
Submitted:
19 June 2023
Posted:
20 June 2023
You are already at the latest version
Abstract
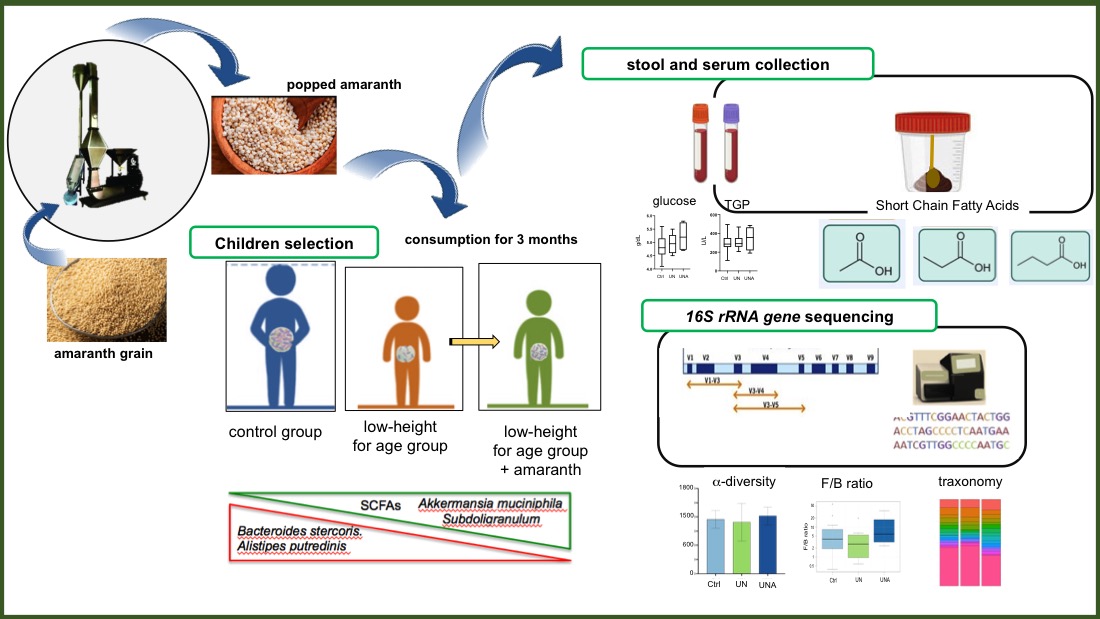
Popped amaranth is a nutritious food consumed since pre-Hispanic times, currently its consumption has been associated with recovery of malnourished children. However, there is no information on the impact that popped amaranth consumption has on gut microbiota composition. A non-randomized pilot trial was conducted to evaluate the changes on composition, structure, and function of gut microbiota of stunted children who received four grams of popped amaranth daily for three months. Stool and serum were collected at the beginning and at the end of the trial. Short-chain-fatty acids were quantified and gut bacterial composition was analyzed by 16S rRNA gene sequencing. Biome-try and hematology results showed that children have no other pathology more than to be low-height-for age. Increase of total SCFAs levels in feces was ob-served as well as a decrease in relative abundance of Alistipes putredinis, Bac-teroides coprocola, and Bacteroides stercoris, bacteria related with inflammation and colitis. On the contrary, an increase in relative abundance of Akkermansia muciniphila and Streptococcus thermophiles, bacteria associated with health and longevity, was observed. Results have proven that popped amaranth, a nutri-tious food, help to combat children malnutrition through gut microbiota modu-lation.
Keywords:
16S rRNA sequencing
; diet
; popped amaranth
; gut microbiota
; short-chain fatty acids
1. Introduction
Malnutrition is a term that encompasses many different manifestations of inadequate nutrition, including both undernutrition and obesity, which are characterized by an imbalance in energy intake and energy expenditure [1]. Undernutrition, defined as a deficiency of calories or a shortage of one or more essential nutrients, is a significant pediatric health problem pressing and overwhelming global health issue contributing to nearly half of all deaths in children under five years of age [2], where mostly occur in low- and middle-income countries where at the same time rates of childhood overweight and obesity are rising [3]. Child´s health has been based on measurements of the body mass index (weight [kg]/height2 [m2]), but to recognize clinical signs of certain undernutrition problems, weight length, height, or Z-score values have been used to classify the nutritional status of children. In this sense, undernutrition is classified as stunting when measured at a low height-for-age <-2 standard deviations (SD); wasting, when is low weight-for-age (<-2 SD), and underweight, for low weight-for-height (<-2 SD). Malnutrition is also classified as moderate when it falls between -2 and -3 SD and severe it when falls <-3 SD [3]. According to the Z-score, 165 million children under five years of age are stunted and 50 million children are wasting [4]. Stunted is caused by some factors such as poor nutrition, poor sanitation and no access to clean drinking water, and inadequate psychosocial stimulation. Stunted is a key risk factor for diminished survival, poor child and adult health, decreased learning capacity, and lost future productivity [5].
Persistent childhood malnutrition is considered part of a vicious cycle of recurrent infections, impaired immunity, and worsening malnutrition [6]. All these factors have been linked to gut microbiota dysbiosis, which can affect the immune response, the susceptibility to infection, and the nutritional status, resulting in the main consequences of undernutrition [1,6,7]. Disruption of the normal gut microbiota altered gut barrier function, impaired mucosal immunity, and increased the risk of gram-negative bacteremia [7]; furthermore, diarrhea is considered as a main contributor to malnutrition, which is induced by pathogenic bacteria of gut microbiota [5].
Growing evidences have shown that dietary patterns and the adequate food processing conditions, are factors that affect the food digestibility, functionality properties, and influence the composition, structure, and function of gut microbiota, and therefore, human health [8,9]. It is also known that the quality and quantity of protein intake affects the metabolites produced by the gut microbiota [9,1011,12]. Carbohydrates (resistant starch and dietary fiber) that escape digestion and absorption in the small intestine are also used by microbiota and through saccharolytic fermentation leads to the generation of short-chain fatty acids (SCFAs). SCFAs, mainly acetate, propionate, and butyrate, are primary end products of fermentation and represent the major flow of carbon from the diet, through the microbiome to the host. SCFAs have a positive influence on gut integrity and nutritional health by improving energy yield, modulation of colonic pH, production of vitamins, and stimulation of gut homeostasis, including anti-pathogenic activities [7].
Amaranth has been recognized as nutraceutical food because it contains higher amounts of proteins (compared with traditional cereals such as corn, wheat, and rice), but most important because of its high nutritive value due to its adequate amino acid composition, which covers the requirements recommended by children and adults [13]. Amaranth grains contains also several encrypted peptides with antidiabetic, antihypertensive, and antioxidant functions [14], they are rich in lipids containing several sterols, including tocopherols, also they are rich source of squalene, a key metabolite in sterol pathway. In relation to micronutrients, amaranth grains are a good source of minerals, including phosphorus, potassium, magnesium, calcium iron, zinc, manganese, and selenium, and also are rich in vitamins (B2, B6, and E), niacin, and thiamine [13]. Popped amaranth grain has been consumed since pre-Hispanic times. Popping is achieved by applying a heat treatment to the amaranth grains resulting in a pre-cooked food with a nutty flavor [15]. Currently, popped amaranth has been proposed as minimally processed healthy food snack due to its high quality and quantity of proteins [16]. As other plant-based food products, heat treatment processing does not altered amaranth grain properties but popping increases the digestibility of amaranth grain nutrients [17], enhances its antioxidative properties [18,19], and reduces adverse antinutritional compounds such as tannins, lectins, and trypsin inhibitors [17]. Consumption of popped amaranth grain has been associated with health benefits in humans, including recovery of severely malnourished children [17,20,21]. Despite the vast evidence that supports the beneficial effects on health of amaranth consumption, to date, there are no studies that analyze the effects of popped amaranth has on gut microbiota. Therefore, the present study aimed to carry out a nonrandomized pilot trial design to explore the effect of popped amaranth consumption on the changes in the structure and abundance of gut microbiota of children classified as low height-for-age (stunted children). Results have shown that popped amaranth consumption help to combat children malnutrition through gut microbiota modulation.
2. Materials and Methods
2.1. Recruitment of participants
Children living in San Antonio Huichimal, Lima, and Vista Hermosa rural area of Tenek, Ciudad Valles, San Luis Potosi, S.L.P., Mexico, were recruited for this research through screening based on inclusion and exclusion criteria. Sick children or those children with some reported diseases were excluded. The selected children had taken no antibiotics in the three months before to the study. Body weight was measured using digital weight scale with the infant wearing a light cloth and no shoes (accuracy: 0.1 kg). Meanwhile, body height was measured using a 2-m-long microtoise without shoes (accuracy: 0.1 cm). Children were eligible for inclusion if they were aged between 6 and 7 years old and they were grouped into two groups: control group (Ctrl) those that presented normal height-for-age with a mean of HAZ=-0.03 ±0.5 (25 children), and the stunted or low height-for-age (HAZ <-2 SD) group (9 children).
2.2. Research design
The study was approved by the by the ethics committees of Health Services of San Luis Potosi (approval reference: SLP/006-2018), and DIF-Ciudad Valles, San Luis Potosí. This research was conducted following the applicable regulations and guidelines in accordance with the Helsinki Declaration, revised in 2000. Informed consent was signed by children participant, as well as by children’s parents/legal guardians.
Stunted children consumed four grams of popped amaranth daily for three months. Serum samples from participant children were obtained before and after the trial for blood chemistry and liver function. Stool samples were collected before and after the trial for the gut microbiota composition and SCFAs analyses. The research design is shown in Figure 1.
2.3. Preparation of amaranth popped grain
Popped seeds were obtained by heating the seeds in an industrial hot air fluidized bed machine popper (AmarantaR, San Miguel de proyectos Agropecuarios, Hidalgo, México). Popped amaranth complies with the Official Mexican Standards (NOM-051-SCFI/SSA1-2010) for food and drinks for human consumption. The protein, fat, fiber, and ash contents were determined by standard methods [22]. Total carbohydrate was calculated by subtracting protein, fat, ash and fiber from 100 [23].
2.4. Research outcome
During the intervention period, the daily popped amaranth intake was supervised. Sera and stool sample were collected before and after the study. Sera were used for biochemical profile and stool was used for the gut microbiota and SCFA profile analyses. The primary outcomes were gut microbiota composition and SCFAs profile, while the secondary outcomes was the hematic biochemical profile.
2.5. Serum collection and biochemical profile analysis
Blood samples (10 mL) were collected in two BD Vacutainer® Venous Blood Collection systems without anticoagulant. Samples were centrifuged at 400 x g for 10 min; the serum was separated and frozen at −70 °C until analysis. Serum concentrations of glucose, triglycerides, total cholesterol, urea, acid uric, creatinine, total protein, albumin, aspartate aminotransferase (AST), alanine aminotransferase (ALT), gamma-glutamyl transferase (GGT), alkaline phosphatase (ALKP), and lactate dehydrogenase (LDH) were determined by spectrophotometry using commercial kits (SPINREACT, Girona, Spain) and read on MultiskanGO microplate spectrophotometer (ThermoFischer Scientific, Waltham, Massachusetts, USA).
2.6. Stool sample collection
Stool samples were obtained immediately after defecation and collected aseptically in a sterile stool container. Stool samples were taken at the beginning of the trial and after three months of diet supplementation with amaranth. Samples were transported to the laboratory using ice packs, aliquoted, and immediately stored at −70 °C until further processing.
