
Submitted:
20 June 2023
Posted:
20 June 2023
You are already at the latest version
Abstract
The basic helix-loop-helix (bHLH) transcription factor family is the second-largest transcription factor in plants. Members of this family are involved in the process of growth and development, secondary metabolic biosynthesis, signal transduction, and plant resistance. Loropetalum chinense var. rubrum is a critical woody plant with higher ornamental and economic values, which used as ornamental architecture and traditional Chinese herbal medicine plant. However, the bHLH transcription factor in the L. chinense var. rubrum (Loropetalum chinense var. Rubrum) have yet to be systematically demonstrated, and their role in anthocyanin biosynthesis remains secret. Here, we identified 165 potential LcbHLHs genes by two methods, and they were unequally distributed on chromosomes 1 to 12 of the L. chinense genome. Based on a phylogenetic comparison with proteins from Arabidopsis, these bHLH proteins were categorized into 21 subfamilies. Most LcbHLHs in a given subfamily had similar gene structures and conserved motifs. The gene ontology annotation and cis-elements predicted that the LcbHLHs had many molecular functions and were involved in plant growth processes, including flavonoid and anthocyanin biosynthetic processes, etc. Transcriptomic analysis revealed different expression patterns among different tissues and cultivars in L. chinense. Many LcbHLHs were expressed in leaves, and only a few genes were highly expressed in flowers. Six LcbHLH genes differentially expressed between species and periods in color variation, which may play a key role in anthocyanin synthesis. These further functional predictions of LcbHLHs were established by quantitative real-time PCR analysis and protein-protein interaction network. This study identified the six potential bHLH genes related to anthocyanin in L. chinense and created a solid framework for future research on the function and evolution of bHLHs.
Keywords:
L. chinense var. rubrum.
; bHLH transcription factor
; expression analyses
; anthocyanin
1. Introduction
Transcription factors (TFs) serve as a central regulators to regulate the expression of target genes and they can form intricate networks through protein–protein interactions to control or affect many biological processes at the transcription level [1]. Among the various TFs, the bHLH TFs family is a relatively large family of transcription factors, which have been widely found in various eukaryotes. The bHLH domain often makes up of 50-60 amino acids and has two functionally distinct regions: a length of 10-15 basic amino acids (The basic region) and a stretch of roughly 40 amino acids(the helix-loop-helix region), which can form two amphipathic α-helices separated by an intervening loop[2,3]. The basic region is located at the N-terminus of the bHLH domain and is the DNA binding region that allows the bHLH TFs to bind to the E-box (CANNTC), while the HLH region is located at the C-terminus of the bHLH domain and acts as a dimerization domain which can promote the formation of homodimers or heterodimers between proteins to alter the expression of target genes involved in various signaling pathways [4,5]. From the bHLH protein domain, only 19 amino acids are conserved and possesses a highly conservative H-E-R motif. Previous studies have typically classified members of the bHLH superfamily into subfamilies or subgroups based on these conserved motifs, evolutionary relationships, DNA binding specificity, and structural domains. Ledent and Atchley have classified the bHLH transcription factors in animals into six subgroups (A-F) based on sequence homology and phylogenetic relationships [3,6]. However, because of the relative independence of the bHLH transcription factors' genealogy between plant and animal, there are differences in the classification and so far no definite categories of plant bHLH transcription factor families criteria have been proposed[7]. Generally, the bHLH transcription factor family in plants has been divided into 15-26 groups [3,8,9]. 147 and 167 bHLH family members were identified from Arabidopsis and rice, respectively, and then classified into 21 and 25 subfamilies [9,10]. Subsequently, the various functions of the bHLH gene also have been validated.
Through previous studies, scientists have classified the function of bHLH genes in Arabidopsis and found that bHLH genes play different roles in different aspects [11]. In terms of mineral nutrition of plants, the AtbHLH121, AtbHLH18, AtbHLH19, AtbHLH20, AtbHLH25 can regulate iron homeostasis by indirectly activating the FER-LIKE IRON DEFICIENCY-INDUCED TRANSCRIPTION FACTOR (FIT) [12]. In growth and development, the AtLP1 and AtLP2 can regulate longitudinal cell elongation[13]. Also, the AtNFL promotes flowering under short-day conditions in Arabidopsis [14]. The PobHLH5 and PobHLH8 of P. ostreatus infestation in Arabidopsis verified that bHLH genes play an important role in growth and development[15]. In addition, bHLHs are also involved in plant abiotic stresses, signal transmission and secondary metabolism, etc.[16] For example, the overexpression of TabHLH39 improved drought tolerance, salt tolerance and frost resistance in transgenic Arabidopsis[17] and The MdCIB1 in apple plays an active role in drought resistance[18]. Researchers also found that expression of CpbHLH1 in transgenic model plants suppressed anthocyanin accumulation [19] and NnTT8 in Lotus had been indicated to be involved in the positive regulation of anthocyanin biosynthesis [20]. It has been further shown that pomegranate fruit anthocyanins can be regulated by a combination of bHLHs and MYB [21], especially the formation of MYB-bHLH-WD40 ternary complex[22,23].
Loropetalum chinense var. rubrum(L. chinense)is a typical and vivid flowering and foliage plant in Asia, Europe, and America. Ornamental plants lovers deeply enjoy its beautiful foliage and pretty plant figure [24]. The ornamental value of Loropetalum chinense var.rubrum mainly includes leaf color, flower color and tree posture, among which leaf color is one of the essential ornamental values. Previous studies have shown that anthocyanin is a crucial compound for the leaf coloration of Loropetalum chinense var.rubrum [25]. Its synthesis is usually affected by the external environment, synthetic structural genes, and regulatory genes. Studies have also shown that short-wavelength light quality is more conducive to improving the activity of phenylalanine ammonia lyase (PAL) in the callosity of Loropetalum chinense var.rubrum and promoting the synthesis of anthocyanin in the callus[26]. High temperature and drought usually cause the regreening of Loropetalum chinense var.rubrum leaves, which is mainly due to the degradation of anthocyanins [27]. In addition, the researchers also further understood the coloration mechanism of L.chinense leaves from the molecular level, identified and cloned structural genes related to anthocyanin synthesis, including LcCHS, LcDFR, LcCHI, LcANS and so on[28]. bHLH gene is one of the essential regulatory genes in the anthocyanin synthesis pathway, which can encode the corresponding transcription factors and then activate or inhibit the spatial and temporal expression of structural genes by forming the MYB-bHLH-WD40 complex, thereby regulating the synthesis of plant anthocyanins. It has been reported that the genome sequencing and identification of the bHLH family have been completed in a variety of plants, such as Arabidopsis, potato, bamboo, and grape etc[11,29,30,31]. However, the characteristics of bHLH transcription factors and their roles in L.chinense are still unclear.
In this study, the bHLH gene family of L. chinense was systematically identified by using bioinformatics methods. Then, potential leaf color-regulated bHLH genes were filtered by a series of biological analyses and expression pattern analysis. These results gave us a further understanding of the structure, function, and evolution of the L. chinense bHLH family, and provided a potential basis for the regulatory network of leaf coloration in Loropetalum chinense var.rubrum.
2. Results
2.1. Identification of LcbHLHs gene members and Phylogenetic Analysis
A total of 165 bHLH members were acquired by the method of homologous blast and HMM rsearch from genomic data of L. chinense and named LcbHLH1 to LcbHLH165 according to the chromosomal localization (Figure 1 Table S1). Prediction of physicochemical properties by ExPasy revealed that the number of amino acids contained in the bHLH protein sequence of L. chinense (Table S2), which ranged from 120 (LcbHLH96) to 1186aa (LcbHLH164), with an average of 375aa. The molecular weight of these protein ranged from 13460.46 (LcbHLH96) to 130429.73Da (LcbHLH84), with an average of 41508.43Da, and the theoretical isoelectric points ranged from 4.70 (LcbHLH62, LcbHLH102, LcbHLH103) to 10.12 (LcbHLH96), with 62.71% of them lower than 7, as predicted to be acidic. It was consistent with the previous study of the isoelectric point pattern in Arabidopsis and Oryza sativa. The Grand average of hydropathicity (GRAVY) of the proteins in the range of -1.037 to -0.102, showing that all LcbHLHs are hydrophilic. The instability index (II) ranged from 36.68 to 71.31, with only two manifested stable proteins (II < 40) and the aliphatic index was between 51.42 and 105.58. Most of the LcbHLHs were localized in the nucleus, and only a few were distributed in the cytoplasm, chloroplasts, plasmodesmata and Golgi apparatus. No signal peptide was found for any of the LcbHLHs by SignalP, indicating that they are non-secretory proteins.
In order to clarify the evolutionary relationship of L. chinense var. rubrum, a phylogenetic tree was constructed with Arabidopsis (Figure S1). The presence of LcbHLHs in 21 of the 24 Arabidopsis bHLH subfamilies and only three subfamilies XIII, XIV and XV did not contain members of LcbHLHs (Figure 2). Subfamily XII with 20 members, was the largest subfamily of LcbHLHs, while subfamilies IVd, VI and X were the smallest, each with only a member. In addition, compared with A. thaliana[5], Ficus carica L[32], P. persica[33] (Table 1), the III(a+b+c), the III(d+e), the IVa, the VII(a+b) and the XII subfamilies were considerably expanded.
2.2. Multiple Sequence Alignment, Motif, Domain and Structure Analysis
To further elucidate the structural features of LcbHLH proteins, a multiple sequence alignment analysis was performed on the bHLH structural domain. As shown in Figure 3, each LcbHLH protein displayed four conserved regions, including one basic region, one loop region and two helix regions. Most LcbHLHs proteins possessed hightly conserved basic region and two helixes, except the LcbHLH120, LcbHLH121, LcbHLH122, LcbHLH123, LcbHLH124 and LcbHLH125. LcbHLH122 and 123 had no loop region and two helix regions. LcbHLH120, LcbHLH121, LcbHLH124 and LcbHLH159 had no loop region and the second helix region. Multiple sequence alignment showed that 24 amino acid residues were highly conserved (> 60% consensus ratio), and two of those were conserved with a 100% consensus ratio. It is noteworthy that most LcbHLH proteins have the highly conserved H-ER-RR structures and Leu-27 in helix 1 and Leu-59 in helix 2 of the HLH region that are considered to play an important role in protein dimerization. Thus, we speculated that LcbHLH proteins may have the capacity to form protein complexes.
The conserved motifs, domain distribution and gene structure analysis of L. chinense were carried out to obtain more information about bHLHs. As is shown in Figure 4, there were ten motifs were identified and marked as 1-10 (Table 2). The different motifs correspond to their bHLH superfamily domains. For example, we can clearly see that motif 8 represents the bHLH-MYC_N domain, and motif 1 and motif 2 can form various bHLH structural domains. In combination with protein sequence alignment, one basic region and one helix region were predicted in motif 1, and another helix region were predicted in motif 2. However, because of the less conserved protein sequence of loop region between helix 1 and helix 2, most of the LcbHLH proteins had a combination of motifs 1 and 2, while LcbHLH122, LcbHLH123, LcbHLH129, LcbHLH128, LcbHLH131, LcbHLH81, LcbHLH76, LcbHLH159, LcbHLH55, LcbHLH54, LcbHLH108, LcbHLH148, LcbHLH146, and LcbHLH93 have only motif 1. The LcbHLH proteins clustered in the same subfamily often have similar motifs. For example, motifs1, 2, 5, 4, 9, 6 and 10 were identified in subfamily II, and Motifs 1, 2, and 8 were identified in subfamily III(d+e+f).
