
Submitted:
26 June 2023
Posted:
27 June 2023
You are already at the latest version
Abstract
Little is known about how differences in water availability within the “super humid” tropics can influence the distribution of understory plant species and the composition of understory plant communities. We investigated variation in the physiological drought tolerances of understory plants and plant communities across a large elevation and precipitation gradient. We established 58 understory plots along a gradient of 400 – 3600 m asl elevation and 1000 – 6000 mm yr-1 rainfall in and around Manu National Park in southeastern Peru. Within the plots, we sampled all understory woody plants and measured three metrics of physiological leaf drought tolerance - turgor loss point (TLP), cuticular conductance (Gmin), and solute leakage (SL) - and assessed how the community-level means of these traits related to mean annual precipitation (MAP) and elevation (in the study gradient temperature decreases linearly and vapor pressure deficit increases monotonically with elevation). We did not find any correlations between the three metrics of leaf drought tolerance, suggesting that they represent independent strategies for coping with low water availability. Despite being widely used metrics of leaf drought tolerance, neither TLP nor Gmin showed any significant relationships with elevation or MAP. In contrast, SL, which has only recently been developed for use in ecological field studies, increased significantly at higher precipitations and at lower elevations (i.e., plants in colder and drier habitats have lower SL, indicating greater drought tolerances). Our results illustrate that differences in water availability may play a strong role in structuring plant communities even in in the wet tropics and highlight the potential for SL assays to be an efficient and effective tool for measuring drought tolerances in the field.
Keywords:
Andes mountains
; drought tolerance
; elevation
; solute leakage
; tropical forest
; understory plants
; turgor loss point
; cuticular conductance
INTRODUCTION
Elevational and environmental gradients are powerful tools for investigating the importance of habitat filtering and community assembly processes. Indeed, the potential value of environmental gradients for demonstrating how different factors drive patterns of plant diversity and vegetation structure has long been recognized [1,2] and previous studies of environmental gradients have provided evidence that many plant species have different distributions based on their tolerances to climate, topography, and soil characteristics [3,4,5,6].
Studies of elevational gradients can also provide important insights into how tropical species, communities, and ecosystems are responding to ongoing anthropogenic climate change [7,8,9,10,11,12]. Elevational gradients provide particular insight into the influence of temperature because air and soil temperatures typically decrease linearly with elevation [12,13]. In the Amazon-Andes region, elevational gradients have been used to assess the effects of temperature on ecosystem productivity and community structure [8,14,15,16,17,18] and to predict how species and communities will be affected by climate change [19,20]. However, much less is known about how changes in rainfall and water availability across elevation and/or through time affect tropical plant communities in these hyper-diverse areas. Some previous studies have shown that changes in precipitation are driving directional changes in the community composition of tropical forests [21,22], but other studies have shown little or no relationship between changes in species composition and changes precipitation [11], and it has even been suggested that observed changes in composition in relation to drought tolerances may be driven primarily by changes in temperature and the inherent relationships between species’ heat and drought tolerances (especially when measured based on geographic occurrence locations) [23]. Given that climate change is altering tropical precipitation regimes [24,25], this incomplete knowledge about how important water availability is for determining species’ current and future distributions greatly limits our ability to predict the fate of these tropical communities and ecosystems.
Since plant water availability is a complex phenomenon involving the interplay between multiple climate variables, soil and environmental factors, and intrinsic species traits related to water uptake and use, the characterization of water availability is nearly impossible for diverse species assemblages spread across large environmental gradients. Water availability is generally believed to decrease at high elevations in the tropics (due to reduced rainfall, increased rainfall seasonality, and thinner soil organic material) [26,27]; however, changes in water availability across elevations are not necessarily monotonic or universal. Lowland rainforests can have heavy rains and high temperatures, but these climate variables are modulated by the soil water capacity [28]. In the sub-montane forests (above lowland rainforests) there is generally higher precipitation but VPD remains high, with increased seasonality at mid-elevations [13]. Cloud forests, which occur above the sub-montane forests, can have high water availability due to high relative humidity and regular cloud and fog immersion, (although there can be seasonal periods with reduced fog, leading to high irradiance and increased vapor pressure deficit, VPD)[13]. As a consequence of regular cloud immersion and high water availability, cloud forests generally have thicker soil organic layers, abundant mosses, and high densities of epiphytes [29,30,31]. Above the cloud forests, the habitat transitions to high-elevation grasslands (in the southern Peruvian Andes these grasslands are referred to as puna) where climate is drier and colder, precipitation is more seasonal, and soils tend to have thinner soil organic layer [32]. Embedded in these high-elevation grasslands, there can be stands of shrubs and small patches of forests where the local climate is more like that of the cloud forests [33]. These changes in water availability across elevations should lead to strong habitat filtering of plant species based on their specific habitat requirements and differences in drought tolerances.
As water availability decreases, plant hydraulic conductance may decrease through different biophysical and physiological mechanisms to prevent embolism or desiccation [34]. In this study, we used turgor loss point, cuticular conductance and solute leakage as tools to characterize relative leaf drought tolerances of individual understory plants and understory plant communities. Turgor loss point (TLP), the leaf water potential at which cell turgor is lost and leaf wilting occurs, provides physiological information pertaining to cell wall integrity, stomatal closure and, more generally, the extent to which plants can maintain metabolism as water availability decreases due to soil drying [35]. As such, plant drought tolerances should be negatively correlated with TLP (i.e., plants that can withstand greater negative water potentials before wilting are considered to be more drought tolerant than plants that wilt at less negative water potentials).
Another mechanism that can promote drought tolerance is to avoid desiccation and maintain high relative water content, thereby resisting the cell damage that could be caused by dehydration. Under dry conditions, plants close their stomata, relying on their cuticle to prevent further water loss. Cuticular conductance (Gmin) is the conductance to vapor diffusion across the leaf epidermis (i.e., through the cuticle and any leaky stomata) once stomata are closed [36]. Plants with low Gmin are considered to be more drought tolerant than plants with high Gmin [37].
Responses at the cellular level may also prevent damage to plants during drought [38,39,40]. The measurement of solute leakage (SL) from plant tissues is a proven method to detect cytorrhysis (damage of cell walls) under laboratory conditions causing desiccation. As such, measurements of SL from leaves exposed to standardized desiccating conditions is a potential metric of relative drought tolerances. Indeed, SL has previously been shown to be an accurate indicator of drought tolerance of crop plants [41,42,43,44]. However, SL has rarely been used in ecological field studies. This is surprising given that SL has many advantages that make it particularly well-suited for studying patterns of plant drought tolerances in biodiverse communities and in remote settings [45,46]. Specifically, the protocol is easy to follow and standardize, large numbers of treatments can be performed relatively quickly, the equipment needed for the experiments are low-cost relative to other physiological measures of drought tolerance, and measurements can be made in areas with little or no existing infrastructure (e.g., measuring SL does not require electricity or compressed gases, and only uses polyethylene glycol, which is widely available as over-the-counter laxatives).
In this study, we measured leaf TLP, Gmin and SL in hundreds of individual understory saplings and shrubs and assessed community-level metrics of leaf drought tolerance in approximately 60 plots located along a 3000+ meter elevation and 5000 mm yr-1 rainfall gradient from lowland rainforests to highland gallery forests and high elevation shrubs. We tested the relationships of these community drought tolerance metrics with elevation (a proxy for temperature and VPD; VPD decreases at higher elevations due to lower air temperatures) and mean annual precipitation. Specifically, we addressed the two following questions and corresponding a priori hypotheses: A) Are the three metrics of drought tolerance coordinated or independent? We hypothesized that TLP, Gmin, and SL would all be correlated due to a coupling of leaf-level mechanisms to resist and tolerate drought [47]. And B) How is drought tolerance related to rainfall and elevation across the gradient? We hypothesized that since adaptation and/or acclimation to drought can incur significant costs, drought-tolerant species would be relatively infrequent in sites with high rainfall and that plant communities in the lower hotter elevations would have greater average drought tolerances than plants at higher elevations. Increasing our understanding of the role that water availability and drought tolerance plays in structuring understory plant communities will improve our ability to predict the responses of tropical ecosystems to environmental changes.
METHODS
Study site. We conducted our study of understory plants and plant communities along an elevational gradient in Manu National Park in Southeastern Peru (Cusco and Madre de Dios regions; Figure 1). The study area ranged from 400 to 3600 m asl and included forests along the Trocha Union ridge in the Kosnipata Valley (900 to 3600 m asl) and around the Cocha Cashu Biological Station (400 m asl; Table 1). Most of the study region is characterized as being “super humid"; the relative orientation of prevailing winds and topography create a zone of frequent cloud immersion when cold Andean winds collide with warm, moist air from the Amazonian lowlands [48]. Mean annual temperature (MAT) ranges from ~23 ºC at 400 m asl to <9 ºC at 3400 m asl (Table 1), with a lapse rate for the understory of 0.53 ºC per 100 m elevation [13]. Mean annual precipitation (MAP) has been measured at a series of meteorological stations along the transect and ranges from ~2000 mm yr−1 at low elevations to 5000 mm yr−1 at the mid-elevations and <1000 mm yr−1 at the very highest elevations. The MAP at each plot location was estimated based on the observed relationship of precipitation vs. elevation in nearby weather stations (adjusted R2 = 0.99) and interpolating to the plot locations based on their elevations as measured with handheld GPS (Table 1). Mean vapor pressure deficit (VPD) in the understory increases monotonically with elevation along the study gradient from 0.01 in the lowlands to 0.12 kPa in the highlands [13].
Vegetation Sampling. We established 58 understory plots of 5 x 5 m (25m2) each. Approximately four plots were installed every 250 m in elevation between 900 and 3600 m asl. Four additional plots were installed at approximately 400 m asl in the lowland rainforest near the Cocha Cashu Biological Station (Figure 1), but these lowland plots were only used to measure SL (see below).