2.6.1. Measurement of fecal Short-Chain Fatty Acids (SCFAs)
Samples (250 mg) were homogenized in one mL of H2SO4 (0.5 mmol/L) and mixed at 1400 rpm for 3 min (Thermomixer, Eppendorf, Hamburg, Germany). The homogenized sample was incubated for 20 min in an ice-water bath and centrifuged at 4 °C and 4800 × g for 15 min. The supernatant was recovered and subsequently centrifuged at 4 °C and 13000 × g for 15 min. This procedure was repeated two times for clarification. The sample was filtered through a 0.22 μm Millipore filter (Merck, Darmstadt, Germany) before injection into the chromatographic system. SCFAs were analyzed using an Agilent 1100 of (Hewlett Packard, Santa Clara, CA, USA) and a Rezex ROA LC Column 150 x 7.8 mm (Phenomenex Inc, Torrance, CA, USA). The mobile phase was composed of H2SO4 (0.5 mmol/L). The column temperature was 60 ºC; the flow rate was 0.5 mL/min; and measurement was using a RID-10A RI detector. Calibration curves were performed from 0.18 to 1.8 mg/mL for acetic acid, 0.08 to 0.8 g/L for propionic acid and 0.11 to 1.1 g/L for butyric acid.
2.6.2. DNA extraction and integrity verification
Stool samples (250 µg) were diluted in 1300 µL of saline solution (0.85%), removing coarse remains. 600 µL of the decanted sample was taken and resuspended in a new tube. The suspension was centrifuged at 10,000xg for 10 min, at 4 °C. Obtained pellet was resuspended in 1 mL of cold PBS and centrifuged at 700 x g, 4 °C for 1 min. The supernatant was collected and centrifuged at 9000 x g for 5 min at 4 °C. The resulting pellet was used for DNA extraction following the specifications of the DNeasy UltraClean Microbial Kit, from QIAGEN (Hilden, Germany). DNA was quantified in NanoDrop One (ThermoFischer Scientific, Waltham, MA, USA), and integrity was verified by visualization on a 1% agarose gel, in Tris-Borate-EDTA (TBE) buffer, for 60 min at 70 volts. DNA extracts were stored at −80 °C until sequenced.
2.7. 16S rRNA gene sequencing and bioinformatics analyses
For microbiome analysis, the V4–V5 region of the 16S ribosomal RNA gene, was amplified once using universal bacterial primers 515FB: 5′-GTGYCAGCMGCCGCGGTAA-3′ and 926R: 5′-CCGYCAATTYMTTTRAGTTT-3′. 51 Samples were sequenced on the Illumina MiSeq platform (Illumina, San Diego, CA, United States) using 300 + 300 bp paired-end according to the protocol described elsewhere [24]. Samples were sequenced at the Integrated Microbiome Resource (IMR) (Dalhousie University, Halifax, NS, Canada).
2.7.1. Amplicon Sequence Variant inference
The R package DADA2 v1.16.0 [25] was used to process the 16S sequencing data. We have trimmed 20 nt to the right side of the forward fragments and 60 nt to the right side of the reverse fragments using the filterAndTrim () function with default parameters. We used the learnErrors() function with default parameters to learn the error rates. We have applied the dada() function to infer the sample composition using default parameters, the filtered fragments, and the calculated error rates. We have merged the fragments with the mergePairs() function and removed the chimeras using the removeBimeraDenovo() function with default parameters. The taxonomy for each sequence was assigned using the assignTaxonomy() function and the silva_nr99_v138 database. The Amplicon Sequence Variant (ASV) quantification and their phylogenetic assignment were obtained.
2.7.2. Diversity quantification and functional prediction
Alpha diversity, Shannon, inverse of Simpson, Fisher, Chao1, and ACE indices were calculated with the diversity() from the R package vegan v2.5.6 [26]. To determine the structural variation of microbial communities, beta diversity and NMD plots were generated with custom R scripts and using the BrayCurtis dissimilarity calculated with the ordinate() function from the vegan package.
The functional prediction of the 16S sequencing data was performed with Tax4Fun2 [27]. RunRefBlast() and makeFunctionalPrediction() functions were used with default parameters, to predict functional profiles of the ASV quantification results. The reference used in both steps runRefBlast and makeFunctionalPrediction was RF99NR. The pathway predictions table (pathway scores per library) was used to perform a sparse Partial Least Squares Discriminant Analysis (sPLS-DA). The function spls-da() from the R package mixOmics with default parameters and ncomp=2 was used [28]. The plots to represent the results of the Discriminative Analysis (DA) were generated with custom R scripts.
Using significant taxa and routes, taxa-pathways networks were created (permutation test spls-da, 1,000 permutations, p-val 0.1). The Spearman correlation between two nodes (taxa or pathways) was used to identify their connection. Using the igraph R package [29]. Only correlations greater than the 30th-percentile were employed to create the networks.
2.8. Statistical analyses
Statistical analysis of results was performed on Prism 8.0 for Mac (GraphPad Software, San Diego, CA, USA). The normal distribution was tested by D’Agostino-Pearson and Shapiro-Wilk tests. One-way ANOVA was performed for data with normal distribution, followed by Bonferroni’s post-hoc test. Kruskall-Wallis, followed by Dunn´s post-hoc test was carried out if data were not normally distributed. A p-value less than 0.05 (p < 0.05) were considered as statistically significant.
3. Results and Discussion
3.1. Popped amaranth as minimally processed food
Popping is a traditional process that has been carried out in Mexico since pre-Hispanic times. At that time popping was obtained by heating the amaranth grains for 15-25 s in covered clay pots at 175 to 195 oC, which resulted in grain expansion [30,31]. Currently, popped amaranth is obtained in industrial hot air fluidized bed machine, at 300 to 330 oC with 9 s of residence, hence that popped amaranth could be considered as a minimally processed food. Under these conditions, popped amaranth showed high protein content (15.8%), fat (6.7%), fiber (3.8%), and ashes (2.2%) (Supplementary Table S1).
3.2. Children´s selection and serum biochemical analysis
Children showing undernutrition and classified as stunted based on the height-for-age age parameters [3] were selected and named as undernutrition group (UN, n= 9). UN group consumed daily four grams of popped amaranth and were namend UNA group), but during the trial and COVID-19 pandemic situation, two children abandoned the study (UNA, n=7). A control group (Ctrl, n=21) represented by children with normal height-for-age living in the same conditions as UN group were was also selected.
The hematic cytometry of participating children are shown in Supplementary Table S2. Baseline values for participants reflected that they do not have any other pathology more than low height-for-age in the UN group. However, although all parameters fall into the biological limits, it was observed that hematocrit and mean globular volume (MCV) values had a tendency to increase in UNA children, even compared with control group. Low values of MCV are related to protein-energy malnutrition or deficiency to iron and folate [32]. Amaranth is a rich source of protein and also contains high amounts of minerals such as iron, calcium, magnesium [13], this could be the reason for the observed trend to increase in those values.
The serum biochemical profile neither shows any pathology. The renal function, tested by creatinine, uric acid, and urea, was normal in all participants. The biomarkers for liver function (ALT, TGO) showed normal levels. No significant changes were observed in cholesterol, triglycerides, and high-density lipoprotein (HDL) amongst groups (Supplementary Table S3).
3.3. Modulation of gut microbiota in undernutrition children group after amaranth consumption
The sequencing of the 16S ribosomal ARN gene (16S rRNA) has generated great interest in studying bacterial communities. Amplicon Sequencing Variants (ASVs) have been proposed as an alternative to Operational Taxonomic Units (OTUs) to analyze microbial communities. The use of ASVs has grown in popularity in part because they reflect a more refined level of taxonomy in that they do not group sequences based on a distance-based threshold; however, its use must be done with caution to avoid the risk of dividing a single bacterial genome into separate clusters [33]. Once the sequences were refined, a total of 90,904 ASVs were obtained for the microbiome of control children and 105,312 ASVs for underweight children and 91,526 for underweight children after amaranth consumption.
Alpha diversity, which reflects the gut microbial richness, is measured as observed species and Shannon index [34]. Although no significant differences amongst groups, a tendency of lower values was observed in UN group, but after amaranth consumption, UNA group showed a tendency to increase at similar values than in Ctrl group (Figure 2A and Figure 2B). Simpson´s inverse index, which indicates the diversity or dominance of species in the sample, also showed a tendency to decrease in UN group, which suggests that the bacterial community was smaller in this group. However, this value also increases after amaranth consumption or UNA group (Figure 2C), and the same tendency was observed for Chao1 (Figure 2D), and for Fisher, ACE, and Simpson values (Supplementary Figure S1).
Reports indicate that children with undernutrition showed lower gut microbiota diversity [12,35,36], which agrees with the observed data in the present work. It was also reported that Shannon index showed a tendency to decrease in malnourished children with protein malnutrition (Kwashiorkor), although no significant values were reported [37]. It is known that results in human are sometimes not statistically significant because of the intrinsic variation of everyone, but they are clinically relevant as they reflect a clear trend [38]. Beta diversity reflects the differences in the gut microbial composition of different groups and is represented by non-metric multidimensional scaling (NMDS) or PCoA, based on Bray-Curtis dissimilarity. As shown in Figure 3, PCoA analysis showed a distinction in gut microbiota between Ctrl and UN groups, but after amaranth consumption, UNA group tend to be more like the Ctrl group. This data are in agreement with recent reports that showed that quinoa protein and its hydrolysates were able to restore the gut microbiota in a mice model of colorectal cancer similar to the control group [39].
The major phylum in gut microbiota of children Ctrl group was Actinobacteriota (28.8%), followed by Bacteroidota and Firmicutes (40.2%), Proteobacteria (12.8), and Verrucomicrobiota (16.8%). It was observed that UN group showed a significant increase of Actinobacteriota (50.9%) with the loss of Verrucomicrobiota phylum, which agrees with data reported by Kamil et al. [40]. After the trial, UNA group showed a significant decrease in abundance of Actinobacteria (20%) with an increase of Bacteriodotes and Firmicutes, interestingly Verrumicrobiota was recovered after amaranth consumption with similar values as control group (13.4%) (Figure 4A). It has been reported that Proteobacteria is the phylum that contributes to dysbiosis [41]; however, in this work this phylum showed not significant changes in abundance amongst groups (12.8 to 15.4%) (Figure 4A).
It is considered that Firmicutes and Bacteroidetes ratio (F/B) is important to determine the health status and known as the dominant phyla in gut microbiota. Changes in F/B ratio has been associated with several diseases; however, it is important to be aware that this ratio could be affected by the change in abundance of all other phyla, as observed in present results. Our data showed that after amaranth consumption these two phyla reach up to 51.6% and F/B was even higher than in control group (Figure 4B). Studies reported that a diet high in sugar and low in fiber in children with undernutrition might cause a high F/B ratio [42]. The abundance of Firmicutes has been associated with the modulation of calorie absorption’s affectivity by increasing the number of lipid droplets [43], and then a higher F/B ratio has been associated with the incidence of obesity [44]. However, other works reported that a relative abundance of Firmicutes was found considerably higher in centenarians concluding that this phylum was related with host longevity and after probiotics administration in undernourished children [45]. Thus, until date no direct relation of F/B ratio with the health status exists.
At family level, high abundance of Eubacterium was observed in UN group with a significant decrease after amaranth consumption (24.8% and 11.1%). On the contrary Akkermansiaceae family was found in very low abundance (3.3%) in UN group, but this family increases after amaranth consumption (9.1%) even at higher abundance than control group (1.0%). Alcaligenaceae family was significanlty increase in abundance after amaranth consumption (11.7%) and Bacteroidaceae family that was almost depleted in UN group (2.6%) increases after amarnath consumption to similar values to control group (5.9 and 7.6%, respectively). Christensenellaceae family was also found in high abundance after amaranth consumption (7.8%) while Coriobacteriaceae decreases from 7.2% in undernoursidhed group to 2.0% after amarnath consumpition (Figure 4C).
3.4. Modulation of gut microbiota at family-genus level was observed in undernourished children after amaranth consumption
As describe above, although the F/B ratio reflects an estimation of persons health, attention should be taken because this ratio could be affected by the change in other phyla, as well as the type of the genus that changes in each phylum. After amaranth consumption most of the species changes in relative abundance were observed in Firmicutes phyla.
