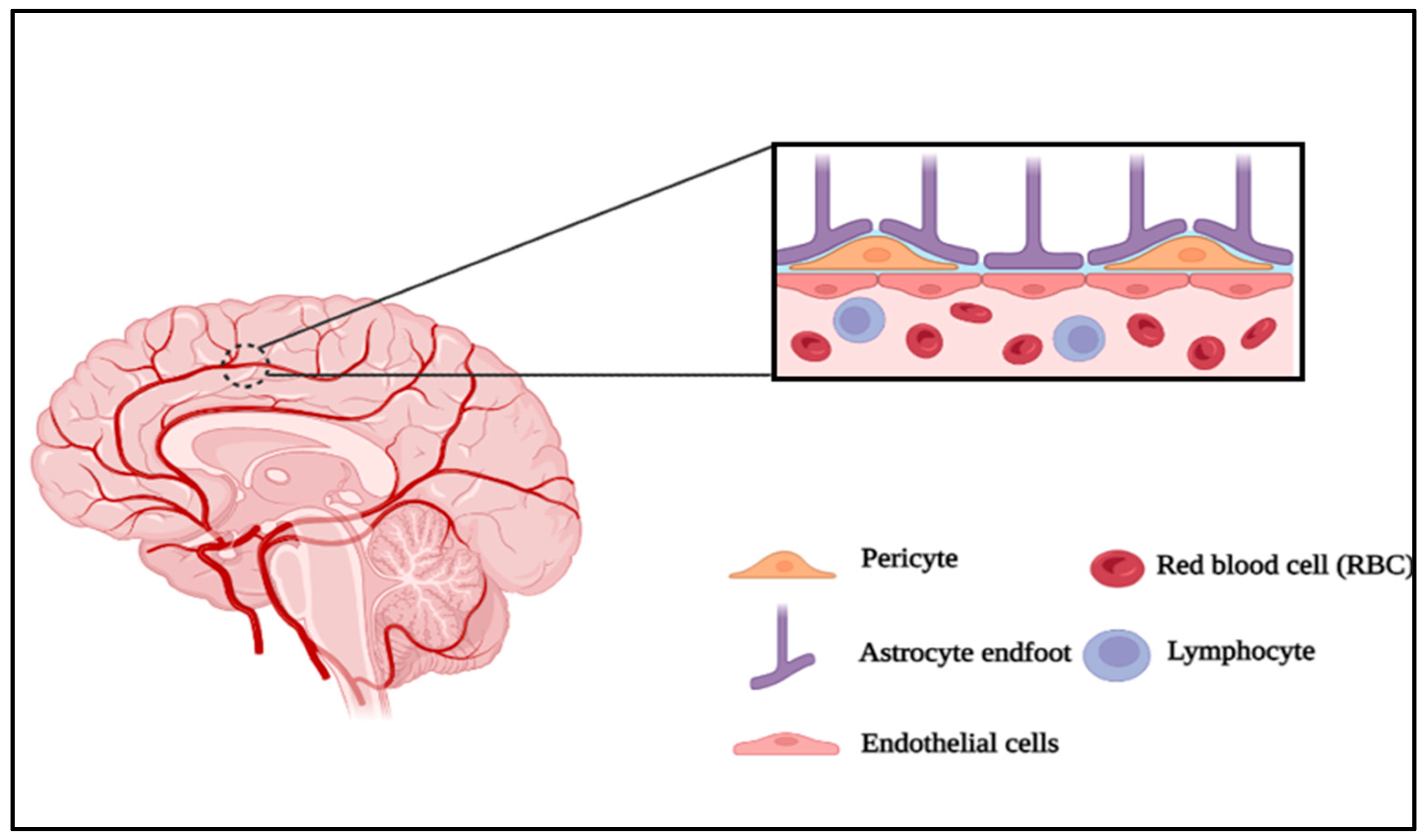
Figure 3.
Various approaches for the integration of multi-cellular device.
