
Submitted:
26 June 2023
Posted:
26 June 2023
You are already at the latest version
Abstract
Adult T-cell leukemia (ATL) is an aggressive hematologic disease caused by human T-cell leukemia virus type 1 (HTLV-1) infection. Various animal models of HTLV-1 infection/ATL have been established to elucidate the pathogenesis of ATL and develop appropriate treatments. For analyses employing murine models, transgenic and immunodeficient mice are used because of the low infectivity of HTLV-1 in mice. HTLV-1 Tax and HBZ transgenic mice spontaneously develop tumors, and roles for both Tax and HBZ in cell transformation and tumor growth have been established. Severely immunodeficient mice were able to engraft ATL cell lines and have been used in preclinical studies of candidate molecules for the treatment of ATL. HTLV-1-infected humanized mice with an established human immune system are a suitable model to characterize cells in the early stages of HTLV-1 infection. This review outlines the characteristics of mouse models of HTLV-1 infection/ATL and describes progress made in elucidating the pathogenesis of ATL and developing related therapies using these mice.
Keywords:
HTLV-1
; ATL
; Tax
; HBZ
; immunodeficient mouse
; humanized mouse
1. Introduction
Human T-cell leukemia virus type 1 (HTLV-1) was first discovered as a pathogenic human retrovirus [1,2]. At least 5 to 10 million people worldwide are infected with HTLV-1, and regions with large numbers of HTLV-1 carriers include Japan, the Caribbean Islands, Latin America, regions of tropical Africa, and Central Australia [3]. HTLV-1 primarily infects CD4+ T cells, and the virus is not eliminated and persists throughout life. Although most infected individuals are asymptomatic, HTLV-1 can cause malignant tumors known as adult T-cell leukemia (ATL) [4,5] and neurodegenerative diseases such as HTLV-1-associated myelopathy (HAM) or tropical spastic paraparesis (TSP) after a latent period of several years to several decades [6]. The lifetime risk of developing ATL is 6%–7% in HTLV-1-infected men and 2%–3% in HTLV-1-infected women in Japan [7], while the risk of developing HAM/TSP is approximately 0.25%–3.8% in HTLV-1 infected individuals [8]. Although new treatments for these diseases are being developed, they remain refractory.
ATL is classified into four disease types: smoldering, chronic, acute, and lymphoma [9]. When ATL progresses to acute or lymphoma types, the 4-year survival rate is approximately 10% [10]. Acute ATL is characterized by increased lymphocytes and eosinophils, the detection of abnormal lymphocytes (called flower cells), hepatosplenomegaly, lymphadenopathy, bone marrow infiltrates, osteolytic lesions resulting in hypercalcemia, and skin lesions. The HTLV-1 genes Tax and HBZ are thought to be involved in the pathogenesis of ATL. Tax promotes viral transcription by activating the 5′ long terminal repeat (LTR). In addition, Tax transforms cells and can induce cell proliferation and immortalization. In vitro, Tax induces immortalization when transfected into T cells and fibroblasts. In vivo, Tax transgenic mice develop tumors. Tax activates nuclear factor (NF)-κB, nuclear factor of activated T cells (NFAT), and activator protein 1 (AP-1) pathways; suppresses p53 function; and regulates various intracellular signaling. Constitutive activation of NF-κB by Tax is important for cell transformation and tumor growth. In vivo, Tax is rarely expressed in HTLV-1-infected cells because of its high immunogenicity. In vitro experiments suggest that the expression pattern of Tax is transient [11], making it possible that transient expression of Tax in vivo allows cells to evade attack from cytotoxic T cells (CTLs). Half of patients with ATL are unable to fully express Tax because of loss of the 5' LTR or loss of transcriptional activity by methylation [12,13]. These findings suggest that Tax is not essential for the survival of infected cells with oncogenic transformation (ATL cells). HBZ is a gene encoded on the minus strand. Unlike Tax, HBZ is less immunogenic and is constitutively expressed in HTLV-1-infected cells and cancer cells. HBZ transgenic mice develop tumors [14]. Moreover, HBZ promotes ATL cell proliferation [15], inhibits apoptosis [16], and modulates DNA damage responses [17].
Treatments for ATL include multidrug chemotherapy, zidovudine/interferon (IFN) combination therapy [18,19], and allogeneic hematopoietic stem cell transplantation [20]. In addition, treatment with mogamulizumab, an anti-CCR4 antibody, has shown some efficacy in the treatment of ATL [21,22]. Tax peptide-pulsed dendritic cell vaccine (Tax-DC), which enhances Tax-specific CTL responses, has also shown efficacy against ATL in clinical trials [23].
Various animal models have been used to elucidate the pathogenesis of ATL and develop relevant therapies. Transgenic mouse models have greatly contributed to the analysis of Tax and HBZ in the transformation of HTLV-1-infected cells and tumor growth. Severely immunodeficient mice can engraft ATL cell lines, and an ATL model was developed to investigate therapeutic agents. Humanized mice with a human immune system can be characterized during the early stages of HTLV-1 infection. In the future, use of humanized mice with acquired immunity in addition to innate immunity will enhance HTLV-1 research, including the development of HTLV-1 vaccines. Here, we introduce the characteristics of each mouse model of HTLV-1 infection and ATL, as well as findings obtained using these mice.
2. Transgenic Mouse Models for HTLV-1 Research
To investigate the role of HTLV-1 in the pathogenesis of ATL, Tax and HBZ transgenic mice were generated. HTLV-1 mainly infects CD4+ T cells in humans, but Tax transgenic mice express Tax in different cells and tissues depending on the promoter used for gene expression. Transgenic mice in which the Tax gene is under control of the LTR promoter (LTR-Tax) express Tax specifically in tissues such as the thymus and muscle, and exhibit mesenchymal tumors [24]. Although LTR-Tax transgenic mice do not display ATL-like symptoms such as lymphoma or leukemia, Tax was proven to have transforming activity and cause tumor formation. Some transgenic mice also developed thymic atrophy [25], neurofibromas composed of perineural fibroblasts [26], exocrine abnormalities involving the salivary and lacrimal glands (similar to Sjogren's syndrome) [27], chronic arthritis similar to rheumatoid arthritis [28,29,30], and myelofibrosis [31]. To investigate Tax expression in detail within tissues in vivo, double-transgenic mice were generated by crossing LTR-Tax transgenic mice with LTR-βgal transgenic mice. These mice express Tax in a tissue-specific manner in muscle, bone, salivary glands, skin, and nerves [32]. Subsequently, transgenic mice have been generated using various promoters to express Tax specifically in lymphocytes. Transgenic mice expressing Tax under control of the human granzyme B promoter (GzmB-Tax) develop large granular lymphocytic (LGL) leukemia [33]. LGL cell lines established from GzmB-Tax transgenic mice exhibit pre-NK cell surface markers [34]. GzmB-Tax transgenic mice were observed to have hypercalcemia and osteolytic bone metastases, which frequently occur in patients with ATL [35]. Transgenic mice expressing Tax under control of the CD3-epsilon promoter display induced mesenchymal tumors, as well as salivary and mammary adenomas [36]. Transgenic mice expressing Tax under control of the proximal promoter of Lck (Lck-Tax), which restricts Tax expression to thymocytes, develop diffuse large cell lymphoma and leukemia after a long latent period. The phenotypes of lymphoma cells from Lck-Tax transgenic mice, which were transferred into severe combined immunodeficiency (SCID) mice, were pre-T cells (CD4-/CD8-/CD44+/CD25+ and cytosolic CD3+) [37]. Lck-distal Tax transgenic mice expressing Tax in mature thymocytes and peripheral T lymphocytes were generated. These mice develop T-cell leukemia and lymphoma, with the major phenotype of leukemic cells being mature CD4+ or CD8+ T cells [38]. In summary, Tax was shown to have cell transforming activity and cause tumors and various clinical manifestations. When Tax expression is restricted to lymphocytes, mice develop leukemias and lymphomas, showing ATL-like clinical symptoms.
The characteristics of tumors and tissues transformed by Tax in Tax transgenic mice have been analyzed. Fibroblast tumors generated in Tax transgenic mice express high levels of granulocyte-macrophage colony-stimulating factor, interleukin (IL)-6, platelet-derived growth factor β, Zif268, and c-fos [39]. Transforming growth factor (TGF)-β1 mRNA and protein are expressed at high levels in tumors of Tax transgenic mice and tissues such as the submandibular gland and skeletal muscle, which express high levels of Tax mRNA [40]. Elevated levels of inflammatory cytokines (e.g., IL-1α, IL-1β, IL-6, tumor necrosis factor α, TGF-β1, IFN-γ, and IL-2) and major histocompatibility complex (MHC) genes were detected in the joints of env-pX transgenic mice, which develop inflammatory arthritis [41]. In primary tumors freshly isolated from Tax transgenic mice, expression of NF-κB-inducible cytokines such as IL-6, IL-10, IL-15, and IFN-γ was elevated [42]. These findings suggest that in Tax transgenic mice, Tax constitutively activates NF-κB to enhance cell activation and inflammatory cytokine expression. The involvement of NF-κB activation in cell transformation, tumor development, and proliferation in Tax transgenic mice was demonstrated by sodium salicylate and prostaglandins, which inhibit NF-κB activity, inhibiting spontaneous proliferation of splenocytes in Tax transgenic mice. In addition, Tax-induced tumor cells resistant to radiation-induced apoptosis showed increased susceptibility to apoptosis in the presence of sodium salicylate and prostaglandin [42]. Mice treated with an antisense NF-κB oligodeoxynucleotide show rapid regression of transplanted Tax-transformed fibrosarcomas [43]. In contrast, transgenic mice with a Tax mutant defective in NF-κB activation fail to immortalize T cells and do not develop tumors [44]. In Tax transgenic mice, constitutive activation of NF-κB by Tax strongly suggests enhanced cell transformation, tumor growth, and resistance to apoptosis.
Tumors from Tax transgenic mice are reportedly characterized by mutations in the p53 tumor suppressor gene and functional inactivation of wild-type p53 protein [45]. Tax transgenic mice heterozygous for the p53 gene show more rapid tumor dissemination and increased mortality. Experiments crossing Tax transgenic mice with p53-deficient mice show little or no acceleration of early tumorigenesis, whereas mice heterozygous for p53 show markedly accelerated disease progression and death. These studies suggest that functional inactivation of p53 by HTLV-1 Tax is unimportant for early tumorigenesis but contributes to late tumor progression [46]. IFN-γ-deficient Tax transgenic mice display accelerated tumor development, metastasis, and death [47]. IFN-γ-deficient Tax transgenic mice also display increased osteolytic bone lesions, soft tissue tumors, and osteoclast formation and activity [48]. The effect of osteoclast inhibition on tumorigenesis is indicated by double-transgenic mice for Tax and the osteoclast activator osteoprotegerin being protected from osteolytic bone disease and developing fewer soft tissue tumors. Similarly, osteoclast suppression by zoledronic acid attenuates bone and soft tissue tumors to prolong the survival of Tax transgenic mice [35].
