
Submitted:
26 June 2023
Posted:
26 June 2023
You are already at the latest version
Abstract
The Botryosphaeriales order are best known for the diseases they cause in woody plants, as primary pathogens or latent pathogens residing in the woody tissue of asymptomatic hosts. In the first instance, Botryosphaeriales species have been identified in Venezuela using morphological de-scriptions in the 80's and 90's, and later, the mid-2000s using molecular techniques. The mor-phological descriptions of the asexual morphs were initially used for the identification of Botry-osphaeriales genera and species. Lasiodiplodia spp., (as L. theobromae) was the most isolated fungus in Venezuela within the Botryosphaeriales and has been found in more than 50% of the hosts in native and non-native plants, followed by Diplodia, Dothiorella, Fusicoccum, Lasiodiplodia, Micro-diplodia, Macrophomina, Neofusicoccum, Sphaeropsis, and Botryosphaeria, considered all of them cosmopolitan group. With molecular studies, that included DNA sequence data from multiple genes, such as the internal transcribed spacer of rDNA (ITS), translation elongation factor-1α (tef1), and β-tubulin (btub) used on the fungi isolated from woody plants, mainly trees or forest species, resulted in the presence of two families within the Botryosphaeriales order for Venezuela. Botryosphaeriaceae family with the genera: Botryosphaeria, Cophinforma, Diplodia, Lasiodiplodia and Neofusicoccum, and the Pseudofusicoccumaceae family that includes the genus Pseudofusicoccum. In Botryosphaeriaceae family was again the Lasiodiplodia genus the most predominant in most hosts, and the specie L. theobromae the most isolated in native and non-native plants; Botryosphaeria dothidea, Cophinforma atrovirens, Diplodia scrobiculata (syn. Diplodia guayanensis), Lasiodiplodia brasi-liensis, L. crassispora, L. pseudotheobromae, Neofusicoccum arbuti (syn. N. andinum), N. parvum, and N. ribis are cosmopolitan species, and they were isolated from native and non-native plants; while Pseudofusicoccum stromaticum was found in plantations non-native of Acacia mangium, E. urophylla x E. grandis, Eucalyptus urophylla, and reported exclusively in South America; Lasiodiplodia venezue-lensis has only been reported in Venezuela, from native and non-native plants. The presence, distribution, diversity, and symptoms of these fungi, mainly of the new genus, new species, and reports found in Venezuela and other parts of the world, were also reviewed.
Keywords:
Botryosphaeriaceae
; DNA sequence
; Forest
; Pseudofusicoccumaceae
; Fungal Taxonomy
1. Introduction
Forest ecosystems are a natural resource of great importance to humanity, since many people depend on them for their survival, in addition to other benefits such as human and environmental health, carbon sequestration, and genetic resources that underpin important wood and wood products-based industries [1]. At present, the health of forests, both natural and managed, is more heavily threatened, and these threats arise from direct and indirect anthropogenic influences on fungal pathogens, and insect pests [1,2]. Plantations in the tropics (planted forests of a single species) are usually of non-native species, such as the genera of Pinus, Eucalyptus, and Acacia, the main forest species planted in Venezuela.
Non-native trees in plantations are in part successful because they have been separated from their natural enemies, but when plantation trees are reunited with their coevolved pests, which may be introduced accidentally, or when they encounter novel pests to which they have no resistance, substantial damage or loss can ensue [3]. The longer non-native trees are planted in an area, the more threatened they become by native pests. Where the trees are of native species, they can be vulnerable to introduced pests. But the relative species uniformity of monoculture stands in intensively managed native plantation forests can make them especially susceptible to the many native pests occurring in the surrounding natural forests [4,5].
An example of an epidemic of native pathogens moving onto an exotic species is provided by the shoot pathogen Gremmeniella abietina (Lagerberg) Morelet, endemic and not particularly damaging on Scots pine in Sweden but causing widespread destruction of Swedish plantations of the extensively planted exotic lodgepole pine [6].
The Botryosphaeriales contains numerous fungal species that occur as saprophytes, parasites, or endophytes on a diverse range of plant hosts [7,8], as well as opportunistic pathogens of woody plants, especially when host plants are stressed [9]. Different species within of order Botryosphaeriales are well known pathogens on forest trees and other woody plants associated with branch and trunk cankers, dieback, decline and mortality, and represent a growing threat to forest ecosystems worldwide [7,8]. An ecological and biological characteristic of the species in the Botryosphaeriales order is the lack of host specificity able to colonize and cause disease in diverse native and introduced plant hosts [7,10].
Different Botryosphaeriales genera can infect multiple hosts, increases the threat that they pose as potential economic and ecological important pathogens of native and cultivated trees around the world. Examples of inter-host exchanges of the Botryosphaeriales, and that include those amongst and between native and non-native trees, we have Botryosphaeriales species have moved between trees in native stands of Eucalyptus (Myrtaceae) and adjacent plantations of these trees [11], between native waterberry trees (Syzygium cordatum; Myrtaceae) and related eucalypt plantations (Myrtaceae) [12], from Pinus resinosa windbreaks to pine nurseries [13], among various tree hosts in the Casuarinaceae, Cupressaceae, Fabaceae, Myrtaceae, Proteaceae, Santalaceae [14], and among native Terminalia spp. (Combretaceae) and between these trees and Theobroma cacao (Malvaceae) [15]. The epidemiology of Botryosphaeriales species is complex. These fungi can be monocyclic or oligocyclic pathogens that cause polyetic epidemics. As monocyclic pathogens, they complete one disease cycle, or even part of one, in one season. Depending on the weather conditions, these species can be oligocyclic pathogens, i.e., polycyclic pathogens with a few (two or three) disease cycles per season [16]. In Venezuela, the Botryosphaeriales species are polycyclic since there are no marked seasons in the tropics as in the temperate regions. In the tropic, high temperatures and humidity are present almost all year round, therefore, these fungi will present several disease cycles and produce constant inoculum or spores throughout the year.
Species identification in Botryosphaeriales has been largely based on the asexual morphs due to the lack of diversity among sexual morph features within this order and the difficulty of finding the sexual morphs in nature or obtaining them under laboratory conditions [17,18]. Different species within the same genera of Botryosphaeriales frequently possess overlapping morphological features [19] that can cause confusion in their accurate identification. In recent decades, several researchers began using identification techniques based on DNA sequencing and phylogenetic analyses to resolve the taxonomic problems associated with overlapping morphological characteristics among the species asexual morphs within Botryosphaeriales genera [17,18,20,21,22]. The phylogenetic analyses of DNA sequence data have significantly impacted all aspects of the systematics and taxonomy of the Botryosphaeriales, including a redefinition of families and genera, identification of new species, cryptic species, and more recently hybrids [23]. Crous et al. [24] defined all genera in the Botryosphaeriales based predominantly on phylogenetic inference and characteristics of their asexual morphs, and without morphological evidence of a sexual morph. In various cases, genera were thus established in the family based on asexual names.
The Botryosphaeriales order has undergone changes in its systematics, mainly at the family level. A total of nine families have been included in the last 5 years within the Botryosphaeriales order, based on phylogenetic, morphological, and ecological differences [23]. These families are: Aplosporellaceae Slippers et al. 2013 [25], Botryosphaeriaceae Theiss. & Syd., 1918 [24], Endomelanopsisaceae TaoYang & Crous, 2016 [26], Melanopsaceae Phillips et al. 2013 [25], Phyllostictaceae Fr., 1849 [27], Planistromellaceae M.E. Barr, 1996 [28], Pseudofusicoccumaceae Tao Yang & Crous, 2016 [26], Saccharataceae Slippers et al. 2013 [25] and Septorioideaceae Wyka & Broders, 2016 [29], being the Botryosphaeriaceae family with the largest number of genera within it.
The purpose of this review is to update all relevant information on morphological descriptions and DNA sequencing data on the Botryoshaeriales fungi that produce different diseases on cultivated and wild plants, as well as their distribution and diversity on woody plants in Venezuela.
2. Genera and species of Botryosphaeriales identified with morphological descriptions in Venezuela
Few general morphological features of within Botryosphaeriales species have been reported in agricultural crops, forest plantations, and natural forests in Venezuela, where the taxonomic identification and associated reports have been initially based on morphological descriptions of the asexual morph. Such morphological descriptions are frequently based on 1) conidial features, such as septation, presence/absence of pigmentation, and wall thickness, and 2) presence/absence of conidiophores, conidiogenous cells, and paraphyses in the conidiomata [30,31,32,33,34,35,36,37].
Lasiodiplodia Ellis & Everh., species are well-known and widespread plant pathogens, occurring mostly in tropical and subtropical regions [38]. Lasiodiplodia theobromae (Pat.) Griffon & Maubl., has been widely reported and commonly occurs on different crops in Venezuela [39]. In a published list of plant diseases in Venezuela [39], L. theobromae was the common fungal pathogen. This list from Urtiaga [39] was updated using website records of fungi from 1998-2001 with specimens from the fungal collection of the Simon Bolivar University, Caracas-Venezuela [40], together with reports from Mohali and other authors during the 1990s through the mid-2000s (Table 1). In addition to Lasiodiplodia, other reported genera include Diplodia Fr., Dothiorella Sacc., Botryosphaeria Ces. & De Not., Microdiplodia Allesch., and Macrophomina Petr. [39,40]. In addition to two Neofusicoccum Crous, Slippers & A.J.L. Phillips species isolated from M. indica in 2012 and 2016, where identified through their morphological descriptions [41,42] (Table 1).
In Venezuela, at least eight genera of the Botryosphaeriales order within two families can be differentiated through the asexual morph. Seven genera belong to Botryosphaeriaceae family, five with dark-conidia when mature age: Diplodia Fr., Dothiorella Sacc., Lasiodiplodia Ellis & Everh., Macrophomina Petr., Sphaeropsis Sacc., and two genera with hyaline conidia: Cophinforma Doilom, J.K. Liu & K.D. Hyde and Neofusicoccum Crous, Slippers & A.J.L. Phillips; and one genus in the Pseudofusicoccumaceae family, Pseudofusicoccum Mohali, Slippers & M.J. Wingf., with hyaline-conidia surrounded by a persistent mucous sheath (Table 2).

Table 1.
Different genera and species within Botryosphaeriales identified by their asexual morph in Venezuela.
Table 1.
Different genera and species within Botryosphaeriales identified by their asexual morph in Venezuela.
| Fungi | Host | Place | Reference |
|---|---|---|---|
| Diplodia Fr. | Ceiba pentandra (L.) Gaertn-old leaves | Buena vista, Lara state | [39] |
| Diplodi a sp. | Cassia L.- root | - | [40] |
| Diplodia ochromae Pat. | Ochroma lagopus Sw. -trunk | - | [40] |
| Diplodia mutila Fr. Apud Mont. |
Pinus caribaea morelet var. hondurensis (Barr. and Golf.)- blue stain on wood |
Chaguaramas, Anzoátegui state | [36] |
| Dothiorella Sacc. | Delonix regia (Bojer ex Hook) Raf. -branches | El Tocuyo, Lara state | [39] |
| Dothiorella sp. | Psidium guajava L.-fruit rot | Merida and Zulia states | [33] |
| Dothiorella dothidea (= Botryosphaeria dothidea) | Prunus pérsica (L.) Batsch- brown rot of fruits | El Arenal, Merida state | [31] |
| Fusicoccum Corda | Echinodorus berteroi (Spreng) Fassett-leaves | Guanare, Portuguesa state | [39] |
| Lasiodiplodia theobromae (Pat.) Griffon & Maubl. | Pachystachys lutea Nees-branches | Barquisimeto, Lara state | [39] |
| L. theobromae | Anacardium occidentale L- terminal branch death | Barquisimeto, Lara state | [39] |
| L. theobromae | Mangifera indica L-branches and seeds | La Calzada de Páez, Barinas state | [39] |
| L. theobromae | Annona reticulata L.-old leaves | Barquisimeto, Lara state | [39] |
| L. theobromae | Catharanthus roseus (L.) G. Don-Leaves | Barquisimeto, Lara state | [39] |
| L. theobromae | Crescentia cujete L.- branches and leaves | Wide distribution in Venezuela | [39] |
| L. theobromae | Citrullus lanatus (Thunb.) Matsum. & Nakai- fruits rot and branches | La Miel, Lara state | [39] |
| L. theobromae | Juniperus lucayana Britton- twigs | Barquisimeto, Lara state | [39] |
| L. theobromae | Curatella Americana L.-old leaves | La Calzada de Páez, Barinas state | [39] |
| L. theobromae | Codiaeum variegatum (L.) Blume var. pictum (Lodd.) Muell | - | [39] |
| L. theobromae | Hura crepitans L.-old leaves | La Calzada de Páez, Barinas state | [39] |
| L. theobromae | Manihot esculenta Crantz-branches | Urachiche, Yaracuy state | [39] |
| Lasiodiplodia theobromae | Duranta repens L.-branches | Ureña, Táchira | [39] |
| L. theobromae | Arachis hypogaea L.-root | Buría Londres, Lara state | [39] |
| L. theobromae | Phaseolus lunatus L.-branches | Sabana de Parra, Yaracuy state | [39] |
| L. theobromae | Sansevieria trifasciata Prain.-old leaves | Barquisimeto, Lara state | [39] |
| L. theobromae | Cedrela odorata L.-branches | Chivacoa, Yaracuy state | [39] |
| L. theobromae | Cecropia peltata L.-branches | Chivacoa, Yaracuy | [39] |
| L. theobromae | Ficus pumila L.-old leaves and galls on the leaves | Barquisimeto, Lara state | [39] |
| L. theobromae | Maxillaria Ruiz & Pavon-old leaves | Duaca, Lara | [39] |
| L. theobromae | Passiflora edulis Sims. form flavicarpa Degener | El Eneal, Lara | [39] |
| L. theobromae | Salix babylonica L.- black root rot | Barquisimeto, Lara state | [39] |
| L. theobromae | Pachystachys lutea Nees-branches | Barquisimeto, Lara state | [39] |
| L. theobromae | Cajanus indicus Spreng.-branches | Lara state | [39] |
| L. theobromae | Duranta repens L.-branches | Tachira state | [39] |
| L. theobromae | Theobroma cacao L. | - | [40] |
| L. theobromae | Vinca rosea L.-leaf and branch | Lara state | [39] |
| L. theobromae | Persea Americana Mill.-fruits | Yaracuy state | [41] |
| L. theobromae | Citrus latifolia Tanaka-fruits | Yaracuy state | [41] |
| L. theobromae | Citrus sinensis (L.) Osbeck-fruits | Yaracuy state | [41] |
|
Lasiodiplodia theobromae |
C. sinensis-Lesion and Gummosis on the branches | Caño Amarillo, Tachira state | [30] |
| L. theobromae |
Citrus aurantiifolia-Lesion and Gummosis on the branches |
Caño Amarillo, Tachira state | [30] |
| L. theobromae |
Passiflora edulis Sims f. flavicarpa-Dieback on the branches |
South of Maracaibo Lake, Zulia and Merida states |
[32] |
| L. theobromae | Pinus caribaea var. hondurensis-blue stain on wood | Uverito plantation and Uverito sawmill, Monagas state | [123] |
| L. theobromae |
Azadirachta indica A. Juss-blue stain on wood |
Cojedes state | [37] |
| L. theobromae | Pinus oocarpa Schiede ex Schltdl | Merida state | [37] |
| L. theobromae | Mangifera indica-branches dieback | Maracay (INIA-CENIAP), Aragua state | [42] |
| Microdiplodia buddleiae Gucevicz | Opuntia caracasana Salm.-spot leaves | Humocaro Bajo, Lara state | [39] |
| Macrophomina phaseolina (Tassi) Goidanich | Begonia sp.- spot on the leaf | Barquisimeto, Lara state | [39] |
| Macrophomina phaseolina | Calendula officinalis L.-stem and inflorescence | Barinas state | [39] |
| M. phaseolina | Ipomoea batata (L.) Lam.-stolons at the roots | Siquisique, Lara state | [39] |
| M. phaseolina | Phaseolus vulgaris L.-stem and basal rot | Moroturo, Lara state | [39] |
| M. phaseolina | Glycine Willd. | - | [40] |
| M. phaseolina | Gossypium L. | - | [40] |
| M. phaseolina | Ipomoea L. | - | [40] |
| M. phaseolina | Nicotiana L. | - | [40] |
| M. phaseolina | Phaseolus L. | - | [40] |
| M. phaseolina | Psidium guajava L.-Fruits | - | [40] |
| M. phaseolina | Solanum melongena L. | - | [40] |
| M. phaseolina | Vigna Savi | - | [40] |
| Neofusicoccum mangiferae (Syd. & P. Syd.) Crous, Slippers & A.J.L. Phillips | Mangifera indica- death of branches | Maracay (INIA-CENIAP), Aragua state | [42] |
| Neofusicoccum parvum (Pennycook & Samuels) Crous, Slippers & A.J.L. Phillips | Mangifera indica- death of branches | Maracay (INIA-CENIAP), Aragua state | [42] |
| Sphaeropsis Sacc. | Cecropia peltata L.-branch and trunk knots | Reserva Forestal de Ticoporo, Mirí, Barinas state | [39] |
| Sphaeropsis sp. | Phthirusa paniculata (Kunth) J.F.Macbr.-leaf | Lara state | [39] |
| Sphaeropsis palmarum Cooke | Cocos nucifera L.-old leaves | Cumanacoa, Sucre state | [39] |
| Sphaeropsis sapinea (Fr.) Dyko & B. Sutton | Pinus caribaea Morelet- Chlorosis in the needles and discoloration lesions on the stem | Nirgua, Yaracuy state | [35] |
| S. sapinea | Pinus oocarpa Schiede-blue stain on wood | Andes region (1600 meters above sea level), Merida state |
[37] |
| Sphaeropsis sapinea | Pinus caribaea var. hondurensis-Shoot blight, dieback and canker on trunks, branches, and roots (Plantations), and death at the tips of the needles (seedlings in nurseries) | Uverito (Monagas state), and Coloradito y Los Hachos (Anzoátegui state) | [34] |
| Sphaeropsis tumefaciens Hedges | Citrus L.-gall | - | [40] |
| Botryosphaeria festucae (Lib.) Arx & E. Müll | Zea mays L.-bract, leaf, and seed | - | [40] |
| Botryosphaeria dothidea (Moug. ex Fr.) Ces. & De Not | Compositae-stem | Aragua state | [40] |
| Botryosphaeria ribis Grossenb. & Duggar | Rosa canina L.-branch | Lara state | [39] |
Table 2.
