
Submitted:
20 June 2023
Posted:
20 June 2023
You are already at the latest version
Abstract
When oxidizing reduced sulfur compounds, the phototrophic sulfur bacterium Allochromatium vinosum forms spectacular sulfur globules as obligatory intracellular – but extracytoplasmic – intermediates. The globule envelope consists of three extremely hydrophobic proteins: SgpA and SgpB are very similar and can functionally replace each other. SgpC is involved in the expansion of the sulfur globules. The presence of a fourth protein, SgpD, has been suggested by comparative transcriptomics and proteomics of enriched sulfur globules. Here, we investigated the in vivo function of SgpD by coupling its carboxy-terminus to mCherry. This fluorescent protein requires oxygen for chromophore maturation, but we were able to use it in anaerobically growing A. vinosum provided the cells were exposed to oxygen for one hour prior to imaging. While mCherry lacking a signal peptide resulted in low fluorescence evenly distributed throughout the cell, fusion with SgpD carrying its original Sec-dependent signal peptide targeted mCherry to the periplasm and co-localized it exactly with the highly light-refractive sulfur deposits seen in sulfide-fed A. vinosum cells. Insertional inactivation of the sgpD gene showed that the protein is not essential for the formation and degradation of sulfur globules.

Keywords:
sulfur globules
; sulfur oxidation
; purple sulfur bacteria
; Allochroamtium vinosum
; fluorescence microscopy
1. Introduction
A large proportion of sulfur-oxidizing bacteria form conspicuous sulfur deposits as intermediates during the oxidation of sulfide, polysulfides or thiosulfate [1,2,3,4]. The sulfur globules are deposited either extracellularly or intracellularly [1,5,6]. The formation of extracellular sulfur globules is characteristic of green sulfur bacteria and sulfur oxidizers of the gammaproteobacterial family Ectothiorhodospiraceae, while intracellular sulfur globules are typical of magnetotactic sulfur oxidizers, purple sulfur bacteria of the family Chromatiaceae, and sulfur-oxidizing bacterial endosymbionts [1]. The general target compartment for intracellular sulfur storage is the bacterial periplasm where the sulfur globules are encased by a protein envelope consisting of one or more structural proteins similar to cytoskeletal keratins or plant cell wall proteins [1,7].
A prominent and well-studied example of intracellular sulfur deposition are the sulfur globules of the anoxygenic phototrophic purple sulfur bacterium Allochromatium vinosum (class Gammaproteobacteria, family Chromatiaceae). In A. vinosum, the sulfur globules reside in the periplasm, i.e. in the same cellular compartment as the periplasmic thiosulfate- and sulfide-oxidizing enzymes. This is evidenced by the presence of signal peptide coding sequences in the genes for the three structural proteins of the sulfur globule envelope that have been studied so far [5,8,9]. In A. vinosum, the so-called rDsr system with the reverse-acting dissimilatory sulfite reductase rDsrAB as the key enzyme is essential for further oxidation of sulfur in the cytoplasm [10,11,12].
The sulfur globules of the Chromatiaceae, including those of A. vinosum, reach up to 1 μm in diameter [5] and account for up to 34% of the total cell dry weight [13,14]. The three established A. vinosum sulfur globule proteins SgpA (Alvin_1905), SgpB (Alvin_0358) and SgpC (Alvin_1325) have in common that they are extremely hydrophobic. SgpA and SgpB are very similar in amino acid sequence and can functionally replace each other. SgpC participates in sulfur globule expansion [7,9]. In 2014, we re-evaluated the A. vinosum sulfur globule proteome and identified the protein Alvin_2515 as a new putative component, which we named SgpD [15]. It resides next to genes for an hydrophobic/amphiphilic exporter and a probable two-component transcriptional regulator. Transcript levels for the sgpD gene dramatically increase on sulfide and thiosulfate compared to growth on malate in the absence of oxidizable sulfur compounds (28-fold and 6-fold, respectively) [16]. SgpD is synthesized with a cleavable Sec-dependent N-terminal signal peptide predicted to mediate transport to the periplasm. The protein could have a coil-coil structure typical of structural proteins, such as bacterial cytoskeletal proteins [15]. However, it should be noted that a recent re-evaluation of coil-coil prediction tools revealed a high number of incorrect predictions, seriously questioning their informative value [17].
At present, direct experimental evidence for a function of SgpD as a sulfur globule protein is lacking. This would require definitive information on the intracellular localization of the protein and its possible association with sulfur deposits in vivo. Fluorescent reporter proteins, such as green fluorescent protein (GFP), are valuable non-invasive molecular tools for real-time in vivo imaging of living specimens and have the greatest potential to address these questions. One limitation of fluorescent proteins in pigmented phototrophic bacteria is signal quenching when they emit light at a wavelength absorbed by the pigments, as discussed for the anoxygenic phototrophic Alphaprotebacteria Rhodobacter capsulatus and Rhodopseudomonas palustris [18,19]. GFP is an example of a fluorescent protein that is incompatible with bacteriochlorophyll a and carotenoids, the pigments that drive photosynthesis not only in R. capsulatus and R. palustris but also in A. vinosum. In addition, commonly used variants of GFP are not suited for investigating the subcellular localization of periplasmic proteins. When exported to the periplasm in an unfolded conformation through the Sec system, they fail to fold properly and do not fluoresce [20]. However, very good alternatives to GFP, such as mCherry, mStrawberry, and tdTomato, derived from screening serial mutants and genetic modification of the original GFP [21,22,23], are now available and widely used for gene expression measurement, protein localization, in situ screening, and multi-omic profiling. Among them, the fluorescent protein mCherry stands out because of its bright signal, rapid maturation, high photostability, high N-terminal fusion tolerance, and excellent pH tolerance [24]. Importantly, red fluorescent protein derivatives such as mCherry do not share the transport difficulties of their GFP relatives and can be effectively transported through the Sec system [25,26].
Unfortunately, despite all its advantages, mCherry shares with almost all fluorescent proteins a strict requirement for molecular oxygen for the maturation of the fluorophore [27], making it inherently difficult to use under anaerobic conditions, such as during the phototrophic growth of A. vinosum. While there is no general limitation to aerobic systems, there is still not much known about the exact conditions that allow full maturation of the fluorophore after exposure of anaerobically grown cells to oxygen [28,29,30]. Flavin-based fluorescent protein presents an alternative, but all available derivatives have two major limitations. They produce only cyan-green fluorescence incompatible with the A. vinosum pigments and the fluorescence emitted is significantly dimmer compared to GFP [31] or newer anaerobic fluorescent reporters such as Fluorescence-Activating Absorption-Shifting Tag, FAST [32]. The latter has so far been used primarily in eukaryotic systems and, when used in bacteria, is best suited for secretion studies [31] and has only been applied under a limited number of conditions, e.g. in E. coli during nitrate or fumarate respiration [31,33]. SNAP-tag and Halo-Tag are further promising reporters for fluorescent labeling in the absence of oxygen, but so far have only been adapted to few anaerobic bacteria, i.e. Clostridium species [34], which belong to the phylum Bacillota, and Bacteroides thetaiotaomicron (phylum Bacteroidota) [35].
After assessment of the available methods, we chose mCherry coupled to aerobic fluorescence recovery of the anaerobically produced protein as the most promising method for in situ fluorescence labeling and protein localization in the anoxygenic phototroph A. vinosum. Using mCherry we localized SgpD to the sulfur globules of the purple sulfur bacterium and collected evidence that its presence is not essential for sulfur globule formation.
