
Submitted:
26 June 2023
Posted:
27 June 2023
You are already at the latest version
Abstract
This study was carried out to evaluate the effect of exogenous proline on growth, biochemical responses, and plant recovery of drought-stressed oilseed rape plants after renewed irrigation. The experiment was conducted under controlled laboratory conditions. After 21 days of cultivation, 3-4 leaf stage seedlings were sprayed with proline (1 mM), then subjected to prolonged drought stress for 8 days to achieve a severe water deficit, then irrigation was resumed and recovery was assessed after 4 days. The results show that exogenous application of proline reduced drought-induced growth inhibition of seedlings while maintaining relative water content (RWC) and growth parameters closer to those of irrigated plants. Proline had a positive effect on chlorophyll accumulation and membrane permeability while decreasing ethylene, H2O2, and MDA levels. Moreover, after 4 days of recovery, the H2O2 content of the proline-treated plants was significantly lower (2-fold) and the MDA content was close to that of continuously irrigated plants. Thus, all these biochemical reactions influenced plant survival: after drought + proline treatment, the number of surviving plants was 2 times higher than that of drought-treated plants. The findings show that exogenous proline has antioxidant, osmotic, and growth-promoting properties that improve the drought tolerance of winter oilseed rape plants and is, therefore, beneficial for drought adaptation in oilseed rape.
Keywords:
Brassica napus
; exogenous proline
; recovered growth
; simulated drought
1. Introduction
Drought stress has detrimental effects on plant growth and development and poses a major threat to sustainable crop production in a rapidly changing environment worldwide. Europe's climate has already become more extreme than previously predicted and rainfall deficits can occur at any time of year [1,2,3]. Plants achieve different physiological and molecular defences to be tolerant to drought stress. It has been estimated that about 80-95% of the fresh biomass of the plant body is water, which plays a vital role in various physiological processes, including many aspects of plant development, and metabolism [4]. One of the adaptive metabolic responses to drought is the accumulation of proline [5]. Proline accumulation is a common physiological response in many plants to a variety of biotic and abiotic stresses [6,7]. Proline accumulates in the cytosol without damaging cellular structures, and it is an essential part of the physiological adaptations to stress in many plant species [8,9]. Farooq et al. [10] reported that the use of proline as an osmotic protection against water deficit in wheat plants resulted in the accumulation of high levels of chlorophyll, proline, glycine betaine, and all soluble phenols. In addition, the use of osmoprotectants has attracted a lot of attention due to their high efficiency, ease of use, low cost, and no need for advanced equipment. Osmoprotectants or compatible solutions are small, highly soluble organic molecules at physiological pH with a neutral charge and low toxicity [11]. It has been shown that proline can protect cells by increasing water uptake potential and facilitating enzyme activation [12]. The relationship between proline content and abiotic stress tolerance in plants is still not clearly understood. However, there is a consensus among plant researchers that the accumulation of proline is beneficial for plants, especially during recovery from stress. In recent years, various studies have suggested that the exogenous application of proline by foliar spraying may play an important role in enhancing abiotic stress tolerance in plants [13,14,15,16,17]. Furthermore, many reports reveal that different plant species have different responses to drought stress, which are generally dependent on the intensity of the stress and the species of plant [18,19,20]. Brassica napus L. is an important oilseed plant in the world and the oil extracted from it is used for human and industrial applications due to its fatty acid composition [21,22].
Drought stress adversely affects germination, seedling establishment, photosynthetic efficiency, mineral uptake, shoot elongation, yield, and quality in rapeseed [23]. It is a problem that needs to be addressed by knowing the targeted pathways and processes. For this reason, the application of exogenous proline in sustainable agriculture practices has emerged as innovative and environment-friendly technology for improving rapeseed productivity. Understanding the role of proline in rapeseed growth and development under drought stress can help the selection of technologies that may be beneficial to regional climate conditions. For example, in Europe, higher temperatures will accelerate the development of winter rapeseed in autumn, making the crop more susceptible to low spring temperatures, but higher temperatures in summer can also cause drought stress and affect plant growth and development [24]. Although the application of exogenous proline effectively reduces the adverse effects of stress, the effect of proline on rapeseed plants depends on the growth stage, the timing and method of application, and the proline concentration. That is why the advanced study on proline metabolism in stress response should be attentive. Thus, the exogenous application of proline may be an effective approach to reducing the adverse effects of water deficit stress, however, the potential role of proline in improving resistance to prolonged drought in rapeseed has not been investigated.
We hypothesize that exogenous proline application will improve drought tolerance and the growth of rapeseed under simulated drought conditions. The objectives of this study were to (1) explore does proline improves shoot growth under water deficit conditions, (2) investigate rape seedling biochemical responses to proline exposure during prolonged drought stress and (3) determine the effect of proline on plant recovery and survival after watering.
2. Results
2.1. Options of Different Concentrations of Proline to Improve RWC and Growth Recovery of Drought-Stressed Oilseed Rape
Differences in RWC of oilseed rape leaves became more pronounced on the 8th day of drought: plants treated with 1 mM proline at 3 and 12.5 ml per pot had RWC of 71% and 73% (mild stress), respectively, whereas drought-only treated plants had RWC of 63% and 52% (high stress). It should be noted that low doses of proline also slightly increased the RWC of leaves of continuously watered plants, i.e. from 83% to 86%. Thus, at 1 mM proline at 12.5 mL per pot, plants were less wilted compared to the drought control (Table 1).
The choice of proline concentration and dose to improve the growth of oilseed rape under simulated prolonged drought conditions showed that the most suitable concentration was 1 mM at 12.5 ml per pot (Table 2). It was observed that this dose of proline enhanced the final average weight of recovered seedlings after 4 days of irrigation.
Thus, treatment with 1 mM proline 12.5 ml per pot, was selected for further experiments because it promotes plant RWC and drought-stressed plants growth recovery (Figure 1).
2.2. Impact of Proline Application on Morphometric Parameters of Oilseed Rape Seedlings Exposed to Drought
Drought stress caused a decrease in fresh and dry weight and length of rapeseed seedlings. Proline treatment had no reliable effect on seedling length both in continuous watering and in drought-stressed. Exogenous application of proline improved the weight of drought-stressed oilseed rape: fresh weight was increased by 31% and 9%; dry weight was increased by 20% and 15% on the 4th and 8th day of drought, respectively (Table 3). The best result was that after 8 days of prolonged drought and recovery after 4 days of irrigation, the length and final average fresh weight of the seedlings treated with 1 mM proline were the highest and close to the continuously watered control.
2.3. Impact of Proline Application on RWC of Prolonged Drought-Stressed Oilseed Rape leaves
After the first 4 days of drought, the plants were highly stressed according to the Hsiao [X] standard, as shown by the reduction of the RWC of oilseed rape leaves by up to 61% (Figure 2). Proline treatment had a positive effect on the RWC of drought-stressed oilseed rape leaves, increasing it to 79% (moderate stress). Even more, after 8 days of prolonged drought, the RWC of the proline-treated seedlings was much closer to the irrigated control than that of the drought-stressed plants, which had only 40% RWC. After 4 days of seedling recovery by irrigation, the RWC of the proline-treated plants was close to that of the continuously irrigated control.
2.4. Impact of Proline Application on Chlorophyll Content of Drought-Stressed Oilseed Rape Leaves
Analysis of chlorophyll a and b content in oilseed rape leaves showed that after 8 days of drought, the concentration of chlorophyll a decreased to 0.57 mg g-1 FW, while the chlorophyll b to 0.12 mg g-1 FW, as compared with watered control 0.99 mg g-1 and 0.23 mg g-1 FW, respectively. Proline significantly increased chlorophyll accumulation after 4 days of drought vs untreated drought control, whereas after 8 days of drought, there was no significant difference between chlorophyll accumulations in leaves of these variants. The highest content of total chlorophylls was found in plants exposed to 8 days of prolonged drought and recovered after 4 days of irrigation (Table 4).
2.5. Impact of Proline Application on Ethylene Emission of Drought-Stressed Oilseed Rape Leaves
In plants treated with proline, ethylene emission was significantly lower by 30% and 19% than only drought treated on the 4th and 8th day, respectively. After 8 days of drought and 4 days of recovery by watering, ethylene levels in proline-treated plants were reduced to those of irrigated control (Figure 3).
2.6. Impact of Proline Application on H2O2 Levels of Drought-Stressed Oilseed Rape Leaves
Spraying with exogenous proline significantly reduced H2O2 content in watered oilseed rape leaf tissues. Impact of exogenous proline manifested on 4-day and 8-day of drought: H2O2 content decreased to 33 % compared to the respective drought control. In addition, after 8 days of drought and 4 days of recovery by watering, H2O2 levels in proline-treated plants were lower by 2-fold (Figure 4).
2.7. Impact of Proline Application on MDA Content of Drought-Stressed Oilseed Rape Leaves
MDA level showed that drought stress significantly increased lipid peroxidation in oilseed rape leaves: after 4 days of drought stress, the amount of MDA in the plants increased by 2 fold, and after 8 days even up to 2.5 fold. In plants treated with proline, the MDA content on the 4th and 8 the day of the drought was significantly lower by 56% and 75% than only drought treated, respectively. In addition, after 8 days of drought and 4 days of water recovery, both proline + drought-treated plants and only drought-treated ones had lower MDA content than continuously irrigated plants (Figure 5).
2.8. Efects of Exogenous Proline on PM ATPase activity of Rapeseed Seedlings Exposed to Drought
After 4 days of drought, PM ATPase activity decreased by 4-fold in oilseed rape leaf cells. Application with exogenous proline increased the H+-ATPase activity of drought-exposed plants by 62% vs only drought-treated plants. After 8 days of drought PM ATPase activity of proline + drought-treated plants increased by 5 fold vs drought test variant, and approached that of the control after recovery of 4 days of irrigation (Figure 6).