Morphological differentiation between the Botryosphaeriaceae genera and a genus in Pseudofusicoccumaceae both belonging to the order Botryosphaeriales found in Venezuela.
Table 2.
Morphological differentiation between the Botryosphaeriaceae genera and a genus in Pseudofusicoccumaceae both belonging to the order Botryosphaeriales found in Venezuela.
| Genera | Conidia | Conidiomata | Conidiophores | Conidiogenesis cells | Paraphyses |
|---|---|---|---|---|---|
| Cophinforma Doilom, J.K. Liu & K.D. Hyde | Hyaline, thin walled, unicellular, aseptate, rarely becoming septate, mostly fusoid to ellipsoidal. Most conidia longer than 30 μm | Material pycnidial, superficial, multilocular, dark brown to black, eustromatic | Absent | Enteroblastic, hyaline, cylindrical | Absent |
| Diplodia Fr. | Initially hyaline, aseptate, thick-walled, becoming 1–septate only rarely becoming 2-septate, pale transluscent brown after discharge from the pycnidia. Some species the conidia become pigmented while still enclosed in the conidioma and these species the conidia rarely become septate. | Pycnidial, ostiolate, formed in uni- or multiloculate stromata | When present: hyaline,simple, occasionally septate, rarely branched, cylindrical, | Holoblastic, hyaline, cylindrical | Absent |
| Dothiorella Sacc. | Initially hyaline, becoming dark brown and one-euseptate within the pycnidial cavity, ellipsoid to ovoid, thick-walled, externally smooth, or striate, internally verruculose | Stromatic, ostiolate, individual or in loose clusters of up to 10 conidiomata, immersed, breaking through the bark when mature. |
Absent | Holoblastic, hyaline, smooth-walled, cylindrical | Absent |
| Lasiodiplodia Ellis & Everh. | Hyaline when young, later becoming medianly 1-euseptate, dark brown with longitudinal striations, thick-walled, oblong to ellipsoid, straight, broadly rounded at the apex, base truncate | Stromatic, immersed or superficial, separate, or aggregated and confluent, globose, dark brown, uni- or multilocular | Often reduced to conidiogenous cells, if present hyaline, simple, sometimes septate, rarely branched | Holoblastic, hyaline, smooth, cylindrical to subobpyriform, discrete, determinate, or indeterminate | Present |
| Macrophomina Petr. | Aseptate, obtuse at each end, straight, cylindrical to fusiform, thin-walled, smooth, guttulate, enclosed in mucoid sheath. Immature conidia hyaline, mature conidia becoming medium to dark brown. | Pycnidial, stromatic, dark brown to black, solitary, or gregarious | Reduced to conidiogenous cells | Enteroblastic, phialidic, determinate, discrete, lageniform to doliiform, hyaline, smooth, with wide aperture and minute collarette, formed from the inner cells of the pycnidial wall, enclosed in mucoid sheath |
Absent |
| Neofusicoccum Crous, Slippers & A.J.L. Phillips | Mostly fusoid to ellipsoidal, hyaline. | Stromatic, pycnidial, solitary or aggregated, often occurring within the same stroma as the ascomata, walls composed of dark brown | When present hyaline, cylindrical, branched at the base, smooth, 0–1 septate | Enteroblastic, integrated, hyaline, smooth, cylindrical | Absent |
| Pseudofusicoccumaceae Tao Yang & Crous, Pseudofusicoccum Mohali, Slippers & M.J. Wingf. | Conidia are more cylindrical than in Noefusicoccum species and surrounded by a persistent mucous sheath, hyaline. | Large, superficial, unilocular or multilocular locule | Reduced to conidiogenous cells | Holoblastic, smooth, cylindrical to subcylindrical, hyaline |
Present or absent |
Notes. Macrophomina has sclerotia black, smooth, hard, formed of dark brown, thick-walled cells [8]. Neofusicoccum was introduced by Crous et al. [24] for species that are morphologically similar to Fusicoccum, but phylogenetically distinct from them, and thus could no longer be accommodated in that genus. The presence of paraphyses in Sphaeropsis differentiates this genus from Diplodia, which does not have pycnidial paraphyses and striate conidia of Lasiodiplodia differentiate it from Sphaeropsis, which has smooth-walled conidia [8]. Also, the absence of septa (aseptate) in mature conidia of Sphaeropsis separates it from the genus Diplodia which is characterized by conidia septate [8].
3. DNA Sequence-based identification of Botryosphaeriales in Venezuela
In the early 2000s, publications began appearing for identifying species within the Botryosphaeriales using DNA sequence data. DNA-based approaches helped to solve the problem of identifying species with overlapping morphology, and the combination of morphological characteristics and DNA sequence data became a powerful tool to separate and identify new genera and species [18,21]. However, single-gene genealogies were not always useful for resolving closely related or cryptic species of the Botryosphaeriales; moreover, comparisons of DNA sequence data from multiple genes or different gene regions were exceptionally useful for discriminating among several closely related species [19,43,44].
From mid-2000s through 2022, different species and genera within the Botryosphaeriales in Venezuela were isolated. Analysis of the morphological characteristics and DNA sequences were used for identifying a new genus and four new species. Multiple DNA loci were used to identify these Botryosphaeriales isolates from Venezuela including, the internal transcribed spacer of rDNA (ITS), translation elongation factor-1α (tef1), and β-tubulin (btub) (Table 3).
3.1. Phylogenetic analysis
For this review, a phylogenetic analysis was carried out for those genera and species of Botryosphaeriales in Venezuela that were identified by partial gene sequences available in NCBI GenBank Database (http://www.ncbi.nlm.nih.gov). For this analysis, Lasiodiplodia genus was analyzed separately from the remaining of the genera and species of Botryosphaeriales because it has the largest number of species reported for different hosts in Venezuela.
For the phylogenetic study were internal transcribed spacers 1 and 2 including the intervening 5.8S nrDNA gene (ITS) [45], the translation elongation factor 1-alpha gene (tef1) [46] and the beta-tubulin gene (tub2) [47]. The 3-loci concatenated alignment contained 1232 characters including gaps for Lasiodiplodia group (526 from ITS, 328 from tef1 and 378 from tub2) and 1295 characters including gaps for Botryosphaeriales remaining (532 from ITS, 333 from tef1 and 430 from tub2) (Table 3).
Phylogenetic analyses were performed for the combined datasets using two different methods: Maximum Likelihood (ML) and Bayesian Inference (BI). A partition homogeneity test (PHT) [48,49] was conducted to determine whether the datasets for the three gene regions could be combined. The PHT performed on the concatenated dataset of three gene regions yielded a P-value = 0.01. The value P-value was significant, datasets for multiple gene regions were combined for phylogenetic analysis. The ML phylogenies were evaluated with a bootstrapping (BS) method. ML phylogenies were performed with MEGA-X [50], and BI phylogenies were performed with MRBAYES v3.2.1 [51]. All sequences from representative isolates were aligned using MUSCLE that along with BI phylogenies were used in association with the Geneious Prime software version 2020.1.2. The best-fit nucleotide substitution models for the combined datasets (ITS, tef1, and btub) were identified separately for ML and BI. For BI analyses, the best-fit nucleotide substitution models were determined with jModeltest 2.1.10 [52] using the Akaike Information Criterion (AIC) and for ML were determined with MEGA-X [50], with HKY+G substitution model used as the best model for both. Phylogenetic species were determined with ML ≥50%, and BI ≥0.6 for Lasiodiplodia group (Figure 1), and ML ≥ 90% and BI ≥ 0.90 for the remaining of Botryosphaeriales (Figure 2).
4. Taxonomy, diversity and distribution of a new genus, new species and reports found in Venezuela and other regions of the world
The taxonomy of a new genus, new species, and reports of Botryosphaeriales identified by DNA sequences and their hosts in Venezuela are discussed below (Table 3, Figure 1, Figure 2 and Figure 3).
Cophinforma atrovirens (Mehl & Slippers) A. Alves & A.J.L. Phillips (Basionym: Fusicoccum atrovirens Mehl & Slippers) was isolated from stems and branches of A. mangium, Eucalyptus urophylla-hybrids, E. urophylla x E. grandis and reported for the first time in Cojedes (CR) and Portuguesa (WCR) states, [53], and from fruits and trees of Theobroma cacao L., in Merida state (AR) [54], Venezuela. Initially, Mohali et al. [53] reported this fungus as Botryosphaeria mamane D.E. Gardner (asexual morph Cophinforma mamane (D.E. Gardner) A.J.L. Phillips & A. Alves), but Phillips et al. [8] found that ITS sequences of the Venezuelan isolates of C. mamane are the same as the ITS sequence of C. atrovirens, therefore they consider the Venezuelan isolates to represent C. atrovirens.
In other regions of the world C. atrovirens was isolated from asymptomatic branches and twigs of Pterocarpus angolensis, in South Africa [55]; dead branch of Eucalyptus sp., in Thailand [56] as Cophinforma eucalypti Doilom, J.K. Liu & K.D. Hyde; it was also isolated from Dimocarpus longan Lour., but producing lesions on inoculated seedlings of Eucalyptus sp., in China [57]; and stem rot and dieback on Cashew tree (Anacardium occidentale) in Brazil [58].
Diplodia scrobiculata J. de Wet, Slippers & M.J. Wingf., (syn. Diplodia guayanensis F. Castro-Medina, J.R. Úrbez-Torres, S.R. Mohali & W.D. Gubler sp. nov., MycoBank 812480) was isolated from the trunk of A. mangium in plantations of Monagas state, North Eastern Region (NER), Venezuela [59]. Diplodia guayanensis was distinguished from D. scrobiculata by its larger conidia [59]. Later, sequence alignment ITS, tef1 and btub of D. scrobiculata and combining two (ITS and tef1) [60] and three loci (ITS, tef1 and btub) [61] for phylogenetic analysis both concluded that D. guayanensis is indistinguishable from D. scrobiculata based on phylogenetic analyses, and considered it to be a synonym for D. scrobiculata, and this was further supported on the basis that Úrbez-Torres et al. [59] used older sequences for D. scrobiculata in their phylogenetic analyses [60], although Zhang et al. [61] used the old sequences and obtained the same results as Linaldeddu et al. [60]. Furthermore, morphological variability is common in these fungi [8,60]; however, distinctive RFLP patterns were obtained for D. guayanensis compared against their closely related species D. scrobiculata and D. sapinea (Fr.) Fuckel (A and B) using Cfol restriction fragments in tef1 PCR products [59]. The PCR-RFLP fingerprinting profiles have been useful in this study to distinguish Botryosphaeriales, although overlapping RFLP patterns may be observed between some species using one, two, or more RE [62].
