
Submitted:
27 June 2023
Posted:
28 June 2023
You are already at the latest version
Abstract
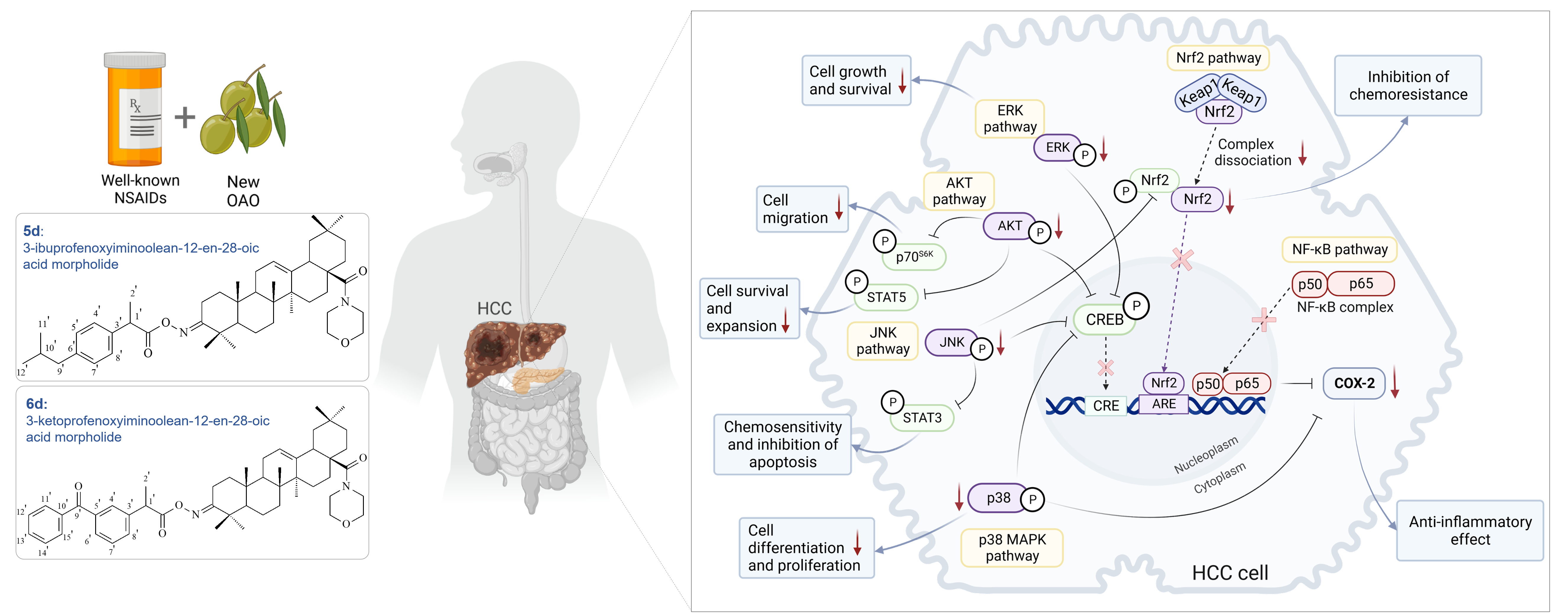
HCC is a highly aggressive malignancy with limited treatment options. In this study, novel conjugates, of NSAIDs - Ibuprofen and Ketoprofen with oleanolic acid oximes derivatives (OAO) were synthesized and their activity as modulators of signaling pathways involved in HCC pathogenesis was evaluated in normal THLE-2 liver cells and HCC derived HepG2 cells. The results demonstrated that conjugation with OAO derivatives reduces the cytotoxicity of parent compounds in both cell lines. In THLE-2 cells treatment with conjugates resulted in increased activation of the Nrf2-ARE pathway. An opposite effect was observed in HepG2 cells. In the later reduction of NF-κB was observed along with modulation of MAPK signaling pathways (AKT, ERK, p38, p70S6K, and JNK). Moreover, STAT3, STAT5, and CREB transcription factors on protein levels were significantly reduced as a result of treatment with IBU- and KET-OAO derivatives conjugates. The most active were conjugates with OAO-morpholide. Overall, the findings of this study demonstrate that IBU-OAO and KET-OAO derivatives conjugates modulate the key signaling pathways involved in hepatic cancer development. Their effect on specific signaling pathways varied depending on the structure of the conjugate. Since the conjugation of IBU and KET with OAO derivatives reduced their cytotoxicity, the conjugates may be considered good candidates for the prevention of liver cancer.
Keywords:
Nrf2
; NF-κB
; MAPK
; HCC
; Ibuprofen
; Ketoprofen
; oleanolic acid oximes
; liver cancer
; NSAIDs
1. Introduction
Hepatocellular carcinoma (HCC) remains a global health burden characterized by high mortality rates and limited treatment options. Chronic liver inflammation, often driven by viral hepatitis, alcohol abuse, or nonalcoholic steatohepatitis, significantly contributes to the development and progression of HCC [1].
Nrf2-ARE and NF-κB signaling pathways play a key role in HCC development. The Nrf2-ARE signaling pathway is responsible for the transcription of genes encoding cytoprotective proteins. Therefore, its activation at the initiation step may protect against cancer development, while NF-κB activated in response to inflammatory stimuli contribute to the promotion of the carcinogenesis process [2,3]. One of the target genes of the latter is COX-2, which overexpression characterizes several cancers including HCC [4,5,6]. Usually, reduced activation and expression of NF-κB results in the opposite effect in the Nrf2 signaling pathway [7]. However, it has also been demonstrated that Nrf2, is often overexpressed in cancer cells, including HCC, and may lead to an enhanced invasiveness potential and chemo-and radio-resistance [8]. Therefore, inhibition of Nrf2 is highly desired at the later stages of cancer development. In addition to NF-κB and Nrf2, MAPK signaling pathways, such as Jak/STAT, CREB, and PI3K/Akt, are involved in liver cancer development and inflammation responses. Dysregulation of MAPK pathways has been implicated in liver cancer progression. Furthermore, these pathways exhibit intricate crosstalk, suggesting that targeting multiple signaling pathways holds promise as a therapeutic approach to HCC [9,10]. Due to the inflammatory background of several cancers, including HCC, non-steroidal anti-inflammatory drugs (NSAIDs) have been proposed for their prophylaxis and/or therapy [11,12,13,14].
However, although efficient, those drugs exhibit unfavorable side effects in particular related to the insurgence of stomach or intestinal bleeding and of circulatory or cardiac deregulation [15]. Thus, natural alternatives are searched. One of them is oleanolic acid. It possesses strong hepatoprotective and anti-inflammatory potential but very low bioavailability [16,17,18,19]. Hence, a reasonable strategy is to modify its chemical structure to obtain derivatives with increased solubility, permeability, and enhanced therapeutic potential. Distinguished strategies for modifying its structure include synthesizing oximes, esters, and amides [20]. Our earlier study showed that oleanolic acid oximes (OAO) substituted at the C-17 with methyl, benzyl ester, or morpholide group significantly increases their ability to modulate the NF-κB signaling pathway [21]. Moreover, conjugation of these derivatives with NSAIDs such as aspirin, indomethacin, or diclofenac enhanced their modulating effect on key cellular signaling pathways ultimately leading to induction of apoptosis and reduced proliferation of HCC cells and reducing tumor burden in vivo [22,23,24].
Ibuprofen and Ketoprofen are other representatives of NSAIDs that are used for many years due to their anti-inflammatory, analgesic, and antipyretic, properties. Like many other NSAIDs, they inhibit the cyclooxygenase enzyme (COX) and act by decreasing the production of prostaglandin inflammatory precursors [25,26]. Their anticancer activity was proven in some types of cancer e.g., of lung cancer [27,28]. In one prospective data analysis performed in 2012 in contrast to aspirin which use was associated with reduced risk of developing HCC and of death due to chronic liver diseases (CLD), Ibuprofen and Ketoprofen were associated only with reduced risk of death due to CLD [29].
Thus, the aim of this study was the synthesis and evaluation of the potential anticancer activity of novel oleanolic acid oximes derivatives (OAO) conjugated with Ibuprofen (IBU-OAO) and Ketoprofen (KET-OAO) on HCC-derived HepG2 cells and THLE-2 normal, immortalized hepatocytes. Their effect on cell viability and ability to modulate the Nrf2-ARE, NF-κB, and MAPK signaling pathways was evaluated in order to compare with the earlier studied conjugates and select the optimal structures of the compounds, for future in vivo studies.
2. Results
2.1. Chemistry
2.2. Spectral characteristics of the OAO conjugates with Ibuprofen and Ketoprofen
2.2.1. Conjugates of OAO and Ibuprofen (5a-d)
In the IR spectra of conjugates 5a-d, the presence of an aromatic ring within the Ibuprofen moiety ([CH3]2CH-CH2-C6H4-CH[CH3]-COON=C-3) was confirmed by the presence of an absorption band located at ν 3335 – 3340 cm-1. The signal observed at ν 1725 – 1735 cm-1 was assigned to the C=O group within the –COO function of the Ibuprofen moiety ([CH3]2CH-CH2-C6H4-CH[CH3]-COON=C-3). The next characteristic absorption band, observed at ν 1720 or 1725 cm-1, was assigned to –N=C-3 of the acyloxyimino function.
The NMR analysis of conjugates 5a-d confirmed the structures of these compounds.
In the 13C NMR spectra of the above conjugates, the signal of the C=O group within the –COO function of the Ibuprofen moiety ([CH3]2CH-CH2-C6H4-CH[CH3]-COON=C-3) was observed at the δ 176 ppm. Signals derived from the aromatic ring of Ibuprofen moiety were present at δ about: 140, 138 (2 x Cq), 137, 129, 128, and 127 (4 x CH) ppm. Other signals characteristic for the Ibuprofen system were observed at δ about 45 ppm (CH2, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3), 41 ppm (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3), 27 ppm (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3), 22 ppm (CH3 x 2, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3) and 15 ppm (CH3, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3). Four signals characteristic for the oleanane system were noticed at δ about: 172 ppm (Cq, C-3), 144 ppm (Cq, C-13), 122 ppm (CH, C-12) and 47 ppm (Cq, C-17).
In the 1H NMR spectra of oleanolic acid oximes conjugates with Ibuprofen, the protons of an aromatic system within Ibuprofen moiety formed two signals: a doublet of doublets (present at δ about 7.2 ppm) and a doublet (present at δ about 7.1 ppm). The other six signals characteristic for the Ibuprofen system were observed at δ about: 3.8 ppm (a quartet for [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3), 2.5 ppm (a doublet for [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3), 2.0 ppm (a triplet of triplets for [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3), 1.5 ppm (a doublet for [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3), 1.2 and 1.1 ppm (two doublets for ([CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3). Two signals characteristic for the oleanane system were present at δ about 5.3 ppm (a triplet for C12-H) and 2.9 ppm (a doublet of doublets for C18-Hβ).
2.2.2. Conjugates of OAO and Ketoprofen (6a-d)
In the IR spectra of conjugates 6a-d, the presence of aromatic rings within the Ketoprofen moiety (C6H5-CO-C6H4-CH[CH3]-COON=C-3) was confirmed by the presence of an absorption band located at ν 3330 – 3335 cm-1. The signal observed at ν 1730 – 17405 cm-1 was assigned to the C=O group within the –COO function of the Ketoprofen moiety (C6H5-CO-C6H4-CH[CH3]-COON=C-3). The next characteristic absorption band, observed at ν 1720 or 1725 cm-1, was assigned to –N=C-3 of the acyloxyimino function.
The NMR analysis of conjugates 6a-d confirmed the structures of these compounds.
In the 13C NMR spectra of the above conjugates, the signal of the C=O group that joined two aromatic rings within the Ketoprofen moiety (C6H5-CO-C6H4-CH[CH3]-COON=C-3) was observed at δ about 197 ppm. The signal of the C=O group within the –COO function of the Ketoprofen moiety (C6H5-CO-C6H4-CH[CH3]-COON=C-3) was observed at the δ 176 ppm. Signals derived from an aromatic ring of Ketoprofen moiety were present at δ about: 141 ppm (Cq), 138 ppm (Cq), 131 ppm (CH), 129 ppm (CH), 129 ppm (CH), 129 ppm (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3) and 137 ppm (Cq), 132 ppm (CH), 130 ppm (2 x CH) and 128 ppm (2 x CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3). The other two signals characteristic for the Ibuprofen system were observed at δ about: 44 ppm (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3) and 18 ppm (CH3, C6H5-CO-C6H4-CH[CH3]-COON=C-3). Four signals characteristic for oleanane system were noticed at δ about: 172 ppm (Cq, C-3), 144 ppm (Cq, C-13), 122 ppm (CH, C-12) and 47 ppm (Cq, C-17).
In the 1H NMR spectra of conjugates 6a-d with Ketoprofen, the protons of aromatic systems within Ketoprofen moiety (C6H5-CO-C6H4-CH-[CH3]-COON=C3), formed four signals: a doublet of doublets located at δ about 7.8 ppm, a doublet observed at δ about 7.7 ppm, a triplet observed at δ about 7.6 ppm and a multiplet at δ about 7.5 – 7.4 ppm. Two more signals derived from Ketoprofen moiety were present at δ about 4 ppm 3.98 (a quartet for C6H5-CO-C6H4-CH-[CH3]-COON=C3) and 1.6 ppm (a doublet for C6H5-CO-C6H4-CH-[CH3]-COON=C3). Two next signals, characteristic for oleanane system, were located at δ about 5.30 (a triplet for C12-H) and δ about 3 ppm (a doublet of doublets for C18-Hβ).