Transgenic mice expressing HBZ under control of the granzyme B promoter (Gzmb-HBZ) develop lymphoproliferative diseases such as tumors, splenomegaly, abnormal white blood cell numbers, and hypercalcemia [49]. Transgenic mice expressing HBZ under control of the CD4 promoter exhibit induced T-cell lymphoma and systemic inflammation [14]. HBZ transgenic mice have proliferating Foxp3+ Treg cells and effector/memory T cells. HBZ directly induces transcription of Foxp3 in T cells, leading to their differentiation into Foxp3+ Treg cells. However, Treg function is disrupted by the interaction of HBZ with Foxp3 and NFAT. Because Foxp3+ T cells in HBZ transgenic mice have unstable Foxp3 expression, Foxp3+ T cells were transformed into Foxp3- T cells with enhanced IFN-γ production [50]. HBZ transgenic mice lacking IFN-γ display reduced incidence of inflammation and lymphoma [51]. These findings implicate increased IFN-γ production in the development of inflammation and lymphoma in HBZ transgenic mice. HBZ transgenic mice display higher expression of T cell immunoglobulin and ITIM domain (TIGIT, a co-suppressor molecule), and increased expression of IL-10 in TIGIT+ CD4+ cells [52]. HBZ was shown to interact with Grb2 and SHP-2-associated THEMIS, inhibiting suppressive signals from programmed death 1 and TIGIT to promote T cell proliferation [53]. Although IL-10 is an immunosuppressive cytokine, HBZ was shown to interact with both signal transducer and activator of transcription (STAT1) and STAT3, and modulate the IL-10/Janus kinase (JAK)/STAT signaling pathway to promote T cell proliferation [54]. Vaccination with recombinant vaccinia virus expressing HBZ induced HBZ-specific T cell responses, although several boosters were required. Transplantation of splenocytes from HBZ-immunized mice into HBZ transgenic mice increased survival, suggesting that anti-HBZ CTLs have a protective effect [55].
HBZ/Tax double-transgenic mice exhibit skin lesions and T-cell lymphoma [56]. ATL stem cells (ATLSCs) were identified in Tax transgenic and HBZ transgenic mice. In Tax transgenic mice, c-kit+/CD38-/CD71- cells were identified as an ATLSC candidate [57]. In HBZ transgenic mice, c-kit+/CD4-/CD8- cells were identified as an ATLSC candidate [58].
3. Development of Severely Immunodeficient Mice
Humanized mice are severely immunodeficient mice into which human cells and tissues are engrafted. To engraft human cells into mice, murine immune cells must be defective or dysfunctional. Various severely immunodeficient mouse models have been developed (Table 1). SCID (severe combined immunodeficiency) mice are severely immunodeficient and lack functional T and B cells [59] because of a nonsense mutation in the PRKDC (Protein Kinase, DNA-Activated, Catalytic polypeptide) gene preventing VDJ recombination of T and B cell receptors [60]. SCID mice were engrafted with mature human T and B cells upon transplantation with human fetal liver hematopoietic cells, human fetal thymus, and human fetal lymph nodes [61]. SCID mice were also short-term engrafted with human peripheral blood lymphocytes (PBL) when inoculated intraperitoneally [62]. However, natural killer (NK) cells and macrophages, which are involved in innate immune responses, have normal functions in SCID mice [63]. SCID mice leak T and B cells as they age [64], making them more sensitive to irradiation because of DNA repair abnormalities. NOD/scid mice were generated by crossing SCID mice with non-obese diabetic (NOD) mice, which have reduced NK cell, dendritic cell (DC), and macrophage activity [65]. NOD/scid mice display loss of T and B cells and reduced NK cell, DC, and macrophage activity. In addition, NOD/scid mice have an improved human peripheral blood mononuclear cells (PBMC) engraftment rate compared with SCID mice [66]. However, NOD/scid mice also display the characteristics of SCID mice, such as leakage of T and B cells and high radiosensitivity. Furthermore, NOD/scid mice develop thymic lymphoma with high frequency and have a short lifespan. NOG (NOD/SCID/IL2Rγnull) and NSG (NOD/SCID/IL2Rγnull) mice were developed by crossing NOD/scid mice with mice lacking the IL-2R common γ chain, which are defective in NK cells [67,68]. The IL-2R common γ chain in NOG mice harbors a mutation that allows cytokines to bind but not elicit signal, while the IL-2R common γ chain in NSG mice is completely defective. NOG and NSG mice display deficient T cell, B cell, NK cell, and complement activity, and have impaired DC and macrophage function. NOG and NSG mice do not develop T or B cell leakage, possibly because signals from the IL-2R common γ chain are responsible for the differentiation and proliferation of T and B cells. NOG and NSG mice have a higher rate of human cell engraftment than NOD/scid mice. Immunodeficient mice with phenotypes similar to NOG and NSG mice have also been developed. For example, NOJ (NOD/scid/jak-3null) mice were developed by crossing NOD/scid mice with mice lacking JAK-3 (jak-3), a tyrosine kinase involved in downstream signaling of the IL-2R common γ chain [69]. As an alternative to SCID mice, NRG (NOD/rag-1null/IL-2Rγnull) mice were developed by crossing mice deficient in recombination activating gene 1 (Rag1), a DNA recombinase essential for genetic reconstitution of T and B cell receptors [70].
4. Immunodeficient Mouse Models for HTLV-1 Research
Immunodeficient mice can be transplanted with HTLV-1-infected cell lines or patient-derived PBL. SCID mice depleted of NK cells by administration of an anti-Asiaro GM-1 antibody formed tumors when transplanted with MT-2 cells, an HTLV-1-infected cell line [71]. Intraperitoneal inoculation of MT-2 cells also resulted in tumor formation in SCID mice [72]. PBL from patients with ATL or HAM were inoculated into SCID mice [73,74]. Characterization of the inoculated cells revealed that ATL-derived PBL developed lymphoblastic lymphoma, while HAM-derived PBL had superior proliferative potential compared with uninfected PBL. T cells immortalized by gene transfer of Tax in vitro could not be maintained when transplanted into SCID mice [75]. Cells derived from Tax transgenic mice were maintained when transplanted into SCID mice, but the viable cells showed no detectable expression of Tax at the protein level and displayed constitutive NF-κB and Akt activity [34,76]. PDLIM2, a ubiquitin E3 ligase, ubiquitinates Tax in the nucleus, allowing Tax to be degraded by the proteasome. Transplantation of cell lines transfected with Tax and PDLIM2 into SCID mice inhibited tumorigenesis compared with transplantation of cell lines transfected with Tax alone [77]. Whether or not Tax is involved in cell proliferation and survival in vivo depended on the cells transplanted into immunodeficient mice. ATL cells from patients with ATL cannot express Tax because of mutations or promoter methylation [78]. These findings suggest that ATL cells eventually no longer require Tax for cell proliferation and survival. HBZ-knockdown HTLV-1 cell lines show markedly reduced tumor formation and organ invasion when transplanted into NOG mice [79]. HBZ was shown to play an important role in the proliferation and survival of HTLV-1-infected T cells.
Immunodeficient mice have contributed to HTLV-1 research as an in vivo model to evaluate candidate molecules for ATL therapeutics. An ATL model has been developed in which HTLV-1-infected and ATL cell lines are transplanted subcutaneously or into the abdominal cavity of immunodeficient mice, allowing measurement of tumor formation and tumor invasion into various organs. Since then, numerous preclinical studies of candidate molecules for ATL therapeutics have been conducted using the ATL model (Table 2). The NF-κB inhibitors Bay 11-7082 [80] and dehydroxymethylpoxyquinomycin (DHMEQ) [81,82,83,84] inhibit tumor formation and invasion derived from HTLV-1-infected cell lines implanted in severely immunodeficient mice. Other drugs that inhibit NF-κB activation have also been used as single agents or in combination with other drugs to treat ATL tumors, including bortezomib (PS-341, a proteasome inhibitor) [85,86], ritonavir (an HIV protease inhibitor) [87], diarsenic trioxide (As2O3, which degrades Tax protein) [88], 17-dimethylaminoethylamino-17-demethoxygeldanamycin hydrochloride (an inhibitor of heat shock protein 90) [89], and 9-aminoacridine [90]. Histone deacetylase inhibitors (HDACi) induce histone hyperacetylation, resulting in chromatin remodeling and reactivation of transcriptional repressor genes, and have shown efficacy against a variety of cancers. The HDACi depsipeptide [91], LBH589 [92], and AR-42 [93] prolonged survival of severely immunodeficient mice transplanted with HTLV-1-infected cell lines either as single agents or in combination with other drugs. The efficacy of apoptosis inhibitors and other inhibitors in the treatment of ATL has also been examined in an ATL model. ABT-737 [a small molecule inhibitor of Bcl-2, Bcl-X(L), and Bcl-w, which act in antiapoptosis] [94], fucoidan (a sulfated polysaccharide that inhibits survivin, an antiapoptotic protein) [95], the autophagy inhibitors chloroquine and hydroxychloroquine [96], the AMPK inhibitor dorsomorphin [97], the IPOα/β1 inhibitor ivermectin [98], the selective JAK inhibitor ruxolitinib in combination with the Bcl-2/Bcl-xL inhibitor navitoclax [99], and the dual SYK/JAK inhibitor cerdulatinib [100] suppressed tumors derived from HTLV-1-infected cells transplanted into immunodeficient mice. Other agents with antitumor activity include incadronate (a bisphosphonate) [101], indole-3-carbinol (a naturally occurring component of cruciferous vegetables) [102], and the biphosphine cyclopalladium complex [Pd2(S-C2,N-dmpa)2(μ-dppe)Cl2] [103]. In addition, the carotenoid peridinin [104], bioactive plant polyphenol butein [105], and organic compound thymoquinone in combination with low concentrations of Dox [106] were shown to be effective in treating ATL using ATL models. ATL cells are characterized by invasion into organs and skin. AMD3100, a CXCR4 antagonist, inhibited lymphoma cell infiltration into liver and lung tissues in vivo in an ATL model [107].
The ATL model with immunodeficient mice is a superior system for evaluating drug activity against ATL cells. However, immunodeficient mice are deficient in host immune cells and cannot be used for experiments involving host immune responses, such as vaccines or immune checkpoint therapy. These experiments require humanized mice/human immune system mice – immunodeficient mice in which human immune cells are constructed.
5. Development of Human Immune System Mice
Several methods are available to reconstitute the human immune system in severely immunodeficient mice, and the humanized mice/human immune system (HIS) mice generated by each method have different characteristics (Figure 1). The simplest is a method for the huPBL-SCID mouse model, which is generated by intravenous or intraperitoneal injection of human PBL into SCID mice [62]. In huPBL-SCID mice, transplanted cells survive for approximately 3 weeks after transplantation. Thereafter, immune cells that react to host mouse cells, mainly effector T cells, and only a few B cells are detected in huPBL-SCID mice [108]. huPBL-SCID mice have low human immune cell viability and reconstitution and develop severe xenogeneic graft-versus-host disease, which can be reduced by utilizing NSG mice deficient in murine MHC class I and class II as hosts [109].
The SCID-hu mouse model is humanized mice in which human fetal thymus and liver are transplanted under the renal capsule of SCID mice [61]. SCID-hu mice display low reconstitution of human immune cells and short T cell survival. The bone marrow liver thymus (BLT) mouse model, a modification of the SCID-hu mouse model, is generated by transplanting autologous human hematopoietic fetal liver CD34+ cells into NOD/SCID mice after engraftment of human fetal thymus and liver [110]. BLT mice display human T cells throughout the body for an extended period of time. In addition, repopulation of T cells, B cells, and DCs was observed in secondary lymphoid tissues. A significant advantage of BLT mice is that thymic transplantation induces differentiation of human HLA-recognizing T cells, which can induce a human MHC-restricted immune response [111,112]. However, the BLT mouse model is difficult to create because the availability of human fetal tissue is ethically and legally restricted in many countries.
hu-HSC mouse models are humanized mice generated by transplanting human hematopoietic stem cells (HSCs) into severely immunodeficient mice. Variations in these models arise from differences in human HSCs used for transplantation and the method of transplantation. Two types of HSCs are used for transplantation: CD34+ cells and CD133+ cells. CD133+ cells are more undifferentiated than CD34+ cells and can differentiate into neural and mesenchymal cells. Transplantation of CD133+ cells may affect the microenvironment of the bone marrow, where immune cells differentiate and are maintained. The first transplantation method of HSCs involves irradiating or administering busulfan to severely immunodeficient mice to create space in the bone marrow niche. Mice are then humanized by intravenous injection or intramedullary inoculation of human HSCs. Intra-bone marrow injection allows for a smaller number of HSCs to be used. The second method of HSC transplantation involves inoculation of the liver of neonatal severely immunodeficient mice. The advantages of this method are that the number of HSCs inoculated can be much less than in adult mice and humanization can be completed earlier. The characteristics of hu-HSC mice generated by these methods are similar, with reconstitution of B cells, DCs, macrophages, and NK cells by approximately 8 weeks after HSC transplantation, and T cells slightly later by approximately 12 weeks after transplantation. The reconstituted human immune cells are maintained for a long period of time.
