Submitted:
22 June 2023
Posted:
22 June 2023
You are already at the latest version
Abstract
It is well-established that plants are sessile and photoautotrophic organisms that rely on light throughout their entire life cycle. Light quality (spectral composition) is especially important as it provides energy for photosynthesis and influences signaling pathways that regulate plant devel-opment in the complex process of photomorphogenesis. During past years, significant progress has been made in the physiological and biochemical effects of light quality on crops. However, understanding how light quality modulates plant growth and development remains a complex challenge. In this review, we provide an overview of the role of light quality in regulating early development of plants, encompassing processes such as seed germination, seedling de-etiolation, and seedling establishment. These insights can be harnessed to improve production planning and crop quality by producing high-quality seedlings in plant factories and improving the theoretical framework for modern agriculture.
Keywords:
light quality
; photomorphogenesis
; seed germination
; seedling de-etiolation
; shoot-root develop-ment
; leaf development
; greenhouse horticulture
1. Introduction
It is widely acknowledged that plants are sessile and photoautotrophic organisms. Light regulates plant physiology through two major functions throughout the plant life cycle [1]. The assimilative function of light provides the energy necessary for photosynthesis, suggesting light is the ultimate energy source for green plant metabolism, and the photosynthetic efficiency depends on the spectral wavelength (light quality) [2,3,4]. On the other hand, the signaling function of light activates and regulates many key signaling pathways related to plant photomorphogenesis [5,6,7,8,9,10,11]. In developmental biology, the response of plant growth patterns to light spectra is known as photomorphogenesis, which occurs during seed germination, seedling development, and the transition from vegetative to anthesis (photoperiodic phenomenon) [12,13,14]. For instance, one seminal study showed that red light promotes the germination of lettuce seeds (Lactuca sativa L.) [15]. Recent studies on Arabidopsis have similarly shown that the process of seed germination is regulated by phytochrome B (PhyB), the primary photoreceptor involved in red-light-induced germination [9]. After germination, the seedlings exhibited an etiolated growth pattern wherein the hypocotyls was elongated and the cotyledons folded to form a hook-like structure [16]. Besides, light quality plays a crucial role in regulating plant photomorphogenesis during seedling de-etiolation [17]. Moreover, light quality also plays an important role in the transition of plants from vegetative to reproductive growth [5] and senescence [13].
There is an increasing consensus suggesting that plants have evolved an array of photoreceptors that function to transduce light cues into biological signals [9,13,18,19,20]. Ample evidence substantiates that the biochemical and physiological features of five classes of photoreceptors in plants have been characterized [21,22]. It has been reported that blue (B)/UV-A light (320-500nm) are absorbed by three distinct classes of photoreceptors, including cryptochromes (CRYs) [23], FLAVIN-BINDING, KELCH REPEAT, F-BOX1 (FKF1), ZEITLUPE (ZTL)/LOV KELCH PROTEIN2 (LKP2) [24], and phototropins (PHOTs) [25]. Red (R) and far-red (FR) light (600-750 nm) are primarily perceived by phytochromes (PHYs) and are involved in many photo-regulatory processes [13]. It has been reported that UVRESISTANCE LOCUS8 (UVR8) could sense UV-B light (280-315 nm) [26]. Overall, these photoreceptors, which perceive different light qualities, have crucial functions throughout the life cycle of plants, starting from seed dormancy and germination to seedling de-etiolation [21,22], flowering [5], and senescence[13] (Figure 1).
Raising and transplanting seedlings is commonly used in agriculture for cultivating vegetables and economic crops. Thus, producing high-quality seedlings is crucial for efficient and successful plant cultivation, providing numerous benefits, including control over growing conditions, disease and pest management, plant breeding opportunities, efficient land use, transplanting flexibility, and precise timing for planting and harvesting [27]. Seedling responses to different wavelengths can have independent effects on light-regulated development [28]. The application of light-emitting diode (LED) light systems in plant factory settings for seedling cultivation have attracted increasing attention, given their ability to flexibly control the spectral composition of light [29,30,31,32]. Therefore, it is of great significance to deeply understand how light quality regulates the growth and development of plant seedlings. Numerous studies and reviews have detailed the effects of light quality on regulating plant growth and development [21,28,33,34,35]. However, few reviews have hitherto assessed the role of light quality on young seedling development. In this review, we summarize the role of light quality in regulating early plant development, including seed germination, seedling de-etiolation, and seedling establishment, providing the foothold to refine production planning and crop quality by producing high-quality seedlings in plant factories, and a theoretical basis for modern agriculture.
2. The light signaling pathway
The light signaling pathway in plants involves the perception of light by photoreceptors, the transduction of the light signal into biochemical changes, and the subsequent regulation of various physiological and developmental responses [33]. It plays a crucial role in plant growth, development, and adaptation to their environment. Plants have specialized photoreceptor molecules that allow them to perceive and respond to light. These photoreceptors can be categorized into five classes based on the wavelength of light they absorb (Figure 1) [21,22,33].
Red and far-red light: PHYs were the first photoreceptor proteins discovered in plants that enable the detection of R and FR light [13,36,37]. PHYs are evolutionarily conserved from bryophytes to angiosperms (except in the chlorophytes). In dicotyledonous plants such as Arabidopsis thaliana, five Phys encoded by small gene families have been identified: PhyA, PhyB, PhyC, PhyD, and PhyE [37,38]. However, in monocots, the phytochrome family consists of three members: PhyA, PhyB, and PhyC [36,39]. Furthermore, based on their stability in light, these phytochromes can be classified into photostable type I (PhyA is the only type I phytochrome) and photostable type II (PhyB to PhyE) [37,40,41]. It has been established that PhyA plays a dominant role in FR light, while PhyB to PhyE regulates R light signaling [36,41].
Current evidence suggests that PHYs exist in Pfr (active form) and Pr (inactive form) forms which are interconvertible. When a PHY molecule absorbs R light, it is converted from Pr to Pfr; when it absorbs FR light, it is converted back to Pr [7,41]. This reversible conversion between Pr and Pfr serves as major molecular switches in the PHYs signaling pathway, and their pathway and downstream components have been extensively studied. First, PHYs are transferred from the cytoplasm to the nucleus through the light-activated Pfr form to interact with transcription factors to regulate the expression of related genes, among which PIFs (Phytochrome-Interacting-Factors) are major transcription factors that interact with PHYs [42,43]. PIFs represent transcription factors of the bHLH (basic-helix-loop-helix) family that primarily function as negative regulators of photomorphogenesis [42,43]. Current evidence suggests that PhyB and PhyA interact with PIF1 to inhibit light-dependent seed germination [44]. In addition, PHYs also regulation of ubiquitin-mediated protein degradation is an important component of the PHYs signaling machinery [33]. COP1-SPA (Constitutive photomorphogenic protein 1-Supressor of PhyA-105) E3 ligase complex is a regulator that plays a central role downstream of various photoreceptors [45,46,47]. It targets several positive regulators of photomorphogenesis, such as HY5 (Elongated- Hypocotyl 5), leading to its destabilization and degradation in the dark through the 26S proteasome pathway [45,48]. In summary, PHYs initiate light signaling pathways through two major negative regulators: interacting with PIFs transcription factors and regulating the stability of COP1-SPA complex protein [39,40,41].
Blue/UV-A light: Blue (B)/UV-A light (320-500nm) is mainly absorbed by three distinct types of photoreceptors, including PHOTs [23], ZTL/FKF1/LKP2 family [24], and CRYs [25].
PHOTs are a class of protein kinases that contains serine/threonine domain and FMN (flavin-mononucleotide) binding LOV (light-oxygen-voltage) domain and widely found in green plants [49,50]. They have been identified in ferns and mosses in addition to higher plants such as Arabidopsis, and their physiological functions are conserved across different species [33,49]. The signaling pathway of PHOTs begins with the absorption of B-light and are primarily involved in mediating phototropism, which responsible for the directional growth or movement of plants in response to light [49,50]. In addition, PHOTs also regulate other B-light-mediated processes, including control of chloroplast movement, stomatal opening, and leaf expansion [50].
The ZTL/FKF1/LKP2 family is another type of blue light receptor, and its protein structure also has the LOV domain [24,50]. It has been shown that ZTL/FKF1/LKP2 family proteins play critical roles in integrating light and circadian signaling pathways to regulate plant development, including flowering time and the circadian clock, by controlling the stability of key light regulatory proteins [24,51].
CRYs are important photosensory receptors that absorb B/UV-A light [52]. Exposure to blue light causes rapid phosphorylation of the cryptochrome molecule, an essential modification for its function. CRYs regulate important physiological processes throughout the plant life cycle, such as seedling photomorphogenesis, photoperiodic flowering, and circadian rhythm. Three CRYs have been identified, with Cry1 and cry2 being homologous and extensively studied to understand their functions. The third CRY, cry3, is a more distinct family member, but its function remains to be determined [23,52]. For instance, in Arabidopsis, Cry1 is involved in blue light activation of the photomorphogenesis pathway, leading to the inhibition of hypocotyl growth, promotion of cotyledon expansion, and stimulation of chloroplast development [52,53,54].