The evolutionary relationships of the members of gene families can be shown through gene structure analysis. The gene structure of LcbHLHs in the same supfamily always were consistent. The exon number of LcbHLHs varied from 1 to 13, whereas the exon-intron organizations were phylogenetically related (Figure 4). The LcbHLHs with one exon were clustered in four subfamilies (III(d+e), VIIIb, Orphans and II) and all the members of subfamily VIIIb and Orphans only have one exon.
2.3. Gene Duplication and Collinear Correlation Analysis
Tandem and fragment chromosome replication are critical means of gene family amplification. The 165 LcbHLHs in our study were unequally distributed among 12 L. chinense var. rubrum chromosomes, with a maximum of 22 on chr3 and a minimum of 2 on chr4. Tandem replication is thought to occur when the distance between genes is less than 100 kb, and 7 pairs of LcbHLHs fell into that category (Table S3). An intraspecific collinearity study revealed that fragment replication produced 56 pairs of LcbHLHs (Table S4). The findings have shown that tandem replication and fragment replication are critical processes for expanding the LcbHLH gene family. In addition, the substitution rate ratio, Ka/Ks, is applicable criterion for gene duplication selective pressure. Ka/Ks value less than 1 often represents negative selection, equal to 1 represents a neutral selection, and greater than 1 represents a positive selection. Among the 56 pairs of fragment replication gene pairs and 7 pairs of tandem replication gene pairs, except that the pair of LcbHLH29/30, LcbHLH140/141, LcbHLH142/144, and LcbHLH102/103 in the fragment replication pair were greater than 1, which showed positive selection. The others' Ka/Ks values were less than 1. It was speculated that it might have been purified and selected in the evolutionary history, indicating that most bHLH genes evolved slowly (Table S5).
In order to better understand the genetic differentiation, gene replication and evolution among bHLH gene families of L. chinense, Arabidopsis, Oryza sativa, Zea mays and Vitis vinifera, MCScanX was used to analyze the homologous bHLH genes among these species. 89, 44, 41 and 123 pairs of orthologous bHLH genes were detected in three comparisons (L. chinense vs. Arabidopsis, L. chinense vs. Oryza sativa, L. chinense vs. Zea mays and L. chinense vs. Vitis vinifera) (Figure 5, Table S6). Therefore, the bHLH genes between L. chinense and Vitis vinifera were considered to be more closely related than the bHLH genes among L. chinense and Oryza sativa, Zea mays or Arabidopsis.
2.4. GO annotation and analyses of cis-regulatory elements
Highly divergent sequences outside the conserved structural domain of bHLH suggested that the LcbHLH protein may be involved in multiple biological processes to some extent. In order to clarify the particular functions of bHLH proteins, GO annotation of LcbHLH was performed. From Table S7, Most LcbHLH proteins are annotated to be associated with transcription factors and protein dimerization activity, regulation of transcription and developmental processes. For the cellular component, 139 of 158 bHLH genes from transcriptome data were located in the nucleus (139/158). Only a small number of genes were predicted distributions in organelles, such as the cytoplasm (4), cytosol (2), chloroplast envelope (1), nucleoplasm (1), vacuole (1), nuclear speck (2), and intracellular membrane-bounded organelle (1) etc. Also, few LcbHLHs are also located in less common membrane structures and complexes, including integral component of membrane (1), membrane (1), vacuolar membrane (1), transcription factor complex (1) and RNA polymerase II transcription factor complex (6). It was anticipated that practically most LcbHLH proteins were engaged in protein dimerization activity (152/158) according to the molecular functions. In addition, the LcbHLHs participated in various biological processes, and many LcbHLH proteins were annotated to be associated with DNA binding, growth regulation and response of stimuli. For example, 127 LcbHLHs associated with DNA binding TFs, part of LcbHLH proteins involved the growth and development including the regulation of growth (6), pollen development (10), anther wall tapetum development (11) and negative regulation of seed germination (6), and several LcbHLHs could react environmental stimulation like cold (7), iron (5) and light (2) etc.
Members of gene families can differ in their expression patterns as well as their function. We looked for cis-elements in the promoter regions of the various LcbHLH genes to see whether there were any distinctions in gene regulation. From this Table S8, Light response elements were the most common cis-elements in LcbHLH promoter sequences, including Box 4, Gap-box, G-Box, AE-box, I-box, GT1-motif, TCCC-motif and so on. In the promoter regions of LcbHLH genes, cis-elements participated in growth and development and related to different stress responses were also discovered, such as MSA-like involved in cell cycle regulation, motif I and RY-element involved in seed-specific regulation, GCN4 motif involved in endosperm development, circadian involved in circadian control etc. Also, the cis-elements related to different stress responses are various, including the abiotic response elements which involved in abscisic acid (ABRE), auxin (AuxRR-core, TGA-element, TGA-box, AuxRE), MeJA (CGTCA-motif, TGACG-motif), gibberellin responsiveness (GARE-motif, TATC-box P-box) and the biological response elements related to salt stress (DRE) and low temperature responsiveness (LTR) etc. We also identified some cis-regulatory elements with MYB recognition sites, which can regulate plant flavonoid synthesis (MBSI) by binding to MYB-related genes, and conduct drought induction (MBS) and light response (MRE).
2.5. Expression profiles of LcbHLHs and Gene Expression Analysis by qRT -PCR
The expression pattern of LcbHLHs was analyzed based on transcriptome data of different Loropetalum chinense cultivars. New varieties of Loropetalum chinense mainly include ‘Xiangnong-Nichang’(XNNC), ‘Xiangnong-Fengjiao’(XNFJ), ‘Xiangnong-Xiangyun’(XNXY) and ‘Xiangnong-Xiaojiao’(XNXJ) (Table S9, Figure 6A). In order to better observe the difference in LcbHLHs expression, we standardized the FPKM value and removed the LcbHLHs with FPKM value less than 1, which is usually considered an invalid expression. From that, an expression pattern map of 105 LcbHLHs was established (Figure 6B).
In Figure 6B, it was found that the expression levels of LcbHLHs in flowers, young leaves (I-period) and mature leaves (II-period) displayed significant differences. Five groups of LcbHLHs were divided according to the cluster analysis. Genes in group IV, V were highly expressed in the flower and lowly expressed in the leaves, mainly predicted to involve floral organ formation and development. For example, LcbHLH133/145/25/102/69/43 in group IV was up-regulated in the flowers of XNXY and XNFJ, while its corresponding leaves showed lower expression levels. LcbHLH84 in group V was highly expressed in flowers of all varieties, LcbHLH43 was significantly expressed in XNFJ flowers, and LcbHLH22/51 was highly expressed in XNXY flowers and lowly expressed in leaves. According to GO enrichment analysis, LcbHLH84 may be involved in photoperiod and flowering processes, LcbHLH69 can regulate pollen development, LcbHLH109 plays a vital role in double-forming fertilization azygote and endosperm and multicellular organismal development. In addition, LcbHLH51/25/102/43 can participate in gibberellin-related anabolism and mediate brassinosteroid signaling pathway. Cis-element analysis showed that all of these genes have light responsive elements and some hormone response elements, such as gibberellin, jasmonic acid, salicylic acid, auxin and abscisic acid. Previous studies have shown that plant flowering is regulated by various plant hormones. Among them, gibberellin plays an essential role in flowering, and other hormones such as abscisic acid, auxin, salicylic acid, and jasmonic acid are also involved in the regulation of flowering. These results were consistent with GO enrichment and transcriptome analysis to some extent.
Since the ornamental value of L. chinense was mainly reflected in leaf color, three genes (LcbHLH4/156/157) from IIIf subfamily were selected to involve in anthocyanin and flavonoid biosynthesis based on structure and function prediction. We performed a preliminary analysis of the gene expression profiles of different leaf color varieties in period I and II (Figure 6B). LcbHLH4 was found distinctly more highly expressed in purple leaves (XNNC) than in green leaves (XNXY) and significantly down-regulated as leaf color becomes green from period I to period II. On the other hand, LcbHLH156 and LcbHLH157 were highly expressed in period I of different dark leaf varieties (XNFJ, XNXJ, XNNC) and decreased in expression as the leaf color became green during II period. In addition, it is also worth noting that LcbHLH149/114 was also highly expressed in XNFJ and LcbHLH117 also showed high expression in dark leaf varieties (XNNC). These expression levels of these genes showed similar trends in leaf phenotype color changes. Thus, we speculated that their function is related to the synthesis of anthocyanins.
To further validate the relationship between LcbHLHs and L. chinense leaf color, we demonstrated the expression levels of candidate genes of XNFJ species from transcriptomic data using qRT-PCR technique. We also investigated the gene expression levels of three different colors of leaves in the natural state of HYJM1. These genes had consistent expression trends at two developmental periods (I and II) (Figure 7, Table S10). We could clearly see that the expression of these six candidate genes was higher in the young leaf stage (I) and significantly decreased in the leaf maturation stage (II), which was consistent with the phenotypic trend. In addition, we compared the gene expression levels of HYJM1 in the three types of leaves using green leaves (GL) as a control group (Figure 8A). The results showed that these six genes were differentially up-regulated in both mixed leaves (ML) and purple leaves (PL), especially the expression was most significantly up-regulated in ML, with the most significant difference in LcbHLH4/156/157 expression (Figure 8B, Table S11).
2.6. The protein-protein interaction network of candidate gene.
Based on the above results, it was preliminarily speculated that the six candidate genes LcbHLH4/156/157/114/117/149 were most likely to participate in anthocyanin biosynthesis. To clarify their functions, we further constructed protein interaction network via Arabidopsis protein database (Figure 9). As shown in Fig. 9, LcbHLH4 (TT8 in Arabidopsis) could coordinate with TTG1 and TT2 to make DIHYDROFLAVONOL 4-REDUCTASE (DFR) and BANYULS (BAN) correctly expressed and then regulated the flavonoid pathway[23,34,35]. Furthermore, LcbHLH156/157(GL3 in Arabidopsis) could interact with TTG1 to regulate trichome development[36]. It was identified as an essential regulator of the anthocyanin pathway in Arabidopsis, together with TTG1 and MYB75 transcription factors, regulating specific genes in the anthocyanin synthesis pathway[37]. LcbHLH117/149 (MYC2 in Arabidopsis) could involve in the jasmonic acid (JA) signaling pathway and control the biosynthesis of anthocyanin by regulating the expression of genes encoding positive regulators such as MYB Domain Protein 75/Production of Anthocyanin Pigment 1 (MYB75/PAP1) and Enhancer of Glabra 3 (EGL3).[38,39,40,41]. LcbHLH114 (AT1G01260 in Arabidopsis) is an ABA-inducible BHLH-type transcription factor, which also involved in JAs signal transduction through the interaction of COI1, JAZ and MYC2 to regulate anthocyanin synthesis indirectly[42]. The protein-protein interaction networks of LcbHLHs further suggested that these six bHLH proteins were involved in anthocyanin biosynthesis in different ways.
