
Submitted:
26 June 2023
Posted:
27 June 2023
You are already at the latest version
Abstract
Piceatannol is a naturally occurring polyphenolic compound that is widely found in grapes, blueberries, mushrooms, passion fruit and other edible fruits and vegetables. It has a variety of biological activities and pharmacological effects, including anti-inflammatory, antioxidation, antiaging, antiviral, antitumor, anticancer, antiatherosclerosis, antiparasitic, immunoregulatory, and cardiovascular protection effects. The aim of this study was to investigate the antiherpesvirus effects of piceatannol. Pseudorabies virus (PRV) belongs to the family Herpesviridae. PRV has a wide host range and can cause cytopathic effects (CPEs) in PK-15 cells. Therefore, PRV was used as a model to study the antiherpesvirus effect of piceatannol. In this study, we evaluated the antiviral activity of piceatannol against PRV in vitro and in vivo. The results showed that piceatannol could reatrain PRV multiplication in PK-15 cells in a dose-dependent manner. The 50% inhibitory concentration (IC50) was 0.0307 mg/mL, the IC50 was 0.0307 mg/mL, and the selectivity index (SI, CC50/IC50) was 3.68. The addition of piceatannol at different stages of PRV infection inhibited the viral adsorption and intracellular replication phases of PRV. Piceatannol significantly reduced the expression levels of the IE180, EP0, UL29, UL44, US6 and UL27 genes of PRV within 48 h, significantly reduced the expression levels of the PRV gB and gD proteins, and reduced PRV-induced apoptosis. Molecular docking showed that piceatannol had good binding activity with the PRV gB and gD proteins. The results of animal experiments in vitro indicated that piceatannol could delay the onset of disease, improve the survival rate of the mice (14.3%), reduce the viral load in the brain and kidney of the mice, alleviate the pathological changes in the mouse tissues and organs, and increase the levels of TNF-α, IEN-γ and IL-4 in the serum of the mice. These data indicate that piceatannol has good anti-PRV activity in vitro and in vivo, indicating that it could be a novel antiherpesvirus infection agent in the future.
Keywords:
Piceatannol
; Pseudorabies virus
; Antiviral activity
; in vivo
; in vitro
1. Introduction
Pseudorabies virus (PRV) is a double-stranded DNA virus belonging to the family Herpesviridae, a member of the genus Varicellovirus, and the causative agent of pseudorabies (also known as Aujeszky disease, AD) [1]. PRV can infect most mammals, mainly affecting the nervous and reproductive systems of infected animals, and can cause the death of piglets and other susceptible animals. In addition, PRV induces a cytopathic effect (CPE) in infected cells [2]. Therefore, PRV is an important model virus for studying herpesvirus biology and effects on neuronal pathways. Recent studies have shown that PRV can infect humans and cause central nervous system diseases, endophthalmitis and encephalitis [3,4]. This suggests that PRV infection is a new and potential public health hazard.
At present, viral diseases are mainly prevented by vaccination. Under the synergistic effect of treatment and vaccination, the diet pathway also plays an important role in the treatment of viral diseases [5]. Studies have shown that phytochemicals in many fruits and vegetables can promote health and reduce the occurrence of chronic diseases [6]. Many studies have shown that the chemical constituents of edible plants have antiviral activity. These natural compounds exist not only in some traditional Chinese medicines but also in some fruits and vegetables [7,8]. Studies have shown that anthocyanins in grapes have inhibitory effects on adenovirus and herpes simplex virus type 1 [9]. The chemical constituents of garlic, namely, quercetin, allicin and thio-sulfenate, have potent antiviral activities against dengue virus, influenza A and B viruses, human immunodeficiency virus (HIV), herpesvirus type 1 and type 2, and Newcastle disease virus [10,11,12,13]. The components of persimmon extracts, such as glycyrrhizin, flavonol, luteolin, apigenin and tannin, have inhibitory activity against Newcastle disease virus, vesicular stomatitis virus, adenovirus and coxsackievirus [14,15]. The linosides and alkaloids in peppers can inhibit hepatitis B virus pathogenesis [16]. The components of guava extracts, namely, tannins and polyphenols, have inhibitory effects on the influenza A H1N1 virus [17,18]. Therefore, these edible plants serve as sources of natural compounds with antiviral effects that can be identified by screening.
Natural compounds have the advantages of being available from a wide range of sources, exhibiting low toxicity, and having a wide range of biological activities [19]. Piceatannol is a bioactive polyphenol substance. It is mainly found in grapes, mushrooms, blueberries, passion fruit and other edible fruits and vegetables [20,21,22]. It has been proven to have anti-inflammatory, antioxidation, antibacterial, antiaging, antileukemia, anti-cell proliferation, immunomodulatory, cardiovascular protection, anticancer and other biological activities [23,24,25]. It is important for human health, nutrition and well-being. In addition, it has been shown that polyphenols can inhibit human and animal viruses. Piceatannol has an inhibitory effect on human cytomegalovirus in vitro [26]. Polyphenol stilbene compounds can inhibit HIV-1 integrase in vitro and thus inhibit HIV-1 proliferation [27]. In this study, we evaluated the inhibitory effect of piceatannol on PRV replication in vivo and in vitro and investigated the mechanism of action of piceatannol against PRV, aiming to find a new drug for the prevention of PRV infection.
2. Materials and methods
2.1. Cells, materials, and piceatannol
PK-15 cells were cultured in DMEM supplemented with 10% fetal bovine serum. The medium used for cytotoxicity and antiviral tests contained 2% fetal bovine serum. PRV-TJ (GenBank accession: KJ789182.1) was provided by Harbin Veterinary Institute of China. PRV was subpropagated in PK-15 cells, and the TCID50 was calculated by the Reed-Muench method to be 106.54/0.1 mL. The stock was stored at -80°C until use. Piceatannol was purchased from Dalian Meilun Biotechnology Co., Ltd., with a purity of 98%, and was dissolved in 1% dimethyl sulfoxide (DMSO).
2.2. Determination of cytotoxicity and inhibitory activity
Cell viability was evaluated using a CCK-8 kit to assess the cytotoxicity of piceatannol. In short, PK-15 cells in 96-well plates were treated with a continuous twofold dilution of piceatannol and incubated at 37°C and 5% CO2 for 48 h. At the same time, a group of cells without drugs was maintained as a control. After 48 h, CCK-8 solution was added according to the instructions, and the cultures were incubated at 37°C for 45 min. The OD value of each well at 450 nm was measured by an plate reader, and the cell survival rate was calculated to evaluate the cytotoxicity of piceatannol. To determine the inhibitory activity of piceatannol against PRV, different concentrations of piceatannol were mixed with an equal volume of 100 TCID50 of PRV solution at a nontoxic concentration and incubated at 37°C for 1 h. The above mixture was added to 96-well cell culture plates, which were incubated for 1 h and then washed with PBS, and cell maintenance solution was then added. Forty-eight hours later, the OD values at 450 nm of each well were determined, and the virus inhibition rate was calculated to evaluate the inhibitory activity of piceatannol against PRV. The half inhibitory concentration (IC50) and half cytotoxic concentration (CC50) of piceatannol were calculated by GraphPad Prism 8.0 software.
2.3. Effect of piceatannol on the PRV life cycle
Virus adsorption phase: PK-15 cells at 80-90% confluence were precooled at 4°C for 1 h, and 100 TCID50 of PRV and an equivolume piceatannol mixture were coincubated with cells at 4°C for 1 h. The solution was discarded, and the cells were washed twice, treated with cell maintenance solution, and cultured at 37°C and with 5% CO2. Forty hours later, cell samples were collected, and viral genomic DNA was extracted. FQ-PCR was used to determine the copy number of viral DNA in infected cells.
Virus entry phase: PK-15 cells at 80-90% confluence were precooled at 4°C for 1 h, and 100 TCID50 of PRV was used to infect cells at 4°C for 1 h. After washing the wells with precooled PBS, cell maintenance solution containing piceatannol was added, incubated at 37°C for 1 h, replaced with cell maintenance solution, and further incubated at 37°C and with 5% CO2. A virus control group was also set up. Forty hours later, cell samples were collected, and viral genomic DNA was extracted. FQ-PCR was used to determine the copy number of viral DNA in infected cells.
Virus replication phase: PRV (100 TCID50) was used to infect PK-15 cells at 37°C for 1 h, the cells were washed twice, cell maintenance solution containing piceatannol was added, the cells were cultured at 37°C and with 5% CO2, and a virus control group was established. After 40 h, the cell samples were collected, and the viral genomic DNA was extracted. The viral DNA copy number of infected cells was determined by FQ-PCR.
A TIANamp Genomic DNA Kit was used to extract the viral genomic DNA, and FQ-PCR was used to analyze the viral DNA copy number in infected cells. The reaction primers were 5’-GCCGAGTACGACCTCTGCC-3’ (forward) and 5’-CGAGACGAACAGCAGCCG-3’ (reverse), and the probe was 5’-HEX-CCGCGTGCACCACGAAGCCT-BHQ1. A standard plasmid containing the gI gene was used to generate a standard curve, and the FQ-PCR conditions were predenaturation at 94°C for 10 min, followed by 45 cycles of 94°C for 35 s and 60°C for 35 s.
2.4. Effects of piceatannol on PRV gene expression
PK-15 cells were grown to a confluent monolayer, washed twice with PBS, and treated with 100 TCID50 PRV solution and 0.063 mg/mL piceatannol mixture at 37°C for 1 h. After 1 h, the solution was discarded, the cells were washed with PBS, and then cell maintenance solution was added for continued culture. Samples of infected cells were collected at 3, 6, 12, 24, and 48 h after viral infection. Total RNA was extracted with an RNA Easy Fast Tissue/Cell Kit. According to the instructions for FastKing gDNA Dispelling RT SuperMix, reverse transcription was carried out, and qPCR primers were designed using Primer 5 software according to the PRV gene and β-actin gene sequence. The RT‒qPCR procedure was as follows: predenaturation at 95°C for 5 min, followed by 95°C for 15 s, 55°C for 30 s, and 72°C for 35 s for a total of 40 cycles. The primer sequences are shown in Table 1.
2.5. Effect of piceatannol on PRV protein expression
The 3D structure of piceatannol was downloaded from PubChem, and the 3D structures of the PRV gB (PDB ID: 5YS6) protein and PRV gD (PDB ID: 5X5V) protein were downloaded from the RCSB PDB online database. PyMOL software was used to optimize the protein structure and remove water and small molecular ligands. AutoDock was used to hydrogenate the PRV gB/gD protein molecular model. GridBox was used to wrap the whole protein molecules for molecular docking. The optimal docking results with the lowest binding energy were screened and visualized by PyMOL.
A monolayer of PK-15 cells was washed twice with PBS and then mixed with 100 TCID50 PRV solution that had been treated at 37°C for 1 h and an equal volume of 0.063 mg/mL piceatannol. After 1 h, the solution was discarded, the cells were washed with PBS, and the wash was replaced with cell maintenance solution to continue the culture. After 6 h, the cells were lysed using protein extraction buffer, and cell samples were harvested for total cellular protein extraction. Proteins were separated by 10% SDS‒PAGE and transferred to NC membranes. The membranes were blocked with 5% skim milk for 2 h at 25°C and then incubated together with different antibodies, including anti-PRV gB, anti-PRV gD and anti-β-actin antibodies, and the membranes were incubated at 4°C overnight. After washing, an HRP-conjugated goat anti-mouse IgG secondary antibody was added and incubated for 1 h at room temperature. Finally, the proteins were visualized using ECL, and the signals were analyzed using ImageJ software.
