
Submitted:
27 June 2023
Posted:
28 June 2023
You are already at the latest version
Abstract
Typhulaceae Jülich is one of the cold-adapted fungal families in basidiomycetes. Typhula (Pers.) Fr. and Pistillaria Fr. representing the family are distinguished from the morphology of a stipe to a hymenium (Fries 1821). The hymenium of Typhula is distinguishable from the stipe, but indistinguishable and consecutive in the genus Pistillaria. This taxonomic criterion is ambiguous, and consequently the opinion of Karsten (1882) has been widely accepted; I the genus Typhula, basidiomata develop from sclerotia, while basidiomata develop directly from substrata in the genus Pistillaris. But, Corner (1970) observed basidiomata of Pistillaria petasitis S. Imai in Hokkaido, Japan, from sclerotia. We collected irregular sclerotia of Typhula hyperborea H. Ekstr. from Upernavik, West Greenland. This specimen had a stipe-like structure on a Poaceae plant, and sclerotia developed on its tip. In August, P. petasitis in Hokkaido, Japan, formed basidiomata directly from substrates on the ground. Similar phenomena were found from other Typhula spp. in Japan. All of these fungi had the potential to produce sclerotia in culture, and as well as the capacity of mycelial growth at ambient air temperatures in each locality when samples were collected. These findings suggest that Typhula spp. developed basidiomata not only from the sclerotia dispersed by the basidiospores but also mycelia generated by the spore germination formed basidiomata multiple times depending on their growth environments.
Keywords:
cryophilic
; ecophysiology
; growth temperature
; life history
; local climate
1. Introduction
Approximately 80% of the biosphere are constantly and seasonally cold and have a temperature below 5°C, and polar regions occupies 14% of the biosphere (e.g. [1]). The biosphere, with an exception of deep sea and stratosphere, is almost identical to the cryosphere. The term cryosphere was proposed by the Polish geophysicist and meteorologist, A.B. Dobrowolski [2,3] and collectively describes the portions of the Earth’s surface where water exists as the frozen state – snow cover, glaciers, ice sheets and shelves, freshwater ice, sea ice, icebergs, permafrost, and ground ice [4].
Several kinds of microorganisms especially fungi were reported from the cryosphere [5,6,7,8,9]. Fungi are eukaryotic organisms without plastids, nutrition absorptive, unicellular or filamentous and consisting of multicellular coenocytic hyphae [10]. The true fungi form kingdom Fungi and the organisms studied by mycologist, fungi are mostly placed here, but others belong to the kingdoms Protozoa and Chromista [10,11]. Fungal species were less frequently recorded from the cryosphere than those of temperate zone despite that all fungal taxa have already been found in the cryosphere. These records suggest a possibility that various fungi are active under various cold environments.
We proposed the term ‘cryophilic fungi’ to denote fungi adapted to the cryosphere [12]. The concept of cryophilic fungi is defined as fungi that are present in the cryosphere, spend a certain life stage or whole life cycle (sexual and/or asexual reproductive stages), and grow under subzero temperature where water remains in the solid state such as snow and ice. The concept of cryophilic fungi also applies to uncultured fungi such as mycorrgizal fungi.
Their life cycles are affected by environmental factors [13]. This group of fungi includes psychrophile and psychrotolerant. The psychrophily is defined by physiological characteristics at each stage of their lifecycle [13]. Snow molds, representing the cryophilic fungi, develop mycelia to attack dormant plants such as forage crops, winter cereals, and conifer seedlings under snow cover [9]. Some of them pass the dormancy from spring to autumn in the form of sclerotia in temperate and frigid zones, as well as the Arctic [14,15,16,17,18,19,20,21,22,23,24,25,26,27] and Antarctica [8,29,30], e.g., Typhula incarnata Lasch, Typhula ishikariensis complex (consisting to T. ishikariensis S. Imai, T. canadensis (J.D. Sm. & Årsvoll) Tam. Hoshino, T. Kasuya & N. Matsumoto and T. hyperborea H. Ekstr.), Sclerotinia boreslis Bubák & Vleugel., and Sclerotinia antarctica.
In 2007, the first author collected aberrant sclerotia of Typhula sp. (Figure 1D–F) from Upernavik (72.7869, -56.1472), West Greenland. They resembled those of T. hyperborea (Figure 1C), but were often formed on the tip of a stem-like structure on the Poaceae host. The stem-like structure was indistinguishable from the stems of Typhula sp. (Figure 1D–F) and seemed identical to secondary sclerotia of hybrids of T. ishikariensis var. ishikariensis S. Imai × T. ishikariensis var. idahoensis (Remsberg) Årsvoll & J.D. Sm. produced under artificial conditions [31,32].
Pistillaria Fr. is close to the genus of Typhula Fr. in Typhulaceae Jülich [33] and saprophytic. The hymenium of Typhula is distinguishable from the stem, but indistinguishable to the stem in the genus Pistillaria [34]. However, this criterion proved unclear to separate both genera. Consequently, Karsten’s view has been widely accepted; Typhula spp. develop basidiomata from sclerotia, and Pistillaris spp., lacking sclerotia, develop basidiomata directly from substrata [35]. But, Corner [36] observed basidiomata of Pistillaria petasitis S. Imai developed from sclerotia in Hokkaido, Japan.
In this study, we aimed to elucidate the plasticity in the genera Typhula and Pistillaria though the interactions between their ecophysiological potential and environmental conditions in their localities. The results here revealed that repeated basidioma production was dependent on both ambient temperatures and the length of time without snow cover after basidioma formation.
2. Materials and Methods
2.1. Fungal materials
Fungal sclerotia were collected from decayed leaves and stems of decayed plants during the surveys. Sclerotium samples were packed in paper envelopes and dried at room temperature during transportation. Sclerotia were surface-sterilized in 70% (v/v) ethanol for 10 sec, in 0.5% (as active chlorine) sodium hypochlorite solution for 30 sec, and thoroughly rinsed in sterilized distilled water. They were then cut with sterilized steel blades, placed on potato dextrose agar (PDA: Difco, Sparks, MD, USA) and incubated at 4°C. Mycelia from growing colony margins were transferred to PDA slants and maintained at 0°C.
Collected basidiomata were put in plastic cases with wet cotton balls and kept in a refrigerator. Basidiomata were attached to the inside of Petri dish lids with double-sided adhesive tape, and spores were collected on PDA plates to incubate at 4°C for 1 or 2 days. Basidiospores subsequently germinated and mated to produce heterokaryons and sclerotia. These sclerotia were transferred to fresh PDA plates and incubated at 4°C for 2 weeks. Isolates were maintained on PDA slants at 0°C.
These specimens were kept in the mycological herbaria of National Museum of Nature and Science, Tokyo (TNS).
2.2. Mating experiments
Monokaryons of T. ishikariensis var. ishikariensis (strains PR7-6-7 and PR9-4-3 from Japan) and T. canadensis (strains 35-8 and 8-2 from Japan) were designated as testers and paired with dikaryons of collected strains (di-mon mating; [37]) on PDA plates and incubated at 4°C for one month. A small agar block was cut from monokaryon colonies near the colony junction and transplanted to another PDA plate. Growth from the block was then examined for the presence of clamp connections on hyphae after incubation for 5 to 7 days at 4°C. The presence of clamp connections on hyphae was the criterion of mating compatibility
2.3. Phylogenic analyses
Fungal strains were cultured for 1 month at 10˚C on PDA or Wort agar (HiMedia Laboratories, Pvt. Ltd., Mumbai, India). Sclerotia of T. hyperborea from Upernavik, West Greenland and S. nivale were harvested and DNA was extracted by the protocol of DNeasy Plant MiniPrep (QIAGEN GmbH, Germany). ITS regions including the 5.8S gene of genomic rDNA were amplified using primer pair ITS1 (5'TCCGTAGGTGAACCTGCGG) and ITS4 (5'-TCCTCCGCTTATTGATATGC), as described by White et al. [38]. PCR products were purified using a QIAquick PCR Purification Kit (QIAGEN GmbH, Germany) and sequenced in one direction on ABI PRISM 3100 Genetic Analyzer (Applied Biosystems, USA) using the ITS1 primer. Multiple alignment of the ITS sequences was performed, and the nucleotide substitution rate (Knuc value) was calculated in CLUSTAL W [39]. A phylogenetic tree was constructed by neighbor-joining method [40,41] using the program CLUSTAL W.