We defined the understory to include all woody shrubs and saplings with basal diameters ≥1 cm at ground level and heights between 0.5 and 5 m (plant height was measured as the vertical distance from ground to the top of the plant canopy; in some cases, stem length was >5 m if the plant was leaning). For every individual, we measured basal diameter, total height, and diameter at breast height (dbh) when height was >1.3m. Most individuals (90%) were identified to family level in the field, and some to genus level. Unidentified individuals were assigned a temporary name in the field. Subsequently, the unidentified individuals were compared with specimens in regional botanical collections and determined to species level or vouchered morphospecies. Vouchers were compared across all plots and taxonomy was standardized. Since all analyses were conducted at the plot-level using the averages of individual-level measurements, the high frequency of morphospecies does not affect the results.
One terminal branch was collected from each individual understory plant in each plot and placed in black plastic bags along with wet paper towels inside and stem ends submerged in plastic bags with water to avoid desiccation during transport to temporary field laboratories established near the top and bottom of the main elevation gradient at 3500 m asl and at 1300m asl, respectively, and at Cocha Cashu Biological Station at 400 m asl. Once in the “laboratory”, branches were placed in buckets, recut under water, and allowed to rehydrate covered with plastic. All traits were measured on leaves from the same branch. Leaves were only included if they looked healthy and were not discolored; leaves that were noticeably affected by pathogens were excluded.
Turgor loss point (TLP). Before measurements of leaf water potential, branches were allowed to rehydrate for 10 hours. After rehydration, three mature leaves were collected from each branch and allowed to slowly desiccate on a table over a period of 6-8 hours between 02:00 and 10:00am local time. During this desiccation period, leaf mass (g) and water potential (Ψ leaf, MPa) were measured periodically. Leaf mass was measured with an analytical balance (±0.002 g, Ohaus, Scout SPX223) and Ψ leaf was measured using a Scholander-type pressure chamber (model 1000, PMS Instruments, Corvallis, Oregon, USA). Plotting the inverse of leaf water potential against relative water content allows determination of the turgor loss point (TLP) as the point of transition between linear and nonlinear portions of the pressure-volume (PV) curve. Lower TLP (i.e., more negative water potentials) indicates higher resistance to leaf wilting [49]. We were not able to construct PV curves or determine TLP for all individuals or species because of short petioles, sap presence in the leaves, or logistical difficulties.
Cuticular conductance (Gmin). For measuring the leaf Gmin, we harvested one to three fully expanded leaves from each individual’s sample branch after ≥4 hours rehydration. We cut the leaf petioles and then sealed the petiole ends with paraffin. Leaves were dried at night using LED lanterns (PPFD 1.73 – 4.6 µmol m-2s-1) on a table for 1 hour in order to induce stomatal closure. The leaves were then massed six times at intervals of 30 minutes. Cuticular conductance was calculated as the rate of water loss (g) over time [36]. At each weighing, we measured the air temperature (°C) and relative humidity (%) with a handheld sensor (Kestrel 2500NV, Nielsen-Kellerman USA) to calculate the saturation vapor pressure (VPsat, kPa). Following standard protocols, the value of Gmin (mmol m-2 s-1) was calculated as the water loss rate divided by VPD (mol mol-1) and twice the total leaf area as measured on scanned leaf images (cm2) [50]. Lower Gmin indicates high drought tolerance since lower cuticular conductance enables maintenance of hydration based on stored water.
Solute leakage (SL). For measuring solute leakage (SL), we harvested three to five leaves from the sample branches of each individual after ≥ 4 hours rehydration. We lightly cleaned each leaf to remove surface debris and cut three pieces from the apical, middle, and bottom area of the leaf with a 1.27 cm diameter hole punch. We submerged the leaf pieces in 20 mL of 50% Polyethylene glycol 3350 (PEG), a hypertonic solution with an osmotic potential of approximately -8.4 MPa, for 12 hours [51]. After the PEG treatment, the leaf pieces were rinsed three times with distilled water (2 µS) to remove any PEG residue. We then transferred the treated leaf pieces to 50 mL falcon tubes with distilled water and measured the initial electrolyte conductivity (C0) using an Oakton CON 6+ Meter conductivity meter. After 12 hours in the distilled water, we took a third conductivity measurement (C1). The leaf pieces and water were then boiled in the falcon tubes for 20 minutes to rupture any remaining intact cells. Twelve hours after boiling, we took a second conductivity measurement (C2). We calculated the relative solute leakage (%) as: (C1 – C0)/(C2 – C0) *100 [45,46]. Higher SL values indicate greater cytorrhysis when exposed to desiccating conditions and thus lower relative leaf drought tolerance. This method assumes that solute leakage is the result of membrane breakage due to water stress within cells, that leaked ions are representative of initial endogenous concentrations in leaf samples before PEG treatment [52], and that differences in leaf nutrition do not influence the concentration of leaked ions [51]. In addition, it assumes that variation in solute leakage is not caused by variation in the capacity of water transport by aquaporins and ion transport by channel and carrier proteins embedded in the plasma membrane. Given that SL is related to several key hydraulic and biochemical parameters, including stomatal resistance and tissue osmotic potential [53], water use efficiency [54], and tissue nitrogen levels [55], we find this a valuable and accessible field tool for assessing relative drought tolerance across many individuals and species.
Statistical Analysis. We used three leaves per individual to calculate TLP and Gmin and five leaves per individual to obtain SL. The values for all individuals in each plot were then averaged to obtain community-weighted mean (CWM) trait values. We measured SL on each individual plant in each plot. In the case of TLP and Gmin, we were not able to sample every individual, and we therefore calculated the CWM trait values as the mean of species-level average trait values weighted by the number of stems per species. Species-level averages were always based on measurements of individuals within each focal plot; in other words, drought tolerance measurements were never generalized between plots. Using plot-specific measurements accounts for any intraspecific variation due to local adaptation or acclimation and eliminates the need for taxonomic standardization across plots. We conducted correlation analysis between the traits to determine possible relationships between the different metrics of physiological drought tolerance. Multiple regression and correlation analyses were used to assess the relationships between the plots’ CWM of SL, TLP, and Gmin with the elevation and MAP. All analyses were conducted in R [56].
RESULTS
Floristic composition. The 58 sample plots contained a total of 834 understory plants with 363 (morpho)species, representing 89 genera and 50 families (Table 1). The most abundant families in the plots were Melastomataceae (170 individuals), Rubiaceae (155 individuals) and Chloranthaceae (55 individuals). The most abundant genera were Miconia (165 individuals), Hedyosmum (55 individuals) and Palicourea (45 individuals). Of the 834 individuals, 501 were saplings and 333 were shrubs (Supplemental Table S1). On average, there were 14 individuals and 10 species per plot, but species richness and stem density varied greatly across elevations and between habitats. Plots at mid-elevations in the sub-montane forest habitat had the greatest density of individuals and the highest number of species (Table 1; Figure 2).
Drought Tolerance metrics. The values of SL, Gmin, and TLP for each individual are reported in Supplemental Table S1 (Table S1 also includes other metrics derived from the pressure volume curves but not included in the analyses reported here). The community weighted mean values of SL, Gmin, and TLP for each plot are reported in Table 2. There were no significant correlations between the CWMs of SL, Gmin, and TLP (Figure 3). Plot mean TLP varied from -0.36 MPa to -1.42 MPa (lower values indicate higher drought tolerance) (Table 2). Due to logistical constraints and the difficulties of measuring TLP in extremely remote locations lacking infrastructure, TLP was only measured in 28 plots from mid and high elevations (937 - 3604 m asl; i.e., only in sub montane forest, cloud forest, and gallery forest habitats). Mean Gmin varied between plots from 4.18 to 99.11 mmol m-2 s-1 (lower values indicate higher drought tolerance) (Table 2). As with TLP, Gmin was only measured in mid and high elevations (1189 - 3604 m asl). Mean SL varied markedly among plots from 13% to 81% (Table 2). SL was measured in all plots 400 - 3615 m asl.
There were no significant relationships between TLP or Gmin with either elevation and/or mean annual precipitation (Figure 4). In contrast, Solute leakage (SL) was significantly related to elevation (SL ~ 65.94 - 0.0067* Elevation (in m); P<0.00005; Adjusted R2 = 0.23) and especially MAP (SL ~ 27.23 + 0.0058 * MAP (in mm); P<0.00005; Adjusted R2 = 0.50) such that SL was lowest (indicating greater drought tolerance) at high elevations and in areas with lower precipitation (Figure 4). If both elevation and MAP are included as explanatory variables, only MAP is significant. The significant relationships between SL and precipitation remains even if we limit the analysis to the subset of plots where we also have information on TLP and Gmin (SL ~ 25.03 + 0.0062 * MAP; P<0.00005; Adjusted R2 = 0.55).
DISCUSSION
We assessed changes in the relative drought tolerance of nearly 60 understory woody plant communities along a 3000+ m elevational gradient in the “super humid” tropics by testing the relationships of three different metrics of physiological leaf drought tolerance with precipitation and elevation. In total, we measured at least one measure of drought tolerance on almost 800 individual plants and measured all three measures of drought tolerance on almost 100 individual plants (Supplemental Table S1). Notably, we did not find covariation between our three metrics of drought tolerance at either the plot level or the individual level (although there was a marginally significant correlation at the individual level between SL and Gmin) (Figure 3). This suggests that plants may develop different independent mechanisms of tolerating drought stress. Additional research is required to understand the interactions and tradeoffs in the different drought tolerance mechanisms employed by understory species and the implications of these mechanisms for plant performance.