2.6. Apoptosis analysis
The samples were analyzed by an Annexin V-FITC apoptosis kit, and cell apoptosis was assessed by flow cytometry. In brief, monolayers of PK-15 cells were washed twice with PBS, and a mixture of 100 TCID50 PRV virus solution and an equal volume of 0.063 mg/mL piceatannol that had been treated at 37°C for 1 h was added. After 1 h, the solution was discarded, the cells were washed with PBS, and then the wash was replaced with cell maintenance solution for continued culture. After 6 h, the cell culture medium was collected, adherent cells were released by digestion, and the samples were centrifuged at 1000 rpm for 5 min. After washing the cell precipitates with PBS, the cells were resuspended by adding 500 μL of binding solution. Then, Annexin V-FITC and propidium iodide were added at 25°C in the dark, and the mixture was mixed. Flow cytometry was performed after passing the samples through a 200 mesh filter.
2.7. Assay of the antiviral activity of piceatannol against PRV in vivo
Thirty female Kunming mice (20 ± 2 g) were purchased from Changsheng Laboratory Animal Center. After one week of acclimated feeding in the animal laboratory of Northeast Agricultural University, the mice were randomly divided into three groups: virus control group, the normal control group and piceatannol treatment group. The mice in the piceatannol treatment group were intraperitoneally injected with 50 mg/kg piceatannol once per day for 3 days, and the mice in the normal control group and the virus control group were injected with the same amount of DMEM. On day 4, the mice in the virus control group and the piceatannol treatment group were injected intramuscularly with 0.1 mL of 100 TCID50 PRV virus solution, and the mice in the normal control group were injected with the same amount of DMEM. Two hours after injection, the mice in the piceatannol administration group were intraperitoneally injected with piceatannol at a dose of 50 mg/kg once per day for 4 days, and the mice in the normal control group and the virus control group were injected with the same volume of DMEM. The health, behavior, and body weight status of the mice were recorded daily. On the third day after challenge, three mice were randomly selected from each group for blood collection and dissection, and the heart, liver, lung, kidney and brain tissues of the mice were collected. At the same time, the remaining mice were monitored to determine the viability during the remaining experimental period. Tissue samples were homogenized in saline for DNA extraction to determine the tissue viral load. The other portion of specimens was fixed in 4% paraformaldehyde and then subjected to tissue section preparation and histopathological examination. The collected blood samples were coagulated at 4°C for 20 min and then centrifuged at 5000 r/min for 10 min to obtain serum, and the concentrations of the, TNF-α, IFN-γ, IL-6 and IL-4 cytokines in mouse serum were measured using ELISA kits.
2.8. Statistical methods
All cell experiments were repeated at least three times, and the results are presented as the mean ± standard deviation (SD). The statistical significance of the data was assessed by two-tailed Student's t test using GraphPad Prism 8.0 software. p < 0.05 was considered to indicate statistical significance. (ns: p > 0.05, * p < 0.05, ** p < 0.01, *** p < 0.001 and **** p < 0.0001)
3. Results
3.1. Cytotoxicity of piceatannol on PK-15 cells
The cytotoxicity of piceatannol on PK-15 cells was evaluated by CCK-8 assay. As shown in Figure 1, piceatannol inhibited the proliferation of PK-15 cells. With an increasing piceatannol concentration, the cell viability decreased gradually. When the concentration of piceatannol was less than 0.063 mg/mL, there was no significant difference in cell viability with the cell control group without treatment (p > 0.05). This concentration was selected as the nontoxic concentration for subsequent antiviral activity assays. The CC50 of piceatannol was 0.1129 mg/mL, as determined by nonlinear regression analysis.
3.2. Piceatannol inhibited PRV proliferation in PK-15 cells
The inhibitory activity of piceatannol against PRV in PK-15 cells was assessed by CCK-8 assay and FQ-PCR assay. The results showed that piceatannol significantly inhibited cell death induced by PRV infection in a dose-dependent manner. When the concentration of piceatannol was 0.063 mg/mL, the inhibition rate was as high as 82.78% (Figure 2B), the IC50 of piceatannol was 0.0307 mg/mL, and the selectivity index (SI, CC50/IC50) was 3.68. In the viral copy number assay, the viral copy number decreased in a dose-dependent manner in the presence of piceatannol. When the concentration of piceatannol was 0.063 mg/mL, the viral copy number decreased by 13.57-fold (Figure 2C). In conclusion, piceatannol significantly inhibited PRV proliferation in PK-15 cells.
3.3. Effect of piceatannol on the replication cycle of PRV
To further explore the effect of piceatannol on the PRV replication cycle, probe-based real-time PCR was used to determine the viral copy number in infected cells during the PRV adsorption, entry and replication phases. For adsorption assays, piceatannol and PRV were added simultaneously to the cells at 4°C. Piceatannol significantly reduced the viral copy number of PRV and inhibited its adsorption (p < 0.0001). For entry assays, cells were infected with PRV at 4 ℃ followed by the addition of maintenance medium containing piceatannol. The results showed that there was no significant difference in viral load between the treatment group and the virus control group (p > 0.05). This result indicates that piceatannol has no significant effect on virus entry. After entering the cell, the virus begins to replicate using the energy and raw materials provided by the cell. In the replication assay, piceatannol at a concentration of 0.063 mg/mL significantly reduced the viral copy number of infected cells and the replication of PRV (p < 0.05).
3.4. Inhibitory effect of piceatannol on PRV gene expression
The expression levels of the PRV immediate early gene IE180, early gene EP0 and the PRV infection-related genes UL6, US27, UL44 and UL29 were measured by real-time PCR at 3, 6, 12, 24 and 48 h after virus infection as described previously. The relative expression levels of the detected genes are shown in Figure 4. The expression levels of the tested genes in the infected cells treated with piceatannol showed an increasing trend within 48 h. That is, with increasing time, the expression of the assayed genes increased. However, compared with that in the virus control group, the expression of all tested genes was significantly inhibited by piceatannol within 48 h (P < 0.05).
3.5. Piceatannol inhibited the expression of the PRV gB and gD proteins
The results of RT-qPCR analysis showed that the expression of the UL27 (gB) and US6 (gD) genes was inhibited by piceatannol. In order to further investigate the effect of piceatannol on the gB and gD proteins, Western blotting was used to measure the expression levels of the gB and gD proteins in PRV-infected cells 36 hours after infection, and ImageJ software was used for grayscale intensity value analysis. The results of Western blotting and grayscale intensity value analysis showed that piceatannol at 0.063 mg/mL and 0.032 mg/mL could significantly inhibit the protein expression of PRV gB and gD (P < 0.05). The results further showed that piceatannol inhibited gB and gD gene transcription to inhibit protein expression and block protein synthesis (Figure 5A–D). Molecular docking technology was used to predict the binding target sites of piceatannol on the PRV gB and gD proteins. The results showed that piceatannol could bind to the PRV gB protein and form hydrogen bonds with amino acid residues GLU-659, ARG-677, TYR-668 and LYS-670 (Figure 5E), and the binding energy was -3.78 kcal/mol. Piceatannol could bind to the PRV gD protein and form hydrogen bonds with amino acid residues PRO-53, LEU-176, ARG-166, ASP-74, and SER-73 (Figure 5F), and the binding energy was -4.88 kcal/mol. The binding energy of piceatannol with the PRV gB and gD proteins was less than 0 kcal/mol, indicating that piceatannol could bind spontaneously and exhibit good binding activity.
3.6. Piceatannol alleviates PRV-induced apoptosis
The apoptosis of PK-15 cells was assessed by flow cytometry. The results showed that 36 h after viral infection, the apoptosis rate of normal control cells was 7.2% (Figure 6A), and that of PRV-infected control cells was 14% (Figure 6B). The apoptosis rate was 9.5% in the 0.063 mg/mL treatment group (Figure 6C). The results showed that PRV infection could induce the apoptosis of PK-15 cells, while leuconotaxol could inhibit apoptosis induced by PRV infection.
3.7. Piceatannol inhibits PRV infection in mice
The mice in the control group began to develop neurological symptoms on the third day after infection, showing abnormal activity, biting at the injection site, skin damage at the injection site, and bleeding. Mice died on the 3rd day after PRV challenge, and all the mice died by the 5th day, with a survival rate of 0%. The mice in the piceatannol group began to show clinical symptoms on the 3rd day after challenge and died on the 4th day, and one mouse survived to the 7th day, with a survival rate of 14.3%. All the mice in the nonchallenged normal control group survived (Figure 7A). The weight change curve of mice showed that compared with those in the normal control group, the mice in the virus control group continued to lose weight after the symptoms appeared on the third day. The mean body weight of the mice treated with piceatannol decreased at days 4 and 5 but increased significantly after day 6 (Figure 7B).
To determine the viral load in tissues and organs, three mice were randomly selected from each group, and brain, liver, kidney, lung, and heart tissues were collected and weighed. Then, total DNA was extracted from tissues and organs, and changes in the viral DNA copy number were determined by FQ-PCR. The results showed that PRV was detectable in the brain, liver, kidney, lung and heart tissues of mice in the virus control group, and the virus loading in brain tissue was the highest, followed by that in the liver, lung, kidney and heart. Compared with that of mice in the viral control group, the viral load in the brain and kidney was significantly reduced in the mice treated with piceatannol (P < 0.05), but there was no significant change in the virus loading in the heart, liver, and lung tissues (Figure 7C).
In order to further explore the protective effect of piceatannol on tissue injury caused by PRV, histopathological examination of the brain, heart, liver, lung and kidney of mice in each group was performed. The results of HE staining are shown in Figure 7D. Compared with those in the normal control group, the mice in the challenge group showed cellular edema and inflammatory cell infiltration in brain tissue. Rupture, rearrangement and slight hemorrhage of myocardial fibers were observed in the heart. The liver showed extensive hepatocyte edema, lymphocyte infiltration, and narrowing of the hepatic sinusoidal space. Pulmonary alveolar wall septal thickening and inflammatory cell infiltration were also observed. The renal tubular epithelial cells were swollen and congested, and the renal interstitium showed hemorrhage. In the mice treated with piceatannol, local lymphocyte infiltration in brain tissue, myocardial hemorrhage in the heart, inflammatory cell infiltration in the liver, and hepatocyte edema were reduced, the lung alveolar septum was thickened, and no inflammatory cell infiltration was observed. Swelling of renal epithelial cells was reduced, and a small amount of inflammatory cell infiltration was observed.