2.4. Morphological observations
Colors of basidiocarps and sclerotia were described according to the color identification chart of the Royal Botanic Garden Edinburgh (Flora of British Fungi) [42].
For light microscope section, aberrant sclerotia of Typhula sp. were fixed with 2% glutaraldehyde (Nisshin EM, Co., Ltd., Tokyo, Japan) in 50 mM phosphate buffer, washed the same buffer. The samples then post-fixed with 1% osmium tetroxide (Nisshin EM, Co., Ltd., Tokyo, Japan), dehydrated with an ethanol series, and embedded in Quetol 651 (Nisshin EM, Co., Ltd., Tokyo, Japan). The sections (0.8 μm thick) were stained with toluidine blue (Wako Ltd., Osaka, Japan) and observed under a light microscope.
2.5. Mycelial growth temperature
Five mm diam discs with mycelia were cut from the margins of actively growing colonies, inoculated to the centers of PDA plates, and incubated at six different temperatures from 0° to 25°C in duplicate. After 1, 2, and 3 weeks of inoculation, colony diameters were determined. Linear mycelial growth rate per week was calculated after the initial lag period.
3. Results
3.1. Typhula hyperborea in Greenland
Fungal cultures from aberrant sclerotia from Upernavik West Greenland had feather-like mycelia when grown on PDA at 4°C (Figure 2A), which were typical physiological reaction of T. hyperborea [28], and our strain did not react when paired with both tester monokaryons of T. ishikariensis var. ishikariensis and T. canadensis. Phylogenic analysis of ITS regions also supported this assumption (data not shown).
When the cultures were kept at 4°C for 2 years, sclerotia were found to have stme-like structures with intercalary, small secondary sclerotia as described by Christen [31,32] (Figure 2B). Cultures of T. ishikariensis var. ishikariensis from Hokkaido, Japan also produced similar aberrant sclerotia. Stem-like structures and aberrant sclerotia of T. hyperborea and T. ishikariensis var. ishikariensis on PDA were dark brown (19 bay) as same color with field samples (Figure 1D,E). However, normal stems of both fungi were white (2 B) to pale yellow (6 E) or pale brown (30 clay pink) [28]. Collected stem-like structures of T. hyperborea from Upernavik West Greenland were gradually connected with aberrant sclerotia (Figure 1F), and similar findings were reported the base of the stem of Typhula sclerotioides (Pers.) Fr. at its origin from the sclerotium [43]. These results suggested that the germination of sclerotia and the secondary sclerotium formation were reversible for T. hyperborea and T. ishikariensis var. ishikariensis.
The strain OUP1811 from Nuuk (64.1666, -57.7500), also in West Greenland was different in culture morphology on PDA plates (Figure 2C). This strain had weak pathogenicity against host [28], and they showed normal growth at 4–10°C and had abundant aerial mycelia without sclerotia on PDA. Sclerotium formation was, however observed on oatmeal agar plates (Figure 2D).
3.2. Typhula sp. and Pistillaria petasitis in Hokkaido, Japan
Basidiomata of Typhula sp. whose hymenium was distinguishable from the stem were found on the dead petioles of Kalopananax septemlobus in the Ashoro Research Forest, Kyushu University, Hokkaido, Japan (43.2507, 143.5498, altitude 114–471m) in October 2010 (Figure 3A,B). Basidioma: ca. 1.5–3.0 cm high, white (2 B). Head: ca. 2.5–8.5 × 0.5– 1.8 mm, clavate to cylindric, obtuse, straight or curved. Stem: ca. 3.5–21.5 × 0.5–1.0 mm, opaque, white (2 B). Although the specimens lacked, resultant cultures produced sclerotia on PDA (Figure 3C). Sclerotium: ca. 1–2.5 mm diam, globose to subglobose, almost black (36 fuscous black).
Basidiomata of P. petasitis without sclerotia were observed on the hillside of Mt. Asahidake (43.6511, 142.7990, altitude 1,100m), Higashikawa, Hokkaido (Figure 3A and Figure 4A,B), in August 2010. Basidioma: ca. 1–3.8 cm high, white (2 B). Head: ca. 2.5–26.5 × 1.5–6.5 mm, clavate to cylindric, obtuse, straight or curved. Stem: ca. 5–12.5 × 0.8–3.5 mm, opaque, white (2 B). The fungus also observed in the forest neighboring agricultural field in Hokkaido Agricultural Research Center, NARO, Sapporo, Hokkaido, Japan in September 2010.
Pistillaria petasitis basidiomata were found on various substrates such as Petasites japonicus, Conioselinum filicinum, and Cirsium kamtschaticum. Basidiomata of P. petasitis were found on substrates on the ground surface to 10cm on 3rd August 2010 (Figure 4D). Up to 80cm on 23rd August 2010 (Figure 4E). All P. petasitis isolates produced sclerotia under culture conditions (Figure 4C). Sclerotia of our isolates were similar as described Corner [36], and ca. 2.5–22.5 × 2.5–5.5 mm, fusiform, somewhat flattened, light brown (15 brick).
3.3. Effect of temperature on mycelial growth
Optimal mycelial growth temperatures of T. hyperborea from Upernavik, West Greenland, Typhula sp. from Ashoro, and P. petasitis from Higashikawa occurred at 5, 15, and 20°C, respectively (Figure 5). Maximal growth temperatures were 15, 25 and 30°C, respectively. The mycelial growth range of T. hyperborea Upernavik, West Greenland, was psyhrophilic and typical in this fungus [28]. According to ranges of their mycelial growth temperature, Typhula sp. from Ashoro and P. petasitis from Higashikawa were psychrotolerant. This is the first record of the mycelial growth range of P. petasitis.
These growth temperature relations agreed with the ambient temperatures when basidiomata samples were collected in each locality. These results suggested that the mycelia of these three fungi could survive and be active without snow in their localities.