Diplodia scrobiculata was isolated and identified for the first time from needles of Pinus banksiana Lamb., P. resinosa Aiton, P. greggii Engelm. ex Parl., in USA (Wisconsin, Minnesota, California), Mexico, and Europe (France, Italy) [63]; wilted twigs, branch dieback, necrosis and stem cankers on Pinus halepensis Mill., trees, in Tunisia [64]; symptomless and die-back on Pinus patula Schiede ex Schltdl. & Cham., in South Africa [65]; asymptomatic trees in Pinus radiata D. Don plantations but producing lesions on inoculated P. radiata seedlings in Spain [66]; Pinus sp., in Canada [67]; and dieback on Coast redwood (Sequoia sempervirens (Lamb. ex. D. Don) Endl.) in California, USA [68].
Lasiodiplodia brasiliensis M.S.B. Netto, M.W. Marques & A.J.L. Phillips was isolated for the first time in Venezuela from T. cacao plantations in the state of Merida (AR), Venezuela [54], although Zhang et al. [61], reported to L. brasiliensis on P. caribaea var. hondurensis, F. insipida and J. copaia wood in Venezuela, these authors taken by mistake these sequences from GenBank that belong to L. theobromae from Venezuela (see Table S1 of these authors).
Lasiodiplodia brasiliensis was identified and reported for the first time in Brazil on stems of Mango (Mangifera indica L.) and fruits of Carica papaya L. [69] and other hosts in Brazil; saprobic on dead branch of teak (Tectona grandis L.f.), in Thailand [70]; Mango dieback, in Peru [71]; Adansonia madagascariensis Baill., in Madagascar [72]; Eucalyptus sp., in China [56]; as endophytic fungus isolated from healthy, brown, and ligaloes tissue of evergreen trees (Aquilaria crassna Pierre ex Lecomte), in Laos [73]; symptoms of gummosis, stem cankers, and dieback on Persian lime (Citrus latifolia Tan.), in Mexico [74]; Gossypium hirsutum L., in Australia [75]; leaf blight of Sansevieria trifasciata Prain (mother-in-law’s tongue or snake plant), ornamental plant, in Malaysia [76]; dieback and corky bark on longan trees (Dimocarpus longan L.), in Puerto Rico [77]; branch dieback, T. cacao, Cameroon and Psychotria tutcheri Dunn fruits, in China [61].
Cruywagen et al. [72] and Farr & Rossman [78] mistakenly cited to L. brasiliensis as the causing of dieback in strawberries (Fragaria x ananassa Duchesne), in Turkey, but the pathogen reported was L. theobromae [79].
Lasiodiplodia crassispora Burgess, Barber sp. nov., was isolated for the first time from wood of living E. urophylla in Acarigua, Portuguesa State (WCR), Venezuela and canker of Santalum album L., (sandalwood) in Western Australia, Australia [80]. The sandalwood is native to southern India, eastern Indonesia, and northern Australia (https://en.wikipedia.org/wiki/Santalum_album), therefore, L. crassispora found in central-western Venezuela (WCR) could have been introduced through imported eucalyptus seeds used for the plantations in Venezuela.
Lasiodiplodia crassispora was associated the internal wood decay symptoms observed in the cordon samples on grapevine (Vitis vinifera L.), in South Africa [81]; E. urophylla, in Uruguay [82]; perennial cankers in the vascular tissue of grapevines, in California, USA [83]; endophytic in Corymbia sp. Hook, and minor lesions in inoculations on 4-month-old baobab seedlings (Adansonia gregorii F.Muell.), in Australia [84]; dieback and stem-end rot of mango, fresh fruit of table grape (Vitis spp.), and causing dieback on Annonaceae in Brazil [85,86,87]; dieback symptoms from trunks and branches on grapevines in Sonora and Baja California, Mexico [88]. Lasiodiplodia crassispora (syn. Lasiodiplodia pyriformis F.J.J. van der Walt, Slippers & G.J.Marais) isolated from the leading edges of lesions on branches of Acacia mellifera (M.Vahl) Benth., in Namibia [61,89].
Lasiodiplodia pseudotheobromae A.J.L. Phillips, A. Alves & Crous was reported for the first time in Uverito plantations, Monagas State (NER), Venezuela in A. mangium [90].
Lasiodiplodia pseudotheobromae was identified for the first time from Gmelina arborea Roxb., (Melina) and A. mangium in Costa Rica, Rosa sp., in Netherlands, Coffea sp., in Zaire and Citrus aurantium L., Suriname [91]; isolated from trees apparently healthy or showing canker and dieback symptoms of Acacia confuse Merr., Albizia falcataria (L.) Fosberg, Eucalyptus sp., Mangifera sylvatica Roxb., and Paulownia fortunei (Seem.) Hemsl., in China [92]; dieback on blackthorn (Acacia mellifera (M.Vahl) Benth.), in Namibia [89]; Adansonia digitata L., in Mozambique and South Africa [71]; Cashew gummosis (Anacardium humile A.St.-Hil.), in Brazil [93]; Annona muricata L., in Australia [75]; Bouea burmanica Griff., Hevea brasiliensis (Willd. ex A.Juss.) Müll.Arg., Persea americana Mill., Coffea arabica L., Mangifera indica, Ficus racemosa L., Syzygium samarangense (Blume) Merr. & L.M.Perry, Dimocarpus longan Lour., in Thailand [94]; stem cankers, gummosis, and branches dieback Citrus latifolia Tan., in Mexico [74]; trunk cankers, Citrus reticulata Blanco, in Pakistan [95]; symptoms of branch dieback, cankers and fruit rot in Citrus sp., in Iran [96]; dieback, Mango, in Egypt, Peru and South Korea [71,97,98]; stem canker on the native Uruguayan tree, Myrcianthes pungens (O.Berg) D. Legrand and pathogenic in inoculated 4 month-old Eucalyptus grandis seedlings, in Uruguay [82]; dieback and fruit rot on Rambutan trees (Nephelium lappaceum L.), in Puerto Rico [77]; symptoms of branch dieback and cankers, and shoot and panicle blight in pistachio (Pistacia sp. and Pistacia vera L.), in Spain, [99]; shoot-dieback, gummosis, and sunken necrotic bark lesions in young nectarine (Prunus persica) trees, in Turkey [100]; Rosa sp., in Netherlands [91]; leaf blight of Sansevieria trifasciata, in Malaysia [76]; die-back disease on Schizolobium parahyba (Vell.) S. F. Blake var. amazonicum (Ducke) Barneby trees, in Ecuador [101]; trunk Diseases in Vitis vinifera, in Tunisia [102]; and post flowering stalk rot of maize (Zea mays L.), in India [103].
Lasiodiplodia theobromae (Pat.) Griffon & Maubl., is a cosmopolitan fungus occurring predominantly throughout tropical and subtropical regions [38,80]. It has also been known as a human pathogen causing keratomycosis and phaeohyphomycosis [103], and as a plant pathogen associated with about 500 plant hosts causing numerous diseases, including dieback, root rot, fruit rots, leaf spot and cankers of many others [38], and it also occurs as an endophyte [104].
Lasiodiplodia theobromae has been reported in Venezuela on A. mangium, and E. urophylla, in Portuguesa State (WCR) [105]; P. caribaea var. hondurensis, E. urophylla x E. grandis, and A. mangium, in Cojedes (CR), Falcon and Portuguesa States (WCR) [53]; Pinus caribaea and A. mangium, in Monagas State (NER) [59]; Ficus insipida, logs yard located within the natural forest of the Imataca Forest Reserve, between the Bolivar and Delta Amacuro States (GR) ([106]; Theobroma cacao, in Merida State (AR) [54].
In Venezuela, regarding the population structure of L. theobromae isolated from forest tree plantations was of a high gene flow between populations and a lack of population differentiation from the three host types considered, A. mangium and Eucalyptus urophylla, in Cojedes and Portuguesa State, and P. caribaea var. hondurensis in Falcon State, therefore the reproduction was predominantly clonal, and all three Venezuelan populations were pooled [104].
Lasiodiplodia venezuelensis Burgess, Barber, Mohali, sp. nov., MB500237 was isolated and described for the first time from wood of living Acacia mangium Willd., in Acarigua, Portuguesa State (WCR), Venezuela. Later, was found causing blue stain on Pinus caribaea Morelet var. hondurensis (Sénécl.) W.H.Barrett & Golfari wood and light-brown cankers with a black exudate on A. mangium in Monagas State (NER), and blue stain on Ficus insipida Willd., wood, Imataca Forest Reserve (natural forests), between the Bolivar and Delta Amacuro States (GR) [59,80,106]. To date, L. venezuelensis has only been reported in Venezuela, and found in the natural forest causing blue stain wood of F. insipida, and as a pathogen in A. mangium plantations. L. venezuelensis could be an endemic native fungus causing blue stain in light wood species native for Venezuela as is the case of F. insipida and moving onto an exotic species as a pathogen in A. mangium plantations [59].
Neofusicoccum arbuti (D.F. Farr & M. Elliott) Crous, Slippers & A.J.L. Phillips (syn. Neofusicoccum andinum (Mohali, Slippers & M.J. Wingf.) Mohali, Slippers & M.J. Wingf. comb. nov. MycoBank MB500871. Basionym: Fusicoccum andinum Mohali, Slippers & M.J. Wingf.) [8,24,105], was isolated from asymptomatic branches of mature Eucalyptus sp., trees in Mucuchies (3140 m), Cordillera Los Andes mountains (AR), Venezuela [105].
Li et al. [57,107] using combination of ITS, tef1, tub2, and rpb2 regions, with maximum parsimony (MP)/maximum likelihood (ML) tests analyses, they could separate cryptic species between N. andinum and N. arbuti. Later, Zhang et al. [61], evaluated the species in Botryosphaeriales, and performed Bayesian analysis of the combined ITS, tef1, tub2 and rpb2 sequence alignment to obtain a new phylogenetic tree of Neofusicoccum spp. They found that the ex-type culture of N. arbuti had nucleotide similarities with the sequences of the ex-type of N. andinum [(ITS: 466/471 (98.94 %), rpb2: 536/537 (99.81 %), tef1: 240/241 (99.59 %) and tub2: 376/376 (100 %), respectively], therefore N. andinum was reduced to synonymy with N. arbuti. Mohali et al. [105] did not include the N. arbuti sequences in the phylogenetic tree because these were not available at that time [61]. Neofusicoccum arbuti was isolated from cankers of Arbutus menziesii Pursh (Pacific madrone), in Washington and California, USA, and Canada [108], and stem canker and dieback of Vaccinium spp. (Blueberry), in Chile [109].
Neofusicoccum parvum (Pennycook & Samuels) Crous, Slippers & A.J.L. Phillips and Neofusicoccum ribis (Slippers, Crous & M.J. Wingf.) Crous, Slippers & A.J.L. Phillips both were isolated on E. urophylla S.T. Blake, and Botryosphaeria dothidea (Moug. ex Fr.) Ces. & De Not., isolated on E. urophylla x E. grandis W. Hill ex Maiden hybrids all from asymptomatic plant tissue, as well as trees exhibiting blue stain and die-back and from entirely dead trees in Portuguesa State, and N. parvum on Psidium guajava L., in Zulia State (ZR) [53].
Inoculation trial was conducted on E. urophylla x E. grandis hybrid stems in Portuguesa State with the fungi B. dothidea, N. parvum and N. ribis, and after 7 weeks lesions development was recorded. Botryosphaeria dothidea produced very small lesions in comparison to N. ribis and N. parvum which produced significantly larger lesions, bark swelling around the inoculation points and in some cases the bark was cracked producing black kino exudation when the outer bark was removed from the points of inoculation [110].
Information on the wide geographic distribution and host range of L. theobromae, N. parvum, N. ribis and B. dothidea can be found in Fungal Database (https://nt.ars-grin.gov/fungaldatabases/) and Mycobank Database (https://www.mycobank.org/).
Pseudofusicoccum Mohali, Slippers & M.J. Wingf. gen. nov. MycoBank MB500884; Pseudofusicoccum stromaticum (Mohali, Slippers & M.J. Wingf.) Mohali, Slippers & M.J. Wingf., comb. nov. MycoBank MB500885, Basionym: Fusicoccum stromaticum Mohali, Slippers & M.J. Wingf., [8,24,105], was isolated from branches of Eucalyptus urophylla S.T.Blake and E. urophylla x E. grandis W. Mill ex Maiden-hybrids, and from branches and stems of Acacia mangium Willd., in Western Central Region (WCR) of Venezuela [105].
Crous et al. [24] introduced to Pseudofusicoccum genus for species that are morphologically similar to Fusicoccum and Neofusicoccum but phylogenetically distinct from both of these genera. Pseudofusicoccum genus resembles species of Fusicoccum but distinct in having conidia encased in a persistent mucous sheath, and conidia are also more cylindrical than in Fusicoccum species [24]. Yang et al. [26] using robust backbone phylogeny for Botryosphaeriales (LSU and rpb2 genes) described and raised this genus as a new family, Pseudofusicoccumaceae Tao Yang & Crous where morphologically the family, is typified by Pseudofusicoccum.
In Venezuela, inoculations with P. stromaticum were made on 2-year-old trees in plantations of E. urophylla x E. grandis hybrid clones. Seven weeks after inoculation produced small lesions on the stems, but at the same time it was observed that the inoculation points had started to heal and produce callus by the end of the trial [110].
Pseudofusicoccum stromaticum has been widely reported in Brazil causing diseases in different hosts such as: dieback on mango (Mangifera indica L.) stems, pathogenic on 5-month-old mango seedlings, and producing the small lesions on inoculated mango fruits [111,112]; dieback, wilting of branches, discoloration of the vascular system, decline and subsequent death of Malay apple (Syzygium malaccense L.) trees [113]; associated with gummosis on native cashew (Anacardium othonianum Rizzinin) [93]; dieback and stem and branch cankers on cashew (Anacardium occidentale L.), guava (Psidium guajava L.) and caja-umbu (Spondias mombin L. x S. tuberosa Arruda) trees [114]; as endophyte in Myracrodruon urundeuva Fr. All. (Anacardiaceae) [115], and dieback of the Annonaceae [87]. In Uruguay, P. stromaticum was associated with cankers showing gummosis in peach shoots and showed moderate virulence on both inoculated apple and peach shoots [116].