3. Discussion
3.1. Systematic and comprehensive genome-wide detection of LcbHLHs in L. chinense
With the completion of whole genome sequencing of a large number of species, a large number of studies have been conducted on the bHLH family of various species. Such as Arabidopsis (n=162)[8,9], rice (n=167)[10] , fig (n=118)[32], strawberry (n=113)[43], lotus (n=115)[44], peach (n=95)[45], bamboo (n=448)[30], potato(n=124)[31], grape (n=94)[31] and maize(n=208)[45] etc (The "n" here refers to the number of bHLH). Compared with the number of these species, the number of LcbHLHs is appropriate. However, the quantity is much smaller than bamboo and maize. Changes in the number of bHLH genes between species were often thought to be possibly associated with gene duplication events or genome size[46]. Our study identified 165 bHLH genes of L. chinense var. rubrum. According to phylogenetic analysis, these 165 bHLHs were divided into 21 subfamilies, and the number distribution of these subfamilies was similar to that of Arabidopsis thaliana. Protein multi-sequence alignment also confirmed that LcbHLHs had a typical bHLH domain. Further structural analysis revealed that most LcbHLHs were linked to DNA-binding and homodimer formation functions. Additionally, the analysis of conserved motifs discovered that most of the LcbHLH family members had the respective conserved motifs (motif1 and motif2), constituting the bHLH structural domain [5]. These results further confirmed the accuracy of bHLH gene family analysis in L.chinense var. rubrum and provided a basis for further application.
3.2. Functional prediction of LcbHLHs
As the second largest gene family in plants, bHLHs always play an irreplaceable role in plant growth and development survival[5,11]. In our study, LcbHLHs are predicted to be widely involved in the growth and development of plants, including the synthesis of floral organs, the regulation of circadian rhythm and the response to various stresses etc. In addition, a few LcbHLHs are closely related to the synthesis and metabolism of some hormones, such as gibberellin, jasmonic acid, auxin and so on. It is predicted that LcbHLHs also play an important role in regulating the synthesis of secondary metabolites, including the biosynthesis of anthocyanins and brassinosteroids. To further explore the accuracy of our functional analysis, we analyzed the transcriptome data and validated some of them by Real-time Quantitative PCR Detecting System.
Many LcbHLHs performed differently in different tissues. Some LcbHLHs were highly expressed in the flowers of the three varieties. LcbHLH106 was annotated as PIF3, which is a phytochrome-interacting factor necessary for photoinduced signal transduction that can further regulate flowering time[47]. LcbHLH13 was highly expressed in leaves, NR annotated as PIL5 (Table S12), which can indirectly regulate DELLA protein content and further participate in gibberellin pathway to regulate leaf elongation and other growth and development processes[48,49]. Moreover, the expression patterns of LcbHLHs in different periods and color leaves were analyzed. We found that the three genes(LcbHLH156/LcbHLH157/4)selected by phylogenetic analysis and GO annotation did show high expression patterns in dark leaves and LcbHLH114/117/149 also were up-regulated in dark leaf varieties. Further qRT-PCR results of the six genes showed the same trend in XNFJ, they all expressed higher in I period than II period that matched the transcriptomic results and leaf color characteristics of varieties. In addition, through the qPCR results of HYJM1 leaves with different colors, we also found that the expression trends of LcbHLH4/156/157 genes in HYJM1 leaves with different colors were generally consistent, and were significantly up-regulated in mixed and purple leaves.
In order to further demonstrate our hypothesis, we further analyzed the interaction relationship of these six bHLH proteins through the protein interaction network diagram. It is found that these proteins have mainly involved in two pathways. LcbHLH4/156/157 play an irreplaceable role in anthocyanin synthesis and trichome development, and have a predicted interaction with TT2/TTG1/MYB75, while LcbHLH117/149/114 mainly involves JA signal transduction. Previous studies have showed the bHLH in subfamily III(d+e) participate in the JA signal pathway, leading to the accumulation of anthocyanin in apples[50,51]and the regulation of plant defense by Arabidopsis thaliana[52,53,54]. In Arabidopsis, JAZ proteins directly interact with bHLH TFs (GL3, EGL3, and TT8) and MYB TFs (MYB75 and GL1). These bHLH and MYB TFs are essential components of the MBW complex to mediate anthocyanin accumulation [39]. In our study, LcbHLH4/156/157 candidate proteins directly regulated the composition of related complexes in the anthocyanin synthesis pathway as well as the expression of structural genes, while LcbHLH114/117/149 were related to the JAZ protein family and mainly mediated the JA signal pathway, and their roles in anthocyanin synthesis deserve further investigation.
4. Materials and Methods
4.1. Plant materials and data resources
Tissue specimens of L. chinense were obtained from the Floral Experimental Station of the College of Horticulture, Hunan Agricultural University, Changsha, China. These experimental materials included the flowers and leaves of some new Loropetalum chinense cultivars 'Xiangnong Xiangyun' (XNXY), 'Xiangnong Fengjiao' (XNFJ), 'Xiangnong Nichang' (XNNC), 'Xiangnong Xiaojiao' (XNXJ) and 'The NO.1 Loropetalum chinense' (HYJM1). The leaf material of HYJM1 is mainly three different colors of leaves in their natural state, while the leaf material of other species was mainly collected at the following developmental stages: a new leaf period (I-period): the leaf is soft and brightly colored, and a mature period (II-period): the leaf is leathery and darkly colored. Samples were obtained as three biological replicates from the four new varieties and immediately frozen in liquid nitrogen for transcriptome sequencing. The sample RNA preparation and library construction are consistent with the previous studies[55,56].
The raw transcriptome data involved in this paper were uploaded in the Genome Sequence Archive (Genomics, Proteomics & Bioinformatics 2021) at the National Genomics Data Center (Nucleic Acids Res 2022) and the Chinese National Center for Biological Information at the Institute of Genomics (GSA: CRA009284 and CRA009285). They are available at https://ngdc.cncb.ac.cn/gsa (accessed on May 10, 2023).
4.2. Identification and Physicochemical Characterization of the bHLH gene family
Genomic data and gene annotation information of L. chinense were obtained from the research group of L. chinense at Hunan Agricultural University[57]. First, downloaded the seed profile of the bHLH signature domain (PF00010) from the PFAM database(https://pfam.xfam.org/) and made use of a Hidden Markov Model HMMER (3.0) to screen the bHLH candidate protein with the E-value cut-off set to 10-5. Then, the Arabidopsis bHLH protein homology blast method was used to search extensively for candidate bHLH proteins. The protein sequences of Arabidopsis were downloaded from TAIR(https://www.arabidopsis.org/). The LcbHLHs candidate proteins were obtained by a combination of two methods. These proteins containing the bHLH domain were screened further in the databases of PFAM(Pfam: Home page (xfam.org) and SMART(https://smart.embl.de/). The subcellular localization of LcbHLHs was predicted by the online website Wolf-psort(https://wolfpsort.hgc.jp/). The sequences of LcbHLHs were analyzed bioinformatically, and the proteins' physicochemical parameters were calculated using ExPASy(https://web.expasy.org/compute_pi/).
4.3. Chromosomal Localization, Tandem Duplication and Collinearity of the bHLH genes
The chromosome distribution was performed on the online website GSDS(http://gsds.gao-lab.org/). Gene duplication analyses for L. chinense var. rubrum was finished by using the Multiple Collinearity Scan Toolkit (MCScanX). Also, through the tools of TBtools [58] and MCScanX, the interspecies collinearity analysis of bHLHs between L. chinense and Arabidopsis, Oryza sativa, Zea mays and Vitis vinifera were performed. All of these genomic data were obtained from EnsemblPlants (https://plants.ensembl.org/). The non-synonymous replacement rate (Ka) and synonymous replacement rate (Ks) of the replicated gene pairs were calculated by KaKs Calculator 2.0, and environmental selection pressure was analyzed by Ka/Ks ratio.
4.4. Phylogeny and Multiple-Sequence Alignment of LcbHLHs
DNAMAN software was used for protein multiple sequence alignment, and SnapGene software was used to plot the amino acid site distribution of the bHLH protein conserved domain. The phylogenetic trees of Arabidopsis thaliana and L. chinense were constructed by using MEGA11 software.
4.5. Analysis of Gene Structure, Conserved Motif and Family Structural Domains
The intron/exon gene structure maps were obtained based on GFF3 files via the online website the Gene Structure Display Server(GSDS)(http://gsds.gao-lab.org/). The analysis of conserved motifs through the MEME online website(https://meme-suite.org/meme/doc/meme.html), and the number of motifs was 10, other parameters were default. The final result graph was showed by TBtools software.
4.6. GO Annotation, Analysis of Cis-acting Components and Protein-Protein Interaction of LcbHLHs
The NCBI database was selected as a reference database for further GO analysis of LcbHLHs by using the Blast2GO program[59]. Then, a 2000 bp promoter upstream of the bHLH gene family member of L. chinense was extracted by TBtools software and uploaded to the online website PlantCARE (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/) for homeopathic element prediction. To better understand the regulation of LcbHLHs on protein-protein interaction networks, STRING database (STRING: functional protein association networks (string-db.org)) was used to predict the protein interaction network of the candidate bHLH gene family in Loropetalum chinense var. rubrum.
4.7. Gene expression patterns and quantitative real-time PCR analysis
All raw transcriptome data were mainly presented by FPKM values (the fragments per kilobase of transcript per million mapped reads). Homogenize raw FPKM values and visualize heatmap using R package.
Plants material RNA was extracted using FastPure Universal Plant Total RNA Isolation Kit. The first standard cDNA was synthesized by Evo M-ML RT Kit for qPCR and stored in a -40°C refrigerator. Primers were designed by Beacon Designer 8. (Table S13), and qRT-PCR was performed on a Bio-Rad CFX384TM with 2X SYBR Green Pro Taq HS Premix (AG, Hunan, China). The cDNA was diluted to 500 ng, and the system was set to 10μl to be prepared as three technical replicates. Each reaction with a 1μl template. The conditions for qRT-PCR are as follows: 95°C for 5 minutes, followed by 40 cycles of 95°C for 15 seconds, 60°C for 1 minute, and 72°C for 5 minutes. Quantitative PCR expression level was calculated by 2−∆∆CT method and expression values of three replicates were normalized using LcActin as the internal control. The error bar represents standard errors from three biological replicates.
5. Conclusions
In summary, we analyzed the physicochemical properties, phylogeny, gene structure, Ka/Ks value and collinearity of different species of bHLH family members of L. chinense var. rubrum. In addition, we also predicted the function of LcbHLHs by using cis-elements, phylogenetic tree clustering and GO annotation. Six candidate genes related to anthocyanin synthesis were selected and verified the function prediction of LcbHLHs based on transcriptome data. Then, further qPCR detection was performed on leaves with different leaf colors in HYJM1. All these results strengthen our understanding of the bHLH gene family in Loropetalum chinense var. rubrum, and provide a reference for further understanding the regulation of anthocyanin synthesis.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: Phylogenetic tree constructed with sequences of the LcbHLH and AtbHLH; Table S1: Gene accession number of all LcbHLH genes; Table S2: Physicochemical properties table; Table S3: The gene tandem replication on L. chinense var. rubrum chromosome; Table S4: The gene fragment replication on L. chinense var. rubrum chromosome; Table S5: The Ka/Ks ratios for duplicate pairs of LcbHLHs; Table S6: The synteny regions of bHLH genes in diverse species; Table S7: GO annotation of the bHLH proteins in L. chinense; Table S8: Cis-elements analyses of the bHLH proteins in L. chinense; Table S9: Transcriptome data FPKM value of LcbHLHs; Table S10: qPCR data of period I and II of XNFJ; Table S11: qPCR data of GL ML PL; Table S12: Nr annotation of LcbHLHs; Table S13: List of primers used for qRT-PCR.