3.8. Changes in cytokine levels in the serum of PRV-infected mice treated with piceatannol
Cytokines are involved in inflammatory and immune responses and play a key role in protecting the mechanism from foreign pathogens. The serum levels of, IL-6, IFN-γ, TNF-α and IL-4 were detected by ELISA. The results are shown in Figure 8. Compared with that in the normal control mice, PRV infection caused significant increases in the serum levels of IL-6, IFN-γ and TNF-α, (P < 0.05). However, the level of the cytokine IL-4 was significantly decreased. Compared with those in the mice in the challenge control group, the levels of TNF-α, IFN-γ and IL-4 cytokines in the serum of the mice in the piceatannol treatment group were significantly increased (P < 0.05). There was no significant difference in serum IL-6 levels between the challenge control group and the piceatannol treatment group (P > 0.05). The cytokine analysis results showed that PRV infection could increase the levels of TNF-α, IFN-γ and IL-6 and reduce the level of IL-4 to regulate the body's immune response to virus infection.
4. Discussion
PRV is a widespread swine pathogen that causes severe neurological symptoms in infected pigs, reproductive disorders such as abortion and stillbirth, and ultimately death in pregnant sows [28]. Once infected with the virus, it is difficult to eliminate the virus from the pig population, which has brought tremendous economic losses to the global pig industry. During this period, PRV vaccines were also developed, but due to the continuous mutation of PRV strains, traditional vaccines cannot provide comprehensive protection, and the disease is still spreading in China [29]. The potential for a threat to humans is also increasing [30]. Accordingly, it is of great importance to public health to develop novel and effective anti-PRV infection drugs. Natural compounds are found in a variety of edible plants with a wide range of sources, and they display few toxic side effects; therefore, edible plants represent an important resource pool for searching for natural antiviral drugs. Therefore, piceatannol, which is widely present in edible plants, was chosen to study its antiherpesvirus effects.
Edible plant-derived compounds have multipathway and multitarget antiviral activities [31]. On the one hand, they can directly inhibit virus activity or replication processes, including blocking virus adsorption, entry, synthesis, mRNA replication or protein synthesis; on the other hand, they can regulate the body's immunity to exert an antiviral activity [32]. Piceatannol is a naturally occurring polyphenolic substance in plants that is widely found in grapes, mushrooms, blueberries, passion fruit and other edible fruits [33]. Studies have reported that polyphenolic compounds have a wide range of antiviral activities [34]. For example, piceatannol has an inhibitory effect on human cytomegalovirus in vitro [26]. Epigallocatechin-3-gallate (EGCG), a bioactive polyphenol found in green tea extract, can inhibit PRV infection by inhibiting viral adsorption, entry and replication [35]. EGCG also inhibited PRRSV proliferation by interfering with lipid metabolism [36]. Moreover, EGCG showed inhibitory effects against influenza A virus, herpes simplex virus and Zika virus [37,38,39]. Curcumin is an acidic polyphenolic compound isolated from Curcuma longa. Yang et al. [40] found that curcumin could protect rat hippocampal neurons against PRV infection by adjusting the BDNF/TrkB pathway. Studies have shown that curcumin can also inhibit the replication of CSFV by interfering with lipid metabolism [41]. In addition, curcumin has an inhibitory effect on infectious gastroenteritis virus, hepatitis B virus and dengue 2 virus [42,43,44]. The above findings indicate that plant-derived polyphenolic compounds have a wide range of antiviral effects.
As a polyphenolic compound, resveratrol can exert anti-PRV activity by inhibiting PRV replication, upregulating IFN-γ and TNF-α cytokine levels in the serum of piglets, reducing the pathological changes in tissues and organs of piglets, and reducing the mortality rate and viral load in the brain of piglets [45]. Resveratrol also has inhibitory effects on herpesvirus [46] and HIV-1 [47]. Zhao et al. [48] showed that resveratrol inhibited viral gene transcription and inhibited PRV replication in a dose-dependent manner. Piceatannol, a derivative of resveratrol, has one additional hydroxyl group in its chemical structure and shares similarities in its mechanism of PRV resistance. In this study, we initially evaluated the anti-PRV effect of piceatannol, which is a widely present in plant species, in vitro and in vivo. In the present study, PRV infection of PK-15 cells in vitro was used as an in vitro model to evaluate the anti-PRV effect of piceatannol. The results showed that piceatannol at 0.063 mg/mL had no significant effect on cell viability in vitro. Its CC50 value was 0.1129 mg/mL, IC50 was 0.0307 mg/mL, and SI was 3.68, which indicated low toxicity and high efficiency of this compound. The inhibition rate of PRV by piceatannol was as high as 82.78% at this concentration. Piceatannol not only inhibited PRV replication as well as resveratrol but also significantly inhibited PRV adsorption. In vivo experiments in mice also showed that piceatannol could delay the onset of disease and improve the survival rate of mice. Piceatannol has a significant inhibitory effect on PRV in vivo and in vitro.
The antiviral activity of natural compounds is mainly demonstrated by their ability to reduce viral titers or inhibit CPE [49]. FQ-PCR is a method used for the quantification of viral DNA. In this study, piceatannol significantly reduced the copy number of PRV, and the killing rate was as high as 82.78%. We then investigated the effect of piceatannol on the replication cycle of PRV. It was found that piceatannol inhibited the adsorption and replication of PRV. This mechanism is similar to the way panax notoginseng polysaccharide exerts its antiviral activity by blocking PRV attachment to the cell surface and inhibiting PRV replication [50].
In the early stage of PRV infection, PRV genes are transcribed, and some early viral proteins are expressed to further promote virus propagation [51]. Early genes are essential for progeny virus production after PRV infection [52]. PRV gD and gB glycoproteins are encoded by the US6 and UL27 genes, respectively, and are indispensable key proteins for the fusion of virions with the host cell membrane [53]. Luteolin can inhibit the proliferation and intercellular transmission of PRV by inhibiting the expression of the gB gene and protein [54]. In this study, we found that piceatannol could also inhibit the expression of the gD and gB genes and the synthesis of the gB and gD proteins of PRV, thereby inhibiting virus proliferation. The expression levels of the PRV immediate early gene IE180 and early gene EP0 and the PRV infection-related genes UL6, US27, UL44 and UL29 were measured by RT‒qPCR. The results showed that piceatannol could reduce the expression level of viral genes. This is consistent with the findings of Men et al. [54].
Innate immunity is an animal's first line of defense against viral invasion. Apoptosis, as a cellular defense mechanism, plays a significant role in preventing virus diffusion in the early stage of virus infection, while it enhances virus replication and release in the late stage of virus infection [55]. Viral infection usually leads to apoptosis and inflammation [56]. Therefore, many viruses promote the production of antiapoptotic factors in the early prriod of infection to delay the death of cells, increase the production of virus, and promote the release of more progeny viruses in the late stage of infection [57]. Studies have shown that both curcumin and myricetin can effectively inhibit viral infection by blocking PRV-induced apoptosis [58,59]. In this study, flow cytometry was used to examine the effect of piceatannol on host cell apoptosis at the late stage of PRV infection. The results confirmed that PRV infection could induce apoptosis of host cells, and piceatannol could inhibit PRV-induced apoptosis by inhibiting apoptosis, thereby limiting the production of progeny virions and inhibiting virus proliferation.
Studies have shown good antiviral activity of piceatannol in vitro. Therefore, we carried out an in vivo experimental study. Mice artificially infected with PRV were used as the animal model. The PRV inhibitory activity of piceatannol in mice was evaluated by administration before and after challenge. The experiment showed that piceatannol had a protective effect on PRV-infected mice. Specifically, compared with those in the untreated virus control group, piceatannol administration delayed the onset of clinical symptoms, prolonged the average survival time, and increased the viability of infected mice. These effects may be due to drug slowing the rate of virus proliferation in mice. Viral load is an important direct parameter used to evaluate antiviral effects in vivo [60]. It can reflect the replication of the virus in different organs [61]. In this study, the highest viral load was found in the brains of the virus control mice, which may be related to the neurotropic properties of alphaherpesviruses, which can invade the center nerve system through the trigeminal and sympathetic nerves [62]. Treatment with piceatannol significantly reduced the viral load in the brain and kidney tissues of the infected mice. Piceatannol reduces renal epithelial cell injury and has a neuroprotective effect [63]. We hypothesized that piceatannol could inhibit the replication of PRV in mouse organs. Meanwhile, the antiviral effect of piceatannol on PRV-infected mice was also confirmed by histopathological observation.
The inflammatory response is the first line of defense against the spread of viral infections. However, ensuring that the inflammation is resolved is necessary to prevent damage to the body. When the body's inflammation is not resolved and not controlled, this condition will usually cause greater damage to the body [64]. After viral infection, the body regulates the expression of cytokines such as interferons, tumor necrosis factor, and interleukins through nonspecific immunity to protect cells from viral infection. IFN-γ can activate macrophages, promote the production of the inflammatory factors TNF-α, IL-1β, and IL-12, and regulate the body’s immunity [65]. TNF-α, IFN-γ and IL-6, as important proinflammatory cytokines, can help the body fight viral infection and prevent tissue damage by regulating the inflammatory response [66,67]. Therefore, in the present study, the serum levels of IFN-γ, IL-6 and TNF-α in mice were increased after challenge, indicating that the innate immunity of mice was activated by PRV. Treatment with piceatannol significantly increased the serum levels of IFN-γ and TNF-α in the infected mice, suggesting that piceatannol could enhance the inflammatory response and inhibit virus replication at the early stage of virus infection by increasing the levels of TNF-α and IFN-γ. As an anti-inflammatory cytokine, IL-4 is mainly produced by Th2 cells. In the immune system, IL-4 not only regulates the immune function of macrophages but also plays a significant role in promoting the development of Th2 cells and inhibiting the growth of Th1 cells. In this study, the IL-4 cytokine level in the serum of infected mice was decreased, and it was significantly increased after treatment with piceatannol. We hypothesized that piceatannol could modulate the Th1/Th2 balance by regulating IL-4 cytokine levels and then inhibit PRV infection.
With the progress and development of science and technology, an increasing number of antiviral components of edible plants will be discovered and scientifically verified. Piceatannol, a natural compound, can be used as a food additive in food or animal feed at safe concentrations to prevent and treat viral diseases. In the future, the development of polyphenols as functional foods should be explored to further explore the antiviral mechanisms of natural compounds in edible plants.
5. Conclusion
Piceatannol showed significant anti-PRV activity both in vitro and in vivo. Piceatannol could exert an anti-PRV effect by reducing the transcription level of viral genes and the expression of the viral gB and gD proteins, inhibiting PRV-induced apoptosis and elevating the levels of IL-4, TNF-α and IFN-γ in the serum of mice. Animal experiments showed that piceatannol could delay the onset of disease, reduce the viral load in the brain and kidney, and reduce the pathological changes in the tissues and organs of the mice to improve the survival rate of the mice (14.3%). In summary, piceatannol is expected to become a commercial antiherpesvirus drug due to its low toxicity, high efficiency and wide availability.
Author Contributions
ZW wrote the manuscript. ZW, XC and ZR carried out experiment design and research. YS , YX and LF carried out statistical analysis of the date. YZ conducted experiments and provided funding. All authors read and approved the final manuscript.
Funding
This work was supported by The Central Government Guides Local Science and Technology Development Fund of China (ZY2022A-HRB-06).
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Review Board of NEAUEC2023 01 14.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, and further inquiries can be directed to the corresponding author(s).
Conflicts of Interest
The authors declare no conflict of interest.