In addition to P. stromaticum, eight species have subsequently been added to the genus, such as Pseudofusicoccum adansoniae Pavlic, T.I. Burgess, M.J. Wingf., on Adansonia gibbosa (A.Cunn.) Guymer ex D.A.Baum, Acacia synchronicia Maslin, Eucalyptus L'Hér., and Ficus opposite Miq., in Australia and, Ficus krishnae L. and Jatropha podagrica Hook, in India [117,118]; P. africanum Marinc., Jami & M.J. Wingf., on twigs of Mimusops caffra E.Mey. ex A.DC. (coastal red milkwood), in Eastern Cape Province, Haga Haga, South Africa [119]; P. ardesiacum Pavlic, T.I. Burgess, M.J. Wingf., on A. gibbosa and Eucalyptus sp., in Australia; P. artocarpi T. Trakunyingcharoen, L. Lombard & Crous, on twigs of Artocarpus heterophyllus Lam., in Chiang Mai Province, Thailand [94]; P. calophylli Jayasiri, E.B.G. Jones & K.D. Hyde on decaying fruit pericarp of Calophyllum inophyllum L., in Krabi Province, Mueang Krabi District, Thailand [120]; P. kimberleyense Pavlic, T.I. Burgess, M.J. Wingf., on Acacia synchronicia Maslin, Adansonia gibbosa, Eucalyptus sp., and Ficus opposite Miq. in Australia [121] and Persea americana Mill., USA [61]; P. olivaceum Mehl & Slippers on asymptomatic branches and twigs of Pterocarpus angolensis (Kiaat), in Mpumalanga Province, Kruger National Park, Pretoriuskop, Terminalia sericea Burch. ex DC., and Terminalia prunioides M.A.Lawson, in South Africa [55,61]; P. violaceum Mehl & Slippers on asymptomatic branch of P. angolensis Mpumalanga Province, Mawewe Nature Reserve, in South Africa [55], and Microcos paniculatus, in Hong Kong, China [61].
This genus is known only as the asexual morph and thus far nine species have been reported [61]. To date, P. stromaticum has been reported exclusively from South America while the remaining of the Pseudofusicoccum species have been reported from other regions, such as South Africa, Australia, Thailand, China, USA, and India [61,117,118].
5. Symptoms associated with species from Botryosphaeriales in Venezuela
Botryosphaeriales species infect plants via wounds or through natural plant openings, such as buds, lenticels, and stomata, resulting in diverse symptoms, such as twig, branch, and main stem cankers; die-back of leaders, shoots, or whole branches; seed capsule abortion; collar rot; damping off or blight of seedlings; root cankers; blue-stain; decline; and death of whole trees in severe cases [7]. The Table 1, different genera within the Botryosphaeriales were found and isolated from different hosts and locations in Venezuela, associated with diverse symptoms, and identified through its asexual morph, and other were identified using DNA sequence data (Table 3, Figure 1 and Figure 2).
Diplodia spp. and Lasiodiplodia spp., have been reported to cause different symptoms, such as blue stain (synonymous sap stain), which is a result of melanin, a pigment produced by the fungal pathogen [122]. The blue color of the wood develops as an optical effect due to refraction of light [123], such as observed in the following examples: Lasiodiplodia theobromae, L. venezuelensis and Diplodia mutila (Fr.) Mont., on Pinus caribaea var. hondurensis (Figure 4a–i); L. theobromae and L. venezuelensis on Ficus insípida (Figure 4j,k). The discolorations in the wood of living trees/woody plants or dead logs are the result of diverse biotic and abiotic causes [124,125]. Wood discoloration and decay are often the result from wounding, such as those caused by animal chewing, branch breaking, pruning, mechanized wood harvest, construction injury, motor traffic, etc. [126], and insects. Further discolorations can result from tree-produced substances, such as deposition of heartwood substances developed by living tree cells, later microbial stains, and finally colored derivatives of wood decay processes [124], examples of tree/wood discolorations include the following: sudden death or die-back in E. urophylla and Eucalyptus hybrid of Portuguesa State caused by Lasiodiplodia crassispora, L. theobromae, Neofusicoccum parvum, N. ribis, Botryosphaeria dothidea, Pseudofusicoccum stromaticum, and Cophinforma atrovirens (Figure 4l,m); discolorations on Acacia mangium in Cojedes and Portuguesa States caused by Lasiodiplodia theobromae, L. venezuelensis, Cophinforma atrovirens, and Pseudofusicoccum stromaticum (Figure 4n,o).
Discolorations and canker in the stem of A. mangium caused by Lasiodiplodia pseudotheobromae, L. theobromae, L. venezuelensis, and Diplodia scrobiculata (syn. D. guayanensis) in plantations of Maderas del Orinoco Company (Figure 5a–d), as well decline symptoms observed in A. mangium, and P. caribaea var. hondurensis. Pathogenicity tests were carried out in commercial plantations at the company Maderas del Orinoco to investigate the status of Botryosphaeriales associated with decline symptoms observed in A. mangium and P. caribaea var. hondurensis. Three Lasiodiplodia spp. and one Diplodia sp., were inoculated in A. mangium, and two Lasiodiplodia spp., on P. caribaea var. hondurensis. A. mangium showed bark swelling, vascular discoloration, necrosis, and cankering around the inoculation points (Figure 5e–g), while in P. caribaea var. hondurensis did not cause any lesions [59,90]. This study showed that Lasiodiplodia spp., and Diplodia sp., are highly virulent to A. mangium, showing. Other pathogenicity tests were carried out in the field, which gave us information about the susceptibility or tolerance to diseases, such as is the case of Eucalyptus spp., a forest species introduced in Venezuela to obtain fibers for cardboard production. These assessments of inoculations were made with different genera and species of Botryosphaeriales in commercial plantations of Eucalyptus at the company Smurfit Kappa Reforestadora Dos, Portuguesa State on different commercial clones of Eucalyptus-hybrids (E. urophylla x E. grandis), and where these clones were shown to be tolerant of Botryosphaeriaceae were observed [110]. Cophinforma atrovirens was isolated from T. cacao fruits with anthracnose and together with Lasiodiplodia theobromae, and L. brasiliensis were found in association with dieback or sudden death symptoms on T. cacao trees [54] in Merida State (Figure 5h), producing discolorations in branches (Figure 5i) and stems (Figure 5j,k). These discolorations were mainly associated with wounds caused by bark beetles- Scolytinae (Figure 5l). Stems, branches, and roots with cankers and dieback on P. caribaea var. hondurensis trees in plantations from 4 to15 years old and in nurseries on 8-month-old seedlings in displaying completely browned needles (Figure 5m,n) were observed at the Maderas del Orinoco Company, and the main fungal pathogen reported as causing these diseases was Sphaeropsis sapinea (Fr.) Dyko & B. Sutton [34]. Cedeño et al. [34] based their identification on the asexual morph, conidia 39,8 (37-45) x 12,7 (11-16) μm, one septum and rarely two or three septa (Figure 5o,p); measurements close to Diplodia sapinea (25.5-) 30.5-52.5 (-54) x (10-) 12.5-20 (-21) μm, D. scrobiculata (37.5-) 39.5 (-41.5) x (13-) 14 (-15.5) μm, and D. scrobiculata (syn. D. guayanensis) (33.5-) 40.6-42.4 (56) x (12-) 15.8-16.7 (-18.5) μm [59]. The absence of septa (aseptate) in mature conidia of Sphaeropsis separates it from the Diplodia genus, which is characterized by septate conidia [8] (Table 2), therefore, the diseases observed by Cedeño et al. [34] in the nurseries and plantations of P. caribaea var. hondurensis could have been caused by a fungal species in the Diplodia genus.
Neofusicoccum arbuti (syn. N. andinum) was collected from asymptomatic branches of mature Eucalyptus sp. trees growing in the Cordillera Los Andes Mountains of Venezuela at an altitude of ca. 3000 m (Figure 6a–d). Photographs of other Botryosphaeriales genera; Diplodia scrobiculata (syn. Diplodia guayanensis) (Figure 6e,f), Pseudofusicoccum stromaticum (Figure 6g–i); Neofusicoccum ribis/Neofusicoccum parvum (Figure 6j,k), Cophinforma atrovirens (Figure 6l, m), and B. dothidea (Figure 6n-p).
6. Conclusions
This is a review and update of information that represents almost 40 years of research work with species pertaining to the order Botryosphaeriales that cause diseases, with special reference to woody plants. The nomenclature of the different species and genera found within the Botryosphaeriales order have been updated, including the identification of the news species of Lasiodiplodia, a new genus and specie of Pseudofusiccocum, and new reports for Venezuela using molecular tools.
At the morphological level, nine genera were isolated and identified within Botryosphaeriales order, where Lasiodiplodia spp is the most abundant of all genera. This was isolated from fruit plantations such as citrus, mango, cacao, avocado, and forest tree plantations of exotic species such as pine, and from native forest species.
With molecular tools, it was possible to define exactly the name of the species that produce or are associated with forest diseases, especially in forest plantations of exotic species. Lasiodiplodia theobromae and L. venezuelensis both didn't cause lesions when they were inoculated in Pinus caribaea var. hondurensis trees, but they were routinely reisolated from asymptomatic wood which indicates the latent pathogen status of these species in this host, as well causing of blue stain on pine wood observed in fallen trees and in log yards at sawmills.
Lasiodiplodia pseudotheobromae, L. theobromae, L. venezuelensis, and Diplodia scrobiculata (=D. guayanensis) were isolated from trunks with symptoms light-brown cankers with a black exudate in Acacia mangium plantations. Inoculation tests carried out on this host showed bark swelling around the inoculation points and necrosis of the vascular system below the bark and black exudation, showing these four species their high virulence on A. mangium.
The fungi B. dothidea, C. atrovirens, L. theobromae, N. arbuti (=N. andinum), N. parvum, N. ribis and P. stromaticum isolated from Eucalyptus spp., plantations, were inoculated on hybrid Eucalyptus trees, where N. ribis and N. parvum produced significantly large lesions (canker) on the trunk, therefore these pathogens can be considered as new emerging diseases on these forest species introduced in the country; B. dothidea produced very small lesions, and the remainder of the fungi did not.
Regarding natural tropical forests in Venezuela, the blue stain of the wood on Ficus insipida in lumber yards was caused by L. theobromae and L. venezuelensis.
Plantations of the non-native forest species, Pinus caribaea var. hondurensis, in the East of Venezuela (between the States of Anzoátegui and Monagas), began in 1961. This plantation had a planted area of approximately 600,000 ha, but currently there are 112,000 ha. Later, and on a smaller scale, non-native species, Acacia mangium, was planted. These forest plantations border one of the largest natural forest reserves in South America, The Imataca Forest Reserve occupying approximately 3.7 million ha, and located between the Bolivar and Delta Amacuro States in Venezuela [106]. The proximity between non-native and native species has allowed native pathogens, such as Lasiodiplodia venezuelensis, found so far only in Venezuela, and together with L. theobromae, L. pseudotheobromae, D. arbuti (=D. guayanensis) to be transferred to these specie exotics causing blue stain on pine wood, and canker in A. mangium plantations.
Batista et al. [2] have assumed that human movement and trade are the main routes of dispersal for all species within the order Botryosphaeriales with worldwide distribution across all continents, with the exception of Antarctica, with climatic variability being the main limitations for the appearance of new stable populations and additionally, they also highlight that the disease expression is mainly due to occasional climatic events that can affect the susceptibility of the host.
Botryosphaeriales are reported as saprophytic, parasites, endophytic, and opportunistic pathogens in different crops, natural forests, and plantations, causing significant losses to the Venezuelan economy, but these losses are not quantified. Information on diseases caused by fungi of the Botryosphaeriales order and their description at the morphological level in Venezuela is very scarce, scattered, and with little information, in addition to future research in plant pathology, phylogenetic studies and fungal taxonomy, and the rest of the other areas of science that are developed in Venezuela is in great uncertainty due to current economic and political problems.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Not applicable
Funding
Not applicable
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
I gratefully acknowledge Dr. Ned B. Klopfenstein of Forest Service, Rocky Mountain Research Station (USDA) and Prof. Leslie Holland Department of Plant Pathology, University of Wisconsin-Madison, USA for their cooperation and comments for the improvement of this paper.
Conflicts of Interest
The author declares no conflict of interest.
References
- Wingfield, M.J.; Brockerhoff, E.G.; Wingfield, B.D.; Slippers, B. Planted forest health: The need for a global strategy. Science 2015, 349, 832–836. [Google Scholar] [CrossRef] [PubMed]
- Batista, E.; Lopes, A.; Alves, A. What Do We Know about Botryosphaeriaceae? An Overview of a Worldwide Cured Dataset. Forests 2021, 12, 313. [Google Scholar] [CrossRef]
- Wingfield, M.J.; Slippers, B.; Hurley,B. P.; Coutinho, T.A.; Wingfield, B.D.; Roux, J. Eucalypt pests and diseases: growing threats to plantation productivity. Southern For. 2008, 70, 139–144. [Google Scholar] [CrossRef]
- Windfield, M.J. Daniel McAlpine Memorial Lecture. Increasing threat of disease to exotic plantation forests in the southern hemisphere: lessons from cryphonectria canker. Australs. Plant Pathol. 2003, 32, 133–139. [Google Scholar] [CrossRef]
- Branco, M.; Brockerhoff, E.G.; Castagneyrol, B.; Orazio, C.; Jactel, H. Host range expansion of native insects to exotic trees increases with area of introduction and the presence of congeneric native trees. J. Appl. Ecol. 2015, 52, 69–77. [Google Scholar] [CrossRef]
- Karlman, M.; Hansson, P.; Witzell, J. Scleroderris canker on lodgepole pine introduced in northern Sweden. Can. J. For. Res. 1994, 24, 1948–1959. [Google Scholar] [CrossRef]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: diversity, ecology, and impact. Fungal Biol. Rev. 2007, 21, 90–106. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Alves,A. ; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef]
- Michailides, T.J.; Morgan, D.P. Spore release by Botryosphaeria dothidea in pistachio orchards and disease control by altering the trajectory angle of sprinklers. Phytopathology 1993, 83, 145–152. [Google Scholar] [CrossRef]
- Zlatković, M.; Keča, N.; Wingfield, M.J.; Jami, F.; Slippers, B. New and unexpected host associations for Diplodia sapinea in the Western Balkans. For. Pathol. 2017, 47, e12328. [Google Scholar] [CrossRef]
- Burgess, T.I.; Sakalidis, M.L.; Hardy, G.E.S.J. Gene flow of the canker pathogen Botryosphaeria australis between Eucalyptus globulus plantations and native eucalypt forests in Western Australia. Austral Ecol. 2006, 31, 559–566. [Google Scholar] [CrossRef]
- Pavlic, D.; Slippers, B.; Coutinho, T.A.; Wingfield, M.J. Botryosphaeriaceae occurring on native Syzygium cordatum in South Africa and their potential threat to Eucalyptus. Plant Pathol. 2007, 56, 624–636. [Google Scholar] [CrossRef]
- Stanosz, G.R.; Smith, D.R.; Leisso, R. Diplodia shoot blight and asymptomatic persistence of Diplodia pinea on or in stems of jack pine nursery seedlings. Forest Pathol. 2007, 37, 145–154. [Google Scholar] [CrossRef]
- Sakalidis, M.L.; Hardy, G.E.S.J.; Burgess, T.I. Class III endophytes, clandestine movement amongst hosts and habitats and their potential for disease; a focus on Neofusicoccum australe. Australasian Plant Pathol. 2011, 40, 510–521. [Google Scholar] [CrossRef]
- Begoude, B.A.D.; Slippers, B.; Perez, G.; Wingfield, M.J.; Roux, J. High gene flow and outcrossing within populations of two cryptic fungal pathogens on a native and non-native host in Cameroon. Fungal Biol. 2012, 116, 343–353. [Google Scholar] [CrossRef]
- Moral, J.; Morgan, D.; Trapero, A.; Michailides, T.J. Ecology and Epidemiology of Diseases of Nut Crops and Olives Caused by Botryosphaeriaceae Fungi in California and Spain. Plant Dis. 2019, 103, 1809–1827. [Google Scholar] [CrossRef]
- Jacobs, K.A.; Rehner, S.A. ; Rehner, S.A. Comparison of cultural and morphological characters and ITS sequences in anamorphs of Botryosphaeria and related taxa. Mycologia 1998, 90, 601–610. [Google Scholar] [CrossRef]
- Denman, S.; Crous, P.W.; Taylor, J.E.; Kang, J.Ch.; Pascoe, I.; Wingfield, M.J. An overview of the taxonomic history of Botryosphaeria, and a re-evaluation of its anamorphs based on morphology and ITS rDNA phylogeny. Stud. Mycol. 2000, 45, 129–140. Available online: https://wwwstudiesinmycologyorg/sim/Sim45/content/pdf/129-140.pdf.
