
Submitted:
16 June 2023
Posted:
16 June 2023
You are already at the latest version
Abstract
Mutations in the Neuroligin-3 (Nlgn3) gene are implicated in autism spectrum disorder (ASD) and gastrointestinal (GI) dysfunction but cellular Nlgn3 expression in the enteric nervous system remains to be characterised. We combined RNAScope in situ hybridization for Nlgn3 mRNA and immunofluorescence for cholinergic and VIP-expressing submucosal neurons, nitrergic and calretinin-containing myenteric neurons and glial cells. We measured Nlgn3 mRNA neuronal and glial expression via quantitative 3-dimensional image analysis. To validate dual RNAScope/immunofluorescence data, we interrogated available single cell RNA sequencing (scRNASeq) data to assess for Nlgn3, Nlgn1, Nlgn2 and their binding partners, Nrxn1-3, MGDA1 and MGDA2, in enteric neural subsets. Most submucosal and myenteric neurons expressed Nlgn3 mRNA. In contrast to other Nlgns and binding partners, Nlgn3 was strongly expressed in enteric glia, suggesting a role for Neuroligin-3 in mediating enteric neuron-glia interactions. The autism-associated R451C mutation reduces Nlgn3 mRNA expression in cholinergic but not VIPergic submucosal neurons. In the myenteric plexus, Nlgn3 mRNA levels are reduced in calretinin and NOS labelled neurons and S100ß labelled glia. We provide a comprehensive cellular profile for neuroligin-3 expression in ileal neuronal subpopulations of mice expressing the R451C autism-associated mutation in Nlgn3, which may contribute to the understanding of the pathophysiology of GI dysfunction in ASD.
Keywords:
neuroligin-3
; autism
; enteric nervous system
; glia
; ileum
; RNAScope
; immunocytochemistry
; scRNASeq
1. Introduction
Neuroligin-3 (NLGN3) is an adhesion molecule expressed at neuronal synapses in the central nervous system (CNS) and is well established as a regulator of brain function [1,2,3,4,5]. Although some studies suggest NLGN3 is important for proper GI function [6,7], its cellular expression profile in the enteric nervous system has not been characterised. In cultured rat hippocampal neurons, NLGN3 is expressed at the postsynaptic membrane of both excitatory and inhibitory neuronal synapses [8]. NLGN3 is also expressed in non-neuronal cells. For example, Venkatesh and colleagues showed that neurally-secreted NLGN3 invades the microenvironment of tumours and induces NLGN3 expression in glioma cells to promote tumour growth [9]. In addition, NLGN3 is expressed in many types of glia during rodent development, including olfactory ensheathing glia, retinal astrocytes, Schwann cells and spinal cord astrocytes [10]. These expression patterns suggest that NLGN3 could also be expressed in enteric glia.
Mutations in the NLGN3 gene including the missense R451C point mutation as well as NLGN3 deletion are implicated in autism [11,12,13]. Patients expressing the NLGN3 R451C mutation show GI dysfunction including diarrhea, faecal incontinence, post-meal regurgitation, oesophageal inflammation, chronic intestinal pain as well as delayed bladder and bowel control [7]. Modification of NLGN3 expression induces GI dysfunction in mice, as shown in both Nlgn3 knockout (KO) and Nlgn3R451C mice [6,7]. Nlgn3 KO mice exhibit distended colons and more rapidly propagating colonic muscle contractions [6]. Nlgn3R451C mice display faster intestinal transit and increased numbers of myenteric neurons in the small intestine as well as GABAA receptor-mediated colonic dysmotility [7]. These findings suggest a role for NLGN3 in gut function.
The R451C mutation impacts Nlgn3 mRNA and protein expression in the brain. In mouse whole brain samples, Nlgn3 mRNA expression levels are unchanged by the Nlgn3 R451C mutation but NLGN3 protein levels are reduced [1]. Although initial studies of NLGN3 expression in the GI tract have been reported in rodents [7,14,15] and humans [14], the distribution of NLGN3 expression in different cell populations of the mouse enteric nervous system has not been profiled. Characterising Nlgn3 expression in enteric cell subtypes is essential for identifying underlying mechanisms contributing to GI activity and to functional GI changes observed in patients and the Nlgn3R451C mouse model of autism.
Localizing the NLGN3 protein in the mouse GI tract is challenging due to non-specific labelling of commercially available antibodies which commonly yield false-positive results in this tissue (Leembruggen et al., unpublished). Therefore, we used the enhanced in situ hybridization technique, RNAScope, to label Nlgn3 mRNA in enteric neurons. We combined RNAScope and immunofluorescence with high-resolution microscopy and 3D image analysis software to compare Nlgn3 mRNA expression in the wild type (WT) and Nlgn3R451C mouse gut.
Overall, we found that Nlgn3 mRNA is expressed in the mouse enteric nervous system (including enteric glia) and that Nlgn3R451C mice have reduced Nlgn3 mRNA expression in subpopulations of submucosal and myenteric plexus neurons. The molecular identities of enteric neuronal populations have now been well described in mice based on scRNASeq combined with immunostaining verification [16,17]. These studies show the presence of 12 enteric neuronal populations with distinct neurochemical signatures and enteric glia. We therefore investigated the cellular expression of Nlgn3 in the mouse enteric nervous system based on previously reported scRNASeq datasets and found that all enteric neuronal subtypes and glia express Nlgn3 at varying levels. In mice, the R451C mutation reduces Nlgn3 mRNA expression in submucosal cholinergic as well as myenteric nitrergic and calretinin neurons. Nlgn3R451C mice also had reduced mRNA expression levels in myenteric (but not submucosal) glia. In contrast, Nlgn3 mRNA expression was unchanged in VIPergic submucosal neurons in Nlgn3R451C mice. Taken together, these findings suggest that NLGN3 is an important component of neuronal and neuron-glial communication in the gut, and that mutations in NLGN3 might play a role in GI dysfunction in individuals with autism.
2. Materials and Methods
Animals
B6;129-Nlgn3tm1Sud/J mice were obtained from The Jackson Laboratory [1] and bred for over 10 generations on a C57BL/6 background at the Howard Florey Institute, Melbourne Australia. Mice were subsequently housed in the Biomedical Science Animal Facility at The University of Melbourne. Male Nlgn3R451C and wild type (WT) mice (age 12-13 weeks) were sacrificed by cervical dislocation in accordance with The University of Melbourne Animal Experimentation Ethics Committee (#1914843). For each set of neuronal markers analysed, a different group of WT and mutant mice were used.
Tissue preparation
Mice were sacrificed and the abdomen was opened using coarse dissection scissors. A 2 cm segment of the distal ileum (1 cm proximal to the caecum) was isolated and immediately placed in oxygenated ice-cold physiological saline solution (composition in mM: 118 NaCl, 4.6 KCl, 2.5 CaCl2, 1.2 MgSO4, 1 NaH2PO4, 25 NaHCO3, 11 D-glucose). The intestinal content was gently flushed clean using a Pasteur pipette. Using small dissecting scissors, the distal ileal tissue sample was cut along the mesenteric border, stretched and pinned flat on a chilled silicon elastomer-lined dish (Sylgard 184, Dow Corning, Australia). The tissue was fixed in 4% formaldehyde for 24 h at 4oC. After fixation, the tissues were washed three times in 1M phosphate buffer saline (PBS) (composition in mM: 137 NaCl, 2.7 KCl, 8 Na2HPO4, 2 KH2PO4). The wholemount submucosal plexus preparation was isolated by peeling the mucosa-submucosal layer and then removing the epithelium using fine forceps (Fine Science Tools, Canada). Myenteric plexus preparations including the longitudinal muscle layer (LMMP) were obtained by microdissection to remove the circular muscle layer.
Dual RNAScope in situ hybridization and immunofluorescence
Using RNAScope, individual RNA molecules are visualized using a novel probe design and amplification system to simultaneously magnify the signal and suppress background noise [18]. RNAScope in situ hybridization in mouse ileal tissue was performed using the ACDBio multiplex RNAScope assay kit (ACD, 320850, USA) according to manufacturer’s instructions. Briefly, wholemount mouse ileal submucosal and LMMP preparations were pre-treated and hybridized with the RNAScope probe of interest. The hybridization signal was subsequently amplified via a sequence of amplifiers and fluorescently labelled probes.
Tissue pre-treatment
Wholemount preparations of submucosal and LMMP preparations were permeabilized via pre-treatment with protease IV prior to RNAScope in situ hybridisation. The tissue samples were incubated in 30 μl of protease IV for 30 min at room temperature (RT) in a sealed humid chamber and washed twice (1 min in each) in 1M PBS.
RNAscope in situ hybridization
Following pre-treatment, ileal tissue preparations were incubated in 20 μl of Nlgn3 RNAScope probe and incubated for 2 h at 40 oC in a humid chamber. The ileal tissues were washed twice in 1x wash buffer for 2 min at RT with agitation. Amplification and detection steps were performed using the RNAScope detection kit reagents. For the signal amplification step, tissues were incubated in 50 μl of amplifier 1 solution (AMP 1) for 30 min at 40 oC and washed twice in 1x wash buffer for 2 min at RT with occasional agitation. Subsequently, the tissue was incubated in 50 μl of AMP 2 solution for 15 min at 40oC and then washed twice in 1x wash buffer for 2 min at RT with occasional agitation. After AMP 2 amplification, 50 μl of AMP 3 solution was added to the tissue preparations and incubated for 30 min at 40oC. The tissue was washed twice in 1x wash buffer for 2 min at RT with occasional agitation. Lastly, the tissue was incubated in 50 μl of AMP 4 solution and incubated for 15 min at 40oC prior to washing the tissue twice in 1x wash buffer for 2 min at RT with occasional agitation. The same protocol was followed for control experiments using the RNAScope universal negative control dapB probe (Supplementary Figure S1).
Immunofluorescence
After washing three times in PBS the wholemount preparations were permeabilised with 1% Triton (ProSciTech, Australia) for 30 min at RT and washed three times in PBS. The tissues were then double labelled with various combinations of primary antibodies at different incubation conditions. Excess primary antibodies were removed by washing in PBS (3 x 10 min) and the tissue samples were subsequently incubated in secondary antibodies for 2 h at 4 0C. After removing excess antisera by washing with 0.1M PBS (3 x 10 min), tissue preparations were mounted using Dakocytomation fluorescence mounting medium (DAKO; Carpenteria, USA).
Image acquisition and analysis
Multi-channel image acquisition was performed with a laser scanning confocal microscope 800 (Carl Zeiss Microscopy, North Ryde, NSW, Australia) using a 40x oil immersion objective lens. The primary and secondary antibodies used in this study are listed in Table 1 and Table 2 respectively. Tissue samples were excited with Diode lasers at 488, 561 and 647 wavelengths. Pinhole diameter, detector gain and laser power parameters were optimised to obtain the highest pixel intensity at the same time eliminating pixel saturation. Consecutive Z stacks on the horizontal plane with a frame size of 1024x1024 pixels and a bit-depth of 16 bits were captured at 1 µm intervals. Z-stacks together with tile scans were performed where necessary.
Quantification of labelling for RNAscope puncta and antisera was performed using 3D reconstruction tools and statistical annotation for quantification purposes in Imaris 9.0 image analysis software (Bitplane, UK). Three-dimensional cell rendering was undertaken using high-resolution Z-stack fluorescent images generated using confocal microscopy. To create a detailed rendered surface, the “Surface” tool in the Imaris software package was used. The minimum diameter of the surface was determined based on the average cell diameter estimated using the “Slice view”. The interactive software histogram within the “Create surface” window was used to set a threshold (the minimum diameter of the neuronal surface) to exclusively include neuronal surfaces and to exclude smaller surfaces generated by the background noise. Aggregated neurons within images were separated using the ‘Split surfaces” tool in Imaris. The Imaris software is additionally equipped with “Filter options” to label specific populations of surfaces either manually or automatically and these were utilized in the current study to identify specific cell populations in any given image. Since the RNAScope signal representing Nlgn3 mRNA appears as puncta, the “spot” analysis tool in Imaris was used to create the 3D structure of individual mRNA molecules. The “average diameter of spots” detected was determined using “slice view” in Imaris. Once spots were generated, they were assigned as being located within the neurons using the “Split spots on to surface” tool. The Imaris 9.0 software additionally generates statistical data yielding the number of spots per surface. This option was used to identify the number of mRNA punctate signals within the neurons for the images analysed in the current study.
Statistical analysis
Data were analysed using GraphPad Prism 8.4.1 (GraphPad software, USA). A different group of animals was used to investigate labelling for each set of enteric cellular subpopulations. Frequency distribution analysis was performed to analyse the distribution of Nlgn3 mRNA expression in enteric neurons and glia and a two-sample Kolmogorov-Smirnov test was conducted to determine statistical significance. In general, to estimate the extent of overlap between two distributions for Nlgn3 mRNA expression between genotypes, the magnitude of the Kolmogorov-Smirnov D statistic (distance; D) relative to the critical value is obtained. D is a measure of the maximum distance between the empirical cumulative distribution function of the sample and the cumulative distribution function of the reference distribution (if the value of D is larger than the critical value, the null hypothesis is rejected). Analysis using the GraphPad Prism algorithm provides a P value calculated based on the maximum distance between the cumulative frequency distributions, considering both the D and D critical value.
Investigation of Nlgn, Nrxn and MDGA gene family expression via single cell RNA sequencing datasets
To further validate the RNAScope findings in the current study, we analysed published scRNA sequencing data identifying molecular and functional subtypes of enteric neurons in mice [17] http://ncbi.nlm.nih.gov/sra/SRP258962. ScRNASeq was performed on isolated neurons from myenteric peels of mice aged postnatal day (P)21 and data were validated using immunofluorescence in the ENS of P21-90 mice [17]. The transcriptome of single enteric glia cells isolated from P21 myenteric plexus was assessed using mousebrain.org (web interface for browsing data presented in [16]).
