
Submitted:
27 June 2023
Posted:
28 June 2023
You are already at the latest version
Abstract
Soil is a crucial component for plant growth, as it provides water, nutrients, and mechanical support. Various factors, such as crop cultivation, microflora, nutrient addition, and water availability, significantly affect soil properties. Maintaining soil health is important, and one approach is the introduction of native organisms with multifaceted activities. In this study, the effects of four native microbes (Trichoderma asperellum strain TAIK 1, Bacillus cabrialesii strain BIK3, Pseudomonas putida strain PIK1, and Pseudomonas otitidis strain POPS1) and their consortia on soil health, plant growth, and the incidence of stem rot disease caused by Sclerotium oryzae in rice are evaluated. Upon bioagent treatment of soil through seed treatment or soil application, variations in chemical properties of the soil were observed, viz., pH, Electrical Conductivity (EC), Organic Carbon (OC), available Soil Nitrogen (SN), Soil Phosphorus (SP), Soil Potassium (SK), and soil enzymes (urease, acid and alkaline phosphatase, dehydrogenase), compared to untreated soils. The treated seeds with the consortia of four native bioagents resulted in a significant increase in plant height (39.16%), number of panicles (30.29%), and average grain yield (41.36%) over control plants. Under controlled conditions, the bioagents-treated plants showed a 69.37% reduction in stem rot disease. The findings of this study indicate a positive correlation between soil properties and plant growth as well as a highly negative association with stem rot disease severity. The results suggest that using native bioagents as a management strategy can control stem rot disease and enhance crop productivity while reducing reliance on chemical interventions. These findings provide valuable insights for the development of sustainable agricultural practices that promote soil health, plant growth, and disease management.
Keywords:
Trichoderma
; Bacillus
; Pseudomonas
; Soil health
; Sclerotium oryzae
; Rice
1. Introduction
Soil is a complex mixture of minerals, organic matter, water, air, and microflora that supports plant growth, water and nutrient storage, and ecosystem processes [1,2,3]. A healthy soil with diverse microflora, a balanced pH, adequate EC, available nutrients, and increased enzymatic activity significantly affects plant health and productivity [4,5]. In order to meet the needs of an increasing population and decreasing land availability, intensive agricultural practices with high inputs of fertilizers and pesticides are being adopted [6,7]. As a result of such disproportionate use of chemicals, soil is negatively influenced, leading to improper soil and reduced microflora [6]. Thus, in the above context, the use of beneficial microbes in agriculture has the potential to maintain soil health and enhance crop production in a sustainable manner by supplying requisite nutrients to the soil and increasing nutrient availability for plants [8,9,10].
Native bioagents, viz., Trichoderma, Bacillus, and Pseudomonas, present in soil play a crucial role in enhancing various soil properties and plant growth [11,12]. These bioagents contribute to soil pH by participating in biological processes such as nutrient cycling and organic matter decomposition, releasing acids or bases that can modify soil pH over time [13,14]. They also regulate EC by facilitating salt leaching and reducing soil salinity, contributing to a more balanced EC [15,16]. In addition, native bioagents aid in enhancing soil OC levels by decomposing organic matter and promoting nutrient availability, contributing to soil fertility and structure [17,18]. These microorganisms also influence soil NPK availability by fixing atmospheric nitrogen, solubilizing phosphorus, and mobilizing potassium, enhancing plant growth and development [19,20].
These bioagents create less favourable conditions for the growth and development of the soil-borne pathogen S. oryzae, which causes stem rot disease in rice, either directly or indirectly by manipulating the soil nutrients and chemical composition, and such soils are called suppressive soils [21]. Stem rot disease of rice is generally managed by the application of chemical fungicides, and these practices are less preferred due to the awareness of the residual effects of fungicides in the grains, damage to human and animal health, and pollution of the environment [22]. Through their combined effects on soil properties and their direct antagonistic activity against stem rot pathogens, these native bioagents create a synergistic effect that ultimately leads to a reduction in the occurrence of stem rot disease in rice. This integrated approach of harnessing the beneficial activities of Trichoderma, Pseudomonas, and Bacillus, which enhance soil health, may contribute to a sustainable disease management strategy and promote the overall productivity of rice crops.
2. Materials and Methods
2.1. Fungal pathogen and Bio-control agents
The pathogen responsible for stem rot was isolated from samples obtained from infected plants in farmers’ fields. A standard tissue isolation procedure was followed as given by Bashyal et al. [23]. The pathogenicity of the pathogen in the rice TN1 cultivar has been established through the application of Koch’s postulates. Potential bioagent pure cultures, viz., TAIK 1 (NCBI Accession No.: MH825714, Whole Genome Sequence: JA1AZZ01), BIK 3 (NCBI Accession No.: MW181668, Whole Genome Sequence: JAHKKH01), and PIK 1 (NCBI Accession No.: ON778610), were obtained from culture collections at ICAR-IIRR, Hyderabad, Telangana, India. Pseudomonas otitidis-strain POPS1 was isolated as part of Ph.D. work characterised and submitted (NCBI Accession No.: ON782043).
2.2. Antagonism Assay
The bio-efficiency of the four native BCAs against stem rot pathogen was analysed using a dual culture assay on PDA plates [24]. For bacterial isolates, their effectiveness was evaluated by streaking a loopful of bacteria on both sides of a pathogen disc positioned at the center of a plate. The plates were then incubated in a BOD (Biological Oxygen Demand) chamber at a temperature of 25 ± 2 °C for a period of 4 days, allowing the pathogen to grow and reach a maximum radial growth of 9 cm on the control plate. The radial growth of the pathogens was measured and recorded. The percent inhibition was calculated using the formula described by Gangwar and Sinha [25], as given below:
where, C = colony growth in control plate (cm)
T = colony growth in treated plate (cm)
2.3. In vivo application of S. oryzae and its severity in plants
The effects of four bioagents and their consortia on plant growth-promoting and yield attributes were investigated. Various growth-promoting parameters, including seedling length, seedling fresh weight, and seedling dry weight, were measured at different time intervals, such as 7, 14, 21, 45, 60, 90, and 120 days after sowing (DAS). Yield-attributing traits, viz., panicle length, test weight, number of tillers, grain size, and yield per hill, were evaluated at the harvesting stage of the treated plants.
To study the impact of individual and consortia of bioagents on stem rot pathogen S. oryzae, plants were inoculated by placing 5 mm-long dried stem bits multiplied with pathogen from the TN1 variety at 45 DAS, specifically during the maximum tillering stage [26]. The inoculation process of S. oryzae on rice plants under glass house conditions is shown in Figure 1. The infection rate was assessed by recording the percentage of infected tillers at the time of maturity. The experiment was conducted in a completely randomized design (CRD) with three replications. Observations were recorded in terms of the percent of infected tillers at the time of maturity. The severity of the disease was assessed using a scale ranging from 0 to 9, as defined by the SES (Standard Evaluation System) by IRRI [27]. Subsequently, the recorded data was transformed into a percent disease index (PDI) using the following formula:
2.4. Seed and soil treatment
Sterile, pre-soaked seeds of rice cultivar TN1 (stem rot susceptible) were treated with suspensions of bioagents, viz., TAIK 1 (22 × 103 spores/ml), BIK 3, PIK 1, and POPS 1 (2.5 × 108 cfu/ml), and incubated for 6 h. Distilled water-treated seeds were considered a negative control. The treated seeds were transferred to pots of size 30 × 25 cm, filled with 5–7 kg of autoclaved soil. The bioagent suspension was applied to each pot at 10 ml/kg of soil and mixed thoroughly.
2.5. Sample collection and estimation of defense-related enzymes
Fresh leaves were harvested at different time intervals like 24 h, 48 h, 72 h, 96 h, and 120 h after pathogen inoculation (hapi), representing different treatments. Randomly selected plants were used, with three replicates per treatment. The harvested leaves were placed in sterile polybags. To ensure aseptic conditions, the collected samples were rinsed with sterile distilled water. Subsequently, the samples were stored in a refrigerator at -80°C until further experimentation.
2.5.1. Phenylalanine ammonia-lyase (PAL) assay
The activity of phenylalanine ammonia-lyase (PAL) was estimated as per the procedure given by Brueske [28]. Fresh leaves (0.5 g) from each replicate were ground in 4 ml of 0.2 M borate buffer containing 1.4 mM β-mercaptoethanol, using a pre-chilled mortar and pestle. The resultant enzyme extract was centrifuged at 16,000 rpm at 4 °C for 15 minutes. The reaction mixture was prepared by combining 2 ml of 0.2 M borate buffer (pH 8.7), 0.2 ml of 0.1M L-phenylalanine, and 2 ml of enzyme extract and incubated for 30 minutes at 32 ± 2°C. Finally, 0.5 ml of 1M trichloroacteic acid was added to stop the reaction, and the absorbance of the whole reaction mixture was measured in a spectrophotometer at 290 nm (µmoles of TCA/mg leaves/min).
2.5.2. Peroxidase (PO) assay
Peroxidase (PO) activity, expressed as µmoles/g leaves/min, was determined from the fresh leaves (0.5 g) from each treatment homogenized in 1 ml of 0.1 M potassium phosphate buffer. The reaction mixture consisted of 2 ml of 0.1 M potassium phosphate buffer (pH 7.0), 100 µl of enzyme extract, and 100 µl of 2 Mm H2O2 was added. The change in absorbance was measured at 436 nm for 2 minutes at 30 second intervals [29].
2.5.3. Polyphenol oxidase (PPO) assay
Leaf samples weighing 0.5 g were homogenized with 5.0 ml of 0.1 M sodium phosphate buffer (pH 6.5) using a pre-chilled mortar and pestle. The homogenate was centrifuged at 16,000 rpm for 15 minutes at 4 °C. The resulting enzyme extract was collected for further analysis. For the PPO activity assay, a reaction mixture was prepared by combining 0.4 ml of the enzyme extract, 0.4 ml of 0.01 M catechol, and 3.0 ml of 0.1 M sodium phosphate buffer (pH 6.5). The mixture was incubated at 28 ± 2 °C for 5 minutes. The absorbance of the reaction mixture was measured at 495 nm. PPO activity was determined by recording the changes in absorbance every 30 seconds for a total of 3 minutes. The recorded PPO activity was expressed as µmoles/g leaves/min [30].
2.5.4. Total phenol content (TPC) assay
The total phenol content (TPC) was measured using a modified methodology outlined by Malick and Singh [31]. Leaf samples weighing 0.1 g from each replicate treatment were crushed in 5 ml of 50% methanol. The samples were incubated for 1 h, and then further centrifuged at 13,000 rpm for 15 minutes. From the obtained methanolic enzyme extract, 0.1 ml was taken in a tube, the final volume was adjusted to 1 ml using distilled water, and 0.5 ml of folin-ciocalteu reagent was thoroughly mixed. To this 1 ml of 20% sodium carbonate was added after 15 minutes and vortexed. The mixture was left to stand for 1 h at room temperature. The absorbance of the changes in the reaction mixture was recorded at 725 nm and expressed as μg gallic acid (GA)/g fresh weight.
2.6. Soil analysis
2.6.1. Estimation of soil chemical properties
Changes in the soil pH, EC and OC were determined according to McLean [32], Rhoades [33], and Walkley and Black [34], respectively. Available nitrogen was estimated by the alkaline permanganate method with the macro Kjeldahl distillation unit [35]. Soil is alkalified with 2.5% NaOH, and 0.32% KMnO4 was added. Ammonia liberated from soil was trapped in a standard boric acid solution. The obtained solution was titrated with 0.02 N H2SO4. In the case of available phosphorus, soil samples were extracted with 0.5 M NaHCO3 buffered at pH 8.5, and the phosphorus in the extract was estimated by an ascorbic acid method using a spectrophotometer at 660 nm [36]. The available potassium in the soil was extracted with 1N neutral ammonium acetate (pH 7.0), and the potassium in the extract was determined by a flame photometer [37], respectively.