2.3. Conjugation with OAO derivatives decreases the cytotoxicity of Ibuprofen and Ketoprofen
The viability of the immortalized hepatocytes line THLE-2 and HCC-derived HepG2 cells was assessed using the MTT assay after treatment with IBU, KET, and their conjugates at a concentration range of 1–150 µM. As depicted in Figure 2A–D, the conjugation of IBU and KET with OAO resulted in a reduction of cytotoxicity in both cell lines. Notably, the majority of the conjugates exhibited lower cytotoxicity compared to the parent compounds (IBU and KET). However, IBU-OAO morpholide conjugate 5d in THLE-2 cells and OAO-KET morpholide conjugate 6d in HepG2 cells displayed a slight deviation from this trend, as they reached the IC50 values, albeit with a marginal difference, 145.8 µM, and 149.2 µM, respectively.
Based on the results obtained from the MTT assay, subsequent assays were performed using a concentration of 30 µM, which ensured more than 70% cell viability. This concentration was chosen to further investigate the effects of the compounds on subsequent molecular and cellular processes.
2.4. Conjugation of Ibuprofen and Ketoprofen with OAO derivatives increases activation and expression of Nrf2 in THLE-2 cells but reduces in HepG2 cells
Activation of Nrf2 requires translocation from the cytosol to the nucleus. In THLE-2 cells, treatment with KET-OAO conjugates 6a and 6d resulted in increased levels of nuclear Nrf2 protein (Figure 3A). Furthermore, treatment with IBU-OAO morpholide 5d and KET-OAO morpholide 6d led to an enhanced binding to the antioxidant response element (ARE) sequence, as measured by the amount of Nrf2 contained in the DNA binding complex to the ARE sequence. The increase in binding was observed to be 25% and 30%, respectively. The oligonucleotides containing ARE consensus-binding site (5′-GTCACAGTGACTCAGCAGAATCTG-3’) for Nrf2 were immobilized on microplates as bait.
To investigate the effect on Nrf2 gene expression at the transcript level, the levels of Nrf2 mRNA were assessed. Treatment with all IBU-OAO conjugates resulted in increased expression of the Nrf2 gene. In normal hepatocytes, IBU-OAO conjugates 5a and 5d demonstrated the most pronounced effect showing a 1.75-fold and 1.65-fold increase in Nrf2 gene expression, respectively. Among the KET-OAO hybrids, the most effective was compound 6d i.e., the morpholide derivative conjugated with KET, showing a 3.5-fold increase compared to the control (Figure 3C).
In contrast, in HepG2 cells, treatment with IBU-OAO 5d, KET-OAO 6a, and 6d led to a reduction in the level of nuclear Nrf2 protein (Figure 3D). Similarly, the binding of Nrf2 to the ARE sequence was reduced by IBU-OAO 5c, 5d, and KET-OAO 6d (Figure 3E). Consistent with the protein level results, the Nrf2 mRNA level was diminished after treatment with IBU-OAO 5b, 5c, 5d, and 6a, ranging from 25% to 57% reduction (Figure 3F). Notably, the most effective modulator of Nrf2 activation and expression in both cell lines was the KET-OAO conjugate 6d.
These findings demonstrate that the conjugation of IBU and KET with OAO increases the activation and expression of Nrf2 in THLE-2 cells while producing opposing effects in HepG2 cells. The KET-OAO conjugate 6d exhibited the most significant modulation of Nrf2 activation and expression.
2.5. Ibuprofen and Ketoprofen - OAO conjugates reduce the activation and expression of NF-κB and COX-2 in HepG2 cells
To assess the effect of IBU-OAO and KET-OAO conjugates on NF-κB activity, the binding of p50 and p65 subunits to their consensus site and their nuclear levels was examined. Treatment with IBU-OAO conjugate 5a resulted in a reduction in p50 binding (~24%), while treatment with KET conjugates 6b, 6c, and 6d decreased p50 binding by approximately 30-35%. The binding of the p65 subunit was not significantly affected (Figure 4B,E). Moreover, compounds 5d, 6c, and 6d led to a reduction in the nuclear level of the p50 subunit protein (Figure 4A), increasing the content of the NF-κB whole protein complex in the cytosol (Figure 4G). Furthermore, treatment with KET-OAO conjugates, specifically compounds 6a, 6c, and 6d, decreased the level of phosphorylated NF-κB (Figure 4H).
At the transcript level, compounds 5d and 6d reduced the expression of the NF-κB p50 subunit gene, while compound 5d decreased the expression of the NF-κB p65 subunit gene by approximately 36% (Figure 4C,F).
Since COX-2 is a target gene of NF-κB and is often overexpressed in hepatic cancer cells, its cytosolic protein and mRNA levels were evaluated. Treatment with compounds 5d, 6c, and 6d resulted in reduced COX-2 transcript and protein levels by approximately 25-30% (Figure 5A). Compound 5c decreased the level of COX-2 transcript without altering its protein level (Figure 5B).
These findings demonstrate that IBU and KET -OAO conjugates in different ways affect NF-κB activity but ultimately reduced its target gene i.e., the COX-2 expression HCC cells.
2.6. Bead-Based Multiplex Immunoassay revealed possible modulation of protein regulating several signaling pathways by Ibuprofen and Ketoprofen - OAO derivatives conjugates
The effect of tested compounds on the total, referring to the overall abundance of an examined protein and phosphorylated (active) protein levels of kinases from the MAPK family, including AKT, JNK, p38, ERK 1/2, and p70S6K, using a bead-based multiplex immunoassay in liver cancer cells lysates. These kinases play crucial roles in various cellular processes and signaling pathways, and their dysregulation has been implicated in liver cancer progression.
As shown in Figure 6A, treatment with compounds 5c, 6c, and 6d led to a significant reduction in the total protein levels of AKT and phospho-AKT (only 6c) in HepG2 cells. The total and phosphorylated protein levels of the extracellular signal-regulated protein kinase (ERK) (Figure 6B) were altered by treatment with KET-OAO derivatives 6b, 6c, and 6d. These changes suggest a potential disruption in ERK-mediated cellular processes, which may also impact liver cancer cell growth and differentiation.
Notably, the levels of p38 (Figure 6C), a stress-activated protein kinase, were strongly diminished by all OAO-KET conjugates, while from OAO-IBU conjugates only morpholide 5d had an impact on its protein level. Additionally, the total protein level of p70S6K (Figure 6D), a key regulator of protein synthesis and cell growth, was diminished by treatment with IBU-OAO conjugates 5b and 5d, as well as OAO-KET derivatives 6c and 6d.
Finally, the protein levels of the c-Jun N-terminal kinase (JNK), which is downstream of AKT and involved in regulating cell proliferation and apoptosis, were also reduced upon treatment with IBU-OAO derivatives 5c and 5d, as well as KET-OAO conjugates 5a, 6b, and 6d (Figure 6E). This decrease in JNK protein levels along with reduction of p70S6K indicates a potential impairment of the signaling cascades involved in liver cancer cell growth and survival.
The effect of IBU-OAO and KET-OAO conjugates on the levels of STAT3, STAT5, and CREB proteins in HepG2 cell lysates was assessed using the same bead-based multiplex immunoassay. The results revealed that compounds 5d and 6d, the morpholide conjugates, led to a reduction in the constitutive/total protein level of STAT3. These compounds, along with compound 5c, also decreased the level of phosphorylated STAT3 (Figure 7A), indicating a potential inhibition of STAT3 activation. Similarly, the phosphorylated level of STAT5 was reduced by compounds 5c and 5d, but there was no significant change in constitutive STAT5 protein level observed (Figure 7B). Both STAT3 and STAT5 transcription factors play a critical role in cell proliferation and survival, and its reduced protein levels indicate a potential disruption of its signaling pathway.
Regarding CREB, the total/constitutive protein level was affected by compound 5a and KET-OAO conjugates 6b and 6d. However, the phosphorylated form of CREB remained unchanged following treatment with any of the tested compounds (Figure 7C). CREB is another transcription factor involved in cell differentiation and survival, and its altered protein levels suggest a potential modulation of CREB-mediated gene expression in liver cancer cells.
3. Discussion
The aim of this study was to investigate the potential of newly synthesized oleanolic acid oxime derivatives (OAO) conjugated with Ibuprofen (IBU-OAO) and Ketoprofen (KET-OAO) as modulators of the signaling pathways, which dysfunction play crucial roles in the liver cancer pathogenesis. Therefore, the study focused on the NF-κB, Nrf2, and MAPK signaling pathways in immortalized normal hepatocytes and the cells derived from HCC.
Generally, the results confirmed our earlier findings that OAO morpholide is not only the most active among the tested OAO derivatives but also forms the most effective hybrids with NSAIDs.
However, it is worth noting the marked differences in the effect on the parameters evaluated between the KET and IBU conjugates and previously tested indomethacin, diclofenac, or aspirin conjugates. While, indomethacin and diclofenac conjugates with OAO, particularly substituted with morpholide, significantly increased the toxicity of these NSAIDs [23,24], the conjugation of IBU and KET with OAO derivatives reduced the cytotoxicity of parent compounds in both normal hepatocytes and liver cancer cells.
To a certain extent, a similar effect was observed as a result of the conjugation of aspirin with OAO derivatives. However, in this case, conjugation with aspirin reduced the cytotoxicity of OAO derivatives [21] suggesting the chemopreventive potential of the conjugates. This suggestion was further supported by the activation of the Nrf2-ARE pathway in normal hepatocytes and the reduction of NF-κB activity in both normal and cancer liver cells [22].
Activation of the Nrf2-ARE pathway in normal immortalized hepatocytes was observed in this study as a result of treatment with KET-OAO conjugates demonstrated by increased levels of nuclear Nrf2 protein and enhanced binding to the antioxidant response element (ARE) sequence. Moreover, the increased Nrf2 gene transcript indicated a possible contribution to enhanced Nrf2 activation upregulation of Nrf2 expression, and ultimately defense against oxidative or electrophilic stress in these cells.
However, in cancer HepG2 cells, treatment with IBU-OAO or KET-OAO conjugates led rather to a reduction in the level of nuclear Nrf2 protein and binding to ARE sequence, suggesting the tendency to downregulation of this pathway. The reduced mRNA transcript level further supports the suggestion.
As overexpression of Nrf2 due to genetic and epigenetic alterations often occur in cancer cells including HCC, its reduced expression and activation may prevent enhanced invasiveness potential and chemo- and radioresistance [30]. It has to be stressed, that the most potent inducers in THLE-2 i.e., immortalized normal hepatocytes and inhibitors in HepG2 cells were the hybrids of IBU and KET with OAO substituted with morpholide group.
Therefore, these compounds might be considered chemopreventive agents applicable in HCC prophylaxis and supporting conventional therapy respectively. Interestingly, recently it was found that IBU can induce ferroptosis of glioblastoma cells via downregulation of the Nrf2 signaling pathway [31]. It is possible that a similar effect may occur in HCC cells as a result of treatment with IBU- and KET-OAO derivatives conjugates.
The impact of IBU-OAO and KET-OAO conjugates on the NF-κB pathway, in terms of binding of its active subunits p50 and p65 into DNA, and their translocation from the cytosol into the nucleus was also evaluated. In contrast to the effect of previously tested NSAIDs, particularly diclofenac [23], p50 binding to DNA and the nuclear level was more affected than the p65 NF-κB subunit. The latter was reduced only as a result of treatment with IBU conjugated with OAO morpholide derivative. The p65 subunit is basically responsible for the transcription initiation, while p50 serves as a helper in NF-κB DNA binding [32,33]. Since the disproportionate increases in the active p65 subunits often occur in cancer this subunit is recognized as an important target for novel drug design [34]. Hence, IBU-OAO morpholide may be worth further studies in this context. However, treatment with KET-OAO conjugates, specifically compounds 6a (KET-OAO carboxylic derivative) and 6d (KET-OAO morpholide derivative), decreased the level of phosphorylated NF-κB.
Several studies have revealed the crucial contribution of NF-κB phosphorylation to controlling NF-κB directed transactivation. Moreover, it was shown that NF-κB phosphorylation controls transcription in a gene-specific manner [35]. Therefore, IBU and KET -OAO derivatives conjugates may affect different mechanisms of NF- κB activation. Ultimately it led to reduced expression of its target COX-2 gene often overexpressed in hepatic cancer cells [36,37]. In this regard, reduced COX-2 transcript and protein levels were observed after treatment with compounds 5d and 6d i.e., conjugates with morpholide OAO derivatives and 6c (KET-OAO benzyl ester).
Interestingly treatment with compound 6d, but also 5c (IBU-OAO benzyl ester) and 6c led to a significant reduction in the total (constitutive) protein levels of AKT and the case of 6c also its active phosphorylated form in HepG2. The modulation of the other elements of MAPK signaling pathways including AKT, ERK, p38, p70S6K, and JNK, was also observed. Notably, the levels of p38 a stress-activated protein kinase, were strongly diminished by all KET-OAO derivatives conjugates and 5d IBU-OAO morpholide conjugate.
The c-Jun N-terminal kinase (JNK), which is downstream of AKT and involved in regulating cell proliferation and apoptosis, protein level was also reduced upon treatment with IBU-OAO derivatives, particularly 5d, as well as KET-OAO conjugates 5a (IBU-OAO), 6b (KET-OAO benzyl ester), and 6d (KET-OAO morpholide).
Diminished AKT activity and related MAPK proteins can have important implications for HCC cells, as AKT signaling is often dysregulated in cancer and contributes to tumor growth and progression. Inhibiting AKT activity, similar to ERK, p38, p70S6K, and JNK, can potentially lead to reduced cell survival, decreased proliferation, and impaired tumor growth [38,39,40]. Interestingly, several studies have correlated the phosphorylation of ERK and Nrf2 activation [41,42,43]. Moreover, it was shown that the activation of MAPK–Akt and ERK is required for OA-induced activation of Nrf2, in primary rat vascular smooth muscle cells [44].
In this study, such correlation was found in the case of KET-OAO morpholide conjugate, which reduced to the most extent both pathways in HepG2 cells. Finally, the results of the current study revealed that morpholide conjugates particularly KET hybrid, affect the STAT3, STAT5 activation, and CREB indicating possible disruption of these signaling pathways associated with cell survival and proliferation [45,46,47].