3.4.1. Changes in Firmicutes phylum at genus/species level
It was reported that Lachnospiraceae family belonging to Eubacteriales order are in over-abundance in undernourished children, which some of members of this family serve as metabolic regulators in undernourished individuals [42]. Our results also showed high abundance of this family in undernutrition group (Figure 4) and changes in relative abundance in Eubacterium hallii, Ruminococcus gauvreauii, Blautia faecis, Blautia obeum, CAG-56, Fusicanilobacter, Roseburia hominis and Roseburia intestinalis, amongst others were observed after amaranth consumption (Figure 5). E. hallii was observed to decrease in undernutrition group, but after amaranth consumption there was a relative increase. E. hallii is an anaerobic bacterium that belongs to the group of butyrate producers [46], and its administration in mice with diabetes changed the function of the intestinal microbiome, improving the metabolic phenotype [47].
At the genus level, Blautia has taken special interest since its establishment in the gastrointestinal tract has been related to alleviate inflammatory and metabolic diseases as well as with antimicrobial activity. This bacterium has also been reported to play a role in the crosstalk with other intestinal microorganisms [48]. Blautia actively participates in intestinal immunomodulation in human health [38,48], improves serum HDL-C, and its relative increase in abundance was reported after a diet intervention in obesity [49]. Although B. obeum has been related to obesity [50], its abundance in undernourished children after amaranth consumption was at the same level as in control children (Figure 5).
Roseburia hominis, and Roseburia intestinalis were identified, both showed low changes in their relative abundances after amranth consumption, while the first decreases the second increases. Roseburia genus are described as benefical organisms as they are butyrate-producing bacteria and both have been found depleted in stool of undernourished children [35]. R. hominis was positively related to colonic mucosal melatonin levels, a pineal hormone that can maintain circadian rhythms and regulate immune, antioxidant, and anti-inflammatory functions, alleviating the symptoms of digestive disorders, such as irritable bowel syndrome and ulcerative colitis [51]. It is also known that R. intestinales prevent inflammation and maintain energy homeostasis [52].
Within the Erysipelotrichaceae family, Holdemanella, was found in a relative increase after amaranth consumption (Figure 5). In malnutrition, it has shown that Holdomanella was depleted [53]. Holdomanella has been isolated from the feces of healthy metabolic volunteer and has been linked to amelioration of hyperglycemia, improvement of oral glucose tolerance, and restoration of gluconeogenesis and insulin signaling in the liver of obese mice [54].
Streptococcaceae family was represented by relative increase in abundance of S. salivarus and S. thermophilus after amaranth consumption (Figure 5). S. thermophilus is a well-known probiotic to which beneficial effects on health have been attributed and it is commonly found in yogurts [55]. S. salivarius is a commensal in humans and is phylogenetically close to S. thermophilus [56]. Within Oscillospiraceae Family, Subdoligranulum showed a high relative abundance after amaranth consumption. Subdoligranulum is a butyrate-producer bacterium with high potential as probiotic to handle metabolic diseases. It has also activity of acetyl-CoA acetyltransferase, acetyl/propionyl-CoA carboxylase, and butanol dehydrogenase, which contribute to butyric acid production [57]. Butyrate has several beneficial properties that are essential to maintain gastrointestinal health. Therefore, the butyrate-producing bacteria are niche-specific next-generation of probiotics and Butyricicoccus species are one of the potential bacteria [58]. Butyricicoccus have also been shown to have a potential as therapeutic target for food allergy [59]. Butyricicoccus showed an increase in relative abundance in undernourished children after amaranth consumption.
3.4.2. Changes in Bacteroidetes phylum at genus/species level
Within Bacteroidota phylum, a relative increased in Bacteroides coprocola and Bacteroides stercoris was observed in undernuntrition group, but after amaranth consumption their relative abundance decreased as well as Alistipes putredinis (Figure 5). Different strains of B. coprocola have been reported, some are related to type 2 diabetes and some of them have been identified in patients with hypertension, and some others have been linked to patients with attention-deficit/hyperactivity disorder [60], thus it is suggested to be cautious about its presence or absence [61,62]. Although B. stercoris is a SCFA producing bacteria, it has been related to ulcerative colitis as well as Alistipes putredinis [63].
A decrease of relative abundance of B. fragilis, B. ovatus, and B. plebeius was observed after amaranth consumption when compared with control group (Figure 5). B. fragilis is a commensal organism but it has also been reported as an opportunistic pathogen in clinical infections and as responsible for a variety of diseases involving the disruption of the intestinal barrier [64]. However, it has been reported that in mice the non-toxigenic B. fragilis strain may mediate beneficial interactions with the host by directing the immune response and suppressing intestinal inflammation [65]; thus, a deep characterization should be carried out to identify the B. fragilis species detected in undernourished children. Some of the B. ovatus strains have been related to the induction of gut IgA and maintenance of tissue homeostasis [66]. On the other hand, B. plebeius was found to be a dominant bacterium in children with the multisystemic inflammatory syndrome [67]. Only B. intestinalis showed a high relative abundance after amaranth consumption (Figure 5), this bacteria has been identified as polyamine-producing bacteria such as putrescine, spermidine, and spermine, all of them are organic cations required for animal cell growth and differentiation, they are also involve in various steps of DNA, RNA, and protein synthesis [68].
3.4.3. Changes in Verrucomicrobiota phylum
Amaranth consumption induces a relative increase in abundance of Akkermansia muciniphila (Figure 5). A. muciniphila, is a colonic mucin-degrading bacterium that has been linked to intestinal health and immune signaling, and has been related with a reduction of the incidence of diabetes when administered as a probiotic [69]. It was reported that A. muciniphila improves metabolic disorders in dietary obese mice and was found to increase after resistant starch intake, then suggested as a new probiotic with the capacity to promote healthy longevity [70]. A. muciniphila can metabolize the mucosal layer as a source of carbon and nitrogen to produce acetate and propionate, playing an important role in maintaining the gut barrier, especially when enteral nutrition intake is low, such as long-term fasting and malnutrition [71].
It has shown that the diversity and richness of the mouse microbiota changed when fed fermented soybeans in the diet, resulting in the enhancement of Lactobacillus, Butyricicoccus, Lachnospiraceae, and A. muciniphila [72], data that is in agreement with our results that have shown that after amaranth consumption those species were also enriched in undernourished children.
3.5. Amaranth consumption promotes gut microbiota-dependent metabolites
Popped amaranth grains, in addition of high-quality proteins, are rich in carbohydrates that escape digestion (resistant starch) as well as dietary fiber from testa [13]. The gut microbiota is predominantly involved in the fermentation of indigestible carbohydrates into SCFAs, which exert multiple effects on energy homeostasis and are crucial for intestinal health. The most abundant SCFA are acetate, butyrate, and propionate, comprising >95% of the SCFA content [73]. Investigations have shown that SCFA concentration in children with severe acute and moderate undernutrition is low, especially propionic and butyric acid and those SCFA are increase during recovery along with gut microbiota number. In this study, it was observed that after amaranth consumption, the levels of acetic, butyric, and propionic acids showed a tendency to increase although only propionic acid was significant as well as the sum of these three SCFAs (Figure 6). Eubacterium hallii, Ruminococcus, Blautia, Roseburia hominis, and Butyricicoccus, known as butyrate-producer species [40], all of them were observed in increased relative abundance after amaranth consumption. A. muciniphila has been described as acetate and propionate producer bacterium [71] and Subdoligranulum sp., as a butyrate-producer bacterium [57], both of them were also found in increased abundance after amaranth consumption. Altogether, these data suggested that SFCAs levels are restored after amaranth consumption with a correlation with the increased abundance of SCFAs producer bacteria.
3.8. Functional prediction of bacterial taxa
The metabolic pathway predictions of gut microbiota composition in undernutrition children compared as control group (UN/Ctrl) (Figure 7), showed increased in starch and sucrose metabolism, which decreased after amaranth consumption (UNA/Ctrl). Biosynthesis of secondary metabolism was increased after amaranth consumption as well as butanoate, pyruvate and propanoate metabolism, which are the routes to produce butyric and propionic acid, which correlates with tendency to increase on these metabolites after amaranth consumption (Figure 7)
Purine and amino sugar and nucleotide metabolism, as well as in pathways related to vitamins metabolism such as biotin (vitamin H), nicotinate and nicotinamide (B3), and riboflavin (B2), showed a tendency to increase after amaranth consumption. Increases of metabolic pathways related to sphingolipid and arachidonic acid metabolism was observed after amaranth consumption. Whereas sphingolipids are considered as a class of bioactive lipids playing key roles in regulation of several cellular processes, while arachidonic acid is part of cell membranes and is important for human health and tissues homeostasis. A correlation between Ruminococcaceae UCG 009 and arachidonic acid has been reported [74]. Several Ruminocococcus species were detected here after amaranth consumption (Figure 5).
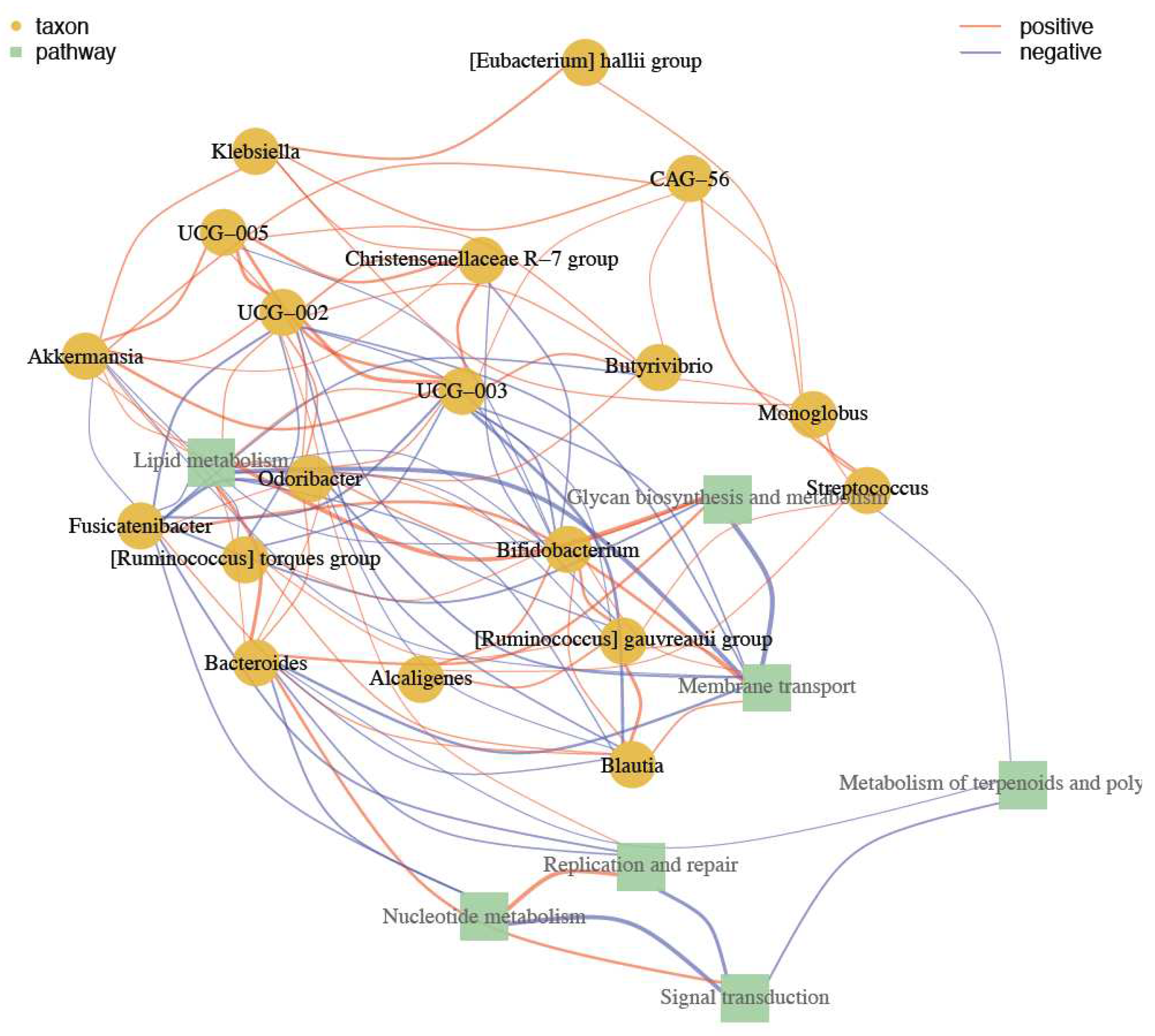
A network of correlation with metabolic activities was obtained and as shown in Figure 8, Akkermansia appear as an important species in controlling Ruminococcus, Bacteroides, and Blautia. Also has a positive relation with Klebsiella and E. hallii. Akkermansia has also a positive correlation with species from Oscillospiracea family such as UCG-005, 003, and 002,. UCG strains regulates to Christensenellaceae R-7 group, Butyrivibrio. These group of bacteria have an action in regulation of nucleotide and terpenoids metabolism, transport, signal transduction and replication and repair.