2. Materials and Methods
2.1. Bacterial Strains, Plasmids, PCR Primers and Growth Conditions
The bacterial strains, plasmids and primers used in this study are listed in Table S1. All fluorescence experiments were carried out in a ΔsgpD strain derived from a spontaneous rifampicin-resistant mutant of the sequenced A. vinosum DSM 180T. A. vinosum was cultivated photoorganoheterotrophically in RCV medium [36] or photolithoautotrophically in Pfennig medium [37] without reduced sulfur compound, referred to as “0 medium”. Sulfide or thiosulfate were added as electron sources at the desired concentration. The cultivation was performed under anoxic conditions and continuous illumination at 30°C in completely filled screw-capped culture bottles or on agar plates. Escherichia coli was cultivated in Luria Bertani medium [38]. Antibiotics used for mutant selection were applied at the following concentrations (in µg ml-1): for E. coli: ampicillin 100, kanamycin 50, gentamycin 25, for A. vinosum: rifampicin 50, ampicillin 10, kanamycin 10, gentamycin 2.
2.2. Recombinant DNA Techniques
Standard methods were used for molecular biological techniques. Chromosomal DNA of A. vinosum strains was obtained by a modified sarcosyl lysis [39]. The genotypes of the A. vinosum recombinants used in this study were confirmed by Southern hybridization or PCR. Southern hybridization was performed over night at 68°C. PCR amplifications with Taq DNA polymerase and Pfu DNA polymerase were done essentially as described previously [40]. DNA probes for Southern hybridization were digoxygenin labelled by PCR. Plasmid DNA from E. coli was purified using the GenJetTM Plasmid Miniprerp Kit (Thermo Scientific).
2.3. Construction of Plasmids for Production of mCherry in A. vinosum
We wanted to clone mCherry under the strong A. vinosum dsr promoter and used the replicative plasmid pBBR1MCS2-L [41] as the basis. This plasmid contains a XbaI-HindIII fragment of the PCR-amplified dsr promoter fused to gene dsrL in XbaI-HindIII of pBBR2MCS-2. The plasmid was first supplied with a gentamycin resistance cartridge which had been amplified from pBRRMCS-5 with primers Gent-CpoI-Fw and Gent-CpoI-rev (Table S1). The amplicon was digested with RsrII and inserted into the RsrII site present within the kanamycin resistance cartridge of pBBRMCS-2, resulting in plasmid pBBR1-L-Gm. The mCherry gene was amplified via PCR with primers for-mCherry-NdeI and rev-mCherry-SalI with plasmid pmCherry [24] as the template. After digestion with NdeI/SalI, the amplicon was cloned between the SalI and NdeI sites of pBBR1-L-Gm, giving plasmid pBBR_dsr_mCherry_Gm that carries the gene for mCherry under the control of the dsr promoter. Fusion of mCherry with the potential signal peptide for SgpD and its first amino acid (tryptophan) a s well as a methionine was achieved by cloning a PCR product generated with primers for-mCher-sig and rev-mCherry-SalI with plasmid pmCherry [24] as the template between NdeI and SalI of pBBR1-L-Gm. The complete sgpD gene was amplified with primers fAlvin2515-NdeI and rAlvin2515-NdeI cloned into the NdeI site of pBBR_dsr_mCherry_Gm, resulting in a fusion of SgpD including its signal peptide and mCherry.
2.4. Fluoresence Microscopy
For microscopy, cells were transferred to a microscopy slide coated with a thin film of 1% (w/vol) agarose. Fluorescence microscopy was carried out at room temperature using a Zeiss Axio Observer Z1 microscope (Zeiss, Jena, Germany) equipped with an HXP 120 V light source, an αPlan-APOCHROMAT 100×/1.46 oil immersion objective and an Axio Cam MR3 camera. Visualization of mCherry was achieved using Carl Zeiss filter set 64 HE (574–599 nm excitation, 605 nm beam splitter and 612-682 nm emission). Image acquisition and analysis were performed with Zen 2 software (Zeiss).
2.5. Construction of A. vinosum ΔsgpD::ΩKm
For the substitution of sgpD (Alvin_2515) in the genome of A. vinosum by an kanamycin cassette, SOE PCR [42] fragments were constructed using primer pairs Del-Alvin2515-fw1/Del-Alvin2515-rev1 and Del-Alvin2515-fw2/Del-Alvin2515-rev2 (Table S1). The resulting fragment was inserted into the mobilizable plasmid pSUP301 by HindIII restriction sites resulting in plasmid pSUP301_ΔsgpD. After digestion with EcoRI, the kanamycin cassette from pHP45ΩKm was ligated into the EcoRI site of pSUP301_ΔsgpD. The final mobilizable construct pSUP301_ΔsgpD_ ΩKm was transferred from E. coli S17.1 to A. vinosum Rif50 by conjugation [43]. Transconjugants were selected on RCV plates containing the appropriate antibiotics under anoxic conditions in the light. Double cross-over recombinants lost the vector encoded ampicillin resistance.
2.6. Characterization of Phenotypes, Detection of Sulfur Species and Protein Determination
A. vinosum wild-type and the ΔsgpD_ΩKm strains were characterized in batch culture experiments essentially as described before [16]. Cells of A. vinosum, grown photoorganoheterotrophically on malate (RCV medium [36]) for three days were used as an inoculum for experiments concerned with transformation of sulfide and thiosulfate. The culture volume of the precultures was 500 ml. Inoculum cells were harvested by centrifugation (10 min, 2680 x g) and washed once in “0” medium. The culture volume for phenotypic characterization was 250 ml. In these experiments, the starting optical density at 690 nm was set to 0.8 to 0.9. Sulfide, polysulfides, elemental sulfur and sulfate were either determined by HPLC [44] or using classical colorimetric or turbidometric methods as previously described [16,40].
3. Results
3.1. Suitability of mCherry for the Purple Sulfur Bacterium Allochromatium Vinosum
Here, we describe the production and validation of a direct fluorescence reporter for cell biological studies in an anoxygenic purple sulfur bacterium of the family Chromatiaceae (class Gammaproteobacteria). First, we show that intrinsic background fluorescence in the applied wavelength range is negligible (Figure 1A, middle panel). The next step was to determine whether mCherry is functional in A. vinosum, i.e. correctly expressed, translationally modified and folded. For this purpose, a reporter plasmid (pBBR_dsr_mCherry_Gm) was constructed in which the strong A. vinosum dsr promoter, which is inducible by sulfide, thiosulfate and elemental sulfur [16,41], was fused to the gene for the red fluorescent protein, mCherry [24,45], directed to the cytoplasm. The plasmid was transferred into A. vinosum via conjugation and the cells were then grown photolithoautotrophically in the light with 4 mM sulfide as electron donor. Figure 1B shows that the cells were fully and homogeneously fluorescent after 70 to 80 min of exposure to air. Our results are fully consistent with experiments in the anoxygenic phototroph R. palustris, in which mCherry also obtained fluorescence when switched from anoxic to oxic conditions [19]. The spectroscopic properties of mCherry are the same irrespective of whether it is originally produced under aerobic or anaerobic conditions [19].