Overall, the findings of this study demonstrate that IBU-OAO and KET-OAO derivatives conjugates, similarly to previously assessed NSAIDs, modulate the key signaling pathways involved in hepatic cancer development. However, although the KET-OAO morpholide seems to be the most potent modulator, their overall effects varied toward specific signaling pathways depending on the OAO derivatives structure. Therefore, selecting the optimal structure for the desired biological is needed.
Importantly, the conjugation of IBU and KET with OAO derivatives reduced their cytotoxicity, which made the conjugates good candidates for the prevention of not only liver cancer but also other diseases with inflammatory origin as safer alternatives to the parent compounds. Further in vivo studies are warranted to validate the efficacy and safety of these conjugates in liver cancer prophylaxis and treatment.
4. Materials and Methods
4.1. Chemistry
General information concerning the performed chemical experiments as well as concerning the elucidation of the synthesized compounds’ chemical structures is presented in our earlier publication [21]. The purity of the obtained oleanolic acid oxime conjugates with IBU (5a-d) and KET (6a-d) was evaluated based on spectra data (NMR) and TLC analysis.
The method involved mixing at room temperature a saturated solution of triterpene oxime (1.0 mmol) in dioxane, with the addition of dicyclohexylcarbodiimide (DCC, 1.5 mmol) and Ibuprofen or Ketoprofen (1.3 mmol). Then the resulting suspension was filtered and the filtrate was poured into about 5 times its volume of water. The resulting sticky product of the consistency of a lubricant was extracted with methylene chloride, the organic solution was washed with water, dried, and the solvent was distilled off. After purification of the product on a silica gel column chromatography, an almost colorless product, still of the consistency of a lubricant, was dissolved in ethanol and poured into water, but did not form a textured precipitate, but a sticky semi-solid. To obtain the product in the form of a solid, the ethanolic solution of the purified conjugate had to be poured into about 1% brine and left for several days at room temperature. After the product solidified, it was ground, filtered, and washed with water, then dried on air. The conjugates thus obtained were in the form of an amorphous solid with low melting points, except for the compound with Ketoprofen moiety and a morpholine ring (6d). This compound had the appearance of an amorphous solid, but at the same time, it was difficult to measure its melting point, because the substance melted slowly and in a fairly wide range (although it was chromatographically pure).
Conjugate of oleanolic acid oxime and Ibuprofen, 3-ibuprofenoxyiminoolean-12-en-28-oic acid (5a):
C43H63NO4. Mol. mass: 657.98. Yield: 580 mg (88.8 %). M.p.: 93 – 96 oC.
IR (ν, cm-1): 3340 (CH, [CH3]2CH-CH2-C6H4-CH[CH3]-COON=C-3), 1730 (C=O, [CH3]2CH-CH2-C6H4-CH[CH3]-COON=C-3), 1725 (N=C, [CH3]2CH-CH2-C6H4-CH[CH3]-COON=C-3), 1695 (C=O, -COOH).
13C NMR (δ, ppm): 183.55 (Cq, -COOH, C-28), 175.78 (Cq, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3), 172.43 (Cq, C-3), 143.24 (Cq, C-13), 140.42 (Cq, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-6′), 137.49 (Cq, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-3′), 137.37 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=, C-5′), 129.57 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-7′), 127.23 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-4′), 126.83 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-8′), 122.65 (CH, C-12), 46.72 (Cq, C-17), 44.90 (CH2, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-9′), 41.34 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-1′), 26.83 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-10′), 22.29 and 22.24 (CH3, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-11′ and C-12′), 14.96 (CH3, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3).
1H NMR (δ, ppm): 7.23 (2H, dd, J = 7.7 and 3.9 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C4′-H and C8′-H), 7.09 (2H, d, J = 7.0 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C5′-H and C7′-H), 5.28 (1H, t, J = 3.4 Hz, C12-H), 3.86 (1H, quart, J = 7.1 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C1′-H), 2.86 (1H, dd, J = 11.7 and 3.2 Hz, C18-Hβ), 2.44 (2H, d, J = 7.0 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C9′-H2), 2.14 (1H, tt, J = 2.5 and 12.1 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C10′-H), 1.54 (3H, d, J = 7.1 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C2′-H3), 1.25 (3H, d, J = 2.4 Hz) and 1.11 (3H, d, J = 3.5 Hz, ([CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C11′-H3, C12′-H3), 0.98, 0.96, 0.92, 0.90, 0.89 x 2, 0.87 (5 x 3H + 1 x 6H, 6 x s, 7 x CH3 groups).
Conjugate of methyl oleanolate oxime and Ibuprofen, 3-ibuprofenoxyiminoolean-12-en-28-oic acid methyl ester (5b):
C44H65NO4. Mol. mass: 672.00. Yield: 620 mg (92.2 %). M.p.: 68 – 73 oC.
IR (ν, cm-1): 3340 (CH, [CH3]2CH-CH2-C6H4-CH[CH3]-COON=C-3), 1735 (C=O, [CH3]2CH-CH2-C6H4-CH[CH3]-COON=C-3), 1725 (N=C, [CH3]2CH-CH2-C6H4-CH[CH3]-COON=C-3), 1720 (C=O, -COO-CH3).
13C NMR (δ, ppm): 178.25 (Cq, -COO-CH3, C-28), 175.81 (Cq, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3), 172.50 (Cq, C-3), 143.93 (Cq, C-13), 140.50 (Cq, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-6′), 137.55 (Cq, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-3′), 137.48 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=, C-5′), 129.27 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-7′), 127.30 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-4′), 127.28 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-8′), 122.04 (CH, C-12), 51.55 (CH3, -COO-CH3), 46.73 (Cq, C-17), 45.01 (CH2, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-9′), 41.41 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-1′), 26.95 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-10′), 22.35 x 2 (CH3, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-11′ and C-12′), 15.02 (CH3, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3).
1H NMR (δ, ppm): 7.24 (2H, dd, J = 7.6 and 3.9 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C4′-H and C8′-H), 7.10 (2H, d, J = 7.1 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C5′-H and C7′-H), 5.29 (1H, t, J = 3.4 Hz, C12-H), 3.85 (1H, quart, J = 7.0 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C1′-H), 3.63 (3H, s, -COO-CH3), 2.87 (1H, dd, J = 11.8 and 3.2 Hz, C18-Hβ), 2.45 (2H, d, J = 7.1 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C9′-H2), 2.16 (1H, tt, J = 2.5 and 12.0 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C10′-H), 1.55 (3H, d, J = 7.0 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C2′-H3), 1.24 (3H, d, J = 2.4 Hz) and 1.11 (3H, d, 3.5 Hz, ([CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C11′-H3, C12′-H3), 1.09, 0.98, 0.94, 0.91, 0.90, 0.89, 0.75 (7 x 3H, 7 x s, 7 x CH3 groups).
Conjugate of benzyl oleanolate oxime and Ibuprofen, 3-ibuprofenoxyiminoolean-12-en-28-oic acid benzyl ester (5c):
C50H69NO4. Mol. mass: 748.10. Yield: 650 mg (87.0 %). M.p.: 75 – 81 oC.
IR (ν, cm-1): 3340 (CH, [CH3]2CH-CH2-C6H4-CH[CH3]-COON=C-3 and -COO-CH2-C6H5), 1735 (C=O, [CH3]2CH-CH2-C6H4-CH[CH3]-COON=C-3), 1725 (N=C, [CH3]2CH-CH2-C6H4-CH[CH3]-COON=C-3), 1705 (C=O, -COO-CH2-C6H5).
13C NMR (δ, ppm): 177.39 (Cq, -COO-CH2-C6H5, C-28); 175.86 (Cq, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3), 172.47 (Cq, C-3), 143.85 (Cq, C-13), 140.48 (Cq, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-6′), 137.59 (Cq, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-3′), 137.49 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-5′), 136.42 (Cq, -COO-CH2-C6H5), 129.28(CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-7′), 128.42 (CH x 2, -COO-CH2-C6H5), 128.01 (CH x 2, -COO-CH2-C6H5), 127.93 (CH, -COO-CH2-C6H5), 127.31 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-4′), 127.30 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-8′), 122.18 (CH, C-12), 65.95 (CH2, -COO-CH2-C6H5), 46.76 (Cq, C-17), 45.03 (CH2, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-9′), 41.45 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-1′), 26.98 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-10′), 22.36 (CH3 x 2, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3), 15.03 (CH3, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3).
1H NMR (δ, ppm): 7.28 (1H, d, J = 3.6 Hz, -COO-CH2-C6H5), 7.23 (2H, dd, J = 7.6 and 3.9 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C4′-H and C8′-H), 7.16 (4H, dd, J = 8.2 and 7.1 Hz, -COO-CH2-C6H5), 7.12 – 7.06 (2H, m, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C5′-H and C7′-H), 5.33 (1H, t, J = 3.5 Hz, C12-H), 5.11 (2H, d, J = 12.6 Hz, -COO-CH2-C6H5), 3.85 (1H, quart, J = 7.0 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C1′-H), 2.95 (1H, dd, J = 4.0 and 13.7 Hz, C18-Hβ), 2.46 (2H, d, J = 7.0 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C9′-H2), 2.14 (1H, tt, J = 2.5 and 12.0 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C10′-H), 1.56 (3H, d, J = 7.0 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C2′-H3), 1.24 (3H, d, J = 2.4 Hz) and 1.12 (3H, d, 3.5 Hz, ([CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C11′-H3, C12′-H3), 0.97, 0.95, 0.92 x 3, 0.90, 0.64 (4 x 3H + 1 x 9H, 5 x s, 7 x CH3 groups).
Conjugate of morpholide of oleanolic acid oxime and Ibuprofen, 3-ibuprofenoxyiminoolean-12-en-28-oic acid morpholide (5d):
C47H70N2O4. Mol. mass: 727.08. Yield: 662 mg (91.1 %). M.p.: 100 – 105 oC.
IR (ν, cm-1): 3335 (CH, [CH3]2CH-CH2-C6H4-CH[CH3]-COON=C-3), 1730 (C=O, [CH3]2CH-CH2-C6H4 CH[CH3]-COON=C-3), 1720 (N=C, [CH3]2CH-CH2-C6H4-CH[CH3]-COON=C-3), 1630 (C=O, -C(O)Mor); 995 (C–N, -C(O)Mor); Mor = morpholine ring. 13C NMR (δ, ppm): 175.79 (Cq, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3), 175.07 (Cq, -C(O)Mor); 172.41 (Cq, C-3), 144.78 (Cq, C-13), 140.43 (Cq, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-6′), 137.49 (Cq, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-3′), 137.39 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-5′), 129.58 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-7′), 127.25 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-4′), 127.20 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-8′), 121.20 (CH, C-12), 66.89 x 2 (CH2 x 2, Mor), 47.35 (Cq, C-17), 46.05 and 41.60 (CH2 x 2, Mor), 44.91 (CH2, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-9′), 41.86 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-1′), 26.90 (CH, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C-10′), 22.29 and 22.22 (CH3 x 2, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3), 15.01 (CH3, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3); Mor = morpholine ring. 1H NMR (δ, ppm): 7.24 (2H, dd, J = 7.7 and 3.9 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C4′-H and C8′-H), 7.11 (2H, d, J = 7.1 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C5′-H and C7′-H), 5.28 (1H, t, J = 3.5 Hz, C12-H), 3.85 (1H, quart, J = 7.1 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C1′-H), 3.70 – 3.58 (8H, m, Morph), 3.10 (2H, d, J = 11.4 Hz, C18-Hβ), 2.45 (2H, d, J = 7.2 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C9′-H2), 2.15 (1H, tt, J = 2.5 and 12.0 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C10′-H), 1.56 (3H, d, J = 7.0 Hz, [CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C2′-H3), 1.24 (3H, d, J = 2.4 Hz) and 1.10 (3H, d, J = 3.5 Hz, ([CH3]2CH-CH2-C6H4-CH-[CH3]-COON=C-3, C11′-H3, C12′-H3), 1.33, 1.21, 1.19, 1.07, 1.00, 0.99, 0.92 (7 x 3H, 7 x s, 7 x CH3 groups); Mor = morpholine ring.
Conjugate of oleanolic acid oxime and Ketoprofen, 3-ketoprofenoxyiminoolean-12-en-28-oic acid (6a):
C46H59NO5. Mol. mass: 705.98. Yield: 659 mg (93.3 %). M.p.: 80 – 84 oC.
IR (ν, cm-1): 3335 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3), 1735 (C=O, C6H5-CO-C6H4-CH[CH3]-COON=C-3), 1725 (N=C, C6H5-CO-C6H4-CH[CH3]-COON=C-3), 1695 (C=O, -COOH).
13C NMR (δ, ppm): 196.50 (Cq, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-9′), 183.55 (Cq, -COOH, C-28), 175.94 (Cq, C6H5-CO-C6H4-CH[CH3]-COON=C-3), 171.84 (Cq, C-3), 143.24 (Cq, C-13), 140.66 (Cq, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-3′), 137.91 (Cq, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-5′), 137.48 (Cq, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-10′), 132.50 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-13′), 132.04 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-8′), 130.05 (CH x 2, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-11′ and C-15′), 129.36 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-7′), 129.13 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-4′), 128.65 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-6′), 128.31 (CH x 2, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-12 and C-14′), 122.64 (CH, C-12), 46.72 (Cq, C-17), 44.53 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-1′), 18.55 (CH3, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-2′)
1H NMR (δ, ppm): 7.80 (2H, dd, J = 3.5 and 1.8 Hz, C6H5-CO-C6H4-CH-[CH3]-COON=C3, C4′-H and C11′-H), 7.69 (1H, d, J = 7.7 Hz, C6H5-CO-C6H4-CH-[CH3]-COON=C3, C6′-H), 7.62 (2H, t, J = 6.1 Hz, C6H5-CO-C6H4-CH-[CH3]-COON=C3, C12′-H and C14′-H), 7.55 – 7.43 (3H, m, C6H5-CO-C6H4-CH-[CH3]-COON=C3, C13′-H, C7′-H and C8′-H), 5.28 (1H, t, J = 3.4 Hz, C12-H), 3.99 (1H, quart., J = 7.0 Hz, C6H5-CO-C6H4-CH-[CH3]-COON=C3), 2.85 (1H, dd, J = 11.7 and 3.2 Hz, C18-Hβ), 1.61 (3H, d, J = 7.1 Hz, C6H5-CO-C6H4-CH-[CH3]-COON=C3), 1.11, 1.00, 0.99, 0.95 x 2, 0.92, 0.76 (5 x 3H + 1 x 6H, 6 x s, 7 x CH3 group).