4. Discussion
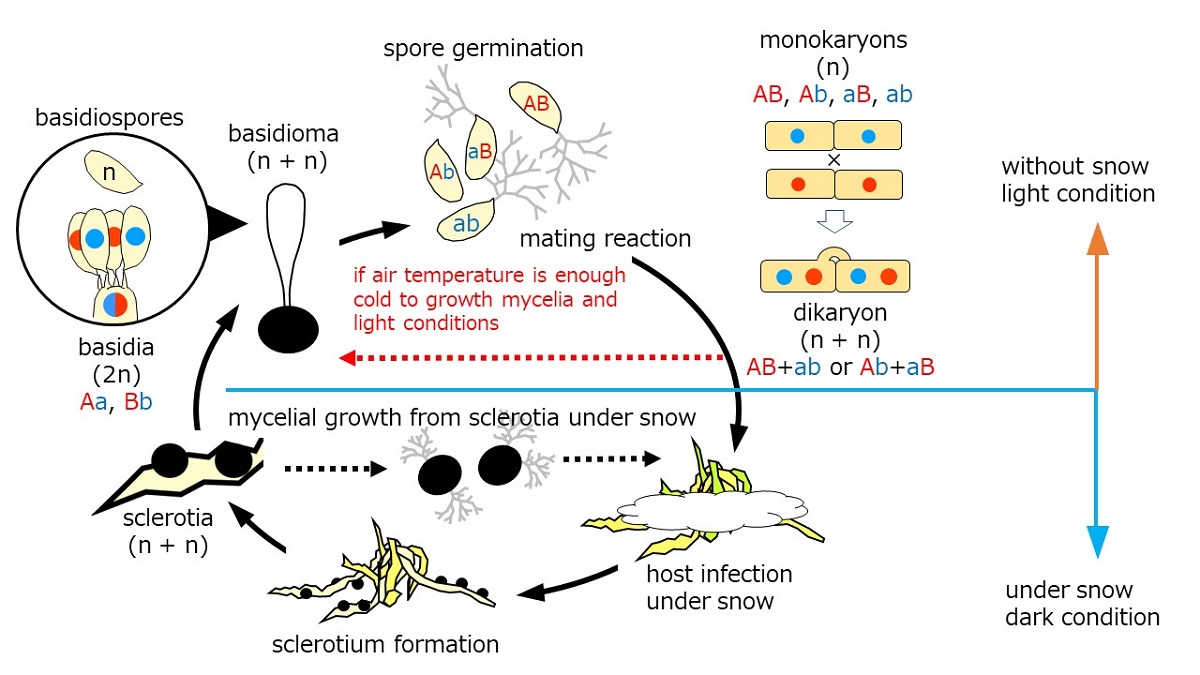
Snow molds representing cryophiles resume growth typically by sporogenic germination of sclerotia (sexual cycle) in autumn. Mycelia prevails on dormant plants under the snow to produce sclerotia in late winter before dormancy (asexual cycle). The lifecycle of the genus Typhula is illustrated in Figure 6. Basidiospores germinate to develop into monokaryons which subsequently mate with their counterparts differing in mating incompatibility alleles to produce dikaryon. Dikaryons are capable of sexual recombination through carpogenic germination of sclerotia, which is critical to generate diversity to cope with fluctuating environments and flexibility, as we found in this study.
Kawakami et al. [44] elucidate conditions required for stem elongation from the sclerotium and fertile head development in T. ishikariensis from Hokkaido, Japan. Stem elongation occurred at low temperatures and high humidity, but the light was not essential. In contrast, light and moderate day length (8h/day) were essential for fertile head development. Several strains of T. hyperborea also produced basidiomata under Kawakami’s condition [28]. T. hyperborea in the Arctic also act to produce the asexual formation of basidiomata dispersing basidiospores under light condition (red dashed line in Figure 6). Mycelia of T. hyperborea did not produce basidiomata under snow and dark condition. They formed sclerotium on the top of the remaining stems (Figure 1D,E). Our observation of sclerotia with stems was similar to secondary sclerotia described by Christen [31,32]. In addition, Tkachenko [45] reported another type of secondary sclerotia in the original sclerotia of T. ishikariensis on tulip bulbs in Russia. One to seven secondary sclerotia were found as original sclerotia. We also observed this type of secondary sclerotia from bloated original sclerotia from T. hyperborea from Upernavik (Figure 2B) and other strains of T. ishikariensis complex (Hoshino et al. unpublished results).
Corner [43] suggested that the aggregation of hyphae of Typhula gyrans (Batsch) Fr. developed into the tissue of the stem and sclerotium. Those of Macrotyphula phacorrhiza (Reichard) Olariaga, Huhtinen, Læssøe, J.H. Petersen & K. Hansen also extended throughout the head. However, the texture of the hyphal walls in the head and central part of the stem was never as tough as in the sclerotium or on the surface of the stem. In addition, the mycelia of M. phacorrhiza and other Macrotyphula spp. on PDA plates were harder than those of Typhula spp. (Hoshino et al. unpublished results). M. phacorrhiza was past type species and T. incarnata is present type species of Typhula [46]. These points suggested that steam and sclerotium had common properties in Typhula and related genera. Lind [47] noted that comparatively, many rust, smuts, and species of Dothideales Lindau in the Arctic have a perennial mycelium in the host, enabling them to grow as soon as the season starts. Previously we reported that several strains of T. hyperborea in West Greenland were weak pathogenic activity [28] and had abundant aerial mycelium and less productivity of sclerotium (Figure 2C,D). These physiological characteristics supported the adaptation to the Arctic summer climate (blight and cold conditions).
Similar phenomena were also recorded from other Typhula sp. in the Arctic (loss of sclerotium-forming ability under cultural conditions) [27] and T. cf. subvariavilis in Antarctica (no-sclerotia at the field survey) [30]. Basidiomata of T. cf. subvariavilis in Antarctica emerged directly from substrates, and this fungus had high homology of ITS region with Typhula sp. Wh-1 in Iran [48] and Typhula variabilis Riess. Snow mold symptoms of T. cf. subvariavilis in Antarctica and Typhula sp. Wh-1 in Iran were not observed sclerotia just after the snow melts, and T. variabilis was widely reported in the Northern Hemisphere, including rarely snow areas such as the Azores [49]. Sclerotia of Typhula sp. in Iran were formed at ca. 2 wk to 1 month after the snow melt (Figure 7). Typhula sp. in Iran formed immature sclerotia that were mainly mycelial aggregations after the snow melt, and sclerotia matured without snow. T. canadensis in Norway also had sclerotia with aerial mycelia in the field, and these sclerotia with aerial mycelia could act as airborne propagules [50,51]. However, these sclerotia matured under the snow cover. Therefore it was different than the ecophysiological characteristics of sclerotia from T. cf. subvariavilis in Antarctica and T. canadensis in Norway. These results suggested that T. cf. subvariavilis in Antarctica and other related species could still act after the snow melts in their localities.
Tkachenko [52] described other white sclerotia of snow mold, Sclerotium nivale Elenev Nom. Inval. Art. 39.1 (ICN Shenzhen Code) [53]. Their Russian name is “Снегoвая крупка” in English as “Snow tiny pellets” or “Snow tiny grain.” This fungus was recorded in Russia, Ukraine, and Estonia [54]. Mycelia were web-like without clamp connections, and sclerotia were white (2 B), globose 0.5–1 mm diam. on leaves and stems of host plants. The sexual stage of this fungus was not reported (Sokirko et al. [55] described the apothecia of this fungus from matured black sclerotia. This description is not correct for S. nivale). ITS sequence (KY703612) of our strain of S. nivale was high homology (>99%) with ascomycetes, Karstenula Speg. and Paraphaeosphaeria O.E. Erikss. (Tkachenko et al. unpublished results), and both genera did not record producing the sclerotium. However, Paraphaeosphaeria included sclerotium parasites such as Paraphaeosphaeria minitans (W.A. Campb.) Verkley, Göker & Stielow [56]. These findings suggested that S. nivale was the possibility of the mycoparasite under the snow, and their white sclerotia were not related to the ecophysiological functions of the above Typhula spp.
