
Submitted:
24 June 2023
Posted:
25 June 2023
You are already at the latest version
Abstract
Canine mammary tumors (CMTs) are among the most common disease in female dogs and share similarities with human breast cancer, which have placed these animals as a model of study in comparative oncology. Metabolic reprogramming is one of them and is known as a hallmark of carcinogenesis whereby cells undergo adjustments to supply the high bioenergetic and biosynthetic demands of rapidly proliferating cells. However, such alterations are also vulnerabilities that may serve as therapeutic strategies which have been tested mostly in human clinical trials, but poorly explored in CMTs. In this dedicated review, we compilated the metabolic changes described for CMTs emphasizing the metabolism of carbohydrates, amino acids, lipids, and mitochondrial function. We observed here key factors associated with the presence and aggressiveness of CMTs, such as the increase in glucose uptake followed by enhanced anaerobic glycolysis via upregulation of glycolytic enzymes, changes in glutamine catabolism due to overexpression of glutaminases, raise in fatty acid oxidation and distinct effects depending on the lipid saturation, in addition to mitochondrial DNA which is a hotspot for mutations. Therefore, more attention should be paid to this topic given that targeting metabolic fragilities could improve the outcome of CMTs.
Keywords:
canine mammary tumors
; metabolism
; cancer
; mitochondria
; metabolic reprogramming
; glucose
; amino acids
; lipids
1. Introduction
Canine mammary tumors (CMTs) are the most common cancer in female dogs and arise spontaneously with increased risk along aging [1]. Several elements contribute to its occurrence and epidemiological, clinical, histological and molecular contexts may drive breast neoplasms formation [2]. The literature frequently reports age, breed, reproductive and hormonal status as the main risk factors for CMT. Such disease generally affects dogs above middle age, with an increased risk between 8 and 11 years. Sexual hormones deregulation, such as exposure to endogenous ovarian hormones, may cause the mammary tumors development in dogs [3,4,5], but others may also influence CMT such as obesity in early stages of life [6], inflammation [7] and the increase in free radicals and reactive oxygen species (ROS) [8]. They may appear as single or multiple nodules, and posterior mammary glands are more frequently affected than anterior [9]. Approximately 50% of CMTs are malignant [10] and the most common tumor type among them is tubular carcinoma (adenocarcinoma), followed by papillary, solid and, complex carcinomas in addition to carcinosarcoma. The other half is benign, being fibroadenomas, ductal papillomas, benign mixed tumors, and simple adenomas the most incidents [11]. Despite such classification, it is also common to find more than one tumor type in different mammary glands of the same patient [12] and CMTs upon metastasis usually show tropism to the lymph nodes area and lungs [6].
CMTs are the leading cause of death in aged female dogs and has a higher incidence compared to human breast cancer, mainly in low-income countries [13,14]. Despite of that, they share similarities like the hormonal influence on carcinogenesis, age of onset, capability to do long-distance metastasis and high heterogeneity [1,15,16]. CMTs are highly prevalent in non-spayed female dogs, with a significant proportion having premalignant lesions. Those with such lesions can progress to invasive cancer within a relatively short period of time. This scenario of high incidence and in most cases with poor prognosis highlights the need for new assertive therapeutic approaches. A remarkable alteration commonly documented among all cancers is the metabolic reprogramming, which supply the bioenergetic demands and the anabolic requirements for cell proliferation and tumor growth. Although it has been widely investigated in human breast cancer, there is a lack of information on metabolic rewiring in CMTs. In this review, we recapitulated metabolic adjustments described in the literature that could serve as therapeutic strategy with focus on lipid, glucose and amino acids metabolism deregulation in CMT.
2. Metabolic features of mammary cancer
Cancer cells undergo several alterations to reach the imbalance between higher proliferative rate and cell death, providing a molecular context whereby neoplastic cells promote tumor growth and even metastasis. Among such adjustments is metabolic reprogramming, one of the hallmarks of carcinogenesis, that for instance supply the bioenergetic and biosynthetic demands of rapid cell proliferation [17,18,19]. Otto Warburg described in 1956 that tumor cells uptake more glucose and use it as energy source through aerobic glycolysis due to the impairment of mitochondria [20]. This was later revisited and demonstrated that the mitochondria has its activity decreased and not impaired, different from originally proposed as the Warburg effect [21]. The increase in glycolysis, a very inefficient ATP source compared to oxidative phosphorylation (OXPHOS) [22], in addition to energy production also provides intermediates to anabolic pathways, such as nucleic acid synthesis and NADPH, all of them required for cell division and tumor growth [21]. Also, glutamine is another carbon source that fuels the tricarboxylic acid cycle (TCA) via reductive carboxylation [23], being often required for cell survival mostly in advanced stages in many cancers [23,24,25]. Glutamine uptake and consumption increase with oxidative stress [26,27] and may be used for fatty acid synthesis which in turn provides phospholipids to membrane synthesis required for cell cycle progression [28]. In addition to glucose and glutamine, lipids are also crucial, if not required, for cancer cell survival and progression to more aggressive stages [29,30,31,32]. Lipids are building blocks to membrane synthesis, post-translational modification, and energy generation. It has been reported that endogenously synthesized fatty acids are required for cancer cell proliferation [28] and the pharmacological inhibition of fatty acid synthase (FASN) has antitumor effects in cell lines, organoids, and xenograft models [31]. Interestingly, not only endogenous but also exogenous fatty acids obtained through diet have been shown to modulate cancer progression [33], including through metabolism regulation [34]. Taken together, this short summary highlights that the metabolism rewiring is a vulnerability that has been explored as a therapeutic opportunity in several cancers.
In the literature, most of available information on tumor metabolism was generated from human breast cancer but given the similarities with CMT is reasonable to consider that some of the alterations can be found in dogs. In breast cancer, tumor cells was shown to display high glucose uptake compared to those exhibiting normal phenotype [35] and may regulate such nutrient availability to favor metastasis [36]. Breast cancer cells have glucose transporters (GLUTs) overexpression [37], being GLUT1 related to poor prognosis and higher proliferation. However, it was reported that GLUTs overexpression has distinct pattern in human cell lines [37,38], suggesting that although glucose uptake is increased, there are adjustments depending on the molecular context. Glucose oxidation changes among breast cancer progression since less aggressive subtypes show increased oxygen consumption rate and decreased extracellular acidification, which was found as opposite in more aggressive [37]. Indeed, several glycolytic enzymes are deregulated in human breast cancer, including PKM2 (pyruvate kinase muscle isozyme 2) [39], PFK (phosphofructokinase) [40,41] and HK2 (hexokinase 2) [42,43], and pharmacological inhibition of HK2 has been investigated as therapeutic strategy [44]. Supporting the glycolytic phenotype in more aggressive subtypes with poor prognosis, it was reported higher expression of lactate dehydrogenase B (LDHB) and lactate transporters (MCT1 and MCT4) [45,46], and inhibition of lactate secretion to the microenvironment suppresses tumor growth [47]. Therefore, this body of evidence shows that the critical point of glucose metabolism is the key enzymes that regulate glycolysis functioning.
Lipids and amino acid metabolism are also dysregulated. Studies have reported that the amino acid profile in tumors and blood is distinct from healthy condition [48,49,50]. Especially, as glucose, glutamine, which is the most abundant amino acid in the bloodstream, is a source of carbon for the generation of reduced coenzymes in the TCA cycle that fuels mitochondrial respiration and biosynthetic pathways. Glutaminolysis is performed mainly by glutaminase 1 (GLS) and glutaminase 2 (GLS2), both with mitochondrial localization [51,52,53]. Glutaminases convert glutamine into glutamate via reductive carboxylation, which enters TCA as α-ketoglutarate [51]. In aggressive stages of breast cancer, GLS is overexpressed and cells display dependency on glutamine to survive [53]. On the other hand, GLS2 is mostly expressed in hormone positive cells and has been investigated as a protumorigenic gene [25]. This scenario served as inspiration for clinical trials with glutaminases inhibitors [54,55,56], but further studies are required. Carbons from glutamine may be channeled to fatty acid synthesis and further processing into phospholipids for biological membranes. Lipid synthesis may occur endogenously from either glutamine or glucose, a biological process named de novo lipogenesis (DNL), which is performed by FASN and Acyl-CoA carboxylase (ACC). In breast cancer, lipid metabolism is deregulated in synthesis, oxidation, and uptake [57,58]. The less aggressive subtype relies mostly on DNL while more aggressive does on exogenous fatty acids, displaying an expression profile of proteins related with triacylglycerol formation and lipid droplets biogenesis [59,60], and rely on fatty acid oxidation (FAO) to supply their bioenergetic demand [61]. However, even FASN inhibition has antitumor effect even at low protein levels of expression [62,63]. Interestingly, DNL is required for brain metastasis in humans [29].
The present scenario built mostly with findings in human breast cancer demonstrates that metabolic reprogramming could be a fragility, therefore becoming a therapeutic opportunity. Moreover, the literature reported several key points in the mammary gland cell metabolism showing that plenty of pathways, either catabolic or anabolic, are deregulated. However, such alterations are poorly explored in the canine mammary gland and put a gap of information in the veterinary community (Figure 1). Therefore, this review aimed to put together the “state-of-the-art” of metabolic adjustments described in CMTs that could be a target in cancer treatment and prevention.
3. Metabolic reprogramming in canine mammary tumors
3.1. Carbohydrates
Glucose metabolism is among the frequently altered metabolic pathways in human breast cancer [36,37,64]. Although information in bitches is not deeply understood, the same occurs in canine nonhematopoietic tumors, including the mammary tumor [65]. Higher blood glucose levels were reported as increased in bitches bearing mammary neoplasms and it seems to be related with the presence of tumor cells, given that upon mastectomy, the levels drop considerably [66]. This has been linked to higher lactate production and release by cancer cells, a well-known feature of the Warburg effect, which is converted to glucose through gluconeogenesis in the liver. Jayasri and colleagues (2016) [67] reported deregulation of glycolysis intermediates and enzymes in mammary tumors compared with control samples which support increase in glucose uptake and processing as well as channeling to biosynthetic pathways. In this study, the authors found that mammary tumors had an increase in the tissue regarding the hexose levels by 1.75-fold and 1.6-fold for glycolytic enzyme HK, both in the tissue. The expression of pyruvate kinase 2 (PKM2), the last enzyme of the glycolytic cascade, was also overexpressed in malignant tissue, being its levels correlated with tumor grade [68]. These findings corroborate previous data that reported increased expression in glycolytic enzymes [69] and also supported the increase in GLUT1 expression observed in complex mammary carcinoma [70]. Interestingly, higher GLUT1 expression is detected under hypoxia, which is often observed in large tumors [71]. The role of glucose metabolism was found to be not only limited to energy supply for tumor growth, but also a signaling pathway among cells within the tumor. The higher glycolysis rate in CMT cells increases lactate release which in turn polarizes macrophages into M2 subtype while promoting vascular endothelial growth factor expression [72]. M2-like macrophages are known for their anti-inflammatory phenotype and immune escape which is correlated with tumor progression and aggressiveness [73,74]. In addition, not only glucose breakdown-related metabolites were altered, but also those related with derivative pathways. Glucose-6-phosphatase and fructose-1,6-bisphosphatase were found increased in CMT [67], both related with gluconeogenesis, in addition to transketolase (TKT) and transketolase-like 1 (TKTL1), key enzymes in the pentose phosphate pathway (PPP) indicating that carbohydrates catabolism and anabolism are essential to supply the demands of tumor growth [75]. Also, TLK and TLK-1 were shown to be differentially expressed over cancer stages, being the former highly expressed in benign and malignant tumors compared to the normal tissue, while TLK-1 levels were higher mainly in hyperplastic lesions, simple adenomas, and simple carcinomas [75]. Indeed, PPP enzymes were show to be deregulated in CMT, but the estrogen context seems to play a role because ER-positive cells had low expression of glucose-6-phosphate dehydrogenase (G6PDH), whereas ER-negative cells had higher levels [76].