6. Improvements in the hu-HSC Mice
Although the hu-HSC mouse model reconstitutes and maintains acquired and innate immune cells for a long time, it does not fully recapitulate human immune responses and has various issues. The most significant issue is that hu-HSC mice have weak acquired immune responses. T cells in hu-HSC mice cannot recognize human MHC (HLA) because they differentiate in the mouse thymus. Therefore, T cells cannot receive antigen stimulation from human antigen-presenting cells or elicit a human immune response. In B cells, antigen-specific IgM can be detected but antigen-specific IgG is rarely detected. It is speculated that the T-B interaction required for antibody class switching in B cells may be insufficient. To address these issues, HLA class II transgenic NSG [NSG Tg(HLA-DR4)] and NOG [NOG Tg(HLA-DR4)] mice were developed to display induction of a B cell class switch [113,114]. NSG mice expressing HLA class I-A2 [NSG Tg(HLA-A2)] and NSG mice expressing HLA class I-A2 and human β2 microglobulin binding domain [NSG Tg(HLA-A2, B2M)] were also developed [115,116]. These mice develop functionally mature CD8+ T cells. hu-HSC mice have fewer innate immune cells, such as DCs, macrophages and NK cells, likely because of a deficiency of cytokines that allow these cells to differentiate and proliferate. Administration of cytokines or cytokine expression vectors to mice or cytokine-transgenic mice improved myeloid and NK cell reconstitution [117,118,119,120,121]. NOG mice have inadequate lymph node development due to deficiency of the IL-2R common γ chain. NOG-pRORγt-γc Tg mice have been developed in which the IL-2R common γ chain gene is expressed in lymphoid tissue of NOG mice under induction of the endogenous promoter of RORγt. Humanized NOG-pRORγt-γc Tg mice have normalized lymph node organogenesis and enhanced antigen-specific IgG responses [122].
7. Human Immune System Mouse Models for HTLV-1 Research
The clonality of HTLV-1-infected cells was examined in the huPBMC-NOG mouse model. Primary infection was established when HTLV-1-uninfected PBMCs were transplanted into NOG mice, which were subsequently inoculated with HTLV-1-infected MT-2 cells [123]. Administration of the reverse transcriptase inhibitors azidothymidine and tenofovir, which suppress de novo infection, to huPBMC-NOG mice prevented primary HTLV-1 infection. However, administration of tenofovir one week after infection did not affect proviral load (PVL) [123]. In the huPBMC-NOG mouse model, PBMCs from asymptomatic HTLV-1 carriers were transplanted into NOG mice; one month after infection, specific clones of HTLV-1-infected cells were selectively proliferating [124]. These results suggest that in the huPBMC-NOG mouse model, de novo infection dominates the early stages of HTLV-1 infection while clonal proliferation dominates as infection progresses.
An HTLV-1-infected hu-HSC mouse model was developed to characterize infected cells. HIS Rag2-/-γc-/- mice were generated by intrahepatic transplantation of human CD34+ cells into neonatal BALB/c Rag2-/-γc-/- mice [125]. Several months after infecting HIS Rag2-/-γc-/- mice with HTLV-1, hepatosplenomegaly, lymphadenopathy, and lymphoma/thymoma were observed in which Tax was detected [125]. IBMI-huNOG mice were generated by intrabonemarrow injection (IBMI) of CD133+ cells into NOG mice [126]. HTLV-1 infection of IBMI-huNOG mice resulted in a rapid increase in peripheral CD4+ T cells and PVL. The mice developed little lymphoma, although enlargement of the spleen and liver was observed. Leukemic symptoms were observed from the early stages of infection, and cells with a laminar nucleus morphologically similar to the flower cells observed in patients with ATL were detected in peripheral blood 4 to 5 months post-infection. Moreover, HTLV-1-infected IBMI-huNOG mice were cytokineemic, similar to patients with ATL [126]. High clonality of cell populations was detected in mice infected for as long as 18 or 23 weeks [126]. A significant feature of HTLV-1-infected IBMI-huNOG mice was the detection of Tax-specific CTLs and production of HTLV-1-specific IgG. HIS NSG mice were generated by engraftment of CD34+CD38-lin- HSCs into the liver of neonatal NSG mice [127]. HTLV-1 infection of HIS NSG mice resulted in a rapid increase of CD4+ T cells in peripheral blood and lymphoid tissues. CD4+ T cells displayed phenotypes of effector memory cells and Th1 cells [127]. HTLV-1-infected HIS NSG mice generated by another group also developed lymphoproliferative transformation by 3 weeks post-infection, which progressed to terminal lymphoproliferative disease by 6–8 weeks (median survival, 5–6 weeks) post-infection [128]. HTLV-1-infected cells in these mice proliferated oligoclonally. These mice also developed osteolytic bone lesions.
HTLV-1-infected hu-HSC mouse models are being evaluated for drugs that target HTLV-1-infected cells. Truncated Pseudomonas exotoxin (PE38) fused with the CCR4 ligand CCL17/thymus and activation-regulated chemokine (TARC) was evaluated to selectively eliminate HTLV-1-infected cells [129]. Full-length PE is known to bind specifically to CD91 on the cell surface via its CD91-binding domain and subsequently be translocated intracellularly by endocytosis, causing cytotoxic effects dependent on the intracellular expression of furin, a protein convertase [130]. TARC-PE38 replaces the CD91-binding domain of PE with TARC, thereby exerting cytotoxic effects on CCR4-expressing cells. In HTLV-1-infected hu-HSC mice, TARC-PE38 markedly inhibits the proliferation of HTLV-1-infected human CD4+CD25+ cells and CD4+CD25+CCR4+ cells, and reduces PVL in PBMC [129]. HTLV-1-infected CD4+ T cells secrete leukotriene B4 (LTB4), a potent chemoattractant [131]. Inhibition of LTB4 secretion by MK886, an inhibitor of the 5-LO cofactor FLAP, in an HTLV-1-infected hu-HSC mouse model reduces both HTLV-1 PVL and the number of infected clones [131]. Recombinant vesicular stomatitis virus (VSV) has also been used to target HTLV-1-infected cells. VSV infects a variety of cell types in vitro, produces many progeny virions, and induces cytolysis of infected cells. VSVΔG-GL, VSVΔG-NP, and VSVΔG-SD were prepared by replacing VSV-G (the envelope glycoprotein of VSV) with GLUT1, NRP1, and SDC1, which HTLV-1 binds as receptors [132]. In the HTLV-1-infected hu-HSC mouse model, VSVΔG-NP efficiently prevented HTLV-1-induced leukocytosis in the periphery and eliminated HTLV-1-infected Env-expressing cells in bone marrow and spleen [132]. The transcription factor myocyte enhancer factor (MEF)-2 family member MEF-2C was enriched in the 3' LTR of HTLV-1 and observed to bind HBZ in this region, along with its cofactors Menin and JunD. Chemical inhibition of the MEF-2 protein reduced PVL in the HTLV-1-infected hu-HSC mouse model [133]. Protein arginine methyltransferase 5 (PRMT5), a type II PRMT enzyme, is directly involved in the pathogenesis of several different lymphomas through transcriptional regulation of related oncogenes [133]. EPZ015666, an inhibitor of PRMT5, enhanced survival of HIS NSG mice inoculated with HTLV-1-producing cells [134].
In the hu-HSC mouse model, mutant HTLV-1 can be transmitted to mice by HTLV-1 transgenic cells with mutations in the HTLV-1 gene as the source of infection. To investigate the in vivo contribution of the Tax PDZ domain-binding motif (PBM) to the lymphocyte proliferation process, cells transfected with HTLV-1 gene harboring a deletion of the Tax PBM were generated [135]. hu-HSC mice inoculated with these cells for HTLV-1 infection showed reduced frequencies of activated CD4+ T cells in the peripheral blood and spleen compared with mice inoculated with wild-type-transfected cells. These results suggest that the PBM of Tax is required for sustained proliferation of HTLV-1-infected T cells. To evaluate the role of HBZ in ATL-associated bone destruction, hu-HSC mice were infected with HTLV-1 lacking HBZ. Compared with wild-type HTLV-1-infected mice, HBZ-deficient HTLV-1-infected mice display a slight delay in lymphoproliferative disease and significant reduction of disease-related bone loss [128]. An enhancer inside the virus was found in the Tax gene near the 3' LTR of HTLV-1. To investigate whether this enhancer affects disease development, HTLV-1-infected cells carrying a mutant enhancer virus (mEnhancer) were transplanted into hu-HSC mice and evaluated. In mEnhancer-infected hu-HSC mice, the enhancer did not affect disease progression, PVL, or viral gene expression [136].
8. Conclusions
Transgenic mice for HTLV-1 Tax and HBZ are useful tools for understanding the role of these genes in the pathogenesis of HTLV-1-related leukemia and lymphoma (ATL). Severely immunodeficient mice allow transplantation of ATL cell lines, advancing the development of ATL therapeutics. Humanized mice with reconstructed human immune systems (HIS mice) have been used, for example, to develop preventive drugs targeting HTLV-1-infected cells. Current HIS mice do not fully recapitulate the human immune system. Therefore, next-generation humanized mice with different reconstructed immune cells have been being developed to improve modeling of the human immune system in mice. It is necessary to select the most appropriate next-generation HIS mice for HTLV-1 research. In the future, animal models for HTLV-1 infection and ATL using next-generation HIS mice are expected to be developed for various purposes, such as elucidating the pathogenesis of ATL, developing methods to prevent the onset of ATL, and developing vaccines against HTLV-1 infection and drugs to prevent or treat ATL.
Author Contributions
writing—original draft preparation, S.N.; writing—review and editing, K.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
We thank Edanz (https://jp.edanz.com/ac) for editing a draft of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Poiesz, B.J.; Ruscetti, F.W.; Gazdar, A.F.; Bunn, P.A.; Minna, J.D.; Gallo, R.C. Detection and Isolation of Type C Retrovirus Particles from Fresh and Cultured Lymphocytes of a Patient with Cutaneous T-Cell Lymphoma. Proc. Natl. Acad. Sci. U. S. A. 1980, 77, 7415–7419. [Google Scholar] [CrossRef]
- Poiesz, B.J.; Ruscetti, F.W.; Reitz, M.S.; Kalyanaraman, V.S.; Gallo, R.C. Isolation of a New Type C Retrovirus (HTLV) in Primary Uncultured Cells of a Patient with Sézary T-Cell Leukaemia. Nature 1981, 294, 268–271. [Google Scholar] [CrossRef]
- Gessain, A.; Cassar, O. Epidemiological Aspects and World Distribution of HTLV-1 Infection. Front. Microbiol. 2012, 3, 388. [Google Scholar] [CrossRef]
- Takatsuki, K.; Uchiyama, T.; Sagawa, K.; Yodoi, J.; Seno, S.; Takaku, F.; Irino, S. Topics in Hematology. In Proceedings of the The 16th International Congress of Hematology; Excerpta Medica: Amsterdam, 1977; pp. 73–77.