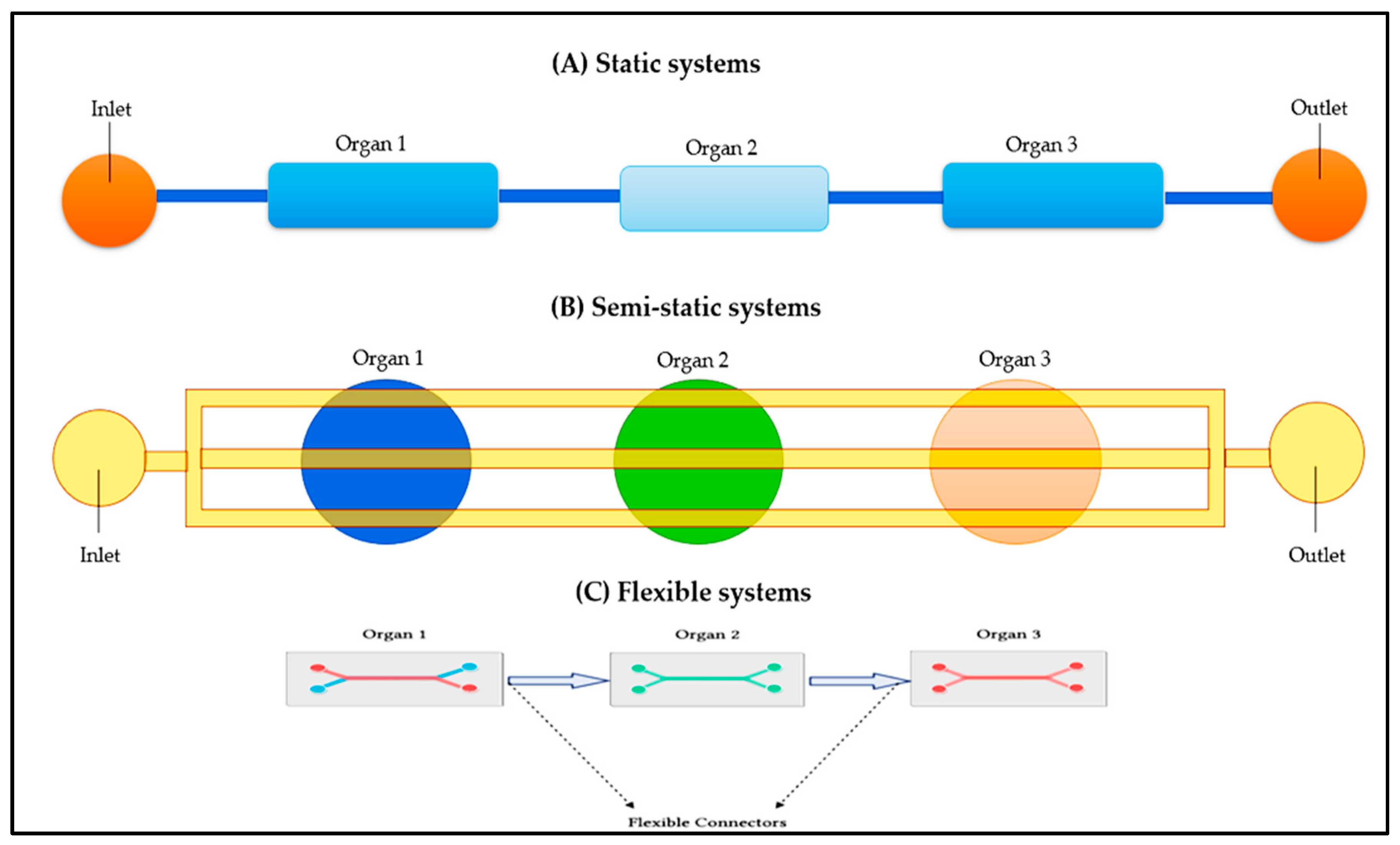
Figure 4.