3. Results
Here we characterized the cellular localization of Nlgn3 mRNA in the enteric nervous system of the mouse distal ileum. Using RNAscope and immunofluorescence in WT mice, we first determined the frequency distribution and the average number of Nlgn3 mRNA copies per cell in neuronal subpopulations and glia. Using scRNASeq datasets, we then identified enteric neuronal subsets expressing Nlgn3 and their binding partners, the neurexins (Nrxns1,2 and 3) and MAM Domain Containing Glycosylphosphatidylinositol Anchor genes (MDGA1 and 2). Subsequently, we investigated whether the R451C missense mutation in Nlgn3 impacts cellular expression profiles in the enteric nervous system of the mouse ileum using RNAscope and immunofluorescence.
Nlgn3 mRNA is expressed in most distal ileal submucosal neurons
All submucosal neurons in distal ileal preparations were labelled with the pan-neuronal marker Hu. We then analysed the distribution of Nlgn3 mRNA expression in somata of individual submucosal neurons using RNAscope (Figure 1 A1-A6). Of 1,658 submucosal neurons assessed in 14 WT mice, 1,555 (94%) expressed Nlgn3 mRNA (Figure 1 A7). In addition to Nlgn3 mRNA localized to the cell soma, Nlgn3 mRNA expression was also observed in neuronal and glial fibers (Supplementary Figure S2).
To assess for differential distribution patterns of Nlgn3 mRNA within the submucosal plexus, the expression profile of Nlgn3 mRNA in cholinergic neurons (labelled using ChAT immunofluorescence) was determined in 298 neurons from 7 WT mice. Almost all cholinergic neurons express Nlgn3 mRNA (273 of 298 neurons; 92%; Figure 1 B1-B8). The distribution profile of Nlgn3 mRNA expression in cholinergic neurons was similar to Nlgn3 mRNA expression in all submucosal neurons (i.e., labelled by the pan-neuronal marker, Hu; p=0.09; Figure 1 B9). When we assessed Nlgn3 mRNA copy number per cell, we found that Nlgn3 mRNA-expressing cholinergic submucosal neurons contain 30.1 ± 3.8 copies of Nlgn3 mRNA and submucosal neurons contain 29.0 ± 1.8 copies (n=298 ChAT and n=1,014 neurons; p=0.7; t=0.29, df=1310; Figure 1 B10).
Next, we measured the expression of Nlgn3 mRNA in 411 non-cholinergic (VIP-containing) submucosal neurons from 7 WT mice (Figure 1 C1-C8). Nearly all (400 of 411, 97%) VIP-positive neurons express Nlgn3 mRNA. The distribution profile of Nlgn3 mRNA expression levels in VIP neurons, however, is skewed to the right and significantly different from the distribution of Nlgn3 mRNA in submucosal neurons (p<0.0001) (Figure 1 C9). Neurons that co-express Nlgn3 mRNA and VIP contain 28.5 ± 1.3 Nlgn3 mRNA copies per neuron (n=411 neurons) which is similar to the number of Nlgn3 mRNA copies expressed in all submucosal neurons sampled from the same preparations overall (25.4 ± 1.0 copies per neuron, n=644 neurons; p=0.06, t=1.8, df=1053) (Figure 1 C10).
Myenteric neurons express Nlgn3 mRNA
The subcellular distribution of Nlgn3 mRNA in the distal ileum was analysed in 3,788 myenteric neurons from 14 WT mice. Most myenteric neurons (3297 of 3788, 87%) expressed Nlgn3 mRNA within the cell soma. (Figure 2 A1-A6). Overall, myenteric neurons expressing Nlgn3 mRNA contained 16.1 ± 0.2 copies of Nlgn3 mRNA in the cell soma (Figure 2 A7).
To determine whether Nlgn3 mRNA is differentially expressed in myenteric neuronal subtypes, we labelled subpopulations using immunofluorescence for cell type-specific markers. Expression data showed that 475 of 550 (86%) of calretinin-immunoreactive neurons express Nlgn3 mRNA. Frequency distribution analysis of Nlgn3 mRNA expression levels in calretinin neurons showed a similar profile to the total myenteric neuronal population (total neurons; n=1484, calretinin-positive neurons; n=550; p=0.2). Nlgn3 mRNA copy numbers in calretinin neurons did not differ significantly from myenteric neuron populations (calretinin-positive neurons; 17.0 ± 0.6 mRNA copies, total neurons; 15.6 ± 0.3 mRNA copies; p=0.3; t=2.1, df=2032; Figure 2 B1-B10).
Co-labelling of NOS-expressing myenteric neurons with RNAScope revealed that 514 of 593 (87%) of NOS neurons express Nlgn3 mRNA in the cell soma. The frequency distribution of Nlgn3 mRNA levels in NOS-containing neurons was similar to that of total neurons in the myenteric plexus (NOS positive neurons; n=593, total neurons; n=2304, D=0.05; p=0.09). Individual NOS-containing neuron contained an average of 17.6 ± 0.9 Nlgn3 mRNA copies (n=593 neurons), which was similar to the number of copies expressed in total myenteric neurons (17.5 ± 0.4 Nlgn3 mRNA copies, n=2304 neurons; p=0.9; t=0.07, df=2895; Figure 2 C1-C10) and in calretinin-positive neurons, as detailed above.
Enteric glia express Nlgn3
Recent studies revealed that Nlgn3 is also expressed in non-neuronal (enteroendocrine) cells in the intestine [15] but whether Nlgn3 is expressed in enteric glia has not been assessed. We therefore labelled submucosal glia using an antiserum against the calcium-binding protein, S100β and assessed for the co-presence of Nlgn3 mRNA (Figure 3 A1-A4). Overall, we found that 31% (70 of a total of 226 cells in 6 WT mice) of submucosal glial cells in the mouse ileum express Nlgn3 mRNA (Figure 3 A1-A9). Similarly, Nlgn3 mRNA expression in ileal myenteric glial cells (1203 glial cells in 4 WT mice) was assessed (Figure 3 B1-B8). In the myenteric plexus, 55% of glial cells (666 of 1203 cells) expressed Nlgn3 mRNA (Figure 3 B9). This higher proportion of myenteric glia represents approximately a 3-fold enrichment of Nlgn3 expression compared to submucosal Nlgn3-expressing glia and may provide a useful way to chemically identify and assist in identifying the function of glial subtypes in the gut.
ScRNASeq confirmation of Nlgn3 expression in enteric neurons and glia
Our dual RNAScope/immunocytochemistry findings show that most enteric neurons express Nlgn3 mRNA in mice. To confirm this finding, we investigated both the distribution and levels of Nlgn3 expression in a published scRNASeq dataset describing molecular cellular subtypes in the juvenile mouse small intestine [17]. In line with our observations using dual RNAScope/immunohistochemistry, scRNASeq data analysis shows Nlgn3 expression in multiple enteric neuronal subtypes including putative intrinsic primary afferent neurons, excitatory and inhibitory motor neurons and interneurons. Given that NLGN3 is thought to form heterodimers, predominantly with NLGN1 in the brain [19], expression patterns for Nlgn1 and Nlgn2 in the scRNASeq atlas were also analysed. Interestingly, unlike in the brain, Nlgn2 and Nlgn3 have largely overlapping expression profiles in the enteric nervous system (Figure 4A). Nlgn1 is expressed in most enteric neuron types, but this expression is less prominent in the ENC4, 11, 12 cell classes. Analysis of the scRNASeq atlas showed that although Nlgn3 is detected across all neuronal clusters, Nlgn3 expression levels are highest in ENC6 cells (Figure 4B). Similarly, Nlgn3 is expressed in the highest proportion of cells in ENC6 (approximately 50% of ENC6 cells express Nlgn3) compared to other ENCs (Figure 4C). Based on morphological attributions and projection patterns, ENC6 was validated to correspond to IPANs (intrinsic sensory neurons) of the myenteric plexus [17]. These findings validate our dual RNAScope/immunohistochemistry results to confirm wide-spread expression of Nlgn3 mRNA in the enteric nervous system and suggest that Nlgn3 is highly expressed in some IPANs.
The expression patterns of neuroligin binding partners provide insight into their function in the enteric nervous system. Neurexins are major binding partners of the neuroligin gene family in the brain [19]. Within the enteric nervous system, Nrxn1 and Nrxn2 are almost ubiquitously expressed across all ENCs including robust expression in ENC6 (IPANs). Specifically, Nrxn1 expression levels are highest in ENC11 (interneurons; INs) whereas Nrxn2 expression is highest in ENC6 (IPANs), closely followed by strong expression levels in ENCs 1,2 and 4 (putative motor neurons; MNs), 5 (INs) and 7 (IPANs/INs). Nrxn3 expression levels are highest in ENC 6,7 (IPANs/INs) and 10 (INs) but less prominent in other enteric neuronal subtypes (Figure 4C). As lower affinity binding partners of neuroligins, MAM Domain Containing Glycosylphosphatidylinositol Anchor genes (Mgda1 and Mdga2) are expressed at varying levels in myenteric neuron populations (Supplementary Figure S3).
The distribution and expression of Nlgn3 alongside Nlgn1 and -2 was also examined in myenteric glia based on scRNASeq data. Nlgn1 was sparsely expressed in myenteric glial cells (Figure 5A). Although Nlgn2 was present across all myenteric glia (in a greater proportion of cells than Nlgn1), fewer than 50% of cells assayed contained Nlgn2 (Figure 5B). In stark contrast, Nlgn3 was strongly expressed across each myenteric glial cluster (Figure 5C,D). However, of the 10,535 total myenteric glial cells counted, only 4,054 cells (38%) express Nlgn3 mRNA according to the scRNASeq analyses compared to 65% of glial cells as detected using RNAScope. These differences may be due to the lower sensitivity of the scRNASeq approach utilized [17] compared to RNAScope. None of the Nrxns (Figure 5E-G) nor Mgda1 or Mdga2 (Supplementary Figure S4) were notably expressed in myenteric glia.
The R451C mutation selectively reduces Nlgn3 mRNA expression
The R451C mutation reduces Nlgn3 mRNA expression in cholinergic submucosal neurons but not in VIPergic neurons.
We next investigated Nlgn3 mRNA expression in wild type and Nlgn3R451C mutant mouse ileum using dual RNAScope and immunocytochemistry plus Imaris-based quantification (Figure 6 A1-A6). Frequency distribution analysis of Nlgn3 mRNA expression in the submucosal plexus showed that Nlgn3 mRNA expression is significantly different and the distribution was skewed to the left in Nlgn3R451C mutant mice compared to WT (1985 neurons in 14 WT and 1300 neurons in 10 Nlgn3R451C mice were analysed; p<0.0001; Figure 6 A7). In Nlgn3R451C mutant mice, submucosal neuronal cell bodies (n=1300 neurons) express 21.6 ± 0.8 copies of Nlgn3 mRNA, indicating that significantly less Nlgn3 mRNA is present in this neuronal subset in mutants than in WT mice (24.8 ± 1.0 copies, n=1985 neurons), p=0.02; t=2.2, df=3283 Figure 6 A8).
We then characterised the expression of Nlgn3 mRNA in different submucosal neuronal populations (Figure 6 B1-B8). Of a total of 241 ChAT neurons in 7 Nlgn3R451C mice, only 69% (167 neurons) of neurons expressed Nlgn3 mRNA. The frequency distribution analysis revealed that the distribution of Nlgn3 mRNA in ChAT neurons in mutant mice is skewed to the left compared to WT (n=298 neurons in 5 WT mice, n=241 neurons in Nlgn3R451C mice; D=0.32; p<0.0001; Figure 6 B9). Specifically, in Nlgn3R451C mutant mice Nlgn3 mRNA expression is significantly reduced in ChAT neurons compared to WT (WT; 30.1 ± 3.8 copies, n=298, Nlgn3R451C mutant mice; 16.2 ± 2.9 copies, n=241, p=0.006; t=2.7, df=537; Figure 6 B10).
The impact of the R451C mutation on Nlgn3 mRNA production in VIP-expressing neurons was also evaluated (Figure 6 C1-C8). The frequency distribution of the Nlgn3 mRNA copy numbers in VIP neurons in ileal tissue from Nlgn3R451C mice (n=623 neurons in 7 WT mice were analysed as well as n=449 neurons in 5 Nlgn3R451C mice), is skewed to the left and differs significantly from WT (p<0.0001; Figure 6 C9). In mutant mice, only 82% (370 of 449) of VIP-containing submucosal neurons express Nlgn3 mRNA in the cell soma. But of those VIP-expressing neurons, Nlgn3 mRNA expression levels showed a trend to increase in mutant mice compared to WT (27.1 ± 1.1 versus 30.4 ± 1.2 mRNA copies per cell; n=623 and 449 cells; WT and mutant mice respectively; p=0.05; t=1.9, df=1070 Figure 6 C10).