CRYs interact with the E3 ubiquitin ligase COP1 protein, forming a complex that regulates light-dependent protein degradation [55,56]. COP1 suppresses photomorphogenic development in the dark by targeting transcription factors, including the bZIP protein HY5, for degradation. Upon light activation, CRYs bind to COP1, preventing its interaction with transcription factors, thus promoting photomorphogenesis. In addition, CRYs interact with many other proteins, but the functional significance of these interactions remains unclear. Numerous studies have demonstrated interactions between CRYs and PHYs [55,57].
UV-B light: The UVR8 protein acts as the primary UV-B photoreceptor in plants and triggers a signaling cascade upon UV-B perception [18,26]. UVR8 is a homodimeric protein composed of two identical subunits. The absorption of UV-B photons by UVR8 leads to changes in the protein conformation, which result in the dissociation of the UVR8 homodimer into monomers [26,58]. After dissociation, UVR8 monomers interact with the COP1 protein, forming a complex. This interaction prevents the degradation of UVR8 and allows it to accumulate in the nucleus. Once in the nucleus, the UVR8-COP1 complex regulates the expression of various genes involved in UV-B responses. It interacts with transcription factors, such as HY5, to activate or repress gene expression, leading to the induction of protective responses, such as the synthesis of UV-absorbing compounds and DNA repair enzymes [18,59]. Plants integrate UVR8 signaling with other light signaling pathways, including the PHYs and CRYs pathways [60]. This interplay between pathways enables plants to synchronize their responses to diverse light wavelengths and environmental conditions.
In general, the fundamental mechanism of transducing light signals in photoreceptor-mediated pathways involves direct interactions between photoreceptors and their target proteins. These interactions can occur in a light-dependent or independent manner and regulate various aspects of physiological and developmental processes controlled by light-related genes in plants [43,45,48,61]. In addition, the phytohormone pathways, including primary plant hormones such as auxin, abscisic acid (ABA), and gibberellins (GAs), are reportedly involved in plant photomorphogenesis processes [16,28,35].
3. Roles of light quality in the regulation of seed germination
The effect of light quality in regulating seedling establishment commences with seed germination. It has long been established that the regulation of two hormones, GA and ABA, are required for seed germination [22,35]. Over the years, the roles of ABA and GA in seed germination have been extensively reviewed [22,35,62,63]. ABA acts as a dormancy-inducing hormone, inhibiting seed germination under unfavorable conditions, while GA promotes seed germination by mobilizing stored nutrients, activating enzymes, and overcoming ABA-induced dormancy. The balance between these two hormones is critical for regulating seed germination and ensuring successful plant establishment [21,35,64,65]. Light quality involved in seed germination is mediated by photoreceptors [13,21] and the levels of ABA and GA, which have antagonistic functions [22,66] (Figure 2).
Previous studies have identified the role of B-light in the inhibition of seed germination, especially in two cultivated cereals: wheat (Triticum aestivum) and barley (Hordeum vulgare) [53,67]. In barley, B-light inhibition of grain germination is dependent on Cry1, given that in germinating Cry1a/Cry1b RNAi seeds, B-light results in the downregulation of the ABA biosynthetic gene NCED1 (9-Cis-Epoxycarotenoid Dioxygenase1), and upregulation of the ABA catabolic gene ABA8’OH1 [53]. Growing evidence suggests that B-light enhancement the expression of NCED1 dependent on Cry1, which increases ABA content and inhibits seed germination in dormant barley [21,67]. Extensive literature substantiates that B-light inhibits the germination of other monocotyledonous seeds, such as in imbibed annual ryegrass (Lolium Rigidum), wheat grain (Triticum Aestivum L.), wild grain (Brachypodium disachyon), et al [68,69,70,71]. In summary, B-light suppresses monocot seed germination by enhancing the expression of NCED1 and repressing the expression of ABA8’OH1 to regulate the content of ABA in embryos (Figure 2) [53,63,67]. The role of B-light is mainly discussed in the context of the seed biology of monocot plants. Recent studies have reported that blue LED light could improve the rate and speed of Stevia seeds germination [72]. However, in dicots, the role of B-light in seed germination and its action mechanism have not been well elucidated.
The regulation of R and FR light on seed germination was discovered by Borthwick et al. in lettuce (Lactuca sativa), and the results showed that R light-induced seed germination, whereas FR light inhibited this process [15,22]. Upon exposure to R light, PHYs are transformed into the activated form of Pfr, which promotes seed germination by controlling the content of GA and ABA by directly or indirectly regulating the expression of synthesis or metabolism-related genes. However, the conversion of Pfr to Pr in the presence of FR light counteracts the effect of R light on seed germination [16,35,73]. In dicots, from PhyA to PhyE each member plays a distinct role in mediating seed germination in response to various environmental cues. These PHYs enable the seeds to adjust their timing and location of germination based on specific environmental signals [74]. PhyB occupies a central position in the regulation of seed germination under R/FR light irradiation, whereas phyA plays a role in mediating very low fluence response to R/FR light (YANG). PhyE and phyD are required for seed germination under continuous FR light and very low R/FR ratios [66,74].
At present, the molecular mechanisms underlying PhyB mediated germination are better understood than those of PhyA-mediated germination. It has been established that PhyB is the most crucial protein involved in initiating the early stages of seed germination. When exposed to R light, PhyB is activated and translocated to the nucleus, facilitating the degradation of PIF1. It is well known that the PIFs negatively regulate PHY-mediated light signaling pathways by directly and indirectly regulating GA and ABA signaling [75]. However, under low R/FR ratio conditions, PhyB-induced degradation of PIF1 is dynamically reversible, allowing PIF1 to accumulate in the cell nucleus, resulting in a decrease in GA levels and inhibition of seed germination [22,42].
It has been shown that PIF1 directly and indirectly control GA content. It directly induces the expression of two repressors of GA signaling, RGA (Repressor of Gai-3) and GAI (gibberellic acid insensitive) [64]. It also indirectly controls GA levels by repressing the expression of GA3ox1 (Gibberellin 3-oxidase 1) and GA3ox2, which are GA biosynthetic genes, while also activating the expression of GA catabolic gene GA2ox2 [64,76,77]. The role of PIF1 in regulating ABA level is similar to participating in the GA signaling pathway. It induces the transcription of three ABA biosynthetic genes: ABA1 (ABA deficient1), NCED6 (9-Cis -Epoxycarotenoid Dioxygenase 6), and NCED9 (9-Cis-Epoxycarotenoid Dioxygenase 9), but inhibit the expression of CYP707A2, which is the ABA catabolic gene [22,65,75]. In summary, the modulation of seed germination by R and FR light involves the interplay between phytochromes and PIF1, which regulates the ABA and GA pathways. However, there is still much to be understood in designing appropriate strategies for regulating seed germination mediated by light quality for individual plant species.