The turgor loss point (TLP) values that we measured were generally higher (less drought tolerant) than those reported from other functional groups and habitats. Specifically the TLP of cloud forest vascular epiphytes (-2.71 to -1.67 MPa) [30], understory shrubs from a tropical rainforest (- 2.55 to -1.43 MPa [57]), cloud forest trees (-2.04 to -1.34 MPa [58]), and trees from tropical rainforest (-2.18 to -1.02 MPa [59]) are all lower than what we measured in understory saplings and shrubs along our tropical montane gradient. Other studies have also found TLP to be related to precipitation and VPD such that plants generally have lower TLP (indicating greater drought tolerance) in drier places [60,61]. The relatively high TLP and the absence of a relationship with either elevation or MAP along our gradient (Figure 4) may indicate that our study area was wetter than those of previous studies either due to higher rainfall, frequent cloud immersion, or higher soil water retention. In addition, understories are generally cooler and more humid than canopies, which may explain the lower TLP values for trees and epiphytes. Alternatively, the plant communities that we studied may be relying more heavily on other mechanisms to tolerate periods of low water availability.
Similarly, we found a no relationship of cuticular conductance (Gmin) with either precipitation or elevation (Figure 4). The lowest values of Gmin were in the high elevation gallery forests and the highest values of Gmin occurred in the cloud forest plots at around 2500 m asl. Cloud forests are wet habitats with high rainfall and near-constant fog immersion – conditions that may not select for traits conferring drought tolerance. High cuticular conductance may also facilitate foliar water uptake in the cloud forests [62].
Cell membranes are one of the first victims of many plant stresses and it is generally accepted that the maintenance of their integrity and stability under water stress conditions is a major component of resistance to environmental stresses in plants [51]. As such, measurements of solute leakage (SL) under desiccating conditions should provide valuable information about plant strategies for dealing with low water availability and stress. Indeed, SL has been previously linked to plant tolerances to drought [63], high temperatures [64], and freezing [65]. In our study, we found high SL in plots in wetter areas and at lower elevations, while understory plant communities located in areas with low precipitation and low temperature showed better membrane stability (low values of SL) (Figure 4). High-elevation shrubs had the lowest SL (= highest drought tolerance) in the gradient, possibly due to the combination of the dry and cold (sometimes freezing) conditions that can occur in these habitats. Lowland and high elevation gallery forests (400 and 3548 - 3615 m asl, respectively) had intermediate SL. Gallery forests are located between or below mountain ridges and may receive less fog because of topographical configuration, resulting in relatively dry conditions and potentially explaining their high drought tolerance relative to other habitats. Lowland forests experience less rainfall and thus exhibit values of SL that are comparable with gallery forests >3000 m higher on the gradient. The highest SL value was exhibited by sub montane forests around 1000 m asl. The plots in these sub montane forests receive the highest precipitation (5362-5992 mm yr-1) and can be extremely humid. The high SL of plants in the understory plants of the sub montane forests suggests that these communities are adapted to super wet conditions and may therefore suffer due to any future climate change-driven decreases in rainfall or humidity.
It is particularly noteworthy that SL had the strongest relationships with the environmental variables. Solute leakage has previously been used in laboratory and agricultural assays, but only a few very recent studies have used it to assess drought tolerance of plants in the field. Fadrique et al. (2022) worked along the same elevational gradient in Peru and measured SL in bamboos growing at different elevations. Like our understory dicots, bamboo SL decreased at higher elevations and under wetter conditions [66]. Zuleta et al. (2022) analyzed intra and interspecific differences of SL in trees growing in different topographic habitats in the lowland Amazon rainforest. Consistent with the other results, Zuleta et al. found that trees growing in wetter microhabitats had higher SL than trees growing on the drier ridge tops [46]. Based on these previous studies and the new results presented here, it does appear that differences in water availability lead to predictable changes in SL, indicating that SL may be a powerful tool for assessing plant water relations. The potential power of SL is especially exciting given that it is much cheaper and easier to measure in the field than TLP, Gmin or other standard measures of drought tolerance (e.g., vascular conductance, leaf embolisms) since it does not require any specialized equipment or facilities.
CONCLUSIONS
This study helps to advance our understanding of how variation in water availability drives assembly patterns in tropical understory plant communities. We show that because of habitat filtering and/or local adaptation and acclimation, plant assemblages in areas with different water availabilities have significant differences in drought tolerance as measured by solute leakage (but not by Gmin or TLP). Temperatures in the Andes are increasing rapidly and there is an overall tendency of decreasing annual rainfall through time [24,67]. Increasing temperatures and decreasing rainfall will reduce water availability. In addition, there is a trend of decreasing mean annual and monthly cloud frequency, which could exacerbate water stress and extend the duration of the dry season [68]. Changes in water availability along this elevational gradient will expose the plant communities to different moisture regimes and could potentially lead to changes in species’ distributions and the composition of understory plant communities. Given the importance of the understory in supporting biodiversity, and the fact that saplings and juveniles of all canopy trees must pass through the understory, changes in the understory conditions and composition will have significant consequences for many plant and animal species as well as the valuable ecosystem services that these forests provide.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Table S1. Complete database showing characteristics and leaf drought tolerance metrics measured on 834 individual plants.
Author Contributions
Conceptualization, C.H.B-A. & K.J.F; data collection, C.H.B-A.; analyses, C.H.B-A.; writing—original draft preparation, C.H.B-A. & K.J.F; writing—review and editing, K.J.F. All authors have read and agreed to the published version of the manuscript.
Funding
C.H.B-A. & K.J.F. were supported by NSF DEB 1743249. Field data collection was partially funded by the University of Miami’s Department of Biology's Kushlan Graduate Research Support Fund, and Florida International University's International Center for Tropical Botany.
Data Availability Statement
Data are available in the article’s supplementary material.
Acknowledgments
The authors thank Dr. L. Santiago and members of the junglebology.org lab for suggestions on earlier versions of this manuscript and A. Kullberg for help with the figures. Thanks to Drs. M. Afkhami and C. Baraloto for design and analysis advice. Thanks to A. Ccahuana. M. Montoya, A. Quispe, G. Vargas and C. Alvarez for field support. Thanks to SERNANP and personnel of Manu National Park, Peru, for providing permission and logistical assistance throughout the field campaigns.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Von Humboldt, A. Essai sur la géographie des plantes: accompagne d'un tableau physique des régions équinoxiales, fondé sur des mesures exécutées, depuis le dixième degré la latitude boréale juasqu'au dixième degré de latitude australe, pendant les années 1799, 1800, 1801, 1802 et 1803 par Al. de Humboldt et A. Bonpland; Schoell et Cie.: Chez Levrault, France, 1805. [Google Scholar]
- Whittaker, R.H.; Niering, W.A. Vegetation of the Santa Catalina Mountains, Arizona: a gradient analysis of the south slope. Ecology 1965, 46, 429–452. [Google Scholar] [CrossRef]
- Clark, D.B.; Palmer, M.W.; Clark, D.A. Edaphic factors and the landscape-scale distributions of tropical rain forest trees. Ecology 1999, 80, 2662–2675. [Google Scholar] [CrossRef]
- Harms, K.E.; Condit, R.; Hubbell, S.P.; Foster, R.B. Habitat associations of trees and shrubs in a 50-ha neotropical forest plot. Journal of Ecology 2001, 89, 947–959. [Google Scholar] [CrossRef]
- Zuleta, D.; Russo, S.E.; Barona, A.; Barreto-Silva, J.S.; Cardenas, D.; Castaño, N.; Davies, S.J.; Detto, M.; Sua, S.; Turner, B.L. Importance of topography for tree species habitat distributions in a terra firme forest in the Colombian Amazon. Plant and Soil 2020, 450, 133–149. [Google Scholar] [CrossRef]
- Valencia, R.; Foster, R.B.; Villa, G.; Condit, R.; Svenning, J.-C.; Hernandez, C.; Romoleroux, K.; Losos, E.; Magard, E.; Balslev, H. Tree species distributions and local habitat variation in the Amazon: large forest plot in eastern Ecuador. J Ecology 2004, 92, 214–229. [Google Scholar] [CrossRef]
- Salinas, N.; Malhi, Y.; Meir, P.; Silman, M.; Roman Cuesta, R.; Huaman, J.; Salinas, D.; Huaman, V.; Gibaja, A.; Mamani, M. The sensitivity of tropical leaf litter decomposition to temperature: results from a large-scale leaf translocation experiment along an elevation gradient in Peruvian forests. New phytologist 2011, 189, 967–977. [Google Scholar] [CrossRef]
- Malhi, Y.; Girardin, C.A.; Goldsmith, G.R.; Doughty, C.E.; Salinas, N.; Metcalfe, D.B.; Huaraca Huasco, W.; Silva-Espejo, J.E.; del Aguilla-Pasquell, J.; Farfán Amézquita, F. The variation of productivity and its allocation along a tropical elevation gradient: a whole carbon budget perspective. New Phytologist 2017, 214, 1019–1032. [Google Scholar] [CrossRef]