- Uchiyama, T.; Yodoi, J.; Sagawa, K.; Takatsuki, K.; Uchino, H. Adult T-Cell Leukemia: Clinical and Hematologic Features of 16 Cases. Blood 1977, 50, 481–492. [Google Scholar] [CrossRef]
- Gessain, A.; Barin, F.; Vernant, J.C.; Gout, O.; Maurs, L.; Calender, A.; de Thé, G. Antibodies to Human T-Lymphotropic Virus Type-I in Patients with Tropical Spastic Paraparesis. Lancet (London, England) 1985, 2, 407–410. [Google Scholar] [CrossRef]
- Iwanaga, M.; Watanabe, T.; Yamaguchi, K. Adult T-Cell Leukemia: A Review of Epidemiological Evidence. Front. Microbiol. 2012, 3, 322. [Google Scholar] [CrossRef]
- Yamano, Y.; Sato, T. Clinical Pathophysiology of Human T-Lymphotropic Virus-Type 1-Associated Myelopathy/Tropical Spastic Paraparesis. Front. Microbiol. 2012, 3, 389. [Google Scholar] [CrossRef]
- Shimoyama, M. Diagnostic Criteria and Classification of Clinical Subtypes of Adult T-Cell Leukaemia-Lymphoma. A Report from the Lymphoma Study Group (1984-87). Br. J. Haematol. 1991, 79, 428–437. [Google Scholar] [CrossRef]
- Katsuya, H.; Ishitsuka, K.; Utsunomiya, A.; Hanada, S.; Eto, T.; Moriuchi, Y.; Saburi, Y.; Miyahara, M.; Sueoka, E.; Uike, N.; et al. Treatment and Survival among 1594 Patients with ATL. Blood 2015, 126, 2570–2577. [Google Scholar] [CrossRef]
- Mahgoub, M.; Yasunaga, J.-I.; Iwami, S.; Nakaoka, S.; Koizumi, Y.; Shimura, K.; Matsuoka, M. Sporadic on/off Switching of HTLV-1 Tax Expression Is Crucial to Maintain the Whole Population of Virus-Induced Leukemic Cells. Proc. Natl. Acad. Sci. U. S. A. 2018, 115, E1269–E1278. [Google Scholar] [CrossRef]
- Furukawa, Y.; Kubota, R.; Tara, M.; Izumo, S.; Osame, M. Existence of Escape Mutant in HTLV-I Tax during the Development of Adult T-Cell Leukemia. Blood 2001, 97, 987–993. [Google Scholar] [CrossRef]
- Koiwa, T.; Hamano-Usami, A.; Ishida, T.; Okayama, A.; Yamaguchi, K.; Kamihira, S.; Watanabe, T. 5’-Long Terminal Repeat-Selective CpG Methylation of Latent Human T-Cell Leukemia Virus Type 1 Provirus in Vitro and in Vivo. J. Virol. 2002, 76, 9389–9397. [Google Scholar] [CrossRef]
- Satou, Y.; Yasunaga, J.-I.; Zhao, T.; Yoshida, M.; Miyazato, P.; Takai, K.; Shimizu, K.; Ohshima, K.; Green, P.L.; Ohkura, N.; et al. HTLV-1 BZIP Factor Induces T-Cell Lymphoma and Systemic Inflammation in Vivo. PLoS Pathog. 2011, 7, e1001274. [Google Scholar] [CrossRef]
- Satou, Y.; Yasunaga, J.; Yoshida, M.; Matsuoka, M. HTLV-I Basic Leucine Zipper Factor Gene MRNA Supports Proliferation of Adult T Cell Leukemia Cells. Proc. Natl. Acad. Sci. U. S. A. 2006, 103, 720–725. [Google Scholar] [CrossRef]
- Tanaka-Nakanishi, A.; Yasunaga, J.; Takai, K.; Matsuoka, M. HTLV-1 BZIP Factor Suppresses Apoptosis by Attenuating the Function of FoxO3a and Altering Its Localization. Cancer Res. 2014, 74, 188–200. [Google Scholar] [CrossRef]
- Vernin, C.; Thenoz, M.; Pinatel, C.; Gessain, A.; Gout, O.; Delfau-Larue, M.-H.; Nazaret, N.; Legras-Lachuer, C.; Wattel, E.; Mortreux, F. HTLV-1 BZIP Factor HBZ Promotes Cell Proliferation and Genetic Instability by Activating OncomiRs. Cancer Res. 2014, 74, 6082–6093. [Google Scholar] [CrossRef]
- Gill, P.S.; Harrington, W.J.; Kaplan, M.H.; Ribeiro, R.C.; Bennett, J.M.; Liebman, H.A.; Bernstein-Singer, M.; Espina, B.M.; Cabral, L.; Allen, S. Treatment of Adult T-Cell Leukemia-Lymphoma with a Combination of Interferon Alfa and Zidovudine. N. Engl. J. Med. 1995, 332, 1744–1748. [Google Scholar] [CrossRef]
- Hermine, O.; Bouscary, D.; Gessain, A.; Turlure, P.; Leblond, V.; Franck, N.; Buzyn-Veil, A.; Rio, B.; Macintyre, E.; Dreyfus, F. Brief Report: Treatment of Adult T-Cell Leukemia-Lymphoma with Zidovudine and Interferon Alfa. N. Engl. J. Med. 1995, 332, 1749–1751. [Google Scholar] [CrossRef]
- Okamura, J.; Utsunomiya, A.; Tanosaki, R.; Uike, N.; Sonoda, S.; Kannagi, M.; Tomonaga, M.; Harada, M.; Kimura, N.; Masuda, M.; et al. Allogeneic Stem-Cell Transplantation with Reduced Conditioning Intensity as a Novel Immunotherapy and Antiviral Therapy for Adult T-Cell Leukemia/Lymphoma. Blood 2005, 105, 4143–4145. [Google Scholar] [CrossRef]
- Ishida, T.; Joh, T.; Uike, N.; Yamamoto, K.; Utsunomiya, A.; Yoshida, S.; Saburi, Y.; Miyamoto, T.; Takemoto, S.; Suzushima, H.; et al. Defucosylated Anti-CCR4 Monoclonal Antibody (KW-0761) for Relapsed Adult T-Cell Leukemia-Lymphoma: A Multicenter Phase II Study. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2012, 30, 837–842. [Google Scholar] [CrossRef]
- Shichijo, T.; Nosaka, K.; Tatetsu, H.; Higuchi, Y.; Endo, S.; Inoue, Y.; Toyoda, K.; Kikukawa, Y.; Kawakita, T.; Yasunaga, J.-I.; et al. Beneficial Impact of First-Line Mogamulizumab-Containing Chemotherapy in Adult T-Cell Leukaemia-Lymphoma. Br. J. Haematol. 2022, 198, 983–987. [Google Scholar] [CrossRef]
- Suehiro, Y.; Hasegawa, A.; Iino, T.; Sasada, A.; Watanabe, N.; Matsuoka, M.; Takamori, A.; Tanosaki, R.; Utsunomiya, A.; Choi, I.; et al. Clinical Outcomes of a Novel Therapeutic Vaccine with Tax Peptide-Pulsed Dendritic Cells for Adult T Cell Leukaemia/Lymphoma in a Pilot Study. Br. J. Haematol. 2015, 169, 356–367. [Google Scholar] [CrossRef]
- Nerenberg, M.; Hinrichs, S.H.; Reynolds, R.K.; Khoury, G.; Jay, G. The Tat Gene of Human T-Lymphotropic Virus Type 1 Induces Mesenchymal Tumors in Transgenic Mice. Science 1987, 237, 1324–1329. [Google Scholar] [CrossRef]
- Furuta, Y.; Aizawa, S.; Suda, Y.; Ikawa, Y.; Kishimoto, H.; Asano, Y.; Tada, T.; Hikikoshi, A.; Yoshida, M.; Seiki, M. Thymic Atrophy Characteristic in Transgenic Mice That Harbor PX Genes of Human T-Cell Leukemia Virus Type I. J. Virol. 1989, 63, 3185–3189. [Google Scholar] [CrossRef]
- Hinrichs, S.H.; Nerenberg, M.; Reynolds, R.K.; Khoury, G.; Jay, G. A Transgenic Mouse Model for Human Neurofibromatosis. Science (80-. ). 1987, 237, 1340–1343. [Google Scholar] [CrossRef]
- Green, J.E.; Hinrichs, S.H.; Vogel, J.; Jay, G. Exocrinopathy Resembling Sjögren’s Syndrome in HTLV-1 Tax Transgenic Mice. Nature 1989, 341, 72–74. [Google Scholar] [CrossRef]
- Yoshida, E.M.I.; Takiguchi, M.; Hatanaka, M.; Yamamoto, H. Induction of Inflammatory Arthropathy Resembling Rheumatoid Artbritis. 1983, 253, 4–6.