The glucose metabolism reprogramming is a hallmark in human carcinogenesis and offers a tool for diagnosis, including in the veterinary approach. Since tumors usually uptake higher levels of glucose, 18F-FDG-PET/CT has been adopted for imaging. Sánchez and colleagues (2019) [77] showed that such technique is useful in distinguishing malignant from benign tumors, but not correlating the levels of glucose uptake with tumor grade. Also, it has been a powerful tool in determining the tumor subtype, treatment protocol, improvement of staging and restaging [78,79,80]. Moreover, given that glucose uptake is increased, metformin, a drug to control glucose levels in diabetes, has been tested as an antiproliferative agent in vivo and in vitro in the context of canine mammary carcinomas [81]. Together, this evidence indicates that glucose is the core of carbohydrates metabolism and is sharply related with tumor initiation and progression in CMT but is still poorly explored. In addition, better understanding of carbohydrates metabolism may improve the diagnosis and offer a more personalized treatment to the animal.
4. Lipids
Obesity in dogs is an epidemiological issue that has been widely investigated due to its consequence to the animal quality of life and health [82]. Overweight or obese female dogs was reported with earlier onset of CMTs and higher histologic grade compared to those with optimal body weight or lean [83]. Moreover, obese dogs were associated with more aggressive phenotype of CMT, angiogenesis and tumor-associated macrophages (TAMs) infiltration, being considered a risk factor for the incidence and progression of mammary neoplasms [84,85]. Decreased expression of adiponectin and increased number of macrophages in overweight or obese individuals were significantly correlated with poor prognosis, since were found high histological grade and lymphatic invasion [86]. The proportion of grade I tumors was higher among leptin-positive CMTs than leptin-negative, and a positive correlation was observed with progesterone receptor (PR)-positive tumors. Also, the number of tumors with positive estrogen receptor (ER) expression was higher among CMTs with ObR expression than those without it. The authors suggested that increased adiponectin expression may prevent cancer development and positively affect the prognosis of CMTs, whereas decreased adiponectin expression in obese dogs influences their aggressive behavior [86]. The same authors in another paper find also that the expression of aromatase, leptin, and insulin-like growth factor-1 receptor (IGF-1 R) was associated with the hormone receptor status. The results showed that the incidence of aromatase-positive CMCs was higher in the HR-positive group (any ER- or PR-positive and both ER- and PR-positive CMCs) than in the HR-negative group (P = .025) [83]. However, this is controversial because a case-control study found no correlation between high-fat diet or obesity with CMT occurrence in bitches [87] while others showed that canine fed a high-fat diet have higher survival rate compared to low-fat [88].
In addition to obesity inducing several metabolic alterations, the disparities in literature may be due to the fact that fatty acids acquired through diet do not affect tumor metabolism only by quantity but also due to their quality. In rapidly proliferating cells, lipids are required for membrane synthesis, energy supply, cell signaling, and post-translational modifications. The carnitine shuttle system is a key player in lipid catabolism since it allows the translocation of fatty acids in the cytoplasm into the mitochondria to further oxidation and ATP production. Carnitine Acylcarnitine Translocase (CACT), Carnitine Palmitoyl transferase 2 (CPT2), and Carnitine O-acetyltransferase (CrAT) were found overexpressed in CMT compared to normal condition, but a decrease was observed in poorly undifferentiated stage [89]. In the same manner, Carnitine Palmitoyl Transferase 1 A (CPT1A), a rate-limiting enzyme of FAO located in the outer mitochondrial membrane, has been reported as overexpressed in differentiated canine mammary tumors compared to normal tissue both in vivo and in vitro, whereas a decrease in CPT1A expression was observed in less differentiated tumors [90]. These findings suggest that lipid metabolism is rewired along carcinogenesis, given that cells seem to rely on energy derived from lipids, but they shift probably to glycolytic phenotype. Not so far, more aggressive tumors rely less on fatty acid oxidation, such as the hormone receptor-negative cells, MDA-MB-231 and PC-3 cells, compared to the those hormone receptor-positive, MCF-7 and C4-2B cells [91,92].
In different experimental models as well as in clinical data, saturated fatty acids (SFAs) were shown to drive carcinogenesis due to the MYC program in several cancers [93] which is also observed in human breast cancer [94,95]. Interestingly, in spheroids from canine mammary adenocarcinoma, the levels of palmitoleate, palmitate and dihomo-gamma-linolenic acid are higher compared to adherent cells, suggesting that certain SFAs are required to tumor formation [96]. However, information on unsaturated fatty acids is scarce. It was reported that oleic acid (C18:1), a monounsaturated fatty acid (MUFA), had increased subcutaneous concentrations in dogs bearing mammary tumors [97]. The polyunsaturated fatty acids (PUFAs) have been investigated due to its antitumoral property in different cancers [33,34,98,99,100,101]. Conjugated linoleic acid (CLA), one of the most abundant PUFA available in the diet, decreased the growth of epithelial and stromal CMT cells through COX-2 and prostaglandin E2 (PGE2) suppression [102], both presented with high levels in CMT cell lines [103]. However, the CLA antiproliferative effect seems to depend on its configuration, given that the trans-10, cis-12 increased the expression of cell cycle progression-related genes and the cis-9, trans-11 stimulated apoptotic genes [104]. In human breast cancer, PUFAs from the omega-3 class have been shown to increase patient survival and serve as a preventive agent [105], although this is still inconclusive [106]. In companion animals, fish oil supplementation, the main source of omega-3 PUFAs, has been recommended in several conditions, such as renal disease, cardiac and skin inflammatory disorders, dyslipidemia, and cancer [107]. In in vitro studies, the omega-3 docosahexaenoic acid (DHA) was able to induce cell death in MDA-MB-231 cells [108], but also cell cycle arrest in other cancers such as prostate [34]. In dogs, a clinical trial using long-chain omega-3 fatty acid supplementation through fish oil (eicosapentaenoic acid, EPA, 29 g/kg of diet and DHA 24 g/kg of diet), showed that increase in DHA content was associated with longer disease-free intervals and survival in the dogs with stage III lymphoma [109]. In TRAMP mice, a DHA-enriched diet was able to delay prostate cancer progression [33]. Therefore, this evidence suggests the protective effect of omega-3, but its role in CMT remains still elusive. Tuzlu and colleagues (2021) [110] reported that PUFAs belonging to the omega-3 class were higher in healthy dogs compared to those bearing CMT while an opposite correlation was observed for omega-6 fatty acids, suggesting a protective property of the former. The fish oil supplementation in female dogs after mastectomy showed that only glucose, total protein, and globulin concentrations in the serum increased after treatment (270 mg of EPA, and 180 mg of DHA, one capsule for each 10 kg of body weight per day) for 30 days [84] but the authors did not evaluate the recurrence status.
Despite being rather studied in humans, lipid metabolism is poorly investigated in canine tumors. Several enzymes from DNL are overexpressed in cancer [111]. In human breast cancer, FASN is required for brain metastasis and usually overexpressed along cancer progression [29]. In prostate cancer, DNL is a hallmark of carcinogenesis increasing in advanced stages and FASN pharmacological inhibition or even knockout suppressed tumor growth [31,112]. Therefore, this indicates that not only exogenous fatty acids may affect lipid metabolism and tumor growth, but also endogenously synthesized lipids.
5. Amino acids
Amino acids and their transporters play a plethora of functions in cells, including protein synthesis, oxidative status, protein conformational regulation, excretion through urea cycle, and cell signaling, but this topic will limit their role to cell metabolism, acting directly or indirectly. Plasma free amino acids (PFAA) levels have been investigated in dogs bearing CMT with or without metastasis and compared with healthy animals [113]. Regardless of the pathological status, the amino acids glutamine, alanine, lysine, threonine, glycine, and valine were the most abundant in the plasma [113]. However, compared with healthy animals, methionine, serine, asparagine, glutamine, alanine, taurine, and citrulline decreased in CMT. In the metastatic group, methionine, lysine, histidine, aspartate, serine, asparagine, glutamate, glutamine, alanine, taurine, citrulline, and ornithine were increased compared with control group [113]. This evidence suggests that distinct amino acids may play a role in CMT initiation, progression, and during metastasis; uncovering which amino acids are more consumed over different stages of CMT and how they are metabolized. The alterations in amino acids profile were also observed in xenograft models for human breast cancer [114]. In this in vivo study [114], metabolites were clustered according to time and cancer-bearing mice showed a significant shifting profile of selected amino acids compared to healthy condition, which could be due to liver metabolism alterations. Based on the results, they concluded that different metabolic profile patterns were associated with the light cycle. During the day, four main clusters of amino acids glutamine, alanine, lysine, and glycine were observed, while in the dark phase the main distinct clusters were glutamine, alanine, lysine, glycine, leucine and valine. Additionally, a significant reduction in plasma amino acid concentrations of arginine, aspartic acid, lysine, glycine, serine, and tyrosine was observed during the dark phase. These findings suggest that alterations in amino acid profiles may be associated with the presence of the tumor. The enrichment analysis showed significant associations with three metabolic pathways involved in molecular biosynthesis and degradation [114].
In humans, TNBC is an aggressive tumor subtype and presents a poor prognosis [115,116], lacking efficient therapeutic strategies. In TNBC, cells are dependent on glutamine metabolism and studies have shown that the inhibition of glutaminase A (GAC), an isoform of GLS often overexpressed, leads to reduction of survival and cell proliferation [117]. In this line, Ryu and colleagues (2017) [118] demonstrated by immunohistochemistry and protein expression that GLS follows the same pattern found in humans, being highly expressed in high-grade tumors, which have worse prognosis. Despite there is little information on GLS, GLS2 lacks information in CMT, but could be explored since its protumorigenic property has been documented in human breast cancer [25]. In addition, transglutaminase II (TGase II) has also been reported as overexpressed in canine and feline tumors, including mammary, and this was linked to anti-apoptotic properties [119]. In this study, TGase II expression correlated with the most aggressive phenotype and metastasis. Therefore, this evidence shows that, from the metabolic standpoint, the amino acid profile is altered in CMT and that glutamine metabolism seems to follow the same response in dogs and humans. Although further studies are required to elucidate this issue, this is an important finding since glutaminase inhibitors such as CB-839 have been proposed as therapeutic opportunities [117].