- Feeley, K.J.; Silman, M.R.; Bush, M.B.; Farfan, W.; Cabrera, K.G.; Malhi, Y.; Meir, P.; Revilla, N.S.; Quisiyupanqui, M.N.R.; Saatchi, S. Upslope migration of Andean trees. Journal of Biogeography 2011, 38, 783–791. [Google Scholar] [CrossRef]
- Feeley, K.J.; Hurtado, J.; Saatchi, S.; Silman, M.R.; Clark, D.B. Compositional shifts in Costa Rican forests due to climate-driven species migrations. Global Change Biology 2013, 19, 3472–2480. [Google Scholar] [CrossRef]
- Duque, A.; Stevenson, P.; Feeley, K.J. Thermophilization of adult and juvenile tree communities in the northern tropical Andes. Proceedings of the National Academy of Sciences USA 2015, 112, 10744–10749. [Google Scholar] [CrossRef]
- Girardin, C.A.J.; Malhi, Y.; Aragao, L.; Mamani, M.; Huaraca Huasco, W.; Durand, L.; Feeley, K.; Rapp, J.; SILVA-ESPEJO, J.; Silman, M. Net primary productivity allocation and cycling of carbon along a tropical forest elevational transect in the Peruvian Andes. Global Change Biology 2010, 16, 3176–3192. [Google Scholar] [CrossRef]
- Rapp, J.; Silman, M. Diurnal, seasonal, and altitudinal trends in microclimate across a tropical montane cloud forest. Climate Research 2012, 55, 17–32. [Google Scholar] [CrossRef]
- Malizia, A.; Blundo, C.; Carilla, J.; Osinaga Acosta, O.; Cuesta, F.; Duque, A.; Aguirre, N.; Aguirre, Z.; Ataroff, M.; Baez, S.; et al. Elevation and latitude drives structure and tree species composition in Andean forests: Results from a large-scale plot network. PLOS ONE 2020, 15, e0231553. [Google Scholar] [CrossRef]
- Homeier, J.; Breckle, S.W.; Günter, S.; Rollenbeck, R.T.; Leuschner, C. Tree diversity, forest structure and productivity along altitudinal and topographical gradients in a species-rich Ecuadorian montane rain forest. BIOTROPICA 2010, 42, 140–148. [Google Scholar] [CrossRef]
- Báez, S.; Malizia, A.; Carilla, J.; Blundo, C.; Aguilar, M.; Aguirre, N.; Aquirre, Z.; Álvarez, E.; Cuesta, F.; Duque, Á.; et al. Large-scale patterns of turnover and basal area change in Andean forests. PLoS ONE 2015, 10, e0126594. [Google Scholar] [CrossRef]
- Quiroga, M.P.; Pacheco, S.; Malizia, L.R.; Premoli, A.C. Shrinking forests under warming: evidence of Podocarpus parlatorei (pino del cerro) from the subtropical Andes. Journal of Heredity 2012, 103, 682–691. [Google Scholar] [CrossRef]
- Krömer, T.; Kessler, M.; Gradstein, S.R. Vertical stratification of vascular epiphytes in submontane and montane forest of the Bolivian Andes: the importance of the understory. Plant Ecology 2007, 189, 261–278. [Google Scholar] [CrossRef]
- Fadrique, B.; Báez, S.; Duque, Á.; Malizia, A.; Blundo, C.; Carilla, J.; Osinaga-Acosta, O.; Malizia, L.; Silman, M.; Farfán-Ríos, W.; et al. Widespread but heterogeneous responses of Andean forests to climate change. Nature 2018, 564, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Feeley, K.J.; Silman, M.R. Land-use and climate change effects on population size and extinction risk of Andean plants. Global Change Biology 2010, 16, 3215–3222. [Google Scholar] [CrossRef]
- Feeley, K.J.; Davies, S.J.; Perez, R.; Hubbell, S.P.; Foster, R.B. Directional changes in the species composition of a tropical forest. Ecology 2011, 92, 871–882. [Google Scholar] [CrossRef]
- Esquivel-Muelbert, A.; Baker, T.R.; Dexter, K.G.; Lewis, S.L.; Brienen, R.J.; Feldpausch, T.R.; Lloyd, J.; Monteagudo-Mendoza, A.; Arroyo, L.; Álvarez-Dávila, E. Compositional response of Amazon forests to climate change. Global change biology 2019, 25, 39–56. [Google Scholar] [CrossRef] [PubMed]
- Feeley, K.J.; Bravo-Avila, C.; Fadrique, B.; Perez, T.M.; Zuleta, D. Climate-driven changes in the composition of New World plant communities. Nature Climate Change 2020. [CrossRef]
- Vuille, M.; Bradley, R.S.; Werner, M.; Keimig, F. 20th century climate change in the tropical Andes: observations and model results. Climate variability and change in high elevation regions: Past, present & future 2003, 75–99.
- Urrutia, R.; Vuille, M. Climate change projections for the tropical Andes using a regional climate model: Temperature and precipitation simulations for the end of the 21st century. Journal of Geophysical Research 2009, 114. [Google Scholar] [CrossRef]
- Bruijnzeel, L.; Mulligan, M.; Scatena, F.N. Hydrometeorology of tropical montane cloud forests: emerging patterns. Hydrological Processes 2011, 25, 465–498. [Google Scholar] [CrossRef]
- Martin, P.H.; Sherman, R.E.; Fahey, T.J. Tropical montane forest ecotones: climate gradients, natural disturbance, and vegetation zonation in the Cordillera Central, Dominican Republic. Journal of Biogeography 2007, 34, 1792–1806. [Google Scholar] [CrossRef]
- Santiago, L.S.; Schuur, E.A.; Silvera, K. Nutrient cycling and plant–soil feedbacks along a precipitation gradient in lowland Panama. Journal of Tropical Ecology 2005, 21, 461–470. [Google Scholar] [CrossRef]
- Nottingham, A.T.; Whitaker, J.; Turner, B.L.; Salinas, N.; Zimmermann, M.; Malhi, Y.; Meir, P. Climate warming and soil carbon in tropical forests: insights from an elevation gradient in the Peruvian Andes. Bioscience 2015, 65, 906–921. [Google Scholar] [CrossRef]
- Gotsch, S.G.; Dawson, T.E.; Draguljić, D. Variation in the resilience of cloud forest vascular epiphytes to severe drought. New Phytologist 2018, 219, 900–913. [Google Scholar] [CrossRef]
- Horwath, A.B.; Royles, J.; Tito, R.; Gudiño, J.A.; Salazar Allen, N.; Farfan-Rios, W.; Rapp, J.M.; Silman, M.R.; Malhi, Y.; Swamy, V. Bryophyte stable isotope composition, diversity and biomass define tropical montane cloud forest extent. Proceedings of the Royal Society B 2019, 286, 20182284. [Google Scholar] [CrossRef]
- Zimmermann, M.; Meir, P.; Silman, M.; Fedders, A.; Gibbon, A.; Malhi, Y.; Urrego, D.; Bush, M.; Feeley, K.; Garcia, K.; et al. No differences in soil carbon stocks across the tree line in the Peruvian Andes. Ecosystems 2010, 13, 62–74. [Google Scholar] [CrossRef]
- Rehm, E.M.; Feeley, K.J. Forest patches and the upward migration of timberline in the southern Peruvian Andes. Forest Ecology and Management 2013, 305, 204–211. [Google Scholar] [CrossRef]
- Venturas, M.D.; Sperry, J.S.; Hacke, U.G. Plant xylem hydraulics: what we understand, current research, and future challenges. Journal of integrative plant biology 2017, 59, 356–389. [Google Scholar] [CrossRef]
- Bartlett, M.K.; Klein, T.; Jansen, S.; Choat, B.; Sack, L. The correlations and sequence of plant stomatal, hydraulic, and wilting responses to drought. Proceedings of the National Academy of Sciences 2016, 113, 13098–13103. [Google Scholar] [CrossRef] [PubMed]
- Kerstiens, G. Cuticular water permeability and its physiological significance. Journal of experimental botany 1996, 47, 1813–1832. [Google Scholar] [CrossRef]
- Slot, M.; Nardwattanawong, T.; Hernández, G.G.; Bueno, A.; Riederer, M.; Winter, K. Large differences in leaf cuticle conductance and its temperature response among 24 tropical tree species from across a rainfall gradient. New Phytologist 2021, 232, 1618–1631. [Google Scholar] [CrossRef] [PubMed]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G. Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? New phytologist 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Choat, B.; Brodribb, T.J.; Brodersen, C.R.; Duursma, R.A.; López, R.; Medlyn, B.E. Triggers of tree mortality under drought. Nature 2018, 558, 531–539. [Google Scholar] [CrossRef]
- Volaire, F. A unified framework of plant adaptive strategies to drought: crossing scales and disciplines. Global change biology 2018, 24, 2929–2938. [Google Scholar] [CrossRef]
- Agarie, S.; Hanaoka, N.; Kubota, F.; Agata, W.; Kaufman, P.B. Measurement of cell membrane stability evaluated by electrolyte leakage as a drought and heat tolerance test in rice (Oryza sativa L.). 1995. [CrossRef]
- Blum, A.; Ebercon, A. Cell membrane stability as a measure of drought and heat tolerance in wheat 1. Crop Science 1981, 21, 43–47. [Google Scholar] [CrossRef]
- Premachandra, G.S.; Shimada, T. The measurement of cell membrane stability using polyethylene glycol as a drought tolerance test in wheat. Japanese Journal of Crop Science 1987, 56, 92–98. [Google Scholar] [CrossRef]
- Premachandra, G.S.; Saneoka, H.; Ogata, S. Cell membrane stability, an indicator of drought tolerance, as affected by applied nitrogen in soyabean. The Journal of Agricultural Science 1990, 115, 63–66. [Google Scholar] [CrossRef]
- Fadrique, B.; Baraloto, C.; Bravo-Avila, C.H.; Feeley, K.J. Bamboo climatic tolerances are decoupled from leaf functional traits across an Andean elevation gradient. Oikos 2022, 2022, e09229. [Google Scholar] [CrossRef]
- Zuleta, D.; Muller-Landau, H.C.; Duque, A.; Caro, N.; Cardenas, D.; Castaño, N.; León-Peláez, J.D.; Feeley, K.J. Interspecific and intraspecific variation of tree branch, leaf and stomatal traits in relation to topography in an aseasonal Amazon forest. Functional Ecology 2022, 36, 2955–2968. [Google Scholar] [CrossRef]
- Santiago, L.S.; De Guzman, M.E.; Baraloto, C.; Vogenberg, J.E.; Brodie, M.; Hérault, B.; Fortunel, C.; Bonal, D. Coordination and trade-offs among hydraulic safety, efficiency and drought avoidance traits in Amazonian rainforest canopy tree species. New Phytologist 2018, 218, 1015–1024. [Google Scholar] [CrossRef] [PubMed]
- Killeen, T.J.; Douglas, M.; Consiglio, T.; Jørgensen, P.M.; Mejia, J. Dry spots and wet spots in the Andean hotspot. Journal of Biogeography 2007, 34, 1357–1373. [Google Scholar] [CrossRef]
- Lenz, T.I.; Wright, I.J.; Westoby, M. Interrelations among pressure–volume curve traits across species and water availability gradients. Physiologia Plantarum 2006, 127, 423–433. [Google Scholar] [CrossRef]
- Pearcy, R.W.; Schulze, E.-D.; Zimmermann, R. Measurement of transpiration and leaf conductance. Plant physiological ecology: field methods and instrumentation 2000, 137–160.
