
Submitted:
21 June 2023
Posted:
22 June 2023
You are already at the latest version
Abstract
Lichens are unique extremophilic organisms due to their phenomenal resistance to adverse environmental factors, including ultraviolet (UV) irradiation. Melanization plays a special role in the protection of lichens from UV-B stress. In present study, we analyzed the binding of melanins with the components of cell walls of mycobiont of upper cortex in the melanized lichen thalli Lobaria pulmonaria. Using scanning electron and atomic force microscopy, the morphological and nanomechanical characteristics of melanized layer of mycobiont cells were visualized. Melanization of lichen thalli led to the smoothing the surface relief and thickening mycobiont cell walls and resulted in the increase of deformation values and reduction of adhesion properties of lichen thallus. Treatment of thalli with hydrolytic enzymes, especially chitinase and lichenase, enhanced the yield of melanin from melanized thalli and release of carbohydrates, while treatment with pectinase increased the release of carbohydrates and phenols. Our results suggest that melanin by firmly binding with hyphal cell wall carbohydrates, particularly chitin and 1,4-β-glucans, can form associations, strengthen the melanized upper cortex of lichen thalli, and thereby can contribute to lichen survival under UV stress.
Keywords:
lichen
; UV-induced melanization
; melanin
; topography
; hydrolytic enzymes
; microscopy
1. Introduction
Lichens are symbiotic photosynthesizing organisms, comprising a thallus formed by a fungus (the mycobiont) and an alga and or a cyanobacterium (the photobiont). These two organisms are closely linked morphologically, physiologically and biochemically [1,2,3]. Lichens may cover up to 100% of the ground at the sites where vascular plants are at their physiological limits [4]. Lichens are classified as extremophilic organisms due to their phenomenal resistance to adverse environmental factors, including ultraviolet (UV) stress [5]. Among the various defense mechanisms, the synthesis of secondary metabolites, including melanin pigments, plays a special role [6]. Melanization of the cortical layer of the lichen thallus prevents the damage of intracellular components during the exposure of lichen thallus to UV irradiation and high intensity light. Melanins are dark pigments produced through the oxidation and polymerization of phenolic or indolic precursors [7,8]. The application of a combination of transmission and scanning electron microscopy methods has shown that melanins have a granular structure and occur as dark granules of different sizes [9,10]. The microstructure of melanins has been studied in several pathogenic fungi [11,12] and human melanosomes [13,14]. In fungi, melanins can be either secreted into the external environment or accumulate in the fungal cell wall [15]. High-resolution solid-state NMR has shown that in Cryptococcus neoformans melanins are probably covalently bound to cell wall chitin [16]. A close association between melanin and chitin has also been reported for Aspergillus nidulans [17], Exophilia dermatitides [18], and Candida albicans [19]. Fungi with mutated genes that are involved in the biosynthesis of the cell-wall chitin or chitosan display a “leaky melanin” phenotype or possess enhanced pigment deposition [19]. The incorporation of melanin into the fungal cell wall decreases the pore size, reducing the conductivity of the wall e.g. to water [20,21]. In our earlier study on UV-induced melanization of the lichen Lobaria pulmonaria, we visualized stages in the formation of melanin-like granules, including the formation of melanin vesicles, their transport, aggregation, and deposition of granules in cell walls of hyphae of the upper cortex [8,10]. However, information about the ability of melanin to bind with cell wall components of lichen mycobionts is scarce. This is likely a result of the complexity of the anatomical structure of the lichen thallus, the interactions between the fungal and photosynthetic symbionts, and the diversity of metabolite biosynthesis pathways that occur in lichens. Furthermore, lichen melanins can form complexes with metals and the elements of lichen cell walls, making them difficult to study [6,22]. Previous studies on melanin complexes isolated from two lichen species used infra-red (IR) spectroscopy to reveal the presence aromatic and aliphatic functional groups [23]. It is known that treatment of cell wall with hydrolytic enzymes is a relatively mild but effective method of breaking down cell walls with high specificity [24]. For example, treatment of the fungi Inonotus hispidus with complex hydrolyzing enzymes resulted in an increased yield of extracted melanin [25].
Understanding of structure and properties of melanin associations in the cell walls is important for understanding how melanin strengthens lichen thalli and facilitates tolerance of lichen to UV stress. The aim of present study was to visualize the topography of UV-melanized thalli and to analyze the binding of melanins with the components of cell walls in Lobaria pulmonaria (L.) Hoffm., a large-leafy epiphytic lichen, which predominantly grows at a height of 1–2 m on wet bark of the lower part of the trunks of deciduous and coniferous trees. The species has an extensive range, covering Europe, Asia, Africa, North America and Australia. The thallus of the lichen L. pulmonaria represents an association of mycobiont hyphae (ascomycetes) with photobiont cells (green algae Symbiochloris reticulata and cyanobacteria Nostoc) [26,27]. Cyanobacteria of the genus Nostoc are included in special structures termed cephalodia and carry out biological fixation of atmospheric nitrogen. Sun-exposed thalli of L. pulmonaria can be dark brown, while shade-adapted thalli are normally bright green. Field experiment have shown that exposing shade-adapted L. pulmonaria to normal solar radiation induces L-DOPA melanin synthesis [28].
To study the morphological and nanomechanical characteristics of melanized layer of mycobiont cells, scanning electron microscopy (SEM) and atomic force microscopy (AFM) were applied. SEM reproduces the lateral dimensions of thalli, while AFM can provide direct information on the relief and adhesion of the surface. We hypothesized that the changes in the relief and adhesion of the surface of thalli that have melanized following exposure to UV-B result from the association of melanins with polysaccharides, especially chitin and β-glucans, and proteins in the cell walls. To test this, hydrolyzing enzymes were applied to melanized lichen thalli. The ability of the enzymes to break down the bonds was assessed by measuring the release of carbohydrates, proteins, and phenols, and the yield of melanin. Changes caused in the ultrastructure of the cells in the upper cortex following enzyme treatments were visualized using transmission electron microscopy (TEM).
2. Results
2.1. Morphology
SEM was used to study the morphological changes of the surface topography of cross-sections of L. pulmonaria thalli that accompany melanisation. The relief of the anticlinal cell walls of the pale thallus was heterogeneous and rough and characterized by a large number of depressions and bulges (Figure 1A). The anticlinal cell walls of melanized thallus were smoother, with fewer differences in height. Cell walls of the melanized thallus were visibly thicker than those in the pale thallus (Figure 1A,B).
We hypothesised that the differences in SEM images of lichen thalli may be due the association of melanins with the components of fungal hyphae cell walls, in particular carbohydrates. The presence of carbohydrates, including chitin, was qualitatively visualed using Calcofluor staining and Lugol staining of the cell walls of cortical cells in cross sections of pale and melanized L. pulmonaria thalli. Interestingly, while in the pale thalli Calcofluor staining was very bright (Figure 2B), fluorescence in melanized thallus was much weaker (Figure 2E). Lugol staining of chitin demonstrated the presence of the small light-purple grains in the upper cortex of pale thalli (Figure 2C), whereas pigmentation of the upper cortex of melanized thalli was darker (Figure 2F).