- Yamamoto, H.; Sekiguchi, T.; Yamamoto, I. Histopathological Observation of Joint Lesions of Extremities in Mice Transferred Genome. Exp. Toxicol. Pathol. Off. J. Gesellschaft fur Toxikologische Pathol. 1993, 45, 233–238. [Google Scholar] [CrossRef]
- Yamamoto, H.; Sekiguchi, T.; Itagaki, K.; Saijo, S.; Iwakura, Y. Inflammatory Polyarthritis in Mice Transgenic for Human T Cell Leukemia Virus Type I. Arthritis Rheum. 1993, 36, 1612–1620. [Google Scholar] [CrossRef]
- Ruddle, N.H.; Li, C.B.; Horne, W.C.; Santiago, P.; Troiano, N.; Jay, G.; Horowitz, M.; Baron, R. Mice Transgenic for HTLV-I LTR-Tax Exhibit Tax Expression in Bone, Skeletal Alterations, and High Bone Turnover. Virology 1993, 197, 196–204. [Google Scholar] [CrossRef]
- Bieberich, C.J.; King, C.M.; Tinkle, B.T.; Jay, G. A Transgenic Model of Transactivation by the Tax Protein of HTLV-I. Virology 1993, 196, 309–318. [Google Scholar] [CrossRef]
- Grossman, W.J.; Kimata, J.T.; Wong, F.H.; Zutter, M.; Ley, T.J.; Ratner, L. Development of Leukemia in Mice Transgenic for the Tax Gene of Human T-Cell Leukemia Virus Type I. Proc. Natl. Acad. Sci. U. S. A. 1995, 92, 1057–1061. [Google Scholar] [CrossRef]
- Grossman, W.J.; Ratner, L. Cytokine Expression and Tumorigenicity of Large Granular Lymphocytic Leukemia Cells From Mice Transgenic for the Tax Gene of Human T-Cell Leukemia Virus Type I. Blood 1997, 90, 783–794. [Google Scholar] [CrossRef]
- Gao, L.; Deng, H.; Zhao, H.; Hirbe, A.; Harding, J.; Ratner, L.; Weilbaecher, K. HTLV-1 Tax Transgenic Mice Develop Spontaneous Osteolytic Bone Metastases Prevented by Osteoclast Inhibition. Blood 2005, 106, 4294–4302. [Google Scholar] [CrossRef]
- Hall, A.P.; Irvine, J.; Blyth, K.; Cameron, E.R.; Onions, D.E.; Campbell, M.E. Tumours Derived from HTLV-I Tax Transgenic Mice Are Characterized by Enhanced Levels of Apoptosis and Oncogene Expression. J. Pathol. 1998, 186, 209–214. [Google Scholar] [CrossRef]
- Hasegawa, H.; Sawa, H.; Lewis, M.J.; Orba, Y.; Sheehy, N.; Yamamoto, Y.; Ichinohe, T.; Tsunetsugu-Yokota, Y.; Katano, H.; Takahashi, H.; et al. Thymus-Derived Leukemia-Lymphoma in Mice Transgenic for the Tax Gene of Human T-Lymphotropic Virus Type I. Nat. Med. 2006, 12, 466–472. [Google Scholar] [CrossRef]
- Ohsugi, T.; Kumasaka, T.; Okada, S.; Urano, T. The Tax Protein of HTLV-1 Promotes Oncogenesis in Not Only Immature T Cells but Also Mature T Cells. Nat. Med. 2007, 13, 527–528. [Google Scholar] [CrossRef]
- Shinohara, T. [HTLV-I tax mediated activation of cellular genes in transgenic mice]. Hokkaido Igaku Zasshi. 1991, 66, 534–543. [Google Scholar]
- Kim, S.J.; Winokur, T.S.; Lee, H.D.; Danielpour, D.; Kim, K.Y.; Geiser, A.G.; Chen, L.S.; Sporn, M.B.; Roberts, A.B.; Jay, G. Overexpression of Transforming Growth Factor-Beta in Transgenic Mice Carrying the Human T-Cell Lymphotropic Virus Type I Tax Gene. Mol. Cell. Biol. 1991, 11, 5222–5228. [Google Scholar] [CrossRef]
- Iwakura, Y.; Saijo, S.; Kioka, Y.; Nakayama-Yamada, J.; Itagaki, K.; Tosu, M.; Asano, M.; Kanai, Y.; Kakimoto, K. Autoimmunity Induction by Human T Cell Leukemia Virus Type 1 in Transgenic Mice That Develop Chronic Inflammatory Arthropathy Resembling Rheumatoid Arthritis in Humans. J. Immunol. 1995, 155, 1588–1598. [Google Scholar] [CrossRef]
- Portis, T.; Harding, J.C.; Ratner, L. The Contribution of NF-Kappa B Activity to Spontaneous Proliferation and Resistance to Apoptosis in Human T-Cell Leukemia Virus Type 1 Tax-Induced Tumors. Blood 2001, 98, 1200–1208. [Google Scholar] [CrossRef]
- Kitajima, I.; Shinohara, T.; Bilakovics, J.; Brown, D.A.; Xu, X.; Nerenberg, M. Ablation of Transplanted HTLV-I Tax-Transformed Tumors in Mice by Antisense Inhibition of NF-Kappa B. Science 1992, 258, 1792–1795. [Google Scholar] [CrossRef]
- Kwon, H.; Ogle, L.; Benitez, B.; Bohuslav, J.; Montano, M.; Felsher, D.W.; Greene, W.C. Lethal Cutaneous Disease in Transgenic Mice Conditionally Expressing Type I Human T Cell Leukemia Virus Tax. J. Biol. Chem. 2005, 280, 35713–35722. [Google Scholar] [CrossRef]
- Ratner, L.; Portis, T.; Robek, M.; Harding, J.; Grossman, W. Studies of the Immortalizing Activity of HTLV Type 1 Tax, Using an Infectious Molecular Clone and Transgenic Mice. AIDS Res. Hum. Retroviruses 2000, 16, 1647–1651. [Google Scholar] [CrossRef]
- Portis, T.; Grossman, W.J.; Harding, J.C.; Hess, J.L.; Ratner, L. Analysis of P53 Inactivation in a Human T-Cell Leukemia Virus Type 1 Tax Transgenic Mouse Model. J. Virol. 2001, 75, 2185–2193. [Google Scholar] [CrossRef]
- Mitra-Kaushik, S.; Harding, J.; Hess, J.; Schreiber, R.; Ratner, L. Enhanced Tumorigenesis in HTLV-1 Tax-Transgenic Mice Deficient in Interferon-Gamma. Blood 2004, 104, 3305–3311. [Google Scholar] [CrossRef]
- Xu, Z.; Hurchla, M.A.; Deng, H.; Uluçkan, O.; Bu, F.; Berdy, A.; Eagleton, M.C.; Heller, E.A.; Floyd, D.H.; Dirksen, W.P.; et al. Interferon-Gamma Targets Cancer Cells and Osteoclasts to Prevent Tumor-Associated Bone Loss and Bone Metastases. J. Biol. Chem. 2009, 284, 4658–4666. [Google Scholar] [CrossRef]
- Esser, A.K.; Rauch, D.A.; Xiang, J.; Harding, J.C.; Kohart, N.A.; Ross, M.H.; Su, X.; Wu, K.; Huey, D.; Xu, Y.; et al. HTLV-1 Viral Oncogene HBZ Induces Osteolytic Bone Disease in Transgenic Mice. Oncotarget 2017, 8, 69250–69263. [Google Scholar] [CrossRef]
- Yamamoto-Taguchi, N.; Satou, Y.; Miyazato, P.; Ohshima, K.; Nakagawa, M.; Katagiri, K.; Kinashi, T.; Matsuoka, M. HTLV-1 BZIP Factor Induces Inflammation through Labile Foxp3 Expression. PLoS Pathog. 2013, 9, e1003630. [Google Scholar] [CrossRef]
- Mitagami, Y.; Yasunaga, J.-I.; Kinosada, H.; Ohshima, K.; Matsuoka, M. Interferon-γ Promotes Inflammation and Development of T-Cell Lymphoma in HTLV-1 BZIP Factor Transgenic Mice. PLoS Pathog. 2015, 11, e1005120. [Google Scholar] [CrossRef]
- Yasuma, K.; Yasunaga, J.; Takemoto, K.; Sugata, K.; Mitobe, Y.; Takenouchi, N.; Nakagawa, M.; Suzuki, Y.; Matsuoka, M. HTLV-1 BZIP Factor Impairs Anti-Viral Immunity by Inducing Co-Inhibitory Molecule, T Cell Immunoglobulin and ITIM Domain (TIGIT). PLoS Pathog. 2016, 12, e1005372. [Google Scholar] [CrossRef]
- Kinosada, H.; Yasunaga, J.-I.; Shimura, K.; Miyazato, P.; Onishi, C.; Iyoda, T.; Inaba, K.; Matsuoka, M. HTLV-1 BZIP Factor Enhances T-Cell Proliferation by Impeding the Suppressive Signaling of Co-Inhibitory Receptors. PLoS Pathog. 2017, 13, e1006120. [Google Scholar] [CrossRef]
- Higuchi, Y.; Yasunaga, J.-I.; Mitagami, Y.; Tsukamoto, H.; Nakashima, K.; Ohshima, K.; Matsuoka, M. HTLV-1 Induces T Cell Malignancy and Inflammation by Viral Antisense Factor-Mediated Modulation of the Cytokine Signaling. Proc. Natl. Acad. Sci. U. S. A. 2020, 117, 13740–13749. [Google Scholar] [CrossRef]
- Sugata, K.; Yasunaga, J.-I.; Mitobe, Y.; Miura, M.; Miyazato, P.; Kohara, M.; Matsuoka, M. Protective Effect of Cytotoxic T Lymphocytes Targeting HTLV-1 BZIP Factor. Blood 2015, 126, 1095–1105. [Google Scholar] [CrossRef]
- Zhao, T.; Satou, Y.; Matsuoka, M. Development of T Cell Lymphoma in HTLV-1 BZIP Factor and Tax Double Transgenic Mice. Arch. Virol. 2014, 159, 1849–1856. [Google Scholar] [CrossRef]
- Yamazaki, J.; Mizukami, T.; Takizawa, K.; Kuramitsu, M.; Momose, H.; Masumi, A.; Ami, Y.; Hasegawa, H.; Hall, W.W.; Tsujimoto, H.; et al. Identification of Cancer Stem Cells in a Tax-Transgenic (Tax-Tg) Mouse Model of Adult T-Cell Leukemia/Lymphoma. Blood 2009, 114, 2709–2720. [Google Scholar] [CrossRef]
- Kuribayashi, W.; Takizawa, K.; Sugata, K.; Kuramitsu, M.; Momose, H.; Sasaki, E.; Hiradate, Y.; Furuhata, K.; Asada, Y.; Iwama, A.; et al. Impact of the SCF Signaling Pathway on Leukemia Stem Cell-Mediated ATL Initiation and Progression in an HBZ Transgenic Mouse Model. Oncotarget 2016, 7, 51027–51043. [Google Scholar] [CrossRef]
- Bosma, G.C.; Custer, R.P.; Bosma, M.J. A Severe Combined Immunodeficiency Mutation in the Mouse. Nature 1983, 301, 527–530. [Google Scholar] [CrossRef]
- Araki, R.; Fujimori, A.; Hamatani, K.; Mita, K.; Saito, T.; Mori, M.; Fukumura, R.; Morimyo, M.; Muto, M.; Itoh, M.; et al. Nonsense Mutation at Tyr-4046 in the DNA-Dependent Protein Kinase Catalytic Subunit of Severe Combined Immune Deficiency Mice. Proc. Natl. Acad. Sci. U. S. A. 1997, 94, 2438–2443. [Google Scholar] [CrossRef]
- McCune, J.M.; Namikawa, R.; Kaneshima, H.; Shultz, L.D.; Lieberman, M.; Weissman, I.L. The SCID-Hu Mouse: Murine Model for the Analysis of Human Hematolymphoid Differentiation and Function. Science 1988, 241, 1632–1639. [Google Scholar] [CrossRef]
- Mosier, D.E.; Gulizia, R.J.; Baird, S.M.; Wilson, D.B. Transfer of a Functional Human Immune System to Mice with Severe Combined Immunodeficiency. Nature 1988, 335, 256–259. [Google Scholar] [CrossRef]
- Dorshkind, K.; Pollack, S.B.; Bosma, M.J.; Phillips, R.A. Natural Killer (NK) Cells Are Present in Mice with Severe Combined Immunodeficiency (Scid). J. Immunol. 1985, 134, 3798–3801. [Google Scholar]
- Bosma, G.C.; Fried, M.; Custer, R.P.; Carroll, A.; Gibson, D.M.; Bosma, M.J. Evidence of Functional Lymphocytes in Some (Leaky) Scid Mice. J. Exp. Med. 1988, 167, 1016–1033. [Google Scholar] [CrossRef]
- Shultz, L.D.; Schweitzer, P.A.; Christianson, S.W.; Gott, B.; Schweitzer, I.B.; Tennent, B.; McKenna, S.; Mobraaten, L.; Rajan, T. V; Greiner, D.L. Multiple Defects in Innate and Adaptive Immunologic Function in NOD/LtSz-Scid Mice. J. Immunol. 1995, 154, 180–191. [Google Scholar] [CrossRef]
- Hesselton, R.M.; Greiner, D.L.; Mordes, J.P.; Rajan, T. V; Sullivan, J.L.; Shultz, L.D. High Levels of Human Peripheral Blood Mononuclear Cell Engraftment and Enhanced Susceptibility to Human Immunodeficiency Virus Type 1 Infection in NOD/LtSz-Scid/Scid Mice. J. Infect. Dis. 1995, 172, 974–982. [Google Scholar] [CrossRef]
- Ito, M.; Hiramatsu, H.; Kobayashi, K.; Suzue, K.; Kawahata, M.; Hioki, K.; Ueyama, Y.; Koyanagi, Y.; Sugamura, K.; Tsuji, K.; et al. NOD/SCID/Gamma(c)(Null) Mouse: An Excellent Recipient Mouse Model for Engraftment of Human Cells. Blood 2002, 100, 3175–3182. [Google Scholar] [CrossRef]