Schematic illustration of the three ways siloxane elastomers have been employed in flexible microfluidics. A layer containing the microchannels (b) Substrate for the electrodes; (c) Sealing layer. Created in Unigraphics NX 12 (https://www.plm.automation.siemens.com/global/en/products/nx/).
Figure 4.
Schematic illustration of the three ways siloxane elastomers have been employed in flexible microfluidics. A layer containing the microchannels (b) Substrate for the electrodes; (c) Sealing layer. Created in Unigraphics NX 12 (https://www.plm.automation.siemens.com/global/en/products/nx/).
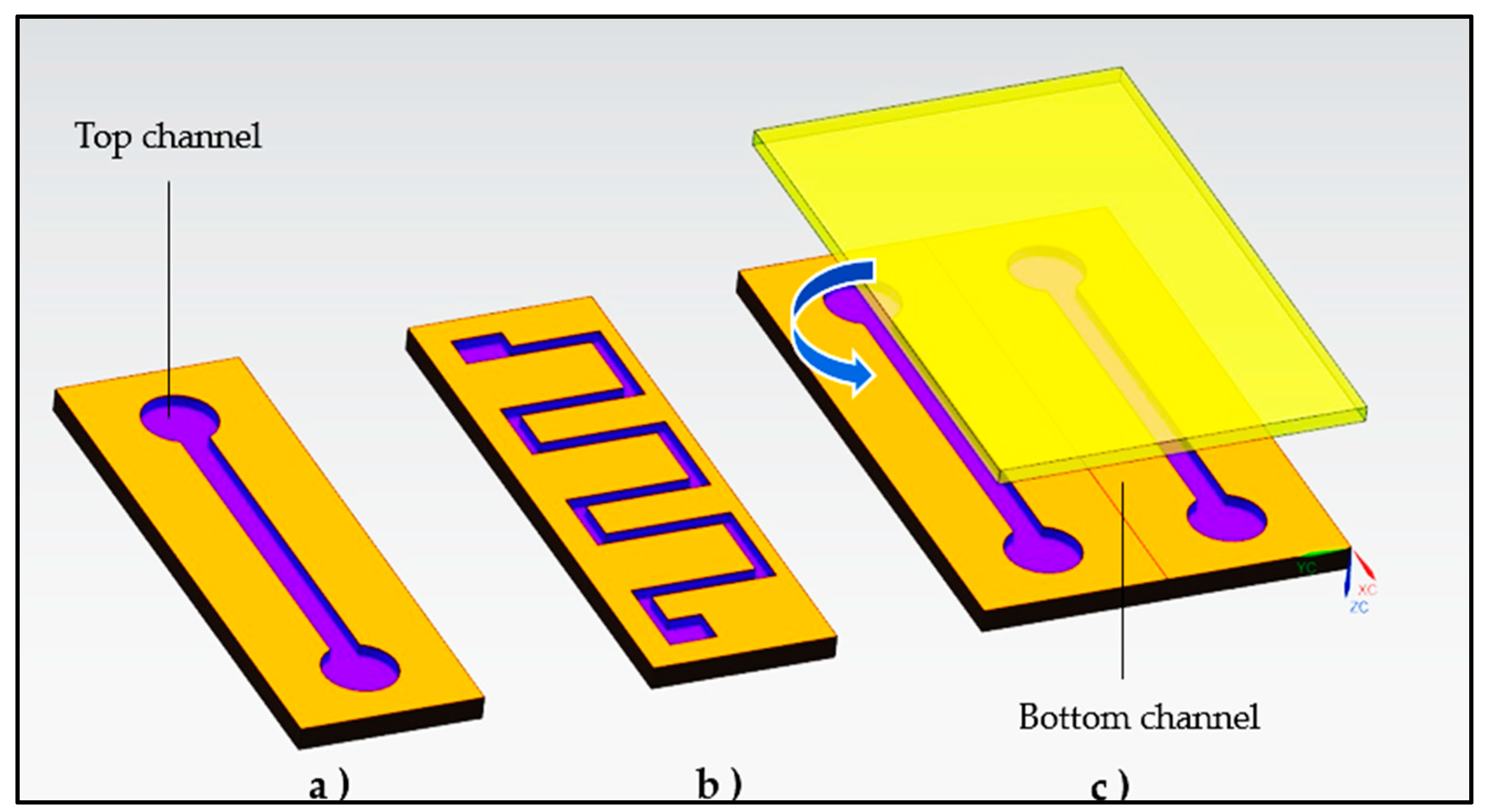
Figure 5.