4. Roles of light quality in the regulation of photomorphogenesis
4.1. Seedling de-etiolation
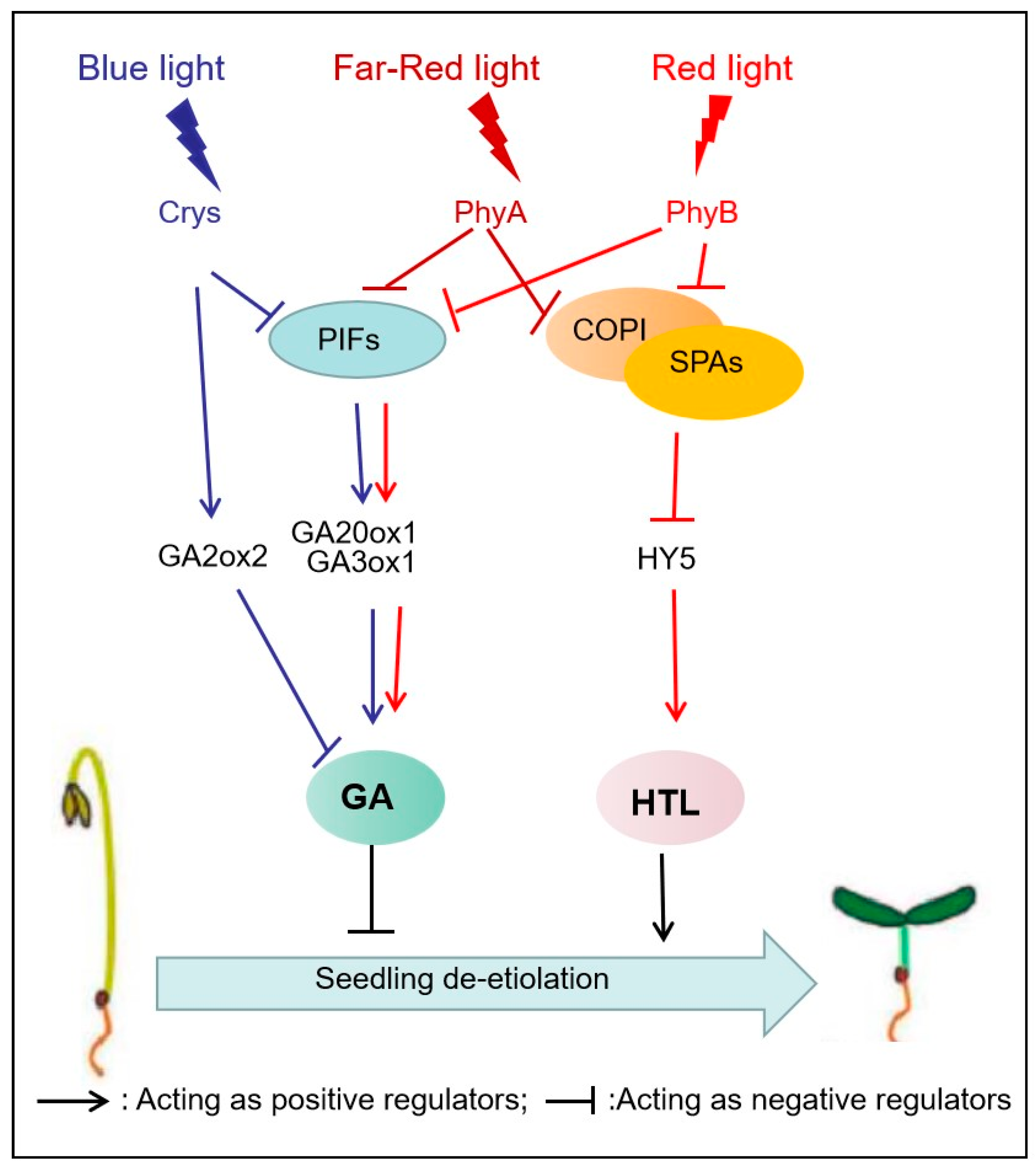
Following germination, seedlings undergo etiolation under the soil, characterized by strong elongation of hypocotyls and closed cotyledons that lack chlorophyll and functional chloroplasts [16]. De-etiolation of the seedlings marks the major developmental switch upon their emergence from the soil as they reach the light. This phase involves the arrest of hypocotyl growth, the opening of the cotyledons, and the biosynthesis of chlorophyll, followed by chloroplast development and eventually autotrophic growth (known as photomorphogenesis) [13,52,78]. Numerous studies have revealed that B-light and UVA induce de-etiolation mainly via CRYs, and PHYs are required for FR and R-light-induced de-etiolation (Figure 3) [17,52] .
The inhibition of hypocotyl elongation may be a characteristic phenotype in de-etiolation studies [52]. The inhibition of hypocotyl elongation in response to B-light is mainly mediated by Cry1, for instance the cry1 (hy4) mutant weakened the inhibitory effect of B-light on hypocotyl elongation [79]. CRYs regulate B-light-induced hypocotyl elongation through mediating the GA inactivation gene (GA2ox2) and the genes involved in GA synthesis (GA20ox1 and GA3ox1) expression [80,81]. Song et al. revealed that CRYs could repress the transcription activity of PIF4 by binding to it, thereby reducing the expression of the downstream GA20ox1 and GA3ox1 genes and upregulating the transcript level of GA2ox1, leading to increased GA inactivation. These processes ultimately result in reduced levels of GA and shorter hypocotyls [80]. Cry2 is also involved in B-light-induced de-etiolation through COP1/SPA-HY5 pathways [52].
PHYs play a central role in FR and R light-regulated de-etiolation events via two main light signaling pathways, COP1/SPA-HY5 and four PIFs members (PIF1, PIF3, PIF4, and PIF5) [13,16,80,82]. The COP1/SPA complex, as an E3 ubiquitin ligase, negatively regulates the levels of several photomorphogenesis-promoting proteins, such as HY5 [82,83]. Specifically, the COP1 complex with SPA1 directly interacts and ubiquitinates HY5, which directly binds to both the C/G box and G box in the promoter of HTL (hypersensitive to light) [13,46]. HTL is a positive regulatory factor of the de-etiolation response mediated by PHYs and CRYs [84]. In addition to the COP1/SPA-HY5 pathway, the PIFs also plays a central role in the de-greening process of seedlings [42]. R and FR light promotes photomorphogenesis due to phosphorylation and rapid degradation of PIFs mediated by PhyA and PhyB [13,42]. UV-B also mediates seedling de-etiolation via its dependence on UVR8 and COP1/SPA-HY5 pathway [85]. However, the mechanism of de-etiolation in monocots has not been thoroughly studied. It is worth noting that in rice, a member of the PIFs family called OsPIL15 is involved in repressing etiolated seedling growth [86].
4.2. Shoot-root development
Despite growing beneath the soil, roots in plants are still impacted by light signaling transmitted from the shoot, which can influence the development of both primary and lateral roots [87]. Several studies have shown that COP1, HY5, and UVR8 play important roles in root and shoot growth and demonstrated how light and photoreceptors regulate root and shoot growth [87,88,89,90]. In shoots, COP1 induces PIN1 (PIN-FORMED 1) transcription to regulate shoot-to-root polar auxin transport and the intracellular distribution of PIN1 and PIN2 in roots to influence growth and development [91]. HY5 is also involved in regulating root growth and development by light quality [87,92]. The reduction in lateral root density induced by the joint application of white and FR light is thought to be mediated by HY5, given that hy5 mutants exhibit a similar reduction in lateral root density compared to wild-type plants, regardless of whether they are exposed to FR light or not [93]. The stability of HY5 in roots is modulated by blue light through its interaction with CRYs, which leads to the activation of miR163 and HY5 and promotes primary root growth [94,95] Further studies revealed that HY5 regulates lateral root emergence by affecting auxin signaling. Recent studies have reported that UVR8 directly interacts with MYB73/MYB77 (MYB domain protein73/77) transcriptional factors, to modulate shoot and root growth in Arabidopsis [95]. In addition to indirectly modulating root growth by regulating shoot growth, light is also directly involved in the regulation of root growth; however, the underlying mechanisms remain unclear. It is widely thought that roots and shoots must coordinate their growth responses to allow better growth of the whole plant. Therefore, further research is warranted to fully understand how light coordinates the development and growth of shoots and roots.
4.3. Leaf development
The shade avoidance response (SAR) is a plant response mechanism that aims to optimize the acquisition of light energy for photosynthesis during vegetative growth. The SAR is characterized by increased hypocotyl, stem, and petiole elongation, a more erect leaf position, increased apical dominance, and early flowering [85,96]. Importantly, it is now understood that shade from the vegetation has a distinct spectral signature. UV-B, F, and B-light in unfiltered sunlight activate their corresponding photoreceptors: UVR8, PHYs, and CRYs. These photoreceptors directly or indirectly converge to inhibit PIFs and the COP1/SPA complex to regulate SAR [97]. UV-B, R, and B-light are depleted in shaded environments, while FR light is relatively abundant. Consequently, UVR8, PHYs, and CRYs activity are greatly reduced, preventing the inhibitory action of these photoreceptors on COP1 and PIFs. Moreover, under low R: FR ratio conditions, the Pfr form converts to the Pr form, leading to re-accumulation and stabilization of PIFs, which promote stem elongation. In addition, under low R: FR ratio conditions, PIF1, PIF5, and PIF7, through regulation auxin biosynthesis involved in hypocotyl elongation [98,99].
The stomata are openings on the leaf surface, which mediate gas and water vapor exchange between the plant and the environment. Numerous studies have indicated that light modulation of stomatal development is another crucial aspect directly related to plant photomorphogenesis [17,100,101]. Light plays a key role in forming mature stomata and ensuring proper stomatal patterning [102]. Consistently, several studies have shown that the formation of M (meristemoid) and stomatal maturation is compromised in phyB, phyA, and cry1cry2, the photoreceptor loss-of-function mutants [55,102]. In Arabidopsis, the components of light signaling (such as COP1 and PIFs) and those of the stomata developmental pathway work synergistically to regulate the whole process of stomatal development [102]. In the dark, single loss-of-function mutants of cop1, cop10, and det1 exhibited constitutive clustered stomata, suggesting that COP1 and SPA proteins all act to repress asymmetric cell division and stomatal fate initiation [55,103,104]. Recently studies have substantiated that COP1 facilitates YDA activity to suppress stomata formation by promoting phosphorylation and subsequent degradation of SPCH (SpchlessP) and ICE1 [47,102,105,106]. Substantial evidence also indicates that PIFs accumulate in the dark and work synergistically with the COP/SPA complex to suppress photomorphogenesis [102,107]. In Arabidopsis, PIF4 directly inhibits SPCH expression, suppressing stomatal development in response to higher temperatures [108]. Another report indicated that the PIF-GNC/GNL module is important in light-mediated stomatal development. PIFs can directly repress the expression of two paralogous Arabidopsis genes, GNC and GNL, which could promote cell division and stomata formation in cotyledons and hypocotyls during dark-to-light transition [109]. Although the past few years have witnessed significant scientific progress, knowledge of the regulatory mechanisms of stomatal development and patterning in cereal grasses like maize, rice, barley, and Brachypodium remains rudimentary, even though significant progress has been made in understanding the mechanisms of light-regulated stomatal development and behavior in Arabidopsis [102].