Author Contributions
X. X, X.Y., and Y.L. (Yanlin Li) proposed the design of the article, performed the experiments, and authored or reviewed drafts of the article; Y.L. (Yang Liu) and L.L. performed the data analysis; Y.L. (Yang Liu) and Q.M. prepared the figures and/or tables; W.L. and D.Z. authored or reviewed drafts of the article Y.L. (Yang Liu) completed the manuscript together. All authors have read and agreed to the published version of the manuscript.
Funding
The work is funded by The Forestry Science and Technology Innovation Foundation of Hunan Province for Distinguished Young Scholarship (XLKJ202205), Open Project of Horticulture Discipline of Hunan Agricultural University (2021YYXK001), Hunan Province key research and development project (2022NK2026), The Found of Changsha Municipal Science and Technology Bureau (KQ2202227), Key project of Hunan Provincial Education Department (22A0155), The Forestry Bureau for Industrialization management of Hunan Province (2130221).
Data Availability Statement
Data are contained within the article and Supplementary Materials.
Acknowledgments
We acknowledge the editors and all anonymous reviewers for their constructive comments on this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Riechmann, J.L.; Ratcliffe, O.J. A Genomic Perspective on Plant Transcription Factors. Curr. Opin. Plant Biol. 2000, 3, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.E. Built by Association: Structure and Function of Helix-Loop-Helix DNA-Binding Proteins. Structure 1994, 2, 1–4. [Google Scholar] [CrossRef]
- Atchley, W.R.; Fitch, W.M. A Natural Classification of the Basic Helix–Loop–Helix Class of Transcription Factors. Proc. Natl. Acad. Sci. 1997, 94, 5172–5176. [Google Scholar] [CrossRef]
- Massari, M.E.; Murre, C. Helix-Loop-Helix Proteins: Regulators of Transcription in Eucaryotic Organisms. Mol. Cell. Biol. 2000, 20, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Heim, M.A. The Basic Helix-Loop-Helix Transcription Factor Family in Plants: A Genome-Wide Study of Protein Structure and Functional Diversity. Mol. Biol. Evol. 2003, 20, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Ledent, V.; Vervoort, M. The Basic Helix-Loop-Helix Protein Family: Comparative Genomics and Phylogenetic Analysis. Genome Res. 2001, 11, 754–770. [Google Scholar] [CrossRef]
- Carretero-Paulet, L.; Galstyan, A.; Roig-Villanova, I.; Martínez-García, J.F.; Bilbao-Castro, J.R.; Robertson, D.L. Genome-Wide Classification and Evolutionary Analysis of the BHLH Family of Transcription Factors in Arabidopsis, Poplar, Rice, Moss, and Algae. Plant Physiol. 2010, 153, 1398–1412. [Google Scholar] [CrossRef] [PubMed]
- Bailey, P.C.; Martin, C.; Toledo-Ortiz, G.; Quail, P.H.; Huq, E.; Heim, M.A.; Jakoby, M.; Werber, M.; Weisshaar, B. Update on the Basic Helix-Loop-Helix Transcription Factor Gene Family in Arabidopsis Thaliana. Plant Cell 2003, 15, 2497–2502. [Google Scholar] [CrossRef]
- Toledo-Ortiz, G.; Huq, E.; Quail, P.H. The Arabidopsis Basic/Helix-Loop-Helix Transcription Factor Family[W]. Plant Cell 2003, 15, 1749–1770. [Google Scholar] [CrossRef]
- Li, X.; Duan, X.; Jiang, H.; Sun, Y.; Tang, Y.; Yuan, Z.; Guo, J.; Liang, W.; Chen, L.; Yin, J.; et al. Genome-Wide Analysis of Basic/Helix-Loop-Helix Transcription Factor Family in Rice and Arabidopsis. Plant Physiol. 2006, 141, 1167–1184. [Google Scholar] [CrossRef]
- Hao, Y.; Zong, X.; Ren, P.; Qian, Y.; Fu, A. Basic Helix-Loop-Helix (BHLH) Transcription Factors Regulate a Wide Range of Functions in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 7152. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Chen, C.-L.; Cui, M.; Zhou, W.-J.; Wu, H.-L.; Ling, H.-Q. Four IVa BHLH Transcription Factors Are Novel Interactors of FIT and Mediate JA Inhibition of Iron Uptake in Arabidopsis. Mol. Plant 2018, 11, 1166–1183. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Zhang, J.; Wu, Y.-W.; Wang, Y.; Zhang, J.; Zheng, Y.; Li, Y.; Li, X.-B. BHLH Transcription Factors LP1 and LP2 Regulate Longitudinal Cell Elongation. Plant Physiol. 2021, 187, 2577–2591. [Google Scholar] [CrossRef]
- Sharma, N.; Xin, R.; Kim, D.-H.; Sung, S.; Lange, T.; Huq, E. NO FLOWERING IN SHORT DAY (NFL) Is a BHLH Transcription Factor That Promotes Flowering Specifically under Short-Day Conditions in Arabidopsis. Development 2016, 143, 682–690. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yang, W.; Jia, Q.; Wang, W.; Zhang, N.; Wang, X.; Wang, S. Pleurotus Ostreatus BHLH Transcription Factors Regulate Plant Growth and Development When Expressed in Arabidopsis. J. Plant Interact. 2017, 12, 542–549. [Google Scholar] [CrossRef]
- Sun, X.; Wang, Y.; Sui, N. Transcriptional Regulation of BHLH during Plant Response to Stress. Biochem. Biophys. Res. Commun. 2018, 503, 397–401. [Google Scholar] [CrossRef]
- Zhai, Y.; Zhang, L.; Xia, C.; Fu, S.; Zhao, G.; Jia, J.; Kong, X. The Wheat Transcription Factor, TabHLH39, Improves Tolerance to Multiple Abiotic Stressors in Transgenic Plants. Biochem. Biophys. Res. Commun. 2016, 473, 1321–1327. [Google Scholar] [CrossRef]
- Ren, Y.-R.; Yang, Y.-Y.; Zhao, Q.; Zhang, T.-E.; Wang, C.-K.; Hao, Y.-J.; You, C.-X. MdCIB1, an Apple BHLH Transcription Factor, Plays a Positive Regulator in Response to Drought Stress. Environ. Exp. Bot. 2021, 188, 104523. [Google Scholar] [CrossRef]
- Zhao, R.; Song, X.; Yang, N.; Chen, L.; Xiang, L.; Liu, X.-Q.; Zhao, K. Expression of the Subgroup IIIf BHLH Transcription Factor CpbHLH1 from Chimonanthus Praecox (L.) in Transgenic Model Plants Inhibits Anthocyanin Accumulation. Plant Cell Rep. 2020, 39, 891–907. [Google Scholar] [CrossRef]
- Deng, J.; Li, J.; Su, M.; Lin, Z.; Chen, L.; Yang, P. A BHLH Gene NnTT8 of Nelumbo Nucifera Regulates Anthocyanin Biosynthesis. Plant Physiol. Biochem. 2021, 158, 518–523. [Google Scholar] [CrossRef]
- Arlotta, C.; Puglia, G.D.; Genovese, C.; Toscano, V.; Karlova, R.; Beekwilder, J.; De Vos, R.C.H.; Raccuia, S.A. MYB5-like and BHLH Influence Flavonoid Composition in Pomegranate. Plant Sci. 2020, 298, 110563. [Google Scholar] [CrossRef]
- Liu, Y.; Hou, H.; Jiang, X.; Wang, P.; Dai, X.; Chen, W.; Gao, L.; Xia, T. A WD40 Repeat Protein from Camellia Sinensis Regulates Anthocyanin and Proanthocyanidin Accumulation through the Formation of MYB–BHLH–WD40 Ternary Complexes. Int. J. Mol. Sci. 2018, 19, 1686. [Google Scholar] [CrossRef] [PubMed]
- Shan, X.; Li, Y.; Yang, S.; Gao, R.; Zhou, L.; Bao, T.; Han, T.; Wang, S.; Gao, X.; Wang, L. A Functional Homologue of Arabidopsis TTG1 from Freesia Interacts with BHLH Proteins to Regulate Anthocyanin and Proanthocyanidin Biosynthesis in Both Freesia Hybrida and Arabidopsis Thaliana. Plant Physiol. Biochem. 2019, 141, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Cai, W.; Zhang, X.; Li, W.; Zhou, Y.; Chen, Y.; Mi, Q.; Jin, L.; Xu, L.; Yu, X.; et al. Different Pruning Level Effects on Flowering Period and Chlorophyll Fluorescence Parameters of Loropetalum Chinense Var. Rubrum. PeerJ 2022, 10, e13406. [Google Scholar] [CrossRef]
- Chen, Q.; Cai, W.; Zhang, X.; Zhang, D.; Zhang, W.; Xu, L.; Yu, X.; Li, Y. The Comparative Studies on Phytochemicals of Leaf Coloration of Loropetalum chinense var. rubrum. Acta Hortic. Sin. 2021, 48, 1969–1982. [Google Scholar] [CrossRef]
- Guo, P.; Deng, S.; Zhang, Y.; Xu, L.; Yu, X.; Li, Y. Effect of Different Light Quality on Callus Growth and Flavonoids Contene of Two Loropetalum,chinense Plants. Acta Bot. Boreali Occident. Sin. 2022, 42, 118–126. [Google Scholar]
- Cai, W.; Zhang, D.; Zhang, X.; Chen, Q.; Liu, Y.; Lin, L.; Xiang, L.; Yang, Y.; Xu, L.; Yu, X.; et al. Leaf Color Change and Photosystem Function Evaluation under Heat Treatment Revealed the Stress Resistance Variation between Loropetalum Chinense and L. Chinense Var. Rubrum. PeerJ 2023, 11, e14834. [Google Scholar] [CrossRef]
- Zhang, X. Cloning, Expression and Transformation of Lc CHI and Lc ANS Genes from Loropetalum Chinense Var. Rubrum. Master, Hunan University of Technology, 2020.