4. Conclusions
Amaranth grains, in addition to high-quality proteins, are rich in vitamins and minerals. Popped amaranth has been claimed as food source with high potential to combat protein-energy malnutrition. Our results showed popped amaranth intervention improves children´s health through gut microbiota modulation. An increased relative abundance of Akkermansia muciniphila, a bacterium that is being important for intestinal health and host longevity, as well as Subdoligranulum, considered as new class of probiotics was observed. Also, was observed the decrease in relative abundance of Bacteroides coprocola and B. stercoris, both related to inflammation and colitis. Streptococcus salivarius and S. thermophilus were detected also in increased abundance in children after amaranth consumption. An increase of SCFAs was correlated with the increased of SCFAs-produced bacteria. In summary, the present work highlights the potential uses of popped amaranth as a source of plant-based proteins, which requires minimum processing to achieve its biological function on health. However, eradication of malnutrition is not only through the with the improvement of infrastructure conditions in rural areas.
Even thought the intervention of popped amaranth shows positive results, mainly in modulating the SCFAs-producing bacteria, the low sample size and short intervention time were limitations of this study. Hence, a second step it is necessary with larger randomized controlled trial to confirm the beneficial effects of popped amaranth consumption, but not only in undernourished children, but also in child with overweight and obesity.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Table S1: Proximal composition of popped amaranth (Amaranthus hyphochondriacus). Supplementary Table S2: Hematic cytometry of participant´s children with low height-for age according to HAZ index. Supplementary Table S3: Serum biochemical profile of participant children. Supplementary Figure S1: Alpha-diversity according to Fisher, ACE , and Simpson values. Supplementary Figure S2: Distribution of gut microbiota at class and order levels.
Author Contributions
OJCC and APBR: Conceptualization, methodology, and investigation. COV: bioinformatics, data curation, and writing review. OJCC and ADLR: SCFAs analysis. OJCC, FV: investigation, blood and feces samples. EER: amaranth popped preparation. OJCC and ST: hematological and biometry analysis. APBR: supervision, funding acquisition, project administration, writing, review & editing
Funding
This work was supported by National Grant from “Fondo Sectorial de Investigación en Salud y Seguridad Social (No. A3-S-37825).
Data Availability Statement
Data will be made available on request
Acknowledgments
The authors acknowledge the Instituto Potosino de Investigación Científica y Tecnológica (IPICYT) and National Supercomputing Center (CNS) computing resources, provided to develop this project under the project grant TKII-R2018- COV1. COV acknowledges the Investigadores por México CONACYT program. Thanks to Rural Schools located at Ciudad Valles, San Luis Potosí, and all participants children and their parents. We thanks to L. Medina-Contreras and C. I. Cano-Gómez for their technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gordon, J.I.; Dewey, K.G.; Mills, D.A.; Medzhitov, R.M. The human gut microbiota and undernutrition. Sci. Transl. Med. 2012, 4, 1–12. [Google Scholar] [CrossRef]
- Blanton, L.V.; Barratt, M.J.; Charbonneau, M.R.; Ahmed, T.; Gordon, J. Childhood undernutrition, the gut microbiota, and microbiota-directed therapeutics. Science 2016, 352(6293), 1533. [Google Scholar] [CrossRef]
- WHO/UNICEF. Levels and trends in child malnutrition: UNICEF/WHO/The World Bank Group joint child malnutrition estimates: key findings of the 2021 edition. Retrieved October 2022. https://www.who.int/publications/i/item/9789240025257.
- Prendergast, A.J.; Humphrey, J.H. The stunting syndrome in developing countries. Paediatr. Int. Child Health 2014, 34, 250–265. [Google Scholar] [CrossRef] [PubMed]
- Iddrisu, I.; Monteagudo-Mera, A.; Poveda, C.; Pyle, S.; Shahzad, M.; Andrews, S.; Walton, G.E. Malnutrition and gut microbiota in children. Nutrients 2021, 13, 2727. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Gallego, C.; García-Mantrana, I.; Martínez-Costa, C.; Salmien, S.; Isolauri, E.; Collado, M.C. The microbiota and malnutrition: Impact of nutritional status during early life. Annu. Rev. Nutr. 2019, 39, 267–290. [Google Scholar] [CrossRef]
- Calder, N.; Walsh, K.; Olupot-Olupt, P.; Ssenyondo, T.; Muhindo, R.; Mpoya, A.; Brignardello, J.; Wang, X.; McKay, E.; Morrison, D.; Holmes, E.; Frost, G.; Maitland, K. Modifying gut integrity and microbiome in children with severe acute malnutrition using legume-based feeds (MIMBLE): A pilot trial. Cell Reports Medicine 2021, 2, 100280. [Google Scholar] [CrossRef]
- Nova, E.; Gómez-Martinez, S.; González-Soltero, R. The influence of dietary factors on the gut microbiota. Microorganisms 2022, 10, 1368. [Google Scholar] [CrossRef]
- Wu, S.; Bhat, Z. F.; Goudner, R.C.; Mohamed Ahmed, I.A.; Al-Juhaimi, F.Y.; Ding, Y.; Bekhit, A.E-D. Effect of dietary protein and processing on gut microbiota- A systematic review. Nutrients 2022, 14, 453. [Google Scholar] [CrossRef]
- Berding, K.; Vlckova, K.; Marx, W.; Schellekens, H.; Stanton, C.; Clarke, G.; Jacka, F.; Dinan, T.G.; Cryan, J.F. Diet and the microbiota-gut-brain-axis: sowing the seeds of good mental health. Adv. Nutr. 2021, 12, 1239–1285. [Google Scholar] [CrossRef]
- Aloo, S.O.; Oh, D.-H. The functional interplay between gut microbiota, protein hydrolysates/bioactive peptides, and obesity: A critical review on the study advances. Antioxidants 2022, 11, 333. [Google Scholar] [CrossRef]
- Gatya, M.; Nur Fibri, D.L.; Utami, T.; Suroto, D.A.; Rahayu, E.S. Gut microbiota composition in undernourished children associated with diet and sociodemographic factors: A case-control study in Indonesia. Microorganisms 2022, 10, 1748. [Google Scholar] [CrossRef]
- Marques Coelho, L.; Silva, P.M.; Martins, J.T.; Pinheiro, A.C.; Vicente, A.A. Emerging opportunities in exploring the nutritional/functional value of amaranth. Food Funct. 2018, 9, 5499. [Google Scholar] [CrossRef]
- Silva-Sánchez, C.; Barba de la Rosa, A.P.; León-Galván, M.F.; de Lumen, B.O.; De León-Rodríguez, A.; González de Mejía, E. Bioactive peptides in amaranth (Amaranthus hypochondriacus) seed. J. Agric. Food Chem. 2008, 56, 1233–1240. [Google Scholar] [CrossRef]
- Kaufam, C.S.; Weber, L.E. Grain amaranth. In J. Janick, and J.E. Simon (Eds). Advances in new crops 1990 pp. 127–139. Portland: Timber Press.
- Solanki, Ch.; Indore, N.; Saha, D.; Kudos, S.K.A. Effect of popping methods on popping characteristic of amaranth grain. Int. J. Chem. Stud. 2018, 6(2), 2779–2782. [Google Scholar]
- Valadez-Vega, C.; Lugo-Magaña, O.; Figueroa-Hernández, C.; Bautista, M.; Betanzos-Cabrera, G.; Bernardino-Nicanor, A.; González-Amaro, R.M.; Alonso-Villegas, R.; Morales-González, J.A.; González-Cruz, L. Effect of germination and popping on the anti-nutritional compounds and the digestibility of Amaranthus hypochondriacus seeds. Foods 2022, 11, 2075. [Google Scholar] [CrossRef]
- Paško, P.; Bartoń, H.; Folta, M.; Gwizdz, J. Evaluation of antioxidant activity of amaranth (Amaranthus cruentus) grain and by-products (flour, popping, cereal). Rocz. Panstw. Zakl. Hig. 2007, 58(1), 35–40. [Google Scholar] [PubMed]
- Muyonga, J.H.; Andabati, B.; Seepuuya, G. Effect of heat processing on selected grain amaranth physicochemical properties. Food Sci. Nutri. 2014, 2, 9–16. [Google Scholar] [CrossRef]
- Aderibigbe, O.R.; Ezekiel, O.O.; Owolade, S.O.; Korese, J.K.; Sturm, B.; Hensel, O. Exploring the potentials of underutilized grain amaranth (Amaranthus spp.) along the value chain for food and nutrition security: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Grundy, M.M.L.; Momanyi, D.K.; Holland, C.; Kawaka, F.; Tan, S.; Salim, M.; Boyd, B.J.; Bajka, B.; Mulet-Cabero, A-I; Bishop, J.; Owino, W,O. Effects of grain source and processing methods on the nutritional profile and digestibility of grain amaranth. J. Funct. Foods 2020, 72, 104065. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis. 18th ed. Association of Official Analytical Chemists, 2007, Washington, D.C.
- Gao, L.; Zhang, L.; Liu, H.; Hu, J. In vitro gastrointestinal digestion of whole grain noodles supplemented with soluble dietary fiber and their effects on children fecal microbiota. Food Biosci. 2023, 53, 102600. [Google Scholar] [CrossRef]
- Comeau, A.M.; Li, W.K.W.; Tremblay, J.E.; Carmack, E.C.; Lovejoy, C. Arctic ocean microbial community structure before and after the 2007 record sea ice minimum. PLoS ONE 2011, 6, e27492. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O´Hara, R.B.; Solymos, P., ... Weedon, J. Vegan: Community ecology package. R Package 2015, Version 2.2-1 2 1-2.