The R451C mutation decreases Nlgn3 mRNA expression in calretinin and nNOS labeled myenteric neurons
We evaluated the impact of the R451C mutation on Nlgn3 mRNA expression distribution profiles and mRNA copy number in the ileal myenteric plexus (Figure 7 A1-A6). Using the same approach, we compared Nlgn3 mRNA expression in total neurons as well as calretinin and NOS-immunoreactive myenteric neurons in WT and Nlgn3R451C mutant mice. We found that the frequency distribution of Nlgn3 mRNA expression in the myenteric plexus was significantly different (skewed to the left) between WT and Nlgn3R451C mice (p<0.0001; Figure 7 A7). Similar to the findings in the submucosal plexus, Nlgn3 mRNA copy numbers in myenteric neurons were significantly reduced in Nlgn3R451C mutant mice compared to WT (WT; 16.8 ± 0.3 mRNA copies per cell, Nlgn3R451C mice; 11.3 ± 0.2 mRNA copies per cell; n=3,788, n=2,825, WT and Nlgn3R451C mutant mice respectively; p<0.0001; t=15.7, df=6611; Figure 7 A8).
We also assessed for differences in Nlgn3 mRNA expression in calretinin-immunoreactive myenteric neurons (Figure 7 B1-B8). The frequency distribution of Nlgn3 mRNA expression in calretinin neurons in Nlgn3R451C mice is significantly different (skewed to the left) from the WT (p<0.0001, Figure 7 B9). In WT, 86% (473 of 550) of calretinin neurons express Nlgn3 mRNA, whereas in Nlgn3R451C mutant mice, 78% of these neurons co-express Nlgn3 mRNA. Significantly fewer Nlgn3 mRNA copies were detected in calretinin-immunoreactive neurons in Nlgn3R451C mutant mice compared to WT (WT; 17.0 ± 0.6 mRNA copies, n=550 neurons, Nlgn3R451C; 12.3 ± 0.42 mRNA copies, n=685 neurons; p<0.0001; t=6.6, df=1233; Figure 7 B10).
We previously reported an increased proportion of NOS neurons in the myenteric plexus of the jejunum and caecum in Nlgn3R451C mice [7,20], suggesting that NLGN3 might play an important role in NOS signalling in the myenteric plexus. We therefore analysed the impact of the R451C mutation on Nlgn3 mRNA expression in NOS-immunoreactive myenteric neurons (Figure 7 C1-C8). The frequency distribution of Nlgn3 mRNA distribution in NOS immunoreactive neurons in Nlgn3R451C mice differs significantly from WT with this distribution being skewed to the left (p<0.0001, Figure 7 C9). In WT, about 87% of NOS-expressing neurons (i.e., 515 of 593 neurons) express Nlgn3. Of a total of 331 NOS-immunoreactive neurons assessed in Nlgn3R451C mutant mice, about 73% (241 neurons) co-express Nlgn3 mRNA. Nlgn3 mRNA copy numbers were significantly reduced in myenteric NOS neurons in mutant mice compared to WT (WT; 17.6 ± 0.9 mRNA copies per cell, n=593 neurons, Nlgn3R451C mutant mice; 9.0 ± 0.4 mRNA copies per cell, n=331 neurons, p<0.0001, t=6.6, df=922; Figure 7 C10).
Reduced Nlgn3 mRNA expression in myenteric but not submucosal glia in Nlgn3R451C mice
As described earlier, Nlgn3 mRNA is expressed in both submucosal and myenteric glia in the WT mouse ileum. Nlgn3 mRNA expression was assessed in of the ileal submucosal plexus in WT (n=6) and Nlgn3R451C (n=5) mutant mice, respectively (Figure 8 A1-A8). In Nlgn3R451C mutant mice, 42% (73 of a total of 173) of submucosal glial cells expressed Nlgn3 mRNA. The frequency distribution data of Nlgn3 mRNA expression in submucosal glial cells show similar levels of Nlgn3 mRNA in WT and Nlgn3R451C (p=0.09, Figure 8 A9). In submucosal glia, there was a trend for an increase in mRNA copy number in Nlgn3R451C mutants (2.8 ± 0.3 mRNA copies and 5.0 ± 1.2 mRNA copies in WT and Nlgn3R451C mice respectively, p=0.05; t=1.9, df=397, Figure 8 A10).
We also compared Nlgn3 mRNA expression in myenteric glia in WT and Nlgn3R451C mutant mice. In Nlgn3R451C mutant mice, only 37% (496 of a total of 1318) of myenteric glial cells expressed Nlgn3 mRNA (Figure 8 B1-B8) and the frequency distribution of Nlgn3 mRNA in myenteric glia in mutant mice was significantly different (skewed to the left) to WT (n=1203 glial cells in 5 WT mice, n=1318 cells in 5 Nlgn3R451C mice; p=0.001, Figure 8 B9). Nlgn3 mRNA expression was reduced in myenteric glia in Nlgn3R451C mice compared to WT (WT; 14.2 ± 1.2 mRNA copies/cell, n=1203 cells, Nlgn3R451C; 4.4 ± 0.1 mRNA copies/cell, n=1,318 cells, p<0.0001, t=8.2, df=2519, Figure 8 B10).

Table 3.
Comparison of Nlgn3 copy number expression in neuronal and glial subtypes in the submucosal and myenteric plexus in Nlgn3R451C and WT mice.
Table 3.
Comparison of Nlgn3 copy number expression in neuronal and glial subtypes in the submucosal and myenteric plexus in Nlgn3R451C and WT mice.
| Enteric-neuronal plexus | Cell type | WT(mean ± SEM) |
Nlgn3R451C(mean± SEM) |
p value |
| Submucosal plexus | Hu | 24.8 ± 1.0 | 21.6 ± 0.8 | 0.02 |
| VIP | 27.1 ± 1.1 | 30.4 ± 1.2 | 0.05 | |
| ChAT | 30.1 ± 3.8 | 16.2 ± 2.9 | 0.006 | |
| S100b | 2.8 ± 0.3 | 5.0 ± 1.2 | 0.05 | |
| Myenteric plexus | Hu | 16.8 ± 0.3 | 11.3 ± 0.2 | <0.0001 |
| Calretinin | 17.0 ± 0.6 | 12.3 ± 0.42 | <0.0001 | |
| NOS | 17.6 ± 0.9 | 9.0 ± 0.4 | <0.0001 | |
| S100b | 14.2 ± 1.2 | 4.4 ± 0.1 | <0.0001 |
4. Discussion
In this study, we showed that most submucosal and myenteric neurons in the mouse ileum express Nlgn3 mRNA. For the first time, we revealed that Nlgn3 mRNA is expressed in ileal submucosal and myenteric glia. In addition, we report that the autism-associated Nlgn3 R451C mutation reduces Nlgn3 mRNA expression in both submucosal and myenteric neurons as well as myenteric glia in the mouse ileum.
Most enteric neurons express Nlgn3 mRNA
Although there is ample evidence for the existence of prominent post-synaptic densities within the ENS [21,22,23,24], the molecular characterization remains to be fully characterized. In the rodent brain, neuroligin-3 is expressed at both excitatory and inhibitory synapses [8] and closely associated with other PDZ-associated molecules to mediate postsynaptic signal transduction. There is evidence that PDZ domain proteins such as PSD95 [25,26], PSD93 [26] as well as the cell adhesion molecules L1 [27] and neuroligin-1 [28] are expressed in the ENS.
Cholinergic transmission
It is well established that NLGN3 is involved in both excitatory and inhibitory synaptic transmission in the brain. However, relatively little is known about the distribution of post-synaptic proteins associated with excitatory (predominantly mediated via nicotinic acetylcholine receptors (nAChRs)) and inhibitory synapses in the ENS. We reveal that most cholinergic submucosal neurons also contain Nlgn3 mRNA. Within the enteric neural circuitry, acetylcholine is the primary excitatory neurotransmitter utilised by cholinergic secretomotor neurons, excitatory muscle motor neurons, ascending interneurons, descending interneurons and intrinsic sensory neurons [29,30,31,32]. Since virtually all submucosal neurons receive synaptic inputs via cholinergic synapses [33], Nlgn3 is likely present in cholinergic synapses in the submucosal plexus. All submucosal neurons receive cholinergic fast excitatory post synaptic potentials hence can be expected to have post-synaptic densities with clusters of nicotinic receptors and NLGNs as a part of the synaptic structure. Although there is no available evidence of NLGN3 expression in the cholinergic system of the central or peripheral nervous systems, expression of other NLGN subtypes in cholinergic synapses has been reported. For example, NLGN2 is expressed at the postsynaptic membrane of cholinergic synapses in the mouse brain [34] and NLGN1 is present in cholinergic synapses in the chick ciliary ganglion [35]. Since NLGN3 and other NLGN isoforms can be colocalized in the same synapse [8], NLGN3 might be co-expressed with NLGN1 and NLGN2 in cholinergic synapses in the submucosal plexus.
VIP-expressing neurons
We also revealed that Nlgn3 mRNA is expressed in VIP-expressing submucosal neurons in the mouse ileum. In the submucosal plexus, VIP is a primary neurotransmitter of secretomotor neurons and stimulates intestinal secretion [33,36]. In the myenteric plexus, VIP is expressed in inhibitory muscle motor neurons and interneurons [37,38]. VIP also acts as a co-transmitter alongside ACh and NO in a subset of descending interneurons in the mouse and guinea pig ileum [38,39,40]. Moreover, an excitatory role for the Vasoactive Intestinal Peptide 1 (VPAC1) receptor on cholinergic neurons has been identified in the myenteric plexus [41]. These findings open a novel avenue for research in Nlgn3R451C mice since the expression profiles of NLGN3 or other NLGN isoforms in VIP-containing synapses have not been reported to date in either the CNS or the peripheral nervous system.
NOS-expressing neurons
One of the major findings of this study is that most NO-containing enteric neurons express Nlgn3 mRNA in the cell soma. Nitric oxide (NO) is the predominant inhibitory neurotransmitter to the smooth muscle in the enteric nervous system [38,39] and is also expressed in descending interneurons together with other neurotransmitters [42]. Given that NLGN complexes anchor postsynaptic densities and PSD95 has at least one nNOS binding PDZ domain, Nlgn3 may contribute to synaptic specialization of enteric NO neurons. However, the distributions of PSD95 and PSD93 have not been well characterized in ENS. It has been shown that PSD93 interacts with nNOS via a PDZ-PDZ domain interaction [26]. Some nNOS expressing myenteric neurons express PSD93 [43]. In agreement with this finding, our data indicating Nlgn3 expression in this neuronal population suggests that Nlgn3 might be expressed at the postsynaptic membrane of nNOS immunoreactive myenteric neurons.
Calretinin-expressing neurons
We also show Nlgn3 mRNA expression in calretinin-expressing myenteric neurons in the mouse ileum. In the enteric nervous system, calretinin is expressed in intrinsic primary afferent neurons, interneurons and excitatory motor neurons innervating the smooth muscle layer [38,39]. In line with these findings, scRNASeq data analysis confirmed expression of Nlgn3 at varying levels in mouse myenteric neurons including putative motor neurons, interneurons and intrinsic primary afferent neurons in the mouse small intestine. It has been reported that the PSD93 protein is localized to calretinin neurons in mouse myenteric plexus [43]. Therefore, it is possible that Nlgn3 is associated with postsynaptic protein complexes within calretinin neurons in the myenteric plexus
Other signalling systems
Given that glutamate and GABA have active roles in the ENS [44,45] alongside the presence of postsynaptic proteins including PSD93 [43], the wide distribution of NLGN3 we observed in the ENS suggests that NLGN3 could also be expressed at glutamatergic and GABAergic synapses within the ENS.
Differential expression of Neuroligin binding partners in the enteric nervous system
When examining scRNASeq data, Nlgn2 and 3 expression profiles are almost identical in all classes of enteric neurons. In contrast with findings in the central nervous system [8] reviewed by [46], this suggests that NLGN3 and NLGN2 co-express in enteric neurons subtypes and we speculate that they may form heterodimers in the enteric nervous system. Expression patterns of the neurexin family of neuroligin binding partners, however, show that Nrxn2 and Nrxn3 are more strongly expressed within the enteric neuronal cell clusters compared with Nrxn1. These findings suggest that Nlgn3 may preferentially bind with Nrxn2 and 3 at synapses in IPANs of the myenteric plexus. In the CNS, NLGNs are present at classical synapses receiving fast EPSPs but most functional evidence suggests that IPANs do not exhibit fast EPSPs. For example, findings from Hibberd and colleagues [47] indicate that IPANs receive fast EPSPs, but in contrast, both [48] and [49] reported that AH neurons do not exhibit fast EPSPs. Nevertheless, it is unclear whether junctions producing slow EPSPs might express neuroligin complexes, therefore, further research is required to identify the role of NLGN3 in IPANs in the ENS.
A role for Nlgn3 in enteric neuronal-glial synapses
For the first time, we reveal that Nlgn3 mRNA is expressed in enteric glia. Intriguingly, scRNAseq analysis showed that most enteric glia express Nlgn3 mRNA, but negligible/low levels of Nlgn1 and Nlgn2. Therefore, we propose that NLGN3 acts as a main adhesion protein in glia for modulating glial-neuron synaptic activity in the enteric nervous system.
Enteric glia play a major role in enteric nervous system-mediated GI functions including mucosal secretion, intestinal permeability, mucosal sensation, GI motility and immune responses [50,51,52] in concert with enteric neurons. Enteric neurons synapse onto enteric glia to regulate enteric nervous system-coordinated GI responses [50,53,54]. Enteric glial-neuronal associations revealed by immunocytochemistry highlight that enteric glia are in close contact with nerve fibers and varicose release sites [55]. Evidence for neuro-glia communication also comes from live imaging experiments reporting the presence of several signalling pathways. Ca2+ imaging studies indicate that neuron-glia communication is regulated by neurally released purines which activate purinergic receptors on enteric glia [54,55,56,57]. In the submucosal plexus, neuron-glial transmission occurs via P2Y1 and P2Y4 receptors [58]. Furthermore, neuronal ATP release via the pannexin-1 channel also influences neuro-glia interactions [59,60]. Based on this evidence, we propose that NLGN3 expressed by both enteric neurons and glia modulate neuro-glia signalling pathways. This hypothesis is well-established in the mouse brain where RNA sequencing of the transcriptome revealed that Nlgn3 transcripts are enriched in glial cell types including astrocytes and oligodendrocytes [61]. Interestingly, NLGNs expressed in astrocytes aid in neuron-astrocyte communication via bi- and tripartite synapses in mice in C. elegans [62] suggesting that a neuronal-glial communication role may not be specific to mice. Similarly, NLGN3 expressed in enteric glia could also be involved in establishing bi-partite and tripartite synapses in the enteric nervous system. Given this evidence, this study strongly suggests that NLGN3 might play a role in synapse formation and modifying synaptic function during neuron-glia communication in the enteric nervous system.