- Pavlic, D.; Slippers, B.; Couthino, T.A.; Wingfield, M.J. Multiple gene genealogies and phenotypic data reveal cryptic species of the Botryosphaeriaceae: A case study on the Neofusicoccum parvum/N. ribis complex. Mol. Phylogenet. Evol. 2009, 51, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.R.; Stanosz, G.R. ; Stanosz, G.R. Molecular and morphological differentiation of Botryosphaeria dothidea (anamorph Fusicoccum aesculi) from some other fungi with Fusicoccum anamorphs. Mycologia 2001, 93, 505–515. [Google Scholar] [CrossRef]
- Zhou, S.; Stanosz, G.R. Relationships among Botryosphaeria species and associated anamorphic fungi inferred from the analysis of ITS and 5.8S rDNA sequences. Mycologia 2001, 93, 516–527. [Google Scholar] [CrossRef]
- Zhou, S.; Smith, D.R.; Stanosz, G.R. Differentiation of Botryosphaeria species and related anamorphic fungi using Inter Simple or Short Sequence repeat (ISSR) fingerprinting. Mycol. Res. 2001, 105, 919–926. [Google Scholar] [CrossRef]
- Slippers, B.; Crous, P.W.; Jami, F.; Groenewald, J.Z.; Wingfield, M.J. Diversity in the Botryosphaeriales: Looking back, looking forward. Fungal Biol. 2017, 121, 307–321. [Google Scholar] [CrossRef]
- Crous, P.W.; Slippers, B.; Wingfield, M.J.; Rheeder, J.; Marasas, W.F.O.; Philips, A.J.L.; Alves, A.; Burgess, T.; Barber, P.; Groenewald, J.Z. Phylogenetic lineages in the Botryosphaeriaceae. Stud. Mycol. 2006, 55, 235–253. [Google Scholar] [CrossRef] [PubMed]
- Slippers, B.; Boissin, E.; Phillips, A.J.L.; Groenewald, J.Z.; Wingfield, M.J.; Postma, A.; Burgess, T.; Crous, P.W. Phylogenetic lineages in the Botryosphaeriales: A systematic and evolutionary framework. Stud. Mycol. 2013, 76, 31–49. [Google Scholar] [CrossRef]
- Yang, T.; Groenewald, J.Z.; Cheewangkoon, R.; Jami, F.; Abdollahzadeh, J.; Lombard, L.; Crous, P.W. Families, genera, and species of Botryosphaeriales. Fungal Biol. 2017, 121, 322–346. [Google Scholar] [CrossRef]
- Wikee, S.; Lombard, L.; Nakashima, C.; Motohashi, K.; Chukeatirote, E.; Cheewangkoon, R.; Mckenzie, E.; Hyde, K.; Crous, P. A phylogenetic re-evaluation of Phyllosticta (Botryosphaeriales). Stud. Mycol. 2013, 76, 1–29. [Google Scholar] [CrossRef]
- Minnis, A.M.; Kennedy, A.H.; Grenier, D.B.; Palm, M.E.; Rossman, A.Y. Phylogeny and taxonomic revision of the Planistromellaceae including its coelomycetous anamorphs: contributions towards a monograph of the genus Kellermania. Persoonia 2012, 29, 11–28. [Google Scholar] [CrossRef]
- Wyka, S.A.; Broders, K.D. The new family Septorioideaceae, within the Botryosphaeriales and Septorioides strobi as a new species associated with needle defoliation of Pinus strobus in the United States. Fungal Biol. 2016, 8, 1030–1040. [Google Scholar] [CrossRef]
- Cedeño, L.; Palacios-Prü, E.; Palacios-Prü, E. ; Identificación de Botryodiplodia theobromae como la causa de las lesiones y gomosis en cítricos. Fitopatol. Venez. 1992, 5, 10–13. [Google Scholar]
- Cedeño, L.; Mohali, S.R.; Carrero, C. Primer reporte en Venezuela de Dothiorella dothidea como la causa de la podredumbre marrón en frutos del duraznero. Fitopatol. Venez. 1994, 7, 34–36. [Google Scholar]
- Cedeño, L.; Carrero, C.; Mohali, S.; Palacios-Pru, E.; Quintero, K. Muerte regresiva en parchita causada por Lasiodiplodia theobromae en Venezuela. Fitopatol. Venez. 1995, 8, 7–10. [Google Scholar]
- Cedeño, L.; Carrero, C.; Santos, R.; Quintero, K. Podredumbre marrón en frutos del guayabo causada por Dothiorella, fase conidial Botryosphaeria dothidea, en los estados Mérida y Zulia. Fitopatol. Venez. 1998, 11, 16–23. [Google Scholar]
- Cedeño, L.; Carrero, C.; Franco, W.; Torres-Lezama, A. Sphaeropsis sapinea asociado con quema del cogollo, muerte regresiva y cáncer en troncos, ramas y raíces del Pino Caribe en Venezuela. Interciencia 2001, 26, 210–215. Available online: https://www.redalyc.org/articulo.oa?id=33905606.
- Mohali, S. First Report in Venezuela of Sphaeropsis sapinea, causal agent of the blue stain on Caribbean pine. Fitopatol. Venez. 1997, 10, 23. [Google Scholar]
- Mohali, S.R.; Encinas, O. Association of Diplodia mutila with blue stain of Caribbean pine in Venezuela. For. Pathol. 2001, 31, 187–189. [Google Scholar] [CrossRef]
- Mohali, S.R.; Encinas, O.; Mora, N. Manchado azul en madera de Pinus oocarpa y Azadirachta indica en Venezuela. Fitopatol. Venez. 2002, 15, 30–32. [Google Scholar]
- Punithalingam, E. Plant diseases attributed to Botryodiplodia theobromae Pat. Commonw. Mycol. Inst., Ferry Lane, Kew, Surrey, UK, 1980; 123 pp.
- Urtiaga, R. Índice de enfermedades en plantas de Venezuela y Cuba. Impresos Nuevo Siglo. S.R.L., Barquisimeto, Venezuela, 1986; pp. 324.
- Iturriaga, T.; Minter, D.W. Fungi of Venezuela. 2006. Available online: http://www.cybertruffle.org.uk/venefung/eng/.
- Hernandez de Parra, J.B.; Ortega, R.; Blanco, G. Diagnóstico de enfermedades en frutales en el estado Yaracuy, Venezuela entre los años 2001-2011. Agronomia Trop. 2012, 62, 111–122. Available online: http://ve.scielo.org/scielo.php?script=sci_arttext&pid=S0002-192X2012000100009.
- Pacheco, C.; Suleima, L.; Manzanilla, E. Diversidad de hongos en cinco cultivares de mango (mangifera indica l.) del banco de germoplasma del inia-ceniap, Maracay. Bioagro 2016, 28, 201–208. Available online: http://ve.scielo.org/scielo.php?script=sci_arttext&pid=S1316-33612016000300007.
- Slippers, B.; Crous, P.W.; Denman, S.; Coutinho, T.A.; Wingfield, B.D. , Wingfield, M.J. Combined multiple gene genealogies and phenotypic characters differentiate several species previously identified as Botryosphaeria dothidea. Mycologia 2004, 96, 83–101. [Google Scholar] [CrossRef]
- Slippers, B.; Fourie, G.; Crous, P.W.; Coutinho, T.A.; Wingfield, B.D.; Carnegie, A.J.; Wingfield, M.J. Multiple gene sequences delimit Botryosphaeria australis sp. nov. from B. lutea. Mycologia 2004, 96, 1030–1041. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J.L. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: a guide to methods and applications: 315-322; Innis, M.A., Gelfand, D.H., Sninsky, J.J., et al., Eds.; Academic Press: San Diego, CA, USA, 1990. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous Ascomycetes. Mycologia 1999, 91, 553. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Farris, J.S.; Kallersjo, M.; Kluge, A.G.; Bult, C. , Testing significance of incongruence. Cladistics 1995, 10, 315–319. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*. Phylogenetic analysis using parsimony (*and other methods). Version 4.0. Sinauer Associates, Sunderland, Massachusetts. 2003. Available online: https://paup.phylosolutions.com/.
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Baysian phylogenetic interference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: more models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Mohali, S.; Slippers, B.; Wingfield, M.J. Identification of Botryosphaeria species from Eucalyptus, Acacia, and Pinus in Venezuela. Fungal Divers. 2007, 25, 103–125. Available online: https://www.fungaldiversity.org/fdp/sfdp/25-7.pdf.
- Mohali, S.R.; Woodward, S.; Klopfenstein, N.B.; Kim, M.-S.; Stewart, J.E. Mycobiota associated with anthracnose and dieback symptoms on Theobroma cacao L. in Merida state, Venezuela. Summa Phytopathol. 2023, 1, 00–00. [Google Scholar]
- Mehl, J.W.M.; Slippers, B.; Roux, J.; Wingfield, M.J. Botryosphaeriaceae associated with Pterocarpus angolensis (kiaat) in South Africa. Mycologia 2011, 103, 534–553. [Google Scholar] [CrossRef]
- Liu, J.K.; Phookamsak, R.; Doilom, M.; Wikee, S.; Li, Y.M.; Ariyawansha, H.; Boonmee, S.; Chomnunti, P.; Dai, D.Q.; Bhat, J.D.; Romero, A.I.; Zhuang, W.Y.; Monkai, J.; Jones, E.B.G.; Chukeatirote, E.; Ko Ko, T.W.; Zhao, Y.C.; Wang, Y.; Hyde, K.D. Towards a natural classification of Botryosphaeriales. Fungal Divers. 2012, 57, 149–210. Available online: https://link.springer.com/article/10.1007/s13225-012-0207-4. [CrossRef]
- Li, G.Q.; Liu, F.F.; Li, J.Q.; Liu, Q.L.; Chen, S.F. Botryosphaeriaceae from Eucalyptus plantations and adjacent plants in China. Persoonia 2018, 40, 63–95. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, J.E.; Fonseca, W.L.; Viana, F.M.P.; Ootani, M.A.; Araújo, F.S.A.; Brasil, S.O.S.; Mesquita, A.L.M.; Lima, C.S. First Report of Cophinforma atrovirens Causing Stem Rot and Dieback of Cashew Plants in Brazil. Plant Dis. 2019, 103, 1772. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R.; Castro-Medina, F.; Mohali, S.R.; Gubler, W.D. Botryosphaeriaceae species associated with cankers and dieback symptoms of Acacia mangium and Pinus caribaea var. hondurensis in Venezuela. Plant Dis. 2016, 100, 2455–2464. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Maddau, L.; Franceschini, A.; Alves, A.; Phillips, A.J.L. Botryosphaeriaceae species associated with lentisk dieback in Italy and description of Diplodia insularis sp. nov. Mycosphere 2016, 7, 962–977. Available online: https://www.mycosphere.org/pdf/Mycosphere_7_7_10-1.pdf. [CrossRef]
- Zhang, W.; Groenewald, J.Z.; Lombard. L.; Schumacher, R.K.; Phillips, A.J.L.; Crous, P.W. Evaluating species in Botryosphaeriales. Persoonia 2021, 46, 63–115. [Google Scholar] [CrossRef]
- Slippers, B.; Fourie, G.; Crous, P.W.; Coutinho, T.A.; Wingfield, B.D.; Carnegie, A.; Wingfield, M.J. Speciation and distribution of Botryosphaeria spp. on native and introduced Eucalyptus trees in Australia and South Africa. Stud. Mycol 2004, 50, 343–358. Available online: https://www.studiesinmycology.org/index.php/issue/52-studies-in-mycology-no-50.
- de Wet, J.; Wingfield, M.J.; Coutinho, T.A.; Wingfield, B.D. Characterization of Sphaeropsis sapinea isolates from South Africa, Mexico, and Indonesia. Plant Dis. 2000, 84, 151–156. [Google Scholar] [CrossRef]
- Hlaiem, S.; Boutiti, M.Z.; Jamaa, M.L.B. First report of shoot blight caused by Diplodia scrobiculata on Pinus halepensis in Tunisia. J. Plant Pathol. 2019, 101, 1237. [Google Scholar] [CrossRef]
- Jami, F.; Wingfield, M.J.; Gryzenhout, M.; Slippers, B. Diversity of tree-infecting Botryosphaeriales on native and non-native trees in South Africa and Namibia. Australasian Plant Pathol. 2017, 46, 529–545. [Google Scholar] [CrossRef]
- Manzanos, T.; Aragones, A.; Iturritxa, E. Diplodia scrobiculata: a latent pathogen of Pinus radiata reported in northern Spain. Phytopathol. Mediterr. 2017, 56, 274–277. Available online: https://www.jstor.org/stable/44809345.