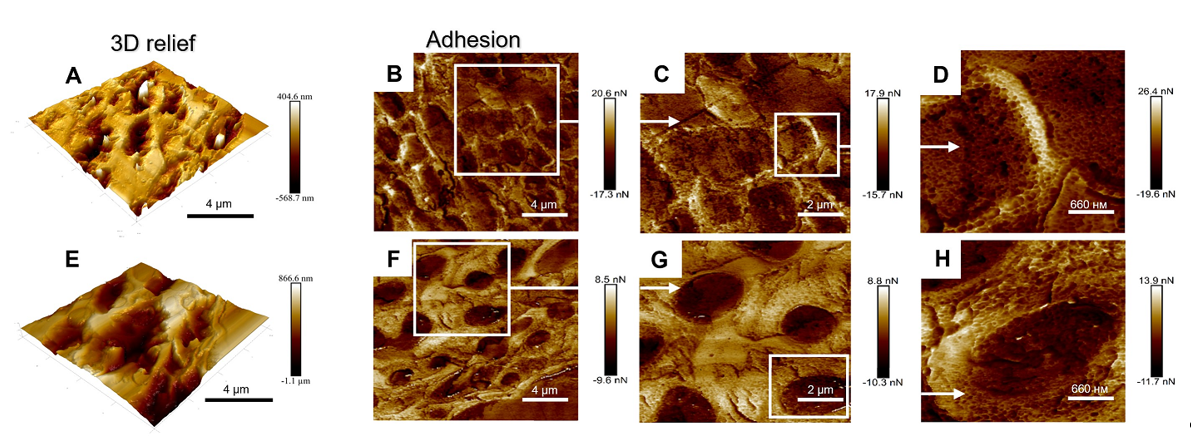
To study the topographical features of the upper cortex, we analyzed the 3D relief and adhesion of the surface of the cross-sections of Lobaria pale and melanized thalli using AFM (Figure 3). In contrast to SEM, which reproduces the lateral dimensions of thalli but does not provide direct information on their relief, the AFM accurately measures the relief or “height” and adhesion. Melanized samples displayed approximately twice the variation in height variations, while adhesion values were reduced by 0.6 compared to those of the pale samples. An increase in height variation of the upper cortex of melanized thalli measured by AFM (vertical dimension, Figure 3A,E) corresponded to an increase in their SEM widths (lateral dimension, Figure 1A,B). Visualized differences in the adhesion in pale (Figure 3B-D) and melanized thalli (Figure 3F-H) corresponded to the differences in the morphology of the cell walls in pale and melanized thalli as shown in Figure 1A,B. Thus, SEM and AFM imaging gave complementary information.
2.2. The effect of enzyme treatment on the release of components
To characterize the binding of melanins with the components of cell walls in L. pulmonaria, discs of pale and melanized lichen thalli were incubated in the solutions with enzymes that hydrolyze the bonds between melanin and carbohydrates, proteins, and phenols. The list of enzymes, their specific activity, corresponding buffers and concentrations are presented in the Table 1. After treatment of pale and melanized thalli with enzymes, the absorbance of the incubation solutions was analyzed at a wavelength 490 nm (Figure 4A,B), which corresponds to the absorbance of melanins [22] and some other phenolic compounds such as quinones [29]. After treatment of thalli with chitinase, lichenase, endoglucanase, and pectinase the absorbance of incubation solutions was low and there was no difference between pale and melanized thalli. However, treatment of the thalli with protease caused a significant increase in absorbance, especially of the incubation solution of melanized lichen (Figure 4B). Interestingly, incubating both types of lichen thalli in the buffer without protease also increased the absorbance. Following incubation in solutions containing buffer only, the absorbance of melanized thalli was more than double that of pale thalli (Figure 4A,B).
Treatment of the pale thalli with pectinase and protease increased the release of phenolic compounds into incubation solutions (Figure 4C). The release of phenolic compounds from melanized thalli was highly stimulated by pectinase (Figure 4D). Significant release of phenolic compounds from melanized thalli was observed following incubation of discs in the solutions with and without protease (Figure 4D). After the incubation of lichen thalli in the solutions containing chitinase, lichenase, and endogluconase, the release of phenolic substances to the incubation solutions was low and similar to their release into solutions containing only buffers.
Treatment of pale thalli with buffers with or without chitinase releases small amounts of carbohydrates (Figure 4E), while the release of total carbohydrates from melanized lichen thalli doubled after treatment with chitinase (Figure 4F). The treatment of pale and melanized lichen thalli with buffers with and without lichenase greatly increased the release of carbohydrates (Figure 4E,F). The release of carbohydrates was not observed after treatment of pale and melanized thalli with endogluconase and protease, while the incubation with pectinase resulted in the significant release of carbohydrate (Figure 4E,F).
Measuring the melanin after enzyme treatments showed that general contents were lower in pale than melanized thalli (Figure 5A,B). After treatment of melanized thalli with chitinase and especially lichenase the yield of melanin was much higher compared to that of solutions that contained only buffer (Figure 5B). Endoglucanase marginally increased the amount of melanin that could be extracted, while pectinase and protease faciliated the increase of more melanins from both pale and melanized thalli.
2.3. The ultrastructural characteristics of the cross sections of melanized lichen thalli
After treatment of melanized L. pulmonaria thalli with chitinase, endoglucanase, pectinase, and protease we analyzed the ultrastructural properties of cross sections of these thalli by TEM (Figure 6). In melanized thallus, the fungal hyphae had thick cell walls with a clearly visualized melanic layer and an intact plasma membrane (Figure 6A).
Treatment of the thalli with enzymes caused significant changes in the cellular ultrastructure. In particular, layering and loosening of the cell walls (Figure 6С-E), and decondensation of interhyphal space (Figure 6С,D) were observed. Furthermore, the integrity of the plasma membrane was disrupted (Figure 6B,E), and it became detached from the cell wall of the hyphae; significant plasmolysis of the cellular contents were visualized (Figure 6B-E).
3. Discussion
Lichens are extremophilic organisms that successfully survive in hostile environments [5]. The ability of these fungi to synthesize darkly pigmented melanins contributes to the high tolerance of lichens to UV and desiccation [30,31]. In present study, by applying advanced imaging techniques such as AFM we discovered that UV-B irradiation causes changes in the topography of thalli of the lichen L. pulmonaria. Following UV irradiation, L. pulmonaria becomes melanized, and compared to pale thalli, anticlinal and periclinal cell walls of melanized upper cortex become visibly thicker and display a smoother relief (Figure 1). In our previous paper, we showed that UV induces the formation of melanin-like granules in the hyphae in the melanized upper cortex of lichen thallus [10]. We suggest that the changes in morphology of thallus surface during melanization result from the formation of melanin complex polymers in the cell walls of the fungal hyphae in the upper cortex.
Studying the structure of melanins is challenging, because they are complex polymers, insoluble, amorphous, and heterogeneous, comprising a mixture of proteins, carbohydrates, and lipid moieties [32,33]. In free-living melanized fungi, NMR, X-ray scattering, electron microscopy and ASM have been used to study the molecular structure of melanins, including the covalent binding of melanins to polysaccharides, chitin/chitosan, and lipids. These compounds appear to serve as scaffolds for melanin synthesis [16,34]. Wall-bound electron-dense melanin granules may occur in the outer or inner part of the cell walls of fungal hyphae [35]. In melanized thalli of Lobaria the upper cortex is visibly much darker than that of pale thalli (Figure 2A,D). To visualize the association of melanin with other components of the hyphal cell walls, qualitative stains such as Calcofluor for carbohydrates and Lugol for chitin, were used. Compared to melanised thalli, pale thalli displayed more intense Calcofluor staining in the upper cortex (Figure 2B,E). It seems likely that bonding between melanins and polysaccharides impedes the normal interaction of the stain with its target molecules. In cross sections of the melanized thalli of L. pulmonaria, staining with Lugol results in a dark purple stippling (Figure 2C,F), indicating the presence of chitin in the upper cortex. Chitin, a long-chain polymer of N-acetylglucosamine, provides mechanical strength to fungal cell walls and is therefore an important structural component of the cell wall [36,37]. As discussed in the Introduction, several studies suggest that chitin is a primary effector for melanin polymer deposition within the fungal cell wall [38,39,40,41]. For example, deletion of a gene involved in chitin synthesis in the black fungus Exophilia (Wangiella) dermatitidis results in significantly reduced melanin deposition within the cell wall [18].
Investigating the mechanical properties of tissues at the cellular level by AFM can greatly assist understanding the processes of growth and morphogenesis [42]. For example, in maize primary roots stiffness and elasticity of the stele vascular parenchyma periclinal cell walls are correlated with cell wall mechanical properties [43]. There have been several studies on the conformation of melanins using ASM, for example the ultrastructural characteristics of eumelanin from Sepia officinalis [44,45]. In present study, imaging the topography and mechanical properties of melanins in the thallus of L. pulmonaria by ASM demonstrates that UV-B induced melanization changes the physical parameters of the upper cortex. The height of the surface of melanized samples increases, while adhesion is reduced compared to that in the pale thalli (Figure 3). Adhesion is an integral parameter comprising both the adherence of the thalli to AFM probe and the contribution of the interfacial water film. Yet, we see that the adhesion force is different for these two types of the thalli. This observation indirectly supports the difference in the topography of two thalli at the nanoscale. In the green alga Enteromorpha linza (L.), AFM was used to show that glycoprotein is a natural adhesive, and provides firm anchorage to the substratum [46]. It is likely that the changes in the height and adhesion of the surface of melanized samples result from the association of melanins with polysaccharides and proteins in the cell walls.