- Wang, R.; Zhao, P.; Kong, N.; Lu, R.; Pei, Y.; Huang, C.; Ma, H.; Chen, Q. Genome-Wide Identification and Characterization of the Potato BHLH Transcription Factor Family. Genes 2018, 9, 54. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Xiong, R.; Liu, H.; Wu, M.; Chen, F.; Hanwei Yan; Xiang, Y. Basic Helix-Loop-Helix Gene Family: Genome Wide Identification, Phylogeny, and Expression in Moso Bamboo. Plant Physiol. Biochem. 2018, 132, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Su, L.; Gao, H.; Jiang, X.; Wu, X.; Li, Y.; Zhang, Q.; Wang, Y.; Ren, F. Genome-Wide Characterization of BHLH Genes in Grape and Analysis of Their Potential Relevance to Abiotic Stress Tolerance and Secondary Metabolite Biosynthesis. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- Song, M.; Wang, H.; Wang, Z.; Huang, H.; Chen, S.; Ma, H. Genome-Wide Characterization and Analysis of BHLH Transcription Factors Related to Anthocyanin Biosynthesis in Fig (Ficus Carica L.). Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Feng, R.; Ma, R.; Shen, Z.; Cai, Z.; Song, Z.; Peng, B.; Yu, M. Genome-Wide Analysis of Basic Helix-Loop-Helix Superfamily Members in Peach. PLOS ONE 2018, 13, e0195974. [Google Scholar] [CrossRef] [PubMed]
- Baudry, A.; Heim, M.A.; Dubreucq, B.; Caboche, M.; Weisshaar, B.; Lepiniec, L. TT2, TT8, and TTG1 Synergistically Specify the Expression of BANYULS and Proanthocyanidin Biosynthesis in Arabidopsis Thaliana. Plant J. 2004, 39, 366–380. [Google Scholar] [CrossRef]
- Gonzalez, A.; Zhao, M.; Leavitt, J.M.; Lloyd, A.M. Regulation of the Anthocyanin Biosynthetic Pathway by the TTG1/BHLH/Myb Transcriptional Complex in Arabidopsis Seedlings. Plant J. 2008, 53, 814–827. [Google Scholar] [CrossRef] [PubMed]
- Payne, C.T.; Zhang, F.; Lloyd, A.M. GL3 Encodes a BHLH Protein That Regulates Trichome Development in Arabidopsis Through Interaction With GL1 and TTG1. Genetics 2000, 156, 1349–1362. [Google Scholar] [CrossRef]
- Wen, J.; Li, Y.; Qi, T.; Gao, H.; Liu, B.; Zhang, M.; Huang, H.; Song, S. The C-Terminal Domains of Arabidopsis GL3/EGL3/TT8 Interact with JAZ Proteins and Mediate Dimeric Interactions. Plant Signal. Behav. 2018, 13, e1422460. [Google Scholar] [CrossRef]
- Niu, Y.; Figueroa, P.; Browse, J. Characterization of JAZ-Interacting BHLH Transcription Factors That Regulate Jasmonate Responses in Arabidopsis. J. Exp. Bot. 2011, 62, 2143–2154. [Google Scholar] [CrossRef]
- Qi, T.; Song, S.; Ren, Q.; Wu, D.; Huang, H.; Chen, Y.; Fan, M.; Peng, W.; Ren, C.; Xie, D. The Jasmonate-ZIM-Domain Proteins Interact with the WD-Repeat/BHLH/MYB Complexes to Regulate Jasmonate-Mediated Anthocyanin Accumulation and Trichome Initiation in Arabidopsis Thaliana. Plant Cell 2011, 23, 1795–1814. [Google Scholar] [CrossRef]
- An, J.-P.; Li, H.-H.; Song, L.-Q.; Su, L.; Liu, X.; You, C.-X.; Wang, X.-F.; Hao, Y.-J. The Molecular Cloning and Functional Characterization of MdMYC2, a BHLH Transcription Factor in Apple. Plant Physiol. Biochem. 2016, 108, 24–31. [Google Scholar] [CrossRef]
- Shan, X.; Zhang, Y.; Peng, W.; Wang, Z.; Xie, D. Molecular Mechanism for Jasmonate-Induction of Anthocyanin Accumulation in Arabidopsis. J. Exp. Bot. 2009, 60, 3849–3860. [Google Scholar] [CrossRef]
- Yu, Q.; Hua, X.; Yao, H.; Zhang, Q.; He, J.; Peng, L.; Li, D.; Yang, Y.; Li, X. Abscisic Acid Receptors Are Involves in the Jasmonate Signaling in Arabidopsis. Plant Signal. Behav. 2021, 16, 1948243. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Li, G.; Hu, P.; Zhao, X.; Li, L.; Wei, W.; Feng, J.; Zhou, H. Identification of Basic/Helix-Loop-Helix Transcription Factors Reveals Candidate Genes Involved in Anthocyanin Biosynthesis from the Strawberry White-Flesh Mutant. Sci. Rep. 2018, 8, 2721. [Google Scholar] [CrossRef] [PubMed]
- Mao, T.-Y.; Liu, Y.-Y.; Zhu, H.-H.; Zhang, J.; Yang, J.-X.; Fu, Q.; Wang, N.; Wang, Z. Genome-Wide Analyses of the BHLH Gene Family Reveals Structural and Functional Characteristics in the Aquatic Plant Nelumbo Nucifera. PeerJ 2019, 7, e7153. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Lv, W.; Zhang, H.; Ma, L.; Li, P.; Ge, L.; Li, G. Genome-Wide Analysis of the Basic Helix-Loop-Helix (BHLH) Transcription Factor Family in Maize. BMC Plant Biol. 2018, 18, 1–14. [Google Scholar] [CrossRef]
- Flagel, L.E.; Wendel, J.F. Gene Duplication and Evolutionary Novelty in Plants. New Phytol. 2009, 183, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Oda, A.; Fujiwara, S.; Kamada, H.; Coupland, G.; Mizoguchi, T. Antisense Suppression of the Arabidopsis PIF3 Gene Does Not Affect Circadian Rhythms but Causes Early Flowering and Increases FT Expression. FEBS Lett. 2004, 557, 259–264. [Google Scholar] [CrossRef]
- Oh, E.; Yamaguchi, S.; Kamiya, Y.; Bae, G.; Chung, W.-I.; Choi, G. Light Activates the Degradation of PIL5 Protein to Promote Seed Germination through Gibberellin in Arabidopsis. Plant J. 2006, 47, 124–139. [Google Scholar] [CrossRef]
- Oh, E.; Yamaguchi, S.; Hu, J.; Yusuke, J.; Jung, B.; Paik, I.; Lee, H.-S.; Sun, T.; Kamiya, Y.; Choi, G. PIL5, a Phytochrome-Interacting BHLH Protein, Regulates Gibberellin Responsiveness by Binding Directly to the GAI and RGA Promoters in Arabidopsis Seeds. Plant Cell 2007, 19, 1192–1208. [Google Scholar] [CrossRef]
- Liu, X.-J.; An, X.-H.; Liu, X.; Hu, D.-G.; Wang, X.-F.; You, C.-X.; Hao, Y.-J. MdSnRK1.1 Interacts with MdJAZ18 to Regulate Sucrose-Induced Anthocyanin and Proanthocyanidin Accumulation in Apple. J. Exp. Bot. 2017, 68, 2977–2990. [Google Scholar] [CrossRef]
- Xie, X.-B.; Li, S.; Zhang, R.-F.; Zhao, J.; Chen, Y.-C.; Zhao, Q.; Yao, Y.-X.; You, C.-X.; Zhang, X.-S.; Hao, Y.-J. The BHLH Transcription Factor MdbHLH3 Promotes Anthocyanin Accumulation and Fruit Colouration in Response to Low Temperature in Apples. Plant Cell Environ. 2012, 35, 1884–1897. [Google Scholar] [CrossRef]
- Qi, T.; Wang, J.; Huang, H.; Liu, B.; Gao, H.; Liu, Y.; Song, S.; Xie, D. Regulation of Jasmonate-Induced Leaf Senescence by Antagonism between BHLH Subgroup IIIe and IIId Factors in Arabidopsis. Plant Cell 2015, 27, 1634–1649. [Google Scholar] [CrossRef] [PubMed]
- Abe, H.; Urao, T.; Ito, T.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis AtMYC2 (BHLH) and AtMYB2 (MYB) Function as Transcriptional Activators in Abscisic Acid Signaling. Plant Cell 2003, 15, 63–78. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Qi, T.; Fan, M.; Zhang, X.; Gao, H.; Huang, H.; Wu, D.; Guo, H.; Xie, D. The BHLH Subgroup IIId Factors Negatively Regulate Jasmonate-Mediated Plant Defense and Development. PLOS Genet. 2013, 9, e1003653. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, L.; Zhang, D.; Su, D.; Li, W.; Wang, X.; Chen, Q.; Cai, W.; Xu, L.; Cao, F.; et al. Comprehensive Analysis of Metabolome and Transcriptome Reveals the Mechanism of Color Formation in Different Leave of Loropetalum Chinense Var. Rubrum. BMC Plant Biol. 2023, 23, 133. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, L.; Zhang, D.; Liu, Y.; Lin, L.; Xiong, X.; Zhang, D.; Sun, M.; Cai, M.; Yu, X.; et al. Transcriptomic and Metabolomic Profiling Provides Insights into Flavonoid Biosynthesis and Flower Coloring in Loropetalum Chinense and Loropetalum Chinense Var. Rubrum. Agronomy 2023, 13, 1296. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Y.; Liu, Y.; Lin, L.; Xiong, X.; Zhang, D.; Li, S.; Yu, X.; Li, Y. Genome-Wide Identification and Characterization of WRKY Transcription Factors and Their Expression Profile in Loropetalum Chinense Var. Rubrum. Plants 2023, 12, 2131. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Conesa, A.; Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A Universal Tool for Annotation, Visualization and Analysis in Functional Genomics Research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef]
Figure 1.
Chromosomal distribution of LcbHLHs. The left scale is used to assess chromosome size. All chromosomes contain the LcbHLH genes. The chr0 represents scaffold.
Figure 1.
Chromosomal distribution of LcbHLHs. The left scale is used to assess chromosome size. All chromosomes contain the LcbHLH genes. The chr0 represents scaffold.
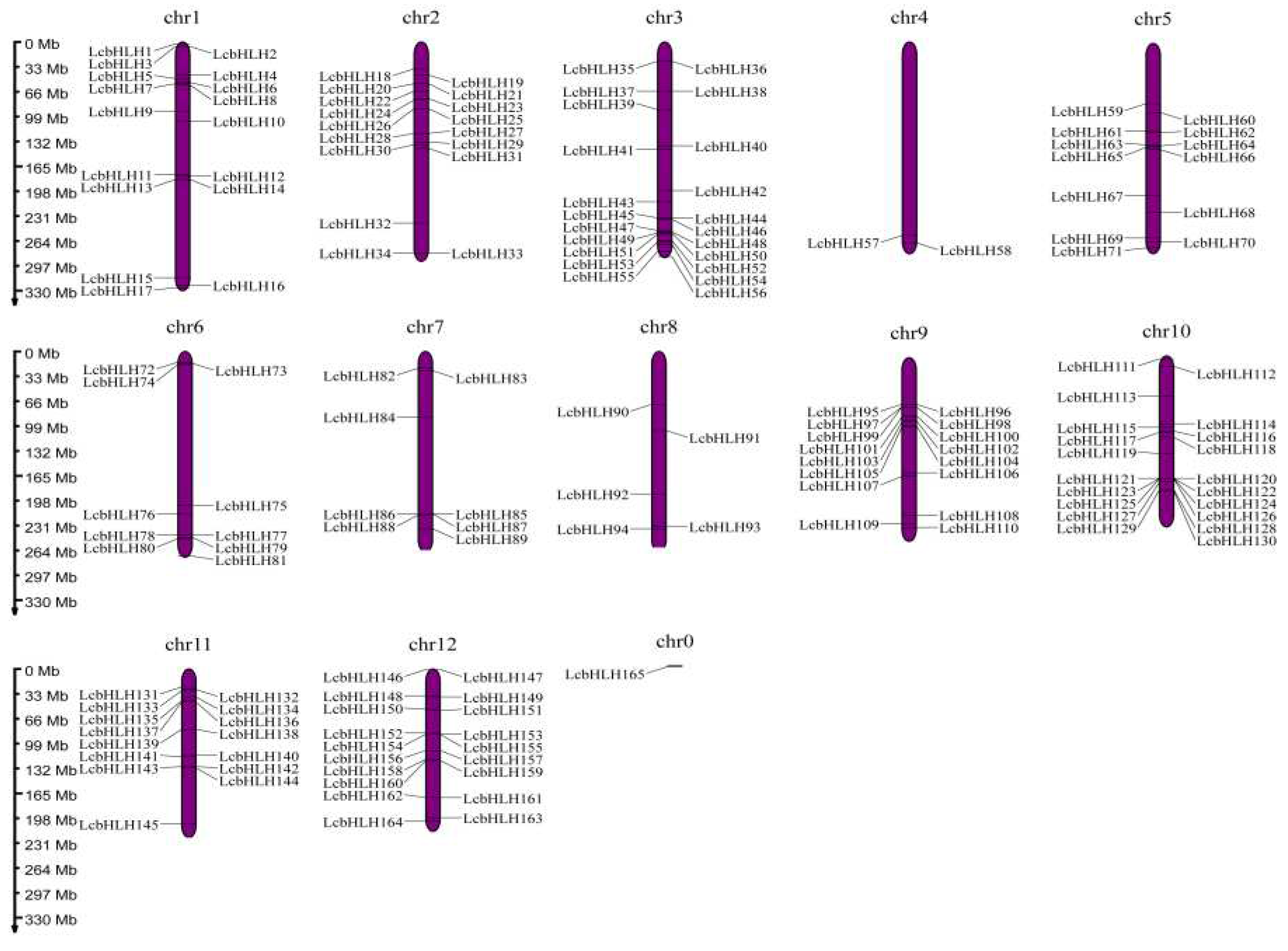
Figure 2.