Ethics Statement
The animal study was reviewed and approved by the Laboratory Animal Ethics Committee of Northeast Agricultural University.
Sample Availability
Not applicable.
References
- Masot:, A.J.; Gil, M.; Risco, D.; Jimenez, O.M.; Nunez, J.I.; Redondo, E. Pseudorabies virus infection (Aujeszky's disease) in an Iberian lynx (Lynx pardinus) in Spain: a case report. Bmc Vet. Res. 2017, 13, 6. [Google Scholar] [CrossRef] [PubMed]
- Katayama, S.; Okada, N.; Ohgitani, T.; Kokubu, T.; Shimizu, Y. Influence of cell surface glycoprotein gC produced by pseudorabies virus on cytopathic effect. J. Vet. Med. Sci. 1998, 60, 905–909. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wang, X.; Xie, C.; Ding, S.; Yang, H.; Guo, S.; Li, J.; Qin, L.; Ban, F.; Wang, D.; et al. A novel human acute encephalitis caused by pseudorabies virus variant strain. Clinical Infectious Diseases : An Official Publication of the Infectious Diseases Society of America. 2020, 73.
- Wang, D.; Tao, X.; Fei, M.; Chen, J.; Guo, W.; Li, P.; Wang, J. Human encephalitis caused by pseudorabies virus infection: a case report. J. Neurovirol. 2020, 26. [Google Scholar] [CrossRef] [PubMed]
- Ghidoli, M.; Colombo, F.; Sangiorgio, S.; Landoni, M.; Giupponi, L.; Nielsen, E.; Pilu, R. Food Containing Bioactive Flavonoids and Other Phenolic or Sulfur Phytochemicals With Antiviral Effect: Can We Design a Promising Diet Against COVID-19? Front. Nutr. 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Casado, A. The Health Potential of Fruits and Vegetables Phytochemicals: Notable Examples. Crit. Rev. Food Sci. Nutr. 2016, 56. [Google Scholar] [CrossRef]
- Maria, R.; Stefania, M.; Carmela, S.; Idolo, T.; Gian, L.R. Roles of flavonoids against coronavirus infection. Chem.-Biol. Interact. 2020, 328.
- Julia, S.; Johannes-Paul, F. Flavonoids: A complementary approach to conventional therapy of COVID-19? Phytochemistry Reviews : Proceedings of the Phytochemical Society of Europe. 2020.
- Mohammadi, P.P.; Fakhri, S.; Asgary, S.; Farzaei, M.H.; Echeverría, J. The Signaling Pathways, and Therapeutic Targets of Antiviral Agents: Focusing on the Antiviral Approaches and Clinical Perspectives of Anthocyanins in the Management of Viral Diseases. Front. Pharmacol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Harazem, R.; Rahman, S.; Kenawy, A. Evaluation of Antiviral Activity of Allium Cepa and Allium Sativum Extracts Against Newcastle Disease Virus. Alexandria Journal of Veterinary Sciences. 2019, 61. [Google Scholar] [CrossRef]
- Alex, H.; Andrea, T.; Berlin, L.; Tonya, M.C.; Curt, H. Garlic Organosulfur Compounds Reduce Inflammation and Oxidative Stress during Dengue Virus Infection. Viruses-Basel. 2017, 9.
- Josling, P. Preventing the common cold with a garlic supplement: a double-blind, placebo-controlled survey. Adv. Ther. 2001, 18. [Google Scholar] [CrossRef]
- Bayan, L.; Koulivand, P.H.; Gorji, A. Garlic: a review of potential therapeutic effects. Avicenna J. Phytomedicine. 2014, 4. [Google Scholar]
- Ueda, K.; Kawabata, R.; Irie, T.; Nakai, Y.; Tohya, Y.; Sakaguchi, T. Inactivation of pathogenic viruses by plant-derived tannins: strong effects of extracts from persimmon (Diospyros kaki) on a broad range of viruses. Plos One. 2013, 8. [Google Scholar] [CrossRef]
- J., L.S.G.; R., C.L.; D., B.O.; S., S.C.; M., M.F.S.M.; M., T.V.R.; N., S.O.S.; M., D.W. In vitro anti-rotavirus activity of some medicinal plants used in Brazil against diarrhea. J. Ethnopharmacol. 2005, 99.
- Zhi-Yong, J.; Wen-Feng, L.; Xue-Mei, Z.; Jie, L.; Yun-Bao, M.; Ji-Jun, C. Anti-HBV active constituents from Piper longum. Bioorg. Med. Chem. Lett. 2013, 23. [Google Scholar]
- Nongluk, S.; Syuichi, F.; Kenji, K.; Hiroaki, H.; Takato, O.; Masato, T.; Yasuo, S. Antiviral effects of Psidium guajava Linn. (guava) tea on the growth of clinical isolated H1N1 viruses: Its role in viral hemagglutination and neuraminidase inhibition. Antiviral Res. 2012, 94.
- Sumra, N.; Shabbir, H.; Naureen, N.; Muhammad, P.; Madiha, R. The phytochemistry and medicinal value of Psidium guajava (guava). Clinical Phytoscience. 2018, 4. [Google Scholar]
- Prochazkova, D.; Bousova, I.; Wilhelmova, N. Antioxidant and prooxidant properties of flavonoids. Fitoterapia. 2011, 82, 513–523. [Google Scholar] [CrossRef]
- Matsui, Y.; Sugiyama, K.; Kamei, M.; Takahashi, T.; Suzuki, T.; Katagata, Y.; Ito, T. Extract of passion fruit (Passiflora edulis) seed containing high amounts of piceatannol inhibits melanogenesis and promotes collagen synthesis. J. Agric. Food Chem. 2010, 58. [Google Scholar] [CrossRef] [PubMed]
- Hanna, P.; Malgorzata, K.; Marek, M. Biological activity of piceatannol: Leaving the shadow of resveratrol. Mutat. Res.-Rev. Mutat. Res. 2011, 750.
- Efstathios, P.V.; Athanassios, K.; Ioannis, G.; Dimitrios, S.; Marilena, E.L. Screening of mushrooms bioactivity: piceatannol was identified as a bioactive ingredient in the order Cantharellales. Eur. Food Res. Technol. 2018, 244. [Google Scholar]
- Shah, U.; Shah, R.; Acharya, S.; Acharya, N. Novel anticancer agents from plant sources:Novel anticancer agents from plant sources. Chin. J. Nat. Med. 2014, 11. [Google Scholar]
- Rimando, A.M.; Kalt, W.; Magee, J.B.; Dewey, J.; Ballington, J.R. Resveratrol, pterostilbene, and piceatannol in vaccinium berries. J. Agric. Food Chem. 2004, 52, 4713–4719. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Smith, W.; Yan, L.; Kong, L. Overview of Cellular Mechanisms and Signaling Pathways of Piceatannol. Curr. Stem Cell Res. Ther. 2020, 15. [Google Scholar]
- Wang, S.Y.; Zhang, J.; Xu, X.G.; Su, H.L.; Xing, W.M.; Zhang, Z.S.; Jin, W.H.; Dai, J.H.; Wang, Y.Z.; He, X.Y.; et al. Inhibitory effects of piceatannol on human cytomegalovirus (hCMV) in vitro. J. Microbiol. 2020, 58, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Pflieger, A.; Waffo, T.P.; Papastamoulis, Y.; Chaignepain, S.; Subra, F.; Munir, S.; Delelis, O.; Lesbats, P.; Calmels, C.; Andreola, M.L.; et al. Natural stilbenoids isolated from grapevine exhibiting inhibitory effects against HIV-1 integrase and eukaryote MOS1 transposase in vitro activities. Plos One. 2013, 8, e81184. [Google Scholar] [CrossRef]
- Li, A.; Lu, G.; Qi, J.; Wu, L.; Tian, K.; Luo, T.; Shi, Y.; Yan, J.; Gao, G.F. Structural basis of nectin-1 recognition by pseudorabies virus glycoprotein D. Plos Pathog. 2017, 13, e1006314. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.H.; Fu, P.F.; Chen, H.Y.; Wang, Z.Y. Pseudorabies Virus: From Pathogenesis to Prevention Strategies. Viruses-Basel. 2022, 14.
- He, W.; Auclert, L.Z.; Zhai, X.; Wong, G.; Zhang, C.; Zhu, H.; Xing, G.; Wang, S.; He, W.; Li, K.; et al. Interspecies Transmission, Genetic Diversity, and Evolutionary Dynamics of Pseudorabies Virus. J. Infect. Dis. 2019, 219, 1705–1715. [Google Scholar] [CrossRef] [PubMed]
- Dey, R.; Samadder, A.; Nandi, S. Exploring the Targets of Novel Corona Virus and Docking-based Screening of Potential Natural Inhibitors to Combat COVID-19. Curr. Top. Med. Chem. 2022, 22, 2410–2434. [Google Scholar] [CrossRef]
- Chen, Z.; Ye, S. Research progress on antiviral constituents in traditional Chinese medicines and their mechanisms of action. Pharm. Biol. 2022, 60. [Google Scholar] [CrossRef]
- Rossi, M.; Caruso, F.; Opazo, C.; Salciccioli, J. Crystal and molecular structure of piceatannol; scavenging features of resveratrol and piceatannol on hydroxyl and peroxyl radicals and docking with transthyretin. J. Agric. Food Chem. 2008, 56, 10557–10566. [Google Scholar] [CrossRef]
- Zakaria, M.Y.; Abd, E.S.M.; Beshay, B.Y.; Zaki, I.; Abourehab, M.A.S. 'Poly phenolic phytoceutical loaded nano-bilosomes for enhanced caco-2 cell permeability and SARS-CoV 2 antiviral activity': in-vitro and insilico studies. Drug Deliv. 2023, 30. [Google Scholar] [CrossRef] [PubMed]
- Huan, C.; Xu, W.; Guo, T.; Pan, H.; Zou, H.; Jiang, L.; Li, C.; Gao, S. (-)-Epigallocatechin-3-Gallate Inhibits the Life Cycle of Pseudorabies Virus In Vitro and Protects Mice Against Fatal Infection. Front. Cell. Infect. Microbiol. 2020, 10, 616895. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.W.; Fu, P.F.; Zeng, L.; Qi, Y.L.; Li, X.Q.; Wang, Q.; Yang, G.Y.; Li, H.W.; Wang, J.; Chu, B.B.; et al. EGCG Restricts PRRSV Proliferation by Disturbing Lipid Metabolism. Microbiol. Spectr. 2022, 10, e227621. [Google Scholar] [CrossRef]
- Mou, Q.; Jiang, Y.; Zhu, L.; Zhu, Z.; Ren, T. EGCG induces beta-defensin 3 against influenza A virus H1N1 by the MAPK signaling pathway. Exp. Ther. Med. 2020, 20, 3017–3024. [Google Scholar]
- Stamos, J.D.; Lee, L.H.; Taylor, C.; Elias, T.; Adams, S.D. In Vitro and In Silico Analysis of the Inhibitory Activity of EGCG-Stearate against Herpes Simplex Virus-2. Microorganisms. 2022, 10. [Google Scholar] [CrossRef]
- Carneiro, B.M.; Batista, M.N.; Braga, A.; Nogueira, M.L.; Rahal, P. The green tea molecule EGCG inhibits Zika virus entry. Virology. 2016, 496, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Luo, G.; Zhang, C.; Feng, L.; Luo, X.; Gan, L. Curcumin protects rat hippocampal neurons against pseudorabies virus by regulating the BDNF/TrkB pathway. Sci Rep. 2020, 10, 22204. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Hu, J.H.; Liang, X.D.; Chen, J.; Liu, C.C.; Liu, Y.Y.; Cheng, Y.; Go, Y.Y.; Zhou, B. Curcumin inhibits classical swine fever virus replication by interfering with lipid metabolism. Vet. Microbiol. 2021, 259, 109152. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, J.; Liu, Y.; Luo, X.; Lei, W.; Xie, L. Antiviral and virucidal effects of curcumin on transmissible gastroenteritis virus in vitro. The Journal of General Virology. 2020, 101. [Google Scholar] [CrossRef]
- Wei, Z.; Zhang, Y.; Ke, C.; Chen, H.; Ren, P.; He, Y.; Hu, P.; Ma, D.; Luo, J.; Meng, Z. Curcumin inhibits hepatitis B virus infection by down-regulating cccDNA-bound histone acetylation. World J. Gastroenterol. 2017, 23. [Google Scholar] [CrossRef]
- Padilla-S, L.; Rodriguez, A.; Gonzales, M.M.; Gallego-G, J.C.; Castano-O, J.C. Inhibitory effects of curcumin on dengue virus type 2-infected cells in vitro. Arch. Virol. 2014, 159, 573–579. [Google Scholar] [CrossRef]
- Zhao, X.; Tong, W.; Song, X.; Jia, R.; Li, L.; Zou, Y.; He, C.; Liang, X.; Lv, C.; Jing, B.; et al. Antiviral Effect of Resveratrol in Piglets Infected with Virulent Pseudorabies Virus. Viruses-Basel. 2018, 10.