The R451C mutation reduces Nlgn3 mRNA expression in the enteric nervous system
We previously demonstrated that mice expressing the Nlgn3 R451C mutation have faster small intestinal transit and increased numbers of myenteric neurons in the small intestine [7]. How this mutation affects NLGN3 expression in the enteric nervous system, however, is not understood. Here we show that the R451C mutation decreases Nlgn3 mRNA expression in both neurons and glia in the mouse ileal submucosal plexus. For example, cholinergic submucosal neurons in mutant mice contain fewer Nlgn3 mRNA copies compared to WT. These changes could potentially alter cholinergic signalling in the submucosal plexus which could affect functions mediated by cholinergic neurons such as secretion, absorption and mucosal barrier functions. Although there are no reports available on changes to the cholinergic system of the enteric nervous system in ASD, CNS studies showed that altered cholinergic neurons are associated with the pathophysiology of autism [63,64]. Specifically, ASD patient basal forebrain tissues show altered cholinergic neuronal numbers, size and structure [65]. In addition, a decreased plasma concentration of choline, a precursor for acetylcholine has been reported in ASD patients [66,67] and reduced levels of hippocampal cytosolic choline have been correlated with autism severity [66]. In the human GI tract, NLGN3-mediated alterations to the cholinergic system could induce GI dysfunction, however further studies are required to determine effects of the R451C mutation on cholinergic signalling in the enteric nervous system.
In the myenteric plexus of the distal ileum, the R451C mutation reduces Nlgn3 mRNA expression levels in neurons. Specifically, Nlgn3 mRNA expression is substantially reduced in both calretinin and nNOS-immunoreactive neuronal populations. Although the contribution of calretinin neurons to ASD or ASD-related GI pathophysiology is unclear, the significance of calretinin neurons in other ENS-related diseases has been highlighted [68]. As mentioned, an increased proportion of NOS1 expressing neurons has previously has previously been observed in Nlgn3R451C mice [7], and our current findings showing reduced Nlgn3 expression in these cells might contribute to altered NO neuronal signalling in the enteric nervous system in these mice.
Finally, the impact of the R451C mutation on Nlgn3 expression in glia has not previously been reported. Here we show that in Nlgn3R451C mice, Nlgn3 mRNA expression is dramatically reduced in myenteric glia but unchanged in glia located within the submucosal plexus. Since glia play an important role in mediating intestinal functions such as mucosal barrier regulation, gut motility, immune responses and neurotransmission, such a reduction in NLGN3 levels could contribute to GI dysfunction in Nlgn3R451C mutant mice. Therefore, in addition to neuronal dysfunction, glial dysfunction could also contribute to GI pathology in individuals diagnosed with ASD.
5. Conclusions
Here we characterised Nlgn3 mRNA expression and identified effects of the autism-associated R451C mutation in this gene in the enteric nervous system of the mouse ileum. We show that Nlgn3 mRNA is expressed in situ in most enteric neuronal and glial populations of the myenteric and submucosal plexuses in the mouse ileum. Our RNAseq analysis confirms the presence of Nlgn3 in myenteric neuronal subtypes and glia in mice. Intriguingly, we found that Nlgn3, but not Nlgn1 or 2, is expressed at high levels in enteric glia, whereas glial expression of major binding partners for Neuroligins, such as Neurexins 1-3 is negligible. We therefore propose that Neuroligin-3 is an important glial adhesion protein in neuronal-glial synapses in the mouse enteric nervous system. We demonstrate that the R451C mutation differentially reduces Nlgn3 mRNA expression in the enteric nervous system; particularly in cholinergic submucosal neurons, myenteric calretinin and NOS1 expressing neurons and myenteric glia in the mouse ileum. Taken together, these findings suggest that Neuroligin-3 plays a role in regulating enteric nervous system-mediated GI physiology and that changes to Nlgn3 mRNA expression could contribute to GI dysfunction in the Nlgn3R451C mouse model of autism.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, M.H, E.H-Y, J.B and A.F.; methodology, M.H.; software, M.H.; validation, M.H., E.H-Y, J.B, E.C and A.F.; formal analysis, M.H.; investigation, E.H-Y, J.B and A.F; resources, M.H.; data curation, M.H.; writing - original draft preparation, M.H., E.H-Y, J.B, E.C and A.F; writing—review and editing, M.H.; visualization, E.H-Y, J.B and A.F.; supervision, E.H-Y, J.B and A.F.; project administration, E.H-Y, J.B and A.F.; funding acquisition, E.H-Y, J.B and A.F. All authors have read and agreed to the published version of the manuscript.
Funding
MH received a Melbourne University PhD Stipend. This work was supported by a National Health and Medical Research Council ideas grant (APP2003848), an Australian Research Council Future Fellowship (FT160100126), an RMIT Vice Chancellor’s Senior Research Fellowship to EH-Y. JB received an NHMRC project grant (APP1158952).
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Ethics Committee of THE UNIVERSITY OF MELBOURNE (protocol code 1914843 and 10/09/2019).
Data Availability Statement
Data supporting the findings of this study are available from the corresponding author upon request.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. ELH-Y and AEF are scientific advisors for Vernx. No funding for this study was provided by Vernx.
References
- Tabuchi, K., et al., A neuroligin-3 mutation implicated in autism increases inhibitory synaptic transmission in mice. Science, 2007. 318(5847): p. 71-6. [CrossRef]
- Etherton, M., et al., Autism-linked neuroligin-3 R451C mutation differentially alters hippocampal and cortical synaptic function. Proc Natl Acad Sci U S A, 2011. 108(33): p. 13764-9. [CrossRef]
- Foldy, C., R.C. Malenka, and T.C. Sudhof, Autism-associated neuroligin-3 mutations commonly disrupt tonic endocannabinoid signaling. Neuron, 2013. 78(3): p. 498-509. [CrossRef]
- Rothwell, P.E., et al., Autism-associated neuroligin-3 mutations commonly impair striatal circuits to boost repetitive behaviors. Cell, 2014. 158(1): p. 198-212. [CrossRef]
- Hosie, S., et al., Altered Amygdala Excitation and CB1 Receptor Modulation of Aggressive Behavior in the Neuroligin-3(R451C) Mouse Model of Autism. Front Cell Neurosci, 2018. 12: p. 234. [CrossRef]
- Leembruggen, A.J.L., et al., Colonic dilation and altered ex vivo gastrointestinal motility in the neuroligin-3 knockout mouse. Autism Res, 2019. [CrossRef]
- Hosie, S., et al., Gastrointestinal dysfunction in patients and mice expressing the autism-associated R451C mutation in neuroligin-3. Autism Res, 2019. 12(7): p. 1043-1056. [CrossRef]
- Budreck, E.C. and P. Scheiffele, Neuroligin-3 is a neuronal adhesion protein at GABAergic and glutamatergic synapses. Eur J Neurosci, 2007. 26(7): p. 1738-48. [CrossRef]
- Venkatesh, H.S., et al., Neuronal Activity Promotes Glioma Growth through Neuroligin-3 Secretion. Cell, 2015. 161(4): p. 803-16. [CrossRef]
- Gilbert, M., et al., Neuroligin 3 is a vertebrate gliotactin expressed in the olfactory ensheathing glia, a growth-promoting class of macroglia. Glia, 2001. 34(3): p. 151-64. [CrossRef]
- Jamain, S., et al., Mutations of the X-linked genes encoding neuroligins NLGN3 and NLGN4 are associated with autism. Nat Genet, 2003. 34(1): p. 27-9. [CrossRef]
- Sanders, S.J., et al., Multiple recurrent de novo CNVs, including duplications of the 7q11.23 Williams syndrome region, are strongly associated with autism. Neuron, 2011. 70(5): p. 863-85. [CrossRef]
- Levy, D., et al., Rare de novo and transmitted copy-number variation in autistic spectrum disorders. Neuron, 2011. 70(5): p. 886-97. [CrossRef]
- Zhang, Q., et al., Expression of neurexin and neuroligin in the enteric nervous system and their down-regulated expression levels in Hirschsprung disease. Mol Biol Rep, 2013. 40(4): p. 2969-75. [CrossRef]
- Bohorquez, D.V., et al., Neuroepithelial circuit formed by innervation of sensory enteroendocrine cells. J Clin Invest, 2015. 125(2): p. 782-6. [CrossRef]
- Zeisel, A., et al., Molecular Architecture of the Mouse Nervous System. Cell, 2018. 174(4): p. 999-1014 e22. [CrossRef]
- Morarach, K., et al., Diversification of molecularly defined myenteric neuron classes revealed by single-cell RNA sequencing. Nat Neurosci, 2021. 24(1): p. 34-46. [CrossRef]
- Wang, F., et al., RNAscope: a novel in situ RNA analysis platform for formalin-fixed, paraffin-embedded tissues. J Mol Diagn, 2012. 14(1): p. 22-9. [CrossRef]
- Sudhof, T.C., Neuroligins and neurexins link synaptic function to cognitive disease. Nature, 2008. 455(7215): p. 903-11. [CrossRef]
- Sharna, S.S., et al., Altered Caecal Neuroimmune Interactions in the Neuroligin-3(R451C) Mouse Model of Autism. Front Cell Neurosci, 2020. 14: p. 85. [CrossRef]
- Gabella, G., Fine structure of the myenteric plexus in the guinea-pig ileum. J Anat, 1972. 111(Pt 1): p. 69-97.
- Llewellyn-Smith, I.J., M. Costa, and J.B. Furness, Light and electron microscopic immunocytochemistry of the same nerves from whole mount preparations. J Histochem Cytochem, 1985. 33(9): p. 857-66. [CrossRef]
- Young, H.M. and J.B. Furness, Ultrastructural examination of the targets of serotonin-immunoreactive descending interneurons in the guinea pig small intestine. J Comp Neurol, 1995. 356(1): p. 101-14.
- Pompolo, S. and J.B. Furness, Ultrastructure and synaptic relationships of calbindin-reactive, Dogiel type II neurons, in myenteric ganglia of guinea-pig small intestine. J Neurocytol, 1988. 17(6): p. 771-82. [CrossRef]
- Chaudhury, A., X.D. He, and R.K. Goyal, Role of PSD95 in membrane association and catalytic activity of nNOSalpha in nitrergic varicosities in mice gut. Am J Physiol Gastrointest Liver Physiol, 2009. 297(4): p. G806-13.
- Brenman, J.E., et al., Cloning and characterization of postsynaptic density 93, a nitric oxide synthase interacting protein. J Neurosci, 1996. 16(23): p. 7407-15. [CrossRef]
- Anderson, R.B., et al., The cell adhesion molecule l1 is required for chain migration of neural crest cells in the developing mouse gut. Gastroenterology, 2006. 130(4): p. 1221-32. [CrossRef]
- Zheng, Y., et al., Down-regulation of fibronectin and the correlated expression of neuroligin in hirschsprung disease. Neurogastroenterol Motil, 2017. 29(12). [CrossRef]
- Brookes, S.J., Classes of enteric nerve cells in the guinea-pig small intestine. Anat Rec, 2001. 262(1): p. 58-70.
- Grider, J.R., Neurotransmitters mediating the intestinal peristaltic reflex in the mouse. J Pharmacol Exp Ther, 2003. 307(2): p. 460-7. [CrossRef]
- Gwynne, R.M. and J.C. Bornstein, Synaptic transmission at functionally identified synapses in the enteric nervous system: roles for both ionotropic and metabotropic receptors. Curr Neuropharmacol, 2007. 5(1): p. 1-17. [CrossRef]
- Brookes, S.J., P.A. Steele, and M. Costa, Identification and immunohistochemistry of cholinergic and non-cholinergic circular muscle motor neurons in the guinea-pig small intestine. Neuroscience, 1991. 42(3): p. 863-78. [CrossRef]
- Foong, J.P., et al., Properties of cholinergic and non-cholinergic submucosal neurons along the mouse colon. J Physiol, 2014. 592(4): p. 777-93. [CrossRef]
- Takacs, V.T., T.F. Freund, and G. Nyiri, Neuroligin 2 is expressed in synapses established by cholinergic cells in the mouse brain. PLoS One, 2013. 8(9): p. e72450. [CrossRef]
- Ross, B.S. and W.G. Conroy, Capabilities of neurexins in the chick ciliary ganglion. Dev Neurobiol, 2008. 68(3): p. 409-19. [CrossRef]
- Mongardi Fantaguzzi, C., et al., Identification of neuron types in the submucosal ganglia of the mouse ileum. Cell Tissue Res, 2009. 336(2): p. 179-89. [CrossRef]
- Keef, K.D., et al., Relationship between nitric oxide and vasoactive intestinal polypeptide in enteric inhibitory neurotransmission. Neuropharmacology, 1994. 33(11): p. 1303-14. [CrossRef]
- Sang, Q. and H.M. Young, Chemical coding of neurons in the myenteric plexus and external muscle of the small and large intestine of the mouse. Cell Tissue Res, 1996. 284(1): p. 39-53. [CrossRef]
- Qu, Z.D., et al., Immunohistochemical analysis of neuron types in the mouse small intestine. Cell Tissue Res, 2008. 334(2): p. 147-61. [CrossRef]
- Costa, M., et al., Neurochemical classification of myenteric neurons in the guinea-pig ileum. Neuroscience, 1996. 75(3): p. 949-67. [CrossRef]
- Fung, C., et al., VPAC1 receptors regulate intestinal secretion and muscle contractility by activating cholinergic neurons in guinea pig jejunum. Am J Physiol Gastrointest Liver Physiol, 2014. 306(9): p. G748-58. [CrossRef]
- Young, H.M., J.B. Furness, and J.M. Povey, Analysis of connections between nitric oxide synthase neurons in the myenteric plexus of the guinea-pig small intestine. J Neurocytol, 1995. 24(4): p. 257-63. [CrossRef]
- Swaminathan, M.F., J. & Hill-Yardin, Elisa, Bornstein, Joel., PSD-93 is expressed in most, but not all, myenteric neurons in the mouse colon. Neurogastroenterology and Motility, 2016. 28: p. 55-55.