- Wemheuer, F.; Taylor, J.A.; Daniel, R.; Johnston, E.; Meinicke, P.; Thomas, T.; Wemheuer, B. Tax4Fun2: prediction of habitat-specific functional profiles and functional redundancy based on 16S rRNA gene sequences. Environ. Microbiome 2020, 15, 11. [Google Scholar] [CrossRef] [PubMed]
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.-A. mixOmics: An R package for “omics” feature selection and multiple data integration. PloS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef]
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. Int. J. Complex Syst. 2006, 1695, 1–9. [Google Scholar]
- Morales, E.; Lembcke, J.; Graham, G.G. Nutritional value for young children of grain amaranth and maize-amaranth mixtures: Effect of processing. J. Nutr. 1988, 118, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Velasco-Lozano, A.M. Los cuerpos divinos: el amarnato, comida ritual y cotidiana. Amarnato fuente de alegría Arqueología Mexicana 2016, 23, 26–33. [Google Scholar]
- Getawa, S.; Getaneh, Z.; Melku, M. Hematological abnormalities and associated factors among undernourished under-five children attending university of Gondar specialized referral hospital, Northwest Ethiopia. J. Blood Med. 2020, 11, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D. Amplicon sequence variants artificially split bacterial genomes into separate clusters. mSphere 2021, 6, e0019–21. [Google Scholar] [CrossRef]
- Qi, Ch.; Cai, Y.; Qian, K.; Li, X.; Ren, J.; Wang, P.; Fu, T.; Zhao, T.; Cheng, L; Shi, L; Zhang, X. gutMDisorder v2.0: a comprehensive database for dysbiosis of gut microbiota in phenotypes and interventions. Nucl. Acids Res. 2022, 51, D717–D722. [Google Scholar] [CrossRef]
- Monira, Sh.; Nakamura, Sh.; Gotoh, K.; Izutsu, K.; Watanabe, H.; Alam, N.H.; Endtz, H.P.; Cravioto, A.; Ali, S.I.; Nakaya, T.; Horii, T.; Lida, T.; Alam, M. Gut microbiota of healthy and malnourished children in Bangladesh. Front. Microbiol. 2011, 2, 228. [Google Scholar] [CrossRef]
- Kamil, R.Z.; Murdiati, A.; Juffrie, M.; Nakayama, J.; Rahayu, E.S. Gut microbiota and shot-chain fatty acid profile between normal and moderate malnutrition children in Yogyakarta, Indonesia. Microorganisms 2021, 9, 127. [Google Scholar] [CrossRef]
- Subramanian, S.; Hug, S.; Yatsunenko, T.; Haque, R.; Mahfuz, M.; Alam, M.A.; Benezra, A.; DeStefano, J.; Meier, M.F.; Muegge, B.D.; Barratt, M.J.; VanArendonk, L.G.; Zhang, Q.; Province, M.A.; Petri Jr, W.A.; Ahmed, T.; Gordon, J.I. Persistent gut microbiota immaturity in malnourished Bangladeshi children. Nature 2014, 510, 417–421. [Google Scholar] [CrossRef]
- Kim, K.; Veerappan, K.; Woo, N.; Park, B.; Natarajan, S.; Chung, H.; Kim, Ch.; Park, J. Ulmus macrocarpa Hance extract modulates intestinal microbiota in healthy adults: a randomized, placebo-controlled clinical trial. J. Microbiol. 2021, 59, 1150–1156. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Guo, H.; Teng, X.; Yang, X.; Qin, P.; Richel, A.; Zhang, L.; Blecker, Ch.; Ren, G. Supplementation of quinoa peptides alleviates colorectal cancer and restores gut microbiota in AOM/DSS-treated mice. Food Chem. 2023, 408, 135196. [Google Scholar] [CrossRef] [PubMed]
- Kamil, R.Z.; Murdiati, A.; Juffrie, M.; Rahayu, E.S. Gut microbiota modulation of moderate undernutrition in infants through gummy Lactobacillus plantarum Dad-13 consumption: A randomized double-blind controlled trial. Nutrients 2022, 14, 1049. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.-R.; Whon, T.W.; Bae, J.-W. Proteobacteria: microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Méndez-Salazar, E.O.; Ortiz-López, M.G.; Granados-Silvestre, M.A.; Palacios-González, B.; Menjivar, M. Altered gut microbiota and compositional changes in Firmicutes and Proteobacteria in Mexican undernourished and obese children. Front. Microbiol. 2018, 9, 2494. [Google Scholar] [CrossRef]
- Sutoyo, D.A.; Atmaka, D.R.; Sidabutar, L.M.G.B. Dietary factors affecting Firmicutes and Bacteroidetes ratio in solving obesity problem: A literature review. J. Media Gizi Indon. 2020, 15, 94. [Google Scholar] [CrossRef]
- Bervoets, L.; Vankerckhoven, K.V.; Kortleven, I.; Noten, C.V.; Hens, N.; Vael, C.; Goossens, H.; Desager, K.N.; Vankerckhoven, V. Differences in gut microbiota composition between obese and lean children: A cross-sectional study. Gut Pathog. 2013, 5, 10. [Google Scholar] [CrossRef]
- Kim, B.-S.; Choi, C.W.; Shin, H.; Jin, S.-P.; Bae, J.-S.; Han, M.; Seo, E.Y.; Chun, J.; Chung, J.H. Comparison of the gut microbiota of centenarians in longevity villages of South Korea with those of other age groups. J. Microbiol. Biotechnol. 2019, 29, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.M.W.; de Souza, R.; Kendall, C.W.C.; Emam, Z.; Jenkins, D.J.A. Colonic health: fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef]
- Udayappan, S.; Manneras-Holm, L.; Chaplin-Scott, A.; Belzer, C.; Herrera, H.; Dallinga-Thie, G.M.; Duncan, S.H.; Stroes, E.S.G.; Groen, A.K.; Flint, H.J.; Backhed, F.; de Vos, W.M.; Nieuwdorp, M. Oral treatment with Eubacterium hallii improves insulin sensitivity in db/db mice. NPJ Biofilms Microbiomes 2016, 6, 16009. [Google Scholar] [CrossRef]
- Liu, X.; Mao, B.; Gu, J.; Wu, J.; Cui, Sh.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Blautia-a new functional genus with potential probiotic properties? Gut Microbes 2021, 13, 1–21. [Google Scholar] [CrossRef]
- Shi, Z.; Zhu, Y.; Teng, C.; Yao, Y.; Ren, G.; Richel, A. Anti-obesity effects of a-amylase inhibitor enriched-extract from white common beans (Phaseolus vulgaris L.) associated with the modulation of gut microbiota composition in high-fat diet-induced obese rats. Food Funct. 2020, 11, 1624–1634. [Google Scholar] [CrossRef]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The influence of probiotics on the Firmicutes/Bacteroides ratio in the treatment of obesity and inflammatory bowel disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef]
- Song, L.; He, M.; Sun, Q.; Wang, Y.; Zhang, J.; Fang, Y.; Liu, Sh.; Duan, L. Roseburia hominis: Increases intestinal melatonin level by activating p-CREB-AANAT pathway. Nutrients 2022, 14, 117. [Google Scholar] [CrossRef]
- Nie, K.; Ma, K.; Luio, W.; Shen, Zh.; Yang, Zh.; Xiao, M.; Tong, T.; Yang, Y.; Wang, X. Roseburia intestinalis: A beneficial gut organism from the discoveries in genus species. Front. Cell. Infect. Microbiol. 2021, 11, 757718. [Google Scholar] [CrossRef] [PubMed]
- Million, M.; Alou, M.T.; Khelaifia, S.; Bachar, D.; Lagier, J.-Ch.; Dione, N.; Brah, S.; Hugon, P.; Lombard, V.; Armougom, F.; Fromonot, J.; Robert, C.; Michelle, C.; Diallo, A.; Fabre, A.; Guieu, R.; Sokhna, Ch.; Henrissat, B.; Parola, P.; Raoult, D. Increased gut redox and depletion of anaerobic and methanogenic prokaryotes in severe acute malnutrition. Sci. Rep. 2016, 6, 26051. [Google Scholar] [CrossRef] [PubMed]
- Romaní-Pérez, M.; López-Almela, I.; Bullich-Vilarrubias, C.; Rueda-Ruzafa, L.; Gómez Del Pulgar, E.M.; Benítez-Páez, A.; Liebish, G.; Lamas, J.A.; Sanz, Y. Holdemanella biformis improves glucose tolerance and regulates GLP-1 signaling in obese mice. FASEB J. 2021, 35, e21734. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, H.; Hollis, J.H.; Jacques, P.F. The associations between yogurt consumption, diet quality, and metabolic profiles in children in the USA. Eur. J. Nutr. 2015, 54, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Delorme, C.; Abraham, A.-L.; Renault, P.; Guédon, E. Genomics of Streptococcus salivarius, a major human commensal. Infec. Genet. Evol. 2015, 33, 381–392. [Google Scholar] [CrossRef]
- Van Hul, M.; Le Roy, T.; Prifti, E.; Dao, M.C.; Paquot, A.; Zucker, J.-D.; Delzenne, N.M.; Muccioli, G.; Clément, K.; Cani, P.D. From correlation to causality: the case of Subdoligranulum. Gut Microbes 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Boesmans, L.; Valles-Colomer, M.; Wang, J.; Eeckhaut, V.; Falony, G.; Ducatelle, R.; Van Immerseel, F.; Raes, J.; Verbeke, K. Butyrate producers as potential next-generation probiots: safety assessment of the administration of Butyricicoccus pullicaecarum to healthy volunteers. mSystems 2018, 3(6), e00094-18. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Ch.; Huang, J.; Xie, M.; Li, X.; Fu, L. Butyricicoccus plays a key role in mediating the antagonism between probiotic and antibiotic on food allergy. Food Agric. Immunol. 2019, 30, 446–461. [Google Scholar] [CrossRef]
- Wang, L.-J.; Yang, Ch.-Y.; Chou, W.-J.; Lee, M.-J.; Chou, M.-Ch.; Kuo, H.-Ch.; Yeh, Y.-M.; Lee, Sh.Y.; Huang, L.-H.; Li, S.-Ch. Gut microbiota and dietary patterns in children with attention-deficit/hyperactivity disorder. Eur. Child Adolesc. Psychiatry 2020, 29, 287–297. [Google Scholar] [CrossRef]
- Chen, Y.; Li, Z.; Hu, Sh.; Zhang, J.; Wu, J.; Shao, N.; Bo, X.; Ni, M.; Ying, X. Gut metagenomes of type 2 diabetic patients have characteristic single-nucleotide polymorphism distribution in Bacteroides coprocola. Microbiome 2017, 5, 15. [Google Scholar] [CrossRef]
- Calderón-Pérez, L.; Gosalbes, M.J.; Yuste, S.; Valls, R.M.; Pedret, A.; Llauradó, E.; Jimenez-Hernandez, N.; Artacho, A.; Pla-Pagà, L.; Companys, J.; Ludwig, I.; Romero, M.P.; Rubió, L.; Solà, R. Gut metagenomic and short chain fatty acids signature in hypertension: a cross-sectional study. Sci. Rep. 2020, 10, 6436. [Google Scholar] [CrossRef]
- Nomura, K.; Ishikawa, D.; Okahara, K.; Ito, Sh.; Haga, K.; Takahashi, M.; Arakawa, A.; Shibuya, T.; Osada, T.; Kuwahara-Arai, K.; Kirikae, T.; Nagahara, A. Bacteroidetes species are correlated with disease activity in ulcerative colitis. J. Clin. Med. 2021, 10, 1749. [Google Scholar] [CrossRef]
- Sun, Y.; Wu, D.; Zeng, W.; Chen, Y.; Guo, M.; Lu, B.; Li, H.; Sun, Ch.; Yang, L.; Jiang, X.; Gao, Q. The role of intestinal dysbacteriosis induced arachidonic acid metabolism disorder in inflammaging in atherosclerosis. Front. Cell. Infect. Microbiol. 2021, 11, 618265. [Google Scholar] [CrossRef]
- Chu, H.; Khosravi, A.; Kusumawardhani, I.P.; Kwon, A.H.K.; Vasconcelos, A.C.; Cunha, L.D.; Mayer, A.E.; Shen, Y.; Wu, W.-L.; Kambla, A.; Targan, S.R.; Xavier, R.J.; Ernst, P.B.; Green, D.R.; McGovern, D.P.B.; Virgin, H.W.; Mazmanian, S.K. Gene-microbiota interactions contribute to the pathogenesis of inflammatory bowel disease. Science 2016, 352, 1116–1120. [Google Scholar] [CrossRef] [PubMed]
- Yang, Ch.; Mogno, I.; Contijoch, E.J.; Borgerding, J.N.; Aggarwala, V.; Li, Zh.; Siu, S.; Grasset, E.K.; Helmus, D.S.; Dubinsky, M.C.; Mehandru, S.; Cerutti, A.; Faith, J.J. Fecal IgA levels are determined by strain-level differences in Bacteroides ovatus and are modifiable by gut microbiota manipulation. Cell Host Microbe 2020, 27, 467–475. [Google Scholar] [CrossRef]
- Suskun, C.; Killic, O.; Ciftdogan, D.Y.; Guven, S.; Karbuz, A.; Parlakay, A.O.; Kara, Y.; Kacmaz, E.; Sahin, A.; Boga, A.; Isancli, D.K.; Gulhan, B.; Kanik-Yuksek, S.; Kiral, E.; Bozan, G.; Arslanoglu, M.O. Kizil, M.C.; Dinleyci, M.; Us, T.; Varis, A.; Kaya, M.; Vandenplas, Y.; Dinleyici, E.C. Intestinal microbiota composition of children with infection with severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and multisystem inflammatory syndrome (MIS-C). Eur. J. Pediatr. 2022, 181, 3175–3191. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Bakir, M.; Kitahara, M.; Sakamoto, M.; Matsumoto, M.; Benno, Y. Bacteroides intestinalis sp. Nov., isolated from human faeces. Int. J. Syst. Evol. Microbiol. 2006, 56(Pt1), 151–154. [Google Scholar] [CrossRef]
- Hänninen, A.; Tivonen, R.; Pöysti, S.; Belzer, C.; Plovier, H.; Ouwerkerk, J.P.; Emani, R.; Cani, P.D.; De Vos, W.M. Akkermansia mucciniphila induces gut microbiota remodeling and controls islet autoimmunity in NOD mice. Gut 2018, 67, 1445–1453. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Del Cerro, E.; Lambea, M.; Félix, J.; Salazar, N.; Gueimonde, M.; De la Fuente, M. Daily ingestion of Akkermansia mucciniphila for one month promotes healthy aging and increases lifespan in old female mice. Biogerontology 2022, 23, 35–52. [Google Scholar] [CrossRef] [PubMed]
- Derrien, M.; van Baarlen, P.; Hooiveld, G.; Norin, E.; Müller, M.; de Vos, W.M. Modulation of mucosal immune response, tolerance, and proliferation in mice colonized by the mucin-degrader Akkermansia muciniphila. Front. Microbiol. 2011, 2, 166. [Google Scholar] [CrossRef]
- Han, W.; Zhuang, X.; Liu, Q.; Sun, B.; Miao, H.; Zhang, X. Fermented soy whey induced changes on intestinal microbiota and metabolic influence in mice. Food Sci. Hum. Well. 2022, 11, 41–48. [Google Scholar] [CrossRef]
- Cummings, J.H. Short chain fatty acids in the human colon. Gut 1981, 22(9), 763–779. [Google Scholar] [CrossRef]
- Sun, Y.; O’Riordan, M.X.D. Regulation of bacterial pathogenesis by intestinal short-chain fatty acids. Adv. Appl. Microbiol. 2013, 85, 93–118. [Google Scholar] [CrossRef]
Figure 1.