- Zhao, X.; Xu, J.; Song, X.; Jia, R.; Yin, Z.; Cheng, A.; Jia, R.; Zou, Y.; Li, L.; Yin, L.; et al. Antiviral effect of resveratrol in ducklings infected with virulent duck enteritis virus. Antiviral Res. 2016, 130, 93–100. [Google Scholar] [CrossRef]
- Krishnan, V.; Zeichner, S.L. Host cell gene expression during human immunodeficiency virus type 1 latency and reactivation and effects of targeting genes that are differentially expressed in viral latency. J. Virol. 2004, 78, 9458–9473. [Google Scholar] [CrossRef]
- Zhao, X.; Cui, Q.; Fu, Q.; Song, X.; Jia, R.; Yang, Y.; Zou, Y.; Li, L.; He, C.; Liang, X.; et al. Antiviral properties of resveratrol against pseudorabies virus are associated with the inhibition of IκB kinase activation. Sci Rep. 2017, 7. [Google Scholar] [CrossRef]
- Jiao, X.; Zhongqiong, Y.; Li, L.; Anchun, C.; Renyong, J.; Xu, S.; Hongke, L.; Shujun, D.; Cheng, L.; Xiaoxia, L.; et al. Inhibitory effect of resveratrol against duck enteritis virus in vitro. Plos One. 2017, 8. [Google Scholar]
- Huan, C.; Zhou, Z.; Yao, J.; Ni, B.; Gao, S. The Antiviral Effect of Panax Notoginseng Polysaccharides by Inhibiting PRV Adsorption and Replication In Vitro. Molecules. 2022, 27. [Google Scholar] [CrossRef]
- Johnson, D.C.; Baines, J.D. Herpesviruses remodel host membranes for virus egress. Nature Reviews. Microbiology. 2011, 9. [Google Scholar] [CrossRef] [PubMed]
- Pomeranz, L.E.; Reynolds, A.E.; Hengartner, C.J. Molecular biology of pseudorabies virus: impact on neurovirology and veterinary medicine. Microbiology and Molecular Biology Reviews : Mmbr. 2005, 69.
- Böhm, S.W.; Backovic, M.; Klupp, B.G.; Rey, F.A.; Mettenleiter, T.C.; Fuchs, W. Functional Characterization of Glycoprotein H Chimeras Composed of Conserved Domains of the Pseudorabies Virus and Herpes Simplex Virus 1 Homologs. J. Virol. 2016, 90. [Google Scholar] [CrossRef] [PubMed]
- Men, X.; Li, S.; Cai, X.; Fu, L.; Shao, Y.; Zhu, Y. Antiviral Activity of Luteolin against Pseudorabies Virus In Vitro and In Vivo. Animals. 2023, 13. [Google Scholar] [CrossRef]
- Ye, G.; Liu, H.; Zhou, Q.; Liu, X.; Huang, L.; Weng, C. A Tug of War: Pseudorabies Virus and Host Antiviral Innate Immunity. Viruses-Basel. 2022, 14.
- Liu, L. Fields Virology, 6th Edition. Clin. Infect. Dis. 2014, 59. [Google Scholar] [CrossRef]
- Teodoro, J.G.; Branton, P.E. Regulation of apoptosis by viral gene products. J. Virol. 1997, 71, 1739–1746. [Google Scholar] [CrossRef]
- Yang, B.; Luo, G.; Zhang, C.; Feng, L.; Luo, X.; Gan, L. Curcumin protects rat hippocampal neurons against pseudorabies virus by regulating the BDNF/TrkB pathway. Sci Rep. 2020, 10. [Google Scholar] [CrossRef]
- Hu, H.; Hu, Z.; Zhang, Y.; Wan, H.; Yin, Z.; Li, L.; Liang, X.; Zhao, X.; Yin, L.; Ye, G.; et al. Myricetin inhibits pseudorabies virus infection through direct inactivation and activating host antiviral defense13. Front. Microbiol. 2022, 13. [Google Scholar]
- Maresch, C.; Lange, E.; Teifke, J.P.; Fuchs, W.; Klupp, B.; Muller, T.; Mettenleiter, T.C.; Vahlenkamp, T.W. Oral immunization of wild boar and domestic pigs with attenuated live vaccine protects against Pseudorabies virus infection. Vet. Microbiol. 2012, 161, 20–25. [Google Scholar] [CrossRef]
- Cai, X.; Shao, Y.; Wang, Z.; Xu, Y.; Ren, Z.; Fu, L.; Zhu, Y. Antiviral activity of dandelion aqueous extract against pseudorabies virus both in vitro and in vivo. Front. Vet. Sci. 2022, 9, 1090398. [Google Scholar] [CrossRef]
- Brittle, E.E.; Reynolds, A.E.; Enquist, L.W. Two modes of pseudorabies virus neuroinvasion and lethality in mice. J. Virol. 2004, 78, 12951–12963. [Google Scholar] [CrossRef]
- Zhang, S.; Ye, J.; Wang, L.; Zhong, X.; Zou, X.; Qiu, F.; Huang, Z. Piceatannol protects rat neuron cells from oxygen-glucose deprivation reperfusion injury via regulation of GSK-3beta/Nrf2 signaling pathway. Zhejiang Da Xue Xue Bao Yi Xue Ban. 2022, 51, 552–562. [Google Scholar] [PubMed]
- Jingyun, W.; Yanmei, M.; Long, W.; Xiaojuan, C.; Ruoxiang, Y.; Song, W.; Xinxin, L.; Xiaoyong, C.; Wenhan, S.; Ji-Long, C. Alpha/beta interferon receptor deficiency in mice significantly enhances susceptibility of the animals to pseudorabies virus infection. Vet. Microbiol. 2017, 203. [Google Scholar]
- Wang, F.; Zhang, S.; Jeon, R.; Vuckovic, I.; Jiang, X.; Lerman, A.; Folmes, C.D.; Dzeja, P.D.; Herrmann, J. Interferon Gamma Induces Reversible Metabolic Reprogramming of M1 Macrophages to Sustain Cell Viability and Pro-Inflammatory Activity. Ebiomedicine. 2018, 30, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.M.; Wolcott, R.M.; Chervenak, R.; Jennings, S.R. Control of acute cutaneous herpes simplex virus infection: T cell-mediated viral clearance is dependent upon interferon-gamma (IFN-gamma). Virology. 1994, 202, 76–88. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harbor Perspect. Biol. 2014, 6, a16295. [Google Scholar] [CrossRef]
Figure 1.
CCK-8 assay was used to determine the toxicity of piceatannol to PK-15 cells at 0.031, 0.063, 0.125, 0.25, 0.5 and 1 mg/mL. Cell viability is expressed as a percentage of the cell viability in the control group (ns P > 0.05, **** P < 0.0001, compared with the control without piceatannol treatment).
Figure 1.
CCK-8 assay was used to determine the toxicity of piceatannol to PK-15 cells at 0.031, 0.063, 0.125, 0.25, 0.5 and 1 mg/mL. Cell viability is expressed as a percentage of the cell viability in the control group (ns P > 0.05, **** P < 0.0001, compared with the control without piceatannol treatment).
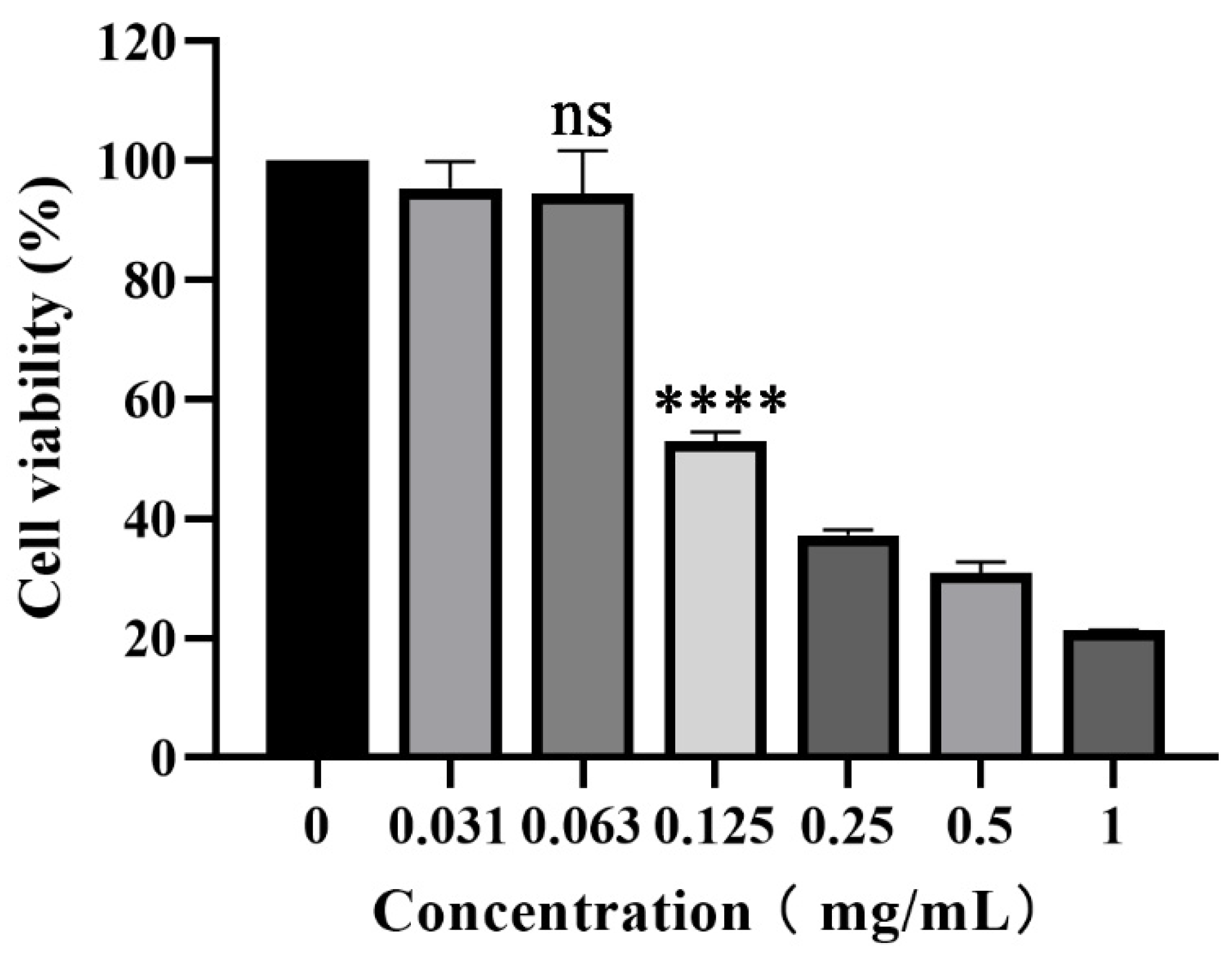
Figure 2.