- Shultz, L.D.; Lyons, B.L.; Burzenski, L.M.; Gott, B.; Chen, X.; Chaleff, S.; Kotb, M.; Gillies, S.D.; King, M.; Mangada, J.; et al. Human Lymphoid and Myeloid Cell Development in NOD/LtSz-Scid IL2R Gamma Null Mice Engrafted with Mobilized Human Hemopoietic Stem Cells. J. Immunol. 2005, 174, 6477–6489. [Google Scholar] [CrossRef]
- Okada, S.; Harada, H.; Ito, T.; Saito, T.; Suzu, S. Early Development of Human Hematopoietic and Acquired Immune Systems in New Born NOD/Scid/Jak3null Mice Intrahepatic Engrafted with Cord Blood-Derived CD34 + Cells. Int. J. Hematol. 2008, 88, 476–482. [Google Scholar] [CrossRef]
- Pearson, T.; Shultz, L.D.; Miller, D.; King, M.; Laning, J.; Fodor, W.; Cuthbert, A.; Burzenski, L.; Gott, B.; Lyons, B.; et al. Non-Obese Diabetic-Recombination Activating Gene-1 (NOD-Rag1 Null) Interleukin (IL)-2 Receptor Common Gamma Chain (IL2r Gamma Null) Null Mice: A Radioresistant Model for Human Lymphohaematopoietic Engraftment. Clin. Exp. Immunol. 2008, 154, 270–284. [Google Scholar] [CrossRef]
- Ishihara, S.; Tachibana, N.; Okayama, A.; Murai, K.; Tsuda, K.; Mueller, N. Successful Graft of HTLV-I-Transformed Human T-Cells (MT-2) in Severe Combined Immunodeficiency Mice Treated with Anti-Asialo GM-1 Antibody. Jpn. J. Cancer Res. 1992, 83, 320–323. [Google Scholar] [CrossRef]
- Ohsugi, T.; Ishibashi, K.; Shingu, M.; Nomura, T. Engraftment of HTLV-I-Transformed Human T-Cell Line into SCID Mice with NK Cell Function. J. Vet. Med. Sci. 1994, 56, 601–603. [Google Scholar] [CrossRef]
- Feuer, G.; Zack, J.A.; Harrington, W.J.J.; Valderama, R.; Rosenblatt, J.D.; Wachsman, W.; Baird, S.M.; Chen, I.S. Establishment of Human T-Cell Leukemia Virus Type I T-Cell Lymphomas in Severe Combined Immunodeficient Mice. Blood 1993, 82, 722–731. [Google Scholar] [CrossRef]
- Kondo, A.; Imada, K.; Hattori, T.; Yamabe, H.; Tanaka, T.; Miyasaka, M.; Okuma, M.; Uchiyama, T. A Model of in Vivo Cell Proliferation of Adult T-Cell Leukemia. Blood 1993, 82, 2501–2509. [Google Scholar] [CrossRef]
- Imada, K.; Takaori-Kondo, A.; Uchiyama, T. [Analysis of in vivo cell proliferation of ATL using SCID mice]. Rinsho. Ketsueki. 1995, 36, 573–577. [Google Scholar]
- Watters, K.M.; Dean, J.; Hasegawa, H.; Sawa, H.; Hall, W.; Sheehy, N. Cytokine and Growth Factor Expression by HTLV-1 Lck-Tax Transgenic Cells in SCID Mice. AIDS Res. Hum. Retroviruses 2010, 26, 593–603. [Google Scholar] [CrossRef]
- Yan, P.; Fu, J.; Qu, Z.; Li, S.; Tanaka, T.; Grusby, M.J.; Xiao, G. PDLIM2 Suppresses Human T-Cell Leukemia Virus Type I Tax-Mediated Tumorigenesis by Targeting Tax into the Nuclear Matrix for Proteasomal Degradation. Blood 2009, 113, 4370–4380. [Google Scholar] [CrossRef]
- Takeda, S.; Maeda, M.; Morikawa, S.; Taniguchi, Y.; Yasunaga, J.-I.; Nosaka, K.; Tanaka, Y.; Matsuoka, M. Genetic and Epigenetic Inactivation of Tax Gene in Adult T-Cell Leukemia Cells. Int. J. cancer 2004, 109, 559–567. [Google Scholar] [CrossRef]
- Arnold, J.; Zimmerman, B.; Li, M.; Lairmore, M.D.; Green, P.L. Human T-Cell Leukemia Virus Type-1 Antisense-Encoded Gene, Hbz, Promotes T-Lymphocyte Proliferation. Blood 2008, 112, 3788–3797. [Google Scholar] [CrossRef]
- Dewan, M.Z.; Terashima, K.; Taruishi, M.; Hasegawa, H.; Ito, M.; Tanaka, Y.; Mori, N.; Sata, T.; Koyanagi, Y.; Maeda, M.; et al. Rapid Tumor Formation of Human T-Cell Leukemia Virus Type 1-Infected Cell Lines in Novel NOD-SCID/Gammac(Null) Mice: Suppression by an Inhibitor against NF-KappaB. J. Virol. 2003, 77, 5286–5294. [Google Scholar] [CrossRef]
- Ohsugi, T.; Horie, R.; Kumasaka, T.; Ishida, A.; Ishida, T.; Yamaguchi, K.; Watanabe, T.; Umezawa, K.; Urano, T. In Vivo Antitumor Activity of the NF-KappaB Inhibitor Dehydroxymethylepoxyquinomicin in a Mouse Model of Adult T-Cell Leukemia. Carcinogenesis 2005, 26, 1382–1388. [Google Scholar] [CrossRef]
- Watanabe, M.; Ohsugi, T.; Shoda, M.; Ishida, T.; Aizawa, S.; Maruyama-Nagai, M.; Utsunomiya, A.; Koga, S.; Yamada, Y.; Kamihira, S.; et al. Dual Targeting of Transformed and Untransformed HTLV-1-Infected T Cells by DHMEQ, a Potent and Selective Inhibitor of NF-KappaB, as a Strategy for Chemoprevention and Therapy of Adult T-Cell Leukemia. Blood 2005, 106, 2462–2471. [Google Scholar] [CrossRef]
- Ohsugi, T.; Kumasaka, T.; Ishida, A.; Ishida, T.; Horie, R.; Watanabe, T.; Umezawa, K.; Yamaguchi, K. In Vitro and in Vivo Antitumor Activity of the NF-KappaB Inhibitor DHMEQ in the Human T-Cell Leukemia Virus Type I-Infected Cell Line, HUT-102. Leuk. Res. 2006, 30, 90–97. [Google Scholar] [CrossRef]
- Ohsugi, T.; Kumasaka, T.; Okada, S.; Ishida, T.; Yamaguchi, K.; Horie, R.; Watanabe, T.; Umezawa, K. Dehydroxymethylepoxyquinomicin (DHMEQ) Therapy Reduces Tumor Formation in Mice Inoculated with Tax-Deficient Adult T-Cell Leukemia-Derived Cell Lines. Cancer Lett. 2007, 257, 206–215. [Google Scholar] [CrossRef]
- Satou, Y.; Nosaka, K.; Koya, Y.; Yasunaga, J.-I.; Toyokuni, S.; Matsuoka, M. Proteasome Inhibitor, Bortezomib, Potently Inhibits the Growth of Adult T-Cell Leukemia Cells Both in Vivo and in Vitro. Leukemia 2004, 18, 1357–1363. [Google Scholar] [CrossRef]
- Shu, S.T.; Nadella, M.V.P.; Dirksen, W.P.; Fernandez, S.A.; Thudi, N.K.; Werbeck, J.L.; Lairmore, M.D.; Rosol, T.J. A Novel Bioluminescent Mouse Model and Effective Therapy for Adult T-Cell Leukemia/Lymphoma. Cancer Res. 2007, 67, 11859–11866. [Google Scholar] [CrossRef]
- Dewan, M.Z.; Uchihara, J.; Terashima, K.; Honda, M.; Sata, T.; Ito, M.; Fujii, N.; Uozumi, K.; Tsukasaki, K.; Tomonaga, M.; et al. Efficient Intervention of Growth and Infiltration of Primary Adult T-Cell Leukemia Cells by an HIV Protease Inhibitor, Ritonavir. Blood 2006, 107, 716–724. [Google Scholar] [CrossRef]
- El Hajj, H.; El-Sabban, M.; Hasegawa, H.; Zaatari, G.; Ablain, J.; Saab, S.T.; Janin, A.; Mahfouz, R.; Nasr, R.; Kfoury, Y.; et al. Therapy-Induced Selective Loss of Leukemia-Initiating Activity in Murine Adult T Cell Leukemia. J. Exp. Med. 2010, 207, 2785–2792. [Google Scholar] [CrossRef]
- Ikebe, E.; Kawaguchi, A.; Tezuka, K.; Taguchi, S.; Hirose, S.; Matsumoto, T.; Mitsui, T.; Senba, K.; Nishizono, A.; Hori, M.; et al. Oral Administration of an HSP90 Inhibitor, 17-DMAG, Intervenes Tumor-Cell Infiltration into Multiple Organs and Improves Survival Period for ATL Model Mice. Blood Cancer J. 2013, 3, e132. [Google Scholar] [CrossRef]
- Ju, W.; Zhang, M.; Petrus, M.; Maeda, M.; Pise-Masison, C.A.; Waldmann, T.A. Combination of 9-Aminoacridine with Campath-1H Provides Effective Therapy for a Murine Model of Adult T-Cell Leukemia. Retrovirology 2014, 11, 43. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, M.; Ju, W.; Waldmann, T.A. Effective Treatment of a Murine Model of Adult T-Cell Leukemia Using Depsipeptide and Its Combination with Unmodified Daclizumab Directed toward CD25. Blood 2009, 113, 1287–1293. [Google Scholar] [CrossRef]
- Hasegawa, H.; Yamada, Y.; Tsukasaki, K.; Mori, N.; Tsuruda, K.; Sasaki, D.; Usui, T.; Osaka, A.; Atogami, S.; Ishikawa, C.; et al. LBH589, a Deacetylase Inhibitor, Induces Apoptosis in Adult T-Cell Leukemia/Lymphoma Cells via Activation of a Novel RAIDD-Caspase-2 Pathway. Leukemia 2011, 25, 575–587. [Google Scholar] [CrossRef]
- Zimmerman, B.; Sargeant, A.; Landes, K.; Fernandez, S.A.; Chen, C.-S.; Lairmore, M.D. Efficacy of Novel Histone Deacetylase Inhibitor, AR42, in a Mouse Model of, Human T-Lymphotropic Virus Type 1 Adult T Cell Lymphoma. Leuk. Res. 2011, 35, 1491–1497. [Google Scholar] [CrossRef]
- Ishitsuka, K.; Kunami, N.; Katsuya, H.; Nogami, R.; Ishikawa, C.; Yotsumoto, F.; Tanji, H.; Mori, N.; Takeshita, M.; Miyamoto, S.; et al. Targeting Bcl-2 Family Proteins in Adult T-Cell Leukemia/Lymphoma: In Vitro and in Vivo Effects of the Novel Bcl-2 Family Inhibitor ABT-737. Cancer Lett. 2012, 317, 218–225. [Google Scholar] [CrossRef]
- Haneji, K.; Matsuda, T.; Tomita, M.; Kawakami, H.; Ohshiro, K.; Uchihara, J.-N.; Masuda, M.; Takasu, N.; Tanaka, Y.; Ohta, T.; et al. Fucoidan Extracted from Cladosiphon Okamuranus Tokida Induces Apoptosis of Human T-Cell Leukemia Virus Type 1-Infected T-Cell Lines and Primary Adult T-Cell Leukemia Cells. Nutr. Cancer 2005, 52, 189–201. [Google Scholar] [CrossRef]
- Fauzi, Y.R.; Nakahata, S.; Chilmi, S.; Ichikawa, T.; Nueangphuet, P.; Yamaguchi, R.; Nakamura, T.; Shimoda, K.; Morishita, K. Antitumor Effects of Chloroquine/Hydroxychloroquine Mediated by Inhibition of the NF-ΚB Signaling Pathway through Abrogation of Autophagic P47 Degradation in Adult T-Cell Leukemia/Lymphoma Cells. PLoS One 2021, 16, e0256320. [Google Scholar] [CrossRef]
- Aikawa, A.; Kozako, T.; Uchida, Y.; Yoshimitsu, M.; Ishitsuka, K.; Ohsugi, T.; Honda, S.-I. Cell Death Induced by Dorsomorphin in Adult T-Cell Leukemia/Lymphoma Is AMPK-Independent. FEBS J. 2020, 287, 4005–4015. [Google Scholar] [CrossRef]
- Ishikawa, C.; Senba, M.; Mori, N. Importin Β1 Regulates Cell Growth and Survival during Adult T Cell Leukemia/Lymphoma Therapy. Invest. New Drugs 2021, 39, 317–329. [Google Scholar] [CrossRef]
- Zhang, M.; Mathews Griner, L.A.; Ju, W.; Duveau, D.Y.; Guha, R.; Petrus, M.N.; Wen, B.; Maeda, M.; Shinn, P.; Ferrer, M.; et al. Selective Targeting of JAK/STAT Signaling Is Potentiated by Bcl-XL Blockade in IL-2-Dependent Adult T-Cell Leukemia. Proc. Natl. Acad. Sci. U. S. A. 2015, 112, 12480–12485. [Google Scholar] [CrossRef]
- Ishikawa, C.; Senba, M.; Mori, N. Anti-Adult T-cell Leukemia/Lymphoma Activity of Cerdulatinib, a Dual SYK/JAK Kinase Inhibitor. Int. J. Oncol. 2018, 53, 1681–1690. [Google Scholar] [CrossRef]
- Ishikawa, C.; Matsuda, T.; Okudaira, T.; Tomita, M.; Kawakami, H.; Tanaka, Y.; Masuda, M.; Ohshiro, K.; Ohta, T.; Mori, N. Bisphosphonate Incadronate Inhibits Growth of Human T-Cell Leukaemia Virus Type I-Infected T-Cell Lines and Primary Adult T-Cell Leukaemia Cells by Interfering with the Mevalonate Pathway. Br. J. Haematol. 2007, 136, 424–432. [Google Scholar] [CrossRef]