2.6.2. Estimation of soil enzymes
2.6.2.1. Assay of dehydrogenase enzyme
Dehydrogenase activity in the soil was assessed by adding 0.2 ml of 3% Triphenyl Tetrazolium Chloride (TTC) solution and 0.5 ml of 1% glucose to 1 g of soil and incubated for 24 h at 28 ± 0.5°C. Then add 10 ml of methanol and stand for 6 h. The amount of dehydrogenase activity was measured and expressed as mg Triphenyl Formazan (TPF)/g of soil ha-1 at 485 nm absorbance [38].
2.6.2.2. Assay of phosphatases enzyme
For detection of acid and alkaline phosphatase activity, 4 ml of expand it Modified Universal Buffer (MUB) phosphate buffer (pH 6.5), 0.2 ml toluene and to a 1 g soil sample, 1 ml of p-nitrophenyl phosphate (disodium salt hexahydrate) solution was added, followed by incubation for 1 h at 37 °C, then add 1 ml of 0.5 M CaCl2 and 4 ml of 0.5 M NaOH. The formation of p-nitrophenol (p-NP) was explored spectrophotometrically at 440 nm and expressed as µg of p-nitrophenol released g−1 soil ha−1 [39,40].
2.6.2.3. Assay of urease enzyme
To determine urease activity in soil, 1 ml of urea solution, 10 ml of 2 M KCI-PMA buffer, and 6 ml of coloring reagent were added to 1 g of soil, and the soil samples were incubated for 5 h at 37 °C. The absorbance of the red color developed at 527 nm was measured using a spectrophotometer and expressed as µg urea hydrolyzed g−1 soil ha−1 [40].
2.7. Estimation of plant nutrients analysis
The plant samples collected at various time intervals were initially air-dried in the shade and then further dried in a hot air oven at 65 °C until a constant weight was achieved. The dried samples were finely ground into a powder using a Willey mill. The nitrogen content in the plant samples was determined using the micro-Kjeldahl distillation method described by Piper [41]. The powdered plant sample was digested using a mixture of three acids (HNO3, H2SO4, and HClO4) in a ratio of 10:4:1. The digested sample was filtered through Whatman No. 42 filter paper, and the residue was washed with double-distilled water until free from chloride, following the method by Bhargava and Raghupathi [42]. The clear extract obtained from the triacid mixture was used to determine the phosphorus (P) and potassium (K) content using a spectrophotometer at 470 nm [41] and a flame photometer [37], respectively.
2.8. Statistical analysis
The experiments were carried out using a completely randomized design (CRD), and the data obtained were analyzed using one-way analysis of variance (ANOVA). A post hoc test was conducted using Duncan’s multiple range test (DMRT) at a significance level of 5% (P ≤ 0.05) using OPSTAT software. During each experiment, three replications were maintained. Correlation and stepwise regression analysis were performed using SAS Version 9.3 software, which was available at ICAR-IIRR. The regression model, represented in matrix notation, can be expressed as follows:
Y = Xβ + e
In this equation, Y represents the response variable, X denotes the vector of exogenous variables, β represents the vector of regression coefficients, and e represents the residual term assumed to follow a normal distribution with e∼N (0, σ^2). Principal Component Analysis (PCA) identifies the variance within a data set and allows to understand the key variables and spot outliers in the data. PCA was carried out using ‘FactoMineR’ [43] and ‘Factoextra’ [44] R packages.
3. Results
3.1. Antagonism assay
Native BCAs under in vitro conditions were evaluated for their antagonistic nature against S. oryzae. TAIK 1 showed the maximum inhibition percentage of 62.7% over the control, followed by PIK 1 (61.56%), and the least inhibition was recorded in POPS 1 with 56.38% (Figure 2).
3.2. In vivo application of S. oryzae and its severity in plants
3.3. Plant growth-promotion and yield attributes
Plants treated with the consortia of four native BCAs resulted in 100% seed germination and an increase of 58.08% in shoot length, 74.79% in root length, 56.66% in fresh weight, and 48.62% in dry weight over the control. The maximum growth promotion was exhibited by the consortia of BCAs, followed by TAIK 1 and PIK 1, and the least was recorded in POPS 1 treated plants (Table 1 and Figure 4).
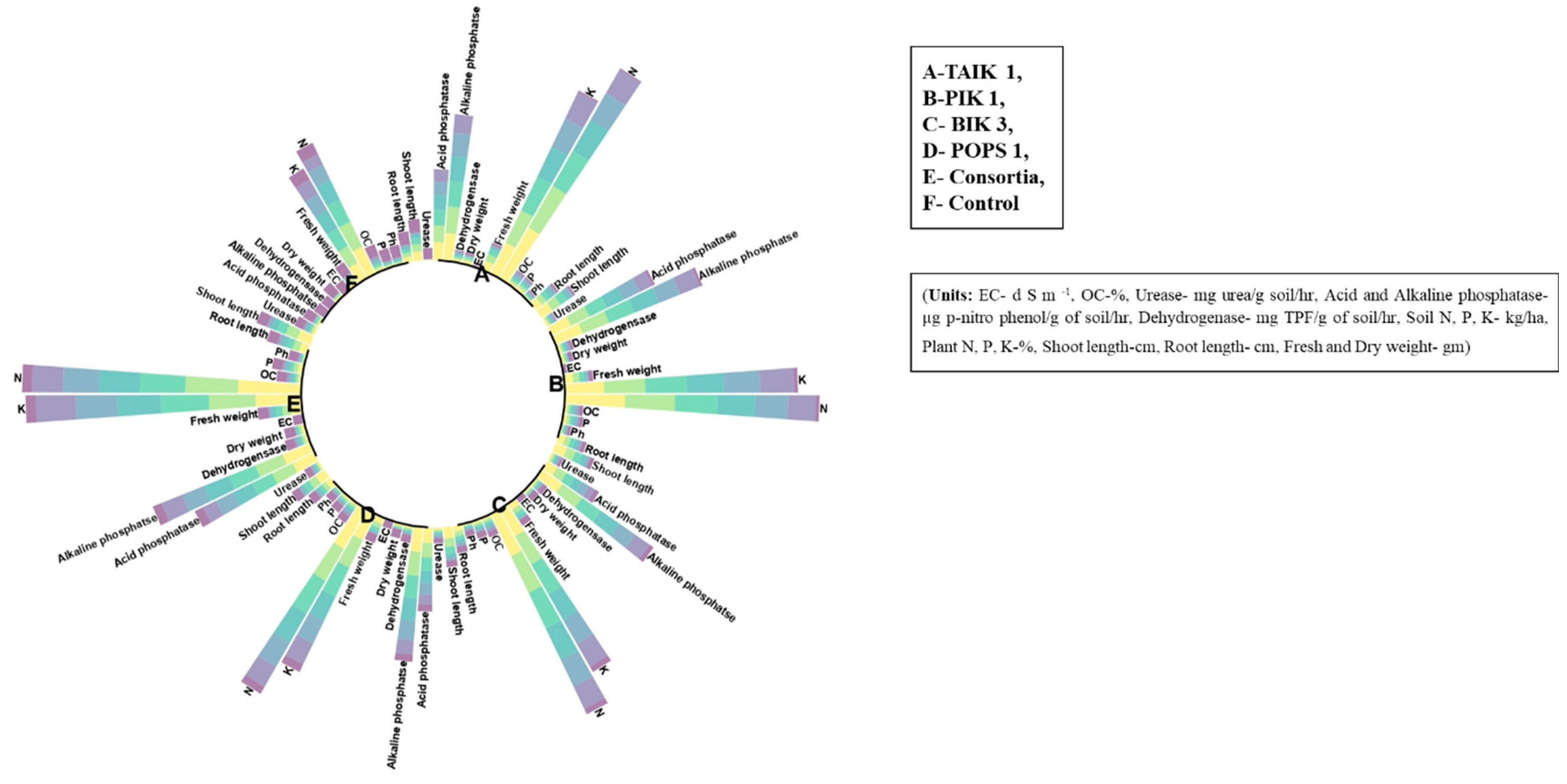
Similar results were recorded in the yield attributes, viz., the bioagents-treated plants showed significantly increased no. of tillers, panicle length, grain size (Kernel length-KL and Kernel breadth-KB), test weight, and yield per hill compared to the control. Consortia of four BCA-treated plants showed a 34.38% percentage increase in panicle length, followed by TAIK 1 (23.96%), PIK 1 (18.75%), BIK 3 (10.42%), and POPS 1 (4.17%) over control plants. Improved grain size in consortia treated plants with KL-6.2 cm and KB-2.9 cm, followed by TAIK 1 (KL-6.0 cm and KB-2.7 cm), PIK 1 (KL-5.8 cm and KB-2.6 cm), BIK 3 (KL-5.7 cm and KB-2.4 cm), and POPS 1 (KL-5.6 cm and KB-2.3 cm), more test weight (24.1 g) and yield/hill (19.1 g), TAIK 1 (17.1 g), PIK 1 (16.1 g), BIK 3 (15.0 g), and POPS 1 (14.0 g). Least was recorded in control untreated plants with a panicle length of 19.2 cm, grain size of KL-5.4 cm and KB-2.1 cm, test weight of 15.2 g, and yield per hill of 11.2 g (Table 1). A stacked circular bar plot depicted the changes in plant growth-promoting parameters and yield attributes over a crop period at regular intervals upon treatment with single and consortia of bioagents (Figure 6).

Table 1.
Descriptive statistics of changes in plant growth, yield, and disease index parameters upon single and consortia of bioagents application. The data represents the mean ± SE of 10 seedlings from 2 experiments collected at different stages of growth, viz., 7, 14, 21, 45, 60, and 90 DAT of bioagents. Numerical values with different letters are significantly different (P < 0.05, DMRT, OPSTAT).
Table 1.
Descriptive statistics of changes in plant growth, yield, and disease index parameters upon single and consortia of bioagents application. The data represents the mean ± SE of 10 seedlings from 2 experiments collected at different stages of growth, viz., 7, 14, 21, 45, 60, and 90 DAT of bioagents. Numerical values with different letters are significantly different (P < 0.05, DMRT, OPSTAT).
| Bioagents | Shoot Length | Root Length | Fresh Weight | Dry Weight | No. of tillers | No. of panicles | Kernel length (mm) | Kernel breadth (mm) | Panicle length (cm) | Test weight (gm) | Grain Yield /hill | PDI (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (gm) | ||||||||||||
| PIK1 | 48.77c | 31.20c | 32.20c | 10.50c | 19.82b | 8.44c | 5.80bc | 2.60ab | 22.80c | 21.50c | 16.10c | 26.38d |
| POPS 1 | 41.60e | 25.20d | 23.30e | 7.83e | 18.99c | 7.99d | 5.60cd | 2.30bc | 20.00e | 18.50e | 14.00e | 38.73b |
| BIK 3 | 44.07d | 25.77d | 26.50d | 8.93d | 19.01c | 8.11d | 5.70c | 2.40bc | 21.20d | 20.00d | 15.00d | 36.88c |
| TAIK 1 | 50.90b | 33.87b | 35.67b | 12.00b | 20.11d | 9.63b | 6.00ab | 2.70ab | 23.80b | 22.20b | 17.10b | 24.06e |
| Consortia | 58.33a | 39.03a | 41.53a | 13.57a | 22.32a | 10.50a | 6.20a | 2.90a | 25.80a | 24.10a | 19.10a | 19.32f |
| Control | 36.90f | 22.33e | 18.00f | 6.97f | 18.54d | 7.32e | 5.40d | 2.10c | 19.20f | 17.70f | 11.20f | 75.24a |
| CD | 0.358 | 0.605 | 0.196 | 0.181 | 0.361 | 0.285 | 0.067 | 0.149 | 0.088 | 0.096 | 0.306 | 2.006 |
| CV | 0.647 | 1.267 | 0.647 | 1.830 | 0.663 | 0.687 | 0.661 | 3.461 | 0.231 | 0.368 | 0.871 | 3.290 |
| SEm | 0.116 | 0.197 | 0.064 | 0.059 | 0.034 | 0.025 | 0.022 | 0.049 | 0.029 | 0.031 | 0.018 | 0.023 |
3.4. Biochemical parameters
The defense-related enzyme activities were assessed in plants treated with individual and consortia of bioagents at different time intervals. The activity of defense enzymes showed an increase after 24 hapi of treatment, reached its peak at 72 hapi, and subsequently declined. The highest activity of PAL, PO, PPO, and TPC was observed in consortia of BCAs + S. oryzae, followed by TAIK 1 + S. oryzae. Specifically, the highest PAL activity was recorded in plants treated with the consortia of BCAs + S. oryzae at 24, 74, and 120 hapi (3.387, 5.961, and 5.602 µmoles/g leaves/min, respectively). In contrast, the control plants exhibited the lowest PAL activity at the same intervals (1.347, 1.764, and 1.474 µmoles/g leaves/min, respectively). The PO activity was highest in the treatment involving the consortia of BCAs + S. oryzae at 24, 74, and 120 hapi (0.693, 0.736, and 0.714 mmoles/g leaves/min, respectively), while the control showed the lowest PO activity at all intervals (0.04, 0.07, and 0.05 mmoles/g leaves/min, respectively). Similarly, the PPO activity was highest in the treatment with BCAs + S. oryzae at 24, 74, and 120 hapi (0.071, 0.093, and 0.078 nmoles/g leaves/min, respectively), whereas the control exhibited the lowest activity at all intervals (0.005, 0.015, and 0.007 nmoles/g leaves/min, respectively). The maximum activity of TPC was observed in the consortia of BCAs + S. oryzae at 24, 74, and 120 hapi (2.932, 4.932, and 3.932 µg phenols/g leaves/min, respectively), while the control showed the lowest activity at all intervals (0.587, 1.587, and 1.087 µg phenols/g leaves/min, respectively) (Figure 5).