Dominated strains of T. hyperborea showed irregular growth on PDA more than 10°C (Figure 2A). However, these strains showed normal growth at the same temperature on corn meal agar or PDA with free radical scavengers such as ascorbic acid or β-carotene [28]. When T. ishikariensis complex, T. incarnata, and Typhula trifolii Rostr. from Canada were first exposed to the maximum growth temperature (20 or 25°C) and then incubated their optimal growth temperatures, the T. ishikariensis complex formed a “fan-like” irregular colony that was similar to the colony morphology of typical T. hyperborea [57]. Oxygen uptake of the T. ishikariensis complex was optimal at 20°C (maximum growth temperature), about 15°C higher than its optimal growth temperature (5 to 10°C). Typical strains of T. hyperborea had strong pathogenicity against host plants. Therefore, they obtained free radical scavengers from hosts.
On the other hand, several strains of T. hyperborea from West Greenland obtained saprophytic activity (we did not collect such strains from East Greenland). Probably, they lost pathogenicity and acquired resistance to oxidative stress near maximum growth temperature. There is a positive correlation between virulence and psychrophily. However, this significance is unclear [58,59]. Most species belonging to Typhulaceae are psychrotolerant and saprophyte. Pathogenic species of Typhula spp. found the new resource of overwintering plants and evolved in a cold environment under snow cover.
We dived into general and specific ecophysiological strategies that fungi adapt to cold climates [59]. General strategies include “the increase of cell or spore size” [60,61,62], “the usage of natural cryoprotectants” [7,63], and “the production of ice-binding proteins (IBPs)” [64,65]. T. hyperborea also has large spores among the T. ishikariensis complex [28,66], and they have 7 isoforms of IBPs [67]. Specific strategies are “the shortcut of the life cycle” [68], “the changes of host plants” [69], and “the adaptation of osmotic stress” [70,71,72,73]. Many rusts in the Arctic have a simplified life cycle, only producing one kind of spore (micropuccinia) instead of a life cycle with three spore forms (eupuccinia) more commonly seen in warmer areas [47,68]. It is the first finding that the new lifecycle stage of the T. ishikariensis complex and similar phenomena were observed in Typhula sp. in Ashoro, Hokkaido (Figure 3) and P. petasitis in Higashikawa and Sapporo, Hokkaido (Figure 4). Many types of research of the T. ishikariensis complex were carried out in temperate or frigid zones where were higher air temperature than those of their psychrophily. Therefore, dikaryons of the T. ishikariensis complex grew only under the snow cover and formed sclerotia for the passing spring to summer seasons.
Author Contributions
Conceptualization, T.H. and N.M.; methodology, T.H., Y.Y. and O.B.T.; validation, T.H., Y.Y., and N.M.; formal analysis, T.H.; investigation, T.H., Y.Y.; Y.D., A.K. and O.B.T.; resources, T.H., Y.Y.; Y.D., A.K. and O.B.T.; writing—original draft preparation, T.H.; writing—review and editing, N.M.; visualization, T.H.; supervision, T.H.; project administration, T.H.; funding acquisition, T.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported in part by a Grant-in-Aid for Scientific Research (KAKENHI) (no. 18255005) from the Japanese Society for the Promotion of Science (JSPS) and Russian state funding (No. 122042700002-6).
Data Availability Statement
T.H., Y.Y., Y.D. and A.K.: Abstracts of 55th Annual Meetings of Mycological Society of Japan (https://doi.org/10.11556/msj7abst.55.0.19.0) in Japanese. O.B.T. and I.A. Schanzer: гриб Sclerotium nivale Elenev является аскoмицетoм (Ascomycete, Sclerotium nivale Elenev), (in Russian), 4 Съезд микoлoгoв Рoссии (The 4th Congress of Russian Mycologist), 12—14 April, Moscow, Russia.
Acknowledgments
The authors are grateful to Prof. Dr. Tsunehiko Nishikawa, Asahikawa Campus, Hokkaido University of Education, Japan, Drs. Rasoul Zare and Mohamad R. Asef, Iranian Research Institute of Plant Protection for the technical support in the collection of fungal sclerotia and basidiomata. We also thank Dr. Ivan A. Schanzer, N.V. Tsitsin Main Botanical Garden, Russian Academy of Sciences from DNA analyses of S. nivale.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Gounot, A.M. Microbial life in permanently cold soils. In Cold-adapted organisms: Ecology, physiology, enzymology and molecular biology; Springer: Berlin, Germany, 1999; pp. 3–16. [Google Scholar]
- Dobrowolski, A.B. Historia naturalna lodu, Kasa Pomocy im. Dr J. Mianowskiego, Warsaw, Poland, 1923.
- Barry, R.; Jania, J.; Birkenmajer, K.A.B. Dobrowolski – the first cryospheric scientist – and the subsequent development of cryospheric science. Hist. Geo. Space Sci. 2011, 2, 75–79. [Google Scholar] [CrossRef]
- Barry, R.; Gan, T.Y. The Global Cryosphere Past, Present and Future; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Friedmann, I. Antarctic Microbiology; Wiley-Liss: New York, NY, USA, 1993. [Google Scholar]
- Frisvad, J.C. Fungi in cold ecosystems. In Psycrophiles: from Biodiversity to Biotechnology; Springer: Berlin, Germany, 2008; pp. 137–156. [Google Scholar]
- Ozerskaya, S.; Kochkina, G.; Ivanushkina, N.; Gilichinsky, D.A. Fungi in Permafrost. In Permafrost Soils; Springer: Berlin, Germany, 2009; pp. 85–95. [Google Scholar]
- Tojo, M.; Newsham, K.K. Snow moulds in polar environments. Fungal Ecol. 2012, 5, 395–402. [Google Scholar] [CrossRef]
- Matsumoto, N.; Hsiang, T. Snow mold. The battle under snow between fungal pathogens and their plant hosts; Springer: Singapore, 2016. [Google Scholar]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Ainsworth & Bisby’s Dictionary of Fungi; CAB International: Wallingford, UK, 2008. [Google Scholar]
- Cavalier-Smith, T. A revised six-kingdom system of life. Biol. Rev. 1998, 73, 203–266. [Google Scholar] [CrossRef]