3.2. Fusions with mCherry Reveals Intracellular Localization of SgpD in A. vinosum
The putative sulfur globule protein SgpD is equipped with a probable Sec-dependent signal peptide proposed to direct its transport across the cytoplasmic membrane into the periplasm of the Gram-negative bacterium A. vinosum [15]. The predicted periplasmic localization of SgpD would have been a further difficulty when using the classical GFP chromophore which is known not to be correctly assembled in the bacterial periplasm [20]. However, this is not the case for mCherry and accordingly brightly fluorescing cells were obtained when mCherry was fused to the signal peptide of SgpD (Figure 1C). A. vinosum is packed with intracytoplasmic membranes that are organized in vesicles (the so-called “chromatophores”). The interior of these chromatophores is extracytoplasmic, i.e. the entire cell body and not only its outermost layer as in other Gram-negative bacteria such as E. coli is filled with periplasmic space [8]. This fully explains the homogeneous distribution of mCherry when exported via the signal peptide (Figure 1C).
In the next step, mCherry was fused to full-length SgpD including its signal peptide. Now, fluorescence was no longer evenly distributed in the cells but concentrated to circular structures inside of the cells (Figure 1C, middle panel). Overlays of the fluorescence micrographs with images obtained by phase contrast microscopy showed that the circular structures exactly matched the position of sulfur globules in the cells and thus demonstrated that SgpD is attached to sulfur globules in vivo. Further evidence for this conclusion was obtained by microscopy of sulfur globules isolated from the A. vinosum strain with the SgpC-mCherry fusion (Figure 2). The sulfur globules were brightly fluorescent due to attachment of mCherry via SgpD.
3.3. Stability of mCherry Fluorescence in Allochromatium Vinosum
We sought to obtain more information about the development and stability of mCherry fluorescence in the purple sulfur bacterium and followed it via fluoresce microscopy over a period of 95 min. The fluorescence intensity gradually increased with oxygen exposure time and reached the maximum after a 60-min-exposure at room temperature. This maximum fluorescence was stably maintained for at least half an hour.
Figure 3.
Development of fluorescence over time in A. vinosum Rif50 carrying pBBR_dsr_SgpD_mCherry. Left, middle and right panels show phase contrast microscopy, fluorescence microscopy and merged phase contrast and fluorescence images, respectively. Excitation times for phase contrast and for the mCherry channel (587 nm) were 49.2 s and 1 s, respectively. Size bar: 2 µm.
Figure 3.
Development of fluorescence over time in A. vinosum Rif50 carrying pBBR_dsr_SgpD_mCherry. Left, middle and right panels show phase contrast microscopy, fluorescence microscopy and merged phase contrast and fluorescence images, respectively. Excitation times for phase contrast and for the mCherry channel (587 nm) were 49.2 s and 1 s, respectively. Size bar: 2 µm.

3.2. In Vivo Role of SgpD
The analysis of the A. vinosum sulfur globule proteome revealed SgpB as the second most abundant sulfur globule protein in this organism while SgpA and SgpC were less frequently detected [15]. The most abundant A. vinosum sulfur globule protein is SgpD (Weissgerber et al. 2014). The presence of a protein envelope around the sulfur inclusions in sulfur-oxidizing bacteria suggests an important structure–function relationship. Indeed, mutants of A. vinosum lacking SgpB and SgpC are no longer able to oxidize sulfide and thiosulfate and to form sulfur inclusions (Prange et al. 2004). SgpA and SgpB can replace each other in the presence of SgpC. SgpC probably plays a role in sulfur globule expansion. Information about the in vivo role of SgpD has so far not been available and we therefore set out to close this knowledge gap.
First, we re-evaluated its distribution within the bacteria and found that it is almost exclusively restricted to purple sulfur bacteria of the family Chromatiaceae with a threshold of e=10-05 in Protein Blast searches. The only exceptions to this rule are a Gammaproteobacteria bacterium and a Chromatiales bacterium as well as the bacterial endosymbiont of Escarpia laminata, all of which cannot be more strictly taxonomically grouped, but could also be members of the Chromatiaceae.
With the other three sulfur globule proteins SgpA, SgpB and SgpC, A. vinosum SgpD shares a somewhat repetitive amino acid sequence with regularly spaced proline residues exemplified by the pattern P-X2-P-X2-P-X4-P-X2-P-X2-P-X2-P-X5-P-X2-PX2-P-X2-X5P-X2P in the central part of the protein. Scanning for repetitive sequences by RADAR [46] revealed four repeats (Figure 4).
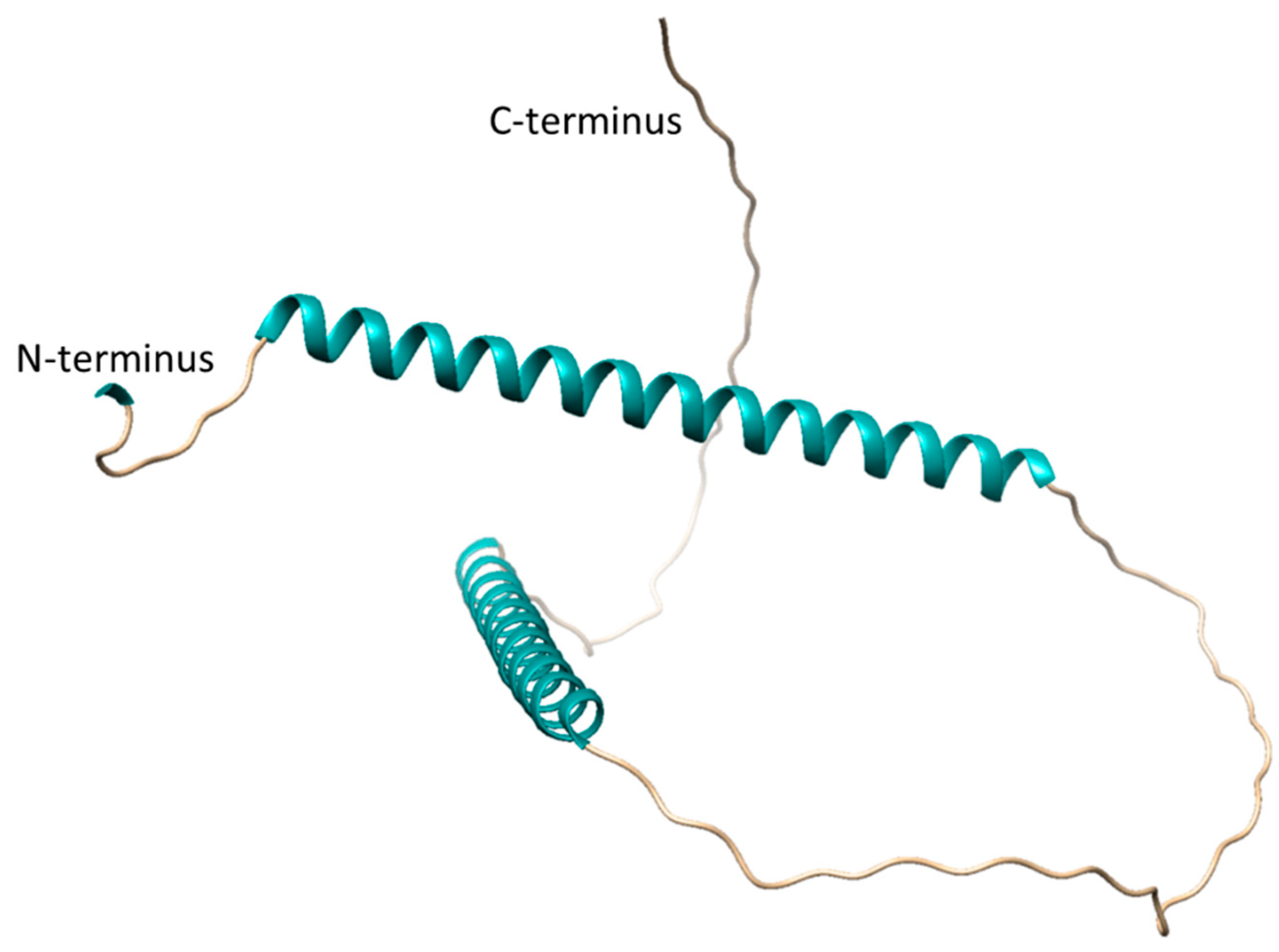
An AlphaFold prediction for SgpD is available at https://alphafold.ebi.ac.uk/entry/D3RP35 and shown in Figure 5. Large predicted loop portions are modelled with low confidence (70 > pLDDT > 50). Two alpha-helices (amino acids 13 to 58 and 126 to 142) are predicted with very high confidence (pLDDT > 90). The first helix consists of the repeats 1 and 2 shown in Figure 4, while the second helix is made up of the residues in repeat 3. Clearly, experimental data is needed to gain further insight into the structural features of the proteins that make up the envelope of the sulfur globules.