3.5. Soil chemical properties
Bioagents treated soil under glass house conditions showed change in the pH, EC, and OC over untreated soils. Soil analysis resulted in significant increase in the bioagents treated soils in comparison with control untreated soil—pH (8.1 to 8.2), EC (0.72 to 0.76 d S m -1), and OC (0.57 to 0.68 %) (Table 2).
Availability of soil N, P, and K was found to be increased in consortia treated soils. Results indicated the consortia of all the four bioagents observed to increase N (199.5 to 315.82 kg/ha), P (13.3 to 24.91 kg/ha) and K (256.48 to 288.28 kg/ha) over a period of 120 DAS. Followed by TAIK 1 showed significant increase in N (179.55 to 297.67 kg/ha), P (11.0 to 19.67 kg/ha), K (218.74 to 249.42 kg/ha) in comparison with the control untreated soil showed N (75.6 to 155.18 kg/ha), P (5.0 to 7.87 kg/ha), and K (107.41 to 121.29 kg/ha) (Table 2). A stacked circular bar plot depicted the changes in soil chemical properties over a crop period at regular intervals upon treatment with single and consortia of bioagents (Figure 6).
3.6. Soil enzymes
Under glass house conditions, the single and combined application of bioagents significantly influenced the activity of soil enzymes, viz., urease, acid phosphatase, alkaline phosphatase, and dehydrogenase, across all time intervals. Urease activity ranged from 0.73 to 7.33 mg of urea released per g of soil/ha, acid phosphatase activity ranged from 0.09 to 1.39 µg of p-nitrophenol released per g of soil/ha, alkaline phosphatase activity ranged from 0.90 to 1.78 µg of p-nitrophenol released per g of soil/ha, and dehydrogenase activity ranged from 0.14 to 16.44 mg of TPF produced per g of soil/ha in all the treatments. Among the treatments, consortia-treated soils exhibited a significant increase in enzymatic activity, followed by TAIK 1 and PIK 1-treated soils, while the control soil showed the lowest enzymatic activity (Table 2). A stacked circular bar plot depicted the changes in soil enzymes over a crop period at regular intervals upon treatment with single and consortia of bioagents (Figure 6).
3.7. Plant nutrients
Under glass house conditions, treatment with bioagents resulted in alterations of plant N, P, and K. Among the treatments, consortia-treated plant samples were found to have significantly higher levels of N (0.58%), P (0.11%), and K (1.58%) in comparison with the control plants recorded with N (0.21%), P (0.02%), and K (0.76%). However, TAIK 1 and PIK 1 treated samples also showed significant increases in plant N, P, and K content; least was observed in POPS 1 treated samples (Table 2). A stacked circular bar plot depicted the changes in plant nutrients over a crop period at regular intervals upon treatment with single and consortia of bioagents (Figure 6).
Table 2.
Descriptive statistics of changes in soil properties upon bioagents application over a crop period. The data represents the mean ±SE of 3 samples from 2 experiments collected at different stages of growth, viz., 7, 14, 21, 45, 60, and 90 DAT of bioagents. Numerical values with different letters are significantly different (P < 0.05, DMRT, OPSTAT).
Table 2.
Descriptive statistics of changes in soil properties upon bioagents application over a crop period. The data represents the mean ±SE of 3 samples from 2 experiments collected at different stages of growth, viz., 7, 14, 21, 45, 60, and 90 DAT of bioagents. Numerical values with different letters are significantly different (P < 0.05, DMRT, OPSTAT).
| Bioagents | Soil pH | EC | OC | Urease | Acid Phosphatase | Alkaline Phosphatase | Dehydrogenase | Soil N | Soil P | Soil K | Plant N | Plant P | Plant K |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PIK1 | 8.24a | 0.75a | 0.64a | 6.18b | 1.25ab | 1.66ab | 12.96c | 283.15c | 17.06c | 216.35c | 0.41b | 0.07ab | 1.23ab |
| POPS 1 | 8.15a | 0.73a | 0.60a | 5.85c | 0.85cd | 1.42b | 11.35d | 235.21e | 12.39e | 181.72e | 0.27c | 0.04b | 1.01ab |
| BIK 3 | 8.20a | 0.74a | 0.61a | 6.10b | 0.64d | 1.46b | 11.58d | 247.63d | 14.01d | 195.27d | 0.38b | 0.05ab | 1.10ab |
| TAIK 1 | 8.25a | 0.75a | 0.65a | 6.32b | 1.00bc | 1.67ab | 14.60b | 297.67b | 19.67b | 249.42b | 0.47b | 0.08ab | 0.93b |
| Consortia | 8.28a | 0.76a | 0.68a | 7.33a | 1.39a | 1.78a | 16.44a | 315.82a | 24.91a | 288.28a | 0.58a | 0.11a | 1.58a |
| Control | 8.08a | 0.72a | 0.57b | 0.73d | 0.09e | 0.90c | 0.14e | 155.18f | 7.87f | 121.29f | 0.21c | 0.02b | 0.76c |
| CD | 0.023 | 0.028 | 0.268 | 0.057 | 0.070 | 0.074 | 0.111 | 0.024 | 0.002 | 0.772 | 0.009 | 0.001 | 1.952 |
| CV | 1.578 | 1.926 | 1.360 | 2.727 | 1.335 | 1.960 | 9.548 | 0.352 | 1.993 | 2.430 | 1.342 | 5.914 | 2.564 |
| SEm | 0.075 | 0.008 | 0.127 | 0.018 | 0.022 | 0.024 | 0.035 | 0.008 | 0.001 | 0.252 | 0.003 | 0.001 | 0.638 |
Figure 6.
Stacked circular bar plot depicts the changes in soil properties and plant growth parameters over a crop period at regular intervals upon treatment with single and consortia of bioagents. Each bar represents the mean ± SE of 6 biological replicate plants from 2 experiments with significant differences (P < 0.05, DMRT, R) at 7, 14, 21, 45, 60, and 90 DAT.
Figure 6.
Stacked circular bar plot depicts the changes in soil properties and plant growth parameters over a crop period at regular intervals upon treatment with single and consortia of bioagents. Each bar represents the mean ± SE of 6 biological replicate plants from 2 experiments with significant differences (P < 0.05, DMRT, R) at 7, 14, 21, 45, 60, and 90 DAT.
3.8. Correlation analysis
Among the variables, plant growth-promotion activities (SL, RL, FW, DW, and Yield-Y) were found to have a highly significant positive correlation (Figure 7, dark blue) with the soil chemical properties, viz., pH, EC, OC, available soil N, P, and K (SN, SP, and SK). Similarly, the yield parameters, viz., panicle length, kernel length and breadth, and test weight, were found to have a moderately significant positive correlation with soil enzymes (Urease-U, Acid Phosphatase-AP, Alkaline Phosphatase-ALP, and Dehydrogenase-D). However, a highly significant negative correlation was observed between the percent disease severity-PDI and all the other parameters studied (Figure 7).
3.9. Stepwise regression analysis
The stepwise regression analysis was carried out to identify the factors influencing shoot length, root length, fresh weight, and dry weight of the plant (dependent variables) and the four bioagent treatments, viz., TAIK1, BIK3, PIK1, POPS1, and one consortia (independent variables), among soil properties. Though the stepwise regression analysis for soil enzymes was carried out over the 5 independent variables, TAIK 1 has retained the variability in SL, RL, FW and DW in plants for urease enzyme. This described 95%, 96%, 93%, and 94% variability in each model. These results depict that for each unit increase in urease enzyme in soil; there will be a change of 0.90 cm SL, 0.65 cm RL, 0.28 g FW, and 0.13 g DW in plants (Table 3, Table 4, Table 5 and Table 6).
Acid phosphatase analysis revealed that POPS1 and consortia are responsible for alterations in SL, RL, FW, and DW of plants. This inferred that for every unit increase in acid phosphatase enzyme in soil, there will be a change in 0.63 cm SL, 0.12 cm RL, 0.66 g, 0.36 g FW, and 0.13 g, 0.80 g DW in plants. In the case of alkaline phosphatase variability, it was retained by POPS 1, BIK 3, and TAIK 1, which explained 97% variation in the SL and RL models, 96% variation in the FW model, and 98% variation in the DW model. The results depict that for a unit increase in alkaline phosphatase activity, there is a 0.4 cm, 0.14 cm, and 0.55 cm variability in SL by POPS1, BIK3, and TAIK1, respectively. 0.96cm, 0.57 cm, and 0.43cm alterations in RL by POPS1, BIK3, and TAIK1, respectively. 0.12 g, 0.67 g, and 0.07 g change in FW by POPS1, BIK3, and TAIK1, respectively. 0.68 g, 0.44 g, and 0.51 g increase in dry weight by POPS1, BIK3, and TAIK1, respectively (Table 3, Table 4, Table 5 and Table 6).
Similarly, stepwise regression analysis for dehydrogenase enzyme was influenced by POPS1 and PIK1, which together explained 96%, 97%, 96%, and 96% of the variability in SL, RL, FW, and DW of plants, respectively. POPS1 and PIK1 have a positive effect on SL, RL, FW, and DW, and with a unit increase in enzyme activity, there was a change in 0.42 cm SL, 0.68 cm RL, 0.61 g FW, and 0.25 g DW (Table 3, Table 4, Table 5 and Table 6).
Regression analysis of plant nutrients N, P, and K has a positive effect on change in SL, RL, FW, and DW. The variation was retained by BIK3 (76% SL, 52% RL, 51% FW, 70% DW), consortia (18% SL, 25% RL, 23% FW), and PIK1 (40% DW). For every 1 percent increase in plant nitrogen, there was a variability of 0.44 cm SL (BIK3), 0.57cm SL (consortia), 0.43 cm RL (BIK3), 0.59 cm RL (consortia), 0.76 g FW (BIK3), 0.80 g FW (consortia), 0.40 g DW (PIK1), and 0.70 g DW (BIK3). For plant P variation, it was retained by POPS1 (0.2% SL, 0.2% DW), BIK3 (96% SL, 93% FW, 95% DW), and PIK1 (94% RL). For every 1 percent increase in plant phosphorus content, there is a change of 0.99 cm SL (POPS1), 0.49 cm SL (BIK3), 0.57 cm RL (PIK1), 0.25 g FW (BIK3), 0.50 g DW (POPS1), and 0.59 g DW (BIK3). For plant K variation, it was retained by PIK1 (0.3% SL, 0.3% RL, 0.6% FW, 0.4% DW), POPS1 (0.3% SL, 0.4% POPS1, 0.3% DW), BIK3 (0.1% SL, 0.1% FW, 0.2% DW), TAIK1 (0.2% RL), and consortia (91% SL, 92% RL, 88% FW, 89% DW). For every 1 percent increase in plant K, there is an increase of 0.70 cm SL (PIK1), 0.51 cm SL (POPS1), 0.48 cm SL (BIK3), 0.35 cm SL (consortia), 0.55 cm RL (PIK1) + 0.70 cm RL (TAIK1) + 0.49 cm RL (consortia), 0.45 g FW (PIK1), 0.72 g FW (POPS1), 0.67 g FW (BIK3), 0.72 g FW (consortia), 0.64 g DW (PIK1), 0.86 g DW (POPS 1), 0.72 g DW (BIK3), 0.95 g DW (consortia) (Table 3, Table 4, Table 5 and Table 6). The other soil parameters affected by bioagent treatment (SL, RL, FW, and DW) are mentioned in the Table 3, Table 4, Table 5 and Table 6.