To study the nature of the link between melanins and cell wall components, we treated pale and melanized lichen thalli with chitinase, lichenase, endoglucanase, pectinase, and protease. These enzymes can hydrolyze the glucosidic bonds of structural carbohydrates of the cell wall, and also proteins (Table 1). Treatment with pectinase causes significant release of carbohydrates and phenols, although pectin substances are not common in lichens (Figure 4). However, some lichens, e.g. Evernia prunastri [47] and Peltigera canina [48], contain pectinases, specifically polygalacturonase, which may indicate that some pectins are present. L. pulmonaria may contain pectin or pectin-like polysaccharides that can be target molecules for pectinase. Interestingly, incubation in buffer solutions with a pH of 7.4 can induce the release of phenolic compounds from melanized lichen thalli, possibly indicating that they are soluble in alkaline solutions [49,50].
Lichen polysaccharides are mainly linear or weakly substituted α- or β-glucans [51]. Therefore, it is not surprising that treatment of melanized lichen thalli with chitinase and lichenase increases the release of carbohydrates (Figure 4E,F). Furthermore, the yield of melanin from melanized thalli was much higher following treatment with chitinase and even more so lichenase, which hydrolyzes 1,4-β-glucosidic bonds (Figure 5). By contrast, endoglucanase, which hydrolyzes 1,3-β-glucosidic bonds, did not increase in the yield of melanin extracted (Figure 5). Therefore, these results suggest the presence of chitin and 1,4-β-glucans in the cell wall of mycobiont. Protease only had a small effect on the release of melanin. It seems likely that melanin is strongly associated with the structural components of the cell wall that are targeted by hydrolytic enzymes. Hydrolytic enzymes also cause changes in the ultrastructure of hyphal cells of the upper cortex, such as layering and loosening of the cell walls, decondensation of interhyphal space and significant plasmolysis of the cellular content (Figure 6). Taken together, the observed ultrastructural changes and the effects of hydrolytic enzymes on the release of compounds from the thalli strongly suggest that bonds exist between melanin and structural polysaccharides in the cell wall. Future research is required to determine the structure, morphology, and physico-chemical properties of isolated associations of lichen melanins with cell wall polysaccharides.
4. Materials and Methods
4.1. Sample Preparation
Pale and melanized thalli of L. pulmonaria (L.) Hoffm. were collected from the bark of poplar trees growing in the outskirts of Syktyvkar, Komi Republic, Russia (latitude 61’34' N, longitude 50’33' E). Lichen material was cleaned, slowly dried at room temperature and then stored at -20oC. Before experiments, 1 cm disks were cut, their dry mass determined and then placed on wet filter paper and hydrated in a climate chamber at +15oС and 12/12 h (light/dark). The thalli were irradiated by UV-B (λ=280–315 nm) fluorescent erythema lamps (3 W m-2) for 80 min daily for 14 d. During this time, the controls were taken out of the climate chamber and kept under regular laboratory conditions without UV irradiation.
4.2. Scanning Electron Microscopy
To image melanin of L. pulmonaria by scanning and transmission electron microscopy thalli were embedded in 3% agarose blocks and cut using a vibratome (Leica VT 1000S, Wetzlar, Germany) resulting in 50 µm cross sections from the upper cortex. The cross sections of pale and melanized lichen thalli were subsequently fixed in 2.5% glutaraldehyde in 0.1 M Na phosphate buffer, pH 7.4 and 1% osmium tetroxide, and further dehydrated as described by Daminova et al. (2022) [10]. Sections were then sputter-coated with gold using the Q150T ES Coater (Quorum Technologies, Lewes, UK) and anticlinal and periclinal cell walls were viewed using a high-resolution scanning electron microscope (Merlin, Carl Zeiss, Oberkochen, Germany) at a voltage of 5 kV.
4.3. Atomic Force Microscopy
Disks of lichen thalli were fixed according to standard protocols [52]. Briefly, the samples were fixed in glutaraldehyde (Sigma, USA), post-fixed in osmium tetroxide (Sigma, USA), dehydrated in a graded aqueous ethanol series and acetone, embedded in LR White resin (Medium Grade Acrylic Resin; Ted Pella, Redding, CA, USA) and polymerized at 60°C for 24 h. Samples were carefully inserted in a vertical orientation in beam capsules, which were then filled with LR White resin before polymerization [53]. The top of the sample blocks was leveled using a diamond knife (Electron Microscopy Sciences, USA). The upper cortex was visualized using the Bruker Dimension FastScan microscope (Bruker, Billerica, MA, USA) in PeakForceQNM (quantitative nanomechanical mapping) mode [54]. To obtain high-quality images of the topography and nanomechanical characteristics of the sections, the standard silicon cantilevers ScanAsyst-Air (Bruker) with curvature 2 nm and stiffness 0.4 N m−1 were used. Images were acquired at a resolution of 512 lines per scan [55].
4.4. Light Microscopy
For light microscopy, pale and melanized lichen thalli were incubated in 2 M NaOH for 1 h at 100°C, the pH adjusted to 7.0 with 5 M sulfuric acid and then thalli rinsed three times with distilled water. Cross-sections were vibratome-cut, stained with Lugol solution and 10% sulfuric acid as described by Yajima et al. (2001) [56], and viewed with a bright field epifluorescence microscope (Leica DM1000, Leica Biosystems, Wetzlar, Germany).
For fluorescence microscopy, cross-sections of pale and melanized thalli were stained with 0.1% Calcofluor White (Sigma-Aldrich, USA) for 15 min in the dark and visualized using an excitation wavelength of 380 nm and an emission wavelength of 475 nm using the epifluorescence microscope Leica DM1000. Images were taken by digital camera at a magnification of x 40.
4.5. Enzyme Treatments
Hydrated discs (on average c. 1.1 g) of L. pulmonaria thalli were vacuum infiltrated in 3 ml solution of the enzymes chitinase, lichenase, endoglucanase, pectinase, and protease (Table 1) three times for 1 min. After that, discs in solutions were shaken in the incubator (ES-20/60, BioSan, Riga, Latvia) at 170 rpm for 12-14 h in the dark at 37-40°С for lichenase and at 25°С for chitinase, endogluconase, pectinase, and protease. As a control, hydrated discs of lichen thalli were incubated in 3 ml of the corresponding buffer without enzyme.
4.6. Determination of the Content of Compounds Released into Incubation Solutions
After enzyme treatment, discs of lichen thalli were removed from the incubation solutions, rinsed with 5 ml of distilled water, and gently blotted with filter paper. Incubation solutions were centrifuged at 8000 rpm for 1.5 min using an Eppendorf MiniSpin (Eppendorf, Germany). The absorbance of the incubation solution was determined at 490 nm using the UV-1900 spectrophotometer (Shimadzu, Kyoto, Japan). The content of the total phenolic compounds was determined using the Folin-Ciocalteu reagent (Sigma-Aldrich, St. Louis, Missouri, USA) [57], calibrated with gallic acid (20-200 µg ml-1) (DiaM, Russia). The content of non-reducing sugars was determined using the anthrone method [58], calibrated with glucose solutions (20-100 µg ml-1) (Reakhim, Russia).
4.7. Content of Melanin in Lichen Thalli
Alkali-soluble metabolites were extracted from the thallus discs of L. pulmonaria after enzyme treatment using the protocol outlined by [50] with modifications. Initially discs were homogenized with liquid N2 and then 2 M NaOH added in the ratio 1:10. After alkaline extraction for 12 h at room temperature the resulting homogenate was centrifuged at 8 000 rpm for 10 min (Hermle, Gosheim, Germany). Melanin was precipitated from the alkaline extracts by adding HCl till pH 2 and incubating for 12 h at room temperature. After centrifugation the resulting precipitate was oven-dried at 60°C for 1 h and weighed. Melanin content was calculated in mg ml-1 of alkali extracts.