In addition to metabolism itself, the expression of amino acid transporters was reported as altered in CMT. The L-type amino acid transporter 1 (LAT1) is related to the uptake of branched or aromatic amino acids, such as leucine, isoleucine, valine, phenylalanine, tyrosine, tryptophan, methionine and histidine, in a sodium-independent manner [120]. While upregulated in CMT, it detains low expression in normal tissue, being LAT1 inhibition, a potential therapeutic strategy under investigation [120]. In addition, citrulline, a non-essential and a non-proteinogenic amino acid involved in the urea cycle, is speculated to also play a role in CMT, since peptidylarginine deiminase 2 (PAD2), an enzyme that converts arginine into citrulline is decreased in human and CMT compared to normal tissue [121]. In this context, PAD2 was shown to be responsive to the epidermal growth factor (EGF), but not estrogen or progesterone, in the canine mammary primary carcinoma cell line CMT25 [122].
6. Mitochondria
Mitochondria is known as the powerhouse of the cell due to its property of ATP production that supplies the bioenergetic needs. According to the revised Warburg effect, its activity is mostly channeled to the biosynthetic pathways and energy production would not be its main role. In CMT, the most frequent mitochondrial alteration reported in literature is on its DNA (mtDNA) which suggests the organelle plasticity to support carcinogenesis. It has a 16727 pb as a reference sequence, encoding 2 ribosomal RNA, 22 transfer RNA, and 13 polypeptides related to the electron transport chain (ETC) [123,124]. The high heterogeneity on mtDNA is observed in all CMT subtypes [125] and might be explained due to lack of histones and repair mechanisms in addition to mitochondria being the main source of ROS. Oxidative stress is related to carcinogenesis and a number of different markers have been identified in CMT, such as increase in lipid oxidation and altered activity of antioxidant enzymes [126,127,128]. Studies have demonstrated that defects on mtDNA are potential risk factors for CMT. Surdyka and Slaska [129] identified a sequence of 26 polymorphic loci and 5 mutations, and revealed that the mitochondrial displacement loop is a hotspot for mutation in CMT, being these alterations correlated with dog size. Also, Surdyka and Slaska [130] reported that ND2 (NADH dehydrogenase subunit 2), COXII (cytochrome c oxidase subunit II), ATP6 (ATP synthase F0 subunit6) and COX3 (cytochrome c oxidase subunit III), mostly of them related with the ETC function, are mutated in CMT, being a hotspot too. Moreover, such alterations were found in both tumor and blood, being ATP6 and COXII exclusively in the last [130]. This is of particular interest because it reveals heteroplasmy, but also that specific mitochondrial mutations might be required for niche formation along metastasis. Surprisingly, such variability was found in the same dog bearing two tumors [125]. Despite this evidence allowing us to infer alterations in mitochondrial metabolism, no study analyzed mitochondrial function, especially among cancer subtypes. However, as aforementioned, CPT1A is overexpressed in CMT [90], suggesting a higher mitochondrial activity compared to normal condition, but further studies are required to address this issue.
High number of alterations in mitochondria indicates its crucial role in CMT carcinogenesis which may also be a vulnerability. In addition to metabolism, mitochondria is tightly related to cell death which has been explored in vitro as therapeutic strategy for CMT [131], including by our research group [132]. Administration of iodine (I2) combined with doxorubicin to dogs with mammary tumors was shown to improve the therapeutic outcome associated with mitochondrial membrane oxidation [133]. Moreover, induced mitochondrial dysfunction has an antitumor effect on CMT cells from metastasis [134].
7. Concluding Remarks and Perspectives
The metabolic reprogramming in CMT is summarized in Figure 2, evidencing most described alterations compared with healthy individuals. However, the study of metabolic rewiring in CMT is challenging because of the reduced number of samples and their high variability, lack of well characterized tumor subtypes and suitable in vitro and in vivo experimental models. Several alterations in CMT are also described in human breast cancer, especially for glucose, amino acids, and lipid metabolism. However, although many clinical trials have attempted to target metabolic vulnerabilities in humans, most of them were not tested on dogs or have very little information reported. Tumor metabolism has gained attention in the last decade since it may be the “Achilles heel” of neoplastic cells and become more complex because of its interaction with the microenvironment. It has been reported that metabolites secreted by tumor cells may regulate immune cell populations such as macrophages. Macrophage polarization, mainly the M2 type, is determinant on inhibition of inflammation, which may favor the immune escape and cancer progression, including metastasis. However, studies concerning the association of these two variables in CMT are scarce, which could improve the outcome of current therapies. Taken together, this body of evidence highlights new venues for mammary cancer therapy in dogs and brings a plethora of vulnerabilities that should be explored metabolically to increase animal disease-free interval and overall survival.
Author Contributions
All authors contributed to manuscript conception. Guilherme Henrique Tamarindo conceptualized and wrote the manuscript with contributions by Adriana Alonso Novais and Debora Aparecida Pires de Campos Zuccari. Luiz Gustavo de Almeida Chuffa drew the figures and revised the manuscript. All authors read and approved the final version.
Funding
The authors are grateful to São Paulo Research Foundation (FAPESP) and the National Council for Scientific and Technological Development (CNPq). Authors received financial support as follows: GHT (FAPESP grant number 2022/00890-5), DAPCZ (FAPESP grant number 2020/12970-8; CNPq Process number 307288/2022-6d) and LGAC (CNPq Process number 304108/2020-0).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gray, M.; Meehan, J.; Martínez-Pérez, C.; Kay, C.; Turnbull, A.K.; Morrison, L.R.; Pang, L.Y.; Argyle, D. Naturally-Occurring Canine Mammary Tumors as a Translational Model for Human Breast Cancer. Front. Oncol. 2020, 10, 617. [Google Scholar] [CrossRef] [PubMed]
- Burrai, G.P.; Gabrieli, A.; Moccia, V.; Zappulli, V.; Porcellato, I.; Brachelente, C.; Pirino, S.; Polinas, M.; Antuofermo, E. A statistical analysis of risk factors and biological behavior in canine mammary tumors: A multicenter study. Animals (Basel) 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Cassali, G.; Jark, P.; Gamba, C.; Damasceno, K.; Estrela-Lima, A.; Nardi, A.; Ferreira, E.; Horta, R.; Firmo, B.; Sueiro, F.; Rodrigues, L.; Nakagaki, K. Consensus Regarding the Diagnosis, Prognosis and Treatment of Canine and Feline Mammary Tumors - 2019. Braz. J. Vet. Pathol. 2020, 13, 555–574. [Google Scholar] [CrossRef]
- Rasotto, R.; Berlato, D.; Goldschmidt, M.H.; Zappulli, V. Prognostic significance of canine mammary tumor histologic subtypes: an observational cohort study of 229 cases. Vet. Pathol. 2017, 54, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Peña, L.; Gama, A.; Goldschmidt, M.H.; Abadie, J.; Benazzi, C.; Castagnaro, M.; Díez, L.; Gärtner, F.; Hellmén, E.; Kiupel, M.; Millán, Y.; Miller, M.A.; Nguyen, F.; Poli, A.; Sarli, G.; Zappulli, V.; de las Mulas, J.M. Canine mammary tumors: a review and consensus of standard guidelines on epithelial and myoepithelial phenotype markers, HER2, and hormone receptor assessment using immunohistochemistry. Vet. Pathol. 2014, 51, 127–145. [Google Scholar] [CrossRef] [PubMed]
- Perez Alenza, M.D.; Peña, L.; del Castillo, N.; Nieto, A.I. Factors influencing the incidence and prognosis of canine mammary tumours. J. Small Anim. Pract. 2000, 41, 287–291. [Google Scholar] [CrossRef]
- Nicolini, A.; Carpi, A.; Rossi, G. Cytokines in breast cancer. Cytokine Growth Factor Rev. 2006, 17, 325–337. [Google Scholar] [CrossRef]
- Machado, V.S.; Crivellenti, L.Z.; Bottari, N.B.; Tonin, A.A.; Pelinson, L.P.; Borin-Crivellenti, S.; Santana, A.E.; Torbitz, V.D.; Moresco, R.N.; Duarte, T.; Duarte, M.M.M.F.; Schetinger, M.R.C.; Morsch, V.M.; Jaques, J.A.; Tinucci-Costa, M.; Da Silva, A.S. Oxidative stress and inflammatory response biomarkers in dogs with mammary carcinoma. Pathol. Res. Pract. 2015, 211, 677–681. [Google Scholar] [CrossRef]
- Benavente, M.A.; Bianchi, C.P.; Aba, M.A. Canine mammary tumors: risk factors, prognosis and treatments. J. Vet. Adv., 2016, 6, 1291–1300. [Google Scholar] [CrossRef]