2.9. Efect of Exogenous Proline on Endogenuos Proline Content of Rapeseed Seedlings Exposed to Drought
The continuously irrigated oilseed rape plants sprayed with proline had an average of 32 % more endogenous free proline than the control plants throughout the study period. Drought stress significantly increased proline levels: during the first 4 days of drought, proline levels increased by a 2 fold, and as the drought progressed by up to 4 fold. Rapeseed plants in the proline-drought variant had 20-23% more free endogenous proline than plants subjected to 8 days of drought alone. In plants treated with proline and exposed to drought for 8 days and recovering after 4 days of irrigation, the endogenous proline content has come closer to that of irrigated plants, still, in the drought test variant, the level of endogenous proline decreased to a lesser extent (Figure 7).
2.10. Efect of Exogenous Proline on Survival of Plants
Survival evaluation of winter oilseed rape seedlings after 8 days of simulated drought stress and 12 days of recovery by watering showed that the number of surviving plants was significantly higher (more than 2 fold) after proline treatment compared to drought test variant plants (Table 5).
3. Discussion
Different crop species respond differently to drought on the intensity of stress and type of crops [26]. Also drought is one of the many factors with a negative effect morphology, physiology and development of the economically important rapeseed plants [27,28]. Rapeseed is highly sensitive to drought stress, and global climate change leading to severe and prolonged drought in some parts of the world is expected to reduce productivity of rapeseed. There are few studies that water deficit induce metabolic changes in plants which led to the accumulation of specific metabolites such as proline [29,30]. Therefore, proline may help plants to regulate the osmotic potential of cells and to improve water absorbance and translocation under drought conditions [31]. Moreover, data from the literature indicate that exogenous application of osmoprotectants has shown positive effects on plants growing under drought stress, promoting growth and antioxidant activity [32,17].In addition, proline plays an essential role in protecting plants from various stresses and helping them recover faster from stress [8]. Previous studies have shown that the relative water content (RWC) of leaves is one of the main indicators of water status and the ability of plants to grow and develop under drought stress [33,34,35]. So, the RWC informs of the relative water content in leaves and is directly influenced by the soil water content. On the other side, Abdelaal et al. [36] showed that the leaf RWC of barley plants during drought was restored by exogenous proline. We found that exogenous proline affected relative water content (RWC) in rapeseed leaves under continuously increasing water deficit (simulated drought). Our data showed that drought-treated plants exposed to 1 mM proline had the best leaf water retention. It should be noted that low doses of proline also slightly increased the RWC of leaves of continuously watered plants compared to drought control.
Many authors have provided evidence that drought stress negatively affected the morphological parameters, including the fresh and dry weight of shoots, which decreased under drought stress. Thus, plant biomass formation can be considered an important indicator for drought stress assessment [27,32,35,37]. This coincides with the data of our study which revealed that drought stress significantly reduced rapeseed plant growth in terms of fresh and dry biomass. According to the literature, exogenous proline increased the growth of maize [38] and wheat [39] plants and the dry weight of barley plants [36] under drought conditions. Our research has shown that application of exogenous proline (1 mM) was effective in increasing fresh and dry mass of shoots, both under normal water supply and under drought stress. The best result was obtained after irrigation recovery of drought-stressed seedlings sprayed with 1 mM proline: the average fresh weight was the highest and close to the continuously irrigated control.
There is no doubt that one of the most important changes in metabolic functions during drought is the loss of photosynthetic pigments or a decrease in their synthesis. Numerous studies have reported that drought-induced oxidative stress significantly reduces chlorophyll content in crop plants [28,34,35,40]. This was also confirmed by the data of our study showing that moderate and severe drought stress significantly reduced the content of chlorophyll a, b, and compared to control plants. It is important to note that exposure of plants to exogenous proline before drought significantly improved photosynthetic pigment levels not only throughout the drought but also during plant recovery after the resumption of watering. Other researchers also noted that exposure to exogenous proline improved chlorophyll content in wheat [10], barley [36], and rice [41] plant leaves under drought conditions.
Literature data suggested that ethylene emission in plants increases during drought stress [42]. In our study, ethylene content significantly increased during drought stress. In addition, application with exogenous proline decreased ethylene accumulation vs only drought-treated plants, indicating reduced stress levels. After growth recovery by watering, phytohormone ethylene emission of proline-treated plants remained close to irrigated control. Our data contribute to the suggestion that ethylene content may help to remove the inhibitory effect of drought stress on plant growth [43].
Draught may also induce stress responses which result in the accumulation of reactive oxygen species (ROS) such as hydrogen peroxide H2O2 [8,44]. Our study showed that H2O2 concentration during 8 days of drought increased 3.3-fold compared to plants grown under irrigation conditions. Similar drought-induced oxidative stress was recorded in oilseed rape plants in studies by other authors [28,34,36]. Excessive concentrations of free radicals, including H2O2, can cause damage to cell membranes, ion leakage, and osmotic imbalance, so maintaining their level is very important [34,40]. Plant spraying with exogenous proline significantly reduced H2O2 content in prolonged drought-stressed rapeseed leaf tissues. In addition, after 4 days of recovery by watering, the H2O2 level in proline + drought-treated plants decreased even by 2-fold. These data agreed with the results obtained by Abdelaal [36] where exposure to exogenous proline significantly reduced H2O2 content in leaves of barley plants under drought conditions. There are reports in the literature [45,46] characterizing the antioxidant properties of proline in detoxifying ROS. In addition, Rejeb and colleagues [47] in their study concluded that the role of proline as a free radical scavenger is more important than its role as a simple osmolyte.
The stability of cell membranes in drought-stressed plants is adversely affected by ROS, which is evident from the increase in MDA [28,36]. In our study, lipid peroxidation results showed significant membrane damage due to drought. MDA content was elevated in drought-stressed plants and increased with increasing drought time from 1.8 to 2.1 fold that of irrigated plants at 4 and 8 days of drought, respectively. Furthermore, after 8 days of drought and 4 days of recovery by watering, MDA content in proline-treated plants was lower than in continuously irrigated plants. Similar results of increased MDA in rapeseed under drought were also obtained by other researchers [28,33,36]. Meanwhile, a significantly lower amount of MDA - 1.5–1.7 fold - was detected in the cells of plants exposed to exogenous proline before experiencing drought stress, which indicates better stability of the cell membranes of these plants. Similar results where exposure to exogenous proline reduced malondialdehyde levels under drought were obtained in wheat [10], barley [36], and rice [41] plants. This reduction in MDA contents could be attributed to the putative role of osmolytes in alleviating the deleterious effects of stress on the structure of cell membranes and activities of PM enzymes as well as reducing the generation of highly destructive free radicals [38,41,48].
The properties, integrity and composition of the cell membrane can have a major impact on the activity of the PM ATPase, which displaces protons from the plant cell, creates an electrochemical gradient across the plasma membrane and plays an important role in various aspects of plant physiology. PM ATPase activity has been reported to be altered by drought in many plant species [49,50]. In our study, PM ATPase activity is significantly reduced by drought stress, a reduction that has been previously reported by other investigators [51,52]. These findings are also in agreement with the results of Mi et al [53], who showed that Na+,K+-ATPase and Ca2+,Mg2+-ATPase activities of winter rapeseed leave cells gradually decreased with increasing drought severity. In our study: the exogenous application of proline significantly increased PM ATPase activity (from 62 to 400%) compared to drought-exposed, proline-untreated plants. These findings are in line with other researchers who have shown that proline not only protects cells by increasing water uptake potential but also facilitates enzyme activation [12]. In the present study, we showed that the PM ATPase activity of proline-treated oilseed rape recovering from drought stress approached that of control continuously watered plants.
The role of the accumulation of the osmoprotectant proline under drought stress is being actively investigated in order to clarify plant resistance to dehydration. Several reports indicate that under drought stress, plants accumulate higher levels of osmolytes such as proline, which prevent cellular water loss and play an important role in maintaining tissue turgor [54,55]. Our study found that prolonged drought stress significantly increased proline levels, with a 2-fold increase in proline levels during the first 4 days of drought, and as much as a 4-fold increase as the drought progressed. Literature data demonstrate that the exogenous application of proline can increase its endogenous levels in plant tissues exposed to drought stress contributing to the maintenance of drought adaptation in plant tissues [17]. In our research, we confirmed that exogenous proline treatment increased the concentration of endogenous proline in rapeseed leaves under prolonged drought. In addition, it is suggested that the higher levels of proline accumulated in plants exposed to severe and moderate stress conditions may be essential in plant recovery from stress [56]. Our research showed that after watering was renewed, endogenous proline levels of proline + drought-treated plants approached those of irrigated plants.
It was also evident that exogenous proline (1 mM) improved the survival of winter canola seedlings upon resumption of irrigation after 8 days of drought stress. We estimated that the number of surviving plants after 12 days of recovery was more than 2 fold higher proline + drought test variant compared to plants exposed only to drought. Thus, the response of plants to drought stress is complex and involves many physiological, biochemical, cellular, and molecular changes to ensure plant survival.
4. Materials and Methods
4.1. Plant Material and Growth Conditions
Oilseed rape seeds (Brassica napus L. cv. 'Visby') were sown in plastic cubic pots with a peat moss substrate (pH 5.5-6.5). Plants were germinated and grown under controlled conditions of a constant temperature of 23 ± 1 °C, a photoperiod of 16/8 h and a fluorescent light photon flux of 60 µmol m-2 s-1 at soil level, and at 65% humidity in a Climacell plant growing chamber (Medcenter Einrichtangen GmbH). Soil moisture was maintained at ~70%.
4.2. Treatments
4.2.1. L-proline aqueous solutions (Roth) was used for seedling spraying at the 3–4 leaf stage (BBCH-scale 13-14 [57].4.2.2. Drought treatment for the drought stress control studies. During the simulated drought, irrigation was stopped to allow gradual drying of the soil. Soil moisture was estimated using a soil moisture meter (Biogrod, China).