3.10. PCA analysis
The effect of individual and consortia of bioagents on soil properties, plant growth, and yield parameters compared with the percent disease index was analysed using principal component analysis. The principal component was generated to explain the statistical variance of 97.0% (Figure 8). The first principal component (Dim 1) had the highest eigenvalue of 20.1, explaining 91.7% of the statistical variance, while the second principal component (Dim 2) had an eigenvalue of 1.14, explaining 5.2% of the statistical variance. A small angle indicates a positive correlation, while a large angle shows a negative correlation. However, the angle of 90° indicates no correlation between the given treatments. Based on the statistical analysis of the biplot (Figure 8), it was observed that all the variables with narrow angles (less than 90°) depict a strong correlation and affect each other directly, whereas PDI lies in another quadrant and depicts a negative correlation. Furthermore, Dim 1 on the positive axis was affected by U, AP, ALP, D, SN, SP, SK, PN, PP, PK, SL, RL, FW, DW, nT, nP, KL, KB, PL, TW, and GY, while Dim 2 on the positive axis was affected by PDI.
The PCA biplot (Figure 8) represents differences in the parameters in terms of bioagents treatment. The bioagents consortia and TAIK 1 lie in the same quadrant on the upper positive side of the Dim1 axis, indicating the maximum variability of plant growth parameters, i.e., PN, PP, PK, SL, RL, FW, DW, nT, nP, KL, KB, PL, and TW. While the bioagent PIK1 located on the lower positive side of Dim 1 indicated the maximum variability of soil properties, i.e., U, AP, ALP, D, SN, SP, and SK. And control located on the positive side of Dim 2 indicated the maximum variability of PDI. PCA results indicated a significant effect of different treatments (bioagents-consortia, TAIK1, and PIK1) on soil and plant growth parameters with a percent disease index.
4. Discussion
Soil-borne pathogens pose a significant threat to rice cultivation in India and elsewhere. The severity and significance of the damage caused have led to the development of effective management strategies. Extensive research has demonstrated that soil antagonistic microflora holds immense potential for suppressing soil-borne pathogens and promoting plant growth [45,46].
The bioagents selected, namely POPS1, PIK1, BIK3, and TAIK1, exhibited promising results in suppressing S. oryzae both in vitro and in vivo. Their suppression mechanism was attributed to the secretion of antimicrobial compounds such as bacteriocins, phenazines, and hydrocynaides (HCN) in the growth media, leading to the inhibition of pathogen growth [47,48]. Additionally, competition for nutrients and the production of enzymes such as chitinases, proteases, pectinases, and glucanases, which degrade pathogen cell walls, were reported as significant modes of action for Trichoderma spp. [49]. Trichoderma spp. was found to produce antimicrobial metabolites like Trichoviridin and Trichodermin. Similarly, Bacillus spp. was found to produce subtilin, bacitracin, bacillin, and bacillomycin, which also contribute to pathogen suppression by degrading cell walls [50]. Pseudomonas spp. are known to secrete secondary metabolites such as 2,4-diacetyl phloroglucinol, phenazine-1-carboxylic acid, phenazine-1-carboxamide, pyoluteorin, and pyrrolnitrin, which effectively inhibit plant pathogens [51].
In addition to their antagonistic activity against S. oryzae, the native BCAs used in this study also significantly increased seedling length, biomass, and yield. The bioagent-treated plants show improvement over the control plants, which may be due to the promotion of nutrient recycling, nitrogen fixation, phosphorus solubilization, and the production of plant growth-promoting hormones and some alkaloids like siderophores [45,52,53]. Further, the plant growth promotion activities of TAIK 1, BIK 3, PIK1, and POPS 1, which are endophytic in nature, and the secretion of major phytohormones IAA, GA, SA, ABA, and Zeatin earlier reported by this group Sowmya et al. [54].
In vivo studies indicated that effective suppression of S. oryzae results in a significant reduction in PDI over the control. The colonization of Trichoderma on the sclerotial bodies and mycelia of S. oryzae resulted in significant mycelial lysis, causing a loss of their viability and inhibiting their ability to germinate. These findings were observed through SEM studies [55,56]. The significant destruction of sclerotia is noteworthy, as these structures are produced by S. oryzae to withstand adverse conditions and survive during the off-season. The effective suppression of these pathogens by TAIK 1, resulting in sclerotial destruction, plays a crucial role in reducing the pathogen’s inoculum in the soil. This provides a sustainable management approach for controlling soil-borne pathogens in rice, both during the crop cycle and in the off-season. These findings have been documented in studies conducted by Halifu et al. [57] and Kannan et al. [45].
Defense enzymes are a significant component of microbial-induced systemic resistance in plants and play a vital role in their ability to fight against invading pathogens [58,59]. Phenylalanine ammonia-lyase (PAL) is the first enzyme to be activated in the plant defense response and is the precursor for various secondary metabolites, including phytoalexins, lignins, and flavonoids. These phytoalexins are toxic to invading pathogens by interfering with their protein and DNA pathways, inhibiting spore germination, and disrupting the cell membrane [60]. PAL effects pathogen development in plants by lignifying the cell wall, producing phenolic compounds, and accumulating reactive oxygen species (ROS) [61]. PO and PPO are involved in the oxidative cross-linking of cell wall components and provide physical barriers to pathogen movements in plants. These enzymes mainly induce defense-related genes and also detoxify pathogen-derived compounds [62]. Total phenolic content includes a wide variety of secondary metabolites, such as tannins and flavonoids that have antimicrobial properties. Phenolic compounds inhibit the enzymes released by the pathogens that are involved in cell wall degradation, leading to blockage of colonization and development [63]. Current study results revealed that consortia-treated plants significantly induced concentrations of defense-related enzymes such as PAL, PPO, PO, and TPC as compared to individual BCA treatments and untreated control.
An increase in soil pH, EC, and OC has a negative impact on soil-borne pathogens. A higher pH value indicates alkaline soil conditions that may be unfavourable to soil-borne fungal pathogens. In alkaline conditions, nutrient imbalance occurs in the soil, which may lead to the unavailability of essential nutrients and make soil-borne pathogens starve [64]. This nutrient imbalance also affects the diversity and abundance of microorganisms, including fungi, thus impacting their growth and survival [65]. In addition, high EC also leads to the accumulation of toxic ions such as chloride and sodium, which may further inhibit the growth and survival of soil-borne pathogens by creating undue osmotic stress [65].
Soil organic carbon improves the suppressive nature of the soil by promoting the growth of beneficial microorganisms that compete with and inhibit the growth and multiplication of fungal pathogens [66]. These beneficial microorganisms also produce compounds that inhibit the growth and reproduction of pathogens, or they can outcompete them for nutrients and other resources in the soil [67]. In our study, BCA-treated soils were found to increase pH, EC, and OC. Keim and Webster [68] proved that the application of native rhizosphere microflora leads to an increase in pH, EC, and OC levels. This increase creates a favorable environment that stimulates the growth and proliferation of beneficial microbes through quorum sensing. The beneficial microbe community secretes phenolic compounds such as coumarins, flavonoids, and tannins, as well as volatile organic compounds (VOCs) and organic acids. These compounds play a crucial role in disrupting the signalling pathways for sclerotial body germination. As a result, the growth and proliferation of pathogens are effectively inhibited [65].
Application of beneficial microorganisms, such as Trichoderma spp., in agricultural soils has positive effects on nutrient availability. Studies conducted by Yadav et al. [69] and Kapri and Tewari [70] reported the ability of Trichoderma spp. to enhance the solubilization of essential nutrients, viz., N, P, and K. These microorganisms release organic acids that acidify the soil, promoting the release of bound nutrients and making them more available to plants. Kucuk et al. [71] have further confirmed that the acidification caused by organic acids released by T. harzianum can provide additional nutrients to plants.
Soil enzymes, viz., urease, acid and alkaline phosphatase, and dehydrogenase, play a key role in sustaining soil health by influencing the soil nutrient cycles. Urease plays a crucial role in the hydrolysis of urea into ammonia and carbon dioxide [72]. Phosphatase plays an important role in the phosphorus cycle and increases phosphorus solubilization in plants, which is a good indicator for soil fertility and quality [73]. The dehydrogenase enzyme catalyzes the oxidation of organic matter in the soil, providing an indication of the microbial activity and metabolic potential of soil microorganisms [74]. Results obtained from our study indicated an increase in soil enzymes upon treatment with BCAs compared to untreated soils, as reported in earlier studies [75]. Soil enzymatic activity is known to have a negative correlation with pathogen development due to the increase in soil pH resulting from the production of ammonia by urease; alkaline conditions have a suppressive effect on some soil-borne pathogens [76]. Phosphorus solubilization by phosphatase makes plants uptake P, which is an essential component for sclerotia germination of the stem rot pathogen, which limits the pathogen’s development [77]. Higher levels of dehydrogenase activity are associated with a more diverse and healthier soil beneficial microbial community, which is better equipped to suppress the growth and survival of soil-borne pathogens.
Correlation analysis revealed the interdependence of plant growth and yield attributes with soil parameters, which are negatively correlated with the disease severity of plants. The PCA clearly showed the role of two principal components, viz., the application of consortia of bioagents and TAIK1, which contributed about 97% of total variability when compared to all the other factors involved. Soil enzymes are more influenced by PIK1 when compared to other bioagents. Stepwise regression analysis inferred that unit change variability in plant growth parameters is due to alterations in soil properties upon treatment with bioagents.
5. Conclusions
The present study reveals the impact of potential BCAs P. otitidis, P. putida, T. asperellum, and B. cabrialesii isolated from the native rice soils of Telangana. They were investigated for their impact on soil health, plant growth promotion, and antagonistic activity against the stem rot pathogen under both in vitro and in vivo conditions. It was observed that all the BCAs had a noticeable effect on soil characteristics such as pH, EC, OC, and soil enzymes, which in turn influenced the development of sclerotia of the soil-borne pathogen S. oryzae. These BCAs demonstrate potential as components of eco-friendly management strategies for stem rot disease. Additionally, they were found to promote plant growth, highlighting their significant role in developing sustainable integrated disease management (IDM) approaches for managing rice stem rot caused by S. oryzae.
Author Contributions
KC: Conceptualization, planning, and editing of manuscript; SV, MP, SK: Conducting of experiments, data analysis, and writing manuscript; GR, SR, SCH: Contributed reagents/materials/analysis tools; UG, RP, TM, ES, CK- Data analysis and Editing the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Department of Agricultural Research (DARE), Ministry of Agriculture and Farmers’ Welfare, Government of India. K.C. acknowledges research support from ICAR-IIRR. C.K. gratefully acknowledges the financial support from the project of the Ministry of Science and Higher Education of Russian Federation on the development of Young Scientist Laboratory within the framework of the Interregional Scientific and Educational Center of the South of Russia (no. LabNOTs-21-01AB, FENW-2021-0014) and to the Strategic Academic Leadership Program of the Southern Federal University (“Priority 2030”).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request from K.C.