Phylogenetic analysis of bHLH proteins in L. chinense. The phylogenetic trees of Arabidopsis thaliana and L. chinense were constructed by using MEGA11 software with the neighbor-joining (NJ) method. The parameters are set as follows: method "p-distance", gaps date treatment "partial deleted", self-expansion value "Bootstrap=1000".
Figure 2.
Phylogenetic analysis of bHLH proteins in L. chinense. The phylogenetic trees of Arabidopsis thaliana and L. chinense were constructed by using MEGA11 software with the neighbor-joining (NJ) method. The parameters are set as follows: method "p-distance", gaps date treatment "partial deleted", self-expansion value "Bootstrap=1000".
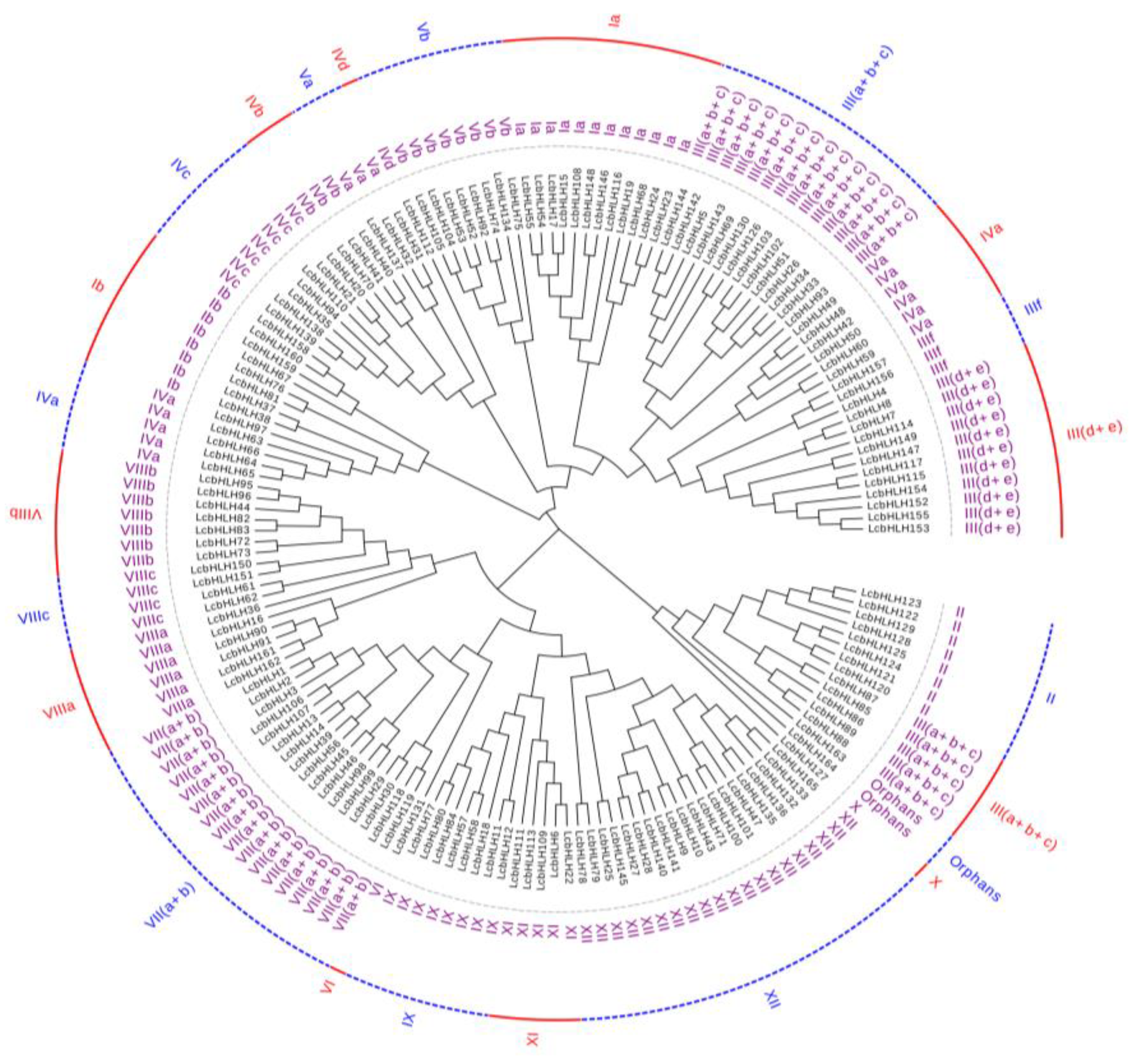
Figure 3.
Multiple alignments of the bHLH domains in the LcbHLH family proteins. Conserved amino acids in the LcbHLHs domain. Sequence identity > 60 % in gray or black shades.
Figure 3.
Multiple alignments of the bHLH domains in the LcbHLH family proteins. Conserved amino acids in the LcbHLHs domain. Sequence identity > 60 % in gray or black shades.

Figure 4.
Phylogenetic relationships, family structural domains, conserved motifs, and gene structural analyses of the LcbHLHs. (A)Phylogenetic tree of LcbHLHs. (B)The family domain distribution and types of LcbHLH. (C)Conserved motifs of LcbHLHs. The blue rectangle represents the exon, the orange rectangle represents the UTR, and the black line represents the intron. The size of these genes is evaluated by the bottom coordinate. (D)Gene structure of LcbHLHs. The 10 predicted motifs are depicted by rectangles of various colors.
Figure 4.
Phylogenetic relationships, family structural domains, conserved motifs, and gene structural analyses of the LcbHLHs. (A)Phylogenetic tree of LcbHLHs. (B)The family domain distribution and types of LcbHLH. (C)Conserved motifs of LcbHLHs. The blue rectangle represents the exon, the orange rectangle represents the UTR, and the black line represents the intron. The size of these genes is evaluated by the bottom coordinate. (D)Gene structure of LcbHLHs. The 10 predicted motifs are depicted by rectangles of various colors.
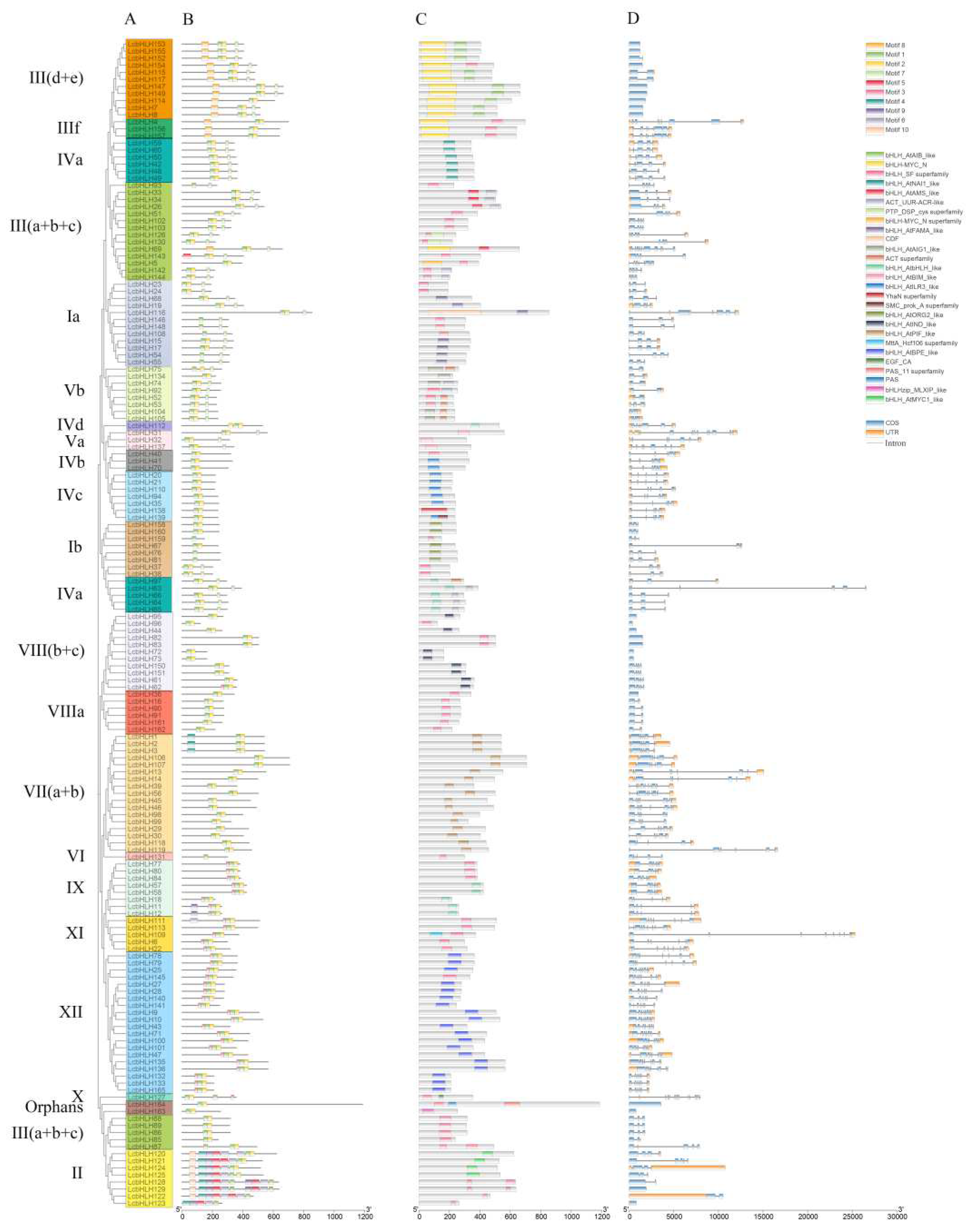
Figure 5.
Syntenic analyses of bHLH genes in L. chinense, Arabidopsis (A), Oryza sativa (B), Zea mays (C) and Vitis vinifera(D).
Figure 5.
Syntenic analyses of bHLH genes in L. chinense, Arabidopsis (A), Oryza sativa (B), Zea mays (C) and Vitis vinifera(D).