4.8. Transmission Electron Microscopy
The discs of melanized lichen thalli of L. pulmonaria after enzyme treatment were fixed according to the standard protocol (see above section AFM). After ultra-thin sectioning of samples using an ultramicrotome (Leica, Wetzlar, Germany), cross-sections were stained with 2% aqueous uranyl acetate (w/v) for 20 min and Reynolds’ lead citrate for 7 min [59]. Finally, the cross-sections were examined using the Excellence transmission electron microscope (Hitachi HT 7700, Tokyo, Japan) at an accelerating voltage of 100 kV.
4.9. Statistical Analysis
Microscopical observations were performed using at least 6 representative samples, while all biochemical measurements were conducted in triplicate (n=3). The values are expressed as mean ± standard deviation (SD). P values less than 0.05 (p < 0.05) were considered as statistically significant.
5. Conclusions
The present study demonstrates that UV-B irradiation induces melanization and changes the topography of the lichen L. pulmonaria thalli. Accumulation of melanin results in a smoother relief with visibly thicker cell walls of the mycobiont upper cortex. Furthermore, melanization increases the height value and reduces the adhesion properties of the thallus. Treatment of thalli with hydrolytic enzymes, especially chitinase, lichenase, and pectinase, enhances the yield of melanin and the release of carbohydrates and phenols from melanized thalli, confirming the existence of bonds between melanins and cell wall components. Results increase our understanding of the structure and properties of melanin associations in the cell wall of fungal hyphae of lichen thalli.
Author Contributions
Conceptualization, A.G.D. and F.V.M.; methodology, A.G.D. and V.R.Kh.; investigation, A.G.D., A.E.R., and V.R.Kh.; writing—original draft preparation, A.G.D.; writing—review and editing, R.P.B. and F.V.M.; project administration, F.V.M.; funding acquisition, A.G.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Russian Science Foundation, grant number 21-74-00153 (for A.G.D).
Acknowledgments
This work was partially performed within the framework of the state assignment of the FRC KazSC RAS. The SEM, ASM, and TEM images were recorded on the equipment of the Interdisciplinary Center for Analytical Microscopy of Kazan Federal University. Dr. Anna Petrova (Kazan Institute of Biochemistry and Biophysics), Dr. Valery Prokhorov (The Institute of Physical Chemistry and Electrochemistry RAS), and Mr. Alexey Rogov (KFU) are thanked for their highly professional consulting on fluorescence microscopy, AFM and SEM. The help of Dr. Mikhail Shelyakin (Institute of Biology, Komi Science Center, Russia) in collecting lichens is much appreciated.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Piercey-Normore, M.D.; Deduke, C. Fungal farmers or algal escorts: lichen adaptation from the algal perspective. Mol. Ecol. 2011, 20, 3708–3710. [Google Scholar] [CrossRef]
- Insarova, I.D.; Blagoveshchenskaya, E.Y. Lichen symbiosis: search and recognition of partners. Izv. Akad. Nauk Ser. Biol. 2016, 5, 479–490. [Google Scholar] [CrossRef]
- Pichler, G.; Muggia, L.; Carniel, F.C.; Grube, M.; Kranner, I. How to build a lichen: from metabolite release to symbiotic interplay. New Phytol. 2023, 238, 1362–1378. [Google Scholar] [CrossRef] [PubMed]
- Honegger, R. Functional aspects of the lichen symbiosis. Annu. Rev. Plant Biol. 1991, 42, 553–578. [Google Scholar] [CrossRef]
- Nash, T.H. , III (Ed.) Lichen Biology; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Mafole, T.C.; Solhaug, K.A.; Minibayeva, F.V.; Beckett, R.P. Tolerance to photoinhibition within a lichen species is higher in melanised thalli. Photosynthetica 2019, 57, 96–102. [Google Scholar] [CrossRef]
- Ndhlovu, N.T.; Solhaug, K.A.; Minibayeva, F.; Beckett, R.P. Melanisation in boreal lichens is accompanied by variable changes in non-photochemical quenching. Plants 2022, 11, 2726. [Google Scholar] [PubMed]
- Mafole, T.C.; Solhaug, K.A.; Minibayeva, F.V.; Beckett, R.P. Occurrence and possible roles of melanic pigments in lichenized ascomycetes. Fungal Biol. Rev. 2019, 33, 159–165. [Google Scholar] [CrossRef]
- Caesar-Tonthat, T.; Van Ommen, K.F.; Geesey, G.G.; Henson, J.M. Melanin production by a filamentous soil fungus in response to copper and localization of copper sulfide by sulfide-silver staining. Appl. Environ. Microbiol. 1995, 61, 1968–1975. [Google Scholar]
- Daminova, A.G.; Rogov, A.M.; Rassabina, A.E.; Beckett, R.P.; Minibayeva, F.V. Effect of melanization on thallus microstructure in the lichen Lobaria pulmonaria. J. Fungi. 2022, 8, 791. [Google Scholar]
- Babitskaya, V.G.; Scherba, V.V.; Ikonnikova, N.V.; Bisko, N.A.; Mitropolskaya, N.Y. Melanin complex from medicinal mushroom Inonotus obliquus (Pers.: Fr.) Pilat (Chaga)(Aphyllophoromycetidae). Int. J. Med. Mushrooms 2002, 4, 139–145. [Google Scholar]
- Eisenman, H.C.; Casadevall, A. Synthesis and assembly of fungal melanin. Appl. Microbiol. Biotechnol. 2012, 93, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, R.L.; Double, K.L.; Gerber, J.P. Using Sepia melanin as a PD model to describe the binding characteristics of neuromelanin–A critical review. J. Chem. Neuroanat. 2015, 64, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Wakamatsu, K.; Fujikawa, K.; Zucca, F.A.; Zecca, L.; Ito, S. The structure of neuromelanin as studied by chemical degradative methods. J. Neurochem. 2003, 86, 1015–1023. [Google Scholar] [CrossRef]
- Butler, M.J.; Day, A.W. Destruction of fungal melanins by ligninases of Phanerochaete chrysosporium and other white rot fungi. Int. J. Plant Sci. 1998, 159, 989–995. [Google Scholar] [CrossRef]
- Camacho, E.; Vij, R.; Chrissian, C.; Prados-Rosales, R.; Gil, D.; O'Meally, R.N.; Cordero, R.J.B.; Cole, R.N.; McCaffery, J.M.; Stark, R.E.; Casadevall, A. The structural unit of melanin in the cell wall of the fungal pathogen Cryptococcus neoformans. J. Biol. Chem. 2019, 294, 10471–10489. [Google Scholar] [CrossRef] [PubMed]
- Bull, A.T. Chemical composition of wild-type and mutant Aspergillus nidulans cell walls. The nature of polysaccharide and melanin constituents. J. Gen. Microbiol. 1970, 63, 75–94. [Google Scholar] [CrossRef]
- Wang, Z.; Zheng, L.; Hauser, M.; Becker, J.M.; Szaniszlo, P.J. WdChs4p, a homolog of chitin synthase 3 in Saccharomyces cerevisiae, alone cannot support growth of Wangiella (Exophiala) dermatitidis at the temperature of infection. Infect. Immun. 1999, 67, 6619–6630. [Google Scholar] [CrossRef]
- Walker, C.A.; Gómez, B.L.; Mora-Montes, H.M.; Mackenzie, K.S.; Munro, C.A.; Brown, A.J.; Gow, N.A.; Kibbler, C.C.; Odds, F.C. Melanin externalization in Candida albicans depends on cell wall chitin structures. Eukaryot. Cell. 2010, 9, 1329–1342. [Google Scholar] [CrossRef]
- Jacobson, E.S.; Ikeda, R. Effect of melanization upon porosity of the cryptococcal cell wall. Med. Mycol. J. 2005, 43, 327–333. [Google Scholar] [CrossRef]
- Chrissian, C.; Camacho, E.; Fu, M.S.; Prados-Rosales, R.; Chatterjee, S.; Cordero, R.J.B.; Lodge, J.K.; Casadevall, A.; Stark, R.E. Melanin deposition in two Cryptococcus species depends on cell-wall composition and flexibility. J. Biol. Chem. 2020, 295, 1815–1828. [Google Scholar] [CrossRef]
- Rassabina, A.; Khabibrakhmanova, V.; Babaev, V.; Daminova, A.; Minibayeva, F. Melanins from the lichens Lobaria pulmonaria and Lobaria retigera as eco-friendly adsorbents of synthetic dyes. Int. J. Mol. Sci. 2022, 23, 15605. [Google Scholar] [CrossRef] [PubMed]