- Swaminathan, M., et al., Endogenous Glutamate Excites Myenteric Calbindin Neurons by Activating Group I Metabotropic Glutamate Receptors in the Mouse Colon. Front Neurosci, 2019. 13: p. 426. [CrossRef]
- Seifi, M., et al., Molecular and functional diversity of GABA-A receptors in the enteric nervous system of the mouse colon. J Neurosci, 2014. 34(31): p. 10361-78. [CrossRef]
- Sudhof, T.C., Synaptic Neurexin Complexes: A Molecular Code for the Logic of Neural Circuits. Cell, 2017. 171(4): p. 745-769. [CrossRef]
- Hibberd, T.J., et al., Synaptic activation of putative sensory neurons by hexamethonium-sensitive nerve pathways in mouse colon. Am J Physiol Gastrointest Liver Physiol, 2018. 314(1): p. G53-G64. [CrossRef]
- Foong, J.P., et al., Myenteric neurons of the mouse small intestine undergo significant electrophysiological and morphological changes during postnatal development. J Physiol, 2012. 590(10): p. 2375-90. [CrossRef]
- Nurgali, K., M.J. Stebbing, and J.B. Furness, Correlation of electrophysiological and morphological characteristics of enteric neurons in the mouse colon. J Comp Neurol, 2004. 468(1): p. 112-24. [CrossRef]
- Gulbransen, B.D. and K.A. Sharkey, Novel functional roles for enteric glia in the gastrointestinal tract. Nat Rev Gastroenterol Hepatol, 2012. 9(11): p. 625-32. [CrossRef]
- Grubisic, V., et al., Enteric glia regulate gut motility in health and disease. Brain Res Bull, 2018. 136: p. 109-117. [CrossRef]
- Broadhead, M.J., et al., Ca2+ transients in myenteric glial cells during the colonic migrating motor complex in the isolated murine large intestine. J Physiol, 2012. 590(2): p. 335-50. [CrossRef]
- Sharkey, K.A., Emerging roles for enteric glia in gastrointestinal disorders. J Clin Invest, 2015. 125(3): p. 918-25. [CrossRef]
- Gulbransen, B.D. and K.A. Sharkey, Purinergic neuron-to-glia signaling in the enteric nervous system. Gastroenterology, 2009. 136(4): p. 1349-58. [CrossRef]
- Boesmans, W., et al., Imaging neuron-glia interactions in the enteric nervous system. Front Cell Neurosci, 2013. 7: p. 183. [CrossRef]
- Gomes, P., et al., ATP-dependent paracrine communication between enteric neurons and glia in a primary cell culture derived from embryonic mice. Neurogastroenterol Motil, 2009. 21(8): p. 870-e62. [CrossRef]
- Delvalle, N.M., et al., Communication Between Enteric Neurons, Glia, and Nociceptors Underlies the Effects of Tachykinins on Neuroinflammation. Cell Mol Gastroenterol Hepatol, 2018. 6(3): p. 321-344. [CrossRef]
- Fung, C. and P. Vanden Berghe, Functional circuits and signal processing in the enteric nervous system. Cell Mol Life Sci, 2020. 77(22): p. 4505-4522. [CrossRef]
- Wang, J. and G. Dahl, Pannexin1: a multifunction and multiconductance and/or permeability membrane channel. Am J Physiol Cell Physiol, 2018. 315(3): p. C290-C299. [CrossRef]
- Hanstein, R., et al., Glial pannexin1 contributes to tactile hypersensitivity in a mouse model of orofacial pain. Sci Rep, 2016. 6: p. 38266. [CrossRef]
- Zhang, Y., et al., An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. J Neurosci, 2014. 34(36): p. 11929-47. [CrossRef]
- Hillen, A.E.J., J.P.H. Burbach, and E.M. Hol, Cell adhesion and matricellular support by astrocytes of the tripartite synapse. Prog Neurobiol, 2018. 165-167: p. 66-86. [CrossRef]
- Perry, E.K., et al., Cholinergic activity in autism: abnormalities in the cerebral cortex and basal forebrain. Am J Psychiatry, 2001. 158(7): p. 1058-66. [CrossRef]
- Deutsch, S.I., et al., Cholinergic abnormalities in autism: is there a rationale for selective nicotinic agonist interventions? Clin Neuropharmacol, 2010. 33(3): p. 114-20.
- Kemper, T.L. and M.L. Bauman, Neuropathology of infantile autism. Mol Psychiatry, 2002. 7 Suppl 2: p. S12-3.
- Sokol, D.K., et al., Hydrogen proton magnetic resonance spectroscopy in autism: preliminary evidence of elevated choline/creatine ratio. J Child Neurol, 2002. 17(4): p. 245-9. [CrossRef]
- Wenk, G.L. and B. Hauss-Wegrzyniak, Altered cholinergic function in the basal forebrain of girls with Rett syndrome. Neuropediatrics, 1999. 30(3): p. 125-9.
- Barshack, I., et al., The loss of calretinin expression indicates aganglionosis in Hirschsprung's disease. J Clin Pathol, 2004. 57(7): p. 712-6.
Figure 1.
The distribution of Nlgn3 mRNA in ileal submucosal neurons. Confocal images of A1: submucosal neurons labelled with Hu pan neuronal marker; A2: Nlgn3 mRNA expression. A3: merged image. A4: 3D reconstruction of submucosal neurons; A5: cellular Nlgn3 mRNA expression; A6: merged image. A7: Frequency distribution of Nlgn3 mRNA expression in submucosal neurons. Confocal images of (B1) submucosal ganglion (B2) Nlgn3 mRNA expression, B3: ChAT neuron, B4: merge. 3D structure of (B5) submucosal neurons; B6: Nlgn3 mRNA, B7: a cholinergic submucosal neuron (B8), merge. B9: The distribution of Nlgn3 mRNA expression in cholinergic neurons is similar to that in total neurons in the submucosal plexus. B10: Average number of Nlgn3 mRNA copies in cholinergic neuronal soma is similar to that in total neurons in the submucosal plexus. Confocal micrographs of: C1: submucosal neurons, C2: Nlgn3 mRNA expression, C3: VIP expression, C4: merge. 3D reconstruction of (C5) submucosal neurons, C6: Nlgn3 mRNA expression C7: VIP-expressing neurons; C8: merge. C9: Frequency distribution of Nlgn3 mRNA expression in VIP submucosal neurons is significantly different to total submucosal neurons. C10: On average, VIP submucosal neurons express similar numbers of Nlgn3 mRNA copies as observed in the total number of neurons in the submucosal plexus. Open arrowheads: Hu staining; arrows: Nlgn3 mRNA expression. Scale bar=10 µm.
Figure 1.
The distribution of Nlgn3 mRNA in ileal submucosal neurons. Confocal images of A1: submucosal neurons labelled with Hu pan neuronal marker; A2: Nlgn3 mRNA expression. A3: merged image. A4: 3D reconstruction of submucosal neurons; A5: cellular Nlgn3 mRNA expression; A6: merged image. A7: Frequency distribution of Nlgn3 mRNA expression in submucosal neurons. Confocal images of (B1) submucosal ganglion (B2) Nlgn3 mRNA expression, B3: ChAT neuron, B4: merge. 3D structure of (B5) submucosal neurons; B6: Nlgn3 mRNA, B7: a cholinergic submucosal neuron (B8), merge. B9: The distribution of Nlgn3 mRNA expression in cholinergic neurons is similar to that in total neurons in the submucosal plexus. B10: Average number of Nlgn3 mRNA copies in cholinergic neuronal soma is similar to that in total neurons in the submucosal plexus. Confocal micrographs of: C1: submucosal neurons, C2: Nlgn3 mRNA expression, C3: VIP expression, C4: merge. 3D reconstruction of (C5) submucosal neurons, C6: Nlgn3 mRNA expression C7: VIP-expressing neurons; C8: merge. C9: Frequency distribution of Nlgn3 mRNA expression in VIP submucosal neurons is significantly different to total submucosal neurons. C10: On average, VIP submucosal neurons express similar numbers of Nlgn3 mRNA copies as observed in the total number of neurons in the submucosal plexus. Open arrowheads: Hu staining; arrows: Nlgn3 mRNA expression. Scale bar=10 µm.
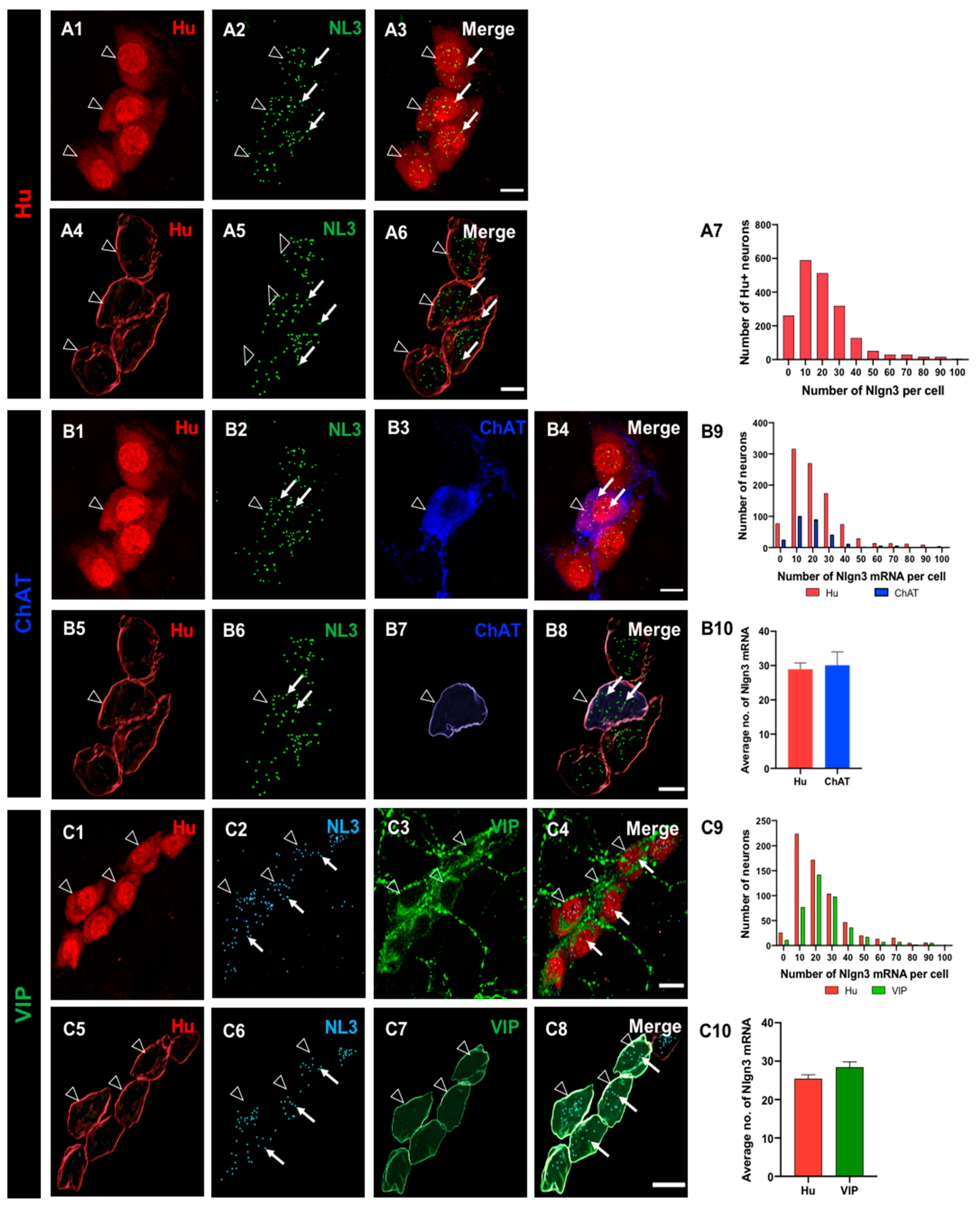
Figure 2.