Besides, the light signal is important for the biogenesis and development of chloroplasts, which are crucial for plant growth [110,111]. Chloroplast development is regulated jointly by nuclear and plastid genes, which are the photosynthesis-associated nuclear-encoded genes (PhANGs) and the photosynthesis-associated plastid-encoded genes (PhAPGs). PhANGs are transcribed by the RNA polymerase NEP (nuclear-encoded RNA polymerase) encoded by the nuclear genome, while PhAPGs are transcribed by the plastid RNA polymerase PEP (plastidial RNA polymerase). It has been established that PHYs could activate these two major classes of genes by inhibiting PIFs [112,113]. Upon light activation, the PHYs induce the PEP complex to complete their assembly in the plastid, thus initiating PhAPG transcription. Conversely, in the dark, the PIFs can suppress the transcription of the nuclear gene PhANGs and inhibit the PEP complex assembly and PhAPG transcription in the plastid [112,113]. It is worth noting that the blue-light induction of the nucleus-encoded SIG5 gene, which acts as a retrograde and light-controlled regulator of chloroplast function, is primarily mediated by CRY1, a photoreceptor that responds to blue light [52,114]. Therefore, both PHYs and CRYs play a pivotal role in the development of etioplasts into chloroplasts.
5. Conclusions and Perspectives
Over the past two decades, the application and impact of light quality in horticulture have become a research hotspot [80,115,116,117]. Herein, we sought to illustrate the molecular mechanism of light quality in regulating early seedling development, including the establishment of seedling and vegetative architecture. Greenhouse horticulture could leverage these insights to improve production schedules and crop quality.
Despite considerable progress in the understanding of molecular events underlying photomorphogenesis, there are still a large number of unresolved issues. Moreover, the influence of light quality on plant growth is relatively complex, and A. thaliana is often used as a model plant in laboratory settings. Therefore, there is an urgent need for research to understand how knowledge gathered in Arabidopsis can be applied to understand the adaptation of greenhouse horticultural plants to their local environment and to improve production schedules and crop quality.
Author Contributions
Conceptualization and writing: Y.W. and S.W.; supervision: D.Y.
Funding
This study was funded by the Shenzhen Peacock Program of Shenzhen (KQTD2017032715165926).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Devlin, P. Photocontrol of Flowering; Blackwell Publ.: Oxford, UK, 2008; pp. 185–210. [Google Scholar]
- Kaiser, E.; Correa Galvis, V.; Armbruster, U. Efficient Photosynthesis in Dynamic Light Environments: A Chloroplast’s Perspective. Biochem. J. 2019, 476, 2725–2741. [Google Scholar] [CrossRef]
- Liu, J.; van Iersel, M.W. Photosynthetic Physiology of Blue, Green, and Red Light: Light Intensity Effects and Underlying Mechanisms. Front. Plant Sci. 2021, 12, 328. [Google Scholar] [CrossRef] [PubMed]
- Trivellini, A.; Toscano, S.; Romano, D.; Ferrante, A. The Role of Blue and Red Light in the Orchestration of Secondary Metabolites, Nutrient Transport and Plant Quality. Plants 2023, 12, 2026. [Google Scholar] [CrossRef]
- Cerdán, P.D.; Chory, J. Regulation of Flowering Time by Light Quality. Nature 2003, 423, 881–885. [Google Scholar] [CrossRef]
- Chen, C.; Huang, M.; Lin, K.; Wong, S.; Huang, W.; Yang, C. Effects of Light Quality on the Growth, Development and Metabolism of Rice Seedlings (Oryza Sativa L.). Res. J. Biotechnol. 2014, 9, 15–24. [Google Scholar]
- Zhao, J.; Bo, K.; Pan, Y.; Li, Y.; Yu, D.; Li, C.; Chang, J.; Wu, S.; Wang, Z.; Zhang, X.; Gu, F.; Weng, Y. Phytochrome-interacting factor PIF3 integrates phytochrome B and UVB signaling pathways to regulate gibberellin-and auxin-dependent growth in cucumber hypocotyls. J. Exp. Bot. 2023, erad181. [Google Scholar] [CrossRef]
- Jiang, H.; Li, Y.; He, R.; Tan, J.; Liu, K.; Chen, Y.; Liu, H. Effect of Supplemental UV-A Intensity on Growth and Quality of Kale under Red and Blue Light. Int. J. Mol. Sci. 2022, 23, 6819. [Google Scholar] [CrossRef]
- Jiang, Z.; Xu, G.; Jing, Y.; Tang, W.; Lin, R. Phytochrome B and REVEILLE1/2-Mediated Signalling Controls Seed Dormancy and Germination in Arabidopsis. Nat. Commun. 2016, 7, 12377. [Google Scholar] [CrossRef]
- Yang, L.; Wang, L.; Ma, J.; Ma, E.; Li, J.; Gong, M. Effects of Light Quality on Growth and Development, Photosynthetic Characteristics and Content of Carbohydrates in Tobacco (Nicotiana Tabacum L.) Plants. Photosynthetica 2016, 55. [Google Scholar] [CrossRef]
- Yavari, N.; Tripathi, R.; Wu, B.-S.; MacPherson, S.; Singh, J.; Lefsrud, M. The Effect of Light Quality on Plant Physiology, Photosynthetic, and Stress Response in Arabidopsis Thaliana Leaves. PloS One 2021, 16, e0247380. [Google Scholar] [CrossRef]
- Batista, D.S.; Felipe, S.H.S.; Silva, T.D.; de Castro, K.M.; Mamedes-Rodrigues, T.C.; Miranda, N.A.; Ríos-Ríos, A.M.; Faria, D.V.; Fortini, E.A.; Chagas, K.; et al. Light Quality in Plant Tissue Culture: Does It Matter? Vitro Cell. Dev. Biol. - Plant 2018, 54, 195–215. [Google Scholar] [CrossRef]
- Tripathi, S.; Hoang, Q.; Han, Y.; Kim, J. Regulation of Photomorphogenic Development by Plant Phytochromes. Int. J. Mol. Sci. 2019, 20, 6165. [Google Scholar] [CrossRef] [PubMed]
- Mohr, H. Lectures on Photomorphogenesis; Springer Science & Business Media, 2012; ISBN 978-3-642-65418-3. [Google Scholar]
- Borthwick, H.A.; Hendricks, S.B.; Parker, M.W.; Toole, E.H.; Toole, V.K. A Reversible Photoreaction Controlling Seed Germination. Proc. Natl. Acad. Sci. U. S. A. 1952, 38, 662–666. [Google Scholar] [CrossRef]
- de Wit, M.; Galvão, V.C.; Fankhauser, C. Light-Mediated Hormonal Regulation of Plant Growth and Development. Annu. Rev. Plant Biol. 2016, 67, 513–537. [Google Scholar] [CrossRef]
- Kami, C.; Lorrain, S.; Hornitschek, P.; Fankhauser, C. Light-Regulated Plant Growth and Development. Current Topics in Developmental Biology 2010, 29–66. [Google Scholar]
- Heijde, M.; Ulm, R. UV-B Photoreceptor-Mediated Signalling in Plants. Trends Plant Sci. 2012, 17, 230–237. [Google Scholar] [CrossRef]
- Pedmale, U.; Huang, S.; Zander, M.; Cole, B.; Hetzel, J.; Ljung, K.; Reis, P.; Sridevi, P.; Nito, K.; Nery, J.; et al. Cryptochromes Interact Directly with PIFs to Control Plant Growth in Limiting Blue Light. Cell 2016, 164, 233–245. [Google Scholar] [CrossRef]
- Xu, P.; Chen, H.; Li, T.; Xu, F.; Mao, Z.; Cao, X.; Miao, L.; Du, S.; Hua, J.; Zhao, J.; et al. Blue Light-Dependent Interactions of CRY1 with GID1 and DELLA Proteins Regulate Gibberellin Signaling and Photomorphogenesis in Arabidopsis. Plant Cell 2021, 33. [Google Scholar] [CrossRef]
- Farooq, M.A.; Ma, W.; Shen, S.; Gu, A. Underlying Biochemical and Molecular Mechanisms for Seed Germination. Int. J. Mol. Sci. 2022, 23, 8502. [Google Scholar] [CrossRef]
- Yang, L.; Liu, S.; Lin, R. The Role of Light in Regulating Seed Dormancy and Germination. J. Integr. Plant Biol. 2020, 62, 1310–1326. [Google Scholar] [CrossRef]
- Cashmore, A. Cryptochromes: Enabling Plants and Animals to Determine Circadian Time. Cell 2003, 114, 537–543. [Google Scholar]
- Ito, S.; Song, Y.H.; Imaizumi, T. LOV Domain-Containing F-Box Proteins: Light-Dependent Protein Degradation Modules in Arabidopsis. Mol. Plant 2012, 5, 573–582. [Google Scholar] [CrossRef]
- Christie, J. Phototropin Blue-Light Receptors. Annu. Rev. Plant Biol. 2007, 58, 21–45. [Google Scholar] [CrossRef] [PubMed]
- Rizzini, L.; Favory, J.; Cloix, C.; Faggionato, D.; O’Hara, A.; Kaiserli, E.; Baumeister, R.; Schäfer, E.; Nagy, F.; Jenkins, G.I.; et al. Perception of UV-B by the Arabidopsis UVR8 Protein. Science 2011, 332, 103–106. [Google Scholar] [CrossRef]
- Balliu, A.; Kacjan Marsic, N.; Gruda, N. Seedling Production. 2017; pp. 189–206.