- Bajji, M.; Kinet, J.-M.; Lutts, S. The use of the electrolyte leakage method for assessing cell membrane stability as a water stress tolerance test in durum wheat. Plant Growth Regulation 2002, 36, 61–70. [Google Scholar] [CrossRef]
- Whitlow, T.H.; Bassuk, N.L.; Ranney, T.G.; Reichert, D.L. An improved method for using electrolyte leakage to assess membrane competence in plant tissues. Plant Physiology 1992, 98, 198–205. [Google Scholar] [CrossRef]
- Premachandra, G.; Saneoka, H.; Ogata, S. Nutrio-physiological evaluation of the polyethylene glycol test of cell membrane stability in maize. Crop science 1989, 29, 1287–1292. [Google Scholar] [CrossRef]
- França, M.G.C.; Thi, A.T.P.; Pimentel, C.; Rossiello, R.O.P.; Zuily-Fodil, Y.; Laffray, D. Differences in growth and water relations among Phaseolus vulgaris cultivars in response to induced drought stress. Environmental and Experimental Botany 2000, 43, 227–237. [Google Scholar] [CrossRef]
- Saneoka, H.; Moghaieb, R.E.; Premachandra, G.S.; Fujita, K. Nitrogen nutrition and water stress effects on cell membrane stability and leaf water relations in Agrostis palustris Huds. Environmental and Experimental Botany 2004, 52, 131–138. [Google Scholar] [CrossRef]
- Team, R.C. R: A language and environment for statistical computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Tobin, M.F.; Lopez, O.R.; Kursar, T.A. Responses of Tropical Understory Plants to a Severe Drought: Tolerance and Avoidance of Water Stress 1. BIOTROPICA 1999, 31, 570–578. [Google Scholar] [CrossRef]
- Jane, G.; Green, T. Patterns of stomatal conductance in six evergreen tree species from a New Zealand cloud forest. Botanical gazette 1985, 146, 413–420. [Google Scholar] [CrossRef]
- Bartlett, M.K.; Scoffoni, C.; Ardy, R.; Zhang, Y.; Sun, S.; Cao, K.; Sack, L. Rapid determination of comparative drought tolerance traits: using an osmometer to predict turgor loss point. Methods in Ecology and Evolution 2012, 3, 880–888. [Google Scholar] [CrossRef]
- Cavelier, J. Tissue water relations in elfin cloud forest tree species of Serrania de Macuira, Guajira, Colombia. Trees 1990, 4, 155–163. [Google Scholar] [CrossRef]
- Lenz, T.I.; Wright, I.J.; Westoby, M. Interrelations among pressure-volume curve traits across species and water availability gradients. Physiologia Plantarum 2006, 127, 423–433. [Google Scholar] [CrossRef]
- Goldsmith, G.R.; Matzke, N.J.; Dawson, T.E. The incidence and implications of clouds for cloud forest plant water relations. Ecology Letters 2013, 16, 307–314. [Google Scholar] [CrossRef]
- Premachandra, G.S.; Saneoka, H.; Fujita, K.; Ogata, S. Leaf water relations, osmotic adjustment, cell membrane stability, epicuticular wax load and growth as affected by increasing water deficits in sorghum. Journal of experimental botany 1992, 43, 1569–1576. [Google Scholar] [CrossRef]
- Yang, G.; Rhodes, D.; Joly, R. Effects of High Temperature on Membrane Stability and Chlorophyll Fluorescence in Glycinebetaine-Deficient and Glycinebetaine-Containing Maize Lines. Functional Plant Biology 1996, 23, 437–443. [Google Scholar] [CrossRef]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Archives of Biochemistry and Biophysics 2005, 444, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Fadrique, B.; Baraloto, C.; Bravo-Avila, C.H.; Feeley, K.J. Bamboo climatic tolerances are decoupled from functional traits across an Andean elevation gradient. Oikos 2022. In Press. [Google Scholar] [CrossRef]
- Haylock, M.R.; Peterson, T.C.; Alves, L.M.; Ambrizzi, T.; Anunciação, Y.M.T.; Baez, J.; Barros, V.R.; Berlato, M.A.; Bidegain, M.; Coronel, G.; et al. Trends in Total and Extreme South American Rainfall in 1960–2000 and Links with Sea Surface Temperature. Journal of Climate 2006, 19, 1490–1512. [Google Scholar] [CrossRef]
- Halladay, K.; Malhi, Y.; New, M. Cloud frequency climatology at the Andes/Amazon transition: 2. Trends and variability. Journal of Geophysical Research: Atmospheres (1984–2012) 2012, 117.
Figure 1.
Map of the study plots along an elevational gradient in and around Manu National Park in Southeastern Peru (Cusco and Madre de Dios regions). The study area ranges from 400 to 3600 m asl and includes forests along the Trocha Union ridge in the Kosnipata Valley (900 to 3600 m asl) with field stations at Tres Cruces and San Pedro, and near the Cocha Cashu Biological Station (400 m asl; Table 1).
Figure 1.
Map of the study plots along an elevational gradient in and around Manu National Park in Southeastern Peru (Cusco and Madre de Dios regions). The study area ranges from 400 to 3600 m asl and includes forests along the Trocha Union ridge in the Kosnipata Valley (900 to 3600 m asl) with field stations at Tres Cruces and San Pedro, and near the Cocha Cashu Biological Station (400 m asl; Table 1).
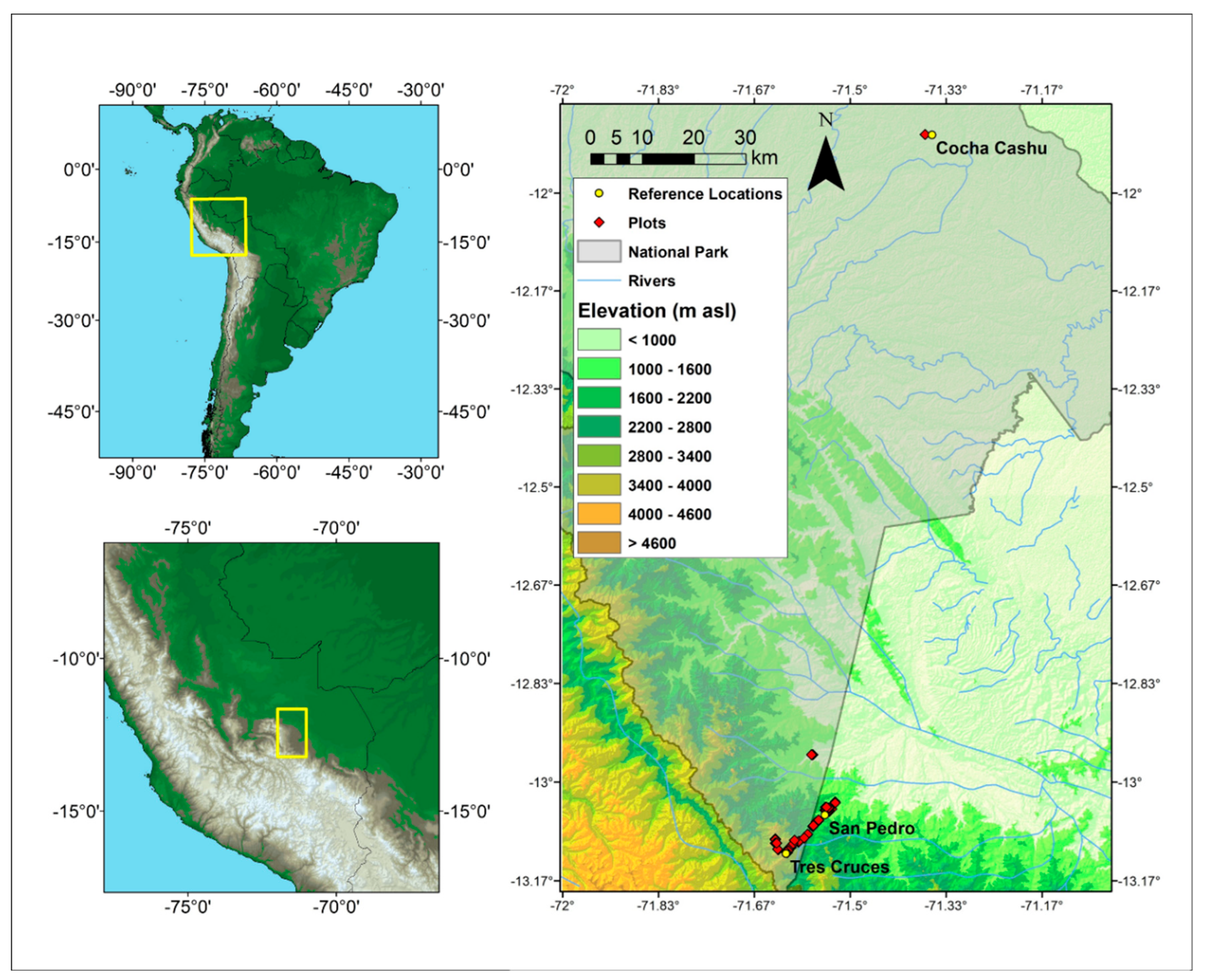
Figure 2.