4.3. Determination of the Active Proline Concentration
To determine the active proline concentration, seeds of oilseed rape were sown in 16x3 pots with peat substrate. Each experimental unit consisted of 14 seeds. Pots without proline served as control. Three treatments of the study of proline impact were used: 1) control, H2O 3 ml per pot; 2) proline 0.1 mmol 3 ml per pot; 3) proline 1 mmol 3 ml per pot; 4) proline 10 mmol·3 ml per pot; 5) control, H2O 12.5 ml per pot; 6) proline 0.1 mmol 12.5 ml per pot; 7) proline 1 mmol 12.5 ml per pot; 8) proline 10 mmol·12.5 ml per pot, other eight treatments were the same plus drought treatment. Seedlings at the BBCH 13–14 third-fourth leaves stage [57] were foliar sprayed with water solutions of proline, control was sprayed with water.
4.4. Experimental Design of Drought Stress Control Studies
Plants were sown in plastic cubic pots (15x35 cm) (12 pots in total), 60 seeds per pot, in a peat moss substrate. Four pots were used for the experiment: two for rational watering and two for drought simulation, repeated three times with four pots each. Aqueous solution of L-proline 1 mmol 12.5 ml per pot was sprayed according to the following scheme: 1) control watering, 2) proline and watering, 3) drought, 4) proline and drought (Figure 8.).
4.5. Sampling
Plant samples were taken for analysis on three occasions: on the 4th day of the drought (soil moisture 40%), on the 8th day of the drought (soil moisture 20%), and on the 4th day of plant recovery after watering when soil moisture was 70%. The watered plants, used as controls, were sampled at the same time (soil moisture 70%). Shoots of thirty wheat seedlings were sampled for morphometrical measurements. For biochemical analysis, three independent replicates were carried out using the third leaves of rapeseed plants. Freshly harvested samples were used for ethylene emission analysis and pigment measurement. For H+-ATPase activity, MDA, H2O2 and proline assays the samples were immediately frozen in liquid nitrogen and stored in a low-temperature freezer (Skadi Green line, EU) at -80 °C until the analysis. Survived seedlings were counted after 41 days of cultivation right away.
4.6. Morphometrical Measurements
Shoot length, fresh and dry mass were taken after 4, 8, and 12 days of growth after treatments using a ruler and balances (Kern EWJ, Germany and Sartorius BP 110S, Korea).
4.7. Recovery Evaluation
The same morphometric and biochemical parameters were measured as after 4 and 8 days of drought.
4.8. Survival Evaluation
The plant survival test was carried out by starting to water the plants after 8 days of prolonged drought stress. The test needs to wait for 5-12 days to confirm that winter oilseed rape seedlings are recovering by generating green shoots at the apical meristem growth point in the middle leaf rosette [58]. Thus, plants in each pot were scored as dead or alive after a 12-day recovery period. Plant survival was presented as a percentage of recovered plants.
4.9. Relative Water Content (RWC)
To determine RWC rapeseed leaves were collected and weighted as fresh weight (FW). Then leaves were allowed to fully hydrate on the surface of pure water for 1 d at 4°C in the dark and their turgid weight (TW) was recorded. The leaves were drying for 2 d at 80°C in a drying chamber and weighted to determine the dry weight (DW). RWC have been calculated according to the formula: RWC = 100 × (FW - DW)/(TW - DW) [59].
4.10. Assessment of Biochemical Parameters
4.10.1. Photosynthetic Pigments.
The photosynthetic pigments were extracted from fresh leaves with N, N'-dimethyl-formamide (DMF) (Sigma-Aldrich). Light absorption was measured at 480, 664, 647 nm. Chlorophyll a/b ratio and chlorophyll a and b contents were calculated according to Wellburn [60].
4.10.2. Ethylene.
Method of Child et al. [61] was used to evaluate ethylene emission from freshly harvested leaves. Samples with known mass were placed in 40 mL clear glass vials (Agilent technologies) sealed with PTFE/Si septa caps and incubated for 24 h at 21 °C in darkness. Following incubation, 1 mL of gas sample from each vial was sampled using a gas-tight syringe (Agilent technologies) and injected into a gas chromatograph equipped with a stainless-steel column (Propac R) and hydrogen flame ionization detector. The temperature of the in-jector, column and detector was 110.90 and 150 °C, respectively. Helium (AGA) was used as the carrier gas. Calibrations were made with ethylene standard (Messer). Results were expressed as nl g-1 FW h-1.
4.10.3. Hydrogen Peroxide (H2O2).
Rapeseed leaves (0.5 g) were homogenized using 5% trichloracetic acid (TCA) (Sigma-Aldrich). H2O2 con-tent in leaves was determined according to [62]. The supernatant was mixed with 10 mM, pH 7.0 potassium phosphate buffer (Alfa Aesar) and 1 M potassium iodide (Alfa Aesar) in a ratio of 1:1:2. The reaction solution was incubated for 30 minutes at 25 ° C in the dark. The absorbance of the supernatant was measured at 390 nm. The amount of H2O2 was calculated using a standard curve. The results expressed in µmol g−1 FW.
4.10.4. MDA content.
For analysis of lipid peroxidation, MDA leaf material (0.5 g) was homogenized using 5% trichloracetic acid (TCA) (Sigma-Aldrich). The method of Hodges et al. with slight modifications, have been used to estimate MDA. The homogenates were centrifuged at 13 g for 17 min (centrifuge MPW-351 R), and supernatant was added to 20% TCA containing 0.5% thiobarbituric acid (TBA) (Alfa Aesar). The homogenate was incubated in a heater at 95 °C for 30 min (Blockthermostat BT 200, Kleinfeld, Labortechnik) and then subsequently cooled on the ice. The optical density was measured at 532 and 660 nm by spectrophotometer (Analytik Jena Specord 210 Plus). The results were expressed in µmol g-1 FW [63].
4.10.5. H+-ATPase activity assay.
Membrane-enriched microsomal fraction was extracted from plant samples. Protein content was measured using the Bradford dye-binding procedure [64] at 595 nm. The H+-ATPase activity of microsomal fraction was evaluated according to re-leased inorganic phosphate (Pi) that accumulates as a result of ATP hydrolysis [65]. The colour reaction for Pi measurement was performed with ammonium molybdate and stannous chloride at 750 nm. The activity of H+-ATPase was expressed as μmol Pi mg-1 of protein h-1.
4.10.6. Proline.
A color reaction of acidified ninhydrin was used to determine the content of proline [66]. The equal volume of supernatant of a ground plant material (0.5 g), acetic acid, and acidified ninhydrin was mixed and heated for 1 h 15 min at 108.5°C in a heater. The formed chromophore was extracted with toluene. The absorbance was read spectrophotometrically at 520 nm using a multi-sample quartz cuvette and Rainbow microplate reader, The corresponding content of proline was determined using the standard curve. Calculations were provided using the SLT program (SLT Labinstruments). Results were expressed as µmol of proline µmol g-1 FW.
4.11. Statistical analysis
The results are presented as mean ± standard deviation (SD) of three independent experiments with at least three replicates. The data were analysed using analysis of variance (ANOVA). Tukey's test was performed to test the statistical significance of differences (p <0.05) between means. Post hoc test Duncan's Multiple Range Test (DMRT) was used for comparing the means of different groups after performing an analysis of variance (ANOVA).
5. Conclusions
In the current study, we determined that exogenous proline improved the tolerance of rapeseed growth to prolonged drought by maintaining water status, the accumulation of photosynthetic pigments, osmoprotective substances, and the stability of the plasma membrane in leaf tissues and thus affected the recovery of irrigated rapeseed seedlings after drought stress. Exogenous proline initiated changes in endogenous proline levels of leave tissues. The accumulation of proline is beneficial for plants, especially during recovery from stress: the number of survived plants was significantly higher (2-fold) after proline treatment than the drought variant. the analysis of the stress-mitigating role of exogenous proline leads to conclude that it could be proposed for improving plant drought-stress resistance and increasing the amount of oilseed rape products.
Author Contributions
Conceptualization, V.G. and S.J..; methodology, R.M.; J.J. and V.Š.; formal analysis, R.M.; E.J-B and M.Z.; investigation, R.M.; M.Z.; T.Ž. and E.J-B; data curation, R.M. and V.Š.; writing—original draft preparation, S.J. and R.M.; writing—review and editing, S.J., V.G. and L.K.; visualization, R.M., T.Ž. and S.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Research Council of Lithuania (LMTLT), agreement No S-LU-22-4.
Data Availability Statement
The data supporting the reported results can be found in the archive of scientific reports of the Nature Research Centre.