- Yadav, A.; Singh, D.; Lingwan, M.; Yadukrishnan, P.; Masakapalli, S.K.; Datta, S. Light Signaling and UV-B-mediated Plant Growth Regulation. J. Integr. Plant Biol. 2020, 62, 1270–1292. [Google Scholar] [CrossRef] [PubMed]
- Bantis, F. Light Emitting Diodes (LEDs) Affect Morphological, Physiological and Phytochemical Characteristics of Pomegranate Seedlings. Sci. Hortic. 2018, 234, 267–274. [Google Scholar] [CrossRef]
- Cavallaro, V.; Muleo, R. The Effects of LED Light Spectra and Intensities on Plant Growth. Plants 2022, 11, 1911. [Google Scholar] [CrossRef]
- Lazzarin, M.; Meisenburg, M.; Meijer, D.; van Ieperen, W.; Marcelis, L.F.M.; Kappers, I.F.; van der Krol, A.R.; van Loon, J.J.A.; Dicke, M. LEDs Make It Resilient: Effects on Plant Growth and Defense. Trends Plant Sci. 2021, 26, 496–508. [Google Scholar] [CrossRef]
- Xu, Y.; Yang, M.; Cheng, F.; Liu, S.; Liang, Y. Effects of LED Photoperiods and Light Qualities on in Vitro Growth and Chlorophyll Fluorescence of Cunninghamia Lanceolata. BMC Plant Biol. 2020, 20, 1–12. [Google Scholar] [CrossRef]
- Chen, M.; Chory, J.; Fankhauser, C. Light Signal Transduction in Higher Plants. Annu. Rev. Genet. 2004, 38, 87–117. [Google Scholar] [CrossRef]
- Möglich, A.; Yang, X.; Ayers, R.A.; Moffat, K. Structure and Function of Plant Photoreceptors. Annu. Rev. Plant Biol. 2010, 61, 21–47. [Google Scholar] [CrossRef] [PubMed]
- Shu, K.; Liu, X.-D.; Xie, Q.; He, Z.-H. Two Faces of One Seed: Hormonal Regulation of Dormancy and Germination. Mol. Plant 2016, 9, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Chory, J. Phytochrome Signaling Mechanisms and the Control of Plant Development. Trends Cell Biol. 2011, 21, 664–671. [Google Scholar] [CrossRef]
- Cheng, M.; Kathare, P.; Paik, I.; Huq, E. Phytochrome Signaling Networks. Annu. Rev. Plant Biol. 2021, 72, 217–244. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Samtani, H.; Sahu, K.; Sharma, A.K.; Khurana, J.P.; Khurana, P. International Journal of Biological Macromolecules Functions of Phytochrome-Interacting Factors (PIFs) in the regulation of plant growth and development: A comprehensive review. Int. J. Biol. Macromol. 2023, 244, 125234. [Google Scholar] [CrossRef]
- Pham, V.N.; Kathare, P.K.; Huq, E. Phytochromes and Phytochrome Interacting Factors. Plant Physiol. 2018, 176, 1025–1038. [Google Scholar] [CrossRef]
- Paik, I.; Huq, E. Plant Photoreceptors: Multi-Functional Sensory Proteins and Their Signaling Networks. Semin. Cell Dev. Biol. 2019, 92, 114–121. [Google Scholar] [CrossRef]
- Rockwell, N.C.; Su, Y.-S.; Lagarias, J.C. Phytochrome Structure and Signaling Mechanisms. Annu. Rev. Plant Biol. 2006, 57, 837–858. [Google Scholar] [CrossRef]
- Leivar, P.; Monte, E. PIFs: Systems Integrators in Plant Development. Plant Cell 2014, 26, 56–78. [Google Scholar] [CrossRef]
- Wang, P.; Abid, M.A.; Qanmber, G.; Askari, M.; Zhou, L.; Song, Y.; Liang, C.; Meng, Z.; Malik, W.; Wei, Y.; et al. Photomorphogenesis in plants: The central role of phytochrome interacting factors (PIFs). Environ. Exp. Bot. 2022, 194, 104704. [Google Scholar] [CrossRef]
- Lee, K.P.; Piskurewicz, U.; Turečková, V.; Carat, S.; Chappuis, R.; Strnad, M.; Fankhauser, C.; Lopez-Molina, L. Spatially and Genetically Distinct Control of Seed Germination by Phytochromes A and B. Genes Dev. 2012, 26, 1984–1996. [Google Scholar] [CrossRef]
- Bian, Y.; Chu, L.; Lin, H.; Qi, Y.; Fang, Z.; Xu, D. PIFs- and COP1-HY5-Mediated Temperature Signaling in Higher Plants. Stress Biol. 2022, 2, 35. [Google Scholar] [CrossRef]
- Kim, J.; Song, J.; Seo, H. COP1 Regulates Plant Growth and Development in Response to Light at the Post-Translational Level. J. Exp. Bot. 2017, 68, 4737–4748. [Google Scholar] [CrossRef]
- Lee, J.; Jung, J.; Park, C. Light Inhibits COP1-Mediated Degradation of ICE Transcription Factors to Induce Stomatal Development in Arabidopsis. Plant Cell 2017, 29, 2817–2830. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Chu, L.; Zhang, Y.; Bian, Y.; Xiao, J.; Xu, D. HY5: A Pivotal Regulator of Light-Dependent Development in Higher Plants. Front. Plant Sci. 2022, 12, 3294. [Google Scholar] [CrossRef] [PubMed]
- Hart, J.E.; Gardner, K.H. Lighting the way: Recent insights into the structure and regulation of phototropin blue light receptors. J. Biol. Chem. 2021, 296, 100594. [Google Scholar] [CrossRef] [PubMed]
- Paik, I.; Huq, E. Plant Photoreceptors: Multi-Functional Sensory Proteins and Their Signaling Networks. Semin. Cell Dev. Biol. 2019, 92, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Banaś, A.K.; Aggarwal, C.; Łabuz, J.; Sztatelman, O.; Gabryś, H. Blue Light Signalling in Chloroplast Movements. J. Exp. Bot. 2012, 63, 1559–1574. [Google Scholar] [CrossRef]
- Pipitone, R.; Eicke, S.; Pfister, B.; Glauser, G.; Falconet, D.; Uwizeye, C.; Pralon, T.; Zeeman, S.C.; Kessler, F.; Demarsy, E. A multifaceted analysis reveals two distinct phases of chloroplast biogenesis during de-etiolation in arabidopsis. Elife 2021, 10, 1–32. [Google Scholar] [CrossRef]
- Barrero, J.M.; Downie, A.B.; Xu, Q.; Gubler, F. A Role for Barley CRYPTOCHROME1 in Light Regulation of Grain Dormancy and Germination. Plant Cell 2014, 26, 1094–1104. [Google Scholar] [CrossRef]
- Mao, Z.; He, S.; Xu, F.; Wei, X.; Jiang, L.; Liu, Y.; Wang, W.; Li, T.; Xu, P.; Du, S.; et al. Photoexcited CRY1 and PhyB Interact Directly with ARF6 and ARF8 to Regulate Their DNA-Binding Activity and Auxin-Induced Hypocotyl Elongation in Arabidopsis. New Phytol. 2020, 225, 848–865. [Google Scholar] [CrossRef]
- Gao, L.; Liu, Q.; Zhong, M.; Zeng, N.; Deng, W.; Li, Y.; Wang, D.; Liu, S.; Wang, Q. Blue light-induced phosphorylation of Arabidopsis cryptochrome 1 is essential for its photosensitivity. J. Integr. Plant Biol. 2022, 64, 1724–1738. [Google Scholar] [CrossRef] [PubMed]
- Martínez, C.; Nieto, C.; Prat, S. Convergent Regulation of PIFs and the E3 Ligase COP1/SPA1 Mediates Thermosensory Hypocotyl Elongation by Plant Phytochromes. Curr. Opin. Plant Biol. 2018, 45, 188–203. [Google Scholar] [CrossRef]
- Stawska, M.; Oracz, K. PhyB and HY5 Are Involved in the Blue Light-Mediated Alleviation of Dormancy of Arabidopsis Seeds Possibly via the Modulation of Expression of Genes Related to Light, GA, and ABA. Int. J. Mol. Sci. 2019, 20, 5882. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, L.; Chen, P.; Liang, T.; Li, X.; Liu, H. UV-B Photoreceptor UVR8 Interacts with MYB73/MYB77 to Regulate Auxin Responses and Lateral Root Development. EMBO J. 2020, 39, e101928. [Google Scholar] [CrossRef] [PubMed]
- Biever, J.J.; Gardner, G. The Relationship between Multiple UV-B Perception Mechanisms and DNA Repair Pathways in Plants. Environ. Exp. Bot. 2016, 124, 89–99. [Google Scholar] [CrossRef]