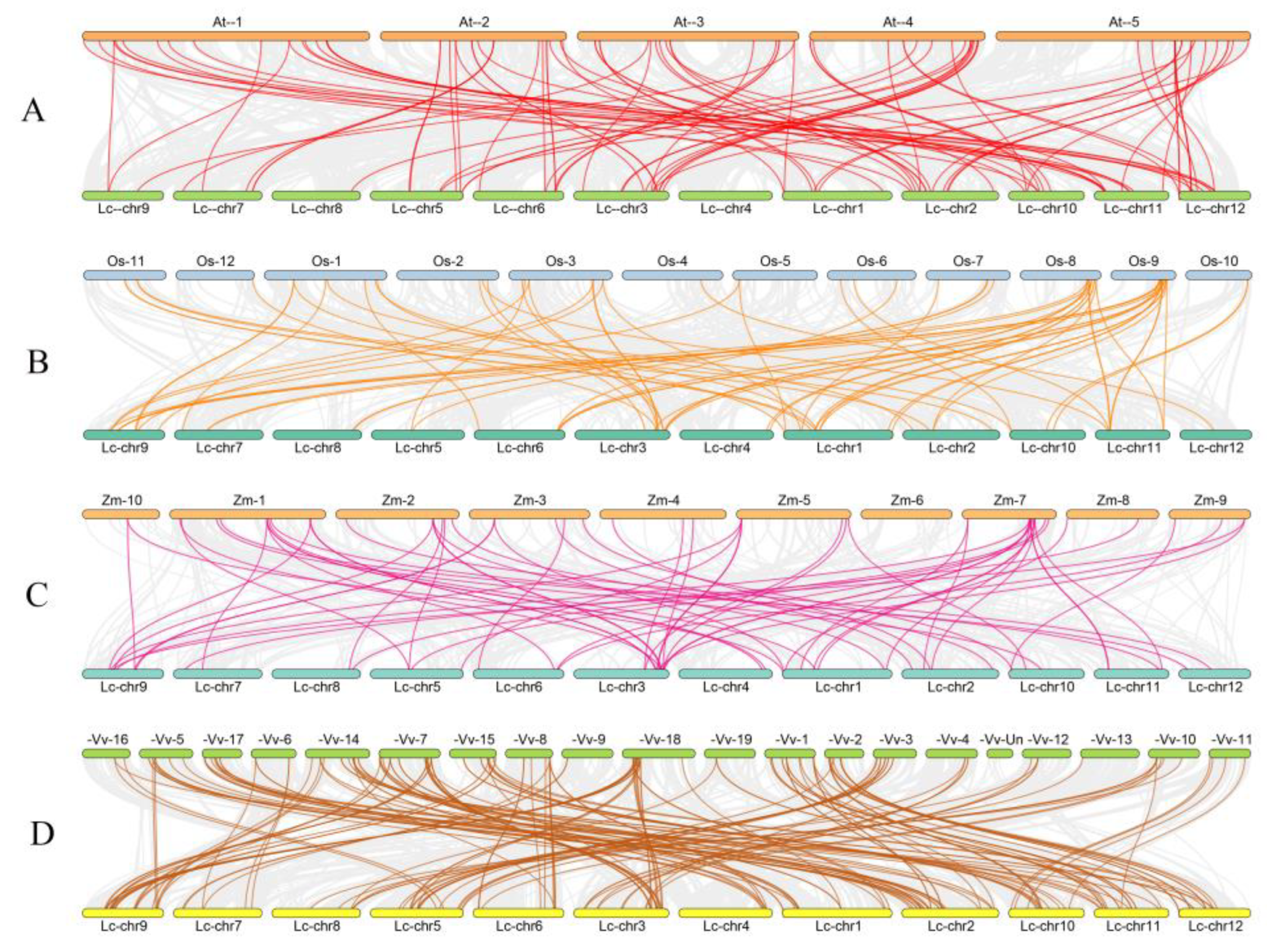
Figure 6.
Expression pattern of the LcbHLHs (A) Four varieties with different colored leaves and flowers. The "F" after the name indicates the flower, and"I" and "II" represent the first period and the second period leaves respectively. The first period is the young leaf, the second period indicates the mature leaf period. (B) Expression pattern of the LcbHLHs. Expression profiles of LcbHLHs in different tissues. "XNFJ", "XNXY","XNXJ" and "XNNC" are the names of our selected materials.
Figure 6.
Expression pattern of the LcbHLHs (A) Four varieties with different colored leaves and flowers. The "F" after the name indicates the flower, and"I" and "II" represent the first period and the second period leaves respectively. The first period is the young leaf, the second period indicates the mature leaf period. (B) Expression pattern of the LcbHLHs. Expression profiles of LcbHLHs in different tissues. "XNFJ", "XNXY","XNXJ" and "XNNC" are the names of our selected materials.
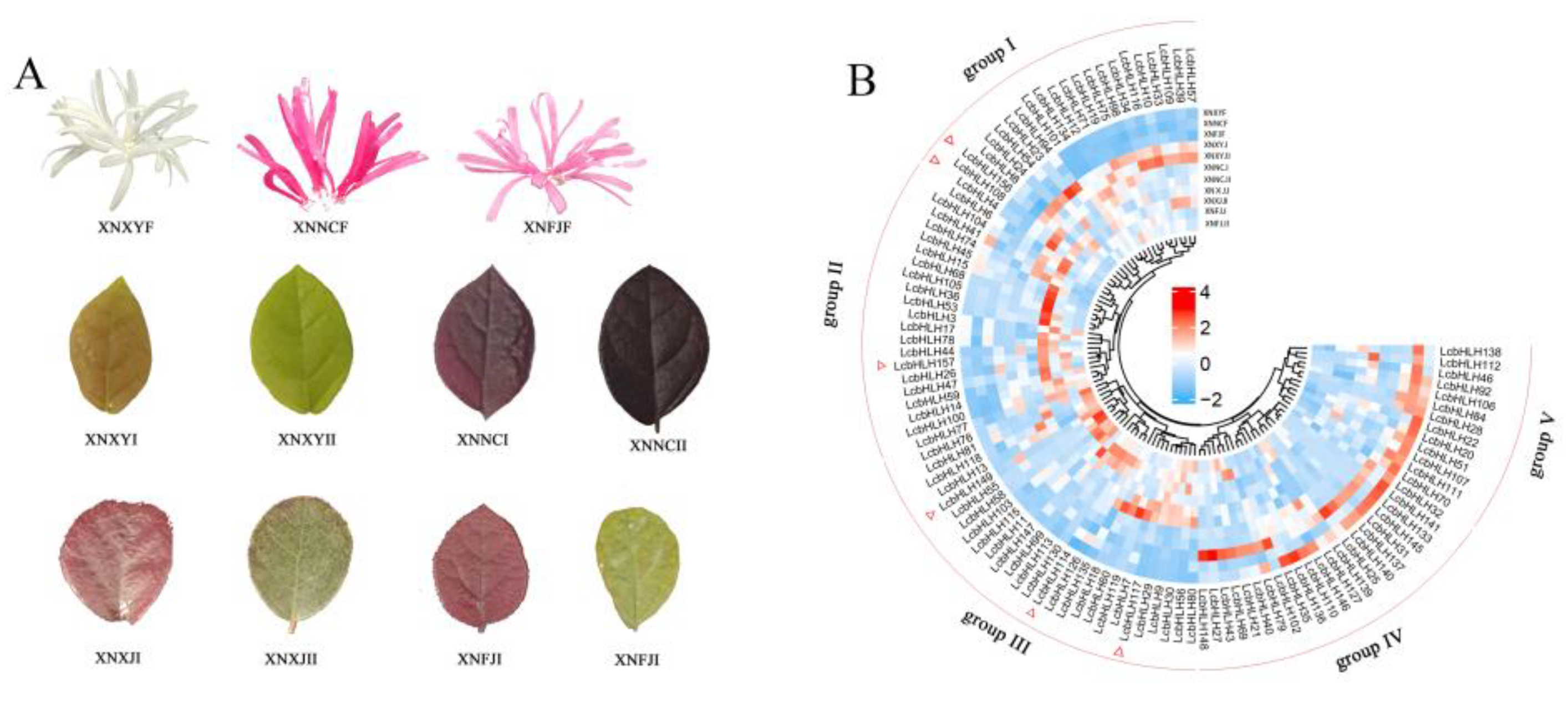
Figure 7.
Expression of six differential LcbHLHs in XNFJ variety during I and II period.
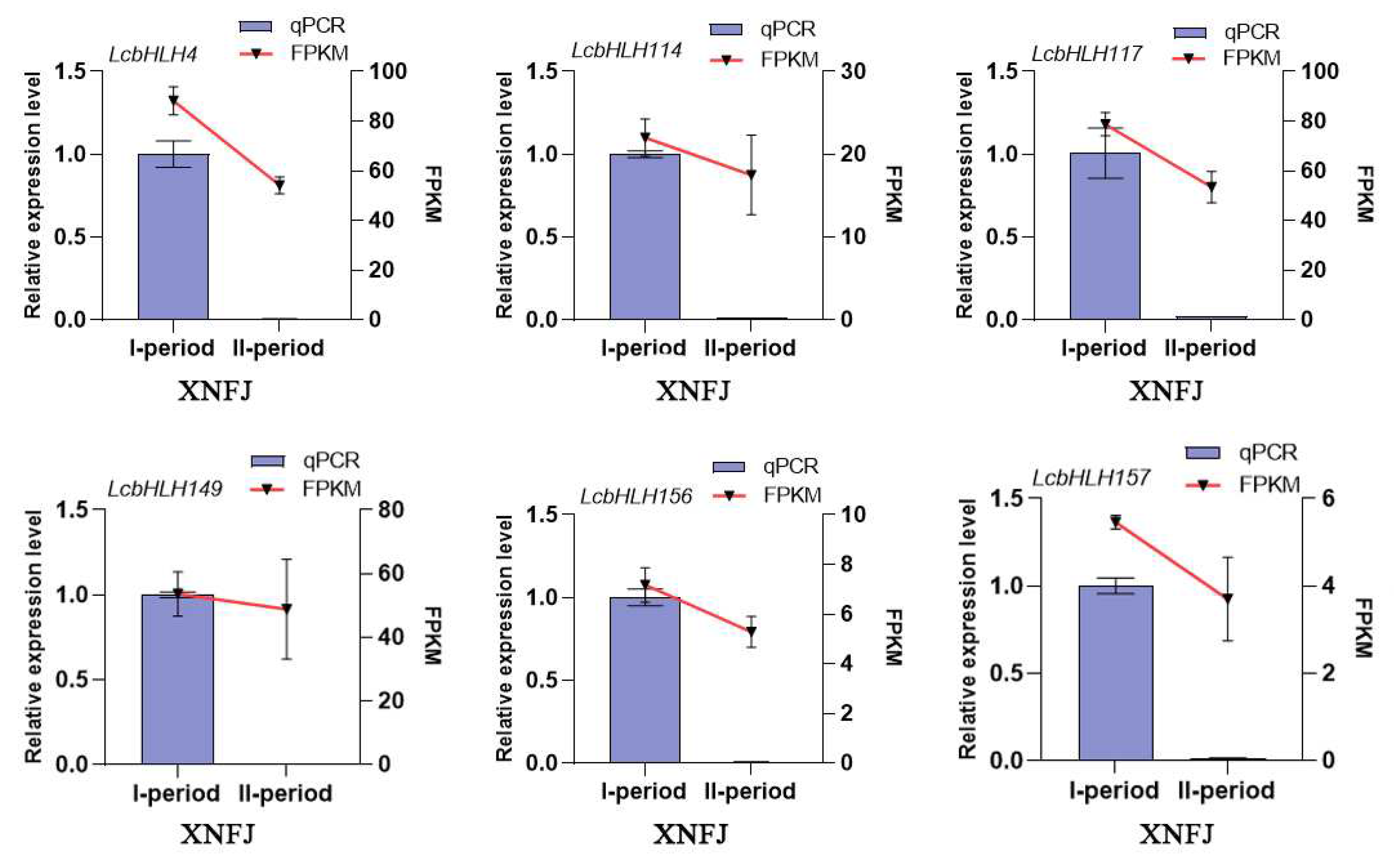
Figure 8.