- Machijima, Y.; Ishikawa, C.; Sawada, S.; Okudaira, T.; Uchihara, J.; Tanaka, Y.; Taira, N.; Mori, N. Anti-Adult T-Cell Leukemia/Lymphoma Effects of Indole-3-Carbinol. Retrovirology 2009, 6, 7. [Google Scholar] [CrossRef]
- Guimaraes-Correa, A.B.; Crawford, L.B.; Figueiredo, C.R.; Gimenes, K.P.; Pinto, L.A.; Grassi, M.F.R.; Feuer, G.; Travassos, L.R.; Caires, A.C.F.; Rodrigues, E.G.; et al. C7a, a Biphosphinic Cyclopalladated Compound, Efficiently Controls the Development of a Patient-Derived Xenograft Model of Adult T Cell Leukemia/Lymphoma. Viruses 2011, 3, 1041–1058. [Google Scholar] [CrossRef]
- Ishikawa, C.; Jomori, T.; Tanaka, J.; Senba, M.; Mori, N. Peridinin, a Carotenoid, Inhibits Proliferation and Survival of HTLV-1-Infected T-Cell Lines. Int. J. Oncol. 2016, 49, 1713–1721. [Google Scholar] [CrossRef]
- Ishikawa, C.; Senba, M.; Mori, N. Butein Inhibits NF-ΚB, AP-1 and Akt Activation in Adult T-Cell Leukemia/Lymphoma. Int. J. Oncol. 2017, 51, 633–643. [Google Scholar] [CrossRef]
- Fatfat, M.; Fakhoury, I.; Habli, Z.; Mismar, R.; Gali-Muhtasib, H. Thymoquinone Enhances the Anticancer Activity of Doxorubicin against Adult T-Cell Leukemia in Vitro and in Vivo through ROS-Dependent Mechanisms. Life Sci. 2019, 232, 116628. [Google Scholar] [CrossRef]
- Kawaguchi, A.; Orba, Y.; Kimura, T.; Iha, H.; Ogata, M.; Tsuji, T.; Ainai, A.; Sata, T.; Okamoto, T.; Hall, W.W.; et al. Inhibition of the SDF-1alpha-CXCR4 Axis by the CXCR4 Antagonist AMD3100 Suppresses the Migration of Cultured Cells from ATL Patients and Murine Lymphoblastoid Cells from HTLV-I Tax Transgenic Mice. Blood 2009, 114, 2961–2968. [Google Scholar] [CrossRef]
- Tary-Lehmann, M.; Saxon, A.; Lehmann, P. V The Human Immune System in Hu-PBL-SCID Mice. Immunol. Today 1995, 16, 529–533. [Google Scholar] [CrossRef]
- King, M.A.; Covassin, L.; Brehm, M.A.; Racki, W.; Pearson, T.; Leif, J.; Laning, J.; Fodor, W.; Foreman, O.; Burzenski, L.; et al. Human Peripheral Blood Leucocyte Non-Obese Diabetic-Severe Combined Immunodeficiency Interleukin-2 Receptor Gamma Chain Gene Mouse Model of Xenogeneic Graft-versus-Host-like Disease and the Role of Host Major Histocompatibility Complex. Clin. Exp. Immunol. 2009, 157, 104–118. [Google Scholar] [CrossRef]
- Lan, P.; Tonomura, N.; Shimizu, A.; Wang, S.; Yang, Y.-G. Reconstitution of a Functional Human Immune System in Immunodeficient Mice through Combined Human Fetal Thymus/Liver and CD34+ Cell Transplantation. Blood 2006, 108, 487–492. [Google Scholar] [CrossRef]
- Melkus, M.W.; Estes, J.D.; Padgett-Thomas, A.; Gatlin, J.; Denton, P.W.; Othieno, F.A.; Wege, A.K.; Haase, A.T.; Garcia, J.V. Humanized Mice Mount Specific Adaptive and Innate Immune Responses to EBV and TSST-1. Nat. Med. 2006, 12, 1316–1322. [Google Scholar] [CrossRef]
- Tonomura, N.; Habiro, K.; Shimizu, A.; Sykes, M.; Yang, Y.-G. Antigen-Specific Human T-Cell Responses and T Cell-Dependent Production of Human Antibodies in a Humanized Mouse Model. Blood 2008, 111, 4293–4296. [Google Scholar] [CrossRef]
- Danner, R.; Chaudhari, S.N.; Rosenberger, J.; Surls, J.; Richie, T.L.; Brumeanu, T.-D.; Casares, S. Expression of HLA Class II Molecules in Humanized NOD.Rag1KO.IL2RgcKO Mice Is Critical for Development and Function of Human T and B Cells. PLoS One 2011, 6, e19826. [Google Scholar] [CrossRef]
- Suzuki, M.; Takahashi, T.; Katano, I.; Ito, R.; Ito, M.; Harigae, H.; Ishii, N.; Sugamura, K. Induction of Human Humoral Immune Responses in a Novel HLA-DR-Expressing Transgenic NOD/Shi-Scid/Γcnull Mouse. Int. Immunol. 2012, 24, 243–252. [Google Scholar] [CrossRef]
- Strowig, T.; Gurer, C.; Ploss, A.; Liu, Y.-F.; Arrey, F.; Sashihara, J.; Koo, G.; Rice, C.M.; Young, J.W.; Chadburn, A.; et al. Priming of Protective T Cell Responses against Virus-Induced Tumors in Mice with Human Immune System Components. J. Exp. Med. 2009, 206, 1423–1434. [Google Scholar] [CrossRef]
- Shultz, L.D.; Saito, Y.; Najima, Y.; Tanaka, S.; Ochi, T.; Tomizawa, M.; Doi, T.; Sone, A.; Suzuki, N.; Fujiwara, H.; et al. Generation of Functional Human T-Cell Subsets with HLA-Restricted Immune Responses in HLA Class I Expressing NOD/SCID/IL2r Gamma(Null) Humanized Mice. Proc. Natl. Acad. Sci. U. S. A. 2010, 107, 13022–13027. [Google Scholar] [CrossRef]
- Willinger, T.; Rongvaux, A.; Takizawa, H.; Yancopoulos, G.D.; Valenzuela, D.M.; Murphy, A.J.; Auerbach, W.; Eynon, E.E.; Stevens, S.; Manz, M.G.; et al. Human IL-3/GM-CSF Knock-in Mice Support Human Alveolar Macrophage Development and Human Immune Responses in the Lung. Proc. Natl. Acad. Sci. U. S. A. 2011, 108, 2390–2395. [Google Scholar] [CrossRef]
- Billerbeck, E.; Barry, W.T.; Mu, K.; Dorner, M.; Rice, C.M.; Ploss, A. Development of Human CD4+FoxP3+ Regulatory T Cells in Human Stem Cell Factor-, Granulocyte-Macrophage Colony-Stimulating Factor-, and Interleukin-3-Expressing NOD-SCID IL2Rγ(Null) Humanized Mice. Blood 2011, 117, 3076–3086. [Google Scholar] [CrossRef]
- Huntington, N.D.; Legrand, N.; Alves, N.L.; Jaron, B.; Weijer, K.; Plet, A.; Corcuff, E.; Mortier, E.; Jacques, Y.; Spits, H.; et al. IL-15 Trans-Presentation Promotes Human NK Cell Development and Differentiation in Vivo. J. Exp. Med. 2009, 206, 25–34. [Google Scholar] [CrossRef]
- Chen, Q.; Khoury, M.; Chen, J. Expression of Human Cytokines Dramatically Improves Reconstitution of Specific Human-Blood Lineage Cells in Humanized Mice. Proc. Natl. Acad. Sci. U. S. A. 2009, 106, 21783–21788. [Google Scholar] [CrossRef]
- Katano, I.; Takahashi, T.; Ito, R.; Kamisako, T.; Mizusawa, T.; Ka, Y.; Ogura, T.; Suemizu, H.; Kawakami, Y.; Ito, M. Predominant Development of Mature and Functional Human NK Cells in a Novel Human IL-2-Producing Transgenic NOG Mouse. J. Immunol. 2015, 194, 3513–3525. [Google Scholar] [CrossRef]
- Takahashi, T.; Katano, I.; Ito, R.; Goto, M.; Abe, H.; Mizuno, S.; Kawai, K.; Sugiyama, F.; Ito, M. Enhanced Antibody Responses in a Novel NOG Transgenic Mouse with Restored Lymph Node Organogenesis. Front. Immunol. 2017, 8, 2017. [Google Scholar] [CrossRef]
- Miyazato, P.; Yasunaga, J.; Taniguchi, Y.; Koyanagi, Y.; Mitsuya, H.; Matsuoka, M. De Novo Human T-Cell Leukemia Virus Type 1 Infection of Human Lymphocytes in NOD-SCID, Common Gamma-Chain Knockout Mice. J. Virol. 2006, 80, 10683–10691. [Google Scholar] [CrossRef]
- Takajo, I.; Umeki, K.; Morishita, K.; Yamamoto, I.; Kubuki, Y.; Hatakeyama, K.; Kataoka, H.; Okayama, A. Engraftment of Peripheral Blood Mononuclear Cells from Human T-Lymphotropic Virus Type 1 Carriers in NOD/SCID/Gammac(Null) (NOG) Mice. Int. J. cancer 2007, 121, 2205–2211. [Google Scholar] [CrossRef]
- Villaudy, J.; Wencker, M.; Gadot, N.; Gillet, N.A.; Scoazec, J.-Y.; Gazzolo, L.; Manz, M.G.; Bangham, C.R.M.; Dodon, M.D. HTLV-1 Propels Thymic Human T Cell Development in “Human Immune System” Rag2−/− Gamma C−/− Mice. PLoS Pathog. 2011, 7, e1002231. [Google Scholar] [CrossRef]
- Tezuka, K.; Xun, R.; Tei, M.; Ueno, T.; Tanaka, M.; Takenouchi, N.; Fujisawa, J. An Animal Model of Adult T-Cell Leukemia: Humanized Mice with HTLV-1-Specific Immunity. Blood 2014, 123, 346–355. [Google Scholar] [CrossRef]
- Espíndola, O. de M.; Siteur-van Rijnstra, E.; Frankin, E.; Weijer, K.; van der Velden, Y.U.; Berkhout, B.; Blom, B.; Villaudy, J. Early Effects of HTLV-1 Infection on the Activation, Exhaustion, and Differentiation of T-Cells in Humanized NSG Mice. Cells 2021, 10. [Google Scholar] [CrossRef]
- Xiang, J.; Rauch, D.A.; Huey, D.D.; Panfil, A.R.; Cheng, X.; Esser, A.K.; Su, X.; Harding, J.C.; Xu, Y.; Fox, G.C.; et al. HTLV-1 Viral Oncogene HBZ Drives Bone Destruction in Adult T Cell Leukemia. JCI insight 2019, 4. [Google Scholar] [CrossRef]
- Hiyoshi, M.; Okuma, K.; Tateyama, S.; Takizawa, K.; Saito, M.; Kuramitsu, M.; Araki, K.; Morishita, K.; Okada, S.; Yamamoto, N.; et al. Furin-Dependent CCL17-Fused Recombinant Toxin Controls HTLV-1 Infection by Targeting and Eliminating Infected CCR4-Expressing Cells in Vitro and in Vivo. Retrovirology 2015, 12, 73. [Google Scholar] [CrossRef]
- Inocencio, N.M.; Moehring, J.M.; Moehring, T.J. Furin Activates Pseudomonas Exotoxin A by Specific Cleavage in Vivo and in Vitro. J. Biol. Chem. 1994, 269, 31831–31835. [Google Scholar] [CrossRef]
- Percher, F.; Curis, C.; Pérès, E.; Artesi, M.; Rosewick, N.; Jeannin, P.; Gessain, A.; Gout, O.; Mahieux, R.; Ceccaldi, P.-E.; et al. HTLV-1-Induced Leukotriene B4 Secretion by T Cells Promotes T Cell Recruitment and Virus Propagation. Nat. Commun. 2017, 8, 15890. [Google Scholar] [CrossRef]
- Tezuka, K.; Okuma, K.; Kuramitsu, M.; Matsuoka, S.; Tanaka, R.; Tanaka, Y.; Hamaguchi, I. Control of Human T-Cell Leukemia Virus Type 1 (HTLV-1) Infection by Eliminating Envelope Protein-Positive Cells with Recombinant Vesicular Stomatitis Viruses Encoding HTLV-1 Primary Receptor. J. Virol. 2018, 92. [Google Scholar] [CrossRef]
- Karkhanis, V.; Hu, Y.-J.; Baiocchi, R.A.; Imbalzano, A.N.; Sif, S. Versatility of PRMT5-Induced Methylation in Growth Control and Development. Trends Biochem. Sci. 2011, 36, 633–641. [Google Scholar] [CrossRef]
- Ernzen, K.; Melvin, C.; Yu, L.; Phelps, C.; Niewiesk, S.; Green, P.L.; Panfil, A.R. The PRMT5 Inhibitor EPZ015666 Is Effective against HTLV-1-Transformed T-Cell Lines in Vitro and in Vivo. Front. Microbiol. 2023, 14, 1101544. [Google Scholar] [CrossRef]
- Pérès, E.; Blin, J.; Ricci, E.P.; Artesi, M.; Hahaut, V.; Van den Broeke, A.; Corbin, A.; Gazzolo, L.; Ratner, L.; Jalinot, P.; et al. PDZ Domain-Binding Motif of Tax Sustains T-Cell Proliferation in HTLV-1-Infected Humanized Mice. PLoS Pathog. 2018, 14, e1006933. [Google Scholar] [CrossRef]
- Maksimova, V.; Smith, S.; Seth, J.; Phelps, C.; Niewiesk, S.; Satou, Y.; Green, P.L.; Panfil, A.R. HTLV-1 Intragenic Viral Enhancer Influences Immortalization Phenotype in Vitro, but Is Dispensable for Persistence and Disease Development in Animal Models. Front. Immunol. 2022, 13, 954077. [Google Scholar] [CrossRef]
Figure 1.