- Yadav, A.; Singh, D.; Lingwan, M.; Yadukrishnan, P.; Masakapalli, S.K.; Datta, S. Light Signaling and UV-B-Mediated Plant Growth Regulation. J. Integr. Plant Biol. 2020, 62, 1270–1292. [Google Scholar] [CrossRef]
- Huché-Thélier, L.; Crespel, L.; Gourrierec, J.L.; Morel, P.; Sakr, S.; Leduc, N. Light Signaling and Plant Responses to Blue and UV Radiations—Perspectives for Applications in Horticulture. Environ. Exp. Bot. 2016, 121, 22–38. [Google Scholar] [CrossRef]
- Ravindran, P.; Kumar, P.P. Regulation of Seed Germination: The Involvement of Multiple Forces Exerted via Gibberellic Acid Signaling. Mol. Plant 2019, 12, 24–26. [Google Scholar] [CrossRef]
- Tuan, P.A.; Kumar, R.; Rehal, P.K.; Toora, P.K.; Ayele, B.T. Molecular Mechanisms Underlying Abscisic Acid/Gibberellin Balance in the Control of Seed Dormancy and Germination in Cereals. Front. Plant Sci. 2018, 9, 668. [Google Scholar] [CrossRef]
- de Wit, M.; Galvão, V.C.; Fankhauser, C. Light-Mediated Hormonal Regulation of Plant Growth and Development. Annu. Rev. Plant Biol. 2016, 67, 513–537. [Google Scholar] [CrossRef]
- Née, G.; Xiang, Y.; Soppe, W. The Release of Dormancy, a Wake-up Call for Seeds to Germinate. Curr. Opin. Plant Biol. 2017, 35, 8–14. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, Y.; Zheng, Y. Integration of ABA, GA, and light signaling in seed germination through the regulation of ABI5. Front. Plant Sci. 2022, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fantini, E.; Facella, P. Cryptochromes in the Field: How Blue Light Influences Crop Development. Physiol. Plant. 2020, 169, 336–346. [Google Scholar] [CrossRef] [PubMed]
- El-Keblawy, A.; Elgabra, M.; Mosa, K.A.; Fakhry, A.; Soliman, S. Roles of hardened husks and membranes surrounding Brachypodium hybridum grains on germination and seedling growth. Plants 2019, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Goggin, D.E.; Steadman, K.J.; Powles, S.B. Green and Blue Light Photoreceptors Are Involved in Maintenance of Dormancy in Imbibed Annual Ryegrass (Lolium Rigidum) Seeds. New Phytol. 2008, 180, 81–89. [Google Scholar] [CrossRef]
- Yang, L.; Liu, S.; Lin, R. The role of light in regulating seed dormancy and germination. J. Integr. Plant Biol. 2020, 62, 1310–1326. [Google Scholar] [CrossRef]
- Jacobsen, J.V.; Barrero, J.M.; Hughes, T.; Julkowska, M.; Taylor, J.M.; Xu, Q.; Gubler, F. Roles for Blue Light, Jasmonate and Nitric Oxide in the Regulation of Dormancy and Germination in Wheat Grain (Triticum Aestivum L.). Planta 2013, 238, 121–138. [Google Scholar] [CrossRef]
- Simlat, M.; Ślęzak, P.; Moś, M.; Warchoł, M.; Skrzypek, E.; Ptak, A. The Effect of Light Quality on Seed Germination, Seedling Growth and Selected Biochemical Properties of Stevia Rebaudiana Bertoni. Sci. Hortic. 2016, 211, 295–304. [Google Scholar] [CrossRef]
- Lymperopoulos, P.; Msanne, J.; Rabara, R. Phytochrome and Phytohormones: Working in Tandem for Plant Growth and Development. Front. Plant Sci. 2018, 9, 1037. [Google Scholar] [CrossRef]
- Arana, M.V.; Sánchez-Lamas, M.; Strasser, B.; Ibarra, S.E.; Cerdán, P.D.; Botto, J.F.; Sánchez, R.A. Functional Diversity of Phytochrome Family in the Control of Light and Gibberellin-Mediated Germination in Arabidopsis. Plant Cell Environ. 2014, 37, 2014–2023. [Google Scholar] [CrossRef]
- Vaistij, F.E.; Barros-Galvão, T.; Cole, A.F.; Gilday, A.D.; He, Z.; Li, Y.; Harvey, D.; Larson, T.R.; Graham, I.A. MOTHER-OF-FT-AND-TFL1 Represses Seed Germination under Far-Red Light by Modulating Phytohormone Responses in Arabidopsis Thaliana. Proc. Natl. Acad. Sci. U. S. A. 2018, 115, 8442–8447. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.; Ryu, J.; Jeong, Y.; Park, J.; Song, J.; Amasino, R.M.; Noh, B.; Noh, Y. Control of Seed Germination by Light-Induced Histone Arginine Demethylation Activity. Dev. Cell 2012, 22, 736–748. [Google Scholar] [CrossRef] [PubMed]
- Albertos, P.; Wlk, T.; Griffiths, J.; Pimenta Lange, M.J.; Unterholzner, S.J.; Rozhon, W.; Lange, T.; Jones, A.M.; Poppenberger, B. Brassinosteroid-regulated bHLH transcription factor CESTA induces the gibberellin 2-oxidase GA2ox7. Plant Physiol. 2022, 188, 2012–2025. [Google Scholar] [CrossRef]
- Jiang, Z.; Xu, G.; Jing, Y.; Tang, W.; Lin, R. Phytochrome B and REVEILLE1/2-Mediated Signalling Controls Seed Dormancy and Germination in Arabidopsis. Nat. Commun. 2016, 7, 12377. [Google Scholar] [CrossRef]
- Ahmad, M.; Cashmore, A.R. HY4 Gene of A. Thaliana Encodes a Protein with Characteristics of a Blue-Light Photoreceptor. Nature 1993, 366, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Cao, K.; Hao, Y.; Song, S.; Su, W.; Liu, H. Hypocotyl Elongation Is Regulated by Supplemental Blue and Red Light in Cucumber Seedling. Gene 2019, 707, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Zhong, M.; Zeng, B.; Tang, D.; Yang, J.; Qu, L.; Yan, J.; Wang, X.; Li, X.; Liu, X.; Zhao, X. The blue light receptor CRY1 interacts with GID1 and DELLA proteins to repress GA signaling during photomorphogenesis in Arabidopsis. Mol. Plant 2021, 14, 1328–1342. [Google Scholar] [CrossRef] [PubMed]
- Ponnu, J.; Hoecker, U. Illuminating the COP1/SPA Ubiquitin Ligase: Fresh Insights into Its Structure and Functions During Plant Photomorphogenesis. Front. Plant Sci. 2021, 12, 1–19. [Google Scholar] [CrossRef]
- Dong, J.; Tang, D.; Gao, Z.; Yu, R.; Li, K.; He, H.; Terzaghi, W.; Deng, X.W.; Chen, H. Arabidopsis DE-ETIOLATED1 Represses Photomorphogenesis by Positively Regulating Phytochrome-Interacting Factors in the Dark. Plant Cell 2014, 26, 3630–3645. [Google Scholar]
- Sun, X.; Ni, M. HYPOSENSITIVE TO LIGHT, an Alpha/Beta Fold Protein, Acts Downstream of ELONGATED HYPOCOTYL 5 to Regulate Seedling De-Etiolation. Mol. Plant 2011, 4, 116–126. [Google Scholar] [CrossRef]
- Yadav, A.; Singh, D.; Lingwan, M.; Yadukrishnan, P.; Masakapalli, S.K.; Datta, S. Light signaling and UV-B-mediated plant growth regulation. J. Integr. Plant Biol. 2020, 62, 1270–1292. [Google Scholar] [CrossRef]
- Zhou, J.; Liu, Q.; Zhang, F.; Wang, Y.; Zhang, S.; Cheng, H.; Yan, L.; Li, L.; Chen, F.; Xie, X. Overexpression of OsPIL15, a Phytochrome-Interacting Factor-like Protein Gene, Represses Etiolated Seedling Growth in Rice. J. Integr. Plant Biol. 2014, 56, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, H. Coordinated Shoot and Root Responses to Light Signaling in Arabidopsis. Plant Commun. 2020, 1, 100026. [Google Scholar] [CrossRef] [PubMed]
- Delker, C.; Sonntag, L.; James, G.V.; Janitza, P.; Ibañez, C.; Ziermann, H.; Peterson, T.; Denk, K.; Mull, S.; Ziegler, J.; et al. The DET1-COP1-HY5 Pathway Constitutes a Multipurpose Signaling Module Regulating Plant Photomorphogenesis and Thermomorphogenesis. Cell Rep. 2014, 9, 1983–1989. [Google Scholar] [CrossRef] [PubMed]