- Rassabina, A.E.; Gurjanov, O.P.; Beckett, R.P.; Minibayeva, F.V. Melanin from the lichens Cetraria islandica and Pseudevernia furfuracea: structural features and physicochemical properties. Biochemistry (Moscow) 2020, 85, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Hou, R.; Liu, X.; Xiang, K.; Chen, L.; Wu, X.; Lin, W.; Zheng, M.; Fu, J. Characterization of the physicochemical properties and extraction optimization of natural melanin from Inonotus hispidus mushroom. Food Chem. 2019, 277, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Xue, F.; Xu, H.; Yuan, Y.; Wu, X.; Zhang, J.; Fu, J. Optimization of solid-state fermentation extraction of Inonotus hispidus fruiting body melanin. Foods 2021, 10, 2893. [Google Scholar] [CrossRef] [PubMed]
- McEvoy, M. , Gauslaa, Y. , Solhaug, K.A. Changes in pools of depsidones and melanins, and their function, during growth and acclimation under contrasting natural light in the lichen Lobaria pulmonaria. New Phytol. 2007, 175, I 271–282. [Google Scholar]
- Gauslaa, Y.; Solhaug, K.A. Fungal melanins as a sun screen for symbiotic green algae in the lichen Lobaria pulmonaria. Oecologia 2001, 126, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Matee, L.P.; Beckett, R.P.; Solhaug, K.A.; Minibayeva, F.V. Characterization and role of tyrosinases in the lichen Lobaria pulmonaria (L.) Hoffm. Lichenologist 2016, 48, 311–322. [Google Scholar] [CrossRef]
- Nguyen, K.H.; Chollet-Krugler, M.; Gouault, N.; Tomasi, S. UV-protectant metabolites from lichens and their symbiotic partners. Nat. Prod. Rep. 2013, 30, 1490–1508. [Google Scholar] [CrossRef]
- Butler, M.J.; Gardiner, R.B.; Day, A.W. Melanin synthesis by Sclerotinia sclerotiorum. Mycologia 2009, 101, 296–304. [Google Scholar] [CrossRef]
- Toledo, A.V.; Franco, M.E.E.; Lopez, S.M.Y.; Troncozo, M.I.; Saparrat, M.C.N.; Balatti, P.A. Melanins in fungi: Types, localization and putative biological roles. Physiol. Mol. Plant Pathol. 2017, 99, 2–6. [Google Scholar] [CrossRef]
- Bell, A.A.; Wheeler, M.H. Biosynthesis and functions of fungal melanins. Ann. Rev. Phytopathol. 1986, 24, 411–451. [Google Scholar] [CrossRef]
- Zajac, G.W.; Gallas, J.M.; Cheng, J.; Eisner, M.; Moss, S.C.; Alvarado-Swaisgood, A.E. The fundamental unit of synthetic melanin: a verification by tunneling microscopy of X-ray scattering results. Biochim. Biophys. Acta 1994, 1199, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Eisenman, H.C.; Nosanchuk, J.D.; Webber, J.B.W.; Emerson, R.J.; Camesano, T.A.; Casadevall, A. Microstructure of cell wall-associated melanin in the human pathogenic fungus Cryptococcus neoformans. Biochem. 2005, 44, 3683–3693. [Google Scholar] [CrossRef]
- Tudor, D.; Robinson, S.C.; Krigstin, T.L.; Cooper, P.A. Microscopic investigation on fungal pigment formation and its morphology in wood substrates. Open Microbiol. J. 2014, 8, 174–186. [Google Scholar]
- Abo Elsoud, M.M.; El Kady, E.M. Current trends in fungal biosynthesis of chitin and chitosan. Bull. Natl. Res. Cent. 2019, 43, 59. [Google Scholar] [CrossRef]
- Brown, H.E.; Esher, S.K.; Alspaugh, J.A. Chitin: A "Hidden figure" in the fungal cell wall. Curr. Top. Microbiol. Immunol. 2020, 425, 83–111. [Google Scholar] [PubMed]
- Banks, S.C.; Lindenmayer, D.B.; Ward, S.J.; Taylor, A.C. The effects of habitat fragmentation via forestry plantation establishment on spatial genotypic structure in the small marsupial carnivore, Antechinus agilis. Mol. Ecol. 2005, 14, 1667–1680. [Google Scholar] [CrossRef] [PubMed]
- Walton, F.J.; Idnurm, A.; Heitman, J. Novel gene functions required for melanization of the human pathogen Cryptococcus neoformans. Mol. Microbiol. 2005, 57, 1381–1396. [Google Scholar] [CrossRef]
- Baker, L.G.; Specht, C.A.; Donlin, M.J.; Lodge, J.K. Chitosan, the deacetylated form of chitin, is necessary for cell wall integrity in Cryptococcus neoformans. Eukaryot. Cell 2007, 6, 855–867. [Google Scholar] [CrossRef] [PubMed]
- Nosanchuk, J.D.; Stark, R.E.; Casadevall, A. Fungal melanin: what do we know about structure? Front. Microbiol. 2015, 6, 1463. [Google Scholar] [CrossRef]
- Routier-Kierzkowska, A.L.; Smith, R.S. Measuring the mechanics of morphogenesis. Curr. Opin. Plant Biol. 2013, 16, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Petrova, A.A.; Kozlova, L.V.; Gaifullina, I.Z.; Ananchenko, B.A.; Martinson, E.A.; Mikshina, P.V.; Gorshkova, T.A. AFM analysis reveals polymorphism of purified flax rhamnogalacturonans I of distinct functional types. Carbohyd. Polym. 2019, 216, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Clancy, C.M.; Simon, J.D. Ultrastructural organization of eumelanin from Sepia officinalis measured by atomic force microscopy. Biochem. 2001, 40, 13353–13360. [Google Scholar] [CrossRef] [PubMed]
- Watt, A.A.R.; Bothma, J.P.; Meredith, P. The supramolecular structure of melanin. Soft Matter 2009, 19, 3754–3760. [Google Scholar] [CrossRef]
- Callow, M.E.; Callow, J.A.; Ista, L.K.; Coleman, S.E.; Nolasco, A.C.; López, G.P. Use of self-assembled monolayers of different wettabilities to study surface selection and primary adhesion processes of green algal (Enteromorpha) zoospores. Appl. Environ. Microbiol. 2000, 66, 3249–3254. [Google Scholar] [CrossRef]
- Yagüe, E.; Orus, M.I.; Estevez, M.P. Extracellular polysaccharidases synthesized by the epiphytic lichen Evernia prunastri (L.) Ach. Planta 1984, 160, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Beckett, R.P.; Zavarzina, A.G.; Liers, C. Oxidoreductases and cellulases in lichens: possible roles in lichen biology and soil organic matter turnover. Fungal Biol. 2013, 117, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Zhang, X.; Sun, S.; Zhang, L.; Shan, S.; Zhu, H. Production of natural melanin by Auricularia auricula and study on its molecular structure. Food Chem. 2016, 190, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Pralea, I.E.; Moldovan, R.C.; Petrache, A.M.; Ilieș, M.; Hegheș, S.C.; Ielciu, I.; Nicoară, R.; Moldovan, M.; Ene, M.; Radu, M.; Uifălean, A.; Iuga, C.A. From extraction to advanced analytical methods: the challenges of melanin analysis. Int. J. Mol. Sci. 2019, 20, 3943. [Google Scholar] [CrossRef]
- Olafsdottir, E.S.; Ingólfsdottir, K. Polysaccharides from lichens: structural characteristics and biological activity. Planta Med. 2001, 67, 199–208. [Google Scholar] [CrossRef]
- Mattson, J.C.; Borgerding, P.J.; Craft, D.L. Fixation of platelets for scanning and transmission electron microscopy. Stain Technol. 1977, 52, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Petrova, A.; Kozlova, L. Characterizing mechanical properties of primary cell wall in living plant organs using atomic force microscopy. J. Vis. Exp. 2022, 183, e63904. [Google Scholar]
- Nikolaeva, V.; Kamalov, M.; Abdullin, T.I.; Salakhieva, D.; Chasov, V.; Rogov, A.; Zoughaib, M. Evaluation of GHK peptide–heparin interactions in multifunctional liposomal covering. J. Liposome Res. 2023, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Akhatova, F.; Ishmukhametov, I.; Fakhrullina, G.; Fakhrullin, R. Nanomechanical atomic force microscopy to probe cellular microplastics uptake and distribution. Int. J. Mol. Sci. 2022, 23, 806. [Google Scholar] [CrossRef]
- Yajima, H.; Morita, M.; Hashimoto, M.; Sashiwa, H.; Kikuchi, T.; Ishii, T. Complex formation of chitosan with iodine and its structure and spectroscopic properties--molecular assembly and thermal hysteresis behavior. Int. J. Thermophys. 2001, 22, 1265–1283. [Google Scholar] [CrossRef]
- Henríquez, C.; Almonacid, S.; Chiffelle, I.; Valenzuela, T.; Araya, M.; Cabezas, L.; Simpson, R.; Speisky, H. Determination of antioxidant capacity, total phenolic content and mineral composition of different fruit tissue of five apple cultivars grown in Chile. Chil. J. Agric. Res. 2010, 70, 523–536. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Nazarova, A.V. Polysaccharides from Arctostaphyllos uva-ursi. Chem. Nat. Compd. 2009, 45, 702–704. [Google Scholar] [CrossRef]
- Reynolds, E.S. The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J. Cell Biol. 1963, 17, 208–212. [Google Scholar] [CrossRef]
Figure 1.