- Sorenmo, K. Canine mammary gland tumors. Vet. Clin. North Am. Small Anim. Pract. 2003, 33, 573–596. [Google Scholar] [CrossRef]
- Salas, Y.; Márquez, A.; Diaz, D.; Romero, L. Epidemiological Study of Mammary Tumors in Female Dogs Diagnosed during the Period 2002-2012: A Growing Animal Health Problem. PLoS ONE 2015, 10, e0127381. [Google Scholar] [CrossRef] [PubMed]
- Kaszak, I.; Ruszczak, A.; Kanafa, S.; Kacprzak, K.; Król, M.; Jurka, P. Current biomarkers of canine mammary tumors. Acta Vet. Scand. 2018, 60, 66. [Google Scholar] [CrossRef] [PubMed]
- Dobson, J.M.; Samuel, S.; Milstein, H.; Rogers, K.; Wood, J.L.N. Canine neoplasia in the UK: estimates of incidence rates from a population of insured dogs. J. Small Anim. Pract. 2002, 43, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Moe, L. Population-based incidence of mammary tumours in some dog breeds. J. Reprod. Fertil. Suppl. 2001, 57, 439–443. [Google Scholar] [PubMed]
- Abdelmegeed, S.M.; Mohammed, S. Canine mammary tumors as a model for human disease. Oncol. Lett. 2018, 15, 8195–8205. [Google Scholar] [CrossRef]
- Queiroga, F.L.; Raposo, T.; Carvalho, M.I.; Prada, J.; Pires, I. Canine mammary tumours as a model to study human breast cancer: most recent findings. In Vivo 2011, 25, 455–465. [Google Scholar]
- Schiliro, C.; Firestein, B.L. Mechanisms of metabolic reprogramming in cancer cells supporting enhanced growth and proliferation. Cells 2021, 10. [Google Scholar] [CrossRef]
- Sun, L.; Suo, C.; Li, S.-T.; Zhang, H.; Gao, P. Metabolic reprogramming for cancer cells and their microenvironment: Beyond the Warburg Effect. Biochim. Biophys. Acta Rev. Cancer 2018, 1870, 51–66. [Google Scholar] [CrossRef]
- Ohshima, K.; Morii, E. Metabolic Reprogramming of Cancer Cells during Tumor Progression and Metastasis. Metabolites 2021, 11. [Google Scholar] [CrossRef]
- Warburg, O. On the origin of cancer cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef]
- DeBerardinis, R.J.; Chandel, N.S. We need to talk about the Warburg effect. Nat. Metab. 2020, 2, 127–129. [Google Scholar] [CrossRef]
- Devic, S. Warburg Effect - a Consequence or the Cause of Carcinogenesis? J. Cancer 2016, 7, 817–822. [Google Scholar] [CrossRef] [PubMed]
- Altman, B.J.; Stine, Z.E.; Dang, C.V. From Krebs to clinic: glutamine metabolism to cancer therapy. Nat. Rev. Cancer 2016, 16, 619–634. [Google Scholar] [CrossRef]
- Zacharias, N.M.; McCullough, C.; Shanmugavelandy, S.; Lee, J.; Lee, Y.; Dutta, P.; McHenry, J.; Nguyen, L.; Norton, W.; Jones, L.W.; Bhattacharya, P.K. Metabolic differences in glutamine utilization lead to metabolic vulnerabilities in prostate cancer. Sci. Rep. 2017, 7, 16159. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.M.; Adamoski, D.; Dos Reis, L.M.; Ascenção, C.F.R.; de Oliveira, K.R.S.; Mafra, A.C.P.; da Silva Bastos, A.C.; Quintero, M.; de G Cassago, C.; Ferreira, I.M.; Fidelis, C.H.V.; Rocco, S.A.; Bajgelman, M.C.; Stine, Z.; Berindan-Neagoe, I.; Calin, G.A.; Ambrosio, A.L.B.; Dias, S.M.G. GLS2 is protumorigenic in breast cancers. Oncogene 2020, 39, 690–702. [Google Scholar] [CrossRef]
- Deep, G.; Schlaepfer, I.R. Aberrant lipid metabolism promotes prostate cancer: role in cell survival under hypoxia and extracellular vesicles biogenesis. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Mullen, A.R.; Wheaton, W.W.; Jin, E.S.; Chen, P.-H.; Sullivan, L.B.; Cheng, T.; Yang, Y.; Linehan, W.M.; Chandel, N.S.; DeBerardinis, R.J. Reductive carboxylation supports growth in tumour cells with defective mitochondria. Nature 2011, 481, 385–388. [Google Scholar] [CrossRef]
- Scaglia, N.; Tyekucheva, S.; Zadra, G.; Photopoulos, C.; Loda, M. De novo fatty acid synthesis at the mitotic exit is required to complete cellular division. Cell Cycle 2014, 13, 859–868. [Google Scholar] [CrossRef]
- Ferraro, G.B.; Ali, A.; Luengo, A.; Kodack, D.P.; Deik, A.; Abbott, K.L.; Bezwada, D.; Blanc, L.; Prideaux, B.; Jin, X.; Posada, J.M.; Chen, J.; Chin, C.R.; Amoozgar, Z.; Ferreira, R.; Chen, I.X.; Naxerova, K.; Ng, C.; Westermark, A.M.; Duquette, M.; Roberge, S.; Lindeman, N.I.; Lyssiotis, C.A.; Nielsen, J.; Housman, D.E.; Duda, D.G.; Brachtel, E.; Golub, T.R.; Cantley, L.C.; Asara, J.M.; Davidson, S.M.; Fukumura, D.; Dartois, V.A.; Clish, C.B.; Jain, R.K.; Vander Heiden, M.G. FATTY ACID SYNTHESIS IS REQUIRED FOR BREAST CANCER BRAIN METASTASIS. Nat. Cancer 2021, 2, 414–428. [Google Scholar] [CrossRef] [PubMed]
- Migita, T.; Ruiz, S.; Fornari, A.; Fiorentino, M.; Priolo, C.; Zadra, G.; Inazuka, F.; Grisanzio, C.; Palescandolo, E.; Shin, E.; Fiore, C.; Xie, W.; Kung, A.L.; Febbo, P.G.; Subramanian, A.; Mucci, L.; Ma, J.; Signoretti, S.; Stampfer, M.; Hahn, W.C.; Finn, S.; Loda, M. Fatty acid synthase: a metabolic enzyme and candidate oncogene in prostate cancer. J Natl Cancer Inst 2009, 101, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Zadra, G.; Ribeiro, C.F.; Chetta, P.; Ho, Y.; Cacciatore, S.; Gao, X.; Syamala, S.; Bango, C.; Photopoulos, C.; Huang, Y.; Tyekucheva, S.; Bastos, D.C.; Tchaicha, J.; Lawney, B.; Uo, T.; D’Anello, L.; Csibi, A.; Kalekar, R.; Larimer, B.; Ellis, L.; Butler, L.M.; Morrissey, C.; McGovern, K.; Palombella, V.J.; Kutok, J.L.; Mahmood, U.; Bosari, S.; Adams, J.; Peluso, S.; Dehm, S.M.; Plymate, S.R.; Loda, M. Inhibition of de novo lipogenesis targets androgen receptor signaling in castration-resistant prostate cancer. Proc Natl Acad Sci USA 2019, 116, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y. Fatty acid oxidation is a dominant bioenergetic pathway in prostate cancer. Prostate Cancer Prostatic Dis. 2006, 9, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Amaro, G.M.; da Silva, A.D.T.; Tamarindo, G.H.; Lamas, C. de A.; Taboga, S.R.; Cagnon, V.H.A.; Góes, R.M. Differential effects of omega-3 PUFAS on tumor progression at early and advanced stages in TRAMP mice. Prostate 2022. [CrossRef]
- Tamarindo, G.H.; Góes, R.M. Docosahexaenoic acid differentially modulates the cell cycle and metabolism- related genes in tumor and pre-malignant prostate cells. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158766. [Google Scholar] [CrossRef]
- Shin, E.; Koo, J.S. Glucose metabolism and glucose transporters in breast cancer. Front. Cell Dev. Biol. 2021, 9, 728759. [Google Scholar] [CrossRef]
- Fong, M.Y.; Zhou, W.; Liu, L.; Alontaga, A.Y.; Chandra, M.; Ashby, J.; Chow, A.; O’Connor, S.T.F.; Li, S.; Chin, A.R.; Somlo, G.; Palomares, M.; Li, Z.; Tremblay, J.R.; Tsuyada, A.; Sun, G.; Reid, M.A.; Wu, X.; Swiderski, P.; Ren, X.; Shi, Y.; Kong, M.; Zhong, W.; Chen, Y.; Wang, S.E. Breast-cancer-secreted miR-122 reprograms glucose metabolism in premetastatic niche to promote metastasis. Nat. Cell Biol. 2015, 17, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Farhadi, P.; Yarani, R.; Valipour, E.; Kiani, S.; Hoseinkhani, Z.; Mansouri, K. Cell line-directed breast cancer research based on glucose metabolism status. Biomed. Pharmacother. 2022, 146, 112526. [Google Scholar] [CrossRef]
- Medina, R.A.; Owen, G.I. Glucose transporters: expression, regulation and cancer. Biol. Res. 2002, 35, 9–26. [Google Scholar] [CrossRef]
- Guan, M.; Tong, Y.; Guan, M.; Liu, X.; Wang, M.; Niu, R.; Zhang, F.; Dong, D.; Shao, J.; Zhou, Y. Lapatinib inhibits breast cancer cell proliferation by influencing PKM2 expression. Technol. Cancer Res. Treat. 2018, 17, 1533034617749418. [Google Scholar] [CrossRef]
- O’Neal, J.; Clem, A.; Reynolds, L.; Dougherty, S.; Imbert-Fernandez, Y.; Telang, S.; Chesney, J.; Clem, B.F. Inhibition of 6-phosphofructo-2-kinase (PFKFB3) suppresses glucose metabolism and the growth of HER2+ breast cancer. Breast Cancer Res. Treat. 2016, 160, 29–40. [Google Scholar] [CrossRef]
- Peng, F.; Li, Q.; Sun, J.-Y.; Luo, Y.; Chen, M.; Bao, Y. PFKFB3 is involved in breast cancer proliferation, migration, invasion and angiogenesis. Int. J. Oncol. 2018, 52, 945–954. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Ren, C.; Qiao, P.; Han, X.; Wang, L.; Lv, S.; Sun, Y.; Liu, Z.; Du, Y.; Yu, Z. PIM2-mediated phosphorylation of hexokinase 2 is critical for tumor growth and paclitaxel resistance in breast cancer. Oncogene 2018, 37, 5997–6009. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.S.; Goodman, T.M.; Zasadny, K.R.; Greenson, J.K.; Wahl, R.L. Expression of hexokinase II and Glut-1 in untreated human breast cancer. Nucl. Med. Biol. 2002, 29, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Tao, L.; Wei, L.; Liu, Y.; Ding, Y.; Liu, X.; Zhang, X.; Wang, X.; Yao, Y.; Lu, J.; Wang, Q.; Hu, R. Gen-27, a newly synthesized flavonoid, inhibits glycolysis and induces cell apoptosis via suppression of hexokinase II in human breast cancer cells. Biochem. Pharmacol. 2017, 125, 12–25. [Google Scholar] [CrossRef]