Generation of human immune system mouse models.
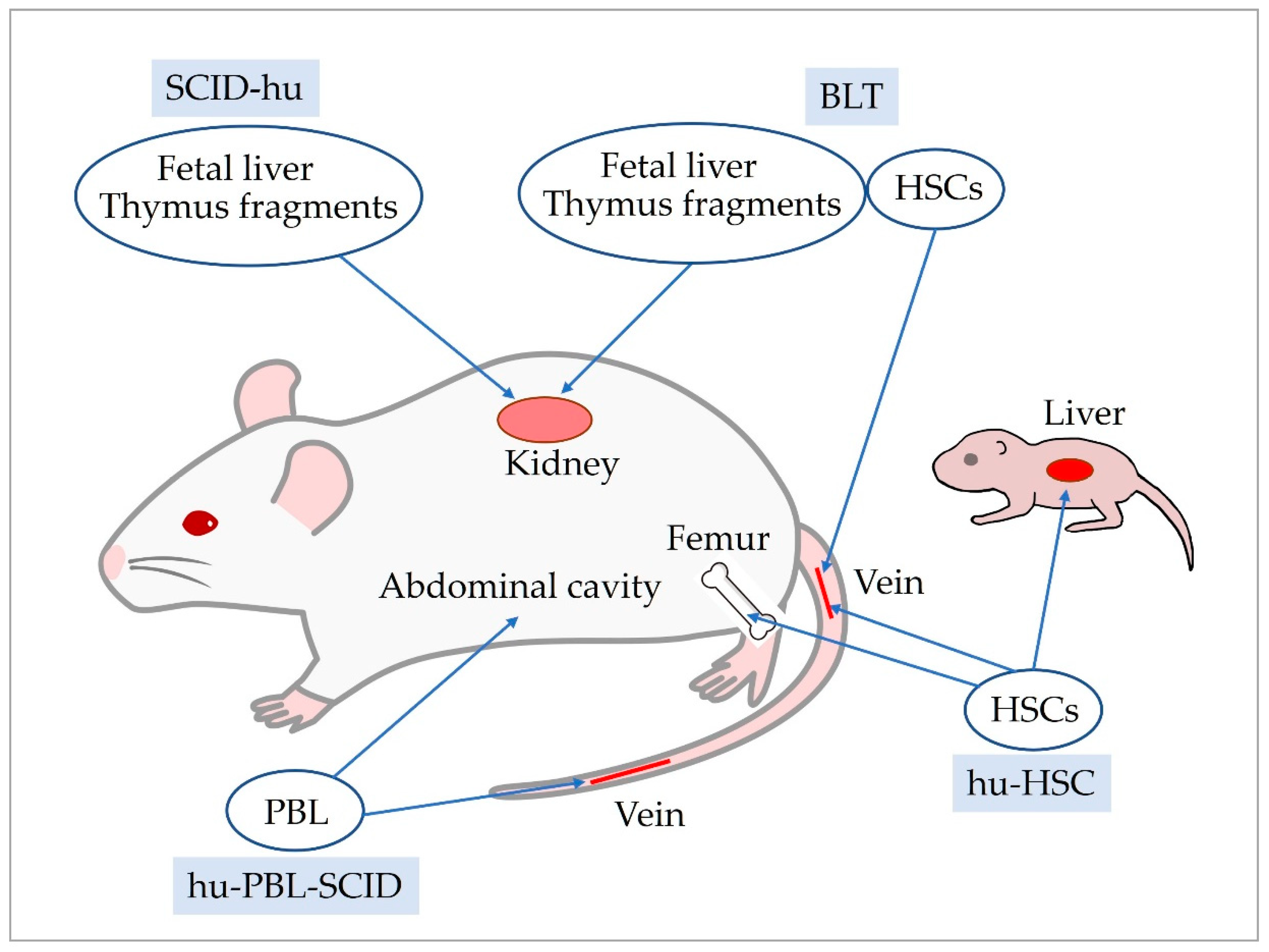

Table 1.
Characteristics of severely immunodeficient mice.
| Strain | Genotype | Characteristics | Limitation | Reference |
|---|---|---|---|---|
| SCID | scid | T cell and B cell defect | T cell and B cell leakage Radiosensitive |
[59] |
| NOD/SCID | nod/scid | T cell and B cell defect Decreased NK cell, DC and macrophage activity Complement deficiency |
T cell and B cell leakage Radiosensitive Spontaneous lymphoma |
[65] |
| NOG | nod/scid/IL-2Rγnull | T cell, B cell and NK cell defect Decreased DC and macrophage activity Complement deficiency |
Radiosensitive | [67] |
| NSG | nod/scid/IL-2Rγnull | T cell, B cell and NK cell defect Decreased DC and macrophage activity Complement deficiency |
Radiosensitive | [68] |
| NOJ | nod/scid/jak-3null | T cell, B cell and NK cell defect Decreased DC and macrophage activity Complement deficiency |
Radiosensitive | [69] |
| NRG | nod/rag-1null/IL-2Rγnull | T cell, B cell and NK cell defect Decreased DC and macrophage activity Complement deficiency |
- | [70] |
Table 2.
ATL therapeutic candidates evaluated in severely immunodeficient mice.
| Drug name | Target | Mouse | Efficacy | Reference |
|---|---|---|---|---|
| Bay 11-7082 | NF-κB inhibitor | NOG mouse | Inhibits tumor growth and invasion | [80] |
| DHMEQ | NF-κB inhibitor | NK(-)SCID mouse SCID mouse |
Inhibits tumor growth and invasion | [81,82,83,84] |
| Ritonavir | HIV protease inhibitor | NOG mouse | Inhibits tumor growth and invasion | [87] |
| Fucoidan | Survivin inhibitor | SCID mouse | Partially inhibits tumor growth | [95] |
| Incadronate | Mevalonate pathway inhibitor | SCID mouse | Reduces tumor formation | [101] |
| PS-341 | Proteasome inhibitor | SCID mouse | Inhibits tumor growth | [85] |
| PS-341 Zoledronic acid |
Proteasome inhibitor Osteoclast inhibitor |
NOD/SCID mouse | Inhibits tumor growth | [86] |
| 9-aminoacridine (9AA) Campath-1H |
Increase p53 transcription activity NF-κB activation inhibitor Humanized anti-CD52 antibody |
NOD/SCID mouse | Inhibits tumor growth Extends survival |
[90] |
| Depsipeptide Daclizumab |
HDAC inhibitor Anti-IL-2Rα antibody |
NOD/SCID mouse | Extends survival | [91] |
| LBH589 | HDAC inhibitor | SCID mouse | Induces tumor cell apoptosis Extends survival |
[92] |
| AR-42 | HDAC inhibitor | NOD/SCID mouse | Extends survival | [93] |
| ABT-737 | Bcl-2, Bcl-X(L), and Bcl-w inhibitor | SCID mouse | Inhibits tumor growth | [94] |
| 17-DMAG | HSP90 inhibitor | NOG mouse | Inhibits tumor invasion Extends survival |
[89] |
| As(2)O(3) IFN-α |
Proteolysis of Tax Antiviral |
SCID mouse | Inhibits tumor cell immortality | [88] |
| C7a | Antitumor effect | NSG mouse | Extends survival | [103] |
| Indole-3-carbinol | Antitumor effect | SCID mouse | Inhibits tumor growth | [102] |
| AMD3100 | CXCR4 antagonist | SCID mouse | Inhibits tumor cell infiltration into liver and lung tissue | [107] |
| Chloroquine Hydroxychloroquine |
Autophagy inhibitor | NOG mouse | Inhibits tumor growth Extends survival |
[96] |
| Dorsomorphin | AMPK inhibitor | NOD/SCID mouse | Inhibits tumor growth | [97] |
| Ivermectin | IPOα/β1 inhibitor | SCID mouse | Inhibits tumor growth | [98] |
| Ruxolitinib Navitoclax |
JAK inhibitor Bcl-2/Bcl-xL inhibitor |
NSG mouse | Inhibits tumor growth | [99] |
| Cerdulatinib | Dual SYK/JAK inhibitor | SCID mouse | Inhibits tumor growth | [100] |
| Peridinin | Antitumor effect | SCID mouse | Inhibits tumor growth | [104] |
| Butein | Antitumor effect | SCID mouse | Inhibits tumor growth | [105] |
| Thymoquinone Doxorubicin |
Antitumor effect Anticancer drug |
NOD/SCID mouse | Inhibits tumor growth | [106] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