Figure 5.
Alphafold [47] prediction of A. vinosum SgpD. The signal peptide (amino acids 1-22) that is cleaved off after transport to the cytoplasm is hidden here.
Figure 5.
Alphafold [47] prediction of A. vinosum SgpD. The signal peptide (amino acids 1-22) that is cleaved off after transport to the cytoplasm is hidden here.
In another approach to obtain information on the in vivo function of SgpD, we generated an A. vinosum ΔsgpD strain in which the gene was inactivated by insertion of a kanamycin Ω interposon [48]. The nucleotide sequence between the ATG and the TAA stop codons of sgpD was completely replaced by a kanamycin Ω interposon. Just as the other sgp genes, sgpD is preceded by a promoter sequence predicted by BPROM [49] (TTGATA-N13-TTATATACT, ending 79 nucleotides upstream of the ATG start codon). An inverted repeat (CGGCCCCATCGACGATGGGGCCG) corresponding to the features of rho-independent transcription terminators [50] is found 45 nt downstream of the TAA stop codon. Thus, sgpD appears to form a separate transcription unit. Downstream polar effects of the Ω cassette should therefore be minimal. It should also be noted that the gene Alvin_2516, which follows sgpD at a distance of 216 nucleotides in the same direction of transcription, is under a completely different transcriptional control, as its transcript abundance is not affected by the presence of sulfide or thiosulfate in the medium [16].
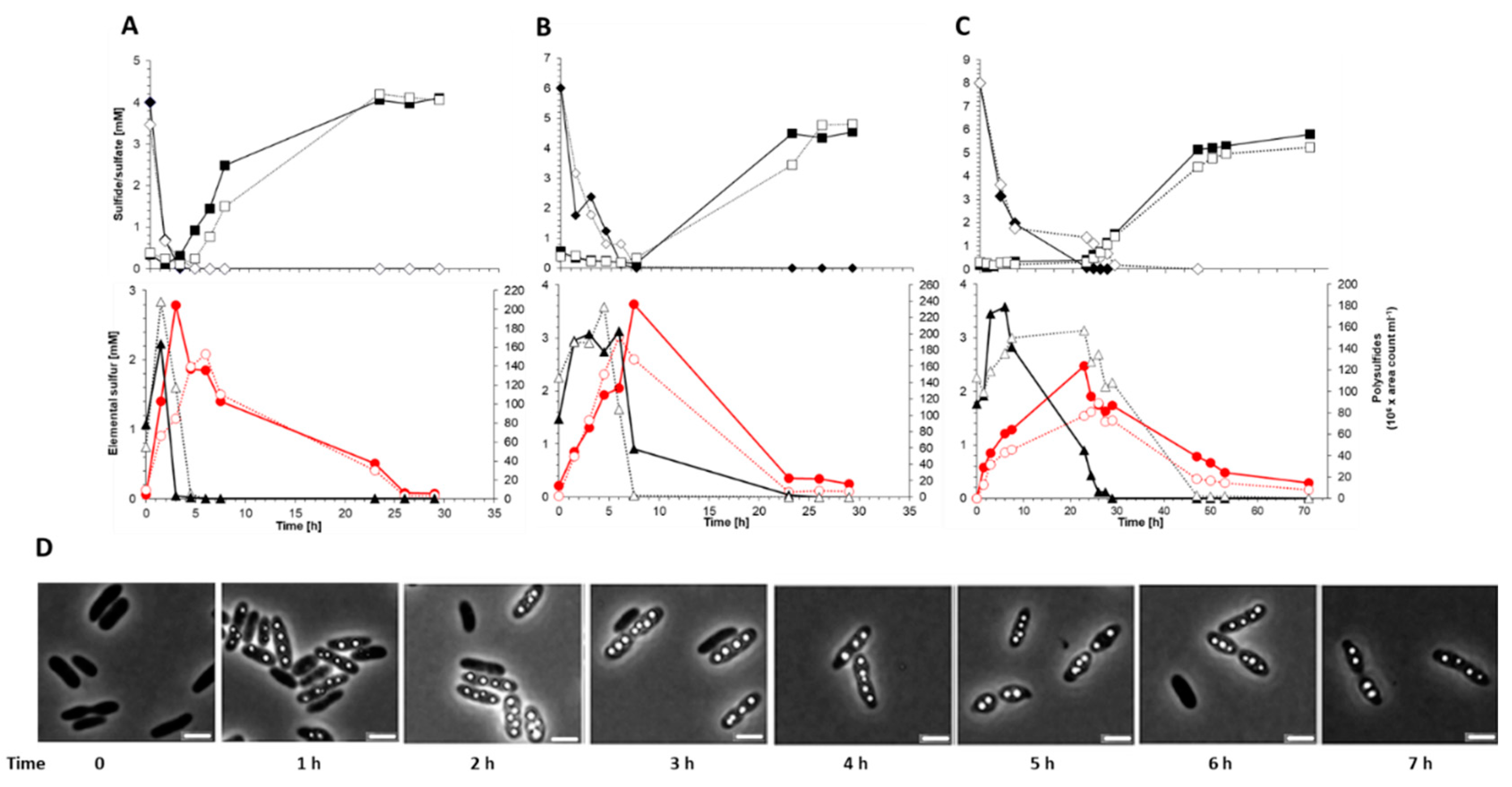
Photoorganoheterotrophic growth of the sgpD-deficient mutant strain on malate-contianing medium with sulfate as the sole sulfur source was unaffected, thus excluding general growth defects. In addition, the ability of the mutant to grow photolithoautotrophically on sulfide-containing media and to store sulfur as intracellular sulfur globules was not affected (Figure 5). Similar to the wildtype, the mutant oxidized sulfide first to polysulfide and then to “zero-valent” polysulfane sulfur stored in sulfur globules, which was finally oxidized to sulfate (Figure 5A–C). The pattern of sulfur compounds in the growth experiments remained essentially the same regardless of the initial sulfide concentration, 2, 4 or 8 mM (Figure 5A–C,). Lack of SgpD neither influenced the size nor the quantity of sulfur globules (Figure 5D). This result is similar to findings for SgpB and SgpA [5,9].