Acknowledgments
The authors thank Mrs. Anusha for her help with this work. We would also like to acknowledge the help and support received from the Director and Head of Plant Pathology, IIRR and Head of Plant Pathology, PJTSAU, Rajendranagar, Hyderabad for this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Neher, D.A. Soil community composition and ecosystem processes: comparing agricultural ecosystems with natural ecosystems. Agrofor. Systems 1999, 45, 159–185. [Google Scholar] [CrossRef]
- Raj, A.; Jhariya, M.K.; Yadav, D.K.; Banerjee, A.; Meena, R.S. Soil for sustainable environment and ecosystems management. Sustainable agriculture, forest and environmental management 2019, 189–121. [Google Scholar]
- Fan, K.; Chu, H.; Eldridge, D.J.; Gaitan, J.J.; Liu, Y.R.; Sokoya, B.; Wang, J.T.; Hu, H.W.; He, J.Z.; Sun, W.; Cui, H. Soil biodiversity supports the delivery of multiple ecosystem functions in urban greenspaces. Nature Ecol. Evol. 2023, 7, 113–126. [Google Scholar] [CrossRef]
- Chaparro, J.M.; Sheflin, A.M.; Manter, D.K.; Vivanco, J.M. Manipulating the soil microbiome to increase soil health and plant fertility. Biol. Fertil Soils 2012, 48, 489–499. [Google Scholar] [CrossRef]
- Else, K.B.; Giulia, B.; Zhanguo, B.; Rachel, E.C.; Gerlinde, D.D.; Ron, G.; Luuk, F.; Violette, G.; Thom Erdel, E.; Simsek, U.; Kesimci, T.G. Effects of Fungi on Soil Organic Carbon and Soil Enzyme Activity under Agricultural and Pasture Land of Eastern Türkiye. Sustainability 2023, 15, 1765. [Google Scholar]
- Nikita, B.; Puneet, S.C. Excessive and Disproportionate Use of Chemicals Cause Soil Contamination and Nutritional. 2020. [Google Scholar] [CrossRef]
- Jagaba, A.H.; Kutty, S.R.M.; Isa, M.H.; Ghaleb, A.A.S.; Lawal, I.M.; Usman, A.K.; Birniwa, A.H.; Noor, A.; Abubakar, S.; Umaru, I.; Saeed, A.A.H. Toxic effects of xenobiotic compounds on the microbial community of activated sludge. Chem. Bio. Eng. Reviews 2022, 9, 497–535. [Google Scholar] [CrossRef]
- Punam, K.; Mukesh, M.; Pooja, G.; Manish, K.D.; Gopal, N.; Upadhyay, R.S. Plant growth promoting rhizobacteria and their biopriming for growth promotion in mung bean (Vigna radiata (L.) R. Wilczek). Biocatal. Agric. Biotechnol. 2018, 16, 163–171. [Google Scholar]
- Kumar, S.; Chandra, R.; Behera, L.; Keswani, C.; Sansinenea, E. Dual Trichoderma consortium mediated elevation of systemic defense response against early blight in potato. Eur. J. Plant Pathol. 2021, 162, 681–696. [Google Scholar] [CrossRef]
- de Andrade, L.A.; Santos, C.H.B.; Frezarin, E.T.; Sales, L.R.; Rigobelo, E.C. Plant growth-promoting rhizobacteria for sustainable agricultural production. Microorganisms 2023, 11, 1088. [Google Scholar] [CrossRef]
- Kumari, S.; Bharat, N.K.; Thakur, A.K. Role of plant growth-promoting rhizobacteria (PGPR) and bio-control agents (BCAs) in crop production. Int. J. Economic Plants 2020, 7, 144–150. [Google Scholar] [CrossRef]
- Mishra, R.K.; Sahu, P.K.; Mishra, V.; Jamal, H.; Varma, A.; Tripathi, S. Isolation and characterization of halotolerant plant growth promoting rhizobacteria from mangrove region of Sundarbans, India for enhanced crop productivity. Front. Plant Sci. 2023, 14, 1122347. [Google Scholar]
- Neina, D. The role of soil pH in plant nutrition and soil remediation. Appl. Env. Soil Sci. 2019, 1–9. [Google Scholar] [CrossRef]
- Shah, K.K.; Tripathi, S.; Tiwari, I.; Shrestha, J.; Modi, B.; Paudel, N.; Das, B.D. Role of soil microbes in sustainable crop production and soil health: A review. Agricul. Sci. Technol. 2021, 13, 1313–1336. [Google Scholar] [CrossRef]
- Pooja, S.; Rajesh, K. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar]
- Chandran, H.; Meena, M.; Swapnil, P. Plant growth-promoting rhizobacteria as a green alternative for sustainable agriculture. Sustainability 2021, 13, 10986. [Google Scholar] [CrossRef]
- Muhammad, I.R.; Liyakat, H.M.; Tanvir, S.; Talal, A.; Iqbal, M.I.; Ismail, M.O. Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils. Microbiol. Res. 2016, 183, 26–41. [Google Scholar]
- Singh, D.P.; Prabha, R.; Renu, S.; Sahu, P.K.; Singh, V. Agrowaste bioconversion and microbial fortification have prospects for soil health, crop productivity, and eco-enterprising. Int. J. Recycling Organic Waste Agricul. 2019, 8, 457–472. [Google Scholar] [CrossRef]
- Shen, H.; He, X.; Liu, Y.; Chen, Y.; Tang, J.; Guo, T. A Complex Inoculant of N2-Fixing, P- and K-Solubilizing Bacteria from a Purple Soil Improves the Growth of Kiwifruit (Actinidia chinensis) Plantlets. Front. Microbiol. 2016, 7, 841. [Google Scholar] [CrossRef]
- Meena, V.S.; Meena, S.K.; Verma, J.P.; Kumar, A.; Aeron, A.; Mishra, P.K.; Bisht, J.K.; Pattanayak, A.; Naveed, M.; Dotaniya, M.L. Plant beneficial rhizospheric microorganism (PBRM) strategies to improve nutrients use efficiency: a review. Ecol. Engineering 2017, 107, 8–32. [Google Scholar] [CrossRef]
- Gopika, K.; Jagadeeshwar, R.; Rao, V.K.; Vijayalakshmi, K. An overview of stem rot disease of rice (Sclerotium oryzae catt.) and its comprehensive management. Int. J. Appl. Biol. Pharml. Technol. 2016, 7, 111–124. [Google Scholar]
- Pramesh, D.; Alase, S.; Muniraju, K.M.; Kumara, M.K. A combination fungicide for the management of sheath blight, sheath rot and stem rot diseases of paddy. Int. J. Current Microbiol. Appl. Sci. 2017, 6, 3500–3509. [Google Scholar]
- Bashyal, B.M.; Parmar, P.; Zaidi, N.W.; Sunil, K.S.; Prakash, G.; Rashmi, A. Improved methodology for the isolation of false smut pathogen Ustilaginoidea virens of rice. Indian Phytopathol. 2021, 74, 249–252. [Google Scholar] [CrossRef]
- Sinclair, J.B.; Dhingra, O.D. Basic Plant Pathology Methods. CRC Press: Boca Raton, 2017; pp. 287–300. [Google Scholar]
- Gangwar, G.P.; Sinha, A.P. Comparative antagonistic potential of Trichoderma spp. against Xanthomonas oryzae pv. oryzae. Ann. Plant Sci. 2010, 8, 458–463. [Google Scholar]
- Kumar, V.; Singh, R.; Jalali, B.L. Evaluation of resistance to stem rot and yield losses caused by the disease in rice. Indian Phytopathol. 2003, 4, 403–407. [Google Scholar]
- IRRI. Standard Evaluation System for Rice, International Rice Research Institute, Los Banos, Manila, Philippines. 2002, p. 24. Available online: http://www.knowledgebank.irri.org/images/docs/rice-standard-evaluation-system.pdf.
- Brueske, C.H. Phenylalanine ammonia lyase activity in tomato roots infected and resistant to the root-knot nematode, Meloidogyne incognita. Physiol. Plant Pathol. 1980, 16, 409–414. [Google Scholar] [CrossRef]
- Hammerschmidt, R.; Nuckles, E.M.; Kuc, J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol. Plant Pathol. 1982, 20, 73–82. [Google Scholar] [CrossRef]
- Gauillard, F.; Richardforget, F.; Nicolas, J. New spectrophotometric assay for polyphenol oxidase activity. Anal. Biochem. 1993, 215, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Malick, C.P.; Singh, M.B. Plant Enzymology and Histo Enzymology; Kalyani Publishers: New Delhi, India, 1980; p. 289. [Google Scholar]
- McLean, E.O. Soil pH and lime requirement. In Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties; 1983; Volume 9, pp. 199–224. [Google Scholar]
- Rhoades, J.D. Soluble Salts. In Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties; Agronomy Monographs; American Society of Agronomy: Madison, WI, USA, 1983; pp. 167–179. [Google Scholar]
- Walkley, A.; Black, L.A. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Subbaiah, B.V. A rapid procedure for estimation of available nitrogen in soil. Curr. Sci. 1956, 25, 259–260. [Google Scholar]
- Watanabe, F.S.; Olsen, S.R. Test of an ascorbic acid method for determining phosphorus in water and NaHCO3 extracts from soil. Soil Sci. Soc. Am. J. 1965, 29, 677–678. [Google Scholar] [CrossRef]
- Muhr, G.R. Soil Testing in India; USAID Mission to India: New Delhi, India, 1965; p. 120. [Google Scholar]
- Casida, J.L.E.; Klein, D.A.; Santoro, T. Soil dehydrogenase activity. Soil Sci. 1964, 98, 371–376. [Google Scholar] [CrossRef]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil. Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- Eivazi, F.; Tabatabai, M.A. Phosphatases in soils. Soil Biol. Biochem. 1977, 9, 167–172. [Google Scholar] [CrossRef]
- Piper, C.S. Vanado-Molybdo-Phosphoric Yellow Colour Method. Soil Sci. Plant Anal. 1966, 368. [Google Scholar]
- Bhargava, B.S.; Raghupathi, H.B. Analysis of plant materials for macro and micronutrients. Methods of analysis of soils, plants, water and fertilizers 1993, 49–82. [Google Scholar]
- Husson, F.; Josse, J.; Le, S.; Mazet, J. FactoMineR: multivariate exploratory data analysis and data mining. 2020. Available online: https://CRAN.R-project.org/package=FactoMineR.
- Kassambara, A.; Mundt, F. Factoextra: Extract and visualize the results of multivariate data analyses, R package v. 1.0.7. 2020.
- Kannan, C.; Divya, M.; Rekha, G.; Maruthi, P.; Hajira, S.; Sundaram, R.M. Diversity analysis of antagonistic microbes against bacterial leaf and fungal sheath blight diseases of rice. Egypt. J. Biol. Pest Cont. 2021, 31, 115. [Google Scholar] [CrossRef]
- Nawaz, M.E.; Malik, K.; Hassan, M.N. Rice-associated antagonistic bacteria suppress the Fusarium fujikoroi causing rice bakanae disease. Biol. Cont. 2022, 67, 101–109. [Google Scholar] [CrossRef]
- Caulier, S.; Nannan, C.; Gillis, A.; Licciardi, F.; Bragard, C.; Mahillon, J. Overview of the Antimicrobial Compounds Produced by Members of the Bacillus subtilis group. Front. Microbiol. 2019, 10, 003. [Google Scholar] [CrossRef] [PubMed]
- Adrien, B.; Martin, F. Phenazines in plant-beneficial Pseudomonas spp. biosynthesis, regulation, function and genomics. Environ. Microbiol. 2018, 20, 3905–3917. [Google Scholar]
- Pandit, M.A.; Kumar, J.; Gulati, S.; Bhandari, N.; Mehta, P.; Katyal, R.; Rawat, C.D.; Mishra, V.; Kaur, J. Major Biological Control Strategies for Plant Pathogens. Pathogens 2022, 11, 273. [Google Scholar] [CrossRef] [PubMed]
- Moubark, M.Y.; Abdel-Monaim, M.F. Effect of bio-control agents on yield, yield components and root rot control in two wheat cultivars at New Valley region. Not. Sci. Biol. 2011, 3, 79–87. [Google Scholar] [CrossRef]
- Ivica, D.; Tamara, J.; Marija, P.; Giuliano, D.; Djordje, F. Plant-associated Bacillus and Pseudomonas antimicrobial activities in plant disease suppression via biological control mechanisms—A review. Physiol. Mol. Plant Pathol. 2022, 117, 101754. [Google Scholar]
- Stuti, S.; Shweena, K.; Rajni, S. Pseudomonas mediated nutritional and growth promotional activities for sustainable food security. Microbiol. Curr. Res. 2021, 2, 2666–5174. [Google Scholar]
- Tian, J.; Ge, F.; Zhang, D.; Deng, S.; Liu, X. Roles of Phosphate Solubilizing Microorganisms from Managing Soil Phosphorus Deficiency to Mediating Biogeochemical P Cycle. Biol. J. 2021, 10, 158. [Google Scholar] [CrossRef]
- Sowmya, V.; Raja Gopalan, N.S.; Maruthi, P.; Baskar, M.; Umadevi, G.; Ladha lakshmi, D.; Koteshwar, P.; Jesudasu, G.; Rathod, S.; Srinivas Prasad, M.; et al. Native bio-control agents from the rice fields of Telangana, India: Characterization and unveiling the potential against stem rot and false smut diseases of rice. World J Microbiol Biotechnol. 2023, in press.