- Doyen, J.; Trastour, C.; Ettore, F.; Peyrottes, I.; Toussant, N.; Gal, J.; Ilc, K.; Roux, D.; Parks, S.K.; Ferrero, J.M.; Pouysségur, J. Expression of the hypoxia-inducible monocarboxylate transporter MCT4 is increased in triple negative breast cancer and correlates independently with clinical outcome. Biochem. Biophys. Res. Commun. 2014, 451, 54–61. [Google Scholar] [CrossRef] [PubMed]
- McCleland, M.L.; Adler, A.S.; Shang, Y.; Hunsaker, T.; Truong, T.; Peterson, D.; Torres, E.; Li, L.; Haley, B.; Stephan, J.-P.; Belvin, M.; Hatzivassiliou, G.; Blackwood, E.M.; Corson, L.; Evangelista, M.; Zha, J.; Firestein, R. An integrated genomic screen identifies LDHB as an essential gene for triple-negative breast cancer. Cancer Res. 2012, 72, 5812–5823. [Google Scholar] [CrossRef]
- Morais-Santos, F.; Granja, S.; Miranda-Gonçalves, V.; Moreira, A.H.J.; Queirós, S.; Vilaça, J.L.; Schmitt, F.C.; Longatto-Filho, A.; Paredes, J.; Baltazar, F.; Pinheiro, C. Targeting lactate transport suppresses in vivo breast tumour growth. Oncotarget 2015, 6, 19177–19189. [Google Scholar] [CrossRef]
- Nagata, C.; Wada, K.; Tsuji, M.; Hayashi, M.; Takeda, N.; Yasuda, K. Plasma amino acid profiles are associated with biomarkers of breast cancer risk in premenopausal Japanese women. Cancer Causes Control 2014, 25, 143–149. [Google Scholar] [CrossRef]
- Park, J.; Shin, Y.; Kim, T.H.; Kim, D.-H.; Lee, A. Plasma metabolites as possible biomarkers for diagnosis of breast cancer. PLoS ONE 2019, 14, e0225129. [Google Scholar] [CrossRef]
- Barnes, T.; Bell, K.; DiSebastiano, K.M.; Vance, V.; Hanning, R.; Russell, C.; Dubin, J.A.; Bahl, M.; Califaretti, N.; Campbell, C.; Mourtzakis, M. Plasma amino acid profiles of breast cancer patients early in the trajectory of the disease differ from healthy comparison groups. Appl. Physiol. Nutr. Metab. 2014, 39, 740–744. [Google Scholar] [CrossRef]
- Masisi, B.K.; El Ansari, R.; Alfarsi, L.; Rakha, E.A.; Green, A.R.; Craze, M.L. The role of glutaminase in cancer. Histopathology 2020, 76, 498–508. [Google Scholar] [CrossRef]
- Wang, J.-B.; Erickson, J.W.; Fuji, R.; Ramachandran, S.; Gao, P.; Dinavahi, R.; Wilson, K.F.; Ambrosio, A.L.B.; Dias, S.M.G.; Dang, C.V.; Cerione, R.A. Targeting mitochondrial glutaminase activity inhibits oncogenic transformation. Cancer Cell 2010, 18, 207–219. [Google Scholar] [CrossRef]
- Cassago, A.; Ferreira, A.P.S.; Ferreira, I.M.; Fornezari, C.; Gomes, E.R.M.; Greene, K.S.; Pereira, H.M.; Garratt, R.C.; Dias, S.M.G.; Ambrosio, A.L.B. Mitochondrial localization and structure-based phosphate activation mechanism of Glutaminase C with implications for cancer metabolism. Proc Natl Acad Sci USA 2012, 109, 1092–1097. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Kim, S.-H.; Im, C.Y.; Hwang, H.-J. Recent development of small molecule glutaminase inhibitors. Curr. Top. Med. Chem. 2018, 18, 432–443. [Google Scholar] [CrossRef] [PubMed]
- Best, S.A.; Gubser, P.M.; Sethumadhavan, S.; Kersbergen, A.; Negrón Abril, Y.L.; Goldford, J.; Sellers, K.; Abeysekera, W.; Garnham, A.L.; McDonald, J.A.; Weeden, C.E.; Anderson, D.; Pirman, D.; Roddy, T.P.; Creek, D.J.; Kallies, A.; Kingsbury, G.; Sutherland, K.D. Glutaminase inhibition impairs CD8 T cell activation in STK11-/Lkb1-deficient lung cancer. Cell Metab. 2022, 34, 874–887. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Li, X.; Gong, G.; Lu, Y.; Guo, Z.; Chen, R.; Huang, H.; Li, Z.; Bian, J. An updated patent review of glutaminase inhibitors (2019-2022). Expert Opin. Ther. Pat. 2023, 33, 17–28. [Google Scholar] [CrossRef]
- Wang, Z.; Jiang, Q.; Dong, C. Metabolic reprogramming in triple-negative breast cancer. Cancer Biol. Med. 2020, 17, 44–59. [Google Scholar] [CrossRef]
- Wang, W.; Bai, L.; Li, W.; Cui, J. The lipid metabolic landscape of cancers and new therapeutic perspectives. Front. Oncol. 2020, 10, 605154. [Google Scholar] [CrossRef]
- Giudetti, A.M.; De Domenico, S.; Ragusa, A.; Lunetti, P.; Gaballo, A.; Franck, J.; Simeone, P.; Nicolardi, G.; De Nuccio, F.; Santino, A.; Capobianco, L.; Lanuti, P.; Fournier, I.; Salzet, M.; Maffia, M.; Vergara, D. A specific lipid metabolic profile is associated with the epithelial mesenchymal transition program. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 344–357. [Google Scholar] [CrossRef]
- Simeone, P.; Tacconi, S.; Longo, S.; Lanuti, P.; Bravaccini, S.; Pirini, F.; Ravaioli, S.; Dini, L.; Giudetti, A.M. Expanding roles of de novo lipogenesis in breast cancer. Int. J. Environ. Res. Public Health 2021, 18. [Google Scholar] [CrossRef]
- German, N.J.; Yoon, H.; Yusuf, R.Z.; Murphy, J.P.; Finley, L.W.S.; Laurent, G.; Haas, W.; Satterstrom, F.K.; Guarnerio, J.; Zaganjor, E.; Santos, D.; Pandolfi, P.P.; Beck, A.H.; Gygi, S.P.; Scadden, D.T.; Kaelin, W.G.; Haigis, M.C. PHD3 loss in cancer enables metabolic reliance on fatty acid oxidation via deactivation of ACC2. Mol. Cell 2016, 63, 1006–1020. [Google Scholar] [CrossRef] [PubMed]
- Giró-Perafita, A.; Palomeras, S.; Lum, D.H.; Blancafort, A.; Viñas, G.; Oliveras, G.; Pérez-Bueno, F.; Sarrats, A.; Welm, A.L.; Puig, T. Preclinical Evaluation of Fatty Acid Synthase and EGFR Inhibition in Triple-Negative Breast Cancer. Clin. Cancer Res. 2016, 22, 4687–4697. [Google Scholar] [CrossRef] [PubMed]
- Wahdan-Alaswad, R.S.; Cochrane, D.R.; Spoelstra, N.S.; Howe, E.N.; Edgerton, S.M.; Anderson, S.M.; Thor, A.D.; Richer, J.K. Metformin-induced killing of triple-negative breast cancer cells is mediated by reduction in fatty acid synthase via miRNA-193b. Horm. Cancer 2014, 5, 374–389. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Wu, J.; Zhao, Q.; Fu, S.; Jin, J. Emerging roles of aerobic glycolysis in breast cancer. Clin. Transl. Oncol. 2020, 22, 631–646. [Google Scholar] [CrossRef]
- Ogilvie, G.K.; Walters, L.; Salman, M.D.; Fettman, M.J.; Johnston, S.D.; Hegstad, R.L. Alterations in carbohydrate metabolism in dogs with nonhematopoietic malignancies. Am. J. Vet. Res. 1997, 58, 277–281. [Google Scholar]
- Rodigheri, S.M.; Paiva, F.N. de; Firmo, B.F.; Fuchs, T.; Mani, C.B.; Nardi, A.B. de Parameters of metabolic response to surgical trauma induced via unilateral total mastectomy associated or not to ovariohysterectomy in dogs. Animals (Basel) 2023, 13. [Google Scholar] [CrossRef]
- Jayasri, K.; Padmaja, K.; Saibaba, M. Altered oxidative stress and carbohydrate metabolism in canine mammary tumors. Vet. World 2016, 9, 1489–1492. [Google Scholar] [CrossRef]
- Lee, H.-J.; Han, H.-J.; Lee, J.-Y.; Son, W.-C. PKM2 in canine mammary tumors: parallels to human breast cancer. Comp. Med. 2020, 70, 349–354. [Google Scholar] [CrossRef]
- Arai, T.; Ogino, T.; Gunji, M.; Washizu, T.; Komori, S.; Washizu, M. Changes in glucose transport activities in mammary adenocarcinoma of dogs. Res. Vet. Sci. 1997, 62, 85–86. [Google Scholar] [CrossRef]
- Freeman, A.; Hetzel, U.; Cripps, P.; Mobasheri, A. Expression of the plasma membrane markers aquaporin 1 (AQP1), glucose transporter 1 (GLUT1) and Na, K-ATPase in canine mammary glands and mammary tumours. Vet. J. 2010, 185, 90–93. [Google Scholar] [CrossRef]
- Mees, G.; Vangestel, C.; Dierckx, R.; Loomans, S.; Van Damme, N.; Peremans, K.; De Rooster, H.; Van Goethem, B.; Pauwels, P.; Ducatelle, R.; Van de Wiele, C. Metabolic correlates of tumour hypoxia in malignant canine mammary carcinoma. Res. Vet. Sci. 2011, 91, e125–8. [Google Scholar] [CrossRef]
- Beirão, B.C.B.; Raposo, T.; Pang, L.Y.; Argyle, D.J. Canine mammary cancer cells direct macrophages toward an intermediate activation state between M1/M2. BMC Vet. Res. 2015, 11, 151. [Google Scholar] [CrossRef]
- Bao, X.; Shi, R.; Zhao, T.; Wang, Y.; Anastasov, N.; Rosemann, M.; Fang, W. Integrated analysis of single-cell RNA-seq and bulk RNA-seq unravels tumour heterogeneity plus M2-like tumour-associated macrophage infiltration and aggressiveness in TNBC. Cancer Immunol. Immunother. 2021, 70, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Mu, X.; Shi, W.; Xu, Y.; Xu, C.; Zhao, T.; Geng, B.; Yang, J.; Pan, J.; Hu, S.; Zhang, C.; Zhang, J.; Wang, C.; Shen, J.; Che, Y.; Liu, Z.; Lv, Y.; Wen, H.; You, Q. Tumor-derived lactate induces M2 macrophage polarization via the activation of the ERK/STAT3 signaling pathway in breast cancer. Cell Cycle 2018, 17, 428–438. [Google Scholar] [CrossRef] [PubMed]
- Burrai, G.P.; Tanca, A.; Cubeddu, T.; Abbondio, M.; Polinas, M.; Addis, M.F.; Antuofermo, E. A first immunohistochemistry study of transketolase and transketolase-like 1 expression in canine hyperplastic and neoplastic mammary lesions. BMC Vet. Res. 2017, 13, 38. [Google Scholar] [CrossRef]
- Nerurkar, V.R.; Ishwad, C.S.; Seshadri, R.; Naik, S.N.; Lalitha, V.S. Glucose-6-phosphate dehydrogenase and 6-phosphogluconate dehydrogenase activities in normal canine mammary gland and in mammary tumours and their correlation with oestrogen receptors. J. Comp. Pathol. 1990, 102, 191–195. [Google Scholar] [CrossRef]