Piceatannol inhibited PRV proliferation in PK-15 cells. (A) Chemical structure of piceatannol. (B) Rate of PRV inhibition by piceatannol. A CCK8 assay was used to detect the viability of PK-15 cells to calculate the rate of PRV inhibition by piceatannol. Inhibition rate (%) = (piceatannol test group OD450 value - virus control group OD450 value)/(cell control group OD450 value - virus control group OD450 value) × 100%; (C) FQ-PCR was used to measure the viral DNA copy number in PRV-infected cells treated with piceatannol. (ns P > 0.05, *** P < 0.001, **** P < 0.0001, compared with the control without piceatannol treatment).
Figure 2.
Piceatannol inhibited PRV proliferation in PK-15 cells. (A) Chemical structure of piceatannol. (B) Rate of PRV inhibition by piceatannol. A CCK8 assay was used to detect the viability of PK-15 cells to calculate the rate of PRV inhibition by piceatannol. Inhibition rate (%) = (piceatannol test group OD450 value - virus control group OD450 value)/(cell control group OD450 value - virus control group OD450 value) × 100%; (C) FQ-PCR was used to measure the viral DNA copy number in PRV-infected cells treated with piceatannol. (ns P > 0.05, *** P < 0.001, **** P < 0.0001, compared with the control without piceatannol treatment).
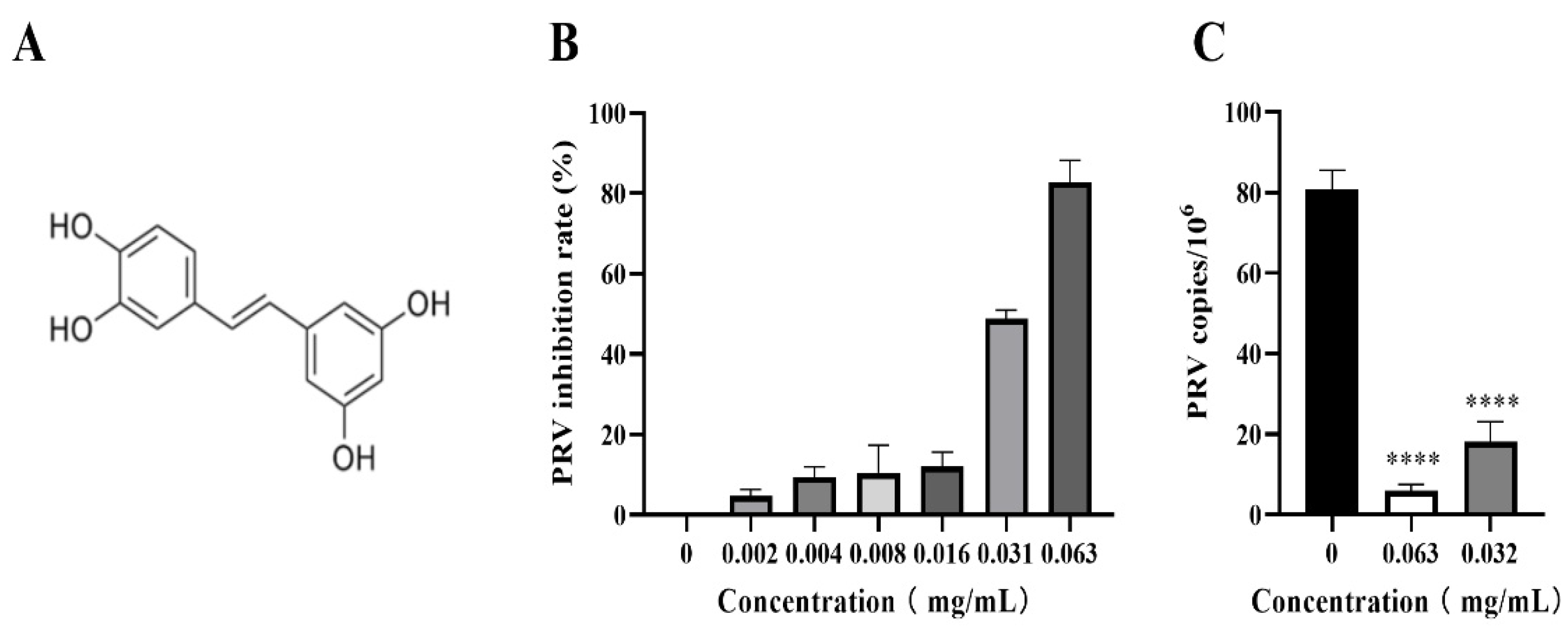
Figure 3.
Piceatannol inhibited the PRV adsorption and replication phases (ns P > 0.05, * P < 0.05, ** P < 0.01, and **** P < 0.0001).
Figure 3.
Piceatannol inhibited the PRV adsorption and replication phases (ns P > 0.05, * P < 0.05, ** P < 0.01, and **** P < 0.0001).
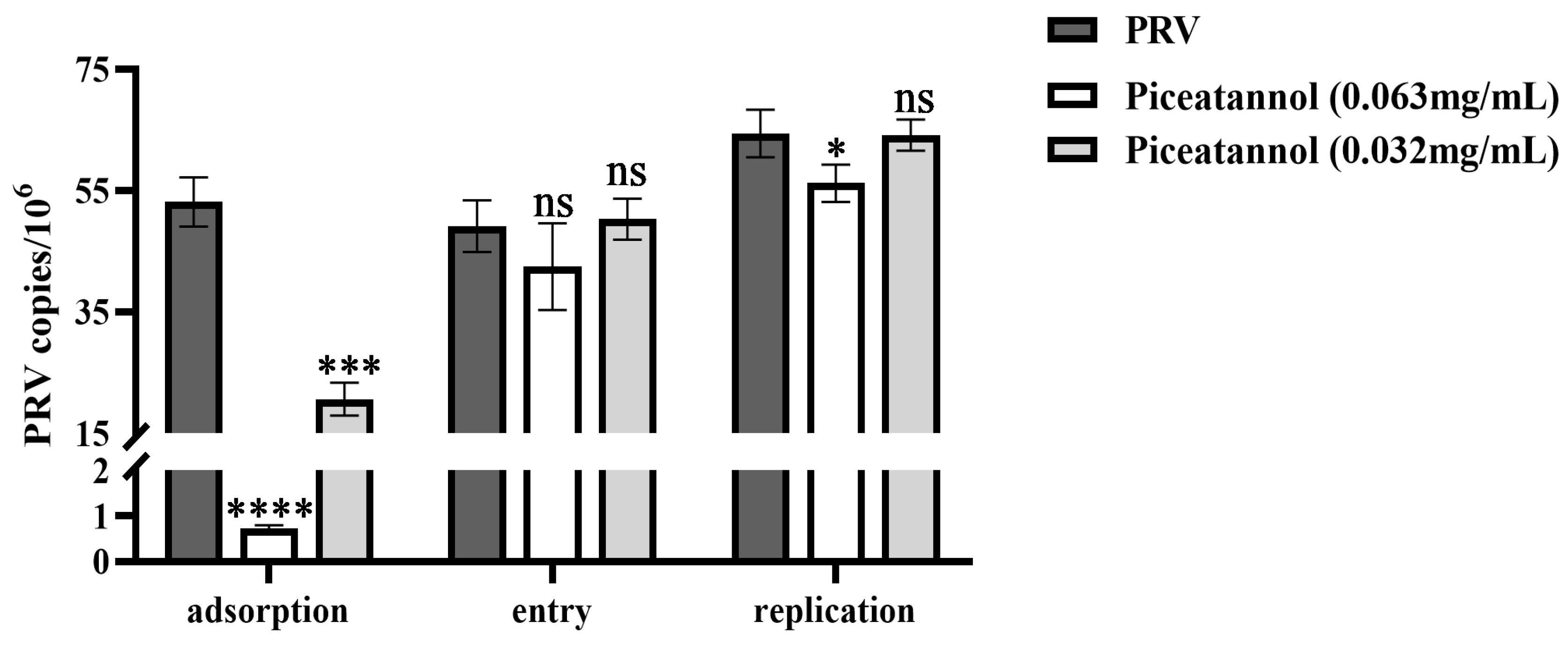
Figure 4.
Piceatannol inhibited PRV gene expression Gene expression levels of PRV in the presence or absence of piceatannol (0.063 mg/mL) at 3, 6, 12, 24 and 48 hpi (** P < 0.01 and **** P < 0.0001).
Figure 4.
Piceatannol inhibited PRV gene expression Gene expression levels of PRV in the presence or absence of piceatannol (0.063 mg/mL) at 3, 6, 12, 24 and 48 hpi (** P < 0.01 and **** P < 0.0001).