Expression and the distribution of Nlgn3 mRNA in distal ileal myenteric neurons. A1: Myenteric neurons labelled using Hu immunofluorescence. A2: Nlgn3 mRNA expression (RNAScope); A3: merge. 3D structure of (A4) myenteric neurons, A5: Nlgn3 mRNA, A6: merge. A7: Frequency distribution of Nlgn3 expression in myenteric neurons. Confocal micrographs of (B1) myenteric neurons labelled with the pan-neuronal marker, Hu; B2: Nlgn3 mRNA, B3; calretinin. B4: Expression of Nlgn3 mRNA in calretinin neurons. 3D rendering of (B5) myenteric neurons, B6: Nlgn3 mRNA, B7: calretinin, B8: merge. B9: Frequency distribution of Nlgn3 mRNA expression in calretinin-positive myenteric neurons. B10: Calretinin-expressing neurons contain similar copy numbers of Nlgn3 mRNA to that of myenteric neurons overall. Triple labelling of (C1) myenteric neurons, C2: Nlgn3 mRNA and C3: nNOS expressing myenteric neurons. C4. Expression of Nlgn3 mRNA in nNOS containing myenteric neurons. 3D structure of (C5) myenteric neurons, C6: Nlgn3 mRNA, C7: NOS-expressing myenteric neurons, C8: merge. C9: Frequency distribution of Nlgn3 mRNA in nNOS containing neurons. C10: Average number of Nlgn3 mRNA copies in myenteric nNOS-containing compared to total myenteric neurons. Open arrowheads: Hu staining; arrows: Nlgn3 mRNA labelling. Scale bar=10 µm.
Figure 2.
Expression and the distribution of Nlgn3 mRNA in distal ileal myenteric neurons. A1: Myenteric neurons labelled using Hu immunofluorescence. A2: Nlgn3 mRNA expression (RNAScope); A3: merge. 3D structure of (A4) myenteric neurons, A5: Nlgn3 mRNA, A6: merge. A7: Frequency distribution of Nlgn3 expression in myenteric neurons. Confocal micrographs of (B1) myenteric neurons labelled with the pan-neuronal marker, Hu; B2: Nlgn3 mRNA, B3; calretinin. B4: Expression of Nlgn3 mRNA in calretinin neurons. 3D rendering of (B5) myenteric neurons, B6: Nlgn3 mRNA, B7: calretinin, B8: merge. B9: Frequency distribution of Nlgn3 mRNA expression in calretinin-positive myenteric neurons. B10: Calretinin-expressing neurons contain similar copy numbers of Nlgn3 mRNA to that of myenteric neurons overall. Triple labelling of (C1) myenteric neurons, C2: Nlgn3 mRNA and C3: nNOS expressing myenteric neurons. C4. Expression of Nlgn3 mRNA in nNOS containing myenteric neurons. 3D structure of (C5) myenteric neurons, C6: Nlgn3 mRNA, C7: NOS-expressing myenteric neurons, C8: merge. C9: Frequency distribution of Nlgn3 mRNA in nNOS containing neurons. C10: Average number of Nlgn3 mRNA copies in myenteric nNOS-containing compared to total myenteric neurons. Open arrowheads: Hu staining; arrows: Nlgn3 mRNA labelling. Scale bar=10 µm.
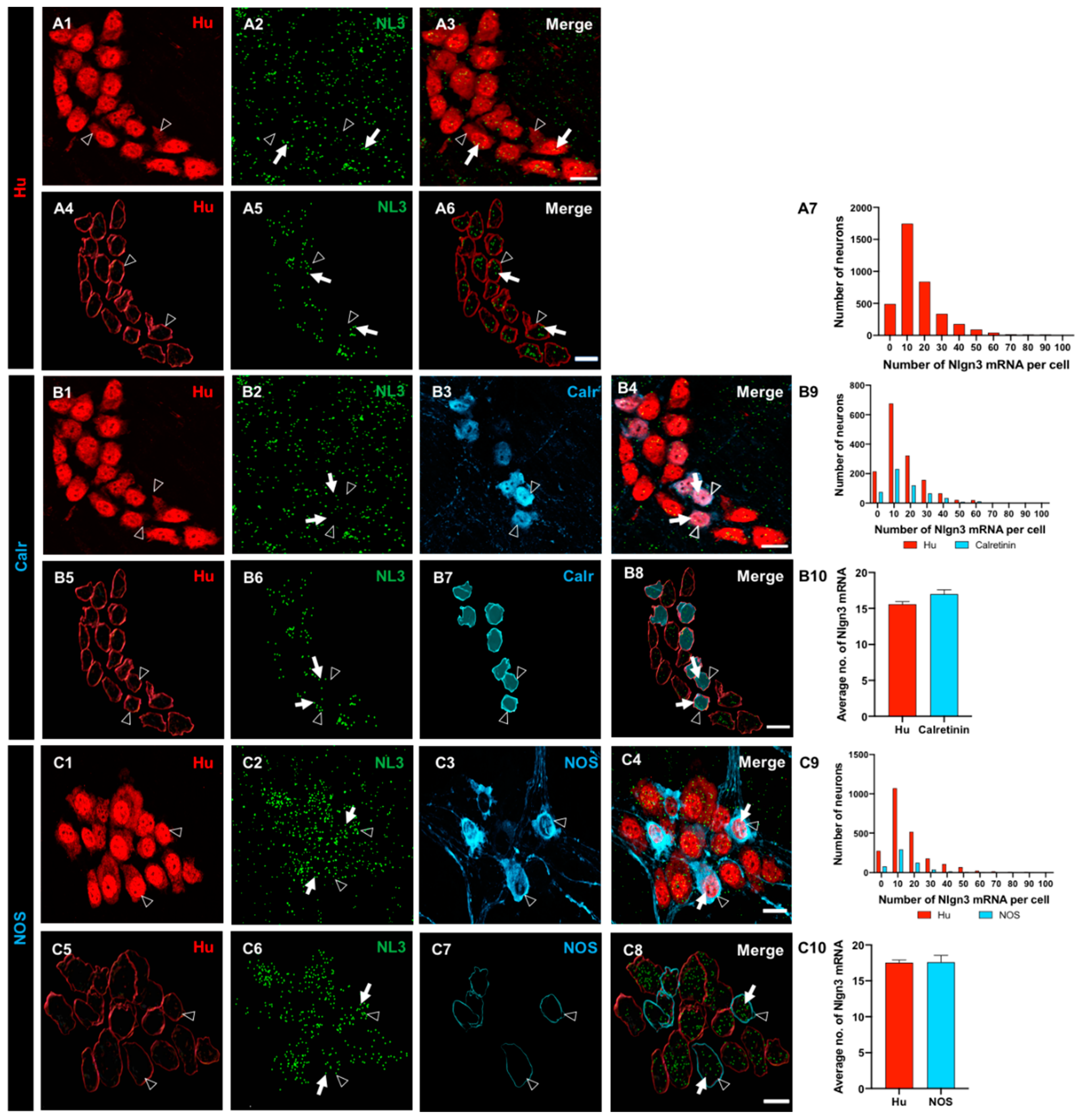
Figure 3.
The expression and distribution of Nlgn3 mRNA in enteric glia. Confocal micrographs of (A1) submucosal neurons labelled with the pan-neuronal marker Hu (A2) Nlgn3 mRNAexpression in the submucosal plexus (A3) submucosal glia labelled with S100β (A4) Nlgn3 mRNA in submucosal glia. Imaris-based 3D rendering of (A5) submucosal neurons; A6: Nlgn3 mRNA, A7: submucosal glia, A8: merge. A9: Frequency distribution analysis demonstrates that Nlgn3 mRNA is expressed in the majority of submucosal glial cells. Triple labelling of (B1) myenteric neurons using the pan-neuronal marker, Hu, B2: Nlgn3 mRNA, B3: myenteric glia (using S100β). B4: Expression of Nlgn3 mRNA in myenteric glia. 3D rendering of (B5) myenteric neurons, B6: Nlgn3 mRNA, B7: myenteric glia, B8: merge. B9: Frequency distribution analysis indicating that the vast majority of myenteric glia do not express Nlgn3 mRNA. Open arrowheads: Hu staining, filled arrows: Nlgn3 mRNA, open arrows: submucosal glia. Scale bar=10 µm.
Figure 3.
The expression and distribution of Nlgn3 mRNA in enteric glia. Confocal micrographs of (A1) submucosal neurons labelled with the pan-neuronal marker Hu (A2) Nlgn3 mRNAexpression in the submucosal plexus (A3) submucosal glia labelled with S100β (A4) Nlgn3 mRNA in submucosal glia. Imaris-based 3D rendering of (A5) submucosal neurons; A6: Nlgn3 mRNA, A7: submucosal glia, A8: merge. A9: Frequency distribution analysis demonstrates that Nlgn3 mRNA is expressed in the majority of submucosal glial cells. Triple labelling of (B1) myenteric neurons using the pan-neuronal marker, Hu, B2: Nlgn3 mRNA, B3: myenteric glia (using S100β). B4: Expression of Nlgn3 mRNA in myenteric glia. 3D rendering of (B5) myenteric neurons, B6: Nlgn3 mRNA, B7: myenteric glia, B8: merge. B9: Frequency distribution analysis indicating that the vast majority of myenteric glia do not express Nlgn3 mRNA. Open arrowheads: Hu staining, filled arrows: Nlgn3 mRNA, open arrows: submucosal glia. Scale bar=10 µm.
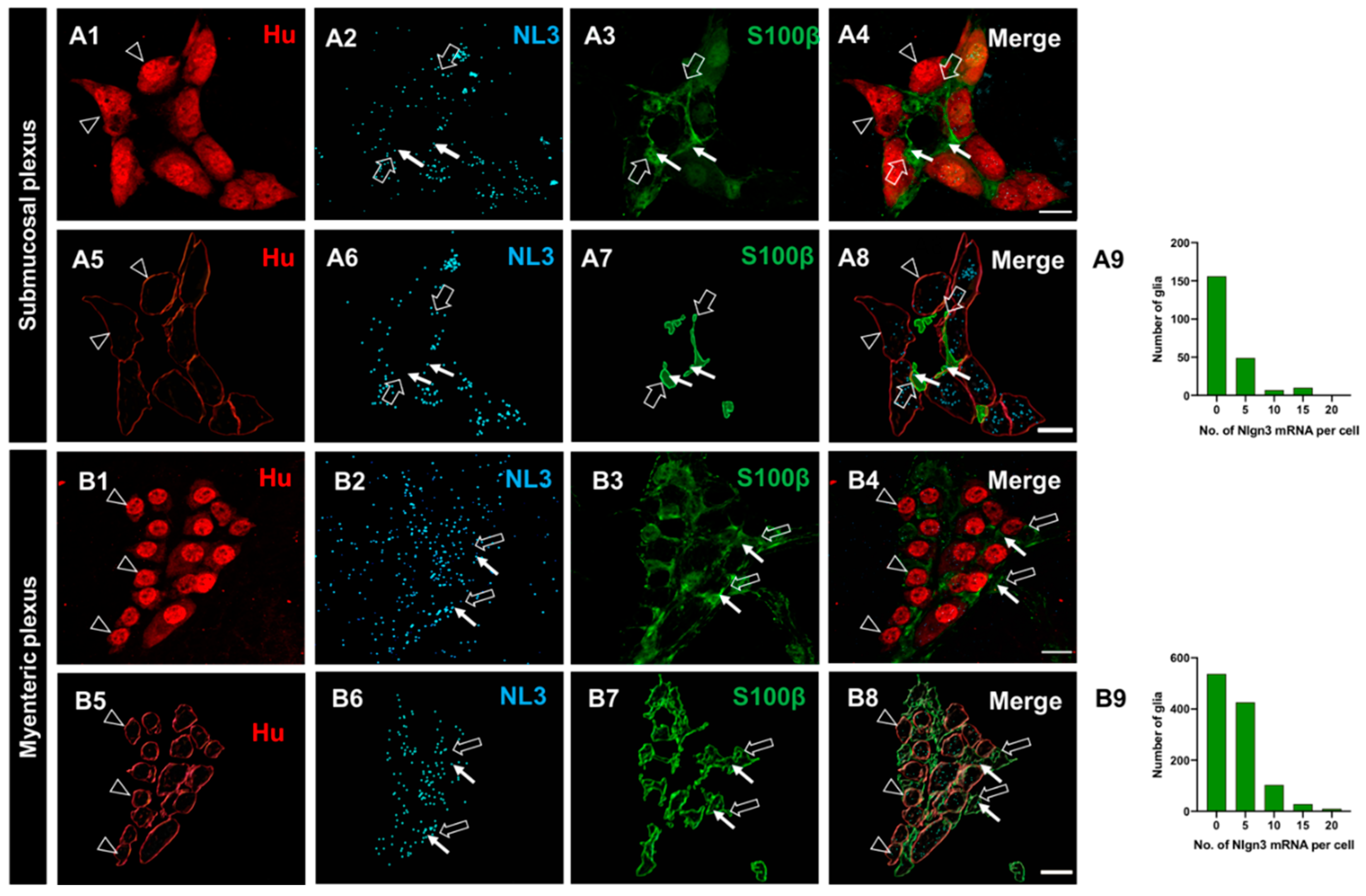
Figure 4.
Single cell RNA Sequencing data analysis reveal broad Nlgn3 expression in enteric neurons. A: UMAP of myenteric neuronal subtypes indicating expression of Nlgn3 in addition to Nlgn1 and Nlgn2 at varying expression levels. Colour bar indicates relative expression level with maximum cut-off at the 90th percentile. B: ENC1-12 displayed on UMAP and schematic indicating plausible functional annotations (modified from [17]). C: Dotblots indicating relative expression of Nrxn and Nlgn genes across ENCs (modified version of figure in [17]) UMAP: Uniform Manifold Approximation and Projection, IN: Interneuron; IPANS: Intrinsic primary afferent neurons. ENC: Enteric Neuron Class.
Figure 4.