- Mukherjee, M.; Mukherjee, P.K.; Horwitz, B.A.; Zachow, C.; Berg, G.; Zeilinger, S. Trichoderma-plant-pathogen interactions: advances in genetics of biological control. Indian J. Microbiol. 2012, 52, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.S.; Koushal, S.; Kumar, A.; Vimal, S.R.; Gupta, V.K. Book review: microbial inoculants in sustainable agricultural Productivity—Vol. II: functional application. Microbiology 2016, 7, 7. [Google Scholar] [CrossRef]
- Halifu, S.; Deng, X.; Song, X.; Song, R.; Liang, X. Inhibitory mechanism of Trichoderma virens ZT05 on Rhizoctonia solani. Plants 2020, 9, 912. [Google Scholar] [CrossRef] [PubMed]
- Rais, A.; Jabeen, Z.; Shair, F.; Hafeez, F.; Hassan, M. Bacillus spp. a bio-control agent enhances the activity of antioxidant defense enzymes in rice against Pyricularia oryzae. PLoS ONE 2017, 12, e0187412. [Google Scholar] [CrossRef]
- Wiesel, L.; Newton, A.C.; Elliott, I.; Booty, D.; Gilroy, E.M.; Birch, P.R.J.; Hein, I. Molecular effects of resistance elicitors from biological origin and their potential for crop protection. Plant Signal Behav. 2014, 5, 655. [Google Scholar] [CrossRef] [PubMed]
- Deice Raasch-Fernandes, L.; Bonaldo, S.M.; Jesus Rodrigues, D.; Magela Vieira-Junior, G.; Regina Freitas Schwan-Estrada, K.; Rocco da Silva, C.; Gabriela Araújo Verçosa, A.; Lopes de Oliveira, D.; Wender Debiasi, B. Induction of phytoalexins and proteins related to pathogenesis in plants treated with extracts of cutaneous secretions of southern Amazonian Bufonidae amphibians. PLoS ONE 2019, 14, e0211020. [Google Scholar] [CrossRef]
- Zandi, P.; Schnug, E. Reactive Oxygen Species, Antioxidant Responses and Implications from a Microbial Modulation Perspective. Biology 2022, 11, 155. [Google Scholar] [CrossRef]
- Kaur, S.; Samota, M.K.; Choudhary, M.; Abhay, K.P.; Anshu, S.; Julie, T. How do plants defend themselves against pathogens-Biochemical mechanisms and genetic interventions. Physiol. Mol. Biol. Plants 2022, 28, 485–504. [Google Scholar] [CrossRef]
- Aryal, S.; Baniya, M.K.; Danekhu, K.; Kunwar, P.; Gurung, R.; Koirala, N. Total Phenolic Content, Flavonoid Content and Antioxidant Potential of Wild Vegetables from Western Nepal. Plants 2019, 8, 96. [Google Scholar] [CrossRef]
- Jacoby, R.; Peukert, M.; Succurro, A.; Koprivova, A.; Kopriva, S. The Role of Soil Microorganisms in Plant Mineral Nutrition-Current Knowledge and Future Directions. Front. Plant Sci. 2017, 8, 1617. [Google Scholar] [CrossRef]
- Ahmad, M.I.; Biswas, A.K.; Sultana, M.S.; Sarker, B.C.; Mondal, C. Effect of Different Media and pH on Mycelial Growth of Sclerotium oryzae Causing Sheath Rot of Rice. South Asian J. Agric. 2019, 18, 67–72. [Google Scholar] [CrossRef]
- Du, J.X.; Li, Y.; Ur-Rehman, S.; Mukhtar, I.; Yin, Z.; Dong, H.; Wang, H.; Zhang, X.; Gao, Z.; Zhao, X.; Xin, X.; Ding, X. Synergistically promoting plant health by harnessing synthetic microbial communities and prebiotics. Iscience 2021, 24, 102918. [Google Scholar] [CrossRef] [PubMed]
- Hibbing, M.E.; Fuqua, C.; Parsek, M.R.; Peterson, S.B. Bacterial competition: surviving and thriving in the microbial jungle. Nat. Rev. Microbiol. 2010, 8, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Keim, R.; Webster, R.K. Fungistasis of sclerotia of Sclerotium oryzae. Phytopathology 1975, 65, 283–287. [Google Scholar] [CrossRef]
- Yadav, R.L.; Shukla, S.K.; Suman, A.; Singh, P.N. Trichoderma inoculation and trash management effects on soil microbial biomass, soil respiration, nutrient uptake and yield of ratoon sugarcane under subtropical conditions. Biol. Fertil. Soils 2009, 45, 461–468. [Google Scholar] [CrossRef]
- Kapri, A.; Tewari, L. Phosphate solubilization potential and phosphatase activity of rhizospheric Trichoderma spp. Braz. J. Microbiol. 2010, 41, 787–795. [Google Scholar] [CrossRef]
- Kucuk, C.; Kivanc, M.; Kinaci, E.; Kinaci, G. Determination of the growth and solubilization capabilities of Trichoderma harzianum T1. Biologia 2008, 63, 167–170. [Google Scholar] [CrossRef]
- Nannipieri, P.; Kandeler, E.; Ruggiero, P. Enzyme activities and microbiological and biochemical processes in soil. Enzymes in the Environment 2002, 1–33. [Google Scholar]
- Gil-Sotres, F.; Trasar-Cepeda, C.; Leirós, M.C.; Seoane, S. Different approaches to evaluating soil quality using biochemical properties. Soil Biol. Biochem. 2005, 37, 877–887. [Google Scholar] [CrossRef]
- Burns, R.G. Enzyme activity in soil: location and a possible role in microbial ecology. Soil Biol Biochem. 1982, 14, 423–427. [Google Scholar] [CrossRef]
- Ng, C.W.W.; Yan, W.H.; Tsim, K.W.K.; So, P.S.; Xia, Y.T.; To, C.T. Effects of Bacillus subtilis and Pseudomonas fluorescens as the soil amendment. Heliyon 2022, 8, e11674. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.P.; Liu, J.; Wang, Y.Q.; Sun, H.M.; He, W.X. Research on relationship between soil enzyme activities and soil fertility. Plant Nut. Fert. Sci. 2004, 10, 277–280. [Google Scholar]
- Zenichi, M.; Fumie, I.; Konomi, T.; Masao, T. The Effects of Phosphorus and Magnesium on Sclerotium formation in Rhizoctonia solani Kühn. Japanese J. Phytopathol. 1991, 57, 649–656. [Google Scholar]
Figure 1.
Inoculation process of S. oryzae and development of stem rot symptoms in glass house conditions (a) S. oryzae pure culture; (b) S. oryzae was multiplied in dried bits of rice stem; (c) S. oryzae infection on the stem of the rice plant; (d) Formation of sclerotial bodies at the base of the stem; (e) Severe infection of the rice plant upon S. oryzae infection; (f) Maintenance of favorable conditions for S. oryzae multiplication.
Figure 1.
Inoculation process of S. oryzae and development of stem rot symptoms in glass house conditions (a) S. oryzae pure culture; (b) S. oryzae was multiplied in dried bits of rice stem; (c) S. oryzae infection on the stem of the rice plant; (d) Formation of sclerotial bodies at the base of the stem; (e) Severe infection of the rice plant upon S. oryzae infection; (f) Maintenance of favorable conditions for S. oryzae multiplication.

Figure 2.
Antagonistic effect of different bioagents on S. oryzae under in vitro conditionsa (a) Control stem rot pathogen; (b) Interaction of TAIK 1 + S. oryzae; (c) Interaction of BIK 3 + S. oryzae; (d) Interaction of PIK1 + S. oryzae; (e) Interaction of POPS 1 with S. oryzae.
Figure 2.
Antagonistic effect of different bioagents on S. oryzae under in vitro conditionsa (a) Control stem rot pathogen; (b) Interaction of TAIK 1 + S. oryzae; (c) Interaction of BIK 3 + S. oryzae; (d) Interaction of PIK1 + S. oryzae; (e) Interaction of POPS 1 with S. oryzae.
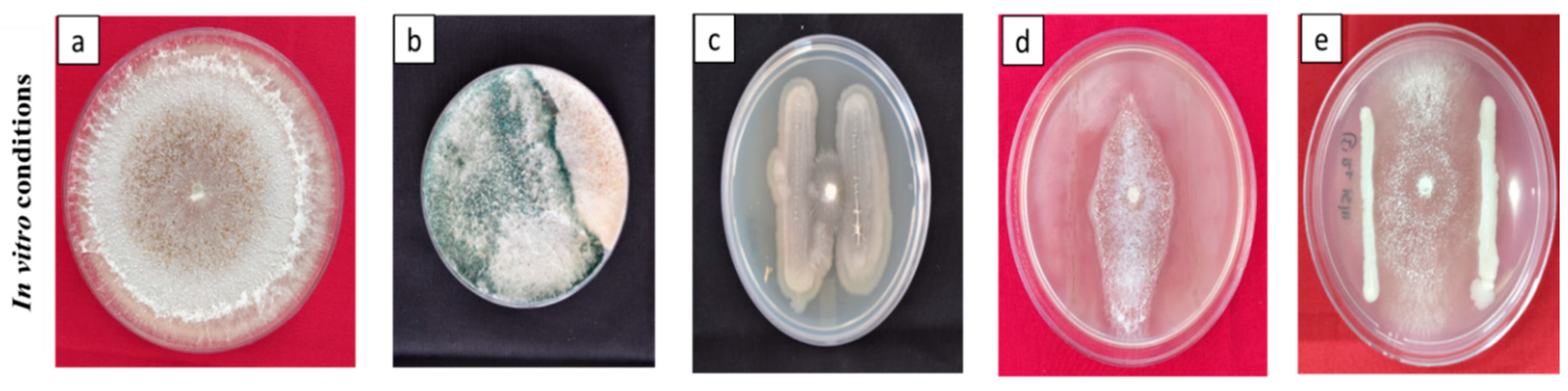
Figure 3.
Antagonistic effect of single and combined application of bioagents on stem rot disease of rice caused by S. oryzae under in vivo conditions (a) Control plant upon challenge inoculation with S. oryzae; (b) TAIK 1 treated plant upon challenge inoculation with S. oryzae; (c) BIK 3 treated plant upon challenge inoculation with S. oryzae; (d) PIK 1 treated plant upon challenge inoculation with S. oryzae; (e) POPS 1 treated plant upon challenge inoculation with S. oryzae; (f) Consortia treated plant upon challenge inoculation with S. oryzae.
Figure 3.
Antagonistic effect of single and combined application of bioagents on stem rot disease of rice caused by S. oryzae under in vivo conditions (a) Control plant upon challenge inoculation with S. oryzae; (b) TAIK 1 treated plant upon challenge inoculation with S. oryzae; (c) BIK 3 treated plant upon challenge inoculation with S. oryzae; (d) PIK 1 treated plant upon challenge inoculation with S. oryzae; (e) POPS 1 treated plant upon challenge inoculation with S. oryzae; (f) Consortia treated plant upon challenge inoculation with S. oryzae.