SEM images of the anticlinal (stars) and periclinal cell walls (triangles) of cross-sections of pale (A) and melanized (B) thalli of L. pulmonaria. Scale bar corresponds to 10 µm.
Figure 1.
SEM images of the anticlinal (stars) and periclinal cell walls (triangles) of cross-sections of pale (A) and melanized (B) thalli of L. pulmonaria. Scale bar corresponds to 10 µm.
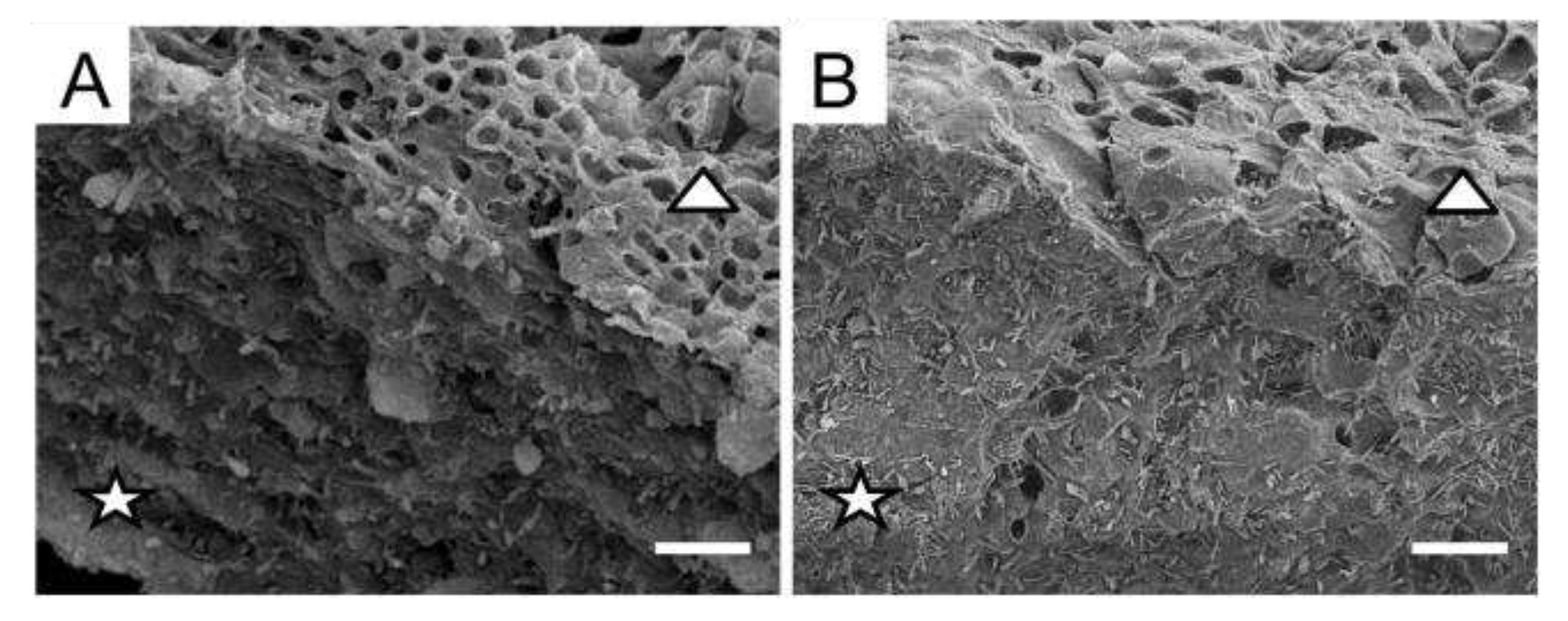
Figure 2.
Cross sections of pale (A-C) and melanized (D-F) thalli of L. pulmonaria: non-stained cross-sections (A,D); Calcofluor staining (B,E); Lugol staining (C,F). Scale bar corresponds to 25 µm.
Figure 2.
Cross sections of pale (A-C) and melanized (D-F) thalli of L. pulmonaria: non-stained cross-sections (A,D); Calcofluor staining (B,E); Lugol staining (C,F). Scale bar corresponds to 25 µm.
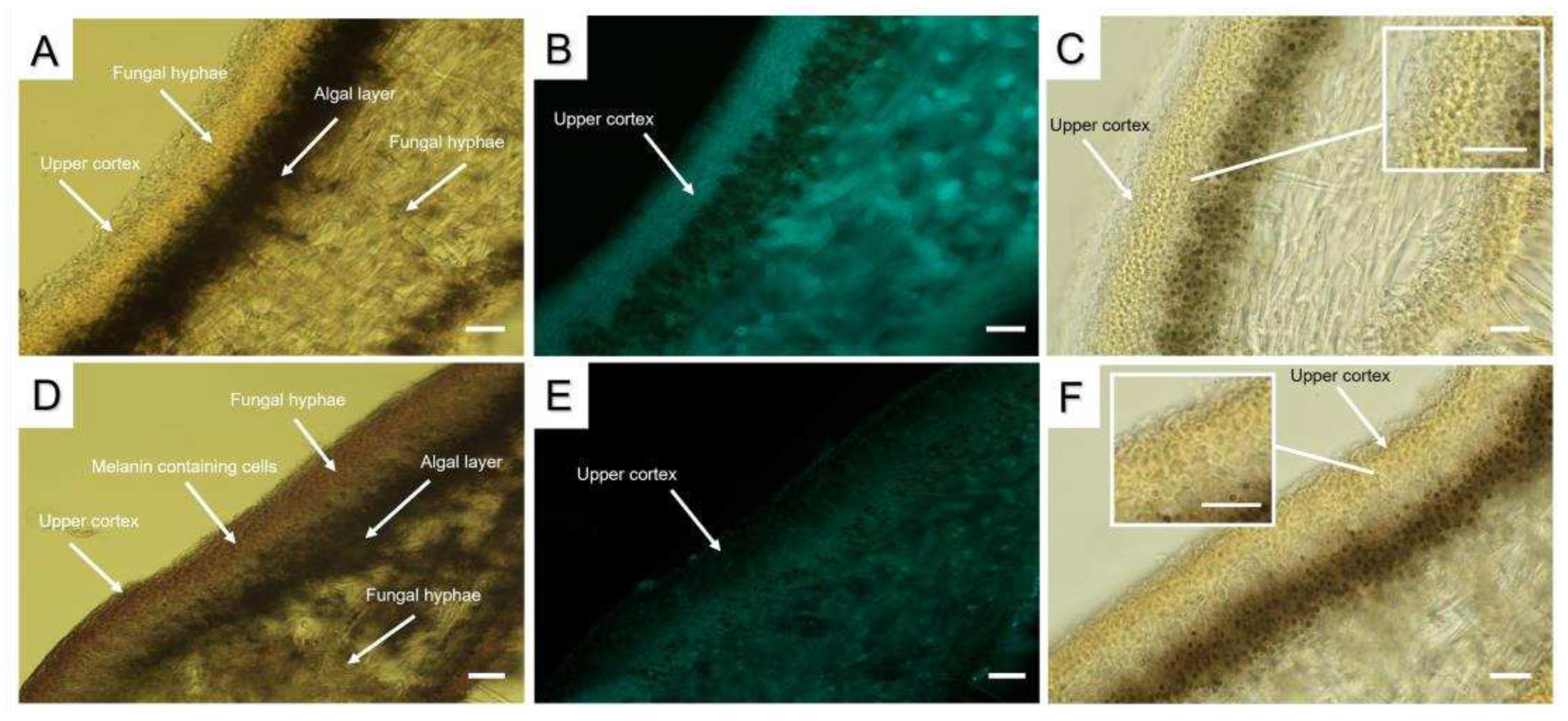
Figure 3.