- Hoecker, U. The Activities of the E3 Ubiquitin Ligase COP1/SPA, a Key Repressor in Light Signaling. Curr. Opin. Plant Biol. 2017, 37, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Yin, R.; Ulm, R. How Plants Cope with UV-B: From Perception to Response. Curr. Opin. Plant Biol. 2017, 37, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Sassi, M.; Lu, Y.; Zhang, Y.; Wang, J.; Dhonukshe, P.; Blilou, I.; Dai, M.; Li, J.; Gong, X.; Jaillais, Y.; et al. COP1 Mediates the Coordination of Root and Shoot Growth by Light through Modulation of PIN1- and PIN2-Dependent Auxin Transport in Arabidopsis. Development 2012, 139, 3402–3412. [Google Scholar] [CrossRef]
- Xiao, L.; Shibuya, T.; Kato, K.; Nishiyama, M.; Kanayama, Y. Effects of Light Quality on Plant Development and Fruit Metabolism and Their Regulation by Plant Growth Regulators in Tomato. Sci. Hortic. 2022, 300, 111076. [Google Scholar] [CrossRef]
- Gelderen, K.; Kang, C.; Paalman, R.; Keuskamp, D.; Hayes, S.; Pierik, R. Far-Red Light Detection in the Shoot Regulates Lateral Root Development through the HY5 Transcription Factor. Plant Cell 2018, 30, 101–116. [Google Scholar] [CrossRef]
- Xiong, J.; Yang, F.; Wei, F.; Yang, F.; Lin, H.; Zhang, D. Inhibition of SIZ1-mediated SUMOylation of HOOKLESS1 promotes light-induced apical hook opening in Arabidopsis. Plant Cell 2023, 35, 2027–2043. [Google Scholar] [CrossRef]
- Xiong, H.; Lu, D.; Li, Z.; Wu, J.; Ning, X.; Lin, W.; Bai, Z.; Zheng, C.; Sun, Y.; Chi, W.; et al. The DELLA-ABI4-HY5 module integrates light and gibberellin signals to regulate hypocotyl elongation. Plant Commun. 2023, 100597. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.G.; Muneer, S.; Jeong, B.R. Morphogenesis, flowering, and gene expression of Dendranthema grandiflorum in response to shift in light quality of night interruption. Int. J. Mol. Sci. 2015, 16, 16497–16513. [Google Scholar] [CrossRef] [PubMed]
- Pierik, R.; Ballaré, C.L. Control of Plant Growth and Defense by Photoreceptors: From Mechanisms to Opportunities in Agriculture. Mol. Plant 2021, 14, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Fraser, D.P.; Hayes, S.; Franklin, K.A. Photoreceptor Crosstalk in Shade Avoidance. Curr. Opin. Plant Biol. 2016, 33, 1–7. [Google Scholar] [CrossRef]
- Gommers, C.M.M.; Visser, E.J.W.; Onge, K.R.S.; Voesenek, L.A.C.J.; Pierik, R. Shade Tolerance: When Growing Tall Is Not an Option. Trends Plant Sci. 2013, 18, 65–71. [Google Scholar] [CrossRef]
- Mukherjee, A.; Dwivedi, S.; Bhagavatula, L.; Datta, S. Integration of light and ABA signaling pathways to combat drought stress in plants. Plant Cell Rep. 2023, 42, 829–841. [Google Scholar] [CrossRef]
- Matthews, J.; Vialet-Chabrand, S.; Lawson, T. Role of Blue and Red Light in Stomatal Dynamic Behaviour. J. Exp. Bot. 2019, 71, 2253–2269. [Google Scholar] [CrossRef]
- Wei, H.; Kong, D.; Yang, J.; Wang, H. Light Regulation of Stomata Development and Patterning: Shifting the Paradigm from Arabidopsis to Grasses. Plant Commun. 2020, 1, 100030. [Google Scholar] [CrossRef]
- Delgado, D.; Ballesteros, I.; Torres-Contreras, J.; Mena, M.; Fenoll, C. Dynamic Analysis of Epidermal Cell Divisions Identifies Specific Roles for COP10 in Arabidopsis Stomatal Lineage Development. Planta 2012, 236, 447–461. [Google Scholar] [CrossRef]
- Kreiss, M.; Haas, F.B.; Hansen, M.; Rensing, S.A.; Hoecker, U. Co-action of COP1, SPA and cryptochrome in light signal transduction and photomorphogenesis of the moss Physcomitrium patens. Plant J. 2023, 159–175. [Google Scholar] [CrossRef]
- Lampard, G.R.; Macalister, C.A.; Bergmann, D.C. Arabidopsis Stomatal Initiation Is Controlled by MAPK-Mediated Regulation of the BHLH SPEECHLESS. Science 2008, 322, 1113–1116. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Wang, P.; Si, T.; Hsu, C.; Wang, L.; Zayed, O.; Yu, Z.; Zhu, Y.; Dong, J.; Tao, W.A.; et al. MAP Kinase Cascades Regulate the Cold Response by Modulating ICE1 Protein Stability. Dev. Cell 2017, 43, 618–629.e5. [Google Scholar] [CrossRef] [PubMed]
- Pham, V.N.; Kathare, P.K.; Huq, E. Phytochromes and Phytochrome Interacting Factors. Plant Physiol. 2018, 176, 1025–1038. [Google Scholar] [CrossRef]
- Lau, O.S.; Song, Z.; Zhou, Z.; Davies, K.A.; Chang, J.; Yang, X.; Wang, S.; Lucyshyn, D.; Tay, I.H.Z.; Wigge, P.A.; et al. Direct Control of SPEECHLESS by PIF4 in the High-Temperature Response of Stomatal Development. Curr. Biol. CB 2018, 28, 1273–1280. [Google Scholar] [CrossRef]
- Klermund, C.; Ranftl, Q.L.; Diener, J.; Bastakis, E.; Richter, R.; Schwechheimer, C. LLM-Domain B-GATA Transcription Factors Promote Stomatal Development Downstream of Light Signaling Pathways in Arabidopsis Thaliana Hypocotyls. Plant Cell 2016, 28, 646–660. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhou, B.; Li, J.; Tang, H.; Tang, J.; Yang, Z. Formation and Change of Chloroplast-Located Plant Metabolites in Response to Light Conditions. Int. J. Mol. Sci. 2018, 19, 654. [Google Scholar] [CrossRef]
- Chory, J. Light Signals in Leaf and Chloroplast Development: Photoreceptors and Downstream Responses in Search of a Transduction Pathway. New Biol. 1991, 3, 538–548. [Google Scholar]
- Yang, E.J.; Yoo, C.Y.; Liu, J.; Wang, H.; Cao, J.; Li, F.; Pryer, K.M.; Sun, T.; Weigel, D.; Zhou, P.; et al. NCP Activates Chloroplast Transcription by Controlling Phytochrome-Dependent Dual Nuclear and Plastidial Switches. Nat. Commun. 2019, 10, 2630. [Google Scholar] [CrossRef]
- Yoo, C.Y.; Pasoreck, E.K.; Wang, H.; Cao, J.; Blaha, G.M.; Weigel, D.; Chen, M. Phytochrome Activates the Plastid-Encoded RNA Polymerase for Chloroplast Biogenesis via Nucleus-to-Plastid Signaling. Nat. Commun. 2019, 10, 2629. [Google Scholar] [CrossRef]
- Mellenthin, M.; Ellersiek, U.; Börger, A.; Baier, M. Expression of the Arabidopsis Sigma Factor SIG5 Is Photoreceptor and Photosynthesis Controlled. Plants 2014, 3, 359–391. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zheng, Y.; Liu, H.; Zhang, Y.; Hao, Y.; Song, S.; Lei, B. Effect of Supplemental Blue Light Intensity on the Growth and Quality of Chinese Kale. Hortic. Environ. Biotechnol. 2019, 60, 49–57. [Google Scholar]
- Li, Y.; Wu, L.; Jiang, H.; He, R.; Song, S.; Su, W.; Liu, H. Supplementary Far-Red and Blue Lights Influence the Biomass and Phytochemical Profiles of Two Lettuce Cultivars in Plant Factory. Molecules 2021, 26, 7405. [Google Scholar] [CrossRef] [PubMed]
- Silva, T.; Batista, D.; Fortini, E.; Castro, K.; Sousa Felipe, S.; Fernandes, A.; De Jesus Sousa, R.M.; Chagas, K.; Da Silva, J.; Correia, L.; et al. Blue and Red Light Affects Morphogenesis and 20-Hydroxyecdisone Content of in Vitro Pfaffia Glomerata Accessions. J. Photochem. Photobiol. B: Biology 2019, 203, 111761. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
A schematic diagram depicting the involvement of light in different stages of photomorphogenesis. UV resistance Locus 8 (UVR8), cryptochromes (CRYs), phototropins (PHOTs), Zeitlupe family proteins (ZTL/FKF1/LKP2), and phytochromes (PHYs).