- Sánchez, D.; Romero, L.; López, S.; Campuzano, M.; Ortega, R.; Morales, A.; Guadarrama, M.; Cesarman-Maus, G.; García-Pérez, O.; Lizano, M. 18F-FDG-PET/CT in Canine Mammary Gland Tumors. Front. Vet. Sci. 2019, 6, 280. [Google Scholar] [CrossRef] [PubMed]
- Seiler, S.M.F.; Baumgartner, C.; Hirschberger, J.; Beer, A.J.; Brühschwein, A.; Kreutzmann, N.; Laberke, S.; Wergin, M.C.; Meyer-Lindenberg, A.; Brandl, J.; von Thaden, A.-K.; Farrell, E.; Schwaiger, M. Comparative Oncology: Evaluation of 2-Deoxy-2-[18F]fluoro-D-glucose (FDG) Positron Emission Tomography/Computed Tomography (PET/CT) for the Staging of Dogs with Malignant Tumors. PLoS ONE 2015, 10, e0127800. [Google Scholar] [CrossRef]
- Borgatti, A.; Winter, A.L.; Stuebner, K.; Scott, R.; Ober, C.P.; Anderson, K.L.; Feeney, D.A.; Vallera, D.A.; Koopmeiners, J.S.; Modiano, J.F.; Froelich, J. Evaluation of 18-F-fluoro-2-deoxyglucose (FDG) positron emission tomography/computed tomography (PET/CT) as a staging and monitoring tool for dogs with stage-2 splenic hemangiosarcoma - A pilot study. PLoS ONE 2017, 12, e0172651. [Google Scholar] [CrossRef] [PubMed]
- Koo, Y.; Yun, T.; Chae, Y.; Lee, D.; Son, M.; Ku, D.; Kim, H.; Yang, M.; Kang, B. Evaluation of a dog with inflammatory mammary carcinoma using18 F-2-deoxy-2-fluoro-d -glucose positron emission tomography/computed tomography. Vet. Med. Sci. 2022, 8, 1361–1365. [Google Scholar] [CrossRef]
- Barbieri, F.; Thellung, S.; Ratto, A.; Carra, E.; Marini, V.; Fucile, C.; Bajetto, A.; Pattarozzi, A.; Würth, R.; Gatti, M.; Campanella, C.; Vito, G.; Mattioli, F.; Pagano, A.; Daga, A.; Ferrari, A.; Florio, T. In vitro and in vivo antiproliferative activity of metformin on stem-like cells isolated from spontaneous canine mammary carcinomas: translational implications for human tumors. BMC Cancer 2015, 15, 228. [Google Scholar] [CrossRef] [PubMed]
- Cortese, L.; Terrazzano, G.; Pelagalli, A. Leptin and immunological profile in obesity and its associated diseases in dogs. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.Y.; Im, K.S.; Kim, N.H.; Kim, H.W.; Shin, J.I.; Yhee, J.Y.; Sur, J.H. Effects of Obesity and Obesity-Related Molecules on Canine Mammary Gland Tumors. Vet. Pathol. 2015, 52, 1045–1051. [Google Scholar] [CrossRef] [PubMed]
- Costa-Santos, K.; Damasceno, K.; Portela, R.D.; Santos, F.L.; Araújo, G.C.; Martins-Filho, E.F.; Silva, L.P.; Barral, T.D.; Santos, S.A.; Estrela-Lima, A. Lipid and metabolic profiles in female dogs with mammary carcinoma receiving dietary fish oil supplementation. BMC Vet. Res. 2019, 15, 401. [Google Scholar] [CrossRef]
- Lim, H.-Y.; Seung, B.-J.; Cho, S.-H.; Kim, S.-H.; Bae, M.-K.; Sur, J.-H. Canine mammary cancer in overweight or obese female dogs is associated with intratumoral microvessel density and macrophage counts. Vet. Pathol. 2022, 59, 39–45. [Google Scholar] [CrossRef]
- Lim, H.Y.; Im, K.S.; Kim, N.H.; Kim, H.W.; Shin, J.I.; Sur, J.H. Obesity, expression of adipocytokines, and macrophage infiltration in canine mammary tumors. Vet. J. 2015, 203, 326–331. [Google Scholar] [CrossRef]
- Sonnenschein, E.G.; Glickman, L.T.; Goldschmidt, M.H.; McKee, L.J. Body conformation, diet, and risk of breast cancer in pet dogs: a case-control study. Am. J. Epidemiol. 1991, 133, 694–703. [Google Scholar] [CrossRef]
- Shofer, F.S.; Sonnenschein, E.G.; Goldschmidt, M.H.; Laster, L.L.; Glickman, L.T. Histopathologic and dietary prognostic factors for canine mammary carcinoma. Breast Cancer Res. Treat. 1989, 13, 49–60. [Google Scholar] [CrossRef]
- Cacciola, N.A.; Sgadari, M.; Sepe, F.; Petillo, O.; Margarucci, S.; Martano, M.; Maiolino, P.; Restucci, B. Metabolic flexibility in canine mammary tumors: implications of the carnitine system. Animals (Basel) 2021, 11. [Google Scholar] [CrossRef]
- Cacciola, N.A.; Sgadari, M.; Petillo, O.; Margarucci, S.; Martano, M.; Cocchia, N.; Maiolino, P.; Restucci, B. Carnitine palmitoyltransferase 1 A expression profile in canine mammary tumors. Vet. J. 2020, 257, 105453. [Google Scholar] [CrossRef]
- Balaban, S.; Lee, L.S.; Varney, B.; Aishah, A.; Gao, Q.; Shearer, R.F.; Saunders, D.N.; Grewal, T.; Hoy, A.J. Heterogeneity of fatty acid metabolism in breast cancer cells underlies differential sensitivity to palmitate-induced apoptosis. Mol. Oncol. 2018, 12, 1623–1638. [Google Scholar] [CrossRef] [PubMed]
- Balaban, S.; Nassar, Z.D.; Zhang, A.Y.; Hosseini-Beheshti, E.; Centenera, M.M.; Schreuder, M.; Lin, H.-M.; Aishah, A.; Varney, B.; Liu-Fu, F.; Lee, L.S.; Nagarajan, S.R.; Shearer, R.F.; Hardie, R.-A.; Raftopulos, N.L.; Kakani, M.S.; Saunders, D.N.; Holst, J.; Horvath, L.G.; Butler, L.M.; Hoy, A.J. Extracellular fatty acids are the major contributor to lipid synthesis in prostate cancer. Mol. Cancer Res. 2019, 17, 949–962. [Google Scholar] [CrossRef] [PubMed]
- Labbé, D.P.; Zadra, G.; Yang, M.; Reyes, J.M.; Lin, C.Y.; Cacciatore, S.; Ebot, E.M.; Creech, A.L.; Giunchi, F.; Fiorentino, M.; Elfandy, H.; Syamala, S.; Karoly, E.D.; Alshalalfa, M.; Erho, N.; Ross, A.; Schaeffer, E.M.; Gibb, E.A.; Takhar, M.; Den, R.B.; Lehrer, J.; Karnes, R.J.; Freedland, S.J.; Davicioni, E.; Spratt, D.E.; Ellis, L.; Jaffe, J.D.; DʼAmico, A.V.; Kantoff, P.W.; Bradner, J.E.; Mucci, L.A.; Chavarro, J.E.; Loda, M.; Brown, M. High-fat diet fuels prostate cancer progression by rewiring the metabolome and amplifying the MYC program. Nat. Commun. 2019, 10, 4358. [Google Scholar] [CrossRef] [PubMed]
- Casciano, J.C.; Perry, C.; Cohen-Nowak, A.J.; Miller, K.D.; Vande Voorde, J.; Zhang, Q.; Chalmers, S.; Sandison, M.E.; Liu, Q.; Hedley, A.; McBryan, T.; Tang, H.-Y.; Gorman, N.; Beer, T.; Speicher, D.W.; Adams, P.D.; Liu, X.; Schlegel, R.; McCarron, J.G.; Wakelam, M.J.O.; Gottlieb, E.; Kossenkov, A.V.; Schug, Z.T. MYC regulates fatty acid metabolism through a multigenic program in claudin-low triple negative breast cancer. Br. J. Cancer 2020, 122, 868–884. [Google Scholar] [CrossRef] [PubMed]
- Camarda, R.; Zhou, A.Y.; Kohnz, R.A.; Balakrishnan, S.; Mahieu, C.; Anderton, B.; Eyob, H.; Kajimura, S.; Tward, A.; Krings, G.; Nomura, D.K.; Goga, A. Inhibition of fatty acid oxidation as a therapy for MYC-overexpressing triple-negative breast cancer. Nat. Med. 2016, 22, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Michishita, M.; Saito, N.; Nozawa, S.; Furumoto, R.; Nakagawa, T.; Sato, T.; Ochiai, K.; Azakami, D.; Katayama, K.; Nakahira, R.; Tazaki, H.; Machida, Y.; Ishiwata, T. Metabolite profiling in sphere-forming cells from canine mammary adenocarcinoma cell lines using gas chromatography-mass spectrometry. J. Vet. Med. Sci. 2019, 81, 1238–1248. [Google Scholar] [CrossRef] [PubMed]
- Pérez Alenza, D.; Rutteman, G.R.; Peña, L.; Beynen, A.C.; Cuesta, P. Relation between habitual diet and canine mammary tumors in a case-control study. J. Vet. Intern. Med. 1998, 12, 132–139. [Google Scholar] [CrossRef]
- Rahman, M.M.; Veigas, J.M.; Williams, P.J.; Fernandes, G. DHA is a more potent inhibitor of breast cancer metastasis to bone and related osteolysis than EPA. Breast Cancer Res. Treat. 2013, 141, 341–352. [Google Scholar] [CrossRef]
- Bai, X.; Shao, J.; Zhou, S.; Zhao, Z.; Li, F.; Xiang, R.; Zhao, A.Z.; Pan, J. Inhibition of lung cancer growth and metastasis by DHA and its metabolite, RvD1, through miR-138-5p/FOXC1 pathway. J. Exp. Clin. Cancer Res. 2019, 38, 479. [Google Scholar] [CrossRef]
- Shekari, N.; Javadian, M.; Ghasemi, M.; Baradaran, B.; Darabi, M.; Kazemi, T. Synergistic Beneficial Effect of Docosahexaenoic Acid (DHA) and Docetaxel on the Expression Level of Matrix Metalloproteinase-2 (MMP-2) and MicroRNA-106b in Gastric Cancer. J. Gastrointest. Cancer 2020, 51, 70–75. [Google Scholar] [CrossRef]
- Abdi, J.; Garssen, J.; Faber, J.; Redegeld, F.A. Omega-3 fatty acids, EPA and DHA induce apoptosis and enhance drug sensitivity in multiple myeloma cells but not in normal peripheral mononuclear cells. J. Nutr. Biochem. 2014, 25, 1254–1262. [Google Scholar] [CrossRef]
- Wang, L.-S.; Huang, Y.-W.; Liu, S.; Chang, H.-L.; Ye, W.; Shu, S.; Sugimoto, Y.; Funk, J.A.; Smeaks, D.D.; Hill, L.N.; Lin, Y.C. Conjugated linoleic acid (CLA) modulates prostaglandin E2 (PGE2) signaling in canine mammary cells. Anticancer Res. 2006, 26, 889–898. [Google Scholar]
- Brunelle, M.; Sartin, E.A.; Wolfe, L.G.; Sirois, J.; Doré, M. Cyclooxygenase-2 expression in normal and neoplastic canine mammary cell lines. Vet. Pathol. 2006, 43, 656–666. [Google Scholar] [CrossRef]
- Da Silva, E.D.; Sandri, E.C.; Fernandes, L.A.; Carraro, P.C.; De Oliveira, D.E. Conjugated linoleic acid trans-10, cis-12 increases the expression of genes from cell cycle progression and cis-9, trans-11 stimulates apoptotic genes in different mammary tumor explants of female dogs. Mol. Biol. Rep. 2023. [Google Scholar] [CrossRef] [PubMed]