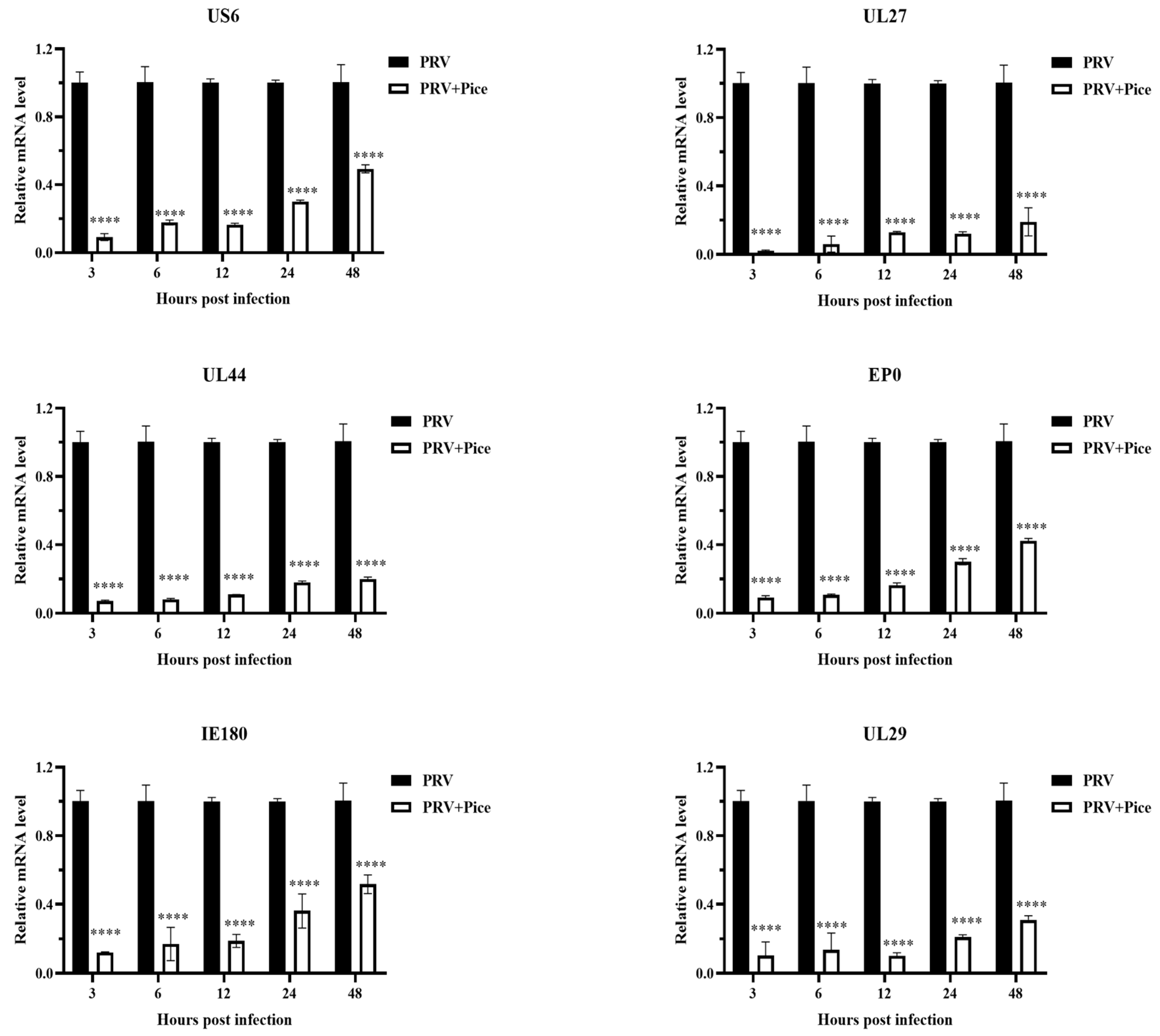
Figure 5.
Piceatannol inhibited the protein expression of PRV gB and gD PK15 cells treated with or without piceatannol after PRV infection. After 36 h of culture, cells were harvested and subjected to protein extraction for Western blotting to calculate the relative expression of the (A) gB protein and (C) gD protein. (B) Quantification of gB protein grayscale intensity value; (D) quantification of gD protein grayscale intensity value; (E) the best binding results between the PRV gB protein and piceatannol molecule; (F) optimal binding of the PRV gD protein to the piceatannol molecule. The green bars represent piceatannol, and the PRV gB protein is represented by a cartoon. Key amino acid residues are shown as rods and labeled. Hydrogen bonds formed between piceatannol and the gB and gD amino acids are indicated by yellow discontinuous lines (** P< 0.01, **** P < 0.0001).
Figure 5.
Piceatannol inhibited the protein expression of PRV gB and gD PK15 cells treated with or without piceatannol after PRV infection. After 36 h of culture, cells were harvested and subjected to protein extraction for Western blotting to calculate the relative expression of the (A) gB protein and (C) gD protein. (B) Quantification of gB protein grayscale intensity value; (D) quantification of gD protein grayscale intensity value; (E) the best binding results between the PRV gB protein and piceatannol molecule; (F) optimal binding of the PRV gD protein to the piceatannol molecule. The green bars represent piceatannol, and the PRV gB protein is represented by a cartoon. Key amino acid residues are shown as rods and labeled. Hydrogen bonds formed between piceatannol and the gB and gD amino acids are indicated by yellow discontinuous lines (** P< 0.01, **** P < 0.0001).
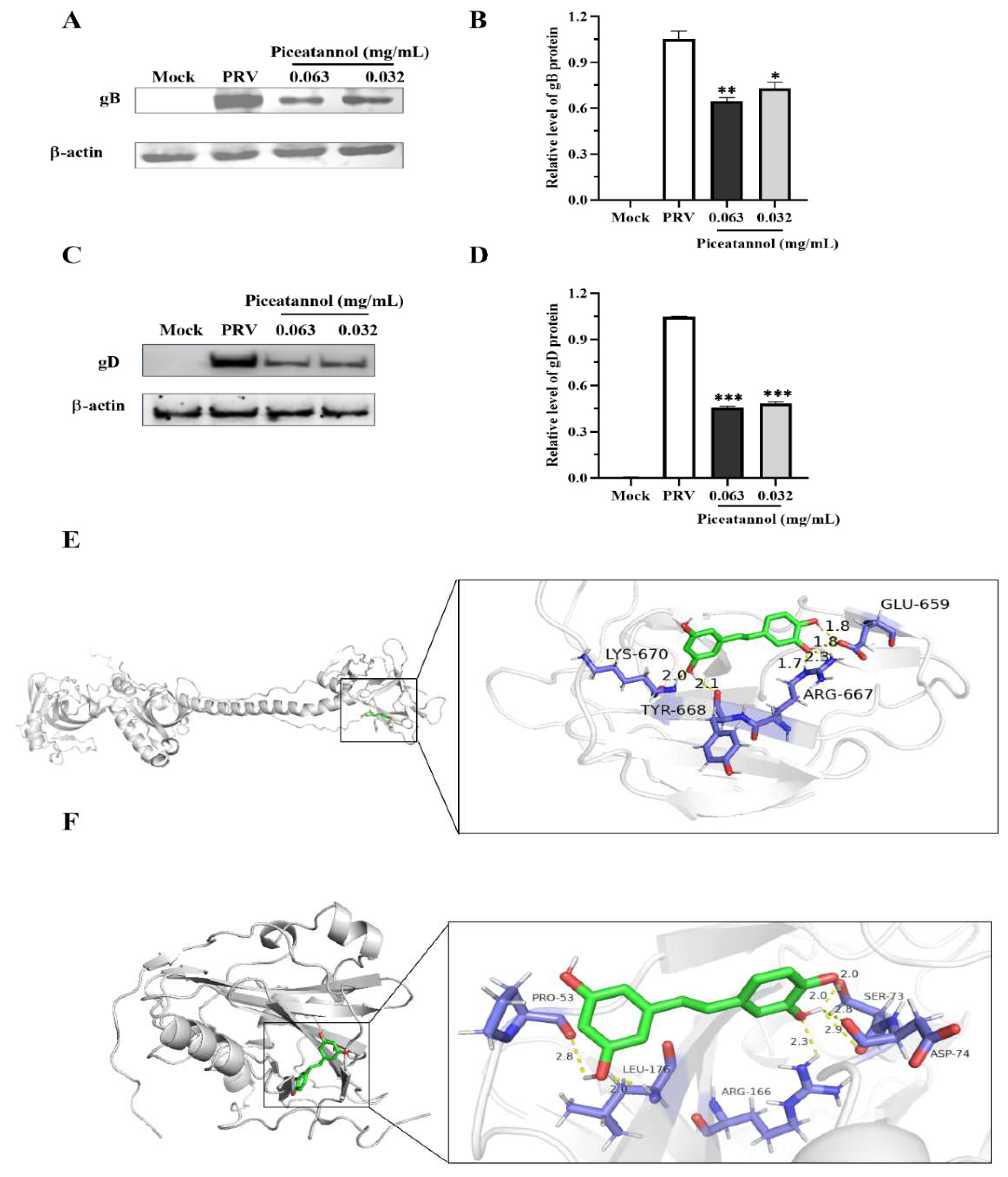
Figure 6.
Piceatannol attenuated PRV infection-induced apoptosis. After treatment with piceatannol (0.0625 mg/mL) for 36 h, the cells were stained with Annexin-V and PI and analyzed by flow cytometry. (A) Apoptosis rate in the cell control group; (B) apoptosis rate in the virus control group; (C) cell apoptosis rate of the piceatannol experimental group; (D) Bar chart of cell apoptosis in each group (*** P < 0.001).
Figure 6.
Piceatannol attenuated PRV infection-induced apoptosis. After treatment with piceatannol (0.0625 mg/mL) for 36 h, the cells were stained with Annexin-V and PI and analyzed by flow cytometry. (A) Apoptosis rate in the cell control group; (B) apoptosis rate in the virus control group; (C) cell apoptosis rate of the piceatannol experimental group; (D) Bar chart of cell apoptosis in each group (*** P < 0.001).
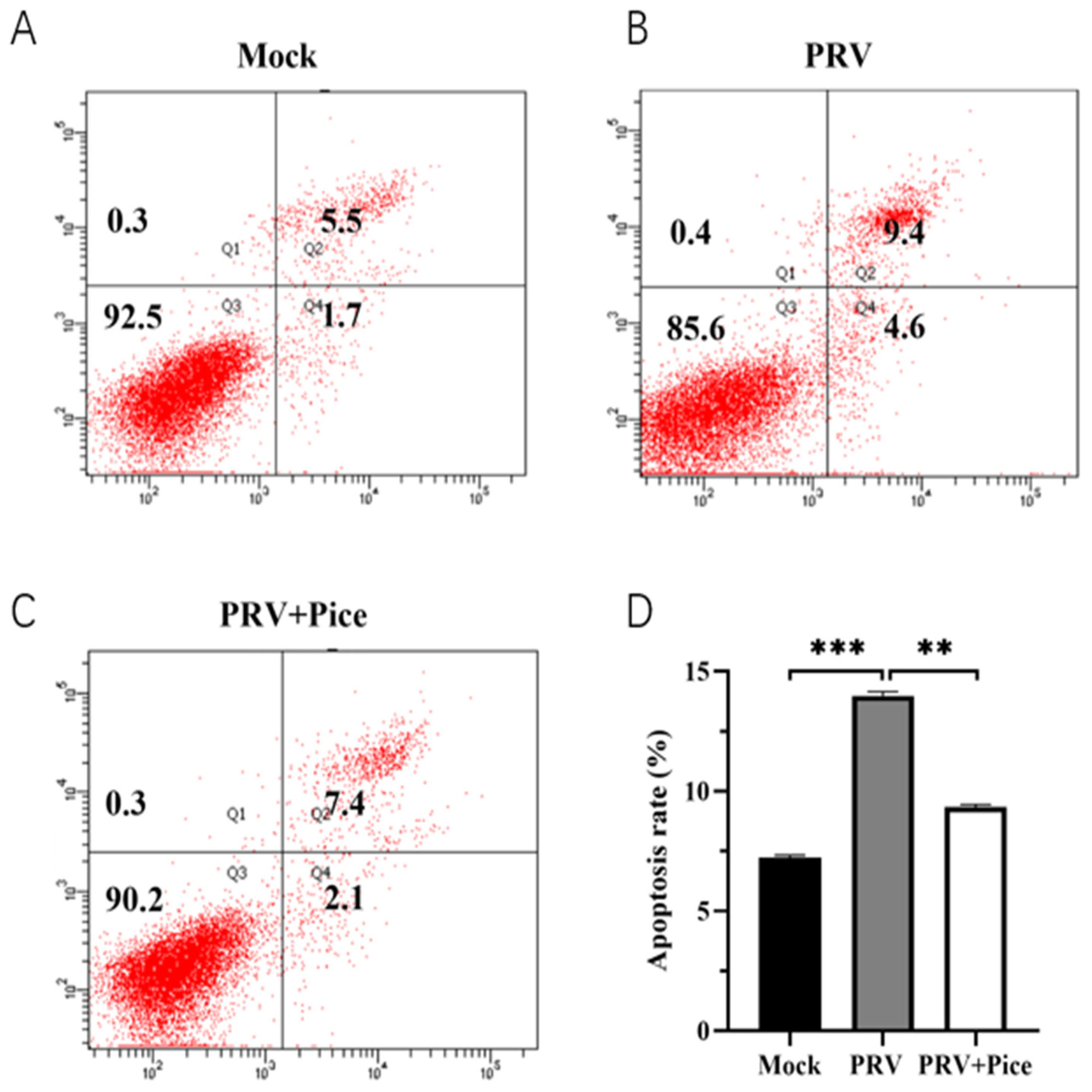
Figure 7.