Figure 4.
Effect of individual and consortia of four bioagents on plant growth-promotion activities under glass house conditions.
Figure 4.
Effect of individual and consortia of four bioagents on plant growth-promotion activities under glass house conditions.

Figure 5.
Box plot illustrates the changes in defense enzymes upon challenge inoculation with S. oryzae. Each box plot represents the mean ±SE of 8 treatments at different intervals upon inoculation of stem rot disease, with a significant difference of (P < 0.05, DMRT, OPSTAT) between treated and control plants.
Figure 5.
Box plot illustrates the changes in defense enzymes upon challenge inoculation with S. oryzae. Each box plot represents the mean ±SE of 8 treatments at different intervals upon inoculation of stem rot disease, with a significant difference of (P < 0.05, DMRT, OPSTAT) between treated and control plants.
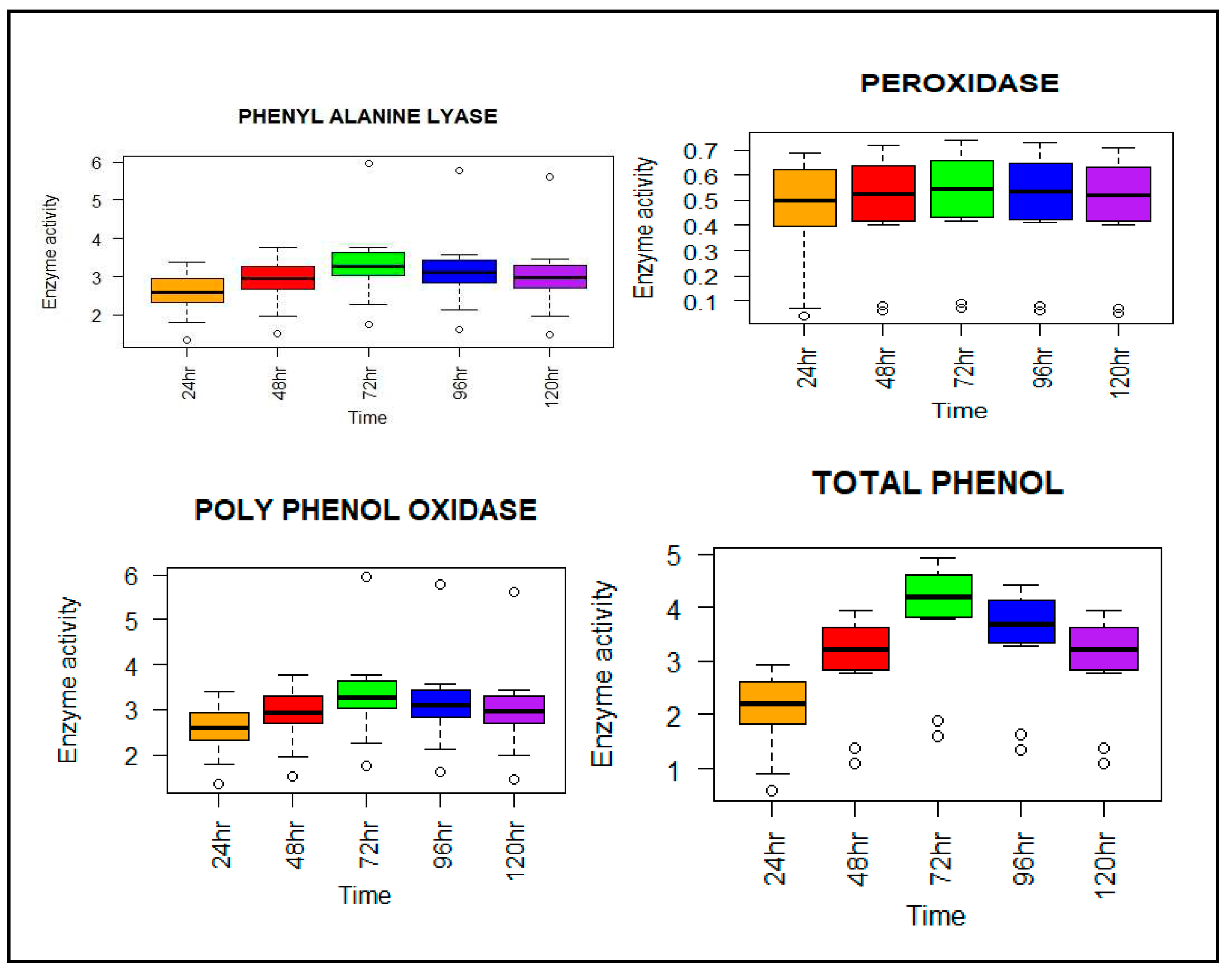
Figure 7.
Graphical interpretation of the relationship between changes in soil properties, PDI, and plant growth parameters of plants treated with bioagents.
Figure 7.
Graphical interpretation of the relationship between changes in soil properties, PDI, and plant growth parameters of plants treated with bioagents.
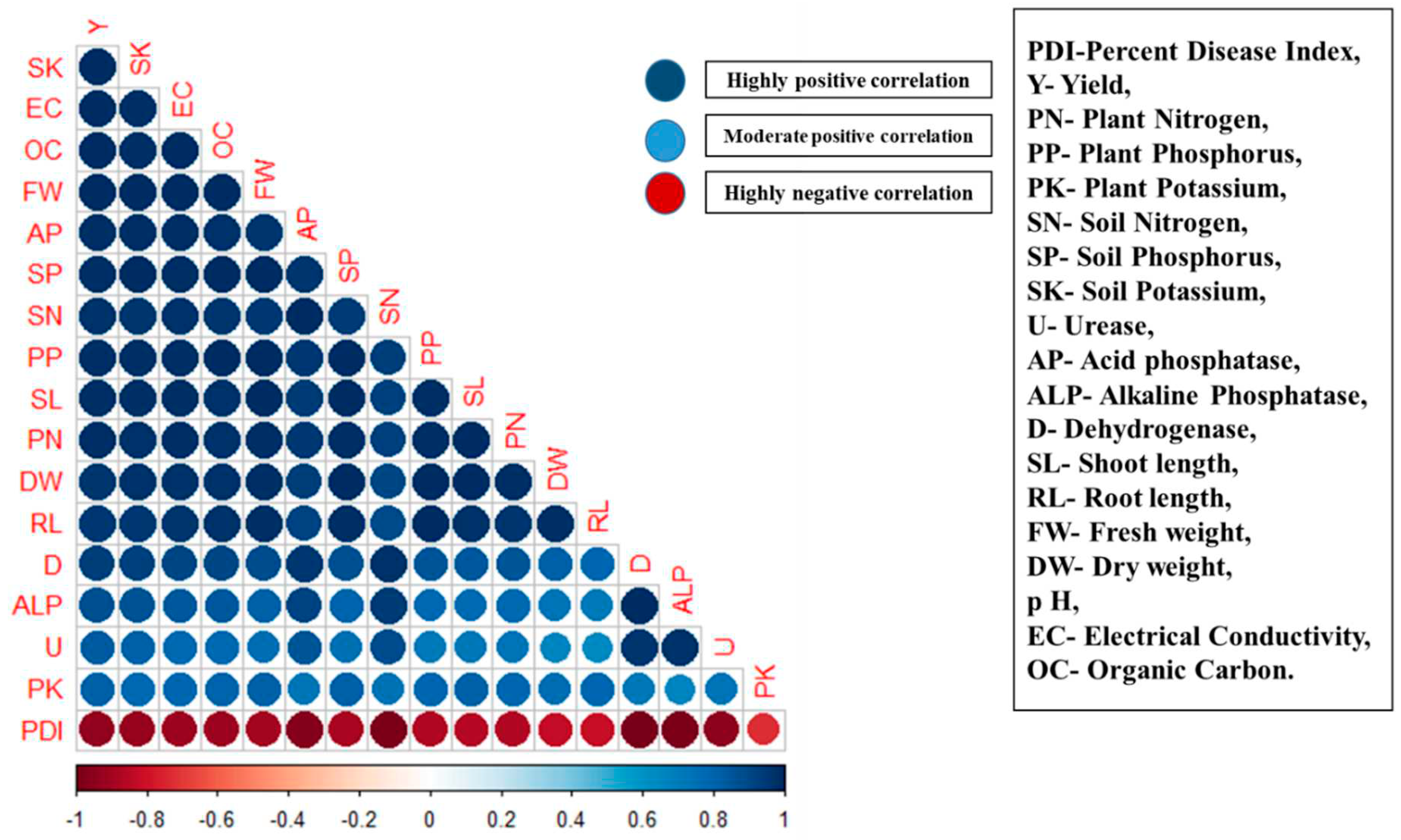
Figure 8.
Principal component analysis (PCA) on the effect of single and consortia of four bioagents on soil and plant growth parameters with percent disease (a) biplot (b) scree plot. PDI-Percent Disease Index, PN- Plant Nitrogen, PP- Plant Phosphorus, PK- Plant Potassium, SN- Soil Nitrogen, SP- Soil Phosphorus, SK- Soil Potassium, U- Urease, AP- Acid phosphatase, ALP- Alkaline Phosphatase, D- Dehydrogenase, SL- Shoot length, RL- Root length, FW- Fresh weight, DW- Dry weight, pH, EC- Electrical Conductivity, OC- Organic Carbon, KL- Kernel length, KB- Kernel breadth, NT- no. of tillers, NP- no. of panicles, PL- Panicle length, GY- Grain yield, and TW- Test weight.
Figure 8.
Principal component analysis (PCA) on the effect of single and consortia of four bioagents on soil and plant growth parameters with percent disease (a) biplot (b) scree plot. PDI-Percent Disease Index, PN- Plant Nitrogen, PP- Plant Phosphorus, PK- Plant Potassium, SN- Soil Nitrogen, SP- Soil Phosphorus, SK- Soil Potassium, U- Urease, AP- Acid phosphatase, ALP- Alkaline Phosphatase, D- Dehydrogenase, SL- Shoot length, RL- Root length, FW- Fresh weight, DW- Dry weight, pH, EC- Electrical Conductivity, OC- Organic Carbon, KL- Kernel length, KB- Kernel breadth, NT- no. of tillers, NP- no. of panicles, PL- Panicle length, GY- Grain yield, and TW- Test weight.