- Burgess, T.I.; Gordon, T.R.; Wingfield, M.J.; Wingfield, B.D. Geographic isolation of Diplodia scrobiculata and its association with native Pinus radiata. Mycol. Res. 2004, 108, 1399–1406. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.A.; Rooney-Latham, S.; Brown, A.A.; McCormick, M.; Baston, D. Pathogenicity of three Botryosphaeriaceae fungi, Diplodia scrobiculata, Diplodia mutila, and Dothiorella californica, isolated from coast redwood (Sequoia sempervirens) in California. Forest Pathol. 2022, 52, 1–11. [Google Scholar] [CrossRef]
- Netto, M.S.B.; Assunção, I.P.; Lima, G.S.A.; Marques, M.W.; Lima, W.G.; Monteiro, J.H.A.; Balbino, V. de Q.; Michereff, S.J.; Phillips, A.J.L.; Câmara, M.P.S. Species of Lasiodiplodia associated with papaya stem-end rot in Brazil. Fungal Divers. 2014, 67, 127–141. [Google Scholar] [CrossRef]
- Doilom, M.; Shuttleworth, L.A.; Roux, J.; Chukeatirote, E.; Hyde, K.D. Botryosphaeriaceae associated with Tectona grandis (teak) in northern Thailand. Phytotaxa 2015, 233, 1–26. [Google Scholar] [CrossRef]
- Rodriguez-Galvez, E.; Guerrero, P.; Barradas, C.; Crous, P.W.; Alves, A. Phylogeny and pathogenicity of Lasiodiplodia species associated with dieback of mango in Peru. Fungal Biol. 2017, 121, 452–465. [Google Scholar] [CrossRef]
- Cruywagen, E.M.; Slippers, B.; Roux, J.; Wingfield, M.J. Phylogenetic species recognition and hybridisation in Lasiodiplodia: A case study on species from baobabs. Fungal Biol. 2017, 121, 420–436. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, S.; Zhao, L.; Sun, X.; He, W.; Zhang, Y.; Dai, Y.C. Lasiodiplodia spp. associated with Aquilaria crassna in Laos. Mycol. Prog. 2019, 18, 683–701. [Google Scholar] [CrossRef]
- Bautista-Cruz, M.A.; Almaguer-Vargas, G.; Leyva-Mir, S.G.; Colinas-Leon, M.T.; Correia, K.C.; Camacho-Tapia, M.; Robles-Yerena, L.; Michereff, S.J.; Tovar-Pedraza, J.M. Phylogeny, distribution, and pathogenicity of Lasiodiplodia species associated with cankers and dieback symptoms of Persian lime in Mexico. Plant Dis. 2019, 103, 1156–1165. [Google Scholar] [CrossRef]
- Tan, Y.P.; Shivas, R.G.; Marney, T.S.; Edwards, J.; Dearnaley, J.; Jami, F.; Burgess, T.I. Australian cultures of Botryosphaeriaceae held in Queensland and Victoria plant pathology herbaria revisited. Australasian Plant Pathol. 2019, 48, 25–34. [Google Scholar] [CrossRef]
- Kee, Y.J.; Zakaria, L.; Mohd, M.H. Lasiodiplodia species associated with Sansevieria trifasciata leaf blight in Malaysia. J. Gen. Plant Pathol. 2019, 85, 66–71. [Google Scholar] [CrossRef]
- Serrato-Diaz, L.M.; Aviles-Noriega, A.; Soto-Bauzo, A.; Rivera-Vargas, L.I.; Goenaga, R.; Bayman, P. Botryosphaeriaceae fungi as causal agents of dieback and corky bark in Rambutan and Longan. Plant Dis. 2020, 104, 105–115. [Google Scholar] [CrossRef]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, U.S. National Fungus Collections, ARS, USDA. Retrieved , 2023. Available online: https://nt.ars-grin.gov/fungaldatabases/ (accessed on 13 June 2023).
- Yildiz, A.; Benlioglu, K.; Benlioglu, H. First report of strawberry dieback caused by Lasiodiplodia theobromae. Plant Dis. 2014, 98, 1579. [Google Scholar] [CrossRef] [PubMed]
- Burgess, T.I.; Barber, P.A.; Mohali, S.; Pegg, G.; de Beer, W.; Wingfield, M.J. Three new Lasiodiplodia spp. from the tropics, recognized based on DNA comparisons and morphology. Mycologia 2006, 98, 423–435. [Google Scholar] [CrossRef] [PubMed]
- van Niekerk, J.M.; Bester, W.; Halleen, F.; Crous, P.W.; Fourie, P.H. First Report of Lasiodiplodia crassispora as a Pathogen of Grapevine Trunks in South Africa. Plant Dis. 2010, 94, 1063. [Google Scholar] [CrossRef] [PubMed]
- Perez, C.A.; Wingfield, M.J.; Slippers, B.; Altier, N.A.; Blanchette, R.A. Endophytic and canker-associated Botryosphaeriaceae occurring on non-native Eucalyptus and native Myrtaceae trees in Uruguay. Fungal Divers. 2010, 41, 53–69. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R.; Peduto, F.; Gubler, W.D. First report of grapevine cankers caused by Lasiodiplodia crassispora and Neofusicoccum mediterraneum in California. Plant Dis. 2010, 94, 785. [Google Scholar] [CrossRef]
- Sakalidis, M.L.; Hardy, G.E.S.J.; Burgess, T.I. Endophytes as potential pathogens of the baobab species Adansonia gregorii: a focus on the Botryosphaeriaceae. Fungal Ecol. 2011, 4, 1–14. [Google Scholar] [CrossRef]
- Marques, M.W.; Lima, N.B.; Morais, M.A. Jr.; Barbosa, M.A.G.; Souza, B.O.; Michereff, S.J.; Phillips, A.J.L.; Câmara, M.P.S. Species of Lasiodiplodia associated with mango in Brazil. Fungal Divers. 2013, 61, 181–193. [Google Scholar] [CrossRef]
- Correia, K.C.; Silva, M.A.; de Morais, M.A. Jr.; Armengold, J.; Phillips, A.J.L.; Camara, M.P.S.; Michereff, S.J. Phylogeny, distribution, and pathogenicity of Lasiodiplodia species associated with dieback of table grape in the main Brazilian exporting region. Plant Pathol. 2015, 65, 92–103. [Google Scholar] [CrossRef]
- Machado, A.R.; Custodio, F.A.; Cabral, P.G.C.; Capucho, A.S.; Pereira, O.L. Botryosphaeriaceae species causing dieback on Annonaceae in Brazil. Plant Pathol. 2019, 68, 1394–1406. [Google Scholar] [CrossRef]
- Rangel-Montoya, E.A.; Paolinelli, M.; Rolshausen, P.E.; Valenzuela-Solano, C.; Hernandez-Martinez, R. Characterization of Lasiodiplodia species associated with grapevines in Mexico. Phytopathol. Mediterr. 2021, 60, 237–251. [Google Scholar] [CrossRef]
- Slippers, B. , Roux, J.; Wingfield, M.J.; van der Walt, F.J.J.; Jami, F.; Mehl, J.W.M.; Marais, G.J. Confronting the constraints of morphological taxonomy in the Botryosphaeriales. Persoonia 2014, 33, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Castro-Medina, F.; Mohali, S.R.; Úrbez-Torres, J.R.; Gubler, W.D. First report of Lasiodiplodia pseudotheobromae causing trunk cankers in Acacia mangium in Venezuela. Plant Dis. 2014, 98, 686. [Google Scholar] [CrossRef] [PubMed]
- Alves, A.; Crous, P.W.; Correia, A.; Phillips, A.J.L. Morphological and molecular data reveal cryptic speciation in Lasiodiplodia theobromae. Fungal Divers. 2008, 28, 1–13. Available online: http://www.fungaldiversity.org/fdp/sfdp/28-1.pdf.
- Zhao, J.P.; Lu, Q.; Liang, J.; Decock, C.; Zhang, X.Y. Lasiodiplodia pseudotheobromae, a new record of pathogenic fungus from some subtropical and tropical trees in southern China. Cryptogam. Mycol. 2010, 31, 431–439. Available online: https://sciencepress.mnhn.fr/en/periodiques/mycologie/31/4/lasiodiplodia-pseudotheobromae-new-record-pathogenic-fungus-some-subtropical-and-tropical-trees-southern-china.
- Netto, M.S.B.; Lima, W.G.; Correia, K.C.; Silva, C.F.B.; Thon, M.; Martins, R.B.; Câmara, M.P.S. Analysis of phylogeny, distribution, and pathogenicity of Botryosphaeriaceae species associated with gummosis of Anacardium in Brazil, with a new species of Lasiodiplodia. Fungal Biol. 2017, 121, 437–451. [Google Scholar] [CrossRef]
- Trakunyingcharoen, T.; Lombard, L.; Groenewald, J.Z.; Cheewangkoon, R.; To-anun, C.; Crous, P.W. Caulicolous Botryosphaeriales from Thailand. Persoonia 2015, 34, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.Z.; Shafique, M.S.; Anwaar, H.A.; Sarfraz, S.; Tufail, M.R.; Fayyaz, A.; Muntaha, S.; Haque, K.; Ghuffar, S.; Amrao, L. First report of Lasiodiplodia pseudotheobromae causing trunk cankers in Citrus reticulata in Pakistan. Plant Dis. 2020, 104, 2522. [Google Scholar] [CrossRef]
- Abdollahzadeh, J.; Javadi, A.; Goltapeh, E.M.; Zare, R.; Phillips, A.J.L. Phylogeny and morphology of four new species of Lasiodiplodia from Iran. Persoonia 2010, 25, 1–10. [Google Scholar] [CrossRef]
- Ismail, A.M.; Cirvilleri, G.; Polizzi, G.; Crous, P.W.; Groenewald, J.Z.; Lombard, L. Lasiodiplodia species associated with dieback disease of mango (Mangifera indica) in Egypt. Australasian Plant Pathol. 2012, 41, 649–660. [Google Scholar] [CrossRef]
- Kwon, J.H.; Choi, O.; Kang, B.; Lee, Y.; Park, J.; Kang, D.W.; Han, I.; Kim, J. Identification of Lasiodiplodia pseudotheobromae causing mango dieback in Korea. Can. J. Plant Pathol. 2017, 39, 241–245. [Google Scholar] [CrossRef]
- Lopez-Moral, A.; del Carmen Raya, M.; Ruiz-Blancas, C.; Medialdea, I.; Lovera, M.; Arquero, O.; Trapero, A.; Agusti-Brisach, C. Aetiology of branch dieback, panicle and shoot blight of pistachio associated with fungal trunk pathogens in southern Spain. Plant Pathol. 2020, 69, 1237–1269. [Google Scholar] [CrossRef]
- Endes, A.; Kayim, M.; Eskalen, A. First report of Lasiodiplodia theobromae, L. pseudotheobromae, and Diplodia seriata causing bot canker and gummosis of nectarines in Turkey. Plant Dis. 2016, 100, 2321. [Google Scholar] [CrossRef]
- Mehl, J.W.M.; Slippers, B.; Roux, J.; Wingfield, M.J. Botryosphaeriaceae associated with die-back of Schizolobium parahyba trees in South Africa and Ecuador. For. Pathol. 2014, 44, 396–408. [Google Scholar] [CrossRef]