Piceatannol inhibits PRV activity in vivo. (A) Survival rate of mice in each group. The survival rate for each group was calculated using the following formula: survival rate = number of surviving mice/total number of mice. (B) Changes in the body weight of mice in each group. (C) Viral load in tissues and organs of mice in each group. The heart, liver, lung, kidney and brain tissues of mice were collected on the third day after PRV challenge, viral DNA was extracted, and the viral copy number in each organ tissue was measured by FQ-PCR (ns P > 0.05, * P < 0.05, *** P< 0.001). (D) Pathological sections of mouse brain, heart, liver, lung and kidney tissues.
Figure 7.
Piceatannol inhibits PRV activity in vivo. (A) Survival rate of mice in each group. The survival rate for each group was calculated using the following formula: survival rate = number of surviving mice/total number of mice. (B) Changes in the body weight of mice in each group. (C) Viral load in tissues and organs of mice in each group. The heart, liver, lung, kidney and brain tissues of mice were collected on the third day after PRV challenge, viral DNA was extracted, and the viral copy number in each organ tissue was measured by FQ-PCR (ns P > 0.05, * P < 0.05, *** P< 0.001). (D) Pathological sections of mouse brain, heart, liver, lung and kidney tissues.
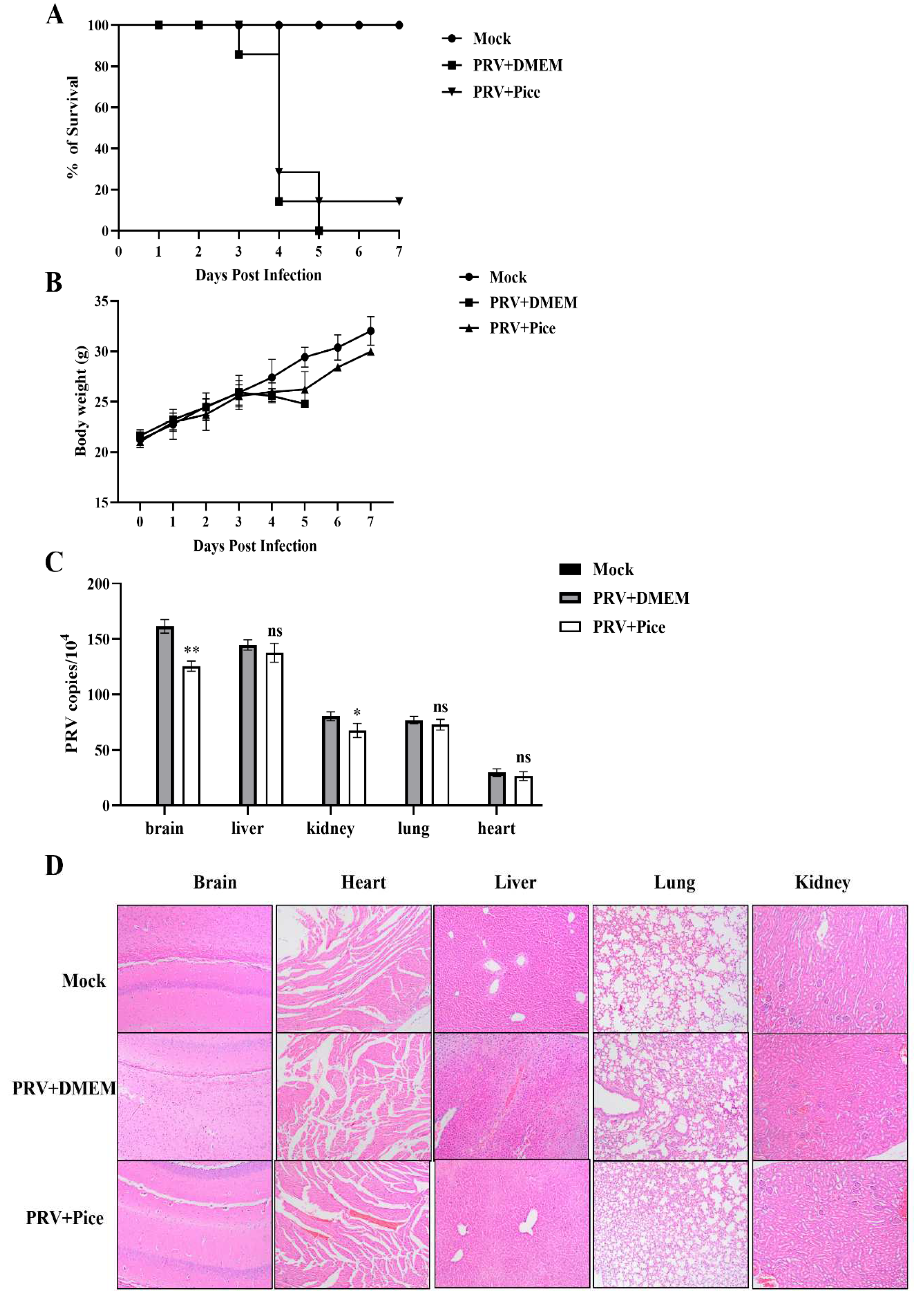
Figure 8.
Serum levels of IFN-γ, TNF-α, IL-6, and IL-4 in each group. On day 3 after PRV infection, three mice from each group were randomly selected for blood collection, and the samples were used to quantify the four cytokines in mouse serum (ns P > 0.05, * P < 0.05, ** P < 0.01, *** P < 0.001, **** P < 0.001).
Figure 8.
Serum levels of IFN-γ, TNF-α, IL-6, and IL-4 in each group. On day 3 after PRV infection, three mice from each group were randomly selected for blood collection, and the samples were used to quantify the four cytokines in mouse serum (ns P > 0.05, * P < 0.05, ** P < 0.01, *** P < 0.001, **** P < 0.001).
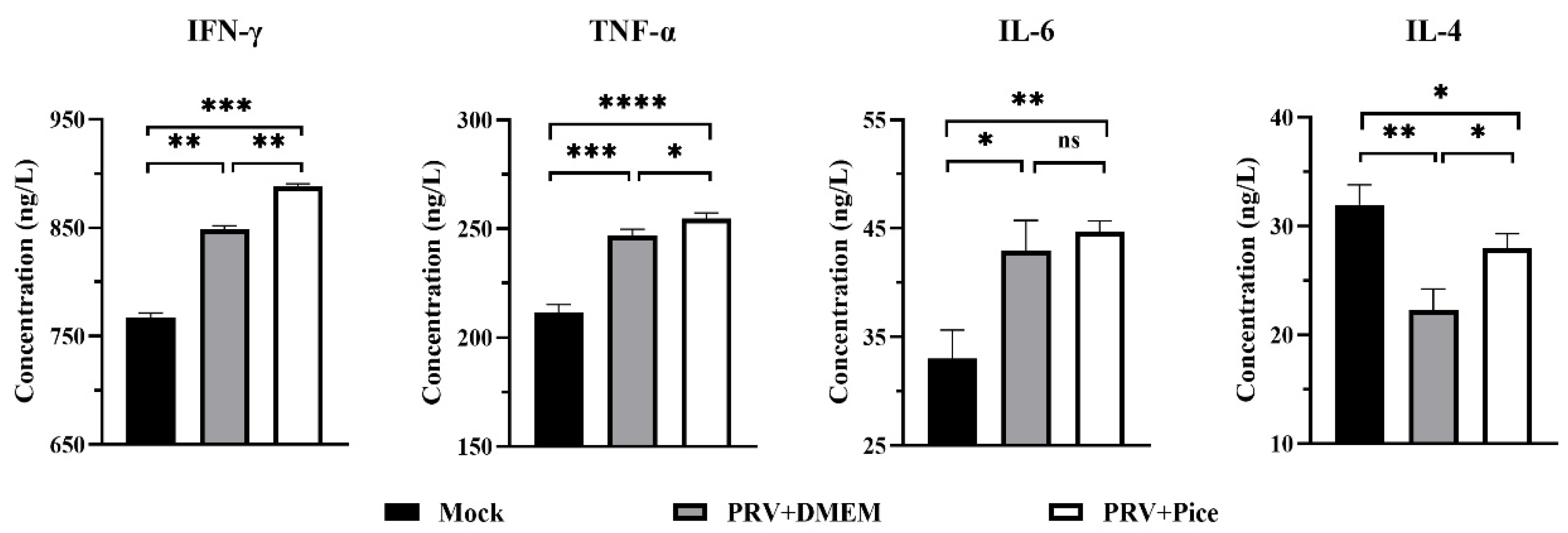

Table 1.
Primer sequences for real-time PCR.
| Gene Name | Sequence (5’-3’) |
|---|---|
| UL44-F | CGTCAGGAATCGCATCA |
| UL44-R | CGCGTCACGTTCACCAC |
| IE180-F | CGCTCCACCAACAACC |
| IE180-R | TCGTCCTCGTCCCAGA |
| UL29-F | AGAAGCCGCACGCCATCACC |
| UL29-R | GGGAACCCGCAGACGGACAA |
| EP0-F | GGGCGTGGGTGTTT |
| EP0-R | GCTTTATGGGCAGGT |
| US6-F | AACATCCTCACCGACTTCA |
| US6-R | CGTCAGGAATCGCATCA |
| UL27-F | TCGTCCACGTCGTCCTCTTCG |
| UL27-R | CGGCATCGCCAACTTCTTCC |
| β-actin-F | TGCGGGACATCAAGGAGAA |
| β-actin-R | AGGAAGGAGGGCTGGAAGA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