Figure 5.
Consumption of 4 mM (A), 6 mM (B) and (8 mM) sulfide by A. vinosum wildtype (filled symbols) compared to A. vinosum ΔsgpD (open symbols). Representative experiments are shown. The upper panels give sulfide (diamonds) and sulfate concentrations in culture supernatants. The lower panels show formation and degradation of the combination of the two main polysulfides (triangles) and of intracellular sulfur (red circles). Part D shows phase contrast micrographs of A. vinosum ΔsgpD at the indicated time points after addition of 4 mM sulfide.
Figure 5.
Consumption of 4 mM (A), 6 mM (B) and (8 mM) sulfide by A. vinosum wildtype (filled symbols) compared to A. vinosum ΔsgpD (open symbols). Representative experiments are shown. The upper panels give sulfide (diamonds) and sulfate concentrations in culture supernatants. The lower panels show formation and degradation of the combination of the two main polysulfides (triangles) and of intracellular sulfur (red circles). Part D shows phase contrast micrographs of A. vinosum ΔsgpD at the indicated time points after addition of 4 mM sulfide.
4. Discussion
The anoxygenic phototrophic bacterium A. vinosum is widely distributed in sulfide-rich, light-penetrated environments and serves as one of the very few genetically accessible model organisms of the purple sulfur bacteria [43,51,52]. Here, we have expanded the genetic toolbox for A. vinosum by establishing a basic cell biological method that now allows protein localization in the living cell using a fluorescent protein reporter, mCherry. We show that mCherry produced in A. vinosum, originally grown in the absence of oxygen under photolithoautotrophic conditions, is fully fluorescent within one hour of exposure of the cells to air. Interference from intrinsic background fluorescence is negligible. When exported to the periplasm, mCherry also develops full fluorescence and specifically accumulates at the cellular target of the protein to which it is fused. In the experiments reported here, mCherry linked to a potential protein of the proteinaceous sulfur globule envelope did indeed appear exclusively in immediate vicinity of the intracellular sulfur deposits. To our knowledge, tagging with fluorescent proteins has so far not been established for any purple sulfur bacterium and we hope that the availability of the technique for A. vinosum will promote studies of the basic cell biology not only of this but also of other bacteria with the same physiology.
In the light, A. vinosum forms sulfur globules only under anaerobic conditions [8], while the red fluorescence protein mCherry needs molecular oxygen for the maturation of its chromophore. As pointed out by Jiang and coworkers [19], oxygen access into the fully folded barrel protein is of great importance, but may also trigger irreversible photobleaching and reduced photostability of a fluorescent protein [53]. In our experiments, the fluorescence signal of A. vinosum was fully recovered after exposure to oxygen for 60 min and was stably maintained at the maximum fluorescence intensity for at least 30 min. For comparison, fluorescence signals in the purple non-sulfur bacterium R. palustris required four hours oxygen exposure for full recovery [19]. In any case, our results, as well as those obtained by others with R. palustris, show that mCherry produced under anaerobic conditions matures and develops fluorescence in the presence of air.
Coupling mCherry to SgpD allowed us to show that SgpD is tightly bound to sulfur globules and represents a novel component of their proteinaceous envelope, although it is neither essential for growth on sulfide nor for sulfur globule formation during its oxidation. Organisms such as A. vinosum, which live in sulfide-rich and oxygen-poor environments, appear to be equipped with an at least partially redundant set of proteins [1,9] that ensure the compatibility of massive sulfur deposition within the cell boundary with energy metabolism and the formation of new biomass. Having a variety of sulfur globule proteins that can compensate for each other to some extent may allow for fine-tuned adaptation to environmental conditions. Possession of SgpD may be advantageous at either low or very high sulfide concentrations that do not allow photolithotrophic growth of pure batch cultures due to insufficient electron supply or sulfide toxicity, respectively, but may be necessary or tolerated in an environment where other members of the community compete for the substrate or remove it rapidly enough to minimize toxic effects. Without question, a lot of further work is needed to elucidate the interaction of all sulfur globule proteins and to gain insights into the structure of these exciting proteins.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org., Table S1: Bacterial strains, primers and plasmids used in this study.
Author Contributions
Conceptualization, C.D. and F. G.; validation, C.D. and F.G.; investigation, C. K..; writing—original draft preparation, C. D.; writing—review and editing, C.D., C. K., and F.G.; supervision, C.D.; project administration, C.D.; funding acquisition, C.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Deutsche Forschungsgemeinschaft, grant number Da 351/6-1 (to CD).
Data Availability Statement
Data are contained within the article and its supplementary material.
Acknowledgments
We thank Jan Wruck and Thomas Weissgerber for help with mutant strain generation and growth experiments.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Dahl, C. Bacterial intracellular sulfur globules. In Bacterial organelles and organelle-like inclusions; Jendrossek, D., Ed.; Springer: Cham, 2020; pp. 19–51. [Google Scholar]
- Marnocha, C.L.; Levy, A.T.; Powell, D.H.; Hanson, T.E.; Chan, C.S. Mechanisms of extracellular S0 globule production and degradation in Chlorobaculum tepidum via dynamic cell-globule interactions. Microbiology 2016, 162, 1125–1134. [Google Scholar] [CrossRef] [PubMed]
- Marnocha, C.L.; Sabanayagam, C.R.; Modla, S.; Powell, D.H.; Henri, P.A.; Steele, A.S.; Hanson, T.E.; Webb, S.M.; Chan, C.S. Insights into the mineralogy and surface chemistry of extracellular biogenic S0 globules produced by Chlorobaculum tepidum. Front. Microbiol. 2019, 10, 271. [Google Scholar] [CrossRef]