Conjugate of methyl oleanolate oxime and Ketoprofen, 3-ketoprofenoxyiminoolean-12-en-28-oic acid methyl ester (6b):
C47H61NO5. Mol. mass: 720.01. Yield: 667 mg (92.7 %). M.p.: 72 – 75 oC.
IR (ν, cm-1): 3330 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3), 1730 (C=O, C6H5-CO-C6H4-CH[CH3]-COON=C-3), 1725 (N=C, C6H5-CO-C6H4-CH[CH3]-COON=C-3), 1720 (C=O, -COO-CH3).
13C NMR (δ, ppm): 196.48 (Cq, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-9′), 178.26 (Cq, -COO-CH3, C-28), 175.94 (Cq, C6H5-CO-C6H4-CH[CH3]-COON=C-3), 171.85 (Cq, C-3), 143.94 (Cq, C-13), 140.69 (Cq, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-3′), 137.91 (Cq, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-5′), 137.50 (Cq, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-10′), 132.53 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-13′), 132.04 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-8′), 130.07 (CH x 2, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-11′ and C-15′), 129.38 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-7′), 129.03 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-4′), 128.55 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-6′), 128.34 (CH x 2, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-12 and C-14′), 122.04 (CH, C-12), 51.56 (CH3, COO-CH3), 46.10 (Cq, C-17), 44.50 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-1′), 18.54 (CH3, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-2′).
1H NMR (δ, ppm): 7.81 (2H, dd, J = 3.5 and 1.9 Hz, C6H5-CO-C6H4-CH-[CH3]-COON=C3, C4′-H and C11′-H), 7.69 (1H, d, J = 7.6 Hz, C6H5-CO-C6H4-CH-[CH3]-COON=C3, C6′-H), 7.61 (2H, t, J = 6.0 Hz, C6H5-CO-C6H4-CH-[CH3]-COON=C3, C12′-H and C14′-H), 7.55 – 7.42 (3H, m, C6H5-CO-C6H4-CH-[CH3]-COON=C3, C13′-H, C7′-H and C8′-H), 5.30 (1H, t, J = 3.4 Hz, C12-H), 3.98 (1H, quart., J = 7.0 Hz, C6H5-CO-C6H4-CH-[CH3]-COON=C3), 3.64 (3H, s, -COO-CH3), 2.88 (1H, dd, J = 13.6 and 3.7 Hz, C18-Hβ), 1.62 (3H, d, J = 7.2 Hz, C6H5-CO-C6H4-CH-[CH3]-COON=C3), 1.11, 1.00, 0.99, 0.95 x 2, 0.92, 0.76 (5 x 3H + 1 x 6H, 6 x s, 7 x CH3 group).
Conjugate of benzyl oleanolate oxime and Ketoprofen, 3-ketoprofenoxyiminoolean-12-en-28-oic acid benzyl ester (6c):
C43H65NO5. Mol. mass: 796.10. Yield: 672 mg (84.4 %). M.p.: 59 – 63 oC.
IR (ν, cm-1): 3335 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3), 2900 (CH, -COO-CH2-C6H5), 1740 (C=O, C6H5-CO-C6H4-CH[CH3]-COON=C-3), 1725 (N=C, C6H5-CO-C6H4-CH[CH3]-COON=C-3), 1705 (C=O, -COO-CH2-C6H5).
13C NMR (δ, ppm): 196.88 (Cq, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-9′), 177.41 (Cq, -COO-CH2-C6H5, C-28); 176.10 (Cq, C6H5-CO-C6H4-CH[CH3]-COON=C-3), 172.02 (Cq, C-3), 143.90 (Cq, C-13), 140.60 (Cq, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-3′), 137.57 (Cq, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-5′), 137.47 (Cq, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-10′), 136.40(Cq, -COO-CH2-C6H5), 132.64 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-13′), 132.08 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-8′), 130.17 (CH x 2, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-11′ and C-15′), 129.37 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-7′), 129.13 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-4′), 128.65 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-6′), 128.38 (CH x 2, -COO-CH2-C6H5), 128.09 (CH x 2, -COO-CH2-C6H5), 127.84 (CH, -COO-CH2-C6H5), 128.29 (CH x 2, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-12 and C-14′), 122.16 (CH, C-12), 65.89 (CH2, -COO-CH2-C6H5), 46.67 (Cq, C-17), 44.53 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-1′), 18.59 (CH3, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-2′).
1H NMR (δ, ppm): 7.80 (2H, dd, J = 3.5 and 1.9 Hz, C6H5-CO-C6H4-CH-[CH3]-COON=C3, C4′-H and C11′-H), 7.67 (1H, d, J = 7.6 Hz, C6H5-CO-C6H4-CH-[CH3]-COON=C3, C6′-H), 7.59 (2H, t, J = 6.0 Hz, C6H5-CO-C6H4-CH-[CH3]-COON=C3, C12′-H and C14′-H), 7.54 – 7.44 (3H, m, C6H5-CO-C6H4-CH-[CH3]-COON=C3, C13′-H, C7′-H and C8′-H), 7.30 (1H, d, J = 3.6 Hz, -COO-CH2-C6H5), 7.15 (4H, dd, J = 8.1 and 7.1 Hz, -COO-CH2-C6H5), 7.12 – 7.06 (2H, m, -COO-CH2-C6H5), 5.33 (1H, t, J = 3.4 Hz, C12-H), 5.10 (2H, d, J = 12.6 Hz, -COO-CH2-C6H5), 3.98 (1H, quart., J = 7.0 Hz, C6H5-CO-C6H4-CH-[CH3]-COON=C3), 2.93 (1H, dd, J = 4.0 and 13.7 Hz, C18-Hβ), 1.62 (3H, d, J = 7.2 Hz, C6H5-CO-C6H4-CH-[CH3]-COON=C3), 1.13, 1.11, 1.04, 0.99, 0.92, 0.91, 0.90 (7 x 3H, 7 x s, 7 CH3 groups).
Conjugate of morpholide of oleanolic acid oxime and Ketoprofen, 3-ketoprofenoxyiminoolean-12-en-28-oic acid morpholide (6d):
C50H66N2O5. Mol. mass: 775.08. Yield: 684 mg (88.2 %). M.p.: --- .
IR (ν, cm-1): 3330 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3), 1735 (C=O, C6H5-CO-C6H4-CH[CH3]-COON=C-3), 1720 (N=C, C6H5-CO-C6H4-CH[CH3]-COON=C-3), 1625 (C=O, -C(O)Mor); 995 (C–N, -C(O)Mor); Mor = morpholine ring.
13C NMR (δ, ppm): 196.36 (Cq, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-9′), 175.93 (Cq, C6H5-CO-C6H4-CH[CH3]-COON=C-3), 175.10 (Cq, -C(O)Mor); 171.81 (Cq, C-3), 144.72 (Cq, C-13), 140.48 (Cq, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-3′), 137.92 (Cq, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-5′), 137.29 (Cq, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-10′), 132.53 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-13′), 131.27 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-8′), 129.98 (CH x 2, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-11′ and C-15′), 129.32 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-7′), 129.01 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-4′), 128.70 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-6′), 128.27 (CH x 2, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-12 and C-14′), 121.18 (CH, C-12), 66.99, 66.87 (CH2 x 2, Mor), 47.33 (Cq, C-17), 46.22, 41.84 (CH2 x 2, Mor), 44.38 (CH, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-1′), 18.48 (CH3, C6H5-CO-C6H4-CH[CH3]-COON=C-3, C-2′); Mor = morpholine ring.
1H NMR (δ, ppm): 7.79 (2H, dd, J = 3.5 and 1.9 Hz, C6H5-CO-C6H4-CH-[CH3]-COON=C3, C4′-H and C11′-H), 7.67 (1H, d, J = 7.7 Hz, C6H5-CO-C6H4-CH-[CH3]-COON=C3, C6′-H), 7.59 (2H, t, J = 6.1 Hz, C6H5-CO-C6H4-CH-[CH3]-COON=C3, C12′-H and C14′-H), 7.54 – 7.46 (3H, m, C6H5-CO-C6H4-CH-[CH3]-COON=C3, C13′-H, C7′-H and C8′-H), 5.28 (1H, t, J = 3.5 Hz, C12-H), 4.04 (1H, quart., J = 7.0 Hz, C6H5-CO-C6H4-CH-[CH3]-COON=C3), 3.70 – 3.58 (8H, m, Morph), 3.08 (2H, d, J = 11.4 Hz, C18-Hβ), 1.60 (3H, d, J = 7.2 Hz, C6H5-CO-C6H4-CH-[CH3]-COON=C3), 1.08, 1.04, 0.98, 0.97, 0.93, 0.90, 0.75 (7 x 3H, 7 x s, 7 x CH3 groups); Mor = morpholine ring.
4.2. Biological assays
4.2.1. Cell culture and viability assay
HepG2 (ATCC HB 8065) and THLE-2 (ATCC CRL-2706) cells were provided by the American Type Culture Collection (ATCC, USA). HepG2 cells were maintained in Dulbecco’s Modified Eagle’s Medium (DMEM, Sigma-Aldrich, USA) containing 10% fetal bovine serum (FBS, EURx, Poland) and 1% antibiotics solution (Sigma-Aldrich, USA) while THLE-2 were cultured in BEGM supplemented with Bullet Kit (Lonza, Germany) and 10% FBS, 5ng/ml EGF, 70ng/ml phosphoethanolamine at 37°C, in a humidified, 5% CO2 atmosphere. To assess the effect of IBU, KET and their OAO conjugates on measured parameters, 1x106 cells were seeded per 100mm culture dish. After 24 hours of initial incubation, the cells were treated with 30µM concentrations of IBU, KET and their OAO conjugates and 0.1% dimethyl sulfoxide (DMSO) control solution. Incubation lasted for 24 hours, and the cells were harvested.
The effect of tested compounds on cell viability was assessed by the MTT assay, following the standard protocol. Briefly, THLE-2 and HepG2 cells were seeded (104 per well) in 96-well plates. After 24 hours of preincubation in a complete medium, compounds were added in various concentrations, and cells were incubated for 24 hours. Later, cells were washed twice with phosphate-buffered saline (PBS) and further incubated for 4 hours with a medium containing 0.5 mg/ml 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide (MTT). Then, the formazan crystals were dissolved in acidic isopropanol, and the absorbance was measured at 570 nm and 690 nm. All experiments were repeated three times. In all following experiments, we used non-toxic concentrations of compounds (with viability level above 70%): 30 μM of IBU, KET, and their OAO conjugates (5a-d and 6a-d).
4.2.2. Nuclear, cytosolic, and total protein lysates preparation
Nuclear and cytosolic fractions were prepared according to the manufacturer’s protocol, and the subcellular extracts from HepG2 and THLE-2 cells were prepared using the Nuclear/Cytosol Fractionation Kit (BioVision Research, USA). Lysates were prepared using Radioimmunoprecipitation assay (RIPA) buffer with the addition of protease inhibitors (Sigma-Aldrich, USA). Protein concentration was assessed, and the samples were stored at –80°C for future downstream applications.
4.2.3. Total RNA isolation, cDNA synthesis, and quantitative real-time PCR (R-T PCR)
The extraction of total RNA was performed by the GeneMatrix Universal DNA/RNA/Protein Purification Kit (EURx, Poland). Subsequently, samples were subjected to reverse transcription by the RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, USA), according to the manufacturer’s instructions.
For the quantitative R-T PCR analyses, the Maxima SYBR Green Kit (Fermentas Inc., USA) and the BioRad Chromo4 thermal cycler (BioRad Laboratories, USA) were used. Protocol started with 5 min enzyme activation at 95°C, followed by 40 cycles of 95°C for 15 s, 56°C for 20 s and 72°C for 40 s, and the final elongation at 72°C for 5 min. Melting curve analysis was used for amplicon verification. The expression of TBP (TATA box binding protein) and PBGD (porphobilinogen deaminase) was used to normalize data. The Pfaffl comparative method was used for fold-change quantification. Primers were designed using the Beacon Designer software and subjected to a BLAST search to minimize unspecific binding. Only the primer pairs that generated intron-spanning amplicons were selected. The primers’ sequences used to analyze Nrf2, NF-кBp65, NF-кBp50, COX-2, TBP, and PBGD genes are listed in Table 1.
4.2.4. Western blot analysis
Cytosolic extracts for COX-2, and β-actin, or nuclear extracts for Nrf2, NF-кB p65, NF-кB p50, and lamin protein detection, were separated on 12% or 10% SDS-PAGE slab gels. β-actin and lamin were used as loading control. Proteins were transferred to the nitrocellulose Immobilon P membrane. After blocking for 2 h with 10% skimmed milk, proteins were probed with primary antibodies against Nrf2, NF-кB p65, NF-кB p50, COX-2, β-actin, and lamin. Alkaline phosphatase AP-labeled anti-rabbit IgG, anti-goat IgG, and anti-mouse IgG secondary antibodies (BioRad Laboratories, USA) were used in the staining reaction. Bands were visualized by AP Conjugate Substrate Kit NBT/BCIP (BioRad Laboratories, USA). The amount of immunoreactive products in each lane was determined using ChemiDoc Imaging System (BioRad Laboratories, USA). Values were calculated as relative absorbance units (RQ) per mg of protein and expressed as a percentage of control.