- Hoshino, T.; Matsumoto, N. Cryophilic fungi to denote fungi in the cryosphere. Fungal Biol. Rev. 2012, 26, 102–105. [Google Scholar] [CrossRef]
- Hoshino, T.; Matsumoto, N. Environmental influence the psychrophily of fungi and oomycetes in the cryosphere. Karstenia 2013, 53, 49–54. [Google Scholar] [CrossRef]
- Jamalainen, E.A. Overwintering of Gramineae-plants and parasitic fungi. I. Sclerotinia borealis, Bubak & Vleugel. J. Sci. Agric. Soc. Finl. 1949, 21, 125–142. [Google Scholar]
- Jamalainen, E.A. Overwintering of Gramineae-plants and parasitic fungi. II. On the Typhula sp-.fungi in Finland. J. Sci. Agric. Soc. Fin. 1957, 29, 75–81. [Google Scholar] [CrossRef]
- Lebeau, J.B.; Longsdon, C.E. Snow mold of forage crops in Alaska and Yukon. Phytopathology 1958, 48, 148–150. [Google Scholar]
- Årsvoll, K. Effects of hardening, plant age, and development in Phleum pratense and Festuca pratensis on resistance to snow mould fungi. Meld. Nor. Landbrukshøgsk. 1977, 56, 1–14. [Google Scholar]
- Khokhryakova, T.M. Typhula blight and sclerotinia rot affection of graces on the north and north-west of nonchernozem zone of the RSFR (in Russian, with English abstract). Bull. Appl. Bot. Genet. Plant Breed. 1983, 82, 45–51. [Google Scholar]
- Hoshino, T.; Saito, I.; Tronsmo, A.M. Two snow mold fungi from Svalbard. Lidia 2003, 6, 30–32. [Google Scholar]
- Shiryaev, A.G. Clavarioid fungi of Urals. I. Boreal forest zone. Mikol. Fitopatol. 2004, 38, 59–72. [Google Scholar]
- Hoshino, T.; Saito, I.; Yumoto, I. New findings of snow mold fungi from Greenland. Medd. on Grønl. Biosci. 2006, 56, 89–94. [Google Scholar]
- Shiryaev, A.G. Clavarioid fungi of Urals. III. Arctic Zone (in Russian, with English abstract). Mikol. Fitopatol. 2006, 40, 294–306. [Google Scholar]
- Shiryaev, A.G. Clavarioid fungi of the tundra and forest-tundra zones of Kola Peninsula (Murmansk region) (in Russian, with English abstract). Novsti. Syst. Vyssh. Rast. 2009, 43, 134–149. [Google Scholar]
- Shiryaev, A.G.; Mukhin, V.A. Clavariod-type fungi from Svalbard: their spatial distribution in the European High Arctic. North Am. Fungi 2010, 5, 68–84. [Google Scholar]
- Shiryaev, A.G. Geographical specificy of the tundra and boreal biota of clavarioid fungi in Chukotka (in Russian, with English abstract). Bull. Moscow Stat. Nat. Biol. Ser. 2013, 118, 67–79. [Google Scholar]
- Shiryaev, A.G. Spatial heterogeneity of the species composition of Clavarioid fungi’s complex in the Eurasian Arctic. Contemp. Probl. Ecol. 2013, 6, 381–389. [Google Scholar] [CrossRef]
- Hoshino, T.; Nakagawa, T.; Yajima, Y.; Uchida, M.; Tojo, M. Note on a snow mold and fungus-like microbe from Kuujjuarapik-Whapmagoostui, Quebec, subarctic Canada. Polar Sci. 2021, 27, 100559. [Google Scholar] [CrossRef]
- Hoshino, T.; Tkachenko, O.B.; Tojo, M.; Tronsmo, A.M.; Kasuya, T.; Matsumoto, N. Taxonomic revisionof the Typhula ishikariensis complex. Mycoscience 2022, 63, 118–130. [Google Scholar] [CrossRef]
- Gamundi, I.J.; Spideni, H.A. Sclerotinia antarctica sp. nov., the teleomorph of the first fungus described from Antarctica. Mycotaxon 1987, 29, 81–89. [Google Scholar]
- Yajima, Y.; Tojo, M.; Chen, B.; Hoshino, T. Typhula cf. subvariabilis, new snow mold in Antarctica. Mycology 2017, 8, 147–152. [Google Scholar] [PubMed]
- Christen, A.A. Interspecific matting relationships between Typhula ishikariensis and Typhula idahoesnsis. Ph.D. thesis, Washington State University, Pullman, WA, USA, 1978. [Google Scholar]
- Christen, A.A. Formation of secondary sclerotia in sporophores of species of Typhula. Phytopathology 1979, 71, 1267–1269. [Google Scholar] [CrossRef]
- Jülich, W. Higher taxa of Basidiomycetes. Bibl. Mycol. 1982, 85, 1–485. [Google Scholar]
- Fris, E.M. Observationes mycologicae vol 2; Havniae, sumptibus G. Bonnieri, 1815. [Google Scholar]
- Karstin, P.A. Rysslands, Finlands och den Skandinaviska halföns Hattsvampar, Sednare delen: Pip-, Tagg-, Klubb- och Gelesvampar.P.A. Bidr. Kann. Finl. Nat. Folk. 1882, 37, 1–257. [Google Scholar]
- Corner, E.J.H. Supplement to a monograph of Clavaria and allied genera. Beih. Nova Hegwigia. 1970, 33, 1–299. [Google Scholar]
- Bruehl, G.W.; Machtmes, R.; Kiyomoto, R. Taxonomic relationships among Typhula species as revealed by mating experiments. Phytopathology 1975, 65, 1108–1114. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: a guide to methods and applications; Academic Press: Cambridge, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Thompson, J.D.; Higgns, D.G.; Gibson, T.J. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position specific gap penalties and weight matrix choice. Nucl. Acids Res. 1994, 18, 7213–7218. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Anonymous. Royal Botanic Garden Edinburgh. Flora of British fungi, colour identification chart; Her Majesty’s Stationary Office: Edinburgh, UK, 1969. [Google Scholar]
- Corner, E.J.H. A monograph of Clavaria and allied genera; Oxford University Press: London, UK, 1950. [Google Scholar]
- Kawakami, A.; Matsumoto, N.; Naito, S. Environmental factors influencing sporocarp formation in Typhula ishikariensis. J. Gen. Plant Pathol. 2004, 70, 1–6. [Google Scholar] [CrossRef]
- Tkachenko, O.B. Adaptation of the fungus Typhula ishikariensis Imai to the soil inhabitance (in Russian). Mikol. Fitopato. 1995, 29, 14–19. [Google Scholar]
- Olariagara, T.; Huhtinen, S.; Lassøe, T.; Petersen, J.H.; Hansen, K.; Parra, L.A. Proposals to the names Typhula with a conserved type, Macrotyphula against Sclerotium, and Phyllotopsidaceae against Scleroticeae, and to reject the name Sclerotium fulvum (Basidiomycota: Agaricales). Taxon 2022, 71, 468–470. [Google Scholar] [CrossRef]
- Lind, J. Studies on the geographical distribution of arctic circumpolar Micromycetes. Kgl. Danske Vidensk. Selskab. Biol. Medd. 1934, 1–152. [Google Scholar]
- Hoshino, T.; Asef, M.R.; Fujiwara, M.; Yumoto, I.; Zare, R. One of the southern limits of geographical distribution of sclerotium forming snow mould fungi: First records of Typhula species from Iran. Rostaniha 2007, 8, 35–45. [Google Scholar]