- Dahl, C. A biochemical view on the biological sulfur cycle. In Environmental technologies to treat sulfur pollution: principles and engineering, 2 ed; Lens, P., Ed.; IWA Publishing: London, 2020; pp. 55–96. [Google Scholar]
- Pattaragulwanit, K.; Brune, D.C.; Trüper, H.G.; Dahl, C. Molecular genetic evidence for extracytoplasmic localization of sulfur globules in Chromatium vinosum. Arch. Microbiol. 1998, 169, 434–444. [Google Scholar] [CrossRef]
- Dahl, C.; Prange, A. Bacterial sulfur globules: occurrence, structure and metabolism. In Inclusions in prokaryotes; Shively, J. M., Ed.; Springer: Berlin Heidelberg, 2006; Vol. 1, pp. 21–51. [Google Scholar]
- Brune, D.C. Isolation and characterization of sulfur globule proteins from Chromatium vinosum and Thiocapsa roseopersicina. Arch. Microbiol. 1995, 163, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Frigaard, N.U.; Dahl, C. Sulfur metabolism in phototrophic sulfur bacteria. Adv. Microb. Physiol. 2009, 54, 103–200. [Google Scholar] [PubMed]
- Prange, A.; Engelhardt, H.; Trüper, H.G.; Dahl, C. The role of the sulfur globule proteins of Allochromatium vinosum: mutagenesis of the sulfur globule protein genes and expression studies by real-time RT PCR. Arch. Microbiol. 2004, 182, 165–174. [Google Scholar] [CrossRef]
- Pott, A.S.; Dahl, C. Sirohaem-sulfite reductase and other proteins encoded in the dsr locus of Chromatium vinosum are involved in the oxidation of intracellular sulfur. Microbiology 1998, 144, 1881–1894. [Google Scholar] [CrossRef]
- Dahl, C.; Engels, S.; Pott-Sperling, A.S.; Schulte, A.; Sander, J.; Lübbe, Y.; Deuster, O.; Brune, D.C. Novel genes of the dsr gene cluster and evidence for close interaction of Dsr proteins during sulfur oxidation in the phototrophic sulfur bacterium Allochromatium vinosum. J. Bacteriol. 2005, 187, 1392–1404. [Google Scholar] [CrossRef]
- Sander, J.; Engels-Schwarzlose, S.; Dahl, C. Importance of the DsrMKJOP complex for sulfur oxidation in Allochromatium vinosum and phylogenetic analysis of related complexes in other prokaryotes. Arch. Microbiol. 2006, 186, 357–366. [Google Scholar] [CrossRef]
- Overmann, J. Mahoney Lake: A case study of the ecological significance of phototrophic sulfur bacteria. Adv. Microb. Ecol. 1997, 15, 251–288. [Google Scholar]
- van Gemerden, H. Growth measurements of Chromatium cultures. Arch. Mikrobiol. 1968, 64, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Weissgerber, T.; Sylvester, M.; Kröninger, L.; Dahl, C. A comparative quantitative proteome study identifies new proteins relevant for sulfur oxidation in the purple sulfur bacterium Allochromatium vinosum. Appl. Environ. Microbiol. 2014, 80, 2279–2292. [Google Scholar] [CrossRef] [PubMed]
- Weissgerber, T.; Dobler, N.; Polen, T.; Latus, J.; Stockdreher, Y.; Dahl, C. Genome-wide transcriptional profiling of the purple sulfur bacterium Allochromatium vinosum DSM 180T during growth on different reduced sulfur compounds. J. Bacteriol. 2013, 195, 4231–4245. [Google Scholar] [CrossRef] [PubMed]
- Simm, D.; Hatje, K.; Waack, S.; Kollmar, M. Critical assessment of coiled-coil predictions based on protein structure data. Sci. Rep. 2021, 11, 12439. [Google Scholar] [CrossRef]
- Fogg, P.C.; Westbye, A.B.; Beatty, J.T. One for all or all for one: heterogeneous expression and host cell lysis are key to gene transfer agent activity in Rhodobacter capsulatus. PLoS One 2012, 7, e43772. [Google Scholar] [CrossRef]
- Jiang, M.; Zeng, Y.; Cui, L.; Wang, M.; Zheng, Y. A red fluorescent protein reporter system developed for measuring gene expression in photosynthetic bacteria under anaerobic conditions. Microorganisms 2022, 10. [Google Scholar] [CrossRef]
- Feilmeier, B.J.; Iseminger, G.; Schroeder, D.; Webber, H.; Phillips, G.J. Green fluorescent protein functions as a reporter for protein localization in Escherichia coli. J. Bacteriol. 2000, 182, 4068–4076. [Google Scholar] [CrossRef]
- Cormack, B.P.; Valdivia, R.H.; Falkow, S. FACS-optimized mutants of the green fluorescent protein (GFP). Gene 1996, 173, 33–38. [Google Scholar] [CrossRef]
- Elsliger, M.A.; Wachter, R.M.; Hanson, G.T.; Kallio, K.; Remington, S.J. Structural and spectral response of green fluorescent protein variants to changes in pH. Biochemistry 1999, 38, 5296–5301. [Google Scholar] [CrossRef]
- Patterson, G.H.; Knobel, S.M.; Sharif, W.D.; Kain, S.R.; Piston, D.W. Use of the green fluorescent protein and its mutants in quantitative fluorescence microscopy. Biophys. J. 1997, 73, 2782–2790. [Google Scholar] [CrossRef]
- Shaner, N.C.; Campbell, R.E.; Steinbach, P.A.; Giepmans, B.N.; Palmer, A.E.; Tsien, R.Y. Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nat. Biotechnol. 2004, 22, 1567–1572. [Google Scholar] [CrossRef]
- Chen, J.C.; Viollier, P.H.; Shapiro, L. A membrane metalloprotease participates in the sequential degradation of a Caulobacter polarity determinant. Mol. Microbiol. 2005, 55, 1085–1103. [Google Scholar] [CrossRef]
- Lewenza, S.; Vidal-Ingigliardi, D.; Pugsley, A.P. Direct visualization of red fluorescent lipoproteins indicates conservation of the membrane sorting rules in the family Enterobacteriaceae. J. Bacteriol. 2006, 188, 3516–3524. [Google Scholar] [CrossRef]
- Carroll, P.; Schreuder, L.J.; Muwanguzi-Karugaba, J.; Wiles, S.; Robertson, B.D.; Ripoll, J.; Ward, T.H.; Bancroft, G.J.; Schaible, U.E.; Parish, T. Sensitive detection of gene expression in mycobacteria under replicating and non-replicating conditions using optimized far-red reporters. PLoS One 2010, 5, e9823. [Google Scholar] [CrossRef]
- Heim, R.; Prasher, D.C.; Tsien, R.Y. Wavelength mutations and posttranslational autoxidation of green fluorescent protein. Proc. Natl. Acad. Sci. USA 1994, 91, 12501–12504. [Google Scholar] [CrossRef]
- Zhang, C.; Xing, X.H.; Lou, K. Rapid detection of a gfp-marked Enterobacter aerogenes under anaerobic conditions by aerobic fluorescence recovery. FEMS Microbiol. Lett. 2005, 249, 211–218. [Google Scholar] [CrossRef]
- Ransom, E.M.; Ellermeier, C.D.; Weiss, D.S. Use of mCherry Red fluorescent protein for studies of protein localization and gene expression in Clostridium difficile. Appl. Environ. Microbiol. 2015, 81, 1652–1660. [Google Scholar] [CrossRef] [PubMed]