Study design for amaranth supplementation in the normal diet of children living in rural areas. Design for the evaluating children between the ages of 6-7 years with moderate acute malnutrition treated with amaranth.
Figure 1.
Study design for amaranth supplementation in the normal diet of children living in rural areas. Design for the evaluating children between the ages of 6-7 years with moderate acute malnutrition treated with amaranth.
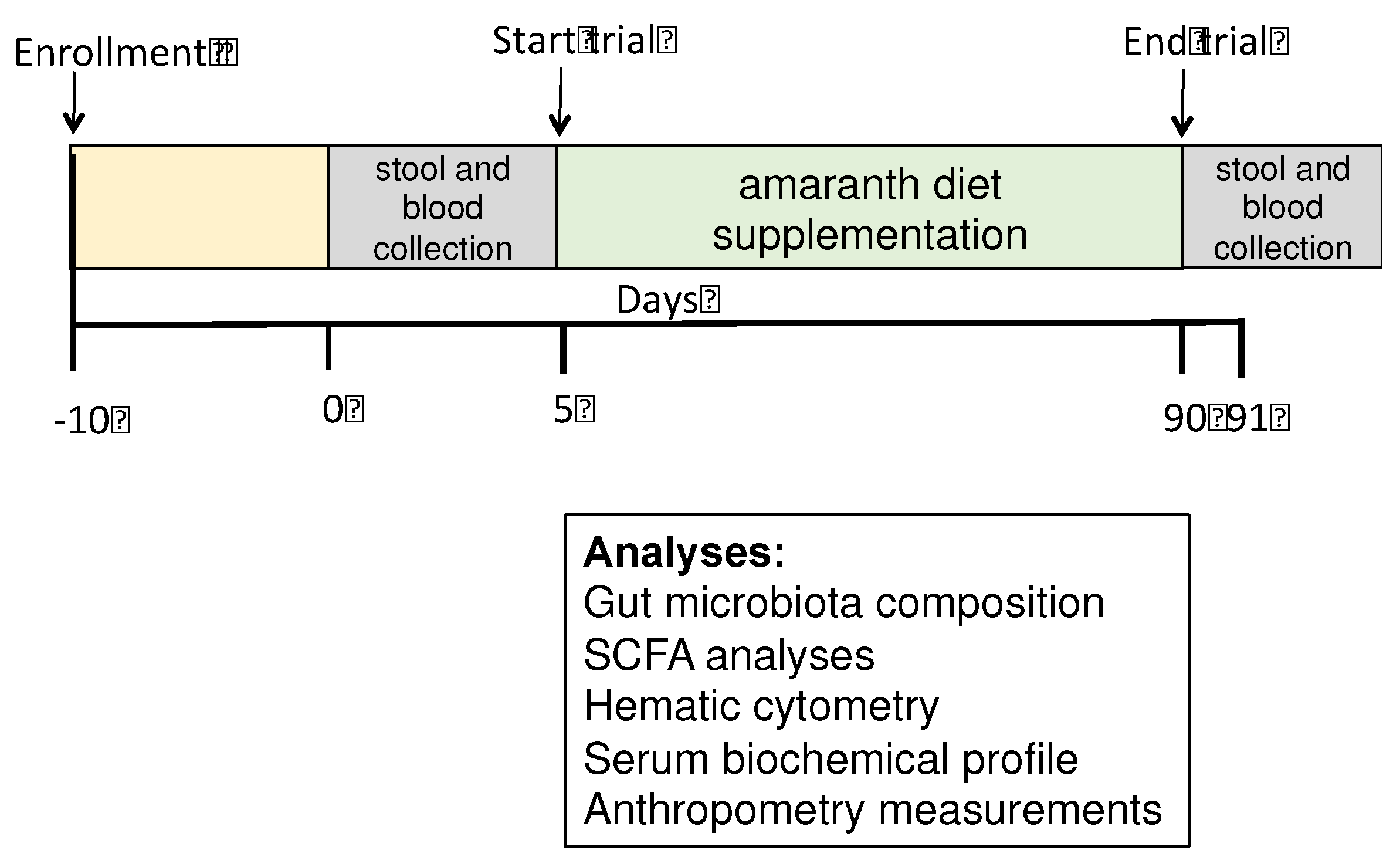
Figure 2.
The effect of amaranth consumption on the gut microbiota composition of children living in rural areas. Alpha-diversity expressed as A) Observed richness, B) Shannon index, C) Inv-Simpson. Boxes express the IQR (interquartile range), bars indicated the minimum and maximum values. A Kruskall-Wallis test and Dunn´s post-hoc analysis (p<0.05) were performed. No significant differences were found amongst groups and different indexes. Ctrl=control group of children with normal height-for-age; UN=undernutrition group with low height-for-age; UNA=UN group after three months of amaranth consumption.
Figure 2.
The effect of amaranth consumption on the gut microbiota composition of children living in rural areas. Alpha-diversity expressed as A) Observed richness, B) Shannon index, C) Inv-Simpson. Boxes express the IQR (interquartile range), bars indicated the minimum and maximum values. A Kruskall-Wallis test and Dunn´s post-hoc analysis (p<0.05) were performed. No significant differences were found amongst groups and different indexes. Ctrl=control group of children with normal height-for-age; UN=undernutrition group with low height-for-age; UNA=UN group after three months of amaranth consumption.
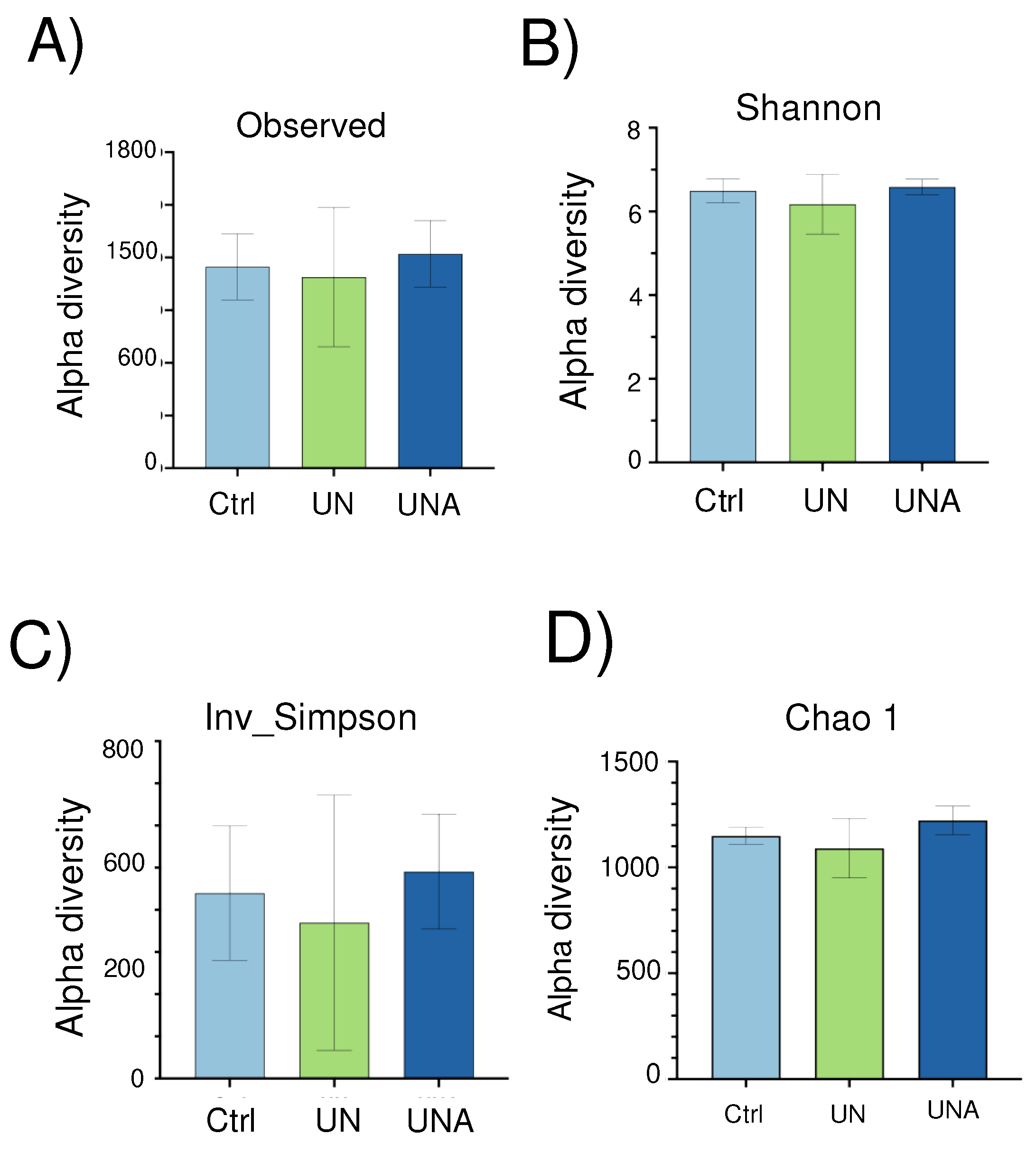
Figure 3.