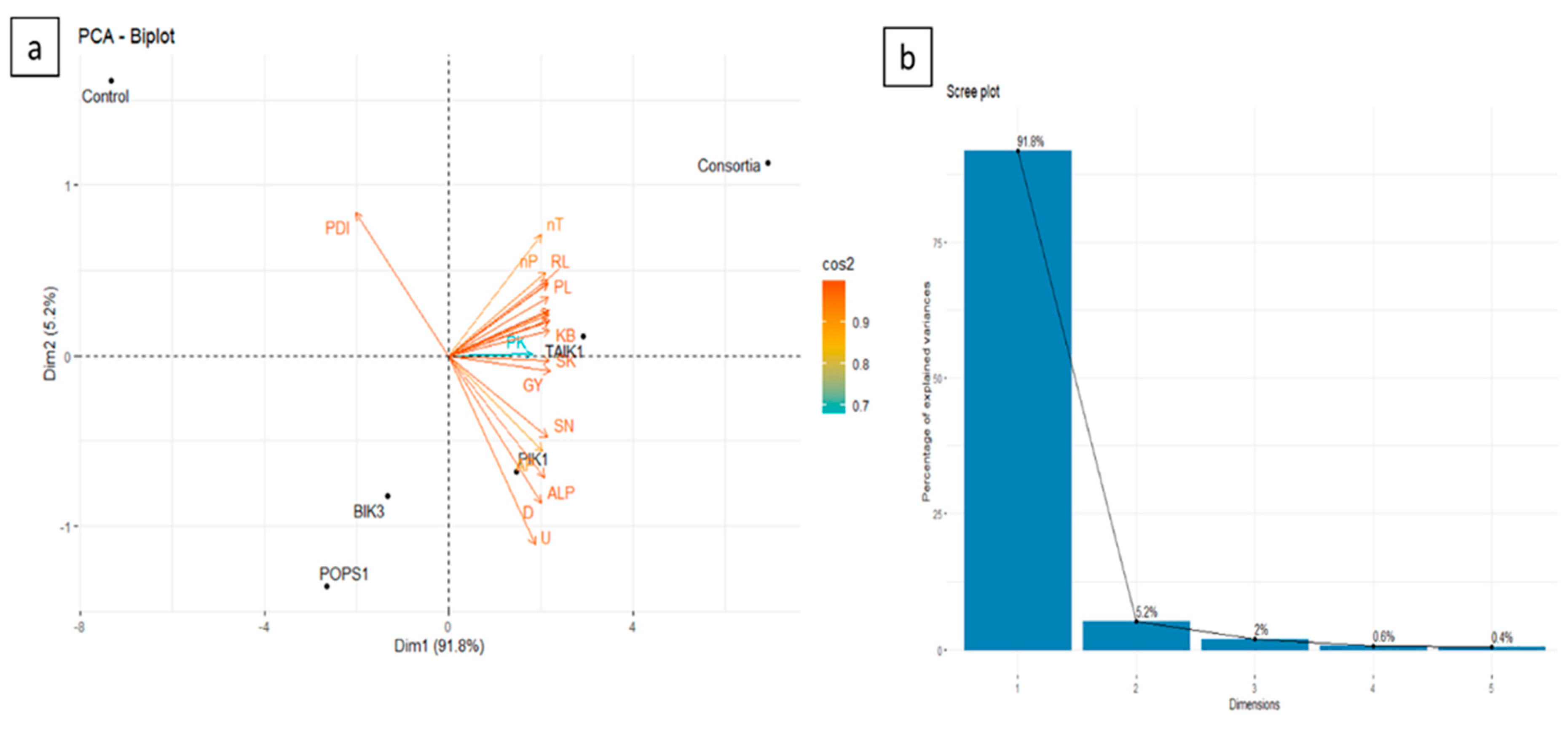
Table 3.
Stepwise regression analysis for shoot length with soil parameters upon bioagents treatment.
Table 3.
Stepwise regression analysis for shoot length with soil parameters upon bioagents treatment.
| Soil and plant characteristics | Parameters | Estimates | Standard Error | Probability | Partial R- Square | Model R- Square |
|---|---|---|---|---|---|---|
| PH | POPS1 | 0.97 | 0.05 | 0.0624 | 0.04 | 0.95 |
| BIK 3 | 0.62 | 0.06 | <.0001 | 0.91 | ||
| EC | BIK 3 | 0.65 | 0.08 | <.0001 | 0.96 | 0.9552 |
| OC | PIK 1 | 0.68 | 0.06 | <.0001 | 0.94 | 0.9414 |
| Available N | TAIK 1 | 0.39 | 0.03 | <.0001 | 0.94 | 0.9403 |
| Available P | PIK 1 | 0.69 | 0.05 | <.0001 | 0.95 | 0.9527 |
| Available K | PIK 1 | 0.69 | 0.05 | 0.0003 | 0.33 | 0.9462 |
| Consortia | 0.89 | 0.1 | <.0001 | 0.61 | ||
| Urease | Consortia | 0.9 | 0.03 | <.0001 | 0.96 | 0.959 |
| Acid Phosphatase | POPS1 | 0.63 | 0.01 | <.0001 | 0.98 | 0.9783 |
| Alkaline Phosphatase | POPS1 | 0.4 | 0.05 | 0.0037 | 0.05 | 0.9702 |
| BIK 3 | 0.14 | 0.02 | 0.0025 | 0.84 | ||
| TAIK 1 | 0.55 | 0.07 | 0.0073 | 0.08 | ||
| Dehydrogenase | POPS1 | 0.42 | 0.08 | <.0001 | 0.96 | 0.9633 |
| Plant N | BIK3 | 0.44 | 0.03 | 0.0098 | 0.59 | 0.76 |
| Consortia | 0.57 | 0.07 | 0.055 | 0.18 | ||
| Plant P | POPS1 | 0.99 | 0.05 | 0.0287 | 0.02 | 0.9785 |
| BIK3 | 0.49 | 0.09 | <.0001 | 0.96 | ||
| Plant K | PIK1 | 0.7 | 0.09 | 0.0994 | 0.03 | 0.9836 |
| POPS1 | 0.51 | 0.02 | 0.0548 | 0.03 | ||
| BIK3 | 0.48 | 0 | 0.0992 | 0.01 | ||
| Consortia | 0.35 | 0.05 | <.0001 | 0.91 |
Table 4.
Stepwise regression analysis for root length with soil parameters upon bioagents treatment.
Table 4.
Stepwise regression analysis for root length with soil parameters upon bioagents treatment.
| Soil and plant characteristics | Parameters | Estimates | Standard Error | Probability | Partial R- Square | Model R- Square |
|---|---|---|---|---|---|---|
| PH | POPS1 | 0.67 | 0.08 | <.0001 | 0.92 | 0.96 |
| BIK 3 | 0.5 | 0.03 | 0.0367 | 0.04 | ||
| EC | BIK 3 | 0.69 | 0.09 | <.0001 | 0.96 | 0.9571 |
| OC | PIK 1 | 0.77 | 0.01 | <.0001 | 0.96 | 0.9608 |
| Available N | TAIK 1 | 0.26 | 0.02 | <.0001 | 0.96 | 0.9643 |
| Available P | PIK 1 | 0.49 | 0.03 | <.0001 | 0.97 | 0.973 |
| Available K | PIK 1 | 0.25 | 0.06 | 0.0011 | 0.58 | 0.9169 |
| Consortia | 0.97 | 0.05 | 0.0104 | 0.34 | ||
| Urease | TAIK 1 | 0.65 | 0.1 | <.0001 | 0.97 | 0.9685 |
| Acid Phosphatase | POPS1 | 0.12 | 0.06 | <.0001 | 0.98 | 0.9785 |
| Alkaline Phosphatase | POPS1 | 0.96 | 0.02 | 0.1215 | 0.04 | 0.9753 |
| BIK3 | 0.57 | 0.03 | 0.0001 | 0.86 | ||
| TAIK1 | 0.43 | 0.03 | 0.0059 | 0.07 | ||
| Dehydrogenase | POPS1 | 0.68 | 0.03 | <.0001 | 0.97 | 0.9726 |
| Plant N | BIK3 | 0.43 | 0.02 | 0.0191 | 0.52 | 0.7677 |
| Consortia | 0.59 | 0.06 | 0.0285 | 0.25 | ||
| Plant P | PIK 1 | 0.57 | 0.04 | <.0001 | 0.94 | 0.9412 |
| Plant K | PIK1 | 0.55 | 0.05 | 0.0527 | 0.03 | 0.9757 |
| TAIK1 | 0.7 | 0.05 | 0.0752 | 0.02 | ||
| Consortia | 0.49 | 0.06 | <.0001 | 0.92 |
Table 5.
Stepwise regression analysis for fresh weight with soil parameters upon bioagents treatment.
Table 5.
Stepwise regression analysis for fresh weight with soil parameters upon bioagents treatment.
| Soil and plant characteristics | Parameters | Estimates | Standard Error | Probability | Partial R- Square | Model R- Square |
|---|---|---|---|---|---|---|
| PH | POPS1 | 0.88 | 0.05 | <.0001 | 0.9 | 0.9383 |
| BIK 3 | 0.67 | 0.05 | 0.0748 | 0.04 | ||
| EC | BIK 3 | 0.82 | 0.03 | <.0001 | 0.94 | 0.9403 |
| OC | PIK 1 | 0.65 | 0.07 | <.0001 | 0.93 | 0.9332 |
| Available N | TAIK 1 | 0.29 | 0.03 | <.0001 | 0.94 | 0.935 |
| Available P | POPS1 | 0.83 | 0.07 | <.0001 | 0.94 | 0.9432 |
| Available K | BIK 3 | 0.08 | 0.03 | 0.1465 | 0.32 | 0.9712 |
| consortia | 0.58 | 0.02 | <.0001 | 0.65 | ||
| Urease | TAIK 1 | 0.28 | 0.05 | <.0001 | 0.94 | 0.9367 |
| Acid Phosphatase | POPS1 | 0.66 | 0.01 | <.0001 | 0.96 | 0.9731 |
| consortia | 0.36 | 0.02 | 0.0929 | 0.01 | ||
| Alkaline Phosphatase | POPS1 | 0.12 | 0.04 | 0.1 | 0.07 | 0.9657 |
| BIK3 | 0.67 | 0.06 | 0.0005 | 0.8 | ||
| TAIK1 | 0.07 | 0.01 | 0.0057 | 0.1 | ||
| Dehydrogenase | PIK 1 | 0.61 | 0.07 | <.0001 | 0.96 | 0.9641 |
| Plant N | BIK3 | 0.76 | 0.03 | 0.0206 | 0.51 | 0.738 |
| Consortia | 0.8 | 0.06 | 0.0424 | 0.23 | ||
| Plant P | BIK3 | 0.25 | 0.03 | <.0001 | 0.93 | 0.9349 |
| Plant K | PIK1 | 0.45 | 0.05 | 0.0438 | 0.06 | 0.9822 |
| POPS1 | 0.72 | 0.07 | 0.0343 | 0.04 | ||
| BIK3 | 0.67 | 0.05 | 0.1244 | 0.01 | ||
| Consortia | 0.72 | 0.1 | <.0001 | 0.88 |
Table 6.
Stepwise regression analysis for dry weight with soil parameters upon bioagents treatment.
| Soil and plant characteristics | Parameters | Estimates | Standard Error | Probability | Partial R- Square | Model R- Square |
|---|---|---|---|---|---|---|
| PH | POPS1 | 0.07 | 0.09 | 0.0877 | 0.9 | 0.93 |
| BIK 3 | 0.05 | 0.07 | <.0001 | 0.04 | ||
| EC | BIK 3 | 0.06 | 0.06 | <.0001 | 0.95 | 0.9624 |
| Consortia | 0.06 | 0.02 | 0.1458 | 0.01 | ||
| OC | PIK 1 | 0.67 | 0.07 | <.0001 | 0.94 | 0.9381 |
| Available N | TAIK 1 | 0.10 | 0.01 | <.0001 | 0.94 | 0.9432 |
| Available P | PIK 1 | 0.07 | 0.01 | <.0001 | 0.95 | 0.9483 |
| Available K | BIK 3 | 0.07 | 0.04 | 0.0965 | 0.58 | 0.9398 |
| Consortia | 0.18 | 0.03 | 0.0113 | 0.36 | ||
| Urease | Consortia | 0.14 | 0.07 | <.0001 | 0.95 | 0.9494 |
| Acid Phosphatase | POPS1 | 0.13 | 0.09 | <.0001 | 0.97 | 0.9827 |
| Consortia | 0.81 | 0.05 | 0.0726 | 0.01 | ||
| Alkaline Phosphatase | POPS1 | 0.68 | 0.01 | 0.0983 | 0.06 | 0.9716 |
| BIK 3 | 0.45 | 0.08 | 0.0003 | 0.82 | ||
| TAIK 1 | 0.52 | 0.07 | 0.0044 | 0.09 | ||
| Dehydrogenase | PIK 1 | 0.25 | 0.02 | <.0001 | 0.97 | 0.9699 |
| Plant N | PIK1 | 0.40 | 0.06 | 0.0472 | 0.21 | 0.7497 |
| BIK3 | 0.70 | 0.10 | 0.015 | 0.54 | ||
| Plant P | POPS1 | 0.51 | 0.03 | 0.0928 | 0.02 | 0.9673 |
| BIK3 | 0.59 | 0.01 | <.0001 | 0.95 | ||
| Plant K | PIK1 | 0.65 | 0.07 | 0.0944 | 0.04 | 0.9807 |
| POPS1 | 0.86 | 0.01 | 0.0702 | 0.03 | ||
| BIK3 | 0.73 | 0.05 | 0.0665 | 0.02 | ||
| Consortia | 0.96 | 0.01 | <.0001 | 0.89 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