4.2.5. Nrf2 and NF-ĸB binding assay
Nrf2, NF-κB p50, and NF-κB p65 activation were assessed by the enzymatic immunoassay (Transcription Factor ELISA Assay Kit Active Motif, Belgium) according to the manufacturer’s instructions. Activated Nrf2 was evaluated based on the amount of Nrf2 contained in the DNA-binding complex to the ARE sequence. The oligonucleotides containing the ARE consensus-binding site (5′-GTCACAGTGACTCAGCAGAATCTG-3′) for Nrf2 were immobilized on microplates as bait. Whereas activated NF-ĸB was measured as the amount of p65 and p50 subunits held in the DNA-binding complex. The oligonucleotides containing (5’-GGGACTTTCC-3’), a consensus site for NF-κB, were immobilized on microplates as bait. Nuclear fractions were incubated with oligonucleotides for 1 h, wells were washed, and DNA-bound subunits were detected by the specific primary antibody and a secondary antibody conjugated with the HRP. The results were expressed as the normalized level of absorbance (OD450 nm per mg of protein).
4.2.6. Bead-based immunoassay on the Luminex MAGPIX instrument
Magnetic bead-based immunoassay was performed on the Luminex-MAGPIX multiplex immunoassay system. Data were analyzed with Milliplex Analyst 5.1 software (EMD Millipore, USA). The panel we performed included quantitation of the following constitutive (total) and phosphorylated (active) proteins in the lysates of HepG2 cells, AKT, ERK, p38, p70S6K, JNK, STAT3, STAT5, and CREB according to the manufacturer’s instructions. The magnetic bead panel with high-sensitivity antibodies was obtained from Merck, Germany. A multiplex test based on microspheres using Luminex® xMAP® technology with different fluorescent colors was detected on a compatible MAGPIX® camera. Lysates were suspended in MILLIPLEX® MAP buffer. The bead suspension was added to each well of a 96-well plate, samples were added into the wells and incubated overnight at 2-8°C on a shaker protected from light. Control samples for HepG2 cells were lysates run as blank wells (containing all assay components minus protein); these were also added to the same plate and mixed with 25 μL of 1× bead mix. The plate was washed with 2X buffer, and then 1X MILLIPLEX® MAP Detection Antibodies were added. After shaking for 1 hour at room temperature, the antibodies were removed, and 1X MILLIPLEX® MAP Streptavidin/Phycoerythrin (SAPE) was added. Then the MILLIPLEX® MAP Amplification Buffer was added to each well and shaken for 15 minutes. The beads were suspended in MILLIPLEX® MAP buffer, and each microsphere was identified with a MAGPIX® Luminex Analyzer, and the results were calculated from the reporters’ fluorescent signals. Mean fluorescence intensities (MFI) were used to measure the relative total protein and relative phosphorylation levels of different targets and quantified with the xPonent 4.2 software (Luminex Corporation, USA). The raw MFI results for the tested protein levels were converted relative to the control of DMSO-treated cells.
4.3. Statistical Analysis
Statistical analysis and graphs were calculated and prepared using GraphPad Prism 9.2.0 (GraphPad Software, San Diego, CA, USA), assuming the significance level of changes as p < 0.05. Student’s t-test was used to assess the statistical significance between the experimental and control groups.
5. Conclusions
Our study highlighted the distinct effects of OAO conjugates with Ibuprofen and Ketoprofen on liver cancer-related signaling pathways, NF-κB, Nrf2, and MAPK. The IBU-OAO and KET-OAO morpholide conjugates (5d and 6d) emerged as the most active and effective hybrids. These conjugates present potential targets for novel drug design and may contribute to reducing invasiveness and chemoresistance in liver cancer cells. Further studies, including in vivo studies, are warranted to explore their application in liver cancer treatment.
Author Contributions
M.N., investigation, biological data collection and interpretation, formal analysis, and preparation of a draft of the manuscript; V.K.-K., experiment planning and designing, methodology, investigation, biological data collection, formal analysis, and data interpretation; B.B.-C., chemical synthesis, and structural data collection and interpretation; W.B.-D., supervision, funding acquisition, project administration, data interpretation, and final writing. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Polish National Science Centre, grant no. 2016/21/B/NZ7/01758.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statements
The data is contained within this article.
Acknowledgments
Graphical abstract and Figure 6 cartoon were created with BioRender.com
Conflicts of Interest
The authors declare no conflict of interest.
References
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikarsky, E.; Roayaie, S.; Lencioni, R.; Koike, K.; Zucman-Rossi, J.; Finn, R.S. Hepatocellular carcinoma. Nat. Rev. Dis. Prim. 2021 71 2021, 7, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Krajka-Kuźniak, V.; Baer-Dubowska, W. Modulation of Nrf2 and NF-κB Signaling Pathways by Naturally Occurring Compounds in Relation to Cancer Prevention and Therapy. Are Combinations Better Than Single Compounds? Int. J. Mol. Sci. 2021, Vol. 22, Page 8223 2021, 22, 8223. [Google Scholar] [CrossRef] [PubMed]
- Sorriento, D.; Campanile, A.; Santulli, G.; Leggiero, E.; Pastore, L.; Trimarco, B.; Iaccarino, G. A new synthetic protein, TAT-RH, inhibits tumor growth through the regulation of NFκB activity. Mol. Cancer 2009, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Malz, M.; Pinna, F.; Schirmacher, P.; Breuhahn, K. Transcriptional regulators in hepatocarcinogenesis – Key integrators of malignant transformation. J. Hepatol. 2012, 57, 186–195. [Google Scholar] [CrossRef]
- Colotta, F.; Allavena, P.; Sica, A.; Garlanda, C.; Mantovani, A. Cancer-related inflammation, the seventh hallmark of cancer: links to genetic instability. Carcinogenesis 2009, 30, 1073–1081. [Google Scholar] [CrossRef]
- Hoesel, B.; Schmid, J.A. The complexity of NF-κB signaling in inflammation and cancer. Mol. Cancer 2013 121 2013, 12, 1–15. [Google Scholar] [CrossRef]
- Ahmed, S.M.U.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta - Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef]
- Raghunath, A.; Sundarraj, K.; Arfuso, F.; Sethi, G.; Perumal, E. Dysregulation of Nrf2 in Hepatocellular Carcinoma: Role in Cancer Progression and Chemoresistance. Cancers (Basel). 2018, 10. [Google Scholar] [CrossRef]
- Moon, H.; Ro, S.W. MAPK/ERK Signaling Pathway in Hepatocellular Carcinoma. Cancers (Basel). 2021, 13. [Google Scholar] [CrossRef]
- Dimri, M.; Satyanarayana, A. Molecular Signaling Pathways and Therapeutic Targets in Hepatocellular Carcinoma. Cancers (Basel). 2020, 12. [Google Scholar] [CrossRef]
- Cuzick, J.; Thorat, M.A.; Bosetti, C.; Brown, P.H.; Burn, J.; Cook, N.R.; Ford, L.G.; Jacobs, E.J.; Jankowski, J.A.; La Vecchia, C.; et al. Estimates of benefits and harms of prophylactic use of aspirin in the general population. Ann. Oncol. 2015, 26, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, P.M.; Fowkes, F.G.R.; Belch, J.F.; Ogawa, H.; Warlow, C.P.; Meade, T.W. Effect of daily aspirin on long-term risk of death due to cancer: Analysis of individual patient data from randomised trials. Lancet 2011, 377, 31–41. [Google Scholar] [CrossRef]
- Cuzick, J.; Otto, F.; Baron, J.A.; Brown, P.H.; Burn, J.; Greenwald, P.; Jankowski, J.; La Vecchia, C.; Meyskens, F.; Senn, H.J.; et al. Aspirin and non-steroidal anti-inflammatory drugs for cancer prevention: an international consensus statement. Lancet Oncol. 2009, 10, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Zappavigna, S.; Cossu, A.M.; Grimaldi, A.; Bocchetti, M.; Ferraro, G.A.; Nicoletti, G.F.; Filosa, R.; Caraglia, M. Anti-Inflammatory Drugs as Anticancer Agents. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Bindu, S.; Mazumder, S.; Bandyopadhyay, U. Non-steroidal anti-inflammatory drugs (NSAIDs) and organ damage: A current perspective. Biochem. Pharmacol. 2020, 180, 114147. [Google Scholar] [CrossRef] [PubMed]
- Ayeleso, T.B.; Matumba, M.G.; Mukwevho, E. Oleanolic Acid and Its Derivatives: Biological Activities and Therapeutic Potential in Chronic Diseases. Mol. 2017, Vol. 22, Page 1915 2017, 22, 1915. [Google Scholar] [CrossRef] [PubMed]
- Parikh, N.R.; Mandal, A.; Bhatia, D.; Siveen, K.S.; Sethi, G.; Bishayee, A. Oleanane triterpenoids in the prevention and therapy of breast cancer: current evidence and future perspectives. Phytochem. Rev. 2014, 13, 793–810. [Google Scholar] [CrossRef]
- Žiberna, L.; Šamec, D.; Mocan, A.; Nabavi, S.F.; Bishayee, A.; Farooqi, A.A.; Sureda, A.; Nabavi, S.M. Oleanolic Acid Alters Multiple Cell Signaling Pathways: Implication in Cancer Prevention and Therapy. Int. J. Mol. Sci. 2017, Vol. 18, Page 643 2017, 18, 643. [Google Scholar] [CrossRef]