The A) number of individual understory saplings and shrubs and B) number of understory species and morphospecies recorded in the 5 × 5m study plots were greatest in sub montane forests around 1500 m asl and then decreased steadily with elevation. Points are colored according to plot habitat: Red = lowland forest, Green = sub montane forest, Blue = cloud forest, Cyan = high-elevation gallery forest, and Purple = high elevation shrubs.
Figure 2.
The A) number of individual understory saplings and shrubs and B) number of understory species and morphospecies recorded in the 5 × 5m study plots were greatest in sub montane forests around 1500 m asl and then decreased steadily with elevation. Points are colored according to plot habitat: Red = lowland forest, Green = sub montane forest, Blue = cloud forest, Cyan = high-elevation gallery forest, and Purple = high elevation shrubs.
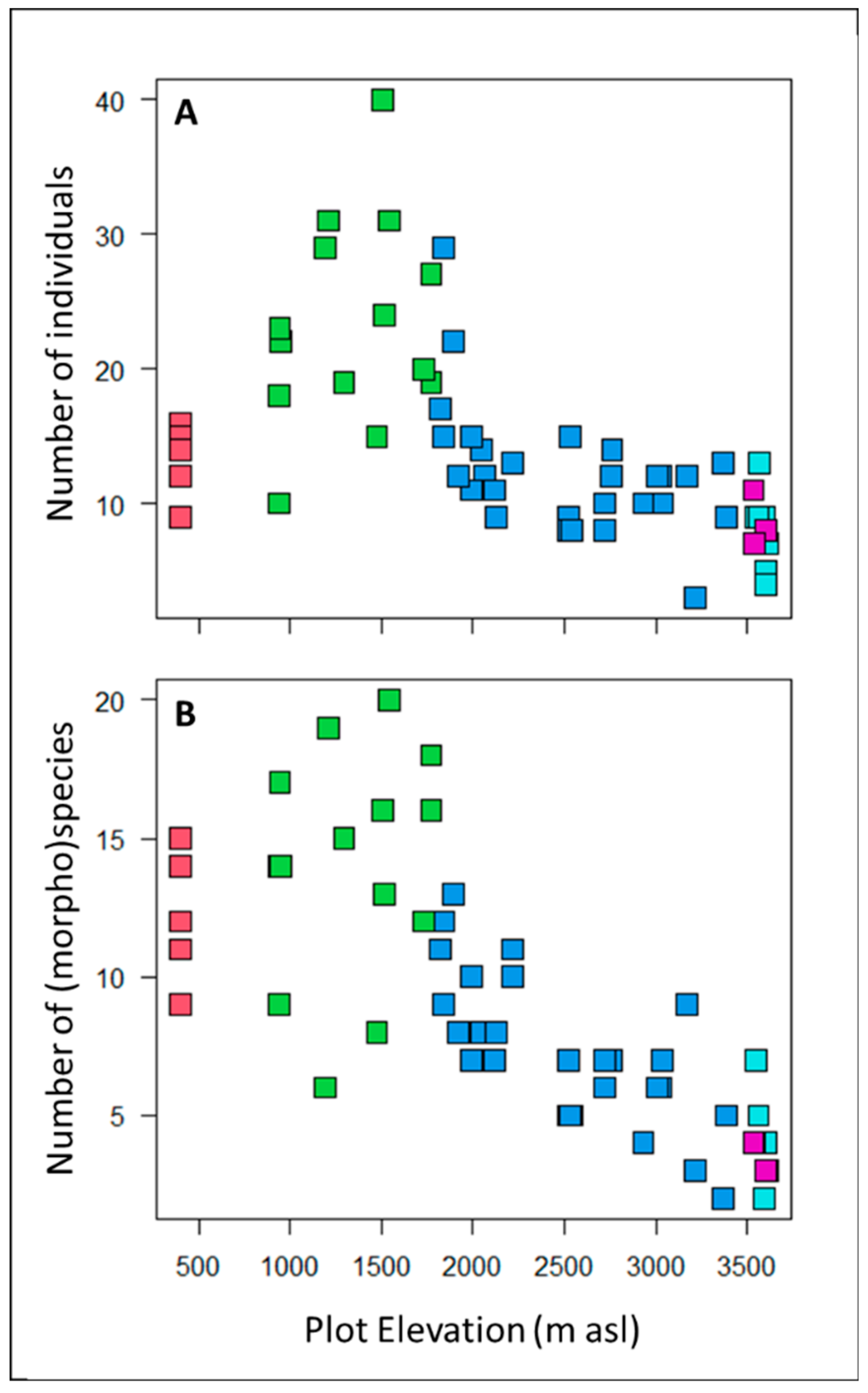
Figure 3.
There were no significant correlations at either the individual level (circles = individual plant measurements) or plot level (squares = community weighted means) between A) Gmin, and TLP, B) Gmin and SL, or C) SL and TLP. Points are colored according to plot habitat: Green = sub montane forest, Blue = cloud forest, and Cyan = high-elevation gallery forest.
Figure 3.
There were no significant correlations at either the individual level (circles = individual plant measurements) or plot level (squares = community weighted means) between A) Gmin, and TLP, B) Gmin and SL, or C) SL and TLP. Points are colored according to plot habitat: Green = sub montane forest, Blue = cloud forest, and Cyan = high-elevation gallery forest.
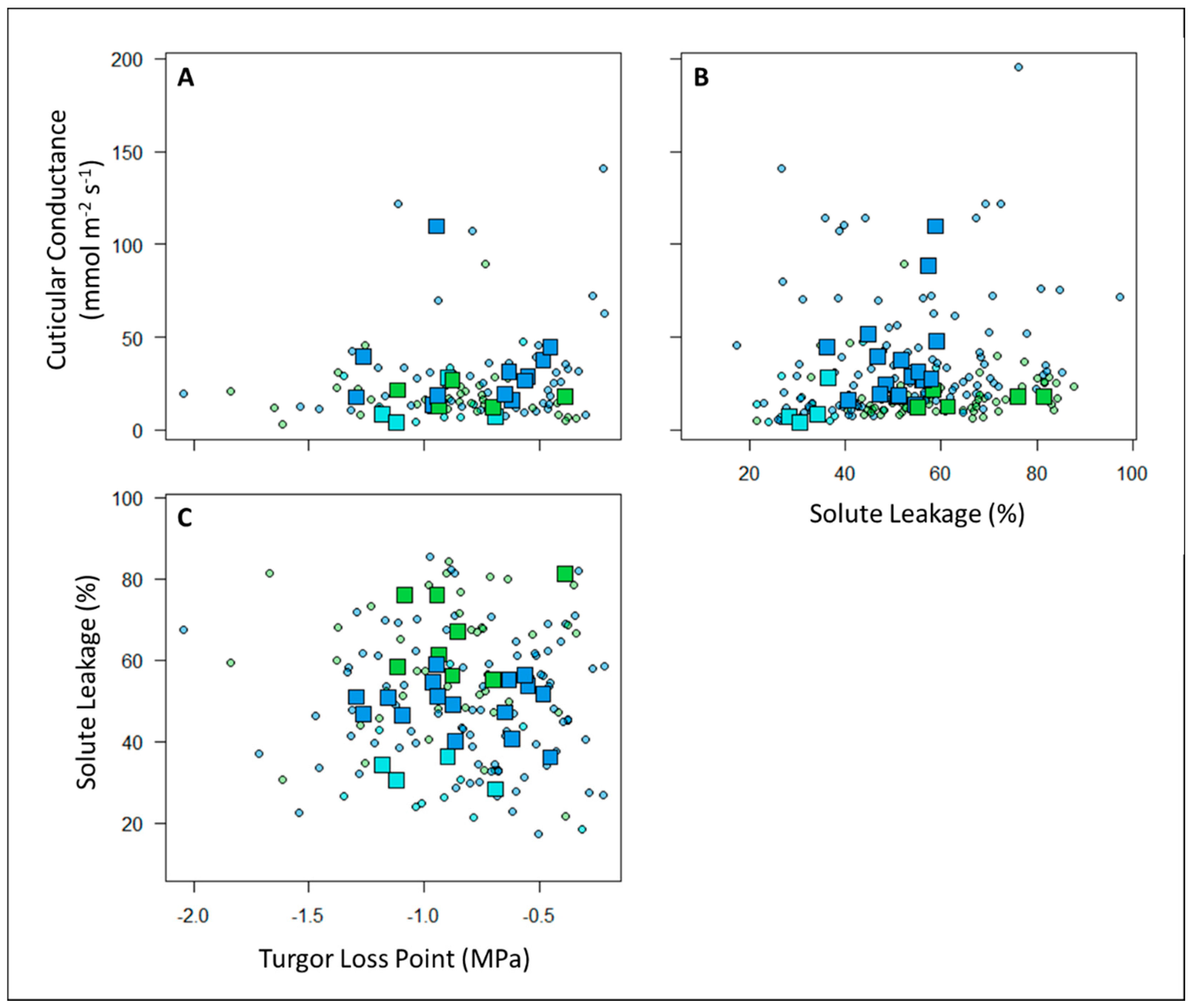
Figure 4.
There were no significant correlations at either the individual level (circles = individual plant measurements) or plot level (squares = community weighted means) between TLP (panels A and B) or Gmin (panels C and D) with either elevation (panels A and C) or Mean Annual Precipitation (panels B and D). In contrast, Solute leakage (SL) was significantly related to elevation (panel E; SL ~ 65.94 - 0.0067* Elevation; P<0.00005; Adjusted R2 = 0.23) and MAP (panel F; SL ~ 27.23 + 0.0058 * MAP; P<0.00005; Adjusted R2 = 0.50). Points are colored according to plot habitat: Red = lowland forest, Green = sub montane forest, Blue = cloud forest, Cyan = high-elevation gallery forest, and Purple = high elevation shrubs.