The effect of amaranth consumption on the gut microbiota composition of children living in rural areas. Beta-diversity measured as principal coordinate analysis (PCoA) based on Bray-Curtis dissimilarity (p<0.05). Ctrl=control group of children with normal height-for-age; UN=undernutrition group with low height-for-age; UNA=UN group after three months of amaranth consumption.
Figure 3.
The effect of amaranth consumption on the gut microbiota composition of children living in rural areas. Beta-diversity measured as principal coordinate analysis (PCoA) based on Bray-Curtis dissimilarity (p<0.05). Ctrl=control group of children with normal height-for-age; UN=undernutrition group with low height-for-age; UNA=UN group after three months of amaranth consumption.
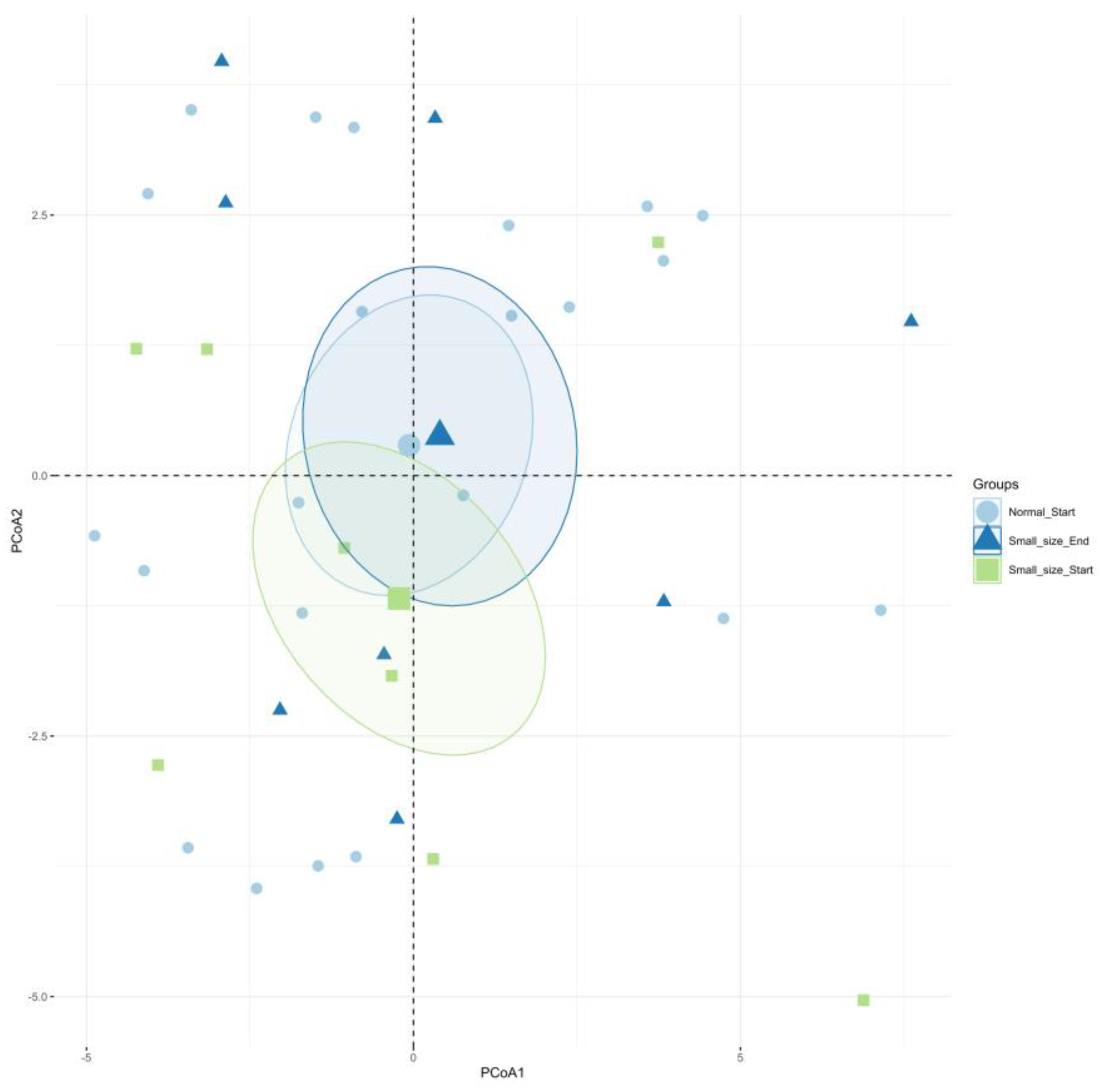
Figure 4.
A) Relative abundance in gut microbiota at phylum level; B) Ratio of Firmicutes to Bacteroidetes (B/F). Significant differences were calculated using the non-parametric Wilcoxon test and not-significant differences were detected. C) Relative abundance in gut microbiota at family level. Ctrl=control group of children with normal weight-height-for-age; UN=undernutrition group with low height-for-age; UNA=undernutrition group after three months of amaranth consumption.
Figure 4.
A) Relative abundance in gut microbiota at phylum level; B) Ratio of Firmicutes to Bacteroidetes (B/F). Significant differences were calculated using the non-parametric Wilcoxon test and not-significant differences were detected. C) Relative abundance in gut microbiota at family level. Ctrl=control group of children with normal weight-height-for-age; UN=undernutrition group with low height-for-age; UNA=undernutrition group after three months of amaranth consumption.
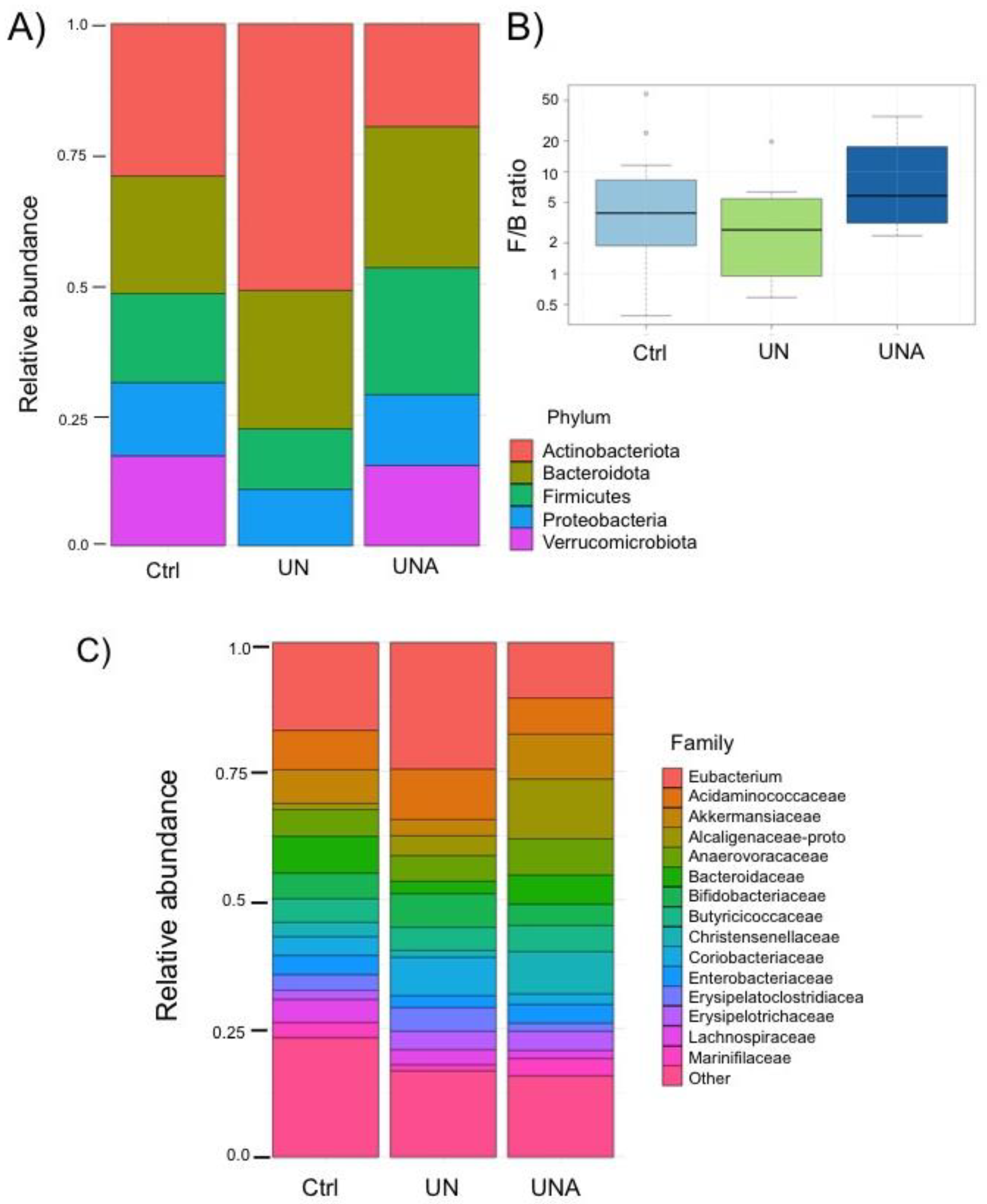
Figure 5.
Heatmap of differentially ASVs observed between: UN/Ctrl, undernutrition group vs control group (Ctrl); UNA/UN, undernutrition group after amaranth consumption (UNA) vs undernutrition group at the beginning of assay (UN); UNA/Ctrl, undernutrition group after amaranth consumption (UNA) vs control group (Ctrl). Values were determined using LogFC, at p<0.05 and FDR <0.1.
Figure 5.
Heatmap of differentially ASVs observed between: UN/Ctrl, undernutrition group vs control group (Ctrl); UNA/UN, undernutrition group after amaranth consumption (UNA) vs undernutrition group at the beginning of assay (UN); UNA/Ctrl, undernutrition group after amaranth consumption (UNA) vs control group (Ctrl). Values were determined using LogFC, at p<0.05 and FDR <0.1.