Figure 1.
A schematic diagram depicting the involvement of light in different stages of photomorphogenesis. UV resistance Locus 8 (UVR8), cryptochromes (CRYs), phototropins (PHOTs), Zeitlupe family proteins (ZTL/FKF1/LKP2), and phytochromes (PHYs).
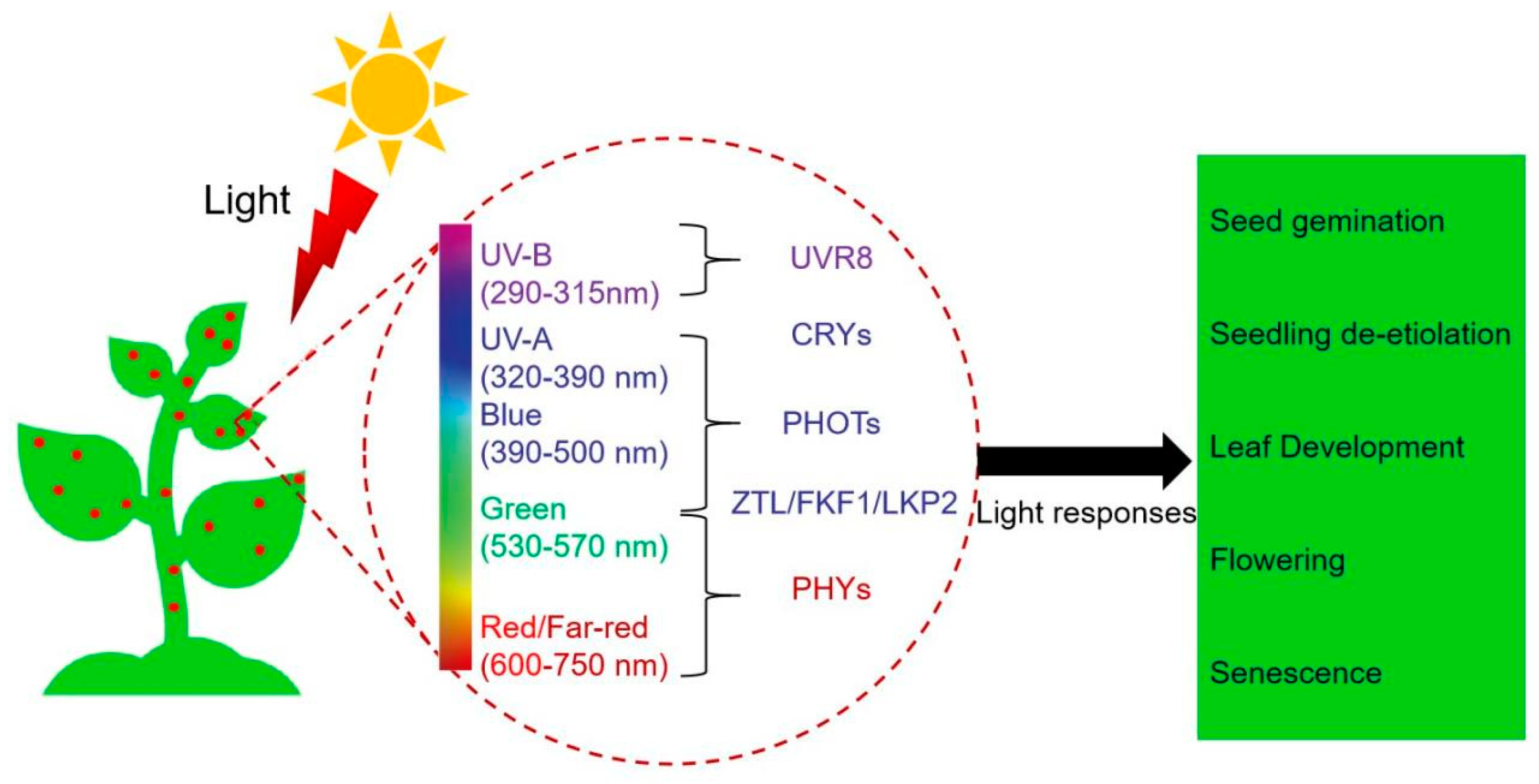
Figure 2.
Light-mediated signaling network in regulating seed germination.
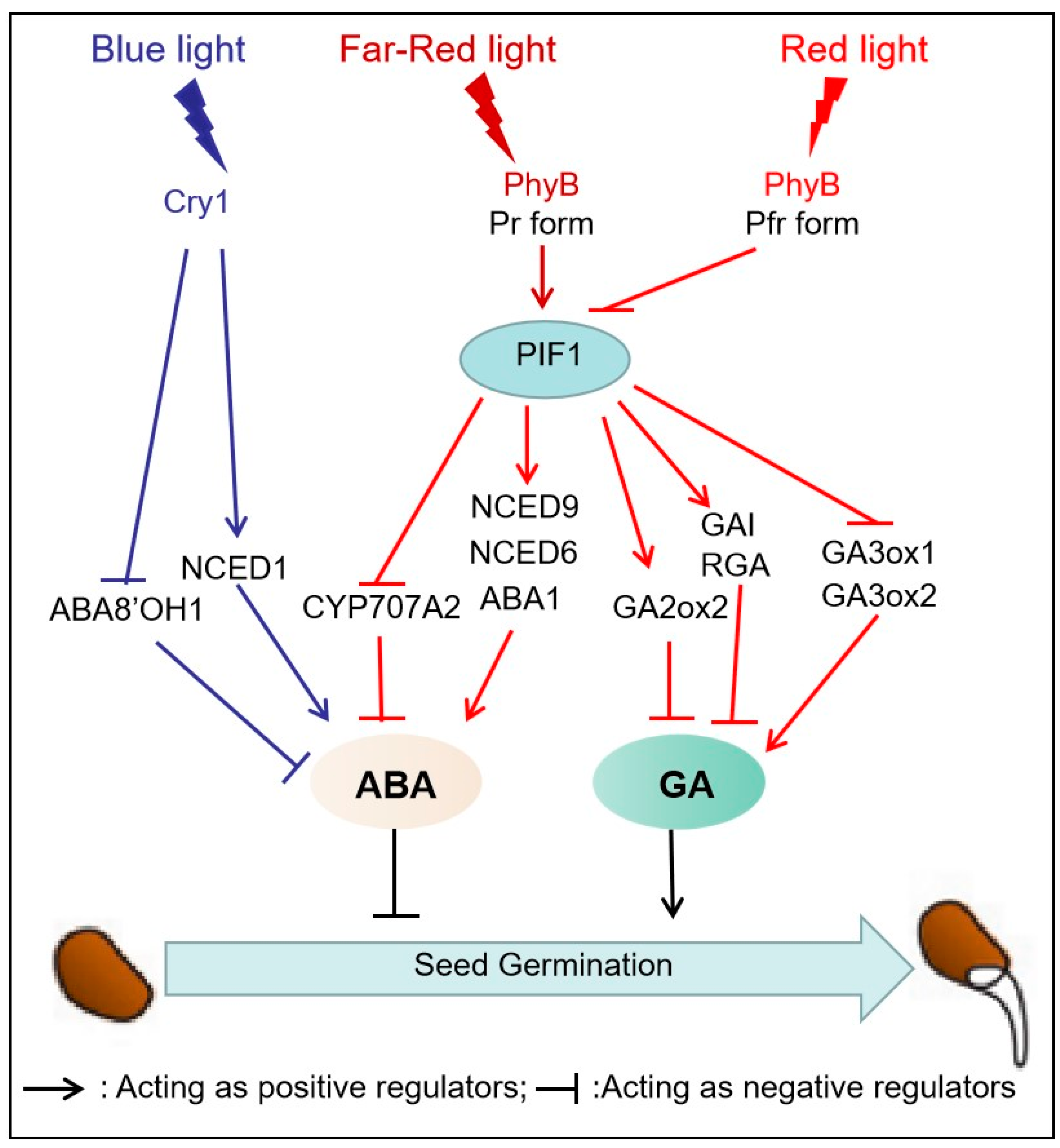
Figure 3.
Light-mediated signaling network in regulating seedling de-etiolation.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