Figure 4.
There were no significant correlations at either the individual level (circles = individual plant measurements) or plot level (squares = community weighted means) between TLP (panels A and B) or Gmin (panels C and D) with either elevation (panels A and C) or Mean Annual Precipitation (panels B and D). In contrast, Solute leakage (SL) was significantly related to elevation (panel E; SL ~ 65.94 - 0.0067* Elevation; P<0.00005; Adjusted R2 = 0.23) and MAP (panel F; SL ~ 27.23 + 0.0058 * MAP; P<0.00005; Adjusted R2 = 0.50). Points are colored according to plot habitat: Red = lowland forest, Green = sub montane forest, Blue = cloud forest, Cyan = high-elevation gallery forest, and Purple = high elevation shrubs.
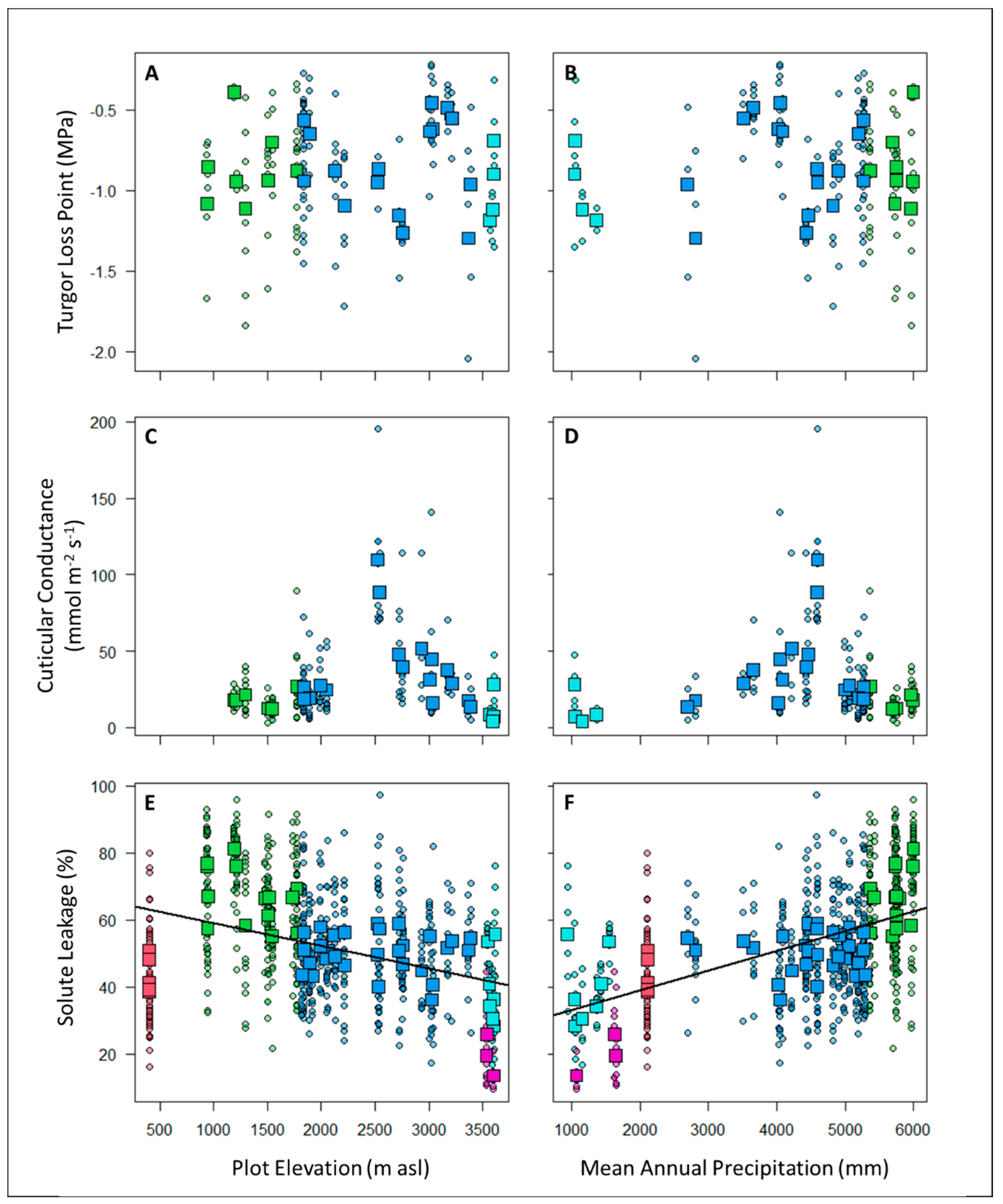

Table 1.
Locations and Characteristics of study plots.
| Plot | Latitude (º) | Longitude (º) | Elevation (m asl) | MAT (º C) | MAP (mm/year) | Habitat | N. Individuals | N. Shrubs | N. Sapling | N. Families | N. Genera | N. (Morpho)Species |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | -13.096 | -71.630 | 3604 | 8.2 | 1617 | gallery forest | 5 | 3 | 2 | 3 | 4 | 4 |
| 2 | -13.097 | -71.630 | 3569 | 8.2 | 1437 | gallery forest | 13 | 5 | 8 | 3 | 3 | 4 |
| 3 | -13.103 | -71.630 | 3603 | 8.0 | 1054 | gallery forest | 4 | 3 | 1 | 2 | 2 | 4 |
| 4 | -13.103 | -71.629 | 3592 | 18.9 | 1157 | gallery forest | 9 | 0 | 9 | 1 | 2 | 2 |
| 5 | -13.114 | -71.625 | 3548 | 18.9 | 1549 | gallery forest | 9 | 5 | 4 | 2 | 1 | 7 |
| 6 | -13.113 | -71.626 | 3562 | 10.1 | 1515 | gallery forest | 9 | 3 | 6 | 3 | 2 | 5 |
| 7 | -13.103 | -71.628 | 3615 | 12.5 | 937 | gallery forest | 7 | 1 | 6 | 3 | 2 | 3 |
| 8 | -13.110 | -71.603 | 3172 | 18.7 | 3654 | cloud forest | 12 | 7 | 5 | 4 | 5 | 9 |
| 9 | -13.110 | -71.604 | 3214 | 18.7 | 3507 | cloud forest | 3 | 3 | 0 | 2 | 3 | 3 |
| 10 | -13.114 | -71.607 | 3366 | 17.7 | 2812 | cloud forest | 13 | 9 | 4 | 2 | 2 | 2 |
| 11 | -13.112 | -71.607 | 3387 | 17.7 | 2692 | cloud forest | 9 | 1 | 8 | 4 | 4 | 5 |
| 12 | -13.033 | -71.526 | 1189 | 10.1 | 5992 | sub-montane | 29 | 21 | 8 | 5 | 5 | 6 |
| 13 | -13.035 | -71.526 | 1212 | 17.7 | 5990 | sub-montane | 31 | 16 | 15 | 5 | 5 | 19 |
| 14 | -13.045 | -71.532 | 1296 | 20.2 | 5989 | sub-montane | 19 | 7 | 12 | 10 | 9 | 15 |
| 15 | -13.047 | -71.544 | 1771 | 19.7 | 5363 | sub-montane | 27 | 9 | 18 | 6 | 8 | 18 |
| 16 | -13.047 | -71.542 | 1837 | 17.7 | 5266 | cloud forest | 29 | 6 | 23 | 5 | 6 | 12 |
| 17 | -13.049 | -71.536 | 1503 | 20.1 | 5744 | sub-montane | 40 | 8 | 32 | 6 | 9 | 16 |
| 18 | -13.048 | -71.537 | 1545 | 8.0 | 5694 | sub-montane | 31 | 19 | 12 | 7 | 8 | 20 |
| 19 | -13.119 | -71.609 | 3601 | 10.1 | 1082 | puna shrubs | 8 | 8 | 0 | 0 | 0 | 3 |
| 20 | -13.116 | -71.608 | 3539 | 12.5 | 1082 | puna shrubs | 7 | 7 | 0 | 0 | 0 | 4 |
| 21 | -13.118 | -71.611 | 3536 | 10.1 | 1082 | puna shrubs | 11 | 11 | 0 | 0 | 0 | 4 |
| 22 | -13.101 | -71.590 | 2754 | 13.3 | 4428 | cloud forest | 12 | 5 | 7 | 4 | 3 | 7 |
| 23 | -13.101 | -71.590 | 2762 | 13.6 | 4421 | cloud forest | 14 | 5 | 9 | 7 | 6 | 7 |
| 24 | -13.100 | -71.589 | 2721 | 11.2 | 4457 | cloud forest | 10 | 5 | 5 | 5 | 5 | 6 |
| 25 | -13.100 | -71.589 | 2724 | 12.5 | 4455 | cloud forest | 8 | 4 | 4 | 5 | 4 | 7 |
| 26 | -13.089 | -71.575 | 2525 | 14.4 | 4595 | cloud forest | 9 | 6 | 3 | 4 | 4 | 7 |
| 27 | -13.089 | -71.575 | 2526 | 14.4 | 4594 | cloud forest | 8 | 4 | 4 | 2 | 1 | 5 |
| 28 | -13.088 | -71.574 | 2542 | 13.6 | 4584 | cloud forest | 8 | 1 | 7 | 4 | 1 | 5 |
| 29 | -13.094 | -71.580 | 2532 | 17.7 | 4590 | cloud forest | 15 | 12 | 3 | 3 | 4 | 5 |
| 30 | -13.104 | -71.599 | 3036 | 22.1 | 4021 | cloud forest | 10 | 7 | 3 | 3 | 2 | 7 |
| 31 | -13.104 | -71.599 | 3025 | 14.5 | 4045 | cloud forest | 12 | 8 | 4 | 3 | 4 | 6 |
| 32 | -13.104 | -71.599 | 3006 | 21.8 | 4084 | cloud forest | 12 | 1 | 11 | 3 | 3 | 6 |
| 33 | -13.098 | -71.597 | 2933 | 17.7 | 4213 | cloud forest | 10 | 4 | 6 | 3 | 3 | 4 |
| 34 | -13.070 | -71.560 | 2064 | 8.7 | 4973 | cloud forest | 12 | 3 | 9 | 6 | 7 | 8 |
| 35 | -13.070 | -71.560 | 2048 | 9.2 | 4991 | cloud forest | 14 | 4 | 10 | 7 | 7 | 8 |
| 36 | -13.068 | -71.559 | 1994 | 8.2 | 5056 | cloud forest | 11 | 6 | 5 | 6 | 6 | 7 |
| 37 | -13.067 | -71.559 | 1992 | 8.2 | 5059 | cloud forest | 15 | 3 | 12 | 6 | 8 | 10 |
| 38 | -13.065 | -71.556 | 1891 | 10.1 | 5191 | cloud forest | 22 | 8 | 14 | 8 | 9 | 13 |
| 39 | -13.066 | -71.556 | 1920 | 8.2 | 5152 | cloud forest | 12 | 7 | 5 | 6 | 6 | 8 |
| 40 | -13.065 | -71.555 | 1839 | 13.3 | 5265 | cloud forest | 15 | 1 | 14 | 7 | 9 | 9 |