Expression of six differential LcbHLHs in different color leaves (A) HYJM1 varieties with different colored leaves. GL represents green leaves, ML represents green and purple mixed leaves, and PL represents purple leaves. (B) Expression of six LcbHLHs in different color leaves (GL, ML, PL). green leaves (GL) as a control group.
Figure 8.
Expression of six differential LcbHLHs in different color leaves (A) HYJM1 varieties with different colored leaves. GL represents green leaves, ML represents green and purple mixed leaves, and PL represents purple leaves. (B) Expression of six LcbHLHs in different color leaves (GL, ML, PL). green leaves (GL) as a control group.
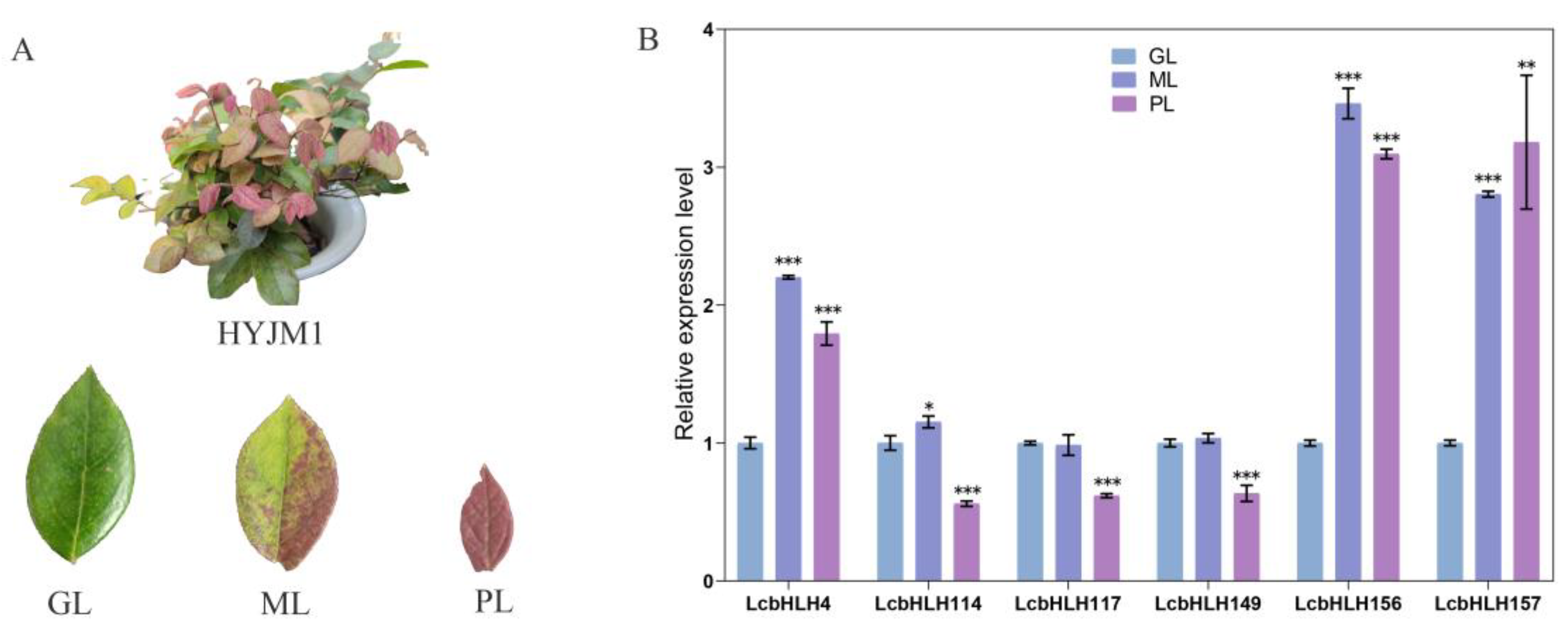
Figure 9.
Interaction network analysis for LcbHLHs. The predicted results in parentheses are based on homologous genes in Arabidopsis.
Figure 9.
Interaction network analysis for LcbHLHs. The predicted results in parentheses are based on homologous genes in Arabidopsis.
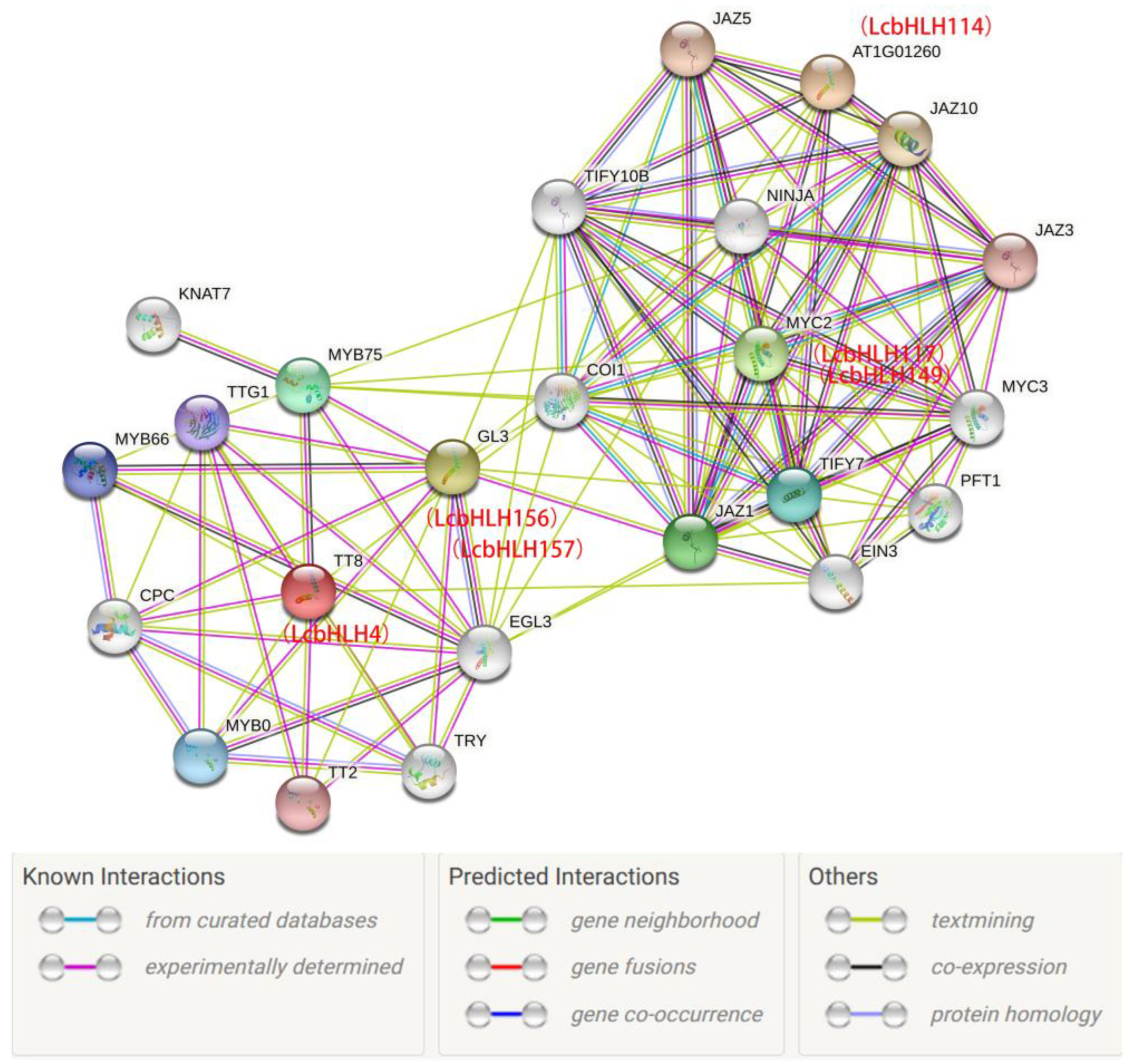

Table 1.
Quantitative distribution of each subfamily of bHLHs in L. chinense, A. thaliana, Ficus carica L. and P. persica Subfamily.
Table 1.
Quantitative distribution of each subfamily of bHLHs in L. chinense, A. thaliana, Ficus carica L. and P. persica Subfamily.
| Subfamilies | Number of LcbHLHs | Number of AtbHLHs | Number of FcbHLHs | Number of PpbHLHs |
| Ia | 12 | 10 | 8 | 8 |
| Ib | 8 | 13 | 7 | 8 |
| II | 8 | 4 | 0 | 1 |
| III(a+b+c) | 19 | 10 | 9 | 10 |
| III(d+e) | 11 | 8 | 6 | 11 |
| IIIf | 3 | 4 | 1 | 2 |
| IVa | 11 | 4 | 4 | 3 |
| IVb | 3 | 3 | 4 | 0 |
| IVc | 7 | 4 | 3 | 0 |
| IVd | 1 | 2 | 1 | 2 |
| Va | 3 | 3 | 3 | 2 |
| Vb | 8 | 5 | 6 | 6 |
| VI | 1 | 2 | 2 | 0 |
| VII(a+b) | 17 | 15 | 7 | 7 |
| VIIIa | 6 | 4 | 3 | 2 |
| VIII(b+c) | 11 | 11 | 14 | 9 |
| IX | 8 | 6 | 5 | 6 |
| X | 1 | 10 | 14 | 2 |
| XI | 5 | 5 | 4 | 4 |
| XII | 20 | 17 | 13 | 12 |
| XIII | 0 | 3 | 4 | 0 |
| XIV | 0 | 3 | 0 | 0 |
| XV | 0 | 5 | 0 | 0 |
| Orphans | 2 | 8 | 0 | 0 |
Table 2.
Sequences of 10 predicted motifs of FcbHLH proteins.
| Motif | Sequence |
|---|---|
| 1 | SHSLAERRRRERJNERFKALRSLVPNCSK |
| 2 | MDKASMLDEAIEYVKELQRQVQELSMKLE |
| 3 | EEPKSDYIHVRARRGQATD |
| 4 | QVMSFEQSNWDASVHEIQGMTSFEHPHNQDQQLHLLHEMQQNGHHHPQSF |
| 5 | FVQKPANFQTSLGFLGDLPTPDNASASSVLYDPLFHLNLPPQPPLFRDLF |
| 6 | YNLPASRTASLFGGGIDEKEGSGGVYQNGVATQFDNGVLEFTGDIGGMGK |
| 7 | RJVSALEKLGLDIVHANVSTF |
| 8 | ERAKLAKSAGIRTLVCIPTASGVVELGSTEIIKEDLGLIQLIKSLF |
| 9 | NSSTLPDTSPYINNPPTQHLLNLFHLPRCSPSSNLLPNSSI |
| 10 | DNIRLSMEELSYHQNPHQEDDAALEQHLGFDMENCYNINNN |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