- Streett, H.; Charubin, K.; Papoutsakis, E.T. Anaerobic fluorescent reporters for cell identification, microbial cell biology and high-throughput screening of microbiota and genomic libraries. Curr. Opin. Biotechnol. 2021, 71, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Plamont, M.A.; Billon-Denis, E.; Maurin, S.; Gauron, C.; Pimenta, F.M.; Specht, C.G.; Shi, J.; Querard, J.; Pan, B.; Rossignol, J.; Moncoq, K.; Morellet, N.; Volovitch, M.; Lescop, E.; Chen, Y.; Triller, A.; Vriz, S.; Le Saux, T.; Jullien, L.; Gautier, A. Small fluorescence-activating and absorption-shifting tag for tunable protein imaging in vivo. Proc. Natl. Acad. Sci. USA 2016, 113, 497–502. [Google Scholar] [CrossRef] [PubMed]
- Hinz, A.J.; Stenzler, B.; Poulain, A.J. Golden gate assembly of aerobic and anaerobic microbial bioreporters. Appl. Environ. Microbiol. 2022, 88, e0148521. [Google Scholar] [CrossRef]
- Charubin, K.; Streett, H.; Papoutsakis, E.T. Development of strong anaerobic fluorescent reporters for Clostridium acetobutylicum and Clostridium ljungdahlii using HaloTag and SNAP-tag proteins. Appl. Environ. Microbiol. 2020, 86. [Google Scholar] [CrossRef] [PubMed]
- Karunatilaka, K.S.; Cameron, E.A.; Martens, E.C.; Koropatkin, N.M.; Biteen, J.S. Superresolution imaging captures carbohydrate utilization dynamics in human gut symbionts. mBio 2014, 5, e02172. [Google Scholar] [CrossRef] [PubMed]
- Weaver, P.F.; Wall, J.D.; Gest, H. Characterization of Rhodopseudomonas capsulata. Arch. Microbiol. 1975, 105, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Pfennig, N.; Trüper, H.G. The family Chromatiaceae. In The Prokaryotes. A handbook on the biology of bacteria: ecophysiology, isolation, identification, applications; Balows, A., Trüper, H. G., Dworkin, M., Harder, W., Schleifer, K. H., Eds.; Springer-Verlag: New York, 1992; Vol. 2, pp. 3200–3221. [Google Scholar]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular cloning: a laboratory manual; Cold Spring Harbor Laboratory: Cold Spring Harbor, N.Y., 1989. [Google Scholar]
- Bazaral, M.; Helinski, D.R. Circular DNA forms of colicinogenic factors E1, E2 and E3 from Escherichia coli J. Mol. Biol. 1968, 36, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Dahl, C. Insertional gene inactivation in a phototrophic sulphur bacterium: APS-reductase-deficient mutants of Chromatium vinosum. Microbiology 1996, 142, 3363–3372. [Google Scholar] [CrossRef] [PubMed]
- Lübbe, Y.J.; Youn, H.S.; Timkovich, R.; Dahl, C. Siro(haem)amide in Allochromatium vinosum and relevance of DsrL and DsrN, a homolog of cobyrinic acid a,c diamide synthase for sulphur oxidation. FEMS Microbiol. Lett. 2006, 261, 194–202. [Google Scholar] [CrossRef]
- Horton, R.M. PCR mediated recombination and mutagenesis: SOEing together tailor-made genes. Mol. Biotechnol. 1995, 3, 93–99. [Google Scholar] [CrossRef]
- Pattaragulwanit, K.; Dahl, C. Development of a genetic system for a purple sulfur bacterium: conjugative plasmid transfer in Chromatium vinosum. Arch. Microbiol. 1995, 164, 217–222. [Google Scholar] [CrossRef]
- Rethmeier, J.; Rabenstein, A.; Langer, M.; Fischer, U. Detection of traces of oxidized and reduced sulfur compounds in small samples by combination of different high- performance liquid chromatography methods. J.Chromatogr.A 1997, 760, 295–302. [Google Scholar] [CrossRef]
- Shu, X.; Shaner, N.C.; Yarbrough, C.A.; Tsien, R.Y.; Remington, S.J. Novel chromophores and buried charges control color in mFruits. Biochemistry 2006, 45, 9639–9647. [Google Scholar] [CrossRef]
- Heger, A.; Holm, L. Rapid automatic detection and alignment of repeats in protein sequences. Proteins 2000, 41, 224–237. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; Bridgland, A.; Meyer, C.; Kohl, S.A.A.; Ballard, A.J.; Cowie, A.; Romera-Paredes, B.; Nikolov, S.; Jain, R.; Adler, J.; Back, T.; Petersen, S.; Reiman, D.; Clancy, E.; Zielinski, M.; Steinegger, M.; Pacholska, M.; Berghammer, T.; Bodenstein, S.; Silver, D.; Vinyals, O.; Senior, A.W.; Kavukcuoglu, K.; Kohli, P.; Hassabis, D. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Fellay, R.; Frey, J.; Krisch, H.M. Interposon mutagenesis of soil and water bacteria: a family of DNA fragments designed for in vitro insertional mutagenesis of Gram-negative bacteria. Gene 1987, 52, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Solovyev, V.; Salamov, A. Automatic annotation of microbial genomes and metagenomic sequences. In Metagenomics and its applications in agriculture, biomedicine and environmental studies; Li, R. W., Ed.; Nova Science Publishers: Hauppage, N.Y, 2010; pp. 71–78. [Google Scholar]
- Reynolds, R.; Bermudez-Cruz, R.M.; Chamberlin, M.J. Parameters affecting transcription termination by Escherichia coli RNA. I. Analysis of 13 rho-independent terminators. J. Mol. Biol. 1992, 224, 31–51. [Google Scholar] [CrossRef]
- Fodor, B.; Rákhely, G.; Kovács, A.T.; Kovács, K.L. Transposon mutagenesis in purple sulfur photosynthetic bacteria: identification of hypF, encoding a protein capable of processing [NiFe] hydrogenases in α, β and γ subdivisions of the proteobacteria. Appl. Environ. Microbiol. 2001, 67, 2476–2483. [Google Scholar] [CrossRef]
- Dahl, C. Sulfur metabolism in phototrophic bacteria. In Modern topics in the phototrophic prokaryotes: Metabolism, bioenergetics and omics; Hallenbeck, P. C., Ed.; Springer International Publishing: Cham, 2017; pp. 27–66. [Google Scholar]
- Regmi, C.K.; Bhandari, Y.R.; Gerstman, B.S.; Chapagain, P.P. Exploring the diffusion of molecular oxygen in the red fluorescent protein mCherry using explicit oxygen molecular dynamics simulations. J. Phys. Chem. B 2013, 117, 2247–2253. [Google Scholar] [CrossRef]
Figure 1.
Phase contrast microscopy (left panels), fluorescence microscopy (middle panel) and merged phase contrast and fluorescence images of A. vinosum Rif50 wild type (A) and strains carrying pBBT_dsr_mCherry_Gm (B), pBBR_dsr_SecSgpD, (C) or pBBR_dsr_SgpD_mCherry. Excitation times for phase contrast and for the mCherry channel (587 nm) were 49.2 s and 1 s, respectively. All cultures were grown photolithoautotrophically in the light in the presence of 4 mM sulfide. Cells were collected by centrifugation when sulfur globule formation had set in, and kept on ice until transfer to microscope slides on which the cells were exposed to air for 60 to 80 min before microscopy. Size bar: 2 µm
Figure 1.