Acknowledgments
We thank Nijolė Bareikienė and the staff of the Laboratory of Plant Physiology of the Nature Research Centre for the support and help provided.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Della Marta, P. M.; Haylock, M. R.; Luterbacher, J.; Wanner, H. Doubled length of western European summer heat waves since 1880. J. Geophys. Res. Atmos. 2007, 112, D15. [Google Scholar] [CrossRef]
- Peña-Ortiz, C.; Barriopedro, D.; García-Herrera, R. Multidecadal Variability of the Summer Length in Europe, J. of Climat. 2015, 28, 5375–5388. [Google Scholar] [CrossRef]
- Basarin, B.; Lukić, T.; Matzarakis, A. Review of Biometeorology of Heatwaves and Warm Extremes in Europe. Atmosphere 2020, 11, 1276. [Google Scholar] [CrossRef]
- Petrov, P.; Petrova, A.; Dimitrov, I.; Tashev, T.; Olsovska, K.; Brestic, M.; Misheva, S. Relationships between leaf morpho-anatomy, water status and cell membrane stability in leaves of wheat seedlings subjected to severe soil drought. J. Agron. Crop Sci. 2018, 204, 219–227. [Google Scholar] [CrossRef]
- Bandurska, H.; Niedziela, J.; Pietrowska-Borek, M.; Nuc, K.; Chadzinikolau, T.; Radzikowska, D. Regulation of proline biosynthesis and resistance to drought stress in two barley (Hordeum vulgare L.) genotypes of different origin. Plant Physiol Biochem. 2017, 118, 427–437. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: a multifunctional amino acid. Trends Plant Sci. 2009, 15, 89–97. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, O.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef]
- Dar, M.I.; Naikoo, M.I.; Rehman, F.; Naushin, F.; Khan, F.A. Proline accumulation in plants: Roles in stress tolerance and plant development. In Osmolytes and Plants Acclimation to Changing Environment: Emerging Omics Technologies; Springer: Berlin/Heidelberg, Germany, 2016; pp. 155–166. [Google Scholar]
- Farooq, M.; Nawaz, A.; Chaudhry, M.; Indrasti, R.; Rehman, A. Improving resistance against terminal drought in bread wheat by exogenous application of proline and gamma-aminobutyric acid. J. Agron. Crop Sci. 2017, 203, 464–472. [Google Scholar] [CrossRef]
- Hossain, M.A.; Kumar, V.; Burritt, D.J.; Fujita, M.; Mäkelä, P. Osmoprotectant-mediated abiotic stress tolerance in plants. In Proline Metabolism and Its Functions in Development and Stress Tolerance; Springer Nature: Cham, Switzerland, 2019; pp. 41–72. [Google Scholar]
- Caverzan, A.; Casassola, A.; Patussi Brammer, S. Reactive oxygen species and antioxidant enzymes involved in plant tolerance to stress. In Abiotic and biotic stress in plants—recent advances and future perspectives; Shanker, A.K., Shanker, C., Eds., Eds.; InTech, 2016; pp. 463–480. [Google Scholar]
- Orsini, F.; Pennisi, G.; Mancarella, S.; Al Nayef, M. , Sanoubar R.; Nicola, S.; Gianquinto, G. Hydroponic lettuce yields are improved under salt stress by utilizing white plastic film and exogenous applications of proline. Sci. Hortic. 2018, 233, 283–293. [Google Scholar] [CrossRef]
- El Moukhtari, A.; Cabassa-Hourton, C.; Farissi, M.; Savouré, A. How Does Proline Treatment Promote Salt Stress Tolerance During Crop Plant Development? Front. Plant Sci. 2020, 11, 1127. [Google Scholar] [CrossRef]
- Zhang, G.; Wang, Y.; Wu, K.; Zhang, Q.; Feng, Y.; Miao, Y.; Yan, Z. Exogenous Application of Chitosan Alleviate Salinity Stress in Lettuce (Lactuca sativa L.). Horticulturae 2021, 7, 342. [Google Scholar] [CrossRef]
- Godoy, F.; Olivos-Hernández, K.; Stange, C.; Handford, M. Abiotic Stress in Crop Species: Improving Tolerance by Applying Plant Metabolites. Plants 2021, 10, 186. [Google Scholar] [CrossRef]
- Hosseinifard, M.; Stefaniak, S.; Ghorbani Javid, M.; Soltani, E.; Wojtyla, Ł.; Garnczarska, M. Contribution of Exogenous Proline to Abiotic Stresses Tolerance in Plants : A Review. Int J. Mol. Sci. 2022, 23, 5186. [Google Scholar] [CrossRef]
- Kaur, G.; Asthir, B. Molecular responses to drought stress in plants. Biol. Plant. 2017, 61, 201–209. [Google Scholar] [CrossRef]
- Bukhari, S.A.H.; Peerzada, A.M.; Javed, M.H.; Dawood, M.; Hussain, N.; Ahmad, S. Growth and Development Dynamics in Agronomic Crops under Environmental Stress. In Agronomic Crops; Springer: Berlin/Heidelberg, Germany, 2019; pp. 83–114. [Google Scholar]
- Daryanto, S.; Wang, L.; Jacinthe, P.A. Global synthesis of drought effects on cereal, legume, tuber and root crops production. Agric. Water Manag. 2020, 179, 18–33. [Google Scholar] [CrossRef]
- Cartea, E.; De Haro-Bailón, A.; Padilla, G.; Obregón-Cano, S.; del Rio-Celestino, M.; Ordás, A. Seed Oil Quality of Brassica napus and Brassica rapa Germplasm from Northwestern Spain. Foods 2019, 8, 292. [Google Scholar] [CrossRef]
- Kordrostami, M.; Mafakheri, M. Rapeseed: Biology and Physiological Responses to Drought Stress. In The Plant Family Brassicaceae; Hasanuzzaman, M., Ed.; Springer: Singapore, 2020; pp. 263–276. [Google Scholar]
- Batool, M.; El-Badri, A.M.; Hassan, M.U.; Haiyun, Y.; Chunyun, W.; Zhenkun, Y.; Jie, K.; Wang, B.; Zhou, G. Drought Stress in Brassica napus: Effects, Tolerance Mechanisms, and Management Strategies. J. Plant Growth Regul. 2022, 1–25. [Google Scholar] [CrossRef]
- Pullens, J.W.M.; Sharif, B.; Trnka, M.; Balek, J.; Semenov, M.A.; Olesen, J.E. Risk factors for European winter oilseed rape production under climate change. Agric. For. Meteorol. 2019, 272, 30–39. [Google Scholar] [CrossRef]
- Hsiao, T.C. Plant responses to water stress. Ann. Rev. Plant Physiol. 1973, 24, 519–570. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought Stress Impacts on Plants and Different Approaches to Alleviate Its Adverse Effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- El Sabagh, A.; Hossain, A.; Barutcular, C.; Gormus, O.; Ahmad, Z.; Hussain, S.; Islam, M.; Alharby, H.; Bamagoos, A.; Kumar, N.; et al. Effects of drought stress on the quality of major oilseed crops: Implications and possible mitigation strategies—A review. Appl. Ecol. Environ. Res.. 2019, 17, 4019–4043. [Google Scholar] [CrossRef]
- Khan, M.N.; Zhang, J.; Luo, T.; Liu, J.; Ni, F.; Rizwan, M.; Fahad, S.; Hu, L. Morpho-physiological and biochemical responses of tolerant and sensitive rapeseed cultivars to drought stress during early seedling growth stage. Acta Physiol. Plant. 2019, 41, 25. [Google Scholar] [CrossRef]
- Signorelli, S.; Dewi, J.R.; Considine, M.J. Soil Water Content Directly Affects Bud Burst Rate in Single-Node Cuttings of Perennial Plants. Agronomy 2022, 12, 360. [Google Scholar] [CrossRef]
- Radzikowska, D.; Sulewska, H.; Bandurska, H.; Ratajczak, K.; Szymańska, G.; Kowalczewski, P.Ł.; Głowicka-Wołoszyn, R. Analysis of Physiological Status in Response to Water Deficit of Spelt (Triticum aestivum ssp. spelta) Cultivars in Reference to Common Wheat (Triticum aestivum ssp. vulgare). Agronomy 2022, 12, 1822. [Google Scholar]
- Forlani, G.; Trovato, M.; Funck, D.; Signorelli, S. Regulation of Proline Accumulation and Its Molecular and Physiological Fctions in Stress Defence. In Osmoprotectant-Mediated Abiotic Stress Tolerance in Plants: Recent Advances and Future Perspectives; Hossain, M.A., Kumar, V., Burritt, D.J., Fujita, M., Mäkelä, P.S.A., Eds.; Springer International Publishing: Cham, 2019; pp. 73–97. [Google Scholar]
- Semida, W.M.; Abdelkhalik, A.; Rady, M.O.A.; Marey, R.A.; Abd El-Mageed, T.A. Exogenously applied proline enhances growth and productivity of drought stressed onion by improving photosynthetic efficiency, water use efficiency and up-regulating osmoprotectants. Sci. Hortic. 2020, 272, 109580. [Google Scholar] [CrossRef]
- Akram, N.A.; Iqbal, M.; Muhammad, A.; Ashraf, M.; Al-Qurainy, F.; Shafiq, S. Aminolevulinic acid and nitric oxide regulate oxidative defense and secondary metabolisms in canola (Brassica napus L.) under drought stress. In Protoplasma.; 2018; Volume 255, pp. 163–174. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Anee, T. I.; Khan, M. I. R.; Fujita, M. Silicon-mediated regulation of antioxidant defense and glyoxalase systems confers drought stress tolerance in Brassica napus L. South African Journal of Botany 2018, 115, 50–57. [Google Scholar] [CrossRef]