- Rezgui, A.; Vallance, J.; Ghnaya-Chakroun, A.B.; Bruez, E.; Dridi, M.; Demasse, R.D.; Rey, P.; Sadfi-Zouaoui, N. Study of Lasiodiplodia pseudotheobromae, Neofusicoccum parvum and Schizophyllum commune, three pathogenic fungi associated with the grapevine trunk diseases in the north of Tunisia. Eur. J. Plant Pathol. 2018, 152, 127–142. [Google Scholar] [CrossRef]
- Summerbell, R.C.; Krajden, S.; Levine, R.; Fuksa, M. Subcutaneous phaeohyphomycosis caused by Lasiodiplodia theobromae and successfully treated surgically. Med. Mycol. 2004, 42, 543–547. [Google Scholar] [CrossRef]
- Mohali, S.; Burgess, T.I.; Wingfield, M.J. Diversity and host association of the tropical tree endophyte Lasiodiplodia theobromae revealed using simple sequence repeat markers. Forest Pathol. 2005, 35, 385–396. [Google Scholar] [CrossRef]
- Mohali, S.; Slippers, B.; Wingfield, M.J. Two new Fusicoccum species from Acacia and Eucalyptus in Venezuela, recognized based on morphology and DNA sequences data. Mycol. Res. 2006, 110, 405–413. [Google Scholar] [CrossRef]
- Mohali, S.R.; Castro-Medina, F.; Úrbez-Torres, J.R.; Gubler, W.D. First report of Lasiodiplodia theobromae and L. venezuelensis associated with blue stain on Ficus insipida wood from the Natural Forest of Venezuela. For. Pathol. 2017, 47, 1–5. [Google Scholar] [CrossRef]
- Li, G.; Slippers, B.; Wingfield, M.J.; Chen, S. Variation in Botryosphaeriaceae from Eucalyptus plantations in Yun Nan Province in southwestern China across a climatic gradient. IMA Fungus 2020, 11, 22. [Google Scholar] [CrossRef]
- Farr, D.F.; Elliot, M.; Rossman, A.Y.; Edmonds, R.L. Fusicoccum arbuti sp. nov. causing cankers on Pacific madrone in western North America with notes on Fusicoccum dimidiatum, the correct name for Scytalidium dimidiatum and Nattrassia mangiferae. Mycologia 2005, 97, 730–741. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, J.G.; Briceño, E.X.; Chávez, E.R.; Úrbez-Torres, J.R.; Latorre, B.A. Neofusicoccum spp. associated with stem canker and dieback of blueberry in Chile. Plant Dis. 2009, 93, 1187–1194. [Google Scholar] [CrossRef] [PubMed]
- Mohali, S.; Slippers, B.; Wingfield, M.J. Pathogenicity of seven species of the Botryosphaeriaceae on Eucalyptus clones in Venezuela. Australasian Plant Pathol. 2009, 38, 135–140. [Google Scholar] [CrossRef]
- Marques, M.W.; Lima, N.B.; Michereff, S.J.; Câmara, M.P.S.; Souza, C.R.B. First report of mango dieback caused by Pseudofusicoccum stromaticum in Brazil. Plant Dis. 2012, 96, 144. [Google Scholar] [CrossRef] [PubMed]
- Marques, M.W.; Lima, N.B.; de Morais, N.A.; Michereff, S.J.; Phillips, A.J.L.; Câmara, M.P.S. Botryosphaeria, Neofusicoccum, Neoscytalidium and Pseudofusicoccum species associated with mango in Brazil. Fungal divers 2013, 61, 195–208. [Google Scholar] [CrossRef]
- Silveira, G.F.; Melo, M.P.; Teixeira, J.W.M.; Viana, D.C.; Silva, J.D.C.; Beserra, J.E.A. First report of Lasiodiplodia theobromae and Pseudofusicoccum stromaticum causing dieback in Syzygium malaccense tree in Brazil. For. Pathol. 2017, 48. [Google Scholar] [CrossRef]
- Coutinho, I.B. L.; Cardoso, J.E.; Lima, C.S.; Lima, J.S.; Goncalves, F.J.T.; Silva, A.M.S.; Freire, F.C.O. An emended description of Neofusicoccum brasiliense and characterization of Neoscytalidium and Pseudofusicoccum species associated with tropical fruit plants in northeastern Brazil. Phytotaxa 2018, 358, 251–264. [Google Scholar] [CrossRef]
- Sobreira, A.C.M.; Pinto, F.C.L.; Florencio, K.G.D.; Wilke, D.V.; Staats, C.C.; Streit, R.A.S.; de O Freire, F.C.; Pessoa, O.D.L.; Trindade-Silva, A.E.; Canuto, K.M. Endophytic fungus Pseudofusicoccum stromaticum produces cyclopeptides and plantrelated bioactive rotenoids. R. Soc. Chem. 2018, 8, 35575–35586. [Google Scholar] [CrossRef]
- Sessa, L.; Abreo, E.; Lupo, S. Pseudofusicoccum sp. causing shoot canker in peach in Uruguay. Australasian Plant Dis. Notes 2021, 16, 5. [Google Scholar] [CrossRef]
- Sharma, R.; Kulkarni, G.; Shouche, Y.S. Pseudofusicoccum adansoniae isolated as an endophyte from Jatropha podagrica: new record for India. Mycotaxon 2013, 123, 39–45. [Google Scholar] [CrossRef]
- Prasher, I.B.; Dhanda, R.K. First record of Pseudofusicoccum adonsoniae Pavlic, T.I. Burgess and M.J. Wingf. from Ficus krishnae (as endophyte) and new record for North India. J. New Biol. Rep. 2017, 6, 112–116. [Google Scholar]
- Jami, F.; Marincowitz, S.; Slippers, B.; Wingfield, M.J. New Botryosphaeriales on native red milkwood (Mimusops caffra). Australasian Plant Pathol. 2018, 47, 475–484. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Jeewon, R.; Phillips, A.J.L.; Bhat, D.J.; Wanasinghe, D.N.; Liu, J.K.; Lu, Y.Z.; Kang, J.C.; Xu, J.; Karunarathna, S.C. Diversity, morphology and molecular phylogeny of Dothideomycetes on decaying wild seed pods and fruits. Mycosphere 2019, 10, 1–186. [Google Scholar] [CrossRef]
- Pavlic, D.; Wingfield, M.J.; Barber, P.; Slippers, B.; Hardy, G.E.S.T.J.; Burgess, T.I. Seven new species of the Botryosphaeriaceae from baobab and other native trees in Western Australia. Mycologia 2008, 100, 851–866. [Google Scholar] [CrossRef]
- Zink, P.; Fengel, D. Studies on the colouring matter of blue-stain fungi. Part 2. Electron microscopic observations of the hyphae walls. Holzforschung 1989, 43, 371–374. [Google Scholar] [CrossRef]
- Mohali, S. Estudio histológico de madera de pino caribe con manchado azul causado por Botryodiplodia theobromae. Fitopatol. Venez. 1993, 6, 14–17. [Google Scholar]
- Bauch, J. Development and characteristics of discolored wood. IAWA Bull ns, 1984, 5, 91–98. [Google Scholar] [CrossRef]
- Kreber, B.; Byrne, A. Discolorations of hem-fir wood: a review of the mechanisms. Forest Prod. J. 1994, 44, 35–42. [Google Scholar]
- Tattar, T.A. Diseases of shade trees; Academic Press: New York, 1978; p. 361. [Google Scholar] [CrossRef]
- de Wet, J.; Burgess, T.; Slippers, B.; Preisig, O.; Wingfield, B.D.; Wingfield, M.J. Multiple gene genealogies and microsatellite markers reflect relationships between morphotypes of Sphaeropsis sapinea and distinguish a new species of Diplodia. Mycol. Res. 2003, 107, 557–566. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Alves, A.; Pennycook, S.R.; Johnston, P.R.; Ramaley, A.; Akulov, A.; Crous, P.W. Resolving the phylogenetic and taxonomic status of dark-spored teleomorph genera in the Botryosphaeriaceae. Persoonia 2008, 21, 29–55. [Google Scholar] [CrossRef] [PubMed]
- Phillips, A.J.L.; Alves, A.; Correia, A.; Luque, J. Two new species of Botryosphaeria with brown, 1-septate ascospores and Dothiorella anamorphs. Mycologia 2005, 97, 513–529. [Google Scholar] [CrossRef] [PubMed]
- Cedeño, L.; Mohali, S.; Palacios-Pru, E. Ultrastructure of Lasiodiplodia theobromae causal agent of Caribbean Pine blue stain in Venezuela. Interciencia 1996, 21, 264–271. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions, or products referred to in the content. |
Figure 1.
Phylogenetic tree of Lasiodiplodia genus in Venezuela results from Bayesian analysis (BI) of the combined ITS, tef1, and tub2 sequence alignment. Maximum likelihood (ML) bootstrap support values (ML ≥50%) and Bayesian posterior probabilities (BI ≥0.6) are shown at the nodes (ML/BI). Ex-type strains are indicated and all hosts named in the tree belong to Venezuela. The tree was rooted to Botryosphaeria dothidea CMW8000 Ex-type.
Figure 1.
Phylogenetic tree of Lasiodiplodia genus in Venezuela results from Bayesian analysis (BI) of the combined ITS, tef1, and tub2 sequence alignment. Maximum likelihood (ML) bootstrap support values (ML ≥50%) and Bayesian posterior probabilities (BI ≥0.6) are shown at the nodes (ML/BI). Ex-type strains are indicated and all hosts named in the tree belong to Venezuela. The tree was rooted to Botryosphaeria dothidea CMW8000 Ex-type.
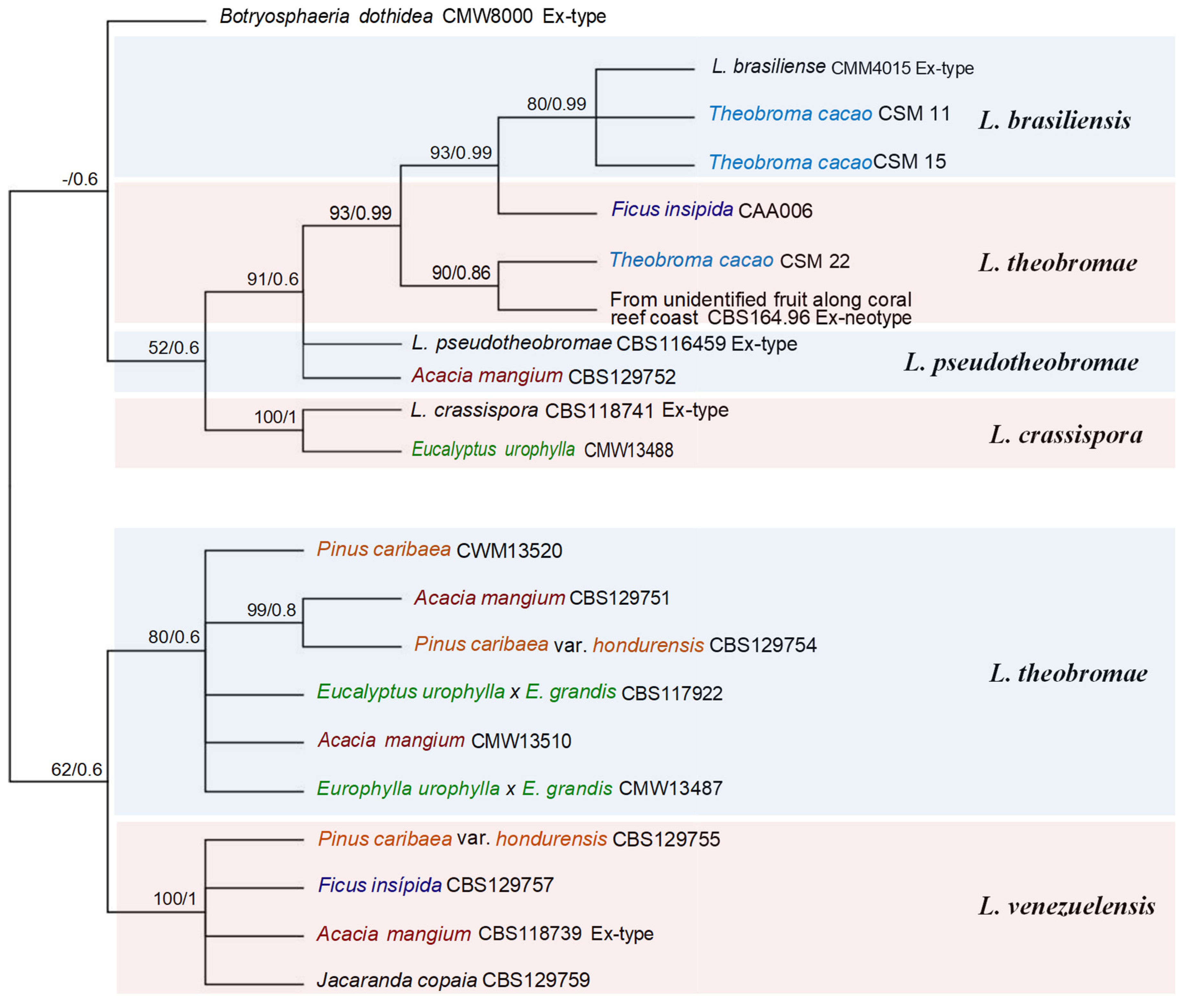
Figure 2.
Phylogenetic tree of Botryosphaeriales remaining group in Venezuela results from Bayesian analysis (BI) of the combined ITS, tef1, and tub2 sequence alignment. Maximum likelihood (ML) bootstrap support values (ML ≥ 90%) and Bayesian posterior probabilities (BI ≥ 0.90) are shown at the nodes (ML/BI). Ex-type strains are indicated and all hosts named in the tree belong to Venezuela. The tree was rooted to Lasiodiplodia brasiliense CMM4015 Ex-type.
Figure 2.
Phylogenetic tree of Botryosphaeriales remaining group in Venezuela results from Bayesian analysis (BI) of the combined ITS, tef1, and tub2 sequence alignment. Maximum likelihood (ML) bootstrap support values (ML ≥ 90%) and Bayesian posterior probabilities (BI ≥ 0.90) are shown at the nodes (ML/BI). Ex-type strains are indicated and all hosts named in the tree belong to Venezuela. The tree was rooted to Lasiodiplodia brasiliense CMM4015 Ex-type.
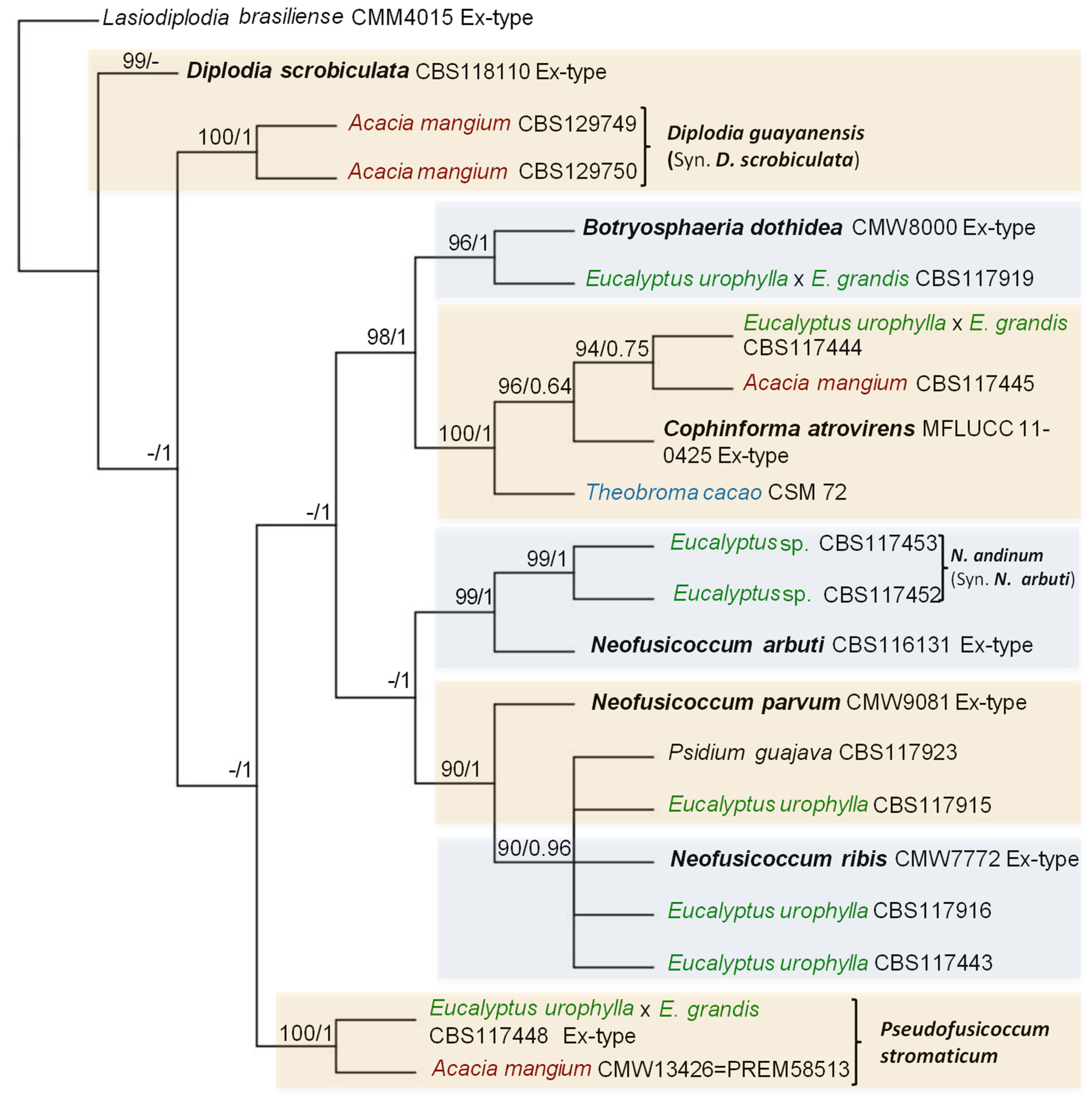
Figure 3.
Localities or states in Venezuela where Botryosphaeriales has been reported using sequences data (bold) and morphological methods (dark red).
Figure 3.
Localities or states in Venezuela where Botryosphaeriales has been reported using sequences data (bold) and morphological methods (dark red).
Figure 4.