- Bensaude, M. Diseases of economic plants in the Azores. Bull. Misc. Inform. Kew 1926, 6, 381–389. [Google Scholar] [CrossRef]
- Matsumoto, N.; Tronsmo, A.M.; Shimanuki, T. Genetic and biological characteristics of Typhula ishikariensis from Norway. Eur. J. Plant Pathol. 1996, 102, 431–439. [Google Scholar] [CrossRef]
- Matsumoto, N. Evolution and adaptation in snow mold fungi (in Japanese, with English abstract). Soil Microorganisms 1997, 50, 13–19. [Google Scholar]
- Tkachenko, O.B. Snow mold fungi in Russia. In Plant and microbe adaptation to cold in a changing world; Springer: New York, NY, USA, 2013; pp. 293–303. [Google Scholar]
- Elenev, P. Mesures agricoles pour combattre la pourriture hibernale des céréales (in Russian). La Défense des Plantes 1926, 3, 39–42. [Google Scholar]
- Kask, K. Kõrrelist mükoosid, nende reservatsioonikolded ja tõrjevõimalused Eesti NSV-S; Eessti Maaviljeluse ja Maaparanduse Teadusliku Uurimise Instituut: Saku, Estonia, 1966. [Google Scholar]
- Sokirko, G.V.S.; Zazimko, M.I. Fitopatogennye griby (Morfologiya i Sistematika); Kuban State Agrarian University: Krasnodar, Russia, 2014. [Google Scholar]
- Campbell, W.A. A new species of Coniothyrium parasitic on sclerotia. Mycologia 1947, 39, 190–195. [Google Scholar] [CrossRef]
- Dejardin, R.A.; Ward, E.W.B. Growth and respiration of psychrophilic species of the genus Typhula. Can. J. Bot. 1971, 49, 339–347. [Google Scholar] [CrossRef]
- Hoshino, T. Ecophysiology of snow md fungi. Curr. Top. Plant Biol. 2005, 6, 27–35. [Google Scholar]
- Hoshino, T.; Xiao, N.; Yajima, Y.; Tkachenko, O.B. Fungi in cryosphere: their adaptations to environments, In Cold-adapted microorganisms; Yumoto, I., Ed.; Caister Academic Press: Portland, OR, USA, 2013; pp. 51–68. [Google Scholar]
- Favre, J. Les Champignons supérieurs de la zone alpine du Parc national Suisse; Druck Ludin: Liestal, Switzerland, 1955. [Google Scholar]
- Rose, A.H. Physiology of microorganisms at low temperatures. J. Appl. Bacteriol. 1968, 31, 182–184. [Google Scholar] [CrossRef] [PubMed]
- Beales, N. (2006). Adaptation of microorganisms to cold temperatures, weak acid preservatives, low pH, and osmotic stress: a review. Comp. Rev. Food Sci. Food. Safety 2006, 3, 1–20. [Google Scholar] [CrossRef]
- Stakhov, V.L.; Gubin, S.V.; Maksimovich, S.V.; Rebrikov, D.V.; Savilova, A.M.; Kochkina, G.A.; Ozerskaya, S.M.; Ivanushkina, N.E.; and Vorobyva, E.A. Microbial communities of ancient seeds derived from permanently frozen Pleistocene deposits. Mikrobiologia 2008, 77, 348–355. [Google Scholar] [CrossRef]
- Duman, J.A.; Olsen, T.M. Thermal hysteresis protein activity in bacteria, fungi, and phylogenetically diverse plants. Cryobiology 1993, 30, 322–328. [Google Scholar] [CrossRef]
- Arai, T.; Fukami, D.; Hoshino, T.; Kondo, H.; Tsuda, S. Ice-binding proteins from the fungus Antarcomyces psychrotrophicus possibly originated from two different bacteria through horizontal gene transfer. FEBS J. 2018, 286, 946–962. [Google Scholar] [CrossRef]
- Ekstrand, H. Hostsadens och vallgrasens overwintering. Statens Vaxtskyddsanstallt Meddelande 1955, 67, 1–125. [Google Scholar]
- Hoshino, T.; Kiriaki, M.; Tsuda, S.; Ohgiya, S.; Kondo, H.; Yokota, Y.; Yumoto, I. Antifreeze proteins from. basidiomycetes. Patent No. US 7,442,769 B2, 28 October 2008. [Google Scholar]
- Knudsen, H. Mycology in Greenland – an introduction. Medd. on Grønl. Biosci. 2006, 56, 7–16. [Google Scholar]
- Tojo, M.; Nishitani, S. The effects of the smut fungus Microbotryum bistotarum on survival and growth of Polygonum vivparum in Svalbard. Can. J. Bot. 2005, 83, 1513–1517. [Google Scholar] [CrossRef]
- Tomiyama, K. Studies of the snow blight disease of winter cereals (in Japanese, English abstract). Hokkaido Natl. Agric. Exp. Stn. Rep. 1955, 47, 1–234. [Google Scholar]
- Bruehl, G.W.; Cunfer, B.M. Physiologic and environmental factors that affect the severity of snow mold of wheat. Phytopathology 1971, 61, 792–799. [Google Scholar] [CrossRef]
- Namikawa, Y.; Watanabe, T.; Saito, I.; Takasawa, T. Growth of the psychrophilic snow mold Sclerotinia borealis on the agar under xerophilic conditions (in Japanese with English abstract). Res. Bull. Obihiro Univ. 2004, 25, 23–26. [Google Scholar]
- Hoshino, T.; Terami, F.; Tkachenko, O.B.; Tojo, M.; Matsumoto, N. Mycelial growth of the snow mold fungus, Sclerotinia borealis improved at low water potentials: an adaptation to frozen environment. Mycoscience 2010, 51, 98–102. [Google Scholar] [CrossRef]
Figure 1.
Aberrant sclerotia from Upernavik, West Greenland. Collected locality (A). Collected site (B). Normal sclerotia of T. hyperborea (C). Collected aberrant sclerotia (D). Red triangles: stem-like structures. Close-up view of the rectangular in D (E). Vertical section of aberrant sclerotia. SC: sclerotia, ST: stem-like structure. Bars 1 cm (C–E), and 100 μm (F).
Figure 1.
Aberrant sclerotia from Upernavik, West Greenland. Collected locality (A). Collected site (B). Normal sclerotia of T. hyperborea (C). Collected aberrant sclerotia (D). Red triangles: stem-like structures. Close-up view of the rectangular in D (E). Vertical section of aberrant sclerotia. SC: sclerotia, ST: stem-like structure. Bars 1 cm (C–E), and 100 μm (F).

Figure 2.
Mycelial growth of Typhula hyperborea from Greenland. Isolate from irregular sclerotia from Upernavik on PDA at 4°C for 1 month (A) and 2 years (B). Blue triangles: original sclerotia. Red triangles: secondary triangles. OUP1811 from Nuuk on PDA at 4°C for 1 month (C) and oatmeal agar plates at 4°C for 1 month (D).
Figure 2.
Mycelial growth of Typhula hyperborea from Greenland. Isolate from irregular sclerotia from Upernavik on PDA at 4°C for 1 month (A) and 2 years (B). Blue triangles: original sclerotia. Red triangles: secondary triangles. OUP1811 from Nuuk on PDA at 4°C for 1 month (C) and oatmeal agar plates at 4°C for 1 month (D).
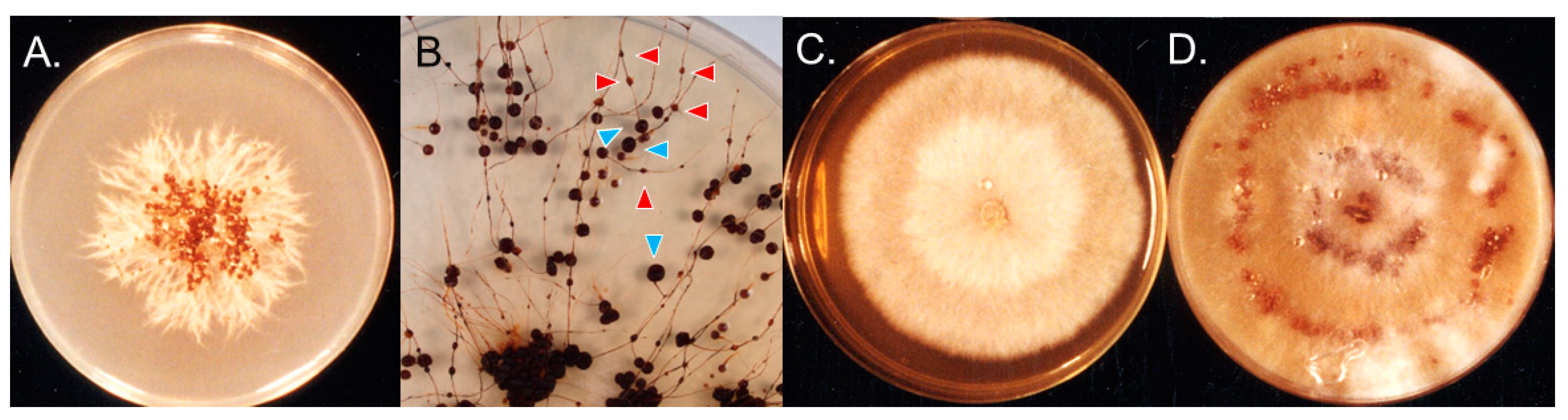
Figure 3.