Single cell RNA Sequencing data analysis reveal broad Nlgn3 expression in enteric neurons. A: UMAP of myenteric neuronal subtypes indicating expression of Nlgn3 in addition to Nlgn1 and Nlgn2 at varying expression levels. Colour bar indicates relative expression level with maximum cut-off at the 90th percentile. B: ENC1-12 displayed on UMAP and schematic indicating plausible functional annotations (modified from [17]). C: Dotblots indicating relative expression of Nrxn and Nlgn genes across ENCs (modified version of figure in [17]) UMAP: Uniform Manifold Approximation and Projection, IN: Interneuron; IPANS: Intrinsic primary afferent neurons. ENC: Enteric Neuron Class.
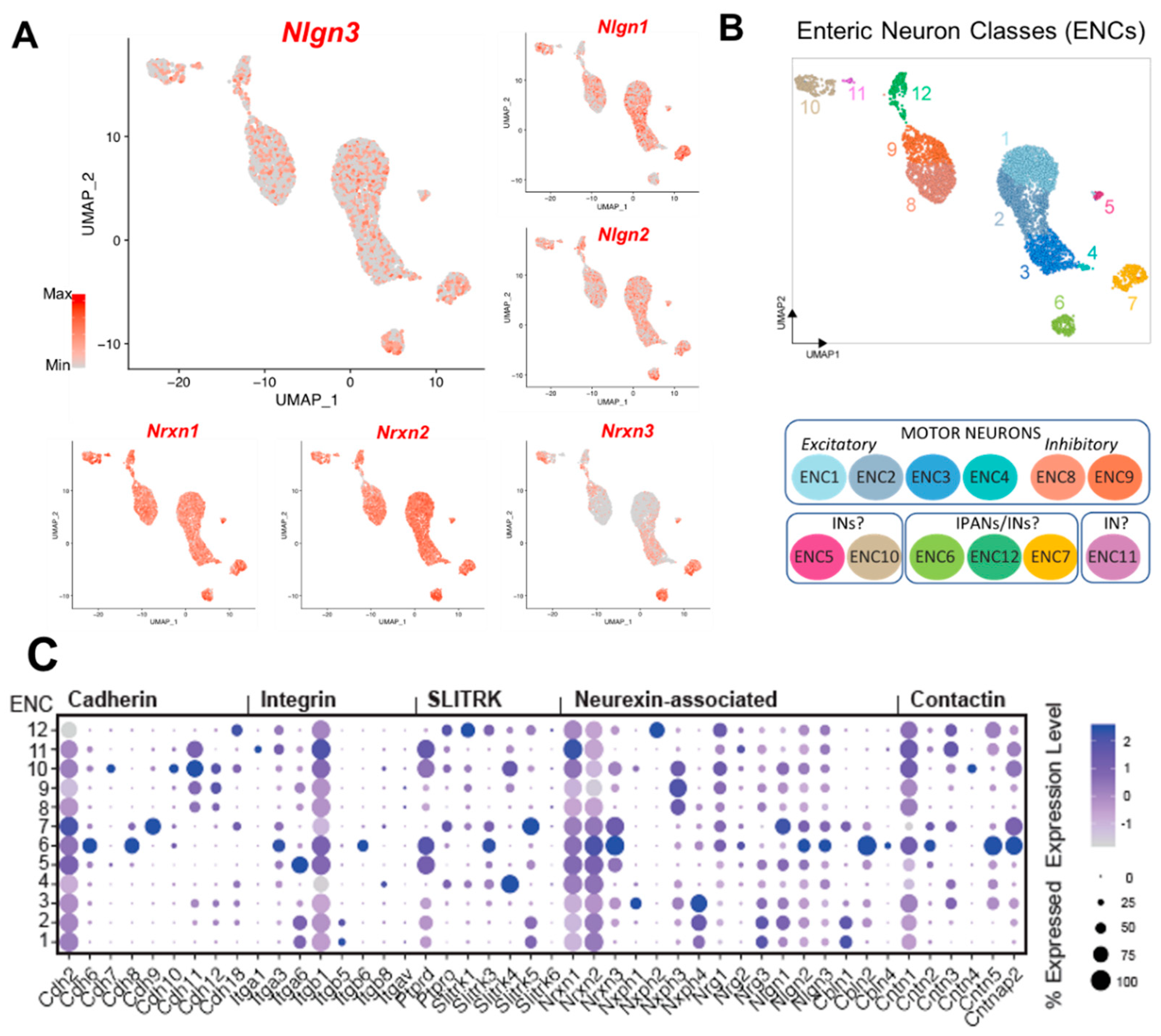
Figure 5.
Differential expression of Neuroligins and Neurexins in enteric glia. A: Nlgn1 is negligible expressed in EGCs B: Nlgn2 is expressed in a low proportion of all ENTGs. C: Nlgn3 is strongly expressed compared to Nlgn1 and Nlgn2 across all ENTGs. D: Representation of enteric glia clusters. E-G: Nrxn1, 2 and 3 are minimally expressed in enteric glia. Colour bar indicates relative gene expression. ENTG: enteric glia cluster; ENMFB: enteric mesothelial fibroblasts. Pseudocolored scale bar refers to maximum and minimum expression levels within ENTGs and ENMFB.
Figure 5.
Differential expression of Neuroligins and Neurexins in enteric glia. A: Nlgn1 is negligible expressed in EGCs B: Nlgn2 is expressed in a low proportion of all ENTGs. C: Nlgn3 is strongly expressed compared to Nlgn1 and Nlgn2 across all ENTGs. D: Representation of enteric glia clusters. E-G: Nrxn1, 2 and 3 are minimally expressed in enteric glia. Colour bar indicates relative gene expression. ENTG: enteric glia cluster; ENMFB: enteric mesothelial fibroblasts. Pseudocolored scale bar refers to maximum and minimum expression levels within ENTGs and ENMFB.
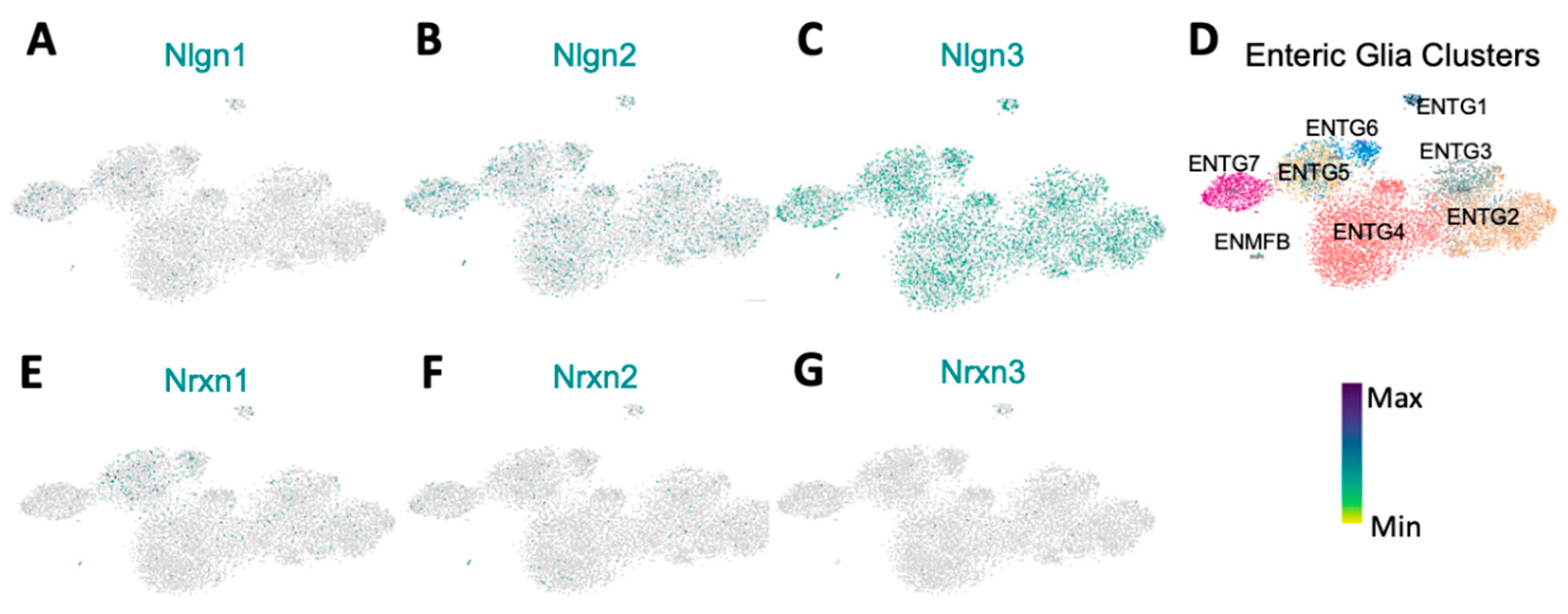
Figure 6.
Expression of Nlgn3 mRNA in the ileal submucosal plexus of Nlgn3R451C mutant mice. Confocal micrographs of submucosal neurons labeled with the pan-neuronal marker Hu (A1), Nlgn3 mRNA expression (A2); merge (A3). The same submucosal neurons shown following 3D reconstruction (A4), corresponding cellular Nlgn3 mRNA expression (A5); merge (A6). A7: The distribution of Nlgn3 mRNA in WT is significantly different compared to Nlgn3R451C mutant mice. A8: Nlgn3R451C mutant mice express fewer copies of Nlgn3 mRNA in submucosal neurons compared to WT. Submucosal neurons labelled with Hu pan-neuronal marker (B1) Nlgn3 mRNA (B2) and the cholinergic marker, ChAT (B3); merge (B4). 3D rendering of (B5) Hu neurons (B6) and corresponding Nlgn3 mRNA expression (B7) and ChAT labelling (B8). B9: Frequency distribution of Nlgn3 mRNA in cholinergic submucosal neurons. B10: Cholinergic neurons express significantly fewer copies of Nlgn3 mRNA in Nlgn3R451C mutants compared to WT. Confocal micrographs of submucosal neurons labelled with C1: the pan-neuronal marker, Hu; C2: Nlgn3 mRNA labelled using RNAScope and C3: non-cholinergic neurons labelled with VIP; C4: merge. 3D reconstruction of C5: submucosal neurons, C6: corresponding Nlgn3 mRNA puncta, C7: VIPergic neurons, C8; merge. C9: Frequency distribution of Nlgn3 mRNA in non-cholinergic submucosal neurons C10: Nlgn3R451C mice express similar numbers of Nlgn3 mRNA copies in non-cholinergic neurons compared to WT in the submucosal plexus. Nlgn3 mRNA is indicated by the filled arrow and submucosal glia are labelled with open arrows **p<0.01, ****p<0.0001, Scale bar=10 µm.
Figure 6.
Expression of Nlgn3 mRNA in the ileal submucosal plexus of Nlgn3R451C mutant mice. Confocal micrographs of submucosal neurons labeled with the pan-neuronal marker Hu (A1), Nlgn3 mRNA expression (A2); merge (A3). The same submucosal neurons shown following 3D reconstruction (A4), corresponding cellular Nlgn3 mRNA expression (A5); merge (A6). A7: The distribution of Nlgn3 mRNA in WT is significantly different compared to Nlgn3R451C mutant mice. A8: Nlgn3R451C mutant mice express fewer copies of Nlgn3 mRNA in submucosal neurons compared to WT. Submucosal neurons labelled with Hu pan-neuronal marker (B1) Nlgn3 mRNA (B2) and the cholinergic marker, ChAT (B3); merge (B4). 3D rendering of (B5) Hu neurons (B6) and corresponding Nlgn3 mRNA expression (B7) and ChAT labelling (B8). B9: Frequency distribution of Nlgn3 mRNA in cholinergic submucosal neurons. B10: Cholinergic neurons express significantly fewer copies of Nlgn3 mRNA in Nlgn3R451C mutants compared to WT. Confocal micrographs of submucosal neurons labelled with C1: the pan-neuronal marker, Hu; C2: Nlgn3 mRNA labelled using RNAScope and C3: non-cholinergic neurons labelled with VIP; C4: merge. 3D reconstruction of C5: submucosal neurons, C6: corresponding Nlgn3 mRNA puncta, C7: VIPergic neurons, C8; merge. C9: Frequency distribution of Nlgn3 mRNA in non-cholinergic submucosal neurons C10: Nlgn3R451C mice express similar numbers of Nlgn3 mRNA copies in non-cholinergic neurons compared to WT in the submucosal plexus. Nlgn3 mRNA is indicated by the filled arrow and submucosal glia are labelled with open arrows **p<0.01, ****p<0.0001, Scale bar=10 µm.