Schematic representation of signaling regulating the blood-brain barrier in health and disease state. Created in biorender.com 2022 (https://app.biorender.com/illustrations/637c94a1c9e4cb9d5f4f9c.
Figure 5.
Schematic representation of signaling regulating the blood-brain barrier in health and disease state. Created in biorender.com 2022 (https://app.biorender.com/illustrations/637c94a1c9e4cb9d5f4f9c.
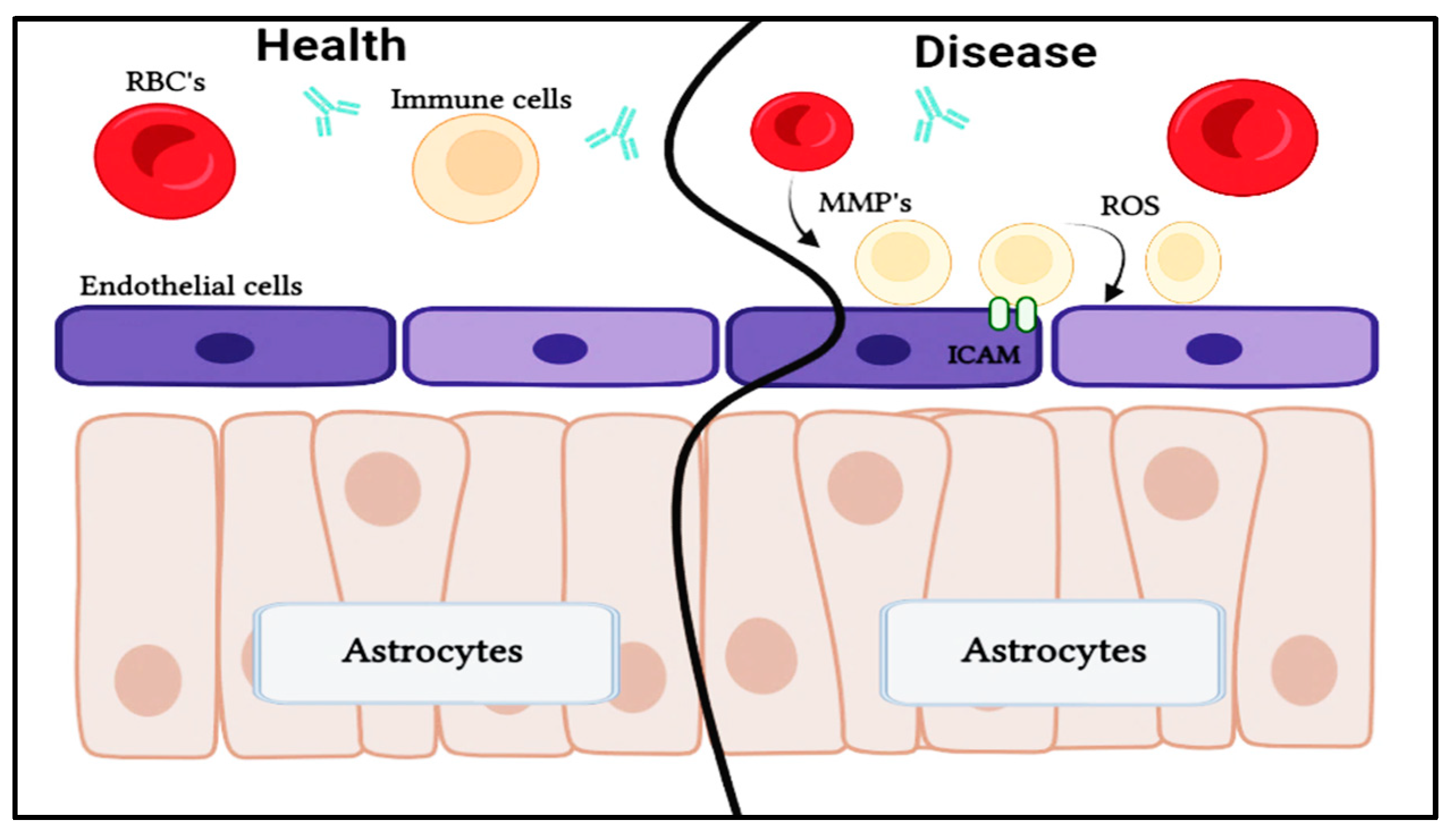
Table 2.