Phase contrast microscopy (left panels), fluorescence microscopy (middle panel) and merged phase contrast and fluorescence images of A. vinosum Rif50 wild type (A) and strains carrying pBBT_dsr_mCherry_Gm (B), pBBR_dsr_SecSgpD, (C) or pBBR_dsr_SgpD_mCherry. Excitation times for phase contrast and for the mCherry channel (587 nm) were 49.2 s and 1 s, respectively. All cultures were grown photolithoautotrophically in the light in the presence of 4 mM sulfide. Cells were collected by centrifugation when sulfur globule formation had set in, and kept on ice until transfer to microscope slides on which the cells were exposed to air for 60 to 80 min before microscopy. Size bar: 2 µm
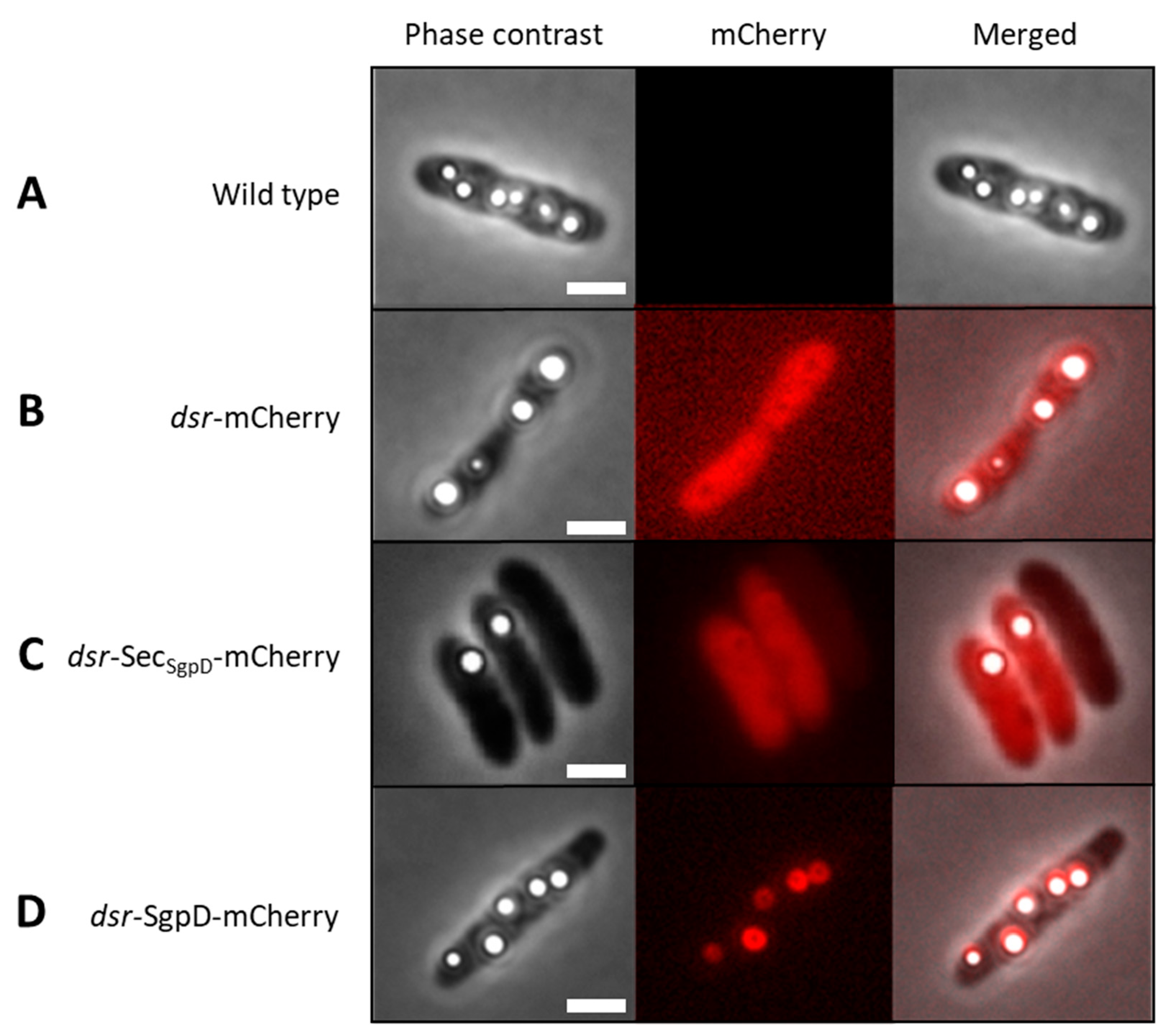
Figure 2.
Phase contrast microscopy (left panel), fluorescence microscopy (middle panel) and merged phase contrast and fluorescence images of sulfur globules isolated from A. vinosum Rif50 carrying pBBR_dsr_SgpD_mCherry. Excitation times for phase contrast and for the mCherry channel (587 nm) were 49.2 s and 5 s, respectively. Size bar: 2 µm.
Figure 2.
Phase contrast microscopy (left panel), fluorescence microscopy (middle panel) and merged phase contrast and fluorescence images of sulfur globules isolated from A. vinosum Rif50 carrying pBBR_dsr_SgpD_mCherry. Excitation times for phase contrast and for the mCherry channel (587 nm) were 49.2 s and 5 s, respectively. Size bar: 2 µm.
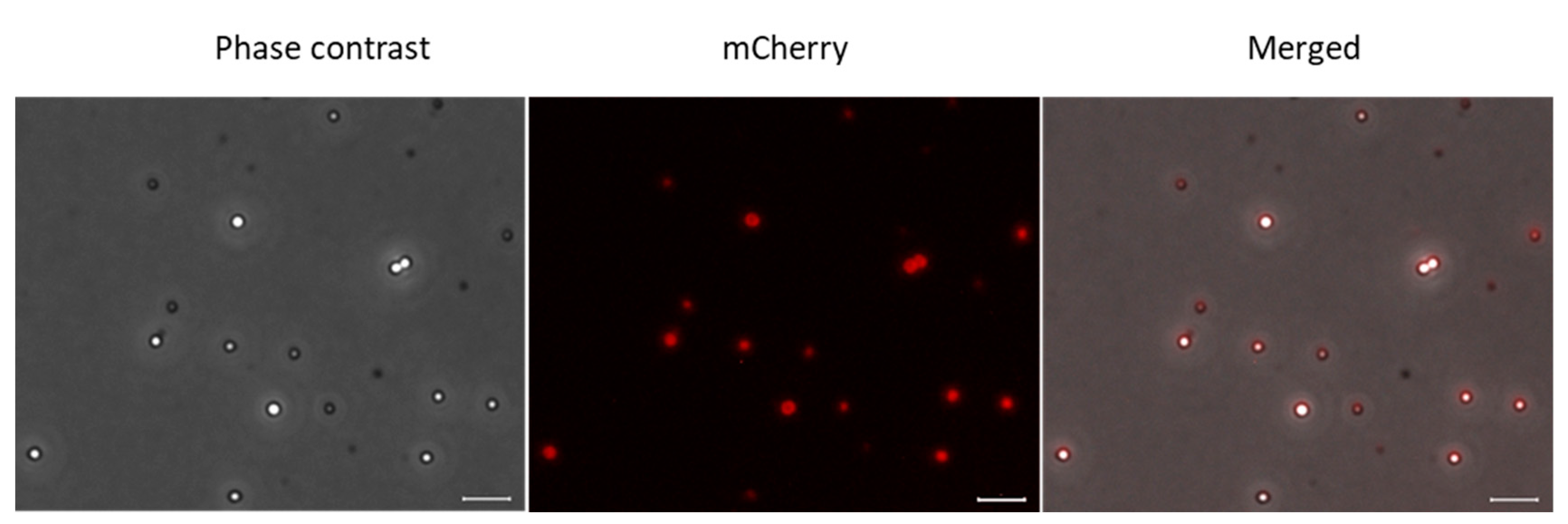
Figure 4.
Alignment of repeats in SgpD without its signal peptide generated by RADAR. Notation: BW, best window; Level, depth of recursion; the alignment row reports first and last residue and, in parentheses, the alignment score and Z-score of the repeat.
Figure 4.
Alignment of repeats in SgpD without its signal peptide generated by RADAR. Notation: BW, best window; Level, depth of recursion; the alignment row reports first and last residue and, in parentheses, the alignment score and Z-score of the repeat.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