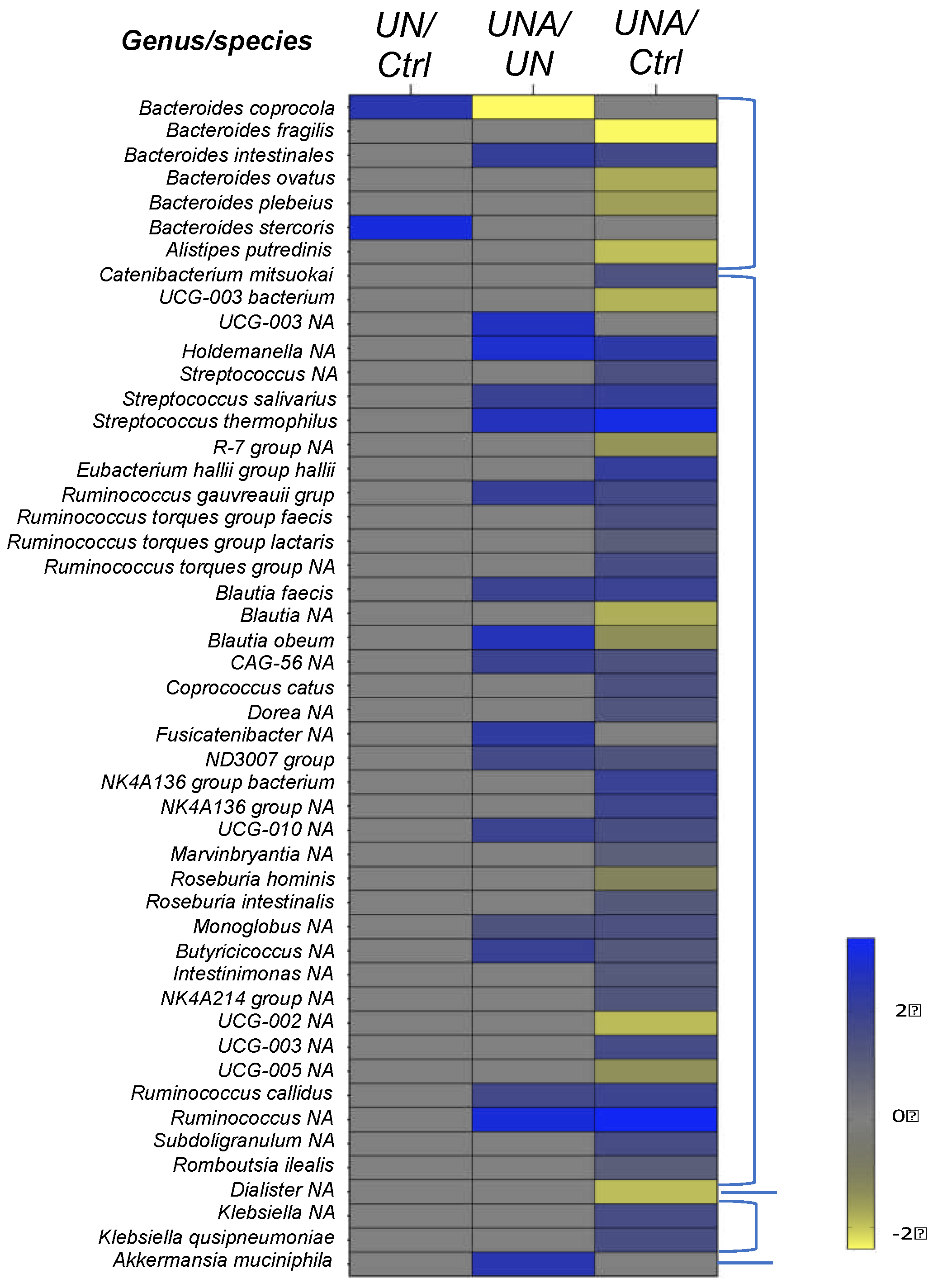
Figure 6.
Short Chain Fatty Acids (SCFA) levels in children feces samples. One-way ANOVA was carried out followed by Tukey´s post-hoc test. Data is presented as mean ±standard deviation. Asterisk shows significant differences at p<0.05. Ctrl=control group of children normal weight-high-for-age; UN=undernutrition children group with low height-for-age; UNA=undernutrition children group after three months of amaranth consumption.
Figure 6.
Short Chain Fatty Acids (SCFA) levels in children feces samples. One-way ANOVA was carried out followed by Tukey´s post-hoc test. Data is presented as mean ±standard deviation. Asterisk shows significant differences at p<0.05. Ctrl=control group of children normal weight-high-for-age; UN=undernutrition children group with low height-for-age; UNA=undernutrition children group after three months of amaranth consumption.
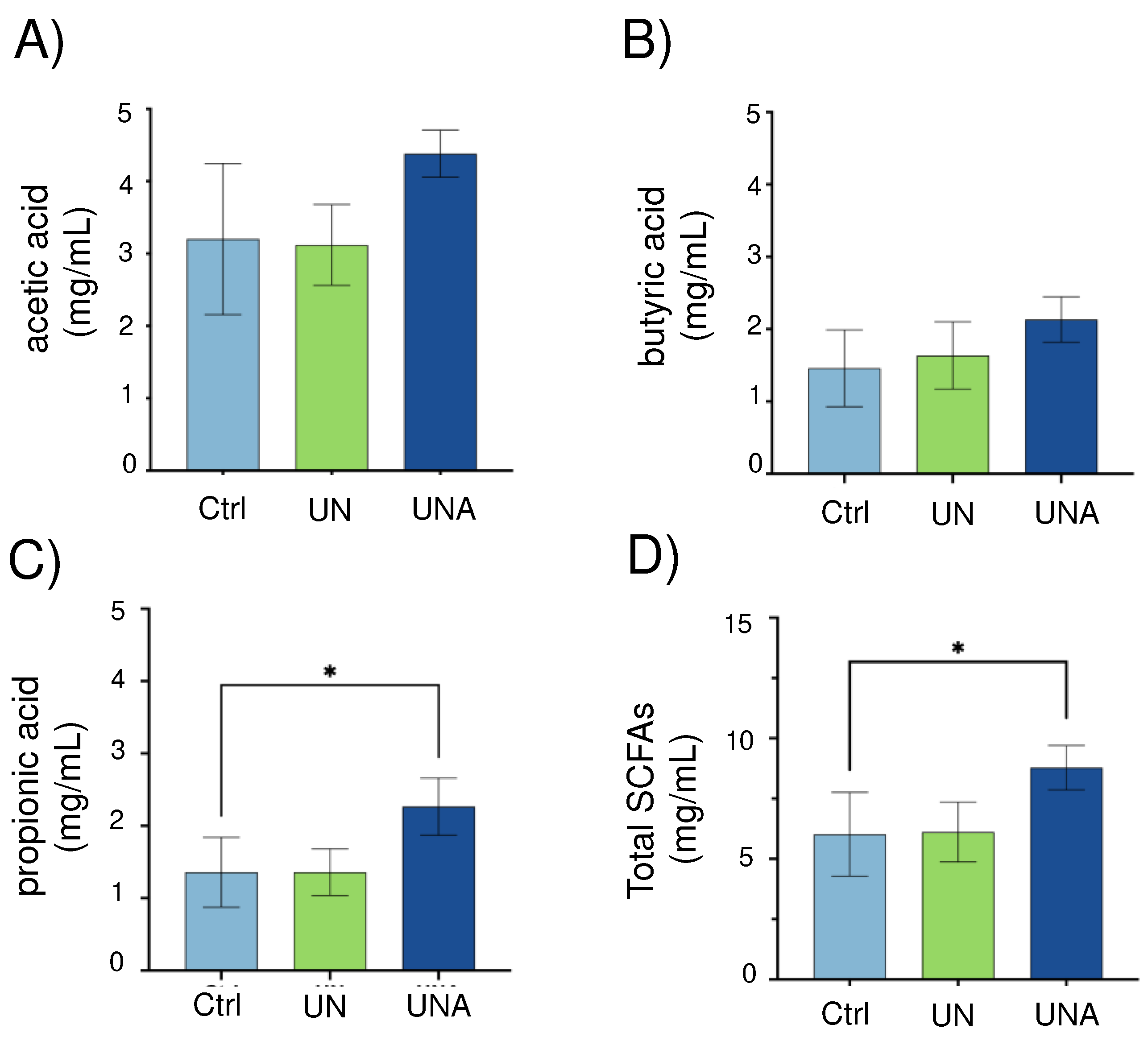
Figure 7.
Heatmap of predicted functions of the gut microbiome by Tax4fun2 evaluated the differences in the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway. The differences in KEGG pathway: A) undernutrition group (UN) vs control children (Ctrl); B) undernutrition group after amaranth consumption (UNA) vs undernutrition group at the beginning of assay (UN); C) undernutrition group before amaranth consumption (UN) vs control group (Ctrl).
Figure 7.
Heatmap of predicted functions of the gut microbiome by Tax4fun2 evaluated the differences in the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway. The differences in KEGG pathway: A) undernutrition group (UN) vs control children (Ctrl); B) undernutrition group after amaranth consumption (UNA) vs undernutrition group at the beginning of assay (UN); C) undernutrition group before amaranth consumption (UN) vs control group (Ctrl).
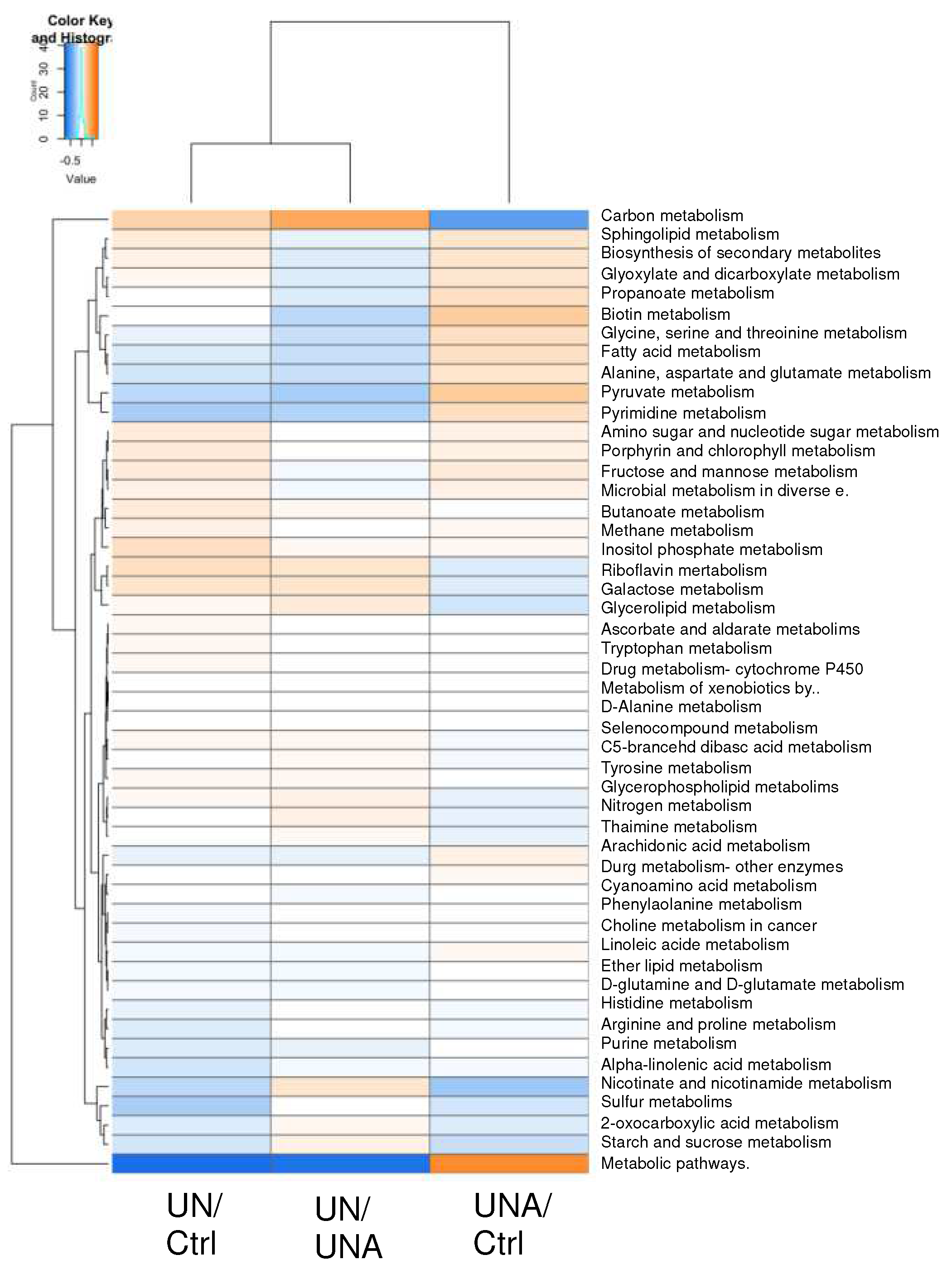
Figure 8.
Association between taxa and pathways. Taxa are represented by yellow circles and pathways by green squares. The network's nodes are taxa and pathways. The node correlation is represented by the vertex. Red and blue, respectively, are the vertex colors for positive and negative correlations.
Figure 8.
Association between taxa and pathways. Taxa are represented by yellow circles and pathways by green squares. The network's nodes are taxa and pathways. The node correlation is represented by the vertex. Red and blue, respectively, are the vertex colors for positive and negative correlations.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