- Sharif, P.; Seyedsalehi, M.; Paladino, O.; Van Damme, P.; Sillanpää, M.; Sharifi, A. Effect of drought and salinity stresses on morphological and physiological characteristics of canola. Int. J. Environ. Sci. Technol. 2018, 15, 1859–1866. [Google Scholar] [CrossRef]
- Abdelaal, K.A.; Attia, K.A.; Alamery, S.F.; El-Afry, M.M.; Ghazy, A.I.; Tantawy, D.S.; Hafez, Y.M. Exogenous application of proline and salicylic acid can mitigate the injurious impacts of drought stress on barley plants associated with physiological and histological characters. Sustainability 2020, 12, 1736. [Google Scholar] [CrossRef]
- Li, J.J.; Zeng, L.; Cheng, Y.; Lu, G.Y.; Fu, G.P.; Ma, H.Q.; Liu, Q.Y.; Zhang, X.K.; Zou, X.L.; Li, C.H. Exogenous melatonin alleviates damage from drought stress in Brassica napus L. (rapeseed) seedlings. Acta Physiol. Plant. 2018, 40, 3. [Google Scholar] [CrossRef]
- Ali, Q.; Ashraf, M.; Athar, H.U. R. Exogenously applied proline at different growth stages enhances growth of two maize cultivars grown under water deficit conditions. Pak. J. Bot 2007, 39, 1133–44. 57. [Google Scholar]
- Kamran, M.; Shahbaz, M.; Ashraf, M.; Akram, N. A. Alleviation of drought-induced adverse effects in spring wheat (Triticum aestivum L.) using proline as a pre-sowing seed treatment. Pak. J. Bot, 2009, 41, 621–632. [Google Scholar]
- Aslam, M.M.; Farhat, F.; Siddiqui, M.A.; Yasmeen, S.; Khan, M.T.; Sial, M.A.; Khan, I.A. Exploration of physiological and biochemical processes of canola with exogenously applied fertilizers and plant growth regulators under drought stress. PLoS One. 2021, 16. [Google Scholar] [CrossRef]
- Hanif, S.; Saleem, M.F.; Sarwar, M.; Irshad, M.; Shakoor, A.; Wahid, M.A.; Khan, H.Z. Biochemically Triggered Heat and Drought Stress Tolerance in Rice by Proline Application. J Plant Growth Regul. 2021, 40, 305–312. [Google Scholar] [CrossRef]
- Valluru, R.; Davies, W.J.; Reynolds, M.P.; Dodd, I.C. Foliar Abscisic acid-to-ethylene accumulation and response regulate shoot growth sensitivity to mild drought in wheat. Front. Plant Sci. 2016, 7, 461. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M. ; Van den Broeck, L.; Inzé, D. The Pivotal Role of Ethylene in Plant Growth. Trends Plant Sci. 2018, 23, 311–323. [Google Scholar] [CrossRef]
- Todorova, D.; Sergiev, I.; Katerova, Z.; Shopova, E.; Dimitrova, L.; Brankova, L. Assessment of the Biochemical Responses of Wheat Seedlings to Soil Drought after Application of Selective Herbicide. Plants 2021, 10, 733. [Google Scholar] [CrossRef]
- Signorelli, S.; Coitiño, E. .L.; Borsani,O.; Monza, J. Molecular mechanisms for the reaction between OH radicals and proline: insights on the role as reactive oxygen species scavenger in plant stress. J.of Physical Chemistr. B 2014, 118, 37–47. [Google Scholar] [CrossRef]
- Raza, A.; Charagh, S.; Abbas, S.; Hassan, M.U.; Saeed, F., Haider; Sharif, R.; Anand, A.; Corpas, F.J.; Jin, W.; Varshney, R.K. Assessment of proline function in higher plants under extreme temperature. Plant Biol J 2023, 25, 379–395. [Google Scholar] [CrossRef]
- Rejeb, K.B.; Abdelly, C.; Savouré, A. How Reactive Oxygen Species and Proline Face Stress Together. Plant Physiol. Biochem. 2014, 80, 278–284. [Google Scholar] [CrossRef]
- Dawood, M.G.; Sh Sadak, M。. Physiological role of glycinebetaine in alleviating the deleterious effects of drought stress on canola plants (Brassica napus L.). Middle East J Agric Res. 2014, 3, 943–954. [Google Scholar]
- Gong, H.J.; Chen, K.M.; Chen, G.C.; Wang, S.M.; Zhang, C.L. Drought stress stimulates p-nitrophenyl phosphate hydrolysis rate of the plasma membrane H+-ATPase from wheat leaves. Plant Growth Regul. 2003, 40, 139–145. [Google Scholar] [CrossRef]
- Feng, X.; Liu, W.; Zeng, F.; Chen, Z.; Zhang, G.; Wu, F. K+ uptake, H+-ATPase pumping activity and Ca2+ efflux mechanism are involved in drought tolerance of barley. Environ. Exp. Bot. 2016, 129, 57–66. [Google Scholar] [CrossRef]
- Michalak, A.; Wdowikowska, A.; Janicka, M. Plant Plasma Membrane Proton Pump: One Protein with Multiple Functions. Cells. 2022, 11, 4052. [Google Scholar] [CrossRef]
- Chang, N.; Ziwen, Z.; Yeyun, L.; Xianchen, Z. Exogenously applied Spd and Spm enhance drought tolerance in tea plants by increasing fatty acid desaturation and plasma membrane H+-ATPase activity. Plant Physiol. Biochem. 2022, 170, 225–233. [Google Scholar]
- Mi, C.; Wang, Q.; Zhao, Y.A.; Zhang, C.L.; Sun, C.; Liu, Z.G.; Lin, L.B. Changes in the Differentially Expressed Proteins and Total Fatty Acid Contents in Winter Rapeseed (Brassica rapa L.) Leaves under Drought Stress. Russ J Plant Physiol 2022, 69, 31. [Google Scholar] [CrossRef]
- Blum, A. Osmotic adjustment is a prime drought stress adaptive engine in support of plant production. Plant Cell Environ. 2017, 40, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Mao, Z.; Zhang, J.; Hemat, M.; Huang, M.; Cai, J.; Jiang, D. Osmolyte accumulation plays important roles in the drought priming induced tolerance to post-anthesis drought stress in winter wheat (Triticum aestivum L.). Envir. and Experiment. Bot. 2019, 166, 103804. [Google Scholar] [CrossRef]
- Kauer, G.; Asthir, B. Proline: a key player in plant abiotic stress tolerance. Biol. Plantarum. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Meier, U. Growth Stages of Mono and Dicotyledonous Plants. In BBCH Monograph; Meier, U., Ed. Quedlinburg: Julius Kühn-Institut, 2018; pp. 85–8. [Google Scholar]
- Fiebelkorn, D.; Rahman, M. Development of a protocol for frost-tolerance evaluation in rapeseed/canola (Brassica napus L.). The Crop Journal 2016, 4, 147–152. [Google Scholar] [CrossRef]
- Weng, M.; Cui, L.; Liu, F.; Zhang, M.; Shan, L.; Yang, S.; Deng, X.-P. Effects of Drought stress on antioxidant enzymes in seedlings of different wheat genotypes. Pak. J. Bot. 2015, 47, 49–56. [Google Scholar]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Child, R.D.; Chauvaux, N.; John, K.; Van Onckelen, H.A.; Ulvskov, P. Ethylene biosynthesis in oilseed rape pods in relation to pod shatter. J. Exp. Bot. 1998, 49, 829–838. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Hodges, D.; DeLong, J.; Forney, C.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid method for the quantification of microgram quantities of proteins utilising the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Darginavičienė, J.; Pašakinskienė, I.; Maksimov, G.; Rognli, O.A.; Jurkonienė, S.; Šveikauskas, V.; Bareikienė, N. Changes in plasmalemma K+ Mg2+-ATPase dephosphorylating activity and H+ transport in relation to freezing tolerance and seasonal growth of Festuca pratensis Huds. J. Plant Physiol. 2008, 165, 825–832. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
Figure 1.
Photograph of Brassica napus seedlings exposed to proline (12.5 ml) after 8 days of drought compared to continuously watered plants.
Figure 1.
Photograph of Brassica napus seedlings exposed to proline (12.5 ml) after 8 days of drought compared to continuously watered plants.

Figure 2.
Impact of proline application on RWC of oilseed rape leaves after simulated prolonged drought and recovery by watering. The horizontal axis indicates the duration of prolonged drought and recovery by irrigation. Vertical error bars represent the standard deviation of the mean of three replications (n = 3). Different lowercase letters indicate statistically significant differences (p < 0.05).
Figure 2.
Impact of proline application on RWC of oilseed rape leaves after simulated prolonged drought and recovery by watering. The horizontal axis indicates the duration of prolonged drought and recovery by irrigation. Vertical error bars represent the standard deviation of the mean of three replications (n = 3). Different lowercase letters indicate statistically significant differences (p < 0.05).
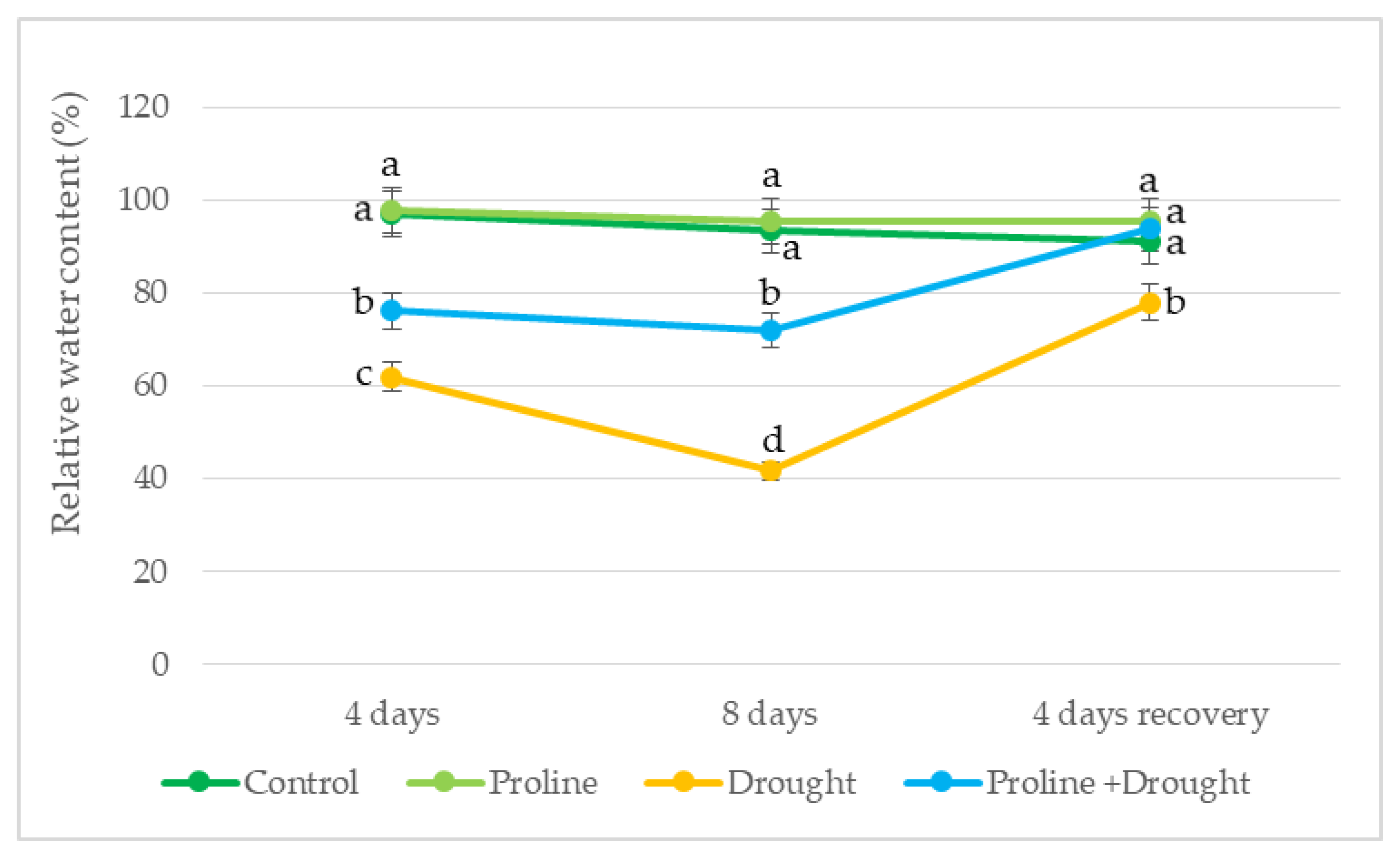
Figure 3.