| 41 | -13.064 | -71.555 | 1820 | 14.4 | 5292 | cloud forest | 17 | 8 | 9 | 4 | 6 | 11 |
| 42 | -13.074 | -71.565 | 2217 | 17.7 | 4789 | cloud forest | 13 | 6 | 7 | 7 | 6 | 10 |
| 43 | -13.075 | -71.565 | 2121 | 17.7 | 4789 | cloud forest | 11 | 1 | 10 | 5 | 5 | 7 |
| 44 | -13.074 | -71.565 | 2217 | 18.7 | 4789 | cloud forest | 13 | 10 | 3 | 4 | 5 | 11 |
| 45 | -13.073 | -71.564 | 2130 | 18.7 | 4789 | cloud forest | 9 | 3 | 6 | 7 | 7 | 8 |
| 46 | -13.044 | -71.536 | 1474 | 17.7 | 5754 | sub-montane | 15 | 2 | 13 | 5 | 6 | 8 |
| 47 | -13.043 | -71.537 | 1514 | 17.7 | 5754 | sub-montane | 24 | 8 | 16 | 10 | 10 | 13 |
| 48 | -13.042 | -71.543 | 1772 | 18.9 | 5395 | sub-montane | 19 | 11 | 8 | 13 | 12 | 16 |
| 49 | -13.042 | -71.541 | 1732 | 18.9 | 5395 | sub-montane | 20 | 2 | 18 | 7 | 11 | 12 |
| 50 | -12.954 | -71.565 | 937 | 17.7 | 5849 | sub-montane | 18 | 4 | 14 | 9 | 10 | 14 |
| 51 | -12.954 | -71.566 | 938 | 18.9 | 5849 | sub-montane | 10 | 6 | 4 | 6 | 6 | 9 |
| 52 | -12.954 | -71.567 | 946 | 22.1 | 5849 | sub-montane | 22 | 13 | 9 | 7 | 8 | 14 |
| 53 | -12.954 | -71.567 | 944 | 21.8 | 5849 | sub-montane | 23 | 13 | 10 | 8 | 11 | 17 |
| 54 | -11.900 | -71.370 | 400 | 25.0 | 1366 | lowland forest | 9 | 0 | 9 | 0 | 0 | 9 |
| 55 | -11.900 | -71.370 | 400 | 25.0 | 1366 | lowland forest | 16 | 0 | 16 | 11 | 11 | 11 |
| 56 | -11.900 | -71.370 | 400 | 25.0 | 1366 | lowland forest | 15 | 0 | 15 | 0 | 0 | 15 |
| 57 | -11.900 | -71.370 | 400 | 25.0 | 1366 | lowland forest | 14 | 0 | 14 | 0 | 0 | 14 |
| 58 | -11.900 | -71.370 | 400 | 25.0 | 1366 | lowland forest | 12 | 0 | 12 | 11 | 12 | 12 |
Table 2.
Community Weighted Means of leaf drought tolerance metrics (se = standard error).
| Plot | Leakage (%) | L_se | TLP (MPa) | TLP_se | Gmin (mmol m-2 s-1) | Gmin_se |
|---|---|---|---|---|---|---|
| 1 | 36.40 | 6.74 | -0.95 | 0.15 | 26.43 | 5.00 |
| 2 | 34.26 | 2.30 | -1.20 | 0.02 | 8.87 | 2.04 |
| 3 | 28.37 | 4.07 | -0.69 | 0.12 | 7.06 | 0.22 |
| 4 | 30.47 | 3.77 | -1.04 | 0.04 | 4.18 | 0.00 |
| 5 | 53.59 | 1.90 | NA | NA | NA | NA |
| 6 | 41.06 | 1.28 | NA | NA | NA | NA |
| 7 | 55.77 | 6.78 | NA | NA | NA | NA |
| 8 | 51.76 | 4.23 | -0.44 | 0.04 | 33.88 | 6.53 |
| 9 | 53.79 | 20.95 | -0.55 | 0.10 | 28.86 | 3.21 |
| 10 | 50.96 | 2.07 | -1.42 | 0.31 | 21.11 | 4.92 |
| 11 | 54.59 | 4.64 | -0.96 | 0.22 | 12.73 | 1.65 |
| 12 | 81.36 | 1.17 | -0.36 | 0.01 | 23.20 | 2.61 |
| 13 | 76.05 | 2.06 | -0.94 | 0.03 | 22.48 | 1.99 |
| 14 | 58.32 | 4.62 | -0.96 | 0.15 | 26.02 | 3.02 |
| 15 | 56.19 | 3.21 | -0.93 | 0.09 | 20.46 | 4.26 |
| 16 | 56.33 | 2.65 | -0.51 | 0.03 | 17.98 | 3.37 |
| 17 | 61.42 | 2.02 | -0.89 | 0.06 | 11.60 | 0.84 |
| 18 | 55.12 | 2.16 | -0.65 | 0.07 | 11.97 | 1.10 |
| 19 | 13.62 | 1.24 | NA | NA | NA | NA |
| 20 | 26.02 | 4.38 | NA | NA | NA | NA |
| 21 | 19.50 | 2.62 | NA | NA | NA | NA |
| 22 | 46.82 | 2.68 | -1.28 | 0.02 | 35.85 | 7.35 |
| 23 | 52.47 | 3.55 | NA | NA | NA | NA |
| 24 | 50.92 | 5.19 | -1.16 | 0.07 | NA | NA |
| 25 | 59.04 | 5.72 | NA | NA | 47.17 | 8.20 |
| 26 | 58.89 | 6.56 | -0.96 | 0.04 | 99.11 | 14.28 |
| 27 | 49.76 | 6.76 | NA | NA | NA | NA |
| 28 | 57.37 | 8.18 | NA | NA | 96.76 | 8.34 |
| 29 | 40.11 | 2.95 | -0.87 | 0.00 | NA | NA |
| 30 | 40.69 | 5.38 | -0.62 | 0.06 | 15.63 | 2.25 |
| 31 | 36.21 | 3.47 | -0.37 | 0.07 | 74.28 | 22.50 |
| 32 | 55.23 | 3.52 | -0.60 | 0.06 | 31.21 | 0.00 |
| 33 | 44.89 | 2.93 | NA | NA | 57.21 | 19.77 |
| 34 | 50.80 | 4.98 | NA | NA | NA | NA |
| 35 | 48.40 | 2.65 | NA | NA | 21.30 | 5.44 |
| 36 | 52.36 | 3.51 | NA | NA | NA | NA |
| 37 | 57.97 | 3.21 | NA | NA | 23.95 | 3.87 |
| 38 | 47.25 | 2.53 | -0.70 | 0.06 | 17.71 | 3.38 |
| 39 | 43.25 | 2.94 | NA | NA | NA | NA |
| 40 | 51.18 | 4.49 | -0.96 | 0.09 | 17.76 | 2.39 |
| 41 | 43.68 | 2.71 | NA | NA | NA | NA |
| 42 | 46.59 | 4.25 | -1.02 | 0.09 | NA | NA |
| 43 | 55.56 | 3.60 | NA | NA | NA | NA |
| 44 | 56.27 | 4.06 | NA | NA | NA | NA |
| 45 | 49.21 | 4.06 | -0.81 | 0.11 | NA | NA |
| 46 | 66.56 | 3.82 | NA | NA | NA | NA |
| 47 | 66.67 | 3.39 | NA | NA | NA | NA |
| 48 | 69.45 | 3.56 | NA | NA | NA | NA |
| 49 | 66.79 | 4.22 | NA | NA | NA | NA |
| 50 | 75.95 | 3.47 | -1.01 | 0.11 | NA | NA |
| 51 | 76.98 | 3.78 | NA | NA | NA | NA |
| 52 | 66.99 | 4.14 | -0.84 | 0.03 | NA | NA |
| 53 | 57.47 | 3.80 | NA | NA | NA | NA |
| 54 | 46.46 | 5.40 | NA | NA | NA | NA |
| 55 | 38.61 | 3.47 | NA | NA | NA | NA |
| 56 | 48.30 | 3.35 | NA | NA | NA | NA |
| 57 | 41.27 | 3.77 | NA | NA | NA | NA |
| 58 | 39.19 | 3.52 | NA | NA | NA | NA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