- Fabian, C.J.; Kimler, B.F.; Hursting, S.D. Omega-3 fatty acids for breast cancer prevention and survivorship. Breast Cancer Res. 2015, 17, 62. [Google Scholar] [CrossRef]
- Lee, K.H.; Seong, H.J.; Kim, G.; Jeong, G.H.; Kim, J.Y.; Park, H.; Jung, E.; Kronbichler, A.; Eisenhut, M.; Stubbs, B.; Solmi, M.; Koyanagi, A.; Hong, S.H.; Dragioti, E.; de Rezende, L.F.M.; Jacob, L.; Keum, N.; van der Vliet, H.J.; Cho, E.; Veronese, N.; Grosso, G.; Ogino, S.; Song, M.; Radua, J.; Jung, S.J.; Thompson, T.; Jackson, S.E.; Smith, L.; Yang, L.; Oh, H.; Choi, E.K.; Shin, J.I.; Giovannucci, E.L.; Gamerith, G. Consumption of Fish and ω-3 Fatty Acids and Cancer Risk: An Umbrella Review of Meta-Analyses of Observational Studies. Adv. Nutr. 2020, 11, 1134–1149. [Google Scholar] [CrossRef] [PubMed]
- Bauer, J.E. Therapeutic use of fish oils in companion animals. J. Am. Vet. Med. Assoc. 2011, 239, 1441–1451. [Google Scholar] [CrossRef] [PubMed]
- Pizato, N.; Luzete, B.C.; Kiffer, L.F.M.V.; Corrêa, L.H.; de Oliveira Santos, I.; Assumpção, J.A.F.; Ito, M.K.; Magalhães, K.G. Omega-3 docosahexaenoic acid induces pyroptosis cell death in triple-negative breast cancer cells. Sci. Rep. 2018, 8, 1952. [Google Scholar] [CrossRef]
- Ogilvie, G.K.; Fettman, M.J.; Mallinckrodt, C.H.; Walton, J.A.; Hansen, R.A.; Davenport, D.J.; Gross, K.L.; Richardson, K.L.; Rogers, Q.; Hand, M.S. Effect of fish oil, arginine, and doxorubicin chemotherapy on remission and survival time for dogs with lymphoma: a double-blind, randomized placebo-controlled study. Cancer 2000, 88, 1916–1928. [Google Scholar] [CrossRef]
- Tuzlu, T.; Saribay, M.K.; KOLDAS ÜRER, E.; KÖSE, A.M.; GÖZER, A.; YAKAN, A.; ÖZSOY, Ş.Y. Evaluation of blood omega-3 and omega-6 levels in healthy female dogs and. J HELLENIC VET MED SOC 2021, 72, 2925–2934. [Google Scholar]
- Cheng, C.; Geng, F.; Cheng, X.; Guo, D. Lipid metabolism reprogramming and its potential targets in cancer. Cancer Commun (Lond) 2018, 38, 27. [Google Scholar] [CrossRef] [PubMed]
- Bastos, D.C.; Ribeiro, C.F.; Ahearn, T.; Nascimento, J.; Pakula, H.; Clohessy, J.; Mucci, L.; Roberts, T.; Zanata, S.M.; Zadra, G.; Loda, M. Genetic ablation of FASN attenuates the invasive potential of prostate cancer driven by Pten loss. J. Pathol. 2021, 253, 292–303. [Google Scholar] [CrossRef] [PubMed]
- Azuma, K.; Osaki, T.; Tsuka, T.; Imagawa, T.; Minami, S.; Okamoto, Y. Plasma free amino acid profiles of canine mammary gland tumors. J. Vet. Sci. 2012, 13, 433. [Google Scholar] [CrossRef] [PubMed]
- Junior, R.P.; Sonehara, N.M.; Jardim-Perassi, B.V.; Pal, A.; Asad, Y.; Almeida Chuffa, L.G.; Chammas, R.; Raynaud, F.I.; Zuccari, D.A.P.C. Presence of human breast cancer xenograft changes the diurnal profile of amino acids in mice. Sci. Rep. 2022, 12, 1008. [Google Scholar] [CrossRef]
- Derakhshan, F.; Reis-Filho, J.S. Pathogenesis of Triple-Negative Breast Cancer. Annu. Rev. Pathol. 2022, 17, 181–204. [Google Scholar] [CrossRef]
- Yin, L.; Duan, J.-J.; Bian, X.-W.; Yu, S.-C. Triple-negative breast cancer molecular subtyping and treatment progress. Breast Cancer Res. 2020, 22, 61. [Google Scholar] [CrossRef]
- Gross, M.I.; Demo, S.D.; Dennison, J.B.; Chen, L.; Chernov-Rogan, T.; Goyal, B.; Janes, J.R.; Laidig, G.J.; Lewis, E.R.; Li, J.; Mackinnon, A.L.; Parlati, F.; Rodriguez, M.L.M.; Shwonek, P.J.; Sjogren, E.B.; Stanton, T.F.; Wang, T.; Yang, J.; Zhao, F.; Bennett, M.K. Antitumor activity of the glutaminase inhibitor CB-839 in triple-negative breast cancer. Mol. Cancer Ther. 2014, 13, 890–901. [Google Scholar] [CrossRef]
- Ryu, J.E.; Park, H.K.; Choi, H.J.; Lee, H.B.; Lee, H.J.; Lee, H.; Yu, E.S.; Son, W.C. Expression of the glutamine metabolism-related proteins glutaminase 1 and glutamate dehydrogenase in canine mammary tumours. Vet. Comp. Oncol. 2018, 16, 239–245. [Google Scholar] [CrossRef]
- Wakshlag, J.J.; McNeill, C.J.; Antonyak, M.A.; Boehm, J.E.; Fuji, R.; Balkman, C.E.; Zgola, M.; Cerione, R.A.; Page, R.L. Expression and activity of transglutaminase II in spontaneous tumours of dogs and cats. J. Comp. Pathol. 2006, 134, 202–210. [Google Scholar] [CrossRef]
- Fukumoto, S.; Hanazono, K.; Komatsu, T.; Ueno, H.; Kadosawa, T.; Iwano, H.; Uchide, T. L-type amino acid transporter 1 (LAT1): a new therapeutic target for canine mammary gland tumour. Vet. J. 2013, 198, 164–169. [Google Scholar] [CrossRef]
- Cherrington, B.D.; Mohanan, S.; Diep, A.N.; Fleiss, R.; Sudilovsky, D.; Anguish, L.J.; Coonrod, S.A.; Wakshlag, J.J. Comparative analysis of peptidylarginine deiminase-2 expression in canine, feline and human mammary tumours. J. Comp. Pathol. 2012, 147, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Cherrington, B.D.; Morency, E.; Struble, A.M.; Coonrod, S.A.; Wakshlag, J.J. Potential role for peptidylarginine deiminase 2 (PAD2) in citrullination of canine mammary epithelial cell histones. PLoS ONE 2010, 5, e11768. [Google Scholar] [CrossRef]
- Kim, K.S.; Lee, S.E.; Jeong, H.W.; Ha, J.H. The complete nucleotide sequence of the domestic dog (Canis familiaris) mitochondrial genome. Mol. Phylogenet. Evol. 1998, 10, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Bertagnolli, A.C.; Soares, P.; van Asch, B.; Amorim, A.; Cirnes, L.; Máximo, V.; Cassali, G.D. An assessment of the clonality of the components of canine mixed mammary tumours by mitochondrial DNA analysis. Vet. J. 2009, 182, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Kowal, K.; Tkaczyk-Wlizło, A.; Pierzchała, M.; Gawor, J.; Ślaska, B. Molecular differences in mitochondrial DNA genomes of dogs with malignant mammary tumours. Vet. Comp. Oncol. 2022, 20, 256–264. [Google Scholar] [CrossRef]
- Karayannopoulou, M.; Fytianou, A.; Assaloumidis, N.; Psalla, D.; Constantinidis, T.C.; Kaldrymidou, E.; Koutinas, A.F. Markers of lipid peroxidation and α-tocopherol levels in the blood and neoplastic tissue of dogs with malignant mammary gland tumors. Vet. Clin. Pathol. 2013, 42, 323–328. [Google Scholar] [CrossRef]
- Szczubiał, M.; Kankofer, M.; Łopuszyński, W.; Dabrowski, R.; Lipko, J. Oxidative stress parameters in bitches with mammary gland tumours. J. Vet. Med. A Physiol. Pathol. Clin. Med. 2004, 51, 336–340. [Google Scholar] [CrossRef]
- Kumaraguruparan, R.; Balachandran, C.; Manohar, B.M.; Nagini, S. Altered oxidant-antioxidant profile in canine mammary tumours. Vet. Res. Commun. 2005, 29, 287–296. [Google Scholar] [CrossRef]
- Surdyka, M.; Slaska, B. Defect of the mitochondrial DNA hypervariable region as a risk factor for canine mammary tumour. Vet. Comp. Oncol. 2017, 15, 820–828. [Google Scholar] [CrossRef]
- Surdyka, M.; Slaska, B. Defect in ND2, COX2, ATP6 and COX3 mitochondrial genes as a risk factor for canine mammary tumour. Vet. Comp. Oncol. 2017, 15, 1062–1072. [Google Scholar] [CrossRef]
- Yang, Y.; Mei, C.; Xian, H.; Zhang, X.; Li, J.; Liang, Z.-X.; Zhi, Y.; Ma, Y.; Wang, H.-J. Toosendanin-induced apoptosis of CMT-U27 is mediated through the mitochondrial apoptotic pathway. Vet. Comp. Oncol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Lopes, J.R.; Maschio, L.B.; Jardim-Perassi, B.V.; Moschetta, M.G.; Ferreira, L.C.; Martins, G.R.; Gelaleti, G.B.; De Campos Zuccari, D.A.P. Evaluation of melatonin treatment in primary culture of canine mammary tumors. Oncol. Rep. 2015, 33, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Zambrano-Estrada, X.; Landaverde-Quiroz, B.; Dueñas-Bocanegra, A.A.; De Paz-Campos, M.A.; Hernández-Alberto, G.; Solorio-Perusquia, B.; Trejo-Mandujano, M.; Pérez-Guerrero, L.; Delgado-González, E.; Anguiano, B.; Aceves, C. Molecular iodine/doxorubicin neoadjuvant treatment impair invasive capacity and attenuate side effect in canine mammary cancer. BMC Vet. Res. 2018, 14, 87. [Google Scholar] [CrossRef] [PubMed]
- Li, H.T.; Song, X.Y.; Yang, C.; Li, Q.; Tang, D.; Tian, W.R.; Liu, Y. Effect of hematoporphyrin monomethyl ether-mediated PDT on the mitochondria of canine breast cancer cells. Photodiagnosis Photodyn. Ther. 2013, 10, 414–421. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Alterations in canine mammary tumor energy metabolism. Metabolic rewiring in female dogs is frequently observed involves as changes in glucose demands (Warburg effect), mitochondrial metabolism, amino acid profile and lipid metabolism. Tumors commonly increase glucose uptake to supply their anabolic demands, being the energy metabolism adjusted concerning amino acids and fatty acid usage. It is not clear the role of distinct fatty acids regarding their unsaturation status, which requires further investigation. Legend: CMT – canine mammary tumor; AAs – amino acids; FA – fatty acid; ETC – electron transport chain.
Figure 1.