- Bishayee, A.; Mandal, A.; Thoppil, R.J.; Darvesh, A.S.; Bhatia, D. Chemopreventive effect of a novel oleanane triterpenoid in a chemically induced rodent model of breast cancer. Int. J. Cancer 2013, 133, 1054–1063. [Google Scholar] [CrossRef]
- Schepetkin, I.A.; Plotnikov, M.B.; Khlebnikov, A.I.; Plotnikova, T.M.; Quinn, M.T. Oximes: Novel Therapeutics with Anticancer and Anti-Inflammatory Potential. Biomolecules 2021, 11. [Google Scholar] [CrossRef]
- Krajka-Kuźniak, V.; Bednarczyk-Cwynar, B.; Paluszczak, J.; Szaefer, H.; Narożna, M.; Zaprutko, L.; Baer-Dubowska, W. Oleanolic acid oxime derivatives and their conjugates with aspirin modulate the NF-κB-mediated transcription in HepG2 hepatoma cells. Bioorg. Chem. 2019, 93, 103326. [Google Scholar] [CrossRef] [PubMed]
- Narożna, M.; Krajka-Kuźniak, V.; Kleszcz, R.; Bednarczyk-Cwynar, B.; Szaefer, H.; Baer-Dubowska, W. Activation of the Nrf2 response by oleanolic acid oxime morpholide (3-hydroxyiminoolean-12-en-28-oic acid morpholide) is associated with its ability to induce apoptosis and inhibit proliferation in HepG2 hepatoma cells. Eur. J. Pharmacol. 2020, 883, 173307. [Google Scholar] [CrossRef]
- Narożna, M.; Krajka-Kuźniak, V.; Bednarczyk-Cwynar, B.; Kucińska, M.; Kleszcz, R.; Kujawski, J.; Piotrowska-Kempisty, H.; Plewiński, A.; Murias, M.; Baer-Dubowska, W. Conjugation of diclofenac with novel oleanolic acid derivatives modulate nrf2 and nf-κb activity in hepatic cancer cells and normal hepatocytes leading to enhancement of its therapeutic and chemopreventive potential. Pharmaceuticals 2021, 14, 688. [Google Scholar] [CrossRef] [PubMed]
- Narożna, M.; Krajka-Kuźniak, V.; Bednarczyk-Cwynar, B.; Kleszcz, R.; Baer-Dubowska, W. The Effect of Novel Oleanolic Acid Oximes Conjugated with Indomethacin on the Nrf2-ARE And NF-κB Signaling Pathways in Normal Hepatocytes and Human Hepatocellular Cancer Cells. Pharm. 2021, Vol. 14, Page 32 2020, 14, 32. [Google Scholar] [CrossRef] [PubMed]
- Gunaydin, C.; Bilge, S.S. Effects of Nonsteroidal Anti-Inflammatory Drugs at the Molecular Level. Eurasian J. Med. 2018, 50, 116–121. [Google Scholar] [CrossRef]
- Baek, S.J.; Eling, T.; Kolawole, O.R.; Kashfi, K. NSAIDs and Cancer Resolution: New Paradigms beyond Cyclooxygenase. Int. J. Mol. Sci. 2022, Vol. 23, Page 1432 2022, 23, 1432. [Google Scholar] [CrossRef]
- Bittoni, M.A.; Carbone, D.P.; Harris, R.E. Ibuprofen and fatal lung cancer: A brief report of the prospective results from the Third National Health and Nutrition Examination Survey (NHANES III). Mol. Clin. Oncol. 2017, 6, 917. [Google Scholar] [CrossRef]
- Moore, B.; Simmons, D. COX-2 Inhibition, Apoptosis, and Chemoprevention by Nonsteroidal Anti-inflammatory Drugs. Curr. Med. Chem. 2012, 7, 1131–1144. [Google Scholar] [CrossRef]
- Sahasrabuddhe, V. V.; Gunja, M.Z.; Graubard, B.I.; Trabert, B.; Schwartz, L.M.; Park, Y.; Hollenbeck, A.R.; Freedman, N.D.; Mcglynn, K.A. Nonsteroidal Anti-inflammatory Drug Use, Chronic Liver Disease, and Hepatocellular Carcinoma. JNCI J. Natl. Cancer Inst. 2012, 104, 1808–1814. [Google Scholar] [CrossRef]
- Raghunath, A.; Sundarraj, K.; Arfuso, F.; Sethi, G.; Perumal, E. Dysregulation of Nrf2 in Hepatocellular Carcinoma: Role in Cancer Progression and Chemoresistance. Cancers 2018, Vol. 10, Page 481 2018, 10, 481. [Google Scholar] [CrossRef]
- Gao, X.; Guo, N.; Xu, H.; Pan, T.; Lei, H.; Yan, A.; Mi, Y.; Xu, L. Ibuprofen induces ferroptosis of glioblastoma cells via downregulation of nuclear factor erythroid 2-related factor 2 signaling pathway. Anticancer. Drugs 2020, 31, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wan, Y.; Huang, C. The Biological Functions of NF-κB1 (p) and its Potential as an Anti-Cancer Target. Curr. Cancer Drug Targets 2009, 9, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Giridharan, S.; Srinivasan, M. Mechanisms of NF-κB p65 and strategies for therapeutic manipulation. J. Inflamm. Res. 2018, 11, 407–419. [Google Scholar] [CrossRef]
- Younus, H. Therapeutic potentials of superoxide dismutase. Int. J. Health Sci. (Qassim). 2018, 12, 88. [Google Scholar]
- Christian, F.; Smith, E.L.; Carmody, R.J. The Regulation of NF-κB Subunits by Phosphorylation. Cells 2016, Vol. 5, Page 12 2016, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Cai, W.; Chu, E.S.H.; Tang, J.; Wong, C.C.; Wong, S.H.; Sun, W.; Liang, Q.; Fang, J.; Sun, Z.; et al. Hepatic cyclooxygenase-2 overexpression induced spontaneous hepatocellular carcinoma formation in mice. Oncogene 2017 3631 2017, 36, 4415–4426. [Google Scholar] [CrossRef] [PubMed]
- Cervello, M.; Montalto, G. Cyclooxygenases in hepatocellular carcinoma. World J. Gastroenterol. 2006, 12, 5113. [Google Scholar] [CrossRef]
- Lu, M.; Wang, Y.; Zhan, X. The MAPK pathway-based drug therapeutic targets in pituitary adenomas. Front. Endocrinol. (Lausanne). 2019, 10, 446999. [Google Scholar] [CrossRef]
- Papa, S.; Choy, P.M.; Bubici, C. The ERK and JNK pathways in the regulation of metabolic reprogramming. Oncogene 2018 3813 2018, 38, 2223–2240. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, X.; Cao, S.; Sun, Y.; He, X.; Jiang, B.; Yu, Y.; Duan, J.; Qiu, F.; Kang, N. Berberine represses human gastric cancer cell growth in vitro and in vivo by inducing cytostatic autophagy via inhibition of MAPK/mTOR/p70S6K and Akt signaling pathways. Biomed. Pharmacother. 2020, 128, 110245. [Google Scholar] [CrossRef]
- Kim, J.K.; Jang, H.D. Nrf2-Mediated HO-1 Induction Coupled with the ERK Signaling Pathway Contributes to Indirect Antioxidant Capacity of Caffeic Acid Phenethyl Ester in HepG2 Cells. Int. J. Mol. Sci. 2014, 15, 12149. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Huang, Z.; Zhang, D.D. Phosphorylation of Nrf2 at Multiple Sites by MAP Kinases Has a Limited Contribution in Modulating the Nrf2-Dependent Antioxidant Response. PLoS One 2009, 4, e6588. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.Y.; Pyo, M.C.; Nam, M.H.; Lee, K.W. ERK/Nrf2 pathway activation by caffeic acid in HepG2 cells alleviates its hepatocellular damage caused by t-butylhydroperoxide-induced oxidative stress. BMC Complement. Altern. Med. 2019, 19, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Zhang, P.; Chen, X.; He, G. PI3K and ERK/Nrf2 pathways are involved in oleanolic acid-induced heme oxygenase-1 expression in rat vascular smooth muscle cells. J. Cell. Biochem. 2011, 112, 1524–1531. [Google Scholar] [CrossRef]
- Yang, S.; Jiang, W.; Yang, W.; Yang, C.; Yang, X.; Chen, K.; Hu, Y.; Shen, G.; Lu, L.; Cheng, F.; et al. Epigenetically modulated miR-1224 suppresses the proliferation of HCC through CREB-mediated activation of YAP signaling pathway. Mol. Ther. - Nucleic Acids 2021, 23, 944–958. [Google Scholar] [CrossRef]
- Dimri, M.; Satyanarayana, A. Molecular Signaling Pathways and Therapeutic Targets in Hepatocellular Carcinoma. Cancers 2020, Vol. 12, Page 491 2020, 12, 491. [Google Scholar] [CrossRef]
- Wingelhofer, B.; Neubauer, H.A.; Valent, P.; Han, X.; Constantinescu, S.N.; Gunning, P.T.; Müller, M.; Moriggl, R. Implications of STAT3 and STAT5 signaling on gene regulation and chromatin remodeling in hematopoietic cancer. Leuk. 2018 328 2018, 32, 1713–1726. [Google Scholar] [CrossRef]
Figure 1.
The brief scheme of synthesis and chemical structures of oleanolic acid oximes (2a–d) and the investigated conjugates with Ibuprofen (IBU-OAO) (5a–d) and Ketoprofen (KET-OAO) (6a-d).
Figure 1.
The brief scheme of synthesis and chemical structures of oleanolic acid oximes (2a–d) and the investigated conjugates with Ibuprofen (IBU-OAO) (5a–d) and Ketoprofen (KET-OAO) (6a-d).
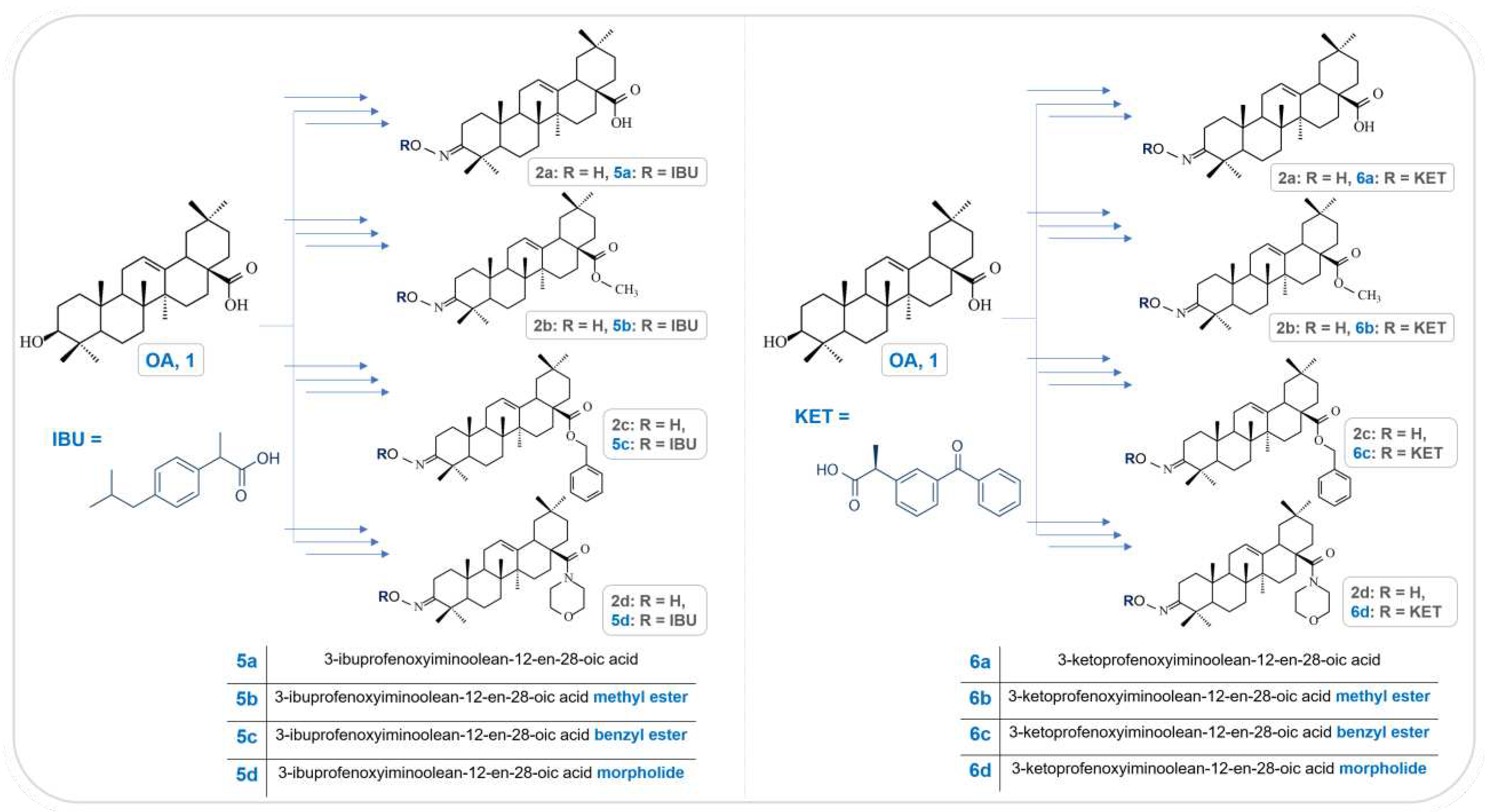
Figure 2.
The effect of the IBU, KET, IBU–OAO conjugates (5a–d) and KET–OAO conjugates (6a–d) on the viability of THLE-2 (A, B) and HepG2 (C,D) cells (after 24 h incubation). The red dashed line on the graphs represents the IC50 level (half maximal inhibitory concentration), the concentration at which the compound is able to reduce the viability of cells by half. The blue dashed line in the graphs represents the concentration of compounds that ensures approximately 70% cell viability. Data (mean ± SEM) from three separate experiments are presented.
Figure 2.
The effect of the IBU, KET, IBU–OAO conjugates (5a–d) and KET–OAO conjugates (6a–d) on the viability of THLE-2 (A, B) and HepG2 (C,D) cells (after 24 h incubation). The red dashed line on the graphs represents the IC50 level (half maximal inhibitory concentration), the concentration at which the compound is able to reduce the viability of cells by half. The blue dashed line in the graphs represents the concentration of compounds that ensures approximately 70% cell viability. Data (mean ± SEM) from three separate experiments are presented.
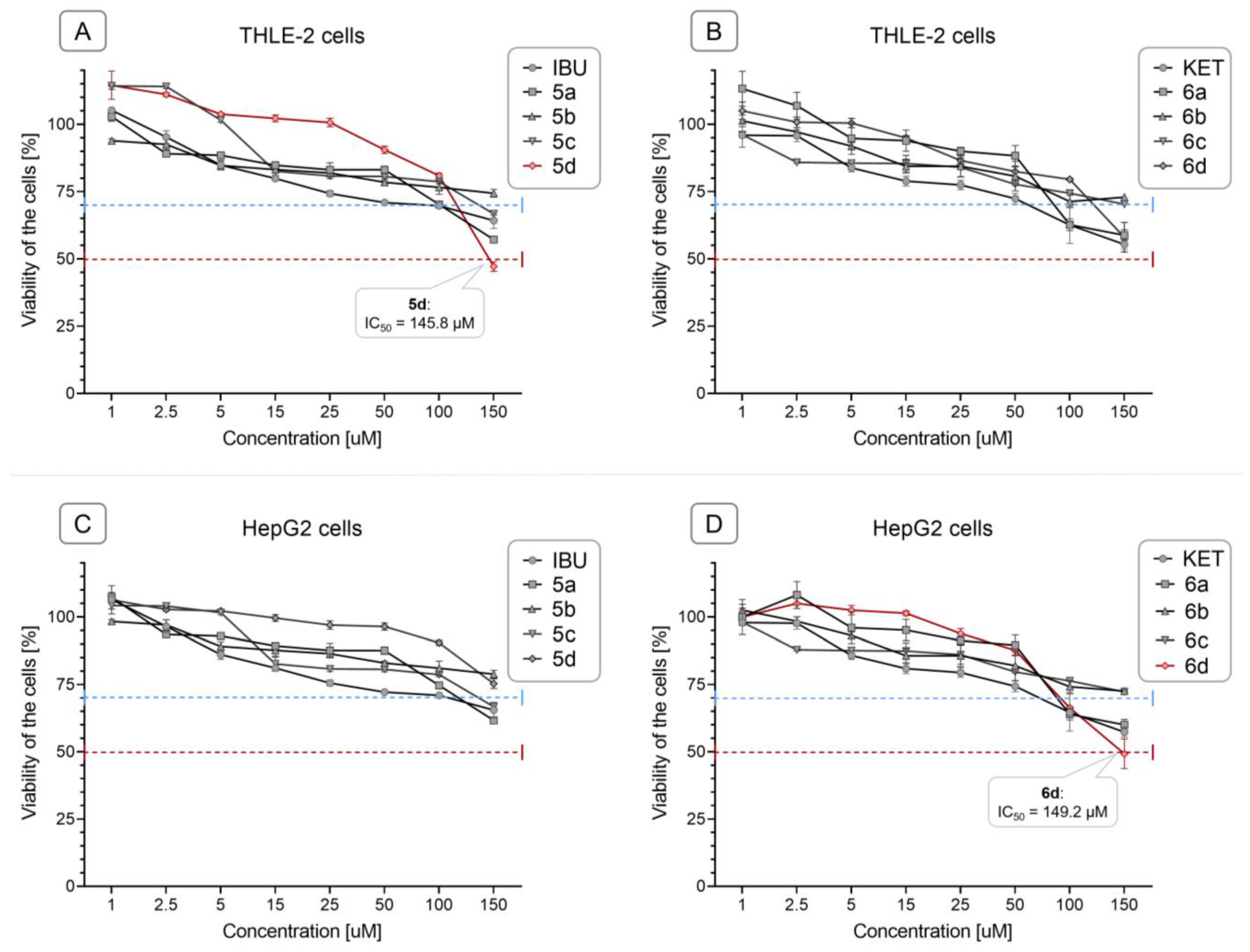
Figure 3.