3D relief (A), adhesion (B-D) of the surface of cross sections of non-melanized (A-D) and 3D relief (E), adhesion (F-H) of the surface of cross sections of melanized (E-H) L. pulmonaria thalli.
Figure 3.
3D relief (A), adhesion (B-D) of the surface of cross sections of non-melanized (A-D) and 3D relief (E), adhesion (F-H) of the surface of cross sections of melanized (E-H) L. pulmonaria thalli.
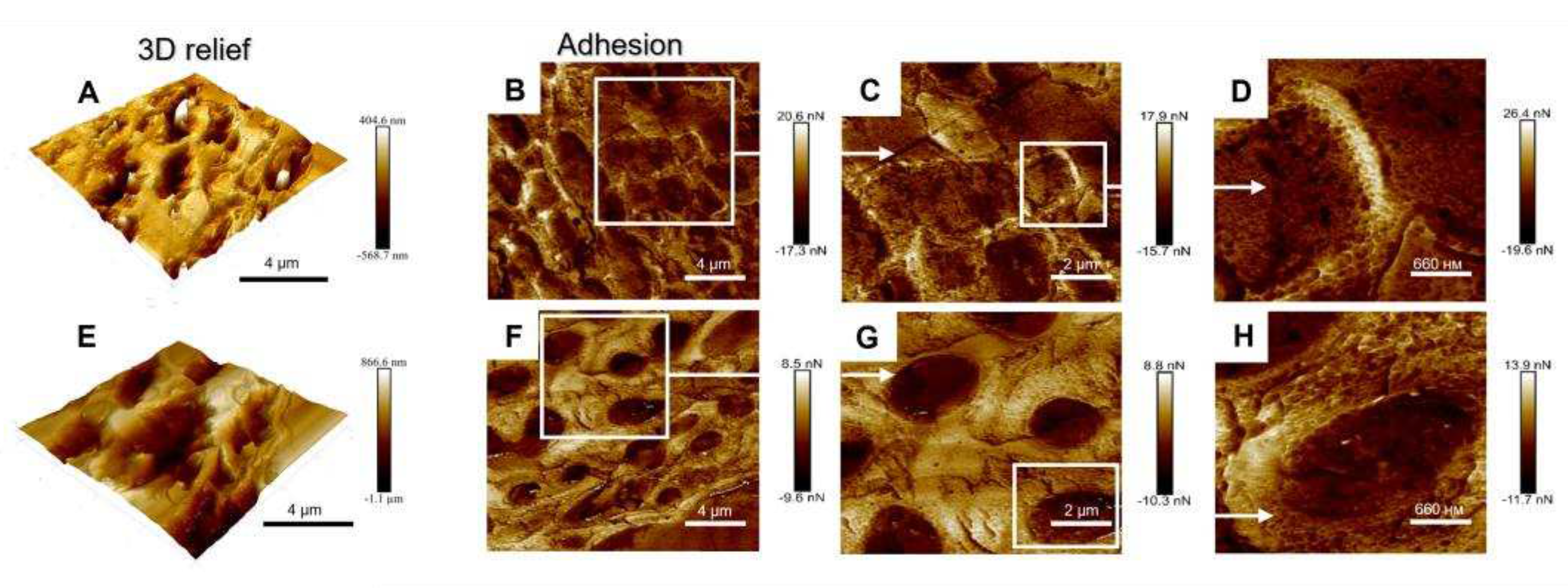
Figure 4.
Absorbance of the incubation solutions (λ490, A,B), the content of total phenolic compounds (C,D) and total carbohydrates (E,F) of pale (A,C,E) and melanized (B,D,F) thalli of lichen L. pulmonaria treated with buffers solutions with and without enzymes (n = 3). Significance: * = p < 0.05, ** = p < 0.01, *** = p < 0.001.
Figure 4.
Absorbance of the incubation solutions (λ490, A,B), the content of total phenolic compounds (C,D) and total carbohydrates (E,F) of pale (A,C,E) and melanized (B,D,F) thalli of lichen L. pulmonaria treated with buffers solutions with and without enzymes (n = 3). Significance: * = p < 0.05, ** = p < 0.01, *** = p < 0.001.
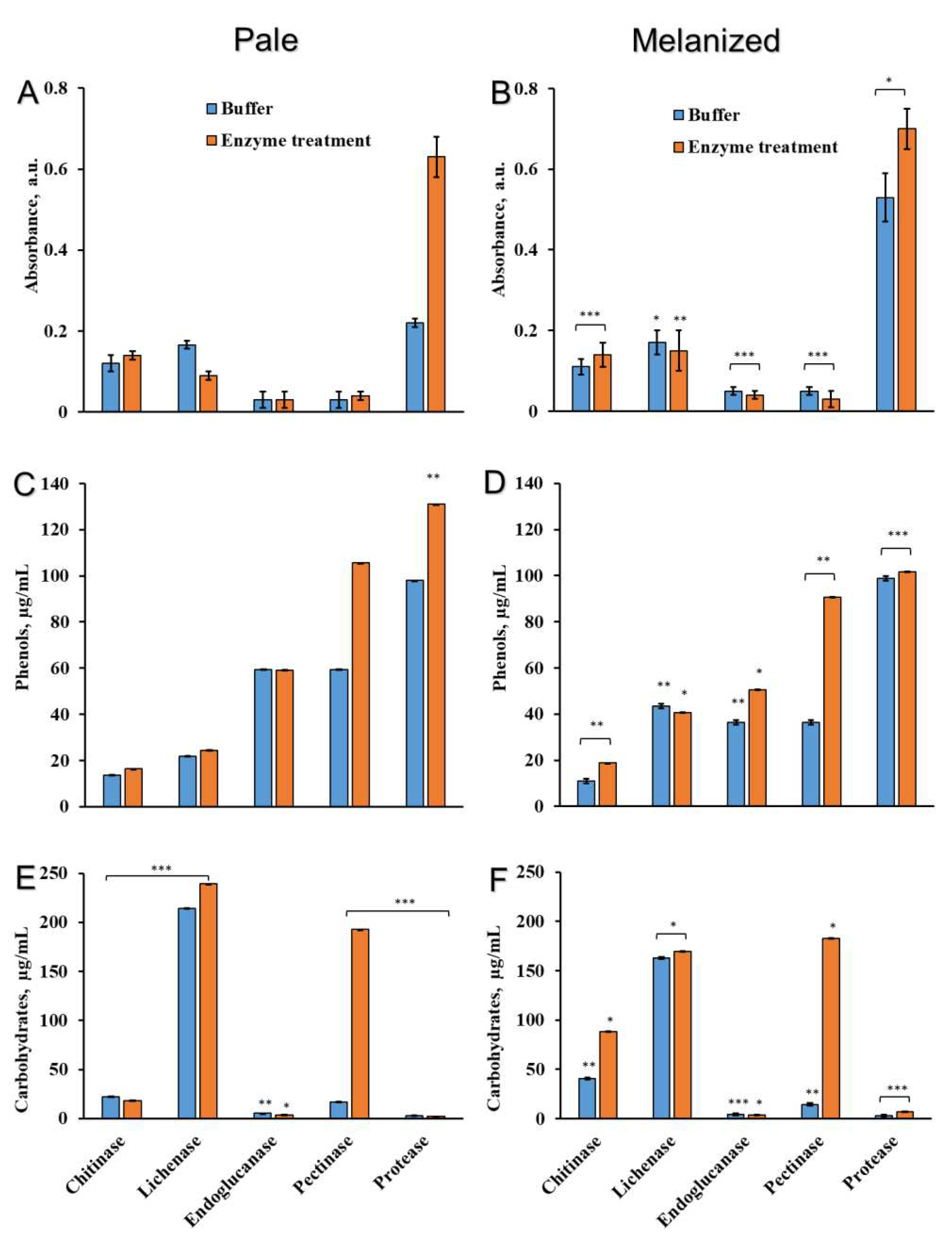
Figure 5.