Different Microfluidic Fabrication Methods and Applications - Obstacles and Prospects.
| Method | Advantages | Disadvantages | References |
| Manufacturing molds by soft lithography | Resolution to mm scale is feasible, which enables the development of multi-layered designs or curved channels. | Expensive and time-consuming to acquire the necessary equipment. | [58] |
| Paper based microfluidics | Simple, low-cost devices made of paper or other porous membranes that wick fluids through capillary action. | Low resolution (mm), patterned shape variation | [59] |
| Manufacturing molds by 3D printing | Resolution to tens of mm is possible, inexpensive and simple. | Channel height must be at least 50 mm. | [60] |
| Inkjet Printing | Simple, quick prototyping, and low cost. | It must be treated with a solvent. | [61] |
| 3D-Laser lithography | True 3D-feature generation, including gradually changing channel dimensions. | Expensive, slow, and unsuitable for deep channels (greater than 100 mm). | [62] |
| Continuous-flow microfluidics | High temporal and spatial precision of flow conditions is possible. | The removal of substances from cells or tissues limits temporal resolution. | [63] |
| Droplet microfluidics | High-throughput screening is possible, and commercial systems for some applications are available. | The development of new approaches necessitates the use of costly laboratory equipment and technically skilled personnel. | [64] |
Table 5.
Characteristics of the main neurological diseases and genes involved.
| Dysfunction | Characteristic features | Symptoms | Genes involved |
|---|---|---|---|
| Alzheimer’s disease | The gradual loss of most of the normal brain activity is caused by abnormal alterations in the brain. Senile plaques and neurofibrillary tangles. | Gradual loss of control over mental and physical abilities, dementia & mood swings | Presenilin 1 and 2 are located on chromosome 14 and 1, respectively, and APP is located on chromosome 21. |
| Parkinson’s disease | Loss of pigmented dopaminergic neurons & Lewy bodies | Both motor and nonmotor symptoms. | X PINK1, PRKN, or SNCA gene PARK7 & LRRK2 |
| Multiple Sclerosis | Axons that are demyelinated and transected, inflammatory cells and their byproducts, and astrogliosis form CNS plaques. | Neurological problems as well as severe physical or cognitive impairment | HLA-DRB1 |
| Huntington’s disease | Chorea, dystonia and Slow or unusual eye movements | Impatience, melancholy, and other mood problems, as well as erratic involuntary movements and a dramatic loss in thinking and reasoning skills | Faulty gene on chromosome 4 |
| Autism spectrum disorder | Challenges with social interaction, verbal and nonverbal communication, the appearance of repetitive behavior, and narrowed interests. | Struggling to make friendships and odd reactions to sensory stimuli | ASH1L, CHD2, SHANK3, SYNGAP1 CHD8 & DYRK1A |
| Neurofibromatosis | Meningiomas and bilateral vestibular schwannomas | Tumors on the skin and in the nerve system. | Neurofibromin 1 (NF1) gene |
| Guillain-Barre syndrome | Autoimmune destruction of peripheral nervous system nerves | Numbness, tingling, and weakness that can develop into paralysis | ICAM1, HLA genes, CD-1A, FcGR, NOD, TNF-α & TLR4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