The effect of the IBU, KET, IBU–OAO conjugates (5a–d) and KET–OAO conjugates (6a–d) at concentrations of 30 µM on Nrf2 activation and expression in THLE-2 cells (A–C) and HepG2 cells (D-F) after 24 hours treatment. Panels (A,D) present nuclear protein levels of Nrf2, panels (B,E) Nrf2 binding to DNA, and panels (C,F) mRNA transcript panels. Representative Western immunoblots are presented above the graphs. The sequence of the bands corresponds to the sequence of the bars in the graph. Lamin A/C, a major component of the nuclear lamina, was used for normalizing nuclear protein levels in Western blot analysis. The values (means ± SEM) were calculated as protein levels compared to control cells (expression equal to 100%). Activated Nrf2 (B,E) was assessed in terms of the amount of Nrf2 contained in the DNA-binding complexes extracted from the nuclei isolated from the cells and calculated compared to control cells set to 100%. The values of mRNA level (C,F) were calculated as the relative change in comparison to control cells (expression equal to 1). * Significantly different from control: DMSO-treated cells, p < 0.05. Student’s t-test was used to assess the statistical significance.
Figure 3.
The effect of the IBU, KET, IBU–OAO conjugates (5a–d) and KET–OAO conjugates (6a–d) at concentrations of 30 µM on Nrf2 activation and expression in THLE-2 cells (A–C) and HepG2 cells (D-F) after 24 hours treatment. Panels (A,D) present nuclear protein levels of Nrf2, panels (B,E) Nrf2 binding to DNA, and panels (C,F) mRNA transcript panels. Representative Western immunoblots are presented above the graphs. The sequence of the bands corresponds to the sequence of the bars in the graph. Lamin A/C, a major component of the nuclear lamina, was used for normalizing nuclear protein levels in Western blot analysis. The values (means ± SEM) were calculated as protein levels compared to control cells (expression equal to 100%). Activated Nrf2 (B,E) was assessed in terms of the amount of Nrf2 contained in the DNA-binding complexes extracted from the nuclei isolated from the cells and calculated compared to control cells set to 100%. The values of mRNA level (C,F) were calculated as the relative change in comparison to control cells (expression equal to 1). * Significantly different from control: DMSO-treated cells, p < 0.05. Student’s t-test was used to assess the statistical significance.
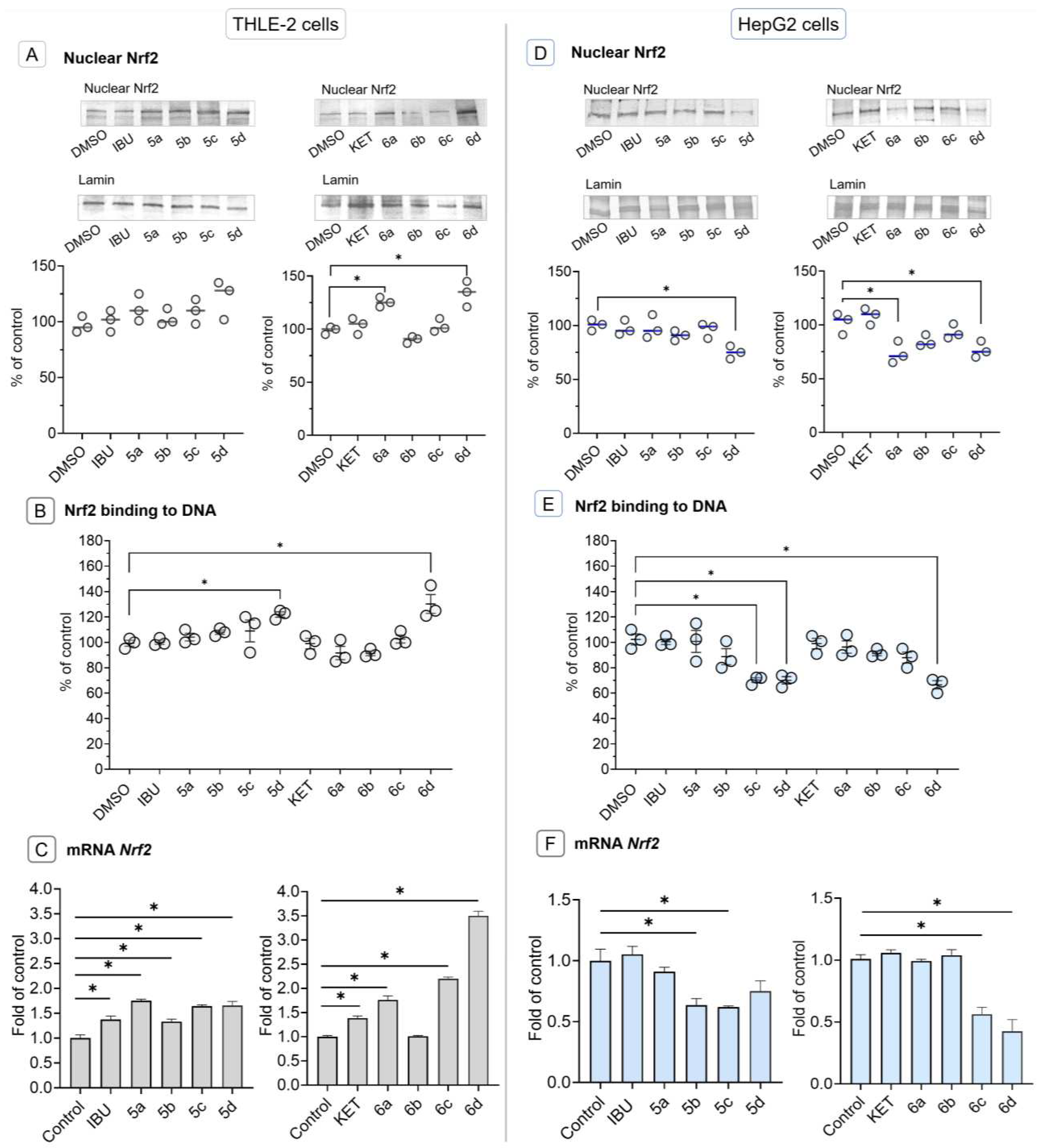
Figure 4.
The effect of the IBU, KET, IBU–OAO conjugates (5a–d) and KET–OAO conjugates (6a–d) at concentrations of 30 µM on NF-κB activation and expression in HepG2 cells after 24 hours treatment. Panels (A,D) show NF-κB p50 and p65 subunits nuclear protein levels, binding to DNA (B,E) and mRNA transcripts panels (C,F). Panels (G,H) show the total cytosolic NF-κB and the total phosphorylated NF-κB protein levels. Representative Western immunoblots are presented above the graphs. The sequence of the bands corresponds to the sequence of the bars in the graph. Lamin A/C was used for normalizing nuclear protein levels in Western blot analysis. The values (means ± SEM) were calculated as protein levels compared to control cells (expression equal to 100%). Activated NF-κB p50 and p65 (B,E) were assessed in terms of the amount of NF-κB subunits contained in the DNA-binding complexes extracted from the nuclei isolated from the cells and calculated compared to control cells set to 100%. The values of mRNA level (C,F) were calculated as the relative change in comparison to control cells (expression equal to 1). The cytosolic NF-κB and phospho-NF-κB (G,H) were assessed using the MAGPIX® system, and the results from five independent measurements are presented as a fold of fluorescence intensity obtained in control cells. * Significantly different from control: DMSO-treated cells, p < 0.05. Student’s t-test was used to assess the statistical significance.
Figure 4.
The effect of the IBU, KET, IBU–OAO conjugates (5a–d) and KET–OAO conjugates (6a–d) at concentrations of 30 µM on NF-κB activation and expression in HepG2 cells after 24 hours treatment. Panels (A,D) show NF-κB p50 and p65 subunits nuclear protein levels, binding to DNA (B,E) and mRNA transcripts panels (C,F). Panels (G,H) show the total cytosolic NF-κB and the total phosphorylated NF-κB protein levels. Representative Western immunoblots are presented above the graphs. The sequence of the bands corresponds to the sequence of the bars in the graph. Lamin A/C was used for normalizing nuclear protein levels in Western blot analysis. The values (means ± SEM) were calculated as protein levels compared to control cells (expression equal to 100%). Activated NF-κB p50 and p65 (B,E) were assessed in terms of the amount of NF-κB subunits contained in the DNA-binding complexes extracted from the nuclei isolated from the cells and calculated compared to control cells set to 100%. The values of mRNA level (C,F) were calculated as the relative change in comparison to control cells (expression equal to 1). The cytosolic NF-κB and phospho-NF-κB (G,H) were assessed using the MAGPIX® system, and the results from five independent measurements are presented as a fold of fluorescence intensity obtained in control cells. * Significantly different from control: DMSO-treated cells, p < 0.05. Student’s t-test was used to assess the statistical significance.
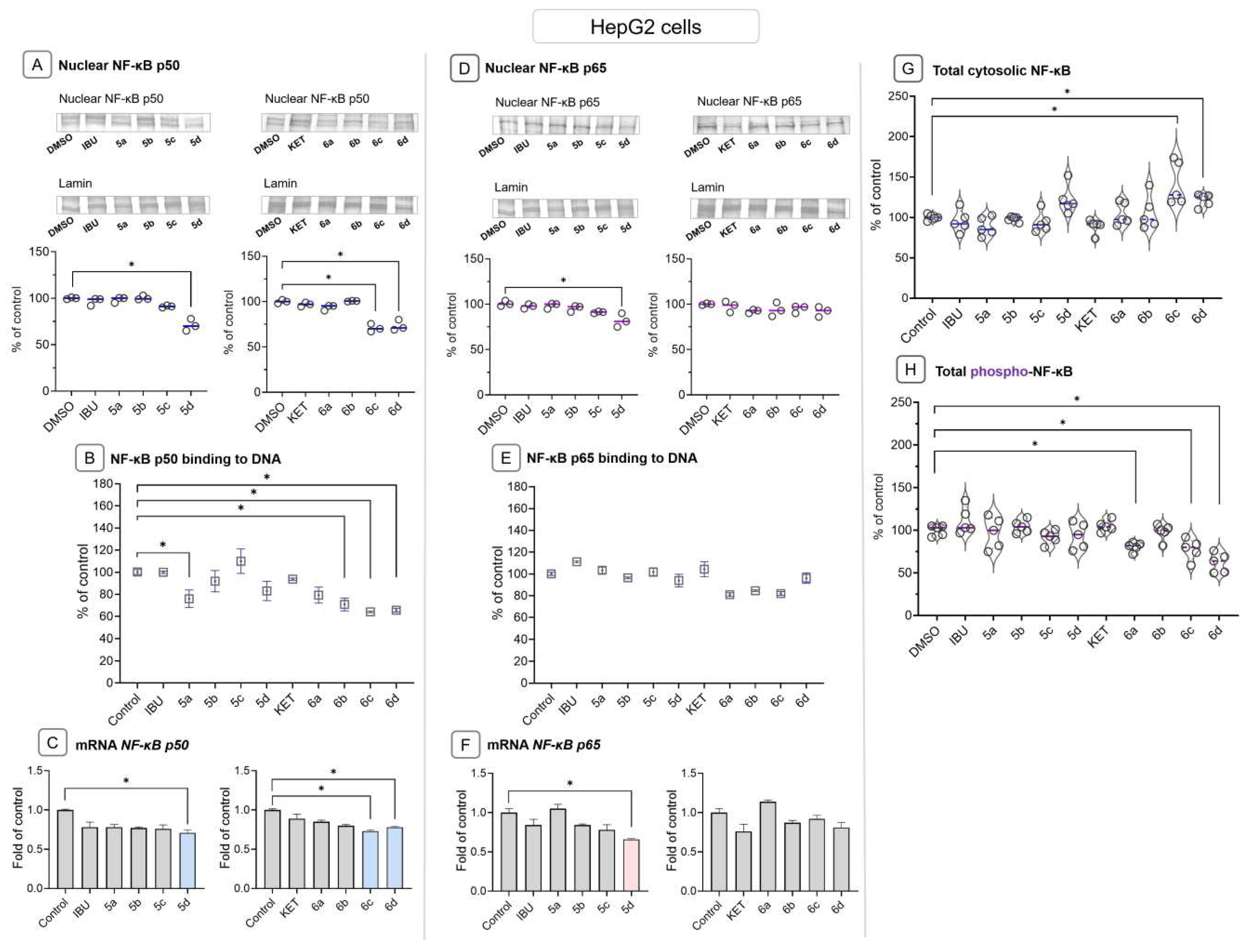
Figure 5.
The effect of the IBU, KET, IBU–OAO conjugates (5a–d) and KET–OAO conjugates (6a–d) at concentrations of 30 µM on the COX-2 protein (A) and COX-2 mRNA levels (B) in HepG2 cells after 24 hours treatment. Representative immunoblots of the cytosolic content of COX-2 in HepG2 cells (A) from three separate experiments are shown. The sequence of the bands corresponds to the sequence of the bars in the graph. β-actin was used as a loading control. The values were calculated as protein levels in comparison to control cells set to 100%. The values (mean ± SEM) for the mRNA levels in HepG2 cells (B) were calculated from three separate experiments in comparison to control cells set to 1. * Significantly different from control: DMSO-treated cells, p < 0.05. Student’s t-test was used to assess the statistical significance.
Figure 5.
The effect of the IBU, KET, IBU–OAO conjugates (5a–d) and KET–OAO conjugates (6a–d) at concentrations of 30 µM on the COX-2 protein (A) and COX-2 mRNA levels (B) in HepG2 cells after 24 hours treatment. Representative immunoblots of the cytosolic content of COX-2 in HepG2 cells (A) from three separate experiments are shown. The sequence of the bands corresponds to the sequence of the bars in the graph. β-actin was used as a loading control. The values were calculated as protein levels in comparison to control cells set to 100%. The values (mean ± SEM) for the mRNA levels in HepG2 cells (B) were calculated from three separate experiments in comparison to control cells set to 1. * Significantly different from control: DMSO-treated cells, p < 0.05. Student’s t-test was used to assess the statistical significance.