Content of melanin (mg dry melanin/mL alkaline solution) in the pale (A) and melanized (B) thalli of the lichen L. pulmonaria following enzyme treatments. Significance: * = p < 0.05, ** = p < 0.01, *** = p < 0.001.
Figure 5.
Content of melanin (mg dry melanin/mL alkaline solution) in the pale (A) and melanized (B) thalli of the lichen L. pulmonaria following enzyme treatments. Significance: * = p < 0.05, ** = p < 0.01, *** = p < 0.001.
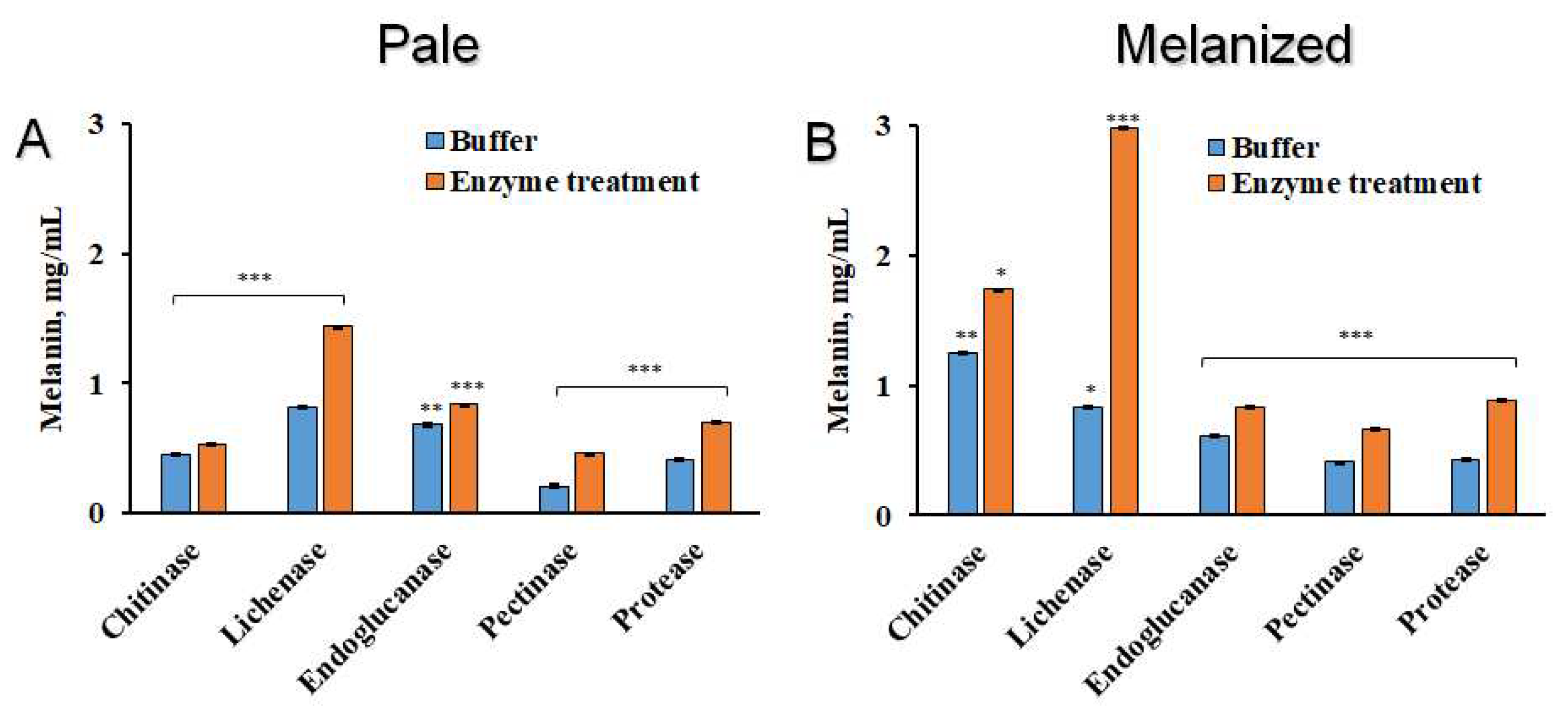
Figure 6.
TEM images of cross sections of melanized L. pulmonaria thalli: without enzyme treatment (A), after treatment with chitinase (B), endoglucanase (С), pectinase (D), and protease (E). Cell wall (CW), plasma membrane (PM), nucleus (N). Scale bar corresponds 0.5 µm.
Figure 6.
TEM images of cross sections of melanized L. pulmonaria thalli: without enzyme treatment (A), after treatment with chitinase (B), endoglucanase (С), pectinase (D), and protease (E). Cell wall (CW), plasma membrane (PM), nucleus (N). Scale bar corresponds 0.5 µm.
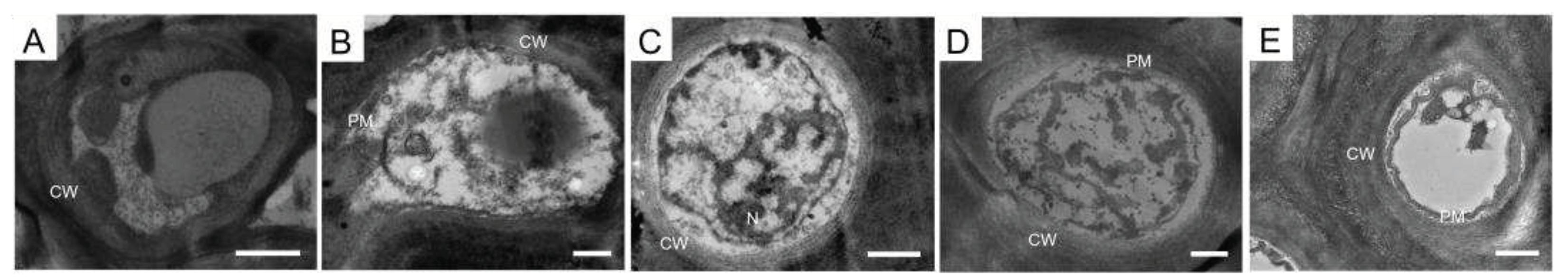

Table 1.
List of enzymes.
| Enzyme | Specific activity | Source | Buffer | Units ml-1 |
|---|---|---|---|---|
| Chitinase 1 | Hydrolysis of N-acetyl-beta-D-glucosaminide (1-4)-beta-linkages in chitin and chitodextrins | Trichoderma viride | 50 mМ sodium phosphate, pH 6.0 |
0.5 |
| Lichenase 2 |
Hydrolysis of (1,4)-β-D-glucosidic linkages in β-D-glucans containing (1,3)- and (1,4)-bonds | Bacillus subtilis | 10 mМ sodium phosphate, pH 6.0 |
5 |
| Endoglucanase 2 |
Hydrolysis of (1,3)-β-D-glucosidic linkages in (1,3)-β-D-glucans | Trichoderma sp. | 100 mМ sodium acetate, pH 4.5 |
0.5 |
| Pectinase 1 | Hydrolysis of pectin-containing substances | Aspergillus aculeatus | 100 mМ sodium acetate, pH 4.5 |
1 |
| Protease 1 | Hydrolysis of peptide bonds in proteins with conversion to shorter polypeptides and amino acids | Bacillus licheniformis | 100 mМ sodium phosphate, pH 7.4 |
1 |
1 Sigma-Aldrich, USA; 2 Megazyme, USA.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