Impact of proline application on ethylene emission of oilseed rape leaves after simulated prolonged drought and recovery by watering. The horizontal axis indicates the duration of prolonged drought and recovery by irrigation. Vertical error bars represent the standard deviation of the mean of three replications (n = 3). Different lowercase letters indicate statistically significant differences (p < 0.05).
Figure 3.
Impact of proline application on ethylene emission of oilseed rape leaves after simulated prolonged drought and recovery by watering. The horizontal axis indicates the duration of prolonged drought and recovery by irrigation. Vertical error bars represent the standard deviation of the mean of three replications (n = 3). Different lowercase letters indicate statistically significant differences (p < 0.05).
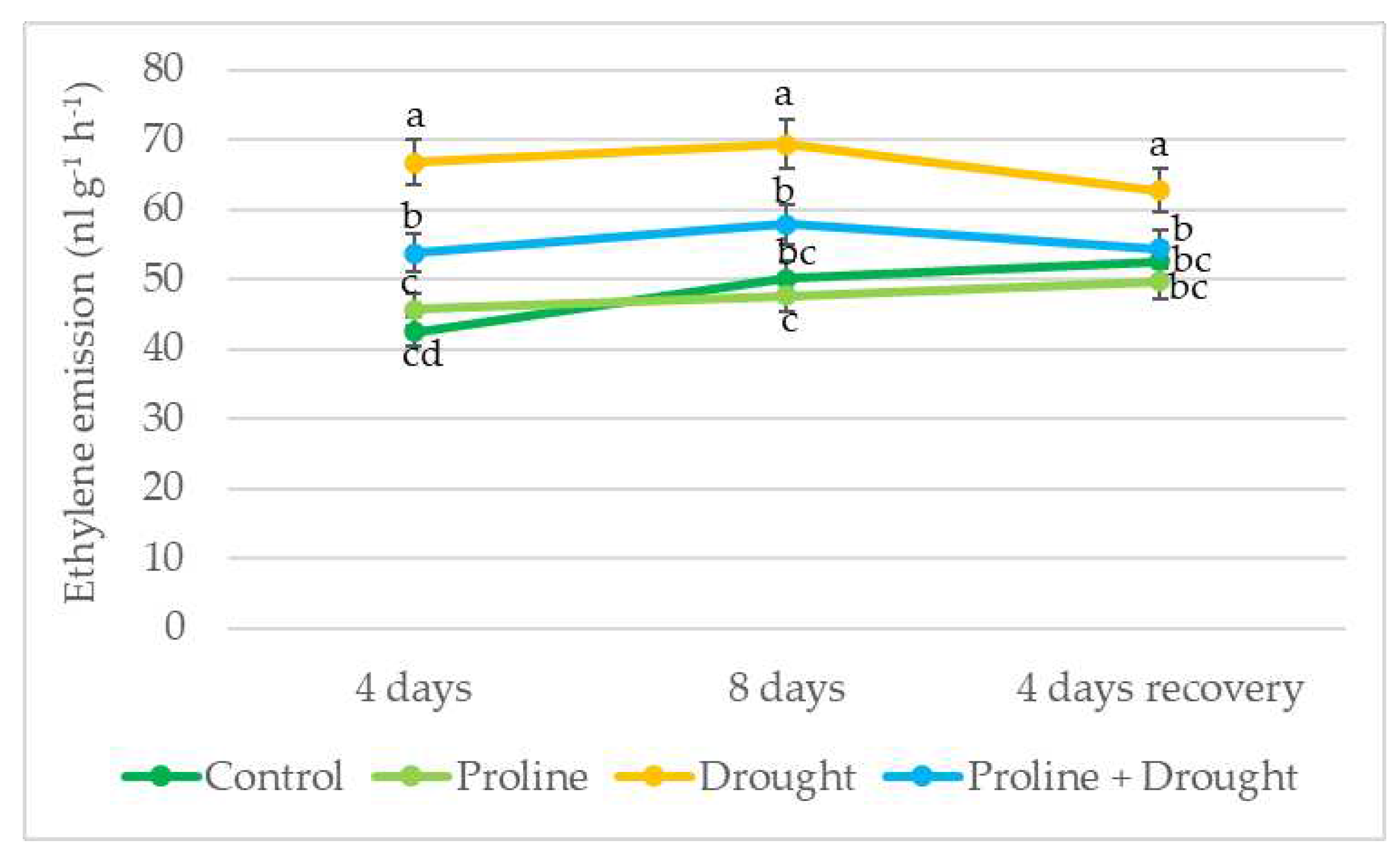
Figure 4.
Impact of proline application on H2O2 level of winter oilseed rape leaves after simulated prolonged drought and recovery by watering. The horizontal axis indicates the duration of prolonged drought and recovery by irrigation. Vertical error bars represent the standard deviation of the mean of three replications (n = 3). Different lowercase letters indicate statistically significant differences (p < 0.05).
Figure 4.
Impact of proline application on H2O2 level of winter oilseed rape leaves after simulated prolonged drought and recovery by watering. The horizontal axis indicates the duration of prolonged drought and recovery by irrigation. Vertical error bars represent the standard deviation of the mean of three replications (n = 3). Different lowercase letters indicate statistically significant differences (p < 0.05).
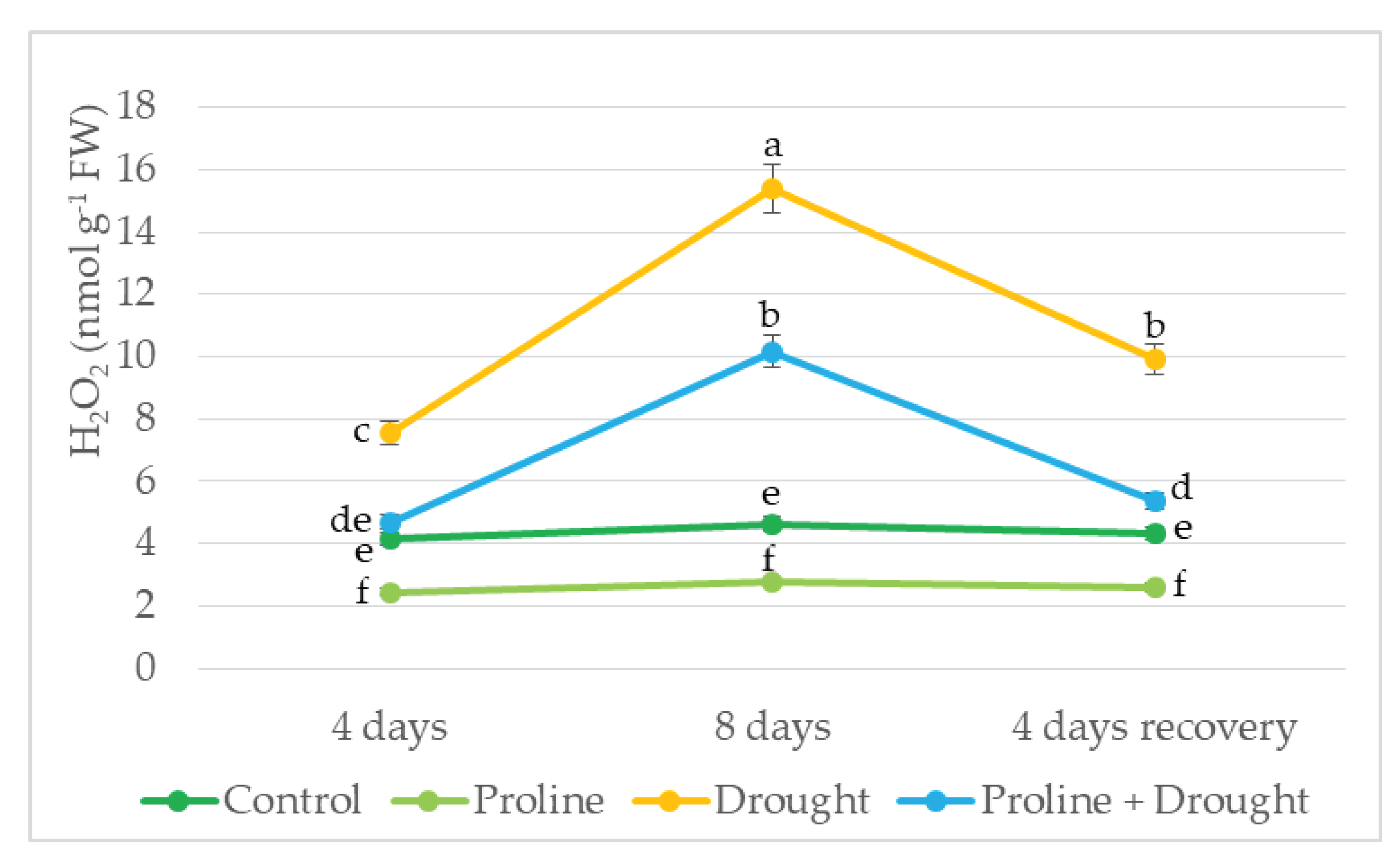
Figure 5.