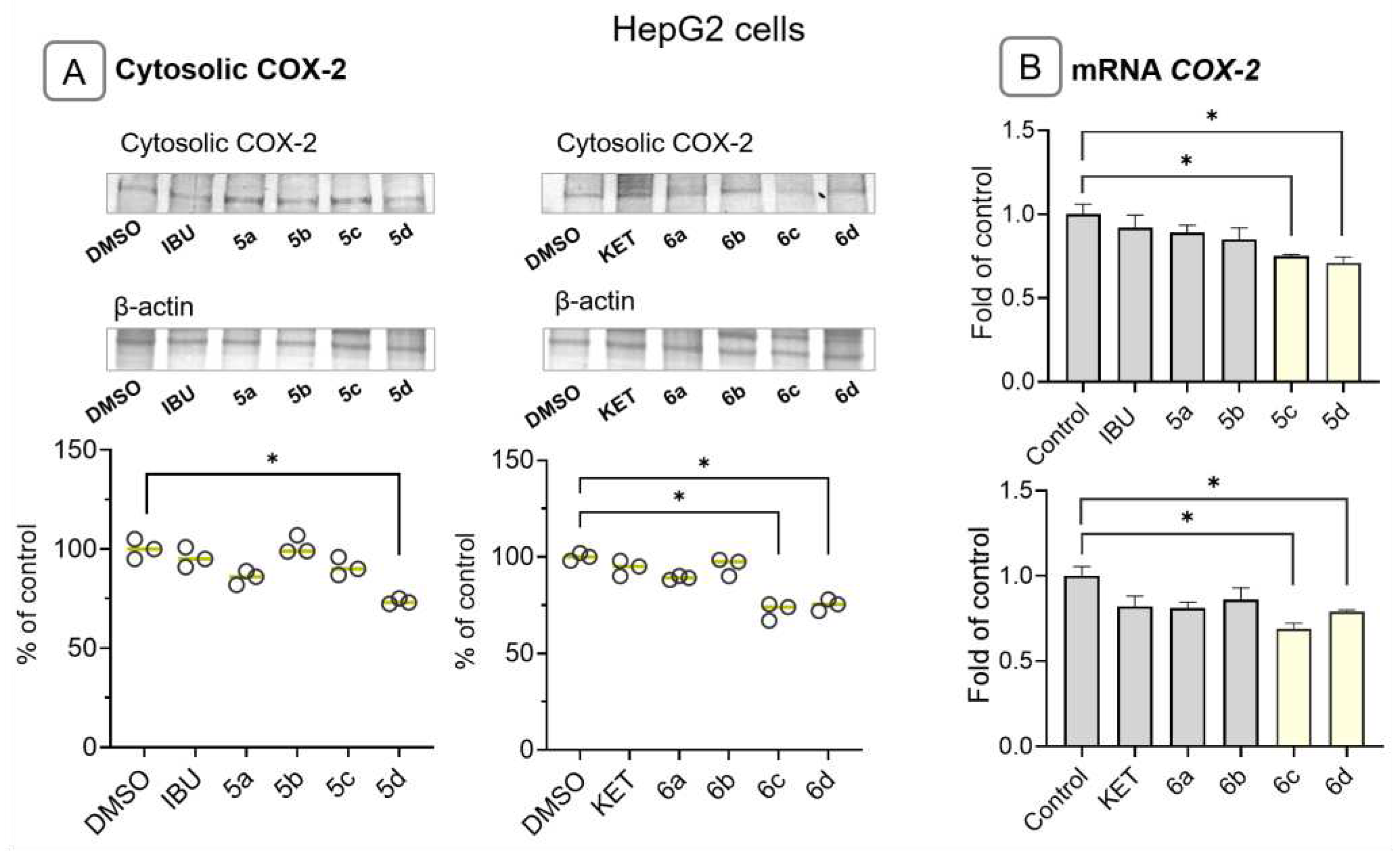
Figure 6.
The effect of the IBU, KET, IBU–OAO conjugates (5a–d) and KET–OAO conjugates (6a–d) on the total (constitutive) and phosphorylated (active) levels of AKT (A), ERK (B), p38 (C), p70S6K (D) and JNK (E), measured and calculated on the MAGPIX® system in HepG2 cells, after 24 h incubation with tested compounds at concentrations of 30 µM. The mean values (mean± SEM) of fluorescence intensity are presented as the fold of control from five independent measurements. * Significantly different from control: DMSO-treated cells, p < 0.05. Student’s t-test was used to assess the statistical significance.
Figure 6.
The effect of the IBU, KET, IBU–OAO conjugates (5a–d) and KET–OAO conjugates (6a–d) on the total (constitutive) and phosphorylated (active) levels of AKT (A), ERK (B), p38 (C), p70S6K (D) and JNK (E), measured and calculated on the MAGPIX® system in HepG2 cells, after 24 h incubation with tested compounds at concentrations of 30 µM. The mean values (mean± SEM) of fluorescence intensity are presented as the fold of control from five independent measurements. * Significantly different from control: DMSO-treated cells, p < 0.05. Student’s t-test was used to assess the statistical significance.
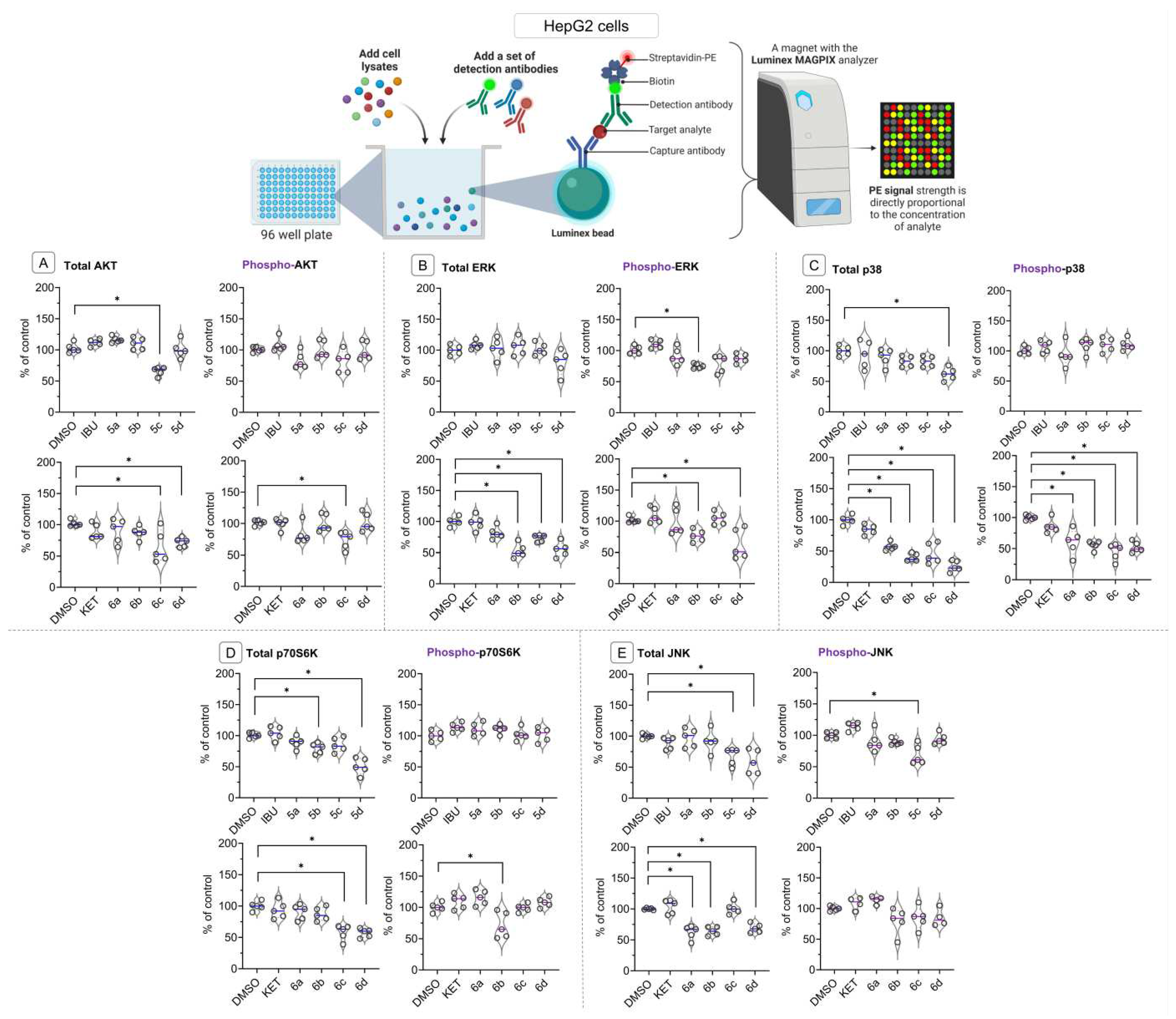
Figure 7.
The effect of the IBU, KET, IBU–OAO conjugates (5a–d) and KET–OAO conjugates (6a–d) on the total (constitutive) and phosphorylated (active) levels of STAT3 (A), STAT5 (B) and CREB (C) measured and calculated on the MAGPIX® system in HepG2 cells, after 24 h incubation with tested compounds at concentrations of 30 µM. The mean values (mean± SEM) of fluorescence intensity are presented as the fold of control from five independent measurements. * Significantly different from control: DMSO-treated cells, p < 0.05. Student’s t-test was used to assess the statistical significance.
Figure 7.
The effect of the IBU, KET, IBU–OAO conjugates (5a–d) and KET–OAO conjugates (6a–d) on the total (constitutive) and phosphorylated (active) levels of STAT3 (A), STAT5 (B) and CREB (C) measured and calculated on the MAGPIX® system in HepG2 cells, after 24 h incubation with tested compounds at concentrations of 30 µM. The mean values (mean± SEM) of fluorescence intensity are presented as the fold of control from five independent measurements. * Significantly different from control: DMSO-treated cells, p < 0.05. Student’s t-test was used to assess the statistical significance.
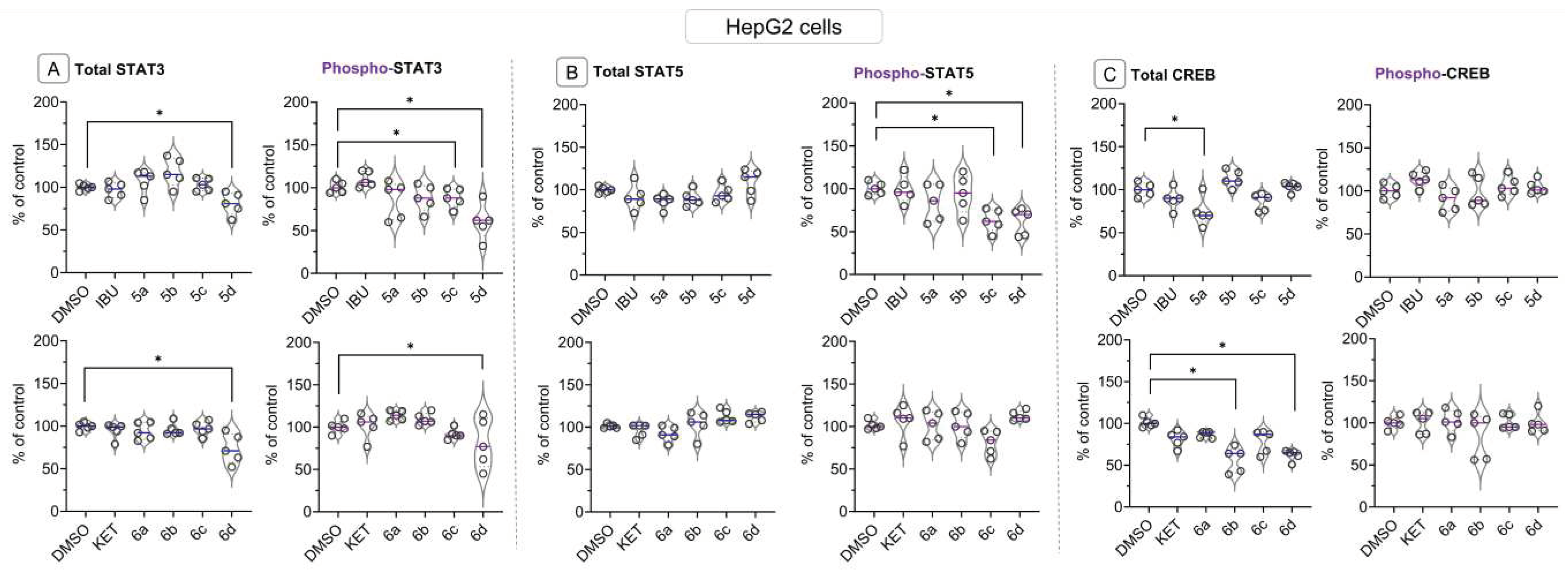

Table 1.
Primers used in R-T PCR.
| Gene | Forward primer | Reverse primer |
| Nrf2 | 5′ATTGCTACTAATCAGGCTCAG | 5′GTTTGGCTTCTGGACTTGG |
| NF-ĸB p50 | 5′ATCATCCACCTTCATTCTCAA | 5′AATCCTCCACCACATCTTCC |
| NF-ĸB p65 | 5′CGCCTGTCCTTTCTCATC | 5′ACCTCAATGTCCTCTTTCTG |
| COX-2 | 5′CCTGTGCCTGATGATTGC | 5′CAGCCCGTTGGTGAAAGC |
| PBGD | 5′TCAGATAGCATACAAGAGACC | 5′TGGAATGTTACGAGCAGTG |
| TBP | 5′GGCACCACTCCACTGTATC | 5′GGGATTATATTCGGCGTTTCG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