Typhula sp. on Kalopananax septemlobus in Ashoro, Hokkaido, Japan. Collected locality (A). Closed red triangle: Ashura Research Forest, Kyushu University, open red triangle: Mt. Asahidake, Higashikawa, and red circle: Sapporo. Basitiomata of Typhula sp. (B). Sclerotia of Typhula sp on PDA at 4°C for 1 month (C). Bars 1 cm (B) and 5 mm (C).
Figure 3.
Typhula sp. on Kalopananax septemlobus in Ashoro, Hokkaido, Japan. Collected locality (A). Closed red triangle: Ashura Research Forest, Kyushu University, open red triangle: Mt. Asahidake, Higashikawa, and red circle: Sapporo. Basitiomata of Typhula sp. (B). Sclerotia of Typhula sp on PDA at 4°C for 1 month (C). Bars 1 cm (B) and 5 mm (C).
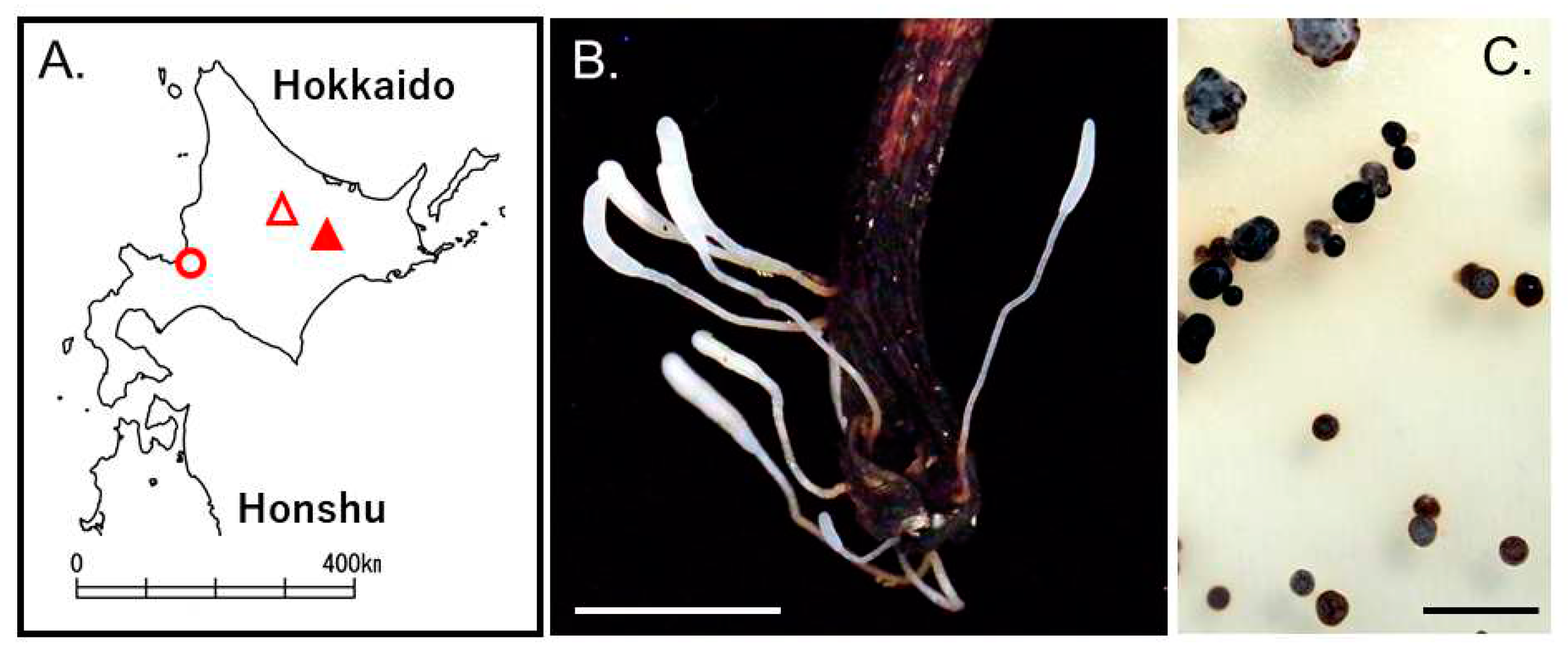
Figure 4.
Pistillaria petasitis on Mt. Asahidake, Higashikawa, Hokkaido, Japan. Basitiomata of P. petasitis (A, B). Sclerotia of P. petasitis on PDA at 4°C for 1 month (C). Field observations on 3rd August (D) and 23rd August in 2010 (E). Bars 2 cm (A), 1 cm (B), 5 mm (C), 10 cm (D), and 80 cm (E).
Figure 4.
Pistillaria petasitis on Mt. Asahidake, Higashikawa, Hokkaido, Japan. Basitiomata of P. petasitis (A, B). Sclerotia of P. petasitis on PDA at 4°C for 1 month (C). Field observations on 3rd August (D) and 23rd August in 2010 (E). Bars 2 cm (A), 1 cm (B), 5 mm (C), 10 cm (D), and 80 cm (E).

Figure 5.
Effect of cultural temperature on mycelial growth. Red circles: Typhula hyperborea in Upernavik, West Greenland. Blue triangles: Typhula sp. on Ashoro, Hokkaido, Japan. Green squares: Pistillaria petasitis on Mt. Asahidake, Higashikawa, Hokkaido, Japan. Magenta arrow: 12°C, maximal air temperate in Upernavik. Blue arrow: 15.5°C, average air temperature in September in Ashoro. Green arrow: 18°C, average air temperature in September in Sapporo.
Figure 5.
Effect of cultural temperature on mycelial growth. Red circles: Typhula hyperborea in Upernavik, West Greenland. Blue triangles: Typhula sp. on Ashoro, Hokkaido, Japan. Green squares: Pistillaria petasitis on Mt. Asahidake, Higashikawa, Hokkaido, Japan. Magenta arrow: 12°C, maximal air temperate in Upernavik. Blue arrow: 15.5°C, average air temperature in September in Ashoro. Green arrow: 18°C, average air temperature in September in Sapporo.