Impact of proline application on MDA content of winter oilseed rape leaves after simulated prolonged drought and recovery by watering. The horizontal axis indicates the duration of prolonged drought and recovery by irrigation. Vertical error bars represent the standard deviation of the mean of three replications (n = 3). Different lowercase letters indicate statistically significant differences (p < 0.05).
Figure 5.
Impact of proline application on MDA content of winter oilseed rape leaves after simulated prolonged drought and recovery by watering. The horizontal axis indicates the duration of prolonged drought and recovery by irrigation. Vertical error bars represent the standard deviation of the mean of three replications (n = 3). Different lowercase letters indicate statistically significant differences (p < 0.05).
Figure 6.
Impact of proline application on PM ATPase activity of winter oilseed rape leaves after simulated prolonged drought and recovery by watering. The horizontal axis indicates the duration of prolonged drought and recovery by irrigation. Vertical error bars represent the standard deviation of the mean of three replications (n = 3). Different lowercase letters indicate statistically significant differences (p < 0.05).
Figure 6.
Impact of proline application on PM ATPase activity of winter oilseed rape leaves after simulated prolonged drought and recovery by watering. The horizontal axis indicates the duration of prolonged drought and recovery by irrigation. Vertical error bars represent the standard deviation of the mean of three replications (n = 3). Different lowercase letters indicate statistically significant differences (p < 0.05).
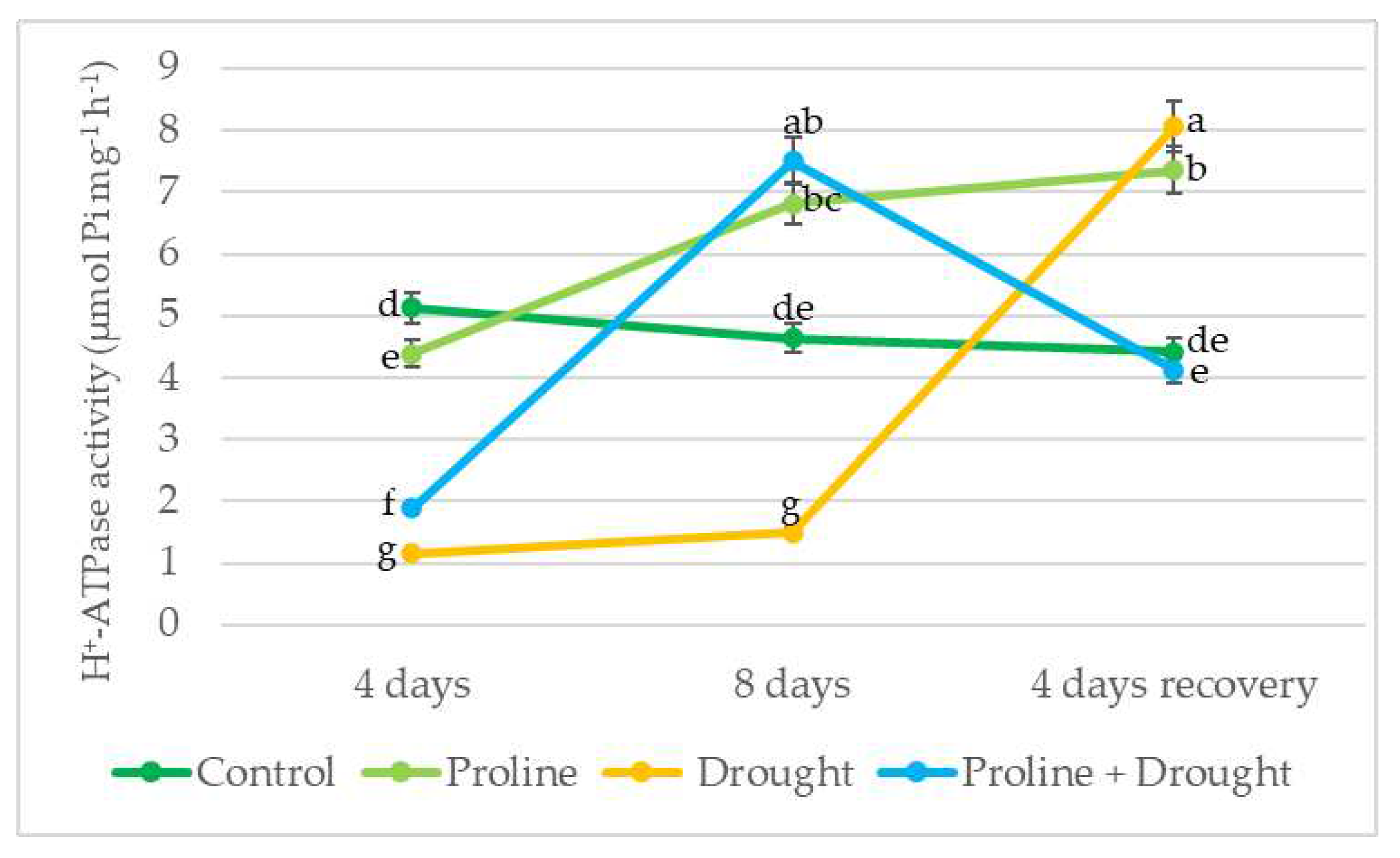
Figure 7.
The effect of exogenous proline application and drought stress, on winter rapeseed proline content. Vertical error bars represent the standard deviation of the mean of three replications (n = 3). Different lowercase letters indicate statistically significant urvived plantsdifferences (p < 0.05).
Figure 7.
The effect of exogenous proline application and drought stress, on winter rapeseed proline content. Vertical error bars represent the standard deviation of the mean of three replications (n = 3). Different lowercase letters indicate statistically significant urvived plantsdifferences (p < 0.05).
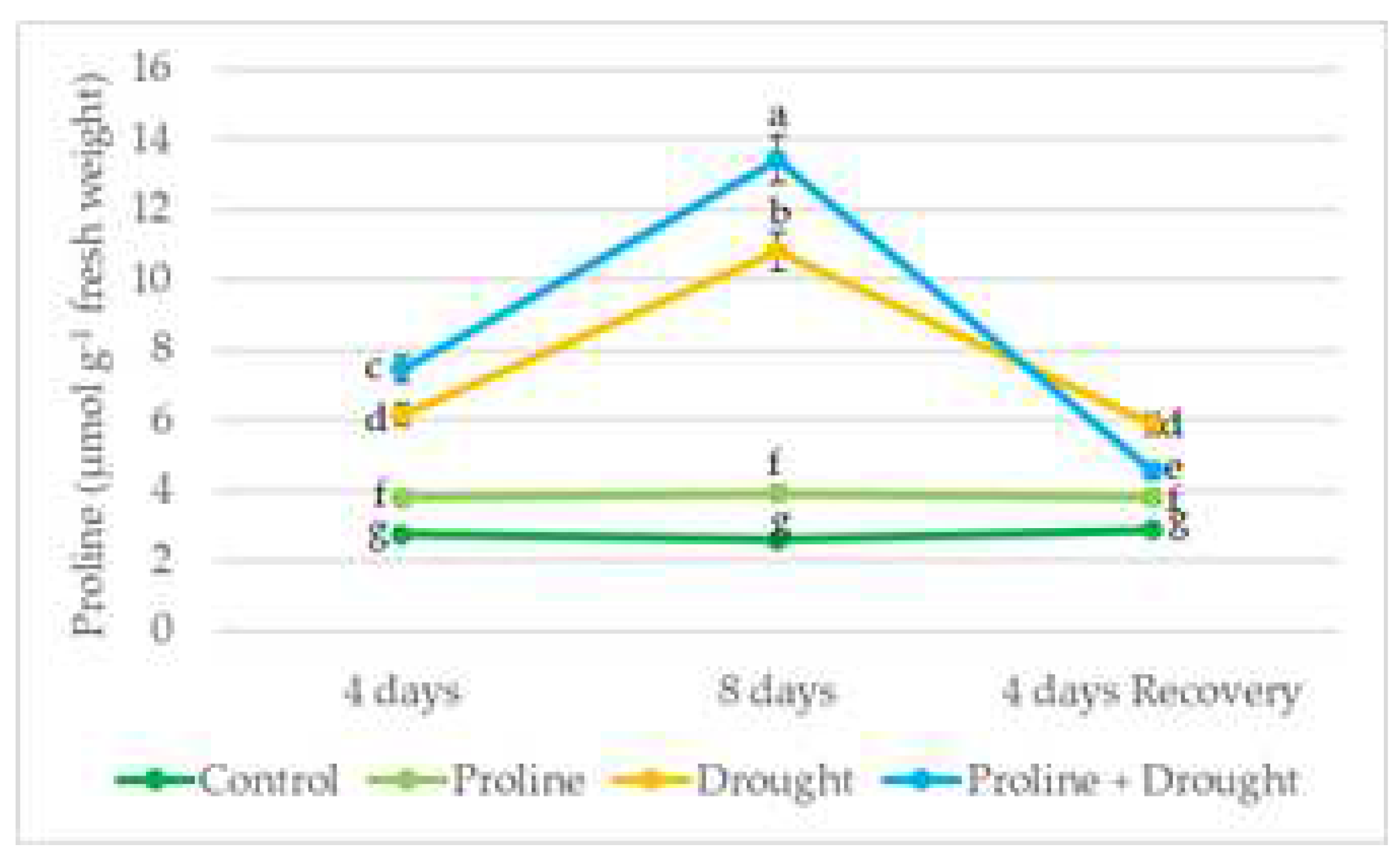
Figure 8.
Experimental design of simulated drought stress control studies.