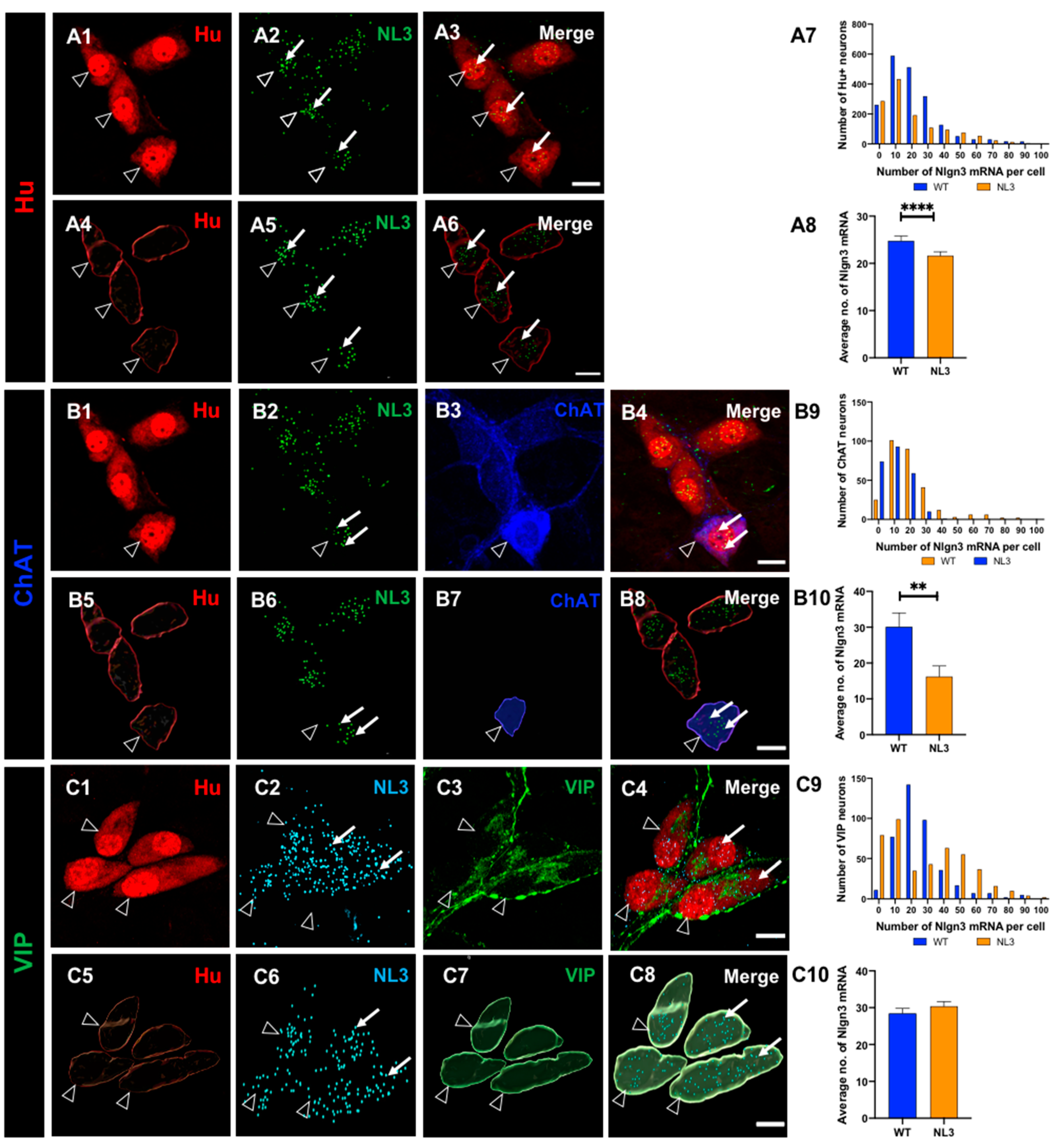
Figure 7.
Effects of the Nlgn3 R451C mutation on Nlgn3 mRNA expression in myenteric neurons. Confocal images of (A1) myenteric neurons (A2) Nlgn3 mRNA expression (A3) Nlgn3 mRNA expression in myenteric neurons. The 3D structure of (A4) myenteric neurons (A5) Nlgn3 mRNA (A6) Nlgn3 mRNA expression in myenteric neurons. A7. The frequency distribution of Nlgn3 mRNA in Nlgn3R451C mutant mice compared WT A8. In the Nlgn3R451C mouse ileum, myenteric neurons express fewer copies of Nlgn3 mRNA compared to WT. Triple labelling of (B1) myenteric neurons using B2: the pan-neuronal marker Hu, B3: Nlgn3 mRNA, B3: calretinin; B4: merge. 3D reconstruction of (B5) myenteric neurons, B6: Nlgn3 mRNA, B7: calretinin expressing neurons and B8: merge. B9. The frequency distribution of Nlgn3 mRNA in calretinin positive myenteric neurons in Nlgn3R451C mutant mice and WT. B10: The Nlgn3 R451C mutation reduces Nlgn3 mRNA expression in mutant mice compared to WT. Triple labelling of (C1) myenteric neurons, C2: Nlgn3 mRNA, C3: nNOS expressing neurons, C4: merge. 3D rendering of (C5) myenteric neurons; C6: nNOS positive neurons, C7: Nlgn3 mRNA and C8: merge. C9: Frequency distribution of Nlgn3 mRNA expression in NOS-positive neurons in Nlgn3R451C mutant mice. C10: In Nlgn3R451C mutant mice, NOS neurons contain fewer Nlgn3 mRNA copies compared to WT. Filled arrows: Nlgn3 mRNA. Open arrows: submucosal glia. ****p<0.0001, Scale bar=10 µm.
Figure 7.
Effects of the Nlgn3 R451C mutation on Nlgn3 mRNA expression in myenteric neurons. Confocal images of (A1) myenteric neurons (A2) Nlgn3 mRNA expression (A3) Nlgn3 mRNA expression in myenteric neurons. The 3D structure of (A4) myenteric neurons (A5) Nlgn3 mRNA (A6) Nlgn3 mRNA expression in myenteric neurons. A7. The frequency distribution of Nlgn3 mRNA in Nlgn3R451C mutant mice compared WT A8. In the Nlgn3R451C mouse ileum, myenteric neurons express fewer copies of Nlgn3 mRNA compared to WT. Triple labelling of (B1) myenteric neurons using B2: the pan-neuronal marker Hu, B3: Nlgn3 mRNA, B3: calretinin; B4: merge. 3D reconstruction of (B5) myenteric neurons, B6: Nlgn3 mRNA, B7: calretinin expressing neurons and B8: merge. B9. The frequency distribution of Nlgn3 mRNA in calretinin positive myenteric neurons in Nlgn3R451C mutant mice and WT. B10: The Nlgn3 R451C mutation reduces Nlgn3 mRNA expression in mutant mice compared to WT. Triple labelling of (C1) myenteric neurons, C2: Nlgn3 mRNA, C3: nNOS expressing neurons, C4: merge. 3D rendering of (C5) myenteric neurons; C6: nNOS positive neurons, C7: Nlgn3 mRNA and C8: merge. C9: Frequency distribution of Nlgn3 mRNA expression in NOS-positive neurons in Nlgn3R451C mutant mice. C10: In Nlgn3R451C mutant mice, NOS neurons contain fewer Nlgn3 mRNA copies compared to WT. Filled arrows: Nlgn3 mRNA. Open arrows: submucosal glia. ****p<0.0001, Scale bar=10 µm.
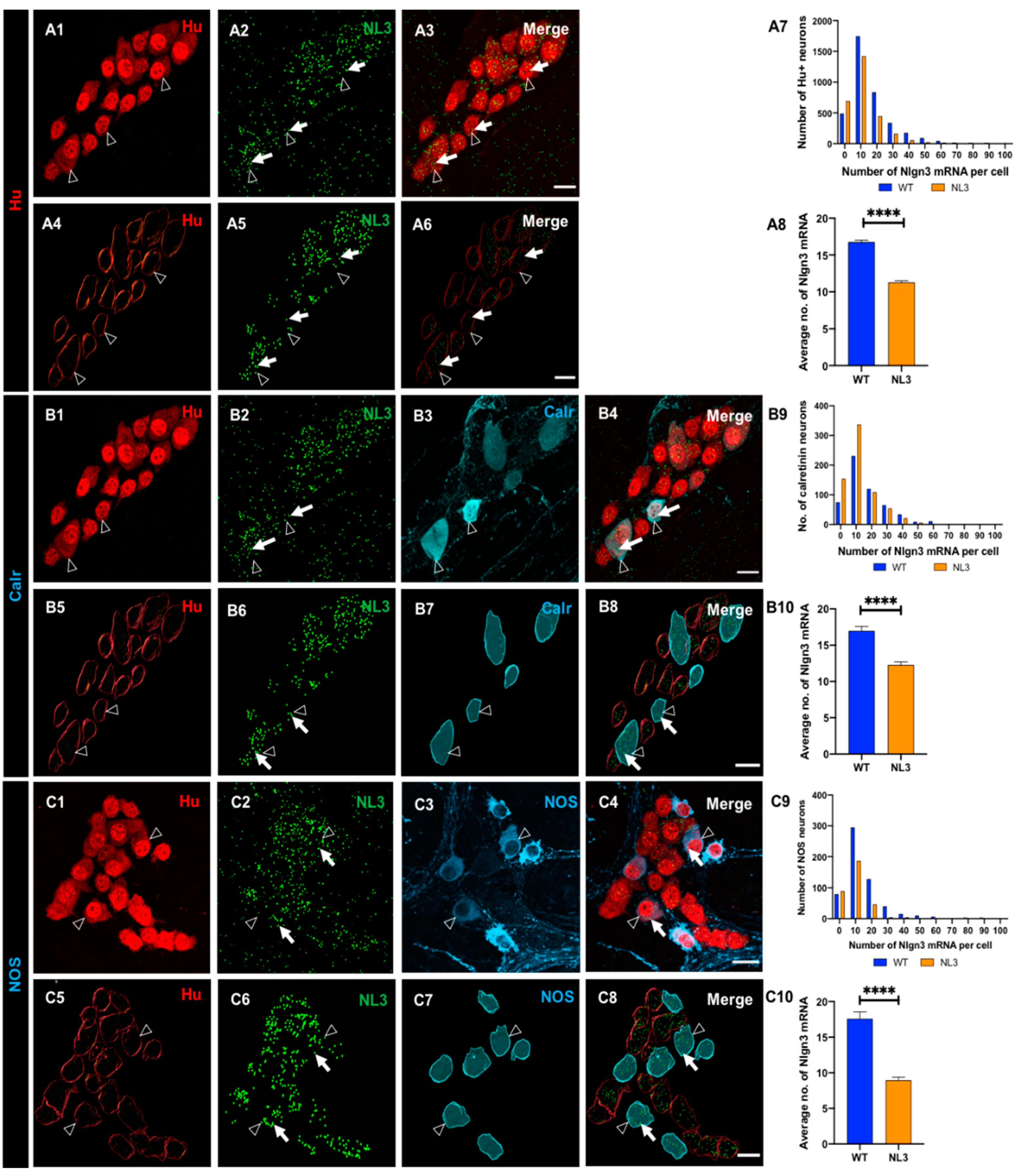
Figure 8.
Effects of Nlgn3 R451C mutation on Nlgn3 mRNA expression in enteric glia. Confocal images of (A1) submucosal neurons labelled with A2: Hu, A3: Nlgn3 mRNA, A4: merge. 3D structure of (A5) submucosal neurons; A6: Nlgn3 mRNA, A7: submucosal glia, A8: merge. A9: Frequency distribution of Nlgn3 mRNA in submucosal glia in Nlgn3R451C mutant mice compared to WT. A10: Nlgn3 mRNA copy numbers per cell in WT compared to Nlgn3R451C mutant mice. Immunofluorescent labelling of (B1) myenteric neurons (Hu), B2: Nlgn3 mRNA, B3: myenteric glia (S100β), B4: merge. 3D reconstruction of (B5) myenteric neurons; B6: Nlgn3 mRNA, B7: myenteric glia, B8: merge. B9: Frequency distribution of Nlgn3 mRNA expression in WT and Nlgn3R451C mutant mice. B10: Mutant mice express significantly fewer Nlgn3 mRNA copies in myenteric glia compared to WT. ****p<0.0001, Scale bar=10 µm.
Figure 8.
Effects of Nlgn3 R451C mutation on Nlgn3 mRNA expression in enteric glia. Confocal images of (A1) submucosal neurons labelled with A2: Hu, A3: Nlgn3 mRNA, A4: merge. 3D structure of (A5) submucosal neurons; A6: Nlgn3 mRNA, A7: submucosal glia, A8: merge. A9: Frequency distribution of Nlgn3 mRNA in submucosal glia in Nlgn3R451C mutant mice compared to WT. A10: Nlgn3 mRNA copy numbers per cell in WT compared to Nlgn3R451C mutant mice. Immunofluorescent labelling of (B1) myenteric neurons (Hu), B2: Nlgn3 mRNA, B3: myenteric glia (S100β), B4: merge. 3D reconstruction of (B5) myenteric neurons; B6: Nlgn3 mRNA, B7: myenteric glia, B8: merge. B9: Frequency distribution of Nlgn3 mRNA expression in WT and Nlgn3R451C mutant mice. B10: Mutant mice express significantly fewer Nlgn3 mRNA copies in myenteric glia compared to WT. ****p<0.0001, Scale bar=10 µm.
Table 1.
Primary antibodies used for immunocytochemistry.
| Primary antisera | Raised in | Dilution | Source |
|---|---|---|---|
| ANNA-1(anti HuC/D)* | Human | 1:5000 | Gift from Dr. V. Lennon |
| VIP | Rabbit | 1:1000 | Merck Millipore |
| ChAT | Goat | 1:100 | Chemicon |
| nNOS | Sheep | 1:1000 | Gift from Dr P. Emson |
| Calretinin | Goat | 1:1000 | SWANT |
| S100β | Rabbit | 1:1000 | DAKO |
* Referred to as Hu throughout the text.
Table 2.
Secondary antibodies used for immunocytochemistry.
| Secondary antisera | Raised in | Dilution | Source |
|---|---|---|---|
| Anti-human AF 594 | Donkey | 1:500 | Jackson Immuno Labs |
| Anti-sheep AF 647 | Donkey | 1:500 | Molecular Probes |
| Anti-rabbit AF 647 | Donkey | 1:400 | Molecular Probes |
| Anti-sheep AF 594 | Donkey | 1:100 | Molecular Probes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