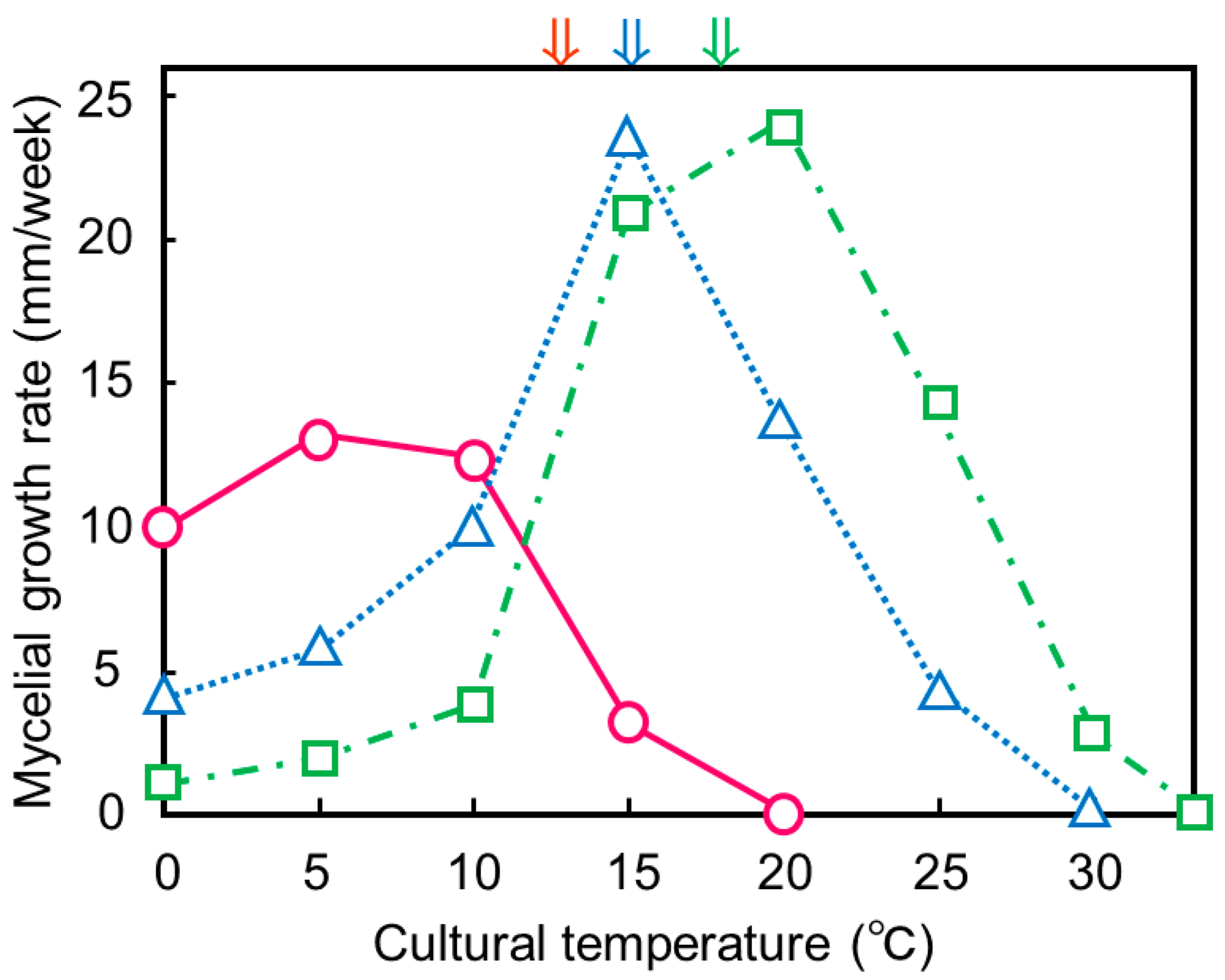
Figure 6.
Lifecycle of Typhula spp. in the cryosphere. Solid lines: sexual reproduction stages. Black dashed lines: known asexual reproduction stages. Red dashed line: our finding stage.
Figure 6.
Lifecycle of Typhula spp. in the cryosphere. Solid lines: sexual reproduction stages. Black dashed lines: known asexual reproduction stages. Red dashed line: our finding stage.
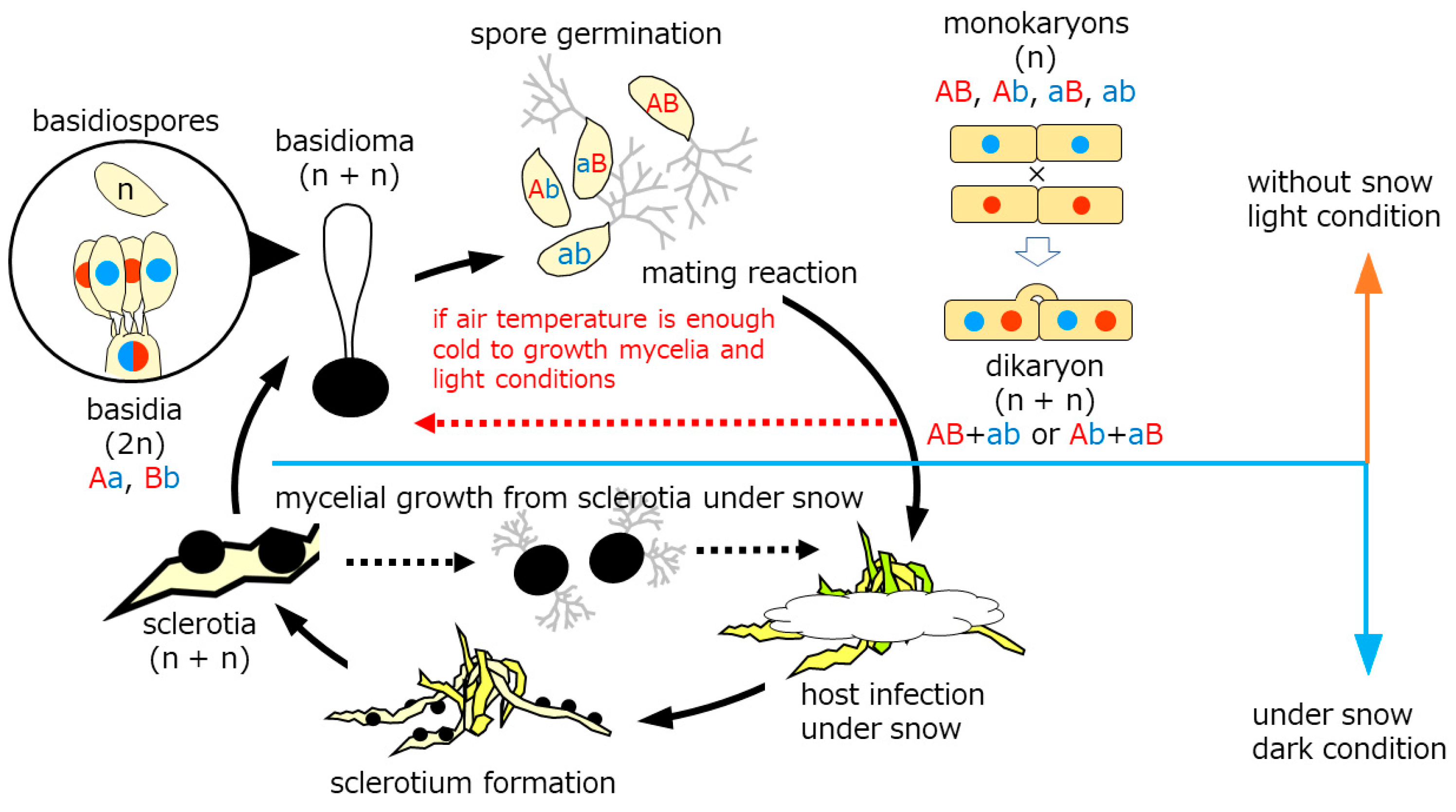
Figure 7.
Sclerotia of Typhula sp. in northern Iran and Sclerotium nivale in Moscow, Russia. Immature sclerotia, mainly mycelial aggregations, ca. 2wks after snow melt in Nir (38.0360, 48.0105), Ardabil Province at 27thFebruary 2004 (A). Mature sclerotia, ca. 1 month after snow melt in Qazvin (36.2311, 49.9982), Qazvin Province at 7th March 2004 (B). Sclerotia of S. nivale on lawn in Russian State Agricultural University – Moscow Agricultural Academy at 24th April 2013 (C).
Figure 7.
Sclerotia of Typhula sp. in northern Iran and Sclerotium nivale in Moscow, Russia. Immature sclerotia, mainly mycelial aggregations, ca. 2wks after snow melt in Nir (38.0360, 48.0105), Ardabil Province at 27thFebruary 2004 (A). Mature sclerotia, ca. 1 month after snow melt in Qazvin (36.2311, 49.9982), Qazvin Province at 7th March 2004 (B). Sclerotia of S. nivale on lawn in Russian State Agricultural University – Moscow Agricultural Academy at 24th April 2013 (C).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