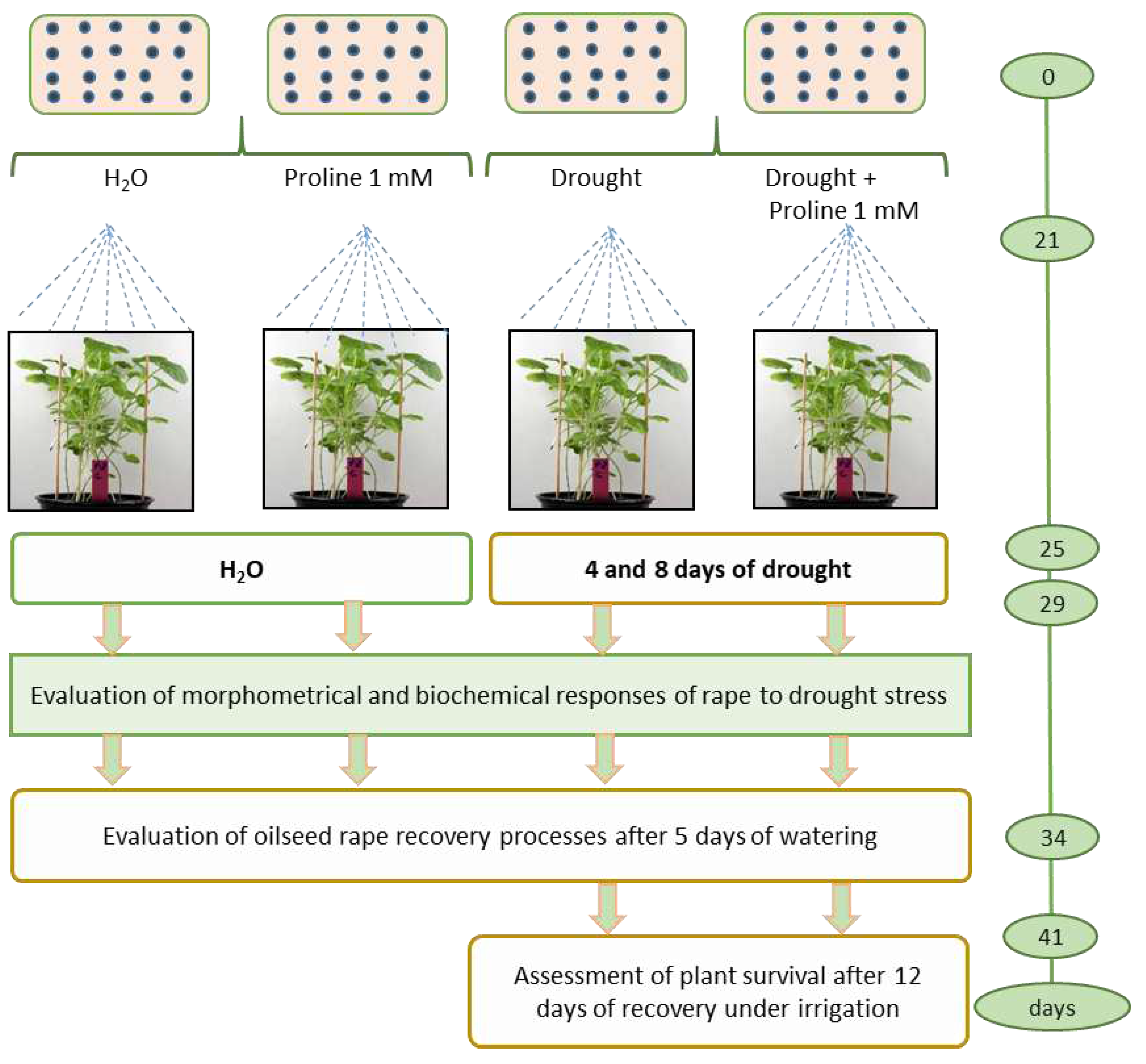

Table 1.
Effect of Proline Treatment on RWC of Leaves with a Continuous Increase in Water Deficit (Simulating Drought).
Table 1.
Effect of Proline Treatment on RWC of Leaves with a Continuous Increase in Water Deficit (Simulating Drought).
| Treatment (3 ml) | RWC, % | |
|---|---|---|
| 4 days | 8 days | |
| Control, H2O | 82.3 a | 82.5a |
| Proline 0.1 mM | 81.5 a | 81.7a |
| Proline 1 mM | 82.3 a | 80.7a |
| Proline 10 mM | 84.0 a | 85.6 a |
| Drought | 70.1 c | 63.2 d |
| Proline 0.1 mM + Drought | 74.7 b | 58.8 e |
| Proline 1 mM + Drought | 73.6 b | 70.7 c |
| Proline 10 mM + Drought | 70.9 c | 59.8 f |
| Treatment (12.5 ml) | ||
| Control, H2O | 84.2 a | 83.5 a |
| Proline 0.1 mM | 85.2 a | 87.2 a |
| Proline 1 mM | 80.7 a | 87.3 a |
| Proline 10 mM | 80.1 a | 86.8 a |
| Drought | 63.1 d | 51.9 e |
| Proline 0.1 mM + Drought | 73.7 b | 60.1 d |
| Proline 1 mM + Drought | 74.6 b | 72.6 bc |
| Proline 10 mM + Drought | 70.4 c | 58.7 d |
* Mean (±SE) was calculated from three replicates for each treatment. Values in a column with different letters are significantly different at p ≤ 0.05 applying Duncan's Multiple Range Test (DMRT).
Table 2.
Effect of Proline on the Weight of Oilseed Rape Recovering from Prolonged Drought after 4 Days of Irrigation.
Table 2.
Effect of Proline on the Weight of Oilseed Rape Recovering from Prolonged Drought after 4 Days of Irrigation.
| Treatment (3 ml) | Average Weight (g) | |
|---|---|---|
| Fresh | Dry | |
| Control, H2O | 0.91 ab | 0.051 a |
| Proline 0.1 mM | 0.88 b | 0.052 a |
| Proline 1 mM | 1.01 a | 0.055 a |
| Proline 10 mM | 1.24 a | 0.052 a |
| Drought | 0.84 c | 0.041 c |
| Proline 0.1 mM + Drought | 0.75 cd | 0.045 ab |
| Proline 1 mM + Drought | 0.78 c | 0.049 ab |
| Proline 10 mM + Drought | 0.97 ab | 0.044 b |
| reatment (12.5 ml) | ||
| Control, H2O | 1.13 a | 0.057 a |
| Proline 0.1 mM | 1.26 a | 0.056 a |
| Proline 1 mM | 1.47 a | 0.059 a |
| Proline 10 mM | 1.35 a | 0.057 a |
| Drought | 1.02 b | 0.043 c |
| Proline 0.1 mM + Drought | 1.07 ab | 0.043 c |
| Proline 1 mM + Drought | 1.18 a | 0.055 ab |
| Proline 10 mM + Drought | 1.11 a | 0.051 b |
* Mean (±SE) was calculated from three replicates for each treatment. Values in a column with different letters are significantly different at p ≤ 0.05 applying DMRT.
Table 3.
Effect of Proline (1 mM) on Morphometric Parameters of Oilseed Rape Seedlings (per Plant) under Simulated Drought Conditions.
Table 3.
Effect of Proline (1 mM) on Morphometric Parameters of Oilseed Rape Seedlings (per Plant) under Simulated Drought Conditions.
| Treatment | Average Length (cm) | Average Weight (g) | |||||||
| Fresh | Dry | ||||||||
| 4 days | 8 days | 4 days recovery | 4 days | 8 days | 4 days recovery | 4 days | 8 days | 4 days recovery | |
| Control H2O | 15.26 a | 17.27 a | 18.00 b | 0.68 b | 0.74 b | 0.77 b | 0.045 a | 0.045 a | 0.046 a |
| Proline | 15.76 a | 17.93 a | 19.03 a | 0.74 a | 0.87 a | 0.89 a | 0.048 a | 0.048 a | 0.049 a |
| Drought | 14.64 b | 14.74 c | 15.71 c | 0.51 c | 0.45 d | 0.46 c | 0.030 c | 0.041 b | 0.038 b |
| Proline + Drought | 14.89 ab | 15.29 b | 17.75 b | 0.67 b | 0.49 c | 0.70 b | 0.036 b | 0.047 a | 0.039 b |
* Mean (±SE) was calculated from three replicates for each treatment. Values in a column with different letters are significantly different at p ≤ 0.05 applying DMRT.
Table 4.
Effect of Proline Application on Chlorophyll Content of Oilseed Rape Seedlings under Drought Stress.
Table 4.
Effect of Proline Application on Chlorophyll Content of Oilseed Rape Seedlings under Drought Stress.
| Treatment | Chlorophyll contents (mg g–1 FW) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Chlorophyll a | Chlorphyll b | Chlorophyll a+b | |||||||
| 4 days | 8 days | 4 days recovery | 4 days | 8 days | 4 days recovery | 4 days | 8 days | 4 days recovery | |
| Control, H2O | 0.98 a | 0.99 a | 0.85 a | 0.24 a | 0.23 a | 0.24 a | 1.22 a | 1.22 a | 1.09 a |
| Proline | 0.97 a | 1.02 a | 0.85 a | 0.25 a | 0.24 a | 0.25 a | 1.22 a | 1.23 a | 1.11 a |
| Drought | 0.78 c | 0.57 c | 0.67 b | 0.21 a | 0.12 b | 0.18 b | 0.98 c | 0.69 c | 0.85 b |
| Proline + Drought | 0.93 b | 0.62 b | 0.89 a | 0.21 a | 0.13 b | 0.22 a | 1.13 b | 0.75 b | 1.10 a |
* Mean (±SE) was calculated from three replicates for each treatment. Different letters in columns designate statistically significant difference at p ≤ 0.05 applying DMRT.
Table 5.
Effect of proline on Brassica napus plant survival after 8 days of prolonged drought recovered after 12 days of irrigation.
Table 5.
Effect of proline on Brassica napus plant survival after 8 days of prolonged drought recovered after 12 days of irrigation.
| Treatment | Number of survived plants (%) |
|---|---|
| Control H2O | 100.00 a |
| Proline | 100.00 a |
| Drought | 19.79±2.21 b |
| Proline + Drought | 44.10 ±3.15 c |
Mean (±SE) was calculated from three replicates for each treatment. Values in a column with different letters are significantly different at p ≤ 0.05 applying DMRT.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