(a) Pinus caribaea var. hondurensis plantations (Maderas del Orinoco C.A) located in Uverito, Monagas state, Venezuela. (b) Blue stain of the wood, observed in fallen trees inside the plantations. (c) Log yard to be processed at the sawmill in Maderas del Orinoco company. (d) Blue stain in the logs at the sawmill. (e) Pycnidium. (f) Conidia of Lasiodiplodia spp. (g, h) Hyphae of Lasiodiplodia spp. invading the medullary rays of Pinus caribaea var. hondurensis (red arrows), g=80X and h=Scanning Electron Microscopy. (i) Hypha found between tracheid or intercellular space of the wood of Caribbean pine (red arrow), Transmission Electron Microscope (19000X). (j) Blue stain in lumber of Ficus insipida; (k) Cross section Ficus insipida lumber with blue stain; (l) Eucalyptus urophylla trees exhibiting dieback or entirely dead trees (sudden death) in Portuguesa state. (m) Eucalyptus urophylla tree dead with blue stain. (n) Acacia mangium plantations in Portuguesa State. (o) Discoloration (red arrow) on Acacia mangium trees in Portuguesa State. Pictures e, f [32]; g [123]; h, i [130]; j, k [106].
Figure 4.
(a) Pinus caribaea var. hondurensis plantations (Maderas del Orinoco C.A) located in Uverito, Monagas state, Venezuela. (b) Blue stain of the wood, observed in fallen trees inside the plantations. (c) Log yard to be processed at the sawmill in Maderas del Orinoco company. (d) Blue stain in the logs at the sawmill. (e) Pycnidium. (f) Conidia of Lasiodiplodia spp. (g, h) Hyphae of Lasiodiplodia spp. invading the medullary rays of Pinus caribaea var. hondurensis (red arrows), g=80X and h=Scanning Electron Microscopy. (i) Hypha found between tracheid or intercellular space of the wood of Caribbean pine (red arrow), Transmission Electron Microscope (19000X). (j) Blue stain in lumber of Ficus insipida; (k) Cross section Ficus insipida lumber with blue stain; (l) Eucalyptus urophylla trees exhibiting dieback or entirely dead trees (sudden death) in Portuguesa state. (m) Eucalyptus urophylla tree dead with blue stain. (n) Acacia mangium plantations in Portuguesa State. (o) Discoloration (red arrow) on Acacia mangium trees in Portuguesa State. Pictures e, f [32]; g [123]; h, i [130]; j, k [106].

Figure 5.
(a) Acacia mangium tree with canker in the stem (red arrow) in Maderas del Orinoco plantations. (b, c) Discoloration in the A. mangium stem. (d) Discoloration from A. mangium tree base. (e) Pathogenicity test with different Botryosphaeria species using standing trees of A. mangium in a commercial plantation, and where A. mangium tree showed bark swelling around the inoculation points and necrosis of the vascular system below the bark (canker) 12 weeks after inoculation. (f, g) Black exudation was observed when the outer bark was removed from inoculation points (red arrows). (h) Dieback or sudden death symptoms in Theobroma cacao tree. (i) Discoloration in branch; (j, k) Discolorations in stems of T. cacao with dieback or sudden death symptoms in Merida State. (l) Bark beetle (Scolytinae) collected from cacao tree stem with discoloration. (m) Control seedling (left) and, tip death symptom in a seedling inoculated with Sphaeropsis sapinea (right) in Caribbean pine three weeks after inoculation. (n) Seedling with longitudinal stem section inoculated with S. sapinea where the discoloration within the stem can be observed. (o) Pycnidium and, (p) S. sapinea Conidia (possible Diplodia sp.). Pictures e-g [59]; m-p [34].
Figure 5.
(a) Acacia mangium tree with canker in the stem (red arrow) in Maderas del Orinoco plantations. (b, c) Discoloration in the A. mangium stem. (d) Discoloration from A. mangium tree base. (e) Pathogenicity test with different Botryosphaeria species using standing trees of A. mangium in a commercial plantation, and where A. mangium tree showed bark swelling around the inoculation points and necrosis of the vascular system below the bark (canker) 12 weeks after inoculation. (f, g) Black exudation was observed when the outer bark was removed from inoculation points (red arrows). (h) Dieback or sudden death symptoms in Theobroma cacao tree. (i) Discoloration in branch; (j, k) Discolorations in stems of T. cacao with dieback or sudden death symptoms in Merida State. (l) Bark beetle (Scolytinae) collected from cacao tree stem with discoloration. (m) Control seedling (left) and, tip death symptom in a seedling inoculated with Sphaeropsis sapinea (right) in Caribbean pine three weeks after inoculation. (n) Seedling with longitudinal stem section inoculated with S. sapinea where the discoloration within the stem can be observed. (o) Pycnidium and, (p) S. sapinea Conidia (possible Diplodia sp.). Pictures e-g [59]; m-p [34].

Figure 6.
(a, b) Eucalyptus sp., at the Cordillera Los Andes Mountains, Merida state, Venezuela at an altitude of approx. 3140 meters above sea level. The black arrows show old Eucalyptus trees without apparent damage. (c) Conidiogenous cells and (d) Conidia of Neofusicoccum arbuti (syn. Neofusicoccum andinum) isolated from Eucalyptus sp., asymptomatic branches at the Cordillera Los Andes Mountains. (e) Conidia immature and mature of Diplodia scrobiculata (syn. Diplodia guayanensis). (f) Mature conidia of D. guayanensis with two and three septa. (g) Pseudofusicoccum stromaticum producing big conidioma on 2 % Malt Extract Agar. (h) Multilocular conidiomata of P. stromaticum without ostioles and embedded locule. (i) Pseudofusicoccum stromaticum conidia encased in a persistent mucous sheath (red arrow). (j) Neofusicoccum ribis/parvum complex conidiogenous cells. (k) Neofusicoccum ribis/parvum complex conidia. (l) Cophinforma atrovirens, asci bitunicate with ascospores aseptate, hyaline, with granular textured contents. (m) Cophinforma atrovirens conidia with one and two septa. (n-p) Botryosphaeria dothidea conidia with 0-2 septa. Pictures e, f [59]; g-I [105]; j-p [53].
Figure 6.
(a, b) Eucalyptus sp., at the Cordillera Los Andes Mountains, Merida state, Venezuela at an altitude of approx. 3140 meters above sea level. The black arrows show old Eucalyptus trees without apparent damage. (c) Conidiogenous cells and (d) Conidia of Neofusicoccum arbuti (syn. Neofusicoccum andinum) isolated from Eucalyptus sp., asymptomatic branches at the Cordillera Los Andes Mountains. (e) Conidia immature and mature of Diplodia scrobiculata (syn. Diplodia guayanensis). (f) Mature conidia of D. guayanensis with two and three septa. (g) Pseudofusicoccum stromaticum producing big conidioma on 2 % Malt Extract Agar. (h) Multilocular conidiomata of P. stromaticum without ostioles and embedded locule. (i) Pseudofusicoccum stromaticum conidia encased in a persistent mucous sheath (red arrow). (j) Neofusicoccum ribis/parvum complex conidiogenous cells. (k) Neofusicoccum ribis/parvum complex conidia. (l) Cophinforma atrovirens, asci bitunicate with ascospores aseptate, hyaline, with granular textured contents. (m) Cophinforma atrovirens conidia with one and two septa. (n-p) Botryosphaeria dothidea conidia with 0-2 septa. Pictures e, f [59]; g-I [105]; j-p [53].

Table 3.
Genera and species within Botryosphaeriales order identified with DNA sequence data in Venezuela.
Table 3.
Genera and species within Botryosphaeriales order identified with DNA sequence data in Venezuela.
| Species | Accession number | Host | Locality | GenBank accession number | References | |||
|---|---|---|---|---|---|---|---|---|
| ITS | ITS | BTUB | ||||||
| Botryosphaeria dothidea | CMW8000 Ex-type | Prunus sp. | Switzerland | AY236949 | AY236898 | AY236927 | [43] | |
| B. dothidea | CMW13390= CBS117919 |
Eucalyptus urophylla x E.grandis |
CR and WCR | EF118044 | - | - | [53] | |
| Cophinforma atrovirens | CMW13416= CBS117444 |
E.urophylla x E.grandis |
CR and WCR | EF118050 | GU134938 | - | [53] | |
| C. atrovirens | CMW13425= CBS117445 | Acacia mangium | CR and WCR | EF118046 | GU134939 | - | [53] | |
| C. atrovirens | CSM 72 | Theobroma cacao | AR | MF436087 | MF436099 | MF436111 | [54] | |
| C. atrovirens | MFLUCC 11-0425 Ex-type |
Eucalyptus sp | Thailand | JX646800 | JX646865 | JX646848 | [56] | |
| Diplodia scrobiculata(syn. D. guayanensis) | CBS129749 | Acacia mangium | NER | JX545106 | JX545126 | JX545146 | [59] | |
| D. scrobiculata(syn. D. guayanensis) | CBS129750 | Acacia mangium | NER | JX545108 | JX545128 | JX545148 | [59] | |
| D. scrobiculata | CMW189 = CBS 118110 Ex-type | Pinus banksiana | United States | KF766160 | KF766399 | AY624258 | [60,127] | |
| Lasiodiplodia brasiliense | CMM4015 Ex-type | Mangifera indica | Brazil | JX464063 | JX464049 | - | [69] | |
| L. brasiliensis | CSM 11 | Theobroma cacao | AR | MF436018 | MF436006 | MF435998 | [54] | |
| L. brasiliensis | CSM 15 | Theobroma cacao | AR | MF436019 | MF436007 | MF435997 | [54] | |
| L. crassispora | WAC 12533= CBS118741 Ex-type | Santalum album | Australia | DQ103550 | EU673303 | KU887506 | [72,80] | |
| L. crassispora | CMW 13488 | Eucalyptus urophylla | CR and WCR | DQ103552 | DQ103559 | KU887507 | [72,80] | |
| L. pseudotheobromae | CBS 129752 | Acacia mangium | NER | JX545091 | JX545111 | JX545131 | [90] | |
| L. pseudotheobromae | CBS116459 Ex-type | Gmelina arborea | Costa Rica | KF766193 | EF622057 | EU673111 | [25,128] | |
| Lasiodiplodia theobromae | CBS 164.96 Ex-neotype | From unidentified fruit along coral reef coast | Papua New Guinea, Madang | AY640255 | AY640258 | KU887532 | [72,129] | |
| L. theobromae | CSM 22 | Theobroma cacao | AR | MF436023 | MF436011 | MF436005 | [54] | |
| L. theobromae | CBS129751 | Acacia mangium | NER | JX545096 | JX545116 | JX545136 | [59] | |
| L. theobromae | CMW13487 |
Europhylla urophylla x E. grandis |
CR and WCR | EF118053 | - | - | [53] | |
| L. theobromae | CBS129754 | Pinus caribaea var. hondurensis | NER | JX545099 | JX545119 | JX545139 | [59] | |
| L. theobromae | CMW13489= CBS117922 |
Eucalyptus urophylla x E. grandis | CR and WCR | DQ103525 | - | - | [53] | |
| L. theobromae | CMW13510 | Acacia mangium | CR and WCR | DQ103526 | - | - | [43] | |
| L. theobromae | CMW13520 | Pinus caribaea | CR and WCR | DQ103527 | - | - | [43] | |
| L. theobromae | CAA006 | Ficus insipida | GR | DQ458891 | DQ458876 | DQ458859 | [106] | |
| L. venezuelensis | CBS129755 |
Pinus caribaea var. hondurensis |
NER | JX545104 | JX545124 | JX545144 | [59] | |
| L. venezuelensis | CBS129757 | Ficus insípida | GR | JX545102 | JX545122 | - | [106] | |
| L. venezuelensis | WAC12539= CBS118739 Ex-type |
Acacia mangium | CR and WCR | DQ103547 | DQ103568 | KU887533 | [72,80] | |
| L. venezuelensis | CBS 129759 | Jacaranda copaia | GR | JX545101 | JX545121 | JX545141 | F. Castro-Medina/ S.R. Mohali-unpublished |
|
| Neofusicoccum arbuti | CBS 116131=AR 4014 Ex-type | Arbutus menziesii | USA | AY819720 | KF531792 | KF531793 | [8,108] | |
| Neofusicoccum arbuti(syn. N. andinum) | CMW13455= CBS117453 |
Eucalyptus sp. | AR | AY693976 | AY693977 | KX464923 | [26,105] | |
| Neofusicoccum arbuti(syn. N. andinum) | CMW13446= CBS117452 |
Eucalyptus sp. | AR | DQ306263 | DQ306264 | KX464922 | [26,105] | |
| N. parvum | CMW9081 Ex-type | Eucalyptus grandis | South Africa | AY236943 | AY236888 | [43] | ||
| N. parvum | CMW13350= CBS117923 | Psidium guajava | ZR | EF118036 | - | - | [53] | |
| N. parvum | CMW13355= CBS117915 | Eucalyptus urophylla | CR and WCR | EF118035 | - | - | [53] | |
| N. ribis | CMW7772 Ex-type | Ribes sp. | New York, United States | AY236935 | AY236877 | - | [43] | |
| N. ribis | CMW13360= CBS117916 | Eucalyptus urophylla | CR and WCR | EF118037 | - | - | [53] | |
| N. ribis | CMW13410= CBS117443 | Eucalyptus urophylla | CR and WCR | EF118038 | - | - | [53] | |
| Pseudofusicoccum stromaticum | CMW13434= CBS117448 Ex-type |
Eucalyptus urophylla x E. grandis |
CR and WCR | AY693974 | AY693975 | EU673094 | [105,128] | |
| P. stromaticum | CMW13426= PREM 58513 |
Acacia mangium | CR and WCR | EF118041 | - | - | [53] | |
Acronyms of culture collections: CBS: Centraalbureau voor Schimmelcultures, Fungal Biodiversity Centre, Utrecht, The Netherlands; IBL: Independent Biological Laboratories Israel. KEFAR MALAL; CMW: Tree Patholgy Co-operative Program, Forestry and Agricultural Biotechnology Institute, University of Pretoria, South Africa; WAC: Department of Agriculture, Western Australia Plant Pathogen Collection, South Perth, Western Australia; CSM: Personal culture collection deposited in the Department of Bioagricultural Sciences & Pest Management, Colorado State University, USA. MFLUCC: Mae Fah Luang University Culture Collection, ChiangRai, Thailand. CAA: A. Alves, Universidade de Aveiro, Portugal. Locality in Venezuela (see map-Figure 3): Central Region=CR (Cojedes state) and Western Central Region=CR and WCR (Falcon, Lara, and Portuguesa states). Los Andes Region (Mérida state) = AR. Northeastern Region (Anzoátegui and Monagas states) = NER. Guayana Region (Bolívar and Delta Amacuro states) = GR. Zulia Region (Zulia state) = ZR. (-) = No sequences.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