Alterations in canine mammary tumor energy metabolism. Metabolic rewiring in female dogs is frequently observed involves as changes in glucose demands (Warburg effect), mitochondrial metabolism, amino acid profile and lipid metabolism. Tumors commonly increase glucose uptake to supply their anabolic demands, being the energy metabolism adjusted concerning amino acids and fatty acid usage. It is not clear the role of distinct fatty acids regarding their unsaturation status, which requires further investigation. Legend: CMT – canine mammary tumor; AAs – amino acids; FA – fatty acid; ETC – electron transport chain.
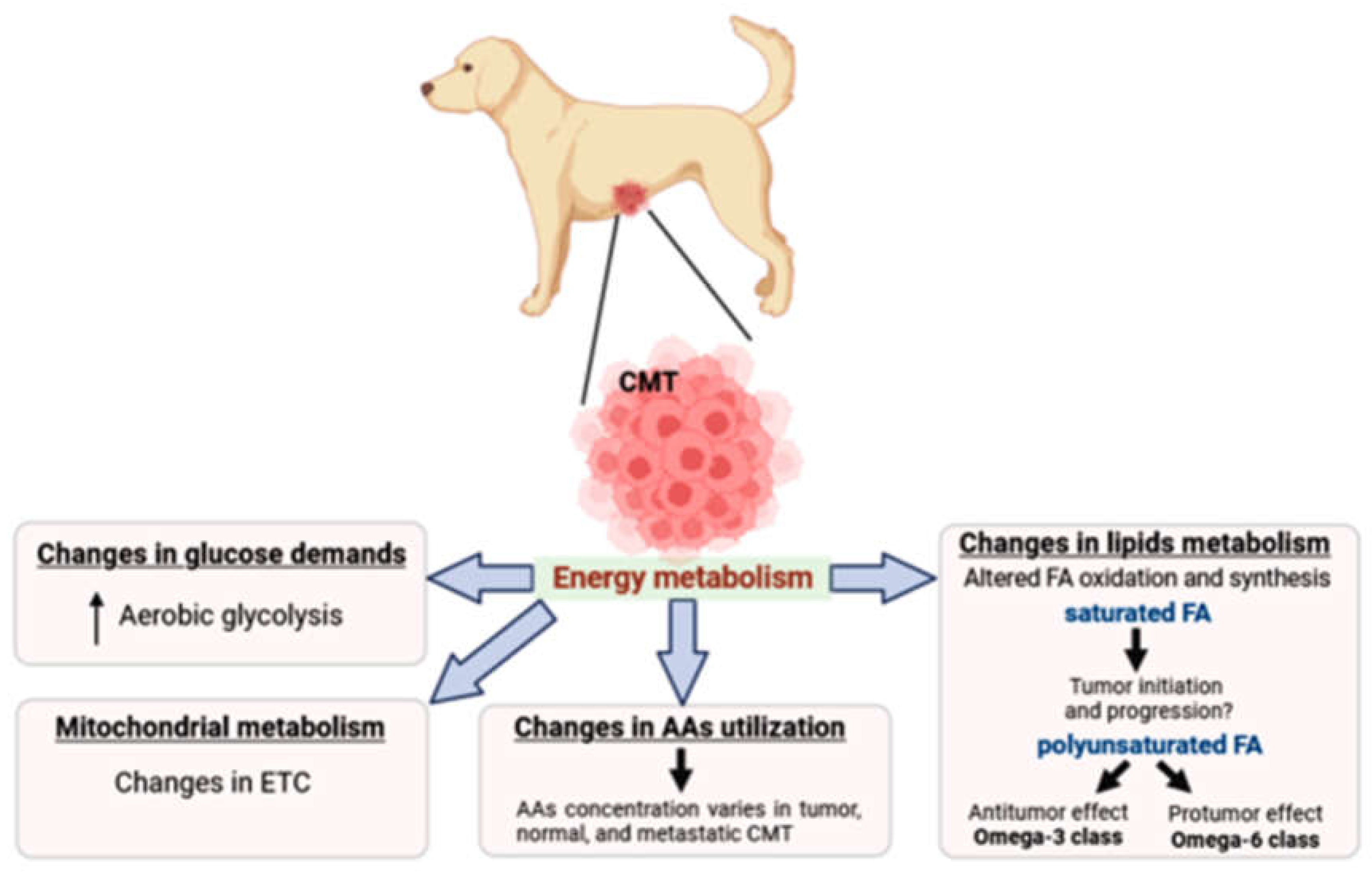
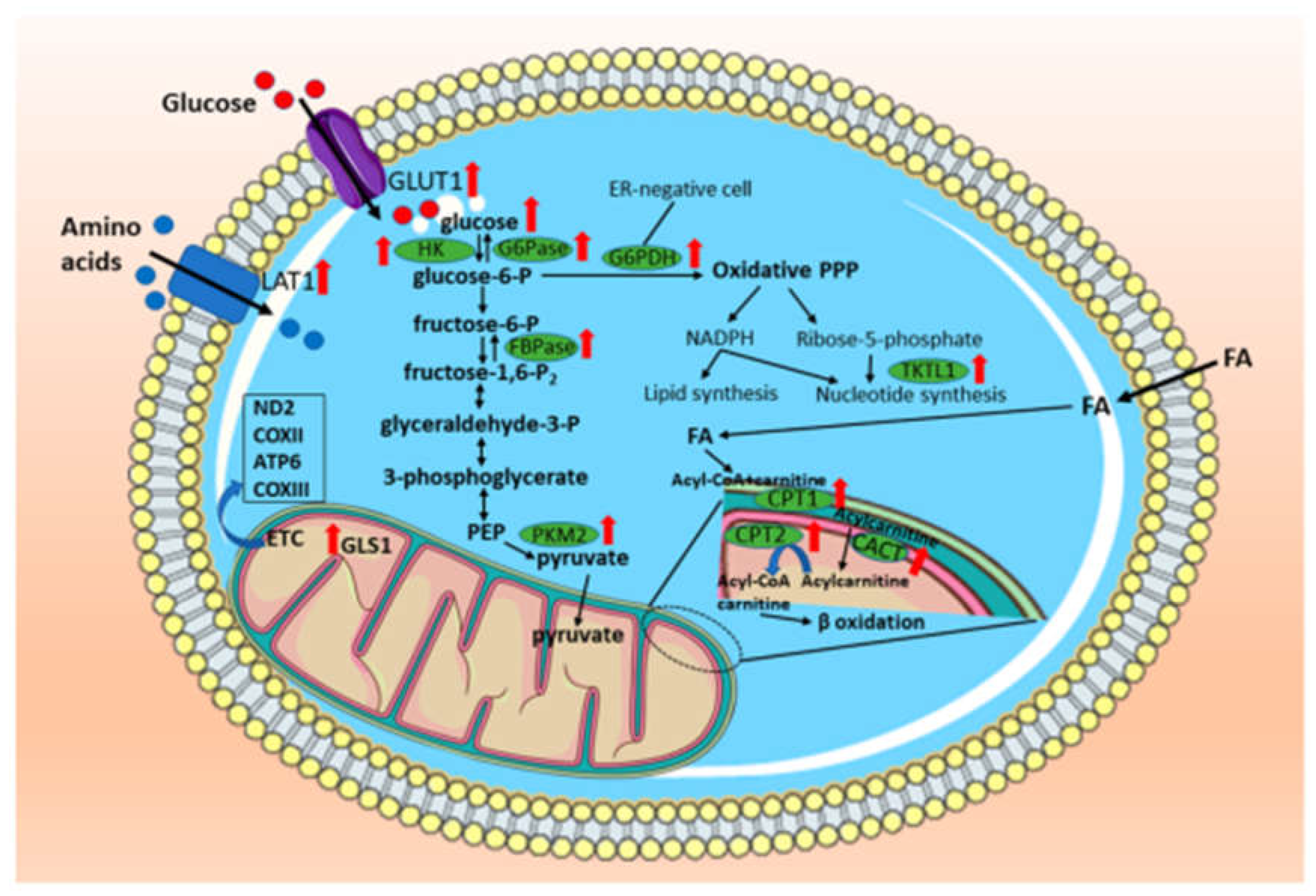
Figure 2.
Summary of metabolic alterations in canine mammary tumors. It was found changes in carbohydrate, amino acids, lipid, and mitochondrial metabolism. Glucose uptake and glycolysis enzymes HK and PKM2 are increased in CMTs. This pattern is also observed for gluconeogenesis and the pentose phosphate pathway. Amino acid utilization was reported to be altered, especially the glutamine metabolism via glutaminase 1 and the amino acids transport. In addition, fatty acid oxidation is increased due to the overexpression of enzymes related to carnitine system. Such alterations involve mitochondrial metabolism which is affected in CMTs mainly the ETC genes given that they are described as hotspot of mutations in mtDNA. Legend: GLUT1 – glucose transporter 1; HK – hexokinase; G6Pase – glucose-6-phosphatase; FBPase – fructose-1,6-bisphosphatase; PKM2 – pyruvate kinase muscle 2; LAT1 - L-type amino acid transporter 1; TKTL1 - transketolase-like 1 – CPT1 – carnitine palmitoyl transferase 1; CPT2 – carnitine palmitoyl transferase 2; CACT – Carnitine Acylcarnitine Translocase; ND2 – NADH dehydrogenase subunit 2; COXII – cytochrome c oxidase subunit II; ATP6 – ATP synthase F0 subunit 6; COXIII – cytochrome c oxidase subunit III; ER – estrogen receptor; FA – fatty acid; GLS1 – glutaminase 1; PPP – pentose phosphate pathway; ETC – electron transport chain; mtDNA – mitochondrial DNA.
Figure 2.
Summary of metabolic alterations in canine mammary tumors. It was found changes in carbohydrate, amino acids, lipid, and mitochondrial metabolism. Glucose uptake and glycolysis enzymes HK and PKM2 are increased in CMTs. This pattern is also observed for gluconeogenesis and the pentose phosphate pathway. Amino acid utilization was reported to be altered, especially the glutamine metabolism via glutaminase 1 and the amino acids transport. In addition, fatty acid oxidation is increased due to the overexpression of enzymes related to carnitine system. Such alterations involve mitochondrial metabolism which is affected in CMTs mainly the ETC genes given that they are described as hotspot of mutations in mtDNA. Legend: GLUT1 – glucose transporter 1; HK – hexokinase; G6Pase – glucose-6-phosphatase; FBPase – fructose-1,6-bisphosphatase; PKM2 – pyruvate kinase muscle 2; LAT1 - L-type amino acid transporter 1; TKTL1 - transketolase-like 1 – CPT1 – carnitine palmitoyl transferase 1; CPT2 – carnitine palmitoyl transferase 2; CACT – Carnitine Acylcarnitine Translocase; ND2 – NADH dehydrogenase subunit 2; COXII – cytochrome c oxidase subunit II; ATP6 – ATP synthase F0 subunit 6; COXIII – cytochrome c oxidase subunit III; ER – estrogen receptor; FA – fatty acid; GLS1 – glutaminase 1; PPP – pentose phosphate pathway; ETC – electron transport chain; mtDNA – mitochondrial DNA.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



